resting breathing pattern and its peripheral … · intrinsic mechanical properties of the...
TRANSCRIPT

1
RESTING BREATHING PATTERN and ITS PERIPHERAL MODULATION -Neural basis of the respiratory output -Descending pathways -Peripheral control of the spirogram : Inspiration 1. Intrinsic mechanical properties of the respiratory muscles 2. Chest wall ventilatory reflexes 3. Vagal control of the breathing pattern 3.1. Vagal reflexes a. maintained lung inflation b. maintained lung deflation c. airways occlusion at end-expiration 3.2. Vagal receptors a. slowly adapting stretch receptors b. rapidly adapting ‘irritant’ receptors c. bronchial C-fibers 4. Upper airways ventilatory reflexes 5. Ventilatory reflexes of cardiac or visceral origin Peripheral control of the spirogram: Expiration At a first glance, the mammalian resting breathing pattern appears to be an oscillatory event approximating a sinusoidal function, of which the amplitude is tidal volume (VT) and the period is the total breath duration (Ttot). In reality, a close look reveals major deviations from the sinusoidal function. For example, Ttot is not equally divided between the inspiratory and expiratory phases of the cycle, the inspiratory time (TI) being most commonly shorter than the expiratory time (TE). Also, the inspiratory phase of the spirogram is usually concave toward the time axis, whereas the expiratory phase is convex, occasionally with sudden interruptions of the expiratory flow [Fig.1]. Nevertheless, considering the

2
Fig.1. Change in volume as function of time during breathing: Spirogram. On top, the corresponding changes in airflow (pneumotachogram). TI, inspiratory time. TE, expiratory time. TTOT, total cycle duration. V T, tidal volume. V, airflow. enormous variety in shape and size among mammals, one cannot avoid being amazed by the similarities of the spirogram [Fig.2]. Equally astonishing is the facts that the spirogram is not drastically modified by changes in body postures, from supine

3
Fig.2. Spirograms obtained in different species during resting condition. From top to bottom, left and right: dog (vertical bar = 500 ml), goat (500 ml), horse (2 l), 3-day old human infant (30 ml), 1-day old rabbit (1 ml), 3-day old mouse (0.01 ml), 1-day old puppy (10 ml), 1-day old piglet (20 ml), 3-day old seal (300 ml), 2-day old kitten (1 ml), mule (1 l). Horizontal bar, in all cases, 5 sec. to prone, sitting or standing, extension and flexion of the neck, or switching from nose to mouth breathing, or even by a modest exercise as slow walking, all conditions which should imply substantial changes in the mechanical impedance of the respiratory system. Experimentally, it is possible to further explore the 'stability' of the spirogram. For example, an experiment can be designed to suddenly decrease the compliance of the respiratory system (elastic load), by having a subject breathing from a container of limited capacity, instead of breathing from the environment. Equally, a resistive load can be imposed to the respiratory system by asking the subject to breathe through a narrow tube. The size of the elastic load could be chosen to decrease the compliance of the subject's respiratory system by half, and the size of the tube could be chosen to double his respiratory resistance. In either case, one would expect that the imposed load should decrease VT to half of the unloaded control breath. However, this is not the case; the loaded VT will be only slightly less than control, and invariably more than the predicted half. In other words, the respiratory system is capable of recognizing the mechanical load, and is able to respond to it so quickly that volume compensation occurs within the breath itself. Load compensation is a fundamental aspect of the breathing act. The respiratory pump, designed to be in the middle of the body, shares muscles and structures with multiple non-respiratory functions, including locomotion, and is therefore subjected to many continuously changing loads. If there was no mechanism to control the mechanical result of the neural output, breathing would probably be a chaotic and energetically demanding act, with catastrophic effects on the distribution of ventilation and blood gas homeostasis. Hence, mechanisms have evolved to carefully monitor the changes in lung volume, and to promptly signal to the neural networks responsible for integration any deviations from the desirable pattern. The respiratory system can therefore be seen as a neuro-mechanical unit of which the mechanical component is a crucial element conditioning the neural motor output itself.

4
The functional organization of the neuro-mechanical unit is based on the 'feedback loop criteria': the respiratory neurons send impulses to the respiratory muscles and, from the periphery, information is conveyed back, mostly from specific receptors placed in the lungs and the chest [Fig.3].
Fig.3. The main components of the neuro-mechanical unit translating the neural output into a mechanical event. At left, some examples of mechanical constraints. Information about the changes in lung volume is fed back to the respiratory pattern generator mostly via activation of muscle proprioceptors and airways receptors ('chest wall' and 'lung' reflexes). Humoral stimuli, which include blood gases, also reach and influence neurons controlling the respiratory motor output, but because of their nature they can only act on a longer time scale, and are aimed to the control of ventilation, rather than the control of tidal volume.
NEURAL BASIS OF THE RESPIRATORY OUTPUT Since the beginning of the last century, it was noticed on anaesthetised preparations that respiration was not affected by removal of the cerebrum and cerebellum, whereas breathing ceased when the medulla was cut at a level which included the nerve roots of the vagus. At the end of the last century it was believed that rhythmic respiration resulted from periodic inhibition of the "inspiratory center" located in the medulla by neural signals originating in the inferior colliculi (upper pons) and the vagal afferents. In the 1920s, Lumsden, through careful brainstem transections in different mammalian species concluded that the central respiratory system can be divided into three centers: pneumotaxic , apneustic and inspiratory centers, the latter further

5
subdivided into the expiratory and gasping centers. Indeed, the term 'center' in referring to nuclei or pools of neurons involved in respiratory regulation, albeit often criticized, is still commonly used. The pattern of breathing which follows the transection of the brainstem at the level indicated by Lumsden before (left) and after bilateral section of the vagi nerves (right) is schematically represented in Fig.4. A transection above the inferior colliculi (A) does not affect the pattern of breathing. However, if the sections were done just above the cerebral peduncle (B) or if the vagi nerves were cut, breathing would become deep and slow. If the section in B were accompanied by vagotomy, breathing would then be apneustic, which is not, however, a permanent state since some rhythmic breathing returns as hypoxia and respiratory acidosis progress. Finally, a section at the acoustic striae (C) determines an irregular pattern of breathing: nevertheless, even in these conditions, the medulla is still capable of rhythmically driving respiration, even in the absence of afferents from the cranial nerves.
Fig.4. Schematic view of the brain stem, and of the breathing pattern following transections at the levels indicated by the dashed lines. Left: vagi intact. Right, vagi cut. NA, Nucleus Ambiguus . NRA, Nucleus Retroambigualis. DRG, Dorsal Respiratory Group. IV, fourth ventricle. Through electrical stimulation and neural recordings, it has been found that the Lumsden's pneumotaxic center, presently referred to as "pneumotaxic mechanism ", corresponds to the nucleus parabrachialis, in the dorsolateral rostral pons. Its function is to modulate, and possibly drive, the rhythmic respiratory neurons in the medulla. It is however, important to acknowledge that even the isolated medulla can generate an intrinsic respiratory rhythm. The observation that rhythmic breathing continues when the brainstem has been transected several millimetres rostral to the obex has led many investigators to concentrate their attention on the respiratory nuclei of the medulla as the potential site for respiratory rhythmogenesis.

