research article the cholesterol-lowering effect of alisol
TRANSCRIPT

Research ArticleThe Cholesterol-Lowering Effect of Alisol Acetates Based onHMG-CoA Reductase and Its Molecular Mechanism
Fei Xu1 Hui Yu2 Cai Lu1 Jun Chen1 and Wei Gu1
1School of Pharmacy Nanjing University of Chinese Medicine Nanjing 210023 China2Jiangsu Hospital of Traditional Chinese Medicine (TCM) The First Affiliated Hospital of Nanjing University ofTraditional Chinese Medicine (TCM) Nanjing 210029 China
Correspondence should be addressed to Fei Xu 6612386qqcom
Received 20 June 2016 Revised 24 September 2016 Accepted 9 October 2016
Academic Editor Hong-Cai Shang
Copyright copy 2016 Fei Xu et al This is an open access article distributed under the Creative Commons Attribution License whichpermits unrestricted use distribution and reproduction in any medium provided the original work is properly cited
This study measured the impact of alisol B 23-acetate and alisol A 24-acetate the main active ingredients of the traditional Chinesemedicine Alismatis rhizoma on total cholesterol (TC) triglyceride (TG) high density lipoprotein-cholesterol (HDL-C) and lowdensity lipoprotein-cholesterol (LDL-C) levels of hyperlipidemic mice The binding of alisol B 23-acetate and alisol A 24-acetate tothe key enzyme involved in the metabolism of TC 3-hydroxy-3-methylglutary-coenzyme A (HMG-CoA) reductase was studiedusing the reagent kit method and the western blotting technique combined with a molecular simulation technique According tothe results alisol acetates significantly lower the TC TG and LDL-C concentrations of hyperlipidemic mice while raising HDL-Cconcentrations Alisol acetates lower HMG-CoA reductase activity in a dose-dependent fashion both in vivo and in vitro Neitherof these alisol acetates significantly lower the protein expression of HMG-CoAThis suggests that alisol acetates lower the TC levelvia inhibiting the activity of HMG-CoA reductase by its prototype drug which may exhibit an inhibition effect via directly andcompetitively binding to HMG-CoA The side chain of the alisol acetate was the steering group via molecular simulation
1 Introduction
Alismatis rhizoma is the rhizome of Alisma orientale (Sam)Juzepcz and belongs to the Alismataceae family Alismatisrhizoma is a diuretic agent of key importance whose diureticeffect is related to the season of harvest the medicinal partsthe processing method the route of administration and thespecies of the tested organism Moreover the diuretic effectof genuine Alismatis rhizoma is strongest when collectedin winter while spring collection results in slightly reducedeffect In addition to its salt solution other processedproducts have an evident diuretic effect Furthermore anethanol extract of Alismatis rhizoma and its triterpenespossess diuretic functions that reduce urinary protein Thetriterpenes in Alismatis rhizoma are alisol A Alisol B alisol B23-acetate alisol A 24-acetate and so on [1 2] AdditionallyAlismatis rhizoma is a lipid-regulating Chinese traditionalmedicine that is commonly used as treatment for hyper-lipidemia Studies have shown that its main lipid-regulating
active ingredients are alisol acetates predominately alisol B23-acetate and alisol A 24-acetate The clinical application ofAlismatis rhizome has been limited by a lack of molecularstudies of its molecular interaction mechanism [3ndash9] Thestudies described in this paper investigated the differenteffects of alisol A 24-acetate and alisol B 23-acetate on totalcholesterol (TC) triglyceride (TG) high density lipoprotein-cholesterol (HDL-C) and low density lipoprotein-cholesterol(LDL-C) of hyperlipidemic mice and obtained the lipid-regulating macroscopic rule of alisol acetates In this studywe investigated the mechanism of alisol acetates on loweringTC level firstly and the regulatory mechanisms of alisolacetates on TG HDL-C and LDL-C levels would bethe future research content of our group The mainenzyme involved in the metabolism of TC is 3-hydroxy-3-methylglutaryl-coenzyme A (HMG-CoA) reductase Thisenzyme facilitates cholesterol synthesis from the beginningby catalyzing the synthesis ofmevalonate (MVA) fromHMG-CoA further generating TC via squalene The decreased
Hindawi Publishing CorporationEvidence-Based Complementary and Alternative MedicineVolume 2016 Article ID 4753852 11 pageshttpdxdoiorg10115520164753852
2 Evidence-Based Complementary and Alternative Medicine
O
Me
MeMe
Me
Me
Me
MeMe
HO
H
H
H
OAc
O
(a)
MeMe
Me
Me Me
Me
MeMe
HO
OAc
O
OH OH
H
H
(b)
Figure 1 Molecular structures of alisol ((a) alisol B 23-acetate (b) alisol A 24-acetate)
HMG-CoA reductase activity can effectively reduce TCgeneration [10ndash13] Using the reagent kit method HMG-CoA reductase activity was measured in vivo and in vitrousing liver homogenates of hyperlipidemic mice before andafter the addition of alisol acetates The results indicatethat alisol acetates might lower TC via inhibition of HMG-CoA reductase activity and that the prototype drug ofalisol acetate inhibits HMG-CoA activity instead of its invivo metabolites Western blotting was used to measure theeffects of alisol acetates on the HMG-CoA reductase proteinexpression of hyperlipidemic mice The results showed thatalisol acetatesmight not inhibit HMG-CoA reductase activityvia downregulation of its protein expression Instead itmight inhibit the HMG-CoA reductase effect by directly andcompetitively binding with it The binding interaction ofalisol acetates and HMG-CoA reductase was studied usinga molecular simulation technique This technique yieldedthe following parameters the binding constant bindingenergy hydrogen bonding hydrophobichydrophilic groupselectrostatic energy and van derWaals forcesThe interactionmodel of alisol acetate and HMG-CoA reductase was builtThese experimental results were compared with the pharma-cological results in order to determine the steering groupsof this type of compound and the key amino acid residuesof the enzyme This could shed light on the cholesterol-lowering mechanism of alisol acetates at the molecular levelThe results were invaluable for promoting the applications ofAlismatis rhizoma in the clinical practice
2 Materials and Methods
21 Reagents and Instruments Themain reagents were as fol-lows simvastatin (MSD Pharmaceutical Co Ltd HangzhouChina) cholesterol (National Group Chemical Reagent CoLtd Beijing China) sodium deoxycholate (National GroupChemical Reagent Co Ltd Beijing China) propylthiouraciltablets (Jinghua Pharmaceutical Co Ltd Nantong China)lard (refined from the leaf lard purchased at the localmarket)polysorbate 80 (Tween 80 Qiangshun Chemical ReagentCo Ltd Shanghai China) propylene glycol (QiangshunChemical Reagent Co Ltd Shanghai China) basal diet (Slac
Laboratory Animal Co Ltd Shanghai China) radioim-munoprecipitation assay (RIPA) tissue cell lysis buffer (Solar-bio Beijing China cat number R0010) bicinchoninic acid(BCA) protein quantitative reagent kit (Thermo MA US catnumber PICPI23223) alisol A 24-acetate standard (MansiteBiological Technology Co Ltd Chengdu China batch num-ber MUST-15022104 98 purity) and alisol B 23-acetatestandard (Mansite Biological Technology Co Ltd ChengduChina batch number MUST-149091210 with 98 purity)The structures of the alisol acetates are shown in Figure 1[2 14]
Further reagents were as follows mice TG enzyme-linked immunosorbent assay (ELISA) reagent kits (Fengx-iang Biotechnology Co Ltd Shanghai China batch num-ber 201207) mice HDL-C ELISA reagent kits (FengxiangBiotechnology Co Ltd Shanghai China batch number201302) mice LDL-C ELISA reagent kits (Fengxiang Biotech-nology Co Ltd Shanghai China batch number 201301)mice TC ELISA reagent kits (Fengxiang Biotechnology CoLtd Shanghai China batch number 201202) HMG-CoAreductase activity assay kits (Genmed Scientifics Inc Shang-hai China batch number 20080717) HMG-CoA reductaseantibodies (Ray Biotech Inc Atlanta US batch number168-10733) internal standard (glyceraldehyde-3-phosphatedehydrogenase [GAPDH]) protein antibody (CST BostonUS batch number 5174) and nitrocellulose membrane (Mil-lipore Boston US batch number HATF00010) All otherchemicals used were of analytical grade and were obtainedfrom China
The following instruments were employed in this studya high-speed ben centrifuge (Medical Analytical InstrumentFactory Shanghai China H1850) an electronic analyticalbalance (Sunny Hengping Scientific Instrument Co LtdShanghai China FA1104) pipettes (Jiarsquoan Analytical Instru-ment Factory Shanghai China WKYIII) a UV-Vis spec-trophotometer (Persee General Instrument Co Ltd BeijingChina TU-1901) a precision digital high-speed homoge-nizer (Shuoguang Electronic Technology Co Ltd ShanghaiChina SG-3048A) an electrophoresis apparatus (BIO-RADCalifornia US mini protean 3 cell) an electroporator (Jing-mai Technology Co Ltd Dalian China PS-9) a microplate
Evidence-Based Complementary and Alternative Medicine 3
Table 1 Blood lipid levels of each group of mice (119909 plusmn 119904 119899 = 11)
Group Animal count TC (ngmL) TG (ngmL) HDL-C (ngmL) LDL-C (ngmL)Blank 11 894 plusmn 137 951 plusmn 152 6893 plusmn 791 1422 plusmn 457Model 11 2905 plusmn 149998771998771 3059 plusmn 302998771998771 1786 plusmn 559998771998771 5171 plusmn 317998771998771
Positive 11 977 plusmn 144lowastlowast 1098 plusmn 123lowastlowast 4511 plusmn 572lowastlowast 2111 plusmn 637lowastlowast
23B high 11 1450 plusmn 191lowastlowast 1408 plusmn 197lowastlowast 4623 plusmn 848lowastlowast 2639 plusmn 401lowastlowast
23B medium 11 2310 plusmn 404lowastlowast 2300 plusmn 167lowastlowast 3756 plusmn 661lowastlowast 3357 plusmn 372lowastlowast
23B low 11 2430 plusmn 218lowastlowast 2456 plusmn 418lowastlowast 2908 plusmn 781lowastlowast 3610 plusmn 463lowastlowast
24A high 11 1870 plusmn 244lowastlowast 1744 plusmn 224lowastlowast 4289 plusmn 543lowastlowast 2923 plusmn 633lowastlowast
24A medium 11 2527 plusmn 531lowast 2679 plusmn 434lowast 2813 plusmn 553lowastlowast 3692 plusmn 529lowastlowast
24A low 11 2757 plusmn 168lowast 2715 plusmn 324lowast 2698 plusmn 377lowastlowast 3857 plusmn 591lowastlowast
Note 24A alisol A 24-acetate 23B alisol B 23-acetate compared to the blank group 998771998771119875 lt 001 and 998771119875 lt 005 compared to the model group lowastlowast119875 lt 001and lowast119875 lt 005
reader (Labsystems Multiskan MS Finland Model 352) awater bath (Leica Germany HI1210) and an imaging system(Tanon Co Ltd Shanghai China Tanon-5200)
22 Animals Male clean-grade ICR mice (18ndash22 g) werepurchased from the Animal Core Facility Nanjing MedicalUniversity (license number SCXK (Su)-2013-0005) Theanimals were placed in a clean-grade animal room at 20ndash24∘C and 55 plusmn 5 humidity The light and dark cycles weretwelve hours eachThe mice were free-fed for one week priorto the experimentsTheAnimal Ethics Committee of NanjingUniversity of Chinese Medicine approved all protocols ofanimal experiments
23 Measurements of TC TG HDL-C and LDL-C Levels
231 The Induction of an Animal Model of Hyperlipidemia[15ndash19] A total of 99 male mice were randomly dividedinto nine groups according to their weight Mice in onegroup the normal blank group were intragastrically feddistilled water (10mLkg) every morning Mice in the othergroupwere intragastrically fed lipid emulsion (10mLkg)Theformulation of the lipid emulsion was as follows 25 lard15 cholesterol 10 polysorbate 80 30 propylene glycol2 propylthiouracil 2 sodium cholate and 1 sugar Forfive consecutive weeks the mice were free-fed a basal dietalongside the intragastric administration of distilled water orlipid emulsion The mice were fasted for twelve hours (waterwas permitted) after the last intragastric administrationBlood samples of mice were collected and their serums wereseparated The TG TC HDL-C and LDL-C concentrationsin serums were measured according to the instructions of thecorresponding reagent kits making it possible to comparethe blood lipid levels of the model and the blank groupsAccording to the results in Table 1 the TG TC and LDL-Cof themodel group increasedwhile theHDL-C droppedThissuggests successful establishment of the hyperlipidemiamod-elsThemodel animals were evenly divided into eight groupsmodel simvastatin alisol B 23-acetate (high medium andlow dosage) and alisol A 24-acetate (high medium and lowdosage)
232 Drug Supply to Hyperlipidemic Mice [19] The appro-priate quantities of alisol A 24-acetate alisol B 23-acetateand simvastatin were weighed dissolved in 03 sodium car-boxymethylcellulose (CMC-Na) solution and diluted to26mL
The quantities administered intragastrically to eachgroup were as follows positive control group (simvastatin7mgkgday) alisol A 24-acetate low-dose group (064mgkgday) alisol A 24-acetate medium-dose group (128mgkgday) alisol A 24-acetate high-dose group (256mgkgday)alisol B 23-acetate low-dose group (064mgkgday) alisol B23-acetatemedium-dose group (128mgkgday) and alisol B23-acetate high-dose group (256mgkgday)
Each group was intragastrically fed 10mLkg of lipidemulsion every morning and treated with the correspondingdrugs every afternoon for three weeks
233 Measurements of TC TG HDL-C and LDL-C LevelsThemice were fasted for twelve hours (water was permitted)following the last dose of drug Blood samples were collectedand left to sit for 20ndash30minutesThe serumswere obtained bycentrifuging the blood samples at 3000 rmin for 15 minutesThe collected serums were measured in accordance with theinstruction of the TC TG HDL-C and LDL-C reagent kits
24 Measurement of In VivoIn Vitro HMG-CoAReductase Activity [19ndash25]
241 Measurement of In Vitro HMG-CoA Reductase ActivityThe establishment of themousemodel of hyperlipidemia wasdone according to Section 231 After the last intragastricadministration (procedure described in Section 231) themice were fasted for twelve hours (water was permitted)and then decapitated Immediately after decapitation theperitoneal cavities of the mice were rinsed with 10mL of coldsaline while the livers of the mice were rapidly frozen inliquid nitrogen and stored at minus70∘C for further analysis
At a temperature of 4∘C 05 g of each liver was weighedplaced in phosphate buffered saline (PBS pH 72 025 gmL)and homogenized Subsequently themixturewas centrifugedfor tenminutes at 4000 rmin and at a temperature of 4∘CThe
4 Evidence-Based Complementary and Alternative Medicine
supernatant was then collected On three separate occasions1mL of the supernatant of the liver homogenate was collectedand extracted with 10mL of 95 ethanol at 60∘C This wasfollowed by vacuum filtration The residues were dissolvedin PBS The samples were adjusted to a certain concentrationand stored at minus70∘C
We added 004mgmL of simvastatin 016mgmL008mgmL and 004mgmL (high medium and lowdosage) of alisol A 24-acetate and alisol B 23-acetate to 5mLof liver homogenate samples respectively After leaving thesamples to stand for 24 hours theHMG-CoA reductase activ-ity was measured using the HMG-CoA reductase activityassay kit in accordance with the instructions
242 Measurement of In Vivo HMG-CoA Reductase ActivityThe induction of the animal model of hyperlipidemia wasdone according to Section 231 The intragastric adminis-tration was performed as described in Section 232 Themice were fasted for twelve hours (water was permitted) anddecapitated after the last dose of the drug The peritonealcavities of the mice were immediately rinsed with 10mL ofcold saline whereas the livers of the mice were rapidly frozenin liquid nitrogen and then stored for further analysis at atemperature of minus70∘C At 4∘C 05 g of each liver were col-lected placed in PBS (pH 72 025 gmL) and homogenizedThe sample was subsequently centrifuged for ten minutes at4000 rmin and at a temperature of 4∘CThe supernatant wasthen collected For each mouse 1mL of liver homogenatesupernatant was extracted with 10mL of 95 ethanol at 60∘Con three occasions This was followed by vacuum filtrationThe residues were dissolved in PBSThe solutionwas adjustedto a certain concentration and stored at minus70∘C The HMG-CoA reductase activity of the liver homogenate samples wasmeasured in accordance with the instructions given in theHMG-CoA reductase activity assay kit
25 Measurement of HMG-CoA Reductase Protein Expres-sion Level via Western Blotting [26ndash29] The hyperlipidemicinduction of animal model of hyperlipidemia was builtaccording to Section 231 The intragastric administrationfollowed the procedure described in Section 232 whereasthe liver homogenate samples were prepared as described inSection 242 The samples were stored at minus70∘C for furtheruse
In order to detect the protein concentration of the super-natant of each sample the A
595 nm absorbance value wasmea-sured via the Bradford method and the calibration curve ofthe bovine serum albumin protein was plotted An equivalentvolume of 2 times sodium dodecyl sulfate (SDS) buffer was addedto the liver homogenate sampleThemixture was denaturizedat 100∘C for three minutes Subsequently the protein was iso-lated using SDS-polyacrylamide gel electrophoresis (PAGE)and electrotransferred to the nitrocellulose membrane Thenitrocellulose membrane was removed and blocked betweenone and two hours in the tris-buffered saline with Tween 20(TBST) solution with 5 skim-milk powder Subsequentlythe membrane was washed with TBST and placed intothe primary antibody (1 2500 rabbit anti-rat HMG-CoAreductase polyclonal antibody) at 4∘C overnight The next
day themembranewas washedwith TBST and blocked in theTBST solution with 5 skim-milk powder for two hoursThemembrane was washed thrice with TBST for 10ndash20 minutesper wash The membrane was incubated for one hour withalkaline phosphatase-conjugated mouse anti-rabbit IgG at adilution of 1 500 Following incubation in the secondaryantibody the membrane was washed again and visualizedusing electrochemiluminescence (ECL) The membrane wasplaced in the antibody elution buffer for ten minutes andrinsed in TBST solution for five minutes It was then blockedin the TBST solution with 5 skim-milk powder and shakenat room temperature for 30 minutes The membrane wasrinsed with TBST repeatedly and then incubated with pri-mary antibody of internal standard GAPDH at a dilution of1 1000 The membrane was subsequently washed and incu-bated with secondary antibody at a dilution of 1 250 Follow-ing washing the membrane was visualized The gray valuesof the filmstrips were measured using a gel imaging ana-lysis system The calculated gray level ratio (HMG-CoAreductaseGAPDH) represents the relative amount of HMG-CoA reductase protein expression
26 Investigation of the Interaction between Alisol Acetatesand HMG-CoA Reductase [30ndash35] A molecular simulationwas conducted The initial molecular structures of the alisolacetates were generated using Discovery Studio 21 (DS21Accelrys US) The structure of HMG-CoA reductase wasobtained from the crystal structure in the protein data bank(PDB) database (PDB code 1HW9) HMG-CoA reductaseis a glycoprotein with N-linked high-mannose oligosaccha-rides consisting of four identical subunits with a molecularweight of 97 ku Each subunit comprised 888 amino acidresidues with the phosphorylation site located at the 872ndamino acid Each subunit featured three different domainsthe N-terminal 339 (1ndash339) amino acids composed the trans-membrane domain the 110 (340ndash449) amino acids composedthe junction domain and the C-terminal 439 (450ndash888)amino acids composed the catalytic domainThe N-terminalcrossed the endoplasmic reticulum membrane eight timesand anchored to the membrane through short rings Thetransmembrane domain was composed of 167 amino acidresidues As the four subunits were identical according to itsinteraction with the drug the A chain was selected as thereceptor for docking simulation Following the removal ofwatermolecules heteroatoms and themulticonformations ofthe amino acid residues in the crystal structure flexible dock-ing in theChemistry atHARvardMacromolecularMechanics(CHARMM) force field was performed Subsequently to thesolvation calculation the compensation ions Na+ and Clminuswere added to the system in order to simulate the environ-ment in the human bodyThemost likely confirmation of theresulting complex was confirmed via minimization calcula-tion
261 Molecular Simulation of Alisol B 23-Acetate and HMG-CoA Reductase The alisol B 23-acetate was defined as aligand After docking the LibDock score the CDockerenergy the CDocker interaction energy and the hydro-gen bond formation of the ligand-receptor complex were
Evidence-Based Complementary and Alternative Medicine 5
comprehensively considered so as to determine the finalsteady conformation This conformation was selected forfurther molecular dynamic simulation
262 Molecular Simulation of Alisol A 24-Acetate andHMG-CoA Reductase Applying the method of Section 261resulted in the final steady conformation of alisol A 24-acetateThis conformationwas selected for furthermoleculardynamic simulation
27 Data Analysis The experimental data were analyzedusing one-way analysis of variance (ANOVA) in SPSS version150 (SPSS Inc Chicago USA) 119875 values of less than 005(119875 lt 005) indicated statistically significant differences Theresults were expressed in the form of mean (119909) plusmn standarddeviation (SD)
3 Results
31 Measurements of TC TG HDL-C and LDL-C LevelsThe blood lipid levels of the blank group positive groupmodel group alisol B 23-acetate high- medium- and low-dose groups and alisol A 24-acetate high- medium- andlow-dosage groups are shown in Table 1 This study revealedthat the TC TG LDL-C and HDL-C levels of the alisol A24-acetate high- medium- and low-dose groups as well asthe alisol B 23-acetate high- medium- and low-dose groupssignificantly differed from those of the model group Thissuggests that alisol acetates could have a lipid-regulatingeffect and significantly decrease the TC TG and LDL-Cconcentrations of hyperlipidemic mice while increasing theHDL-C concentrations
32 Measurement of In VivoIn Vitro HMG-CoAReductase Activity
321 Measurement of In Vitro HMG-CoA Reductase ActivityAs Table 2 indicates alisol acetates lower the HMG-CoAreductase activity dose dependently The extent to whichalisol B 23-acetate lowered HMG-CoA reductase activityexceeded the extent to which alisol A 24-acetate lowered theactivity of this enzyme
322 Measurement of In Vivo HMG-CoA Reductase ActivityThe results in Table 3 illustrate that alisol acetates lower thein vivo HMG-CoA reductase activity in a dose-dependentfashion with a greater degree of enzyme inhibition for alisolB 23-acetate than for alisol A 24-acetate
33 Measurement of HMG-CoA Reductase Protein Expres-sion Level via Western Blotting The western blotting resultssuggest that alisol acetates did not significantly decrease theprotein expression of HMG-CoA reductase (Figure 2)
34 Investigation of the Interaction between the Alisol Acetatesand HMG-CoA Reductase
341 Molecular Simulation Results for Alisol B 23-Acetate andHMG-CoA Reductase The molecular simulation results forthe interaction between alisol B 23-acetate and HMG-CoA
Table 2 The activity of HMG-CoA reductase in vitro (119909 plusmn s 119899 =11)
Group HMG-CoA reductase (UL)Blank 13467 plusmn 3991Positive 16429 plusmn 2832lowastlowast
Model 49788 plusmn 2881998771998771
24A high 19042 plusmn 3092lowastlowast
24A medium 25079 plusmn 1635lowastlowast
24A low 33515 plusmn 2332lowastlowast
23B high 17912 plusmn 4159lowastlowast
23B medium 23079 plusmn 1664lowastlowast
23B low 32759 plusmn 1915lowastlowast
Note 24A alisol A 24-acetate and 23B alisol B 23-acetate compared to theblank group 998771998771119875 lt 001 and 998771119875 lt 005 compared to the model grouplowastlowast119875 lt 001 and lowast119875 lt 005
Table 3The activity of HMG-CoA reductase in vivo (119909 plusmn s 119899 = 11)
Group HMG-CoA reductase (UL)Blank 12345 plusmn 2534Positive 20021 plusmn 3104lowastlowast
Model 49012 plusmn 4117998771998771
24A high 280123 plusmn 2915lowastlowast
24A medium 34554 plusmn 3234lowastlowast
24A low 40131 plusmn 4221lowastlowast
23B high 25203 plusmn 2931lowastlowast
23B medium 29614 plusmn 3218lowastlowast
23B low 39082 plusmn 2323lowastlowast
Note 24A alisol A 24-acetate and 23B alisol B 23-acetate compared to theblank group 998771998771119875 lt 001 and 998771119875 lt 005 compared to the model grouplowastlowast119875 lt 001 and lowast119875 lt 005
reductase are depicted in Figure 3 Figure 3(a) illustratesthe overall pattern of the interaction The interaction pocketwhere alisol B 23-acetate bound to HMG-CoA reductasecontained the following amino acids Leu521 Val522Met523Gly524 Ala525 Cys527 Glu528 Asn529 Val530 Met588Thy589 Arg590 Gly591 Asp653 Ala654 Met655 Gly656Met657 Asn658 Met659 Ile660 Ser661 Lys662 Gly663Thy664 Glu665 Cys688 Asp690 Lys691 Lys692 Arg702Lys704 Ile760 Try761 Ile762Ala763 Cys764Glu801 Ile802Gly803 Thr804 Val805 Gly806 Gly807 Gly808 Thr809Asn810 and Leu811Theywere located in the catalytic domainconstituted by the 439 (450ndash888) amino acids at the C-terminal After entering the interaction pocket alisol B 23-acetate interacted with the peripheral amino acid residues viahydrogen bonding and hydrophobic and hydrophilic effectsThe hydrogen bond graph in Figure 3(b) shows two hydrogenbonds between alisol B 23-acetate and the amino acids Lys691and Asp767 Table 4 presents detailed information aboutthe hydrogen bonding The interaction energy between thehydrophobic amino acid residues and the small molecule wascalculated to be minus128 KJmol while the interaction energybetween hydrophilic amino acid residues and the smallmolecule was calculated to be minus1353 KJmol This indicatesthat the hydrophilic interaction between the small moleculeand the peripheral amino acids was stronger
6 Evidence-Based Complementary and Alternative Medicine
HMG-CoAreductase
GAPDH
05
04
03
02
01
00HM
G-C
oA re
duct
ase
GA
PDH
Blan
k
Mod
el
Posit
ive
24
A-H
24
A-M
24
A-L
23
B-H
23
B-M
23
B-L
Blan
k
Mod
el
Posit
ive
24
A-H
24
A-M
24
A-L
23
B-H
23
B-M
23
B-L
lowast
Figure 2 Results of the protein expression of HMG-CoA reductasein mice liver tissue protein (119899 = 11) Note compared to the blankgroup 998771998771119875 lt 001 and 998771119875 lt 005 compared to the model grouplowastlowast119875 lt 001 and lowast119875 lt 005 24A-H alisol A 24-acetate high 24A-
M alisol A 24-acetate medium and 24A-L alisol A 24-acetate low23B-H alisol B 23-acetate high 23B-M alisol B 23-acetate mediumand 23B-L alisol B 24-acetate low
Table 4 The length (gt) and the angle of hydrogen bond formedbetween alisol acetates and HMG-CoA reductase
Name XndashHsdot sdot sdotYd
(XndashH)
d(Hsdot sdot sdotY)
d(Xsdot sdot sdotY) Angle XHY
Alisol B
23-acetate
Lys691 NZ-HZ1sdot sdot sdotO37 104 237 298 11594
Asp767OD2-
HH88sdot sdot sdotO2096 240 324 14490
Alisol A24-acetate
Asn658 OD1-H5Zsdot sdot sdotO5 095 201 286 14897
After docking the conformation of alisol B 23-acetatechanged as shown in Figure 3(c) The calculated root-mean-square deviation (RMSD) was 289gt The conformationof the small molecule changed from the clustered sheetstructure to the open structure Before docking the sidechain and the ring structure of the small molecule foldedas shown in Figures 3(d) and 3(e) The angle between C
2ndash
C10ndashC19was 1095∘ 1095∘ between C
10ndashC19ndashC20 and minus599∘
between C2ndashC10ndashC19ndashC20 As shown in Figures 3(f) and
3(g) after docking the angle between C2ndashC10ndashC19was 1136∘
1156∘ between C10ndashC19ndashC20 and 1260∘ between C
2ndashC10ndash
C19ndashC20 The angle between the parent ring and the side
chain was significantly increasedThe twisting of the dihedralangles resulted in the opening of side chains and insertioninto the protein which led to interactions There was ahydrophilic ether bond on the side chain and the peripheralamino acids were mainly hydrophilic (Figures 3(h) and 3(i))
This created a hydrophilic environment around the sidechains culminating in stable binding The energy of thevan der Waals forces between them was calculated to beminus1519 KJmol whereas the electrostatic energy was calcu-lated to be minus2366 KJmol As the electrostatic energy waslower than the energy of the van derWaals forces between thesystems the electrostatic interactionwas themain interactionbetween this small molecule and the macromolecule Thebinding energy (ΔG∘) between the alisol B 23-acetate and theHMG-CoA reductase was calculated to be minus1033 KJmol
342 Molecular Simulation Results of Alisol A 24-Acetateand HMG-CoA Reductase Themolecular simulation resultsfor the interaction between alisol A 24-acetate and HMG-CoA reductase are shown in Figure 4 Figure 4(a) shows theoverall interaction pattern of this compound and HMG-CoAreductase The interaction pocket where it bound to HMG-CoA reductase contained the following amino acids Tyr519Ser520 Leu521 Val522 Met523 Gly524 Ala525 Cys527Glu528 Asn529 Val530 Ile531 Gly532 Try533 Met588Thy589 Arg590 Gly591 Asp653 Ala654 Met655 Gly656Met657 Asn658 Met659 Ile660 Ser661 Lys662 Gly663Thy664 Glu665 Cys688 The689 Asp690 Lys691 Lys692Cys764 Gly765 Gln766 Asp767 Ala768 Ala769 Gln770Asn771 Val772 Gly773 Ser774 Glu801 Ile802 Gly803Thr804Val805Gly806Gly807Gly808Thr809 andAsn810There were 439 (450ndash888) amino acids in the catalyticdomain at the C-terminal After entering the interactionpocket alisol A 24-acetate exhibited hydrogen bonding andhydrophobic and hydrophilic interactions with peripheralamino acid residues Figure 4(b) shows the hydrogen bondcontact map There was one hydrogen bond between alisol A24 acetate and amino acid Asn658 (Table 4) The interactionenergy of the hydrophobic amino acid residues and thesmall molecule was calculated to be 465 KJmol whereas theinteraction energy of the hydrophilic amino acid residues andthe small molecule was calculated to be minus1023 KJmol Thissuggests that the hydrophilic groups had a much strongereffect than the hydrophobic groups The small molecule andthe peripheral amino acids exhibited a stronger hydrophilicinteraction as indicated by the interface graphs (Figures 4(f)and 4(g))
After docking the alisol A 24-acetate conformationchanged (Figure 4(c)) The RMSD was calculated to be334gt The conformation angle C
3ndashC18ndashC26ndashC28
changedfrom 600∘ before docking to 1242∘ after docking (Figures4(d) and 4(e)) This suggests a large torsion of the smallmolecule side chain after docking whereby the stretchedopen state changed to a ring-like sheet state Consequentlyit was difficult to insert the side chain into the protein andinteract with the active pocket of the protein This resultedin considerably lower binding relative to alisol B 23-acetateThe electrostatic energy of alisol A 24-acetate and HMG-CoA reductase was determined to be minus886 KJmol whilethe energy of the van der Waals forces was determined tobe minus1649 KJmol The energy of the van der Waals forceswas lower than the electrostatic energy showing that themain interaction between this small molecule and themacro-molecule was the van derWaals interactionThe ΔG∘ of alisol
Evidence-Based Complementary and Alternative Medicine 7
(a) (b) (c)
(d) (e) (f)
(g) (h) (i)
Figure 3 Interaction between alisol B 23-acetate and HMG-CoA reductase (a) Overall pattern of interaction (alisol B 23-acetate is denotedby a bar chart) (b) Hydrogen bond graph of alisol B 23-acetate with Lys691 and Asp767 (the hydrogen bond is denoted by a dotted line) (c)Stacking chart of alisol B 23-acetate before and after docking (the initial structure is shown in green and the structure after docking is shown inpurple) (d) Initial angle of alisol B 23-acetate C
2ndashC10ndashC19and C
10ndashC19ndashC20 (e) Initial dihedral angle of alisol B 23-acetate C
2ndashC10ndashC19ndashC20
(f) The angle of alisol B 23-acetate C2ndashC10ndashC19and C
10ndashC19ndashC20after stacking (g)The dihedral angle of alisol B 23-acetate C
2ndashC10ndashC19ndashC20
after stacking (h) Hydrophobic interface of the alisol B 23-acetate side chain and the peripheral amino acid residues after stacking (alisol B23-acetate is denoted by yellow bars) (i) Hydrophilic interface of the alisol B 23-acetate side chain and the peripheral amino acid residuesafter stacking (alisol B 23-acetate is denoted by yellow bars)
A 24-acetate and HMG-CoA reductase was calculated to beminus23 KJmol
4 Discussion
Various experimental and clinical studies focus on the bloodlipid reducing effects of different solvent extracts (especially
water and alcohol extracts) fromAlisma orientale and its pro-cessed products [36ndash38] However the mechanisms under-lying the effects of Alisma orientale constituents remain to berevealed Alisma orientalemay interfere with exogenous cho-lesterol absorption and endogenous cholesterol metabolismthus reducing the level of TC Moreover alisol acetates mayinterfere with themetabolism of endogenous cholesterol [39]
8 Evidence-Based Complementary and Alternative Medicine
(a) (b) (c)
(d) (e) (f)
(g)
Figure 4 The interaction between alisol A 24-acetate and HMG-CoA reductase (a) Overall pattern of interaction (alisol A 24-acetate isdenoted by a bar chart) (b) Hydrogen bond graph of alisol A 24-acetate and Asn658 (the hydrogen bond is denoted by a dotted line) (c)Stacking chart of alisol A 24-acetate before and after docking (the initial structure is shown in green and the structure after docking is shownin purple) (d) The initial dihedral angle of alisol A 24-acetate angle C
3ndashC18ndashC26ndashC28 (e) The dihedral angle of alisol A 24-acetate angle
C3ndashC18ndashC26ndashC28after stacking (f) Hydrophobic interface of the alisol A 24-acetate side chain and the peripheral amino acid residues after
stacking (alisol A 24-acetate is denoted by yellow bars) (g) Hydrophilic interface of the alisol A 24-acetate side chain and the peripheralamino acid residues after stacking (alisol A 24-acetate is denoted by yellow bars)
Therefore we studied the TC-lowering effect of alisolacetates based on HMG-CoA reductase and the underly-ing molecular mechanism As the main source of endoge-nous cholesterol and one of the key enzymes in the TCmetabolism HMG-CoA reductase is the first described rate-limiting enzyme in cholesterol synthesis in vivo Moreover its
activation directly affects the speed of cholesterol synthesisand the level of TC in vivo
The TC-lowering effect and molecular mechanism ofalisol acetates the main active ingredient of Alismatis rhi-zoma were investigated using the reagent kit method andwestern blotting technique combined with the molecular
Evidence-Based Complementary and Alternative Medicine 9
simulation technique An interaction model was built foralisol acetate and the key enzyme involved in the metabolismof TC was HMG-CoA reductase The present study revealsthat alisol acetates can significantly lower the TC level ofhyperlipidemic miceThe alisol B 23-acetate showed a higherextent of TC lowering than alisol A 24-acetate
Consistent with the in vitro results alisol acetates couldalso lower the in vivoHMG-CoA reductase activity in a dose-dependent fashion with a higher degree of enzyme inhibitionfor alisol B 23-acetate compared to alisol A 24-acetate Theseresults suggest that alisol acetates might lower TC levels viaregulation of HMG-CoA reductase activity The consistencybetween the results of the in vivo and in vitro experimentsindicates that the alisol acetate prototype drug inhibitsHMG-CoA reductase activity instead of the in vivometabolites
Revealed by the western blotting results alisol acetatesmight inhibit the activity of HMG-CoA reductase by directlyand competitively binding to HMG-CoA reductase ratherthan by downregulating its protein expression
The molecular simulation technique was employed inorder to investigate the interaction of alisol acetates andHMG-CoA reductase The results (Figures 3 and 4 andTable 4) revealed themolecularmechanism of the interactionof alisol acetates and HMG-CoA reductase Both alisolacetates bound to HMG-CoA reductase in their catalyticdomain at the C-terminalThe alisol B 23-acetate formed twohydrogen bonds with HMG-CoA reductase The hydrophilicinteraction was stronger than the hydrophobic interactionwhereas the electrostatic interaction was stronger than thevan der Waals forces After the interaction the structure ofthe small molecule dramatically changed from the originalfolded sheet to an open structure Additionally the side chainstretched and was inserted into the protein The peripheralamino acids formed a hydrophilic environment and stablybound with it In general a hydrogen bond with a lengthof less than 35 A and an angle of 90ndash180∘ is consideredto be strong These two hydrogen bonds were relativelystrong and might be partly responsible for the torsion ofthe small molecule The alisol A 24-acetate formed onehydrogen bond with the HMG-CoA reductase and this bondwas weaker compared to those of alisol B 23-acetate Thehydrophilic interaction was stronger than the hydrophobicinteraction and the van derWaals force was stronger than theelectrostatic interaction After the interaction the moleculeside chain had large torsion and changed from the originalstretched open state to a ring-like sheet state which wasdifficult to insert into the proteinThe binding energy of alisolB 23-acetate andHMG-CoA reductase was lower than that ofalisol A 24-acetate and HMG-CoA reductase indicating thatthe binding of alisol B 23-acetate and HMG-CoA reductasewas stable According to the pharmacological results alisolB 23-acetate lowered the HMG-CoA reductase activity morethan alisol A 24-acetate This suggests that the active centerwas the interaction pocket constituted by these amino acidsand that the hydrogen bonds involving Lys691 Asp767 andAsn658 were the key amino acid residues for the interactionof this protein with alisol acetates
The binding of alisol B 23-acetate and HMG-CoA reduc-tase was considerably stronger than the binding of alisol
