relation of leaf area and position to quality of fruit …
TRANSCRIPT

TECHNICAL BULLETIN NO. 338 FEBBUARY, 1933
UNITED STATES DEPARTMENT OF AGRICULTURE WASHINGTON, D. C.
RELATION OF LEAF AREA AND POSITION TO QUALITY OF FRUIT AND TO BUD
DIFFERENTIATION IN APPLES" By M. H. HALLEB, Assistant Pomologistj and J. R. MAGNESS, Principal Pomolo
gisty Division of Horticultural Crops and Diseases, Bureau of Plant Industry
CONTENTS
Page Introduction ._-_ 1 Material and methods 2 Relation of leaf area to growth and composi-
tion of apples... 4 Increase in size of fruit 5 Chemical composition 10 Dessert; quality . — 12 Firmness 13 Color development 13
Relation of distance and direction of fruit from leaves to size and composition of apples 15
Relation of leaf distance to growth of fruit . Relation of direction of apples from
leaves to growth of fruit. 19
Page Relation of distance and direction of fruit from
leaves to size and composition of apples- Continued.
Relation between number of seeds per apple and size of fruit 20
Relation between leaf distance and chem- ical composition of fruit 20
Relation of leaf area per apple to bud differ- entiation 22
Review of previous work.. ._ 22 Results of present investigations 25
Discussion 29 Summary 32 Literature cited....— 33
INTRODUCTION
The average mature apple consists of about 85 per cent of water and 15 per cent of dry matter, the latter consistmg primarily of organic matter elaborated in the leaves. It is natural to suppose, therefore, that the growth and composition of the fruit would depend largely on the amount of water supplied to the fruit and on the elaborated foods available in the leaves. Obviously a liberal supply of moisture is essential for the proper development of the fruit. However, the rôle of elaborated foods supplied by the leaves is not so apparent. In this regard the relationstup of leaf area to growth and composition of the fruit appears to be of importance, particularly with reference to the carbohydrate supplied to the fruit. The purpose of the investi- gation reported herein was to obtain definite information as to the relation of the carbohydrate supply to the growth and composition of the fruit. Another phase of this investigation, closely associated with carbohydrate supply, was the effect of fruiting on bud differen- tiation.
The supply of carbohydrates available to the fruit depends pri- marily on two factors—the amount of carbohydrates elaborated per fruit, and the transport of the carbohydrates to the fruit.
1 Presented by the senior writer to the faculty of the graduate school of the University of Maryland in partial fulfillment of the requirements for the degree of doctor of philosophy.
146102—33 1

2 TECHNICAL BULLETIN 338, U. S. DEPT. OF AGRICULTXJEE
The amount of carbohydrates elaborated per fruit will depend first upon the leaf area per fruit. The leaf area per fruit may be influenced by certain orchard practices such as thinning, pruning, and fertilizing. Pruning usually increases the leaf area per fruit by reducing the num- ber of fruiting spurs and thus stimulating the growth and leaf devel- opment in those that remain, but it may reduce the leaf area per fruit by increasing the set of fruit per spur. Nitrogen fertilizers also increase the leaf area by increasing vegetative vigor and thus the size of the leaves, but on the other hand they may reduce the leaf area per fruit by increasing the set of fruit. Thinning the fruit is the most effective method of increasing leaf area per fruit.
With a given leaf area per fruit the amount of carbohydrates formed will depend on the efficiency of the leaves, which in turn may be influenced by such factors as temperature, moisture, amount of chlorophyll, mineral nutrients, the amount of carbon dioxide in the surrounding atmosphere, and the duration and intensity of simlight.
Factors that might influence the transport of carbohydrates to the fruit are (1) the diffusion gradient to the fruit and to other parts of the tree, (2) the distance and direction of the leaves from the fruit, and (3) the cross-sectional diameter of the conducting tissue at its narrowest point. The seed content of the fruit also seems to have some effect on the accumulation of elaborated foods in the fruit.
This bulletin deals with (1) the growth and composition of the fruit in relation to the carbohydrate supply as influenced by the leaf area per apple, (2) the transport of the carbohydrates to the fruit as influenced particiilarly by the distance and direction of the leaves from the fruit, and (3) the effect of fruiting on bud differentiation.
MATERIAL AND METHODS
The trees used in this investigation were located in the variety orchard at the Arlington Experiment Farm, Kosslyn, Va., near Wash- ington, D. C. The experiments usually were started in June or the latter part of May, when the apples were about one-half to 1 inch in diameter.
Treatments were applied to both ringed and unringed branches. In ringing the branches a strip of bark about one-quarter inch wide was removed, the cambium was scraped, and melted paraffin was applied to the wound. Scraping the cambium prevented regeneration of phloem across the ring; this treatment would also prevent the de- velopment of new xylem tissue at the ring. Apparently greater injury to the branches resulted when the ringing was done very early in the season than when it was done later, probably because of the reduction in the amount of new xylem tissue, which would probably be more active in the translocation of water to the branch. Similarly, the smaller the branch the greater was the injury. For this reason, after the first year fairly large branches, at least 2.5 cm in circumference, were used in order to reduce this injury as much as possible. Larger branches, at least 6 cm in circumference, were used for the unringed branches.
^ To study the effect of leaf area on the fruit and on bud differentia- tion, leaves or fruit w^re removed from both ringed and imringed branches until the desired number of leaves per fruit was obtained.

KELATION OF LEAF AKEA AND POSITION TO FRUIT QUALITY 6
To study the effect of the leaf position on the fruit, unringed branches from which all of the leaves were removed were used. On such branches the fruit was at varying distances from the leaves on the main limbs to which the branches were attached. These leaves would be the nearest source of elaborated food. The distances from the main limbs to the fruits were measured along the shortest possible paths through the branches. Kinged branches were also used, on winch 1 fruit and 20 leaves were left above the ring on each branch, the leaves being left at varying distances from the fruit. In some cases the leaves were left as close as possible to the fruit, as in Figure 1, A; in others they were left 50 to 100 cm or more from the fruit. In some cases the leaves were above the fruit, as in Figure 1, B, while in others they were below the fruit or on separate branches, as in Figure 1, C. In another experiment ringed branches were used on which 20 leaves and 2 fruits were left above the ring, one of the fruits being distant from the leaves, whereas the other was close, as shown in Figure 1, D.
FIGURE 1.—Treatment of ringed branches: A, Leaves adjacent to apple; B, leaves distant from and above apple; C, leaves distant on separate branch from apple; D, leaves distant from one apple and adjacent to another
Small or deformed leaves were stripped off, leaving only the medium-sized and large ones. The area of at least 100 leaves similar to those on the test branches was measured by means of a plani- meter, and the leaf area per.apple was approximated from these measurements. In most cases all treatments for each variety were applied to a single tree or to two adjacent trees.
The circumference of the fruit at the widest transverse diameter was determined at the beginning of the experiments and at intervals thereafter until harvest. The volume of the fruit, assumed to be a sphere, was computed from the circumference measurements, and the increase in volume was obtained by subtracting the volume of each apple at the beginning of the experiment from the volume of the same apple when mature.

4 TECHNICAL BULLETIN 338, XT. S. DEPT. OF AGBICÜLTUKE
The fruit was harvested in the fall and grouped according to the number of leaves with which the apples were grown or according to the distance they were from the leaves. After it had been stored for a period in order to allow the starch to be changed to sugar, samples were taken, and sugar and acid determinations were made according to the methods described in a previous publication {30)? Dry- weight déterminations were made in 1925 and 1926 by drying 25-g ^ samples of ground tissue to constant weight at 85° to 90° C. In 1927 direct determinations were made by distilling with toluol according to the method described by BidweU and Sterling {10).
In the spring following the year in which the thinning treatments were made, blossom and leaf buds were counted on the spurs of the branches that were used for the leaf-area studies. Spurs that fruited during the year of treatment were tagged in 1926 and 1927 but not in 1925. Spurs that had blossomed but from which the fruit had dropped or had been removed were determined by the fruit scars on the spurs, and those that were leaf spurs during the year of treatment were determined by the lack of scars.
RELATION OF LEAF AREA TO GROWTH AND COMPOSITION OF APPLES
A preliminary report {21) of part of the material presented in this bulletin showed that the size of the fruit increased with an increase in leaf area per apple up to a point beyond which further increase in leaf area had little or no effect. Dry weight and sugar concentration also increased as the leaf area increased. Under the conditions of the experiments and with the varieties studied at the Arlington Experiment Farm, it was found that about 30 leaves are required to produce a commercial-sized apple of good quality. Magness {28) and Magness and Overley {31) conducted similar experiments in Washington State with both apples and pears and obtained similar results. They found that under Washington conditions the same leaf area produced larger fruit but with a slightly lower sugar concen- tration than under Virginia conditions.
According to the work done by Curtis {14^ 15^ 16), Gardner {19), and Mason and Maskell {32, 33), the removal of a ring of bark prevents the translocation of carbohydrates and possibly of mineral nutrients in either direction past the ring. Thus, fruit grown above the ring would depend entirely on the leaves above the ring or on food stored above the ring for organic nutrients. Likewise, the food elaborated in leaves above the ring would be confined above the ring, and any excess material would not move out of the branch as it would in unringed branches. However, fruits on unringed branches are not necessarily dependent on food elaborated in adjacent leaves, but may draw upon jfood elaborated in adjacent branches, and food elaborated in leaves on the unringed branch may be translocated to other parts of the tree. For these reasons ringing is necessary in order to obtain a critical reading of the leaf area that is required to build the nutrient material utilized in the development of a fruit. Results obtained from experiments on ringed branches, however, are not directly comparable with those obtained from experiments on unringed branches.
2 Italic numbers in parentheses refer to Literature Cited, p. 33. 3 g is the abbreviation for gram or grams, and cm» and cms are the abbreviations for square and cubic
centimeters recently adopted by the Style Manual for U. S. Government printing.

RELATION OF LEAF AREA AND POSITION TO FRUIT QUALITY Ö
INCREASE IN SIZE OF FRUIT
Table 1 shows the coefficients of correlation between the increase in volume of the fruit and the number of leaves per apple for fruit grown on ringed branches. The correlations are positive and sig- nificant in all cases and are even higher when the fruit grown with the greater leaf areas is not considered. For example, in the Ben Davis apples for 1925 a correlation of +0.743 ±0.028 was obtained when all the fruit was considered, whereas a correlation of + 0.933 ± 0.01 was obtained when only the fruit grown with 5 to 30 leaves each was considered, indicating less increase in size of the fruit when grown with the larger leaf areas. This is brought out more clearly in Figure 2, in which the increase in volume per 10-day interval is plotted against the leaf area per apple. Figure 2 shows a rapid increase in volume with an increase in leaf area up to about 300 to 500 cm^ per fruit, depending on the variety. Above this range the fruit continues to increase in volume with an increase in leaf area, but the increase is relatively slight.
^ûo eoo aço /ooo /200 /wo ¿oo(y
FIGURE 2.~Average increase in volume of apples at 10-day intervals in relation to the leaf area per apple on ringed branches. ("Sec. D" and "Sec. E" refer to trees in different sections of the orchard to which treatments were applied at different times)
TABLE 1.—Coefficients of correlation between the increase in volume of different varieties of apples and the number of leaves per apple on ringed branches
Variety Year Num- ber of leaves
Correlation Variety Year Num- ber of leaves
Correlation
Rome Beauty Ben Davis ^
1924 1924
1925 1925
lto20 4 to 20 4 to 20 5 to 75 Oto 75 Oto 80
+0.975±0.002 + . 594± . 038 + .423± .044 + . 743± . 028 + . 805± . 017 + .829± .009
Ben Davis.-_. 1926 1926 1926 1927 1927
5 to 75 Oto 75 5 to 75 0 to 75 Oto 75
+0. 696±0.033 Do --_- + .760± .031
Delicious York Imperial Baldwin...-
+ . 673db . 031 Ben Davis.- -.... + . 704± . 029 Delicious Jonathan + . 800± . 029 Grimes Golden

6 TECHNICAL BULLETIN 338, U. S. DEPT. OF AGRICULTURE
In most cases increase in volume of fruit is not directly propor- tional to the increase in leaf area. Thus, doubling the leaf area from 5 leaves to 10 or from 10 leaves to 20 does not double the size of the fruit. This indicates that the practice of thinning, although increas- ing the size of fruit, would reduce the total crop per tree. This is shown more clearly in Table 2, which gives the percentage increase in the volume of the fruit in relation to the percentage of increase in leaf area. With 100 per cent increase in leaf area, that is, from 5 leaves to 10 leaves per fruit, there was an average increase of only 56.4 per cent in the volume of the fruit for all varieties. Increasing the leaf area from 10 to 20 leaves per fruit increased the average volume of all varieties 71.5 per cent. In the York Imperial and Grimes Golden varieties the percentage of increase in the volume of the fruit was greater than the percentage of increase in the leaf area, being 110.1 per cent for York Imperial and 103.5 per cent for Grimes Golden. The latter is not shown in Table 2. These two cases are probably due to experimental error. Why the percentage of increase in volume of the fruit should be greater with an increase in leaf area from 10 to 20 leaves per fruit than from 5 to 10 leaves per fruit is not Imown. With an increase in leaf area above 20 leaves per apple there is a reduction in the percentage of increase in volume of the fruit. The results are rather variable but show clearly that the increase in volume of the fruit is not proportional to the increase in leaf area, and that beyor^¿ 20 to 30 leaves per fruit a further increase in leaf area has less eflFect on the size of the fruit in most cases.
TABLE 2.—Percentage of increase in volume of fruit in relation to percentage of increase in leaf area on ringed branches of different varieties of apple
Percentage of increase in volume of fruit Increas e in leaves per
fruit Ben Davis
Delicious, 1925
York Imperial,
1926 Baldwin,
1927 Jonathan,
1927 Aver- age
From— To- Per cent 1925 1926, first lot 1
1926, sec- ond lot 1
5 lo 100 93.3 47.9 49.1 27.2 72.5 57.0 47.6 56.4 10 20 100 56.9 53.2 47.5 74.0 110.1 78.2 80.4 71.6 20 30 50 26.8 25.6 8.0 5.4 15.0 2.9 27.6 16.9 30 50 67 9.3 20.2 17.2 12.7 8.0 7.1 1.7 10.9 50 75 60 29.2 2.8 19.4 9.4 5.0 9.8 2.6 11.2
1 Treatments applied to two trees at different times.
The total average increase in the volume of fruit both on ringed branches and on those not ringed is shown in Table 3. The increase in volume, by 10-day intervals, of fruit on branches that were not ringed is shown in Figure 3. It will be observed that while the tend- ency for fruit on unringed branches is to increase in size with an increase in leaf area, the increase in size was not nearly so decided nor consistent as on ringed branches, and in a few cases there was a de- crease in size with an increase in leaf area. Fruit on unringed branches with a very small leaf area was usually larger than fruit with the same leaf area on ringed branches, whereas frmt on unringed branches with a large leaf area was usually smaller than the correspond- ing fruit on ringed branches. In all cases fruit grown on unringed

EELATION OF LEAF AREA AND POSITION TO FRUIT QUALITY 7
branches from which all the leaves had been stripped was as large as, or larger than, fruit on ringed branches with 10 leaves per fruit. With the exception of the Delicious variety in 1925, fruit grown on
B^tLoyy/Nf ¡927 2 —JON/iTHAN^ ¡927 3—OCUCIOUSJ92S 4—B€N a/1l//S, 1326, SCO. E S-Gfí/MCS OOLO€N,/92S G-YORK IMPCRIALJ926 7—B£.N a^i^f3,/92S &-3CN OAyt-S,f326, -SeLCO
/^OO
FiGUBE 3.—Average increase in volume of apples at 10-day intervals in relation to the leaf area per apple on unringed branches. ("Sec. D" and "Sec. E" refer to trees in different sections of the orchard to which the treatments were applied at different times)
unringed branches with 50 leaves per fruit was as small as, or smaller than, fruit grown on ringed branches with 20 leaves per fruit.
TABLE 3.—Relation of leaf area per apple to increase in volume of fruit
Variety and date of ringing
Leaves per fruit
Num- ber
Volume of fruit
at begin- ning of
test
Period of
growth
Increase in volume of fruit on-
Ringed branches
Unringed branches
Delicious, May 8 to June 4,1925..
Grimes Golden, June 17-18,1925..
Ben Davis: June 3-6,1925_
May 15-19,1926.
0 80.3
160.6 321.2 481.8 803<0
1,204.5 353.3 803.0
0 65.8
131.6 263.2 526.4
1,052.8 579.0 736.9 92.6
185.2 370.4 555.6 926.0
1,389.0 222.2 314.8 481.5 777.8 69.3
138.7 277.4 416.1 693.5
1,040.2 83.2
346.7 693.5
Crm 13.8 12.7 14.3 13.9 16.1 14.9 17.9 13.7 17.5 24.3 23.1 22.4 21.6 22.3 22.3 23.6 21.4 11.8 11.5 12.2 11.6 13.2 12.7 12.7 11.2 12.7 14.0 1.3 1.0 1.0 .9 .8
1.1 1.1 1.2 1.0
Days 85 85 85 85 85 85 85 85 85
68
107 107 107 107 107 107 107 107 107 107 112 112 112 112 112 112 112 112 112
5.6=fc0.6 28.3±1.4 54.7±1.7 85.8rfc2.4
108.8d=2.5 118.9±4.0 153.6±9.4
7.2=h .3 25.3± . 7 43.1±1.6 87.7±1.8
124.2=b2.8 128.4±4.3
43.4±3.1 64.2±2.7 98.4=k2.6
123.6±2.4 148.6±3.6 152.8±4.5
54.0=fc2.2 80.5=b2.0
118.7±4.3 128.2±4.7 150.3=b6.8 179.4±6.4
Cm»
99.4±3.9 138.3d=l. 6
97.3±1.6 88.8=fcl. 8
94.0±3.4 87.2d=2.6
100.0±2.3 129.8:1:2.3
76.8±1.6 109.4±3.4 109.2db3.3

8 TECHNICAL BULLETIN 338, Ü. S. DEPT. OF AGRICULTURE
TABLE 3.—Relation of leaf area per apple to increase in volume of fruit—Con.
Variety and date of ringing
Leaves per fruit Volume of fruit
at begin- ning of
test
Period of
growth
Increase in volume of fruit on-
Num- ber Area Ringed
branches Unringed branches
0 6
10 20 30 60 76 0
10 20 30
I 61 I 5
10 20 30 60 75 0
10 20 30
I 60 ( 0
6 10 20 30 60 75 0 8
I 51 ( 0
5 10 20 30 50 75 0 0
10 30
\ 50
0 69.3
138.7 277.4 416.1 693.5
1,040.2 0
138.7 277.4 416.1 707.4 64.8
129.7 259.4 389.1 648.5 972.7
0 129.7 259.4 389.1 648.5
0 127.1 254.2 508.4 762.6
1,271.0 1,906.5
0 203.4
1,296.4 0
71.9 143.9 287.8 431.7 719.6
1,079.2 0 0
143.9 431.7 719.5
C7TO3 21.7 22.6 23.9 23.2 23.5 24.9 25.0 27.8 27.6 23.9 24.5 31.4 7.7 8.6
10.2 10.1 9.6 9.1
10.6 9.3 9.6 9.6
10.3 25.2 27.6 27.9 31.9 27.3 24.7 23.3 28.6 22.6 39.7 34.7 35.3 34.6 40.8 43.0 42.4 42.0 33.4 36.5 32.1 35.6 38.1
76 76 76 76 76 76 76 76 76 76 76 73 73 73 73 73 73 73 73 73 73 73 91 91 91 91 91 91 91 91 91 91 64 64 64 64 64 64 64 64 64 64 64 64
9.1 ±0.4 52.3±2.3 66.5±3.7
115.7±3.0 122.0±4.9 137. 5±5.1 150.4±6.0
CT»«
Ben Davis—Continued. Jnnft 24 2.*» 1926
65.5±1.8 81.5d=2.3 82.5±2.0 85. Oil. 3
113. 0±2.8 27.6±1. 0 47.6±1.9
100.0±2.4 115.0±4.6 124.2d=3.8 130.4±4.6 York Imperial, May 27 to June 2,1926
62.3±2.3 78.2ifcl. 9 63.9±1.1 72.3±1.4 84.3±1.8
36.4±3.6 66.3±3.7
104.1±3.6 l«6,5tfc3.9 190.8±5.2 204.4±9.0 224.4±8. 6
■Rftlflwin TiiTip %-'lñ 1027
121.2±1.9 128. 5±2.5 176.5±3.4
8.6±0.9 42.4±1. 5 62.6±2.0
112.9±3.1 143.9±3.9 146.4±3.5 150.2±3.6
JnnathftTi JIITIA 20-24 1927
69. 6±1.6 71.9±1.5 87.7d=2.7 97.6±2.3 94.6±2.8
In general, trees bearing a medium to heavy crop were used for these investigations, so that the average leaf area for the tree as a whole was probably about 20 leaves per apple. The fruit on un- ringed branches seemed to respond, therefore, according to the num- ber of leaves per apple on the ti:ee as a whole rather than according to the number of leaves per apple on the experimental branches.
The discrepancies between the results obtained on ringed and on unringed branches can be explained by the data presented in the portion of this bulletin deahng with the relation of the distance and direction of the fruit from the leaves to the size and composition of apples (pp. 15 to 22), in which it is shown that there may be con- siderable translocation from a branch with a Hght crop of fruit to one with a heavy crop. There is also a relatively large use of syn- thesized materials in building trunk and root tissue, much of which probably comes from leaves on Ughtly loaded branches. Thus, the fruit grown on unringed branches with few or no leaves per fruit undoubtedly drew upon elaborated foods in adjacent branches.
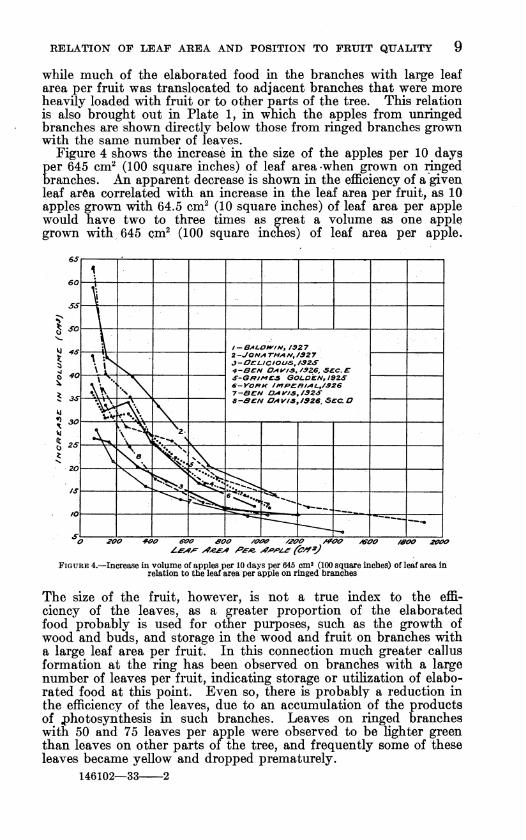
RELATION OF LEAF AREA AND POSITION TO FRUIT QUALITY 9
while much of the elaborated food in the branches with large leaf area per fruit was translocated to adjacent branches that were more heavily loaded with fruit or to other parts of the tree. This relation is also brought out in Plate 1, in which the apples from imringed branches are shown directly below those from ringed branches grown with the same niunber of leaves.
Figure 4 shows the increase in the size of the apples per 10 days per 645 cm^ (100 square inches) of leaf area-when grown on ringed branches. An apparent decrease is shown in the efficiency of a given leaf area correlated with an increase in the leaf area per fruit, as 10 apples grown with 64.5 cm^ (10 square inches) of leaf area per apple would have two to three times as great a volume as one apple grown with 645 çm^ (100 square inches) of leaf area per apple.
^^OO 600 SOO /O^û /^ÛO A^ÛO /SOO 2000
FIGURE 4. -Increase in volume of apples per 10 days per 645 cm^ (lOO square inches) of leaf area in relation to the leaf area per apple on ringed branches
The size of the fruit, however, is not a true index to the effi- ciency of the leaves, as a greater proportion of the elaborated food probably is used for other purposes, such as the growth of wood and buds, and storage in the wood and fruit on branches with a large leaf area per fruit. In this connection much greater callus formation at the ring has been observed on branches with a large number of leaves per fruit, indicating storage or utiHzation of elabo- rated food at this point. Even so, there is probably a reduction in the efficiency of the leaves, due to an accumulation of the products of photosynthesis in such branches. Leaves on ringed branches with 50 and 75 leaves per apple were observed to be lighter green than leaves on other parts of the tree, and frequently some of these leaves became yellow and dropped prematurely.
146102—33 2
10 TECHNICAL BULLETIN 338, U. S. DEPT. OF AGRICULTXJKE
CHEMICAL COMPOSITION
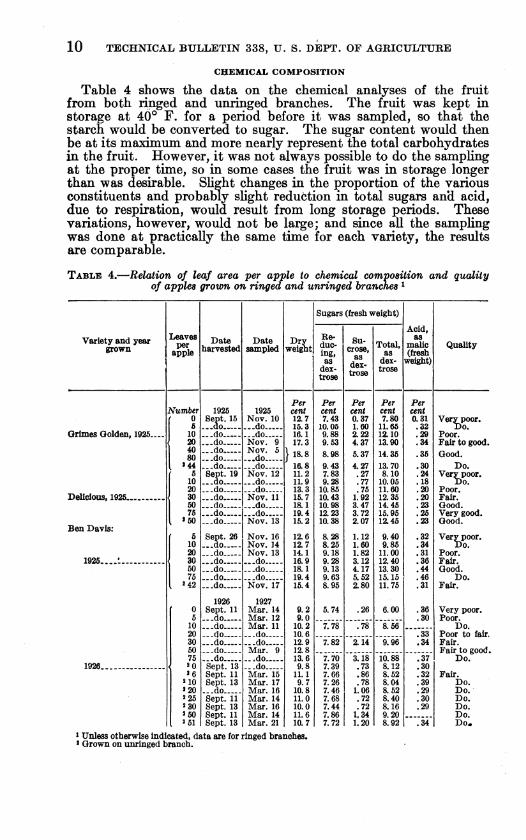
Table 4 shows the data on the chemical analyses of the fruit from both ringed and imringed branches. The fruit was kept in storage at 40° F. for a period before it was sampled, so that the starch would be converted to sugar. The sugar content would then be at its maximum and more nearly represent the total carbohydrates in the fruit. However, it was not always possible to do the sampling at the proper time, so in some cases the fruit was in storage longer than was desirable. Slight changes in the proportion of the various constituents and probably slight reduction in total sugars and acid, due to respiration, would result from long storage periods. These variations, however, would not be large; and since all the sampling was done at practically the same time for each variety, the results are comparable.
TABLE 4.—Relation of leaf area per apple to chemical composition and quality of apples grown on ringed and unringed branches ^
Sugars (fresh weight)
Acid, Variety and year
grown Leaves
per apple
Date Iiarvested
Date sampled
Dry weight
Re- duc- ing,
Su- crose,
as dex- trose
Total, as
as malic (fresh
Quality
as dex- weight) dex- trose
trose
Per Per Per Per Per Number 1926 1926 cent cent cent cent cent ( 0 Sept. 16 Nov. 10 12.7 7.43 0.37 7.80 a 31 Very poor.
5 ...do..... 15.3 10.05 1.60 n.65 .32 Grimes Golden, 1925_— 10 IlldoIII" ...do 16.1 9.88 2.22 12.10 .29 Poor.
20 ...do Nov. 9 17.3 9.63 4.37 13.90 .34 Fair to good. 40 80
...do
...do Nov. 6
...do } 18.8 8.98 6.37 14.36 .36 Good. 144 —do ...do 16.8 9.43 4.27 13.70 .30 Do.
6 Sept. 19 Nov. 12 11.2 7.83 .27 8.10 .24 Very poor. 10 ...do 11.9 9.28 .77 10.06 .18 Do. 20 "IdoIIIII ...do 13.3 10.86 .76 11.60 .20 Poor.
Delicious, 1926- 30 60
...do
...do Nov. 11
...do 15.7 18.1
10.43 10.98
1.92 3.47
12.36 14.46
.20
.23 Fair. Good.
76 ...do ...do 19.4 12.23 3.72 16.96 .26 Very good. , «60 ...do Nov. 13 15.2 10.38 2.07 12.46 .23 Good.
Ben Davis; Í ß Sept. 26 Nov. 16 12.6 8.28 L12 9.40 .32 Very poor.
10 ...do Nov. 14 12.7 8.25 L60 9.86 .34 20 ...do Nov. 13 14.1 9.18 1.82 11.00 .31 Poor.
1926 1 30 60
...do
...do..... ...do ...do
16.9 18.1
9.28 9.13
3.12 4.17
12.40 13.30
.36
.44 Fair. Good.
75 ...do ...do 19.4 9.63 6.62 16.16 .46 Do. . 242 —do
1926
Nov. 17
1927
15.4 8.95 2.80 11.76 .31 Fair.
0 Sept. 11 Mar. 14 9.2 6.74 .26 6.00 .36 Very poor. 6
10 —do —do
Mar. 12 Mar. 11
9.0 10.2
.30 Poor. "7."78" ""."78" ""8."66" Do.
20 30
--do —do
...do —do
10.6 12.9
■""."33" .34
Poor to fair. "7.12' ""¿"14" ""9."96" Fair.
60 ...do Mar. 9 12.8 Fair to good. 76 ...do ...do 13.6 "7.'7Ô' "3." is" "ÍÓ.'88" ""."37" Do.
1926 20 26
Sept. 13 Sept. 11
...do-.— Mar. 15
9.8 11.1
7.39 7.66
.73
.86 8.12 8.62
.30
.32 Fair. 2 10 Sept. 13 Mar. 17 9.7 7.26 .78 8.04 .39 Do. 220 ...do Mar. 16 10.8 7.46 L06 8.52 .29 Do. 2 25 Sept. 11 Mar. 14 11.0 7.68 .72 8.40 .30 Do. 2 30 Sept. 13 Mar. 16 10.0 7.44 .72 8.16 .29 Do. 250 Sept. 11 Mar. 14 11.6 7.86 1.34 9.20 Do. 2 61 Sept. 13 Mar. 21 10.7 7.72 1.20 8.92 "'"."34" Do*
1 Unless otherwise indicated, data are for ringed branches. 2 Grown on unringed branch.
EELATION OF LEAF ABEA AND POSITION TO FRUIT QUALITY 11
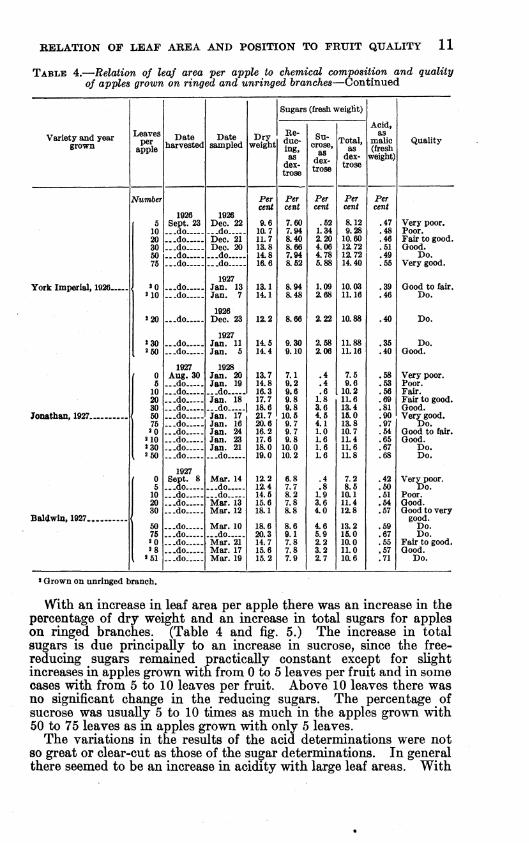
TABLE 4.—Relation of leaf area per apple to chemical composition and quality of apples grown on ringed and unringed branches—Continued
Sugars (fresh weight)
Acid,
Variety and year grown
Leaves per
apple Date
harvested Date
sampled Dry
weight Re- duc- ing»
Su- crose,
as dex- trose
Total, as
as maUc (fredi
Quality
as dex- weight) dex- trose
trose
Number Per Per Per Per Per cent cent cent cent cent
1926 1926 5 Sept. 23 Dec. 22 9.6 7.60 .62 8.12 .47 Very poor.
10 ...do —do 10.7 7.94 1.34 9.28 .48 Poor. 20 ...do Dec. 21 11.7 8.40 2.20 10.60 .46 Fair to good. 30 ...do Dec. 20 13.8 8.66 4.06 12.72 .51 Good. 60 ...do ...do 14.8 7.94 4.78 12.72 .49 Do. 75 ...do ...do
1927
16.6 8.62 6.88 14.40 .55 Very good.
York Imperial, 1926 Î0 —do Jan. 13 13.1 8.94 1,09 10.03 .39 Good to fair. UO ...do Jan. 7
1926
14.1 8.48 2.68 11.16 .46 Do.
Î20 ...do Dec. 23
1927
12.2 8.66 2.22 10.88 .40 Do.
»30 ...do Jan. 11 14.5 9.30 2.68 11.88 .35 Do. 2 60 ...do
1927
Jan. 6
1928
14.4 9.10 2.06 11.16 .40 Good.
0 Aug. 30 Jan. 20 13.7 7.1 .4 7.6 .58 Very poor. 5 ...do Jan. 19 14.8 9.2 .4 9.6 .63 Poor.
10 ...do ...do 16.3 9.6 .6 10.2 .56 Fair. 20 ...do Jan. 18 17.7 9.8 1.8 11.6 .69 Fair to good. 30 ...do ...do 18.6 9.8 3.6 13.4 .81 Good.
Jonathan, 1927 60 76
...do.-...
...do Jan. 17 Jan. 16
21.7 20.6
10.6 9.7
4.6 4.1
16.0 13.8
.90
.97 Verygood.
»0 ...do Jan. 24 16.2 9.7 1.0 10.7 .64 Good to fair. s 10 ...do Jan. 23 17.6 9.8 1.6 11.4 .65 Good. »30 -..do Jan. 21 18.0 10.0 1.6 11.6 .67 Do. »60 ...do
1927
—do 19.0 10.2 1.6 11.8 .68 Do.
I 0 'X-J.. Mar. 14 12.2 6.8 .4 7.2 .42 Very poor. 6 ...do....- 12.4 7.7 .8 8.6 .60
10 ...do ...do 14.6 8.2 1.9 10.1 .51 Poor. 20 ...do Mar. 13 15.6 7.8 3.6 11.4 .54 Good. 30 ...do Mar. 12 18.1 8.8 4.0 12.8 .57 Good to very
Baldwin, 1927 60 ...do Mar. 10 18.6 8.6 4.6 13.2 .69
good. Do.
76 ...do —do 20.3 9.1 6.9 16.0 .67 Do. »0 ...do Mar. 21 14.7 7.8 2.2 10.0 .55 Fair to good. »8 ...do Mar. 17 15.6 7.8 3.2 11.0 .57 Good.
, «61 ...do Mar. 19 15.2 7.9 2.7 10.6 .71 Do.
» Grown on unringed branch.
With au increase in leaf area per apple there was an increase in the percentage of dry weight and an increase in total sugars for apples on ringed branches. (Table 4 and fig. 5.) The increase in total sugars is due principaUy to an increase in sucrose, since the free- reducing sugars remained practically constant except for shght increases in apples grown with from 0 to 5 leaves per fruit and in some cases with from 5 to 10 leaves per fruit. Above 10 leaves there was no significant change in the reducing sugars. The percentage of sucrose was usually 5 to 10 times as much in the apples grown with 50 to 75 leaves as in apples grown with only 5 leaves.
The variations in the results of the acid determinations were not so great or clear-cut as those of the sugar determinations. In general there seemed to be an increase in acidity with large leaf areas. With

12 TECHNICAL BULLETIN 338, U. S. DEPT. OF AGRICULTURE
very small leaf areas, however, there was in some cases an increase in acidity with a decrease in leaf area.
On imringed branches there was no consistent variation in any of the constituents determined, associated with the leaf area per apple on the branch. In general the composition of fruit on unringed branches, regardless of the number of leaves per apple, was similar to that on ringed branches with 20 to 30 leaves per apple.
DESSERT QUALITY
The dessert quality of the fruit grown on ringed branches varied greatly, depending on the leaf area. All fruit grown on ringed branches without leaves was bitter and sour, with a disagreeable flavor and a coarse, woody texture. Such fruit was small and green and apparently had changed little in either appearance or eating
,c::^r^o/?K
OCUCIQUS, /9ZS
B^L orrtH, f9Z 7 \^ '
-JO^./^ THA N,/9Z7
DCLtCtOU'Sf /92S
B£N 0AI//S,/32€
-JONA TH/^N, /327 ~^CN aAy/S,/92S
SALO WIN, /azr
B€N OAÍI//S /926
-YOPK /ATP£/f/y9C, /92a 8Ai.OkyiN, /927 3CN OAVtS,/92S
JONA THAN i /327 ■OCL f ClOUS, /S2S BCN O A i^/S, /S26
-^ TOTAL. SUGAFIS ^ REiouciNO SUGARS O SUCROSE:
L£>U^£.S- /^£ñ y4PPí£ C^i/A73eñ)
FIGURE 5.—Leaf area per apple in relation to percentage of sugar in apples on ringed branches
quality from its condition at the time the treatments were applied. (PI. 1.) Fruit grown with only five leaves per apple was very little better than that grown without leaves; however, the texture of the flesh was usually less woody and the flavor somewhat less disagreeable. When the fruit was grown with 10 leaves per apple the quaUty was still poor, the flavor being rather bitter and disagreeable, although the texture was no longer woody. When grown with 20 leaves per apple, the fruit usually showed considerable improvement in quahty, although in most cases it still lacked the sweetness and good flavor characteristic of the variety. This was particularly true of Delicious, which was still rather disagreeable in flavor, whereas Jonathan was fairly good. Baldwin also showed good quality with 20 leaves per fruit, but the leaves were so large that 20 of them gave a leaf area as large as that of 30 leaves of the other varieties. With 30 leaves per

;i!¡iIZIZI7TTTrii,IllIiXXIj„Xai^^^
TI r > -{ m
Ben Davis apples grown in 1926 with various numbers of leaves per fruit as indicated by the number on the apple. Those marked "A" grown on ringed branches; those marked "B" grown on unringed branches. The apple marked "ÁB" represents the size of the fruit at the beginning of the experiment

RELATION OF LEAF AREA AND POSITION TO FRUIT QUALITY 13
fruit on ringed branches, the apples were usually of good average quality for the variety, and with 50 and 75 leaves per fruit the apples were of very good quahty, being sweet and highly flavored and usually having a firm, crisp texture.
On unringed branches the number of leaves per fruit had no per- ceptible effect on the quality of the fruit.
FIRMNESS
The differences in the texture of the fruit are shown in Table 5, which gives the firmness of the fruit when sampled as measured by the pressure tester. In the case of Ben Davis there was a gradual decrease in firmness with an increase in leaf area. In the other three varieties (York Imperial, Baldwin, and Jonathan), however, the firmness decreased to about 20 leaves per fruit and then increased with a further increase in leaf area per fruit. The texture of fruit grown with a small leaf area differs greatly from that grown with a large leaf area even though the firmness as measured by the pressure tester may be about the same. In the case of fruit grown with a small number of leaves the flesh is woody and tough as in very green fruit, whereas the flesh of fruit grown with a large number of leaves was firm but crisp. The firmness of fruit grown with a very large leaf area would seem to indicate better storage quaUty for such fruit as compared with that grown with a smaller leaf area.
TABLE 5.—Firmness of apples grown on ringed branches in relation to number of leaves per fruit
Number of
Pressure test (in pounds) at date of sampling Number
of leaves
per fruit
Pressure test (in pounds) at date of sampling
leaves per
fruit York
Imperial (1926)
Ben Davis (1926)
Baldwin (1927)
Jonathan (1927)
York Imperial
(1926)
Ben Davis (1926)
Baldwin (1927)
Jonathan (1927)
0.. 12.5 10.3 10.1 10.2
12.1 8.0 7.7 7.1
30 .- 50 -. 75
1 1 15. 3 18.2
r 13.6 I 13.1
12.6
11.6 12.5 14.3
7.7 5 10.._..- 20 _-
22.8 19.8 14.0
19.1 15.7 14.1
• 8.5 9.0
1 Fruit with 30-50 leaves became mixed and 15.3 represents pressure test on mixture of two lots.
COLOR DEVELOPMENT
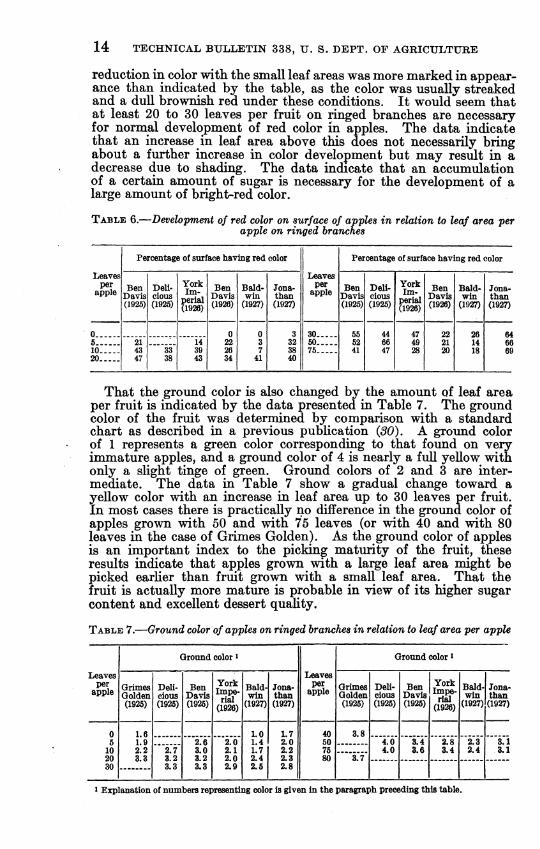
The importance of an accumulation of carbohydrates in apples in the development of red color has been pointed out in a previous report {29), in which an increase in solid-red color with an increase in leaf area has been shown for Delicious. Fletcher {18) found that 20 and 40 leaves for Kome Beauty and York Imperial, respectively, were necessary on ringed branches to produce normal color development. He also found increased color and sugar in apples when sugar in the form of ^'cerelose'^ was added to the soil. The development of red color in relation to leaf area is shown in Table 6 for apples grown on ringed branches. In general the data show an increase in red color witn 9.n increase in leaf area. There was considerable variation, however, and only in the case of Jonathan was there a consistent increase for all leaf areas. In most cases there is a decrease in red color wdth very large leaf areas. This may be due to such fruit being less exposed to the Tight owing to the large leaf area. The
14 TECHNICAL BULLETIN 338, U. S. DEPT. OF AGRICULTÜEE
reduction in color with the small leaf areas was more marked in appear- ance than indicated by the table, as the color was usually streaked and a dull brownish red under these conditions. It would seem that at least 20 to 30 leaves per fruit on ringed branches are necessary for normal development of red color in apples. The data indicate that an increase in leaf area above this does not necessarily bring about a further increase in color development but may result in a decrease due to shading. The data indicate that an accumulation of a certain amount of sugar is necessary for the development of a large amount of bright-red color.
TABLE 6.—Development of red color on surface of apples in relation to leaf area per apple on ringed branches
Percentage of surface having red color
Leaves per
apple
Percentage of surface having red color
Leaves per
apple Ben Davis (1925)
Deli- cious (1925)
York Im-
perial (1926)
Ben Davis (1926)
Bald- win
(1927)
Jona- than (1927)
Ben Davis (1925)
Deli- cious (1925)
York Im-
perial (1926)
Ben Davis (1926)
Bald- win
(1927)
Jona- than (1927)
0 0 22 26 34
0 3 7
41
3 32 38 40
30 60 75
66 62 41
44 66 47
47 49 28
22 21 20
26 14 18
64 6 10 20
21 43 47
"""33' 38
14 39 43
66 69
That the ground color is also changed by the amoimt of leaf area per fruit is indicated by the data presented in Table 7. The ground color of the fruit was determined by comparison with a standard chart as described in a previous pubUcation (SO), A ground color of 1 represents a green color corresponding to that found on very immature apples, and a ground color of 4 is nearly a full yellow with only a shght tinge of green. Ground colors of 2 and 3 are inter- mediate. The data in Table 7 show a gradual change toward a yellow color with an increase in leaf area up to 30 leaves per fruit. In most cases there is practically no difference in the groxmd color of apples grown with 50 and with 75 leaves (or with 40 and with 80 leaves in the case of Grimes Golden). As the ground color of apples is an important index to the picking maturity of the fruit, these results indicate that apples grown with a large leaf area might be picked earher than fruit grown with a small leaf area. ^ That the fruit is actually more mature is probable in view of its higher sugar content and excellent dessert quahty.
TABLE 7.—Ground color of apples on ringed branches in relation to leaf area per apple
Ground color i
Leaves per
apple
Ground color »
Leaves per
apple Grimes Golden (1925)
DeU- cious (1925)
Ben Davis (1926)
York Impe- ril
(1926)
Bald- win
(1927)
Jona- than (1927)
Grimes Golden (1926)
DeH- cious (1926)
Ben Davis (1926)
York Impe-
rial (1926)
Bald- win
(1927)
Jona-
(1927)
0 1.6 1.9 2.2 3.3
1.0 1.4 1.7 2.4 2.6
1.7 2.0 2.2 2.3 2.8
40 60 76 80
3.8
"""3.'7"
6 10 20 3.2
3.3
2.6 3.0 3.2 3.3
2.0 2.1 2.0 2.9
4.0 4.0
3.4 3.6
2.8 3.4
2.3 2.4
3.1 3.1
30
1 Explanation of numbers representing color is given in the paragraph preceding this table.

RELATION OF LEAF AREA AND POSITION TO FRUIT QUALITY 15
There were no consistent differences in either ground color or red color in fruit grown on branches that were not ringed, except that fruit grown on branches from which the leaves were all removed usually showed a higher percentage of red color, due to the greater exposure to light. The sugar content of such fruit was usually equal to that of other fruit on unringed branches with a larger leaf area.
RELATION OF DISTANCE AND DIRECTION OF FRUIT FROM LEAVES TO SIZE AND COMPOSITION OF APPLES
In the study of the relation of leaf area to the growth of apples it was foimd that the size of the fruit increased with an increase m leaf area up to^ 30 to 50 leaves per fruit even though it was often necessary to use leaves at considerable distance from the fruit to obtain this number of fully developed leaves. It was considered important, therefore, to determine to what extent the fruit could draw elaborated foods from leaves that were various distances away, and to determine the maximimi distance from which foods might be transported to the fruit. The results of such an investigation should have a bearing on fruit-bud differentiation and on the practice of thinning. Incidental to this work a study was also made of the increase in size of the fruit in relation to (1) the direction of the leaves from the fruit, (2) the size of the fruit at the beginning of the season, and (3) the seed content of the fruit.
In a prehminary report (21) it was found that apples on ringed branches could apparently draw with equal facihty upon elaborated foods from leaves 100 cm distant and from leaves adjacent to the fruit. Coit (12) showed that there was no significant difference in the size and sugar content of oranges grown on the ends of twigs that were defoliated for 8 inches back and of similar oranges grown on twigs that were not defoHated.
There seems to be no information as to the abiUty of the fruit to draw upon leaves that are above or below the fruit or that are on separate branches.
Whitehouse (42) has found that the size and shape of apples at the beginning of the season are fairly accurate indexes to their size and shape at the end of the season.
RELATION OF LEAP DISTANCE TO GROWTH OP PRUIT
The data on the growth of apples on branches that were not ringed but from which aU the leaves had been stripped are presented in Table 8. In the column headed ri2 are given the correlations between the increase in volume and the distance to the main limb; r2z repre- sents the correlation between the volume at the beginning and the distance to the main limb; ria represents the correlation between the increase in volume and the volume at the beginning; and ri2,z repre- sents the partial correlation between the increase in volume and the distance to the main limb with the volume at the beginning constant. A correlation that is four times the probable error is considered significant.

16 TECHNICAL BULLETIN 338, U. S. DEPT. OF AGRICULTURE
TABLE 8.—Growth of apples on unringed branches from which all the leaves were removed, as shown by correlations between (1) increase in volume of fruit, {2) distance from fruit to main limb, and (3) volume of fruit at the beginning of the experiment
Year Fruits
on branch
Dis- tance
be- tween apples
and main limb
Correlations i
Variety ri2 r23 ri3 ri2.3
Ben Davis 1926 1926 1927 1927 1927 1927
Number 21 25 47 45 55 33
Cm 40-150 40-180 50-310
170-475 80-280 60-320
+0.088±0.145 -. 209db . 129 -l-.449d= .078 -.714db .049 +.202± .087 -.033± .117
+0.041±0.146 -. 293± . 123 -i-.446db .079 -.315± .091 +.499± .068 -H. 230± . Ill
-hO. 831±0.045 -H.839± .040 +.500d= .074 +.595d= .065 -f-.780db .036 +.693± .061
-i-0.096±0.145 +. 071zb . 134 +.292± .090 -.690zt .053 -.346± .080 -.275± .108
York Imperial Baldwin
Do Jonathan
Do
1 ri2 denotes correlation between items 1 and 2; r23, between 2 and 3; ri3, between 1 and 3; and ri2.3, between 1 and 2 with 3 constant, viz: na a=—-^"''^^'''^^
In 1926 there were used one Ben Davis branch on which were 21 apples that were 40 to 150 cm from the main limb and one York Imperial branch on which were 25 apples 40 to 180 cm from the main limb. In neither case was there a significant correlation between the distance from the leaves or main limb and the increase in the volume of the fruit. (Table 8, r^.)
In 1927 larger branches were used on which some of the fruits were as much as 300 to 475 cm from the main limb. No correlation was found on the two Jonathan branches between the leaf distance and the increase in the volume of the fruit. On the two Baldwin branches the correlation was significant in both cases, being positive, however, in one and negative in the other.
These apparently conflicting results led to a closer study of the data in an effort to determine why a negative correlation was found in one case and a positive correlation in the other. The correlation between the size of the fruit at the beginning of the experiment and the increase in size was calculated to see whether the rate of growth had been changed by stripping the leaves from the branch. The correlations thus obtained (Table 8, ris) were positive and significant in all cases, showing that the relative rate of growth of the different apples had not been greatly changed. Although these results agree with those of Whitehouse (4^), it is rather surprising to find this relationship maintaiaed even after the removal of all the surrounding leaves.
Since a significant correlation was found between the increase in the size of the fruit and the distance from the main limb without a material change in the relative rate of growth of the fruit, there must have existed a similar correlation between the size of the fruit at the beginmng of the experiment and the distance from the maia limb. This correlation was computed and is shown in Table 8, r2z. On the Baldwia branch in which the correlation between the increase in volume and the distance to the main limb was +0.449 ±0.078, a correlation of +0.446 ±0.079 was found to exist at the beginning of the experiment between the size of the fruit and the distance to the main limb, while on the other Baldwiu branch correlations of —0.714

RELATION OF LEAF AREA AND POSITION TO FRUIT QUALITY 17
±0.049 and —0.315 ±0.091, respectively, obtained. Thus the corre- lations found on the Baldwin branches between the size and the leaf distance appear to be due to a correlation existing before any treat- ment was given. Therefore, in order to obtain the true effect of the leaf distance on the growth of the apples it was necessary to compute the partial correlation between the increase in size and the distance to the main limb, with the size at the beginning of the experiment constant. This is shown in the column headed ri2.3.
The corrected results thus obtained show significant negative correlations in the case of one Baldwin branch and one Jonathan branch, while on the other four branches the correlations are not significant. On the Baldwin branch, on which the fruit was 170 to 475 cm from any leaves, a correlation of — 0.690 ± 0.053 was obtained. Although this correlation is significant, it is somewhat questionable in view of the fact that a positive correlation of over three times the probable error was obtained on the other Baldwin branch, and also because of the negative correlation already existing at the beginning of the experiment, which may not have been entirely corrected by the use of the partial correlation.
In the case of the Jonathan branches the correlations were negative in both cases and significantly so in one case. This would seem to indicate that in the case of Jonathan as well as Baldwin the extreme distance was becoming a limiting factor in the growth of the fruit. In no case, however, was the reduction in size sufficiently marked to be noticeable by simply looking at the fruit on the branch.
The residts on unringed branches indicate that apples can, if necessary, readily obtain elaborated foods from leaves that are at a considerable distance. With the varieties used, no appreciable reduc- tion in size occurred when the leaves were 310 cm distant in the case of Baldwin, about 280 cm in the case of Jonathan, and at least 180 cm and 150 cm in the case of York Imperial and Ben Davis, respectively.
The data on the growth of apples on ringed branches with 1 fruit and 20 leaves at varying distances from the fruit are presented in Table 9. In view of the results obtained on unringed branches, it was deemed advisable to compute the partial correlations between the increase in volume and the leaf distance in order to correct for any correlation that might exist at the beginning of the experiment between the size of the fruit and the leaf distance. This was done, although no significant correlation was found (7-23) between the size of the fruit at the beginning and the leaf distance. Neither was there a significant correlation in the case of Grimes Golden between the increase in volume and the volume at the beginning (vi^). Con- trary to expectations, an apparently significant positive correlation (^12.3) was obtaÎQed with Grimes Golden in the first year of the exper- iment (1925). This would indicate that the farther the apples were from the leaves up to 110 cm the larger they would become, which seemed obviously incorrect. It was somewhat difíicult to find branches on which the fruit could be separated widely from the leaves; conse- quently the larger and longer the branch the farther the fruit was likely to be separated from the leaves, while small branches were used where the leaves were adjacent to the frait. The explanation for the positive correlation obtained the first year is probably that the large branches with the more distantly separated fruit and leaves
146102—33 3

18 TECHNICAL BULLETIN 338, XJ. S. DEPT. OF AGRICULTURE
Wore less injured by the ringing operation than were the small branches. In the later experiments an effort was made to use branches of the same size for the small distances as for the large. The first year's results indicated that at least there was no decrease in the size of the fruit even though there were no leaves within 100 cm of the fruit.
TABLE 9.—Growth of apples on ringed branches with 1 fruit and 20 leaves^ as shown by correlations between (1) increase in volume of fruit, {2) distance from fruit to leaves, and (3) volume of fruit at the beginning of the experiment
Year Branches
Dis- tance
between apples and
leaves
Correlations i
Variety r» ra n» ria.i
Grimes Golden— Do
1925 1926 1926 1927 1927 1927
Number 125 70 63 71 43 35
Cm Q-llO 0-135 0-180 0-200 0-240 O-170
+0.519±0.044 H-.265=h .075 -.092± .084 +.036± .107 -. 482± . 079 -. 038± . 114
+0.129±0.059 +.176± .078 -.158± .084 -.204± .081 -.106± .102 +. 054± . 114
+0.131±0.059 +. 117± . 079 +.904=b .016 +.672± .058 +.405± .086 +.318± .102
+0.511±0.045 +.289=b .074 +.121± .084 +. 194± . 077 --.483=t: .080
Ben Davis Baldwin... Johathan.-
Do - -.058± .114
1 For explanation see footnote to Table 8.
The experiment was repeated in 1926 with the Grimes Golden and Ben Davis varieties and in 1927 with Baldwin and Jonathan. Again in 1926 the corrected correlation (ri2.3) for Grimes Golden was posi- tive and practically significant, being 3.9 times the probable error. In this case the correlation was not due to differences in the size of the branches, as an effort was made to use as large branches for the short as for the long distances. The circumference of the branch at the ring was measured in each case, and the correlation between the circumference of the branches and the increase in the volume of the fruit was +0.064 ±0.12, showing that the size of the branches had not affected the growth of the fruit.
In the case of Ben Davis and Baldwin the correlations (^2.3) were not significant. In the Jonathan variety, in which the leaves were in some cases as much as 240 cm from the fruit, a significant negative correlation was obtained. When the branches on which the leaves were more than 170 cm from the fruit were left out, the correlation wasr= —0.059 ±0.114, indicating that the distance was not a Hmit- ing factor up to 170 cm.
In general, the results on ringed branches indicate that, if necessary^ apples can secure food materials with equal facility from leaves that are a considerable distance away and from those that are immediately adjacent. With Grimes Golden, there was no decrease in the growth of the apples up to 135 cm, with Ben Davis up to 180 cm, with Baldwin up to 200 cm, and with Jonathan up to at least 170 cm.
Although the foregoing results show that an apple will grow as large with 20 leaves that are at a considerable distance as will an apple with 20 leaves immediately adjacent, it was thought that the distant fruit might not be able to grow so large if two apples, one of which was distant, were competing for elaborated foods from the same leaves. To determine this point, branches were ringed with 20 leaves and 2 apples above the ring, one of the apples being adjacent to the leaves and the other at a considerable distance from them. (Fig. 1, D.) The data for this experiment are presented in Table 10.

BELATION OF LEAF AREA 4ND POSITION TO FRUIT QUALITY 19
TABLE 10.—Growth of apples on ringed branches in relation to distance of fruit from leaves
[Branches with 20 leaves and 2 apples]
Variety and year Distance from leaves
Volume at start Increase in volume Relative increase in volume
Average Relative Average Relative Relative volume at start
Jonathan, 1927.. Adjacent^ Cmf
42.1 43.1 44.4 38.9
Per cent 100 102.4 100 87.6
71.8 61.6
144.0 96.7
Per cent 100 85.7
100 67.2
Do- - Distant (112 cm)-.„ Adjacent--
83.7 per cent. Baldwin, 1927—
Do- Distant (109 cm).... 76.7 per cent.
In the case of the Jonathan variety seven branches with the dis- tant apples averaging 112.4 cm from the leaves were treated in this way. In the Baldwin variety 15 branches with the distant apples averaging 109.2 cm from the leaves were treated. At the beginning of the experiment the distant apples averaged slightly larger (2.4 per cent) than those adjacent to the leaves in the case of the Jona- than variety, while with the Baldwin variety the distant apples were considerably smaller (12.4 per cent). In both cases the increase in the size of the distant apples was less than that of those adjacent to the leaves, being 85.7 per cent as ^eat in the case of Jonathan and 67.2 per cent in the case of Baldwin. Since it was found in the previous experiments that the increase in volume was correlated with the volume at the beginniQg of the experiment, it would have been expected that the distant Baldwin apples would have increased in volume only 87.6 per cent as much as the apples close to the leaves if they were able to draw with equal facility upon the leaves. Since the distant fruit grew only 67.2 per cent, it appears that while they were able to obtain considerable elaborated foods from the leaves, they were not able to get as much as the adjacent fruit. The last column of Table 10 shows the amount of growth they made relative to the amount of growth they would have been expected to make from their relative volume at the start of the experiment, and shows that the rate of growth of the distant apples was decreased to 83.7 per cent in the case of Jonathan and 76.7 per cent in the case of Baldwin. Thus the distant apples apparently received respectively 83.7 and 76.7 per cent as much of the elaborated foods as did the apples adjacent to the leaves.
RELATION OF DIRECTION OF APPLES FROM LEAVES TO GROWTH OF FRUIT
In the experiment in which branches were ringed and 1 fruit and 20 leaves left above the ring at varying distances from the fruit, the leaves were in some cases above the fruit, as in Figure 1, B, and in other cases they were below the fruit or on separated branches, as in Figure 1, C. In order to determine whether the direction of the fruit from the leaves had any effect on its ability to draw on the elaborated foods, the average increase in the size of the fruit in the various positions was determined and is shown in Table 11. When the growth of the apples with the leaves adjacent was used as a standard, it was found that in no case was there a significant differ- ence in the increase in the volume of the fruit whether the leaves were below the fruit, above the fruit, or on separate branches.

20 TECHNICAL BULLETIN 338, U. S. DEPT. OF AGKICULTURE
TABLE 11.—Growth of apples on ringed branches in relation to relative position of leaves and fruit
Variety Year
With leaves adja- cent to fruit
With leaves below fruit
With leaves above fruit
With leaves on separate branch
Apples Increase in volume Apples Increase
in volume Apples Increase in volume Apples Increase
in volume
Grimes Golden. Ben Davis Baldwin Jonathan
1926 1926 1927 1927
Number 14 14 11 10
CTO3 68.6±2.7
100.7±2.9 165.0±2.4 98. Oil. 8
Number 13 7 2
Cm3 72.1±2.7 93.9±2.9
170.5
Number 28 30 14 13
CTO3 78.6±1.3 97.2±1.8
167.9±5.5 99.8±1.8
Number 15 11 13 20
Cmi 71.8±2.1
107.0±4.2 168.1±5.1 93.2±2.8
RELATION BETWEEN NUMBER OF SEEDS PER APPLE AND SIZE OF FRUIT
In the experiments herein reported it was found that there was some variation in the increase in the size of the fruit even though it was growni with the same number of leaves. This variation was not correlated wdth the distance from the leaves but seemed to depend on the size of the fruit at the beginning of the experiment. In view of the results obtained by Morris (5^), Auchter (4), Asami (2), and others, it was thought that the variation in the size of the fruit at the beginning of the experiment might be due to the number of seeds per fruit. At the time of sampling the fruit for chemical analysis the seeds in each apple were counted and the number correlated with the size of the fruit at the beginning of the experiment. The results are presented in Table 12. Slight positive correlations were obtained in the case of York Imperial and Jonathan. In the case of the former, however, the correlation is not significant, owdng to the high probable error. While the number of seeds per apple may be partly respon- sible for the variation in the size of the Jonathan apples, it apparently had little effect on the growth of the fruit in the other varieties studied.
TABLE 12.—Correlation between number of seeds and volume of apples at the begin- ning of the experiment
Variety Year Fruits Seeds per fruit
Correlation between num-
ber of seeds and volume of
fruit
Ben Davis.- 1926 1926 1927 1927
Number 57 52
259 186
Number 3 to 10 4 to 10 ItolO lto7
+0. llliO. 088 York Imperial __ +.298db .085 Jonathan _ - +.229db .040 Baldwin _ -_. -.002± .049
RELATION BETWEEN LEAF DISTANCE AND CHEMICAL COMPOSITION OF FRUIT
Following harvest, the apples from each of the experiments were arbitrarily grouped according to their distance from the leaves, and analyses for sugars and acid were then made on each group. For the fruit from ringed branches the results are presented in Table 13 and Figure 6, and for the branches that were not ringed but from which all the leaves had been stripped the results are shown in Table 14 and Figure 7.

RELATION OF LEAF AREA AND POSITION TO FRUIT QUALITY 21
TABLE 13.—Relation between leaf distance and chemical composition of fruit on ringed branches each with 1 fruit and 20 leaves above ring
Group 1 Average leaf dis- tance
Average increase in vol- ume
Sugars (percentage of fresh weight) Acid ,as
malic (percent- age of fresh
weight)
Variety and year Reduc- ing, as
dextrose
Sucrose, as dex- trose
Total, as dextrose
Ben Davis, 1926..
a b c d e f a b c d a b c
2A 2B
Cm 10 40 70 90
113 169
10 86
144 208
11 80
133 12
109
C7»3 100.5 112.2 90.7
103.6 94.0 99.8 97.7
101.6 97.2 79.4
165.2 160.9 167.2 144.0 96.7
7.7 7.9 7.5 7.6 7.5 7.2 9.8
10.4 9.7 9.6 8.4 8.4 8.2 8.2 8.2
1.8 1.6 1.6 1.8 1.4 2.2 2.1 1.8 1.9 1.1 3.1 3.6 4.5 2.6 1.4
9.5 9.5 9.0 9.4 8.9 9.4
11.9 12.2 11.6 10.7 11.5 12.0 12.7 10.8 9.6
0.30 .28 .28
Jonathan, 1927
.28
.28
.32
.52
.50
Baldwhi, 1927
.57
.47
.47
.48
.52
.45
.43
1 Groups represent fruits grown in the same range of distances from the leaves. 2 Branches with 20 leaves and 2 apples: A, Adjacent to leaves; B, distance from leaves.
TABLE 14.—Relation between leaf distance and chemical composition of fruit on unringed branches from which all leaves were stripped
Variety and year Branch No. Group Í
Average leaf dis- tance
Average increase in vol- ume
Sugars (percentage of fresh weight)
Acidas malic
(percent-
Reducing Sucrose Total
age of fresh
weight)
York Imperial, 1926. _
Í J-5 J-5 J-5 J-5 J-5 J-4 J-4 J-4 J-4 J-1 J-1 J-1 J-2 J-2 J-2 J-2 J-2 J-5 J-5 J-5
a b c d e a b c d a b c a b a b c a b c
Cm 51 67 90
124 176 57 84
100 138 114 176 229 166 271 133 205 249 233 351 443
Cm3 63.9 57.9 64.6 56.9 54.1 59.3 64.0 69.4 57.3 67.0 71.3 72.0 70.8 71.2
109.8 112.6 130.8 140.2 124.9 107.0
9.0 9.0 9.1 8.9 8.6 7.3 7.4 7.4
li 9.7
10.0 9.7 9.7 7.5 7.5 8.0 8.0 7.6 8.1
1.4 LO 1.1 L2 .8 .6 .6 .5
1.2 1.2 1.1 1.0 .8 .7
1.9 1.9 1.1 2.5 3.1 2.9
10.4 10.0 10.2 10.1 9.4 7.9 8.0 7.9 8.7
10.7 10.8 11.0 10.5 10.4 9.4 9.4 9.1
10.5 10.7 n.o
0.38 .38 .39 .40 .43
Ben Davis, 1926
Jonathan, 1927
Baldwin, 1927
.31
.29
.29
.57
.55
.60
.51
.48
.61
.61
.61
.52
.47
.48
1 Groups represent fruits grown in the same range of distances from the leaves.
In no case was there a consistent or significant variation in the acid content. There was practically no variation in the sugar content of Ben Davis apples from ringed branches whether near to or far from leaves. (Table 13.) In the case of Jonathan there was a decrease in sucrose and total sugars in the most distant group, associated with a decrease in the size of the fruit. In the case of Baldwin, however, there was an increase in sucrose and total sugars with an increase in the distance between the fruit and the leaves. Groups A and B under Baldwin (Table 13) represent apples from ringed branches on

22 TECHNICAL BULLETIN 338, V. S. DEPT. OF AGRICXJLTUKE
which there were 20 leaves and 2 apples, one of which (A) was adjacent to the leaves and the other (B) distant from the leaves. (Fig. 1, D.) In this case the apples in the B group averaged much smaUer than those in the A group and also showed a lower sucrose and total sugar content.
There was very little variation in the sugar content of the apples from the branches that were not ringed but from which the leaves had been stripped. (Table 14.) In general, a decrease or an increase in total sugar content seems to be associated to some extent with a correspondmg decrease or increase in the volume of the fruit on ringed branches (ñgi 6) rather than with the distance between the fruit and the leaves. Fruit on unringed branches, however, does not seem to show this relation. (Fig. 7.) In all cases the differences are slight.
RELATION OF LEAF AREA PER APPLE TO BUD DIFFERENTIATION
It has been shown previously that apples require a rather large leaf area of 30 or more leaves çer fruit for the development of good size and quality. Since a fruiting spur usually does not have nearly so many leaves as this, it is necessary for the fruit to draw upon other leaves. This it can do to a considerable degree. It would seem, therefore, that when trees are producing a heavy crop the fruit might utilize nearly aU of the material synthesized not only in the fruit- bearing spurs but also in spurs and twigs at considerable distances from them, so that a condition of high carbohydrate concentration apparently necessary for the formation of blossom buds would not exist in the sçurs. That heavy fruit production inhibits blossom- bud formation is well recognized. Theoretically, by thinning the fruit previous to bud diflferentiation it should be possible to reduce the carbohydrate consumption by the fruit to such an extent that there would be sufficient accumulation of carbohydrates in the spurs to bring about conditions favorable to blossom-bud differentiation. It is probable, however, that there would be little accumulation of carbo- hydrates in the spurs until not only the major requirements of the fruit but also the major requirements for the growth of the tree as a whole have been met»
REVIEW OF PREVIOUS WORK
Thompson (41) found an average increase in dry weight for two trees each of five varieties of 39.7 pounds in growth of leaves, branches, trunk, and roots during the ninth year. In a more mature tree this figure would be much larger if the tree were making a good vegetative growth. On the other hand, a 400-pound crop of apples with a dry weight of 15 per cent of the fresh weight would require 60 pounds of dry matter. It seems reasonable to suppose, therefore, that the dry weight added to the tree in the growth of leaves, shoots, and roots, and increase in girth of trunk and limbs, woidd almost equal the dry weight of a fairly heavy crop of fruit, provided the tree is m a thrifty, growing condition.
^ Since the work of Kraus and Eiaybill {26) with the tomato, con- siderable attention has been given by horticulturists to the relation of the carbohydrate-nitrogen ratio to blossom-bud differentiation in

RELATION OF LEAF AREA AND POSITION TO FRUIT QUALITY 23
FIGURE 6.—Effect of leaf distance on size and composition of apples on ringed branches. (A and B refer to apples grown on the same branch with 20 leaves, A being adjacent to and B distant from the leaves)
JO 70 90 no 130 ISO no I90 2IO 230 ZSO 270 Z90 310 330 3SO 370 3SO tlO -fSO -fSP Lc^r Oisr/tNCC fOM)
FIGURE 7.--Eflect of leaf distance on size and composition of apples on tmringed branches. (J-2 and J-5 refer to branch numbers as given in Table 14)

24 TECHNICAL BULLETIN 338 U. S. DEPT. OF AGRICULTURE
the apple. Analyses of spurs at the time of bud differentiation by various investigators—Hooker (^4), Harvey and Murneek {23) ^ Harley {22), and Auchter and Schrader {6)—have shown that blossom- bud formation is associated with a relatively high carbohydrate- nitrogen ratio. It also has been sliown that factors that reduce the carbohydrate supply, such as heavy fruit production, shading (7, 20)^ and defoliation {27y 40)j inhibit blossom-bud formation.
The time of blossom-bud differentiation varies somewhat with the location and the season. In general, however, differentiation has been found to begin during the last week of June and the first two weeks of July. Although the time at which bud differentiation begins has been well established, the literature does not clearly define the period during which differentiation may take place or the time at which maximum differentiation has occurred. Rasmussen {39) has shown for two varieties under Pennsylvania conditions that differentiation began the latter part of July during the two years studied and extended over a period of about 10 to 20 days. This would seem to indicate a rather short period for blossom-bud formation. Normally maximum differ- entiation probably occurs before the first of August. For maximum results, therefore, any^ treatment designed to influence blossom-bud formation should be given early enough to be effective during July.
That blossom thinning greatly influences blossom-bud formation has been demonstrated by several investigators. Auchter and Schrader {6) obtained blossom-bud formation on 32 to 37 per cent of the spurs from which the blossoms were removed, as compared with 1 per cent of the spurs that matured fruit and 50 per cent of the nonblossoming spurs. This work was done on a Stayman Winesap tree which was apparently bearing annually. With a biennially bear- ing Oldenburg tree, Crow {13) was able to get blossom-bud formation in the bearing year by blossom thinning previous to petal fall but not after the fruit was well set. Drain {17) obtained similar results with Wealthy trees, but thinned his blossoms by spraying with chemicals previous to fertilization. Bailey {8) removed the blossoms from 50, 75, and 100 per cent of the spurs of biennially bearing Wealthy, Oldenburg, and Yellow Transparent trees. The removal of 100 per cent of the blossoms resulted in blossom-bud formation, whereas 75 and 50 per cent removal had practically no effect.
Whfle blossom thinning has been found to favor blossom-bud for- mation, there has been little evidence that fruit thinning following the June drop, or even previous to that, affects blossom-bud forma- tion. Beach {9), working with Baldwin, Rhode Island Greening, and Hubbardston, found no effect of thinning on blossom-bud formation in New York. The time at which the thinning was done was usually between July 1 and 15, though this was not always stated. The maximum distance to which he thinned was 6 inches, and the thinned trees produced only slightly less total crop than the unthinned. It would appear, therefore, that the thinning was done too late and was not sufficiently severe to be expected to have any effect on blos- som-bud formation. Auchter (5, 4) studied the effect of thinning on several varieties, including Baldwin, York Imperial, Rome Beauty, Ben Davis, and Delicious, in West Virginia. He thinned at three different periods—just previous to the June drop (about June 15), just after the June drop (about July 1), and in midsummer (August 15). The maximum distance the fruit was thinned was 9 to 10

KELATION OF LEAF AREA AND POSITION TO FRUIT QUALITY 25
inches. The thinniag in most cases did nofc materially reduce the total crop per tree, aor did it have any effect on blossom-bud formation.
Auchter and Schrader (6) obtained blossom-bud formation on 5 per eéñt of the spurs from which the fruit was thinned at the time of the June drop, as compared with 1 per cent of the spurs that matured fruit. Auchter and Whitehouse ^ showed that complete removal of the fruit from the entire tree or from half of a tree resulted in blossom- bud differentiation and successive blossoming. Magness and Overley (81) j reporting on investigations similar to those presented herein, showed an increase in blossom-bud formation with an increase in leaf area on ringed branches under conditions in Washington State.
Aldrich (1) thinned several varieties of biennially bearing trees to 50 and 100 leaves per apple. He found that the thinning of vigorous trees, if sufficiently heavy, usually resulted in increased blossom-bud formation, even if the thinning was done as late as July 1.
From the investigations reviewed above it would seem that blossom-bud formation is normally associated with an accumulation of carbohydrates in spurs sufficient to give a fairly high carbohydrate- nitrogen ratio, and that heavy fruit production prevents this. The thinning investigations have shown that reduction of fruiting by blossom thinning may bring about blossom-bud formation, but that fruit thinning as usually practiced does not. From this it has been concluded by some that the critical period for blossom-bud differentia- tion is at or immediately following blossoming. In most fruit-thin- ning investigations, however, the thinning has been done about the time of the initiation of bud differentiation, which may not be early enough to allow any material carbohydrate accumulation before much of the differentiation is complete. Furthermore, since most thinning has not been so severe as to reduce materially the total crop, and since it must require as much carbohydrate to produce a given weight of fruit, whether the fruits are large or small, it is difficult to see how such thinning could be of much benefit to the carbohydrate accumu- lation of the tree as a whole.
RESULTS OF PRESENT INVESTIGATIONS
The data on the bud counts in 1926 following the treatments given in 1925 are presented in Table 15; those for 1927 following treatment in 1926 are presented in Table 16, and those for 1928 following treat- ment in 1927 in Table 17. The results are shown graphically in Figure 8.
In 1925 treatments were applied to three varieties—Grimes Golden, Delicious, and Ben Davis. Of these varieties. Grimes Golden and Ben Davis are normally regular annual bearers, and Delicious under some conditions tends to alternate. During this year the number of leaves per apple on the unringed branches was the same previous to the date of treatment as after, since neither leaves nor fruit were removed from such branches at the beginning of the experiment. As the fruiting spurs were not labeled, it was some- times difficult at the time the bud counts were made to distinguish between those that had fruited and those that had only blossomed in the year previous.
* AUCHTER, E. C, and WHITEHOUSE, W. E. Unpublished data. Maryland Agricultural Experiment Station. 1926.

26 TECHNICAL BULLETIN 338, U. S. DEFT. OF AGRICULTUKE
It will be seen from Table 15 and Figure 8 that on ringed branches variations in the leaf area per apple had a decided effect on the for- mation of blossom buds. In general, with 10 or less leaves per apple, blossom-bud formation was inhibited in all varieties; with 20 leaves per apple slight to moderate blossom-bud formation occurred; and with 30 or more leaves per apple a large percentage of blossom buds were formed even on spurs that bore fruit the previous season. Treatments given as late as June 17 and 18 on Grimes Golden greatly influenced the formation of blossom buds.
-ff so S6
FIGURE 8.—Effect of leaf area per apple on blossom-bud formation on ringed and unringed branches
On unringed branches there was an increased formation of blossom buds with increased leaf area, with the exception of Grimes Golden, in which no branches were used with small leaf areas per apple.
In 1926 two varieties were used—^Ben Davis, which is normally regular in "bearing but in this case was bearing biennially, and York Imperial, which tends to bear alternately and was doing so in this case. One Ben Davis tree was treated between May 15 and 19 and another was treated June 24 and 25. When bud counts were made in 1927 there were no fruit buds on either the York Imperial or the Ben Davis trees except on some of the branches that were treated, as recorded in Table 16.

RELATION OF LEAF AREA AND POSITION TO FRUIT QUALITY 27
TABLE 15.—Effect of leaf area per apple on blossom-bud formation (bud counts made in 1926 following treatments in 1925)
[Branches ringed except as noted]
Variety
G r i m € Golden.
Delicious..
Ben Davfe.
Date of treatment
June 17-18 -—do —.do —do _.-. -.-do —do.i :„. ,--do.i May 8 to June 2— May 9 to June 2—. —do May 27 to June 2- May 29 to June 2.. May 29 to June 3.. June 11 .—do.i /June 3 June 4 —do -..-do -. June 5—- - —-do _ -—do.i —.do.i —do.i -. .-..do.i
Leaf spurs of 1925
No. 8Í 64 64 51 21
121 192 88 87
30 122 117 73 34
245 68
6 10 26 66 39 29
127 76 13
-I No.
28Í 131 26 0 0
12 19
327 166 42
5 0 0
29 0
243 160 47 13 0 0
97 5 0
. 0
P.ct 76.4 67.2 2a9 0 0 9.0 9.0
78.8 65.6 25.6 4.1 0 0
10. 0
97.6 93.8 64.4 18.8 0 0
43.3 6.2 0 0
Blossoming spurs of 1925
No. 17 22 19 12 3 25 21 7 10 19 20 26 10 16 17 0 0 4 8 1 8 9
32 12 1
No. 75 29 15 1 0 2 1
61 33 4 0 0 0 1 0
25 7 3 3 0 0 7 1 1 0
P.ct 81.6 66. 44.1 7.7 0 7.4 4.5
87.9 76.7 17.3 0 0 0 6.3 0
100 100 42.8 27.3 0 0
43.8 3.0 7.7 0
Fruiting spurs of 1925
No. 24 23 25 24 27 21 31 3 10 28 43 42 33 29 14 0 2 16 21 22 26 34 26 24 11
PQ.O
No. 13 7 0 0 0 0 0
29 21 9 1 0 0 0 0
24 27 9 1 0 0 5 0 0 0
p.ct 36.1 23.3 0 0 0 0 0 9a6 67.7 24.3 2.3 0 0 0 0
100 93 37.6 4.6 0 0 12.8 0 0 0
All spurs of 1925
No. 130 109 108 87 61 167 244 98 107 169 189 141 77
289 89 6 12 45 85 62 63 170 134 49 18
No. 376 167 41 1 0 14 20
407 220 66 6 0 0
30 0
292 184 59 17 0 0
109 6 1 0
P.ct. 74.3 60.6 27.6 1.1 0 7.7 7.6
80.6 67.3 24.6 3.1 0 0 9.4 0 98.0 93.9 66.7 16.7 0 0
39.1 4.3 2.0 0
1 Unringed branch.
On ringed branches a higher percentage of blossom buds formed on the Ben Davis tree treated in May than on the one treated in Jime. Only 10 leaves per apple were necessary for the formation of blossom buds on the ringed branches of the Ben Davis tree treated in May, whereas 30 leaves per apple were necessary on the York Imperial tree treated about 10 days later.
No blossom buds were formed on unringed branches of York Imperial or Ben Davis except on the Ben Davis branch thinned to 50 leaves per apple in May.
In 1927 two varieties were used—Jonathan, which was bearing regularly, and Baldwin, which tends to bear alternately and was doing so in this case.
On ringed branches (Table 17) 20 leaves per apple caused over a third of the spurs to form blossom buds, and practically all spurs formed blossom buds with 50 or 75 leaves per apple in the case of Jonathan, whereas with Baldwin 30 leaves were necessary for the formation of blossom buds on 25 per cent of the spurs, and 75 leaves resulted in blossom buds on only 53 per cent of the spurs.
On unringed Jonathan branches there was an increased blossom- bud formation with increased leaf area per fruit, whereas with Bald- win no blossom buds were formed on any of the unringed branches, even though all the fruit was removed from one of the branches. This branch was over 3 inches in diameter at the base and about 25 feet long, so that it was thought that it would act to a large extent independently of the rest of the tree.

28 TECHNICAL BULLETIN 338, V. S. DEPT. OF AGRICULTURE
TABLE 16.—Effect of leaf area per apple on hlossom-hud formation (bud counts made in 1927 following treatments in 1926)
[Branches ringed except as noted]
Date of treatment
Leaves per
apple
Leaf spurs of 1926
Blossoming spurs of 1926
Fruiting spurs of 1926
All spurs of 1926
Variety
¡25
SS <
Is 5S
L 03
No. 0 2 5
24 24 16 9 7
16 22
a 20
102 61 30 39 42 64
9 17 6
12 50 72 38 62
Mi-H
CSC
55
íMay 15-.. 75 50 30 20 10 5
75 50 30 20 10 5
50 25
6 51 30 20 10 0
75 50 30 20 10 6
60 30 20 10 0
No. 0 2
17 49 39
8 38 24 16
II 5
145 30 24 39 59 22 26 50 47 92
110 75 66 66
No. 109 102 77 35 i 0
40 17 14 0 0 0 9 0 0 0 0 0 0 0
363 134 28
0 0 0 0 0 0 0 0
p. ct. 100 98.1 81.9 41.7 2.5 0
51,3 41.6 46.7 0 0 0 5.8 0 0 0 0 0 0 0
88.5 59.3 20.3
• 0 0 0 0 0 0 0 0
No. 72 39 20 10
1 0
50 27 8 4 0 0
17 0 0 0 0 0 0 0
71 69 30
0 0 0 0 0 0 0 0
P. ci. 100 95.1 80.0 29.4 4.0 0
84.7 79.4 34.8 15.4 0 0
14.3 0 0 0 0 0 0 0
93.4 83.1 37.5
0 0 0 0 0 0 0 0
No. 0 1 5
14 20 16 5 6
10 10 8
12 27 15 56 13 20 22 24 19 11 8
13 25 23 19
No. 10 14 11 8 1 0 5 1 0 0 0 0 1 0 0 0 0 0 0 0 6 6 2 0 0 0 0 0 0 0 0
P.cL 100 93.3 68.7 36.4 4.8 0
50.0 14.3 0 0 0 0 3.6 0 0 0 0 0 0 0
35.3 38.6 13.3 0 0 0 0 0 0 0 0
No. 0 5
27 87 83 38 62 37 41 69 38 37
274 114 110 91
121 108 69 86 63
112 173 172 127 147
No. 191 155 108 53
3 0
95 45 22 4 0 0
27 0 0 0 0 0 0 0
440 198 60
0 0 0 0 0 0 0 0
Pet. 100
May 17 96.9 May 18 80.0 ...-do 37.9 May 19 3.5
June 24 .... 0
64.« ..-.do.. June 25
.... 54.9 34,9
... do 5.5 Ben Davis __ — .do-, ....
do — - 0
0 May 191 --- 9.0 do.i. 0 ... do.i 0 June 251 ... 11
20 10 11 11
0 ....do.i 0 —-do.i-. 0 ....do.i 0 -—do.i
/May 27-28 - 0
87.5 May 28 .-- 63.9 May 28-29 -. 25.8 May 29- 0 -.--do._... 0
York Imperial-.-. May 29 to June 1— June 3 i._ - "'48
34 16 20
0 0
.-- do.i - - 0 ---do.i. 0 ..--do.i.. 0 -- do.i . - . 0
^ Unringed branch.
It is of particular interest to note that in the case of the two varieties that tend to bear alternately, York Imperial and Baldwin, practically no blossom-bud formation occurred with 20 leaves per fruit on ringed branches, whereas with Jonathan, Ben Davis, and Grimes Golden, which are normally annual bearers, fairly good blossom-bud formation occurred with 20 leaves. Blossom-bud formation in Dehcious, which tends to bear alternately but not to so great an extent as York Imperial and Baldwin, was low with 20 leaves per apple. This would seem to indicate that varieties that tend to bear alternately require a higher leaf area per apple for the development of blossom buds.
While analyses of the spurs were not made, the analyses of the fruit should be to some extent an index to the concentration of sugars in the spurs. The concentration of sugars in the spurs would be expected to be proportional to that in the fruit. In this connec- tion it is of interest to note (Table 4) that the concentration of total sugars in apples on unringed branches that failed to form blossom buds was lower than the concentration of sugars in fruit on ringed branches that formed blossom buds. In the case of Baldwin, for example, 30 leaves per apple on ringed branches were required to form

EELATION OF LEAF AREA AND POSITION TO FRUIT QUALITY 29
TABLE 17.—Effect of leaf area per apple on hlossom-bud formation (hud counts made in 1928 following treatments in 1927)
[Branches ringed except as noted]
Variety Date of treat- ment
Leaf spurs of 1927
Blossoming spurs of 1927
Fruiting spurs of 1927
Oa>
All spurs of 1927
Jonathan..
Baldwin.
June 20-22 June 21-22. —.do.... -.-do.... June 22-23. June 23-24. June 24 June 25 L. .....do.L.- —-do.i... June 24 i.. June 301.. June 3-8... June 3-9... June 6-9...
-do June 7-9...
-do June 71 ....do.i... -..do.i... -.-.do.i.... do.»...
10 5 0
60 30 10 0 0
75 50 30 20 10 5
50 8 0 0
No,
0 5
49 75 84 .54 14 13 19 75
131 75 80 75
124 55
No. P.ct
143 75 27 0 0 0
22 16 0 0 0
126 13 26
3 0 0 0 0 0 0 0
100 93.7 35.6 0 0 0
61.1 55.2 0 0 0
62.7 14.0 25.7 2.4 0 0 0 0 0 0 0
No. ( 1 7
31 76 62 72 16 31 53 156 110 42 3 2
23 29 16
No, 190 116 100 26 0 0 0
29 18 1 0 0 19 1 0 0 0 0 0 0 0 0 0
p.ct. 100 99.1 93.5 45.6 0 0 0
64.4 36.7 1.9 0 0
31.1 25.0 0 0 0 0 0 0 0 0 0
No. 0 1
11 22 14 23 20 7
16 13 46 25 10 7 8
26 25 24
No, 20 20 15
1 0 0 0 1 0 0 0 0 2 0 1 0 0 0 0 0 0 0 0
P.ct. 100 95.2 57.7 4.3 0 0 0
12.5 0 0 0 0
16.7 0
11.1 0 0 0 0 0 0 0 0
No.
23 102 165 159 146 52 60 85
277 266 127 90 85
173 109 67
57
No. 470 279 190 54 0 0 0
37 34
1 0 0
147 14 27
3 0 0 0 0 0 0 0
P.ct. 99.8 99.7 89.2 34.6
0 0 0
41.6 36.2
1.2 0 0
53.6 13.5 24.1 1.7 0 0 0 0 0 0 0
I Unringed branch. * Apples removed.
blossom buds and the fruit had a total sugar concentration of 12.8 per cent; fruit on the unringed branches had a sugar concentration of only 10 to 11 per cent and failed to form blossom buds. Similarly, York Imperial formed blossom buds on ringed branches with 30 leaves per fruit with a total sugar concentration of 12.7 per cent in the fruit; no blossom buds were formed on the unringed branches, and the total sugar concentration in the fruit in all cases was less than 12 per cent. In the case of Jonathan, blossom buds were formed on ringed branches with 20 leaves per fruit and a sugar concentration of 11.6 per cent. On the two unringed branches, in which the fruit had a similar sugar concentration of 11.6 and 11.8 per cent, blossom buds were formed to practically the same extent as on the ringed branches with 20 leaves per fruit, whereas practically no blossom buds were forined on the other unringed branches in which the sugar concen- tration of the fruit was less than 11.6 per cent. A similar relation obtained in the other varieties studied.
DISCUSSION
Varying the leaf area per apple on ringed branches greatly influ- enced the size and quahty of the fruit, whereas similar treatment on branches that were not ringed had relatively much less effect on the fruit. The question arises, then, which of these treatments more nearly represents what would happen if entire trees were treated in a similar manner? When individual branches are treated without ringing there seems to be a translocation of food materials away from

30 TECHNICAL BULLETIN 338, U. S. DEFT. OF AGRICULTURE
or to the branch, depending on whether there are more or fewer leaves per apple on the branch than on the tree as a whole. If an enth'e tree were treated so that there was a very small leaf area per apple, there would be no other soiu-ce of elaborated food supply to draw upon, except possibly reserve or storage materials that inight be present in the branches, trunk, and roots; so that under conditions of small leaf area the fruit probably would respond as on ringed branches. Thus, Auchter (4) found that when practically all of the leaves dropped early in the smnmer because of cedar rust, the fruit ceased to develop and became no larger than walnuts. On the other hand, in case an entire tree were thinned so that there was a very large leaf area per apple, there would probably be considerable translocation of elabo- rated foods away from the fruit to the trunk and roots. In the latter case a large leaf area per fruit on an entire tree would not have so great an effect on the fruit as it would on ringed branches. It prob- ably would have a much greater eflfect, however, than a similar large leaf area per fruit on an unringed branch, since the roots probably would not draw upon the synthesized foods to the same extent as fruit on adjacent branches with a smaller leaf area.
The evidence presented seems to indicate that with a Umited supply of elaborated foods the fruit may utihze the available material to a greater extent than other parts of the plant. This is brought out by the fact that fruit grown at a considerable distance from any leaves became practically as large as fruit grown adjacent to leaves and its sugar and acid content was not materially changed. For example, fruit on unringed branches from which all the leaves were reinoved grew as large as, and in most cases larger than, similar fruit on ringed branches with 10 leaves per fruit. Thus, in the case of Baldwin in 1927, there were, respectively, 47 and 45 apples on each of the two branches used. (Table 8.) Since each apple apparently used at least as much food as was elaborated in 10 leaves, the amount of food translocated into each of the branches must have been as much as was elaborated by 470 and 450 leaves, respectively. This amount of food was translocated a distance of 50 to 310 cm in one case and 170 to 475 cm in the other. A further indication that the fruit tends to monopolize the available synthesized foods is that the principal move- ment seems to be in the direction of the fruit, and may be either up- ward or downward or from one branch to another, depending on whether the fruit is above the leaves, below the leaves, or on a separate branch.
In this connection, Murneek {S5) found that vegetative growth in the tomato was inhibited by the development of the fruit and was associated with an accumulation of carbohydrates and nitrogen in the fruit. The accumulation of nitrogen, however, was thought to be the factor responsible for the decreased vegetative growth. Simi- lar results were also found for the apple {S6, 37), in which both nitro- gen and carbohydrate material accumulated in the fruit to a much greater extent than in any other part of the bearing spur system.
It would seem therefore that the fruit requires a large amount of synthesized food for proper development, and that there would be little available for vegetative growth and the development of buds unless there is suflâcient leaf area to more than supply the major re- quirements of the fruit. In this connection it has been found by Chandler and Heinicke \ll) that fruiting reduced the growth of Olden-

KBLATION OF LEAF AREA AND POSITION TO FRUIT QUALITY 31
berg apple trees. This effect is due in part to the fact that fewer and smaller leaves are produced on fruiting spurs than on nonfruiting spurs. The principal factor responsible for reduced vegetation, however, would seem to be the withdrawal of food materials from the tree into the fruit. Unless thinning were suJBSciently severe to reduce materi- ally the total crop, there would be Httle benefit to the tree, as the re- maining fruit might still use most of the available elaborated materials.
Auchter (5), working with young nonbearing peach and apple trees from one side of which the leaves had been stripped, concluded that the foods manufactured on one side of the tree were used or stored mainly in that side or in the roots directly beneath. The re- sults presented herewith seem to indicate that under certain conditions there may be a cross transfer of elaborated food materials. Thus, if one branch were bearing a very heavy crop and an adjacent branch a very Hght crop, there would be, to some extent at least, a tendency toward equahzation of the food materials.
From defoliation experiments Magness {27), Roberts {40), and Harvey and Murneek {23) have shown that there is considerable individuahty in buds and spurs and that they depend largely if not entirely upon their own leaves for development and blossom-bud formation. On the other hand. Hooker and Bradford {2ö) found with bearing trees that under some conditions there seemed to be no such localization of factors. Since apples require considerably inore leaves than are normally present on one spur to supply sufficient material for normal development, and since they can apparently utilize material elaborated at some distance for additional foods, it would seem that the presence of fruit would have an effect on the food supply of spurs and buds for some distance from it.
It has been recognized that blossom thinning may influence fruit- bud formation, but there has been little previous evidence to show that thinning of the fruit later in the season would effect blossom-bud differentiation. Paddock and Charles {38) conclude from experiinents in shading that fruit buds are differentiated previous to the time indi- cated by microscopic examination and that treatments following blossoming have no effect on differentiation except on secondary growth. The data presented in this bulletin conclusively show that treatments even as late as the middle of June, if sufficiently severe, may inhibit or greatly increase blossom-bud differentiation in the apple.
The results obtained have a direct bearing on the practice of thin- ning. They indicate that the leaves do not need to be directly adja- cent to the fruit in order for it to obtain satisfactory size and quaUty. Thus, one branch might be allowed to bear a somewhat heavier crop provided an adjacent branch had a very light crop or no crop. Fur- thermore, the fruit could be thinned from the outer ends of the branches and be left correspondingly thick farther back on the branches where the weight would be less likely to cause breakage. Such a practice, however, would probably result in less color on red varieties, owing to less exposure to light. Similarly, from the stand- point of size and quality two apples might be allowed to grow on one spur provided there was sufficient leaf area on other parts of the branch to develop them properljr.
In pubKcations dealing with thinning it is usually recommended that fruit be thinned to a certain number of inches between fruits. In

32 TECHNICAL BULLETIN 338, U. S. DEPT. OF AGRICULTURE
view of the results given here it would seem more desirable to base thinning operations on the number and size of leaves per fruit. By this it is not meant that the leaves should be counted or measured, but that the size of the leaves and their thickness on the branches or trees should be considered. Thus, when a tree is bearing heavily and the leaves are small and scattered, the fruits should be thinned to a much greater distance apart than on a tree where the leaves are large^ vigorous, and close together.
It has long been recognized that thinning apples and thus increasing the leaf area per fruit increased the size of the apples. The importance of thinning on dessert quality and color has not been so well recog- nized. Fruit grown with a small number of leaves in these tests was of very poor quality. Of the varieties studied this was particularly true of Delicious and least so of Jonathan. >
In most thinning experiments the objective has been to increase the size of the fruit without materially decreasing the total yield of ihß tree. It is improbable that any benefit can be obtained by the tree from thinning that does not materially reduce the total crop of fruit; In most cases, however, thinning experiments have resulted in more or less reduction in total crop. The maximum distance between fruits has usually been not more than 10 to 12 inches. This probably would not give as many leaves per apple as seem to be necessary for the formation of blossom buds as well as the growth of the fruit of varie- ties. 3p«.rkedlybieûni«.l in habit.
It is not claimed from the evidence presented that a biennially bear-» ing tree can be brought into annual bearing by thinning. The evir dence strongly indicates, however, that thinning of the fruit may be an important factor in maintaining trees in regular annual bearing- It also indicates that severe thinning to reduce the fruit crop, as well as pruning and fertilizing to stimulate growth and leaf area, should be resorted to in any efl*ort to change biennial trees into annual bearers.
SUMMARY
Apples were grown on both ringed and unringed branches with dif- ferent leaf areas per apple and with a given leaf area at different dis- tances and different directions from the fruit. Growth measurements were made of the fruit during the growing season, and the chemical composition was determined following harvest. Bud differentiation was determined on the branches with the different leaf areas per apple.
On unringed branches the variation in leaf area had no appreciable effect on the composition of the fruit and very little effect on the size.
On ringed branches there was an increase in size of fruit with an increase in leaf area. With the varieties studied nearly maximum growth was usually obtained with about 30 leaves per apple, and increase in leaf area above this point had relatively little effect on the size of the fruit. Increase in size even with small leaf areas was not directly proportional to the increase in leaf area per apple. With a given leaf area the size of the fruit at the end of the season was corre- lated with the size of the fruit early in the season.
The percentage of dry weight and total sugars increased with increased leaf area on ringed branches. The increases in total sugars were due primarily to increases in nonreducing sugars.
Good dessert quality was associated with the high concentration of total sugars found in fruit grown with a large leaf area. About 3Ö

RELATION OF LEAF AREA AND POSITION TO FRUIT QUALITY 33
leaves per apple were necessary to produce good quality for the variety. The ground color of fruit grown with a small number of leaves was greener and the fruit was lacking in red color as compared with fruit grown with leaves sufficient to produce good size and quality.
In the case of the Ben Davis variety the firmness of the fruit after storage decreased with an increase in the leaf area per ai)ple. In the case of the York Imperial, Baldwin, and Jonathan varieties there was a decrease in firmness of the fruit with an increase in leaf area to about 20 leaves per apple. Above 20 leaves per apple there was an increase in firmness with a further increase in leaf area.
Although the fruit requires a rather large leaf area for full develop- ment, the leaves may be a considerable distance from the fruit. Prac- tically no reduction in the size and composition of the fruit occurred when no leaves were less than 135 cm (4K feet) from Grimes Golden, 180 cm (6 feet) from Ben Davis and York Imperial, 200 cm (6K feet) from Jonathan, and over 300 cm (10 feet) from Baldwin, as compared with fruit grown with the same number of leaves that were immedi- ately adjacent to the fruit.
Apples may obtain elaborated foods with equal facility from leaves that are above the fruit, from leaves that are below the fruit, or from leaves that are on separate branches.
On ringed branches a small leaf area per apple inhibited blossom- bud formation, whereas a large leaf area caused most of the spurs to form blossom buds. About 20 to 30 leaves per fruit, depending on the variety and time of thinning, were necessary to produce sufficient blossom buds for a normal crop. Blossom-bud formation was asso- ciated with a relatively high sugar content in adjacent apples. Bud differentiation was greatly influenced by thinning treatments applied as late as June 25.
LITERATURE CITED
(1) ALDRICH, W. W. 1932. EFFECT OF FKUIT THINNING UPON CARBOHYDRATE ACCUMULATION,
FORMATION OF FRUIT BUDS, AND SET OF BLOOM IN APPLE TREES.
Amer. Soc. Hort. Sei. Proc. (1931) 28:599-604. (2) AsAMi, Y., and CHANG CHIH HU.
1928. STUDIES ON THE CORRELATION BETWEEN THE SIZE OF THE FRUIT AND SOME CHARACTERS IN THE JAPANESE PEAR. Nogaku Kwaihô (Jour. Sei. Agr. Soc. [Japan]) 303:55-66, illus. [English abstract in Japan. Jour. Bot. 4: (31), 1928.]
(3) AUCHTER, E. C. 1917. FIVE YEARS' INVESTIGATIONS IN APPLE THINNING. W. Va. Agr.
Expt. Sta. Bui. 162, 56 p., iUus. (4)
[1920]. SOME INFLUENCES OF THINNING, POLLINATION, AND FRUIT SPUR GROWTH ON THE YEARLY PERFORMANCE RECORD OF FRUIT »SPURS AND ON THE SIZE OF FRUIT PRODUCED. Amer. Soc. Hort. Sei. Proc. (1919) 16:11^131.
(5) 1923. IS THERE NORMALLY A CROSS TRANSFER OF FOODS, WATER, AND
MINERAL NUTRIENTS IN WOODY PLANTS? Md. Agr. Expt. Sta. Bul. 257, 60 p.
(6) and SCHRADER, A. L. [1924]. FRUIT SPUR GROWTH AND FRUIT BUD PRODUCTION IN THE APPLE.
Amer. Soc. Hort. Sei. Proc. (1923) 20:127-144. (7) SCHRADER, A. L., LAGASSE, F. S., and ALDRICH, W. W.
[1927]. THE EFFECT OF SHADE ON THE GROWTH, FRUIT BUD FORMATION, AND CHEMICAL COMPOSITION OF APPLE TREES. Amer. SoC. Hort. Sei. Proc. (1926) 23:368-382.

34 TECHNICAL BULLETIN 338, TJ. S. DEPT. OF AGKICULTURE
(8) BAILEY, J, S. 1929. THE EFFECT OF APPLE BLOSSOM REMOVAL ON FLOWER BUD FORMA-
TION. Amer. Soc. Hort. Sei. Proc. (1928) 25:198-201. (9) BEACH, S. A.
1903. THINNING APPLES. N. Y. State Agr. Expt. Sta. Bui. 239, p. [197]-224, iUus.
(10) BID WELL, G. L., and STERLING, W. F. 1925. PRELIMINARY NOTES ON THE DIRECT DETERMINATION OF MOISTURE.
Indus, and Engin. Chem. 17:147-149, illus. (11) CHANDLER, W. H., and HEINICKE, A. J.
[1927]. THE EFFECT OF FRUITING ON THE GROWTH OF OLDENBURG TREES. Amer. Soc. Hort. Sei. Proe. (1926) 23:36-46.
(12) CoiT,J.E. 1918. EFFECT OF ADJACENT LEAF AREA ON THE SUGAR CONTENT OF
ORANGES. Amer. Soe. Hort. Sei. Proc. (1917) 14:92-93. (13) CROW, J. W.
[1921]. BIENNIAL FRUIT BEARING IN THE APPLE. Amer. SoC. Hort. Sci. Proc. (1920) 17:52-54.
(14) CURTIS, O. F. 1920. THE UPWARD TRANSLOCATION OF FOODS IN WOODY PLANTS. I,
TISSUES CONCERNED IN TRANSLOCATION. Amer. Jour. Bot. 7 :101-124, iUus.
(15)
(16)
1923. THE EFFECT OF RINGING A STEM ON THE UPWARD TRANSFER OF NITROGEN AND ASH CONSTITUENTS. Amer. JoUF. Bot. 10:361-382, iUus.
1925. STUDIES ON THE TISSUES CONCERNED IN THE TRANSFER OF SOLUTES IN PLANTS. THE EFFECT ON THE UPWARD TRANSFER OF SOLUTES OF CUTTING THE XYLEM AS COMPARED WITH THAT OF CUTTING THE PHLOEM. Ann. Bot. [London] 39: [573J-585.
(17) DRAIN, B.D. [1925]. ANNUAL CROPS FROM BIENNIAL BEARING APPLE TREES. Amer.
Soc. Hort. Sei. Proe. (1924) 21: 300-302. (18) FLETCHER, L. A.
1930. A PRELIMINARY STUDY OF THE FACTORS AFFECTING THE RED COLOR ON APPLES. Amer. Soe. Hort. Sei. Proc. (1929) 26:191-196.
(19) GARDNER, F.E. 1925. A STUDY OF THE CONDUCTIVE TISSUES IN SHOOTS OF THE BARTLETT
PEAR AND THE RELATIONSHIP OF FOOD MOVEMENT TO DOMINANCE OF THE APICAL BUDS. Calif. Agr. Expt. Sta. Tech. Paper 20, 4? p., illus.
(20) GouRLEY, J. H., and NIGHTINGALE, G. T. 1921. THE EFFECT OF SHADING SOME HORTICULTURAL PLANTS. A PRE-
LIMINARY REPORT. N. H. Agr. Expt. Sta. Tech. Bui. 18, 22 p., illus.
(21) HALLER, M. H., and MAGNESS, J. R. [1926]. THE RELATION OF LEAF AREA TO THE GROWTH AND COMPOSITION
OF APPLES. Amer. Soe. Hort. Sei. Proe. (1925) 22:189-196. (22) HARLEY, C. P.
[1926]. NORMAL VARIATION IN THE CHEMICAL COMPOSITION OF FRUIT SPURS AND THE RELATION OF COMPOSITION TO FRUIT-BUD FOR- MATION. Amer. Soc. Hort. Sei. Proe. (1925) 22:13^146.
(23) HARVEY, E. M., and MURNEEK, A. E. 1921. THE RELATION OF CARBOHYDRATES AND NITROGEN TO THE BEHAVIOR
OF APPLE SPURS. Oreg. Agr. Expt. Sta. Bui. 176, 47 p., illus. (24) HOOKER, H. D., JR.
1920. SEASONAL CHANGES IN THE CHEMICAL COMPOSITION OF APPLE SPURS. Missouri Agr. Expt. Sta. Research Bui. 40, 51 p., illus.
(25) and BRADFORD, F. C. 1921. LOCALIZATION OF THE FACTORS DETERMINING FRUIT BUD FORMA-
TION. Missouri Agr. Expt. Sta. Research Bui. 47, 19 p. (26) KRAUS, E. J., and KRAYBILL, H. R.
1918. VEGETATION AND REPRODUCTION WITH SPECIAL REFERENCE TO THE TOMATO. Oreg. Agr. Expt. Sta. Bui. 149:5-90.
(27) MAGNESS, J. R. 1| 1917. STUDIES IN FRUIT-BUD FORMATION. Oreg. Agr. Expt. Sta. Bui.
146:3-27.

RELATION OF LEAF AREA AND POSITION TO FRUIT QUALITY 35
(28) MAGNESS, J. R. 1929. RELATION OF LEAF AREA TO SIZE AND QUALITY IN APPLES. AMER.
Soc. Hort. Sei. Proc. (1928) 25:285-288. (29)
1929. OBSERVATIONS ON COLOR DEVELOPMENT IN APPLES. Amer. SoC. Hort. Sei. Proe. (1928) 25:289-292.
(30) DiEHL, H. C, H ALLER, M. H., and GRAHAM, W. S., with the coopera- tion of CARRICK, D. B., HOWLBTT, F. S., MARSHALL, R. E., PLAGGE, H. H., and RALEIGH, G. J.
1926. THE RIPENING, STORAGE, AND HANDLING OF APPLES. U. S. Dept. Agr. Bui. 1406, 64 p., iUus.
(31) and OVERLEY, F. L. 1930. RELATION OF LEAF AREA TO SIZE AND QUALITY OF APPLES AND
PEARS. Amer. Soe. Hort. Sei. Proe. (1929) 26:160-162. (32) MASON, T. G., and MASKELL, E. J.
1928. STUDIES ON THE TRANSPORT OF CARBOHYDRATES IN THE COTTON PLANT. I. A STUDY OF DIURNAL VARIATION IN THE CARBOHY- DRATES OF LEAF, BARK, AND WOOD, AND OF THE EFFECTS OF RINGING. Ann. Bot. [London] 42 : [189]-253, illus.
(33) and MASKELL, E. J. 1928. STUDIES ON THE TRANSPORT OF CARBOHYDRATES IN THE COTTON
PLANT. II. THE FACTORS DETERMINING THE RATE AND THE DIREC- TION OF MOVEMENT OF SUGARS. Ann. Bot. [London] 42 : [571]- 636, illus.
(34) MORRIS, 0. M. 1921. STUDIES IN APPLE POLLINATION. Wash. Agr. Expt. Sta. Bui. 163,
32 p., iUus. (35) MURNEEK, A. E.
1926. EFFECTS OF CORRELATION BETWEEN VEGETATIVE AND REPRODUC- TIVE FUNCTIONS IN THE TOMATO (LYCOPERSICON ESCULENTUM MILL.) Plant Physiol. 1:3-56, illus.
(36)
(37)
[1926]. IS FRUITING OF THE APPLE AN EXHAUSTIVE PROCESS? Amer. SoC. Hort. Sei. Proc. (1925) 22:19&-200.
1928. NITROGEN AND CARBOHYDRATE DISTRIBUTION IN ORGANS OF THE BEARING APPLE SPUR. Missouri AgT. Expt. Sta. Research Bui. 119, 50 p., illus.
(38) PADDOCK, W., and CHARLES, F. G. 1929. THE EFFECT OF SHADE UPON FRUIT-BUD DIFFERENTIATION. Amer.
Soc. Hort. Sei. Proc. (1928) 25:195-197. (39) RASMUSSEN, E. J.
1930. THE PERIOD OF BLOSSOM-BUD DIFFERENTIATION IN THE BALDWIN AND MciNTOSH APPLES. Amer. Soc. Hort. Sei. Proc. (1929) 26:255-260, iUus.
(40) ROBERTS, R. H. 1923. EFFECT OF DEFOLIATION UPON BLOSSOM BUD FORMATION: AMERICAN
PLUM SPECIES. Wis. Agr. Expt. Sta. Research Bui. 56, 15 p., illus. (41) THOMPSON, R. C.
1916. THE RELATION OF FRUIT GROWING TO SOIL FERTILITY. ARK. AGR. Expt. Sta. Bui. 123, 20 p.
(42) WHITEHOUSE, W. E. 1916. A STUDY OF VARIATION IN APPLES DURING THE GROWING SEASON.
Oreg. Agr. Expt. Sta. Bui. 134, 13 p.

ORGANIZATION OF THE UNITED STATES DEPARTMENT OF AGRICULTURE WHEN THIS PUBLICATION WAS LAST PRINTED
Secretary of Agriculture. ARTHUR M. HYDE. Assistant Secretary _- R. W. DUNLAP.
Director of Scientific Work A. F. WOODS.
Director of Regulatory Work WALTER G. CAMPBELL.
Director of Extension Work C. W. WARBURTON.
Director of Personnel and Business Adminis- W. W. STOGKBERGER.
tration. Director of Information M.S. EISENHOWER.
Solicitor E. L. MARSHALL.
Bureau of Agricultural Economics NILS A. OLSEN, Chief. Bureau of Agricultural Engineering S. H. MCCRORY, Chief. Bureau of Animal Industry JOHN R. MOHLER, Chief. Bureau of Biological Survey PAUL G. REDINGTON, Chief. Bureau of Chemistry and Soils H. G. KNIGHT, Chief. Office of Cooperative Extension Work C. B. SMITH, Chief. Bureau of Dairy Industry O.E. REED, Chief. Bureau of Entomology C. L. MARLATT. Chief. Office of Experiment Stations JAMES T. JARDINE, Chief. Food and Drug Administration WALTER G. CAMPBELL, Director of
Regulatory Woi^k, in Charge. Forest Service R. Y. STUART, Chief. Grain Futures Administration J. W. T. DUVEL, Chief. Bureau of Home Economics LOUISE STANLEY, Chief. Library CLARIBEL R. BARNETT, Librarian. Bureau of Plant Industry WILLIAM A. TAYLOR, Chief. Bureau of Plant Quarantine LEE A. STRONG, Chief. Bureau of Public Roads THOMAS H. MACDONALD, Chief. Weather Bureau CHARLES F. MARVIN, Chief.
This bulletin is a contribution from
Bureau of Plant Industry WILLIAM A. TAYLOR, Chief. Division of Horticultural Crops and
Diseases E. C. AUCHTER, Principal Horticul- turist, in Charge.
36
U. S. GOVERNMENT PRINTING OFFICE: 1933
For sale by the Superintendent of Documents, Washington, D. C. Price 5 cents