6
Several main aggregations of rhythmic respiratory neurons have been localized in the medulla [Fig.5], placed dorsally and ventrally (VRG). The dorsal respiratory group (DRG) is associated with the Nucleus of the Tractus Solitarius (NTS), and would include exclusively inspiratory respiratory units, i.e. units firing in phase with the phrenic burst. The ventral groups are associated with the region of the Nucleus Ambiguus (NA) and the Nucleus Retroambigualis (NRA), and contain both inspiratory and expiratory neurons. Some evidence suggests that the Bötzinger complex, located rostral to NA, plays an important role in respiratory rhythmogenesis. The upper VRG (NA) consists of motoneurons of cranial nerves which innervate the ipsilateral accessory muscles of respiration. The caudal component of VRG (NRA) is the source of rhythmic drive to inspiratory and expiratory intercostals and probably abdominal expiratory motoneurons, with controlateral as well as ipsilateral projections.
Fig.5. Respiratory nuclei in the medulla. Schematic representation of the medullary center. Dorsal respiratory group (DRG); Inferior olivar nucleus (ION); Nucleus Ambiguus (NA); Nucleus Retroambigualis (NRA); Nucleus Tractus Solitarius (NTS); Obex (OB); Ventral horn (VH); Ventral respiratory group (VRG); Hypoglossus nucleus (XII), Vagus Nerve (X); Glossopharingeal Nerve (IX). The descending axons seem to functionally separate within the spinal pathways into ‘voluntary’ cortico-spinal tracts and automatic pathways for expiratory and inspiratory motoneurons.

7
The nature of the cellular organization of the respiratory nuclei involved in the generation of respiratory activity, including those presumed involved in breathing rhythmicity, still remains one of the major mysteries of the neurophysiology of breathing. Two main possibilities can be considered in the mechanisms generating a rhythmic pattern of neuronal activity, the pacemaker cell and the neural network. The pacemaker cell does not require interactions with other neurons and its basic pattern can be ascribed to the intrinsic properties of the cell, as the pacemaker cells of the sinoatrial node of the heart. Cells characterized by a maintained output pattern that is rhythmic or continuous and not dependent on interactions with other cells have been described, e.g. in the abdominal ganglion of Aplysia and in the sub-esophageal ganglion of both hermit crabs and lobsters. The key test in demonstrating the presence of a pacemaker cell is to show that its rhythmic pattern continues in the absence of inputs to the cell; this demonstration has not yet been provided for any neuron of the medullary area in vivo. The neural network possibility would imply interactions among cells of a network resulting in a rhythmic output pattern. In the absence of neuronal interactions, no single cell of the network is capable of generating a rhythmic pattern. Many models of neural networks have been proposed, either based on reciprocal inhibition and mutual excitation between inspiratory and expiratory cells ("Bistable Oscillator") or on phasic inhibition of inspiratory cells through action of interneurons ("Inhibitory Phasing"). These models are refined and updated according to the most recent findings about the properties and interconnections of the medullary respiratory cells, as the search for a satisfactory explanation about the generation of respiratory rhythmicity continues. Finally, it cannot be excluded that respiratory activity can be originated by more than one network capable of originating and sustaining respiratory activity. For example, some evidence indicates that in severe hypoxia or anoxia, after termination of eupnea, gasping activity may be the result of activation of neural pools not normally involved with the eupneic breathing pattern.
DESCENDING PATHWAYS It is a common experience that the breathing pattern can be voluntarily controlled, although in the large majority of cases it is proceeding automatically. In other words, cortical (voluntary) control can compete and override the automatic control. The separation between automatic and voluntary control seems to continue along the spinal axis, with separate pathways of the descending neura l signals that drive the respiratory motoneurons. The extent of the anatomical separation between the two control systems is difficult to establish with certainty, being based mostly on rare clinical observations and postmortem analysis of unusual pathological conditions. Some breathing acts associated with consciousness, such as speaking, ventilatory responses to anxiety and fear, breath holding and voluntary hyperventilation, could involve alterations in the respiratory pattern by bypassing entirely the respiratory network, and overriding the automatic drive at the level of the spinal motoneurons. In addition, evidence is accumulating that spinal respiratory neurons are not simple relay stations, and significant integration of neural information occurs at these levels. This 'cortical' control of breathing would project through pathways located in the dorsolateral columns of the spinal cord [Fig.5]. The automatic 'involuntary' pathways of the rhythmic pattern as well as of reflex involuntary ventilatory acts such as cough, hiccup and yawning lie in the ventral and lateral columns of the cord [Fig.5]. Furthermore, the rhythmic expiratory pathway takes up a more medial position than the rhythmic inspiratory pathway. This separation of neural descending pathways should imply the possibilities of losing the voluntary control of breathing, retaining its automatic aspect, as well as the opposite situation, whereby breathing can continue only as long as it is voluntarily (cortically) controlled. Cases of patients with long periods of apnea, even during wakefulness, but capable of breathing upon command, have been described*.

8
[*Ondine's course, after the german mythological story of the nymph Ondine, who, betrayed by her mortal lover, condemned him to 'think' about all his automatic functions, including breathing: when he eventually fell asleep, he died] In practice, these conditions would require well-defined bilateral lesions. Hence, they are extremely rare. The 'primary alveolar hypoventilation ' is a somewhat general term which comprises these cases as well as a number of other clinical conditions of reduced resting ventilation and reduced ventilatory response to chemical stimuli ( i.e. the 'locked-in syndrome).
PERIPHERAL CONTROL OF THE SPIROGRAM: INSPIRATION 1. Intrinsic mechanical properties of the respiratory muscles. The relation between the nervous stimuli to a respiratory muscle and the force developed depends on the length of the muscle, with maximal force generated at the optimal length (lo). For the same degree of activation, a muscle develops progressively less force as it shortens (Force-Length relationship). In addition, at any muscle length, the force generated is inversely proportional to the speed of fiber shortening (Force-velocity relationship) [Fig.6].