A 24-acetate and HMG-CoA reductase The relatively weakbinding of alisol A 24-acetate and HMG-CoA reductasemight be due to its distinct side-chain structure The sidechain of alisol B 23-acetate contained an ether bond whichwas strongly hydrophilic and could be inserted into thehydrophilic environment of the protein to form strong inter-actions Previous studies found that the side chains of alisolacetates were their active groups [40] In the present studythis conclusionwas validated A folded side chainparent ringbound more strongly to the macromolecule than an openside-chainparent ring did Thus the side chain acts as thesteering group in that it steers the interactions between alisolacetates and macromolecules This might be the key groupfor interactions with macromolecules as it had a decisiveeffect on the interactions of this type of compound withmacromolecules Our previous studies indicated that alisol B23-acetate was partially converted to alisol A 24-acetate in theenvironment of the human stomach However the bindingof alisol A 24-acetate and HMG-CoA reductase was weaker[14 41 42] Therefore it is advisable for drug developers todevelop a dosage form that circumvents the conversion ofalisol B 23-acetate in the stomach
Previously published studies indicate that [43 44] themechanism of reducing the level of TC is that two acetylCoA molecules condense to acetyl CoA under the catalysisof acetoacetyl-coenzyme A thiolase (AACT) acetyl CoAand acetyl CoA condense to HMG-CoA and HMG-CoAreduces to MVA under the catalytic action of HMG-CoAreductase MVA further generates TC with squalene Thehigh intracellular free fatty acid (FC) level stimulates micro-somal acyl coenzyme A-cholesterol acyltransferase (ACAT)and promotes esterification of the cholesterol Subsequentlycholesterol is stored in the form of cholesterol esters (CE)which is a feedback inhibition of HMG-CoA reductasesynthesis so that cholesterol synthesis rate-limiting enzymeswill be inhibited thus reducing intracellular cholesterolsynthesis This process features three important enzymesAACT HMG-CoA reductase and ACAT Whether AACTand ACAT are involved in the effect of alisol acetates will bethe subject of intensive research in the future
The results of the study indicate that alisol acetates cannot only decrease TC level but also decrease TG and LDL-C levels and increase HDL-C level Furthermore a follow-upstudy is designed to investigate the regulatory mechanisms ofalisol acetates on TG HDL-C and LDL-C
5 Conclusion
The present study reveals that alisol acetates could signifi-cantly lower the TC level of hyperlipidemic mice The effectof alisol B 23-acetate on HMG-CoA reductase was strongerthan that of alisol A 24-acetate Alisol acetates could lowerHMG-CoA reductase activity in a dose-dependent fashionboth in vivo and in vitro Neither of these alisol acetatescould significantly lower the protein expression of HMG-CoA This suggests that alisol acetates lower the TC levelvia inhibiting the activity of HMG-CoA reductase via itsprototype drug which may have an inhibitory effect bydirectly and competitively binding with HMG-CoA In the
10 Evidence-Based Complementary and Alternative Medicine
binding interaction of alisol acetate andHMG-CoA reductasevia molecular simulation the key amino acid residues ofHMG-CoA reductase might be Lys691 Asp767 and Asn658The side chain of the alisol acetatewas the steering group Fur-thermore a folded side chainparent ring boundmoreweaklyto HMG-CoA reductase than an open side chainparent ringThis study explored the TC-lowering mechanism of alisolacetates at the molecular level and presented guidance for itsclinical use
Competing Interests
The authors declare that they have no competing interests
Acknowledgments
This work was financially supported by the National NatureScience Foundation of China (81303173 81673534) the Nat-ural Science Foundation of Jiangsu Province (BK20161576)the Natural Science Fund for Colleges and Universities inJiangsu Province (14KJB360001) and the Top-Notch Aca-demic Programs Project of Jiangsu Higher Education Insti-tutions (PPZY2015A070)
References
[1] L X Wang Q N Wu Q Zhang G P Peng and A W DingldquoBasic study of diuretic active compounds in Rhizoma Alis-matisrdquo West China Journal of Pharmaceutical Sciences vol 23no 6 pp 670ndash672 2008
[2] S S Wu Pharmaceutical and Clinical Research of Alisma Ori-entalis Chinese Press Beijing China 2007
[3] M Miyazawa S Yoshinaga and Y Kashima ldquoChemical com-position and characteristic odor compounds in essential oilfrom Alismatis Rhizoma (Tubers of Alisma orientale)rdquo DiatomResearch vol 23 no 1 pp 133ndash141 2016
[4] M-K Jang Y-R Han J S Nam et al ldquoProtective effects ofAlisma orientale extract against hepatic steatosis via inhibitionof endoplasmic reticulum stressrdquo International Journal ofMolec-ular Sciences vol 16 no 11 pp 26151ndash26165 2015
[5] Z-P Mai K Zhou G-B Ge et al ldquoProtostane triterpenoidsfrom the rhizome of Alisma orientale exhibit inhibitory effectson human carboxylesterase 2rdquo Journal of Natural Products vol78 no 10 pp 2372ndash2380 2015
[6] T Tian H Chen andY-Y Zhao ldquoTraditional uses phytochem-istry pharmacology toxicology and quality control of Alismaorientale (Sam) Juzep a reviewrdquo Journal of Ethnopharmacologyvol 158 pp 373ndash387 2014
[7] KHKimM J Kwun J-Y Choi et al ldquoTherapeutic effect of thetuber of Alisma orientale on lipopolysaccharide-induced acutelung injuryrdquo Evidence-Based Complementary and AlternativeMedicine vol 2013 Article ID 863892 10 pages 2013
[8] H-G Jin Q Jin A R Kim et al ldquoA new triterpenoid fromAlisma orientale and their antibacterial effectrdquoArchives of Phar-macal Research vol 35 no 11 pp 1919ndash1926 2012
[9] J H Lee O S Kwon H-G Jin E-R Woo Y S Kim and H PKim ldquoThe rhizomes of Alisma orientale and alisol derivativesinhibit allergic response and experimental atopic dermatitisrdquoBiological and Pharmaceutical Bulletin vol 35 no 9 pp 1581ndash1587 2012
[10] WGu C GengW Xue et al ldquoCharacterization and function ofthe 3-hydroxy-3-methylglutaryl-CoA reductase gene in Alismaorientale (Sam) Juz and its relationship with protostane triter-pene productionrdquo Plant Physiology and Biochemistry vol 97 pp378ndash389 2015
[11] W Gu B L Xi Q N Wu et al ldquoCloning and distribution pat-tern of HMGR gene conserved fragment in Alisma orientalerdquoJournal of Chinese Medicinal Materials vol 34 no 9 pp 1335ndash1339 2011
[12] T Kobayashi S Kato-Emori K Tomita and H Ezura ldquoDetec-tion of 3-hydroxy-3-methylglutaryl-coenzymeA reductase pro-tein Cm-HMGR during fruit development in melon (Cucumismelo L)rdquo Theoretical and Applied Genetics vol 104 no 5 pp779ndash785 2002
[13] R Tommer A Rachel P C Sylvie and R Jerry ldquoImpaired regu-lation of 3-hydroxy-3-methylglutaryl-coenzyme A reductasedegradation in lovastatin-resistant cellsrdquo The Journal of Bioog-ical Chemistry vol 274 no 41 pp 29341ndash29351 1999
[14] F Xu Q-N Wu J Chen et al ldquoThe binding mechanisms ofplasma protein to active compounds in Alismaorientale rhi-zomes (Alismatis Rhizoma)rdquo Bioorganic and Medicinal Chem-istry Letters vol 24 no 17 pp 4099ndash4105 2014
[15] G Gong Y Qin W Huang et al ldquoProtective effects of dios-genin in the hyperlipidemic rat model and in human vascularendothelial cells against hydrogen peroxide-induced apoptosisrdquoChemico-Biological Interactions vol 184 no 3 pp 366ndash3752010
[16] T F Tzeng H J Lu S S Lion C J Chang and IM Liu ldquoLipid-lowering effects of zerumbone a natural cyclic sesquiterpeneof Zingiber zerumbet Smith in high-fat diet-induced hyperlipi-demic hamstersrdquoFood andChemical Toxicology vol 69 pp 132ndash139 2014
[17] Y Xie H Zhang H Liu et al ldquoHypocholesterolemic effectsof Kluyveromyces marxianus M3 isolated from Tibetan mush-rooms on diet-induced hypercholesterolemia in ratrdquo BrazilianJournal of Microbiology vol 46 no 2 pp 389ndash395 2015
[18] I Karam N Ma X-W Liu et al ldquoRegulation effect of AspirinEugenol Ester on blood lipids in Wistar rats with hyperlipi-demiardquo BMC Veterinary Research vol 11 article 217 2015
[19] G Jiao J B Wei M H Yin and P T Chun ldquoA new TCMformula FTZ lowers serum cholesterol by regulating HMG-CoA reductase and CYP7A1 in hyperlipidemic ratsrdquo Journal ofEthnopharmacology vol 135 no 2 pp 299ndash307 2011
[20] Y Geng W L Tu J J Zhang L Zhang and J Zhang ldquoEffectsof extract from cole pollen on lipidmetabolism in experimentalhyperlipidemic ratsrdquo The Scientific World Journal vol 2014Article ID 982498 6 pages 2014
[21] K Sueyoshi Y Nogawa K Sugawara T Endo and HHisamoto ldquoHighly sensitive and multiple enzyme activity assayusing reagent-release capillary-isoelectric focusing with rhod-amine 110-based substratesrdquo Analytical Sciences vol 31 no 11pp 1155ndash1161 2015
[22] D Fteita E Kononen M Gursoy E Soderling and U KGursoy ldquoDoes estradiol have an impact on the dipeptidylpeptidase IV enzyme activity of the Prevotella intermedia groupbacteriardquo Anaerobe vol 36 pp 14ndash18 2015
[23] L L Zhuo LWang and N Zhou ldquoRandomly arrayed G-quad-ruplexes for label-free and real-time assay of enzyme activityrdquoChemical Communications vol 50 no 52 pp 6875ndash6878 2014
[24] M Garelnabi and A Younis ldquoParaoxonase-1 enzyme activityassay for clinical samples validation and correlation studiesrdquoMedical Science Monitor vol 21 pp 902ndash908 2015
Evidence-Based Complementary and Alternative Medicine 11
[25] X Bi andZ Liu ldquoEnzyme activity assay of glycoprotein enzymesbased on a boronate affinity molecularly imprinted 96-wellmicroplaterdquo Analytical Chemistry vol 86 no 24 pp 12382ndash12389 2014
[26] A Walls S Dermody R Kumaran N Krishnan and E HHarley ldquoCharacterization of B-Cells in tonsils of patients diag-nosed with pediatric autoimmune neuropsychiatric disorderassociated streptococcusrdquo International Journal of PediatricOtorhinolaryngology vol 80 pp 49ndash52 2016
[27] I Karam N Ma X-W Liu et al ldquoRegulation effect of AspirinEugenol Ester on blood lipids in Wistar rats with hyperlipi-demiardquo BMC Veterinary Research vol 11 no 1 article 217 2015
[28] C Sunil S Ignacimuthu and C Kumarappan ldquoHypolipidemicactivity of Symplocos cochinchinensis S Moore leaves in hyper-lipidemic ratsrdquo Journal of Natural Medicines vol 66 no 1 pp32ndash38 2012
[29] W Shen Y Han B Huang et al ldquoMicroRNA-483-3p inhibitsextracellular matrix production by targeting smad4 in humantrabecular meshwork cellsrdquo Investigative Ophthalmology amp Vis-ual Science vol 56 no 13 pp 8419ndash8427 2015
[30] Z Wu Z Yi L Dong and A Zhang ldquoMolecular simulationstudy of the specific combination between four kinds of phthalicacid esters and human serum albuminrdquo Environmental Toxicol-ogy and Pharmacology vol 41 pp 259ndash265 2016
[31] K-J Hwang M Yoon H-J Hwang S-M Choi and S-MJeong ldquoMolecular dynamics simulation of oxygen ion conduc-tion in orthorhombic perovskite Ba-doped LaInO
3using cubic
and orthorhombic modelrdquo Journal of Nanoscience and Nan-otechnology vol 15 no 11 pp 8947ndash8950 2015
[32] H Yu W Han W Ma and K Schulten ldquoTransient 120573-hairpinformation in 120572-synuclein monomer revealed by coarse-grainedmolecular dynamics simulationrdquo The Journal of ChemicalPhysics vol 143 no 24 Article ID 243142 2015
[33] M Breuer K M Rosso and J Blumberger ldquoFlavin binding tothe deca-heme cytochrome mtrc insights from computationalmolecular simulationrdquo Biophysical Journal vol 109 no 12 pp2614ndash2624 2015
[34] V Rastija D Agic S Tomic et al ldquoSynthesis QSAR and mole-cular dynamics simulation of amidino-substituted benzimi-dazoles as dipeptidyl peptidase III inhibitorsrdquo Acta ChimicaSlovenica vol 62 no 4 pp 867ndash878 2015
[35] M Enciso C Schutte and L Delle Site ldquoInfluence of pH andsequence in peptide aggregation via molecular simulationrdquoTheJournal of Chemical Physics vol 143 no 24 Article ID 2431302015
[36] L XWangQWu andG P Peng ldquoA Study on determination ofcontent of 23-acetyl alisol B in Rhizoma Alismatis orientalitisrdquoJournal of Nanjing University of Traditional Chinese Medicine(Natural Science) vol 18 no 2 pp 105ndash107 2002
[37] G P Peng andY L Zhu ldquoProgress in the study on chemical con-stituents of Alisma orientalisrdquo Natural Product Research andDevelopment vol 18 no 2 pp 348ndash351 2006
[38] D Y Liu S J Gu J Wu et al ldquoStudies on fractions of zexiedecoction with antihyperlipidemia bioactivity in micerdquo ChinaPharmacist vol 13 no 6 pp 763ndash766 2010
[39] Q-F Luo L Sun and G-H Du ldquoProgress in new drugs tar-geting reverse cholesterol transportrdquo Chinese PharmacologicalBulletin vol 22 no 8 pp 904ndash907 2006
[40] M Yoshikawa and Y Fukuda ldquoHatakeayamardquo Chemical andPharmaceutical Bulletin vol 41 no 6 pp 1194ndash1196 1993
[41] F Xu L Zhang LHe et al ldquoMolecularmechanismon the inter-action between alisols and human serum albuminrdquo Acta Chim-ica Sinica vol 69 no 19 pp 2228ndash2234 2011
[42] F Xu J ChenW Gu et al ldquoStudy on transformation of effectivecomponents of Alisma under simulated in vivo conditionsrdquoChinese Journal of Pharmaceutical Analysis vol 31 no 11 pp2035ndash2039 2011
[43] H Yang Y G Gao and L X Zhang ldquoResearch progress on bio-synthetic pathway of terpenoids containing ginsenoside and theHMGRrdquo China Journal of Biological Engineering vol 28 no 10pp 130ndash135 2008
[44] G Z Cui Y L Ben N Li and D L Liu ldquoCatalytic mechanismand characteristics of HMG-COA recuctaserdquo Journal ofMedicalMolecular Biology vol 5 no 6 p 542 2008
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom

2 Evidence-Based Complementary and Alternative Medicine
O
Me
MeMe
Me
Me
Me
MeMe
HO
H
H
H
OAc
O
(a)
MeMe
Me
Me Me
Me
MeMe
HO
OAc
O
OH OH
H
H
(b)
Figure 1 Molecular structures of alisol ((a) alisol B 23-acetate (b) alisol A 24-acetate)
HMG-CoA reductase activity can effectively reduce TCgeneration [10ndash13] Using the reagent kit method HMG-CoA reductase activity was measured in vivo and in vitrousing liver homogenates of hyperlipidemic mice before andafter the addition of alisol acetates The results indicatethat alisol acetates might lower TC via inhibition of HMG-CoA reductase activity and that the prototype drug ofalisol acetate inhibits HMG-CoA activity instead of its invivo metabolites Western blotting was used to measure theeffects of alisol acetates on the HMG-CoA reductase proteinexpression of hyperlipidemic mice The results showed thatalisol acetatesmight not inhibit HMG-CoA reductase activityvia downregulation of its protein expression Instead itmight inhibit the HMG-CoA reductase effect by directly andcompetitively binding with it The binding interaction ofalisol acetates and HMG-CoA reductase was studied usinga molecular simulation technique This technique yieldedthe following parameters the binding constant bindingenergy hydrogen bonding hydrophobichydrophilic groupselectrostatic energy and van derWaals forcesThe interactionmodel of alisol acetate and HMG-CoA reductase was builtThese experimental results were compared with the pharma-cological results in order to determine the steering groupsof this type of compound and the key amino acid residuesof the enzyme This could shed light on the cholesterol-lowering mechanism of alisol acetates at the molecular levelThe results were invaluable for promoting the applications ofAlismatis rhizoma in the clinical practice
2 Materials and Methods
21 Reagents and Instruments Themain reagents were as fol-lows simvastatin (MSD Pharmaceutical Co Ltd HangzhouChina) cholesterol (National Group Chemical Reagent CoLtd Beijing China) sodium deoxycholate (National GroupChemical Reagent Co Ltd Beijing China) propylthiouraciltablets (Jinghua Pharmaceutical Co Ltd Nantong China)lard (refined from the leaf lard purchased at the localmarket)polysorbate 80 (Tween 80 Qiangshun Chemical ReagentCo Ltd Shanghai China) propylene glycol (QiangshunChemical Reagent Co Ltd Shanghai China) basal diet (Slac
Laboratory Animal Co Ltd Shanghai China) radioim-munoprecipitation assay (RIPA) tissue cell lysis buffer (Solar-bio Beijing China cat number R0010) bicinchoninic acid(BCA) protein quantitative reagent kit (Thermo MA US catnumber PICPI23223) alisol A 24-acetate standard (MansiteBiological Technology Co Ltd Chengdu China batch num-ber MUST-15022104 98 purity) and alisol B 23-acetatestandard (Mansite Biological Technology Co Ltd ChengduChina batch number MUST-149091210 with 98 purity)The structures of the alisol acetates are shown in Figure 1[2 14]
Further reagents were as follows mice TG enzyme-linked immunosorbent assay (ELISA) reagent kits (Fengx-iang Biotechnology Co Ltd Shanghai China batch num-ber 201207) mice HDL-C ELISA reagent kits (FengxiangBiotechnology Co Ltd Shanghai China batch number201302) mice LDL-C ELISA reagent kits (Fengxiang Biotech-nology Co Ltd Shanghai China batch number 201301)mice TC ELISA reagent kits (Fengxiang Biotechnology CoLtd Shanghai China batch number 201202) HMG-CoAreductase activity assay kits (Genmed Scientifics Inc Shang-hai China batch number 20080717) HMG-CoA reductaseantibodies (Ray Biotech Inc Atlanta US batch number168-10733) internal standard (glyceraldehyde-3-phosphatedehydrogenase [GAPDH]) protein antibody (CST BostonUS batch number 5174) and nitrocellulose membrane (Mil-lipore Boston US batch number HATF00010) All otherchemicals used were of analytical grade and were obtainedfrom China
The following instruments were employed in this studya high-speed ben centrifuge (Medical Analytical InstrumentFactory Shanghai China H1850) an electronic analyticalbalance (Sunny Hengping Scientific Instrument Co LtdShanghai China FA1104) pipettes (Jiarsquoan Analytical Instru-ment Factory Shanghai China WKYIII) a UV-Vis spec-trophotometer (Persee General Instrument Co Ltd BeijingChina TU-1901) a precision digital high-speed homoge-nizer (Shuoguang Electronic Technology Co Ltd ShanghaiChina SG-3048A) an electrophoresis apparatus (BIO-RADCalifornia US mini protean 3 cell) an electroporator (Jing-mai Technology Co Ltd Dalian China PS-9) a microplate
Evidence-Based Complementary and Alternative Medicine 3
Table 1 Blood lipid levels of each group of mice (119909 plusmn 119904 119899 = 11)
Group Animal count TC (ngmL) TG (ngmL) HDL-C (ngmL) LDL-C (ngmL)Blank 11 894 plusmn 137 951 plusmn 152 6893 plusmn 791 1422 plusmn 457Model 11 2905 plusmn 149998771998771 3059 plusmn 302998771998771 1786 plusmn 559998771998771 5171 plusmn 317998771998771
Positive 11 977 plusmn 144lowastlowast 1098 plusmn 123lowastlowast 4511 plusmn 572lowastlowast 2111 plusmn 637lowastlowast
23B high 11 1450 plusmn 191lowastlowast 1408 plusmn 197lowastlowast 4623 plusmn 848lowastlowast 2639 plusmn 401lowastlowast
23B medium 11 2310 plusmn 404lowastlowast 2300 plusmn 167lowastlowast 3756 plusmn 661lowastlowast 3357 plusmn 372lowastlowast
23B low 11 2430 plusmn 218lowastlowast 2456 plusmn 418lowastlowast 2908 plusmn 781lowastlowast 3610 plusmn 463lowastlowast
24A high 11 1870 plusmn 244lowastlowast 1744 plusmn 224lowastlowast 4289 plusmn 543lowastlowast 2923 plusmn 633lowastlowast
24A medium 11 2527 plusmn 531lowast 2679 plusmn 434lowast 2813 plusmn 553lowastlowast 3692 plusmn 529lowastlowast
24A low 11 2757 plusmn 168lowast 2715 plusmn 324lowast 2698 plusmn 377lowastlowast 3857 plusmn 591lowastlowast
Note 24A alisol A 24-acetate 23B alisol B 23-acetate compared to the blank group 998771998771119875 lt 001 and 998771119875 lt 005 compared to the model group lowastlowast119875 lt 001and lowast119875 lt 005
reader (Labsystems Multiskan MS Finland Model 352) awater bath (Leica Germany HI1210) and an imaging system(Tanon Co Ltd Shanghai China Tanon-5200)
22 Animals Male clean-grade ICR mice (18ndash22 g) werepurchased from the Animal Core Facility Nanjing MedicalUniversity (license number SCXK (Su)-2013-0005) Theanimals were placed in a clean-grade animal room at 20ndash24∘C and 55 plusmn 5 humidity The light and dark cycles weretwelve hours eachThe mice were free-fed for one week priorto the experimentsTheAnimal Ethics Committee of NanjingUniversity of Chinese Medicine approved all protocols ofanimal experiments
23 Measurements of TC TG HDL-C and LDL-C Levels
231 The Induction of an Animal Model of Hyperlipidemia[15ndash19] A total of 99 male mice were randomly dividedinto nine groups according to their weight Mice in onegroup the normal blank group were intragastrically feddistilled water (10mLkg) every morning Mice in the othergroupwere intragastrically fed lipid emulsion (10mLkg)Theformulation of the lipid emulsion was as follows 25 lard15 cholesterol 10 polysorbate 80 30 propylene glycol2 propylthiouracil 2 sodium cholate and 1 sugar Forfive consecutive weeks the mice were free-fed a basal dietalongside the intragastric administration of distilled water orlipid emulsion The mice were fasted for twelve hours (waterwas permitted) after the last intragastric administrationBlood samples of mice were collected and their serums wereseparated The TG TC HDL-C and LDL-C concentrationsin serums were measured according to the instructions of thecorresponding reagent kits making it possible to comparethe blood lipid levels of the model and the blank groupsAccording to the results in Table 1 the TG TC and LDL-Cof themodel group increasedwhile theHDL-C droppedThissuggests successful establishment of the hyperlipidemiamod-elsThemodel animals were evenly divided into eight groupsmodel simvastatin alisol B 23-acetate (high medium andlow dosage) and alisol A 24-acetate (high medium and lowdosage)
232 Drug Supply to Hyperlipidemic Mice [19] The appro-priate quantities of alisol A 24-acetate alisol B 23-acetateand simvastatin were weighed dissolved in 03 sodium car-boxymethylcellulose (CMC-Na) solution and diluted to26mL
The quantities administered intragastrically to eachgroup were as follows positive control group (simvastatin7mgkgday) alisol A 24-acetate low-dose group (064mgkgday) alisol A 24-acetate medium-dose group (128mgkgday) alisol A 24-acetate high-dose group (256mgkgday)alisol B 23-acetate low-dose group (064mgkgday) alisol B23-acetatemedium-dose group (128mgkgday) and alisol B23-acetate high-dose group (256mgkgday)
Each group was intragastrically fed 10mLkg of lipidemulsion every morning and treated with the correspondingdrugs every afternoon for three weeks
233 Measurements of TC TG HDL-C and LDL-C LevelsThemice were fasted for twelve hours (water was permitted)following the last dose of drug Blood samples were collectedand left to sit for 20ndash30minutesThe serumswere obtained bycentrifuging the blood samples at 3000 rmin for 15 minutesThe collected serums were measured in accordance with theinstruction of the TC TG HDL-C and LDL-C reagent kits
24 Measurement of In VivoIn Vitro HMG-CoAReductase Activity [19ndash25]
241 Measurement of In Vitro HMG-CoA Reductase ActivityThe establishment of themousemodel of hyperlipidemia wasdone according to Section 231 After the last intragastricadministration (procedure described in Section 231) themice were fasted for twelve hours (water was permitted)and then decapitated Immediately after decapitation theperitoneal cavities of the mice were rinsed with 10mL of coldsaline while the livers of the mice were rapidly frozen inliquid nitrogen and stored at minus70∘C for further analysis
At a temperature of 4∘C 05 g of each liver was weighedplaced in phosphate buffered saline (PBS pH 72 025 gmL)and homogenized Subsequently themixturewas centrifugedfor tenminutes at 4000 rmin and at a temperature of 4∘CThe
4 Evidence-Based Complementary and Alternative Medicine
supernatant was then collected On three separate occasions1mL of the supernatant of the liver homogenate was collectedand extracted with 10mL of 95 ethanol at 60∘C This wasfollowed by vacuum filtration The residues were dissolvedin PBS The samples were adjusted to a certain concentrationand stored at minus70∘C
We added 004mgmL of simvastatin 016mgmL008mgmL and 004mgmL (high medium and lowdosage) of alisol A 24-acetate and alisol B 23-acetate to 5mLof liver homogenate samples respectively After leaving thesamples to stand for 24 hours theHMG-CoA reductase activ-ity was measured using the HMG-CoA reductase activityassay kit in accordance with the instructions
242 Measurement of In Vivo HMG-CoA Reductase ActivityThe induction of the animal model of hyperlipidemia wasdone according to Section 231 The intragastric adminis-tration was performed as described in Section 232 Themice were fasted for twelve hours (water was permitted) anddecapitated after the last dose of the drug The peritonealcavities of the mice were immediately rinsed with 10mL ofcold saline whereas the livers of the mice were rapidly frozenin liquid nitrogen and then stored for further analysis at atemperature of minus70∘C At 4∘C 05 g of each liver were col-lected placed in PBS (pH 72 025 gmL) and homogenizedThe sample was subsequently centrifuged for ten minutes at4000 rmin and at a temperature of 4∘CThe supernatant wasthen collected For each mouse 1mL of liver homogenatesupernatant was extracted with 10mL of 95 ethanol at 60∘Con three occasions This was followed by vacuum filtrationThe residues were dissolved in PBSThe solutionwas adjustedto a certain concentration and stored at minus70∘C The HMG-CoA reductase activity of the liver homogenate samples wasmeasured in accordance with the instructions given in theHMG-CoA reductase activity assay kit
25 Measurement of HMG-CoA Reductase Protein Expres-sion Level via Western Blotting [26ndash29] The hyperlipidemicinduction of animal model of hyperlipidemia was builtaccording to Section 231 The intragastric administrationfollowed the procedure described in Section 232 whereasthe liver homogenate samples were prepared as described inSection 242 The samples were stored at minus70∘C for furtheruse
In order to detect the protein concentration of the super-natant of each sample the A
595 nm absorbance value wasmea-sured via the Bradford method and the calibration curve ofthe bovine serum albumin protein was plotted An equivalentvolume of 2 times sodium dodecyl sulfate (SDS) buffer was addedto the liver homogenate sampleThemixture was denaturizedat 100∘C for three minutes Subsequently the protein was iso-lated using SDS-polyacrylamide gel electrophoresis (PAGE)and electrotransferred to the nitrocellulose membrane Thenitrocellulose membrane was removed and blocked betweenone and two hours in the tris-buffered saline with Tween 20(TBST) solution with 5 skim-milk powder Subsequentlythe membrane was washed with TBST and placed intothe primary antibody (1 2500 rabbit anti-rat HMG-CoAreductase polyclonal antibody) at 4∘C overnight The next
day themembranewas washedwith TBST and blocked in theTBST solution with 5 skim-milk powder for two hoursThemembrane was washed thrice with TBST for 10ndash20 minutesper wash The membrane was incubated for one hour withalkaline phosphatase-conjugated mouse anti-rabbit IgG at adilution of 1 500 Following incubation in the secondaryantibody the membrane was washed again and visualizedusing electrochemiluminescence (ECL) The membrane wasplaced in the antibody elution buffer for ten minutes andrinsed in TBST solution for five minutes It was then blockedin the TBST solution with 5 skim-milk powder and shakenat room temperature for 30 minutes The membrane wasrinsed with TBST repeatedly and then incubated with pri-mary antibody of internal standard GAPDH at a dilution of1 1000 The membrane was subsequently washed and incu-bated with secondary antibody at a dilution of 1 250 Follow-ing washing the membrane was visualized The gray valuesof the filmstrips were measured using a gel imaging ana-lysis system The calculated gray level ratio (HMG-CoAreductaseGAPDH) represents the relative amount of HMG-CoA reductase protein expression
26 Investigation of the Interaction between Alisol Acetatesand HMG-CoA Reductase [30ndash35] A molecular simulationwas conducted The initial molecular structures of the alisolacetates were generated using Discovery Studio 21 (DS21Accelrys US) The structure of HMG-CoA reductase wasobtained from the crystal structure in the protein data bank(PDB) database (PDB code 1HW9) HMG-CoA reductaseis a glycoprotein with N-linked high-mannose oligosaccha-rides consisting of four identical subunits with a molecularweight of 97 ku Each subunit comprised 888 amino acidresidues with the phosphorylation site located at the 872ndamino acid Each subunit featured three different domainsthe N-terminal 339 (1ndash339) amino acids composed the trans-membrane domain the 110 (340ndash449) amino acids composedthe junction domain and the C-terminal 439 (450ndash888)amino acids composed the catalytic domainThe N-terminalcrossed the endoplasmic reticulum membrane eight timesand anchored to the membrane through short rings Thetransmembrane domain was composed of 167 amino acidresidues As the four subunits were identical according to itsinteraction with the drug the A chain was selected as thereceptor for docking simulation Following the removal ofwatermolecules heteroatoms and themulticonformations ofthe amino acid residues in the crystal structure flexible dock-ing in theChemistry atHARvardMacromolecularMechanics(CHARMM) force field was performed Subsequently to thesolvation calculation the compensation ions Na+ and Clminuswere added to the system in order to simulate the environ-ment in the human bodyThemost likely confirmation of theresulting complex was confirmed via minimization calcula-tion
261 Molecular Simulation of Alisol B 23-Acetate and HMG-CoA Reductase The alisol B 23-acetate was defined as aligand After docking the LibDock score the CDockerenergy the CDocker interaction energy and the hydro-gen bond formation of the ligand-receptor complex were
Evidence-Based Complementary and Alternative Medicine 5
comprehensively considered so as to determine the finalsteady conformation This conformation was selected forfurther molecular dynamic simulation
262 Molecular Simulation of Alisol A 24-Acetate andHMG-CoA Reductase Applying the method of Section 261resulted in the final steady conformation of alisol A 24-acetateThis conformationwas selected for furthermoleculardynamic simulation
27 Data Analysis The experimental data were analyzedusing one-way analysis of variance (ANOVA) in SPSS version150 (SPSS Inc Chicago USA) 119875 values of less than 005(119875 lt 005) indicated statistically significant differences Theresults were expressed in the form of mean (119909) plusmn standarddeviation (SD)
3 Results
31 Measurements of TC TG HDL-C and LDL-C LevelsThe blood lipid levels of the blank group positive groupmodel group alisol B 23-acetate high- medium- and low-dose groups and alisol A 24-acetate high- medium- andlow-dosage groups are shown in Table 1 This study revealedthat the TC TG LDL-C and HDL-C levels of the alisol A24-acetate high- medium- and low-dose groups as well asthe alisol B 23-acetate high- medium- and low-dose groupssignificantly differed from those of the model group Thissuggests that alisol acetates could have a lipid-regulatingeffect and significantly decrease the TC TG and LDL-Cconcentrations of hyperlipidemic mice while increasing theHDL-C concentrations
32 Measurement of In VivoIn Vitro HMG-CoAReductase Activity
321 Measurement of In Vitro HMG-CoA Reductase ActivityAs Table 2 indicates alisol acetates lower the HMG-CoAreductase activity dose dependently The extent to whichalisol B 23-acetate lowered HMG-CoA reductase activityexceeded the extent to which alisol A 24-acetate lowered theactivity of this enzyme
322 Measurement of In Vivo HMG-CoA Reductase ActivityThe results in Table 3 illustrate that alisol acetates lower thein vivo HMG-CoA reductase activity in a dose-dependentfashion with a greater degree of enzyme inhibition for alisolB 23-acetate than for alisol A 24-acetate
33 Measurement of HMG-CoA Reductase Protein Expres-sion Level via Western Blotting The western blotting resultssuggest that alisol acetates did not significantly decrease theprotein expression of HMG-CoA reductase (Figure 2)
34 Investigation of the Interaction between the Alisol Acetatesand HMG-CoA Reductase
341 Molecular Simulation Results for Alisol B 23-Acetate andHMG-CoA Reductase The molecular simulation results forthe interaction between alisol B 23-acetate and HMG-CoA
Table 2 The activity of HMG-CoA reductase in vitro (119909 plusmn s 119899 =11)
Group HMG-CoA reductase (UL)Blank 13467 plusmn 3991Positive 16429 plusmn 2832lowastlowast
Model 49788 plusmn 2881998771998771
24A high 19042 plusmn 3092lowastlowast
24A medium 25079 plusmn 1635lowastlowast
24A low 33515 plusmn 2332lowastlowast
23B high 17912 plusmn 4159lowastlowast
23B medium 23079 plusmn 1664lowastlowast
23B low 32759 plusmn 1915lowastlowast
Note 24A alisol A 24-acetate and 23B alisol B 23-acetate compared to theblank group 998771998771119875 lt 001 and 998771119875 lt 005 compared to the model grouplowastlowast119875 lt 001 and lowast119875 lt 005
Table 3The activity of HMG-CoA reductase in vivo (119909 plusmn s 119899 = 11)
Group HMG-CoA reductase (UL)Blank 12345 plusmn 2534Positive 20021 plusmn 3104lowastlowast
Model 49012 plusmn 4117998771998771
24A high 280123 plusmn 2915lowastlowast
24A medium 34554 plusmn 3234lowastlowast
24A low 40131 plusmn 4221lowastlowast
23B high 25203 plusmn 2931lowastlowast
23B medium 29614 plusmn 3218lowastlowast
23B low 39082 plusmn 2323lowastlowast
Note 24A alisol A 24-acetate and 23B alisol B 23-acetate compared to theblank group 998771998771119875 lt 001 and 998771119875 lt 005 compared to the model grouplowastlowast119875 lt 001 and lowast119875 lt 005
reductase are depicted in Figure 3 Figure 3(a) illustratesthe overall pattern of the interaction The interaction pocketwhere alisol B 23-acetate bound to HMG-CoA reductasecontained the following amino acids Leu521 Val522Met523Gly524 Ala525 Cys527 Glu528 Asn529 Val530 Met588Thy589 Arg590 Gly591 Asp653 Ala654 Met655 Gly656Met657 Asn658 Met659 Ile660 Ser661 Lys662 Gly663Thy664 Glu665 Cys688 Asp690 Lys691 Lys692 Arg702Lys704 Ile760 Try761 Ile762Ala763 Cys764Glu801 Ile802Gly803 Thr804 Val805 Gly806 Gly807 Gly808 Thr809Asn810 and Leu811Theywere located in the catalytic domainconstituted by the 439 (450ndash888) amino acids at the C-terminal After entering the interaction pocket alisol B 23-acetate interacted with the peripheral amino acid residues viahydrogen bonding and hydrophobic and hydrophilic effectsThe hydrogen bond graph in Figure 3(b) shows two hydrogenbonds between alisol B 23-acetate and the amino acids Lys691and Asp767 Table 4 presents detailed information aboutthe hydrogen bonding The interaction energy between thehydrophobic amino acid residues and the small molecule wascalculated to be minus128 KJmol while the interaction energybetween hydrophilic amino acid residues and the smallmolecule was calculated to be minus1353 KJmol This indicatesthat the hydrophilic interaction between the small moleculeand the peripheral amino acids was stronger
6 Evidence-Based Complementary and Alternative Medicine
HMG-CoAreductase
GAPDH
05
04
03
02
01
00HM
G-C
oA re
duct
ase
GA
PDH
Blan
k
Mod
el
Posit
ive
24
A-H
24
A-M
24
A-L
23
B-H
23
B-M
23
B-L
Blan
k
Mod
el
Posit
ive
24
A-H
24
A-M
24
A-L
23
B-H
23
B-M
23
B-L
lowast
Figure 2 Results of the protein expression of HMG-CoA reductasein mice liver tissue protein (119899 = 11) Note compared to the blankgroup 998771998771119875 lt 001 and 998771119875 lt 005 compared to the model grouplowastlowast119875 lt 001 and lowast119875 lt 005 24A-H alisol A 24-acetate high 24A-
M alisol A 24-acetate medium and 24A-L alisol A 24-acetate low23B-H alisol B 23-acetate high 23B-M alisol B 23-acetate mediumand 23B-L alisol B 24-acetate low
Table 4 The length (gt) and the angle of hydrogen bond formedbetween alisol acetates and HMG-CoA reductase
Name XndashHsdot sdot sdotYd
(XndashH)
d(Hsdot sdot sdotY)
d(Xsdot sdot sdotY) Angle XHY
Alisol B
23-acetate
Lys691 NZ-HZ1sdot sdot sdotO37 104 237 298 11594
Asp767OD2-
HH88sdot sdot sdotO2096 240 324 14490
Alisol A24-acetate
Asn658 OD1-H5Zsdot sdot sdotO5 095 201 286 14897
After docking the conformation of alisol B 23-acetatechanged as shown in Figure 3(c) The calculated root-mean-square deviation (RMSD) was 289gt The conformationof the small molecule changed from the clustered sheetstructure to the open structure Before docking the sidechain and the ring structure of the small molecule foldedas shown in Figures 3(d) and 3(e) The angle between C
2ndash
C10ndashC19was 1095∘ 1095∘ between C
10ndashC19ndashC20 and minus599∘
between C2ndashC10ndashC19ndashC20 As shown in Figures 3(f) and
3(g) after docking the angle between C2ndashC10ndashC19was 1136∘
1156∘ between C10ndashC19ndashC20 and 1260∘ between C
2ndashC10ndash
C19ndashC20 The angle between the parent ring and the side
chain was significantly increasedThe twisting of the dihedralangles resulted in the opening of side chains and insertioninto the protein which led to interactions There was ahydrophilic ether bond on the side chain and the peripheralamino acids were mainly hydrophilic (Figures 3(h) and 3(i))
This created a hydrophilic environment around the sidechains culminating in stable binding The energy of thevan der Waals forces between them was calculated to beminus1519 KJmol whereas the electrostatic energy was calcu-lated to be minus2366 KJmol As the electrostatic energy waslower than the energy of the van derWaals forces between thesystems the electrostatic interactionwas themain interactionbetween this small molecule and the macromolecule Thebinding energy (ΔG∘) between the alisol B 23-acetate and theHMG-CoA reductase was calculated to be minus1033 KJmol
342 Molecular Simulation Results of Alisol A 24-Acetateand HMG-CoA Reductase Themolecular simulation resultsfor the interaction between alisol A 24-acetate and HMG-CoA reductase are shown in Figure 4 Figure 4(a) shows theoverall interaction pattern of this compound and HMG-CoAreductase The interaction pocket where it bound to HMG-CoA reductase contained the following amino acids Tyr519Ser520 Leu521 Val522 Met523 Gly524 Ala525 Cys527Glu528 Asn529 Val530 Ile531 Gly532 Try533 Met588Thy589 Arg590 Gly591 Asp653 Ala654 Met655 Gly656Met657 Asn658 Met659 Ile660 Ser661 Lys662 Gly663Thy664 Glu665 Cys688 The689 Asp690 Lys691 Lys692Cys764 Gly765 Gln766 Asp767 Ala768 Ala769 Gln770Asn771 Val772 Gly773 Ser774 Glu801 Ile802 Gly803Thr804Val805Gly806Gly807Gly808Thr809 andAsn810There were 439 (450ndash888) amino acids in the catalyticdomain at the C-terminal After entering the interactionpocket alisol A 24-acetate exhibited hydrogen bonding andhydrophobic and hydrophilic interactions with peripheralamino acid residues Figure 4(b) shows the hydrogen bondcontact map There was one hydrogen bond between alisol A24 acetate and amino acid Asn658 (Table 4) The interactionenergy of the hydrophobic amino acid residues and thesmall molecule was calculated to be 465 KJmol whereas theinteraction energy of the hydrophilic amino acid residues andthe small molecule was calculated to be minus1023 KJmol Thissuggests that the hydrophilic groups had a much strongereffect than the hydrophobic groups The small molecule andthe peripheral amino acids exhibited a stronger hydrophilicinteraction as indicated by the interface graphs (Figures 4(f)and 4(g))
After docking the alisol A 24-acetate conformationchanged (Figure 4(c)) The RMSD was calculated to be334gt The conformation angle C
3ndashC18ndashC26ndashC28
changedfrom 600∘ before docking to 1242∘ after docking (Figures4(d) and 4(e)) This suggests a large torsion of the smallmolecule side chain after docking whereby the stretchedopen state changed to a ring-like sheet state Consequentlyit was difficult to insert the side chain into the protein andinteract with the active pocket of the protein This resultedin considerably lower binding relative to alisol B 23-acetateThe electrostatic energy of alisol A 24-acetate and HMG-CoA reductase was determined to be minus886 KJmol whilethe energy of the van der Waals forces was determined tobe minus1649 KJmol The energy of the van der Waals forceswas lower than the electrostatic energy showing that themain interaction between this small molecule and themacro-molecule was the van derWaals interactionThe ΔG∘ of alisol
Evidence-Based Complementary and Alternative Medicine 7
(a) (b) (c)
(d) (e) (f)
(g) (h) (i)
Figure 3 Interaction between alisol B 23-acetate and HMG-CoA reductase (a) Overall pattern of interaction (alisol B 23-acetate is denotedby a bar chart) (b) Hydrogen bond graph of alisol B 23-acetate with Lys691 and Asp767 (the hydrogen bond is denoted by a dotted line) (c)Stacking chart of alisol B 23-acetate before and after docking (the initial structure is shown in green and the structure after docking is shown inpurple) (d) Initial angle of alisol B 23-acetate C
2ndashC10ndashC19and C
10ndashC19ndashC20 (e) Initial dihedral angle of alisol B 23-acetate C
2ndashC10ndashC19ndashC20
(f) The angle of alisol B 23-acetate C2ndashC10ndashC19and C
10ndashC19ndashC20after stacking (g)The dihedral angle of alisol B 23-acetate C
2ndashC10ndashC19ndashC20
after stacking (h) Hydrophobic interface of the alisol B 23-acetate side chain and the peripheral amino acid residues after stacking (alisol B23-acetate is denoted by yellow bars) (i) Hydrophilic interface of the alisol B 23-acetate side chain and the peripheral amino acid residuesafter stacking (alisol B 23-acetate is denoted by yellow bars)
A 24-acetate and HMG-CoA reductase was calculated to beminus23 KJmol
4 Discussion
Various experimental and clinical studies focus on the bloodlipid reducing effects of different solvent extracts (especially
water and alcohol extracts) fromAlisma orientale and its pro-cessed products [36ndash38] However the mechanisms under-lying the effects of Alisma orientale constituents remain to berevealed Alisma orientalemay interfere with exogenous cho-lesterol absorption and endogenous cholesterol metabolismthus reducing the level of TC Moreover alisol acetates mayinterfere with themetabolism of endogenous cholesterol [39]
8 Evidence-Based Complementary and Alternative Medicine
(a) (b) (c)
(d) (e) (f)
(g)
Figure 4 The interaction between alisol A 24-acetate and HMG-CoA reductase (a) Overall pattern of interaction (alisol A 24-acetate isdenoted by a bar chart) (b) Hydrogen bond graph of alisol A 24-acetate and Asn658 (the hydrogen bond is denoted by a dotted line) (c)Stacking chart of alisol A 24-acetate before and after docking (the initial structure is shown in green and the structure after docking is shownin purple) (d) The initial dihedral angle of alisol A 24-acetate angle C
3ndashC18ndashC26ndashC28 (e) The dihedral angle of alisol A 24-acetate angle
C3ndashC18ndashC26ndashC28after stacking (f) Hydrophobic interface of the alisol A 24-acetate side chain and the peripheral amino acid residues after
stacking (alisol A 24-acetate is denoted by yellow bars) (g) Hydrophilic interface of the alisol A 24-acetate side chain and the peripheralamino acid residues after stacking (alisol A 24-acetate is denoted by yellow bars)
Therefore we studied the TC-lowering effect of alisolacetates based on HMG-CoA reductase and the underly-ing molecular mechanism As the main source of endoge-nous cholesterol and one of the key enzymes in the TCmetabolism HMG-CoA reductase is the first described rate-limiting enzyme in cholesterol synthesis in vivo Moreover its
activation directly affects the speed of cholesterol synthesisand the level of TC in vivo
The TC-lowering effect and molecular mechanism ofalisol acetates the main active ingredient of Alismatis rhi-zoma were investigated using the reagent kit method andwestern blotting technique combined with the molecular
Evidence-Based Complementary and Alternative Medicine 9
simulation technique An interaction model was built foralisol acetate and the key enzyme involved in the metabolismof TC was HMG-CoA reductase The present study revealsthat alisol acetates can significantly lower the TC level ofhyperlipidemic miceThe alisol B 23-acetate showed a higherextent of TC lowering than alisol A 24-acetate
Consistent with the in vitro results alisol acetates couldalso lower the in vivoHMG-CoA reductase activity in a dose-dependent fashion with a higher degree of enzyme inhibitionfor alisol B 23-acetate compared to alisol A 24-acetate Theseresults suggest that alisol acetates might lower TC levels viaregulation of HMG-CoA reductase activity The consistencybetween the results of the in vivo and in vitro experimentsindicates that the alisol acetate prototype drug inhibitsHMG-CoA reductase activity instead of the in vivometabolites
Revealed by the western blotting results alisol acetatesmight inhibit the activity of HMG-CoA reductase by directlyand competitively binding to HMG-CoA reductase ratherthan by downregulating its protein expression
The molecular simulation technique was employed inorder to investigate the interaction of alisol acetates andHMG-CoA reductase The results (Figures 3 and 4 andTable 4) revealed themolecularmechanism of the interactionof alisol acetates and HMG-CoA reductase Both alisolacetates bound to HMG-CoA reductase in their catalyticdomain at the C-terminalThe alisol B 23-acetate formed twohydrogen bonds with HMG-CoA reductase The hydrophilicinteraction was stronger than the hydrophobic interactionwhereas the electrostatic interaction was stronger than thevan der Waals forces After the interaction the structure ofthe small molecule dramatically changed from the originalfolded sheet to an open structure Additionally the side chainstretched and was inserted into the protein The peripheralamino acids formed a hydrophilic environment and stablybound with it In general a hydrogen bond with a lengthof less than 35 A and an angle of 90ndash180∘ is consideredto be strong These two hydrogen bonds were relativelystrong and might be partly responsible for the torsion ofthe small molecule The alisol A 24-acetate formed onehydrogen bond with the HMG-CoA reductase and this bondwas weaker compared to those of alisol B 23-acetate Thehydrophilic interaction was stronger than the hydrophobicinteraction and the van derWaals force was stronger than theelectrostatic interaction After the interaction the moleculeside chain had large torsion and changed from the originalstretched open state to a ring-like sheet state which wasdifficult to insert into the proteinThe binding energy of alisolB 23-acetate andHMG-CoA reductase was lower than that ofalisol A 24-acetate and HMG-CoA reductase indicating thatthe binding of alisol B 23-acetate and HMG-CoA reductasewas stable According to the pharmacological results alisolB 23-acetate lowered the HMG-CoA reductase activity morethan alisol A 24-acetate This suggests that the active centerwas the interaction pocket constituted by these amino acidsand that the hydrogen bonds involving Lys691 Asp767 andAsn658 were the key amino acid residues for the interactionof this protein with alisol acetates
The binding of alisol B 23-acetate and HMG-CoA reduc-tase was considerably stronger than the binding of alisol
A 24-acetate and HMG-CoA reductase The relatively weakbinding of alisol A 24-acetate and HMG-CoA reductasemight be due to its distinct side-chain structure The sidechain of alisol B 23-acetate contained an ether bond whichwas strongly hydrophilic and could be inserted into thehydrophilic environment of the protein to form strong inter-actions Previous studies found that the side chains of alisolacetates were their active groups [40] In the present studythis conclusionwas validated A folded side chainparent ringbound more strongly to the macromolecule than an openside-chainparent ring did Thus the side chain acts as thesteering group in that it steers the interactions between alisolacetates and macromolecules This might be the key groupfor interactions with macromolecules as it had a decisiveeffect on the interactions of this type of compound withmacromolecules Our previous studies indicated that alisol B23-acetate was partially converted to alisol A 24-acetate in theenvironment of the human stomach However the bindingof alisol A 24-acetate and HMG-CoA reductase was weaker[14 41 42] Therefore it is advisable for drug developers todevelop a dosage form that circumvents the conversion ofalisol B 23-acetate in the stomach
Previously published studies indicate that [43 44] themechanism of reducing the level of TC is that two acetylCoA molecules condense to acetyl CoA under the catalysisof acetoacetyl-coenzyme A thiolase (AACT) acetyl CoAand acetyl CoA condense to HMG-CoA and HMG-CoAreduces to MVA under the catalytic action of HMG-CoAreductase MVA further generates TC with squalene Thehigh intracellular free fatty acid (FC) level stimulates micro-somal acyl coenzyme A-cholesterol acyltransferase (ACAT)and promotes esterification of the cholesterol Subsequentlycholesterol is stored in the form of cholesterol esters (CE)which is a feedback inhibition of HMG-CoA reductasesynthesis so that cholesterol synthesis rate-limiting enzymeswill be inhibited thus reducing intracellular cholesterolsynthesis This process features three important enzymesAACT HMG-CoA reductase and ACAT Whether AACTand ACAT are involved in the effect of alisol acetates will bethe subject of intensive research in the future
The results of the study indicate that alisol acetates cannot only decrease TC level but also decrease TG and LDL-C levels and increase HDL-C level Furthermore a follow-upstudy is designed to investigate the regulatory mechanisms ofalisol acetates on TG HDL-C and LDL-C
5 Conclusion
The present study reveals that alisol acetates could signifi-cantly lower the TC level of hyperlipidemic mice The effectof alisol B 23-acetate on HMG-CoA reductase was strongerthan that of alisol A 24-acetate Alisol acetates could lowerHMG-CoA reductase activity in a dose-dependent fashionboth in vivo and in vitro Neither of these alisol acetatescould significantly lower the protein expression of HMG-CoA This suggests that alisol acetates lower the TC levelvia inhibiting the activity of HMG-CoA reductase via itsprototype drug which may have an inhibitory effect bydirectly and competitively binding with HMG-CoA In the
10 Evidence-Based Complementary and Alternative Medicine
binding interaction of alisol acetate andHMG-CoA reductasevia molecular simulation the key amino acid residues ofHMG-CoA reductase might be Lys691 Asp767 and Asn658The side chain of the alisol acetatewas the steering group Fur-thermore a folded side chainparent ring boundmoreweaklyto HMG-CoA reductase than an open side chainparent ringThis study explored the TC-lowering mechanism of alisolacetates at the molecular level and presented guidance for itsclinical use
Competing Interests
The authors declare that they have no competing interests
Acknowledgments
This work was financially supported by the National NatureScience Foundation of China (81303173 81673534) the Nat-ural Science Foundation of Jiangsu Province (BK20161576)the Natural Science Fund for Colleges and Universities inJiangsu Province (14KJB360001) and the Top-Notch Aca-demic Programs Project of Jiangsu Higher Education Insti-tutions (PPZY2015A070)
References
[1] L X Wang Q N Wu Q Zhang G P Peng and A W DingldquoBasic study of diuretic active compounds in Rhizoma Alis-matisrdquo West China Journal of Pharmaceutical Sciences vol 23no 6 pp 670ndash672 2008
[2] S S Wu Pharmaceutical and Clinical Research of Alisma Ori-entalis Chinese Press Beijing China 2007
[3] M Miyazawa S Yoshinaga and Y Kashima ldquoChemical com-position and characteristic odor compounds in essential oilfrom Alismatis Rhizoma (Tubers of Alisma orientale)rdquo DiatomResearch vol 23 no 1 pp 133ndash141 2016
[4] M-K Jang Y-R Han J S Nam et al ldquoProtective effects ofAlisma orientale extract against hepatic steatosis via inhibitionof endoplasmic reticulum stressrdquo International Journal ofMolec-ular Sciences vol 16 no 11 pp 26151ndash26165 2015
[5] Z-P Mai K Zhou G-B Ge et al ldquoProtostane triterpenoidsfrom the rhizome of Alisma orientale exhibit inhibitory effectson human carboxylesterase 2rdquo Journal of Natural Products vol78 no 10 pp 2372ndash2380 2015
[6] T Tian H Chen andY-Y Zhao ldquoTraditional uses phytochem-istry pharmacology toxicology and quality control of Alismaorientale (Sam) Juzep a reviewrdquo Journal of Ethnopharmacologyvol 158 pp 373ndash387 2014
[7] KHKimM J Kwun J-Y Choi et al ldquoTherapeutic effect of thetuber of Alisma orientale on lipopolysaccharide-induced acutelung injuryrdquo Evidence-Based Complementary and AlternativeMedicine vol 2013 Article ID 863892 10 pages 2013
[8] H-G Jin Q Jin A R Kim et al ldquoA new triterpenoid fromAlisma orientale and their antibacterial effectrdquoArchives of Phar-macal Research vol 35 no 11 pp 1919ndash1926 2012
[9] J H Lee O S Kwon H-G Jin E-R Woo Y S Kim and H PKim ldquoThe rhizomes of Alisma orientale and alisol derivativesinhibit allergic response and experimental atopic dermatitisrdquoBiological and Pharmaceutical Bulletin vol 35 no 9 pp 1581ndash1587 2012
[10] WGu C GengW Xue et al ldquoCharacterization and function ofthe 3-hydroxy-3-methylglutaryl-CoA reductase gene in Alismaorientale (Sam) Juz and its relationship with protostane triter-pene productionrdquo Plant Physiology and Biochemistry vol 97 pp378ndash389 2015
[11] W Gu B L Xi Q N Wu et al ldquoCloning and distribution pat-tern of HMGR gene conserved fragment in Alisma orientalerdquoJournal of Chinese Medicinal Materials vol 34 no 9 pp 1335ndash1339 2011
[12] T Kobayashi S Kato-Emori K Tomita and H Ezura ldquoDetec-tion of 3-hydroxy-3-methylglutaryl-coenzymeA reductase pro-tein Cm-HMGR during fruit development in melon (Cucumismelo L)rdquo Theoretical and Applied Genetics vol 104 no 5 pp779ndash785 2002
[13] R Tommer A Rachel P C Sylvie and R Jerry ldquoImpaired regu-lation of 3-hydroxy-3-methylglutaryl-coenzyme A reductasedegradation in lovastatin-resistant cellsrdquo The Journal of Bioog-ical Chemistry vol 274 no 41 pp 29341ndash29351 1999
[14] F Xu Q-N Wu J Chen et al ldquoThe binding mechanisms ofplasma protein to active compounds in Alismaorientale rhi-zomes (Alismatis Rhizoma)rdquo Bioorganic and Medicinal Chem-istry Letters vol 24 no 17 pp 4099ndash4105 2014
[15] G Gong Y Qin W Huang et al ldquoProtective effects of dios-genin in the hyperlipidemic rat model and in human vascularendothelial cells against hydrogen peroxide-induced apoptosisrdquoChemico-Biological Interactions vol 184 no 3 pp 366ndash3752010
[16] T F Tzeng H J Lu S S Lion C J Chang and IM Liu ldquoLipid-lowering effects of zerumbone a natural cyclic sesquiterpeneof Zingiber zerumbet Smith in high-fat diet-induced hyperlipi-demic hamstersrdquoFood andChemical Toxicology vol 69 pp 132ndash139 2014
[17] Y Xie H Zhang H Liu et al ldquoHypocholesterolemic effectsof Kluyveromyces marxianus M3 isolated from Tibetan mush-rooms on diet-induced hypercholesterolemia in ratrdquo BrazilianJournal of Microbiology vol 46 no 2 pp 389ndash395 2015
[18] I Karam N Ma X-W Liu et al ldquoRegulation effect of AspirinEugenol Ester on blood lipids in Wistar rats with hyperlipi-demiardquo BMC Veterinary Research vol 11 article 217 2015
[19] G Jiao J B Wei M H Yin and P T Chun ldquoA new TCMformula FTZ lowers serum cholesterol by regulating HMG-CoA reductase and CYP7A1 in hyperlipidemic ratsrdquo Journal ofEthnopharmacology vol 135 no 2 pp 299ndash307 2011
[20] Y Geng W L Tu J J Zhang L Zhang and J Zhang ldquoEffectsof extract from cole pollen on lipidmetabolism in experimentalhyperlipidemic ratsrdquo The Scientific World Journal vol 2014Article ID 982498 6 pages 2014
[21] K Sueyoshi Y Nogawa K Sugawara T Endo and HHisamoto ldquoHighly sensitive and multiple enzyme activity assayusing reagent-release capillary-isoelectric focusing with rhod-amine 110-based substratesrdquo Analytical Sciences vol 31 no 11pp 1155ndash1161 2015
[22] D Fteita E Kononen M Gursoy E Soderling and U KGursoy ldquoDoes estradiol have an impact on the dipeptidylpeptidase IV enzyme activity of the Prevotella intermedia groupbacteriardquo Anaerobe vol 36 pp 14ndash18 2015
[23] L L Zhuo LWang and N Zhou ldquoRandomly arrayed G-quad-ruplexes for label-free and real-time assay of enzyme activityrdquoChemical Communications vol 50 no 52 pp 6875ndash6878 2014
[24] M Garelnabi and A Younis ldquoParaoxonase-1 enzyme activityassay for clinical samples validation and correlation studiesrdquoMedical Science Monitor vol 21 pp 902ndash908 2015
Evidence-Based Complementary and Alternative Medicine 11
[25] X Bi andZ Liu ldquoEnzyme activity assay of glycoprotein enzymesbased on a boronate affinity molecularly imprinted 96-wellmicroplaterdquo Analytical Chemistry vol 86 no 24 pp 12382ndash12389 2014
[26] A Walls S Dermody R Kumaran N Krishnan and E HHarley ldquoCharacterization of B-Cells in tonsils of patients diag-nosed with pediatric autoimmune neuropsychiatric disorderassociated streptococcusrdquo International Journal of PediatricOtorhinolaryngology vol 80 pp 49ndash52 2016
[27] I Karam N Ma X-W Liu et al ldquoRegulation effect of AspirinEugenol Ester on blood lipids in Wistar rats with hyperlipi-demiardquo BMC Veterinary Research vol 11 no 1 article 217 2015
[28] C Sunil S Ignacimuthu and C Kumarappan ldquoHypolipidemicactivity of Symplocos cochinchinensis S Moore leaves in hyper-lipidemic ratsrdquo Journal of Natural Medicines vol 66 no 1 pp32ndash38 2012
[29] W Shen Y Han B Huang et al ldquoMicroRNA-483-3p inhibitsextracellular matrix production by targeting smad4 in humantrabecular meshwork cellsrdquo Investigative Ophthalmology amp Vis-ual Science vol 56 no 13 pp 8419ndash8427 2015
[30] Z Wu Z Yi L Dong and A Zhang ldquoMolecular simulationstudy of the specific combination between four kinds of phthalicacid esters and human serum albuminrdquo Environmental Toxicol-ogy and Pharmacology vol 41 pp 259ndash265 2016
[31] K-J Hwang M Yoon H-J Hwang S-M Choi and S-MJeong ldquoMolecular dynamics simulation of oxygen ion conduc-tion in orthorhombic perovskite Ba-doped LaInO
3using cubic
and orthorhombic modelrdquo Journal of Nanoscience and Nan-otechnology vol 15 no 11 pp 8947ndash8950 2015
[32] H Yu W Han W Ma and K Schulten ldquoTransient 120573-hairpinformation in 120572-synuclein monomer revealed by coarse-grainedmolecular dynamics simulationrdquo The Journal of ChemicalPhysics vol 143 no 24 Article ID 243142 2015
[33] M Breuer K M Rosso and J Blumberger ldquoFlavin binding tothe deca-heme cytochrome mtrc insights from computationalmolecular simulationrdquo Biophysical Journal vol 109 no 12 pp2614ndash2624 2015
[34] V Rastija D Agic S Tomic et al ldquoSynthesis QSAR and mole-cular dynamics simulation of amidino-substituted benzimi-dazoles as dipeptidyl peptidase III inhibitorsrdquo Acta ChimicaSlovenica vol 62 no 4 pp 867ndash878 2015
[35] M Enciso C Schutte and L Delle Site ldquoInfluence of pH andsequence in peptide aggregation via molecular simulationrdquoTheJournal of Chemical Physics vol 143 no 24 Article ID 2431302015
[36] L XWangQWu andG P Peng ldquoA Study on determination ofcontent of 23-acetyl alisol B in Rhizoma Alismatis orientalitisrdquoJournal of Nanjing University of Traditional Chinese Medicine(Natural Science) vol 18 no 2 pp 105ndash107 2002
[37] G P Peng andY L Zhu ldquoProgress in the study on chemical con-stituents of Alisma orientalisrdquo Natural Product Research andDevelopment vol 18 no 2 pp 348ndash351 2006
[38] D Y Liu S J Gu J Wu et al ldquoStudies on fractions of zexiedecoction with antihyperlipidemia bioactivity in micerdquo ChinaPharmacist vol 13 no 6 pp 763ndash766 2010
[39] Q-F Luo L Sun and G-H Du ldquoProgress in new drugs tar-geting reverse cholesterol transportrdquo Chinese PharmacologicalBulletin vol 22 no 8 pp 904ndash907 2006
[40] M Yoshikawa and Y Fukuda ldquoHatakeayamardquo Chemical andPharmaceutical Bulletin vol 41 no 6 pp 1194ndash1196 1993
[41] F Xu L Zhang LHe et al ldquoMolecularmechanismon the inter-action between alisols and human serum albuminrdquo Acta Chim-ica Sinica vol 69 no 19 pp 2228ndash2234 2011
[42] F Xu J ChenW Gu et al ldquoStudy on transformation of effectivecomponents of Alisma under simulated in vivo conditionsrdquoChinese Journal of Pharmaceutical Analysis vol 31 no 11 pp2035ndash2039 2011
[43] H Yang Y G Gao and L X Zhang ldquoResearch progress on bio-synthetic pathway of terpenoids containing ginsenoside and theHMGRrdquo China Journal of Biological Engineering vol 28 no 10pp 130ndash135 2008
[44] G Z Cui Y L Ben N Li and D L Liu ldquoCatalytic mechanismand characteristics of HMG-COA recuctaserdquo Journal ofMedicalMolecular Biology vol 5 no 6 p 542 2008
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom
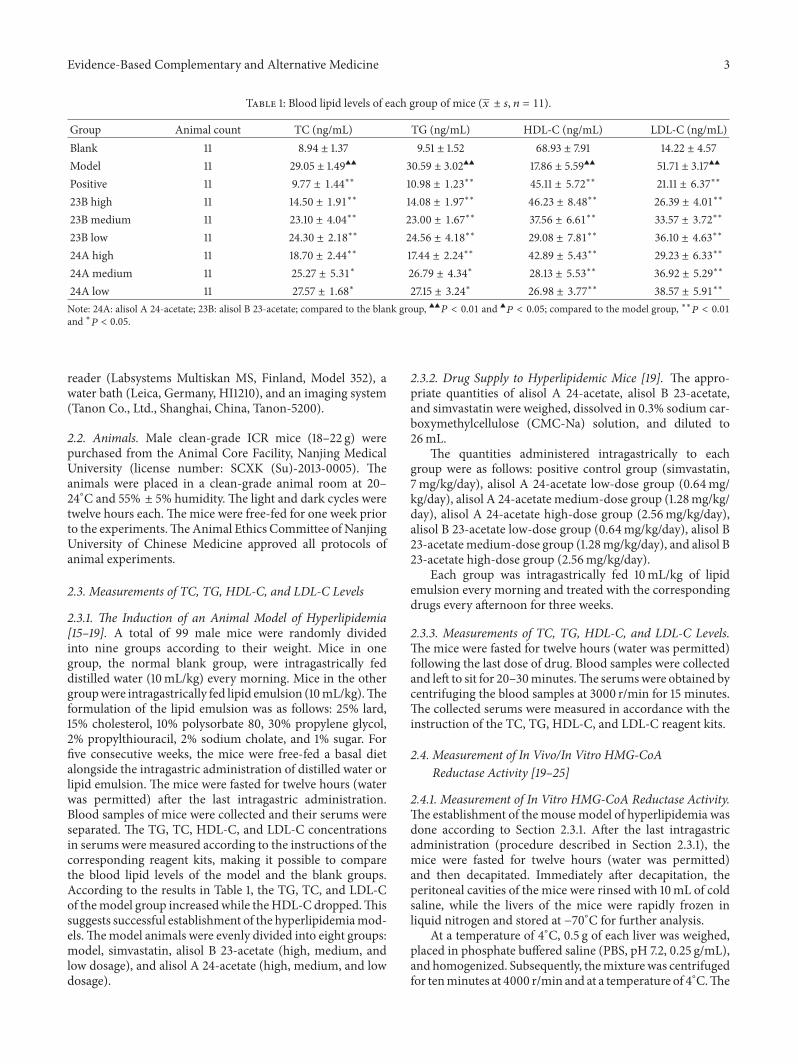
Evidence-Based Complementary and Alternative Medicine 3
Table 1 Blood lipid levels of each group of mice (119909 plusmn 119904 119899 = 11)
Group Animal count TC (ngmL) TG (ngmL) HDL-C (ngmL) LDL-C (ngmL)Blank 11 894 plusmn 137 951 plusmn 152 6893 plusmn 791 1422 plusmn 457Model 11 2905 plusmn 149998771998771 3059 plusmn 302998771998771 1786 plusmn 559998771998771 5171 plusmn 317998771998771
Positive 11 977 plusmn 144lowastlowast 1098 plusmn 123lowastlowast 4511 plusmn 572lowastlowast 2111 plusmn 637lowastlowast
23B high 11 1450 plusmn 191lowastlowast 1408 plusmn 197lowastlowast 4623 plusmn 848lowastlowast 2639 plusmn 401lowastlowast
23B medium 11 2310 plusmn 404lowastlowast 2300 plusmn 167lowastlowast 3756 plusmn 661lowastlowast 3357 plusmn 372lowastlowast
23B low 11 2430 plusmn 218lowastlowast 2456 plusmn 418lowastlowast 2908 plusmn 781lowastlowast 3610 plusmn 463lowastlowast
24A high 11 1870 plusmn 244lowastlowast 1744 plusmn 224lowastlowast 4289 plusmn 543lowastlowast 2923 plusmn 633lowastlowast
24A medium 11 2527 plusmn 531lowast 2679 plusmn 434lowast 2813 plusmn 553lowastlowast 3692 plusmn 529lowastlowast
24A low 11 2757 plusmn 168lowast 2715 plusmn 324lowast 2698 plusmn 377lowastlowast 3857 plusmn 591lowastlowast
Note 24A alisol A 24-acetate 23B alisol B 23-acetate compared to the blank group 998771998771119875 lt 001 and 998771119875 lt 005 compared to the model group lowastlowast119875 lt 001and lowast119875 lt 005
reader (Labsystems Multiskan MS Finland Model 352) awater bath (Leica Germany HI1210) and an imaging system(Tanon Co Ltd Shanghai China Tanon-5200)
22 Animals Male clean-grade ICR mice (18ndash22 g) werepurchased from the Animal Core Facility Nanjing MedicalUniversity (license number SCXK (Su)-2013-0005) Theanimals were placed in a clean-grade animal room at 20ndash24∘C and 55 plusmn 5 humidity The light and dark cycles weretwelve hours eachThe mice were free-fed for one week priorto the experimentsTheAnimal Ethics Committee of NanjingUniversity of Chinese Medicine approved all protocols ofanimal experiments
23 Measurements of TC TG HDL-C and LDL-C Levels
231 The Induction of an Animal Model of Hyperlipidemia[15ndash19] A total of 99 male mice were randomly dividedinto nine groups according to their weight Mice in onegroup the normal blank group were intragastrically feddistilled water (10mLkg) every morning Mice in the othergroupwere intragastrically fed lipid emulsion (10mLkg)Theformulation of the lipid emulsion was as follows 25 lard15 cholesterol 10 polysorbate 80 30 propylene glycol2 propylthiouracil 2 sodium cholate and 1 sugar Forfive consecutive weeks the mice were free-fed a basal dietalongside the intragastric administration of distilled water orlipid emulsion The mice were fasted for twelve hours (waterwas permitted) after the last intragastric administrationBlood samples of mice were collected and their serums wereseparated The TG TC HDL-C and LDL-C concentrationsin serums were measured according to the instructions of thecorresponding reagent kits making it possible to comparethe blood lipid levels of the model and the blank groupsAccording to the results in Table 1 the TG TC and LDL-Cof themodel group increasedwhile theHDL-C droppedThissuggests successful establishment of the hyperlipidemiamod-elsThemodel animals were evenly divided into eight groupsmodel simvastatin alisol B 23-acetate (high medium andlow dosage) and alisol A 24-acetate (high medium and lowdosage)
232 Drug Supply to Hyperlipidemic Mice [19] The appro-priate quantities of alisol A 24-acetate alisol B 23-acetateand simvastatin were weighed dissolved in 03 sodium car-boxymethylcellulose (CMC-Na) solution and diluted to26mL
The quantities administered intragastrically to eachgroup were as follows positive control group (simvastatin7mgkgday) alisol A 24-acetate low-dose group (064mgkgday) alisol A 24-acetate medium-dose group (128mgkgday) alisol A 24-acetate high-dose group (256mgkgday)alisol B 23-acetate low-dose group (064mgkgday) alisol B23-acetatemedium-dose group (128mgkgday) and alisol B23-acetate high-dose group (256mgkgday)
Each group was intragastrically fed 10mLkg of lipidemulsion every morning and treated with the correspondingdrugs every afternoon for three weeks
233 Measurements of TC TG HDL-C and LDL-C LevelsThemice were fasted for twelve hours (water was permitted)following the last dose of drug Blood samples were collectedand left to sit for 20ndash30minutesThe serumswere obtained bycentrifuging the blood samples at 3000 rmin for 15 minutesThe collected serums were measured in accordance with theinstruction of the TC TG HDL-C and LDL-C reagent kits
24 Measurement of In VivoIn Vitro HMG-CoAReductase Activity [19ndash25]
241 Measurement of In Vitro HMG-CoA Reductase ActivityThe establishment of themousemodel of hyperlipidemia wasdone according to Section 231 After the last intragastricadministration (procedure described in Section 231) themice were fasted for twelve hours (water was permitted)and then decapitated Immediately after decapitation theperitoneal cavities of the mice were rinsed with 10mL of coldsaline while the livers of the mice were rapidly frozen inliquid nitrogen and stored at minus70∘C for further analysis
At a temperature of 4∘C 05 g of each liver was weighedplaced in phosphate buffered saline (PBS pH 72 025 gmL)and homogenized Subsequently themixturewas centrifugedfor tenminutes at 4000 rmin and at a temperature of 4∘CThe
4 Evidence-Based Complementary and Alternative Medicine
supernatant was then collected On three separate occasions1mL of the supernatant of the liver homogenate was collectedand extracted with 10mL of 95 ethanol at 60∘C This wasfollowed by vacuum filtration The residues were dissolvedin PBS The samples were adjusted to a certain concentrationand stored at minus70∘C
We added 004mgmL of simvastatin 016mgmL008mgmL and 004mgmL (high medium and lowdosage) of alisol A 24-acetate and alisol B 23-acetate to 5mLof liver homogenate samples respectively After leaving thesamples to stand for 24 hours theHMG-CoA reductase activ-ity was measured using the HMG-CoA reductase activityassay kit in accordance with the instructions
242 Measurement of In Vivo HMG-CoA Reductase ActivityThe induction of the animal model of hyperlipidemia wasdone according to Section 231 The intragastric adminis-tration was performed as described in Section 232 Themice were fasted for twelve hours (water was permitted) anddecapitated after the last dose of the drug The peritonealcavities of the mice were immediately rinsed with 10mL ofcold saline whereas the livers of the mice were rapidly frozenin liquid nitrogen and then stored for further analysis at atemperature of minus70∘C At 4∘C 05 g of each liver were col-lected placed in PBS (pH 72 025 gmL) and homogenizedThe sample was subsequently centrifuged for ten minutes at4000 rmin and at a temperature of 4∘CThe supernatant wasthen collected For each mouse 1mL of liver homogenatesupernatant was extracted with 10mL of 95 ethanol at 60∘Con three occasions This was followed by vacuum filtrationThe residues were dissolved in PBSThe solutionwas adjustedto a certain concentration and stored at minus70∘C The HMG-CoA reductase activity of the liver homogenate samples wasmeasured in accordance with the instructions given in theHMG-CoA reductase activity assay kit
25 Measurement of HMG-CoA Reductase Protein Expres-sion Level via Western Blotting [26ndash29] The hyperlipidemicinduction of animal model of hyperlipidemia was builtaccording to Section 231 The intragastric administrationfollowed the procedure described in Section 232 whereasthe liver homogenate samples were prepared as described inSection 242 The samples were stored at minus70∘C for furtheruse
In order to detect the protein concentration of the super-natant of each sample the A
595 nm absorbance value wasmea-sured via the Bradford method and the calibration curve ofthe bovine serum albumin protein was plotted An equivalentvolume of 2 times sodium dodecyl sulfate (SDS) buffer was addedto the liver homogenate sampleThemixture was denaturizedat 100∘C for three minutes Subsequently the protein was iso-lated using SDS-polyacrylamide gel electrophoresis (PAGE)and electrotransferred to the nitrocellulose membrane Thenitrocellulose membrane was removed and blocked betweenone and two hours in the tris-buffered saline with Tween 20(TBST) solution with 5 skim-milk powder Subsequentlythe membrane was washed with TBST and placed intothe primary antibody (1 2500 rabbit anti-rat HMG-CoAreductase polyclonal antibody) at 4∘C overnight The next
day themembranewas washedwith TBST and blocked in theTBST solution with 5 skim-milk powder for two hoursThemembrane was washed thrice with TBST for 10ndash20 minutesper wash The membrane was incubated for one hour withalkaline phosphatase-conjugated mouse anti-rabbit IgG at adilution of 1 500 Following incubation in the secondaryantibody the membrane was washed again and visualizedusing electrochemiluminescence (ECL) The membrane wasplaced in the antibody elution buffer for ten minutes andrinsed in TBST solution for five minutes It was then blockedin the TBST solution with 5 skim-milk powder and shakenat room temperature for 30 minutes The membrane wasrinsed with TBST repeatedly and then incubated with pri-mary antibody of internal standard GAPDH at a dilution of1 1000 The membrane was subsequently washed and incu-bated with secondary antibody at a dilution of 1 250 Follow-ing washing the membrane was visualized The gray valuesof the filmstrips were measured using a gel imaging ana-lysis system The calculated gray level ratio (HMG-CoAreductaseGAPDH) represents the relative amount of HMG-CoA reductase protein expression
26 Investigation of the Interaction between Alisol Acetatesand HMG-CoA Reductase [30ndash35] A molecular simulationwas conducted The initial molecular structures of the alisolacetates were generated using Discovery Studio 21 (DS21Accelrys US) The structure of HMG-CoA reductase wasobtained from the crystal structure in the protein data bank(PDB) database (PDB code 1HW9) HMG-CoA reductaseis a glycoprotein with N-linked high-mannose oligosaccha-rides consisting of four identical subunits with a molecularweight of 97 ku Each subunit comprised 888 amino acidresidues with the phosphorylation site located at the 872ndamino acid Each subunit featured three different domainsthe N-terminal 339 (1ndash339) amino acids composed the trans-membrane domain the 110 (340ndash449) amino acids composedthe junction domain and the C-terminal 439 (450ndash888)amino acids composed the catalytic domainThe N-terminalcrossed the endoplasmic reticulum membrane eight timesand anchored to the membrane through short rings Thetransmembrane domain was composed of 167 amino acidresidues As the four subunits were identical according to itsinteraction with the drug the A chain was selected as thereceptor for docking simulation Following the removal ofwatermolecules heteroatoms and themulticonformations ofthe amino acid residues in the crystal structure flexible dock-ing in theChemistry atHARvardMacromolecularMechanics(CHARMM) force field was performed Subsequently to thesolvation calculation the compensation ions Na+ and Clminuswere added to the system in order to simulate the environ-ment in the human bodyThemost likely confirmation of theresulting complex was confirmed via minimization calcula-tion
261 Molecular Simulation of Alisol B 23-Acetate and HMG-CoA Reductase The alisol B 23-acetate was defined as aligand After docking the LibDock score the CDockerenergy the CDocker interaction energy and the hydro-gen bond formation of the ligand-receptor complex were
Evidence-Based Complementary and Alternative Medicine 5
comprehensively considered so as to determine the finalsteady conformation This conformation was selected forfurther molecular dynamic simulation
262 Molecular Simulation of Alisol A 24-Acetate andHMG-CoA Reductase Applying the method of Section 261resulted in the final steady conformation of alisol A 24-acetateThis conformationwas selected for furthermoleculardynamic simulation
27 Data Analysis The experimental data were analyzedusing one-way analysis of variance (ANOVA) in SPSS version150 (SPSS Inc Chicago USA) 119875 values of less than 005(119875 lt 005) indicated statistically significant differences Theresults were expressed in the form of mean (119909) plusmn standarddeviation (SD)
3 Results
31 Measurements of TC TG HDL-C and LDL-C LevelsThe blood lipid levels of the blank group positive groupmodel group alisol B 23-acetate high- medium- and low-dose groups and alisol A 24-acetate high- medium- andlow-dosage groups are shown in Table 1 This study revealedthat the TC TG LDL-C and HDL-C levels of the alisol A24-acetate high- medium- and low-dose groups as well asthe alisol B 23-acetate high- medium- and low-dose groupssignificantly differed from those of the model group Thissuggests that alisol acetates could have a lipid-regulatingeffect and significantly decrease the TC TG and LDL-Cconcentrations of hyperlipidemic mice while increasing theHDL-C concentrations
32 Measurement of In VivoIn Vitro HMG-CoAReductase Activity
321 Measurement of In Vitro HMG-CoA Reductase ActivityAs Table 2 indicates alisol acetates lower the HMG-CoAreductase activity dose dependently The extent to whichalisol B 23-acetate lowered HMG-CoA reductase activityexceeded the extent to which alisol A 24-acetate lowered theactivity of this enzyme
322 Measurement of In Vivo HMG-CoA Reductase ActivityThe results in Table 3 illustrate that alisol acetates lower thein vivo HMG-CoA reductase activity in a dose-dependentfashion with a greater degree of enzyme inhibition for alisolB 23-acetate than for alisol A 24-acetate
33 Measurement of HMG-CoA Reductase Protein Expres-sion Level via Western Blotting The western blotting resultssuggest that alisol acetates did not significantly decrease theprotein expression of HMG-CoA reductase (Figure 2)
34 Investigation of the Interaction between the Alisol Acetatesand HMG-CoA Reductase
341 Molecular Simulation Results for Alisol B 23-Acetate andHMG-CoA Reductase The molecular simulation results forthe interaction between alisol B 23-acetate and HMG-CoA
Table 2 The activity of HMG-CoA reductase in vitro (119909 plusmn s 119899 =11)
Group HMG-CoA reductase (UL)Blank 13467 plusmn 3991Positive 16429 plusmn 2832lowastlowast
Model 49788 plusmn 2881998771998771
24A high 19042 plusmn 3092lowastlowast
24A medium 25079 plusmn 1635lowastlowast
24A low 33515 plusmn 2332lowastlowast
23B high 17912 plusmn 4159lowastlowast
23B medium 23079 plusmn 1664lowastlowast
23B low 32759 plusmn 1915lowastlowast
Note 24A alisol A 24-acetate and 23B alisol B 23-acetate compared to theblank group 998771998771119875 lt 001 and 998771119875 lt 005 compared to the model grouplowastlowast119875 lt 001 and lowast119875 lt 005
Table 3The activity of HMG-CoA reductase in vivo (119909 plusmn s 119899 = 11)
Group HMG-CoA reductase (UL)Blank 12345 plusmn 2534Positive 20021 plusmn 3104lowastlowast
Model 49012 plusmn 4117998771998771
24A high 280123 plusmn 2915lowastlowast
24A medium 34554 plusmn 3234lowastlowast
24A low 40131 plusmn 4221lowastlowast
23B high 25203 plusmn 2931lowastlowast
23B medium 29614 plusmn 3218lowastlowast
23B low 39082 plusmn 2323lowastlowast
Note 24A alisol A 24-acetate and 23B alisol B 23-acetate compared to theblank group 998771998771119875 lt 001 and 998771119875 lt 005 compared to the model grouplowastlowast119875 lt 001 and lowast119875 lt 005
reductase are depicted in Figure 3 Figure 3(a) illustratesthe overall pattern of the interaction The interaction pocketwhere alisol B 23-acetate bound to HMG-CoA reductasecontained the following amino acids Leu521 Val522Met523Gly524 Ala525 Cys527 Glu528 Asn529 Val530 Met588Thy589 Arg590 Gly591 Asp653 Ala654 Met655 Gly656Met657 Asn658 Met659 Ile660 Ser661 Lys662 Gly663Thy664 Glu665 Cys688 Asp690 Lys691 Lys692 Arg702Lys704 Ile760 Try761 Ile762Ala763 Cys764Glu801 Ile802Gly803 Thr804 Val805 Gly806 Gly807 Gly808 Thr809Asn810 and Leu811Theywere located in the catalytic domainconstituted by the 439 (450ndash888) amino acids at the C-terminal After entering the interaction pocket alisol B 23-acetate interacted with the peripheral amino acid residues viahydrogen bonding and hydrophobic and hydrophilic effectsThe hydrogen bond graph in Figure 3(b) shows two hydrogenbonds between alisol B 23-acetate and the amino acids Lys691and Asp767 Table 4 presents detailed information aboutthe hydrogen bonding The interaction energy between thehydrophobic amino acid residues and the small molecule wascalculated to be minus128 KJmol while the interaction energybetween hydrophilic amino acid residues and the smallmolecule was calculated to be minus1353 KJmol This indicatesthat the hydrophilic interaction between the small moleculeand the peripheral amino acids was stronger
6 Evidence-Based Complementary and Alternative Medicine
HMG-CoAreductase
GAPDH
05
04
03
02
01
00HM
G-C
oA re
duct
ase
GA
PDH
Blan
k
Mod
el
Posit
ive
24
A-H
24
A-M
24
A-L
23
B-H
23
B-M
23
B-L
Blan
k
Mod
el
Posit
ive
24
A-H
24
A-M
24
A-L
23
B-H
23
B-M
23
B-L
lowast
Figure 2 Results of the protein expression of HMG-CoA reductasein mice liver tissue protein (119899 = 11) Note compared to the blankgroup 998771998771119875 lt 001 and 998771119875 lt 005 compared to the model grouplowastlowast119875 lt 001 and lowast119875 lt 005 24A-H alisol A 24-acetate high 24A-
M alisol A 24-acetate medium and 24A-L alisol A 24-acetate low23B-H alisol B 23-acetate high 23B-M alisol B 23-acetate mediumand 23B-L alisol B 24-acetate low
Table 4 The length (gt) and the angle of hydrogen bond formedbetween alisol acetates and HMG-CoA reductase
Name XndashHsdot sdot sdotYd
(XndashH)
d(Hsdot sdot sdotY)
d(Xsdot sdot sdotY) Angle XHY
Alisol B
23-acetate
Lys691 NZ-HZ1sdot sdot sdotO37 104 237 298 11594
Asp767OD2-
HH88sdot sdot sdotO2096 240 324 14490
Alisol A24-acetate
Asn658 OD1-H5Zsdot sdot sdotO5 095 201 286 14897
After docking the conformation of alisol B 23-acetatechanged as shown in Figure 3(c) The calculated root-mean-square deviation (RMSD) was 289gt The conformationof the small molecule changed from the clustered sheetstructure to the open structure Before docking the sidechain and the ring structure of the small molecule foldedas shown in Figures 3(d) and 3(e) The angle between C
2ndash
C10ndashC19was 1095∘ 1095∘ between C
10ndashC19ndashC20 and minus599∘
between C2ndashC10ndashC19ndashC20 As shown in Figures 3(f) and
3(g) after docking the angle between C2ndashC10ndashC19was 1136∘
1156∘ between C10ndashC19ndashC20 and 1260∘ between C
2ndashC10ndash
C19ndashC20 The angle between the parent ring and the side
chain was significantly increasedThe twisting of the dihedralangles resulted in the opening of side chains and insertioninto the protein which led to interactions There was ahydrophilic ether bond on the side chain and the peripheralamino acids were mainly hydrophilic (Figures 3(h) and 3(i))
This created a hydrophilic environment around the sidechains culminating in stable binding The energy of thevan der Waals forces between them was calculated to beminus1519 KJmol whereas the electrostatic energy was calcu-lated to be minus2366 KJmol As the electrostatic energy waslower than the energy of the van derWaals forces between thesystems the electrostatic interactionwas themain interactionbetween this small molecule and the macromolecule Thebinding energy (ΔG∘) between the alisol B 23-acetate and theHMG-CoA reductase was calculated to be minus1033 KJmol
342 Molecular Simulation Results of Alisol A 24-Acetateand HMG-CoA Reductase Themolecular simulation resultsfor the interaction between alisol A 24-acetate and HMG-CoA reductase are shown in Figure 4 Figure 4(a) shows theoverall interaction pattern of this compound and HMG-CoAreductase The interaction pocket where it bound to HMG-CoA reductase contained the following amino acids Tyr519Ser520 Leu521 Val522 Met523 Gly524 Ala525 Cys527Glu528 Asn529 Val530 Ile531 Gly532 Try533 Met588Thy589 Arg590 Gly591 Asp653 Ala654 Met655 Gly656Met657 Asn658 Met659 Ile660 Ser661 Lys662 Gly663Thy664 Glu665 Cys688 The689 Asp690 Lys691 Lys692Cys764 Gly765 Gln766 Asp767 Ala768 Ala769 Gln770Asn771 Val772 Gly773 Ser774 Glu801 Ile802 Gly803Thr804Val805Gly806Gly807Gly808Thr809 andAsn810There were 439 (450ndash888) amino acids in the catalyticdomain at the C-terminal After entering the interactionpocket alisol A 24-acetate exhibited hydrogen bonding andhydrophobic and hydrophilic interactions with peripheralamino acid residues Figure 4(b) shows the hydrogen bondcontact map There was one hydrogen bond between alisol A24 acetate and amino acid Asn658 (Table 4) The interactionenergy of the hydrophobic amino acid residues and thesmall molecule was calculated to be 465 KJmol whereas theinteraction energy of the hydrophilic amino acid residues andthe small molecule was calculated to be minus1023 KJmol Thissuggests that the hydrophilic groups had a much strongereffect than the hydrophobic groups The small molecule andthe peripheral amino acids exhibited a stronger hydrophilicinteraction as indicated by the interface graphs (Figures 4(f)and 4(g))
After docking the alisol A 24-acetate conformationchanged (Figure 4(c)) The RMSD was calculated to be334gt The conformation angle C
3ndashC18ndashC26ndashC28
changedfrom 600∘ before docking to 1242∘ after docking (Figures4(d) and 4(e)) This suggests a large torsion of the smallmolecule side chain after docking whereby the stretchedopen state changed to a ring-like sheet state Consequentlyit was difficult to insert the side chain into the protein andinteract with the active pocket of the protein This resultedin considerably lower binding relative to alisol B 23-acetateThe electrostatic energy of alisol A 24-acetate and HMG-CoA reductase was determined to be minus886 KJmol whilethe energy of the van der Waals forces was determined tobe minus1649 KJmol The energy of the van der Waals forceswas lower than the electrostatic energy showing that themain interaction between this small molecule and themacro-molecule was the van derWaals interactionThe ΔG∘ of alisol
Evidence-Based Complementary and Alternative Medicine 7
(a) (b) (c)
(d) (e) (f)
(g) (h) (i)
Figure 3 Interaction between alisol B 23-acetate and HMG-CoA reductase (a) Overall pattern of interaction (alisol B 23-acetate is denotedby a bar chart) (b) Hydrogen bond graph of alisol B 23-acetate with Lys691 and Asp767 (the hydrogen bond is denoted by a dotted line) (c)Stacking chart of alisol B 23-acetate before and after docking (the initial structure is shown in green and the structure after docking is shown inpurple) (d) Initial angle of alisol B 23-acetate C
2ndashC10ndashC19and C
10ndashC19ndashC20 (e) Initial dihedral angle of alisol B 23-acetate C
2ndashC10ndashC19ndashC20
(f) The angle of alisol B 23-acetate C2ndashC10ndashC19and C
10ndashC19ndashC20after stacking (g)The dihedral angle of alisol B 23-acetate C
2ndashC10ndashC19ndashC20
after stacking (h) Hydrophobic interface of the alisol B 23-acetate side chain and the peripheral amino acid residues after stacking (alisol B23-acetate is denoted by yellow bars) (i) Hydrophilic interface of the alisol B 23-acetate side chain and the peripheral amino acid residuesafter stacking (alisol B 23-acetate is denoted by yellow bars)
A 24-acetate and HMG-CoA reductase was calculated to beminus23 KJmol
4 Discussion
Various experimental and clinical studies focus on the bloodlipid reducing effects of different solvent extracts (especially
water and alcohol extracts) fromAlisma orientale and its pro-cessed products [36ndash38] However the mechanisms under-lying the effects of Alisma orientale constituents remain to berevealed Alisma orientalemay interfere with exogenous cho-lesterol absorption and endogenous cholesterol metabolismthus reducing the level of TC Moreover alisol acetates mayinterfere with themetabolism of endogenous cholesterol [39]
8 Evidence-Based Complementary and Alternative Medicine
(a) (b) (c)
(d) (e) (f)
(g)
Figure 4 The interaction between alisol A 24-acetate and HMG-CoA reductase (a) Overall pattern of interaction (alisol A 24-acetate isdenoted by a bar chart) (b) Hydrogen bond graph of alisol A 24-acetate and Asn658 (the hydrogen bond is denoted by a dotted line) (c)Stacking chart of alisol A 24-acetate before and after docking (the initial structure is shown in green and the structure after docking is shownin purple) (d) The initial dihedral angle of alisol A 24-acetate angle C
3ndashC18ndashC26ndashC28 (e) The dihedral angle of alisol A 24-acetate angle
C3ndashC18ndashC26ndashC28after stacking (f) Hydrophobic interface of the alisol A 24-acetate side chain and the peripheral amino acid residues after
stacking (alisol A 24-acetate is denoted by yellow bars) (g) Hydrophilic interface of the alisol A 24-acetate side chain and the peripheralamino acid residues after stacking (alisol A 24-acetate is denoted by yellow bars)
Therefore we studied the TC-lowering effect of alisolacetates based on HMG-CoA reductase and the underly-ing molecular mechanism As the main source of endoge-nous cholesterol and one of the key enzymes in the TCmetabolism HMG-CoA reductase is the first described rate-limiting enzyme in cholesterol synthesis in vivo Moreover its
activation directly affects the speed of cholesterol synthesisand the level of TC in vivo
The TC-lowering effect and molecular mechanism ofalisol acetates the main active ingredient of Alismatis rhi-zoma were investigated using the reagent kit method andwestern blotting technique combined with the molecular
Evidence-Based Complementary and Alternative Medicine 9
simulation technique An interaction model was built foralisol acetate and the key enzyme involved in the metabolismof TC was HMG-CoA reductase The present study revealsthat alisol acetates can significantly lower the TC level ofhyperlipidemic miceThe alisol B 23-acetate showed a higherextent of TC lowering than alisol A 24-acetate
Consistent with the in vitro results alisol acetates couldalso lower the in vivoHMG-CoA reductase activity in a dose-dependent fashion with a higher degree of enzyme inhibitionfor alisol B 23-acetate compared to alisol A 24-acetate Theseresults suggest that alisol acetates might lower TC levels viaregulation of HMG-CoA reductase activity The consistencybetween the results of the in vivo and in vitro experimentsindicates that the alisol acetate prototype drug inhibitsHMG-CoA reductase activity instead of the in vivometabolites
Revealed by the western blotting results alisol acetatesmight inhibit the activity of HMG-CoA reductase by directlyand competitively binding to HMG-CoA reductase ratherthan by downregulating its protein expression
The molecular simulation technique was employed inorder to investigate the interaction of alisol acetates andHMG-CoA reductase The results (Figures 3 and 4 andTable 4) revealed themolecularmechanism of the interactionof alisol acetates and HMG-CoA reductase Both alisolacetates bound to HMG-CoA reductase in their catalyticdomain at the C-terminalThe alisol B 23-acetate formed twohydrogen bonds with HMG-CoA reductase The hydrophilicinteraction was stronger than the hydrophobic interactionwhereas the electrostatic interaction was stronger than thevan der Waals forces After the interaction the structure ofthe small molecule dramatically changed from the originalfolded sheet to an open structure Additionally the side chainstretched and was inserted into the protein The peripheralamino acids formed a hydrophilic environment and stablybound with it In general a hydrogen bond with a lengthof less than 35 A and an angle of 90ndash180∘ is consideredto be strong These two hydrogen bonds were relativelystrong and might be partly responsible for the torsion ofthe small molecule The alisol A 24-acetate formed onehydrogen bond with the HMG-CoA reductase and this bondwas weaker compared to those of alisol B 23-acetate Thehydrophilic interaction was stronger than the hydrophobicinteraction and the van derWaals force was stronger than theelectrostatic interaction After the interaction the moleculeside chain had large torsion and changed from the originalstretched open state to a ring-like sheet state which wasdifficult to insert into the proteinThe binding energy of alisolB 23-acetate andHMG-CoA reductase was lower than that ofalisol A 24-acetate and HMG-CoA reductase indicating thatthe binding of alisol B 23-acetate and HMG-CoA reductasewas stable According to the pharmacological results alisolB 23-acetate lowered the HMG-CoA reductase activity morethan alisol A 24-acetate This suggests that the active centerwas the interaction pocket constituted by these amino acidsand that the hydrogen bonds involving Lys691 Asp767 andAsn658 were the key amino acid residues for the interactionof this protein with alisol acetates
The binding of alisol B 23-acetate and HMG-CoA reduc-tase was considerably stronger than the binding of alisol
A 24-acetate and HMG-CoA reductase The relatively weakbinding of alisol A 24-acetate and HMG-CoA reductasemight be due to its distinct side-chain structure The sidechain of alisol B 23-acetate contained an ether bond whichwas strongly hydrophilic and could be inserted into thehydrophilic environment of the protein to form strong inter-actions Previous studies found that the side chains of alisolacetates were their active groups [40] In the present studythis conclusionwas validated A folded side chainparent ringbound more strongly to the macromolecule than an openside-chainparent ring did Thus the side chain acts as thesteering group in that it steers the interactions between alisolacetates and macromolecules This might be the key groupfor interactions with macromolecules as it had a decisiveeffect on the interactions of this type of compound withmacromolecules Our previous studies indicated that alisol B23-acetate was partially converted to alisol A 24-acetate in theenvironment of the human stomach However the bindingof alisol A 24-acetate and HMG-CoA reductase was weaker[14 41 42] Therefore it is advisable for drug developers todevelop a dosage form that circumvents the conversion ofalisol B 23-acetate in the stomach
Previously published studies indicate that [43 44] themechanism of reducing the level of TC is that two acetylCoA molecules condense to acetyl CoA under the catalysisof acetoacetyl-coenzyme A thiolase (AACT) acetyl CoAand acetyl CoA condense to HMG-CoA and HMG-CoAreduces to MVA under the catalytic action of HMG-CoAreductase MVA further generates TC with squalene Thehigh intracellular free fatty acid (FC) level stimulates micro-somal acyl coenzyme A-cholesterol acyltransferase (ACAT)and promotes esterification of the cholesterol Subsequentlycholesterol is stored in the form of cholesterol esters (CE)which is a feedback inhibition of HMG-CoA reductasesynthesis so that cholesterol synthesis rate-limiting enzymeswill be inhibited thus reducing intracellular cholesterolsynthesis This process features three important enzymesAACT HMG-CoA reductase and ACAT Whether AACTand ACAT are involved in the effect of alisol acetates will bethe subject of intensive research in the future
The results of the study indicate that alisol acetates cannot only decrease TC level but also decrease TG and LDL-C levels and increase HDL-C level Furthermore a follow-upstudy is designed to investigate the regulatory mechanisms ofalisol acetates on TG HDL-C and LDL-C
5 Conclusion
The present study reveals that alisol acetates could signifi-cantly lower the TC level of hyperlipidemic mice The effectof alisol B 23-acetate on HMG-CoA reductase was strongerthan that of alisol A 24-acetate Alisol acetates could lowerHMG-CoA reductase activity in a dose-dependent fashionboth in vivo and in vitro Neither of these alisol acetatescould significantly lower the protein expression of HMG-CoA This suggests that alisol acetates lower the TC levelvia inhibiting the activity of HMG-CoA reductase via itsprototype drug which may have an inhibitory effect bydirectly and competitively binding with HMG-CoA In the
10 Evidence-Based Complementary and Alternative Medicine
binding interaction of alisol acetate andHMG-CoA reductasevia molecular simulation the key amino acid residues ofHMG-CoA reductase might be Lys691 Asp767 and Asn658The side chain of the alisol acetatewas the steering group Fur-thermore a folded side chainparent ring boundmoreweaklyto HMG-CoA reductase than an open side chainparent ringThis study explored the TC-lowering mechanism of alisolacetates at the molecular level and presented guidance for itsclinical use
Competing Interests
The authors declare that they have no competing interests
Acknowledgments
This work was financially supported by the National NatureScience Foundation of China (81303173 81673534) the Nat-ural Science Foundation of Jiangsu Province (BK20161576)the Natural Science Fund for Colleges and Universities inJiangsu Province (14KJB360001) and the Top-Notch Aca-demic Programs Project of Jiangsu Higher Education Insti-tutions (PPZY2015A070)
References
[1] L X Wang Q N Wu Q Zhang G P Peng and A W DingldquoBasic study of diuretic active compounds in Rhizoma Alis-matisrdquo West China Journal of Pharmaceutical Sciences vol 23no 6 pp 670ndash672 2008
[2] S S Wu Pharmaceutical and Clinical Research of Alisma Ori-entalis Chinese Press Beijing China 2007
[3] M Miyazawa S Yoshinaga and Y Kashima ldquoChemical com-position and characteristic odor compounds in essential oilfrom Alismatis Rhizoma (Tubers of Alisma orientale)rdquo DiatomResearch vol 23 no 1 pp 133ndash141 2016
[4] M-K Jang Y-R Han J S Nam et al ldquoProtective effects ofAlisma orientale extract against hepatic steatosis via inhibitionof endoplasmic reticulum stressrdquo International Journal ofMolec-ular Sciences vol 16 no 11 pp 26151ndash26165 2015
[5] Z-P Mai K Zhou G-B Ge et al ldquoProtostane triterpenoidsfrom the rhizome of Alisma orientale exhibit inhibitory effectson human carboxylesterase 2rdquo Journal of Natural Products vol78 no 10 pp 2372ndash2380 2015
[6] T Tian H Chen andY-Y Zhao ldquoTraditional uses phytochem-istry pharmacology toxicology and quality control of Alismaorientale (Sam) Juzep a reviewrdquo Journal of Ethnopharmacologyvol 158 pp 373ndash387 2014
[7] KHKimM J Kwun J-Y Choi et al ldquoTherapeutic effect of thetuber of Alisma orientale on lipopolysaccharide-induced acutelung injuryrdquo Evidence-Based Complementary and AlternativeMedicine vol 2013 Article ID 863892 10 pages 2013
[8] H-G Jin Q Jin A R Kim et al ldquoA new triterpenoid fromAlisma orientale and their antibacterial effectrdquoArchives of Phar-macal Research vol 35 no 11 pp 1919ndash1926 2012
[9] J H Lee O S Kwon H-G Jin E-R Woo Y S Kim and H PKim ldquoThe rhizomes of Alisma orientale and alisol derivativesinhibit allergic response and experimental atopic dermatitisrdquoBiological and Pharmaceutical Bulletin vol 35 no 9 pp 1581ndash1587 2012
[10] WGu C GengW Xue et al ldquoCharacterization and function ofthe 3-hydroxy-3-methylglutaryl-CoA reductase gene in Alismaorientale (Sam) Juz and its relationship with protostane triter-pene productionrdquo Plant Physiology and Biochemistry vol 97 pp378ndash389 2015
[11] W Gu B L Xi Q N Wu et al ldquoCloning and distribution pat-tern of HMGR gene conserved fragment in Alisma orientalerdquoJournal of Chinese Medicinal Materials vol 34 no 9 pp 1335ndash1339 2011
[12] T Kobayashi S Kato-Emori K Tomita and H Ezura ldquoDetec-tion of 3-hydroxy-3-methylglutaryl-coenzymeA reductase pro-tein Cm-HMGR during fruit development in melon (Cucumismelo L)rdquo Theoretical and Applied Genetics vol 104 no 5 pp779ndash785 2002
[13] R Tommer A Rachel P C Sylvie and R Jerry ldquoImpaired regu-lation of 3-hydroxy-3-methylglutaryl-coenzyme A reductasedegradation in lovastatin-resistant cellsrdquo The Journal of Bioog-ical Chemistry vol 274 no 41 pp 29341ndash29351 1999
[14] F Xu Q-N Wu J Chen et al ldquoThe binding mechanisms ofplasma protein to active compounds in Alismaorientale rhi-zomes (Alismatis Rhizoma)rdquo Bioorganic and Medicinal Chem-istry Letters vol 24 no 17 pp 4099ndash4105 2014
[15] G Gong Y Qin W Huang et al ldquoProtective effects of dios-genin in the hyperlipidemic rat model and in human vascularendothelial cells against hydrogen peroxide-induced apoptosisrdquoChemico-Biological Interactions vol 184 no 3 pp 366ndash3752010
[16] T F Tzeng H J Lu S S Lion C J Chang and IM Liu ldquoLipid-lowering effects of zerumbone a natural cyclic sesquiterpeneof Zingiber zerumbet Smith in high-fat diet-induced hyperlipi-demic hamstersrdquoFood andChemical Toxicology vol 69 pp 132ndash139 2014
[17] Y Xie H Zhang H Liu et al ldquoHypocholesterolemic effectsof Kluyveromyces marxianus M3 isolated from Tibetan mush-rooms on diet-induced hypercholesterolemia in ratrdquo BrazilianJournal of Microbiology vol 46 no 2 pp 389ndash395 2015
[18] I Karam N Ma X-W Liu et al ldquoRegulation effect of AspirinEugenol Ester on blood lipids in Wistar rats with hyperlipi-demiardquo BMC Veterinary Research vol 11 article 217 2015
[19] G Jiao J B Wei M H Yin and P T Chun ldquoA new TCMformula FTZ lowers serum cholesterol by regulating HMG-CoA reductase and CYP7A1 in hyperlipidemic ratsrdquo Journal ofEthnopharmacology vol 135 no 2 pp 299ndash307 2011
[20] Y Geng W L Tu J J Zhang L Zhang and J Zhang ldquoEffectsof extract from cole pollen on lipidmetabolism in experimentalhyperlipidemic ratsrdquo The Scientific World Journal vol 2014Article ID 982498 6 pages 2014
[21] K Sueyoshi Y Nogawa K Sugawara T Endo and HHisamoto ldquoHighly sensitive and multiple enzyme activity assayusing reagent-release capillary-isoelectric focusing with rhod-amine 110-based substratesrdquo Analytical Sciences vol 31 no 11pp 1155ndash1161 2015
[22] D Fteita E Kononen M Gursoy E Soderling and U KGursoy ldquoDoes estradiol have an impact on the dipeptidylpeptidase IV enzyme activity of the Prevotella intermedia groupbacteriardquo Anaerobe vol 36 pp 14ndash18 2015
[23] L L Zhuo LWang and N Zhou ldquoRandomly arrayed G-quad-ruplexes for label-free and real-time assay of enzyme activityrdquoChemical Communications vol 50 no 52 pp 6875ndash6878 2014
[24] M Garelnabi and A Younis ldquoParaoxonase-1 enzyme activityassay for clinical samples validation and correlation studiesrdquoMedical Science Monitor vol 21 pp 902ndash908 2015
Evidence-Based Complementary and Alternative Medicine 11
[25] X Bi andZ Liu ldquoEnzyme activity assay of glycoprotein enzymesbased on a boronate affinity molecularly imprinted 96-wellmicroplaterdquo Analytical Chemistry vol 86 no 24 pp 12382ndash12389 2014
[26] A Walls S Dermody R Kumaran N Krishnan and E HHarley ldquoCharacterization of B-Cells in tonsils of patients diag-nosed with pediatric autoimmune neuropsychiatric disorderassociated streptococcusrdquo International Journal of PediatricOtorhinolaryngology vol 80 pp 49ndash52 2016
[27] I Karam N Ma X-W Liu et al ldquoRegulation effect of AspirinEugenol Ester on blood lipids in Wistar rats with hyperlipi-demiardquo BMC Veterinary Research vol 11 no 1 article 217 2015
[28] C Sunil S Ignacimuthu and C Kumarappan ldquoHypolipidemicactivity of Symplocos cochinchinensis S Moore leaves in hyper-lipidemic ratsrdquo Journal of Natural Medicines vol 66 no 1 pp32ndash38 2012
[29] W Shen Y Han B Huang et al ldquoMicroRNA-483-3p inhibitsextracellular matrix production by targeting smad4 in humantrabecular meshwork cellsrdquo Investigative Ophthalmology amp Vis-ual Science vol 56 no 13 pp 8419ndash8427 2015
[30] Z Wu Z Yi L Dong and A Zhang ldquoMolecular simulationstudy of the specific combination between four kinds of phthalicacid esters and human serum albuminrdquo Environmental Toxicol-ogy and Pharmacology vol 41 pp 259ndash265 2016
[31] K-J Hwang M Yoon H-J Hwang S-M Choi and S-MJeong ldquoMolecular dynamics simulation of oxygen ion conduc-tion in orthorhombic perovskite Ba-doped LaInO
3using cubic
and orthorhombic modelrdquo Journal of Nanoscience and Nan-otechnology vol 15 no 11 pp 8947ndash8950 2015
[32] H Yu W Han W Ma and K Schulten ldquoTransient 120573-hairpinformation in 120572-synuclein monomer revealed by coarse-grainedmolecular dynamics simulationrdquo The Journal of ChemicalPhysics vol 143 no 24 Article ID 243142 2015
[33] M Breuer K M Rosso and J Blumberger ldquoFlavin binding tothe deca-heme cytochrome mtrc insights from computationalmolecular simulationrdquo Biophysical Journal vol 109 no 12 pp2614ndash2624 2015
[34] V Rastija D Agic S Tomic et al ldquoSynthesis QSAR and mole-cular dynamics simulation of amidino-substituted benzimi-dazoles as dipeptidyl peptidase III inhibitorsrdquo Acta ChimicaSlovenica vol 62 no 4 pp 867ndash878 2015
[35] M Enciso C Schutte and L Delle Site ldquoInfluence of pH andsequence in peptide aggregation via molecular simulationrdquoTheJournal of Chemical Physics vol 143 no 24 Article ID 2431302015
[36] L XWangQWu andG P Peng ldquoA Study on determination ofcontent of 23-acetyl alisol B in Rhizoma Alismatis orientalitisrdquoJournal of Nanjing University of Traditional Chinese Medicine(Natural Science) vol 18 no 2 pp 105ndash107 2002
[37] G P Peng andY L Zhu ldquoProgress in the study on chemical con-stituents of Alisma orientalisrdquo Natural Product Research andDevelopment vol 18 no 2 pp 348ndash351 2006
[38] D Y Liu S J Gu J Wu et al ldquoStudies on fractions of zexiedecoction with antihyperlipidemia bioactivity in micerdquo ChinaPharmacist vol 13 no 6 pp 763ndash766 2010
[39] Q-F Luo L Sun and G-H Du ldquoProgress in new drugs tar-geting reverse cholesterol transportrdquo Chinese PharmacologicalBulletin vol 22 no 8 pp 904ndash907 2006
[40] M Yoshikawa and Y Fukuda ldquoHatakeayamardquo Chemical andPharmaceutical Bulletin vol 41 no 6 pp 1194ndash1196 1993
[41] F Xu L Zhang LHe et al ldquoMolecularmechanismon the inter-action between alisols and human serum albuminrdquo Acta Chim-ica Sinica vol 69 no 19 pp 2228ndash2234 2011
[42] F Xu J ChenW Gu et al ldquoStudy on transformation of effectivecomponents of Alisma under simulated in vivo conditionsrdquoChinese Journal of Pharmaceutical Analysis vol 31 no 11 pp2035ndash2039 2011
[43] H Yang Y G Gao and L X Zhang ldquoResearch progress on bio-synthetic pathway of terpenoids containing ginsenoside and theHMGRrdquo China Journal of Biological Engineering vol 28 no 10pp 130ndash135 2008
[44] G Z Cui Y L Ben N Li and D L Liu ldquoCatalytic mechanismand characteristics of HMG-COA recuctaserdquo Journal ofMedicalMolecular Biology vol 5 no 6 p 542 2008
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom

4 Evidence-Based Complementary and Alternative Medicine
supernatant was then collected On three separate occasions1mL of the supernatant of the liver homogenate was collectedand extracted with 10mL of 95 ethanol at 60∘C This wasfollowed by vacuum filtration The residues were dissolvedin PBS The samples were adjusted to a certain concentrationand stored at minus70∘C
We added 004mgmL of simvastatin 016mgmL008mgmL and 004mgmL (high medium and lowdosage) of alisol A 24-acetate and alisol B 23-acetate to 5mLof liver homogenate samples respectively After leaving thesamples to stand for 24 hours theHMG-CoA reductase activ-ity was measured using the HMG-CoA reductase activityassay kit in accordance with the instructions
242 Measurement of In Vivo HMG-CoA Reductase ActivityThe induction of the animal model of hyperlipidemia wasdone according to Section 231 The intragastric adminis-tration was performed as described in Section 232 Themice were fasted for twelve hours (water was permitted) anddecapitated after the last dose of the drug The peritonealcavities of the mice were immediately rinsed with 10mL ofcold saline whereas the livers of the mice were rapidly frozenin liquid nitrogen and then stored for further analysis at atemperature of minus70∘C At 4∘C 05 g of each liver were col-lected placed in PBS (pH 72 025 gmL) and homogenizedThe sample was subsequently centrifuged for ten minutes at4000 rmin and at a temperature of 4∘CThe supernatant wasthen collected For each mouse 1mL of liver homogenatesupernatant was extracted with 10mL of 95 ethanol at 60∘Con three occasions This was followed by vacuum filtrationThe residues were dissolved in PBSThe solutionwas adjustedto a certain concentration and stored at minus70∘C The HMG-CoA reductase activity of the liver homogenate samples wasmeasured in accordance with the instructions given in theHMG-CoA reductase activity assay kit
25 Measurement of HMG-CoA Reductase Protein Expres-sion Level via Western Blotting [26ndash29] The hyperlipidemicinduction of animal model of hyperlipidemia was builtaccording to Section 231 The intragastric administrationfollowed the procedure described in Section 232 whereasthe liver homogenate samples were prepared as described inSection 242 The samples were stored at minus70∘C for furtheruse
In order to detect the protein concentration of the super-natant of each sample the A
595 nm absorbance value wasmea-sured via the Bradford method and the calibration curve ofthe bovine serum albumin protein was plotted An equivalentvolume of 2 times sodium dodecyl sulfate (SDS) buffer was addedto the liver homogenate sampleThemixture was denaturizedat 100∘C for three minutes Subsequently the protein was iso-lated using SDS-polyacrylamide gel electrophoresis (PAGE)and electrotransferred to the nitrocellulose membrane Thenitrocellulose membrane was removed and blocked betweenone and two hours in the tris-buffered saline with Tween 20(TBST) solution with 5 skim-milk powder Subsequentlythe membrane was washed with TBST and placed intothe primary antibody (1 2500 rabbit anti-rat HMG-CoAreductase polyclonal antibody) at 4∘C overnight The next
day themembranewas washedwith TBST and blocked in theTBST solution with 5 skim-milk powder for two hoursThemembrane was washed thrice with TBST for 10ndash20 minutesper wash The membrane was incubated for one hour withalkaline phosphatase-conjugated mouse anti-rabbit IgG at adilution of 1 500 Following incubation in the secondaryantibody the membrane was washed again and visualizedusing electrochemiluminescence (ECL) The membrane wasplaced in the antibody elution buffer for ten minutes andrinsed in TBST solution for five minutes It was then blockedin the TBST solution with 5 skim-milk powder and shakenat room temperature for 30 minutes The membrane wasrinsed with TBST repeatedly and then incubated with pri-mary antibody of internal standard GAPDH at a dilution of1 1000 The membrane was subsequently washed and incu-bated with secondary antibody at a dilution of 1 250 Follow-ing washing the membrane was visualized The gray valuesof the filmstrips were measured using a gel imaging ana-lysis system The calculated gray level ratio (HMG-CoAreductaseGAPDH) represents the relative amount of HMG-CoA reductase protein expression
26 Investigation of the Interaction between Alisol Acetatesand HMG-CoA Reductase [30ndash35] A molecular simulationwas conducted The initial molecular structures of the alisolacetates were generated using Discovery Studio 21 (DS21Accelrys US) The structure of HMG-CoA reductase wasobtained from the crystal structure in the protein data bank(PDB) database (PDB code 1HW9) HMG-CoA reductaseis a glycoprotein with N-linked high-mannose oligosaccha-rides consisting of four identical subunits with a molecularweight of 97 ku Each subunit comprised 888 amino acidresidues with the phosphorylation site located at the 872ndamino acid Each subunit featured three different domainsthe N-terminal 339 (1ndash339) amino acids composed the trans-membrane domain the 110 (340ndash449) amino acids composedthe junction domain and the C-terminal 439 (450ndash888)amino acids composed the catalytic domainThe N-terminalcrossed the endoplasmic reticulum membrane eight timesand anchored to the membrane through short rings Thetransmembrane domain was composed of 167 amino acidresidues As the four subunits were identical according to itsinteraction with the drug the A chain was selected as thereceptor for docking simulation Following the removal ofwatermolecules heteroatoms and themulticonformations ofthe amino acid residues in the crystal structure flexible dock-ing in theChemistry atHARvardMacromolecularMechanics(CHARMM) force field was performed Subsequently to thesolvation calculation the compensation ions Na+ and Clminuswere added to the system in order to simulate the environ-ment in the human bodyThemost likely confirmation of theresulting complex was confirmed via minimization calcula-tion
261 Molecular Simulation of Alisol B 23-Acetate and HMG-CoA Reductase The alisol B 23-acetate was defined as aligand After docking the LibDock score the CDockerenergy the CDocker interaction energy and the hydro-gen bond formation of the ligand-receptor complex were
Evidence-Based Complementary and Alternative Medicine 5
comprehensively considered so as to determine the finalsteady conformation This conformation was selected forfurther molecular dynamic simulation
262 Molecular Simulation of Alisol A 24-Acetate andHMG-CoA Reductase Applying the method of Section 261resulted in the final steady conformation of alisol A 24-acetateThis conformationwas selected for furthermoleculardynamic simulation
27 Data Analysis The experimental data were analyzedusing one-way analysis of variance (ANOVA) in SPSS version150 (SPSS Inc Chicago USA) 119875 values of less than 005(119875 lt 005) indicated statistically significant differences Theresults were expressed in the form of mean (119909) plusmn standarddeviation (SD)
3 Results
31 Measurements of TC TG HDL-C and LDL-C LevelsThe blood lipid levels of the blank group positive groupmodel group alisol B 23-acetate high- medium- and low-dose groups and alisol A 24-acetate high- medium- andlow-dosage groups are shown in Table 1 This study revealedthat the TC TG LDL-C and HDL-C levels of the alisol A24-acetate high- medium- and low-dose groups as well asthe alisol B 23-acetate high- medium- and low-dose groupssignificantly differed from those of the model group Thissuggests that alisol acetates could have a lipid-regulatingeffect and significantly decrease the TC TG and LDL-Cconcentrations of hyperlipidemic mice while increasing theHDL-C concentrations
32 Measurement of In VivoIn Vitro HMG-CoAReductase Activity
321 Measurement of In Vitro HMG-CoA Reductase ActivityAs Table 2 indicates alisol acetates lower the HMG-CoAreductase activity dose dependently The extent to whichalisol B 23-acetate lowered HMG-CoA reductase activityexceeded the extent to which alisol A 24-acetate lowered theactivity of this enzyme
322 Measurement of In Vivo HMG-CoA Reductase ActivityThe results in Table 3 illustrate that alisol acetates lower thein vivo HMG-CoA reductase activity in a dose-dependentfashion with a greater degree of enzyme inhibition for alisolB 23-acetate than for alisol A 24-acetate
33 Measurement of HMG-CoA Reductase Protein Expres-sion Level via Western Blotting The western blotting resultssuggest that alisol acetates did not significantly decrease theprotein expression of HMG-CoA reductase (Figure 2)
34 Investigation of the Interaction between the Alisol Acetatesand HMG-CoA Reductase
341 Molecular Simulation Results for Alisol B 23-Acetate andHMG-CoA Reductase The molecular simulation results forthe interaction between alisol B 23-acetate and HMG-CoA
Table 2 The activity of HMG-CoA reductase in vitro (119909 plusmn s 119899 =11)
Group HMG-CoA reductase (UL)Blank 13467 plusmn 3991Positive 16429 plusmn 2832lowastlowast
Model 49788 plusmn 2881998771998771
24A high 19042 plusmn 3092lowastlowast
24A medium 25079 plusmn 1635lowastlowast
24A low 33515 plusmn 2332lowastlowast
23B high 17912 plusmn 4159lowastlowast
23B medium 23079 plusmn 1664lowastlowast
23B low 32759 plusmn 1915lowastlowast
Note 24A alisol A 24-acetate and 23B alisol B 23-acetate compared to theblank group 998771998771119875 lt 001 and 998771119875 lt 005 compared to the model grouplowastlowast119875 lt 001 and lowast119875 lt 005
Table 3The activity of HMG-CoA reductase in vivo (119909 plusmn s 119899 = 11)
Group HMG-CoA reductase (UL)Blank 12345 plusmn 2534Positive 20021 plusmn 3104lowastlowast
Model 49012 plusmn 4117998771998771
24A high 280123 plusmn 2915lowastlowast
24A medium 34554 plusmn 3234lowastlowast
24A low 40131 plusmn 4221lowastlowast
23B high 25203 plusmn 2931lowastlowast
23B medium 29614 plusmn 3218lowastlowast
23B low 39082 plusmn 2323lowastlowast
Note 24A alisol A 24-acetate and 23B alisol B 23-acetate compared to theblank group 998771998771119875 lt 001 and 998771119875 lt 005 compared to the model grouplowastlowast119875 lt 001 and lowast119875 lt 005
reductase are depicted in Figure 3 Figure 3(a) illustratesthe overall pattern of the interaction The interaction pocketwhere alisol B 23-acetate bound to HMG-CoA reductasecontained the following amino acids Leu521 Val522Met523Gly524 Ala525 Cys527 Glu528 Asn529 Val530 Met588Thy589 Arg590 Gly591 Asp653 Ala654 Met655 Gly656Met657 Asn658 Met659 Ile660 Ser661 Lys662 Gly663Thy664 Glu665 Cys688 Asp690 Lys691 Lys692 Arg702Lys704 Ile760 Try761 Ile762Ala763 Cys764Glu801 Ile802Gly803 Thr804 Val805 Gly806 Gly807 Gly808 Thr809Asn810 and Leu811Theywere located in the catalytic domainconstituted by the 439 (450ndash888) amino acids at the C-terminal After entering the interaction pocket alisol B 23-acetate interacted with the peripheral amino acid residues viahydrogen bonding and hydrophobic and hydrophilic effectsThe hydrogen bond graph in Figure 3(b) shows two hydrogenbonds between alisol B 23-acetate and the amino acids Lys691and Asp767 Table 4 presents detailed information aboutthe hydrogen bonding The interaction energy between thehydrophobic amino acid residues and the small molecule wascalculated to be minus128 KJmol while the interaction energybetween hydrophilic amino acid residues and the smallmolecule was calculated to be minus1353 KJmol This indicatesthat the hydrophilic interaction between the small moleculeand the peripheral amino acids was stronger
6 Evidence-Based Complementary and Alternative Medicine
HMG-CoAreductase
GAPDH
05
04
03
02
01
00HM
G-C
oA re
duct
ase
GA
PDH
Blan
k
Mod
el
Posit
ive
24
A-H
24
A-M
24
A-L
23
B-H
23
B-M
23
B-L
Blan
k
Mod
el
Posit
ive
24
A-H
24
A-M
24
A-L
23
B-H
23
B-M
23
B-L
lowast
Figure 2 Results of the protein expression of HMG-CoA reductasein mice liver tissue protein (119899 = 11) Note compared to the blankgroup 998771998771119875 lt 001 and 998771119875 lt 005 compared to the model grouplowastlowast119875 lt 001 and lowast119875 lt 005 24A-H alisol A 24-acetate high 24A-
M alisol A 24-acetate medium and 24A-L alisol A 24-acetate low23B-H alisol B 23-acetate high 23B-M alisol B 23-acetate mediumand 23B-L alisol B 24-acetate low
Table 4 The length (gt) and the angle of hydrogen bond formedbetween alisol acetates and HMG-CoA reductase
Name XndashHsdot sdot sdotYd
(XndashH)
d(Hsdot sdot sdotY)
d(Xsdot sdot sdotY) Angle XHY
Alisol B
23-acetate
Lys691 NZ-HZ1sdot sdot sdotO37 104 237 298 11594
Asp767OD2-
HH88sdot sdot sdotO2096 240 324 14490
Alisol A24-acetate
Asn658 OD1-H5Zsdot sdot sdotO5 095 201 286 14897
After docking the conformation of alisol B 23-acetatechanged as shown in Figure 3(c) The calculated root-mean-square deviation (RMSD) was 289gt The conformationof the small molecule changed from the clustered sheetstructure to the open structure Before docking the sidechain and the ring structure of the small molecule foldedas shown in Figures 3(d) and 3(e) The angle between C
2ndash
C10ndashC19was 1095∘ 1095∘ between C
10ndashC19ndashC20 and minus599∘
between C2ndashC10ndashC19ndashC20 As shown in Figures 3(f) and
3(g) after docking the angle between C2ndashC10ndashC19was 1136∘
1156∘ between C10ndashC19ndashC20 and 1260∘ between C
2ndashC10ndash
C19ndashC20 The angle between the parent ring and the side
chain was significantly increasedThe twisting of the dihedralangles resulted in the opening of side chains and insertioninto the protein which led to interactions There was ahydrophilic ether bond on the side chain and the peripheralamino acids were mainly hydrophilic (Figures 3(h) and 3(i))
This created a hydrophilic environment around the sidechains culminating in stable binding The energy of thevan der Waals forces between them was calculated to beminus1519 KJmol whereas the electrostatic energy was calcu-lated to be minus2366 KJmol As the electrostatic energy waslower than the energy of the van derWaals forces between thesystems the electrostatic interactionwas themain interactionbetween this small molecule and the macromolecule Thebinding energy (ΔG∘) between the alisol B 23-acetate and theHMG-CoA reductase was calculated to be minus1033 KJmol
342 Molecular Simulation Results of Alisol A 24-Acetateand HMG-CoA Reductase Themolecular simulation resultsfor the interaction between alisol A 24-acetate and HMG-CoA reductase are shown in Figure 4 Figure 4(a) shows theoverall interaction pattern of this compound and HMG-CoAreductase The interaction pocket where it bound to HMG-CoA reductase contained the following amino acids Tyr519Ser520 Leu521 Val522 Met523 Gly524 Ala525 Cys527Glu528 Asn529 Val530 Ile531 Gly532 Try533 Met588Thy589 Arg590 Gly591 Asp653 Ala654 Met655 Gly656Met657 Asn658 Met659 Ile660 Ser661 Lys662 Gly663Thy664 Glu665 Cys688 The689 Asp690 Lys691 Lys692Cys764 Gly765 Gln766 Asp767 Ala768 Ala769 Gln770Asn771 Val772 Gly773 Ser774 Glu801 Ile802 Gly803Thr804Val805Gly806Gly807Gly808Thr809 andAsn810There were 439 (450ndash888) amino acids in the catalyticdomain at the C-terminal After entering the interactionpocket alisol A 24-acetate exhibited hydrogen bonding andhydrophobic and hydrophilic interactions with peripheralamino acid residues Figure 4(b) shows the hydrogen bondcontact map There was one hydrogen bond between alisol A24 acetate and amino acid Asn658 (Table 4) The interactionenergy of the hydrophobic amino acid residues and thesmall molecule was calculated to be 465 KJmol whereas theinteraction energy of the hydrophilic amino acid residues andthe small molecule was calculated to be minus1023 KJmol Thissuggests that the hydrophilic groups had a much strongereffect than the hydrophobic groups The small molecule andthe peripheral amino acids exhibited a stronger hydrophilicinteraction as indicated by the interface graphs (Figures 4(f)and 4(g))
After docking the alisol A 24-acetate conformationchanged (Figure 4(c)) The RMSD was calculated to be334gt The conformation angle C
3ndashC18ndashC26ndashC28
changedfrom 600∘ before docking to 1242∘ after docking (Figures4(d) and 4(e)) This suggests a large torsion of the smallmolecule side chain after docking whereby the stretchedopen state changed to a ring-like sheet state Consequentlyit was difficult to insert the side chain into the protein andinteract with the active pocket of the protein This resultedin considerably lower binding relative to alisol B 23-acetateThe electrostatic energy of alisol A 24-acetate and HMG-CoA reductase was determined to be minus886 KJmol whilethe energy of the van der Waals forces was determined tobe minus1649 KJmol The energy of the van der Waals forceswas lower than the electrostatic energy showing that themain interaction between this small molecule and themacro-molecule was the van derWaals interactionThe ΔG∘ of alisol
Evidence-Based Complementary and Alternative Medicine 7
(a) (b) (c)
(d) (e) (f)
(g) (h) (i)
Figure 3 Interaction between alisol B 23-acetate and HMG-CoA reductase (a) Overall pattern of interaction (alisol B 23-acetate is denotedby a bar chart) (b) Hydrogen bond graph of alisol B 23-acetate with Lys691 and Asp767 (the hydrogen bond is denoted by a dotted line) (c)Stacking chart of alisol B 23-acetate before and after docking (the initial structure is shown in green and the structure after docking is shown inpurple) (d) Initial angle of alisol B 23-acetate C
2ndashC10ndashC19and C
10ndashC19ndashC20 (e) Initial dihedral angle of alisol B 23-acetate C
2ndashC10ndashC19ndashC20
(f) The angle of alisol B 23-acetate C2ndashC10ndashC19and C
10ndashC19ndashC20after stacking (g)The dihedral angle of alisol B 23-acetate C
2ndashC10ndashC19ndashC20
after stacking (h) Hydrophobic interface of the alisol B 23-acetate side chain and the peripheral amino acid residues after stacking (alisol B23-acetate is denoted by yellow bars) (i) Hydrophilic interface of the alisol B 23-acetate side chain and the peripheral amino acid residuesafter stacking (alisol B 23-acetate is denoted by yellow bars)
A 24-acetate and HMG-CoA reductase was calculated to beminus23 KJmol
4 Discussion
Various experimental and clinical studies focus on the bloodlipid reducing effects of different solvent extracts (especially
water and alcohol extracts) fromAlisma orientale and its pro-cessed products [36ndash38] However the mechanisms under-lying the effects of Alisma orientale constituents remain to berevealed Alisma orientalemay interfere with exogenous cho-lesterol absorption and endogenous cholesterol metabolismthus reducing the level of TC Moreover alisol acetates mayinterfere with themetabolism of endogenous cholesterol [39]
8 Evidence-Based Complementary and Alternative Medicine
(a) (b) (c)
(d) (e) (f)
(g)
Figure 4 The interaction between alisol A 24-acetate and HMG-CoA reductase (a) Overall pattern of interaction (alisol A 24-acetate isdenoted by a bar chart) (b) Hydrogen bond graph of alisol A 24-acetate and Asn658 (the hydrogen bond is denoted by a dotted line) (c)Stacking chart of alisol A 24-acetate before and after docking (the initial structure is shown in green and the structure after docking is shownin purple) (d) The initial dihedral angle of alisol A 24-acetate angle C
3ndashC18ndashC26ndashC28 (e) The dihedral angle of alisol A 24-acetate angle
C3ndashC18ndashC26ndashC28after stacking (f) Hydrophobic interface of the alisol A 24-acetate side chain and the peripheral amino acid residues after
stacking (alisol A 24-acetate is denoted by yellow bars) (g) Hydrophilic interface of the alisol A 24-acetate side chain and the peripheralamino acid residues after stacking (alisol A 24-acetate is denoted by yellow bars)
Therefore we studied the TC-lowering effect of alisolacetates based on HMG-CoA reductase and the underly-ing molecular mechanism As the main source of endoge-nous cholesterol and one of the key enzymes in the TCmetabolism HMG-CoA reductase is the first described rate-limiting enzyme in cholesterol synthesis in vivo Moreover its
activation directly affects the speed of cholesterol synthesisand the level of TC in vivo
The TC-lowering effect and molecular mechanism ofalisol acetates the main active ingredient of Alismatis rhi-zoma were investigated using the reagent kit method andwestern blotting technique combined with the molecular
Evidence-Based Complementary and Alternative Medicine 9
simulation technique An interaction model was built foralisol acetate and the key enzyme involved in the metabolismof TC was HMG-CoA reductase The present study revealsthat alisol acetates can significantly lower the TC level ofhyperlipidemic miceThe alisol B 23-acetate showed a higherextent of TC lowering than alisol A 24-acetate
Consistent with the in vitro results alisol acetates couldalso lower the in vivoHMG-CoA reductase activity in a dose-dependent fashion with a higher degree of enzyme inhibitionfor alisol B 23-acetate compared to alisol A 24-acetate Theseresults suggest that alisol acetates might lower TC levels viaregulation of HMG-CoA reductase activity The consistencybetween the results of the in vivo and in vitro experimentsindicates that the alisol acetate prototype drug inhibitsHMG-CoA reductase activity instead of the in vivometabolites
Revealed by the western blotting results alisol acetatesmight inhibit the activity of HMG-CoA reductase by directlyand competitively binding to HMG-CoA reductase ratherthan by downregulating its protein expression
The molecular simulation technique was employed inorder to investigate the interaction of alisol acetates andHMG-CoA reductase The results (Figures 3 and 4 andTable 4) revealed themolecularmechanism of the interactionof alisol acetates and HMG-CoA reductase Both alisolacetates bound to HMG-CoA reductase in their catalyticdomain at the C-terminalThe alisol B 23-acetate formed twohydrogen bonds with HMG-CoA reductase The hydrophilicinteraction was stronger than the hydrophobic interactionwhereas the electrostatic interaction was stronger than thevan der Waals forces After the interaction the structure ofthe small molecule dramatically changed from the originalfolded sheet to an open structure Additionally the side chainstretched and was inserted into the protein The peripheralamino acids formed a hydrophilic environment and stablybound with it In general a hydrogen bond with a lengthof less than 35 A and an angle of 90ndash180∘ is consideredto be strong These two hydrogen bonds were relativelystrong and might be partly responsible for the torsion ofthe small molecule The alisol A 24-acetate formed onehydrogen bond with the HMG-CoA reductase and this bondwas weaker compared to those of alisol B 23-acetate Thehydrophilic interaction was stronger than the hydrophobicinteraction and the van derWaals force was stronger than theelectrostatic interaction After the interaction the moleculeside chain had large torsion and changed from the originalstretched open state to a ring-like sheet state which wasdifficult to insert into the proteinThe binding energy of alisolB 23-acetate andHMG-CoA reductase was lower than that ofalisol A 24-acetate and HMG-CoA reductase indicating thatthe binding of alisol B 23-acetate and HMG-CoA reductasewas stable According to the pharmacological results alisolB 23-acetate lowered the HMG-CoA reductase activity morethan alisol A 24-acetate This suggests that the active centerwas the interaction pocket constituted by these amino acidsand that the hydrogen bonds involving Lys691 Asp767 andAsn658 were the key amino acid residues for the interactionof this protein with alisol acetates
The binding of alisol B 23-acetate and HMG-CoA reduc-tase was considerably stronger than the binding of alisol
A 24-acetate and HMG-CoA reductase The relatively weakbinding of alisol A 24-acetate and HMG-CoA reductasemight be due to its distinct side-chain structure The sidechain of alisol B 23-acetate contained an ether bond whichwas strongly hydrophilic and could be inserted into thehydrophilic environment of the protein to form strong inter-actions Previous studies found that the side chains of alisolacetates were their active groups [40] In the present studythis conclusionwas validated A folded side chainparent ringbound more strongly to the macromolecule than an openside-chainparent ring did Thus the side chain acts as thesteering group in that it steers the interactions between alisolacetates and macromolecules This might be the key groupfor interactions with macromolecules as it had a decisiveeffect on the interactions of this type of compound withmacromolecules Our previous studies indicated that alisol B23-acetate was partially converted to alisol A 24-acetate in theenvironment of the human stomach However the bindingof alisol A 24-acetate and HMG-CoA reductase was weaker[14 41 42] Therefore it is advisable for drug developers todevelop a dosage form that circumvents the conversion ofalisol B 23-acetate in the stomach
Previously published studies indicate that [43 44] themechanism of reducing the level of TC is that two acetylCoA molecules condense to acetyl CoA under the catalysisof acetoacetyl-coenzyme A thiolase (AACT) acetyl CoAand acetyl CoA condense to HMG-CoA and HMG-CoAreduces to MVA under the catalytic action of HMG-CoAreductase MVA further generates TC with squalene Thehigh intracellular free fatty acid (FC) level stimulates micro-somal acyl coenzyme A-cholesterol acyltransferase (ACAT)and promotes esterification of the cholesterol Subsequentlycholesterol is stored in the form of cholesterol esters (CE)which is a feedback inhibition of HMG-CoA reductasesynthesis so that cholesterol synthesis rate-limiting enzymeswill be inhibited thus reducing intracellular cholesterolsynthesis This process features three important enzymesAACT HMG-CoA reductase and ACAT Whether AACTand ACAT are involved in the effect of alisol acetates will bethe subject of intensive research in the future
The results of the study indicate that alisol acetates cannot only decrease TC level but also decrease TG and LDL-C levels and increase HDL-C level Furthermore a follow-upstudy is designed to investigate the regulatory mechanisms ofalisol acetates on TG HDL-C and LDL-C
5 Conclusion
The present study reveals that alisol acetates could signifi-cantly lower the TC level of hyperlipidemic mice The effectof alisol B 23-acetate on HMG-CoA reductase was strongerthan that of alisol A 24-acetate Alisol acetates could lowerHMG-CoA reductase activity in a dose-dependent fashionboth in vivo and in vitro Neither of these alisol acetatescould significantly lower the protein expression of HMG-CoA This suggests that alisol acetates lower the TC levelvia inhibiting the activity of HMG-CoA reductase via itsprototype drug which may have an inhibitory effect bydirectly and competitively binding with HMG-CoA In the
10 Evidence-Based Complementary and Alternative Medicine
binding interaction of alisol acetate andHMG-CoA reductasevia molecular simulation the key amino acid residues ofHMG-CoA reductase might be Lys691 Asp767 and Asn658The side chain of the alisol acetatewas the steering group Fur-thermore a folded side chainparent ring boundmoreweaklyto HMG-CoA reductase than an open side chainparent ringThis study explored the TC-lowering mechanism of alisolacetates at the molecular level and presented guidance for itsclinical use
Competing Interests
The authors declare that they have no competing interests
Acknowledgments
This work was financially supported by the National NatureScience Foundation of China (81303173 81673534) the Nat-ural Science Foundation of Jiangsu Province (BK20161576)the Natural Science Fund for Colleges and Universities inJiangsu Province (14KJB360001) and the Top-Notch Aca-demic Programs Project of Jiangsu Higher Education Insti-tutions (PPZY2015A070)
References
[1] L X Wang Q N Wu Q Zhang G P Peng and A W DingldquoBasic study of diuretic active compounds in Rhizoma Alis-matisrdquo West China Journal of Pharmaceutical Sciences vol 23no 6 pp 670ndash672 2008
[2] S S Wu Pharmaceutical and Clinical Research of Alisma Ori-entalis Chinese Press Beijing China 2007
[3] M Miyazawa S Yoshinaga and Y Kashima ldquoChemical com-position and characteristic odor compounds in essential oilfrom Alismatis Rhizoma (Tubers of Alisma orientale)rdquo DiatomResearch vol 23 no 1 pp 133ndash141 2016
[4] M-K Jang Y-R Han J S Nam et al ldquoProtective effects ofAlisma orientale extract against hepatic steatosis via inhibitionof endoplasmic reticulum stressrdquo International Journal ofMolec-ular Sciences vol 16 no 11 pp 26151ndash26165 2015
[5] Z-P Mai K Zhou G-B Ge et al ldquoProtostane triterpenoidsfrom the rhizome of Alisma orientale exhibit inhibitory effectson human carboxylesterase 2rdquo Journal of Natural Products vol78 no 10 pp 2372ndash2380 2015
[6] T Tian H Chen andY-Y Zhao ldquoTraditional uses phytochem-istry pharmacology toxicology and quality control of Alismaorientale (Sam) Juzep a reviewrdquo Journal of Ethnopharmacologyvol 158 pp 373ndash387 2014
[7] KHKimM J Kwun J-Y Choi et al ldquoTherapeutic effect of thetuber of Alisma orientale on lipopolysaccharide-induced acutelung injuryrdquo Evidence-Based Complementary and AlternativeMedicine vol 2013 Article ID 863892 10 pages 2013
[8] H-G Jin Q Jin A R Kim et al ldquoA new triterpenoid fromAlisma orientale and their antibacterial effectrdquoArchives of Phar-macal Research vol 35 no 11 pp 1919ndash1926 2012
[9] J H Lee O S Kwon H-G Jin E-R Woo Y S Kim and H PKim ldquoThe rhizomes of Alisma orientale and alisol derivativesinhibit allergic response and experimental atopic dermatitisrdquoBiological and Pharmaceutical Bulletin vol 35 no 9 pp 1581ndash1587 2012
[10] WGu C GengW Xue et al ldquoCharacterization and function ofthe 3-hydroxy-3-methylglutaryl-CoA reductase gene in Alismaorientale (Sam) Juz and its relationship with protostane triter-pene productionrdquo Plant Physiology and Biochemistry vol 97 pp378ndash389 2015
[11] W Gu B L Xi Q N Wu et al ldquoCloning and distribution pat-tern of HMGR gene conserved fragment in Alisma orientalerdquoJournal of Chinese Medicinal Materials vol 34 no 9 pp 1335ndash1339 2011
[12] T Kobayashi S Kato-Emori K Tomita and H Ezura ldquoDetec-tion of 3-hydroxy-3-methylglutaryl-coenzymeA reductase pro-tein Cm-HMGR during fruit development in melon (Cucumismelo L)rdquo Theoretical and Applied Genetics vol 104 no 5 pp779ndash785 2002
[13] R Tommer A Rachel P C Sylvie and R Jerry ldquoImpaired regu-lation of 3-hydroxy-3-methylglutaryl-coenzyme A reductasedegradation in lovastatin-resistant cellsrdquo The Journal of Bioog-ical Chemistry vol 274 no 41 pp 29341ndash29351 1999
[14] F Xu Q-N Wu J Chen et al ldquoThe binding mechanisms ofplasma protein to active compounds in Alismaorientale rhi-zomes (Alismatis Rhizoma)rdquo Bioorganic and Medicinal Chem-istry Letters vol 24 no 17 pp 4099ndash4105 2014
[15] G Gong Y Qin W Huang et al ldquoProtective effects of dios-genin in the hyperlipidemic rat model and in human vascularendothelial cells against hydrogen peroxide-induced apoptosisrdquoChemico-Biological Interactions vol 184 no 3 pp 366ndash3752010
[16] T F Tzeng H J Lu S S Lion C J Chang and IM Liu ldquoLipid-lowering effects of zerumbone a natural cyclic sesquiterpeneof Zingiber zerumbet Smith in high-fat diet-induced hyperlipi-demic hamstersrdquoFood andChemical Toxicology vol 69 pp 132ndash139 2014
[17] Y Xie H Zhang H Liu et al ldquoHypocholesterolemic effectsof Kluyveromyces marxianus M3 isolated from Tibetan mush-rooms on diet-induced hypercholesterolemia in ratrdquo BrazilianJournal of Microbiology vol 46 no 2 pp 389ndash395 2015
[18] I Karam N Ma X-W Liu et al ldquoRegulation effect of AspirinEugenol Ester on blood lipids in Wistar rats with hyperlipi-demiardquo BMC Veterinary Research vol 11 article 217 2015
[19] G Jiao J B Wei M H Yin and P T Chun ldquoA new TCMformula FTZ lowers serum cholesterol by regulating HMG-CoA reductase and CYP7A1 in hyperlipidemic ratsrdquo Journal ofEthnopharmacology vol 135 no 2 pp 299ndash307 2011
[20] Y Geng W L Tu J J Zhang L Zhang and J Zhang ldquoEffectsof extract from cole pollen on lipidmetabolism in experimentalhyperlipidemic ratsrdquo The Scientific World Journal vol 2014Article ID 982498 6 pages 2014
[21] K Sueyoshi Y Nogawa K Sugawara T Endo and HHisamoto ldquoHighly sensitive and multiple enzyme activity assayusing reagent-release capillary-isoelectric focusing with rhod-amine 110-based substratesrdquo Analytical Sciences vol 31 no 11pp 1155ndash1161 2015
[22] D Fteita E Kononen M Gursoy E Soderling and U KGursoy ldquoDoes estradiol have an impact on the dipeptidylpeptidase IV enzyme activity of the Prevotella intermedia groupbacteriardquo Anaerobe vol 36 pp 14ndash18 2015
[23] L L Zhuo LWang and N Zhou ldquoRandomly arrayed G-quad-ruplexes for label-free and real-time assay of enzyme activityrdquoChemical Communications vol 50 no 52 pp 6875ndash6878 2014
[24] M Garelnabi and A Younis ldquoParaoxonase-1 enzyme activityassay for clinical samples validation and correlation studiesrdquoMedical Science Monitor vol 21 pp 902ndash908 2015
Evidence-Based Complementary and Alternative Medicine 11
[25] X Bi andZ Liu ldquoEnzyme activity assay of glycoprotein enzymesbased on a boronate affinity molecularly imprinted 96-wellmicroplaterdquo Analytical Chemistry vol 86 no 24 pp 12382ndash12389 2014
[26] A Walls S Dermody R Kumaran N Krishnan and E HHarley ldquoCharacterization of B-Cells in tonsils of patients diag-nosed with pediatric autoimmune neuropsychiatric disorderassociated streptococcusrdquo International Journal of PediatricOtorhinolaryngology vol 80 pp 49ndash52 2016
[27] I Karam N Ma X-W Liu et al ldquoRegulation effect of AspirinEugenol Ester on blood lipids in Wistar rats with hyperlipi-demiardquo BMC Veterinary Research vol 11 no 1 article 217 2015
[28] C Sunil S Ignacimuthu and C Kumarappan ldquoHypolipidemicactivity of Symplocos cochinchinensis S Moore leaves in hyper-lipidemic ratsrdquo Journal of Natural Medicines vol 66 no 1 pp32ndash38 2012
[29] W Shen Y Han B Huang et al ldquoMicroRNA-483-3p inhibitsextracellular matrix production by targeting smad4 in humantrabecular meshwork cellsrdquo Investigative Ophthalmology amp Vis-ual Science vol 56 no 13 pp 8419ndash8427 2015
[30] Z Wu Z Yi L Dong and A Zhang ldquoMolecular simulationstudy of the specific combination between four kinds of phthalicacid esters and human serum albuminrdquo Environmental Toxicol-ogy and Pharmacology vol 41 pp 259ndash265 2016
[31] K-J Hwang M Yoon H-J Hwang S-M Choi and S-MJeong ldquoMolecular dynamics simulation of oxygen ion conduc-tion in orthorhombic perovskite Ba-doped LaInO
3using cubic
and orthorhombic modelrdquo Journal of Nanoscience and Nan-otechnology vol 15 no 11 pp 8947ndash8950 2015
[32] H Yu W Han W Ma and K Schulten ldquoTransient 120573-hairpinformation in 120572-synuclein monomer revealed by coarse-grainedmolecular dynamics simulationrdquo The Journal of ChemicalPhysics vol 143 no 24 Article ID 243142 2015
[33] M Breuer K M Rosso and J Blumberger ldquoFlavin binding tothe deca-heme cytochrome mtrc insights from computationalmolecular simulationrdquo Biophysical Journal vol 109 no 12 pp2614ndash2624 2015
[34] V Rastija D Agic S Tomic et al ldquoSynthesis QSAR and mole-cular dynamics simulation of amidino-substituted benzimi-dazoles as dipeptidyl peptidase III inhibitorsrdquo Acta ChimicaSlovenica vol 62 no 4 pp 867ndash878 2015
[35] M Enciso C Schutte and L Delle Site ldquoInfluence of pH andsequence in peptide aggregation via molecular simulationrdquoTheJournal of Chemical Physics vol 143 no 24 Article ID 2431302015
[36] L XWangQWu andG P Peng ldquoA Study on determination ofcontent of 23-acetyl alisol B in Rhizoma Alismatis orientalitisrdquoJournal of Nanjing University of Traditional Chinese Medicine(Natural Science) vol 18 no 2 pp 105ndash107 2002
[37] G P Peng andY L Zhu ldquoProgress in the study on chemical con-stituents of Alisma orientalisrdquo Natural Product Research andDevelopment vol 18 no 2 pp 348ndash351 2006
[38] D Y Liu S J Gu J Wu et al ldquoStudies on fractions of zexiedecoction with antihyperlipidemia bioactivity in micerdquo ChinaPharmacist vol 13 no 6 pp 763ndash766 2010
[39] Q-F Luo L Sun and G-H Du ldquoProgress in new drugs tar-geting reverse cholesterol transportrdquo Chinese PharmacologicalBulletin vol 22 no 8 pp 904ndash907 2006
[40] M Yoshikawa and Y Fukuda ldquoHatakeayamardquo Chemical andPharmaceutical Bulletin vol 41 no 6 pp 1194ndash1196 1993
[41] F Xu L Zhang LHe et al ldquoMolecularmechanismon the inter-action between alisols and human serum albuminrdquo Acta Chim-ica Sinica vol 69 no 19 pp 2228ndash2234 2011
[42] F Xu J ChenW Gu et al ldquoStudy on transformation of effectivecomponents of Alisma under simulated in vivo conditionsrdquoChinese Journal of Pharmaceutical Analysis vol 31 no 11 pp2035ndash2039 2011
[43] H Yang Y G Gao and L X Zhang ldquoResearch progress on bio-synthetic pathway of terpenoids containing ginsenoside and theHMGRrdquo China Journal of Biological Engineering vol 28 no 10pp 130ndash135 2008
[44] G Z Cui Y L Ben N Li and D L Liu ldquoCatalytic mechanismand characteristics of HMG-COA recuctaserdquo Journal ofMedicalMolecular Biology vol 5 no 6 p 542 2008
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom

Evidence-Based Complementary and Alternative Medicine 5
comprehensively considered so as to determine the finalsteady conformation This conformation was selected forfurther molecular dynamic simulation
262 Molecular Simulation of Alisol A 24-Acetate andHMG-CoA Reductase Applying the method of Section 261resulted in the final steady conformation of alisol A 24-acetateThis conformationwas selected for furthermoleculardynamic simulation
27 Data Analysis The experimental data were analyzedusing one-way analysis of variance (ANOVA) in SPSS version150 (SPSS Inc Chicago USA) 119875 values of less than 005(119875 lt 005) indicated statistically significant differences Theresults were expressed in the form of mean (119909) plusmn standarddeviation (SD)
3 Results
31 Measurements of TC TG HDL-C and LDL-C LevelsThe blood lipid levels of the blank group positive groupmodel group alisol B 23-acetate high- medium- and low-dose groups and alisol A 24-acetate high- medium- andlow-dosage groups are shown in Table 1 This study revealedthat the TC TG LDL-C and HDL-C levels of the alisol A24-acetate high- medium- and low-dose groups as well asthe alisol B 23-acetate high- medium- and low-dose groupssignificantly differed from those of the model group Thissuggests that alisol acetates could have a lipid-regulatingeffect and significantly decrease the TC TG and LDL-Cconcentrations of hyperlipidemic mice while increasing theHDL-C concentrations
32 Measurement of In VivoIn Vitro HMG-CoAReductase Activity
321 Measurement of In Vitro HMG-CoA Reductase ActivityAs Table 2 indicates alisol acetates lower the HMG-CoAreductase activity dose dependently The extent to whichalisol B 23-acetate lowered HMG-CoA reductase activityexceeded the extent to which alisol A 24-acetate lowered theactivity of this enzyme
322 Measurement of In Vivo HMG-CoA Reductase ActivityThe results in Table 3 illustrate that alisol acetates lower thein vivo HMG-CoA reductase activity in a dose-dependentfashion with a greater degree of enzyme inhibition for alisolB 23-acetate than for alisol A 24-acetate
33 Measurement of HMG-CoA Reductase Protein Expres-sion Level via Western Blotting The western blotting resultssuggest that alisol acetates did not significantly decrease theprotein expression of HMG-CoA reductase (Figure 2)
34 Investigation of the Interaction between the Alisol Acetatesand HMG-CoA Reductase
341 Molecular Simulation Results for Alisol B 23-Acetate andHMG-CoA Reductase The molecular simulation results forthe interaction between alisol B 23-acetate and HMG-CoA
Table 2 The activity of HMG-CoA reductase in vitro (119909 plusmn s 119899 =11)
Group HMG-CoA reductase (UL)Blank 13467 plusmn 3991Positive 16429 plusmn 2832lowastlowast
Model 49788 plusmn 2881998771998771
24A high 19042 plusmn 3092lowastlowast
24A medium 25079 plusmn 1635lowastlowast
24A low 33515 plusmn 2332lowastlowast
23B high 17912 plusmn 4159lowastlowast
23B medium 23079 plusmn 1664lowastlowast
23B low 32759 plusmn 1915lowastlowast
Note 24A alisol A 24-acetate and 23B alisol B 23-acetate compared to theblank group 998771998771119875 lt 001 and 998771119875 lt 005 compared to the model grouplowastlowast119875 lt 001 and lowast119875 lt 005
Table 3The activity of HMG-CoA reductase in vivo (119909 plusmn s 119899 = 11)
Group HMG-CoA reductase (UL)Blank 12345 plusmn 2534Positive 20021 plusmn 3104lowastlowast
Model 49012 plusmn 4117998771998771
24A high 280123 plusmn 2915lowastlowast
24A medium 34554 plusmn 3234lowastlowast
24A low 40131 plusmn 4221lowastlowast
23B high 25203 plusmn 2931lowastlowast
23B medium 29614 plusmn 3218lowastlowast
23B low 39082 plusmn 2323lowastlowast
Note 24A alisol A 24-acetate and 23B alisol B 23-acetate compared to theblank group 998771998771119875 lt 001 and 998771119875 lt 005 compared to the model grouplowastlowast119875 lt 001 and lowast119875 lt 005
reductase are depicted in Figure 3 Figure 3(a) illustratesthe overall pattern of the interaction The interaction pocketwhere alisol B 23-acetate bound to HMG-CoA reductasecontained the following amino acids Leu521 Val522Met523Gly524 Ala525 Cys527 Glu528 Asn529 Val530 Met588Thy589 Arg590 Gly591 Asp653 Ala654 Met655 Gly656Met657 Asn658 Met659 Ile660 Ser661 Lys662 Gly663Thy664 Glu665 Cys688 Asp690 Lys691 Lys692 Arg702Lys704 Ile760 Try761 Ile762Ala763 Cys764Glu801 Ile802Gly803 Thr804 Val805 Gly806 Gly807 Gly808 Thr809Asn810 and Leu811Theywere located in the catalytic domainconstituted by the 439 (450ndash888) amino acids at the C-terminal After entering the interaction pocket alisol B 23-acetate interacted with the peripheral amino acid residues viahydrogen bonding and hydrophobic and hydrophilic effectsThe hydrogen bond graph in Figure 3(b) shows two hydrogenbonds between alisol B 23-acetate and the amino acids Lys691and Asp767 Table 4 presents detailed information aboutthe hydrogen bonding The interaction energy between thehydrophobic amino acid residues and the small molecule wascalculated to be minus128 KJmol while the interaction energybetween hydrophilic amino acid residues and the smallmolecule was calculated to be minus1353 KJmol This indicatesthat the hydrophilic interaction between the small moleculeand the peripheral amino acids was stronger
6 Evidence-Based Complementary and Alternative Medicine
HMG-CoAreductase
GAPDH
05
04
03
02
01
00HM
G-C
oA re
duct
ase
GA
PDH
Blan
k
Mod
el
Posit
ive
24
A-H
24
A-M
24
A-L
23
B-H
23
B-M
23
B-L
Blan
k
Mod
el
Posit
ive
24
A-H
24
A-M
24
A-L
23
B-H
23
B-M
23
B-L
lowast
Figure 2 Results of the protein expression of HMG-CoA reductasein mice liver tissue protein (119899 = 11) Note compared to the blankgroup 998771998771119875 lt 001 and 998771119875 lt 005 compared to the model grouplowastlowast119875 lt 001 and lowast119875 lt 005 24A-H alisol A 24-acetate high 24A-
M alisol A 24-acetate medium and 24A-L alisol A 24-acetate low23B-H alisol B 23-acetate high 23B-M alisol B 23-acetate mediumand 23B-L alisol B 24-acetate low
Table 4 The length (gt) and the angle of hydrogen bond formedbetween alisol acetates and HMG-CoA reductase
Name XndashHsdot sdot sdotYd
(XndashH)
d(Hsdot sdot sdotY)
d(Xsdot sdot sdotY) Angle XHY
Alisol B
23-acetate
Lys691 NZ-HZ1sdot sdot sdotO37 104 237 298 11594
Asp767OD2-
HH88sdot sdot sdotO2096 240 324 14490
Alisol A24-acetate
Asn658 OD1-H5Zsdot sdot sdotO5 095 201 286 14897
After docking the conformation of alisol B 23-acetatechanged as shown in Figure 3(c) The calculated root-mean-square deviation (RMSD) was 289gt The conformationof the small molecule changed from the clustered sheetstructure to the open structure Before docking the sidechain and the ring structure of the small molecule foldedas shown in Figures 3(d) and 3(e) The angle between C
2ndash
C10ndashC19was 1095∘ 1095∘ between C
10ndashC19ndashC20 and minus599∘
between C2ndashC10ndashC19ndashC20 As shown in Figures 3(f) and
3(g) after docking the angle between C2ndashC10ndashC19was 1136∘
1156∘ between C10ndashC19ndashC20 and 1260∘ between C
2ndashC10ndash
C19ndashC20 The angle between the parent ring and the side
chain was significantly increasedThe twisting of the dihedralangles resulted in the opening of side chains and insertioninto the protein which led to interactions There was ahydrophilic ether bond on the side chain and the peripheralamino acids were mainly hydrophilic (Figures 3(h) and 3(i))
This created a hydrophilic environment around the sidechains culminating in stable binding The energy of thevan der Waals forces between them was calculated to beminus1519 KJmol whereas the electrostatic energy was calcu-lated to be minus2366 KJmol As the electrostatic energy waslower than the energy of the van derWaals forces between thesystems the electrostatic interactionwas themain interactionbetween this small molecule and the macromolecule Thebinding energy (ΔG∘) between the alisol B 23-acetate and theHMG-CoA reductase was calculated to be minus1033 KJmol
342 Molecular Simulation Results of Alisol A 24-Acetateand HMG-CoA Reductase Themolecular simulation resultsfor the interaction between alisol A 24-acetate and HMG-CoA reductase are shown in Figure 4 Figure 4(a) shows theoverall interaction pattern of this compound and HMG-CoAreductase The interaction pocket where it bound to HMG-CoA reductase contained the following amino acids Tyr519Ser520 Leu521 Val522 Met523 Gly524 Ala525 Cys527Glu528 Asn529 Val530 Ile531 Gly532 Try533 Met588Thy589 Arg590 Gly591 Asp653 Ala654 Met655 Gly656Met657 Asn658 Met659 Ile660 Ser661 Lys662 Gly663Thy664 Glu665 Cys688 The689 Asp690 Lys691 Lys692Cys764 Gly765 Gln766 Asp767 Ala768 Ala769 Gln770Asn771 Val772 Gly773 Ser774 Glu801 Ile802 Gly803Thr804Val805Gly806Gly807Gly808Thr809 andAsn810There were 439 (450ndash888) amino acids in the catalyticdomain at the C-terminal After entering the interactionpocket alisol A 24-acetate exhibited hydrogen bonding andhydrophobic and hydrophilic interactions with peripheralamino acid residues Figure 4(b) shows the hydrogen bondcontact map There was one hydrogen bond between alisol A24 acetate and amino acid Asn658 (Table 4) The interactionenergy of the hydrophobic amino acid residues and thesmall molecule was calculated to be 465 KJmol whereas theinteraction energy of the hydrophilic amino acid residues andthe small molecule was calculated to be minus1023 KJmol Thissuggests that the hydrophilic groups had a much strongereffect than the hydrophobic groups The small molecule andthe peripheral amino acids exhibited a stronger hydrophilicinteraction as indicated by the interface graphs (Figures 4(f)and 4(g))
After docking the alisol A 24-acetate conformationchanged (Figure 4(c)) The RMSD was calculated to be334gt The conformation angle C
3ndashC18ndashC26ndashC28
changedfrom 600∘ before docking to 1242∘ after docking (Figures4(d) and 4(e)) This suggests a large torsion of the smallmolecule side chain after docking whereby the stretchedopen state changed to a ring-like sheet state Consequentlyit was difficult to insert the side chain into the protein andinteract with the active pocket of the protein This resultedin considerably lower binding relative to alisol B 23-acetateThe electrostatic energy of alisol A 24-acetate and HMG-CoA reductase was determined to be minus886 KJmol whilethe energy of the van der Waals forces was determined tobe minus1649 KJmol The energy of the van der Waals forceswas lower than the electrostatic energy showing that themain interaction between this small molecule and themacro-molecule was the van derWaals interactionThe ΔG∘ of alisol
Evidence-Based Complementary and Alternative Medicine 7
(a) (b) (c)
(d) (e) (f)
(g) (h) (i)
Figure 3 Interaction between alisol B 23-acetate and HMG-CoA reductase (a) Overall pattern of interaction (alisol B 23-acetate is denotedby a bar chart) (b) Hydrogen bond graph of alisol B 23-acetate with Lys691 and Asp767 (the hydrogen bond is denoted by a dotted line) (c)Stacking chart of alisol B 23-acetate before and after docking (the initial structure is shown in green and the structure after docking is shown inpurple) (d) Initial angle of alisol B 23-acetate C
2ndashC10ndashC19and C
10ndashC19ndashC20 (e) Initial dihedral angle of alisol B 23-acetate C
2ndashC10ndashC19ndashC20
(f) The angle of alisol B 23-acetate C2ndashC10ndashC19and C
10ndashC19ndashC20after stacking (g)The dihedral angle of alisol B 23-acetate C
2ndashC10ndashC19ndashC20
after stacking (h) Hydrophobic interface of the alisol B 23-acetate side chain and the peripheral amino acid residues after stacking (alisol B23-acetate is denoted by yellow bars) (i) Hydrophilic interface of the alisol B 23-acetate side chain and the peripheral amino acid residuesafter stacking (alisol B 23-acetate is denoted by yellow bars)
A 24-acetate and HMG-CoA reductase was calculated to beminus23 KJmol
4 Discussion
Various experimental and clinical studies focus on the bloodlipid reducing effects of different solvent extracts (especially
water and alcohol extracts) fromAlisma orientale and its pro-cessed products [36ndash38] However the mechanisms under-lying the effects of Alisma orientale constituents remain to berevealed Alisma orientalemay interfere with exogenous cho-lesterol absorption and endogenous cholesterol metabolismthus reducing the level of TC Moreover alisol acetates mayinterfere with themetabolism of endogenous cholesterol [39]
8 Evidence-Based Complementary and Alternative Medicine
(a) (b) (c)
(d) (e) (f)
(g)
Figure 4 The interaction between alisol A 24-acetate and HMG-CoA reductase (a) Overall pattern of interaction (alisol A 24-acetate isdenoted by a bar chart) (b) Hydrogen bond graph of alisol A 24-acetate and Asn658 (the hydrogen bond is denoted by a dotted line) (c)Stacking chart of alisol A 24-acetate before and after docking (the initial structure is shown in green and the structure after docking is shownin purple) (d) The initial dihedral angle of alisol A 24-acetate angle C
3ndashC18ndashC26ndashC28 (e) The dihedral angle of alisol A 24-acetate angle
C3ndashC18ndashC26ndashC28after stacking (f) Hydrophobic interface of the alisol A 24-acetate side chain and the peripheral amino acid residues after
stacking (alisol A 24-acetate is denoted by yellow bars) (g) Hydrophilic interface of the alisol A 24-acetate side chain and the peripheralamino acid residues after stacking (alisol A 24-acetate is denoted by yellow bars)
Therefore we studied the TC-lowering effect of alisolacetates based on HMG-CoA reductase and the underly-ing molecular mechanism As the main source of endoge-nous cholesterol and one of the key enzymes in the TCmetabolism HMG-CoA reductase is the first described rate-limiting enzyme in cholesterol synthesis in vivo Moreover its
activation directly affects the speed of cholesterol synthesisand the level of TC in vivo
The TC-lowering effect and molecular mechanism ofalisol acetates the main active ingredient of Alismatis rhi-zoma were investigated using the reagent kit method andwestern blotting technique combined with the molecular
Evidence-Based Complementary and Alternative Medicine 9
simulation technique An interaction model was built foralisol acetate and the key enzyme involved in the metabolismof TC was HMG-CoA reductase The present study revealsthat alisol acetates can significantly lower the TC level ofhyperlipidemic miceThe alisol B 23-acetate showed a higherextent of TC lowering than alisol A 24-acetate
Consistent with the in vitro results alisol acetates couldalso lower the in vivoHMG-CoA reductase activity in a dose-dependent fashion with a higher degree of enzyme inhibitionfor alisol B 23-acetate compared to alisol A 24-acetate Theseresults suggest that alisol acetates might lower TC levels viaregulation of HMG-CoA reductase activity The consistencybetween the results of the in vivo and in vitro experimentsindicates that the alisol acetate prototype drug inhibitsHMG-CoA reductase activity instead of the in vivometabolites
Revealed by the western blotting results alisol acetatesmight inhibit the activity of HMG-CoA reductase by directlyand competitively binding to HMG-CoA reductase ratherthan by downregulating its protein expression
The molecular simulation technique was employed inorder to investigate the interaction of alisol acetates andHMG-CoA reductase The results (Figures 3 and 4 andTable 4) revealed themolecularmechanism of the interactionof alisol acetates and HMG-CoA reductase Both alisolacetates bound to HMG-CoA reductase in their catalyticdomain at the C-terminalThe alisol B 23-acetate formed twohydrogen bonds with HMG-CoA reductase The hydrophilicinteraction was stronger than the hydrophobic interactionwhereas the electrostatic interaction was stronger than thevan der Waals forces After the interaction the structure ofthe small molecule dramatically changed from the originalfolded sheet to an open structure Additionally the side chainstretched and was inserted into the protein The peripheralamino acids formed a hydrophilic environment and stablybound with it In general a hydrogen bond with a lengthof less than 35 A and an angle of 90ndash180∘ is consideredto be strong These two hydrogen bonds were relativelystrong and might be partly responsible for the torsion ofthe small molecule The alisol A 24-acetate formed onehydrogen bond with the HMG-CoA reductase and this bondwas weaker compared to those of alisol B 23-acetate Thehydrophilic interaction was stronger than the hydrophobicinteraction and the van derWaals force was stronger than theelectrostatic interaction After the interaction the moleculeside chain had large torsion and changed from the originalstretched open state to a ring-like sheet state which wasdifficult to insert into the proteinThe binding energy of alisolB 23-acetate andHMG-CoA reductase was lower than that ofalisol A 24-acetate and HMG-CoA reductase indicating thatthe binding of alisol B 23-acetate and HMG-CoA reductasewas stable According to the pharmacological results alisolB 23-acetate lowered the HMG-CoA reductase activity morethan alisol A 24-acetate This suggests that the active centerwas the interaction pocket constituted by these amino acidsand that the hydrogen bonds involving Lys691 Asp767 andAsn658 were the key amino acid residues for the interactionof this protein with alisol acetates
The binding of alisol B 23-acetate and HMG-CoA reduc-tase was considerably stronger than the binding of alisol
A 24-acetate and HMG-CoA reductase The relatively weakbinding of alisol A 24-acetate and HMG-CoA reductasemight be due to its distinct side-chain structure The sidechain of alisol B 23-acetate contained an ether bond whichwas strongly hydrophilic and could be inserted into thehydrophilic environment of the protein to form strong inter-actions Previous studies found that the side chains of alisolacetates were their active groups [40] In the present studythis conclusionwas validated A folded side chainparent ringbound more strongly to the macromolecule than an openside-chainparent ring did Thus the side chain acts as thesteering group in that it steers the interactions between alisolacetates and macromolecules This might be the key groupfor interactions with macromolecules as it had a decisiveeffect on the interactions of this type of compound withmacromolecules Our previous studies indicated that alisol B23-acetate was partially converted to alisol A 24-acetate in theenvironment of the human stomach However the bindingof alisol A 24-acetate and HMG-CoA reductase was weaker[14 41 42] Therefore it is advisable for drug developers todevelop a dosage form that circumvents the conversion ofalisol B 23-acetate in the stomach
Previously published studies indicate that [43 44] themechanism of reducing the level of TC is that two acetylCoA molecules condense to acetyl CoA under the catalysisof acetoacetyl-coenzyme A thiolase (AACT) acetyl CoAand acetyl CoA condense to HMG-CoA and HMG-CoAreduces to MVA under the catalytic action of HMG-CoAreductase MVA further generates TC with squalene Thehigh intracellular free fatty acid (FC) level stimulates micro-somal acyl coenzyme A-cholesterol acyltransferase (ACAT)and promotes esterification of the cholesterol Subsequentlycholesterol is stored in the form of cholesterol esters (CE)which is a feedback inhibition of HMG-CoA reductasesynthesis so that cholesterol synthesis rate-limiting enzymeswill be inhibited thus reducing intracellular cholesterolsynthesis This process features three important enzymesAACT HMG-CoA reductase and ACAT Whether AACTand ACAT are involved in the effect of alisol acetates will bethe subject of intensive research in the future
The results of the study indicate that alisol acetates cannot only decrease TC level but also decrease TG and LDL-C levels and increase HDL-C level Furthermore a follow-upstudy is designed to investigate the regulatory mechanisms ofalisol acetates on TG HDL-C and LDL-C
5 Conclusion
The present study reveals that alisol acetates could signifi-cantly lower the TC level of hyperlipidemic mice The effectof alisol B 23-acetate on HMG-CoA reductase was strongerthan that of alisol A 24-acetate Alisol acetates could lowerHMG-CoA reductase activity in a dose-dependent fashionboth in vivo and in vitro Neither of these alisol acetatescould significantly lower the protein expression of HMG-CoA This suggests that alisol acetates lower the TC levelvia inhibiting the activity of HMG-CoA reductase via itsprototype drug which may have an inhibitory effect bydirectly and competitively binding with HMG-CoA In the
10 Evidence-Based Complementary and Alternative Medicine
binding interaction of alisol acetate andHMG-CoA reductasevia molecular simulation the key amino acid residues ofHMG-CoA reductase might be Lys691 Asp767 and Asn658The side chain of the alisol acetatewas the steering group Fur-thermore a folded side chainparent ring boundmoreweaklyto HMG-CoA reductase than an open side chainparent ringThis study explored the TC-lowering mechanism of alisolacetates at the molecular level and presented guidance for itsclinical use
Competing Interests
The authors declare that they have no competing interests
Acknowledgments
This work was financially supported by the National NatureScience Foundation of China (81303173 81673534) the Nat-ural Science Foundation of Jiangsu Province (BK20161576)the Natural Science Fund for Colleges and Universities inJiangsu Province (14KJB360001) and the Top-Notch Aca-demic Programs Project of Jiangsu Higher Education Insti-tutions (PPZY2015A070)
References
[1] L X Wang Q N Wu Q Zhang G P Peng and A W DingldquoBasic study of diuretic active compounds in Rhizoma Alis-matisrdquo West China Journal of Pharmaceutical Sciences vol 23no 6 pp 670ndash672 2008
[2] S S Wu Pharmaceutical and Clinical Research of Alisma Ori-entalis Chinese Press Beijing China 2007
[3] M Miyazawa S Yoshinaga and Y Kashima ldquoChemical com-position and characteristic odor compounds in essential oilfrom Alismatis Rhizoma (Tubers of Alisma orientale)rdquo DiatomResearch vol 23 no 1 pp 133ndash141 2016
[4] M-K Jang Y-R Han J S Nam et al ldquoProtective effects ofAlisma orientale extract against hepatic steatosis via inhibitionof endoplasmic reticulum stressrdquo International Journal ofMolec-ular Sciences vol 16 no 11 pp 26151ndash26165 2015
[5] Z-P Mai K Zhou G-B Ge et al ldquoProtostane triterpenoidsfrom the rhizome of Alisma orientale exhibit inhibitory effectson human carboxylesterase 2rdquo Journal of Natural Products vol78 no 10 pp 2372ndash2380 2015
[6] T Tian H Chen andY-Y Zhao ldquoTraditional uses phytochem-istry pharmacology toxicology and quality control of Alismaorientale (Sam) Juzep a reviewrdquo Journal of Ethnopharmacologyvol 158 pp 373ndash387 2014
[7] KHKimM J Kwun J-Y Choi et al ldquoTherapeutic effect of thetuber of Alisma orientale on lipopolysaccharide-induced acutelung injuryrdquo Evidence-Based Complementary and AlternativeMedicine vol 2013 Article ID 863892 10 pages 2013
[8] H-G Jin Q Jin A R Kim et al ldquoA new triterpenoid fromAlisma orientale and their antibacterial effectrdquoArchives of Phar-macal Research vol 35 no 11 pp 1919ndash1926 2012
[9] J H Lee O S Kwon H-G Jin E-R Woo Y S Kim and H PKim ldquoThe rhizomes of Alisma orientale and alisol derivativesinhibit allergic response and experimental atopic dermatitisrdquoBiological and Pharmaceutical Bulletin vol 35 no 9 pp 1581ndash1587 2012
[10] WGu C GengW Xue et al ldquoCharacterization and function ofthe 3-hydroxy-3-methylglutaryl-CoA reductase gene in Alismaorientale (Sam) Juz and its relationship with protostane triter-pene productionrdquo Plant Physiology and Biochemistry vol 97 pp378ndash389 2015
[11] W Gu B L Xi Q N Wu et al ldquoCloning and distribution pat-tern of HMGR gene conserved fragment in Alisma orientalerdquoJournal of Chinese Medicinal Materials vol 34 no 9 pp 1335ndash1339 2011
[12] T Kobayashi S Kato-Emori K Tomita and H Ezura ldquoDetec-tion of 3-hydroxy-3-methylglutaryl-coenzymeA reductase pro-tein Cm-HMGR during fruit development in melon (Cucumismelo L)rdquo Theoretical and Applied Genetics vol 104 no 5 pp779ndash785 2002
[13] R Tommer A Rachel P C Sylvie and R Jerry ldquoImpaired regu-lation of 3-hydroxy-3-methylglutaryl-coenzyme A reductasedegradation in lovastatin-resistant cellsrdquo The Journal of Bioog-ical Chemistry vol 274 no 41 pp 29341ndash29351 1999
[14] F Xu Q-N Wu J Chen et al ldquoThe binding mechanisms ofplasma protein to active compounds in Alismaorientale rhi-zomes (Alismatis Rhizoma)rdquo Bioorganic and Medicinal Chem-istry Letters vol 24 no 17 pp 4099ndash4105 2014
[15] G Gong Y Qin W Huang et al ldquoProtective effects of dios-genin in the hyperlipidemic rat model and in human vascularendothelial cells against hydrogen peroxide-induced apoptosisrdquoChemico-Biological Interactions vol 184 no 3 pp 366ndash3752010
[16] T F Tzeng H J Lu S S Lion C J Chang and IM Liu ldquoLipid-lowering effects of zerumbone a natural cyclic sesquiterpeneof Zingiber zerumbet Smith in high-fat diet-induced hyperlipi-demic hamstersrdquoFood andChemical Toxicology vol 69 pp 132ndash139 2014
[17] Y Xie H Zhang H Liu et al ldquoHypocholesterolemic effectsof Kluyveromyces marxianus M3 isolated from Tibetan mush-rooms on diet-induced hypercholesterolemia in ratrdquo BrazilianJournal of Microbiology vol 46 no 2 pp 389ndash395 2015
[18] I Karam N Ma X-W Liu et al ldquoRegulation effect of AspirinEugenol Ester on blood lipids in Wistar rats with hyperlipi-demiardquo BMC Veterinary Research vol 11 article 217 2015
[19] G Jiao J B Wei M H Yin and P T Chun ldquoA new TCMformula FTZ lowers serum cholesterol by regulating HMG-CoA reductase and CYP7A1 in hyperlipidemic ratsrdquo Journal ofEthnopharmacology vol 135 no 2 pp 299ndash307 2011
[20] Y Geng W L Tu J J Zhang L Zhang and J Zhang ldquoEffectsof extract from cole pollen on lipidmetabolism in experimentalhyperlipidemic ratsrdquo The Scientific World Journal vol 2014Article ID 982498 6 pages 2014
[21] K Sueyoshi Y Nogawa K Sugawara T Endo and HHisamoto ldquoHighly sensitive and multiple enzyme activity assayusing reagent-release capillary-isoelectric focusing with rhod-amine 110-based substratesrdquo Analytical Sciences vol 31 no 11pp 1155ndash1161 2015
[22] D Fteita E Kononen M Gursoy E Soderling and U KGursoy ldquoDoes estradiol have an impact on the dipeptidylpeptidase IV enzyme activity of the Prevotella intermedia groupbacteriardquo Anaerobe vol 36 pp 14ndash18 2015
[23] L L Zhuo LWang and N Zhou ldquoRandomly arrayed G-quad-ruplexes for label-free and real-time assay of enzyme activityrdquoChemical Communications vol 50 no 52 pp 6875ndash6878 2014
[24] M Garelnabi and A Younis ldquoParaoxonase-1 enzyme activityassay for clinical samples validation and correlation studiesrdquoMedical Science Monitor vol 21 pp 902ndash908 2015
Evidence-Based Complementary and Alternative Medicine 11
[25] X Bi andZ Liu ldquoEnzyme activity assay of glycoprotein enzymesbased on a boronate affinity molecularly imprinted 96-wellmicroplaterdquo Analytical Chemistry vol 86 no 24 pp 12382ndash12389 2014
[26] A Walls S Dermody R Kumaran N Krishnan and E HHarley ldquoCharacterization of B-Cells in tonsils of patients diag-nosed with pediatric autoimmune neuropsychiatric disorderassociated streptococcusrdquo International Journal of PediatricOtorhinolaryngology vol 80 pp 49ndash52 2016
[27] I Karam N Ma X-W Liu et al ldquoRegulation effect of AspirinEugenol Ester on blood lipids in Wistar rats with hyperlipi-demiardquo BMC Veterinary Research vol 11 no 1 article 217 2015
[28] C Sunil S Ignacimuthu and C Kumarappan ldquoHypolipidemicactivity of Symplocos cochinchinensis S Moore leaves in hyper-lipidemic ratsrdquo Journal of Natural Medicines vol 66 no 1 pp32ndash38 2012
[29] W Shen Y Han B Huang et al ldquoMicroRNA-483-3p inhibitsextracellular matrix production by targeting smad4 in humantrabecular meshwork cellsrdquo Investigative Ophthalmology amp Vis-ual Science vol 56 no 13 pp 8419ndash8427 2015
[30] Z Wu Z Yi L Dong and A Zhang ldquoMolecular simulationstudy of the specific combination between four kinds of phthalicacid esters and human serum albuminrdquo Environmental Toxicol-ogy and Pharmacology vol 41 pp 259ndash265 2016
[31] K-J Hwang M Yoon H-J Hwang S-M Choi and S-MJeong ldquoMolecular dynamics simulation of oxygen ion conduc-tion in orthorhombic perovskite Ba-doped LaInO
3using cubic
and orthorhombic modelrdquo Journal of Nanoscience and Nan-otechnology vol 15 no 11 pp 8947ndash8950 2015
[32] H Yu W Han W Ma and K Schulten ldquoTransient 120573-hairpinformation in 120572-synuclein monomer revealed by coarse-grainedmolecular dynamics simulationrdquo The Journal of ChemicalPhysics vol 143 no 24 Article ID 243142 2015
[33] M Breuer K M Rosso and J Blumberger ldquoFlavin binding tothe deca-heme cytochrome mtrc insights from computationalmolecular simulationrdquo Biophysical Journal vol 109 no 12 pp2614ndash2624 2015
[34] V Rastija D Agic S Tomic et al ldquoSynthesis QSAR and mole-cular dynamics simulation of amidino-substituted benzimi-dazoles as dipeptidyl peptidase III inhibitorsrdquo Acta ChimicaSlovenica vol 62 no 4 pp 867ndash878 2015
[35] M Enciso C Schutte and L Delle Site ldquoInfluence of pH andsequence in peptide aggregation via molecular simulationrdquoTheJournal of Chemical Physics vol 143 no 24 Article ID 2431302015
[36] L XWangQWu andG P Peng ldquoA Study on determination ofcontent of 23-acetyl alisol B in Rhizoma Alismatis orientalitisrdquoJournal of Nanjing University of Traditional Chinese Medicine(Natural Science) vol 18 no 2 pp 105ndash107 2002
[37] G P Peng andY L Zhu ldquoProgress in the study on chemical con-stituents of Alisma orientalisrdquo Natural Product Research andDevelopment vol 18 no 2 pp 348ndash351 2006
[38] D Y Liu S J Gu J Wu et al ldquoStudies on fractions of zexiedecoction with antihyperlipidemia bioactivity in micerdquo ChinaPharmacist vol 13 no 6 pp 763ndash766 2010
[39] Q-F Luo L Sun and G-H Du ldquoProgress in new drugs tar-geting reverse cholesterol transportrdquo Chinese PharmacologicalBulletin vol 22 no 8 pp 904ndash907 2006
[40] M Yoshikawa and Y Fukuda ldquoHatakeayamardquo Chemical andPharmaceutical Bulletin vol 41 no 6 pp 1194ndash1196 1993
[41] F Xu L Zhang LHe et al ldquoMolecularmechanismon the inter-action between alisols and human serum albuminrdquo Acta Chim-ica Sinica vol 69 no 19 pp 2228ndash2234 2011
[42] F Xu J ChenW Gu et al ldquoStudy on transformation of effectivecomponents of Alisma under simulated in vivo conditionsrdquoChinese Journal of Pharmaceutical Analysis vol 31 no 11 pp2035ndash2039 2011
[43] H Yang Y G Gao and L X Zhang ldquoResearch progress on bio-synthetic pathway of terpenoids containing ginsenoside and theHMGRrdquo China Journal of Biological Engineering vol 28 no 10pp 130ndash135 2008
[44] G Z Cui Y L Ben N Li and D L Liu ldquoCatalytic mechanismand characteristics of HMG-COA recuctaserdquo Journal ofMedicalMolecular Biology vol 5 no 6 p 542 2008
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom
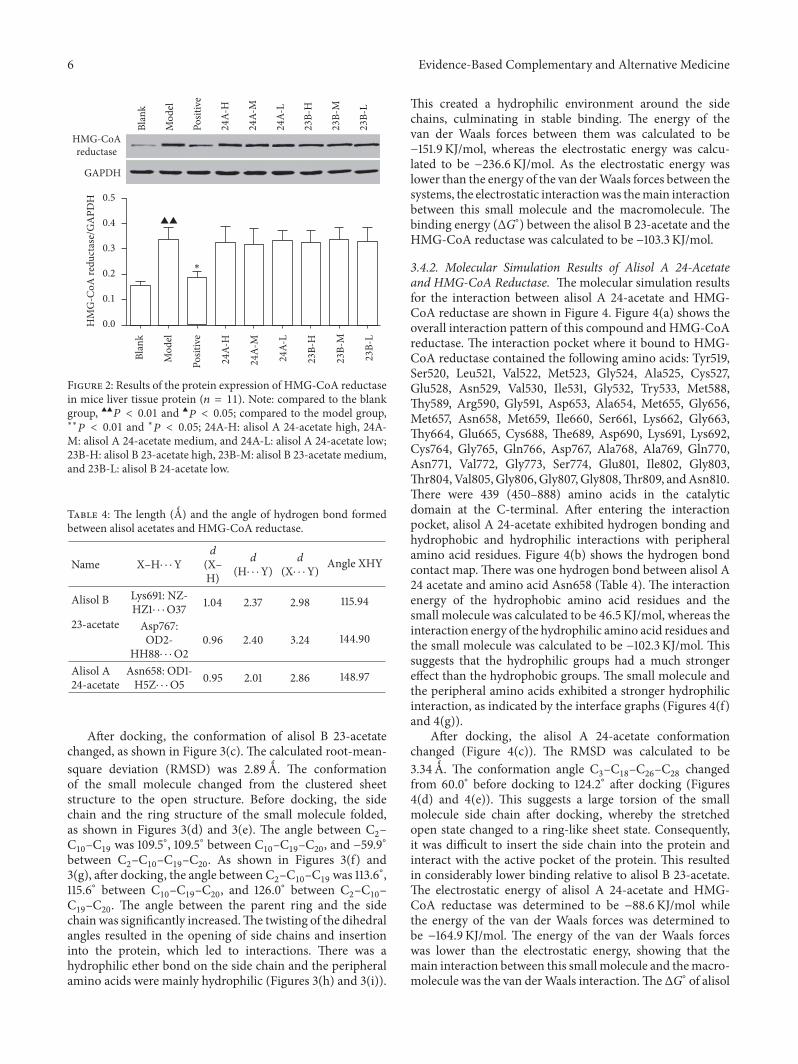
6 Evidence-Based Complementary and Alternative Medicine
HMG-CoAreductase
GAPDH
05
04
03
02
01
00HM
G-C
oA re
duct
ase
GA
PDH
Blan
k
Mod
el
Posit
ive
24
A-H
24
A-M
24
A-L
23
B-H
23
B-M
23
B-L
Blan
k
Mod
el
Posit
ive
24
A-H
24
A-M
24
A-L
23
B-H
23
B-M
23
B-L
lowast
Figure 2 Results of the protein expression of HMG-CoA reductasein mice liver tissue protein (119899 = 11) Note compared to the blankgroup 998771998771119875 lt 001 and 998771119875 lt 005 compared to the model grouplowastlowast119875 lt 001 and lowast119875 lt 005 24A-H alisol A 24-acetate high 24A-
M alisol A 24-acetate medium and 24A-L alisol A 24-acetate low23B-H alisol B 23-acetate high 23B-M alisol B 23-acetate mediumand 23B-L alisol B 24-acetate low
Table 4 The length (gt) and the angle of hydrogen bond formedbetween alisol acetates and HMG-CoA reductase
Name XndashHsdot sdot sdotYd
(XndashH)
d(Hsdot sdot sdotY)
d(Xsdot sdot sdotY) Angle XHY
Alisol B
23-acetate
Lys691 NZ-HZ1sdot sdot sdotO37 104 237 298 11594
Asp767OD2-
HH88sdot sdot sdotO2096 240 324 14490
Alisol A24-acetate
Asn658 OD1-H5Zsdot sdot sdotO5 095 201 286 14897
After docking the conformation of alisol B 23-acetatechanged as shown in Figure 3(c) The calculated root-mean-square deviation (RMSD) was 289gt The conformationof the small molecule changed from the clustered sheetstructure to the open structure Before docking the sidechain and the ring structure of the small molecule foldedas shown in Figures 3(d) and 3(e) The angle between C
2ndash
C10ndashC19was 1095∘ 1095∘ between C
10ndashC19ndashC20 and minus599∘
between C2ndashC10ndashC19ndashC20 As shown in Figures 3(f) and
3(g) after docking the angle between C2ndashC10ndashC19was 1136∘
1156∘ between C10ndashC19ndashC20 and 1260∘ between C
2ndashC10ndash
C19ndashC20 The angle between the parent ring and the side
chain was significantly increasedThe twisting of the dihedralangles resulted in the opening of side chains and insertioninto the protein which led to interactions There was ahydrophilic ether bond on the side chain and the peripheralamino acids were mainly hydrophilic (Figures 3(h) and 3(i))
This created a hydrophilic environment around the sidechains culminating in stable binding The energy of thevan der Waals forces between them was calculated to beminus1519 KJmol whereas the electrostatic energy was calcu-lated to be minus2366 KJmol As the electrostatic energy waslower than the energy of the van derWaals forces between thesystems the electrostatic interactionwas themain interactionbetween this small molecule and the macromolecule Thebinding energy (ΔG∘) between the alisol B 23-acetate and theHMG-CoA reductase was calculated to be minus1033 KJmol
342 Molecular Simulation Results of Alisol A 24-Acetateand HMG-CoA Reductase Themolecular simulation resultsfor the interaction between alisol A 24-acetate and HMG-CoA reductase are shown in Figure 4 Figure 4(a) shows theoverall interaction pattern of this compound and HMG-CoAreductase The interaction pocket where it bound to HMG-CoA reductase contained the following amino acids Tyr519Ser520 Leu521 Val522 Met523 Gly524 Ala525 Cys527Glu528 Asn529 Val530 Ile531 Gly532 Try533 Met588Thy589 Arg590 Gly591 Asp653 Ala654 Met655 Gly656Met657 Asn658 Met659 Ile660 Ser661 Lys662 Gly663Thy664 Glu665 Cys688 The689 Asp690 Lys691 Lys692Cys764 Gly765 Gln766 Asp767 Ala768 Ala769 Gln770Asn771 Val772 Gly773 Ser774 Glu801 Ile802 Gly803Thr804Val805Gly806Gly807Gly808Thr809 andAsn810There were 439 (450ndash888) amino acids in the catalyticdomain at the C-terminal After entering the interactionpocket alisol A 24-acetate exhibited hydrogen bonding andhydrophobic and hydrophilic interactions with peripheralamino acid residues Figure 4(b) shows the hydrogen bondcontact map There was one hydrogen bond between alisol A24 acetate and amino acid Asn658 (Table 4) The interactionenergy of the hydrophobic amino acid residues and thesmall molecule was calculated to be 465 KJmol whereas theinteraction energy of the hydrophilic amino acid residues andthe small molecule was calculated to be minus1023 KJmol Thissuggests that the hydrophilic groups had a much strongereffect than the hydrophobic groups The small molecule andthe peripheral amino acids exhibited a stronger hydrophilicinteraction as indicated by the interface graphs (Figures 4(f)and 4(g))
After docking the alisol A 24-acetate conformationchanged (Figure 4(c)) The RMSD was calculated to be334gt The conformation angle C
3ndashC18ndashC26ndashC28
changedfrom 600∘ before docking to 1242∘ after docking (Figures4(d) and 4(e)) This suggests a large torsion of the smallmolecule side chain after docking whereby the stretchedopen state changed to a ring-like sheet state Consequentlyit was difficult to insert the side chain into the protein andinteract with the active pocket of the protein This resultedin considerably lower binding relative to alisol B 23-acetateThe electrostatic energy of alisol A 24-acetate and HMG-CoA reductase was determined to be minus886 KJmol whilethe energy of the van der Waals forces was determined tobe minus1649 KJmol The energy of the van der Waals forceswas lower than the electrostatic energy showing that themain interaction between this small molecule and themacro-molecule was the van derWaals interactionThe ΔG∘ of alisol
Evidence-Based Complementary and Alternative Medicine 7
(a) (b) (c)
(d) (e) (f)
(g) (h) (i)
Figure 3 Interaction between alisol B 23-acetate and HMG-CoA reductase (a) Overall pattern of interaction (alisol B 23-acetate is denotedby a bar chart) (b) Hydrogen bond graph of alisol B 23-acetate with Lys691 and Asp767 (the hydrogen bond is denoted by a dotted line) (c)Stacking chart of alisol B 23-acetate before and after docking (the initial structure is shown in green and the structure after docking is shown inpurple) (d) Initial angle of alisol B 23-acetate C
2ndashC10ndashC19and C
10ndashC19ndashC20 (e) Initial dihedral angle of alisol B 23-acetate C
2ndashC10ndashC19ndashC20
(f) The angle of alisol B 23-acetate C2ndashC10ndashC19and C
10ndashC19ndashC20after stacking (g)The dihedral angle of alisol B 23-acetate C
2ndashC10ndashC19ndashC20
after stacking (h) Hydrophobic interface of the alisol B 23-acetate side chain and the peripheral amino acid residues after stacking (alisol B23-acetate is denoted by yellow bars) (i) Hydrophilic interface of the alisol B 23-acetate side chain and the peripheral amino acid residuesafter stacking (alisol B 23-acetate is denoted by yellow bars)
A 24-acetate and HMG-CoA reductase was calculated to beminus23 KJmol
4 Discussion
Various experimental and clinical studies focus on the bloodlipid reducing effects of different solvent extracts (especially
water and alcohol extracts) fromAlisma orientale and its pro-cessed products [36ndash38] However the mechanisms under-lying the effects of Alisma orientale constituents remain to berevealed Alisma orientalemay interfere with exogenous cho-lesterol absorption and endogenous cholesterol metabolismthus reducing the level of TC Moreover alisol acetates mayinterfere with themetabolism of endogenous cholesterol [39]
8 Evidence-Based Complementary and Alternative Medicine
(a) (b) (c)
(d) (e) (f)
(g)
Figure 4 The interaction between alisol A 24-acetate and HMG-CoA reductase (a) Overall pattern of interaction (alisol A 24-acetate isdenoted by a bar chart) (b) Hydrogen bond graph of alisol A 24-acetate and Asn658 (the hydrogen bond is denoted by a dotted line) (c)Stacking chart of alisol A 24-acetate before and after docking (the initial structure is shown in green and the structure after docking is shownin purple) (d) The initial dihedral angle of alisol A 24-acetate angle C
3ndashC18ndashC26ndashC28 (e) The dihedral angle of alisol A 24-acetate angle
C3ndashC18ndashC26ndashC28after stacking (f) Hydrophobic interface of the alisol A 24-acetate side chain and the peripheral amino acid residues after
stacking (alisol A 24-acetate is denoted by yellow bars) (g) Hydrophilic interface of the alisol A 24-acetate side chain and the peripheralamino acid residues after stacking (alisol A 24-acetate is denoted by yellow bars)
Therefore we studied the TC-lowering effect of alisolacetates based on HMG-CoA reductase and the underly-ing molecular mechanism As the main source of endoge-nous cholesterol and one of the key enzymes in the TCmetabolism HMG-CoA reductase is the first described rate-limiting enzyme in cholesterol synthesis in vivo Moreover its
activation directly affects the speed of cholesterol synthesisand the level of TC in vivo
The TC-lowering effect and molecular mechanism ofalisol acetates the main active ingredient of Alismatis rhi-zoma were investigated using the reagent kit method andwestern blotting technique combined with the molecular
Evidence-Based Complementary and Alternative Medicine 9
simulation technique An interaction model was built foralisol acetate and the key enzyme involved in the metabolismof TC was HMG-CoA reductase The present study revealsthat alisol acetates can significantly lower the TC level ofhyperlipidemic miceThe alisol B 23-acetate showed a higherextent of TC lowering than alisol A 24-acetate
Consistent with the in vitro results alisol acetates couldalso lower the in vivoHMG-CoA reductase activity in a dose-dependent fashion with a higher degree of enzyme inhibitionfor alisol B 23-acetate compared to alisol A 24-acetate Theseresults suggest that alisol acetates might lower TC levels viaregulation of HMG-CoA reductase activity The consistencybetween the results of the in vivo and in vitro experimentsindicates that the alisol acetate prototype drug inhibitsHMG-CoA reductase activity instead of the in vivometabolites
Revealed by the western blotting results alisol acetatesmight inhibit the activity of HMG-CoA reductase by directlyand competitively binding to HMG-CoA reductase ratherthan by downregulating its protein expression
The molecular simulation technique was employed inorder to investigate the interaction of alisol acetates andHMG-CoA reductase The results (Figures 3 and 4 andTable 4) revealed themolecularmechanism of the interactionof alisol acetates and HMG-CoA reductase Both alisolacetates bound to HMG-CoA reductase in their catalyticdomain at the C-terminalThe alisol B 23-acetate formed twohydrogen bonds with HMG-CoA reductase The hydrophilicinteraction was stronger than the hydrophobic interactionwhereas the electrostatic interaction was stronger than thevan der Waals forces After the interaction the structure ofthe small molecule dramatically changed from the originalfolded sheet to an open structure Additionally the side chainstretched and was inserted into the protein The peripheralamino acids formed a hydrophilic environment and stablybound with it In general a hydrogen bond with a lengthof less than 35 A and an angle of 90ndash180∘ is consideredto be strong These two hydrogen bonds were relativelystrong and might be partly responsible for the torsion ofthe small molecule The alisol A 24-acetate formed onehydrogen bond with the HMG-CoA reductase and this bondwas weaker compared to those of alisol B 23-acetate Thehydrophilic interaction was stronger than the hydrophobicinteraction and the van derWaals force was stronger than theelectrostatic interaction After the interaction the moleculeside chain had large torsion and changed from the originalstretched open state to a ring-like sheet state which wasdifficult to insert into the proteinThe binding energy of alisolB 23-acetate andHMG-CoA reductase was lower than that ofalisol A 24-acetate and HMG-CoA reductase indicating thatthe binding of alisol B 23-acetate and HMG-CoA reductasewas stable According to the pharmacological results alisolB 23-acetate lowered the HMG-CoA reductase activity morethan alisol A 24-acetate This suggests that the active centerwas the interaction pocket constituted by these amino acidsand that the hydrogen bonds involving Lys691 Asp767 andAsn658 were the key amino acid residues for the interactionof this protein with alisol acetates
The binding of alisol B 23-acetate and HMG-CoA reduc-tase was considerably stronger than the binding of alisol
A 24-acetate and HMG-CoA reductase The relatively weakbinding of alisol A 24-acetate and HMG-CoA reductasemight be due to its distinct side-chain structure The sidechain of alisol B 23-acetate contained an ether bond whichwas strongly hydrophilic and could be inserted into thehydrophilic environment of the protein to form strong inter-actions Previous studies found that the side chains of alisolacetates were their active groups [40] In the present studythis conclusionwas validated A folded side chainparent ringbound more strongly to the macromolecule than an openside-chainparent ring did Thus the side chain acts as thesteering group in that it steers the interactions between alisolacetates and macromolecules This might be the key groupfor interactions with macromolecules as it had a decisiveeffect on the interactions of this type of compound withmacromolecules Our previous studies indicated that alisol B23-acetate was partially converted to alisol A 24-acetate in theenvironment of the human stomach However the bindingof alisol A 24-acetate and HMG-CoA reductase was weaker[14 41 42] Therefore it is advisable for drug developers todevelop a dosage form that circumvents the conversion ofalisol B 23-acetate in the stomach
Previously published studies indicate that [43 44] themechanism of reducing the level of TC is that two acetylCoA molecules condense to acetyl CoA under the catalysisof acetoacetyl-coenzyme A thiolase (AACT) acetyl CoAand acetyl CoA condense to HMG-CoA and HMG-CoAreduces to MVA under the catalytic action of HMG-CoAreductase MVA further generates TC with squalene Thehigh intracellular free fatty acid (FC) level stimulates micro-somal acyl coenzyme A-cholesterol acyltransferase (ACAT)and promotes esterification of the cholesterol Subsequentlycholesterol is stored in the form of cholesterol esters (CE)which is a feedback inhibition of HMG-CoA reductasesynthesis so that cholesterol synthesis rate-limiting enzymeswill be inhibited thus reducing intracellular cholesterolsynthesis This process features three important enzymesAACT HMG-CoA reductase and ACAT Whether AACTand ACAT are involved in the effect of alisol acetates will bethe subject of intensive research in the future
The results of the study indicate that alisol acetates cannot only decrease TC level but also decrease TG and LDL-C levels and increase HDL-C level Furthermore a follow-upstudy is designed to investigate the regulatory mechanisms ofalisol acetates on TG HDL-C and LDL-C
5 Conclusion
The present study reveals that alisol acetates could signifi-cantly lower the TC level of hyperlipidemic mice The effectof alisol B 23-acetate on HMG-CoA reductase was strongerthan that of alisol A 24-acetate Alisol acetates could lowerHMG-CoA reductase activity in a dose-dependent fashionboth in vivo and in vitro Neither of these alisol acetatescould significantly lower the protein expression of HMG-CoA This suggests that alisol acetates lower the TC levelvia inhibiting the activity of HMG-CoA reductase via itsprototype drug which may have an inhibitory effect bydirectly and competitively binding with HMG-CoA In the
10 Evidence-Based Complementary and Alternative Medicine
binding interaction of alisol acetate andHMG-CoA reductasevia molecular simulation the key amino acid residues ofHMG-CoA reductase might be Lys691 Asp767 and Asn658The side chain of the alisol acetatewas the steering group Fur-thermore a folded side chainparent ring boundmoreweaklyto HMG-CoA reductase than an open side chainparent ringThis study explored the TC-lowering mechanism of alisolacetates at the molecular level and presented guidance for itsclinical use
Competing Interests
The authors declare that they have no competing interests
Acknowledgments
This work was financially supported by the National NatureScience Foundation of China (81303173 81673534) the Nat-ural Science Foundation of Jiangsu Province (BK20161576)the Natural Science Fund for Colleges and Universities inJiangsu Province (14KJB360001) and the Top-Notch Aca-demic Programs Project of Jiangsu Higher Education Insti-tutions (PPZY2015A070)
References
[1] L X Wang Q N Wu Q Zhang G P Peng and A W DingldquoBasic study of diuretic active compounds in Rhizoma Alis-matisrdquo West China Journal of Pharmaceutical Sciences vol 23no 6 pp 670ndash672 2008
[2] S S Wu Pharmaceutical and Clinical Research of Alisma Ori-entalis Chinese Press Beijing China 2007
[3] M Miyazawa S Yoshinaga and Y Kashima ldquoChemical com-position and characteristic odor compounds in essential oilfrom Alismatis Rhizoma (Tubers of Alisma orientale)rdquo DiatomResearch vol 23 no 1 pp 133ndash141 2016
[4] M-K Jang Y-R Han J S Nam et al ldquoProtective effects ofAlisma orientale extract against hepatic steatosis via inhibitionof endoplasmic reticulum stressrdquo International Journal ofMolec-ular Sciences vol 16 no 11 pp 26151ndash26165 2015
[5] Z-P Mai K Zhou G-B Ge et al ldquoProtostane triterpenoidsfrom the rhizome of Alisma orientale exhibit inhibitory effectson human carboxylesterase 2rdquo Journal of Natural Products vol78 no 10 pp 2372ndash2380 2015
[6] T Tian H Chen andY-Y Zhao ldquoTraditional uses phytochem-istry pharmacology toxicology and quality control of Alismaorientale (Sam) Juzep a reviewrdquo Journal of Ethnopharmacologyvol 158 pp 373ndash387 2014
[7] KHKimM J Kwun J-Y Choi et al ldquoTherapeutic effect of thetuber of Alisma orientale on lipopolysaccharide-induced acutelung injuryrdquo Evidence-Based Complementary and AlternativeMedicine vol 2013 Article ID 863892 10 pages 2013
[8] H-G Jin Q Jin A R Kim et al ldquoA new triterpenoid fromAlisma orientale and their antibacterial effectrdquoArchives of Phar-macal Research vol 35 no 11 pp 1919ndash1926 2012
[9] J H Lee O S Kwon H-G Jin E-R Woo Y S Kim and H PKim ldquoThe rhizomes of Alisma orientale and alisol derivativesinhibit allergic response and experimental atopic dermatitisrdquoBiological and Pharmaceutical Bulletin vol 35 no 9 pp 1581ndash1587 2012
[10] WGu C GengW Xue et al ldquoCharacterization and function ofthe 3-hydroxy-3-methylglutaryl-CoA reductase gene in Alismaorientale (Sam) Juz and its relationship with protostane triter-pene productionrdquo Plant Physiology and Biochemistry vol 97 pp378ndash389 2015
[11] W Gu B L Xi Q N Wu et al ldquoCloning and distribution pat-tern of HMGR gene conserved fragment in Alisma orientalerdquoJournal of Chinese Medicinal Materials vol 34 no 9 pp 1335ndash1339 2011
[12] T Kobayashi S Kato-Emori K Tomita and H Ezura ldquoDetec-tion of 3-hydroxy-3-methylglutaryl-coenzymeA reductase pro-tein Cm-HMGR during fruit development in melon (Cucumismelo L)rdquo Theoretical and Applied Genetics vol 104 no 5 pp779ndash785 2002
[13] R Tommer A Rachel P C Sylvie and R Jerry ldquoImpaired regu-lation of 3-hydroxy-3-methylglutaryl-coenzyme A reductasedegradation in lovastatin-resistant cellsrdquo The Journal of Bioog-ical Chemistry vol 274 no 41 pp 29341ndash29351 1999
[14] F Xu Q-N Wu J Chen et al ldquoThe binding mechanisms ofplasma protein to active compounds in Alismaorientale rhi-zomes (Alismatis Rhizoma)rdquo Bioorganic and Medicinal Chem-istry Letters vol 24 no 17 pp 4099ndash4105 2014
[15] G Gong Y Qin W Huang et al ldquoProtective effects of dios-genin in the hyperlipidemic rat model and in human vascularendothelial cells against hydrogen peroxide-induced apoptosisrdquoChemico-Biological Interactions vol 184 no 3 pp 366ndash3752010
[16] T F Tzeng H J Lu S S Lion C J Chang and IM Liu ldquoLipid-lowering effects of zerumbone a natural cyclic sesquiterpeneof Zingiber zerumbet Smith in high-fat diet-induced hyperlipi-demic hamstersrdquoFood andChemical Toxicology vol 69 pp 132ndash139 2014
[17] Y Xie H Zhang H Liu et al ldquoHypocholesterolemic effectsof Kluyveromyces marxianus M3 isolated from Tibetan mush-rooms on diet-induced hypercholesterolemia in ratrdquo BrazilianJournal of Microbiology vol 46 no 2 pp 389ndash395 2015
[18] I Karam N Ma X-W Liu et al ldquoRegulation effect of AspirinEugenol Ester on blood lipids in Wistar rats with hyperlipi-demiardquo BMC Veterinary Research vol 11 article 217 2015
[19] G Jiao J B Wei M H Yin and P T Chun ldquoA new TCMformula FTZ lowers serum cholesterol by regulating HMG-CoA reductase and CYP7A1 in hyperlipidemic ratsrdquo Journal ofEthnopharmacology vol 135 no 2 pp 299ndash307 2011
[20] Y Geng W L Tu J J Zhang L Zhang and J Zhang ldquoEffectsof extract from cole pollen on lipidmetabolism in experimentalhyperlipidemic ratsrdquo The Scientific World Journal vol 2014Article ID 982498 6 pages 2014
[21] K Sueyoshi Y Nogawa K Sugawara T Endo and HHisamoto ldquoHighly sensitive and multiple enzyme activity assayusing reagent-release capillary-isoelectric focusing with rhod-amine 110-based substratesrdquo Analytical Sciences vol 31 no 11pp 1155ndash1161 2015
[22] D Fteita E Kononen M Gursoy E Soderling and U KGursoy ldquoDoes estradiol have an impact on the dipeptidylpeptidase IV enzyme activity of the Prevotella intermedia groupbacteriardquo Anaerobe vol 36 pp 14ndash18 2015
[23] L L Zhuo LWang and N Zhou ldquoRandomly arrayed G-quad-ruplexes for label-free and real-time assay of enzyme activityrdquoChemical Communications vol 50 no 52 pp 6875ndash6878 2014
[24] M Garelnabi and A Younis ldquoParaoxonase-1 enzyme activityassay for clinical samples validation and correlation studiesrdquoMedical Science Monitor vol 21 pp 902ndash908 2015
Evidence-Based Complementary and Alternative Medicine 11
[25] X Bi andZ Liu ldquoEnzyme activity assay of glycoprotein enzymesbased on a boronate affinity molecularly imprinted 96-wellmicroplaterdquo Analytical Chemistry vol 86 no 24 pp 12382ndash12389 2014
[26] A Walls S Dermody R Kumaran N Krishnan and E HHarley ldquoCharacterization of B-Cells in tonsils of patients diag-nosed with pediatric autoimmune neuropsychiatric disorderassociated streptococcusrdquo International Journal of PediatricOtorhinolaryngology vol 80 pp 49ndash52 2016
[27] I Karam N Ma X-W Liu et al ldquoRegulation effect of AspirinEugenol Ester on blood lipids in Wistar rats with hyperlipi-demiardquo BMC Veterinary Research vol 11 no 1 article 217 2015
[28] C Sunil S Ignacimuthu and C Kumarappan ldquoHypolipidemicactivity of Symplocos cochinchinensis S Moore leaves in hyper-lipidemic ratsrdquo Journal of Natural Medicines vol 66 no 1 pp32ndash38 2012
[29] W Shen Y Han B Huang et al ldquoMicroRNA-483-3p inhibitsextracellular matrix production by targeting smad4 in humantrabecular meshwork cellsrdquo Investigative Ophthalmology amp Vis-ual Science vol 56 no 13 pp 8419ndash8427 2015
[30] Z Wu Z Yi L Dong and A Zhang ldquoMolecular simulationstudy of the specific combination between four kinds of phthalicacid esters and human serum albuminrdquo Environmental Toxicol-ogy and Pharmacology vol 41 pp 259ndash265 2016
[31] K-J Hwang M Yoon H-J Hwang S-M Choi and S-MJeong ldquoMolecular dynamics simulation of oxygen ion conduc-tion in orthorhombic perovskite Ba-doped LaInO
3using cubic
and orthorhombic modelrdquo Journal of Nanoscience and Nan-otechnology vol 15 no 11 pp 8947ndash8950 2015
[32] H Yu W Han W Ma and K Schulten ldquoTransient 120573-hairpinformation in 120572-synuclein monomer revealed by coarse-grainedmolecular dynamics simulationrdquo The Journal of ChemicalPhysics vol 143 no 24 Article ID 243142 2015
[33] M Breuer K M Rosso and J Blumberger ldquoFlavin binding tothe deca-heme cytochrome mtrc insights from computationalmolecular simulationrdquo Biophysical Journal vol 109 no 12 pp2614ndash2624 2015
[34] V Rastija D Agic S Tomic et al ldquoSynthesis QSAR and mole-cular dynamics simulation of amidino-substituted benzimi-dazoles as dipeptidyl peptidase III inhibitorsrdquo Acta ChimicaSlovenica vol 62 no 4 pp 867ndash878 2015
[35] M Enciso C Schutte and L Delle Site ldquoInfluence of pH andsequence in peptide aggregation via molecular simulationrdquoTheJournal of Chemical Physics vol 143 no 24 Article ID 2431302015
[36] L XWangQWu andG P Peng ldquoA Study on determination ofcontent of 23-acetyl alisol B in Rhizoma Alismatis orientalitisrdquoJournal of Nanjing University of Traditional Chinese Medicine(Natural Science) vol 18 no 2 pp 105ndash107 2002
[37] G P Peng andY L Zhu ldquoProgress in the study on chemical con-stituents of Alisma orientalisrdquo Natural Product Research andDevelopment vol 18 no 2 pp 348ndash351 2006
[38] D Y Liu S J Gu J Wu et al ldquoStudies on fractions of zexiedecoction with antihyperlipidemia bioactivity in micerdquo ChinaPharmacist vol 13 no 6 pp 763ndash766 2010
[39] Q-F Luo L Sun and G-H Du ldquoProgress in new drugs tar-geting reverse cholesterol transportrdquo Chinese PharmacologicalBulletin vol 22 no 8 pp 904ndash907 2006
[40] M Yoshikawa and Y Fukuda ldquoHatakeayamardquo Chemical andPharmaceutical Bulletin vol 41 no 6 pp 1194ndash1196 1993
[41] F Xu L Zhang LHe et al ldquoMolecularmechanismon the inter-action between alisols and human serum albuminrdquo Acta Chim-ica Sinica vol 69 no 19 pp 2228ndash2234 2011
[42] F Xu J ChenW Gu et al ldquoStudy on transformation of effectivecomponents of Alisma under simulated in vivo conditionsrdquoChinese Journal of Pharmaceutical Analysis vol 31 no 11 pp2035ndash2039 2011
[43] H Yang Y G Gao and L X Zhang ldquoResearch progress on bio-synthetic pathway of terpenoids containing ginsenoside and theHMGRrdquo China Journal of Biological Engineering vol 28 no 10pp 130ndash135 2008
[44] G Z Cui Y L Ben N Li and D L Liu ldquoCatalytic mechanismand characteristics of HMG-COA recuctaserdquo Journal ofMedicalMolecular Biology vol 5 no 6 p 542 2008
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom
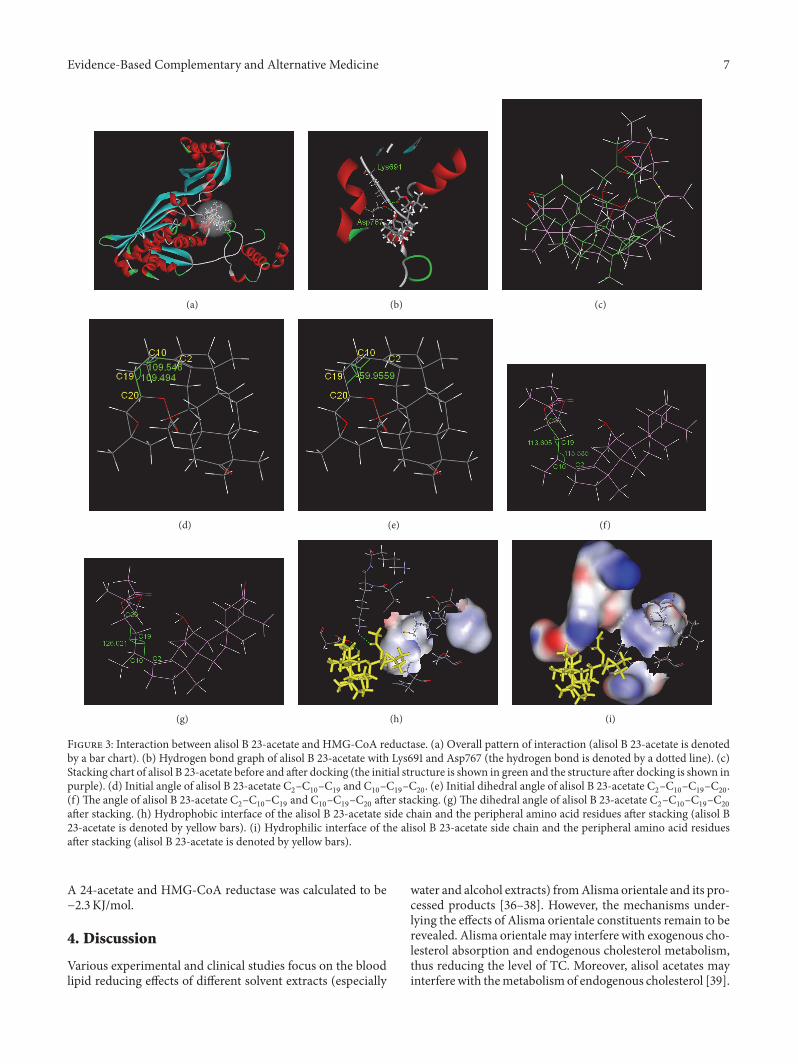
Evidence-Based Complementary and Alternative Medicine 7
(a) (b) (c)
(d) (e) (f)
(g) (h) (i)
Figure 3 Interaction between alisol B 23-acetate and HMG-CoA reductase (a) Overall pattern of interaction (alisol B 23-acetate is denotedby a bar chart) (b) Hydrogen bond graph of alisol B 23-acetate with Lys691 and Asp767 (the hydrogen bond is denoted by a dotted line) (c)Stacking chart of alisol B 23-acetate before and after docking (the initial structure is shown in green and the structure after docking is shown inpurple) (d) Initial angle of alisol B 23-acetate C
2ndashC10ndashC19and C
10ndashC19ndashC20 (e) Initial dihedral angle of alisol B 23-acetate C
2ndashC10ndashC19ndashC20
(f) The angle of alisol B 23-acetate C2ndashC10ndashC19and C
10ndashC19ndashC20after stacking (g)The dihedral angle of alisol B 23-acetate C
2ndashC10ndashC19ndashC20
after stacking (h) Hydrophobic interface of the alisol B 23-acetate side chain and the peripheral amino acid residues after stacking (alisol B23-acetate is denoted by yellow bars) (i) Hydrophilic interface of the alisol B 23-acetate side chain and the peripheral amino acid residuesafter stacking (alisol B 23-acetate is denoted by yellow bars)
A 24-acetate and HMG-CoA reductase was calculated to beminus23 KJmol
4 Discussion
Various experimental and clinical studies focus on the bloodlipid reducing effects of different solvent extracts (especially
water and alcohol extracts) fromAlisma orientale and its pro-cessed products [36ndash38] However the mechanisms under-lying the effects of Alisma orientale constituents remain to berevealed Alisma orientalemay interfere with exogenous cho-lesterol absorption and endogenous cholesterol metabolismthus reducing the level of TC Moreover alisol acetates mayinterfere with themetabolism of endogenous cholesterol [39]
8 Evidence-Based Complementary and Alternative Medicine
(a) (b) (c)
(d) (e) (f)
(g)
Figure 4 The interaction between alisol A 24-acetate and HMG-CoA reductase (a) Overall pattern of interaction (alisol A 24-acetate isdenoted by a bar chart) (b) Hydrogen bond graph of alisol A 24-acetate and Asn658 (the hydrogen bond is denoted by a dotted line) (c)Stacking chart of alisol A 24-acetate before and after docking (the initial structure is shown in green and the structure after docking is shownin purple) (d) The initial dihedral angle of alisol A 24-acetate angle C
3ndashC18ndashC26ndashC28 (e) The dihedral angle of alisol A 24-acetate angle
C3ndashC18ndashC26ndashC28after stacking (f) Hydrophobic interface of the alisol A 24-acetate side chain and the peripheral amino acid residues after
stacking (alisol A 24-acetate is denoted by yellow bars) (g) Hydrophilic interface of the alisol A 24-acetate side chain and the peripheralamino acid residues after stacking (alisol A 24-acetate is denoted by yellow bars)
Therefore we studied the TC-lowering effect of alisolacetates based on HMG-CoA reductase and the underly-ing molecular mechanism As the main source of endoge-nous cholesterol and one of the key enzymes in the TCmetabolism HMG-CoA reductase is the first described rate-limiting enzyme in cholesterol synthesis in vivo Moreover its
activation directly affects the speed of cholesterol synthesisand the level of TC in vivo
The TC-lowering effect and molecular mechanism ofalisol acetates the main active ingredient of Alismatis rhi-zoma were investigated using the reagent kit method andwestern blotting technique combined with the molecular
Evidence-Based Complementary and Alternative Medicine 9
simulation technique An interaction model was built foralisol acetate and the key enzyme involved in the metabolismof TC was HMG-CoA reductase The present study revealsthat alisol acetates can significantly lower the TC level ofhyperlipidemic miceThe alisol B 23-acetate showed a higherextent of TC lowering than alisol A 24-acetate
Consistent with the in vitro results alisol acetates couldalso lower the in vivoHMG-CoA reductase activity in a dose-dependent fashion with a higher degree of enzyme inhibitionfor alisol B 23-acetate compared to alisol A 24-acetate Theseresults suggest that alisol acetates might lower TC levels viaregulation of HMG-CoA reductase activity The consistencybetween the results of the in vivo and in vitro experimentsindicates that the alisol acetate prototype drug inhibitsHMG-CoA reductase activity instead of the in vivometabolites
Revealed by the western blotting results alisol acetatesmight inhibit the activity of HMG-CoA reductase by directlyand competitively binding to HMG-CoA reductase ratherthan by downregulating its protein expression
The molecular simulation technique was employed inorder to investigate the interaction of alisol acetates andHMG-CoA reductase The results (Figures 3 and 4 andTable 4) revealed themolecularmechanism of the interactionof alisol acetates and HMG-CoA reductase Both alisolacetates bound to HMG-CoA reductase in their catalyticdomain at the C-terminalThe alisol B 23-acetate formed twohydrogen bonds with HMG-CoA reductase The hydrophilicinteraction was stronger than the hydrophobic interactionwhereas the electrostatic interaction was stronger than thevan der Waals forces After the interaction the structure ofthe small molecule dramatically changed from the originalfolded sheet to an open structure Additionally the side chainstretched and was inserted into the protein The peripheralamino acids formed a hydrophilic environment and stablybound with it In general a hydrogen bond with a lengthof less than 35 A and an angle of 90ndash180∘ is consideredto be strong These two hydrogen bonds were relativelystrong and might be partly responsible for the torsion ofthe small molecule The alisol A 24-acetate formed onehydrogen bond with the HMG-CoA reductase and this bondwas weaker compared to those of alisol B 23-acetate Thehydrophilic interaction was stronger than the hydrophobicinteraction and the van derWaals force was stronger than theelectrostatic interaction After the interaction the moleculeside chain had large torsion and changed from the originalstretched open state to a ring-like sheet state which wasdifficult to insert into the proteinThe binding energy of alisolB 23-acetate andHMG-CoA reductase was lower than that ofalisol A 24-acetate and HMG-CoA reductase indicating thatthe binding of alisol B 23-acetate and HMG-CoA reductasewas stable According to the pharmacological results alisolB 23-acetate lowered the HMG-CoA reductase activity morethan alisol A 24-acetate This suggests that the active centerwas the interaction pocket constituted by these amino acidsand that the hydrogen bonds involving Lys691 Asp767 andAsn658 were the key amino acid residues for the interactionof this protein with alisol acetates
The binding of alisol B 23-acetate and HMG-CoA reduc-tase was considerably stronger than the binding of alisol
A 24-acetate and HMG-CoA reductase The relatively weakbinding of alisol A 24-acetate and HMG-CoA reductasemight be due to its distinct side-chain structure The sidechain of alisol B 23-acetate contained an ether bond whichwas strongly hydrophilic and could be inserted into thehydrophilic environment of the protein to form strong inter-actions Previous studies found that the side chains of alisolacetates were their active groups [40] In the present studythis conclusionwas validated A folded side chainparent ringbound more strongly to the macromolecule than an openside-chainparent ring did Thus the side chain acts as thesteering group in that it steers the interactions between alisolacetates and macromolecules This might be the key groupfor interactions with macromolecules as it had a decisiveeffect on the interactions of this type of compound withmacromolecules Our previous studies indicated that alisol B23-acetate was partially converted to alisol A 24-acetate in theenvironment of the human stomach However the bindingof alisol A 24-acetate and HMG-CoA reductase was weaker[14 41 42] Therefore it is advisable for drug developers todevelop a dosage form that circumvents the conversion ofalisol B 23-acetate in the stomach
Previously published studies indicate that [43 44] themechanism of reducing the level of TC is that two acetylCoA molecules condense to acetyl CoA under the catalysisof acetoacetyl-coenzyme A thiolase (AACT) acetyl CoAand acetyl CoA condense to HMG-CoA and HMG-CoAreduces to MVA under the catalytic action of HMG-CoAreductase MVA further generates TC with squalene Thehigh intracellular free fatty acid (FC) level stimulates micro-somal acyl coenzyme A-cholesterol acyltransferase (ACAT)and promotes esterification of the cholesterol Subsequentlycholesterol is stored in the form of cholesterol esters (CE)which is a feedback inhibition of HMG-CoA reductasesynthesis so that cholesterol synthesis rate-limiting enzymeswill be inhibited thus reducing intracellular cholesterolsynthesis This process features three important enzymesAACT HMG-CoA reductase and ACAT Whether AACTand ACAT are involved in the effect of alisol acetates will bethe subject of intensive research in the future
The results of the study indicate that alisol acetates cannot only decrease TC level but also decrease TG and LDL-C levels and increase HDL-C level Furthermore a follow-upstudy is designed to investigate the regulatory mechanisms ofalisol acetates on TG HDL-C and LDL-C
5 Conclusion
The present study reveals that alisol acetates could signifi-cantly lower the TC level of hyperlipidemic mice The effectof alisol B 23-acetate on HMG-CoA reductase was strongerthan that of alisol A 24-acetate Alisol acetates could lowerHMG-CoA reductase activity in a dose-dependent fashionboth in vivo and in vitro Neither of these alisol acetatescould significantly lower the protein expression of HMG-CoA This suggests that alisol acetates lower the TC levelvia inhibiting the activity of HMG-CoA reductase via itsprototype drug which may have an inhibitory effect bydirectly and competitively binding with HMG-CoA In the
10 Evidence-Based Complementary and Alternative Medicine
binding interaction of alisol acetate andHMG-CoA reductasevia molecular simulation the key amino acid residues ofHMG-CoA reductase might be Lys691 Asp767 and Asn658The side chain of the alisol acetatewas the steering group Fur-thermore a folded side chainparent ring boundmoreweaklyto HMG-CoA reductase than an open side chainparent ringThis study explored the TC-lowering mechanism of alisolacetates at the molecular level and presented guidance for itsclinical use
Competing Interests
The authors declare that they have no competing interests
Acknowledgments
This work was financially supported by the National NatureScience Foundation of China (81303173 81673534) the Nat-ural Science Foundation of Jiangsu Province (BK20161576)the Natural Science Fund for Colleges and Universities inJiangsu Province (14KJB360001) and the Top-Notch Aca-demic Programs Project of Jiangsu Higher Education Insti-tutions (PPZY2015A070)
References
[1] L X Wang Q N Wu Q Zhang G P Peng and A W DingldquoBasic study of diuretic active compounds in Rhizoma Alis-matisrdquo West China Journal of Pharmaceutical Sciences vol 23no 6 pp 670ndash672 2008
[2] S S Wu Pharmaceutical and Clinical Research of Alisma Ori-entalis Chinese Press Beijing China 2007
[3] M Miyazawa S Yoshinaga and Y Kashima ldquoChemical com-position and characteristic odor compounds in essential oilfrom Alismatis Rhizoma (Tubers of Alisma orientale)rdquo DiatomResearch vol 23 no 1 pp 133ndash141 2016
[4] M-K Jang Y-R Han J S Nam et al ldquoProtective effects ofAlisma orientale extract against hepatic steatosis via inhibitionof endoplasmic reticulum stressrdquo International Journal ofMolec-ular Sciences vol 16 no 11 pp 26151ndash26165 2015
[5] Z-P Mai K Zhou G-B Ge et al ldquoProtostane triterpenoidsfrom the rhizome of Alisma orientale exhibit inhibitory effectson human carboxylesterase 2rdquo Journal of Natural Products vol78 no 10 pp 2372ndash2380 2015
[6] T Tian H Chen andY-Y Zhao ldquoTraditional uses phytochem-istry pharmacology toxicology and quality control of Alismaorientale (Sam) Juzep a reviewrdquo Journal of Ethnopharmacologyvol 158 pp 373ndash387 2014
[7] KHKimM J Kwun J-Y Choi et al ldquoTherapeutic effect of thetuber of Alisma orientale on lipopolysaccharide-induced acutelung injuryrdquo Evidence-Based Complementary and AlternativeMedicine vol 2013 Article ID 863892 10 pages 2013
[8] H-G Jin Q Jin A R Kim et al ldquoA new triterpenoid fromAlisma orientale and their antibacterial effectrdquoArchives of Phar-macal Research vol 35 no 11 pp 1919ndash1926 2012
[9] J H Lee O S Kwon H-G Jin E-R Woo Y S Kim and H PKim ldquoThe rhizomes of Alisma orientale and alisol derivativesinhibit allergic response and experimental atopic dermatitisrdquoBiological and Pharmaceutical Bulletin vol 35 no 9 pp 1581ndash1587 2012
[10] WGu C GengW Xue et al ldquoCharacterization and function ofthe 3-hydroxy-3-methylglutaryl-CoA reductase gene in Alismaorientale (Sam) Juz and its relationship with protostane triter-pene productionrdquo Plant Physiology and Biochemistry vol 97 pp378ndash389 2015
[11] W Gu B L Xi Q N Wu et al ldquoCloning and distribution pat-tern of HMGR gene conserved fragment in Alisma orientalerdquoJournal of Chinese Medicinal Materials vol 34 no 9 pp 1335ndash1339 2011
[12] T Kobayashi S Kato-Emori K Tomita and H Ezura ldquoDetec-tion of 3-hydroxy-3-methylglutaryl-coenzymeA reductase pro-tein Cm-HMGR during fruit development in melon (Cucumismelo L)rdquo Theoretical and Applied Genetics vol 104 no 5 pp779ndash785 2002
[13] R Tommer A Rachel P C Sylvie and R Jerry ldquoImpaired regu-lation of 3-hydroxy-3-methylglutaryl-coenzyme A reductasedegradation in lovastatin-resistant cellsrdquo The Journal of Bioog-ical Chemistry vol 274 no 41 pp 29341ndash29351 1999
[14] F Xu Q-N Wu J Chen et al ldquoThe binding mechanisms ofplasma protein to active compounds in Alismaorientale rhi-zomes (Alismatis Rhizoma)rdquo Bioorganic and Medicinal Chem-istry Letters vol 24 no 17 pp 4099ndash4105 2014
[15] G Gong Y Qin W Huang et al ldquoProtective effects of dios-genin in the hyperlipidemic rat model and in human vascularendothelial cells against hydrogen peroxide-induced apoptosisrdquoChemico-Biological Interactions vol 184 no 3 pp 366ndash3752010
[16] T F Tzeng H J Lu S S Lion C J Chang and IM Liu ldquoLipid-lowering effects of zerumbone a natural cyclic sesquiterpeneof Zingiber zerumbet Smith in high-fat diet-induced hyperlipi-demic hamstersrdquoFood andChemical Toxicology vol 69 pp 132ndash139 2014
[17] Y Xie H Zhang H Liu et al ldquoHypocholesterolemic effectsof Kluyveromyces marxianus M3 isolated from Tibetan mush-rooms on diet-induced hypercholesterolemia in ratrdquo BrazilianJournal of Microbiology vol 46 no 2 pp 389ndash395 2015
[18] I Karam N Ma X-W Liu et al ldquoRegulation effect of AspirinEugenol Ester on blood lipids in Wistar rats with hyperlipi-demiardquo BMC Veterinary Research vol 11 article 217 2015
[19] G Jiao J B Wei M H Yin and P T Chun ldquoA new TCMformula FTZ lowers serum cholesterol by regulating HMG-CoA reductase and CYP7A1 in hyperlipidemic ratsrdquo Journal ofEthnopharmacology vol 135 no 2 pp 299ndash307 2011
[20] Y Geng W L Tu J J Zhang L Zhang and J Zhang ldquoEffectsof extract from cole pollen on lipidmetabolism in experimentalhyperlipidemic ratsrdquo The Scientific World Journal vol 2014Article ID 982498 6 pages 2014
[21] K Sueyoshi Y Nogawa K Sugawara T Endo and HHisamoto ldquoHighly sensitive and multiple enzyme activity assayusing reagent-release capillary-isoelectric focusing with rhod-amine 110-based substratesrdquo Analytical Sciences vol 31 no 11pp 1155ndash1161 2015
[22] D Fteita E Kononen M Gursoy E Soderling and U KGursoy ldquoDoes estradiol have an impact on the dipeptidylpeptidase IV enzyme activity of the Prevotella intermedia groupbacteriardquo Anaerobe vol 36 pp 14ndash18 2015
[23] L L Zhuo LWang and N Zhou ldquoRandomly arrayed G-quad-ruplexes for label-free and real-time assay of enzyme activityrdquoChemical Communications vol 50 no 52 pp 6875ndash6878 2014
[24] M Garelnabi and A Younis ldquoParaoxonase-1 enzyme activityassay for clinical samples validation and correlation studiesrdquoMedical Science Monitor vol 21 pp 902ndash908 2015
Evidence-Based Complementary and Alternative Medicine 11
[25] X Bi andZ Liu ldquoEnzyme activity assay of glycoprotein enzymesbased on a boronate affinity molecularly imprinted 96-wellmicroplaterdquo Analytical Chemistry vol 86 no 24 pp 12382ndash12389 2014
[26] A Walls S Dermody R Kumaran N Krishnan and E HHarley ldquoCharacterization of B-Cells in tonsils of patients diag-nosed with pediatric autoimmune neuropsychiatric disorderassociated streptococcusrdquo International Journal of PediatricOtorhinolaryngology vol 80 pp 49ndash52 2016
[27] I Karam N Ma X-W Liu et al ldquoRegulation effect of AspirinEugenol Ester on blood lipids in Wistar rats with hyperlipi-demiardquo BMC Veterinary Research vol 11 no 1 article 217 2015
[28] C Sunil S Ignacimuthu and C Kumarappan ldquoHypolipidemicactivity of Symplocos cochinchinensis S Moore leaves in hyper-lipidemic ratsrdquo Journal of Natural Medicines vol 66 no 1 pp32ndash38 2012
[29] W Shen Y Han B Huang et al ldquoMicroRNA-483-3p inhibitsextracellular matrix production by targeting smad4 in humantrabecular meshwork cellsrdquo Investigative Ophthalmology amp Vis-ual Science vol 56 no 13 pp 8419ndash8427 2015
[30] Z Wu Z Yi L Dong and A Zhang ldquoMolecular simulationstudy of the specific combination between four kinds of phthalicacid esters and human serum albuminrdquo Environmental Toxicol-ogy and Pharmacology vol 41 pp 259ndash265 2016
[31] K-J Hwang M Yoon H-J Hwang S-M Choi and S-MJeong ldquoMolecular dynamics simulation of oxygen ion conduc-tion in orthorhombic perovskite Ba-doped LaInO
3using cubic
and orthorhombic modelrdquo Journal of Nanoscience and Nan-otechnology vol 15 no 11 pp 8947ndash8950 2015
[32] H Yu W Han W Ma and K Schulten ldquoTransient 120573-hairpinformation in 120572-synuclein monomer revealed by coarse-grainedmolecular dynamics simulationrdquo The Journal of ChemicalPhysics vol 143 no 24 Article ID 243142 2015
[33] M Breuer K M Rosso and J Blumberger ldquoFlavin binding tothe deca-heme cytochrome mtrc insights from computationalmolecular simulationrdquo Biophysical Journal vol 109 no 12 pp2614ndash2624 2015
[34] V Rastija D Agic S Tomic et al ldquoSynthesis QSAR and mole-cular dynamics simulation of amidino-substituted benzimi-dazoles as dipeptidyl peptidase III inhibitorsrdquo Acta ChimicaSlovenica vol 62 no 4 pp 867ndash878 2015
[35] M Enciso C Schutte and L Delle Site ldquoInfluence of pH andsequence in peptide aggregation via molecular simulationrdquoTheJournal of Chemical Physics vol 143 no 24 Article ID 2431302015
[36] L XWangQWu andG P Peng ldquoA Study on determination ofcontent of 23-acetyl alisol B in Rhizoma Alismatis orientalitisrdquoJournal of Nanjing University of Traditional Chinese Medicine(Natural Science) vol 18 no 2 pp 105ndash107 2002
[37] G P Peng andY L Zhu ldquoProgress in the study on chemical con-stituents of Alisma orientalisrdquo Natural Product Research andDevelopment vol 18 no 2 pp 348ndash351 2006
[38] D Y Liu S J Gu J Wu et al ldquoStudies on fractions of zexiedecoction with antihyperlipidemia bioactivity in micerdquo ChinaPharmacist vol 13 no 6 pp 763ndash766 2010
[39] Q-F Luo L Sun and G-H Du ldquoProgress in new drugs tar-geting reverse cholesterol transportrdquo Chinese PharmacologicalBulletin vol 22 no 8 pp 904ndash907 2006
[40] M Yoshikawa and Y Fukuda ldquoHatakeayamardquo Chemical andPharmaceutical Bulletin vol 41 no 6 pp 1194ndash1196 1993
[41] F Xu L Zhang LHe et al ldquoMolecularmechanismon the inter-action between alisols and human serum albuminrdquo Acta Chim-ica Sinica vol 69 no 19 pp 2228ndash2234 2011
[42] F Xu J ChenW Gu et al ldquoStudy on transformation of effectivecomponents of Alisma under simulated in vivo conditionsrdquoChinese Journal of Pharmaceutical Analysis vol 31 no 11 pp2035ndash2039 2011
[43] H Yang Y G Gao and L X Zhang ldquoResearch progress on bio-synthetic pathway of terpenoids containing ginsenoside and theHMGRrdquo China Journal of Biological Engineering vol 28 no 10pp 130ndash135 2008
[44] G Z Cui Y L Ben N Li and D L Liu ldquoCatalytic mechanismand characteristics of HMG-COA recuctaserdquo Journal ofMedicalMolecular Biology vol 5 no 6 p 542 2008
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom
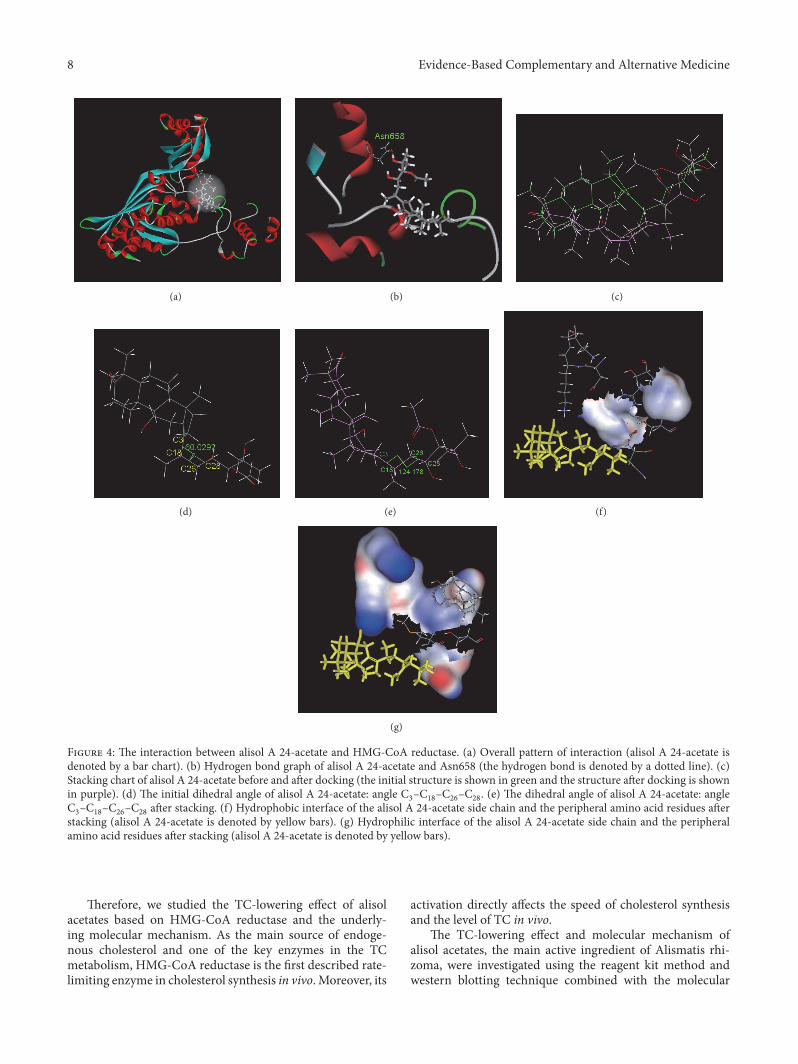
8 Evidence-Based Complementary and Alternative Medicine
(a) (b) (c)
(d) (e) (f)
(g)
Figure 4 The interaction between alisol A 24-acetate and HMG-CoA reductase (a) Overall pattern of interaction (alisol A 24-acetate isdenoted by a bar chart) (b) Hydrogen bond graph of alisol A 24-acetate and Asn658 (the hydrogen bond is denoted by a dotted line) (c)Stacking chart of alisol A 24-acetate before and after docking (the initial structure is shown in green and the structure after docking is shownin purple) (d) The initial dihedral angle of alisol A 24-acetate angle C
3ndashC18ndashC26ndashC28 (e) The dihedral angle of alisol A 24-acetate angle
C3ndashC18ndashC26ndashC28after stacking (f) Hydrophobic interface of the alisol A 24-acetate side chain and the peripheral amino acid residues after
stacking (alisol A 24-acetate is denoted by yellow bars) (g) Hydrophilic interface of the alisol A 24-acetate side chain and the peripheralamino acid residues after stacking (alisol A 24-acetate is denoted by yellow bars)
Therefore we studied the TC-lowering effect of alisolacetates based on HMG-CoA reductase and the underly-ing molecular mechanism As the main source of endoge-nous cholesterol and one of the key enzymes in the TCmetabolism HMG-CoA reductase is the first described rate-limiting enzyme in cholesterol synthesis in vivo Moreover its
activation directly affects the speed of cholesterol synthesisand the level of TC in vivo
The TC-lowering effect and molecular mechanism ofalisol acetates the main active ingredient of Alismatis rhi-zoma were investigated using the reagent kit method andwestern blotting technique combined with the molecular
Evidence-Based Complementary and Alternative Medicine 9
simulation technique An interaction model was built foralisol acetate and the key enzyme involved in the metabolismof TC was HMG-CoA reductase The present study revealsthat alisol acetates can significantly lower the TC level ofhyperlipidemic miceThe alisol B 23-acetate showed a higherextent of TC lowering than alisol A 24-acetate
Consistent with the in vitro results alisol acetates couldalso lower the in vivoHMG-CoA reductase activity in a dose-dependent fashion with a higher degree of enzyme inhibitionfor alisol B 23-acetate compared to alisol A 24-acetate Theseresults suggest that alisol acetates might lower TC levels viaregulation of HMG-CoA reductase activity The consistencybetween the results of the in vivo and in vitro experimentsindicates that the alisol acetate prototype drug inhibitsHMG-CoA reductase activity instead of the in vivometabolites
Revealed by the western blotting results alisol acetatesmight inhibit the activity of HMG-CoA reductase by directlyand competitively binding to HMG-CoA reductase ratherthan by downregulating its protein expression
The molecular simulation technique was employed inorder to investigate the interaction of alisol acetates andHMG-CoA reductase The results (Figures 3 and 4 andTable 4) revealed themolecularmechanism of the interactionof alisol acetates and HMG-CoA reductase Both alisolacetates bound to HMG-CoA reductase in their catalyticdomain at the C-terminalThe alisol B 23-acetate formed twohydrogen bonds with HMG-CoA reductase The hydrophilicinteraction was stronger than the hydrophobic interactionwhereas the electrostatic interaction was stronger than thevan der Waals forces After the interaction the structure ofthe small molecule dramatically changed from the originalfolded sheet to an open structure Additionally the side chainstretched and was inserted into the protein The peripheralamino acids formed a hydrophilic environment and stablybound with it In general a hydrogen bond with a lengthof less than 35 A and an angle of 90ndash180∘ is consideredto be strong These two hydrogen bonds were relativelystrong and might be partly responsible for the torsion ofthe small molecule The alisol A 24-acetate formed onehydrogen bond with the HMG-CoA reductase and this bondwas weaker compared to those of alisol B 23-acetate Thehydrophilic interaction was stronger than the hydrophobicinteraction and the van derWaals force was stronger than theelectrostatic interaction After the interaction the moleculeside chain had large torsion and changed from the originalstretched open state to a ring-like sheet state which wasdifficult to insert into the proteinThe binding energy of alisolB 23-acetate andHMG-CoA reductase was lower than that ofalisol A 24-acetate and HMG-CoA reductase indicating thatthe binding of alisol B 23-acetate and HMG-CoA reductasewas stable According to the pharmacological results alisolB 23-acetate lowered the HMG-CoA reductase activity morethan alisol A 24-acetate This suggests that the active centerwas the interaction pocket constituted by these amino acidsand that the hydrogen bonds involving Lys691 Asp767 andAsn658 were the key amino acid residues for the interactionof this protein with alisol acetates
The binding of alisol B 23-acetate and HMG-CoA reduc-tase was considerably stronger than the binding of alisol
A 24-acetate and HMG-CoA reductase The relatively weakbinding of alisol A 24-acetate and HMG-CoA reductasemight be due to its distinct side-chain structure The sidechain of alisol B 23-acetate contained an ether bond whichwas strongly hydrophilic and could be inserted into thehydrophilic environment of the protein to form strong inter-actions Previous studies found that the side chains of alisolacetates were their active groups [40] In the present studythis conclusionwas validated A folded side chainparent ringbound more strongly to the macromolecule than an openside-chainparent ring did Thus the side chain acts as thesteering group in that it steers the interactions between alisolacetates and macromolecules This might be the key groupfor interactions with macromolecules as it had a decisiveeffect on the interactions of this type of compound withmacromolecules Our previous studies indicated that alisol B23-acetate was partially converted to alisol A 24-acetate in theenvironment of the human stomach However the bindingof alisol A 24-acetate and HMG-CoA reductase was weaker[14 41 42] Therefore it is advisable for drug developers todevelop a dosage form that circumvents the conversion ofalisol B 23-acetate in the stomach
Previously published studies indicate that [43 44] themechanism of reducing the level of TC is that two acetylCoA molecules condense to acetyl CoA under the catalysisof acetoacetyl-coenzyme A thiolase (AACT) acetyl CoAand acetyl CoA condense to HMG-CoA and HMG-CoAreduces to MVA under the catalytic action of HMG-CoAreductase MVA further generates TC with squalene Thehigh intracellular free fatty acid (FC) level stimulates micro-somal acyl coenzyme A-cholesterol acyltransferase (ACAT)and promotes esterification of the cholesterol Subsequentlycholesterol is stored in the form of cholesterol esters (CE)which is a feedback inhibition of HMG-CoA reductasesynthesis so that cholesterol synthesis rate-limiting enzymeswill be inhibited thus reducing intracellular cholesterolsynthesis This process features three important enzymesAACT HMG-CoA reductase and ACAT Whether AACTand ACAT are involved in the effect of alisol acetates will bethe subject of intensive research in the future
The results of the study indicate that alisol acetates cannot only decrease TC level but also decrease TG and LDL-C levels and increase HDL-C level Furthermore a follow-upstudy is designed to investigate the regulatory mechanisms ofalisol acetates on TG HDL-C and LDL-C
5 Conclusion
The present study reveals that alisol acetates could signifi-cantly lower the TC level of hyperlipidemic mice The effectof alisol B 23-acetate on HMG-CoA reductase was strongerthan that of alisol A 24-acetate Alisol acetates could lowerHMG-CoA reductase activity in a dose-dependent fashionboth in vivo and in vitro Neither of these alisol acetatescould significantly lower the protein expression of HMG-CoA This suggests that alisol acetates lower the TC levelvia inhibiting the activity of HMG-CoA reductase via itsprototype drug which may have an inhibitory effect bydirectly and competitively binding with HMG-CoA In the
10 Evidence-Based Complementary and Alternative Medicine
binding interaction of alisol acetate andHMG-CoA reductasevia molecular simulation the key amino acid residues ofHMG-CoA reductase might be Lys691 Asp767 and Asn658The side chain of the alisol acetatewas the steering group Fur-thermore a folded side chainparent ring boundmoreweaklyto HMG-CoA reductase than an open side chainparent ringThis study explored the TC-lowering mechanism of alisolacetates at the molecular level and presented guidance for itsclinical use
Competing Interests
The authors declare that they have no competing interests
Acknowledgments
This work was financially supported by the National NatureScience Foundation of China (81303173 81673534) the Nat-ural Science Foundation of Jiangsu Province (BK20161576)the Natural Science Fund for Colleges and Universities inJiangsu Province (14KJB360001) and the Top-Notch Aca-demic Programs Project of Jiangsu Higher Education Insti-tutions (PPZY2015A070)
References
[1] L X Wang Q N Wu Q Zhang G P Peng and A W DingldquoBasic study of diuretic active compounds in Rhizoma Alis-matisrdquo West China Journal of Pharmaceutical Sciences vol 23no 6 pp 670ndash672 2008
[2] S S Wu Pharmaceutical and Clinical Research of Alisma Ori-entalis Chinese Press Beijing China 2007
[3] M Miyazawa S Yoshinaga and Y Kashima ldquoChemical com-position and characteristic odor compounds in essential oilfrom Alismatis Rhizoma (Tubers of Alisma orientale)rdquo DiatomResearch vol 23 no 1 pp 133ndash141 2016
[4] M-K Jang Y-R Han J S Nam et al ldquoProtective effects ofAlisma orientale extract against hepatic steatosis via inhibitionof endoplasmic reticulum stressrdquo International Journal ofMolec-ular Sciences vol 16 no 11 pp 26151ndash26165 2015
[5] Z-P Mai K Zhou G-B Ge et al ldquoProtostane triterpenoidsfrom the rhizome of Alisma orientale exhibit inhibitory effectson human carboxylesterase 2rdquo Journal of Natural Products vol78 no 10 pp 2372ndash2380 2015
[6] T Tian H Chen andY-Y Zhao ldquoTraditional uses phytochem-istry pharmacology toxicology and quality control of Alismaorientale (Sam) Juzep a reviewrdquo Journal of Ethnopharmacologyvol 158 pp 373ndash387 2014
[7] KHKimM J Kwun J-Y Choi et al ldquoTherapeutic effect of thetuber of Alisma orientale on lipopolysaccharide-induced acutelung injuryrdquo Evidence-Based Complementary and AlternativeMedicine vol 2013 Article ID 863892 10 pages 2013
[8] H-G Jin Q Jin A R Kim et al ldquoA new triterpenoid fromAlisma orientale and their antibacterial effectrdquoArchives of Phar-macal Research vol 35 no 11 pp 1919ndash1926 2012
[9] J H Lee O S Kwon H-G Jin E-R Woo Y S Kim and H PKim ldquoThe rhizomes of Alisma orientale and alisol derivativesinhibit allergic response and experimental atopic dermatitisrdquoBiological and Pharmaceutical Bulletin vol 35 no 9 pp 1581ndash1587 2012
[10] WGu C GengW Xue et al ldquoCharacterization and function ofthe 3-hydroxy-3-methylglutaryl-CoA reductase gene in Alismaorientale (Sam) Juz and its relationship with protostane triter-pene productionrdquo Plant Physiology and Biochemistry vol 97 pp378ndash389 2015
[11] W Gu B L Xi Q N Wu et al ldquoCloning and distribution pat-tern of HMGR gene conserved fragment in Alisma orientalerdquoJournal of Chinese Medicinal Materials vol 34 no 9 pp 1335ndash1339 2011
[12] T Kobayashi S Kato-Emori K Tomita and H Ezura ldquoDetec-tion of 3-hydroxy-3-methylglutaryl-coenzymeA reductase pro-tein Cm-HMGR during fruit development in melon (Cucumismelo L)rdquo Theoretical and Applied Genetics vol 104 no 5 pp779ndash785 2002
[13] R Tommer A Rachel P C Sylvie and R Jerry ldquoImpaired regu-lation of 3-hydroxy-3-methylglutaryl-coenzyme A reductasedegradation in lovastatin-resistant cellsrdquo The Journal of Bioog-ical Chemistry vol 274 no 41 pp 29341ndash29351 1999
[14] F Xu Q-N Wu J Chen et al ldquoThe binding mechanisms ofplasma protein to active compounds in Alismaorientale rhi-zomes (Alismatis Rhizoma)rdquo Bioorganic and Medicinal Chem-istry Letters vol 24 no 17 pp 4099ndash4105 2014
[15] G Gong Y Qin W Huang et al ldquoProtective effects of dios-genin in the hyperlipidemic rat model and in human vascularendothelial cells against hydrogen peroxide-induced apoptosisrdquoChemico-Biological Interactions vol 184 no 3 pp 366ndash3752010
[16] T F Tzeng H J Lu S S Lion C J Chang and IM Liu ldquoLipid-lowering effects of zerumbone a natural cyclic sesquiterpeneof Zingiber zerumbet Smith in high-fat diet-induced hyperlipi-demic hamstersrdquoFood andChemical Toxicology vol 69 pp 132ndash139 2014
[17] Y Xie H Zhang H Liu et al ldquoHypocholesterolemic effectsof Kluyveromyces marxianus M3 isolated from Tibetan mush-rooms on diet-induced hypercholesterolemia in ratrdquo BrazilianJournal of Microbiology vol 46 no 2 pp 389ndash395 2015
[18] I Karam N Ma X-W Liu et al ldquoRegulation effect of AspirinEugenol Ester on blood lipids in Wistar rats with hyperlipi-demiardquo BMC Veterinary Research vol 11 article 217 2015
[19] G Jiao J B Wei M H Yin and P T Chun ldquoA new TCMformula FTZ lowers serum cholesterol by regulating HMG-CoA reductase and CYP7A1 in hyperlipidemic ratsrdquo Journal ofEthnopharmacology vol 135 no 2 pp 299ndash307 2011
[20] Y Geng W L Tu J J Zhang L Zhang and J Zhang ldquoEffectsof extract from cole pollen on lipidmetabolism in experimentalhyperlipidemic ratsrdquo The Scientific World Journal vol 2014Article ID 982498 6 pages 2014
[21] K Sueyoshi Y Nogawa K Sugawara T Endo and HHisamoto ldquoHighly sensitive and multiple enzyme activity assayusing reagent-release capillary-isoelectric focusing with rhod-amine 110-based substratesrdquo Analytical Sciences vol 31 no 11pp 1155ndash1161 2015
[22] D Fteita E Kononen M Gursoy E Soderling and U KGursoy ldquoDoes estradiol have an impact on the dipeptidylpeptidase IV enzyme activity of the Prevotella intermedia groupbacteriardquo Anaerobe vol 36 pp 14ndash18 2015
[23] L L Zhuo LWang and N Zhou ldquoRandomly arrayed G-quad-ruplexes for label-free and real-time assay of enzyme activityrdquoChemical Communications vol 50 no 52 pp 6875ndash6878 2014
[24] M Garelnabi and A Younis ldquoParaoxonase-1 enzyme activityassay for clinical samples validation and correlation studiesrdquoMedical Science Monitor vol 21 pp 902ndash908 2015
Evidence-Based Complementary and Alternative Medicine 11
[25] X Bi andZ Liu ldquoEnzyme activity assay of glycoprotein enzymesbased on a boronate affinity molecularly imprinted 96-wellmicroplaterdquo Analytical Chemistry vol 86 no 24 pp 12382ndash12389 2014
[26] A Walls S Dermody R Kumaran N Krishnan and E HHarley ldquoCharacterization of B-Cells in tonsils of patients diag-nosed with pediatric autoimmune neuropsychiatric disorderassociated streptococcusrdquo International Journal of PediatricOtorhinolaryngology vol 80 pp 49ndash52 2016
[27] I Karam N Ma X-W Liu et al ldquoRegulation effect of AspirinEugenol Ester on blood lipids in Wistar rats with hyperlipi-demiardquo BMC Veterinary Research vol 11 no 1 article 217 2015
[28] C Sunil S Ignacimuthu and C Kumarappan ldquoHypolipidemicactivity of Symplocos cochinchinensis S Moore leaves in hyper-lipidemic ratsrdquo Journal of Natural Medicines vol 66 no 1 pp32ndash38 2012
[29] W Shen Y Han B Huang et al ldquoMicroRNA-483-3p inhibitsextracellular matrix production by targeting smad4 in humantrabecular meshwork cellsrdquo Investigative Ophthalmology amp Vis-ual Science vol 56 no 13 pp 8419ndash8427 2015
[30] Z Wu Z Yi L Dong and A Zhang ldquoMolecular simulationstudy of the specific combination between four kinds of phthalicacid esters and human serum albuminrdquo Environmental Toxicol-ogy and Pharmacology vol 41 pp 259ndash265 2016
[31] K-J Hwang M Yoon H-J Hwang S-M Choi and S-MJeong ldquoMolecular dynamics simulation of oxygen ion conduc-tion in orthorhombic perovskite Ba-doped LaInO
3using cubic
and orthorhombic modelrdquo Journal of Nanoscience and Nan-otechnology vol 15 no 11 pp 8947ndash8950 2015
[32] H Yu W Han W Ma and K Schulten ldquoTransient 120573-hairpinformation in 120572-synuclein monomer revealed by coarse-grainedmolecular dynamics simulationrdquo The Journal of ChemicalPhysics vol 143 no 24 Article ID 243142 2015
[33] M Breuer K M Rosso and J Blumberger ldquoFlavin binding tothe deca-heme cytochrome mtrc insights from computationalmolecular simulationrdquo Biophysical Journal vol 109 no 12 pp2614ndash2624 2015
[34] V Rastija D Agic S Tomic et al ldquoSynthesis QSAR and mole-cular dynamics simulation of amidino-substituted benzimi-dazoles as dipeptidyl peptidase III inhibitorsrdquo Acta ChimicaSlovenica vol 62 no 4 pp 867ndash878 2015
[35] M Enciso C Schutte and L Delle Site ldquoInfluence of pH andsequence in peptide aggregation via molecular simulationrdquoTheJournal of Chemical Physics vol 143 no 24 Article ID 2431302015
[36] L XWangQWu andG P Peng ldquoA Study on determination ofcontent of 23-acetyl alisol B in Rhizoma Alismatis orientalitisrdquoJournal of Nanjing University of Traditional Chinese Medicine(Natural Science) vol 18 no 2 pp 105ndash107 2002
[37] G P Peng andY L Zhu ldquoProgress in the study on chemical con-stituents of Alisma orientalisrdquo Natural Product Research andDevelopment vol 18 no 2 pp 348ndash351 2006
[38] D Y Liu S J Gu J Wu et al ldquoStudies on fractions of zexiedecoction with antihyperlipidemia bioactivity in micerdquo ChinaPharmacist vol 13 no 6 pp 763ndash766 2010
[39] Q-F Luo L Sun and G-H Du ldquoProgress in new drugs tar-geting reverse cholesterol transportrdquo Chinese PharmacologicalBulletin vol 22 no 8 pp 904ndash907 2006
[40] M Yoshikawa and Y Fukuda ldquoHatakeayamardquo Chemical andPharmaceutical Bulletin vol 41 no 6 pp 1194ndash1196 1993
[41] F Xu L Zhang LHe et al ldquoMolecularmechanismon the inter-action between alisols and human serum albuminrdquo Acta Chim-ica Sinica vol 69 no 19 pp 2228ndash2234 2011
[42] F Xu J ChenW Gu et al ldquoStudy on transformation of effectivecomponents of Alisma under simulated in vivo conditionsrdquoChinese Journal of Pharmaceutical Analysis vol 31 no 11 pp2035ndash2039 2011
[43] H Yang Y G Gao and L X Zhang ldquoResearch progress on bio-synthetic pathway of terpenoids containing ginsenoside and theHMGRrdquo China Journal of Biological Engineering vol 28 no 10pp 130ndash135 2008
[44] G Z Cui Y L Ben N Li and D L Liu ldquoCatalytic mechanismand characteristics of HMG-COA recuctaserdquo Journal ofMedicalMolecular Biology vol 5 no 6 p 542 2008
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom

Evidence-Based Complementary and Alternative Medicine 9
simulation technique An interaction model was built foralisol acetate and the key enzyme involved in the metabolismof TC was HMG-CoA reductase The present study revealsthat alisol acetates can significantly lower the TC level ofhyperlipidemic miceThe alisol B 23-acetate showed a higherextent of TC lowering than alisol A 24-acetate
Consistent with the in vitro results alisol acetates couldalso lower the in vivoHMG-CoA reductase activity in a dose-dependent fashion with a higher degree of enzyme inhibitionfor alisol B 23-acetate compared to alisol A 24-acetate Theseresults suggest that alisol acetates might lower TC levels viaregulation of HMG-CoA reductase activity The consistencybetween the results of the in vivo and in vitro experimentsindicates that the alisol acetate prototype drug inhibitsHMG-CoA reductase activity instead of the in vivometabolites
Revealed by the western blotting results alisol acetatesmight inhibit the activity of HMG-CoA reductase by directlyand competitively binding to HMG-CoA reductase ratherthan by downregulating its protein expression
The molecular simulation technique was employed inorder to investigate the interaction of alisol acetates andHMG-CoA reductase The results (Figures 3 and 4 andTable 4) revealed themolecularmechanism of the interactionof alisol acetates and HMG-CoA reductase Both alisolacetates bound to HMG-CoA reductase in their catalyticdomain at the C-terminalThe alisol B 23-acetate formed twohydrogen bonds with HMG-CoA reductase The hydrophilicinteraction was stronger than the hydrophobic interactionwhereas the electrostatic interaction was stronger than thevan der Waals forces After the interaction the structure ofthe small molecule dramatically changed from the originalfolded sheet to an open structure Additionally the side chainstretched and was inserted into the protein The peripheralamino acids formed a hydrophilic environment and stablybound with it In general a hydrogen bond with a lengthof less than 35 A and an angle of 90ndash180∘ is consideredto be strong These two hydrogen bonds were relativelystrong and might be partly responsible for the torsion ofthe small molecule The alisol A 24-acetate formed onehydrogen bond with the HMG-CoA reductase and this bondwas weaker compared to those of alisol B 23-acetate Thehydrophilic interaction was stronger than the hydrophobicinteraction and the van derWaals force was stronger than theelectrostatic interaction After the interaction the moleculeside chain had large torsion and changed from the originalstretched open state to a ring-like sheet state which wasdifficult to insert into the proteinThe binding energy of alisolB 23-acetate andHMG-CoA reductase was lower than that ofalisol A 24-acetate and HMG-CoA reductase indicating thatthe binding of alisol B 23-acetate and HMG-CoA reductasewas stable According to the pharmacological results alisolB 23-acetate lowered the HMG-CoA reductase activity morethan alisol A 24-acetate This suggests that the active centerwas the interaction pocket constituted by these amino acidsand that the hydrogen bonds involving Lys691 Asp767 andAsn658 were the key amino acid residues for the interactionof this protein with alisol acetates
The binding of alisol B 23-acetate and HMG-CoA reduc-tase was considerably stronger than the binding of alisol
A 24-acetate and HMG-CoA reductase The relatively weakbinding of alisol A 24-acetate and HMG-CoA reductasemight be due to its distinct side-chain structure The sidechain of alisol B 23-acetate contained an ether bond whichwas strongly hydrophilic and could be inserted into thehydrophilic environment of the protein to form strong inter-actions Previous studies found that the side chains of alisolacetates were their active groups [40] In the present studythis conclusionwas validated A folded side chainparent ringbound more strongly to the macromolecule than an openside-chainparent ring did Thus the side chain acts as thesteering group in that it steers the interactions between alisolacetates and macromolecules This might be the key groupfor interactions with macromolecules as it had a decisiveeffect on the interactions of this type of compound withmacromolecules Our previous studies indicated that alisol B23-acetate was partially converted to alisol A 24-acetate in theenvironment of the human stomach However the bindingof alisol A 24-acetate and HMG-CoA reductase was weaker[14 41 42] Therefore it is advisable for drug developers todevelop a dosage form that circumvents the conversion ofalisol B 23-acetate in the stomach
Previously published studies indicate that [43 44] themechanism of reducing the level of TC is that two acetylCoA molecules condense to acetyl CoA under the catalysisof acetoacetyl-coenzyme A thiolase (AACT) acetyl CoAand acetyl CoA condense to HMG-CoA and HMG-CoAreduces to MVA under the catalytic action of HMG-CoAreductase MVA further generates TC with squalene Thehigh intracellular free fatty acid (FC) level stimulates micro-somal acyl coenzyme A-cholesterol acyltransferase (ACAT)and promotes esterification of the cholesterol Subsequentlycholesterol is stored in the form of cholesterol esters (CE)which is a feedback inhibition of HMG-CoA reductasesynthesis so that cholesterol synthesis rate-limiting enzymeswill be inhibited thus reducing intracellular cholesterolsynthesis This process features three important enzymesAACT HMG-CoA reductase and ACAT Whether AACTand ACAT are involved in the effect of alisol acetates will bethe subject of intensive research in the future
The results of the study indicate that alisol acetates cannot only decrease TC level but also decrease TG and LDL-C levels and increase HDL-C level Furthermore a follow-upstudy is designed to investigate the regulatory mechanisms ofalisol acetates on TG HDL-C and LDL-C
5 Conclusion
The present study reveals that alisol acetates could signifi-cantly lower the TC level of hyperlipidemic mice The effectof alisol B 23-acetate on HMG-CoA reductase was strongerthan that of alisol A 24-acetate Alisol acetates could lowerHMG-CoA reductase activity in a dose-dependent fashionboth in vivo and in vitro Neither of these alisol acetatescould significantly lower the protein expression of HMG-CoA This suggests that alisol acetates lower the TC levelvia inhibiting the activity of HMG-CoA reductase via itsprototype drug which may have an inhibitory effect bydirectly and competitively binding with HMG-CoA In the
10 Evidence-Based Complementary and Alternative Medicine
binding interaction of alisol acetate andHMG-CoA reductasevia molecular simulation the key amino acid residues ofHMG-CoA reductase might be Lys691 Asp767 and Asn658The side chain of the alisol acetatewas the steering group Fur-thermore a folded side chainparent ring boundmoreweaklyto HMG-CoA reductase than an open side chainparent ringThis study explored the TC-lowering mechanism of alisolacetates at the molecular level and presented guidance for itsclinical use
Competing Interests
The authors declare that they have no competing interests
Acknowledgments
This work was financially supported by the National NatureScience Foundation of China (81303173 81673534) the Nat-ural Science Foundation of Jiangsu Province (BK20161576)the Natural Science Fund for Colleges and Universities inJiangsu Province (14KJB360001) and the Top-Notch Aca-demic Programs Project of Jiangsu Higher Education Insti-tutions (PPZY2015A070)
References
[1] L X Wang Q N Wu Q Zhang G P Peng and A W DingldquoBasic study of diuretic active compounds in Rhizoma Alis-matisrdquo West China Journal of Pharmaceutical Sciences vol 23no 6 pp 670ndash672 2008
[2] S S Wu Pharmaceutical and Clinical Research of Alisma Ori-entalis Chinese Press Beijing China 2007
[3] M Miyazawa S Yoshinaga and Y Kashima ldquoChemical com-position and characteristic odor compounds in essential oilfrom Alismatis Rhizoma (Tubers of Alisma orientale)rdquo DiatomResearch vol 23 no 1 pp 133ndash141 2016
[4] M-K Jang Y-R Han J S Nam et al ldquoProtective effects ofAlisma orientale extract against hepatic steatosis via inhibitionof endoplasmic reticulum stressrdquo International Journal ofMolec-ular Sciences vol 16 no 11 pp 26151ndash26165 2015
[5] Z-P Mai K Zhou G-B Ge et al ldquoProtostane triterpenoidsfrom the rhizome of Alisma orientale exhibit inhibitory effectson human carboxylesterase 2rdquo Journal of Natural Products vol78 no 10 pp 2372ndash2380 2015
[6] T Tian H Chen andY-Y Zhao ldquoTraditional uses phytochem-istry pharmacology toxicology and quality control of Alismaorientale (Sam) Juzep a reviewrdquo Journal of Ethnopharmacologyvol 158 pp 373ndash387 2014
[7] KHKimM J Kwun J-Y Choi et al ldquoTherapeutic effect of thetuber of Alisma orientale on lipopolysaccharide-induced acutelung injuryrdquo Evidence-Based Complementary and AlternativeMedicine vol 2013 Article ID 863892 10 pages 2013
[8] H-G Jin Q Jin A R Kim et al ldquoA new triterpenoid fromAlisma orientale and their antibacterial effectrdquoArchives of Phar-macal Research vol 35 no 11 pp 1919ndash1926 2012
[9] J H Lee O S Kwon H-G Jin E-R Woo Y S Kim and H PKim ldquoThe rhizomes of Alisma orientale and alisol derivativesinhibit allergic response and experimental atopic dermatitisrdquoBiological and Pharmaceutical Bulletin vol 35 no 9 pp 1581ndash1587 2012
[10] WGu C GengW Xue et al ldquoCharacterization and function ofthe 3-hydroxy-3-methylglutaryl-CoA reductase gene in Alismaorientale (Sam) Juz and its relationship with protostane triter-pene productionrdquo Plant Physiology and Biochemistry vol 97 pp378ndash389 2015
[11] W Gu B L Xi Q N Wu et al ldquoCloning and distribution pat-tern of HMGR gene conserved fragment in Alisma orientalerdquoJournal of Chinese Medicinal Materials vol 34 no 9 pp 1335ndash1339 2011
[12] T Kobayashi S Kato-Emori K Tomita and H Ezura ldquoDetec-tion of 3-hydroxy-3-methylglutaryl-coenzymeA reductase pro-tein Cm-HMGR during fruit development in melon (Cucumismelo L)rdquo Theoretical and Applied Genetics vol 104 no 5 pp779ndash785 2002
[13] R Tommer A Rachel P C Sylvie and R Jerry ldquoImpaired regu-lation of 3-hydroxy-3-methylglutaryl-coenzyme A reductasedegradation in lovastatin-resistant cellsrdquo The Journal of Bioog-ical Chemistry vol 274 no 41 pp 29341ndash29351 1999
[14] F Xu Q-N Wu J Chen et al ldquoThe binding mechanisms ofplasma protein to active compounds in Alismaorientale rhi-zomes (Alismatis Rhizoma)rdquo Bioorganic and Medicinal Chem-istry Letters vol 24 no 17 pp 4099ndash4105 2014
[15] G Gong Y Qin W Huang et al ldquoProtective effects of dios-genin in the hyperlipidemic rat model and in human vascularendothelial cells against hydrogen peroxide-induced apoptosisrdquoChemico-Biological Interactions vol 184 no 3 pp 366ndash3752010
[16] T F Tzeng H J Lu S S Lion C J Chang and IM Liu ldquoLipid-lowering effects of zerumbone a natural cyclic sesquiterpeneof Zingiber zerumbet Smith in high-fat diet-induced hyperlipi-demic hamstersrdquoFood andChemical Toxicology vol 69 pp 132ndash139 2014
[17] Y Xie H Zhang H Liu et al ldquoHypocholesterolemic effectsof Kluyveromyces marxianus M3 isolated from Tibetan mush-rooms on diet-induced hypercholesterolemia in ratrdquo BrazilianJournal of Microbiology vol 46 no 2 pp 389ndash395 2015
[18] I Karam N Ma X-W Liu et al ldquoRegulation effect of AspirinEugenol Ester on blood lipids in Wistar rats with hyperlipi-demiardquo BMC Veterinary Research vol 11 article 217 2015
[19] G Jiao J B Wei M H Yin and P T Chun ldquoA new TCMformula FTZ lowers serum cholesterol by regulating HMG-CoA reductase and CYP7A1 in hyperlipidemic ratsrdquo Journal ofEthnopharmacology vol 135 no 2 pp 299ndash307 2011
[20] Y Geng W L Tu J J Zhang L Zhang and J Zhang ldquoEffectsof extract from cole pollen on lipidmetabolism in experimentalhyperlipidemic ratsrdquo The Scientific World Journal vol 2014Article ID 982498 6 pages 2014
[21] K Sueyoshi Y Nogawa K Sugawara T Endo and HHisamoto ldquoHighly sensitive and multiple enzyme activity assayusing reagent-release capillary-isoelectric focusing with rhod-amine 110-based substratesrdquo Analytical Sciences vol 31 no 11pp 1155ndash1161 2015
[22] D Fteita E Kononen M Gursoy E Soderling and U KGursoy ldquoDoes estradiol have an impact on the dipeptidylpeptidase IV enzyme activity of the Prevotella intermedia groupbacteriardquo Anaerobe vol 36 pp 14ndash18 2015
[23] L L Zhuo LWang and N Zhou ldquoRandomly arrayed G-quad-ruplexes for label-free and real-time assay of enzyme activityrdquoChemical Communications vol 50 no 52 pp 6875ndash6878 2014
[24] M Garelnabi and A Younis ldquoParaoxonase-1 enzyme activityassay for clinical samples validation and correlation studiesrdquoMedical Science Monitor vol 21 pp 902ndash908 2015
Evidence-Based Complementary and Alternative Medicine 11
[25] X Bi andZ Liu ldquoEnzyme activity assay of glycoprotein enzymesbased on a boronate affinity molecularly imprinted 96-wellmicroplaterdquo Analytical Chemistry vol 86 no 24 pp 12382ndash12389 2014
[26] A Walls S Dermody R Kumaran N Krishnan and E HHarley ldquoCharacterization of B-Cells in tonsils of patients diag-nosed with pediatric autoimmune neuropsychiatric disorderassociated streptococcusrdquo International Journal of PediatricOtorhinolaryngology vol 80 pp 49ndash52 2016
[27] I Karam N Ma X-W Liu et al ldquoRegulation effect of AspirinEugenol Ester on blood lipids in Wistar rats with hyperlipi-demiardquo BMC Veterinary Research vol 11 no 1 article 217 2015
[28] C Sunil S Ignacimuthu and C Kumarappan ldquoHypolipidemicactivity of Symplocos cochinchinensis S Moore leaves in hyper-lipidemic ratsrdquo Journal of Natural Medicines vol 66 no 1 pp32ndash38 2012
[29] W Shen Y Han B Huang et al ldquoMicroRNA-483-3p inhibitsextracellular matrix production by targeting smad4 in humantrabecular meshwork cellsrdquo Investigative Ophthalmology amp Vis-ual Science vol 56 no 13 pp 8419ndash8427 2015
[30] Z Wu Z Yi L Dong and A Zhang ldquoMolecular simulationstudy of the specific combination between four kinds of phthalicacid esters and human serum albuminrdquo Environmental Toxicol-ogy and Pharmacology vol 41 pp 259ndash265 2016
[31] K-J Hwang M Yoon H-J Hwang S-M Choi and S-MJeong ldquoMolecular dynamics simulation of oxygen ion conduc-tion in orthorhombic perovskite Ba-doped LaInO
3using cubic
and orthorhombic modelrdquo Journal of Nanoscience and Nan-otechnology vol 15 no 11 pp 8947ndash8950 2015
[32] H Yu W Han W Ma and K Schulten ldquoTransient 120573-hairpinformation in 120572-synuclein monomer revealed by coarse-grainedmolecular dynamics simulationrdquo The Journal of ChemicalPhysics vol 143 no 24 Article ID 243142 2015
[33] M Breuer K M Rosso and J Blumberger ldquoFlavin binding tothe deca-heme cytochrome mtrc insights from computationalmolecular simulationrdquo Biophysical Journal vol 109 no 12 pp2614ndash2624 2015
[34] V Rastija D Agic S Tomic et al ldquoSynthesis QSAR and mole-cular dynamics simulation of amidino-substituted benzimi-dazoles as dipeptidyl peptidase III inhibitorsrdquo Acta ChimicaSlovenica vol 62 no 4 pp 867ndash878 2015
[35] M Enciso C Schutte and L Delle Site ldquoInfluence of pH andsequence in peptide aggregation via molecular simulationrdquoTheJournal of Chemical Physics vol 143 no 24 Article ID 2431302015
[36] L XWangQWu andG P Peng ldquoA Study on determination ofcontent of 23-acetyl alisol B in Rhizoma Alismatis orientalitisrdquoJournal of Nanjing University of Traditional Chinese Medicine(Natural Science) vol 18 no 2 pp 105ndash107 2002
[37] G P Peng andY L Zhu ldquoProgress in the study on chemical con-stituents of Alisma orientalisrdquo Natural Product Research andDevelopment vol 18 no 2 pp 348ndash351 2006
[38] D Y Liu S J Gu J Wu et al ldquoStudies on fractions of zexiedecoction with antihyperlipidemia bioactivity in micerdquo ChinaPharmacist vol 13 no 6 pp 763ndash766 2010
[39] Q-F Luo L Sun and G-H Du ldquoProgress in new drugs tar-geting reverse cholesterol transportrdquo Chinese PharmacologicalBulletin vol 22 no 8 pp 904ndash907 2006
[40] M Yoshikawa and Y Fukuda ldquoHatakeayamardquo Chemical andPharmaceutical Bulletin vol 41 no 6 pp 1194ndash1196 1993
[41] F Xu L Zhang LHe et al ldquoMolecularmechanismon the inter-action between alisols and human serum albuminrdquo Acta Chim-ica Sinica vol 69 no 19 pp 2228ndash2234 2011
[42] F Xu J ChenW Gu et al ldquoStudy on transformation of effectivecomponents of Alisma under simulated in vivo conditionsrdquoChinese Journal of Pharmaceutical Analysis vol 31 no 11 pp2035ndash2039 2011
[43] H Yang Y G Gao and L X Zhang ldquoResearch progress on bio-synthetic pathway of terpenoids containing ginsenoside and theHMGRrdquo China Journal of Biological Engineering vol 28 no 10pp 130ndash135 2008
[44] G Z Cui Y L Ben N Li and D L Liu ldquoCatalytic mechanismand characteristics of HMG-COA recuctaserdquo Journal ofMedicalMolecular Biology vol 5 no 6 p 542 2008
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom

10 Evidence-Based Complementary and Alternative Medicine
binding interaction of alisol acetate andHMG-CoA reductasevia molecular simulation the key amino acid residues ofHMG-CoA reductase might be Lys691 Asp767 and Asn658The side chain of the alisol acetatewas the steering group Fur-thermore a folded side chainparent ring boundmoreweaklyto HMG-CoA reductase than an open side chainparent ringThis study explored the TC-lowering mechanism of alisolacetates at the molecular level and presented guidance for itsclinical use
Competing Interests
The authors declare that they have no competing interests
Acknowledgments
This work was financially supported by the National NatureScience Foundation of China (81303173 81673534) the Nat-ural Science Foundation of Jiangsu Province (BK20161576)the Natural Science Fund for Colleges and Universities inJiangsu Province (14KJB360001) and the Top-Notch Aca-demic Programs Project of Jiangsu Higher Education Insti-tutions (PPZY2015A070)
References
[1] L X Wang Q N Wu Q Zhang G P Peng and A W DingldquoBasic study of diuretic active compounds in Rhizoma Alis-matisrdquo West China Journal of Pharmaceutical Sciences vol 23no 6 pp 670ndash672 2008
[2] S S Wu Pharmaceutical and Clinical Research of Alisma Ori-entalis Chinese Press Beijing China 2007
[3] M Miyazawa S Yoshinaga and Y Kashima ldquoChemical com-position and characteristic odor compounds in essential oilfrom Alismatis Rhizoma (Tubers of Alisma orientale)rdquo DiatomResearch vol 23 no 1 pp 133ndash141 2016
[4] M-K Jang Y-R Han J S Nam et al ldquoProtective effects ofAlisma orientale extract against hepatic steatosis via inhibitionof endoplasmic reticulum stressrdquo International Journal ofMolec-ular Sciences vol 16 no 11 pp 26151ndash26165 2015
[5] Z-P Mai K Zhou G-B Ge et al ldquoProtostane triterpenoidsfrom the rhizome of Alisma orientale exhibit inhibitory effectson human carboxylesterase 2rdquo Journal of Natural Products vol78 no 10 pp 2372ndash2380 2015
[6] T Tian H Chen andY-Y Zhao ldquoTraditional uses phytochem-istry pharmacology toxicology and quality control of Alismaorientale (Sam) Juzep a reviewrdquo Journal of Ethnopharmacologyvol 158 pp 373ndash387 2014
[7] KHKimM J Kwun J-Y Choi et al ldquoTherapeutic effect of thetuber of Alisma orientale on lipopolysaccharide-induced acutelung injuryrdquo Evidence-Based Complementary and AlternativeMedicine vol 2013 Article ID 863892 10 pages 2013
[8] H-G Jin Q Jin A R Kim et al ldquoA new triterpenoid fromAlisma orientale and their antibacterial effectrdquoArchives of Phar-macal Research vol 35 no 11 pp 1919ndash1926 2012
[9] J H Lee O S Kwon H-G Jin E-R Woo Y S Kim and H PKim ldquoThe rhizomes of Alisma orientale and alisol derivativesinhibit allergic response and experimental atopic dermatitisrdquoBiological and Pharmaceutical Bulletin vol 35 no 9 pp 1581ndash1587 2012
[10] WGu C GengW Xue et al ldquoCharacterization and function ofthe 3-hydroxy-3-methylglutaryl-CoA reductase gene in Alismaorientale (Sam) Juz and its relationship with protostane triter-pene productionrdquo Plant Physiology and Biochemistry vol 97 pp378ndash389 2015
[11] W Gu B L Xi Q N Wu et al ldquoCloning and distribution pat-tern of HMGR gene conserved fragment in Alisma orientalerdquoJournal of Chinese Medicinal Materials vol 34 no 9 pp 1335ndash1339 2011
[12] T Kobayashi S Kato-Emori K Tomita and H Ezura ldquoDetec-tion of 3-hydroxy-3-methylglutaryl-coenzymeA reductase pro-tein Cm-HMGR during fruit development in melon (Cucumismelo L)rdquo Theoretical and Applied Genetics vol 104 no 5 pp779ndash785 2002
[13] R Tommer A Rachel P C Sylvie and R Jerry ldquoImpaired regu-lation of 3-hydroxy-3-methylglutaryl-coenzyme A reductasedegradation in lovastatin-resistant cellsrdquo The Journal of Bioog-ical Chemistry vol 274 no 41 pp 29341ndash29351 1999
[14] F Xu Q-N Wu J Chen et al ldquoThe binding mechanisms ofplasma protein to active compounds in Alismaorientale rhi-zomes (Alismatis Rhizoma)rdquo Bioorganic and Medicinal Chem-istry Letters vol 24 no 17 pp 4099ndash4105 2014
[15] G Gong Y Qin W Huang et al ldquoProtective effects of dios-genin in the hyperlipidemic rat model and in human vascularendothelial cells against hydrogen peroxide-induced apoptosisrdquoChemico-Biological Interactions vol 184 no 3 pp 366ndash3752010
[16] T F Tzeng H J Lu S S Lion C J Chang and IM Liu ldquoLipid-lowering effects of zerumbone a natural cyclic sesquiterpeneof Zingiber zerumbet Smith in high-fat diet-induced hyperlipi-demic hamstersrdquoFood andChemical Toxicology vol 69 pp 132ndash139 2014
[17] Y Xie H Zhang H Liu et al ldquoHypocholesterolemic effectsof Kluyveromyces marxianus M3 isolated from Tibetan mush-rooms on diet-induced hypercholesterolemia in ratrdquo BrazilianJournal of Microbiology vol 46 no 2 pp 389ndash395 2015
[18] I Karam N Ma X-W Liu et al ldquoRegulation effect of AspirinEugenol Ester on blood lipids in Wistar rats with hyperlipi-demiardquo BMC Veterinary Research vol 11 article 217 2015
[19] G Jiao J B Wei M H Yin and P T Chun ldquoA new TCMformula FTZ lowers serum cholesterol by regulating HMG-CoA reductase and CYP7A1 in hyperlipidemic ratsrdquo Journal ofEthnopharmacology vol 135 no 2 pp 299ndash307 2011
[20] Y Geng W L Tu J J Zhang L Zhang and J Zhang ldquoEffectsof extract from cole pollen on lipidmetabolism in experimentalhyperlipidemic ratsrdquo The Scientific World Journal vol 2014Article ID 982498 6 pages 2014
[21] K Sueyoshi Y Nogawa K Sugawara T Endo and HHisamoto ldquoHighly sensitive and multiple enzyme activity assayusing reagent-release capillary-isoelectric focusing with rhod-amine 110-based substratesrdquo Analytical Sciences vol 31 no 11pp 1155ndash1161 2015
[22] D Fteita E Kononen M Gursoy E Soderling and U KGursoy ldquoDoes estradiol have an impact on the dipeptidylpeptidase IV enzyme activity of the Prevotella intermedia groupbacteriardquo Anaerobe vol 36 pp 14ndash18 2015
[23] L L Zhuo LWang and N Zhou ldquoRandomly arrayed G-quad-ruplexes for label-free and real-time assay of enzyme activityrdquoChemical Communications vol 50 no 52 pp 6875ndash6878 2014
[24] M Garelnabi and A Younis ldquoParaoxonase-1 enzyme activityassay for clinical samples validation and correlation studiesrdquoMedical Science Monitor vol 21 pp 902ndash908 2015
Evidence-Based Complementary and Alternative Medicine 11
[25] X Bi andZ Liu ldquoEnzyme activity assay of glycoprotein enzymesbased on a boronate affinity molecularly imprinted 96-wellmicroplaterdquo Analytical Chemistry vol 86 no 24 pp 12382ndash12389 2014
[26] A Walls S Dermody R Kumaran N Krishnan and E HHarley ldquoCharacterization of B-Cells in tonsils of patients diag-nosed with pediatric autoimmune neuropsychiatric disorderassociated streptococcusrdquo International Journal of PediatricOtorhinolaryngology vol 80 pp 49ndash52 2016
[27] I Karam N Ma X-W Liu et al ldquoRegulation effect of AspirinEugenol Ester on blood lipids in Wistar rats with hyperlipi-demiardquo BMC Veterinary Research vol 11 no 1 article 217 2015
[28] C Sunil S Ignacimuthu and C Kumarappan ldquoHypolipidemicactivity of Symplocos cochinchinensis S Moore leaves in hyper-lipidemic ratsrdquo Journal of Natural Medicines vol 66 no 1 pp32ndash38 2012
[29] W Shen Y Han B Huang et al ldquoMicroRNA-483-3p inhibitsextracellular matrix production by targeting smad4 in humantrabecular meshwork cellsrdquo Investigative Ophthalmology amp Vis-ual Science vol 56 no 13 pp 8419ndash8427 2015
[30] Z Wu Z Yi L Dong and A Zhang ldquoMolecular simulationstudy of the specific combination between four kinds of phthalicacid esters and human serum albuminrdquo Environmental Toxicol-ogy and Pharmacology vol 41 pp 259ndash265 2016
[31] K-J Hwang M Yoon H-J Hwang S-M Choi and S-MJeong ldquoMolecular dynamics simulation of oxygen ion conduc-tion in orthorhombic perovskite Ba-doped LaInO
3using cubic
and orthorhombic modelrdquo Journal of Nanoscience and Nan-otechnology vol 15 no 11 pp 8947ndash8950 2015
[32] H Yu W Han W Ma and K Schulten ldquoTransient 120573-hairpinformation in 120572-synuclein monomer revealed by coarse-grainedmolecular dynamics simulationrdquo The Journal of ChemicalPhysics vol 143 no 24 Article ID 243142 2015
[33] M Breuer K M Rosso and J Blumberger ldquoFlavin binding tothe deca-heme cytochrome mtrc insights from computationalmolecular simulationrdquo Biophysical Journal vol 109 no 12 pp2614ndash2624 2015
[34] V Rastija D Agic S Tomic et al ldquoSynthesis QSAR and mole-cular dynamics simulation of amidino-substituted benzimi-dazoles as dipeptidyl peptidase III inhibitorsrdquo Acta ChimicaSlovenica vol 62 no 4 pp 867ndash878 2015
[35] M Enciso C Schutte and L Delle Site ldquoInfluence of pH andsequence in peptide aggregation via molecular simulationrdquoTheJournal of Chemical Physics vol 143 no 24 Article ID 2431302015
[36] L XWangQWu andG P Peng ldquoA Study on determination ofcontent of 23-acetyl alisol B in Rhizoma Alismatis orientalitisrdquoJournal of Nanjing University of Traditional Chinese Medicine(Natural Science) vol 18 no 2 pp 105ndash107 2002
[37] G P Peng andY L Zhu ldquoProgress in the study on chemical con-stituents of Alisma orientalisrdquo Natural Product Research andDevelopment vol 18 no 2 pp 348ndash351 2006
[38] D Y Liu S J Gu J Wu et al ldquoStudies on fractions of zexiedecoction with antihyperlipidemia bioactivity in micerdquo ChinaPharmacist vol 13 no 6 pp 763ndash766 2010
[39] Q-F Luo L Sun and G-H Du ldquoProgress in new drugs tar-geting reverse cholesterol transportrdquo Chinese PharmacologicalBulletin vol 22 no 8 pp 904ndash907 2006
[40] M Yoshikawa and Y Fukuda ldquoHatakeayamardquo Chemical andPharmaceutical Bulletin vol 41 no 6 pp 1194ndash1196 1993
[41] F Xu L Zhang LHe et al ldquoMolecularmechanismon the inter-action between alisols and human serum albuminrdquo Acta Chim-ica Sinica vol 69 no 19 pp 2228ndash2234 2011
[42] F Xu J ChenW Gu et al ldquoStudy on transformation of effectivecomponents of Alisma under simulated in vivo conditionsrdquoChinese Journal of Pharmaceutical Analysis vol 31 no 11 pp2035ndash2039 2011
[43] H Yang Y G Gao and L X Zhang ldquoResearch progress on bio-synthetic pathway of terpenoids containing ginsenoside and theHMGRrdquo China Journal of Biological Engineering vol 28 no 10pp 130ndash135 2008
[44] G Z Cui Y L Ben N Li and D L Liu ldquoCatalytic mechanismand characteristics of HMG-COA recuctaserdquo Journal ofMedicalMolecular Biology vol 5 no 6 p 542 2008
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom

Evidence-Based Complementary and Alternative Medicine 11
[25] X Bi andZ Liu ldquoEnzyme activity assay of glycoprotein enzymesbased on a boronate affinity molecularly imprinted 96-wellmicroplaterdquo Analytical Chemistry vol 86 no 24 pp 12382ndash12389 2014
[26] A Walls S Dermody R Kumaran N Krishnan and E HHarley ldquoCharacterization of B-Cells in tonsils of patients diag-nosed with pediatric autoimmune neuropsychiatric disorderassociated streptococcusrdquo International Journal of PediatricOtorhinolaryngology vol 80 pp 49ndash52 2016
[27] I Karam N Ma X-W Liu et al ldquoRegulation effect of AspirinEugenol Ester on blood lipids in Wistar rats with hyperlipi-demiardquo BMC Veterinary Research vol 11 no 1 article 217 2015
[28] C Sunil S Ignacimuthu and C Kumarappan ldquoHypolipidemicactivity of Symplocos cochinchinensis S Moore leaves in hyper-lipidemic ratsrdquo Journal of Natural Medicines vol 66 no 1 pp32ndash38 2012
[29] W Shen Y Han B Huang et al ldquoMicroRNA-483-3p inhibitsextracellular matrix production by targeting smad4 in humantrabecular meshwork cellsrdquo Investigative Ophthalmology amp Vis-ual Science vol 56 no 13 pp 8419ndash8427 2015
[30] Z Wu Z Yi L Dong and A Zhang ldquoMolecular simulationstudy of the specific combination between four kinds of phthalicacid esters and human serum albuminrdquo Environmental Toxicol-ogy and Pharmacology vol 41 pp 259ndash265 2016
[31] K-J Hwang M Yoon H-J Hwang S-M Choi and S-MJeong ldquoMolecular dynamics simulation of oxygen ion conduc-tion in orthorhombic perovskite Ba-doped LaInO
3using cubic
and orthorhombic modelrdquo Journal of Nanoscience and Nan-otechnology vol 15 no 11 pp 8947ndash8950 2015
[32] H Yu W Han W Ma and K Schulten ldquoTransient 120573-hairpinformation in 120572-synuclein monomer revealed by coarse-grainedmolecular dynamics simulationrdquo The Journal of ChemicalPhysics vol 143 no 24 Article ID 243142 2015
[33] M Breuer K M Rosso and J Blumberger ldquoFlavin binding tothe deca-heme cytochrome mtrc insights from computationalmolecular simulationrdquo Biophysical Journal vol 109 no 12 pp2614ndash2624 2015
[34] V Rastija D Agic S Tomic et al ldquoSynthesis QSAR and mole-cular dynamics simulation of amidino-substituted benzimi-dazoles as dipeptidyl peptidase III inhibitorsrdquo Acta ChimicaSlovenica vol 62 no 4 pp 867ndash878 2015
[35] M Enciso C Schutte and L Delle Site ldquoInfluence of pH andsequence in peptide aggregation via molecular simulationrdquoTheJournal of Chemical Physics vol 143 no 24 Article ID 2431302015
[36] L XWangQWu andG P Peng ldquoA Study on determination ofcontent of 23-acetyl alisol B in Rhizoma Alismatis orientalitisrdquoJournal of Nanjing University of Traditional Chinese Medicine(Natural Science) vol 18 no 2 pp 105ndash107 2002
[37] G P Peng andY L Zhu ldquoProgress in the study on chemical con-stituents of Alisma orientalisrdquo Natural Product Research andDevelopment vol 18 no 2 pp 348ndash351 2006
[38] D Y Liu S J Gu J Wu et al ldquoStudies on fractions of zexiedecoction with antihyperlipidemia bioactivity in micerdquo ChinaPharmacist vol 13 no 6 pp 763ndash766 2010
[39] Q-F Luo L Sun and G-H Du ldquoProgress in new drugs tar-geting reverse cholesterol transportrdquo Chinese PharmacologicalBulletin vol 22 no 8 pp 904ndash907 2006
[40] M Yoshikawa and Y Fukuda ldquoHatakeayamardquo Chemical andPharmaceutical Bulletin vol 41 no 6 pp 1194ndash1196 1993
[41] F Xu L Zhang LHe et al ldquoMolecularmechanismon the inter-action between alisols and human serum albuminrdquo Acta Chim-ica Sinica vol 69 no 19 pp 2228ndash2234 2011
[42] F Xu J ChenW Gu et al ldquoStudy on transformation of effectivecomponents of Alisma under simulated in vivo conditionsrdquoChinese Journal of Pharmaceutical Analysis vol 31 no 11 pp2035ndash2039 2011
[43] H Yang Y G Gao and L X Zhang ldquoResearch progress on bio-synthetic pathway of terpenoids containing ginsenoside and theHMGRrdquo China Journal of Biological Engineering vol 28 no 10pp 130ndash135 2008
[44] G Z Cui Y L Ben N Li and D L Liu ldquoCatalytic mechanismand characteristics of HMG-COA recuctaserdquo Journal ofMedicalMolecular Biology vol 5 no 6 p 542 2008
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom

Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom