relation entre les deux souches bactériennes principales d ...composant de fertilisants agricoles...
TRANSCRIPT

i
UniversitéduQuébec
INRS-InstitutArmand-Frappier
Relationentrelesdeuxsouchesbactériennesprincipales
d’unbiofilmdénitrifiant
Par
JonathanCloutier
Mémoireprésentépourl’obtentiondugrade
Maîtredessciences(M.Sc.)enmicrobiologieappliquée
Juryd’évaluation
Présidentdujuryetexaminateurinterne
Examinateurexterne
Directeurderecherce
ÉricDéziel
INRS-InstitutArmand-Frappier
FranceDaigleUniversitédeMontréal
RichardVillemurINRS-InstitutArmand-Frappier

ii
Résumé
Lorsd’étudesportantsurunbiofilmprovenantd'unbioprocédéde
dénitrification,ilaétédéterminéqu’ilyavaitdeuxsouchesbactériennesquiétaient
lesplusabondantesentermedeproportiondelabiomasse.Cesdeuxsouches,NL23
etJAM1,ontéventuellementétédésignéescommenouvellesespècesbactériennes
HyphomicrobiumnitrativoransNL23etMethylophaganitratireduscenticrescens
JAM1.TandisquelasoucheNL23estenmesured'effectuertouteslesétapesde
dénitrification,lasoucheJAM1estincapablederéduirelenitriteetdoncnepeutpas
réduirecomplètementlenitrateenazotegazeux.Plusieursfacteursdansleur
relationsuggèrentunecertainecoopération.Cemémoireprésentelestravaux
effectuésafind’enapprendredavantagesurl’interactionentrecesdeuxbactéries
Dansunpremiertemps,desconditionsoptimalesdeco-culturesentrelesdeux
souchesontétédéveloppées.Lacaractérisationdeladynamiquedeconsommation
dunitrateetdunitriteadémontréquelesco-culturesdessouchesNL23etJAM1
n’étaientpasnécessairementplusrapidespoureffectuerladénitrificationquela
soucheNL23enculturepure.Parcontre,desco-culturesdelasoucheNL23avecun
mutantdelasoucheJAM1(inactifpourréduirelenitrate)ontdémontréune
dénitrificationplusrapidequelasoucheNL23enculturepure.
LarechercheaaussipermisdedémontrerquelasoucheJAM1apporteune
aidedecroissanceàlasoucheNL23enconditionsaérobiesetuneaideenréduction
denitrateenconditionsanaérobies.Parcontre,iln’apasétédémontréquela
soucheNL23procureunavantageàlasoucheJAM1.

iii
Remerciements
J’aimeraisremerciermondirecteurderecherche,RichardVillemur,de
m’avoirpermisdecomplétermamaîtrisedanssonlaboratoire,luiquiatoujoursété
disponiblepouroffrirsonaideetsesconseils.Sanssapatience,sacompréhension
etsagénérositéjen’auraispaspufinirleprojet.
J’aimeraisaussiremerciertouslesmembresdulaboratoiredeRichard
Villemurpourleuraide,enparticulierChristineMartineau,poursonaide
indispensabledansmesdébutsnerveux;KarlaVasquezpoursonsoutien,son
expérienceetsaluttepourl’entretienduIC;etFlorianMauffrey,poursesconseils
(etsesmutants).MerciaussiàGeneviève,Marc-AndréetValérie,ainsiqu’aux
membresduGRMEquiontrendul’expérienceencoreplusagréable.
Sanslesoutienmoraldemafamilleetdemesamis,cetteentrepriseaurait
étéimpossible.Merciàmesparents,àmasœuretàtouslesgensquiontfaitpartie
demaviedurantcettepériode.

iv
Tabledesmatières
1. Introduction..............................................................................................12. Revuedelittérature...............................................................................42.1. Dénitrification..........................................................................................................................................42.2. Biofilms.......................................................................................................................................................72.3. Osmoprotection.......................................................................................................................................82.4. Bioprocédés...............................................................................................................................................92.5. BiodômedeMontréal..........................................................................................................................102.6. Methylophaganitratireduscenticrescens.....................................................................................132.7. Hyphomicrobiumnitrativorans.......................................................................................................142.8. Co-cultures,partagederessources...............................................................................................152.9. Hypothèseetobjectifs........................................................................................................................17
3. Matérieletméthodes...........................................................................183.1. Souchesbactériennes..........................................................................................................................183.1.1. Mutants.............................................................................................................................................18
3.2. Culturedesbactéries...........................................................................................................................193.2.1. MilieuoptimaldelasoucheJAM1.........................................................................................193.2.2. MilieuoptimaldelasoucheNL23.........................................................................................213.2.3. Milieudecroissanceadaptéàlaco-culture......................................................................223.2.4. Milieuxsousformedegéloses................................................................................................23
3.3. Préparationdescultures...................................................................................................................233.3.1. Ensemencement............................................................................................................................243.3.2. Conditionsdeculture..................................................................................................................24
3.4. Récoltededonnées..............................................................................................................................253.4.1. Spectrophotométrie....................................................................................................................263.4.2. Chromatographieionique.........................................................................................................263.4.3. Extractiond’ADN..........................................................................................................................263.4.4. qPCR...................................................................................................................................................27
4. Résultats...................................................................................................304.1. Déterminationdesconditionsoptimalesdeculture.............................................................304.1.1. Suividecroissancepréliminaireenconditionsaérobies...........................................304.1.2. Suividecroissanceenconditionsanaérobies..................................................................314.1.3. Variationsproblématiques.......................................................................................................404.1.4. Réductiondelaquantitéinitialedenitrate......................................................................46
4.2. CaractérisationdelarelationentrelessouchesNL23etJAM1........................................494.2.1. Croissancedeculturespuresenprésencedesurnageant..........................................494.2.2. Recherched’unosmoprotecteur...........................................................................................534.2.3. Caractérisationapprofondiedelaconsommationdunitrate/nitriteàl’aidedemutantsdelasoucheJAM1......................................................................................................................57
5. Discussion................................................................................................645.1. Paramètresoptimauxpourlacroissancedesco-culturesdessouchesNL23etJAM1 645.2. Profildeconsommationdenitrateetdenitritelorsdeladénitrification...................665.3. LessouchesNL23etJAM1enbiofilm..........................................................................................70

v
5.4. Échangederessources.......................................................................................................................73
6. Conclusion................................................................................................75

vi
Listedestableaux
Tableau3.1:compositiondumélangedanschaqueréactiondeqPCR.28Tableau3.2:détailssurlatechniqueduqPCR.................................................29Tableau4.1:concentrationdeNaClpartubedanslarecherched’un
osmoprotecteur..............................................................................................................54

vii
ListedesfiguresFigure2.1:Représentationschématiquesimplifiéeducycledel’azote..............................................................4
Figure2.2:Voiemétaboliquedeladénitrification........................................................................................................6
Figure2.3:BioréacteurdubiodômedeMontréal.......................................................................................................11
Figure3.1:LessouchesNL23etJAM1.............................................................................................................................15
Figure3.2:Exempledecultureenanaérobiose...........................................................................................................25
Figure4.1:Croissanceenconditionsaérobies(InstantOcean15g/L).............................................................31
Figure4.2:Lacroissancedesco-culturesdessouchesNL23etJAM1enconditionsanaérobies...........33
Figure4.3:Consommationdenitratedesco-culturesdessouchesNL23etJAM1enconditionsanaérobies.....................................................................................................................................................................................35
Figure4.4:concentrationdenitritedanslesco-culturesdessoucheNL23etJAM1enconditionsanaérobies.....................................................................................................................................................................................36
Figure4.5:Concentrationd’azotetotaledanslesco-culturesdessouchesNL23etJAM1enconditionsanaérobie........................................................................................................................................................................................38
Figure4.6:Consommationdenitratepourchaqueréplicatdesco-cultures1:5........................................40
Figure4.7:Concentrationdenitritepourchaqueréplicatdesco-cultures1:5...........................................41
Figure4.8:Concentrationd’azotetotalpourchaqueréplicatdesco-cultures1:5.....................................41
Figure4.9:Consommationdenitratepourchaqueréplicatdesco-cultures1:10......................................42
Figure4.10:Concentrationdenitritepourchaqueréplicatdesco-cultures1:10......................................43
Figure4.11:Concentrationd’azotetotalpourchaqueréplicatdesco-cultures1:10...............................43
Figure4.12:QuantitésrelativesdesgènesnapAetnarG1danslesco-cultures1:5..................................44
Figure4.13:QuantitésrelativesdesgènesnapAetnarG1danslesco-cultures1:10................................45
Figure4.14:Consommationdenitratedesco-culturesdessoucheNL23etJAM1enconditionsanaérobies.....................................................................................................................................................................................47
Figure4.15:Concentrationdenitritedanslesco-culturesdessoucheNL23etJAM1enanaérobie....47
Figure4.16:Concentrationd’azotetotaldanslesco-culturesdessouchesNL23etJAM1enconditionsanaérobies.....................................................................................................................................................................................48
Figure4.17:CroissanceenconditionsaérobiesdelasoucheNL23enprésencedesurnageant...........50
Figure4.18:CroissanceenconditionsaérobiesdelasoucheJAM1enprésencedesurnageant...........52
Figure4.19:CroissancedelasoucheNL23enprésencedemétabolitesdelasoucheJAM1(0%deNaCl)................................................................................................................................................................................................55
Figure4.20:CroissancedelasoucheNL23enprésencedemétabolitesdelasoucheJAM1(1%deNaCl)................................................................................................................................................................................................55
Figure4.21:CroissancedelasoucheNL23enprésencedemétabolitesdelasoucheJAM1(1,5%deNaCl)................................................................................................................................................................................................56
Figure4.22:CroissancedelasoucheNL23enprésencedemétabolitesdelasoucheJAM1(2%deNaCl)................................................................................................................................................................................................56
Figure4.23:Consommationdunitrateparlesco-culturesdessouchesJAM1non-mutantesoumutantesaveclasoucheNL23(conditionsanaérobies)..........................................................................................59

viii
Figure4.24:Concentrationdenitritedanslesco-culturesdessouchesJAM1non-mutantesoumutantesaveclasoucheNL23(conditionsanaérobies)..........................................................................................60
Figure4.25:Concentrationd’azotetotaldanslesco-culturesdessouchesJAM1non-mutantesoumutantesaveclasoucheNL23(conditionsanaérobies)..........................................................................................61
Figure5.1:ProfildeconsommationdunitrateetdunitriteparlasoucheNL23lorsdeladénitrification..............................................................................................................................................................................66
Figure5.2:ProfildeconsommationdunitrateetdunitriteparlasoucheJAM1lorsdeladénitrification..............................................................................................................................................................................67
Figure5.3:Profildeconsommationdunitrateetdunitriteparuneco-culturedessouchesNL23etJAM1lorsdeladénitrification.............................................................................................................................................67
Figure5.4:Profildeconsommationdunitrateetdunitriteparuneco-culturedesespècesBacillussubtilisetPseudomonasdenitrificanslorsdeladénitrification.............................................................................68
Figure5.5:Supportenplastiquegarnid’unbiofilm...................................................................................................71
Figure7.1:Schémadelaméthodedecréationd’unbiofilm...................................................................................77

ix
Listedessiglesetdesabbréviations˚C:degrésCelsius
ADN:acidedésoxyribonucléique
DO600:densitéoptiqueàunelongueurd’ondede600nm
EPS:extracellularpolymericsubstances
FISH:fluorescentin-situhybridization
TSA:trypticasesoyagar
R2A:Reasoner’s2Aagar
ATCC:americantypeculturecollection
Pb:pairesdebases
FAM:carboxylfluorescein
BHQ:blackholequencher
rpm:rotationsparminute

x

1
1. Introduction
Lenitratereprésenteunpolluantpotentieldanslecontexte
d’environnementsaquatiquesnaturelsousimulés.Lamoléculeesttoxiquepourles
animauxàdesconcentrationsexcédantenviron30ppm(0,49mMd’azote).La
limiteimposéeàl’eaupotableauCanadaetauxÉtats-Unisestde10mg/Ld’azote-
nitrate(45mg/Ldenitrate),concentrationàlaquellel’eauestconsidérée
définitivementsécuritairepourleshumainslesplusvulnérables.Lenitriteest
encoreplustoxique:quelquesmilligrammesparlitresontsuffisantspourla
manifestationdeproblèmeschezlespoissonsetlalimiteacceptabledansl’eau
potableestde1mg/Ld’azotenitrite(3mg/Ldenitrite).Lenitrateestsouventun
composantdefertilisantsagricolesets’accumuledoncàhautesconcentrationsà
l’embouchuredecoursd’eau.Outreleseffetsdirectssurlasantédespopulations
humainesetanimales,ilpeutyavoirproliférationd’algues,hypoxieetautreseffets
reliéscausantdeschangementsnocifsàl’écosystèmedumilieu(Camargoetal.,
2005).Dansunsystèmeaquatiquefermé,commeunaquarium,l’accumulationdu
nitratepeutseproduiredemanièreinexorablepuisquel’eaudumilieuestrecyclée
etquelesorganismesvivantsensontdessourcescontinuelles.Unesolutionnon-
biologiqueestleremplacementtotaldel’eau,cequipeutserévélerêtredispendieux
etpeupratique.Unesolutionpotentielleestladénitrificationbiologiquepardes
organismesbactériens.
UndesécosystèmesenexhibitionauBiodômedeMontréalestlemésocosme
dufleuveSaint-Laurent,quicomporteungrandaquariummarindanslequelvivent
plusieursespècesanimalesoriginairedel’estuairedufleuveSaint-Laurent.
L’accumulationdunitrateétantproblématique,unbioréacteurfutinstallé,dans
l’espoirquedesespècesbactériennesdénitrifiantespuissentréduirelamolécule
toxiqueenazotegazeux.Cebioréacteurétaitcomposéd’unmécanismesupportant
lacroissanced’unbiofilmexhibantdel’activitédénitrifiante;cebiofilmestétudié
pardesmembresdulaboratoireduPr.RichardVillemurdepuis2000.Plusieurs

2
souchesbactériennesdistinctesenontétéisolées,etplusieursorganismesnon-
bactérienssontnotésd’ycohabiter.Deuxsouchesbactériennes,cependant,
composentlagrandemajoritédelacommunautébiologiquedubiofilm,entermes
debiomasseetd’activitédénitrifiante:lessouchesJAM1etNL23,touteslesdeux
méthylotrophesetanaérobesfacultatives.Depuisleurdécouverteetleur
caractérisationcommedesespècesdistinctes,ellessontdénomméesMethylophaga
nitratireducenticrescensetHyphomicrobiumnitrativorans,respectivement.
Ladénitrificationestunprocessusderéductiondunitrateetdecomposés
intermédiairesenquatreétapesprincipales.Lenitrateestd’abordréduitennitrite,
quiestréduitenacidenitrique,etensuiteenacidenitreuxpourfinalementproduire
del’azotegazeux.Àchacunedesétapesestassociéuneenzyme.LasoucheJAM1
n’effectuequetroisdesquatreétapesdedénitrification,nepossédantpasdegène
codantpourlanitriteréductase.Elleréduitlenitratedefaçonefficacegrâceàdeux
nitrateréductasesmembranaires,Nar1etNar2.Lasoucheestaussitoléranteaux
conditionssalées,exhibantunecroissancenormaledansunmilieucontenant
jusqu’à8%deNaCl.LasoucheNL23,quantàelle,effectuetouteslesétapesdela
dénitrification.Toutefois,sanitrateréductasepériplasmiqueexécutelaréduction
dunitratebeaucoupmoinsrapidementqueleduod’enzymesdelasoucheJAM1.
Contrairementàcettedernière,lasoucheNL23netolèrepasleseldanssonmilieu,
etuneconcentrationdeplusde1%NaClinhibesacroissance.Notamment,ceci
sembleparticuliercarlemilieudanslequelétaitcultivélebiofilmétaitmarinet
incluaitprèsde3%desel.
Puisquecesdeuxsouchesprincipalesétaientdeloinlesplusabondanteset
activesdubiofilm,ilestnaturellementintéressantdelesétudierenplusdedétails
defaçonindividuelle.Parcontre,unbiofilmestunecommunautécomplexede
microorganismes,etlessouchesNL23etJAM1interagissentdoncpresque
certainementàuncertainniveau.Ilestdoncégalementintéressantd’étudierla
relationentrelesdeuxsouches,spécialementlorsdeladénitrification.Ceprojetde
recherchetentededéterminers’ilexisteunecoopérationentrelesdeuxespèces

3
principalesdubiofilm.Cetterechercheserasurtoutaxéesurlepartagedesoxydes
d’azotemaisviseraégalementdetrouverd’autresélémentspouvantcontribuerà
unesynergie,telsquedesfacteursdecroissance,desmoléculesessentielles(ex:
vitamineB12),etc.

4
2. Revuedelittérature
2.1. Dénitrification
Leprocessusdedénitrificationestunedescomposantesmajeuresducycle
del’azote,oùl’azotecirculedansl’atmosphère,lesoletl’eausousdiversesformes
moléculaires.L’importancedecettecirculationprovientengrandepartiedela
dépendanced’organismesvivantsàl’azotepourlastructureetlaformationde
moléculesprotéiquesetnucléiques.Eneffet,aucunevieterrestrenepeutexister
sanscetélément,etlesorganismesexistantauseindemilieuxpauvresenazote
doiventdévelopperdesstratégiesspécialesdesurvie.
Figure2.1:Représentationschématiquesimplifiéeducycledel’azote Lecycledel’azoteestunsystèmecomplexederéactionschimiquesengrandepartiecatalyséespardesorganismesmicrobiens.(1)Fixationd’azote;(2)nitrification;(3)dénitrification;(4)co-dénitrification;(5)anammox;(6)l’oxydationd’ammoniacdurantlanitrification.(Hayatsuetal.,2008)

5
Ladénitrificationréfèrespécifiquementàtouteslesétapesdelaréductiondu
nitrateenazote(Fig.2.2).Danslaplupartdescas,cetteréductionestexécutéepar
desespècesbactériennesprofitantdupotentielénergétiquedesréactionschimiques
impliquées;leprocessusréduitséquentiellementlenitrateennitrite,enoxyde
nitrique,enoxydenitreuxetfinalementenazotegazeux,chaqueétapeoffrantun
avantageénergétiquedifférent.Chacunedesréactionsestcatalyséeparuneenzyme
spécifique:
- Lanitrateréductasecatalyselaréductiondunitrateennitrite,cequioffrele
plusd’énergiepotentielleparmilesétapesdeladénitrification.Deux
versionsdissimilatoiresdecetteenzymeexistent:uneversionmembranaire,
codéeparlesgènesnar,etuneversionpériplasmique,codéeparlesgènes
nap.Ilexisteégalementdesvariétésdenitrateréductasesassimilatoires.
(Zumft,1997)
- Lanitriteréductaseàsontourpermetlaréductiondunitriteenoxyde
nitrique,lapremièreformegazeusedesmoléculesazotéesdela
dénitrification.Ilyaaussideuxversionsdecetteenzyme:Cu-Nir,qui
contientunatomedecuivre,estcodéparlegènenirK;cd1-Nir,quicontient
uncytochromecd1,estcodéparlegènenirS.Cesdeuxvariétésd’enzymes
n’ontjamaisétéretrouvéesensembledanslamêmeespèceetaucuneétude
n’apurévélerunedifférencefonctionnelleentreelles.(Priemeetal.,2002,
Zumft,1997)
- L’oxydenitriqueréductasecatalyselaréductiondel’oxydenitriqueenoxyde
nitreux.Laversiondel’enzymequiestprésentedanslesbactéries
dénitrifiantesestcNor,quicontientuncytochromecetestcodéparlesgènes
nor(unitécatalytiquecodéeparnorB).Ilexisteégalementd’autresversions
del’oxydenitriqueréductasedanscertainesespècesbactériennesquifont
partiedusystèmedeleursdéfensescontrelatoxicitédel’oxydenitrique.
(Hinoetal.,2012)

6
- L’oxydenitreuxréductasecatalyselaréductiondel’oxydenitreuxenazote
gazeux.Cetteenzymecontientunatomedecuivreetestcodéeparlesgènes
nos(unitécatalytiquecodéeparlegènenosZ).(Pauletaetal.,2013)
Figure2.2:Voiemétaboliquedeladénitrification Lavoiecomplètededénitrificationtransformelenitrateenazotegazeuxparunesériederéactionschimiquescatalyséespardesenzymes(surlignées).Lesmoléculessontlenitrate(NO3),lenitrite(NO2),l’oxydenitrique(NO),l’oxydenitreux(N2O)etl’azotegazeux(N2).
Cesenzymespeuventsemanifestersousdesformesdifférentesetavoirdes
activitésvariablesselonl’espèce.Puisqu’elleoffreunavantageénergétique
inférieuràlarespirationaérobie,ladénitrificationestprincipalementutilisépardes
espècesstrictementanaérobiesoucommealternativepardesbactériesanaérobies
facultatives.Seuleunefractiond’espècesbactériennespossèdelacapacité
d’effectuerladénitrificationetcelles-cin’eneffectuentpasnécessairementtoutes
lesétapeselles-mêmes(Zumft,1997).Plusieursespècesbactériennessecontentent
d’effectuerseulementlarespirationdunitrate,tellequeSerratiamarcescens,quine
peutproduiredel’oxydenitriquequeparlaréductionnon-enzymatiquedunitrite
(Andersonetal.,1986).Celles-ciconstituentleplusimportantsous-groupede
bactériesprofitantdelaréductiond’oxydesd’azote(Gregoryetal.,2003).D’autres
espècesseserventplutôtd’enzymesderéductioncommemécanismededéfense
contrelesmoléculestoxiques.L’enzymeqNorestutilisépardesbactériescomme
Neisseriagonorrhoeaepoursedébarrasserd’oxydenitrique(Hinoetal.,2012).De

7
plus,certainesbactériesréduisentdesmoléculesintermédiairesdeladénitrification
enconditionsaérobies.Parexemple,l’espèceParacoccuspantatrophus
(précédemmentnomméeThisphaerapantatropha)estenmesuredeperformerune
dénitrificationcomplèteenprésenced’oxygène.Ilestpossiblequecetteactivité
permetteàlabactériededissiperlesmoléculesréductricesetretrouverun
équilibreredox.(Berksetal.,1995)
2.2. BiofilmsDanslanature,lavastemajoritédelaviemicrobienneexistesousformede
biofilm,etnondiffuseenmilieuliquide.Unbiofilmestcomposéd’organismesde
classificationstaxonomiquesvariéesainsiqued’unematricecomplexede
polymèresbiologiquesgénéralementconnussouslenom«substances
polymériquesextracellulaires»(enAnglais,EPS).Celles-cisontproduites
directementparlesorganismesvivantdanslebiofilmousontmodifiéespardes
enzymesproduitesparcesorganismes.Cependant,lacompositionexactedela
structured’unbiofilmestdifficileàdéterminer.LesEPSsontintimement
impliquéesdanslaviemicrobienned’unbiofilm,etleursfonctionssontmultiples.
LesEPSpeuventagircommesourcedenourriture,faciliterl’échangegénétique
entrebactéries,faciliterlarétentiond’eau,fourniruneprotectioncontredes
facteursantibactériensetbiend’autreschoses.L’architectured’unbiofilmestaussi
hautementvariéeetdépenddel’organisationdesEPS.Ilpeutyavoirdescanauxet
descompartiments,etlaformedubiofilmpeutvarierselonlasituation.Unedes
composantesessentiellesàlaformationd’unbiofilmestlagammedemolécules
(protéines,ADN,polysaccaridesetmoléculesamphiphiles)quiserventàl’adhésion
debactériescolonisatricesàunesurface.Eneffet,lapremièreétapedansla
formationd’unbiofilmestl’attachementàunesurface,suividelaformationd’une
microcoloniepardivisioncellulaire.Cen’estqu’aprèsl’adhésionoriginalequeles
autrescomposantesdelamatricedubiofilmsontproduitesetqued’autresespèces
bactériennesysontrecrutées.Sanslaprésenced’unesurfacepropice,lesbactéries

