renal acid-base balance · but weak buffer acids such as monobasic phosphate, uric acid,...
TRANSCRIPT

THE RENALREGULATIONOF ACID-BASE BALANCEIN MAN.II. FACTORSAFFECTING THE EXCRETIONOF
TITRATABLE ACID BY THE NORMALHUMANSUBJECT1
By W. A. SCHIESS, J. L. AYER, W. D. LOTSPEICH AND R. F. PITTSWITH THE TECHNICAL ASSISTANCE OF PHYLLIS MINER
(From the Department of Physiology, Syracuse University College of Medicine,Syracuse, N. Y.)
(Received for publication August 29, 1947)
Transformation of the slightly alkaline glomeru-lar filtrate into acid urine has been assigned in man(1 ) as in the dog (2) to the exchange of hydrogenions formed within the tubular cells for ions offixed base in the tubular urine. The most acidurine which the kidney can form is of pH 4.5 to 4.7(1, 3). At this reaction negligible quantities ofstrong acids such as hydrochloric or sulfuric canexist in free form. Accordingly the exchange ofhydrogen ions for base bound by chloride and sul-fate in the tubular urine is quantitatively insignifi-cant. But weak buffer acids such as monobasicphosphate, uric acid, creatinine, and beta-hydroxy-butyric and aceto-acetic acids can exist in con-siderable proportion in free titratable form inurine of such acidity (4). As a consequence, theexchange of hydrogen ions for base bound by weakbuffer acids in the tubular urine plays a significantrole in stabilizing the alkali reserve of the body(5).
It has been demonstrated in the dog that themajor determinants of the rate of excretion oftitratable acid are (1) the rate of excretion ofbuffer; (2) the acid strength of the buffer ex-creted; and (3) the degree of acidosis, as reflectedin the bicarbonate content of the plasma (6).Each of these factors appears to play a role inclinical acidosis in man. Thus, in severe diabeticketosis, a high rate of excretion of titratable acidis associated with a low alkali reserve and with ahigh rate of excretion of ketone bodies (4). Theketone bodies are relatively strong buffer acids.Accordingly the kidney can salvage from themonly a portion of the base they bind in the plasmaand glomerular filtrate, and although the rateof titratable acid excretion is high in diabetic
1 Aided by grants from the United States Public HealthService and the John and Mary R. Markle Foundation.
ketosis, a considerable complement of base is lostin the urine.
In the present study the role played by theseseveral factors mentioned above has been investi-gated systematically in normal human subjects inwhom acidosis has been induced by the ingestionof ammonium chloride. By infusing phosphate,creatinine and para-aminohippurate at progres-sively increasing rates to cause the elimination inthe urine of increasing quantities of these buffers,it has been found in man as in the dog that (1)the rate of excretion of titratable acid increasesin proportion to the rate of excretion of buffer;(2) at any given molar rate of excretion of bufferthe quantity of titratable acid eliminated per unitof time is greatest for the buffer of lowest acidstrength (phosphate) and least for the buffer ofgreatest acid strength (para-aminohippurate) ; and(3) the rate of excretion of titratable acid is,within limits, proportional to the degree of reduc-tion of bicarbonate in the plasma.
METHODS
The 12 experiments forming the basis of this reportwere performed on 3 normal adult male subjects. How-ever, the data presented in this paper have been restrictedto those obtained on a single subject on whoma completeset of experiments was performed. Additional experi-ments on the other 2 subjects confirmed the observationsrecorded here. Experimental methods employed werereported in the preceding paper (1).
RESULTS
The relationship between the rate of excretion oftitratable acid and the rate of excretion of buffer
It is generally recognized that the majority ofthe titratable acid of normal human urine is mono-basic phosphate, and that changes in the rate ofexcretion of endogenous phosphate are accom-
57
W. A. SCHIESS, J. L. AYER, W. D. LOTSPEICH, AND R. F. PITTS
5 EPAA730.3
J LJo
9- 02-
16-wa.~ ~ ~~.
0.1 02 0.3 0.4MILLIMOLS PHOSPHATEEXCRETEDPER MINUTE
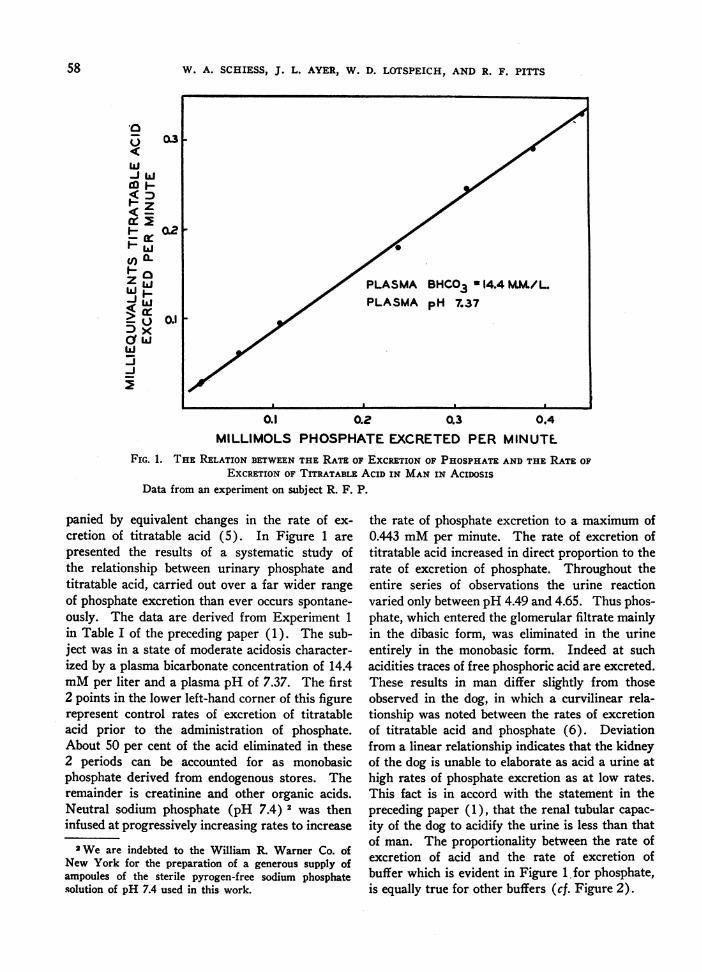
FIG. 1. THE RELATION BETWEENTHE RATE OF EXCRETIONOF PHOSPHATEANDTHE RATE OFEXCRETIONOF TITRATABLE AC SIN MANIN ACIOSIS
Data from an experiment on subject R. F. P.
panied by equivalent changes in the rate of ex-cretion of titratable acid (5). In Figure 1 arepresented the results of a systematic study ofthe relationship between urinary phosphate andtitratable acid, carried out over a far wider rangeof phosphate excretion than ever occurs spontane-ously. The data are derived from Experiment 1in Table I of the preceding paper (1). The sub-ject was in a state of moderate acidosis character-ized by a plasma bicarbonate concentration of 14.4mMper liter and a plasma pH of 7.37. The first2 points in the lower left-hand corner of this figurerepresent control rates of excretion of titratableacid prior to the administration of phosphate.About 50 per cent of the acid eliminated in these2 periods can be accounted for as monobasicphosphate derived from endogenous stores. Theremainder is creatinine and other organic acids.Neutral sodium phosphate (pH 7.4) 2 was theninfused at progressively increasing rates to increase
2We are indebted to the William R. Warner Co. ofNew York for the preparation of a generous supply ofampoules of the sterile pyrogen-free sodium phosphatesolution of pH 7.4 used in this work.
the rate of phosphate excretion to a maximum of0.443 mMper minute. The rate of excretion oftitratable acid increased in direct proportion to therate of excretion of phosphate. Throughout theentire series of observations the urine reactionvaried only between pH 4.49 and 4.65. Thus phos-phate, which entered the glomerular filtrate mainlyin the dibasic form, was eliminated in the urineentirely in the monobasic form. Indeed at suchacidities traces of free phosphoric acid are excreted.These results in man differ slightly from thoseobserved in the dog, in which a curvilinear rela-tionship was noted between the rates of excretionof titratable acid and phosphate (6). Deviationfrom a linear relationship indicates that the kidneyof the dog is unable to elaborate as acid a urine athigh rates of phosphate excretion as at low rates.This fact is in accord with the statement in thepreceding paper (1), that the renal tubular capac-ity of the dog to acidify the urine is less than thatof man. The proportionality between the rate ofexcretion of acid and the rate of excretion ofbuffer which is evident in Figure 1. for phosphate,is equally true for other buffers (cf. Figure 2).