9
Fig.6. Mechanical properties of a skeletal muscle.

10
Top: Force (F) - length (l) diagram, constructed by maximal stimulation of the muscle at various length. Maximal force is obtained at length lo., which approximately corresponds to the resting length of the muscle. Above lo, T is contributed by the passive stretching of muscle (elastic elements ‘in parallel’); hence, the tension developed by the muscle under stimulation is the difference between the curve T and P, represented by the dashed line. Bottom: Force (F) - velocity (v) diagram, at various l / lo , as the muscle is shortening or is elongated. The area underneath the curve has the physical dimensions of work/time, or power. Power is nil at maximal velocity (because the force is zero) and during isometric contraction (because the muscle does not shorten), and varies with the speed of shortening as indicated by the dotted line. Hence, an isometric (constant length) contraction at lo generates the largest force. The lo of the inspiratory muscles is at very low lung volumes, whereas lo of the expiratory muscles is around the largest lung volumes attainable. This can be appreciated by attempting to maximally contract either set of muscles against occlusion of the airways, i.e. with nostrils and lips closed. During this manoeuver, and neglecting phenomena of chest distortion, the muscles can be considered as contracting in isometric (i.e. no-shortening) conditions. During inspiratory efforts the airways pressure is negative, the maximal negative value occurring when the manoeuvre is performed at very low volumes. On the contrary, the largest positive airways pressures during contraction of the expiratory muscles are generated at the largest lung volumes (cf."Mechanics", fig.8)*. [*The implications that changes in lung volume have on the pressure (P) generated by the diaphragm would not differ even if the diaphragm mechanical action was modelled by a dome which generates P according to the Young-La Place relationship (Tension = Presure · radius of curvature of the dome, T=P · r). At high lung volumes the flatter shape of the diaphragm implies a larger dome with a greater r, resulting therefore in lower P; the opposite occurs at low lung volumes] This effect of muscle length on force and pressure generation applies also during resting breathing. Any obstacle (load) to respiratory muscle shortening during inspiration increases the muscle force, therefore providing a mechanism for load compensation. In fact, an elastic load reduces the extent of muscle shortening, and a resistive load reduces the speed of its shortening. In either case, the force will be increased, tending to offset the mechanical impairment produced by the load. The force-length compensation is one of the mechanisms that comes into play in protecting VT against small changes in FRC. Consider, for example, the increase in FRC which occurs in going from the supine to the standing posture (about 1 liter in an adult man). On the one hand, the larger lung volume reduces the length, and therefore should reduce the force, of the inspiratory muscles. On the other hand, because of the shape of the pressure-volume curve of the respiratory system, as lung volume increases the recoil pressure that the muscles have to overcome increases, therefore offering an elastic mechanical load which helps to maintain muscle length and preserve its force. Eventually, as lung volume progressively increases, the load of the low compliance becomes an energetic burden, rather than a mechanical compensation; hence, at high lung volumes, breathing requires a major activation of the inspiratory muscles in order to generate an acceptable VT. This situation, faced by some patients with lung hyperinflation because of chronic obstructive lung disease, is energetically very demanding. One strategy adopted by these patients is that of contracting the abdominal muscles simultaneously with the diaphragm. The manoeuver, by increasing abdominal pressure, effectively represents another mechanical trick:

11
by increasing abdominal pressure during inspiration, the patient attempts to maintain the length of the diaphragm, hence improving its force and mechanical efficiency. It is obvious that even this strategy is only a temporary relief for the hyperinflated patient; it is effective only as long as the hyperinflation is not too extreme, at which point the cost of contraction of the abdominal muscles would more than offset its benefits. 2. Chest wall ventilatory reflexes Afferent activity from respiratory muscles can influence ventilation by projecting to the respiratory controller (supraspinal reflexes), or via segmental and inter-segmental spinal reflexes. These are based upon the proprioceptive control of muscle activation by the muscle spindles (facilitatory on the alpha motoneurons) and the Golgi tendon organs (inhibitory), which send their information through the γ-afferents via the dorsal roots. Their reflex effects are different. Stretching of the intercostal muscles, which contain many spindles, increases muscle force (facilitation), whereas stretching the diaphragm, which contains mainly tendon organs, inhibits its contraction. Supraspinal reflexes include the intercostal-to-phrenic reflex, which consists of an inhibition of phrenic nerve activity by intercostal nerves stimulation. This reflex is mediated by brain stem structures, and inhibits not only bulbo-spinal inspiratory neurons but also motoneurons to the laryngeal abductory muscles, which are normally active in synchrony with the diaphragm. Although the intercostal (and abdominal) muscles afferents have supraspinal ventilatory effects similar to those of the vagal slowly adapting stretch receptors (SAR, see below), some evidences suggest that their central projections differ. 3. Vagal control of the breathing pattern
3.1. Vagal reflexes The vagus, tenth cranial nerve, is a mixed (sensory and motor) bilateral nerve. Its sensory neurons have their body in the nodus and petrosus ganglions, and the afferent fibers, partly myelinated and mostly not-myelinated, innervate a variety of thoracic and abdominal organs [Fig.7].

12
Fig.7. Schematic view of the multiple afferent and efferent fibers contained in the vagi nerves. m, myelinated; nm, not myelinated. (Numbers are based from data provided by Paintal , Ergeb.Physiol. 52:75-156, 1963, and Agostoni et al., J. Physiol. 135:182-205, 1957). It has long been recognized that section of the vagi in the neck (before entrance into the vertebral column) has a major effect on the breathing pattern; breathing frequency is reduced and tidal volume increased (e.g. Fig.4, left). Similar effects are apparent during electric block of the vagal conduction [Fig.8]. VE is little affected or decreased, depending upon the species, age, and state of consciousness. In birds, vagotomy can lead to dyspnea and asphyxia, with low arterial PO2 and pH.

13
Fig.8. Records of airflow (top) and tidal volume (bottom) in an anaesthetised rabbit. The horizontal bar indicates the duration of the anodal vagal block. Time marker, 1 sec. Because these effects on breathing pattern are seen also upon selective intrathoracic denervation of the lung, they are attributed to elimination of the innervation to the lung. Of course, sectioning the nerves eliminates both afferent and efferent pulmonary innervation. One of the effects of abolishing the efferent (motor) innervation is that of reducing the tone of the smooth muscle in the airways (bronchomotor tone). Because in the intact condition there is, normally, some bronchomotor tone, its reduction results in a slight broncho-dilatation, and reduction in airways resistance. Instead of severing the nerves, it is possible to selectively block the conduction of only the larger myelinated fibers, either pharmacologically or with appropriate adjustment of the anodal current. The results on the breathing pattern are similar to those observed with bilateral section, i.e. the pattern becomes deeper and slower. In addition, cough is reduced or eliminated, and the spontaneous deep breaths, or sighs, periodically occurring during undisturbed breathing (e.g. Fig.17), are abolished. Vagotomy, or pulmonary deafferentation, modifies not only the resting breathing pattern but also the pattern during hyperpnea. For example, during asphyxic hyperventilation (a combination of hypercapnia and hypoxia, as it occurs during rebreathing of the expired air) both tidal volume and breathing rate increase. However, after block of the vagal conduction, only tidal volume increases, with little or no changes in breathing rate [Fig.9].