8
nepeuventpasformerdebiofilm,etrestentenphaseplanctoniqueous’agrègenten
d’autresformes(ex:floc).(Flemmingetal.,2010)
Ilestaussipossiblequ’unbiofilmsoitnécessairepourlafonctionde
certainessubstancesrelâchéespardesbactéries.Ilaétédocumentéquecertaines
souchesbactérienneshalophilespouvaientrelâcherdesparticulesosmoprotectrices
(ectoïne,glutamate)dansleurmilieupouraideràlasurviedebactériesdansleur
entourage.(Plakunovetal.,2008)
2.3. OsmoprotectionLorsqu’ellessontconfrontéesàdesfluctuationsdel’osmolaritédedeleur
milieu(comme,parexemple,uneaugmentationensel),lesbactériesdoivent
équilibrerlesgradientsosmotiquescrééspoursurvivre.Pouratteindrecetobjectif,
ellespeuventproduiredesosmoprotecteurs(ousolutéscompatibles)oulesobtenir
dansleurenvironnement.Unosmoprotecteurestunemoléculeavecunpotentiel
osmotiqueélevéquin’interfèrepasaveclesprocessusmétaboliquesd’unecellule,
mêmeenhautesconcentrations.Enl’accumulantdanssoncytoplasme,unebactérie
peutréduirelapressionosmotiqueentrelesdeuxcôtésdesamembrane.(Kempfet
al.,1998)
Ilexisteunevariétéd’osmoprotecteurs.Ceux-ciincluent:desdiholosides
commelesaccharoseetletréhalose;despolyolscommeleglycérol;ainsiquedes
acidesaminés(etdérivés)commelaproline,lesbétaïnes,lacarnitineetl’ectoïne
(Kempfetal.,1998).Plusieursbactériesontdessystèmescomplexesenplacepour
lerèglementfindeleurréponseàunstressosmotique.L’espècebactérienne
Bacillussubtilis,parexemple,faitlasynthèsedelaprolinepourseprotéger,mais
ellepossèdeaussiuntransporteurspécifiqueàlacapturedeprolinedans
l’environnement.Ellepeutaussicapturerdespeptidesetenextrairedelaproline
avecdesprotéases,unestratégiequiestadoptéeparplusieursautresespèces
également(ex:Listeriamonocytogenes,Lactobacilluscasei,Oenococcus

9
oeni)(Zaprasisetal.,2013).Deplus,certainesbactériespeuventfaireusagede
plusieursosmoprotecteursdifférents.Escherichiacoliestcapabledesynthétiserle
tréhaloseetlaglycinebétaïne,ainsiqued’utiliserlaprolineetl’ectoïneretrouvées
danssonenvironnement(Jebbaretal.,1992).L’ectoïne,dérivédela
tetrahydropyrimidine,estl’osmoprotecteurquelasoucheJAM1semblepouvoir
produire(voirsection2.6).Cetosmoprotecteurestparticulièrementintéressant
danslecadredescommunautésbactériennestolérantesauselcarilaétédémontré
quedesbactériesnon-halophilespeuventenfaireusagepourobtenirunetolérance
aupressionsosmotiques(Jebbaretal.,1992).
2.4. BioprocédésUnbioprocédéestuneapplicationdescapacitésdemicroorganismespourla
productiondebiensoupourd’autresprocessusutiles.Cesapplications
biotechnologiquessontcourammentutiliséesdansunepanopliededomaines,
incluantlaproductionalimentaire,laproductiondemoléculespourutilisation
pharmaceutique,etc.
Lapurificationd’eauuséeimpliquesouventdesbioprocédés,dontles
systèmesdeboueactivée,quidépendentdel’activitédebiomasseenflocdansdes
réservoirsaéréspourpurifierl’eauquis’yretrouve(Roediger,1983).Les
bioréacteursmembranaires(«Membranebioreactor»enAnglais,ouMBR)
combinentunsystèmedeboueactivéeavecuneultrafiltrationpourpurifierdel’eau.
Cettetechnologiereprésenteunmarchéévaluéàdescentainesdemillionsde
dollarsetresteunsujetderechercheactif(Le-Clechetal.,2006,Wangetal.,2016).
Labiomasseutiliséedansunbioprocédépeutaussiêtrecultivéesousformede
biofilmfixéàunsupportàl’aided’unbioréacteuràlitfluidisé,commedanslecas
d’untraitementdelixiviatdansunedécharged’ordures(Hajipouretal.,2011)oude
dégradationdesubstanceshuileusesparunbiofilmfongique(Pakulaetal.,1996).
Cependant,lesbiofilmspeuventaussiêtreproblématiquesdanslesbioprocédés.
DansunMBR,laformationd’unbiofilmsurlamembranedefiltrationmèneàla

10
dégradationdecelle-ci.Ceciestconsidérécommeundesproblèmesprincipaux
dansledéveloppementdesMBR(Le-Clechetal.,2006).
Ladénitrificationpeutelleaussiêtreexploitéecommebioprocédé.Lenitrate
etlesautrescomposésintermédiairesdeladénitrificationreprésententtousdes
dangerspotentielspourl’environnementainsiquepourlasantéd’organismes
vivants,incluantleshumains.Pourcetteraison,ledéveloppementdeméthodesde
dénitrificationenbioréacteuresttrèsactif.Undomained’attentionparticulièreest
letraitementd’eaupolluéeparl’industrieagroalimentaire.Plusieursétudes
récentesontinvestiguél’efficacitédebioréacteursavecdifférentessourcesde
carbonesetunevariétédeparamètres.Lesrésultatsmontrentengénéralune
améliorationpotentielleaveccestechniquesparrapportsauxtechniquesde
traitementtraditionnelles(créationdezoneshumides,constructiondefossés
écologiques,etc)(Liangetal.,2015).
2.5. BiodômedeMontréalLeBiodômedeMontréalainstalléen1998unbioréacteuràlitfluidisépour
remédieràunniveaudangereusementélevédenitrate(environ15mM)dansl’eau
deleuraquariumd’eaumarine(Fig.2.3).Àl’intérieurduréacteurseretrouvaitun
biofilmdénitrifiantnourrideméthanoletformédebactériespréalablement
présentesdanslemilieu.L’espoirétaitquelebioréacteurpourraitgarderla
concentrationdunitratedissousàdesvaleursplussécuritaires,danslebutd’éviter
dessolutionsplusdrastiques,commeleremplacementd’eau.Lesystèmede
dénitrificaitonn’apuatteindrecesattentes,etdoncdeseffortsd’optimisationdela
performancedubioréacteurontétéentreprises.(Labbé,2007)
Ilaétédémontréqu’ilétaitpossibledegénérerunbiofilmcapablederéduire
uneconcentrationde50mgd’azoteparlitreàprèsde1mgd’azoteparlitre.Le
biofilmétaitformédebactériesprésentesdansl’eaumarinedumésocosmedu
FleuveSt-LaurentduBiodômeetétaitcultivéenmodecontinudansunréacteuràlit

11
fluidisé,maisàuneéchellepilote,beaucoupmoinsvolumineusequedans
l’aquariumoriginal.Ilaétéconcluquelebioréacteurdénitrifiantpourraitêtre
efficace,maisqueplusieursfacteurs(épaisseurdubiofilm,dégradationdesupports,
zonesmortes)quiétaientfacilesàcontrôlerenlaboratoiredevenaientdesobstacles
importantàl’applicationduprocédéàgrandeéchelle.(Labelleetal.,2005)
Figure2.3:BioréacteurdubiodômedeMontréal Bioréacteurdedésoxygénation(A)etdedénitrification(B)pourlemésocosmeduFleuveSt-LaurentauBiodômedeMontréal.Installéen1998.(Labbé,2007)
Parlasuite,lacompositionmicrobiologiquedubiofilmdubioréacteuraété
investiguée.Delabiomasseissuedubiofilmaétécultivéesurtroistypesd’agar
(TSA,R2AetWinogradsky).TroisisolatsontétéobtenusetleurADNribosomal16S
analysés,cequiapermisdedéterminerqu’ilsétaientdesα-protéobactériesdes
genresHyphomicrobium,Paracoccus,etd’unnouveaugenrebactérien.Lasouchedu

12
genreHyphomicrobiumaéténomméeNL23etaétécaractériséecommeundes
acteursprincipauxdel’activitédénitrifiante(Labbéetal.,2003).L’espècedu
nouveaugenreasubséquemmentétécaractériséecommenouvelleespèce
dénomméeNitratireductoraquibiodomus(Labbéetal.,2004).Lesbactériesnon-
cultivablesdubiofilmontétéétudiéesàl’aided’unegénothèqued’ADNribosomal
16Stotaldubiofilm.Ilaétédéterminéque70%desclonesgénérésavaientla
séquenced’uneγ-protéobactériedugenreMethylophaga.Lesautresprofils
indiquaientlaprésenced’unevariétédeséquencesreliéesàdesespèces
bactériennesdénitrifiantes.DesétudessubséquentesparFISH(«Fluorescentin
situhybridization»)ontconfirmél’abondancedominante(jusqu’à79%)desouches
appartenantaugenreMethylophagaetl’importancedugenreHyphomicrobium(7-
8%).Ilaaussiétédéterminéquelebiofilmincluaitenviron15espèces
bactériennes.(Labbéetal.,2003,Labbéetal.,2007)
Ilétaitsurprenantàl’époquequedesespècesdugenreMethylophaga
puissentcomposeruneaussigrandeproportiond’unbiofilmenconditions
anaérobies;legenreétaitjusque-làcaractérisépardesespècesstrictementaérobes.
Àl’aidedeméthanolmarquépardesisotopesdecarbone(14C)etd’analysedes
séquences16Sdanslesbactériesl’ayantassimilé,ilaétédémontréqu’uneespèce
dugenreMethylophagaétaithautementimpliquéedansleprocessusde
dénitrification.Unefoisisolée,cettesoucheaéténomméeJAM1;celle-ciaété
désignéecommeespècedistincteaprèsdesétudesdecaractérisationapprofondies
(voirsection2.6).LesgènesnarG1etnarG2ontaussiétécorrélésavecl’activitéde
réductiondenitratedelasoucheJAM1.Lesrésultatsd’analysesbioinformatiques
ontsuggéréuneoriginedetransferthorizontalpourlegènenarG1.(Auclairetal.,
2010)

13
2.6. MethylophaganitratireduscenticrescensLorsdesexpériencesexplorantladiversitémicrobienned’unbiofilm
provenantdel’aquariumdumésocosmeduFleuveSt-LaurentauBiodômede
montréal,ilfutdéterminéqu’unedessouchesbactériennesreprésentaitenviron
79%detoutelabiomasseprésente.L’identitédecettesouchen’étaitpasconnueà
l’époque,maisilavaitétédéterminéqu’elleétaitétroitementapparentéeaux
espècesdugenreMethylophaga.(Labbéetal.,2007)Suivantdesrecherchesplus
poussées,cettesouche,initialementnomméeJAM1,futdésignéecommenouvelle
espèceMethylophaganitratireduscenticrescens.(C.Villeneuveetal.,2013)
LegenreMethylophagaestunedivisiondesγ-protéobactériescaractérisée
pardesespècesbactériennesdeformebacille,typiquementlégèrementhalophiles.
Lesespècesmembresdugenresontaussidécritescommeconsommatricesde
composésàuncarbone(saufleméthane)dontleméthanol,qu’ellesassimilentà
l’aidedelavoiemétaboliqueduribulosemonophosphate.(Janvieretal.,1995)
Typiquement,lesespècesdugenreMethylophagasontdesespèceshabitantdes
milieuxmarinsoùellespeuventconsommerleméthanolabondammentproduitpar
desmicroorganismes(phytoplancton)(Minceretal.,2016).Plusieursespècesdu
genresontaussireportéesd’êtrecapabledelaproductiond’osmoprotecteur,
notammentl’ectoïne.CesespècesincluentM.thalassica,M.alcalica,M.murataet
autres(Doroninaetal.,2005,Mustakhimovetal.,2012,Mustakhimovetal.,2009).
LasoucheJAM1esttoléranteaumilieuxsalésjusqu’à8%deNaCl,malgréque
sacroissanceoptimaleestobtenuedansdesmilieuxd’environ3%deNaCl.Elleest
capabledecroissanceentredestempératuresde15˚Cà37˚Cmaispréfèreune
températureautourde30˚C.UnpHentre6et11permetlacroissance,maislepH
optimumestde8.ElleestauxotrophepourlavitamineB12,etrequiertdonc
l’inclusiondecettemoléculedanslemilieupourlacroissance.Surgélosedumilieu
1403(milieuadaptéauxsouchesbactériennesmembresdugenreMethylophaga),
sescoloniessontrondesetontenviron2mmdediamètre.Lasoucheestleseul

14
membredugenreMethylophagaàêtreanaérobefacultative,c’est-à-dired’être
capabledecroissanceenutilisantlenitrate(Auclairetal.,2010,C.Villeneuveetal.,
2013)etl’oxydenitreux(Mauffrey,résultatsnon-publiés)commeaccepteurs
d’électron,maispréférantl’oxygènedanscerôles’ilestdisponible.Ellepeut
égalementréduirel’oxydenitriteenoxydenitreux(Mauffrey,résultatsnon-publiés)
maispaslenitrite(Auclairetal.,2010).
2.7. HyphomicrobiumnitrativoransOutrelasoucheJAM1,uneautrebactériecomposaitunpourcentage
considérable(7-8%)delabiomassetotaledubiofilm.(Labbéetal.,2007)Celle-ci
futdésignéeNL23etparlasuitefutelleaussidécritecommeunenouvelleespèce,
Hyphomicrobiumnitrativorans.(Martineauetal.,2013)
LegenreHyphomicrobiumfaitpartiedesα-protéobactéries.Laparticularité
desespècesmembresestleurproductiond’hypheslorsdelareproduction.Les
bactériessontaussidécritescommebacillesauxboutspointus.Aussi,étantdonné
deleurfaibletoléranceauxenvironnementssalés,ellessontrarementretrouvées
dansdesmilieuxmarins.Plusieursmembresdugenreontétéimpliquésdansla
dénitrification,dontH.zavarziniietH.denitrificans,entreautres(Fesefeldtetal.,
1998,Martineauetal.,2015).Onretrouvecourammentcesbactériesdansles
bioprocédésdetraitementd’eauxusées.(Gliescheetal.,2005)
LasoucheNL23netolèrepasunmilieusalin,cequiestsurprenanten
considérantlecontextedesadécouverteetdesonactivité;lacroissancemaximale
estobtenueenconditionsde0-0,5%deNaCl.Presqueaucunecroissancen’est
possibleenmilieuxavecplusde1%deNaCl.LasoucheNL23peutcroîtreàdes
températuresentre15˚Cet35˚C,maispréfèreunetempératurede30˚à35˚C.LepH
optimalestentre7,5et8,5,maisunpHentre7et9,5estsuffisantpoursupporter
unecertainecroissance.CommelasoucheJAM1,lasoucheNL23estanaérobe
facultative,préférantlesenvironnementsoxygénésmaisétantcapabledecroissance

15
efficaceenutilisantlenitratecommeaccepteurd’électronspourgénérerdel’azote
gazeux.Surgélose,lescoloniesforméessontdepetitdiamètre(~1mm)etblanches.
LasoucheNL23peuttirersoncarboneduméthanol,delaméthylamineoudu
formate.(Martineauetal.,2013)
Figure3.1:LessouchesNL23etJAM1
LesespècesbactériennesHyphomicrobiumnitrativorans(soucheNL23,haut)etMethylophaganitratireduscenticrescens(soucheJAM1,bas)enmicroscopie.
2.8. Co-cultures,partagederessourcesÉtantdonnédelacomplexitédesconditionsdansunmilieunaturel,ilest
probablequelesprocessusdedénitrificationn’ysontnormalementpasperformés
pardesespècesuniquesmaisbienpardescommunautésvariéesimpliquantdes
partagescompliquésderessources.Unerelationsynergiqueentrebactériesestune

16
coopérationquiestbénéfiquepourtouteslesbactériesimpliquées.Ilexistedéjàdes
casdetellesinteractionsenco-culturesdénitrifiantesdocumentéesdansla
littératurescientifique.Parexemple,ilaétédémontréquel’espècebactérienne
Ralstoniapickettiauneactivitédénitrifiantede40à100foissupérieurelorsqu’elle
estcultivéeenprésencedel’espèceStreptomycesgriseus.L’aideétaitfourniepar
cettedernièresousformedeprotéasessecrétéesquiaugmentaientlaconcentration
d’histidinedanslemilieu.Cettehistidineaensuiteétéenmesured’augmenterle
tauxdedénitrificationdeR.pickettipourdesraisonsinconnues.(Takakietal.,2008)
Unautreexempledesynergieestlaco-culturedel’espèceAlcaligenesfaecalis
aveclasouchePseudomonasG9.Lapremièreestunebactériedénitrifiantequiest
incapablederéduirelenitrateennitrite,maisquiperformelesautresétapesdela
dénitrification.Àl’inverse,lasoucheG9exprimesanitrateréductase
constitutivement.Celle-ciperformeaussilesautresétapesdeladénitrificationmais
souffred’uneaccumulationtoxiquedenitriteenculturepure.Deplus,ellenepeut
pascroîtrelorsqu’elleestinoculéeenconditionsanaérobies.Enconditions
dénitrifiantes(anaérobies)avecseullenitratecommeaccepteurd’électronfourni,
chaquebactérieestincapablededémontrerunecroissanceenculturepure.Ila
pourtantétédémontréqu’uneco-culturedesdeuxbactériesaétécapablede
croissanceetd’activitédénitrifianteefficace.(VandePas-Schoonenetal.,2005)

17
2.9. HypothèseetobjectifsCertainesdescaractéristiquesuniquesàchaquesouche(NL23etJAM1)
laissentimaginerqu’ilpuisseexisteruncertainéchangederessources.Lasouche
JAM1nepeutpasréduirelenitrite,tandisquecettecapacitéestdisponibleàla
soucheNL23.Demême,lasoucheNL23nepeutexisterenprésencedehautes
concentrationsdesel;sepourrait-ilquelasoucheJAM1puissepartagerune
certaineosmoprotectiondelaquelleelledispose?Leportraitfinalseraitun
mutualismequiamélioreraitsubséquemmentl’efficacitédeladénitrification.
Donc:
Hypothèse:ilexisteunecoopérationentrelesNL23etJAM1enco-culturesous
formedepartagederessources(nitrate,nitrite,vitamineB12)oud’autresfacteurs
(ex:osmoprotection/ectoïne).
Objectifs:
• l’optimisationdesconditionsdelaco-culturedessouchesNL23etJAM1danslebutd’enfaciliterl’étude;
• lacaractérisationdesinteractionsentrelesdeuxsouches.

18
3. Matérieletméthodes
3.1. SouchesbactériennesLessouchesbactériennesdeceprojetontétéisoléesàpartird’unbiofilm
dontl’origineestlebioréacteurdénitrifiantdumésocosmeduFleuveSt-Laurentdu
BiodômedeMontréal.Celles-cisont:Methylophaganitratireducenticrescens
(soucheJAM1),Hyphomicrobiumnitrativorans(soucheNL23)(Fig.3.1).Plusieurs
échantillonsdechaqueespèceontétéconservéesà-80˚Cdansleursmilieuxde
culturerespectifscomplémentésde25%deglycérol.
3.1.1. Mutants
Enplusdesdeuxsouchesbactériennesci-dessus,troismutantsdetype
«knockout»delasoucheJAM1ontétéutilisés.Ceux-ciexhibenttousdes
déficiencesencequiconcerneleursenzymesdelanitrateréductase:JAM1ΔnarG1
estunmutantsanscopiedugènenarG1;JAM1ΔnarG2estunautresanscopiedu
gènenarG2;etJAM1ΔnarG1narG2estundouble-mutantnepossédantaucunecopie
desgènesnarG1etnarG2.Cessouchesontétécrééesparlaméthodedeladouble
recombinaisonparFlorianMauffrey,étudiantdedoctoratdanslelaboratoireduPr.
Villemur(Mauffreyetal.,2015).

19
3.2. Culturedesbactéries
3.2.1. MilieuoptimaldelasoucheJAM1LemilieuoptimalpourlacroissancedelasoucheJAM1telquedéterminé
danslestestsd’optimisationpréalables(Villeneuveetal.,2013)estlemilieuATCC
1403pourespècesdugenreMethylophaga.Unlitredumilieuliquidecontient:
NaCl----------------------------------------------24g
MgCl2·6H2O--------------------------------------3g
MgSO4·7H2O------------------------------------2g
KCl------------------------------------------------0,5g
CaCl2·2H2O--------------------------------------1g
Bis-Tris------------------------------------------0,5g
SolutionT-------------------------------------20ml
SolutiondeWolf-----------------------------10ml
Eaudistillée---------------------------------970ml
LasolutionTestunesolutionminéraledontlacompositionestlasuivante(pour1L
desolution):
KH2PO4--------------------------------------------7g
NH4Cl------------------------------------------100g
Bis-Tris----------------------------------------100g
(NH4)5Fe(C6H4O7)2------------------------------3g
Eaudistillée-------------------------------1000ml

20
LasolutiondeWolfestuneautresolutionminéraleenutilisationcourante.Unlitre
delasolutioncontient:
N(CH2CO2H)3-----------------------------------1,5g
MgSO4·7H2O------------------------------------3g
MnSO4·H2O------------------------------------0,5g
NaCl------------------------------------------------1g
FeSO4·7H2O-----------------------------------0,1g
CoCl2·6H2O------------------------------------0,1g
CaCl2---------------------------------------------0,1g
ZnSO4·7H2O-----------------------------------0,1g
CuSO4·5H2O---------------------------------0,01g
AlK(SO4)2·12H2O---------------------------0,01g
H3BO3------------------------------------------0,01g
Na2MoO4·2H2O-----------------------------0,01g
Eaudistillée-------------------------------1000ml
Suiteàl’ajoutdesminérauxlemilieuaétéajustéàunpHde7,3etautoclavéà121˚C
pendantaumoins15minutes.Avantutilisation,etseulementlorsquelasolutionest
àtempératuredelapièce,l’ajoutdevitamineB12etdeméthanolaétéeffectué.Pour
100mldemilieu,0,3mldeméthanolstériliséparfiltrationet0,1mldevitamineB12
(0,1mg/ml)(stérilisédelamêmefaçon)ontétéajoutés.

21
3.2.2. MilieuoptimaldelasoucheNL23
PourlasoucheNL23,lemilieudecroissanceoptimalestuneversion
modifiéedumilieuATCC337a,dontl’efficacitéaététestéelorsd’études
précédentes(Martineauetal.,2013).Danschaquelitredumilieuonretrouve:
KH2PO4------------------------------------------1,3g
Na2HPO4--------------------------------------1,13g
(NH4)2SO4---------------------------------------0,5g
MgSO4SO4·7H2O------------------------------0,2g
Solutiond’élémentstraces------------------5ml
Eaudistillée-------------------------------1000ml
Lasolutiond’élémentstracesestspécifiquementadaptéeaumilieu337a;unlitrede
lasolutionestcomposéde:
CaCl2·2H2O--------------------------------0,618g
FeSO4·7H2O-----------------------------------0,4g
Na2MoO4·H2O---------------------------------0,2g
MnSO4·4H2O------------------------------0,176g
Eaudistillé---------------------------------1000ml
Lasolutiond’élémentstracespouvaitcontenirunecertainequantitédeprécipité
suivantuncycleàl’autoclave,maiscecin’apasproduitd’effetsnotablessurla
croissancedesbactéries.Suiteàlacombinaisondesingrédients,lasolutionaété
ajustéeàunpHentre7,5et8etautoclavéeà121˚Cpendantaumoins15minutes.
DelavitamineB12(0,1mlà0,1mg/ml)etduméthanol(0,3ml)ontétéajoutés
avantl’utilisation.

22
3.2.3. Milieudecroissanceadaptéàlaco-culture
Avantledébutduprojet,ilétaitdéjàconnuquelessouchesNL23etJAM1,
ainsiqu’uneco-culturedesdeux,pouvaientsimplementcroitredansuneversion
diluée(15g/Laulieude30g/L)dumilieumarinartificiel«InstantOcean»
retrouvéauBiodôme.Lacompositionexacted’unlitredumilieu«InstantOcean»
pourco-cultureest:
Mélange«InstantOcean»------------------15g
NaNO3---------------------------------------1,214g
KH2PO4(ensolution32,67g/L)----------2,5ml
Solutiond’élémentstraces------------------1ml
Eaudistillée-------------------------------1000ml
Lemélanged’eausaléepouraquarium«InstantOcean»estacheté
commercialementetsacompositionexacteestdoncinconnue(«InstantOcean»est
lenomdufournisseur).
Lasolutiond’élémentstracesestspécifiqueaumilieu«InstantOcean»;unlitrede
lasolutionestcomposéde:
FeSO4·7H2O-----------------------------------0,9g
CuSO4·5H2O-------------------------------0,035g
MnSO4·H2O--------------------------------0,202g
Eaudistillée-------------------------------1000ml
Unefoisquelemilieuestcomplet,lepHaétéajustéentre7,8et8etautoclavéà
121˚Cpendantaumoins15minutes.Avantl’utilisation,0,1mldevitamineB12(0,1
mg/ml)et0,3mldeméthanolontétéajoutés.Laquantitédenitrateajoutéeau
milieuvarieselonlesbesoinsdel’expérienceetlesconnaissancesacquiseslorsdela
recherche.Cesdifférencessontindiquéesoùelless’appliquent.

