regulation of heme pathway in regenerating mouse liver
TRANSCRIPT

Corap. Biochem. Physiol. Vol. 10lIB, No. I/2, pp. 243-246, 1992 0305-0491/92 $5.00 + 0.00 Printed in Great Britain © 1992 Pergamon Press pie
REGULATION OF HEME PATHWAY IN REGENERATING MOUSE LIVER
CESAR F. POLO, ELBA S. VAZQUEZ and ALCIRA M. DEL C. BATLLE*
Centro de Investigaciones sobre Porfirinas y Porfirias (CIPYP), CONICET y FCEN, UBA, Cindad Universitaria, Pabellon II, 2do Piso, 1428 Buenos Aires, Republica Argentina
(Received 20 May 1991)
Abstract--l. 6-Aminolevulinic acid synthetase (ALA-S), rhodanese and microsomai heme oxygenase (MHO), were quantitated in C14C induced regenerating mouse liver.
2. Maximal hepatomegalia was observed at 48 hr after i.p. injection of a single dose of the toxin. 3. ALA-S activity decreased on day 2, and then significantly increased (50%) between days 3 and 7,
returning afterwards to control values. 4. Cytoplasmic rhodanese, as well as MHO activities, exhibited a clear correlation as compared with
the ALA-S activity profile. 5. Porphyrin biosynthesis from precursor t~-aminolevulinic acid (ALA) was significantly increased even
after 15 days of intoxication. 6. Present results would indicate that C14C is acting in a dual fashion.
INTRODUCTION
Biochemical changes in the regenerating rat and mouse liver have been extensively investigated (von der Decken and Hultin, 1960; Presta et al., 1980; Srivastava et al., 1982; Roberts et al., 1983; Stout and Becker, 1986). Partial hepatectomy causes a signifi- cant decrease in the activity of fi-aminolevulinic acid synthetase (ALA-S), while that of heme oxygenase, the rate limiting enzyme in heme catabolism, is increased. These activities return to normal after 3-7 days (Srivastava et al., 1982; Stout and Becker, 1986).
One of the characteristics of the rat preneoplastic liver focus is its relative resistance to hepatotoxins (Hutterer et al., 1969; Farber et al., 1976; Eriksson et al., 1983). This feature would enable these cells to proliferate rapidly in response to chemically induced hepatocellular injury, and ultimately give rise to malignancy (Farber, 1984). This resistance arises partly from metabolic alterations that diminish the ceils capacity to activate xenobiotics to reactive species (Stout and Becker, 1978) and it is largely attributable to reduced availability of cytochrome P-450 (cyt P-450), the terminal electron aeceptor of the monooxygenase system (Oyanagui et aL, 1974; Cameron et aL, 1976; Okita et aL, 1976). It has also been demonstrated that spontaneous mouse liver tumors exhibit the same deficiency (Becker and Stout, 1984). Therefore, chemically induced and spon- taneous liver tumors share some metabolic alter- ations which likely represent intrinsic characteristics of the tumorigenic process and are independent of its etiology (Stout and Becker, 1986). Stout and Becker (1987) suggested that the decline in bemoprotein levels during hepatocarcinogenesis may result from a diminution of the intracellular heme pool, due
*Author to whom correspondence should be addressed at: Viamonte 1881, 10 ° "A", 1056 Buenos Aires, Repub- lica Argentina.
either to reduced heme synthesis or increased heme catabolism or both.
Because diminution of cyt P-450 has been a con- stant feature of preneoplastic liver lesions (Stout and Becker, 1986), and considering that initiation of chemical carcinogenesis is a muiti-step process requir- ing cell proliferation (Cayama et aL, 1978), we under- took a study to determine whether altered heme metabolism is the reason for cyt P-450 deficiency and whether this alteration occurs at the step of cell proliferation enhancement promoted by adminis- tration of a single dose of C14C. We quantitated ALA-S (EC 2.3.1.37), the rate limiting enzyme in the heme pathway, rhodanese (EC2.8.1.1), the enzyme proposed to modulate ALA-S activity, and micro- somal heme oxygenase (MHO) (EC 1.14.99.3), a degradative enzyme. We also measured the bio- synthesis of porphyrins in liver from 8-aminolevulinic acid (ALA) using the in vitro explant tissue culture method (Vazquez et al., 1987a).
