requirement for rho gtpases and pi 3-kinases during apoptotic cell phagocytosis by macrophages
TRANSCRIPT

Brief Communication 195
Requirement for Rho GTPases and PI 3-kinasesduring apoptotic cell phagocytosis by macrophagesYann Leverrier and Anne J. Ridley
In vivo, apoptotic cells are removed by surrounding starvation [4, 5]. Murine bone marrow–derived macro-phages were used as phagocytes. They use the vitronectinphagocytes, a process thought to be essential for
tissue remodeling and the resolution of receptor (an avb3 integrin) as a recognition mechanism toingest a variety of apoptotic cells [6] and apoptotic Baf-3inflammation [1]. Although apoptotic cells are
known to be efficiently phagocytosed by cells (see below).macrophages, the mechanisms whereby theirinteraction with the phagocytes triggers their Phagocytosis is a process used by phagocytes to engulfengulfment have not been described in mammals. large particles. Although phagocytosis is actin dependent,Here, we report that primary murine bone the protrusive activity during the engulfment and themarrow–derived macrophages (using avb3 integrin signaling pathways mediating phagocytosis depends onfor apoptotic cell uptake) extend lamellipodia to the type of receptor stimulated [7, 8]. We used scanningengulf apoptotic cells and form an actin cup where electron microscopy as a first approach to characterize thephosphotyrosine accumulates. Rho GTPases and PI engulfment of apoptotic cells. The binding of apoptotic3-kinases have been widely implicated in the Baf-3 cells to macrophages induced the formation ofregulation of the actin cytoskeleton [2, 3]. We show plasma membrane protrusions from the macrophages atthat inhibition of Rho GTPases by Clostridium the contact points between the two cells (Figure 1A).difficile toxin B prevents apoptotic cell phagocytosis Lamellipodia rose around and above the apoptotic cells,and inhibits the accumulation of both F-actin and and this led to the formation of a cup that eventuallyphosphotyrosine. Importantly, the Rho GTPases surrounded the apoptotic cells. Apoptotic cells induced aRac1 and Cdc42 are required for apoptotic cell dramatic increase in the F-actin concentration at the siteuptake whereas Rho inhibition enhances uptake. of contact with the macrophages (Figure 1B), and protru-The PI 3-kinase inhibitor LY294002 also prevents sions rich in F-actin extended from the macrophage dorsalapoptotic cell phagocytosis but has no effect on the surface to form an actin cup. We obtained similar resultsaccumulation of F actin and phosphotyrosine. using different types of apoptotic cells (primary interleu-These results indicate that both Rho GTPases kin-3-dependent cells starved of growth factors, stauro-and PI 3-kinases are involved in apoptotic cell sporine-treated U937 cells, UV-treated Jurkat cells) andphagocytosis but that they play distinct roles in different types of phagocytes (bone marrow GM-CSF-this process. derived phagocytes, J774 mouse macrophage cell line)
(data not shown). Taken together, these results indicateAddresses: Ludwig Institute for Cancer Research, 91 Riding House that a protrusive activity accompanies apoptotic cell uptake.Street, London W1P 8BT, and the Department of Biochemistry andMolecular Biology, University College London, London WC1E 6BT,United Kingdom. Tyrosine phosphorylated proteins accumulate at nascent
phagosomes during FcgR-mediated and complement-Correspondence: Anne J. Ridley mediated phagocytosis [8]. Similarly, phosphotyrosine ac-E-mail: [email protected]
cumulated at the site of apoptotic cell ingestion withinthe actin cups (Figure 1B). When apoptotic cells wereReceived: 11 September 2000
Revised: 4 December 2000 completely engulfed, they were no longer associated withAccepted: 22 December 2000 phosphotyrosine or F-actin filaments (data not shown).