23
3.2.4. Milieuxsousformedegéloses
Chaquevariétédemilieudécriteci-hautaétéprincipalementutiliséesous
formeliquidepoursoutenirdespopulationsdebactériesensuspensionoupour
permettrelaformationdebiofilmsursupportsadaptés.Cependant,cesmilieuxont
aussiétéutilessousformedegélosesenboîtesdePetri.Pourproduiredesgéloses,
lasolutionminérale(enconcentrationdouble)etlasolutiond’agar(3%)ontété
autoclavéesséparément,puiscombinéesavantd’êtrecouléesdanslesboîtesde
Petri.LavitamineB12etleméthanolontétéajoutésaprèsl’autoclavage.
3.3. Préparationdescultures
PuisquelessouchesJAM1etNL23ontdestempsdecroissancedifférents,la
culturedesbactériesadûsuivreunrégimerigoureux.Uneréserveàlongtermede
chaquesouchebactérienneaétéconservéeà-80˚C.Àpartirdecelle-ci,descolonies
distinctesontétéproduitessurgéloses;lacroissancesedéroulaitsurunepériode
d’environ10joursà30˚C,puiscesculturesétaientconservéesà4˚Cpourune
périoded’utilisationd’environunmois.Àpartirdecoloniesdistinctes,
préférablement,despetitsvolumes(environ15ml)decultureliquideontété
préparés,cequinécessitede2à3joursd’incubation.Cesculturesontàleurtour
étéconservéesà4˚Cpourunepérioded’utilisationde1-2semaines.Également,ces
culturesontétéutiliséespourcréerlesculturesenplusgrosvolumes(variables
selonlesbesoinsdel’expérience)quiservirontd’inoculumpourlesexpériences
elles-mêmes.Parexemple,ajouter0,6mldecultureliquideà10mldemilieu
optimalfraisapermislacroissancejusqu’àdensitémaximaledelasoucheJAM1en
24hetdelasoucheNL23en40h.Danslescasoùl’expérienceenvisagéeimplique
uneco-culture,lestempsd’ensemencementontétéplanifiéspourqueladensité
maximalesoitatteinteenmêmetempspourlesdeuxsouchesaudébutdeladite
expérience.

24
3.3.1. Ensemencement
Àl’ensemencement,ladensitéoptiquedesculturesbactériennesservant
d’inoculumaétémesuréeàunelongueurd’ondede600nm(DO600).Decette
façonlaconcentrationdelabiomasseapuêtreajustéeàunevaleurquiétait
toujourslamêmepourchaqueexpérience;danscecasleDO600viséétaitde0,1
danslacultureexpérimentale.Donc,danslecasd’uneco-culture,chaquesouche
individuelledevaitcontribuerunDO600de0,1danslaco-cultureexpérimentaleau
tempsinitial.Ceciapermisunecertaineconstancedansleprocessusexpérimental
quiaévitéquelesconcentrationsinitialesdebactériesaffectentlerésultatd’une
expérience.Deplus,danslescasoùunchangementdemilieuaeulieuaumoment
del’ensemencement(parexemple,uneco-cultureoùchaquebactériepassed’un
milieuoptimalàunmilieuaccommodantlesdeuxsouches),lesculturesde
l’inoculumontdûd’abordêtrecentrifugéespourendébarrasserlemilieuinitial
(8000rpm,10minutes,25˚C,deuxfois).
3.3.2. Conditionsdeculture
Lesbactériesontétécultivéesenconditionsaérobiesouanaérobies.Les
culturesaérobiesontétéfaitesenflaconsErlenmeyerouenplaquesmulti-puits,
selonlebesoin,etontrequisdel’agitationconstante(entre60et100rpm)pour
faciliterlacirculationd’oxygènedanslemilieu.Lesculturesanaérobiesontété
effectuéesdansdesfiolesdeborosilicatedanslesquellesl’oxygèneaétéévacuéà
l’aided’azotegazeux(Fig3.2).Danscesfioles,l’échantillonnageaétérendupossible
grâceàunbouchonenbutyledanslesquelspeuventêtreinsérésdesaiguilles.Dans
touslescas,lesculturesontétéincubéesàunetempératureconstantede30˚C.

25
Figure3.2:Exempledecultureenanaérobiose
Unefioleferméehermétiquementpourculturedebactériesenconditionsanaérobies.Uneculturecommecelle-cicontienttypiquement30mldemilieuliquideetestincubéeà30˚Cavecseringuedanslebouchonenbutyle.
3.4. Récoltededonnées
Pourprendreunéchantillon,lacultureaétéagitéepourassurerune
distributionégaledescellulesetdoncdelaconcentrationdansleliquide.En
anaérobie,uneseringueaétéinséréedanslebouchondebutylescellantlafiole,et
aumoins1mlaétéextrait.UnefoisquelamesuredeDO600aétépriseavecle
spectrophotomètre,l’échantillonaétéconservéà-20˚Cdanslebutd’effectuer
d’autrestests(principalementdelachromatographieionique).

26
3.4.1. Spectrophotométrie
Pourrécolterdesdonnéesutilessurcescultures,dessuivisdecroissances
ontétéeffectuéesavecplusieurscombinaisonsdeconditions.Enconditions
aérobies,lesuiviaétéeffectuéuniquementparmesuresdeDO600àl’aided’un
spectrophotomètre,ouparmesuresdeturbiditéàl’aided’unappareilautomatisé
d’analysedecroissance(«Bioscreen»).L’avantageprincipald’unappareil
automatiséaétélapossibilitédeprendredesmesuressimultanémentpourchaque
échantillondefaçonrapide,cequiapermisderécolterdesdonnéesàdesintervalles
detempsplusrapprochées.Enconditionsanaérobies,lesuiviaégalementété
effectuéparmesuresdeDO600(manuellesseulement).
3.4.2. Chromatographieionique
Lachromatographieioniqueaétéutiliséepourquantifierlaconsommation
denitrateetdenitritedanslemilieuparlesdeuxsouchesbactériennes.Les
échantillonsontétépréalablementcentrifugés(13000rpm,2min.)dansdes
microtubesdetypeEppendorfpourenretirerlesbactéries.Unefoisquelescellules
seretrouvaientaufonddutube,aumoins0,5mldusurnageantdechaque
échantillonaétéprélevépourêtreanalyséparchromatographieionique.
L’appareilutilisépourlaquantificationdunitrateetdunitriteétaitle850
ProfessionalICdeMetrohm,munid’unecolonneMetroseppAsupp5dedimensions
250X4mm.Lacolonneaétémaintenueà35˚Cdurantl’opération.Lesmesuresde
nitrateetdenitriteontétéeffectuéesparUV.L’éluantutiliséétaitcomposéde3.2
mmol/LdeNa2CO3etde1mmol/LdeNaHCO3.Ledébitétaitde0,7ml/min.
3.4.3. Extractiond’ADN
Labiomassed’intérêt(ensuspensionliquide)ad’abordétécentrifugéeafin
d’enretirerlemilieudeculture.Lescellulesrésultantesontensuiteétédispersées
dansunesolutiontamponTEN(10mMTris-Cl;1mMEDTA;1MNaCl)dansun
microtubedetypeEppendorfde1,5mLcontenant0,2mgdebillesdeverresde0,1

27
mm.Cemélangeasubséquemmentétéplacédansunhomogénéisateurpour
effectuerunbroyage:rapidementagiterlescellulesensuspensionpourqu’elles
éclatentcontrelesbillesdeverre.L’appareilutiliséétaitleFastPrep-24deMPBio.
L’ADNaétéextraitàl’aided’unmélangedephénol,dechloroformeetd’alcool
isoamylique(25:24:1).L’ADNapuensuiteêtreprécipitédansunemixture(9:1)
d’éthanolfroid(90%)etd’acétatedesodium.Letoutaétécentrifugé,etleprécipité
aétédissousdansl’eau.L’ADNaétéquantifiéparspectrophotométrie,cequia
égalementdonnéuneindicationdelapuretédelasolutionobtenue.Le
spectrophotomètreutilisélorsdecesétudesétaitleNanodrop2000deThermo
Scientific.
3.4.4. qPCR
Danslecadredecesétudes,l’applicationprincipalementintéressantedela
techniqueestlaquantificationdecopiesdegènesparticuliersprésentsdansuneco-
culture.Puisquelessouchesbactériennesétudiéesontchacuneleurpropreversion
uniquedelanitrateréductasedontlaséquencegénétiqueestconnue,ilaété
possibled’utiliserleqPCRpourmesurerlaquantitérelativedesbactériesdansle
mélange.Lesnitrateréductasesenquestionsontlanitrateréductasepériplasmique
delasoucheNL23(gènenapA)etunedesdeuxnitrateréductasesmembranairesde
lasoucheJAM1(gènenarG1).Donc,parexemple,siuneanalyseqPCRindiqueque
legènenarG1représente90%descopiesdegènesdenitrateréductase(narG1+
napA)dansl’échantillon,celaindiquequelasoucheJAM1comptaitneuffoisplusde
cellulesquelasoucheNL23danslaco-culture.
LatechniquedeqPCRutiliséelorsdecesétudesétaitleTaqMan.Danscette
variationduqPCR,deuxcourtesséquencesdenucléotidesserventd’amorces,tandis
qu’uneautrecourteséquencesertdesonde.Cettedernièreestconçuepour
s’apparieràunendroitentrelesdeuxamorces.Àlasondesontfixésun
fluorochromeetun«quencher».Lafluorescenceémiseparlefluorochromeest
dissimuléeparle«quencher»lorsquelesdeuxsontàproximité.Lapolymérase
Taqsemetalorsàajouterdesnucléotidesauxamorces,etquandellearriveàla

28
sonde,celle-ciestenlevéeestdégradée,libérantlefluorochromequiémetalorsune
fluorescencequantifiable.LeTaqManaétéutilisédansceprojetàcausedesoncoût
abordablecomparéauSYBRgreen,leprotocolequiétaitutiliséprécédemmentdans
lelaboratoire.Ilaaussiéténotéquedesproblèmesdedimèresd’amorcessont
aussiplusfréquentsavecleSYBRgreen.D’ailleurs,ilaétédéterminéqueleTaqMan
génèredesrésultatscomparablesauSYBRgreen(Blais,2014).
L’instrumentutilisépoureffectuerlesqPCRétaitleRotor-Gene6000de
CorbettLifeSciences(Qiagen).Chaquemicrotube(tubesspécialisés0,1mlavec
couverclesdeQiagen)contenait25µld’unmélangepréparéeenlaboratoire(voir
tableau3.1).L’ADNajoutéaumélangeaétéobtenugrâceàlaméthoded’extraction
décritedanslasection3.4.3.LeprogrammePCRétait:
i) 5minutesà95˚C
ii) 40cyclesdesétapessuivantes:
(1) 15secondesà95˚C
(2) 20secondesà55˚C
(3) 30secondesà65˚C
Tableau3.1:compositiondumélangedanschaqueréactiondeqPCR
Composante Volumeparréaction
H2O 14,88µl
Tampon10Xtaq«hotstart» 2,5µl
BSA(20µg/µl) 0,5µl
dNTP(10mM) 0,5µl
Amorce(f)(10mM) 0,5µl
Amorce(r)(10mM) 0,5µl
SondeTaqMan(5mM) 0,5µl
Taqpolymérase«hotstart»(5U/µl) 0,125µl
ADNgénomique(dilué1/5) 5µl
29
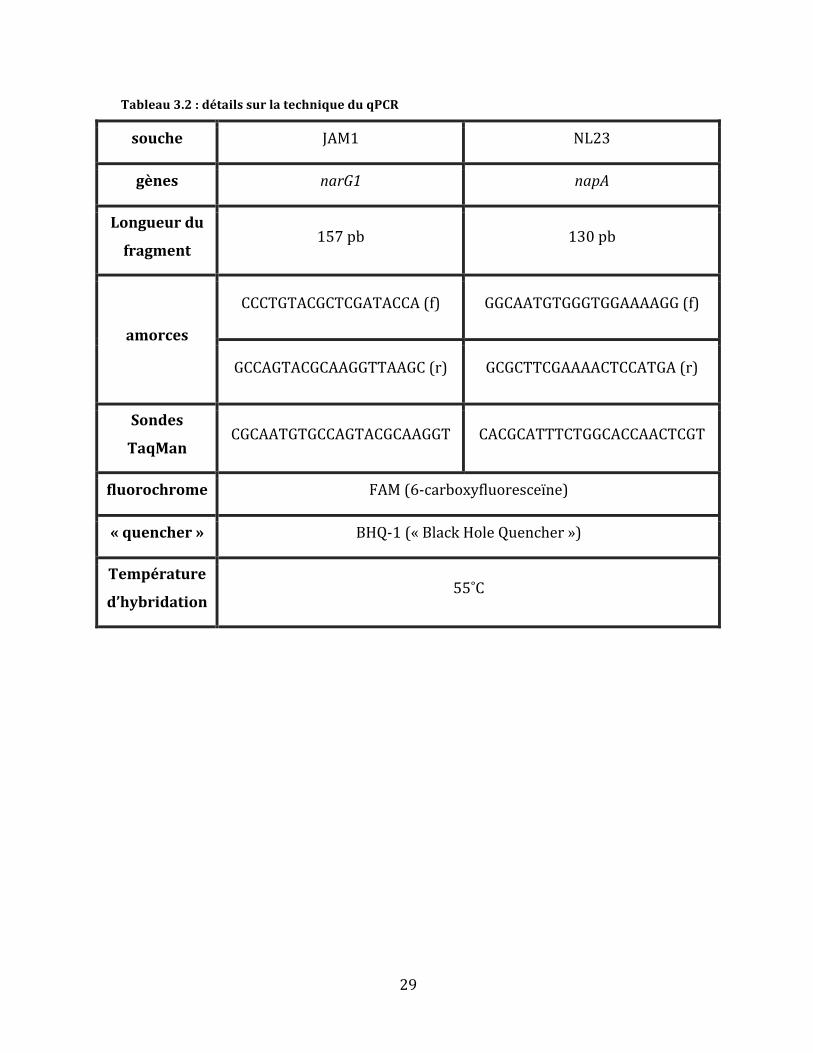
Tableau3.2:détailssurlatechniqueduqPCR
souche JAM1 NL23
gènes narG1 napA
Longueurdu
fragment157pb 130pb
amorces
CCCTGTACGCTCGATACCA(f) GGCAATGTGGGTGGAAAAGG(f)
GCCAGTACGCAAGGTTAAGC(r) GCGCTTCGAAAACTCCATGA(r)
Sondes
TaqManCGCAATGTGCCAGTACGCAAGGT CACGCATTTCTGGCACCAACTCGT
fluorochrome FAM(6-carboxyfluoresceïne)
«quencher» BHQ-1(«BlackHoleQuencher»)
Température
d’hybridation55˚C

30
4. Résultats4.1. Déterminationdesconditionsoptimalesdeculture
Lepremiervoletduprojetétaitconstituéd’expériencesvisantàdéterminer
quellesconditionsdeculturepermettraientlameilleurecroissancedessouches
JAM1etNL23.Unegrandepartiedutravailàcetégardétaitdéjàcomplétépardes
étudespréliminaires.Eneffet,laplupartdesparamètresdebase,incluantla
température,lepH,lasalinité,etlemilieudecroissanceavaientétéétablisavantle
débutdestravauxprésentésdanscemémoire.Lesconditionsoptimalesqu’ilrestait
àdéterminerimpliquaientprincipalementlesconcentrationsdenitrateetles
proportionsdesbactériesenco-cultures.
4.1.1. Suividecroissancepréliminaireenconditionsaérobies
Cespremierssuivisdecroissanceavaientpourbutdevaliderlechoixde
milieudecroissancepourlesco-culturesdessouchesJAM1etNL23ainsiquede
donnerunaperçudescourbesdecroissancetypiquesdecelles-ci.LessouchesJAM1
etNL23ontétécultivéesenconditionsaérobiesouanaérobies,soitsousformede
culturespuresouenco-cultures.Lemilieucommerciald’eaudemer«Instant
Ocean»aétéutilisépourcespremièresexpériencesenprésencedeprèsde40mM
d’azotesousformedenitrate.Ilaétédéterminéauparavantquelemilieu«Instant
Ocean»coupédemoitié(15g/L,aulieude30g/L)permettaitlacroissanceàlafois
dessouchesJAM1etNL23,lasoucheNL23étanttrèssensibleàlaconcentrationen
sel.
31
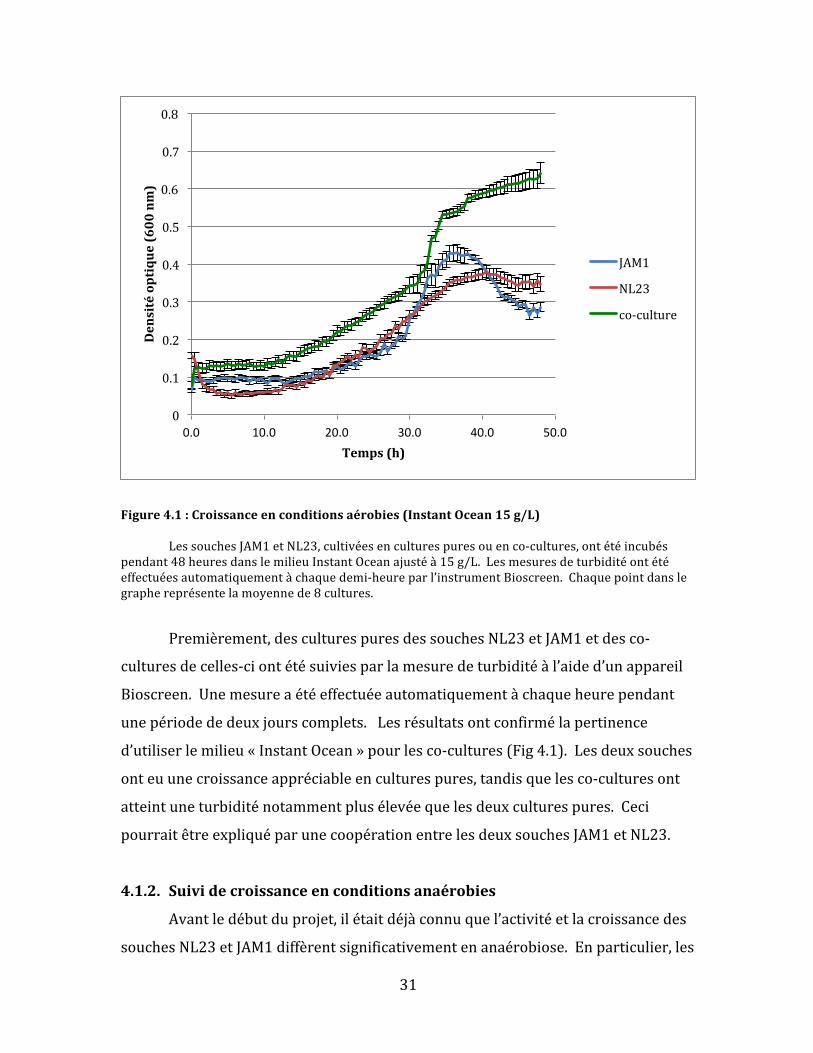
Figure4.1:Croissanceenconditionsaérobies(InstantOcean15g/L) LessouchesJAM1etNL23,cultivéesenculturespuresouenco-cultures,ontétéincubéspendant48heuresdanslemilieuInstantOceanajustéà15g/L.Lesmesuresdeturbiditéontétéeffectuéesautomatiquementàchaquedemi-heureparl’instrumentBioscreen.Chaquepointdanslegraphereprésentelamoyennede8cultures.
Premièrement,desculturespuresdessouchesNL23etJAM1etdesco-
culturesdecelles-ciontétésuiviesparlamesuredeturbiditéàl’aided’unappareil
Bioscreen.Unemesureaétéeffectuéeautomatiquementàchaqueheurependant
unepériodededeuxjourscomplets.Lesrésultatsontconfirmélapertinence
d’utiliserlemilieu«InstantOcean»pourlesco-cultures(Fig4.1).Lesdeuxsouches
onteuunecroissanceappréciableenculturespures,tandisquelesco-culturesont
atteintuneturbiditénotammentplusélevéequelesdeuxculturespures.Ceci
pourraitêtreexpliquéparunecoopérationentrelesdeuxsouchesJAM1etNL23.
4.1.2. Suividecroissanceenconditionsanaérobies
Avantledébutduprojet,ilétaitdéjàconnuquel’activitéetlacroissancedes
souchesNL23etJAM1diffèrentsignificativementenanaérobiose.Enparticulier,les
0
0.1
0.2
0.3
0.4
0.5
0.6
0.7
0.8
0.0 10.0 20.0 30.0 40.0 50.0
Densitéoptique(600nm)
Temps(h)
JAM1
NL23
co-culture

32
nitrateréductasesdelasoucheJAM1sontbeaucoupplusefficaces,etcelle-ci
démontreunecroissanceinitialeplusélevée(Martineauetal.,2015,Mauffreyetal.,
2015).Lespremierssuivisdecroissanceenconditionsanaérobiesontdoncétéfaits
avecdifférentsratiosd’inoculumpourdéterminerlesconditionsd’ensemencement
lesplusfavorables.LessouchesNL23etJAM1requièrentlaprésencedenitrateet
sesdérivéescommesourced’accepteursd’électronspourcroîtreenanaérobiose.
Dansleursmilieuxoptimauxrespectifs,unequantitéinitialedenitrateoptimaleest
d’environ40mM(CélineVilleneuve,2012)(donnéesnon-publiéespourlasouche
NL23),donccetteconcentrationinitialeaétéchoisie.
Lessuivisdecroissanceontétéeffectuéssurunesériedeco-culturesavec
desvariationsdansleratioentrelesdeuxsouchesbactériennes.L’expérience,qui
comptaitégalementcommetémoinslessouchesNL23etJAM1enculturespures,a
étéeffectuéeavecdesco-culturesderatios(JAM1:NL23)1:1,1:2,1:5,et1:10.
Cesratiosréfèrentuniquementauxquantitésrelativesdessouchesaumomentde
l’ensemencement,telsquemesurésparDO600.Lacroissanceétantbeaucoupplus
lenteencesconditions,etàcausedeconsidérationspratiques,desmesuresde
DO600ontétéprisesmanuellementunefoisparjourjusqu’àcomplétiondusuivi.
L’appareil«Bioscreen»nepermetpaslesculturesenconditionsanaérobies.Les
échantillonsainsirecueillisontégalementservipourlaquantificationdenitrateet
denitrite.Leséchantillonnagesjournaliersontétéeffectuéssurunepériodedeprès
de200heures,aprèsquoilesculturesontétéconservéespourdesétudesparqPCR.