MATERIALS AND METHODS
Animals CFI mice weighing 25-30 g were maintained in controlled
conditions and allowed free access to food (Purina 3) and water. Animals were given a single dose of C14C, i.p. (0.2ml/100 g, 1:1, v/v in corn oil) and were sacrificed 1-15 days later. Control animals were injected with corn oil. All animals were fasted 16 hr before death,
Homogenate preparation Mice were killed under ether anaesthesia. A fraction of
the liver (approximately one third) was cut up and immedi- ately homogenized (1:3, w/v) in a solution containing 0.9% NaC1, 0.1 mM Tris-HCl pH 7.4 and 0.5raM EDTA, for ALA-S determination. The remainder organ previously perfused with sterile ice cold saline was removed. Homogen- ates were prepared in ice cold 0.25 M sucrose for 10-15 sec in an Ultraturrax homogenizer. After differential centrifu- gation of the homogenate the resulting 18,000 g supernatant was used for measuring MHO activity and the pellet for
243

244 C-~at F. POLO et al.
measuring mitochondrial rhodanese activity previously solubilized in 0.05 M Tris-HC1 buffer, pH 8.7, containing 0.5 % Triton X- 100. An aliquot of the 18,000 g supernatant was centrifuged at 105,000 g and this supernatant was used for measuring the so-called cytosolic rhodanese activity.
Enzyme assays
ALA-S was measured as described by Marver et al. (1966), rhodanese by the method of S6rbo (1953) and MHO according to Yoshida and Kikuchi (1978). Protein concen- tration was determined by the method of Lowry et al. (1951).
Enzyme units (U) were defined as the amount of enzyme producing 1 nmol of product under the standard incubation conditions. Specific activity (SA) was expressed as U/mg protein.
Explant cultures
A fraction of the perfused liver (ca 50 mg) was placed on stainless steel supports in culture chambers. The explants were incubated at 37°C for 24 hr in darkness as previously described (Vazquez et aL, 1987a). Lactic acid dehydrogenase was measured according to Kornberg (1955).
Determination of porphyrins
Porphyrins were estimated fluorometrically according to Polo et al. (1988), expressed as fluorescence relative units (FRU)/mg tissue, and normalized taking into account the degree of lysis of the explant as described by Buzaleh et al. (1988). All measurements were made in the maintenance medium only.
RESULTS AND DISCUSSION
Clinical observations
Intoxicated animals showed general psychomotor depression immediately after C14C injection lasting for 2 hr, but nearly 10% of these animals died during the first 24 hr. Within 24 and 72 hr after intoxication, the liver showed hypopigmentated and necrotic zones, clearly observed by the naked eye. Maxima hepatomegalia was observed 48 hr after intoxication which reverted to normal values after 15 days (Fig. 1). Control animals showed no alterations.
Enzymic activities and in vitro biosynthesis o f porphyrins
ALA-S activity decreased on day 2 of intoxication and then significantly increased (50%) between days
510
o
I I I 0 5 10 15
Time (days)
Fig. 1. Effect of a single dose of C14C on the ratio of liver to body wts over a period of 15 days. Data shown are the average of three separate experiments run in triplicates and expressed as means + SD. Weight ratio means g of liver/100 g body. Other experimental conditions are as indi-
cated in the text.
200
~j 10C <
0 5 10 15 Time (days)
Fig. 2. Effect of a single dose of C14C on ALA-S activity over a period of 15 days. Data shown are the average of three separate experiments run in triplicates and are expressed relative to mean spec. act. of control animals. Mean control value: 0.147 + 0.016. Experimental conditions
are given in the text.
3 and 7, returning afterwards to control values (Fig. 2).
Cytoplasmic as well as mitochondrial rhodanese activities exhibited a sharp induction peak (more than 200%) between 24 and 48 hr after intoxication, recov- ering its basal level on day 7 (Fig. 3) Total rhodanese activity appeared diminished on day 3.