These results indicate that a tyrosine kinase pathway isPublished: 6 February 2001activated following the binding of apoptotic cells to themacrophages.Current Biology 2001, 11:195–199
0960-9822/01/$ – see front matter We developed a protocol using epifluorescence micros- 2001 Elsevier Science Ltd. All rights reserved.copy to quantify the phagocytosis of apoptotic cells bymacrophages. This protocol allowed the apoptotic cellsthat had been internalized to be distinguished from thosethat had only been bound to the exterior of the macro-Results and discussion
To study the phagocytosis of apoptotic cells by macro- phages (Figure 2A). All apoptotic Baf-3 cells were labeledwith CFSE and readily identified by their intense greenphages, we used as a prey the interleukin-3-dependent
Baf-3 pre-B cell line since these cells undergo a robust fluorescence. Cell surface proteins on Baf-3 cells werealso biotinylated, and following fixation, only those Baf-3and well-characterized apoptosis following growth factor

196 Current Biology Vol 11 No 3
Figure 1
Protrusive activity and tyrosinephosphorylation during apoptotic cellengulfment by macrophages.(A) Scanning electron micrographs are shownof bone marrow CSF-1-derivedmacrophages incubated for 2 hr withapoptotic Baf-3 cells. Different stages ofengulfment are shown: early (left), middle(center), and late (right). Bars 5 5 mm.(B) F-actin and phosphotyrosine colocalizeduring the phagocytosis of apoptotic cells.Macrophages were cultured with apoptoticcells for 45 min. Actin filaments werevisualized using rhodamine-conjugatedphalloidin (left panel). The actin cytoskeletonof apoptotic Baf-3 cells is barely detectable.This may be due to depolymerization orcleavage of F-actin, which occurs duringapoptosis [27–29]. Phosphotyrosine wasdetected with mouse anti-phosphotyrosineantibody followed by Cy5-conjugated anti-mouse antibody (middle panel). Apoptoticcells were labeled with a cell-permeablegreen fluorescent dye (CFSE) to allow theirdetection (right panel). Arrowheads indicatecolocalization of actin cups, phosphotyrosinefoci, and apoptotic cells. Bars 5 10 mm.
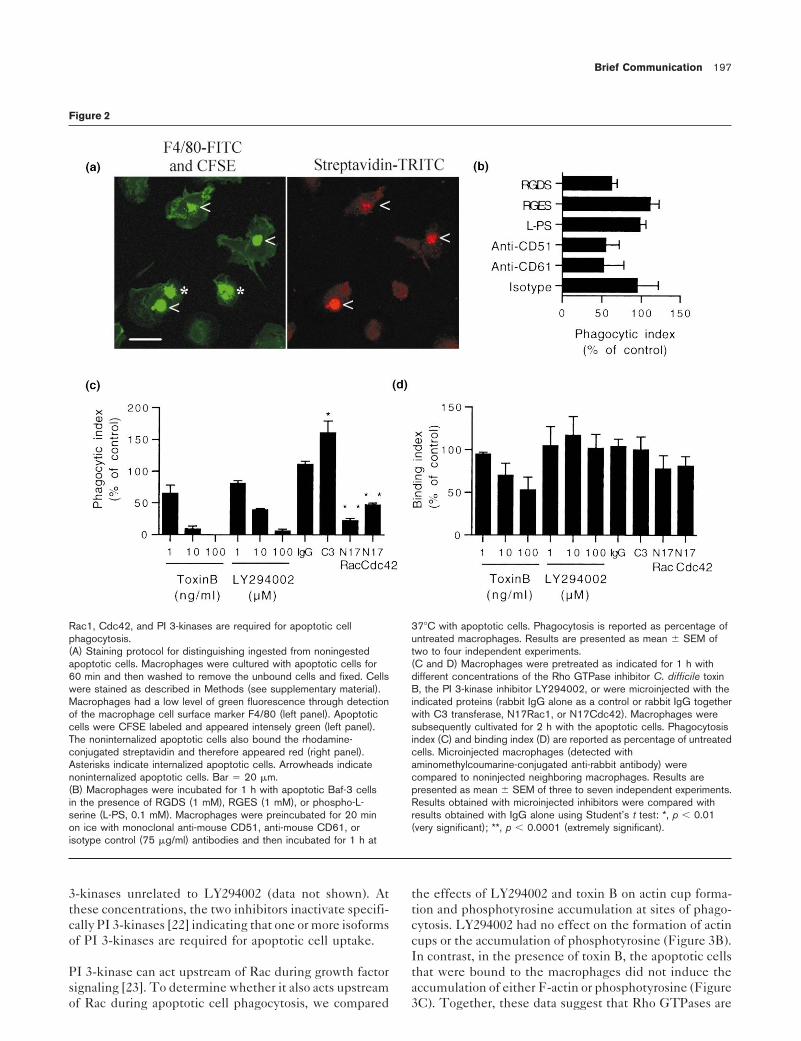
cells that were not internalized were accessible to and ing (IgG control, Figure 2D) or uptake by macrophages(IgG control, Figure 2C). Microinjection of C3 transferasecould be labeled with rhodamine-conjugated streptavidin.
Incompletely or noningested Baf-3 cells were therefore from Clostridium botulinum which ADP ribosylates andinactivates Rho [15] significantly increased apoptotic cellidentified by their red fluorescence (see arrowheads).uptake. In contrast, microinjection of the dominant inhibi-tory mutants of Rac1 (N17Rac1) or Cdc42 (N17Cdc42)We investigated which receptor was used by macrophagesseverely impaired apoptotic cell phagocytosis. None offor apoptotic Baf-3 cell uptake (Figure 2B). Phagocytosisthe inhibitors had any significant effect on apoptotic cellwas significantly inhibited by the integrin binding peptidebinding. Microinjection of N17Rac or N17Cdc42 also in-RGDS but not by control peptide RGES or by phospho-hibited phagocytosis of apoptotic Jurkat cells (UV-treatedL-serine, an inhibitor of the phosphatidylserine receptor-or staurosporine-treated) (data not shown). These resultsmediated uptake [9]. Monoclonal antibodies against CD51show that apoptotic cell phagocytosis required Rac1 andand CD61 (av and b3 chains of the vitronectin receptor)Cdc42, whereas inhibition of Rho enhanced phagocytosis.inhibited uptake. Together, these results indicate thatThe latter is consistent with suggestions that Rho inhibi-bone marrow–derived macrophages use the avb3 integrintion enhances Rac1- and Cdc42-mediated responses infor apoptotic Baf-3 cell uptake in agreement with previousmacrophages [16] and competes with Cdc42 and/or Rac1reports obtained with other types of apoptotic cells [6, 10].in neurite outgrowth [17]. Moreover, our results show thatthe involvement of Rac1 in apoptotic cell uptake has beenRho GTPases have been widely implicated in the regula-conserved through evolution as its homolog in nematodetion of the actin cytoskeleton [3] and recently in theCaenorhabditis elegans is required for engulfment of apo-process of FcgR- and complement-mediated phagocytosisptotic corpses [18].[7, 11–13]. Clostridium difficile toxin B, an inhibitor for
endogenous Rho GTPases [14], inhibited apoptotic cellphagocytosis in a dose-dependent manner (Figure 2C) with PI 3-kinases have been implicated in many cellular re-
sponses downstream of tyrosine kinases such as macro-an IC50 of 1.5 6 0.7 ng/ml. This was not a consequence ofa lack of binding of apoptotic cells to the macrophages phage chemotaxis [19] and FcgR-mediated phagocytosis
[20, 21]. The PI 3-kinase inhibitor LY294002 inhibited(Figure 2D). The inhibition of apoptotic cell phagocytosisby toxin B led us to investigate the identity of the Rho apoptotic cell phagocytosis in a dose-dependent manner
(Figure 2C) with an IC50 of 4.3 6 0.7 mM. The impairmentGTPases required for apoptotic cell uptake. We testedthe ability of macrophages microinjected with inhibitors of phagocytosis by LY294002 was not due to decreased
binding (Figure 2D). Similar results were obtained withof Rho, Rac1, and Cdc42 to engulf apoptotic cells. Theprocess of microinjection did not affect apoptotic cell bind- wortmannin (IC50 < 7 nM), an irreversible inhibitor of PI
Brief Communication 197
Figure 2
Rac1, Cdc42, and PI 3-kinases are required for apoptotic cell 378C with apoptotic cells. Phagocytosis is reported as percentage ofphagocytosis. untreated macrophages. Results are presented as mean 6 SEM of(A) Staining protocol for distinguishing ingested from noningested two to four independent experiments.apoptotic cells. Macrophages were cultured with apoptotic cells for (C and D) Macrophages were pretreated as indicated for 1 h with60 min and then washed to remove the unbound cells and fixed. Cells different concentrations of the Rho GTPase inhibitor C. difficile toxinwere stained as described in Methods (see supplementary material). B, the PI 3-kinase inhibitor LY294002, or were microinjected with theMacrophages had a low level of green fluorescence through detection indicated proteins (rabbit IgG alone as a control or rabbit IgG togetherof the macrophage cell surface marker F4/80 (left panel). Apoptotic with C3 transferase, N17Rac1, or N17Cdc42). Macrophages werecells were CFSE labeled and appeared intensely green (left panel). subsequently cultivated for 2 h with the apoptotic cells. PhagocytosisThe noninternalized apoptotic cells also bound the rhodamine- index (C) and binding index (D) are reported as percentage of untreatedconjugated streptavidin and therefore appeared red (right panel). cells. Microinjected macrophages (detected withAsterisks indicate internalized apoptotic cells. Arrowheads indicate aminomethylcoumarine-conjugated anti-rabbit antibody) werenoninternalized apoptotic cells. Bar 5 20 mm. compared to noninjected neighboring macrophages. Results are(B) Macrophages were incubated for 1 h with apoptotic Baf-3 cells presented as mean 6 SEM of three to seven independent experiments.in the presence of RGDS (1 mM), RGES (1 mM), or phospho-L- Results obtained with microinjected inhibitors were compared withserine (L-PS, 0.1 mM). Macrophages were preincubated for 20 min results obtained with IgG alone using Student’s t test: *, p , 0.01on ice with monoclonal anti-mouse CD51, anti-mouse CD61, or (very significant); **, p , 0.0001 (extremely significant).isotype control (75 mg/ml) antibodies and then incubated for 1 h at
3-kinases unrelated to LY294002 (data not shown). At the effects of LY294002 and toxin B on actin cup forma-tion and phosphotyrosine accumulation at sites of phago-these concentrations, the two inhibitors inactivate specifi-
cally PI 3-kinases [22] indicating that one or more isoforms cytosis. LY294002 had no effect on the formation of actincups or the accumulation of phosphotyrosine (Figure 3B).of PI 3-kinases are required for apoptotic cell uptake.In contrast, in the presence of toxin B, the apoptotic cellsthat were bound to the macrophages did not induce thePI 3-kinase can act upstream of Rac during growth factor
signaling [23]. To determine whether it also acts upstream accumulation of either F-actin or phosphotyrosine (Figure3C). Together, these data suggest that Rho GTPases areof Rac during apoptotic cell phagocytosis, we compared

198 Current Biology Vol 11 No 3
Figure 3
Inhibition of Rho GTPases but not PI 3-kinasesprevents phosphotyrosine and actinaccumulation during apoptotic cellphagocytosis.Macrophages were pretreated for 1 h withDMSO as a control (A), 100 mM LY294002(B), or 100 ng/ml toxin B (C) and thencultivated for 45 min with apoptotic cells. Thecells were costained for phosphotyrosine (leftpanel) and F-actin (middle panel). CFSE-labeled apoptotic cells are shown in the rightpanel. Arrows indicate colocalization ofphosphotyrosine foci, F-actin accumulation,and apoptotic cells. Similar settings wereused to acquire and process the imagesexcept for toxin B–treated specimens wherethe F-actin signal had to be increased to bedetected. Bar 5 20 mm.
necessary for the formation of an actin cup and the accu- difference in the duration of Rac1 and/or Cdc42 activationduring FcgR-mediated and apoptotic cell phagocytosis ismulation of phosphotyrosine while PI 3-kinases act down-
stream of these events, and thus that PI 3-kinases do not responsible for determining whether or not proinflamma-tory mediators are produced.act upstream of Rac.
Supplementary materialThe signaling pathways regulating apoptotic cell phagocy-Supplementary material including detailed methodology is available attosis and FcgR-mediated phagocytosis share a commonhttp://current-biology.com/supmat/supmatin.htm.
requirement for Rho GTPases (including the requirementfor Rac1 and Cdc42 but not Rho) and PI 3-kinases [7, 11, Acknowledgements
We are grateful to R. Garg, A. Entwistle, and M. Tumaine for expert assistance,20]. One important consequence of FcgR cross-linkingK. Aktories and F. Hofmann (Freiburg, Germany) for providing toxin B, andin macrophages is the subsequent secretion of proinflam-B. Vanhaesebroeck and S.J. Leevers for helpful comments. Y. L. was sup-
matory mediators [24]. In contrast, the phagocytosis of ported by a grant from the Association pour la Recherche sur le Cancer anda Medical Research Council fellowship.apoptotic cells by human and mouse macrophages is not
inflammatory but rather antiinflammatory [25, 26]. Simi-Referenceslarly, complement-mediated phagocytosis does not initi- 1. Ren Y, Savill J: Apoptosis: the importance of being eaten. Cell
ate a proinflammatory response [24]. It has been proposed Death Differ 1998, 5:563-568.2. Vanhaesebroeck B, Leevers SJ, Panayotou G, Waterfield MD:that the involvement of Rac1 and Cdc42 in FcgR-medi-
Phosphoinositide 3-kinases: a conserved family of signalated phagocytosis versus Rho in complement-mediated transducers. Trends Biochem Sci 1997, 22:267-272.3. Hall A: Rho GTPases and the actin cytoskeleton. Science 1998,phagocytosis could explain why the first type of phagocy-
279:509-514.tosis is associated with an inflammatory response and not4. Rodriguez-Tarduchy G, Collins M, Lopez-Rivas A: Regulation of
the second [7]. As Rac1 and Cdc42 are required for apo- apoptosis in interleukin-3-dependent hemopoietic cells byinterleukin-3 and calcium ionophores. EMBO J 1990, 9:2997-ptotic cell uptake, a similar mechanism is unlikely to3002.explain the absence of inflammatory response following 5. Leverrier Y, Thomas J, Mathieu AL, Low W, Blanquier B, Marvel J:Role of PI3-kinase in Bcl-X induction and apoptosisapoptotic cell phagocytosis. It is possible, however, that a

Brief Communication 199
inhibition mediated by IL-3 or IGF-1 in Baf-3 cells. Cell Death 28. Brown SB, Bailey K, Savill J: Actin is cleaved during constitutiveapoptosis. Biochem J 1997, 323:233-237.Differ 1999, 6:290-296.
6. Fadok VA, Savill JS, Haslett C, Bratton DL, Doherty DE, Campbell 29. McCarthy NJ, Whyte MK, Gilbert CS, Evan GI: Inhibition of Ced-3/ICE-related proteases does not prevent cell deathPA, et al.: Different populations of macrophages use either
the vitronectin receptor or the phosphatidylserine receptor induced by oncogenes, DNA damage, or the Bcl-2 homologueBak. J Cell Biol 1997, 136:215-227.to recognize and remove apoptotic cells. J. Immunol. 1992,
149:4029-4035.7. Caron E, Hall A: Identification of two distinct mechanisms of
phagocytosis controlled by different Rho GTPases. Science1998, 282:1717-1721.
8. Allen LA, Aderem A: Molecular definition of distinct cytoskeletalstructures involved in complement- and Fc receptor-mediated phagocytosis in macrophages. J Exp Med 1996,184:627-637.
9. Fadok VA, Bratton DL, Rose DM, Pearson A, Ezekewitz RA, HensonPM: A receptor for phosphatidylserine-specific clearance ofapoptotic cells. Nature 2000, 405:85-90.
10. Pradhan D, Krahling S, Williamson P, Schlegel RA: Multiplesystems for recognition of apoptotic lymphocytes bymacrophages. Mol Biol Cell 1997, 8:767-778.
11. Cox D, Chang P, Zhang Q, Reddy PG, Bokoch GM, Greenberg S:Requirements for both Rac1 and Cdc42 in membraneruffling and phagocytosis in leukocytes. J Exp Med 1997,186:1487-1494.
12. Massol P, Montcourrier P, Guillemot JC, Chavrier P: Fc receptor-mediated phagocytosis requires CDC42 and Rac1. EMBO J1998, 17:6219-6229.
13. Hackam DJ, Rotstein OD, Schreiber A, Zhang W, Grinstein S: Rhois required for the initiation of calcium signaling andphagocytosis by Fcgamma receptors in macrophages. J ExpMed 1997, 186:955-966.
14. Just I, Selzer J, Wilm M, von Eichel-Streiber C, Mann M, Aktories K:Glucosylation of Rho proteins by Clostridium difficile toxinB. Nature 1995, 375:500-503.
15. Sekine A, Fujiwara M, Narumiya S: Asparagine residue in the rhogene product is the modification site for botulinum ADP-ribosyltransferase. J Biol Chem 1989, 264:8602-8605.
16. Allen WE, Zicha D, Ridley AJ, Jones GE: A role for Cdc42 inmacrophage chemotaxis. J Cell Biol 1998, 141:1147-1157.
17. Kozma R, Sarner S, Ahmed S, Lim L: Rho family GTPases andneuronal growth cone remodelling: relationship betweenincreased complexity induced by Cdc42Hs, Rac1, andacetylcholine and collapse induced by RhoA andlysophosphatidic acid. Mol Cell Biol 1997, 17:1201-1211.
18. Reddien PW, Horvitz HR: CED-2/CrkII and CED-10/Rac controlphagocytosis and cell migration in Caenorhabditis elegans.Nat Cell Biol 2000, 2:131-136.
19. Vanhaesebroeck B, Jones GE, Allen WE, Zicha D, Hooshmand-RadR, Sawyer C, et al.: Distinct PI(3)Ks mediate mitogenicsignalling and cell migration in macrophages. Nat Cell Biol1999, 1:69-71.
20. Cox D, Tseng CC, Bjekic G, Greenberg S: A requirement forphosphatidylinositol 3-kinase in pseudopod extension. JBiol Chem 1999, 274:1240-1247.
21. Araki N, Johnson MT, Swanson JA: A role for phosphoinositide3-kinase in the completion of macropinocytosis andphagocytosis by macrophages. J Cell Biol 1996, 135:1249-1260.
22. Stein RC, Waterfield MD: PI3-kinase inhibition: a target for drugdevelopment? Mol Med Today 2000, 6:347-357.
23. Leevers SJ, Vanhaesebroeck B, Waterfield MD: Signalling throughphosphoinositide 3-kinases: the lipids take centre stage.Curr Opin Cell Biol 1999, 11:219-225.
24. Aderem A, Underhill DM: Mechanisms of phagocytosis inmacrophages. Annu Rev Immunol 1999, 17:593-623.
25. Fadok VA, Bratton DL, Konowal A, Freed PW, Westcott JY, HensonPM: Macrophages that have ingested apoptotic cells in vitroinhibit proinflammatory cytokine production throughautocrine/paracrine mechanisms involving TGF-beta,PGE2, and PAF. J Clin Invest 1998, 101:890-898.
26. McDonald PP, Fadok VA, Bratton D, Henson PM: Transcriptionaland translational regulation of inflammatory mediatorproduction by endogenous TGF-beta in macrophages thathave ingested apoptotic cells. J Immunol 1999, 163:6164-6172.
27. Bursch W, Hochegger K, Torok L, Marian B, Ellinger A, HermannRS: Autophagic and apoptotic types of programmed celldeath exhibit different fates of cytoskeletal filaments. J CellSci 2000, 113:1189-1198.