33
Figure4.2:Lacroissancedesco-culturesdessouchesNL23etJAM1enconditionsanaérobies Desco-culturesdessouchesNL23etJAM1ontétécultivéesdanslemilieuInstantOcean15g/Ljusqu’àl’atteinteduplateaudecroissanceou200heures.Lesco-culturesavaientdesratiosd’inoculumsdifférents.ChaquepointdanslegraphereprésenteleDO600moyende3cultures,mesuréparspectrophotomètrie.
LesuividuDO600estsouventpeuinformatifdanslecadred’uneexpérience
enconditionsanaérobies.Ceciestdûaucaractéristiquesdelacroissancedela
soucheJAM1.CettedernièreexhibedesDO600trompeursàcausedelaformation
defloc,desaggrégationsdecellulesvisiblesàl’oeilnu,etaussiàcaused’un
“éclaircissement”dumilieuInstantOcean,quiestnormalementtrouble.
Effectivement,aprèsnormalisationavecleDO600dumilieunon-inoculé,ilest
communderecueillirdesDO600àvaleursnégatives,ainsiqueded’obtenirdes
valeurstrèsdifférentespourlemêmeéchantillon.Ilestdoncpeuprobablequ’une
mesureduDO600reflètecequisepasseréellementauseind’uneculturesimplede
lasoucheJAM1oud’uneco-culture.Pourcetteraison,lesuivideDO600sert
-0.15
-0.1
-0.05
0
0.05
0.1
0.15
0.2
0.25
0 50 100 150 200
Densité
op+
que
Temps(h)
NL23
co-culture1:1
co-culture1:2
co-culture1:5
co-culture1:10
JAM1

34
principalementdeméthodepermettantd’observerqualitativementetrapidementle
progrèsdelacroissancedesbactéries.
Danslecasdecetteexpérience,cependant,ilestintéressantdenoterqueles
co-culturesavecunratiopluségal(lesco-cultures1:1et1:2)ontsuiviunecourbe
decroissancequiressemblebeaucoupauxculturespuresdelasoucheJAM1,tandis
quelesautresco-culturesontsuiviunecourbedecroissancequiressembleàcelle
desculturespuresdelasoucheNL23(Fig4.2).Cecipourraitréfletersimplement
quelecomportementtypedelasoucheJAM1devientmoinsvisibledanslesdonnées
lorsquecettedernièreestmoinsabondantedanslesco-cultures.Toutefois,ilest
aussipossiblequ’unedensitéplusimportantedesdeuxsouchesontpuse
développerenco-cultureslorsquel’inoculumdedépartdesoucheJAM1étaitmoins
important.Cetteexplicationseraitcompatibleavecl’hypothèsedudépart.Pour
approfondirlacompréhensiondelasituation,ilafalluinvestiguerauniveaudela
consommationdunitrateetdunitrite(Fig4.3et4.4).
35
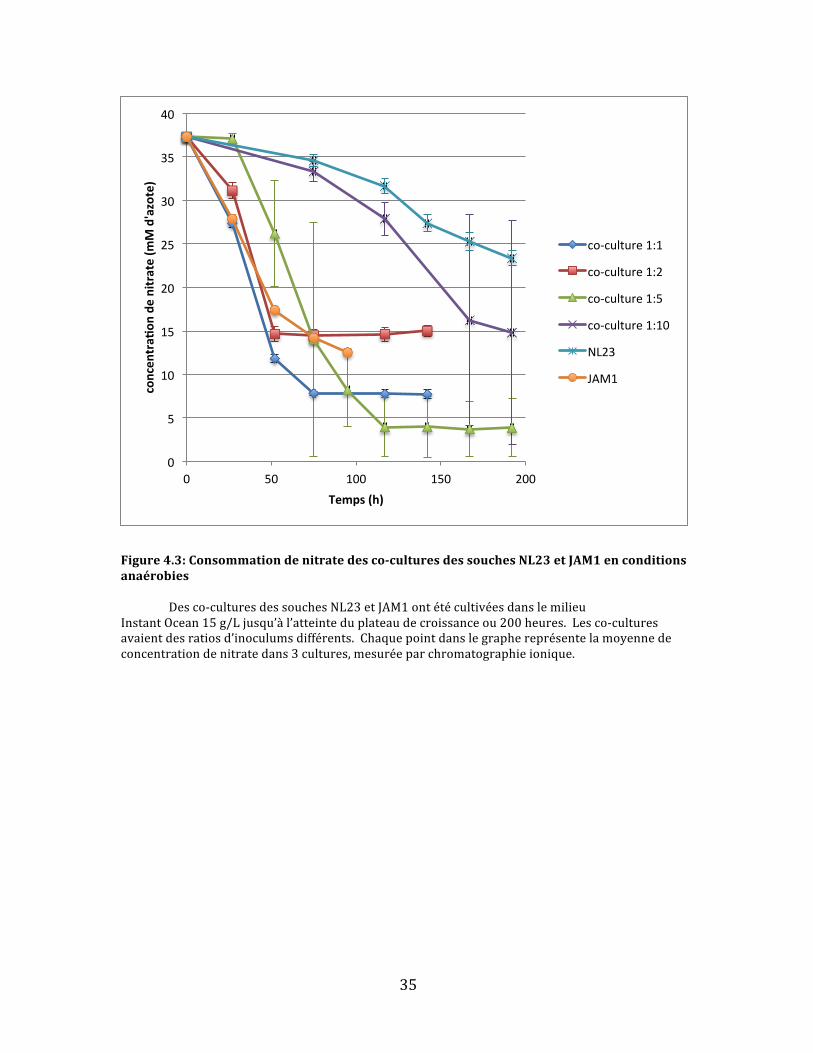
Figure4.3:Consommationdenitratedesco-culturesdessouchesNL23etJAM1enconditionsanaérobies Desco-culturesdessouchesNL23etJAM1ontétécultivéesdanslemilieuInstantOcean15g/Ljusqu’àl’atteinteduplateaudecroissanceou200heures.Lesco-culturesavaientdesratiosd’inoculumsdifférents.Chaquepointdanslegraphereprésentelamoyennedeconcentrationdenitratedans3cultures,mesuréeparchromatographieionique.
0
5
10
15
20
25
30
35
40
0 50 100 150 200
concen
tra+
ondenitrate(m
Md'azote)
Temps(h)
co-culture1:1
co-culture1:2
co-culture1:5
co-culture1:10
NL23
JAM1

36
Figure4.4:concentrationdenitritedanslesco-culturesdessoucheNL23etJAM1enconditionsanaérobies Desco-culturesdessouchesNL23etJAM1ontétéincubéesdanslemilieuInstantOcean15g/Ljusqu’àl’atteinteduplateaudecroissanceou200heures.Lesco-culturesavaientdesratiosd’inoculumsdifférents.Chaquepointdanslegraphereprésentelamoyennedeconcentrationdenitritedans3cultures,mesuréeparchromatographieionique.
Cequiestimmédiatementévidentdanslegraphederéductiondenitrate(fig.
4.3)estquepluslaconcentrationdelasoucheJAM1estélevéeinitialementdansles
co-cultures,plusletauxderéductiondunitrateestrapide.Ladeuxième
observationqu’onpuissefaireestqu’aucunedesco-culturesneparvientàréduire
complètementlenitrate.Danslecasdesco-culturesàconcentrationsélevéesen
soucheJAM1,laconsommationdunitrates’arrêtebrusquementaprèsenviron75
heuresavec5-15mMd’azoterésiduel.Quantauxco-culturesmoinsabondantesen
soucheJAM1,ilsemblequ’ellessoienttroplentespourréduireentièrementle
nitrateavant200heures,laduréedel’expérience.Iln’estpasclairdanslesdonnées
0
5
10
15
20
25
30
0 50 100 150 200
concen
tra+
ondenitrite
(mMd'azote)
Temps(h)
co-culture1:1
co-culture1:2
co-culture1:5
co-culture1:10
NL23
JAM1

37
recueilliessilaréductionauraitpucontinuerjusqu’àépuisementdesréservesde
nitrate.
Lesuividelaconsommationdunitritecorroborel’hypothèse:Lesco-
cultures1:1et1:2ontaccumulélenitriterapidementpendant50-100heures
avantdeplafonner(entre18et26mMd’azote)indéfiniment.Lesco-cultures1:5
ontatteintunniveaudenitritemaximum(environ17mMd’azote)àenviron120
heurespourredescendreverslafindusuivi.Finalement,lesco-cultures1:10n’ont
démontréqu’unepetiteaccumulationdenitritetarddansl’expérience(170heures).
Lestempsoùlespointsmaximauxd’accumulationdenitritesontatteintssemblent
correspondreàpeuprèsàceuxd’arrêtd’activitéderéductiondenitrate.
38
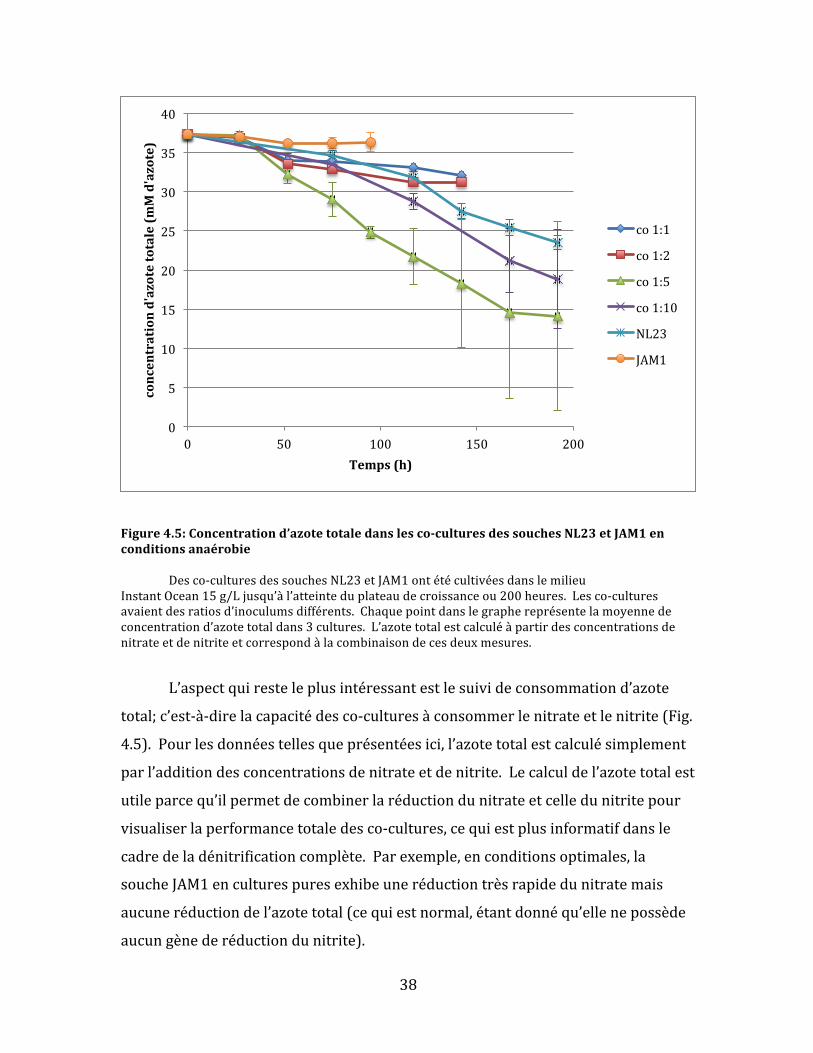
Figure4.5:Concentrationd’azotetotaledanslesco-culturesdessouchesNL23etJAM1enconditionsanaérobie Desco-culturesdessouchesNL23etJAM1ontétécultivéesdanslemilieuInstantOcean15g/Ljusqu’àl’atteinteduplateaudecroissanceou200heures.Lesco-culturesavaientdesratiosd’inoculumsdifférents.Chaquepointdanslegraphereprésentelamoyennedeconcentrationd’azotetotaldans3cultures.L’azotetotalestcalculéàpartirdesconcentrationsdenitrateetdenitriteetcorrespondàlacombinaisondecesdeuxmesures.
L’aspectquiresteleplusintéressantestlesuivideconsommationd’azote
total;c’est-à-direlacapacitédesco-culturesàconsommerlenitrateetlenitrite(Fig.
4.5).Pourlesdonnéestellesqueprésentéesici,l’azotetotalestcalculésimplement
parl’additiondesconcentrationsdenitrateetdenitrite.Lecalculdel’azotetotalest
utileparcequ’ilpermetdecombinerlaréductiondunitrateetcelledunitritepour
visualiserlaperformancetotaledesco-cultures,cequiestplusinformatifdansle
cadredeladénitrificationcomplète.Parexemple,enconditionsoptimales,la
soucheJAM1enculturespuresexhibeuneréductiontrèsrapidedunitratemais
aucuneréductiondel’azotetotal(cequiestnormal,étantdonnéqu’ellenepossède
aucungènederéductiondunitrite).
0
5
10
15
20
25
30
35
40
0 50 100 150 200
concentrationd'azotetotale(m
Md'azote)
Temps(h)
co1:1
co1:2
co1:5
co1:10
NL23
JAM1

39
Pourlaprésenteexpérience,lesco-cultures1:5ontdémontrélaplusgrande
consomptiond’azote,suividesco-cultures1:10.Ilfautnoterquecesdeuxco-
culturesontconsomméplusrapidementlenitrateetlenitritequelesculturespures
delasoucheNL23,cequipourraitêtreunindicedelacoopérationbénéfiquedes
deuxsouchesdanscesco-cultures.Parcontre,lesco-cultures1:1et1:2ontété
moinsefficacesquelasoucheNL23enculturespures.Aucunedesco-culturesn’a
démontréuneréductioncomplètedel’azotetotaleavant200heuresdesuivide
croissance.
Malgrélacroissanceréussiedesdeuxsouchesenculturespuresetenco-
cultures,cessuivisontsoulevéuneproblématiqueimportante:lesco-culturesne
sontjamaisarrivéesàréduirecomplètementlenitrateleurétantfourniaudépart,ni
l’azotetotal.Eneffet,lesco-culturesàratiosplusinégauxontsimplementdémontré
uneréductiontrèslentedunitrate.Lesco-culturesàratiospluségauxsesont
comportéesdefaçonpresqueidentiqueàlasoucheJAM1enculturepure:une
réductionrapide(maisincomplète)dunitratesuivid’unestagnationd’activité
dénitrifiante,incluantuneaccumulationdenitrite,ceciendépitdelaprésencedela
soucheNL23,quialacapacitéderéduirecenitrite.Uneexplicationprobableadonc
étéformulée:étantdonnéquelenitriteestparticulièrementtoxiqueaux
organismesvivants(DeRoosetal.,2003),sonaccumulationenimportante
concentrationinhiberaitlacroissancedelasoucheNL23.CommelasoucheJAM1
estbeaucoupplusrapidepourréduirelenitratequelasoucheNL23,ilestprobable
quecelle-ciaitétéresponsabled’uneaccumulationdenitritesupérieureàcequela
soucheNL23peutsupporter.Enconséquence,sil’abondancedunitritepouvaitêtre
limitée,lasoucheNL23pourraitavoirlachancederéduirecenitriteàmesurequ’il
soitproduit.
40
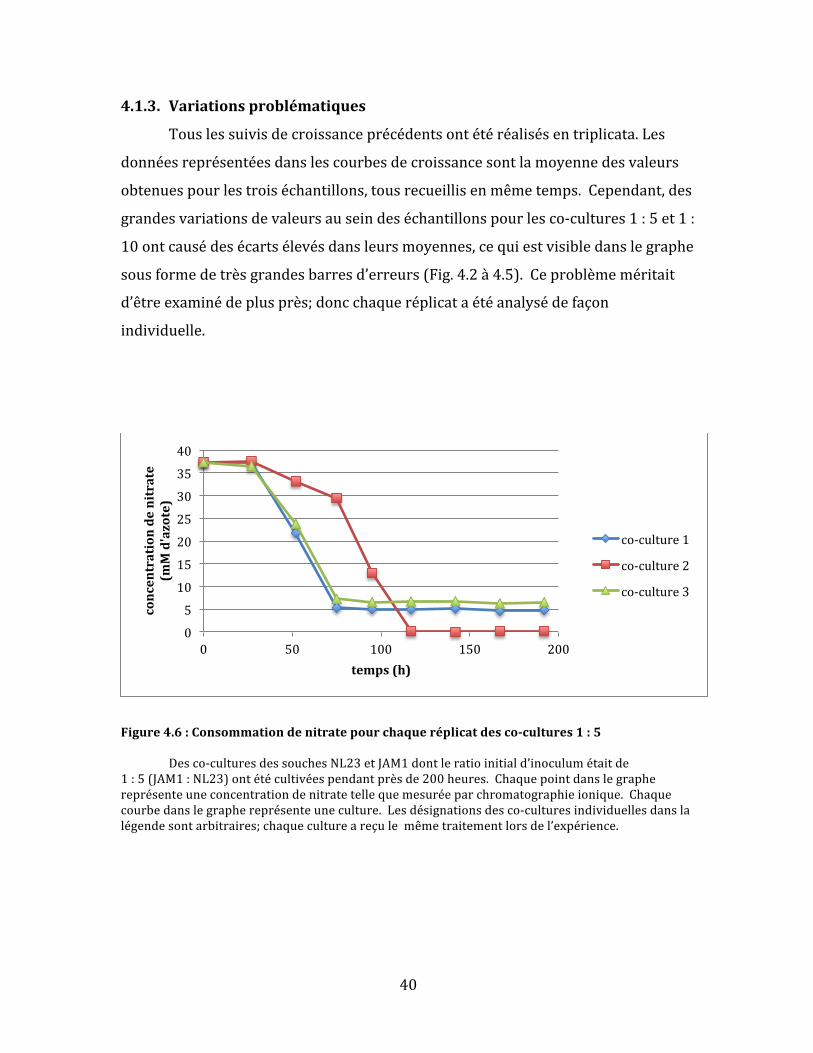
4.1.3. Variationsproblématiques
Touslessuivisdecroissanceprécédentsontétéréalisésentriplicata.Les
donnéesreprésentéesdanslescourbesdecroissancesontlamoyennedesvaleurs
obtenuespourlestroiséchantillons,tousrecueillisenmêmetemps.Cependant,des
grandesvariationsdevaleursauseindeséchantillonspourlesco-cultures1:5et1:
10ontcausédesécartsélevésdansleursmoyennes,cequiestvisibledanslegraphe
sousformedetrèsgrandesbarresd’erreurs(Fig.4.2à4.5).Ceproblèmeméritait
d’êtreexaminédeplusprès;doncchaqueréplicataétéanalysédefaçon
individuelle.
Figure4.6:Consommationdenitratepourchaqueréplicatdesco-cultures1:5 Desco-culturesdessouchesNL23etJAM1dontleratioinitiald’inoculumétaitde1:5(JAM1:NL23)ontétécultivéespendantprèsde200heures.Chaquepointdanslegraphereprésenteuneconcentrationdenitratetellequemesuréeparchromatographieionique.Chaquecourbedanslegraphereprésenteuneculture.Lesdésignationsdesco-culturesindividuellesdanslalégendesontarbitraires;chaquecultureareçulemêmetraitementlorsdel’expérience.
0
5
10
15
20
25
30
35
40
0 50 100 150 200
concentrationdenitrate
(mMd'azote)
temps(h)
co-culture1
co-culture2
co-culture3
41
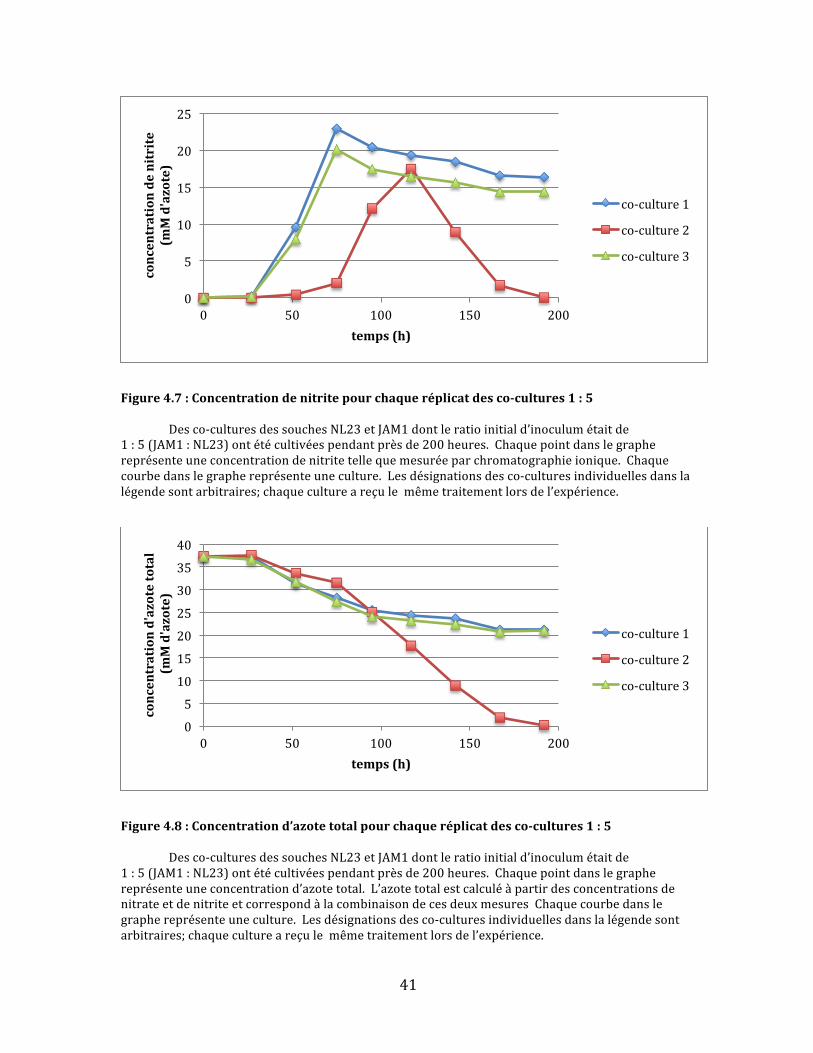
Figure4.7:Concentrationdenitritepourchaqueréplicatdesco-cultures1:5 Desco-culturesdessouchesNL23etJAM1dontleratioinitiald’inoculumétaitde1:5(JAM1:NL23)ontétécultivéespendantprèsde200heures.Chaquepointdanslegraphereprésenteuneconcentrationdenitritetellequemesuréeparchromatographieionique.Chaquecourbedanslegraphereprésenteuneculture.Lesdésignationsdesco-culturesindividuellesdanslalégendesontarbitraires;chaquecultureareçulemêmetraitementlorsdel’expérience.
Figure4.8:Concentrationd’azotetotalpourchaqueréplicatdesco-cultures1:5 Desco-culturesdessouchesNL23etJAM1dontleratioinitiald’inoculumétaitde1:5(JAM1:NL23)ontétécultivéespendantprèsde200heures.Chaquepointdanslegraphereprésenteuneconcentrationd’azotetotal.L’azotetotalestcalculéàpartirdesconcentrationsdenitrateetdenitriteetcorrespondàlacombinaisondecesdeuxmesuresChaquecourbedanslegraphereprésenteuneculture.Lesdésignationsdesco-culturesindividuellesdanslalégendesontarbitraires;chaquecultureareçulemêmetraitementlorsdel’expérience.
0
5
10
15
20
25
0 50 100 150 200
concentrationdenitrite
(mMd'azote)
temps(h)
co-culture1
co-culture2
co-culture3
0
5
10
15
20
25
30
35
40
0 50 100 150 200
concentrationd'azotetotal
(mMd'azote)
temps(h)
co-culture1
co-culture2
co-culture3