M H D activity was noticeably increased during the first 48 hr after C14C administration, but then de- creased on day 3 and reached its basal level on day 7 (Fig. 4).
It is interesting that a clear especular likeness correlation between the profile of ALA-$ activity and those exhibited by MHO and rhodanese was found.
It appears therefore that in this system, as it occurs in others previously described (Vazquez el aL, 1980, 1986, 1987b), rhodanese might exert a regulatory effect on ALA-S activity. We can further postulate that the cellular content of a thio compound which would actually regulate ALA-S activity (Wider et al.,
25¢
20C
15(
.>
10( <
50
I I I 5 10 15
Time (days)
Fig. 3. Effect of a single dose of C14C on cytoplasmic and mitochondrial rhodanese activities over a period of 15 days. Data shown are the average of three separate experiments run in triplicates and are expressed relative to mean spec. acts of control animals. (C)) Citoplasmic rhodanese; (0) mitochondrial rhodanese. Mean control values: 7.26 + 0.50 (cytoplasmic) and 4.60 + 0.25 (mitochondrial).
Experimental conditions are given in the text.
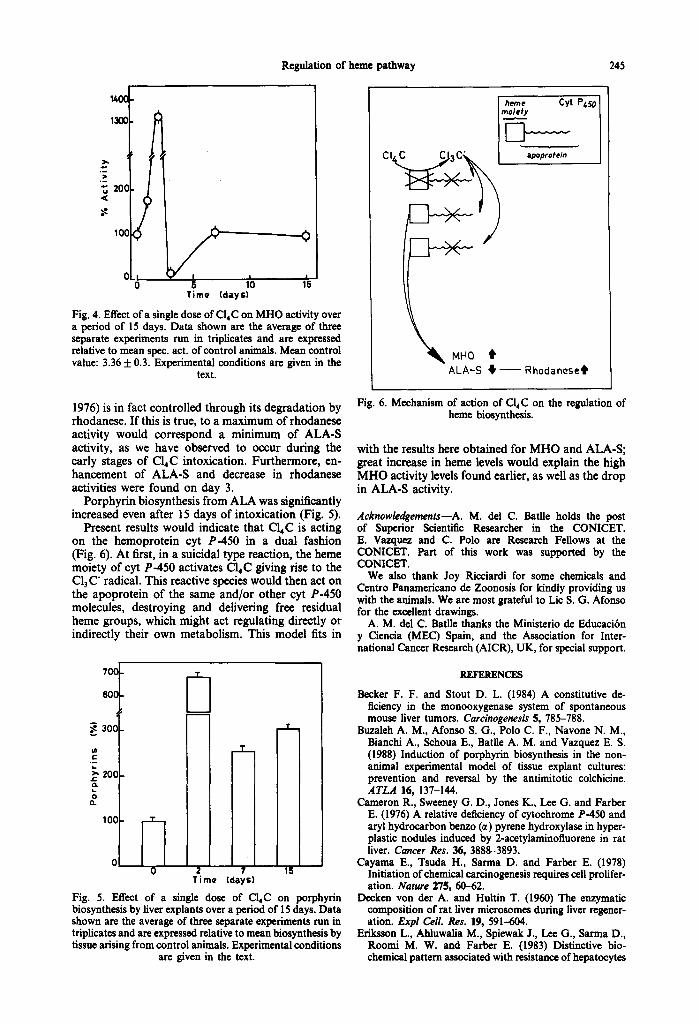
Regulation of heine pathway 245
140G
1300
20q < .<
10¢ --0
I I 5 10 15
Time (days)
Fig. 4. Effect of a single dose of Cl4C on MHO activity over a period of 15 days. Data shown are the average of three separate experiments run in triplicates and are expressed relative to mean spec. act. of control animals. Mean control value: 3.36 + 0.3. Experimental conditions are given in the
text.
I •
~ M H O 'I' ALA-S 4 Rhodaneset
1976) is in fact controlled through its degradation by rhodanese. If this is true, to a maximum of rhodanese activity would correspond a min imum of ALA-S activity, as we have observed to occur during the early stages of C14C intoxication. Furthermore, en- hancement of ALA-S and decrease in rhodanese activities were found on day 3.
Porphyrin biosynthesis from ALA was significantly increased even after 15 days of intoxication (Fig. 5).
Present results would indicate that C14C is acting on the hemoprotein cyt P-450 in a dual fashion (Fig. 6). At first, in a suicidal type reaction, the heme moiety of cyt P-450 activates C14C giving rise to the C13 C" radical. This reactive species would then act on the apoprotein of the same and/or other cyt P-450 molecules, destroying and delivering free residual heme groups, which might act regulating directly or indirectly their own metabolism. This model fits in
70(:
60C
.< 300
ot =..
t,.
zoo Q.
0 (3.
1011
Fig.
D T
T
0 2 7 15 Time (days)
5. Effect of a single dose of CI4C on porphyrin biosynthesis by liver explants over a period of 15 days. Data shown are the average of three separate experiments run in triplicates and are expressed relative to mean biosynthesis by tissue arising from control animals. Experimental conditions
are given in the text.
Fig. 6. Mechanism of action of C14C on the regulation of heine biosynthesis.
with the results here obtained for MHO and ALA-S; great increase in heme levels would explain the high M HO activity levels found earlier, as well as the drop in ALA-S activity.
Acknowledgements--A. M. del C. Batile holds the post of Superior Scientific Researcher in the CONICET. E. Vazquez and C. Polo are Research Fellows at the CONICET. Part of this work was supported by the CONICET.
We also thank Joy Ricciardi for some chemicals and Centro Panamericano de Zoonosis for kindly providing us with the animals. We are most grateful to Lic S. G. Afonso for the excellent drawings.
A. M. del C. Batile thanks the Ministerio de Educaci6n y Ciencia (MEC) Spain, and the Association for Inter- national Cancer Research (AICR), UK, for special support.
REFERENCES
Becker F. F. and Stout D. L. (1984) A constitutive de- ficiency in the monooxygenase system of spontaneous mouse fiver tumors. Carcinogenesis $, 785-788.
Buzaleh A. M., Afonso S. G., Polo C. F., Navone N. M., Bianchi A., Schoua E., BatHe A. M. and Vazquez E. S. (1988) Induction of porphyrin biosynthesis in the non- animal experimental model of tissue explant cultures: prevention and reversal by the antimitotic colchicine. ATLA 16, 137-144.
Cameron R., Swceney G. D., Jones K., Lee G. and Father E. (1976) A relative deficiency of cytochrome P-450 and aryl hydrocarbon benzo (~,) pyrene hydroxylase in hyper- plastic nodules induced by 2-acetylaminofluorene in rat liver. Cancer Res. 36, 3888-3893.
Cayama E., Tsoda H., Sarma D. and Farber E. (1978) Initiation of chemical carcinogenesis requires cell prolifer- ation. Nature 275, 60-62.
Decken yon der A. and Hultin T. (1960) The enzymatic composition of rat liver microsomes during liver regener- ation. Expl Cell. Res. 19, 591--604.
Eriksson L., Ahluwalia M., Spiewak J., Lee G., Sarma D., Roomi M. W. and Farber E. (1983) Distinctive bio- chemical pattern associated with resistance of hepatocytes

246 C"ESAR F. POLO et al.
in hepatocyte nodules during liver carcinogenesis. Environ. Health Perspect. 49, 171-174.
Farber E. (1984) Cellular biochemistry of the stepwise development of cancer with chemicals: G. H. A. Clowes memorial lecture. Cancer Res. 44, 5463-5474.
Farber E., Parker S. and Gruenstein M. (1976) The resist- ance of putative premalignant liver cell populations, hyperplastic nodules, to the acute cytotoxic effects of some hepatocarcinogens. Cancer Res. 36, 3879-3887.
Hutterer F., Klion F. M., Wengraf A., Schaffner F. and Popper H. (1969) Hepatocellular adaptation and injury. Structural and biochemical changes following dieldrin and methyl butter yellow. Lab. Invest. 2, 455-464.
Kornberg A. (1955) Lactic dehydrogenase of muscle. In Methods in Enzymology (Edited by Colowick S. and Kaplar N.), Vol. I, pp. 441-443. Academic Press, New York.
Lowry O. H., Rosebrough N. J., Farr A. L. and Randall R. J. (1951) Protein measurement with the Folin phenol reagent. J. biol. Chem. 193, 265-275.
Marver H,, Tschudy D., Perlroth M. and Collins A. (1966) 6-Aminolevulinic acid synthetase I. Studies in liver hom- ogenates. J. biol. Chem. 241, 2803-2809.
Okita K., Noda K., Fujumoto Y. and Takemoto T. (1976) Cytochrome P-450 in hyperplastic nodules during hepato- carcinogenesis with N-2-fluorenyl-acetamide in rats. Gann 67, 899-902.
Oyanagui Y., Sato N. and Hagihara B. (1974) Spectro- photometric analysis of cytochromes in rat liver during carcinogenesis. Cancer Res. 34, 458-462.
Polo C. F., Frisardi A. L., Resnik E., Schoua E. and Batlle A. M. (1988) Factors influencing free porphyrins fluor- escence spectra. Clin. Chem. 34, 757-760.
Presta M., Aletti M. G. and Ragnotti G. (1980) Decrease of the activity of the mixed function oxidase system in regenerating rat liver. An alternative explanation. Biochem. biophys. Res. Commun. 95, 829-834.
Roberts E., Ahluwalia M. B., Lee C., Chan C., Sarma D. S. R. and Farber E. (1983) Resistance to hepatotoxins acquired by hepatocytes during liver regeneration. Cancer Res. 43, 28-34.
S6rbo B. H. (1953) Crystalline rhodanese. I. Purification and physico-chemical examination. Acta chem. scand. 7, I 129-1136.
Srivastava R. C., Dwivedi R. S., Kaur G. and Srivastava R. (1982) Heme- and drug metabolizing enzymes in regener- ating rat liver. Br. J. exp. Patrol. 63, 1-4.
Stout D. L. and Becker F. F. (1978) Alteration of the ability of liver mierosomes to activate N-2-fluorenylacetamide to a mutagen of Salmonella typhimurium during hepato- carcinogenesis. Cancer Res. 38, 2274-2278.
Stout D. L. and Becker F. F. (1986) Heine enzyme pattern in genetically and chemically induced mouse liver tumors. Cancer Res. 46, 2756-2759.
Stout D. L. and Ikcker F. F. (1987) Heme enzyme pattern in rat liver nodules and tumors. Cancer Res. 47, 963-966.
Vazquez E., Afonso S., Buzaleh A. M., Navone N., Polo C., Schoua E. and Batlle A. M. (1987a) The effect of griseo- fulvin on the heme pathway. Studies on tissue explant cultures. Toxicology 46, 73-82.
Vazquez E., Buzaleh A. M., Wider E. and Batlle A. M. (1987b) Porphyrin biosynthesis in Rp. palustris. X. Purifi- cation and some properties of rhodanese. Int. J. Biochem. 19, 1193-1197.
Vazquez E., Buzaleh A. M., Wider E. and Batlle A. M. (1986) Reduced thiosulphate alfinity for erythrocyte rhodanese: an explanation for increased 6-aminolevulinic synthetase activity in some porphyrias? IRCS Med. Sci. 14, 999-1000.
Vazquez E., Wider E. and Baffle A. M. (1980) Porphyrin biosynthesis in the soybean callus tissue system XVIII. Levels of succinyl CoA synthetase, cystathionase, rhodanese, 6-aminolevulinate synthetase and t~-amino- levulinate dehydratase in clones of different ages. Int. J. Biochem. 12, 721-724.
Wider E., Sandy J., Davies R. and Neuberger A. (1976) Control of 5-aminolevulinate synthetase activity in Rh. spheroides. Phil. Trans. R. Soc. Lond. B 273, 79-98.
Yoshida T. and Kikuchi G. (1978) Purification and proper- ties of heme oxygenase from pig spleen microsomes. J. biol. Chem. 253, 4224--4229.