14
Fig.9. Left. Anaesthetised rabbit. Changes in VE and VT during rebreathing following artificial hyperventilation, before (open symbols) and after (filled symbols) bilateral vagotomy. Oblique lines are iso-frequency lines. Right. VE and breathing rate in an adult man breathing a CO2-enriched gas mixture, before and after bilateral block of the vagi in the neck with a local anaesthetic (from Guz et al., Cli.Sci. 27:293-304, 1964). The available observations indicate that vagal afferents are of importance in regulating the depth and frequency of breathing in most vertebrates, including man, although there could be major quantitative differences among species. For example, in man after block of the vagi the resting breathing pattern is little affected, and its effects become clearly visible only during hyperpnea. Sleep or anaesthesia can magnify the vagal control of the breathing pattern. Several manoeuvres or special conditions, such as those mentioned below, are useful for a better understanding of how vagal afferents control the breathing pattern. a. Maintained lung inflation In an intubated animal or patient, during spontaneous breathing the endotracheal tube can be briefly occluded at end-inspiration. This manoeuver can be easily performed also in infants breathing through a face-mask connected to a pneumotachograph. During the occlusion lung volume is artificially maintained elevated, and it can be readily appreciated that the next inspiratory effort is delayed [Fig.10]. The larger the lung volume, the bigger the delay. If the vagi were cut or blocked, there would be no delay, irrespective of the lung volume. This inhibition of the breathing pattern during maintained lung inflation is known as the Hering-Breüer inflation reflex, although the distinction is sometimes made between the inspiratory inhibitory and the expiratory facilitatory components of this reflex, depending upon the timing of the
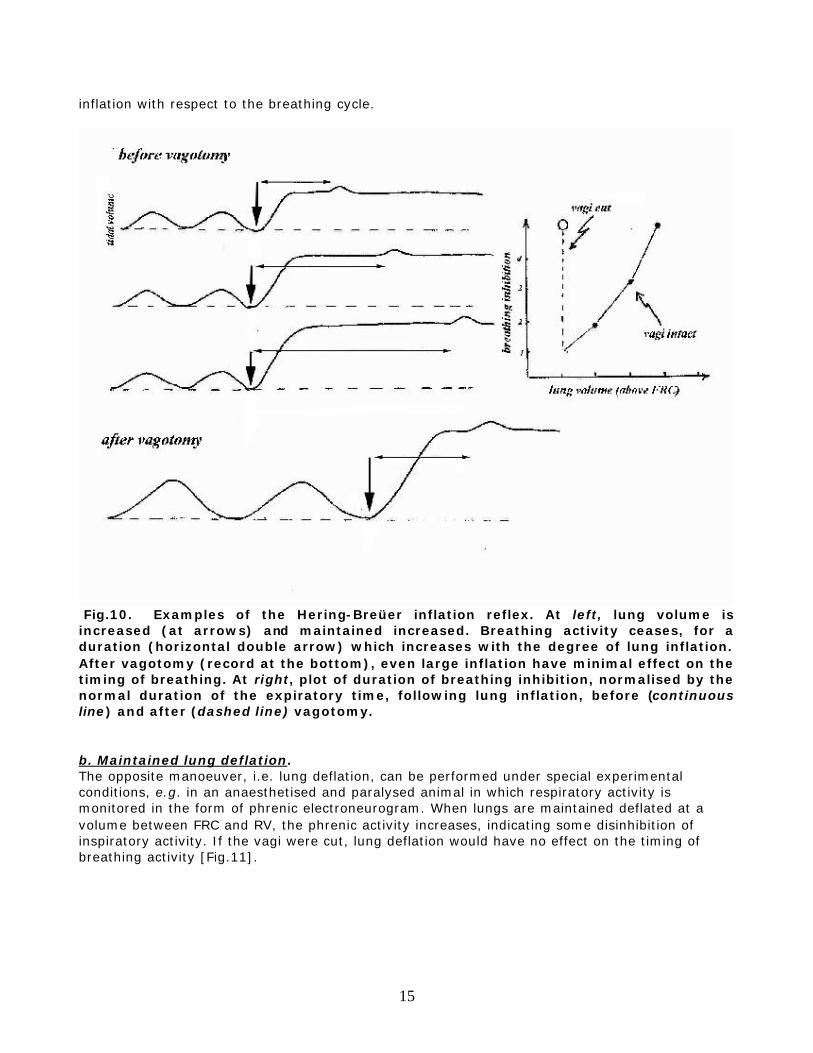
15
inflation with respect to the breathing cycle.
Fig.10. Examples of the Hering-Breüer inflation reflex. At left, lung volume is increased (at arrows) and maintained increased. Breathing activity ceases, for a duration (horizontal double arrow) which increases with the degree of lung inflation. After vagotomy (record at the bottom), even large inflation have minimal effect on the timing of breathing. At right, plot of duration of breathing inhibition, normalised by the normal duration of the expiratory time, following lung inflation, before (continuous line) and after (dashed line) vagotomy. b. Maintained lung deflation. The opposite manoeuver, i.e. lung deflation, can be performed under special experimental conditions, e.g. in an anaesthetised and paralysed animal in which respiratory activity is monitored in the form of phrenic electroneurogram. When lungs are maintained deflated at a volume between FRC and RV, the phrenic activity increases, indicating some disinhibition of inspiratory activity. If the vagi were cut, lung deflation would have no effect on the timing of breathing activity [Fig.11].

16
Fig.11. An example of the breathing response to lung deflation, in an anaesthetised rabbit. With the vagi intact (left) lung deflation (at arrow) increases the respiratory output, as apparent by the longer duration of the electromyographic activity of the diaphragm. At right, the conduction of the vagal fibers has been blocked for the duration of the horizontal bar, as apparent from the longer duration of the EMG during resting breathing. Upon lung deflation (arrow) EMG activity does not increase any further. c. Airways occlusion at end-expiration. When the airways are occluded at end-expiration, the duration of the next inspiratory effort can be determined from the recording of airways pressure or that of the diaphragm or phrenic nerve; the duration of the effort is most commonly longer than the previous control (open airways) inspirations [Fig.12]. If the vagi were cut, or blocked, the inspiratory effort against the end-expiratory occlusion would last as the normal (open airways) inspiratory time.

17
Fig.12. Records of tidal volume (VT) and intra-airways pressure (P) in a spontaneously breathing, anaesthetised rabbit. At the arrow the airways are occluded for the duration of one inspiratory effort. The duration of the effort is longer than the normal (open airways) inspiratory time. After block of the vagi (records at bottom) the difference is no longer apparent. The above observations, and many others, indicated that pulmonary vagal afferents play an inhibitory role on inspiration approximately in proportion to lung volume. When lung volume is maintained elevated (a.), inspiration is inhibited. When lung volume is decreased (b.), inspiratory output increases. When lung volume is not permitted to increase (c.) the inspiratory output, as measured from the duration of the inspiratory effort, increases. Hence, vagotomy or vagal block result in deeper inspiration (e.g. Fig.8), because no lung-volume related inhibition is sent to the respiratory output generator*. [*Eventually, even after vagotomy, inspiration terminates because of the inspiratory inhibition within the respiratory neural network. If also these nuclei were lesioned, inspiratory activity may continue undisturbed for a longer time, resulting in a maintained lung inflation (apneustic breathing pattern, e.g. Fig. 4, at right)]

18
The longer duration of the breathing cycle, hence the lower breathing rate after vagotomy, reflects the longer time needed for the deeper inspiration and the following expiration.
3.2. Vagal receptors A better understanding of the mechanisms behind the vagal ventilatory reflexes, and their implications in the regulation of the breathing pattern, can be appreciated by a close look at the behaviour of the pulmonary vagal afferents during the manoeuvres described above. This can be done by recording vagal afferent activity from the peripheral cut end of the nerve, or, for a more accurate analysis, from only one or a few afferent fibers.
Fig.13. Action potentials obtained by recording from single fibers of the peripheral cut end of the vagus, during spontaneous ventilation (left) and during sustained lung inflation (right). At top a rapidly adapting ("irritant") receptor, at bottom a slowly adapting pulmonary receptor. Fig.13 shows two main patterns of afferent fiber activity that can be recorded during tidal ventilation and when the lung volume is maintained inflated. In the latter case, it is easy to recognize two quite different patterns, a sustained activity, with little adaptation, from the slowly adapting pulmonary receptors, or a short lasting burst, with rapid adaptation, from the rapidly adapting pulmonary receptors. Both are 'stretch' receptors, meaning that they respond to tissue stretch. The study of the spontaneous activities of these receptors, and their responses to

19
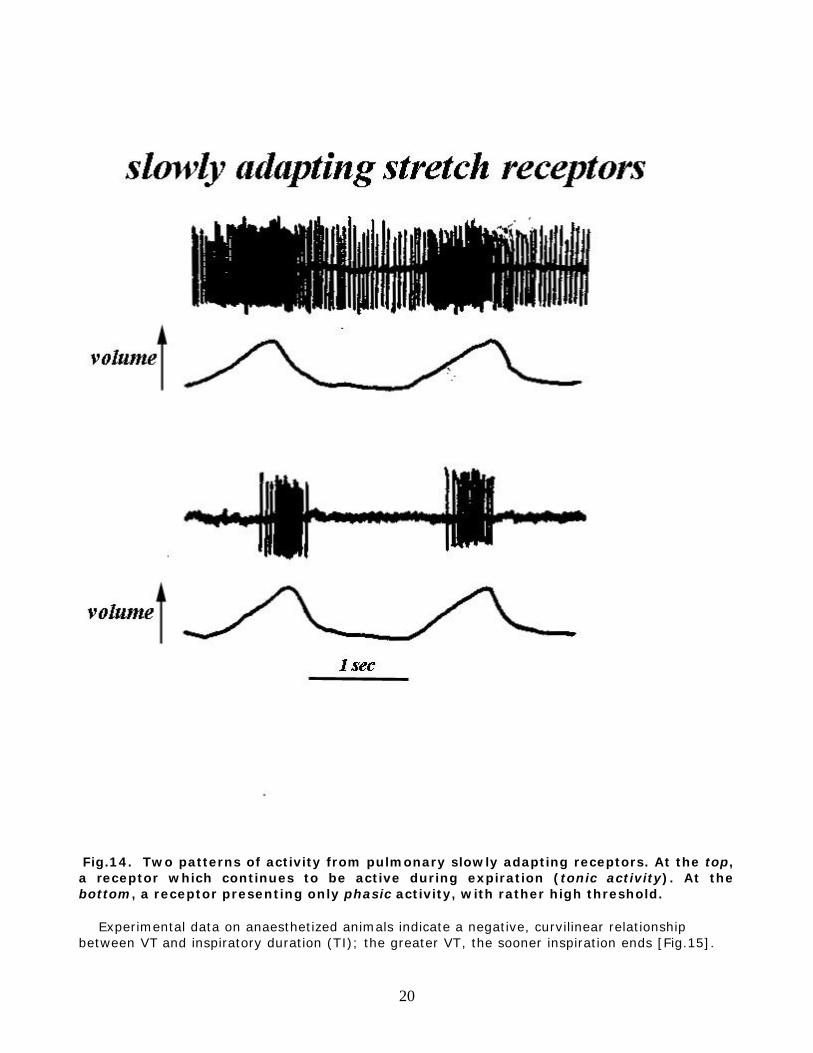
various conditions, is a relatively straightforward task. Much more complex is the interpretation of their reflex effects on breathing. In fact, studies aimed to the latter are often based on approaches which presume a selective inhibition or stimulation of only one group of receptors, with the hope that activities and responses from the other receptors remain unchanged. The correctness of these assumptions is often very difficult to prove. a. Slowly adapting stretch receptors (SAR). They are located in the larger airways of the lungs, with only a minority in the very periphery. With respect to the circumferential location, SAR are found in the posterior wall of the airways; this comprises the layers of soft tissue that complete the horse-shoe cartilaginous ring, the characteristic supporting structure of the larger airways in the tracheobronchial tree of most mammals. Because the SARs sustained activity parallels the prolongation of expiration during lung inflation, SAR are considered the receptors mediating the Hering-Breüer inflation reflex, a view supported by several other experimental approaches. Some SAR are activated by lung expansion when lung volume exceeds FRC, whereas others are also active at end-expiration [Fig.14]. This has led to the separation of a phasic and a tonic SAR inspiratory inhibition, the latter lasting throughout the breathing cycle, whereas the phasic activity appears only when lung volume increases above FRC. In other words, even at end-expiration, some SAR are active. The reduction of this tonic inhibition when lung volume is reduced below FRC may contribute to the facilitation of inspiration observed with this manoeuver (b, above).
20
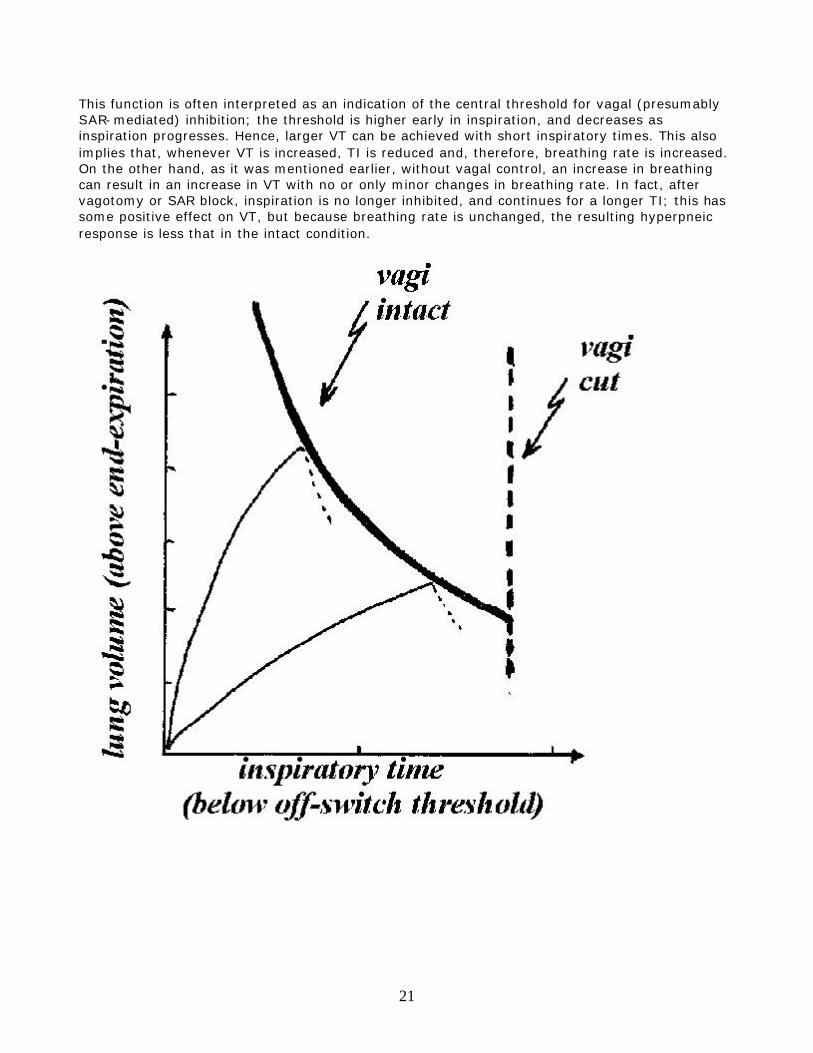
Fig.14. Two patterns of activity from pulmonary slowly adapting receptors. At the top, a receptor which continues to be active during expiration (tonic activity). At the bottom, a receptor presenting only phasic activity, with rather high threshold. Experimental data on anaesthetized animals indicate a negative, curvilinear relationship between VT and inspiratory duration (TI); the greater VT, the sooner inspiration ends [Fig.15].
21
This function is often interpreted as an indication of the central threshold for vagal (presumably SAR-mediated) inhibition; the threshold is higher early in inspiration, and decreases as inspiration progresses. Hence, larger VT can be achieved with short inspiratory times. This also implies that, whenever VT is increased, TI is reduced and, therefore, breathing rate is increased. On the other hand, as it was mentioned earlier, without vagal control, an increase in breathing can result in an increase in VT with no or only minor changes in breathing rate. In fact, after vagotomy or SAR block, inspiration is no longer inhibited, and continues for a longer TI; this has some positive effect on VT, but because breathing rate is unchanged, the resulting hyperpneic response is less that in the intact condition.

22
Fig.15. The tidal volume - inspiratory time (VT-TI) relationship. The curvilinear thick line represents the vagal inspiratory inhibitory threshold, which is high early in inspiration and decreases as TI progresses. Two inspiratory trajectories are represented by the thin lines. When they hit the vagal threshold, inspiration ceases, and expiration begins (dotted lines). Without pulmonary vagal feed back, the duration of TI is fixed, irrespective of the tidal volume (vertical dashed line). This view of the role of SAR in the regulation of the breathing pattern is undoubtedly simplistic, and, to some extent, contradicted by data collected in conscious animals; however, it represents a useful framework, as a starting point for the interpretation of a number of experimental observations. Although for many practical purposes it is useful to think of the SAR as receptors responding to a change in lung volume (or in transpulmonary pressure), in reality it can be demonstrated that their most appropriate stimulus is the change in intra-tissue tension. This notion becomes important when the vagal control of the breathing pattern is interpreted during conditions that change the mechanical properties of the airways, or bronchomotor tone. For example, a reduction in airways compliance, or an increase in bronchomotor tone, increase SAR activity, even if FRC and VT remain unaltered, with predictable effects on the breathing pattern. Carbon dioxide is known to decrease the activity of SAR, although in mammals, differently from birds, only high concentration of CO2 have appreciable inhibitory effects on these receptors. The inhibition of SAR activity by CO2 has been postulated to facilitate the hyperpnea during exercise. b. Rapidly adapting 'irritant' receptors (RAR). As the SARs, also RARs are located mostly in the larger airways. They are thought to be multi-branching endings, placed more superficially than the SAR, possibly in the epithelium, with location all around the airways circumference. RAR are also called 'irritant ' receptors because a number of conditions, including airways noxious agents, can activate them, probably because of their rather superficial location. For example, in addition to pneumothorax, anaphylaxis or histamine injection, lung edema, and pulmonary embolism, their activity is increased by inhalation of dust, cigarette smoke, some gaseous irritants, like ammonia, and some chemicals reaching the airways via blood perfusion, like histamine or diphenyl-diguanide. Most of, if not all, these conditions are characterized by a decrease in total lung, or regional pulmonary, compliance. Indeed, it has been shown that RAR activity is inversely proportional to lung compliance. Hence, it is possible that increased tension in the microenvironment of the receptor represents the common feature of the various stimuli and conditions known to activate these receptors*. [*A decrease in compliance implies, for the same VT, a larger increase in transpulmonary pressure and tissue tension]

23
Fig.16. Spirometric record in an anaesthetised supine rabbit. For the duration of the horizontal bar the animal is maintained in the vertical head-up position. Because of the descent of the abdominal viscera and diaphragm, the lungs are suddenly inflated, triggering the vagal Hering-Breüer inflation reflex. At the moment of the inflation, however, note the facilitation of inspiratory activity (Head’s paradoxical reflex). After vagotomy, the Head’s reflex and the Hering-Breüer inflation reflex would no longer occur. The role played by RAR in the regulation of the breathing pattern is not as clear as that of the SAR. Their short lasting activation seems to promote inspiratory activity. Hence, a sudden large increase in lung volume, at first, could elicit a paradoxical facilitation of inspiratory activity (Head's paradoxical reflex), briefly overcoming the Hering-Breüer inspiratory inhibition [Fig.16]. This effect was thought to play a role in the air expansion of the lungs at birth; the first breath is in fact usually the deepest of the whole neonatal period. The deep breath, or sigh, commonly occurring during steady resting breathing [Fig.17], at a rate which varies among species, is also thought to be mediated by activation of RAR; during resting breathing the airways naturally tend to close, reducing lung compliance and gradually increasing the tidal change in transpulmonary pressure.

24
Fig.17. Records of tidal volume (top), airflow (middle) and diaphragm electromyogram (EMG) during spontaneous resting breathing. Note the deep breath, or sigh. Quite differently from the response to a rapidly occurring and transient stimulus, prolonged stimulation of RAR, as obtained pharmacologically with histamine, would result not in a prolongation, but rather in a shortening of TI, eventually leading to a rapid and shallow pattern. In summary, the RAR could be seen as part of the airways neural defense mechanisms, possibly with the reflex effect of enhancing inspiration when lung compliance drops, and shortening it when the stimulus persists. It may be argued that promoting inspiration in face of a chemical challenge is not a good strategy. However, the inspiratory activity can be followed by fast expiration; in fact, RAR are probably the main receptors controlling cough of intrapulmonary origin. c. Bronchial C-fibers. Although the majority of vagal afferents are small not-myelinated fibers [Fig.7], not much is known about their characteristics and precise location. Some pulmonary C-fibers seem to respond to drugs (i.e. serotonin, capsaicin, phenyldiguanide) injected into systemic circulation, others only to right heart injection, suggesting locations accessible mostly from, respectively, the bronchial or the pulmonary circulation. At least some of these fibers were thought to be located in the alveolar-capillary region, and to respond to increases in local tension. Upon stimulation, their reflex effect would consist in inhibition of breathing at first, followed by rapid and shallow pattern if the stimulus persists. In this respect, they have been considered responsible for the rapid and shallow breathing pattern during interstitial edema. Indeed, it would seem important to have accurate monitoring of any change in alveolar

25
interstitial pressure, given the importance of this area for gas diffusion, but whether this is really the role of the C-fibers is still not conclusively proven. 4. Upper airways ventilatory reflexes. The multiple tasks of the upper airways passages explain the abundance of innervation and fine muscle control of this region. Being anatomically in series with both the digestive and the respiratory systems*, the coordination of breathing with deglutition is [*This is not true for all mammals. In whales, for example, the two systems are anatomically separated] an obvious task. Normally, the latter interrupts breathing, although infants seem capable of sucking and swallowing simultaneously with breathing. The defence and protection of the lower airways passages, including filtering, warming and humidification of the inspired air, phonation in its various forms, postural needs, and thermoregulatory functions are additional responsibilities which make the upper airways an anatomically complex, and functionally busy, region. In addition to the nasal- and oral-pharyngeal areas, with afferent innervation mostly by the trigeminal nerve, the larynx is a particularly important source of ventilatory reflexes. It is innervated by the superior laryngeal and the recurrent (or inferior laryngeal) nerves, all branches of the vagus. Single fiber recording from these nerves has indicated the existence of receptors sensitive to mechanical stimuli (pressure, tension, airflow), osmo- (sensitive to water) and ions-sensitive receptors, as well as temperature-sensitive receptors. Stimuli applied throughout the upper airways, and in the laryngeal area in particular, could have both facilitatory and inhibitory effects on breathing, although, in general, inhibitory effects seem to prevail, both on tidal volume and breathing rate. This response is much more apparent and powerful in newborn and young animals, than in adults. At a superficial look, such respiratory inhibition of upper airways origin may recall the 'diving reflex', classically characterised by the trilogy apnea, bradycardia and peripheral vasoconstriction, and mediated by trigeminal afferents from the facial region, possibly in combination with baro- and chemo-reflexes. The laryngeal ventilatory reflexes are strictly of laryngeal origin, and local anaesthesia or section of the superior laryngeal nerves abolishes the responses.

26
Fig.18. Newborn puppy, anaesthetised, breathing through a tracheotomy cannula. At the arrow, a steady airflow is delivered in the expiratory direction through the larynx and upper airways. Spontaneous breathing ceases, and remains inhibited for some time after the removal of the stimulus. Time bar, 10 sec. Since in newborns a major decrease in breathing rate and apnea [Fig.18] during laryngeal stimulation are not an uncommon reflex response, laryngeal stimuli are under scrutiny as possibly involved in the pathophysiology of the Sudden Infant Death. Many of these studies, by necessity, have been conducted on anaesthetised preparations, which could substantially modify the intensity, and often even the quality of the reflex responses. For example, it is known that the arousal response, with the switch from sleep to awake state, is an important mechanism to stimulate breathing, but it cannot operate under anaesthesia. Nevertheless, the information available would indicate the existence of neural circuitries which, under a special set of circumstances, can lead to cessation of breathing. Undoubtedly, apnea in response to a foreign or noxious stimulus is a protective response, but its maladaptive aspects consist of not being coupled to effective defensive expulsory manoeuvres, and in the possibility of overriding the inspiratory drive, with potential fatal effects. When foreign materials enter the upper airways, the general strategy is to avoid further penetration toward the lower airways, by controlling the breathing pattern and airways resistance, and by triggering reflexes oriented to expel the material (cough or sneeze) or swallow it. These two sets of responses are referred to as ‘defensive’ and ‘protective’, although the distinction is somewhat arbitrary, and the occurrence of one can be dependent upon the other one. Cough is considered a defensive mechanisms in the sense that it is oriented to eliminate the cause of the disorder. It includes a deep inspiration followed by a forced expiratory manoeuver, at first, against partial or total closure of the glottis, during which airways pressure can reach very high values. Differently from sneezing, but in keeping with other aspects of ventilatory control, cough can be both an automatic (involuntary) and a voluntary act. When involuntary, it results from the stimulation of any of several groups of receptors, all along the tracheobronchial tree. Although originally thought to be triggered by receptors with rapidly adapting properties

27
(including the vagal RAR) it is now believed that effective cough requires the activation of several sets of receptors, including the pulmonary SAR, which would help in the activation of the expiratory muscles. In the fetal period, airways are constantly filled by liquid (fetal and amniotic), which does not evoke any defense response. After birth, It is likely that the newborn retains some of the fetal characteristics, in the transition toward the adult pattern. Indeed, in newborns, cough is a defensive mechanism not as prompt and effective as it is in adults. Sneezing is also a defence mechanism, aiming to clear nose and nasopharynx. Expiratory pressure is generated against the pharynx, which partially closes in the first phase of the forced expiration, in contrast to coughing in which positive airways pressure is generated against the closed larynx. The expiratory duration of sneezing is shorter than in coughing, possibly because in the latter the SAR are also activated. 5. Ventilatory reflexes of cardiac or visceral origin Many stimuli applied to the skin or abdominal viscera can modify the breathing pattern. They could be either excitatory or inhibitory, and in some cases both, depending on the intensity and the conditions. It is thought that some of the visceral stimuli could play a role in the intraining between locomotion (e.g. walking or running) and breathing. The physiological meaning and interpretation of the reflex responses, simple in some instances, can be elusive or very complex in others. For example, the paralysis of the peri-esophageal diaphragmatic fibers by esophageal distention is an obvious necessity for the passage of the bolus. On the other hand, the consistent inhibition of breathing by distention of the urinary bladder or stimulation of pelvic nerves, is more difficult to interpret within a physiological context. Indeed, in some cases, the observed responses could represent only a small fraction of a group or sequence of responses the integrity of which is hard to monitor and appreciate. Equally, the response could be the result of a web of interplaying reflexes. As an example, consider the ventilatory effects of changes in blood pressure. An increase in blood pressure, even of moderate degree, decreases ventilation. The opposite is also true, i.e. hypotension stimulates ventilation. However, only the change in tidal volume is strictly a baro-reflex. The effects on breathing timing result from the vagal compensation to the changes in lung volume, via the SAR. Within a few breaths, the hypoxia and hypercapnia of the hypertensive-hypoventilation stimulate breathing, tending to buffer the hypotensive hyperpnea. Also, because hypertension can increase venous return to the heart, this is by itself the source of another reflex of cardiac origin (Bainbridge reflex) which stimulates ventilation. In brief, it can be appreciated that the analysis of what may seem a simple ventilatory response to changes in blood pressure, in reality involves the complex interplay of at least circulatory, vagal and chemo-reflexes.
PERIPHERAL CONTROL OF THE SPIROGRAM: EXPIRATION Expiration during resting breathing is driven by the elastic pressure accumulated by the respiratory system during inspiration (Pel,rs). The expiratory flow (Vexp) generated depends on the expiratory resistance of the respiratory system Rrs Pel,rs = Vexp · Rrs If Rrs were constant, V and volume during expiration should change according to exponential functions. Indeed, in a paralyzed subject, the deflation following mechanical inflation follows an approximately exponential profile. During spontaneous breathing, however, the flow is consistently less than expected from the passive characteristics of the respiratory system, and therefore expiration takes a longer time than passive deflation. In other words, during the expiratory phase of resting breathing the respiratory system behaves as if it had higher

28
resistance than in passive conditions. This pattern results mostly from the combination of two mechanisms, the post-inspiratory activity of the inspiratory muscles and upper airways control of expiratory flow (cf. ‘Mechanical aspects of breathing, Fig.16'). What determines the end of inspiration is the equilibrium between the inspiratory muscle pressure and the recoil pressure of the respiratory system. If the inspiratory muscles were active, but the inflation pressure by them generated was less than recoil, expiration begins. This is indeed what happens in the first part of expiration; in fact, a substantial portion of expiration is accompanied by some inspiratory activity, which acts as a brake to passive expiration. In some cases of resting breathing this activity has been estimated to last for up to 80% of the expiratory flow. The upper airways represent a resistive load to breathing, both in inspiration and in expiration. However, their resistance is not constant, being modulated mostly by laryngeal muscles controlling the opening of the glottis. In inspiration abductory muscles (e.g. the posterior crico-arytenoid) move the vocal folds apart, widening the glottis and lowering its resistance. In expiration, on the contrary, adductory muscles (e.g. the thyro-arytenoid) narrow the glottis opening [Fig.19], increasing the resistance. This reflex control, the efferent limb of which is via the recurrent and superior laryngeal nerves, seems to be related to lung volume, and influenced by vagal information, probably via activation of the SAR.
Fig.19. Left : records of diaphragm (top) and thyro-arytenoid (middle) electromyogram and laryngeal resistance (increase in the upward direction) during spontaneous breathing in a sedated lamb. Time mark 5 sec. (Redrawn and modified from Harding, Sleep 3:307-322, 1980). Right: From top to bottom, records of tidal volume, diaphragm and thyro-arytenoid electromyograms in a spontaneously breathing newborn lamb. Note the contraction of the TA when lung volume is maintained increased during expiration. (Redrawn and modified from Andrews et al., "The Physiological Development of the Fetus and the Newborn, Academic Press, p.821-825, 1985)

29
The purpose of these fine mechanisms controlling expiratory flow is not totally clear. In the adult, they could be vestigial forms of a control which is much more pronounced and important in fetal and neonatal life. In fact, in the fetus laryngeal resistance contributes to the control of the fetal pulmonary fluid, which is essential for lung development. In the newborn, post-inspiratory muscle activity and laryngeal resistance decrease expiratory flow in such a way that expiration is not completed by the time the next inspiration begins. In other words, by effectively increasing the expiratory resistance, the newborn's end-expiratory level (FRC) is maintained above the volume that would be attained without expiratory control. This could be desirable in newborns, since the high chest wall compliance would lead to a small FRC (cfr 'Respiratory Mechanics, Fig.7'). Furthermore, the internal positive end-expiratory pressure could be an additional factor promoting fluid reabsorption in the first hours after birth. This probably explains why the most marked manifestation of this phenomenon, i.e. brief periods of complete glottis closure during expiration, is particularly pronounced in the first hours after birth [Fig.20 and Fig.21], and can be retained longer in infants with respiratory distress.
Fig.20. Human infant, 10 min after birth. Records of respiratory airflow and tidal volume (VT, inspiration upward). Note the interruptions of expiratory flow during expiration, with volume maintained above the end-expiratory level