42
Danslecasdesco-cultures1:5,deuxdestroisculturesontsuiviunecourbe
deréductiondenitratequasimentidentiquequirappellelecomportementdela
soucheJAM1:uneréductionrapidedunitratesuivid’unarrêtbrusqueà5mM
d’azoteaprèsenviron75heuresdecroissance(Fig.4.6).L’autrecultureaétéplus
lentedanslaréductiontotaledunitrate.D’ailleurs,cettedernièreestlaseuledes
troisquifinitparégalementréduirecomplètementlenitrite(Fig.4.7)etàaccomplir
uneconsommationcomplètedel’azotetotal(Fig.4.8).Lenitrites’yestaussi
accumulépluslentement.Ilsemblaitdoncquelarapiditéàlaquellelenitrite
s’accumulepourraiteffectivementjouerunrôledanslaperformanced’uneco-
culture.
Figure4.9:Consommationdenitratepourchaqueréplicatdesco-cultures1:10 Desco-culturesdessouchesNL23etJAM1dontleratioinitiald’inoculumétaitde1:10(JAM1:NL23)ontétécultivéespendantprèsde200heures.Chaquepointdanslegraphereprésenteuneconcentrationdenitratetellequemesuréeparchromatographieionique.Chaquecourbedanslegraphereprésenteuneculture.Lesdésignationsdesco-culturesindividuellesdanslalégendesontarbitraires;chaquecultureareçulemêmetraitementlorsdel’expérience.
0
5
10
15
20
25
30
35
40
0 50 100 150 200
concentrationdenitrate
(mMd'azote)
temps(h)
co-culture1
co-culture2
co-culture3
43
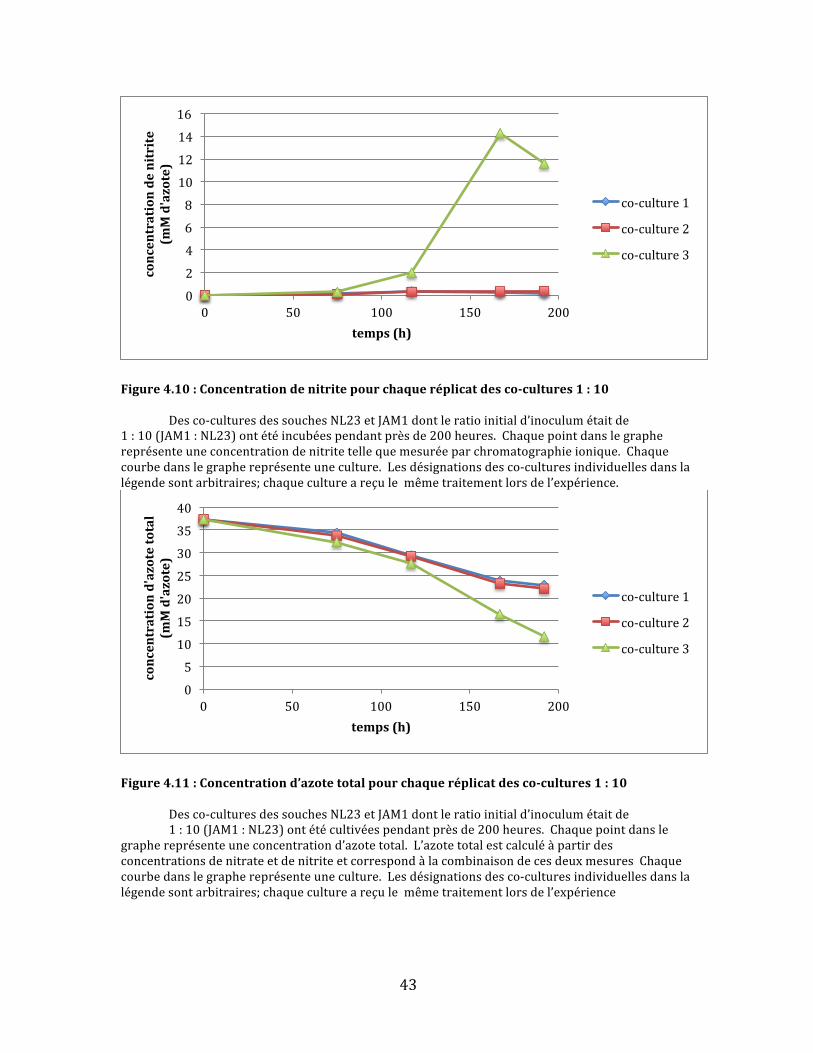
Figure4.10:Concentrationdenitritepourchaqueréplicatdesco-cultures1:10 Desco-culturesdessouchesNL23etJAM1dontleratioinitiald’inoculumétaitde1:10(JAM1:NL23)ontétéincubéespendantprèsde200heures.Chaquepointdanslegraphereprésenteuneconcentrationdenitritetellequemesuréeparchromatographieionique.Chaquecourbedanslegraphereprésenteuneculture.Lesdésignationsdesco-culturesindividuellesdanslalégendesontarbitraires;chaquecultureareçulemêmetraitementlorsdel’expérience.
Figure4.11:Concentrationd’azotetotalpourchaqueréplicatdesco-cultures1:10 Desco-culturesdessouchesNL23etJAM1dontleratioinitiald’inoculumétaitde
1:10(JAM1:NL23)ontétécultivéespendantprèsde200heures.Chaquepointdanslegraphereprésenteuneconcentrationd’azotetotal.L’azotetotalestcalculéàpartirdesconcentrationsdenitrateetdenitriteetcorrespondàlacombinaisondecesdeuxmesuresChaquecourbedanslegraphereprésenteuneculture.Lesdésignationsdesco-culturesindividuellesdanslalégendesontarbitraires;chaquecultureareçulemêmetraitementlorsdel’expérience
0
2
4
6
8
10
12
14
16
0 50 100 150 200
concentrationdenitrite
(mMd'azote)
temps(h)
co-culture1
co-culture2
co-culture3
0
5
10
15
20
25
30
35
40
0 50 100 150 200
concentrationd'azotetotal
(mMd'azote)
temps(h)
co-culture1
co-culture2
co-culture3

44
Danslegroupedesco-cultures1:10,c’estleproblèmeinversequis’est
manifesté:seuleunedesco-culturesapuréduiretotalementlenitrate,tandisque
lesdeuxautresontététroplentespouryarriveravant200heures.Deplus,une
augmentationdenitriten’aétéobservéequedanslaco-culturelaplusefficaceà
partird’environ100heures(Fig.4.9et4.10).Malgrécetteaccumulation,cette
dernièreadémontréunréductiond’azotetotallégèrementplusrapidequelesdeux
autres,maisquin’atoutdemêmepasatteint0mMd’azoteavantlafindusuivi(Fig.
4.11).
Pourliercesrésultatsàlaquantitérelativedesdeuxsouchesdanslesco-
cultures,desessaisenqPCRontétéeffectués.Envisantdesgènesquisontpropres
àchaqueespèce,ilaétépossibledequantifierl’abondancedechaquesoucheen
pourcentageparrapportaunombretotaldegènesdétectés.
Figure4.12:QuantitésrelativesdesgènesnapAetnarG1danslesco-cultures1:5 Troisco-culturesdessouchesNL23etJAM1dontleratioinitiald’inoculumétaitde1:5(JAM1:NL23)ontétécultivéespendantprèsde200heures.Àlafindecesuivi,labiomassetotaledechacunedesculturesaétérecueillie.L’ADNgénomiquedecettebiomasseaensuiteétéextraitetsoumisàdeuxprocessusdeqPCR:unepourquantifierl’abondancedugènenapAetl’autrepourquantifierl’abondancedugènenarG1.Lespourcentagessurlesbandesreprésententlaproportiondechaquegènedanslaquantitétotaledegènesquantifiés(napA+narG1)Dansuncasparticulier,laco-culture3aétéaccidentellementperduelorsdemanipulationsenlaboratoire.
45
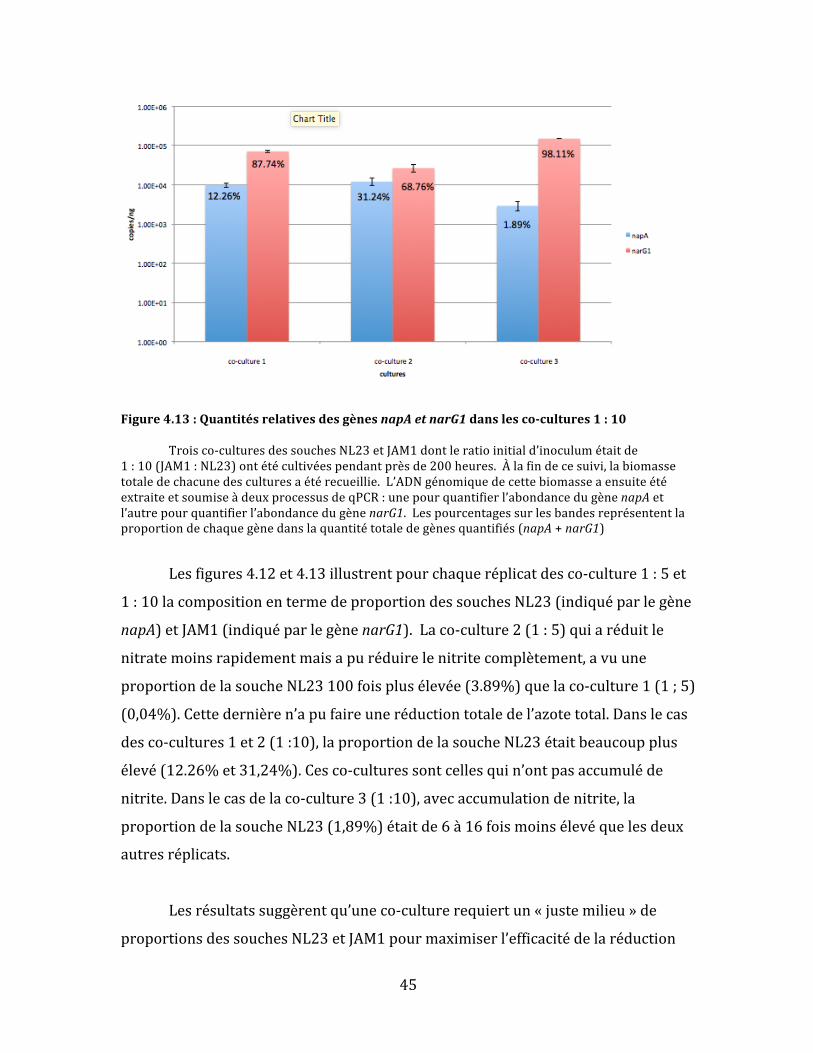
Figure4.13:QuantitésrelativesdesgènesnapAetnarG1danslesco-cultures1:10 Troisco-culturesdessouchesNL23etJAM1dontleratioinitiald’inoculumétaitde1:10(JAM1:NL23)ontétécultivéespendantprèsde200heures.Àlafindecesuivi,labiomassetotaledechacunedesculturesaétérecueillie.L’ADNgénomiquedecettebiomasseaensuiteétéextraiteetsoumiseàdeuxprocessusdeqPCR:unepourquantifierl’abondancedugènenapAetl’autrepourquantifierl’abondancedugènenarG1.Lespourcentagessurlesbandesreprésententlaproportiondechaquegènedanslaquantitétotaledegènesquantifiés(napA+narG1)
Lesfigures4.12et4.13illustrentpourchaqueréplicatdesco-culture1:5et
1:10lacompositionentermedeproportiondessouchesNL23(indiquéparlegène
napA)etJAM1(indiquéparlegènenarG1).Laco-culture2(1:5)quiaréduitle
nitratemoinsrapidementmaisapuréduirelenitritecomplètement,avuune
proportiondelasoucheNL23100foisplusélevée(3.89%)quelaco-culture1(1;5)
(0,04%).Cettedernièren’apufaireuneréductiontotaledel’azotetotal.Danslecas
desco-cultures1et2(1:10),laproportiondelasoucheNL23étaitbeaucoupplus
élevé(12.26%et31,24%).Cesco-culturessontcellesquin’ontpasaccumuléde
nitrite.Danslecasdelaco-culture3(1:10),avecaccumulationdenitrite,la
proportiondelasoucheNL23(1,89%)étaitde6à16foismoinsélevéquelesdeux
autresréplicats.
Lesrésultatssuggèrentqu’uneco-culturerequiertun«justemilieu»de
proportionsdessouchesNL23etJAM1pourmaximiserl’efficacitédelaréduction

46
dunitrateetdunitritetoutenévitantlatoxicitédunitrite.Malheureusement,il
s’estavérédifficiled’obtenirceparfaitmélangedefaçonassezconstantepour
produiredesrésultatsentriplicata,etlavariabilitéimprévisibledesrésultatsa
empêchéleprogrèsduprojetdanscettedirection.
4.1.4. Réductiondelaquantitéinitialedenitrate
Unenouvellestratégieaétéétabliepourcontournerleproblème
d’accumulationdenitrite.L’idéeétaitdediminuerlaconcentrationinitialede
nitratedanslemilieu«InstantOcean»pourréduirel’accumulationmaximalede
nitrite.Danslesessaisprécédents(Auclairetal.,2010),ilsemblaitêtrepossible
pourlesco-culturesdetoléreruncertainniveaudenitrite(jusqu’àenviron15mM
d’azote)avantd’éprouverdesdifficultés.L’hypothèseaétéémisequ’une
accumulationdenitritesousceniveauéviteraitleproblèmedetoxicité.
Pourvalidercettenouvelleapproche,l’expérienceprécédente(voirsection
4.1.2)aétérépétéeentriplicata,maisavecdumilieu«InstantOcean»contenant
environ15mMdenitrateaulieude40mM.Aveccettequantitéinitialedenitrate,
laconcentrationmaximalethéoriquepossibledenitriteestd’environ15mM.De
plus,cetteconcentrationcorrespondà-peu-prèsauniveaumaximalobservédansle
mésocosmedufleuveSaint-Laurentaubiodôme(Labbéetal.,2003).

47
Figure4.14:Consommationdenitratedesco-culturesdessoucheNL23etJAM1enconditionsanaérobies Desco-culturesdessouchesNL23etJAM1ontétécultivéesdanslemilieuInstantOcean15g/Lpendantenviron130heures.Lesco-culturesavaientdesratiosd’inoculumsdifférents.Chaquepointdanslegraphereprésentelamoyennedeconcentrationdenitratedans3cultures,mesuréeparchromatographieionique.
Figure4.15:Concentrationdenitritedanslesco-culturesdessoucheNL23etJAM1enanaérobie Desco-culturesdessouchesNL23etJAM1ontétéincubéesdanslemilieuInstantOcean15g/Lpendantenviron130heures.Lesco-culturesavaientdesratiosd’inoculumsdifférents.Chaquepointdanslegraphereprésentelamoyennedeconcentrationdenitritedans3cultures,mesuréeparchromatographieionique.
0
2
4
6
8
10
12
0 20 40 60 80 100 120 140
concentration(mMd'azote)
temps(h)
NL23
co-culture1:1
co-culture1:2
co-culture1:5
co-culture1:10
0
2
4
6
8
10
12
0 20 40 60 80 100 120 140
concentration(mMd'azote)
temps(h)
NL23
co-culture1:1
co-culture1:2
co-culture1:5
co-culture1:10
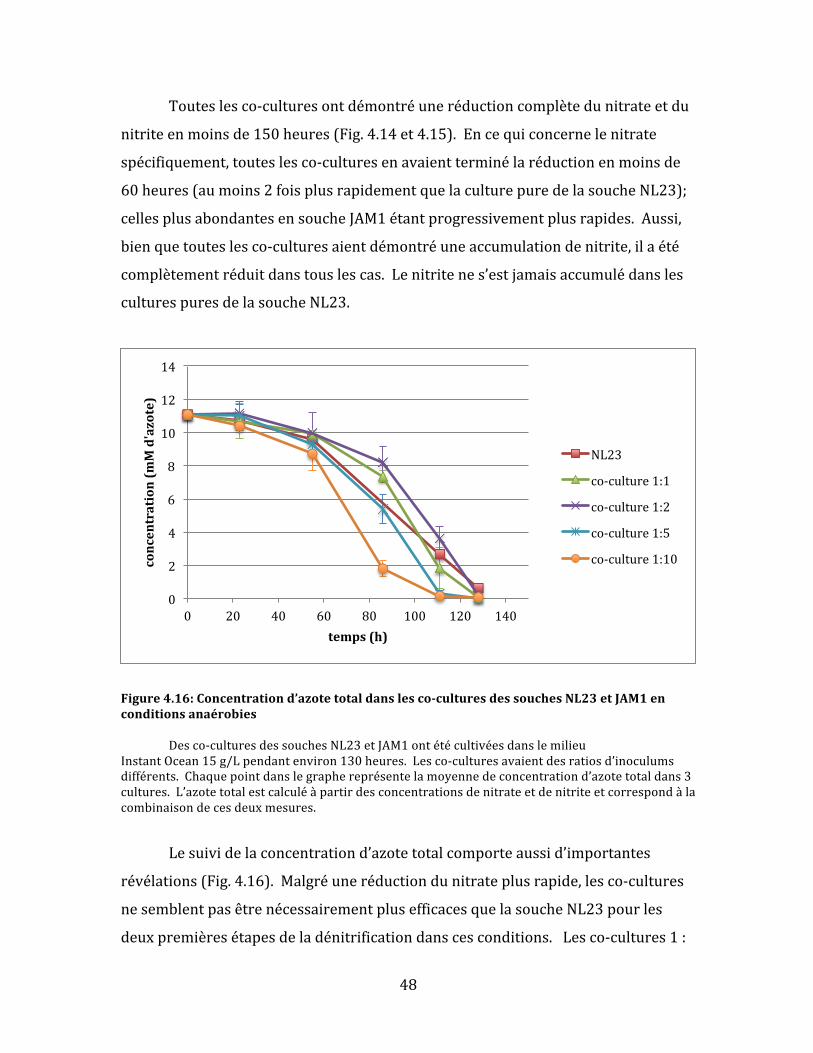
48
Touteslesco-culturesontdémontréuneréductioncomplètedunitrateetdu
nitriteenmoinsde150heures(Fig.4.14et4.15).Encequiconcernelenitrate
spécifiquement,touteslesco-culturesenavaientterminélaréductionenmoinsde
60heures(aumoins2foisplusrapidementquelaculturepuredelasoucheNL23);
cellesplusabondantesensoucheJAM1étantprogressivementplusrapides.Aussi,
bienquetouteslesco-culturesaientdémontréuneaccumulationdenitrite,ilaété
complètementréduitdanstouslescas.Lenitritenes’estjamaisaccumulédansles
culturespuresdelasoucheNL23.
Figure4.16:Concentrationd’azotetotaldanslesco-culturesdessouchesNL23etJAM1enconditionsanaérobies Desco-culturesdessouchesNL23etJAM1ontétécultivéesdanslemilieuInstantOcean15g/Lpendantenviron130heures.Lesco-culturesavaientdesratiosd’inoculumsdifférents.Chaquepointdanslegraphereprésentelamoyennedeconcentrationd’azotetotaldans3cultures.L’azotetotalestcalculéàpartirdesconcentrationsdenitrateetdenitriteetcorrespondàlacombinaisondecesdeuxmesures.
Lesuividelaconcentrationd’azotetotalcomporteaussid’importantes
révélations(Fig.4.16).Malgréuneréductiondunitrateplusrapide,lesco-cultures
nesemblentpasêtrenécessairementplusefficacesquelasoucheNL23pourles
deuxpremièresétapesdeladénitrificationdanscesconditions.Lesco-cultures1:
0
2
4
6
8
10
12
14
0 20 40 60 80 100 120 140
concentration(mMd'azote)
temps(h)
NL23
co-culture1:1
co-culture1:2
co-culture1:5
co-culture1:10

49
5et1:10ontdémontréiciuneréductionlégèrementplusrapide.L’étapelimitante
danscesystèmeestdonclaréductiondunitriteparlasoucheNL23.Ilsemble
probablequesontauxdeconsommationdunitriten’estpasouestpeuaccélérépar
laprésencedelasoucheJAM1ouuneréductionaccéléréedunitrate.
Pourtouteslesexpériencesdesuividecroissancesubséquentes,lesco-
culturesontétépréparéesavecunratioinitialde1:5(JAM1:NL23)etenviron15
mMdenitrateinitial.
4.2. CaractérisationdelarelationentrelessouchesNL23etJAM1
4.2.1. Croissancedeculturespuresenprésencedesurnageant
Enplusdupartagedunitrate/nitrite,ilaétéintéressantdechercherdes
indicesdecoopérationentrelessouchesNL23etJAM1.Lapremièretentativede
trouverunetelleinteractionavaitpourbutdedéterminers’ilyavaitexistencede
moléculessécrétéesdanslemilieuparchaquesouchequipouvaientaiderla
croissancedel’autre.L’expérienceaétéexécutéecommesuit:
- DesculturespuresdessouchesNL23etJAM1ainsiquedesco-culturesdes
deuxontétéeffectuéesenconditionsaérobiesdanslemilieu«Instant
Ocean»15g/L.
- Unefoisquelacroissancemaximaleaitétéatteinte,cesculturesontété
centrifugéespourrecueillirlesurnageant.Lescellulesrestantesdansce
surnageantontensuiteétéenlevéesàl’aided’uneseringuemunied’unfiltre.
- Chaquesoucheaétéinoculéedanslesurnageantrécoltédelasouche
opposée(soucheNL23danslesurnageantdelasoucheJAM1etvice-versa).
- Chaquesoucheaaussiétéinoculéedanssonpropresurnageant,dansle
surnageantdelaco-culturedesdeuxsouches(àproportionségales)etd’un
surnageanttémoinnégatif(milieunon-inoculé).

50
- 50µldechacundecessurnageantsensemencésaensuiteétémélangéà150
µldemilieu«InstantOcean»frais.
- LesDO600ontétérécoltéespendantlacroissancedesculturesrésultantesà
l’aidedel’instrumentBioscreenàchaquedemi-heurependant48heures.
Lesessaisdevaientêtrefaitsavecdesculturespurespouréviterl’addition
desdeuxsouchesdanslesmesuresdesDO600.
Figure4.17:CroissanceenconditionsaérobiesdelasoucheNL23enprésencedesurnageant LasoucheNL23aétécultivéeenconditionsaérobiesdansplusieursconditionspendant48heures.LesmilieuxutilisésétaienttouscomposésdeInstantOcean15g/Làlabase,maischaquevariationcontenaitaussilesurnageantd’unecultureparticulière.LacroissancedelasoucheNL23danssonmilieuoptimal(337a)estaussiincluspourcomparer.Chaquepointdanslegraphereprésentelamoyennedeturbiditéde8cultures,mesuréparl’appareilBioscreenàtouteslesdemi-heures.
-0.2
-0.1
0
0.1
0.2
0.3
0.4
0.5
0.0 10.0 20.0 30.0 40.0 50.0
DensitéOptique(600nm)
Temps(h)
milieuoptimal
srngt.JAM1
srngt.contrôle
srngt.co-culture

51
Lesrésultatsdémontrentclairementqu’en48heureslasoucheNL23a
atteintunDO600nettementplusélevéeenprésencedusurnageantdelasouche
JAM1qu’enprésencedusurnageantdutémoinnégatif(Fig.4.17).Lesurnageant
desco-culturessembleavoireuuneffetpotentielégalement,maismoinssignificatif.
Étantdonnéquelacroissancedesculturesdutémoinnégatifetdusurnageantdes
co-culturesn’avaitpasatteintsonplateau,ilestpossiblequecesdernièresauraient
éventuellementatteintunDO600semblableàcelledelacultureavecsurnageantde
lasoucheJAM1.LacroissancelasoucheNL23enprésencedesonpropre
surnageantadémontréàpeuprèslamêmecourbequeletémoinnégatifetn’estpas
inclusdanslegraphetelqueprésentéici.L’autreélémentdugrapheestlacourbe
decroissancedelasoucheNL23danssonmilieuoptimal(337a)sansmodifications.
Leprofildecroissanceenmilieu337as’alignepresqueparfaitementavecceluidela
culturedanslesurnageantdelasoucheJAM1.Ainsi,lasoucheNL23cultivéeen
milieu«InstantOcean»(quin’estpasunmilieuoptimal)enprésencedusurnageant
d’uneculturedelasoucheJAM1aétécapabled’atteindreunniveaudecroissance
quiressembleàceluidesacultureenmilieuoptimal.Puisquelemilieu337aestun
milieuminéralnon-salin,cecilaissepeut-êtreentrevoirlerôled’unosmoprotecteur.
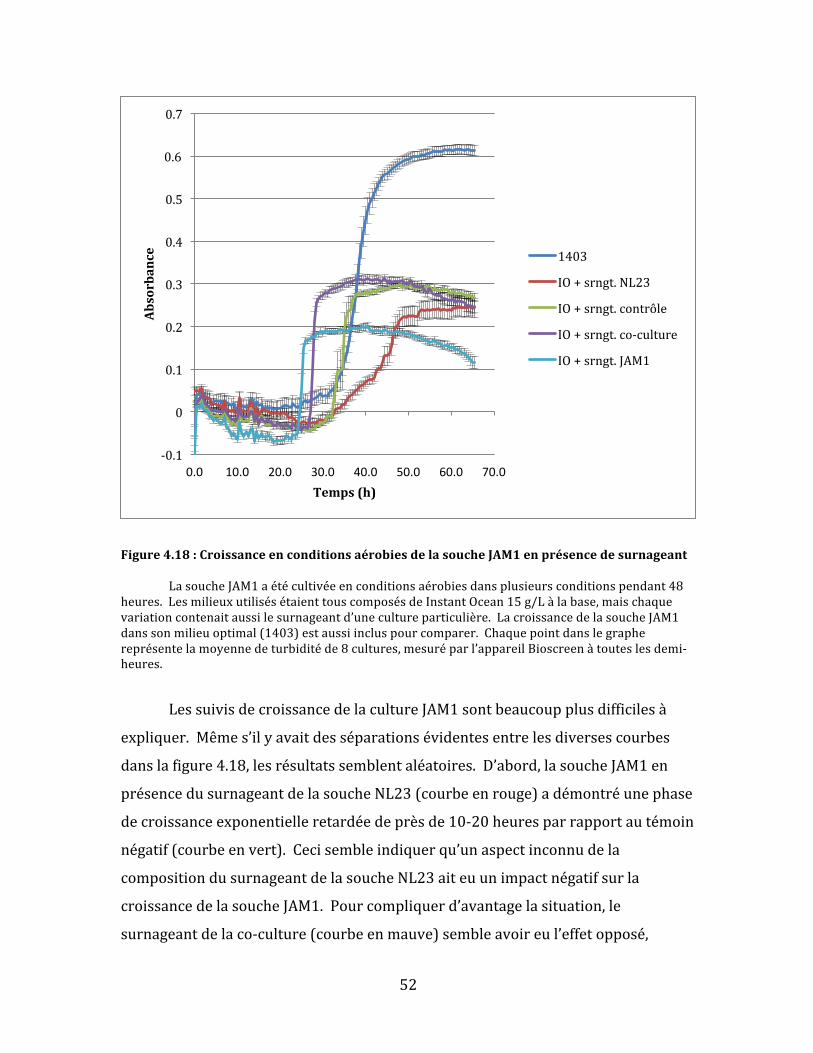
52
Figure4.18:CroissanceenconditionsaérobiesdelasoucheJAM1enprésencedesurnageant LasoucheJAM1aétécultivéeenconditionsaérobiesdansplusieursconditionspendant48heures.LesmilieuxutilisésétaienttouscomposésdeInstantOcean15g/Làlabase,maischaquevariationcontenaitaussilesurnageantd’unecultureparticulière.LacroissancedelasoucheJAM1danssonmilieuoptimal(1403)estaussiincluspourcomparer.Chaquepointdanslegraphereprésentelamoyennedeturbiditéde8cultures,mesuréparl’appareilBioscreenàtouteslesdemi-heures.
LessuivisdecroissancedelacultureJAM1sontbeaucoupplusdifficilesà
expliquer.Mêmes’ilyavaitdesséparationsévidentesentrelesdiversescourbes
danslafigure4.18,lesrésultatssemblentaléatoires.D’abord,lasoucheJAM1en
présencedusurnageantdelasoucheNL23(courbeenrouge)adémontréunephase
decroissanceexponentielleretardéedeprèsde10-20heuresparrapportautémoin
négatif(courbeenvert).Cecisembleindiquerqu’unaspectinconnudela
compositiondusurnageantdelasoucheNL23aiteuunimpactnégatifsurla
croissancedelasoucheJAM1.Pourcompliquerd’avantagelasituation,le
surnageantdelaco-culture(courbeenmauve)sembleavoireul’effetopposé,
-0.1
0
0.1
0.2
0.3
0.4
0.5
0.6
0.7
0.0 10.0 20.0 30.0 40.0 50.0 60.0 70.0
Absorbance
Temps(h)
1403
IO+srngt.NL23
IO+srngt.contrôle
IO+srngt.co-culture
IO+srngt.JAM1