58
FACTORSAFFECTING EXCRETION OF TITRATABLE ACID
0.1 0.2 0.3 0.4 0.5MILLIMOLS BUFFER EXCRETEDPER MINUTE
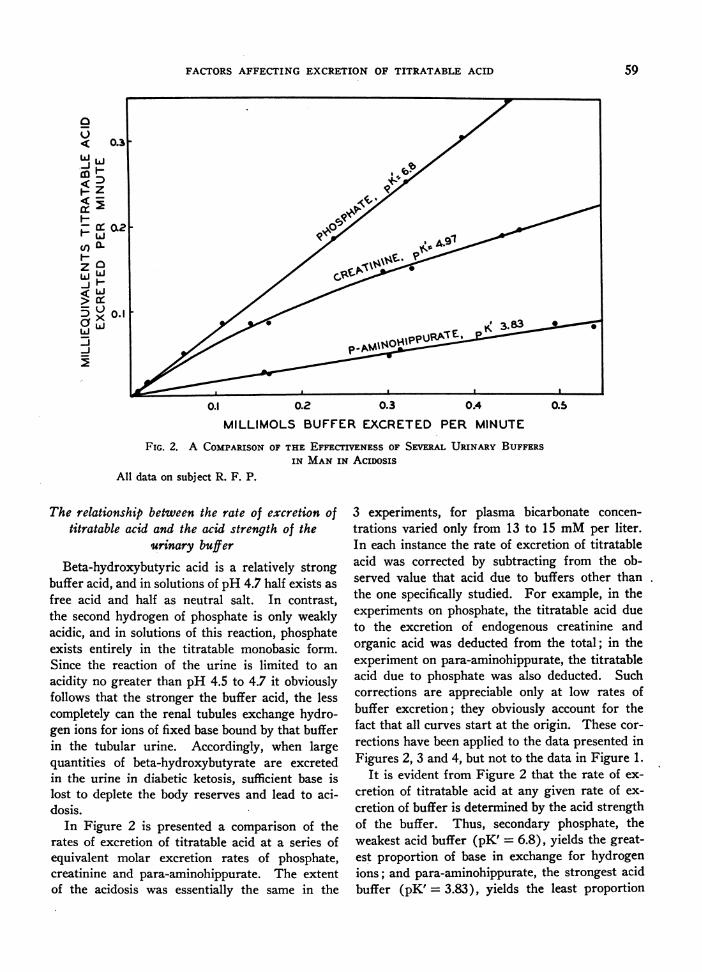
FIG. 2. A COMPARISONOF THE EFFECTIVENESS OF SEVERAL URINARY BUFFERSIN MANIN ACIDOSIS
All data on subject R. F. P.
The relationship between the rate of excretion oftitratable acid and the acid strength of the
urinary buffer
Beta-hydroxybutyric acid is a relatively strongbuffer acid, and in solutions of pH 4.7 half exists as
free acid and half as neutral salt. In contrast,the second hydrogen of phosphate is only weaklyacidic, and in solutions of this reaction, phosphateexists entirely in the titratable monobasic form.Since the reaction of the urine is limited to an
acidity no greater than pH 4.5 to 4.7 it obviouslyfollows that the stronger the buffer acid, the lesscompletely can the renal tubules exchange hydro-gen ions for ions of fixed base bound by that bufferin the tubular urine. Accordingly, when largequantities of beta-hydroxybutyrate are excretedin the urine in diabetic ketosis, sufficient base islost to deplete the body reserves and lead to aci-dosis.
In Figure 2 is presented a comparison of therates of excretion of titratable acid at a series ofequivalent molar excretion rates of phosphate,creatinine and para-aminohippurate. The extentof the acidosis was essentially the same in the
3 experiments, for plasma bicarbonate concen-
trations varied only from 13 to 15 mMper liter.In each instance the rate of excretion of titratableacid was corrected by subtracting from the ob-served value that acid due to buffers other thanthe one specifically studied. For example, in theexperiments on phosphate, the titratable acid dueto the excretion of endogenous creatinine andorganic acid was deducted from the total; in theexperiment on para-aminohippurate, the titratableacid due to phosphate was also deducted. Suchcorrections are appreciable only at low rates ofbuffer excretion; they obviously account for thefact that all curves start at the origin. These cor-
rections have been applied to the data presented inFigures 2, 3 and 4, but not to the data in Figure 1.
It is evident from Figure 2 that the rate of ex-
cretion of titratable acid at any given rate of ex-
cretion of buffer is determined by the acid strengthof the buffer. Thus, secondary phosphate, theweakest acid buffer (pK' = 6.8), yields the great-est proportion of base in exchange for hydrogenions; and para-aminohippurate, the strongest acidbuffer (pK' = 3.83), yields the least proportion
1w LLJ
6 z
I- ~LJ
Z/ a.zo-Ii-< U
5 Ux
-J
59
W. A. SCHIESS, J. L. AYER, W. D. LOTSPEICH, AND R. F. PITTS
of base in exchange for hydrogen ions. Creati-nine, a buffer of intermediate strength (pK' =4.97), occupies an intermediate position in theseries. Beta-hydroxybutyrate (pK' = 4.70) wouldfit into this series between creatinine and para-aminohippurate somewhat closer to the formerthan to the latter. Primary phosphate is such astrongly acid buffer (pK' = 2.11) that the kidneycan exchange hydrogen ions for the base whichit binds only in insignificant trace amounts. Werethey plotted in this diagram, such points wouldfall on or very near the base line.
The relation between the rate of excretion oftitratable acid and the extent of the
acidosisIt has been repeatedly observed that the inges-
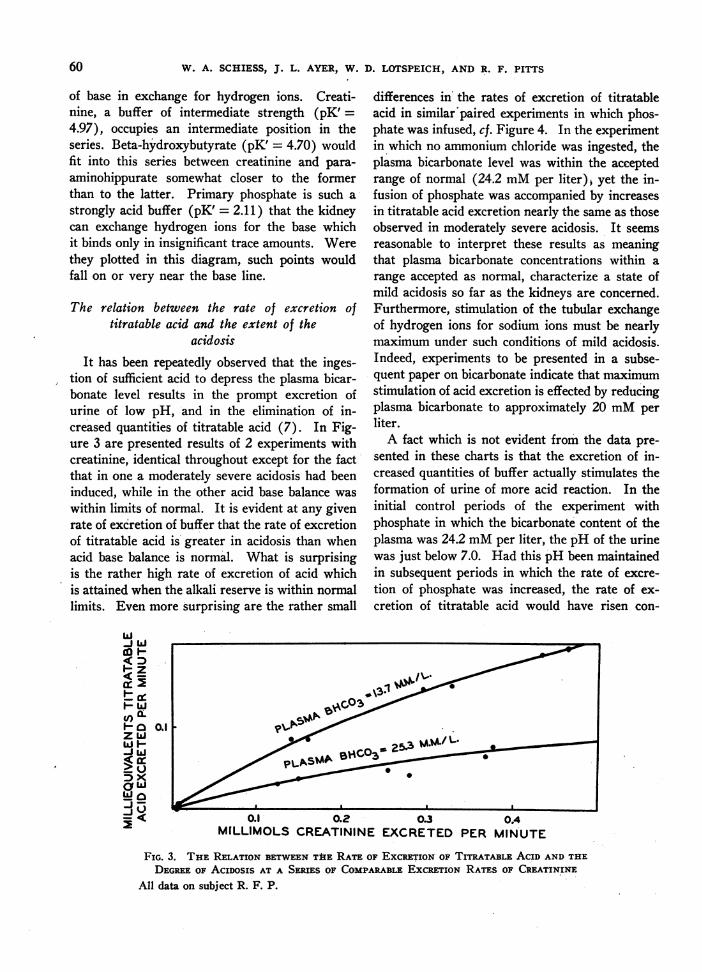
tion of sufficient acid to depress the plasma bicar-bonate level results in the prompt excretion ofurine of low pH, and in the elimination of in-creased quantities of titratable acid (7). In Fig-ure 3 are presented results of 2 experiments withcreatinine, identical throughout except for the factthat in one a moderately severe acidosis had beeninduced, while in the other acid base balance waswithin limits of normal. It is evident at any givenrate of excretion of buffer that the rate of excretionof titratable acid is greater in acidosis than whenacid base balance is normal. What is surprisingis the rather high rate of excretion of acid whichis attained when the alkali reserve is within normallimits. Even more surprising are the rather small
j Idco-z
a- 0.1z 6i
eXai
MlLLIMUL5 CREATINIP
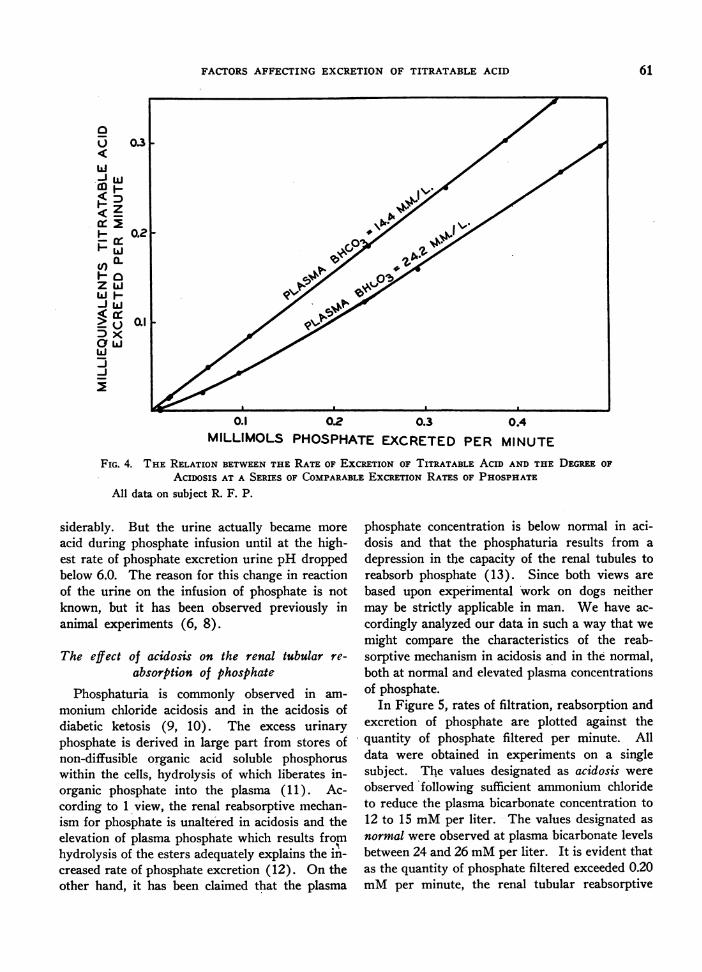
differences in the rates of excretion of titratableacid in similar paired experiments in which phos-phate was infused, cf. Figure 4. In the experimentin which no ammonium chloride was ingested, theplasma bicarbonate level was within the acceptedrange of normal (24.2 mMper liter)+ yet the in-fusion of phosphate was accompanied by increasesin titratable acid excretion nearly the same as thoseobserved in moderately severe acidosis. It seemsreasonable to interpret these results as meaningthat plasma bicarbonate concentrations within arange accepted as normal, characterize a state ofmild acidosis so far as the kidneys are concerned.Furthermore, stimulation of the tubular exchangeof hydrogen ions for sodium ions must be nearlymaximum under such conditions of mild acidosis.Indeed, experiments to be presented in a subse-quent paper on bicarbonate indicate that maximumstimulation of acid excretion is effected by reducingplasma bicarbonate to approximately 20 mMperliter.
A fact which is not evident from the data pre-sented in these charts is that the excretion of in-creased quantities of buffer actually stimulates theformation of urine of more acid reaction. In theinitial control periods of the experiment withphosphate in which the bicarbonate content of theplasma was 24.2 mMper liter, the pH of the urinewas just below 7.0. Had this pH been maintainedin subsequent periods in which the rate of excre-tion of phosphate was increased, the rate of ex-cretion of titratable acid would have risen con-
0.3 0.4NE EXCRETEDPER MINUTE
FIG. 3. THE RELATION BETWEENTHE RATE OF EXCRETIONOF TITRATABLE ACID ANDTHEDEGREEOF AcIDOSIS AT A SERIES OF COMPARABLEEXCRETION RATES OF CREATININE
All data on subject R. F. P.
60
FACTORSAFFECTING EXCRETION OF TITRATABLE ACID
0.1 02 0.3 0.4
MILLIMOLS PHOSPHATEEXCRETEDPER MINUTE
FIG. 4. THE RELATION BETWEENTHE RATE OF ExCRETION OF TITRATABLE ACID AND THE DEGREEOF
ACIDOSIS AT A SERIES OF COMPARABLEEXCRETIONRATES OF PHOSPHATEAll data on subject R. F. P.
siderably. But the urine actually became more
acid during phosphate infusion until at the high-est rate of phosphate excretion urine pH droppedbelow 6.0. The reason for this change in reactionof the urine on the infusion of phosphate is notknown, but it has been observed previously inanimal experiments (6, 8).
The effect of acidosis on the renal tubular re-
absorption of phosphate
Phosphaturia is commonly observed in am-
monium chloride acidosis and in the acidosis ofdiabetic ketosis (9, 10). The excess urinaryphosphate is derived in large part from stores ofnon-diffusible organic acid soluble phosphoruswithin the cells, hydrolysis of which liberates in-organic phosphate into the plasma (11). Ac-cording to 1 view, the renal reabsorptive mechan-ism for phosphate is unaltered in acidosis and theelevation of plasma phosphate which results fromhydrolysis of the esters adequately explains the in-creased rate of phosphate excretion (12). On theother hand, it has been claimed that the plasma
phosphate concentration is below normal in aci-dosis and that the phosphaturia results from a
depression in the capacity of the renal tubules toreabsorb phosphate (13). Since both views are
based upon experimental work on dogs neithermay be strictly applicable in man. Wehave ac-
cordingly analyzed our data in such a way that we
might compare the characteristics of the reab-sorptive mechanism in acidosis and in the normal,both at normal and elevated plasma concentrationsof phosphate.
In Figure 5, rates of filtration, reabsorption andexcretion of phosphate are plotted against thequantity of phosphate filtered per minute. Alldata were obtained in experiments on a singlesubject. The values designated as acidosis were
observed Ifollowing sufficient ammonium chlorideto reduce the plasma bicarbonate concentration to12 to 15 mMper liter. The values designated as
normal were observed at plasma bicarbonate levelsbetween 24 and 26 mMper liter. It is evident thatas the quantity of phosphate filtered exceeded 0.20mMper minute, the renal tubular reabsorptive
0u
ii.J W-iw
- z
ZLJ<a:
OwJ LiF
-J
61
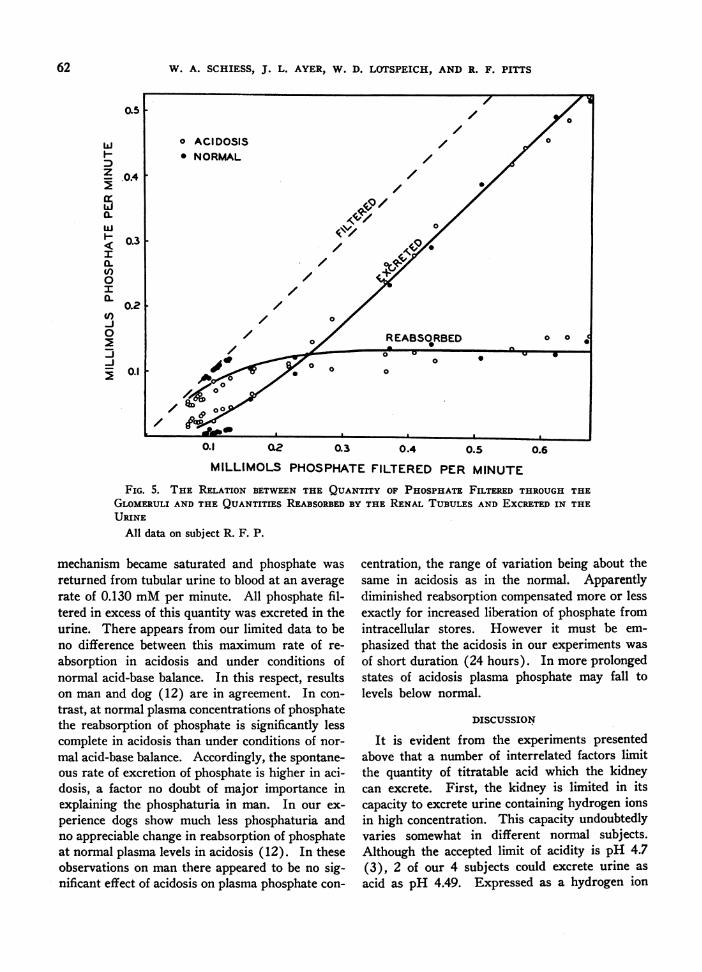
W. A. SCHIESS, J. L. AYER, W. D. LOTSPEICH, AND R. F. PITTS
0.5
LLU
'0.4
I0.3
I0.3CL)
0I0L
0.2V)-J0
-J
0~~~
0.1 02 0.3 0.4 0.5 0.6MILLIMOLS PHOSPHATEFILTERED PER MINUTE
FIG. 5. THE RELATION BETWEENTHE QUANTITY OF PHOSPHATEFILTERED THROUGHTHEGLOMERULIAND THE QUANTITIES REABSORBEDBY THE RENAL TUBULES AND EXCRETEDIN THEURINE
All data on subject R. F. P.
mechanism became saturated and phosphate wasreturned from tubular urine to blood at an averagerate of 0.130 mMper minute. All phosphate fil-tered in excess of this quantity was excreted in theurine. There appears from our limited data to beno difference between this maximum rate of re-absorption in acidosis and under conditions ofnormal acid-base balance. In this respect, resultson man and dog (12) are in agreement. In con-trast, at normal plasma concentrations of phosphatethe reabsorption of phosphate is significantly lesscomplete in acidosis than under conditions of nor-mal acid-base balance. Accordingly, the spontane-ous rate of excretion of phosphate is higher in aci-dosis, a factor no doubt of major importance inexplaining the phosphaturia in man. In our ex-perience dogs show much less phosphaturia andno appreciable change in reabsorption of phosphateat normal plasma levels in acidosis (12). In theseobservations on man there appeared to be no sig-nificant effect of acidosis on plasma phosphate con-
centration, the range of variation being about thesame in acidosis as in the normal. Apparentlydiminished reabsorption compensated more or lessexactly for increased liberation of phosphate fromintracellular stores. However it must be em-phasized that the acidosis in our experiments wasof short duration (24 hours). In more prolongedstates of acidosis plasma phosphate may fall tolevels below normal.
DISCUSSION
It is evident from the experiments presentedabove that a number of interrelated factors limitthe quantity of titratable acid which the kidneycan excrete. First, the kidney is limited in itscapacity to excrete urine containing hydrogen ionsin high concentration. This capacity undoubtedlyvaries somewhat in different normal subjects.Although the accepted limit of acidity is pH 4.7(3), 2 of our 4 subjects could excrete urine asacid as pH 4.49. Expressed as a hydrogen ion
62
-

FACTORSAFFECTING EXCRETION OF TITRATABLE ACID
gradient between urine and plasma, the maximumwhich the renal tubules of these 2 subjects couldestablish is 800 to 1. It is obvious that in theabsence of buffer the exchange of hydrogen ionsfor ions of fixed base in the tubular urine couldproceed only to an insignificant extent before thislimiting gradient would be attained. Second, thequantity of buffer which is eliminated by the nor-mal subject is rather small, and as a consequencethe quantity of base which can be salvaged fromthe urine by hydrogen ion exchange is likewisesmall. In the normal individual sufficient buffer(largely phosphate) is present in the urine topermit recovery of only 10 to 30 mEq of baseeach day. In contrast, in diabetic ketosis as muchas 500 to 1000 mMof beta-hydroxybutyrate andaceto-acetate may be excreted in 24 hours. Be-cause of the increased availability of buffer, up to150 mEqof base may be returned to the body eachday in exchange for hydrogen ions. Third, theacid strength of the buffer is a limiting factor inthe exchange of hydrogen ions for base. Obviouslythe lower the acid strength, the more completelycan the kidney salvage base bound by buffer inurine of maximal acidity. All of the secondarybase can be recovered from phosphate, only halfcan be recovered from beta-hydroxybutyrate. Al-though titratable acid excretion in diabetic ketosismay increase to some 10 times the normal becauseof increased buffer excretion, the loss of avail-able base is likewise increased because of the highacid strength of the buffer. Theoretically at leastthere must be some ceiling to the quantity of hy-drogen ions which the kidney can exchange, evenunder hypothetical conditions of unlimited sup-plies of buffer of optimum acid strength, for theprocess requires energy, and the energy availableto the kidney is not unlimited. However, in ourexperiments we have by no means approached anysuch limiting rate of exchange, although for shortperiods of time during phosphate infusion hydro-gen ions have been transferred at a rate whichwould be equivalent to 0.6 mol per day. This isequal to the excretion of 6000 ml. of one-tenth nor-mal acid, or to the renal manufacture from a bufferprecursor of 50 grams of sodium bicarbonate perday.
As Henderson (14) has so clearly pointed out,the properties of secondary phosphate are suchas t9 render it the ideal urinary buffer. However,
from this fact one must not make the extrapola-tion that the phosphaturia of diabetic acidosis iscompensatory in nature. The kidney can salvageonly the secondary base of phosphate, the primarybound base is lost in the urine. Thus, for each ex-tra mol of phosphate excreted, the body store ofbase is depleted to the extent of 1 equivalent. Theonly point about the process which can be con-sidered as compensatory is the fact that the hy-drolysis of organic intracellular phosphate enablesbase of the cells to be transferred to the plasma.Thus, the cellular phosphate esters bind roughlythe same quantity of base per mol of phosphate asis bound by inorganic phosphate in the plasma.The excretion of this phosphate in inorganic formas monobasic phosphate returns to the plasma thesecondary bound base as bicarbonate. Thus, in-tracellular base replenishes extracellular bicarbon-ate in acidosis, but in the process more than halfthe base is lost. The kidney actually operates alittle more efficiently on beta-hydroxybutyrate thanit does on phosphate, for in severe ketosis less thanhalf the base bound by this hydroxy acid in theplasma is lost in the urine.
The reabsorptive mechanism for phosphate inman appears to be qualitatively similar to that inthe dog in that the tubules are capable of reab-sorbing only a limited quantity of phosphate perunit of time. When as a consequence of an in-crease in plasma concentration the quantity de-livered into the filtrate exceeds the reabsorptivecapacity of the tubules, the excess is excreted inthe urine. The reabsorptive capacity is fixed,limited, and independent of plasma concentrationwhen that concentration is sufficiently high to satu-rate the mechanism. This maximum reabsorptivecapacity appears to be unaffected in either dogor man by moderately severe acidosis of shortduration. However, at low plasma concentra-tions the kinetics of the reabsorptive process ap-pear to be altered in man, for reabsorption is onlypartial and excretion is correspondingly increased.The diminished reabsorption at normal plasmaconcentrations is undoubtedly a factor in explain-ing the phosphaturia of acidosis in man. Ap-parently, in our experiments increased excretionwas balanced more or less exactly by increaseddelivery of inorganic phosphate into the plasma,for we have observed no significant differences inplasma level in man as a consequence of acidosis.
63

W. A. SCHIESS, J. L. AYER, W. D. LOTSPEICH, AND R. F. PITTS
CONCLUSIONS
The rate of excretion of titratable acid in man as
in the dog is largely determined by 3 factors,namely: (1) the rate of excretion of buffer; (2)the acid strength of the buffer; and (3) the ex-
tent of the reduction in the plasma concentrationof bicarbonate. For phosphate and para-amino-hippurate the relationship between buffer andtitratable acid excretion is essentially a linear one
within the range of buffer excretion that we havestudied. For creatinine the relationship is curvi-linear because urine pH increases slightly at highrates of buffer excretion. At any given molarrate of excretion of buffer the rate of eliminationof titratable acid is highest for the buffer of lowestacid strength. The accepted normal range of 24 to26 mMof bicarbonate per liter of plasma con-
stitutes a state of mild acidosis so far as the kidneysare concerned, and titratable acid elimination pro-
ceeds at a nearly maximal rate when a buffer offavorable acid strength is presented to the kidney.Further reduction of plasma bicarbonate to 20 mMper liter maximally stimulates the acid excretorymechanism. The results presented are in accordwith the view that the urine is acidified by theexchange of hydrogen ions formed within the tu-bular cells for ions of fixed base in the tubularurine.
BIBLIOGRAPHY
1. Pitts, R. F., Lotspeich, W. D., Schiess, W. A., andAyer, J. L., The renal regulation of acid-base bal-ance in normal man. I. The nature of the mecha-
nism for acidifying the urine. J. Clin. Invest.,1948, 27, 48.
2. Pitts, R. F., and Alexander, R. S., The nature of therenal tubular mechanism for acidifying the urine.Am. J. Physiol., 1945, 144, 239.
3. Henderson, L. J., and Palmer, W. W., On the in-tensity of urinary acidity in normal and pathologi-cal conditions. J. Biol. Chem., 1913, 13, 393.
4. Peters, J. P., and Van Slyke, D. D., QuantitativeClinical Chemistry. Vol. I, Interpretations. Wil-liams and Wilkins Co., Baltimore, 1932.
5. Henderson, L. J., and Palmer, W. W., On the severalfactors of acid excretion. J. Biol. Chem., 1914,17, 305.
6. Pitts, R. F., and Lotspeich, W. D., Factors governingthe rate of excretion of titratable acid in the dog.Am. J. Physiol., 1946, 147, 481.
7. Gamble, J. L., Blackfan, K. D., and Hamilton, B.,A study of the diuretic action of acid-producingsalts. J. Clin. Invest., 1925, 1, 359.
8. Hendrix, B. M., and Sanders, J. P., The effect ofinjections of sodium phosphates and sodium hip-purate upon the excretion of acid and ammonia bythe kidney. J. Biol. Chem., 1923, 58, 503.
9. Haldane, J. B. S., Experiments on the regulation ofthe blood's alkalinity. J. Physiol., 1921, 55, 265.
10. Guest, G. M., Organic phosphates of the blood andmineral metabolism in diabetic acidosis. Am. J.Dis. Child., 1942, 64, 401.
11. Guest, G. M., and Rapoport, S., Organic acid-solublephosphorus compounds of the blood. Physiol.Rev., 1941, 21, 410.
12. Pitts, R. F., and Alexander, R. S., The renal reabsorp-tive mechanism for inorganic phosphate in normaland acidotic dogs. Am. J. Physiol., 1944, 142, 648.
13. Harrison, H. E., and Harrison, H. C., The effect ofacidosis upon the renal tubular reabsorption ofphosphate. Am. J. Physiol., 1941, 134, 781.
14. Henderson, L. J., A critical study of the process ofacid excretion. J. Biol. Chem., 1911, 9, 403.
64