53
puisquelaphaseexponentielledeJAM1estarrivéelégèrement(environ5heures)
plustôtensaprésence.Bienquecerésultatsoitsurprenant,ilpourraitêtre
expliquéparlapossibilitéquelasoucheJAM1émetdesparticulesdurantsa
croissancequiluisontbénéfiques,tandisquelacroissancedelasoucheNL23
résulteenlasécrétiondeparticulesquisontnéfastesàsacroissance.Lacroissance
delasoucheJAM1enprésencedesonpropresurnageant(courbeenbleupâle)
semblaitcorroborer,enpartie,cettehypothèse,maisaatteintunedensitémaximale
notammentinférieureauxautrescultures.Lestroisautresculturesontatteintun
plateauàpeuprèségal,etlaculturedelasoucheJAM1enmilieuoptimal(1403)
(courbeenbleufoncé)aatteintunDO600quienestdeuxfoissupérieures.Malgré
lanatureinconnuedesdétails,unechoseétaitcertaine:lasoucheNL23nesemble
pasfournird’aideàlacroissancedelasoucheJAM1enconditionsaérobies.
4.2.2. Recherched’unosmoprotecteur
CommelasoucheJAM1estadaptéeàdesconditionsdemilieuxsalins,une
despossibilitésintéressantesdanssoncasseraitlaproductionpotentielled’un
osmoprotecteur,unemoléculecapabledeprotégerunebactériecontrelesstress
osmotiques(parexemple:unehauteconcentrationensel).Avantledébutdece
projet,desrecherchesbioinformatiquesontmontréquelegénomedelasouche
JAM1contientdesgènespermettantlaproductiond’ectoïne,unemolécule
osmoprotectrice.LasoucheNL23aunerésistancelimitéeauxenvironnements
salés,doncl’existencedecettemoléculedansuneco-cultureseraitd’une
importancesignificativepourlacompréhensiondelarelationentrelesdeux
espèces.L’ectoïnepourraitaussiexpliquerenpartiel’aidedecroissancequela
soucheJAM1fournitàlasoucheNL23,tellequ’observédansl’expériencedela
sectionprécédente.
Uneexpérienceadoncétédéveloppéepourvérifierl’existenced’une
protectionosmotiquedelapartdelasoucheJAM1enverslasoucheNL23.Le
protocoled’expérienceaétébasésurleprocessusde«BacterialMilking»(Saueret
al.,1998),quiajustementétédéveloppéepourlarécoltedesolutésincluant

54
l’ectoïne.LasoucheJAM1ad’abordétécultivéedansuneversionmodifiéedeson
milieuoptimal(1403),oùletauxdesalinitéaétéaugmentéà6%NaCl(aulieude
2,4%).Lesculturesontensuiteétécentrifugées,etlesculotsdispersésdansle
milieuoptimaldelasoucheNL23(337a).L’espoirétaitquelaproductionde
moléculesosmoprotectricesinduiteparlemilieuhautementsalinrésulteenla
présencedecesparticulesdanslenouveaumilieu.Aprèsenviron6heuresdansle
milieu337a,lescellulesontétécentrifugéesdenouveau,etcettefoislesurnageanta
étéconservé.Celui-ciaétéséparéenplusieurstubesdanslesquelsdesquantités
progressivementcroissantesdeselontétéajoutées.
Tableau4.1:concentrationdeNaClpartubedanslarecherched’unosmoprotecteur
tube PourcentageNaCL(masse)1 0%(0,00g)2 1%(0,05g)3 1,5%(0,075g)4 2,0%(0,10g)
LasoucheNL23aensuiteétéinoculéedanschacundesnouveauxmilieux,etles
culturesontétésuiviesàl’aidedel’appareilBioscreenpendant48heures.La
soucheNL23aégalementétécultivéedanslemilieu337anon-modifié(pasexposé
auxcellulesdelasoucheJAM1)àtitredecomparaison(Fig.4.19à4.22).
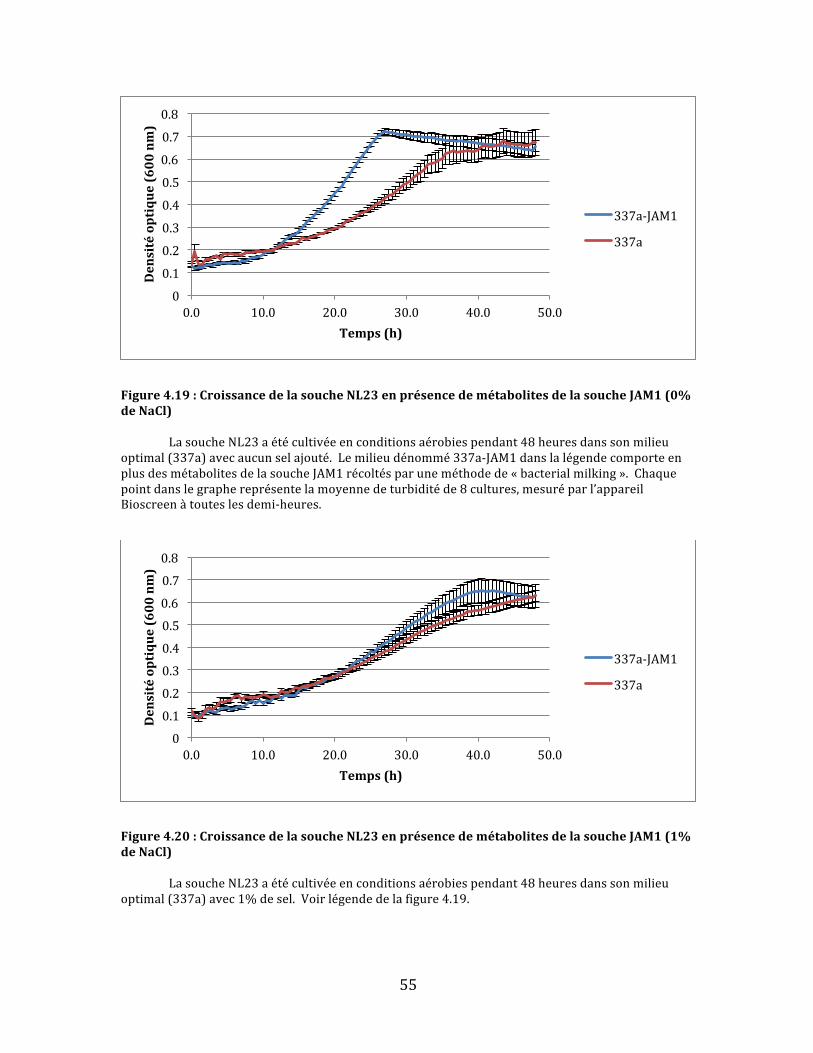
55
Figure4.19:CroissancedelasoucheNL23enprésencedemétabolitesdelasoucheJAM1(0%deNaCl) LasoucheNL23aétécultivéeenconditionsaérobiespendant48heuresdanssonmilieuoptimal(337a)avecaucunselajouté.Lemilieudénommé337a-JAM1danslalégendecomporteenplusdesmétabolitesdelasoucheJAM1récoltésparuneméthodede«bacterialmilking».Chaquepointdanslegraphereprésentelamoyennedeturbiditéde8cultures,mesuréparl’appareilBioscreenàtouteslesdemi-heures.
Figure4.20:CroissancedelasoucheNL23enprésencedemétabolitesdelasoucheJAM1(1%deNaCl) LasoucheNL23aétécultivéeenconditionsaérobiespendant48heuresdanssonmilieuoptimal(337a)avec1%desel.Voirlégendedelafigure4.19.
0
0.1
0.2
0.3
0.4
0.5
0.6
0.7
0.8
0.0 10.0 20.0 30.0 40.0 50.0
Densitéoptique(600nm)
Temps(h)
337a-JAM1
337a
0
0.1
0.2
0.3
0.4
0.5
0.6
0.7
0.8
0.0 10.0 20.0 30.0 40.0 50.0
Densitéoptique(600nm)
Temps(h)
337a-JAM1
337a
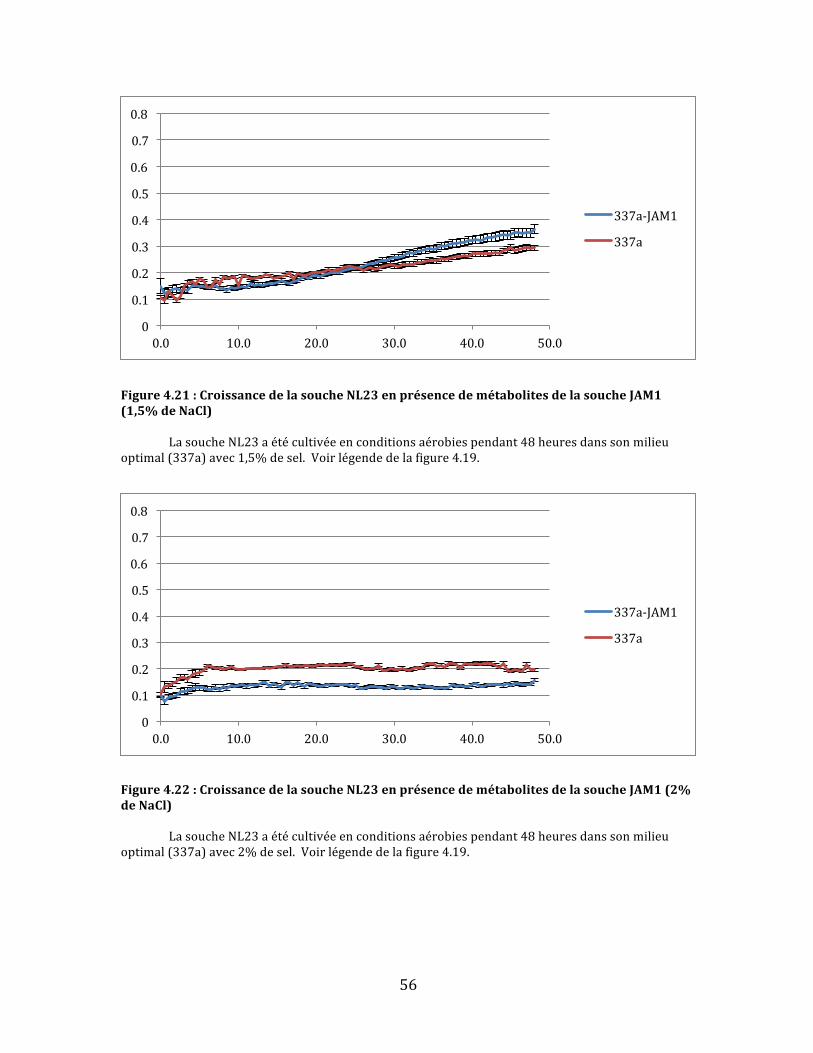
56
Figure4.21:CroissancedelasoucheNL23enprésencedemétabolitesdelasoucheJAM1(1,5%deNaCl) LasoucheNL23aétécultivéeenconditionsaérobiespendant48heuresdanssonmilieuoptimal(337a)avec1,5%desel.Voirlégendedelafigure4.19.
Figure4.22:CroissancedelasoucheNL23enprésencedemétabolitesdelasoucheJAM1(2%deNaCl) LasoucheNL23aétécultivéeenconditionsaérobiespendant48heuresdanssonmilieuoptimal(337a)avec2%desel.Voirlégendedelafigure4.19.
0
0.1
0.2
0.3
0.4
0.5
0.6
0.7
0.8
0.0 10.0 20.0 30.0 40.0 50.0
337a-JAM1
337a
0
0.1
0.2
0.3
0.4
0.5
0.6
0.7
0.8
0.0 10.0 20.0 30.0 40.0 50.0
337a-JAM1
337a

57
LesculturesdelasoucheNL23danslemilieu337amodifiéontmontréune
croissanceplusrapidequeletémoinauxconcentrations0%,1%et1,5%deNaCl.
Cetteobservationauraitpuporteràconfusion,maisuneexpérienceprécédentede
ceprojetavaitdéjàdémontréquelasoucheNL23pouvaitcroîtreplusrapidement
enprésenced’unsurnageantdelasoucheJAM1(voirsection4.2.1).Cetaspectdu
résultatn’estdoncpassurprenant.Cequiétaitréellementlepointmajeurdansles
donnéesrecueilliesétaitl’absencedecroissancedanslegraphepourla
concentration2%(Fig.4.22).Eneffet,cecomportementestexactementdequoion
s’attendaitsilasoucheJAM1n’offraitaucuneprotectionosmotique.Toutefois,ilest
possiblequeleprotocoleutilisénepermettaitpasderécolterdesmolécules
osmoprotectrices.Unerépétitiondel’expérienceaconfirmélesrésultatsjusqu’à
uneconcentrationdeselde2,5%.
4.2.3. Caractérisationapprofondiedelaconsommationdunitrate/nitriteà
l’aidedemutantsdelasoucheJAM1
Lorsd’étudesenparallèlesaveccelles-ci,desmutantsdelasoucheJAM1ont
étédéveloppéspouraideràlacaractérisationdesonprofildedénitrification
(Mauffreyetal.,2015).Lestroismutantsenquestionétaienttousrendusdéficients
dansleurhabiletéderéduirelenitrate:
- LasoucheJAM1ΔnarG1(AbsencedusystèmeNar1)
- LasoucheJAM1ΔnarG2(AbsencedusystèmeNar2)
- LasoucheJAM1ΔnarG1narG2(doublemutant)
Commelenitrateestprincipalementimportantdanslecontextedela
dénitrification,l’applicationexpérimentaledesmutantsétaitsurtoutintéressanteen
conditionsanaérobies.Leplanétaitdoncdefaireunsuividecroissancedesco-
culturesdelasoucheNL23avecchacundesmutantsetdelasoucheJAM1non
mutante(ainsiquedesculturespuresdechaquesoucheimpliquéecommetémoin).
Lesco-culturesdelasoucheNL23avecledoublemutantallaientêtre
particulièrementinformatif,puisquel’effetdelasoucheJAM1surlaperformancede

58
lasoucheNL23seraitvisibledanslesuividedénitrificationparchromatographie
ionique.Leratiod’inoculumdedépartpourtouteslesco-culturesétaitde1:5
(nitrate:nitrite),basésurdesrésultatsprécédents(voirsection4.1.2).Lesuivia
étéeffectuésurunepérioded’environ225heuresetleséchantillonsontété
mesurésenDO600etenconcentrationsdenitrateetdenitrite(Fig.4.21à4.23).
Cependant,seulslesrésultatsdelachromatographieioniquesontprésentés,carle
suiviparDO600nerévèlepasbeaucoupd’informationssurl’activitédescultures.
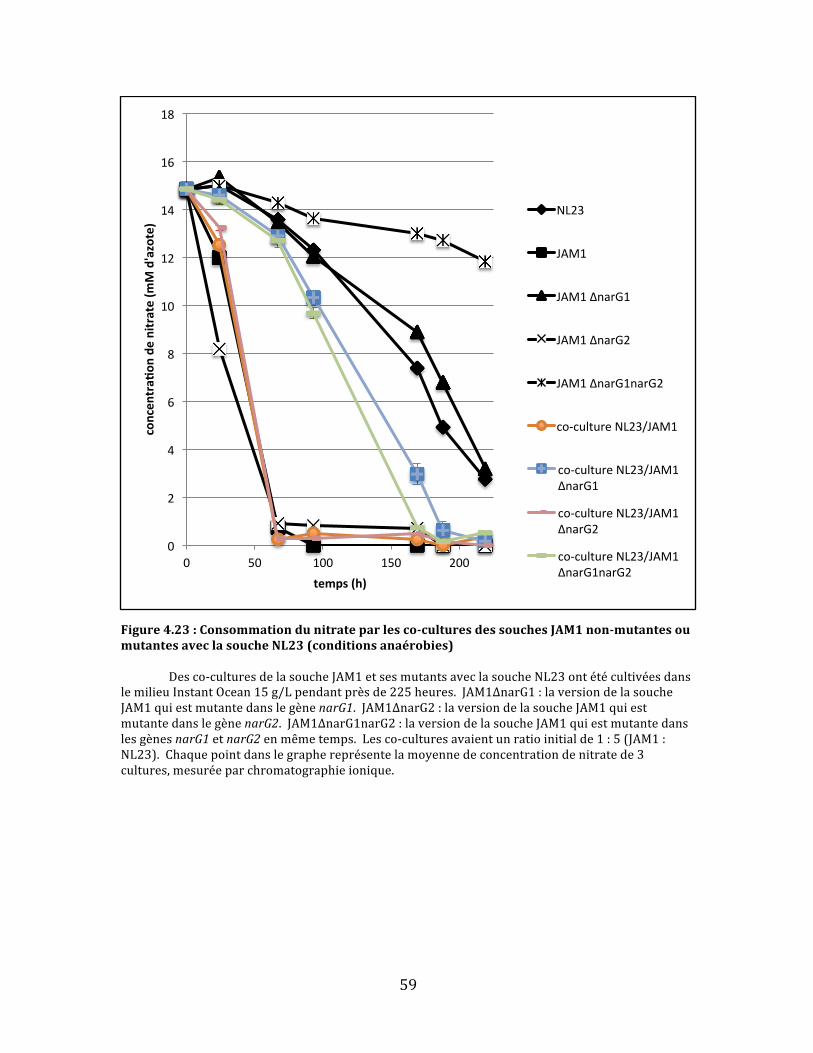
59
Figure4.23:Consommationdunitrateparlesco-culturesdessouchesJAM1non-mutantesoumutantesaveclasoucheNL23(conditionsanaérobies) Desco-culturesdelasoucheJAM1etsesmutantsaveclasoucheNL23ontétécultivéesdanslemilieuInstantOcean15g/Lpendantprèsde225heures.JAM1ΔnarG1:laversiondelasoucheJAM1quiestmutantedanslegènenarG1.JAM1ΔnarG2:laversiondelasoucheJAM1quiestmutantedanslegènenarG2.JAM1ΔnarG1narG2:laversiondelasoucheJAM1quiestmutantedanslesgènesnarG1etnarG2enmêmetemps.Lesco-culturesavaientunratioinitialde1:5(JAM1:NL23).Chaquepointdanslegraphereprésentelamoyennedeconcentrationdenitratede3cultures,mesuréeparchromatographieionique.
0
2
4
6
8
10
12
14
16
18
0 50 100 150 200
concen
tra+
ondenitrate(m
Md'azote)
temps(h)
NL23
JAM1
JAM1∆narG1
JAM1∆narG2
JAM1∆narG1narG2
co-cultureNL23/JAM1
co-cultureNL23/JAM1∆narG1
co-cultureNL23/JAM1∆narG2
co-cultureNL23/JAM1∆narG1narG2

60
Figure4.24:Concentrationdenitritedanslesco-culturesdessouchesJAM1non-mutantesoumutantesaveclasoucheNL23(conditionsanaérobies) Desco-culturesdelasoucheJAM1etsesmutantsaveclasoucheNL23ontétécultivéesdanslemilieuInstantOcean15g/Lpendantprèsde225heures.Voirlalégendedelafigure4.23.Chaquepointdanslegraphereprésentelamoyennedeconcentrationdenitritede3cultures,mesuréeparchromatographieionique.LacourbedelaculturepuredelasoucheJAM1ΔnarG2aétéomisepourdesraisonsdevisibilité.Celle-cidémontraituneaccumulationdenitritesimilaireàlacourbedelasoucheJAM1.
0
2
4
6
8
10
12
0 50 100 150 200
concentrationdenitrite(m
Md'azote)
temps(h)
NL23
JAM1∆narG1narG2
co-cultureNL23/JAM1
co-cultureNL23/JAM1∆narG1narG2
co-cultureNL23/JAM1ΔnarG1
co-cultureNL23/JAM1ΔnarG2
JAM1
JAM1ΔnarG1
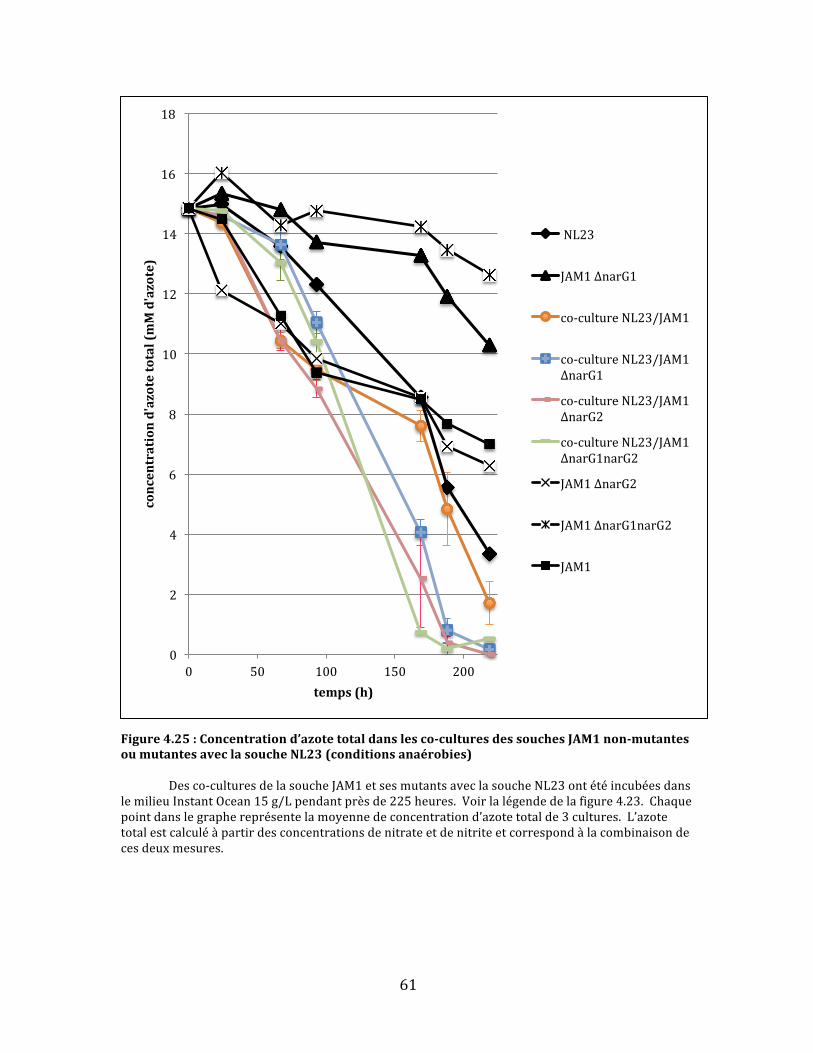
61
Figure4.25:Concentrationd’azotetotaldanslesco-culturesdessouchesJAM1non-mutantesoumutantesaveclasoucheNL23(conditionsanaérobies) Desco-culturesdelasoucheJAM1etsesmutantsaveclasoucheNL23ontétéincubéesdanslemilieuInstantOcean15g/Lpendantprèsde225heures.Voirlalégendedelafigure4.23.Chaquepointdanslegraphereprésentelamoyennedeconcentrationd’azotetotalde3cultures.L’azotetotalestcalculéàpartirdesconcentrationsdenitrateetdenitriteetcorrespondàlacombinaisondecesdeuxmesures.
0
2
4
6
8
10
12
14
16
18
0 50 100 150 200
concentrationd'azotetotal(mMd'azote)
temps(h)
NL23
JAM1∆narG1
co-cultureNL23/JAM1
co-cultureNL23/JAM1∆narG1
co-cultureNL23/JAM1∆narG2
co-cultureNL23/JAM1∆narG1narG2
JAM1∆narG2
JAM1∆narG1narG2
JAM1

62
Lepremieraspectintéressantdesrésultatsestlecomportementdela
dénitrificationdessouchesmutantes.Ledoublemutant(JAM1ΔnarG1narG2)était
évidemmentlemoinsperformantenréductiondunitrate,puisquecelui-cine
possèdepasdenitrateréductasesfonctionnelles(Fig.4.23).Lalégèrebaissede
concentrationdenitrateetd’azotetotalepourraitêtredueàdesfluctuations
causéesparlechromatographe.LemutantJAM1ΔnarG1avaitunprofilderéduction
denitratetrèssimilaireàlasoucheNL23,tandisquelemutantJAM1ΔnarG2aun
profilderéductiondenitratequiressembleplutôtàceluidelasoucheJAM1.Ces
observationsétaientprévisibles;desexpériencesantérieuresavaientdéjà
déterminéquelegènenarG1estleplusactifdelapaire(Mauffreyetal.,2015).
Ilestintéressantdeconstaterquetouteslesco-culturesontréduitlenitrate
plusrapidementquelasoucheNL23enculturespures(Fig.4.23).Pourlesco-
culturesNL23/JAM1,cerésultatn’étaitpasnouveau,puisquecerésultatavaitdéjà
étéobtenu.ÉtantdonnéquelasoucheJAM1ΔnarG2performeexactementcomme
saversionnonmutante,lerésultatpourlesco-culturesNL23/JAM1ΔnarG2n’était
pasétonnantnonplus.Lesco-culturesNL23/JAM1ΔnarG1étaientplusrapidesdans
laréductiondunitratequelemêmemutantenculturespures.Cetauxderéduction
plusélevépourraitêtreattribuéàl’additiondel’activitédeNapAetdeNar2.
Toutefois,cecinepourraitpasexpliquerlemêmecomportementchezlesco-
culturesNL23/JAM1ΔnarG1narG2.Eneffet,ilsemblequelasimpleprésencedela
soucheJAM1double-mutanteaitpermisàlasoucheNL23d’augmentersonactivité
deréductiondenitrate.Ilexisteraitdoncunfacteurd’aideinconnufourniparla
soucheJAM1.Ilestimpossibleparcetteexpériencededéterminercequ’estla
naturedecefacteur.Celui-cipourraitêtrereliéàl’aidedecroissanceobservéeen
conditionsaérobiesdansuneexpérienceprécédente(voirsection4),uneprotection
osmotique,ouunautreélémentjusqu’iciinconnu.
Danslegraphedeconcentrationsdenitrite(Fig.4.24),onvoitquelesseules
co-culturesàavoiraccumulédunitriteétaientlesco-culturesNL23/JAM1et
NL23/JAM1ΔnarG2.Cecirenforcel’idéequelemutantJAM1ΔnarG2secomporte

63
largementcommelasoucheJAM1non-mutante.Aucunesdesautresco-cultures
n’ontaccumulédenitrite.
Commed’habitudelesplusgrandesdécouvertessetrouvaientdanslegraphe
dusuividelaréductiond’azotetotal(Fig.4.25).Premièrement,lesrésultats
corroborentlanotionquelesco-culturesNL23/JAM1nesontpasbeaucoupplus
efficacesàcetégardquelasoucheNL23enculturespures.Deuxièmement,toutes
lesco-culturesavecdesmutantsontréduitl’azotetotalplusrapidementquela
soucheNL23enculturespuresetlesco-culturesNL23/JAM1.Ceciétaitunrésultat
trèssurprenant,surtoutdanslecasdesco-culturesNL23/JAM1ΔnarG1narG2,qui
necomptaientabsolumentpassurlacapacitéderéductiondenitratedelasouche
JAM1.Pourtant,celles-ciontatteint0mMd’azotetotalplusde20heuresavantles
co-culturessansmutantoulasoucheNL23enculturespures.
UnedernièreobservationnotableestquelasoucheJAM1etlemutant
JAM1ΔnarG2ontsembléréduirel’azotetotal.Commeilyapeud’ammoniumdans
lemilieuInstantOcean,l’assimilationdel’azotedoitsefairevialaréductiondu
nitrateennitriteparlavoieassimilatriceprésentedanslasoucheJAM1(Mauffreyet
al.,2016).

64
5. Discussion
5.1. Paramètresoptimauxpourlacroissancedesco-culturesdes
souchesNL23etJAM1
Avantledébutduprojet,unepartiedelarecherchepréliminairesurles
conditionsdesco-culturesdessouchesNL23etJAM1avaitdéjàétéfaite.Lemilieu
InstantOcean15/Lavaitdéjàétédécouvertcommeétantapteàsupporterla
croissancedesdeuxbactériesenculturespuresouenco-cultures.Deplus,la
plupartdesautresparamètresimportants,dontlatempératured’incubation,lepH
dumilieuetlesélémentstracesajoutésétaientdéjàétabliscommeétantlesmêmes
quepourlaculturedubiofilmoriginal.Laproblématiquerestanteétaitsurtoutaxée
surlarecherched’unesolutionauproblèmed’accumulationtoxiquedenitrite.
Lapremièresolutiontentéeétaitd’ajusterlesratiosdebactériesdanslesco-
culturesaumomentdel’ensemencement.Cettesolutionétaitlogiquemaiselleavait
aussiétéutiliséeavecsuccèsensituationdeco-culturesdeClostridiumbutyricumet
Rhodobactersphaeroides.Eneffet,dépendantduratiodesdeuxbactériesdans
l’inoculum,ilpouvaityavoiraméliorationdel’efficacitédelaphotoproduction
d’hydrogènegazeuxparlesco-cultures.Enlimitantl’abondancedeC.butyricum,qui
démontreunecroissancenettementplusrapidequesonhomologue,unéquilibre
bénéfiqueaétéatteint.(Laurinavicheneetal.,2016).Ainsi,pluslaproportiondela
soucheJAM1dansl’inoculumdedépartétaitimportante,plusletauxderéduction
denitrateétaitrapide,étantdonnéquelasoucheJAM1estplusefficacepourréduire
lenitratequelasoucheNL23.Toutefois,enréduisantlaquantitérelativeinitialede
lasoucheJAM1parrapportàlasoucheNL23,ilaétépossibledanscertainscas
d’éviterleniveaufataldenitrite.Lesrésultatsdanscesessaisétaientsouvent
difficilesàreproduireetàanalyserparlasuiteàcausedevariabilitésconsidérables
etimprévisibles.Laplupartdesexpériencescomportaientdestriplicatsdans
lesquelslesculturesavaientuncomportementcomplètementdifférentmalgrédes

65
conditionsdecultureidentiques.Lacausedecettevariabilitédemeureinconnue,
maispourraisêtreattribuéàunparamètretropdifficileàcontrôler,cequiest
parfoislecasdansdessystèmesbiologiques.Desrésultatspréliminairesont
suggéréqueleratioentrelessouchesNL23etJAM1doittendreversuncertain
équilibrepoureffectuerunedénitrificationefficace.Évidemment,lacapacitéde
reproduiredesrésultatsavecconstanceestdésirable.Iladoncétéconcluque
l’ajustementduratioinitialdesbactériesdansl’inoculumnepouvaitêtrequ’une
partiedelasolutionauproblème.
Ladeuxièmesolutionauproblèmed’accumulationdenitriteétaitde
simplementbaisseràuntierslaconcentrationdenitrateinitialedanslemilieude
croissance.Lebutétaitdelimiterlaquantitémaximalepossibledenitriteaccumulé.
Deplus,lesconcentrationsdenitrateobservéesdanslanatureoudansles
aquariumsd’oùprovientlaproblématiquen’approchentjamais40mMd’azotetel
quedanscesexpériences,etleseffetstoxiquessurlesanimauxsontressentisàdes
concentrationsnettementinférieures(DeRoosetal.,2003,Labbé,2007).L’impact
decetteactionaétédécisif.Avecunequantitémoindredenitrateàsadisposition,
lesco-culturesn’ontjamaispuaccumulerassezdenitritepourquelaconcentration
deviennenocive.Touteslesexpériencessubséquentesavaientdoncunratioinitial
de1:5(JAM1:NL23)etenviron15mMdenitrateaulieude40mM,ainsi
combinantlesrésultatsdesdeuxsolutions.

66
5.2. Profildeconsommationdenitrateetdenitritelorsdela
dénitrification
Unegrandepartiedesexpériencesvisantàoptimiserlesconditionsde
culturesaaussipermislacaractérisationduprofildeconsommationdenitrateetde
nitritedesco-culturesdessouchesNL23etJAM1(Fig.5.1à5.3).Lescourbes
généréesdurantl’expériencederéductiondenitratereprésententtrèsclairementle
comportementtypiqued’uneco-cultureréussie(enconditionsanaérobies)des
souchesJAM1etNL23:unebaissedeconcentrationrapidedunitrateavecune
accumulationdenitriteassociée,suivied’uneréductiondecenitrite(Fig5.3).
Figure5.1:ProfildeconsommationdunitrateetdunitriteparlasoucheNL23lorsdeladénitrification ExempleducomportementdelasoucheNL23enconditionsanaérobies.Legrapheaétégénéréàpartirdevraiesdonnéesmaisn’estqu’unereprésentationgénéraleduprofildelasoucheNL23lorsdeladénitrification.
0
2
4
6
8
10
12
0 20 40 60 80 100 120 140
concentrationdenitrateou
nitrite(mMd'azote)
temps(h)
NL23nitrate
NL23nitrite
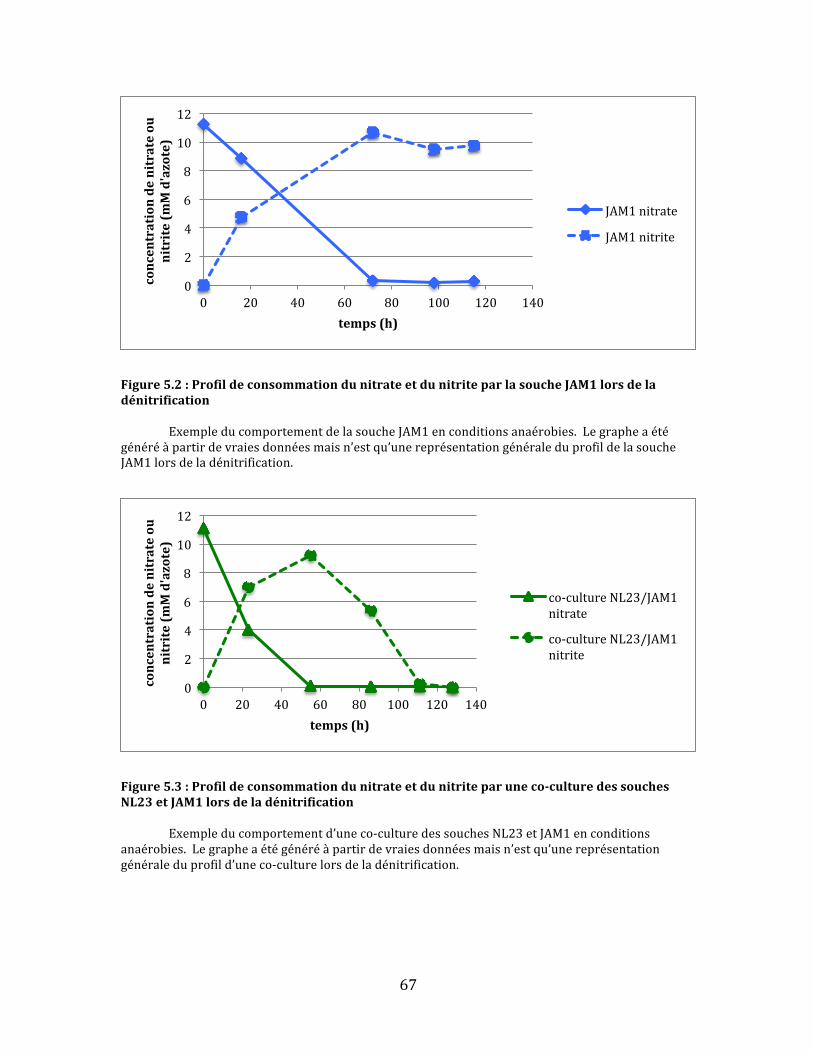
67
Figure5.2:ProfildeconsommationdunitrateetdunitriteparlasoucheJAM1lorsdeladénitrification ExempleducomportementdelasoucheJAM1enconditionsanaérobies.Legrapheaétégénéréàpartirdevraiesdonnéesmaisn’estqu’unereprésentationgénéraleduprofildelasoucheJAM1lorsdeladénitrification.
Figure5.3:Profildeconsommationdunitrateetdunitriteparuneco-culturedessouchesNL23etJAM1lorsdeladénitrification Exempleducomportementd’uneco-culturedessouchesNL23etJAM1enconditionsanaérobies.Legrapheaétégénéréàpartirdevraiesdonnéesmaisn’estqu’unereprésentationgénéraleduprofild’uneco-culturelorsdeladénitrification.
0
2
4
6
8
10
12
0 20 40 60 80 100 120 140
concentrationdenitrateou
nitrite(mMd'azote)
temps(h)
JAM1nitrate
JAM1nitrite
0
2
4
6
8
10
12
0 20 40 60 80 100 120 140
concentrationdenitrateou
nitrite(mMd'azote)
temps(h)
co-cultureNL23/JAM1nitrate
co-cultureNL23/JAM1nitrite
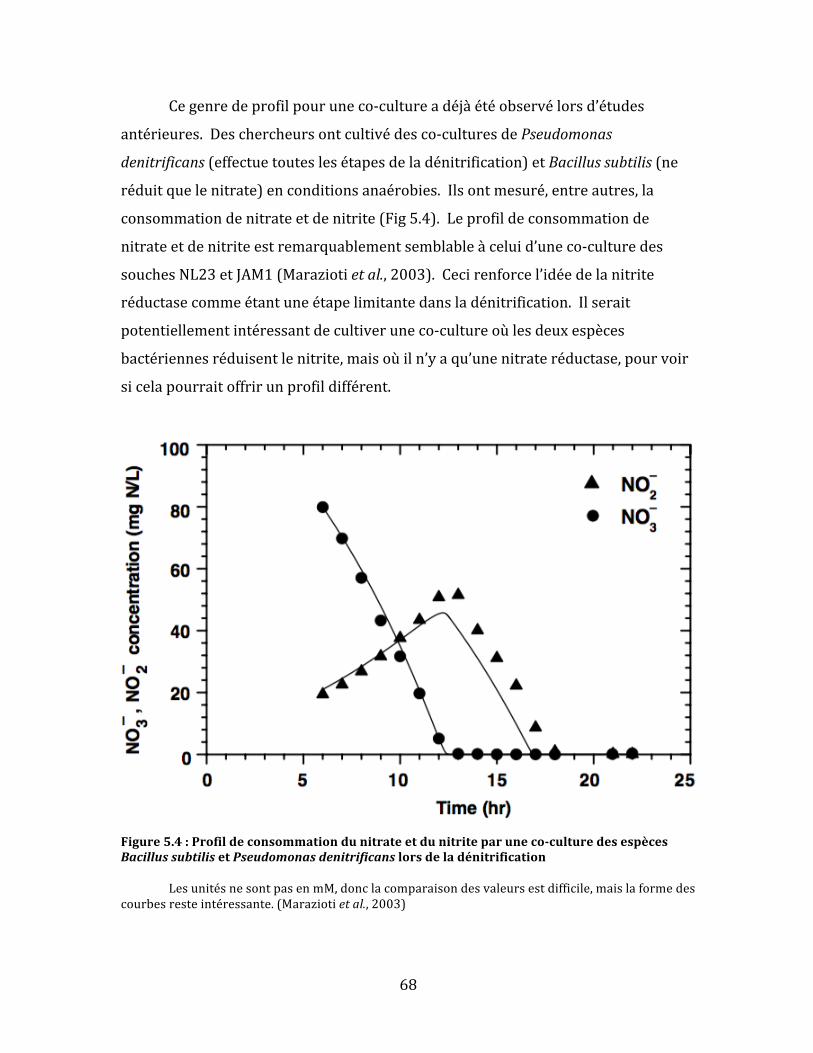
68
Cegenredeprofilpouruneco-cultureadéjàétéobservélorsd’études
antérieures.Deschercheursontcultivédesco-culturesdePseudomonas
denitrificans(effectuetouteslesétapesdeladénitrification)etBacillussubtilis(ne
réduitquelenitrate)enconditionsanaérobies.Ilsontmesuré,entreautres,la
consommationdenitrateetdenitrite(Fig5.4).Leprofildeconsommationde
nitrateetdenitriteestremarquablementsemblableàceluid’uneco-culturedes
souchesNL23etJAM1(Maraziotietal.,2003).Cecirenforcel’idéedelanitrite
réductasecommeétantuneétapelimitantedansladénitrification.Ilserait
potentiellementintéressantdecultiveruneco-cultureoùlesdeuxespèces
bactériennesréduisentlenitrite,maisoùiln’yaqu’unenitrateréductase,pourvoir
sicelapourraitoffrirunprofildifférent.
Figure5.4:Profildeconsommationdunitrateetdunitriteparuneco-culturedesespècesBacillussubtilisetPseudomonasdenitrificanslorsdeladénitrification LesunitésnesontpasenmM,donclacomparaisondesvaleursestdifficile,maislaformedescourbesresteintéressante.(Maraziotietal.,2003)

69
Lorsdel’expérienceaveclesmutantsdelasoucheJAM1,ilyavaitaussi
l’occasiondecorroborerdesnotionsreliéesauxdeuxnitrateréductases(Nar1et
Nar2)delasoucheJAM1.IlaétéconfirméquelanitrateréductaseNar1estla
principaleresponsabledelaréductiondunitrate.Eneffet,unesouchemutantede
lasoucheJAM1oùladeuxièmenitrateréductase(Nar2)aétédésactivée(doncNar1
actifseulement)reproduitpresqueexactementlemêmetauxderéductiondu
nitratequelasouchesauvage.DanslecasinverseoùNar1estinactivé,lesrésultats
ontdémontréquelasouchemutanteavecNar2actifseulementaunedynamiquede
réductiondunitrateéquivalenteàlasoucheNL23quipossèdeunenitrateréductase
detypeNap.
L’aspectleplusimportantengénéraldecetteoptiqueétaitl’efficacitéd’une
co-culturepourréduirel’azotetotal,c’estàdirelavitesseàlaquellelenitrateetle
nitritesontréduitsàzéro.Unedeshypothèsescentralesduprojetétaitquelesdeux
espècessontplusefficacesensemblesqu’individuellement.Incidemment,
l’améliorationdurendementmétaboliqueparl’applicationdeco-culturesestlebut
deplusieurseffortsderecherche(ex:laproductiond’hydrogèneparuneco-culture
deC.butyricumetR.sphaeroides,(Laurinavicheneetal.,2016).Lespremiers
résultatsàcetégardn’étaientpasencourageants:Lesco-culturesnesemblaientpas
êtrenécessairementplusefficacespourréduirel’azotetotal,prenantàpeuprès
autantdetempsquequelasoucheNL23enculturepurepourleréduireàzéro.Les
seulesco-culturesquiontdémontréuneaméliorationdecetaspectétaientcelles
aveclasoucheNL23avecuneversionmutantedeJAM1quiavaitNar1désactivé.
CombinéaveclanotionquelasoucheJAM1fournituneaidedecroissanceàla
soucheNL23(expliquéplusbas),cecipourraitindiquerquelasoucheNL23est
négativementaffectéeparl’activitédénitrifiantedelasoucheJAM1,mais
positivementaffectéeparsaprésence.

70
5.3. LessouchesNL23etJAM1enbiofilm
Unedesgrandesquestionsaudébutduprojetétaitlerôledelastructuredu
biofilmdansl’interactiondesbactériesNL23etJAM1,surtoutdanslecasde
l’osmoprotectiondelasoucheNL23danslesmilieuxconcentrésenNaCl(plusque
1.5%).Ladécisionadoncétéprisedetenterlacréationd’unbiofilmcomposéde
chacunedessouchesNL23etJAM1ouen«co-biofilm».Lacréationetl’observation
d’unbiofilmconsistaientdéjàunprocessusconnudanslelaboratoire.Desétudes
précédentesportaientsouventsurlacompositionetlaperformancedubiofilm
originalrecueilliauBiodômedeMontréal.Lesméthodesdecréationdubiofilm
développéesdanscesétudesprécédentesonttoutdemêmeservid’exempledansla
méthodeemployéeici(unedescriptiondecetteméthodeestdisponibleenannexe).
Unbiofilmcommecelui-ciauraitpuservirdansplusieursexpériences
différentes,maisl’idéeinitialeétaitd’étudierunrôlepossibledelastructuredu
biofilmdanslaprotectiondelasoucheNL23enmilieusalé.Lebiofilmoriginal(au
biodômeetenlaboratoire)étaitcultivéenmilieuavec3%desel.LasoucheNL23
étantunedesespècesprincipalesdecesystème,elledevaitnécessairementrecevoir
uneaideextérieure.CetaidepourraitprovenirdelasoucheJAM1,parexemplepar
l’entremisedel’osmoprotecteurectoïne;ilaétédocumentépourd’autresco-
culturesquel’aided’unosmoprotecteurn’estefficacequ’enbiofilm.Dansunesérie
d’expériences,unesouchebactériennehalophiledugenreChromohalobacteraété
cultivéeenco-cultureavecuneespècehalotolérantedugenreDietza.L’espèce
halophilenedémontreaucunecroissanceenmilieuxcontenantmoinsde5%desel,
tandisquel’espècehalotolérantenedémontreaucunecroissanceenmilieux
contenantplusde12%desel.Enco-culturesplanctoniques(ensuspensionliquide),
iln’yavaitpasd’effetprotecteurd’uncôtéoudel’autre;seulesdescellulesde
l’espècehalotoléranteontétédétectéesdanslemilieusanssel,etseulesdescellules
del’espècehalophileontétédétectéesdanslemilieude15%desel.Quandles
espècesétaientcultivéesen«co-biofilm»,l’espècehalotoléranten’apportaitaucune

71
aideàl’autrebactérieenmilieusanssel.Cependant,unequantitéàpeuprèségale
desdeuxespècesontétédétectéedansle«co-biofilm»à15%desel.Les
chercheursontsuggérélerôledesosmoprotecteursectoïneetglutamatedansle
résultat(Plakunovetal.,2008).Malgrétout,ilneseraitpassurprenantquela
protectiondanslecasdessouchesNL23etJAM1provienneplutôtdirectementdela
structuredubiofilmetdesEPSquilecompose,étantdonnélagrandevariétéde
fonctionsquiluisontattribué.
L’expérienceplanifiéeétaitdecultiverlebiofilmenmilieuInstantOceanavec
1,5%deNaCl,puisdeletransférerdanslemêmemilieuà3%deNaCl,pourune
autresériedetransferts.Lasurvie(ounon)delasoucheNL23,encomparaison
avecuneco-cultureliquide,auraitalorsétéuneindicationduniveaudeprotection
offertparlebiofilm.Uneautreindicationdelaprotectionauraitétéletaux
dénitrification.SilacultureenbiofilmdanslemilieuInstantOcean(3%NaCl)était
arrivéeàréduirelenitrite,celaauraitétéunsigned’activitédelapartdelasouche
NL23.
Figure5.5:Supportenplastiquegarnid’unbiofilm Dessupportsenplastiquecommecelui-ciontétéutiliséspourfournirunesurfaced’accrochageausoucheNL23etJAM1pourlacréationd’unbiofilm.Lessupportssonassezpetitspourêtrefacilementinsérésetretirésdesfiolesdeculture.

72
Pourdesraisonsqu’onignore,nilessouchesindividuelles,nilaco-culture
dessouchesNL23etJAM1n’ontdémontrélacapacitédeformerunbiofilm.Les
raisonsexactesdecerésultatsontévidemmentinconnues,maisonpeutdévelopper
deshypothèses.L’échecpourraitêtredûàl’absenced’uneespècecolonisatrice
importante,puisquedescaractéristiquesspécifiquessontrequispourfaciliter
l’adhérencedecellulesàuneparoi.Lesfacteursassociésauxstagesinitiauxdela
formationd’unbiofilmpourE.coli,parexemple,incluentlamotilité(ex.:flagelles),
laprésencedefimbriae,laprésencedecurlietlaproductiondecertainsEPSclés,
dontlepolysaccaridePGA(β-1,6-N-acetyl-D-glucosamine)(VanHoudtetal.,2005).
Lacapacitédeformationdebiofilmpeutvarierénormémententreespècesetmême
entresouchesdelamêmeespèces,commeilaétédémontrépourCronobacter
sakazakii(Yeetal.,2015).Ilestaussidocumentéquelaformationdebiofilmpour
certainesespècesestfacilitéeparlacolonisationinitialed’unesurfaceparuneautre
espèce.L’espècebactérienneSphingomonasnatatoriaestcapabledeformerdes
biofilmsfragilesenculturepure,maiselleformedesbiofilmsplusrobustes
lorsqu’elleestexposéeàunesurfacerecouvertedebactériesdel’espèceMicrococcus
luteus.Ceciestdûàune«coagrégation»,unereconnaissanceetunattachement
spécifiqueentrelesdeuxbactéries(Minetal.,2009).
Lebiofilmoriginalestunecommunautécomplexedebactériesetd’autres
organismes;ilestpossiblequ’unouplusieursdeceux-ciagissentcomme
spécialistescolonisateursdesurfacesetsontdoncnécessairesàlaformation
efficaced’unbiofilm.Dansunedesanalysesdubiofilmoriginal,ilavaitéténoté
qu’unegrandevariétédecelluleseucaryotescomposaientuneproportion
importante(20%)delabiomasseaudébutdudéveloppementdubiofilm;ces
cellulesdevenaientmoinsabondantesparlasuite(5-6%)(Labbéetal.,2007).Ceci
pourraitêtreindicatifd’unrôledecolonisationounon,maislechangement
dramatiquedelacompositiondelabiomassedubiofilmestcertainementune
indicationquecertainesbactériessontresponsablesdedifférentsélémentsdelavie
microbienneenbiofilm,incluantlacolonisation.Ilestentièrementpossiblequeles
souchesNL23etJAM1nesoientpasaxéessurcettepartiedudéveloppementd’un

73
biofilm.Ilestaussipossiblequelesconditionsdanslesquellesl’expériences’est
dérouléen’étaientpasoptimales.Lasurfaceenplastiquedessupportsn’étaitpeut-
êtrepasapteaubiofilm;dessurfacesenverreontététestéeségalement,sansplus
derésultat.
Éventuellement,cetaspectduprojetaétéabandonnéavecleverdictquela
formationd’unbiofilmcomposédessouchesNL23etJAM1n’estpaspossibleavec
lesméthodesutilisées.
5.4. Échangederessources
Lesexpériencesontdémontréqu’ilyavaitaumoinsexistenced’uneaide
bénéfiquedecroissanceetd’activitédénitrifiantedanslesco-culturesdelapartde
lasoucheJAM1.Eneffet,ilsemblequ’ilexisteunfacteurinconnuquiaidelasouche
NL23àcroîtreplusrapidement(démontréenconditionsaérobies)etàréduirele
nitrateplusrapidement(démontréenconditionsanaérobies).L’identitédece
mécanismeresteindéterminée;Ilpourraits’agird’uneparticuleajoutéeou
transférée,oualorsl’enlèvementd’unélémentlimitant.Deplus,ilseraitpossible
queplusieursfacteurssoientprésents,etmêmequedifférentsfacteurssoient
responsablesdesaméliorationsobservéesenaérobioseetenanaérobiose.L’aspect
unidirectionneldecetaideaaussiétédémontré;lasoucheJAM1n’ayantjamais
montrélamoindreindicationdeprofiterdelaprésencedelasoucheNL23,voire
mêmesaprésencepouvantêtrenocive.Dansl’étudemenéeparTakakietal.(2007)
(voirsection2.8),l’espèceStreptomycesgriseusfournissaituneaidede
dénitrificationàRalstoniapickettiiparl’entremisedeprotéasesrelâchéesdans
l’environnement.Pourarriveràcetteconclusion,leschercheursontpremièrement
cultivéR.pickettiienprésencedesurnageantdeS.griseusenconditionsanaérobies.
Cetteastuceaétéutiliséeiciégalement,maisseulementenconditionsaérobies.
Pourconfirmerquec’estbienunemoléculesecrétéeoul’absenced’unélément
limitantquiestresponsabledel’améliorationdeladénitrificationdelasouche
NL23,ilfaudraitrépéterl’expérienceenconditionsanaérobies.Deuxièmement,le

74
surnageantdeS.griseusaétésoumisàuneultracentrifugationpourcréerune
séparationdescomposantesparmassemolaire.Entestantl’effetdechaquefraction
demoléculesséparément,ilestdevenupossibled’identifierlesplusmassives
commeétantresponsablesdutauxsupérieurdedénitrification,cequiamenéà
l’hypothèsedesprotéases.Ceciestunestratégiequipourraitêtreintéressantepour
identifierlanaturedufacteurd’aidedelasoucheJAM1.Deplus,cettemêmeétudea
concluquel’histidinejouaitpeut-êtreunrôledansl’améliorationdel’activité
dénitrifiante.CecipourraitêtrerapidementtestéaveclasoucheNL23également.
Uncandidatd’unfacteuraidantlasoucheNL23estl’ectoïne.Ilétait
connuquelasoucheJAM1possèdelesgènesnécessairesàlaproductiond’ectoïne,
uneparticuleosmoprotectrice.Ilaétérécemmentdéterminédanslelaboratoire
quelasoucheJAM1produisaitdel’ectoïnedanslemilieudeculture(donnéesnon-
publiées).Lorsdeceprojet,leseffortspourdémontrerl’effetd’osmoprotectionde
lasoucheJAM1ontéchoué(voirsection4.3.2),maiscelaneprouvepasquel’ectoïne
nejouepasderôle.Dessolutionspossiblesseraientdetrouverunmoyende
quantifierl’ectoïnedirectementoudeconcentrerl’ectoïneproduitparlasouche
JAM1pourinduiresoneffet.Ilseraitaussipossiblequel’ectoïnen’estpas
responsabledeseffetsobservésdanslessituationsplanctoniques.Dansuneétudeà
cesujet,ilaétédéterminéquel’ectoïneétaitsimplementtropdiluéedansune
cultureensuspensionliquidepouravoiruneffetnotable(Plakunovetal.,2008).
Danscecas,l’effetdel’ectoïnenepourraitêtreobservéquesiuneméthodeétait
trouvéepourproduireunbiofilmaveclessoucheNL23etJAM1.

75
6. Conclusion
Desconditionsoptimalesdeco-culturesdessouchesNL23etJAM1ontété
développées,cequiapermisd’optimiserlesétudessuivantes.Laproblématique
d’accumulationdenitriteparlaréductiontroprapidedunitrateaétécontournée
parlabaissedenitrateinitialedanslemilieuetl’ajustementduratiodessouchesau
momentdel’ensemencement.Lesmêmesexpériencesontaussidévoiléplusieurs
aspectsducomportementdesdeuxsouchesenconditionsaérobiesetanaérobies,
oùcelles-ciperformentladénitrification.
Ilaétédémontréquelesco-culturesdessouchesNL23etJAM1n’étaientpas
nécessairementplusrapidesendénitrificationquelasoucheNL23encultures
pures.Toutefois,ilaégalementétédémontréquelesco-culturesdelasoucheNL23
aveclessouchesmutantesdeJAM1quiontlanitrateréductaseNar2désactivée
étaientcapablesd’unedénitrificationplusrapidequelasoucheNL23encultures
pures.
Lestentativesdeformationsd’unbiofilmuniquementcomposédessouches
NL23etJAM1ontéchoué.Ilsemblequedanslesconditionsimposées,cesdeux
espècesbactériennesnepeuventpasdévelopperdebiofilm.
IlaétédémontréquelasoucheJAM1apporteuneaidedecroissanceàla
soucheNL23enconditionsaérobiesetuneaidederéductiondenitrateen
conditionsanaérobies.LasoucheNL23n’apasfaitdemêmepourlasoucheJAM1
danslesconditionsd’expérience.Cependant,iln’apuêtredémontréquelleestla
naturedecetaide.

76
Annexe
Protocoledecréationd’unbiofilm
Certainesexpériencesontrequislaformationd’unbiofilm.Pourcefaire,des
supportsenplastiquesontétéinsérésdanslesfiolesdecultureenprésencede
bactériesensuspension.Cesensemencementssontensuiteincubésenconditions
anaérobies.Puisqueledéveloppementd’unbiofilmapuprendreplusieurs
semaines,lemilieudanslequelsetrouvaitlacultureadûêtrerégulièrementchangé
pouryéviterladéplétiondesressources.Lessupportsontdoncététransférés
régulièrementdansunenouvellefiolecontenantdumilieufrais.Pourlesco-
culturesdessouchesJAM1etNL23,lapériodedetempsentrelestransfertsétait
d’environunesemaine.
Pourtransférerlessupportsenévitantlacontaminationexterneetentreles
échantillons,touslesoutilsutilisésontdûêtrestérilesetletravailadûêtrefaitdans
lahottebiologique.Lessupportsontétéenlevésdesfiolesàl’aidedepincespuis
insérésdanslesnouvellesfiolespréalablementautoclavées.Celles-ciontensuiteété
ferméeshermétiquementetl’oxygèneàl’intérieuraétéenlevépourcréer
l’environnementd’anaérobiose.Danslecasdeceprojet,leprocessusaétérépété
jusqu’àcequ’ilestdevenuévidentqu’iln’yauraitpasformationdebiofilm(4
semainesouplus).
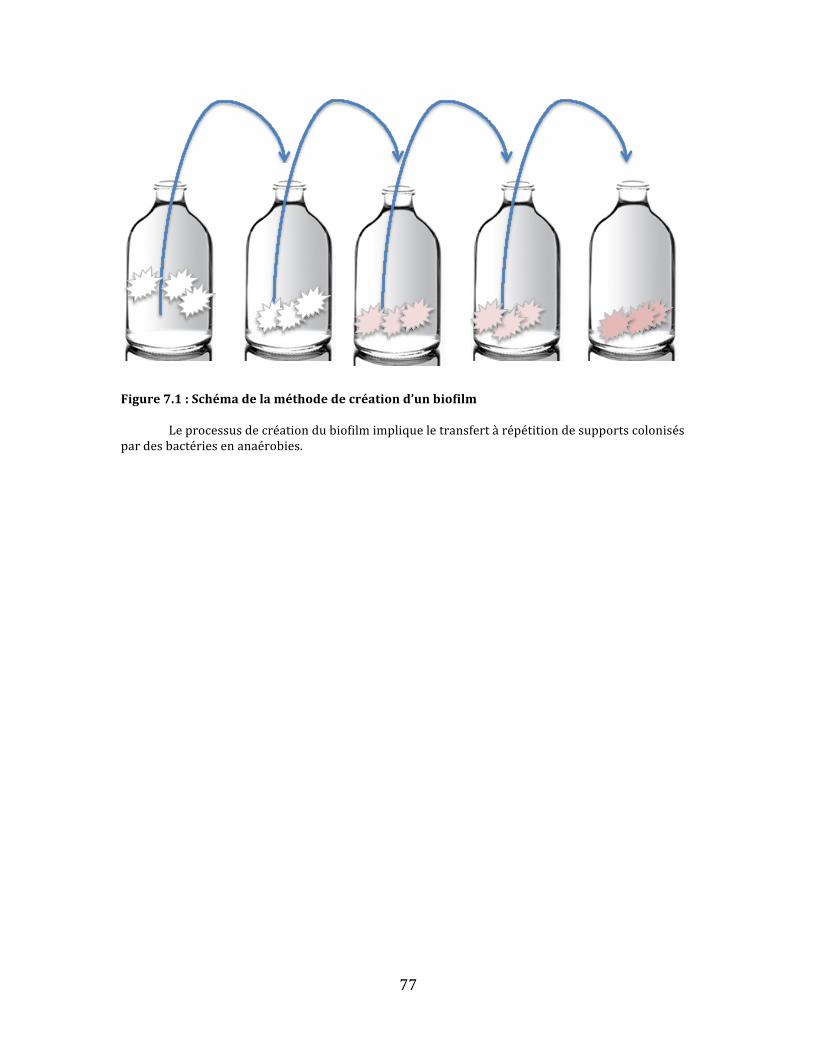
77
Figure7.1:Schémadelaméthodedecréationd’unbiofilm Leprocessusdecréationdubiofilmimpliqueletransfertàrépétitiondesupportscoloniséspardesbactériesenanaérobies.

78
Listedesréférences
AndersonIC&LevineJS(1986)RelativeRatesofNitricOxideandNitrousOxideProductionbyNitrifiers,Denitrifiers,andNitrateRespirers.Appliedandenvironmentalmicrobiology51(5):938-945.
Doi:AuclairJ,LepineF,ParentS&VillemurR(2010)Dissimilatoryreductionofnitrate
inseawaterbyaMethylophagastraincontainingtwohighlydivergentnarGsequences.TheISMEjournal4(10):1302-1313.
Doi:10.1038/ismej.2010.47BerksBC,FergusonSJ,MoirJW&RichardsonDJ(1995)Enzymesandassociated
electrontransportsystemsthatcatalysetherespiratoryreductionofnitrogenoxidesandoxyanions.Biochimicaetbiophysicaacta1232(3):97-173.
Doi:BlaisM-A(2014)SuividelacontaminationfécaledanslagranderégiondeMontréal.
(UniversitéduQuébec,InstitutNationaldelaRechercheScientifique).CamargoJA,AlonsoA&SalamancaA(2005)Nitratetoxicitytoaquaticanimals:a
reviewwithnewdataforfreshwaterinvertebrates.Chemosphere58(9):1255-1267.
Doi:10.1016/j.chemosphere.2004.10.044DeRoosAJ,WardMH,LynchCF&CantorKP(2003)Nitrateinpublicwatersupplies
andtheriskofcolonandrectumcancers.Epidemiology14(6):640-649. Doi:DoroninaN,LeeTD,IvanovaE&TrotsenkoYA(2005)Methylophagamuratasp.
nov.:ahaloalkaliphilicaerobicmethylotrophfromdeterioratingmarble.Microbiology74(4):440-447.
Doi:FesefeldtA,KloosK,BotheH,LemmerH&GliescheC(1998)Distributionof
denitrificationandnitrogenfixationgenesinHyphomicrobiumspp.andotherbuddingbacteria.Canadianjournalofmicrobiology44(2):181-186.
Doi:FlemmingHC&WingenderJ(2010)Thebiofilmmatrix.Naturereviews.
Microbiology8(9):623-633. Doi:10.1038/nrmicro2415GliescheC,FesefeldtA&HirschP(2005)HyphomicrobiumStutzerandHartleb
1898,76AL.Bergey’sManual®ofSystematicBacteriology:476-494. Doi:GregoryLG,BondPL,RichardsonDJ&SpiroS(2003)Characterizationofanitrate-
respiringbacterialcommunityusingthenitratereductasegene(narG)asafunctionalmarker.Microbiology149(1):229-237.
Doi:

79
HajipourA,MoghadamN,NosratiM&ShojasadatiS(2011)Aerobicthermophilictreatmentoflandfillleachateinamoving-bedbiofilmbioreactor.JournalofEnvironmentalHealthScience&Engineering8(1):3-14.
Doi:HayatsuM,TagoK&SaitoM(2008)Variousplayersinthenitrogencycle:Diversity
andfunctionsofthemicroorganismsinvolvedinnitrificationanddenitrification.SoilScienceandPlantNutrition54(1):33-45.
Doi:10.1111/j.1747-0765.2007.00195.xHinoT,NaganoS,SugimotoH,ToshaT&ShiroY(2012)Molecularstructureand
functionofbacterialnitricoxidereductase.Biochimicaetbiophysicaacta1817(4):680-687.
Doi:10.1016/j.bbabio.2011.09.021JanvierM&GrimontP(1995)ThegenusMethylophaga,anewlineofdescentwithin
phylogeneticbranchγofProteobacteria.Researchinmicrobiology146(7):543-550.
Doi:JebbarM,TalibartR,GlouxK,BernardT&BlancoC(1992)Osmoprotectionof
Escherichiacolibyectoine:uptakeandaccumulationcharacteristics.JBacteriol174(15):5027-5035.
Doi:KempfB&BremerE(1998)Uptakeandsynthesisofcompatiblesolutesas
microbialstressresponsestohigh-osmolalityenvironments.Archivesofmicrobiology170(5):319-330.
Doi:LabbéN(2007)Caractérisationdudynamismemicrobiendudénitrificateurmarindu
BiodômedeMontréal.(UniversitéduQuébec,InstitutNationaldelaRechercheScientifique).
LabbéN,JuteauP,ParentS&VillemurR(2003)Bacterialdiversityinamarinemethanol-feddenitrificationreactoratthemontrealbiodome,Canada.Microbialecology46(1):12-21.
Doi:10.1007/s00248-002-1056-6LabbéN,LaurinV,JuteauP,ParentS&VillemurR(2007)Microbiological
communitystructureofthebiofilmofamethanol-fed,marinedenitrificationsystem,andidentificationofthemethanol-utilizingmicroorganisms.Microbialecology53(4):621-630.
Doi:10.1007/s00248-006-9168-zLabbéN,ParentS&VillemurR(2004)Nitratireductoraquibiodomusgen.nov.,sp.
nov.,anovelα-proteobacteriumfromthemarinedenitrificationsystemoftheMontrealBiodome(Canada).Internationaljournalofsystematicandevolutionarymicrobiology54(1):269-273.
Doi:LabelleMA,JuteauP,JolicoeurM,VillemurR,ParentS&ComeauY(2005)Seawater
denitrificationinaclosedmesocosmbyasubmergedmovingbedbiofilmreactor.Waterresearch39(14):3409-3417.
Doi:10.1016/j.watres.2005.06.001

80
LaurinavicheneT&TsygankovA(2016)DifferenttypesofH2photoproductionbystarch-utilizingco-culturesofClostridiumbutyricumandRhodobactersphaeroides.InternationalJournalofHydrogenEnergy41(31):13419-13425.
Doi:Le-ClechP,ChenV&FaneTA(2006)Foulinginmembranebioreactorsusedin
wastewatertreatment.Journalofmembranescience284(1):17-53. Doi:LiangX,LinL,YeY,GuJ,WangZ,XuL,JinY,RuQ&TianG(2015)Nutrientremoval
efficiencyinarice-strawdenitrifyingbioreactor.Bioresourcetechnology198:746-754.
Doi:http://dx.doi.org/10.1016/j.biortech.2015.09.083MaraziotiC,KornarosM&LyberatosG(2003)Kineticmodelingofamixedculture
ofPseudomonasdenitrificansandBacillussubtilisunderaerobicandanoxicoperatingconditions.Waterresearch37(6):1239-1251.
Doi:MartineauC,MauffreyF&VillemurR(2015)Comparativeanalysisofdenitrifying
activitiesofHyphomicrobiumnitrativorans,Hyphomicrobiumdenitrificans,andHyphomicrobiumzavarzinii.Appliedandenvironmentalmicrobiology81(15):5003-5014.
Doi:MartineauC,VilleneuveC,MauffreyF&VillemurR(2013)Hyphomicrobium
nitrativoranssp.nov.,isolatedfromthebiofilmofamethanol-feddenitrificationsystemtreatingseawaterattheMontrealBiodome.Internationaljournalofsystematicandevolutionarymicrobiology63(Pt10):3777-3781.
Doi:10.1099/ijs.0.048124-0MauffreyF,MartineauC&VillemurR(2015)ImportanceoftheTwoDissimilatory
(Nar)NitrateReductasesintheGrowthandNitrateReductionoftheMethylotrophicMarineBacteriumMethylophaganitratireducenticrescensJAM1.Frontiersinmicrobiology6.
Doi:MinK&RickardA(2009)Coaggregationbythefreshwaterbacterium
Sphingomonasnatatoriaaltersdual-speciesbiofilmformation.Appliedandenvironmentalmicrobiology75(12):3987-3997.
Doi:MincerTJ&AicherAC(2016)Methanolproductionbyabroadphylogeneticarrayof
marinephytoplankton.PloSone11(3):e0150820. Doi:MustakhimovI,ReshetnikovA,FedorovD,KhmeleninaV&TrotsenkoYA(2012)
RoleofEctRastranscriptionalregulatorofectoinebiosynthesisgenesinMethylophagathalassica.Biochemistry(Moscow)77(8):857-863.
Doi:MustakhimovI,ReshetnikovA,KhmeleninaV&TrotsenkoYA(2009)EctR—anovel
transcriptionalregulatorofectoinebiosynthesisgenesinthehaloalcaliphilicmethylotrophicbacteriumMethylophagaalcalica.DokladyBiochemistryandBiophysics.Springer,p305-308.

81
PakulaR&FreemanA(1996)Anewcontinuousbiofilmbioreactorforimmobilizedoil-degradingfilamentousfungi.Biotechnologyandbioengineering49(1):20-25.
Doi:PauletaSR,Dell’AcquaS&MouraI(2013)Nitrousoxidereductase.Coordination
ChemistryReviews257(2):332-349. Doi:http://dx.doi.org/10.1016/j.ccr.2012.05.026PlakunovVK,ZhurinaMV&BeliaevSS(2008)[Resistanceofthepetroleum-
oxidizingmicroorganismDietziasp.tohyperosmoticshockinreconstitutedbiofilms].Mikrobiologiia77(5):581-589.
Doi:PriemeA,BrakerG&TiedjeJM(2002)Diversityofnitritereductase(nirKandnirS)
genefragmentsinforesteduplandandwetlandsoils.Appliedandenvironmentalmicrobiology68(4):1893-1900.
Doi:RoedigerM(1983)Activatedsludgesystem.(GooglePatents).SauerT&GalinskiEA(1998)Bacterialmilking:Anovelbioprocessforproductionof
compatiblesolutes.Biotechnologyandbioengineering59(1):128. Doi:TakakiK,FushinobuS,KimS-W,MiyaharaM,WakagiT&ShounH(2008)
StreptomycesgriseusenhancesdenitrificationbyRalstoniapickettiiK50,whichispossiblymediatedbyhistidineproducedduringco-culture.Bioscience,biotechnology,andbiochemistry72(1):163-170.
Doi:VandePas-SchoonenK,Schalk-OtteS,HaaijerS,SchmidM,DenCampHO,StrousM,
KuenenJG&JettenM(2005)Completeconversionofnitrateintodinitrogengasinco-culturesofdenitrifyingbacteria.BiochemicalSocietytransactions33(1):205-209.
Doi:VanHoudtR&MichielsCW(2005)Roleofbacterialcellsurfacestructuresin
Escherichiacolibiofilmformation.Researchinmicrobiology156(5):626-633. Doi:VilleneuveC(2012)Caractérisationetclassificationdesnouvellesespèces
MethylophaganitratireducenticrescensetMethylophagafrappieri..(UniversitéduQuébec,InstitutNationaldelaRechercheScientifique).
VilleneuveC,MartineauC,MauffreyF&VillemurR(2013)Methylophaganitratireducenticrescenssp.nov.andMethylophagafrappierisp.nov.,isolatedfromthebiofilmofthemethanol-feddenitrificationsystemtreatingtheseawaterattheMontrealBiodome.Internationaljournalofsystematicandevolutionarymicrobiology63(Pt6):2216-2222.
Doi:10.1099/ijs.0.044545-0WangX,ChangVW&TangCY(2016)Osmoticmembranebioreactor(OMBR)
technologyforwastewatertreatmentandreclamation:Advances,challenges,andprospectsforthefuture.JournalofMembraneScience504:113-132.
Doi:

82
YeY,LingN,JiaoR,WuQ,HanY&GaoJ(2015)EffectsofcultureconditionsonthebiofilmformationofCronobactersakazakiistrainsanddistributionofgenesinvolvedinbiofilmformation.LWT-FoodScienceandTechnology62(1,Part1):1-6.
Doi:http://dx.doi.org/10.1016/j.lwt.2015.01.035ZaprasisA,BrillJ,ThüringM,WünscheG,HeunM,BarzantnyH,HoffmannT&
BremerE(2013)OsmoprotectionofBacillussubtilisthroughimportandproteolysisofproline-containingpeptides.Appliedandenvironmentalmicrobiology79(2):576-587.
Doi:ZumftWG(1997)Cellbiologyandmolecularbasisofdenitrification.Microbiology
andmolecularbiologyreviews:MMBR61(4):533-616. Doi: