restoration ecology (the new frontier) || restoration of freshwater lakes
TRANSCRIPT

Restoration Ecology: The New Frontier, Second Edition. Edited by Jelte van Andel, James Aronson.© 2012 Blackwell Publishing Ltd. Published 2012 by Blackwell Publishing Ltd.
233
Chapter 18
Restoration of Freshwater Lakes Ramesh D. Gulati , L. Miguel Dionisio Pires and Ellen van Donk

234 Restoration ecology
improve lake conditions designated for human use and health such as recreation, fi shing, water supply, nature conservation and biodiversity protection. As is done throughout this book, we use the term ‘ restoration ’ in a general sense, while recognizing that most lake res-toration projects aim primarily at improving the most important functional attributes rather than a hypo-thetical return to some kind of pristine ecological con-dition. In practice, most such attempts focus primarily on eliminating undesirable consequences of human - induced disturbances.
Restoration is steadily becoming a critical part of national and international efforts to improve both water quality and the ecology of freshwater ecosys-tems (NRC 1992 ), especially in western Europe, the United States and Canada (Cooke et al . 2005 ). Lake restoration work in the United States and western and northern Europe started in the early 1970s (e.g. Bjork 1972 ). By 1975, the US Environmental Protection Agency had initiated the Clean Lakes Program by amending the Federal Pollution Control Act. Subse-quently, more federal funds were provided to clean more than 300 lakes in 47 federal states. Restoration of acidifi ed lakes by liming, as in the north - eastern United States, became a relatively common practice in Scandinavia. In Europe, the Water Framework Direc-tive (WFD) was established in 2000 (European Union 2000 ). It requires that in European member states all inland and coastal waters within defi ned river basin districts achieve at least a ‘ good ecological status ’ by 2015. Moreover, an important aim of the WFD is to initiate water pollution control by reducing emissions of harmful substances into waters (see papers in Solheim & Gulati 2008 ).
Since 1975, some 25 lake restoration measures and techniques have been developed worldwide and tested for their effectiveness, cost per unit area, required fre-quency of employment, range of applicability, etc. (see in Cooke et al . 2005 ). Since the early 1980s, lake biomanipulation as a tool for lake restoration has often been used, both in Europe and in North America (see section 18.3.2 ). Lake restoration is now one of the key topics of water management (Gulati & van Donk 2002 ; Nienhuis & Gulati 2002 ).
In this chapter, we present an overview of published results of ongoing studies of aquatic ecosystem resto-ration and rehabilitation projects of the past four decades, especially those in north - western Europe and North America. Particular emphasis is paid to shallow lakes, which are particularly susceptible to eutrophica-
18.1 INTRODUCTION
The development of modern society, including the demographic explosion and intense industrial and urban developments of the past 150 years, but espe-cially of the last half a century, has caused inland surface waters to become heavily enriched by agricul-tural fertilizers and toxic substances. The human - made alterations to freshwater aquatic ecosystems world-wide – not only lakes but also reservoirs, various kinds of wetlands and rivers of all sizes – have been severe and destructive. Despite recent attempts to restore some lakes and other bodies of freshwater, these eco-systems are arguably among the most damaged and vulnerable of all (Schindler 2006 , among others). In the western world, lakes and reservoirs are recrea-tional sites and tourist attractions (for swimming, boating, fi shing etc.). In addition, they are major sources of water for drinking, irrigation, industry and transportation, and sink areas for fl oodwater storage. Whereas lakes and lagoons also act as fi nal sinks for many of the waste products of human activity, rivers carry human and animal wastes, and other wastewa-ter effl uents (urban, industrial and agricultural), down to the sea. For better or worse, this is a service that society demands of lakes, lagoons, wetlands and rivers. Increasing demand for fresh water has also led to the creation of storage reservoirs in the fl oodplains of many river systems (Moss 1998 ). All too often, conven-tional management plans for these aquatic ecosystems ignore their regional watershed context, their hydrol-ogy, and the socio - economic systems of which they are part.
For sustained and long - term use, many aquatic eco-systems should be not only protected by abatement of pollution, but also restored or rehabilitated. Several restoration studies have been carried out in the United States and Europe on how to facilitate or accelerate the return of disturbed wetland ecosystems to functional-ity similar to the conditions prevailing prior to major disturbance (e.g. National Research Council (NRC) 1992 ; Cooke et al . 2005 ). The goal of ecosystem res-toration is to emulate or reconstitute a more or less natural and self - regulating system that is integrated within its ecological landscape (SER 2004 ; see also Chapter 2 ). In practice, however, what is often called ‘ lake restoration ’ is in fact rehabilitation . It tends to focus on improvements in water quality (using para-meters such as water clarity, dissolved oxygen condi-tions and the amount and species of algae present) to

Restoration of freshwater lakes 235
diffuse agricultural runoff from catchment areas into lakes. Accordingly, the rates of algal production and nutrient accumulation in lake sediments rise and the effects of eutrophication will persist and accumulate over time (Rast & Thornton 1996 ).
In temperate regions, increases in light and water temperature in eutrophic lakes, especially in the months of spring, generally result in increased algal growth rates and production, dominated by cyano-bacteria. Consequently, the underwater light condi-tions deteriorate. In eutrophic lakes, many genera of cyanobacteria (e.g. fi lamentous genera: Planktothrix(Oscillatoria) , Anabaena , and Aphanizomenon ; and the cosmopolitan, colony - forming species Microcystis aeru-ginosa ) gradually come to dominate because they form blooms and scums. The ability of cyanobacteria to dominate phytoplankton over a wide range of nutrient levels, as well as to fi x atmospheric nitrogen, together with their relatively low palatability to zooplankton, allows them to attain high densities and biomass. Thus, they can persist and dominate the phytoplank-ton of shallow, eutrophic lakes throughout the main growth period (spring – autumn), and even overwinter. Lastly, several species of cyanobacteria can produce potent toxins, which constitute a menace to both human and animal health (Codd 2000 ).
Rast and Holland (1988) provide a decision - making protocol for developing and implementing eutrophica-tion control measures in lakes and reservoirs (Figure 18.1 ). This scheme addresses the various concerns and cost - effectiveness to be considered when selecting fea-sible, remedial measures. A practical framework for the management and control measures of lakes and reser-voirs is to (1) assess the available information, (2) iden-tify eutrophication problems, (3) analyse costs and expected benefi ts of the alternative management strat-egies and adequacy of institutional and regulatory frameworks for implementation of a given strategy and then (4) select a control strategy and publicity meas-ures and evolve mechanisms to minimize the recur-rence of eutrophication problems.
18.2.2 Acidifi cation
Fortunately, acidifi cation problems in lakes and reser-voirs in Scandinavian countries and the United Kingdom are less grave today than they were in the late twentieth century. In the United States, however, acidi-fi cation is still an important environmental hazard in
tion. First we shall assess the major types of distur-bances of lakes, identifying the problems and main symptoms, and then discuss lake restoration strategies and techniques in use. Finally we draw some conclu-sions and discuss research and development prospects for the future.
18.2 DISTURBANCE FACTORS AFFECTING LAKES
Continuously rising inputs of nutrients, organic matter, silt and still more toxic contaminants, have led to sharp deterioration of water quality in many lakes and reservoirs over recent decades. This entails increased growth of algae and aquatic plants, and accumulation of organic matter, including detritus, leading to reduced water transparency. Water trans-parency is considered an indicator of lake and reservoir health. The most persistent and widespread water quality problem related to human use of lakes and res-ervoirs is eutrophication , but acidifi cation remains a problem in some parts of the world. Here we discuss both problems, presenting their main causes and symptoms. This will set the stage for discussing restora-tion strategies in the following section.
18.2.1 Eutrophication
Eutrophication is a natural ageing process of lakes that causes a steady increase in production of biomass due to a gradual accumulation of nutrients and a slow decrease in lake depth. In classical terms, eutrophica-tion is the enrichment of water by inorganic plant nutrients, especially N and P. Increasing eutrophica-tion in the early 1970s of the Great Lakes and many other lakes and reservoirs in the United States and Canada, along with many lakes in western Europe, raised widespread public concern. In view of P - enrichment in lakes and related problems of eutroph-ication, many state agencies funded large - scale research projects, such as the Experimental Lake Area in Winnipeg, Canada (e.g. Blanchfi eld et al . 2009 ). The eutrophication models thus developed provided insight into the quantitative relationships between nutrientloading rates and algal biomass and production (Vol-lenweider 1987 ). What emerges clearly is that the main causal factors for eutrophication are direct, point discharges of human and animal wastes, as well as
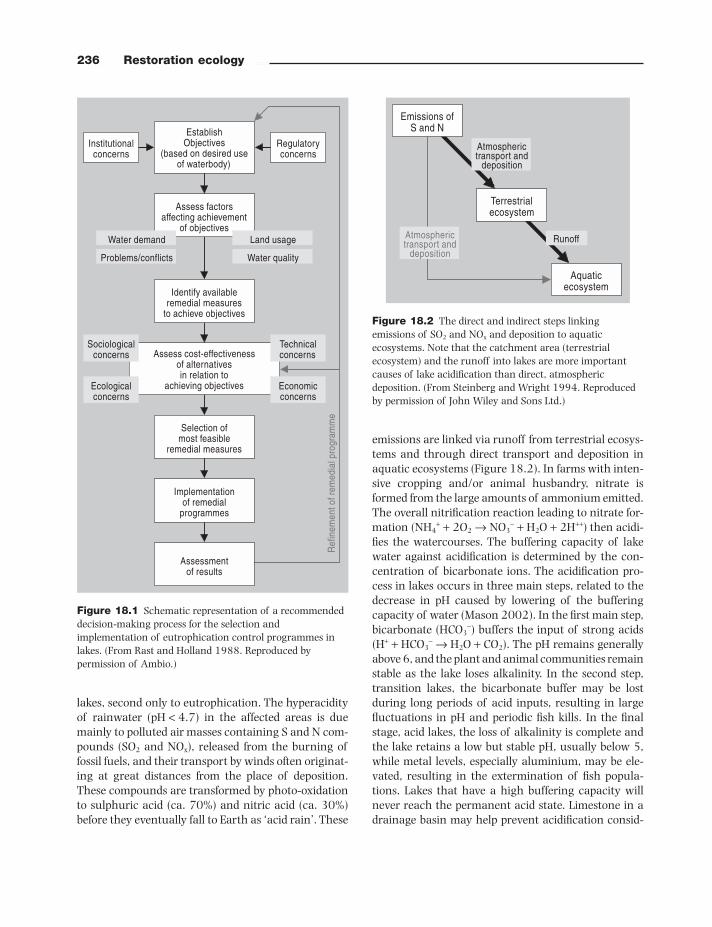
236 Restoration ecology
emissions are linked via runoff from terrestrial ecosys-tems and through direct transport and deposition in aquatic ecosystems (Figure 18.2 ). In farms with inten-sive cropping and/or animal husbandry, nitrate is formed from the large amounts of ammonium emitted. The overall nitrifi cation reaction leading to nitrate for-mation (NH 4+ + 2O 2 → NO 3− + H 2 O + 2H ++ ) then acidi-fi es the watercourses. The buffering capacity of lake water against acidifi cation is determined by the con-centration of bicarbonate ions. The acidifi cation pro-cess in lakes occurs in three main steps, related to the decrease in pH caused by lowering of the buffering capacity of water (Mason 2002 ). In the fi rst main step, bicarbonate (HCO 3− ) buffers the input of strong acids (H+ + HCO 3− → H 2 O + CO 2 ). The pH remains generally above 6, and the plant and animal communities remain stable as the lake loses alkalinity. In the second step, transition lakes, the bicarbonate buffer may be lost during long periods of acid inputs, resulting in large fl uctuations in pH and periodic fi sh kills. In the fi nal stage, acid lakes, the loss of alkalinity is complete and the lake retains a low but stable pH, usually below 5, while metal levels, especially aluminium, may be ele-vated, resulting in the extermination of fi sh popula-tions. Lakes that have a high buffering capacity will never reach the permanent acid state. Limestone in a drainage basin may help prevent acidifi cation consid-
Figure 18.2 The direct and indirect steps linking emissions of SO 2 and NO x and deposition to aquatic ecosystems. Note that the catchment area (terrestrial ecosystem) and the runoff into lakes are more important causes of lake acidifi cation than direct, atmospheric deposition. (From Steinberg and Wright 1994 . Reproduced by permission of John Wiley and Sons Ltd.)
Emissions ofS and N
Atmospherictransport and
deposition
Runoff
Terrestrialecosystem
Aquaticecosystem
Atmospherictransport and
deposition
Figure 18.1 Schematic representation of a recommended decision - making process for the selection and implementation of eutrophication control programmes in lakes. (From Rast and Holland 1988 . Reproduced by permission of Ambio.)
EstablishObjectives
(based on desired useof waterbody)
Assess factorsaffecting achievement
of objectives
Institutionalconcerns
Water demand
Problems/conflicts
Land usage
Water quality
Regulatoryconcerns
Identify availableremedial measures
to achieve objectives
Assess cost-effectivenessof alternativesin relation to
achieving objectives
Selection ofmost feasible
remedial measures
Implementationof remedial
programmes
Sociologicalconcerns
Technicalconcerns
Ecologicalconcerns
Economicconcerns
Ref
inem
ent o
f rem
edia
l pro
gram
me
Assessmentof results
lakes, second only to eutrophication. The hyperacidity of rainwater (pH < 4.7) in the affected areas is due mainly to polluted air masses containing S and N com-pounds (SO 2 and NO x ), released from the burning of fossil fuels, and their transport by winds often originat-ing at great distances from the place of deposition. These compounds are transformed by photo - oxidation to sulphuric acid (ca. 70%) and nitric acid (ca. 30%) before they eventually fall to Earth as ‘ acid rain ’ . These

Restoration of freshwater lakes 237
that is, the nature of water use, and public awareness of the problems on the one hand, and funding, scien-tifi c knowledge and desirable restoration measures on the other (Figure 18.3 ). Most lake restoration tech-niques are directed at reducing external P inputs (e.g. papers in Ryding & Rast 1989 ; Cooke et al . 2005 ), and they can be divided into two main types, namely, exter-nal and in - lake control measures (Figure 18.4 ). Exter-nal measures focus on diverting sewage and wastewater inputs intended to prevent nutrient - rich inputs into the lake or reservoir (section 18.3.1 ). The in - lake measures are designed to decrease internal P loading (P released from retention in lake sediments) by various physico - chemical means, biomanipulation of the lake ’ s food web structure and functioning, or both (section 18.3.2 ). In section 18.3.3 we refer briefl y to a few other science - based measures of lake rehabilitation.
18.3.1 Reduction of external nutrient loads
During the 1970s – 1980s, both in western Europe and in North America, tertiary treatment of sewage and
Figure 18.3 Strategic principles of lake restoration. The choice of restoration measures will depend on both direct and indirect factors related to the nature of water use, problems relating to water quality, and scientifi c studies (thick arrows). (From Gulati 1989 ; see also Vollenweider 1987 .)
Social aspects Technological inputs
Nature of water use(aquatic ecosystems)
RESTORATIONMEASURES
Problems Scientific studies
Public awareness Funding
Politics and economy
erably; regions with a calcareous geology are not sensi-tive to acidifi cation.
Acidifi cation causes marked biotic changes in com-munities of lake organisms. Shifts in the competitive relationships of the algae or the disappearance of key-stone species can alter community structure (Eriks-son et al . 1980 ). When acidifi ed, the solubility of many metals (aluminium, iron, copper, zinc, nickel, lead and cadmium) in water increases, but that of some other metals (mercury and vanadium) decreases. Adverse effects on organisms are invariably attributed to the increased solubility of aluminium and its shift to the toxic Al 3++ form, which on its increased mobilization also causes precipitation of P and humic substances. The decreased availability of PO 4 – P thus leads to a decreased phytoplankton production. Such acid lakes tend to become oligotrophic.
Ecosystem processes such as ion regulation, osmoregulation, acid – base balance, nitrogen excretion and respiration are adversely affected by an increase in acidity (Steinberg & Wright 1994 ). Great damage to fi sh populations in lakes in Scandinavia has occurred already from the 1960s to the late 1980s (Henriksen et al . 1989 ). In Norwegian lakes, Atlantic salmon (Salmo salar ) was wiped out in the early 1900s and brown trout ( Salmo trutta ) had already disappeared in mountain lakes as early as the 1920s and 1930s. In Finland, roach ( Rutilus rutilus ) disappeared from many bodies of water in the 1980s. Hardy species like white-fi sh ( Coregonus peled ) and perch ( Perca fl uviatilis ) expe-rienced high mortality of eggs. Similar harmful effects on fi sh populations are reported from eastern prov-inces of Canada. In conclusion, acidifi cation leads to periodic mortality of fi sh during the early stages of development and growth and causes populations to decrease and disappear.
18.3 LAKE RESTORATION MEASURES
Building on applied research programmes, manage-ment and restoration technologies for lakes and resevoirs developed rapidly during the 1980s in the United States, Canada and Europe, especially in the Netherlands and Denmark, but also in Germany and the United Kingdom (Moss et al . 1996 ). The restoration and management measures applied depend on users ’ interests (i.e. the economic and recreational uses of the body of water in question). Lake restoration strategies combine the social aspects and available technology,

238 Restoration ecology
and thereby improving water transparency. However, P retention in lake sediments, and its subsequent release (internal P loading), retarded the lakes ’ response to P reduction measures (Ibelings et al . 2007 ; Jeppesen et al . 2007a ; Figure 18.5 ). Thus, the observed variation in responses of lakes to P - reduction measures depends on the amount of P that is stored in the sediments.
Phosphorus from external inputs may also be removed by fi rst passing P - rich effl uents into treatment ponds, where much of the P is adsorbed on to sediments and settles there. Also, most of the P released due to lysis and death of algal blooms will be taken up by the sediment. Regular harvesting of aquatic vegetation (e.g. duckweeds, Lemna ; water fern, Azolla ; and other fl oating plants) can signifi cantly reduce P (e.g. Viess-man & Hammer 1993 ). To reduce nonpoint P loading, agricultural farming practices in a lake ’ s catchment area need to be altered radically by preventing erosion, and rationalizing the frequency, amount and timing of fertilizer applications. In addition, creation of pasture land and planting of buffer strips between farmland and watercourses may help reduce P.
The route to recovery or restoration of lakes (i.e. from eutrophication back to nutrient - poor conditions) takes
animal wastes ( > 95% P removal) was the main P - control measure. Most importantly, P - containing laundry detergents were replaced by P - free detergents in most western countries. This led to about 50% reduction of P input into some lakes in the United States. In Barton Broad, Norfolk, United Kingdom, about 90% of the annual P load in the effl uent from sewage treatment works was reduced by adding ferric sulphate (iron binds P and the Fe – P product precipi-tates to the bottom). However, because the bottom con-tained so much residual P as a result of decades of eutrophication, this P started to mobilize from the bottom into the water column blocking the hoped - for improvement in water quality (Phillips et al . 1994 ).
The response of algae to restoration measures can greatly differ among lakes: whereas some lakes improve in water quality (i.e the water transparency depth increases because of a reduction in concentration of algae), the others exhibit a small reduction of algal biomass, and still others show no reduction at all in algal biomass. In lakes in the Netherlands and Denmark, the P - reduction measures undertaken over recent decades have generally reduced concentrations of both P and chlorophyll (indicating algal biomass),
Figure 18.4 Schematic representation of factors causing an increase in nutrient loading via external inputs, which also result in an increase of internal loading, and restoration measures (white boxes) leading to lake recovery. (From Gulati 1989 .)
Industry,agriculture,recreation,
others
Biomanipulation
Humaninfluences:
Lake'smorphometry,
geology
Hydrology
Wind, light,temperature Inlake
restoration
Pointsources
Non-point
sources
Internal nutrientloading
Limnologicalcharacteristics
LAKERECOVERY
Wastewatertreatment,
sewagediversion
Externalsanitationmeasures:
(nutrientdecrease)LAKE
External nutrient inputs

Restoration of freshwater lakes 239
18.3.2 In-lake techniques for P-reduction
External P - load reductions do not guarantee improve-ments in water quality of lakes in the short term (van Liere & Gulati 1992 ), primarily because of accumu-lated P in lake sediments. In shallow lakes high stocks of benthivorous fi sh (foraging at the bottom of the lake), like large bream ( Abramis brama ), resuspend the sediments and therefore contribute to P mobilization from sediment to the water column. Apparently, this recycled P from the sediments increases if P loading in infl ows is reduced. Therefore, high P concentrations, often observed in lakes during summer, and algal pho-tosynthesis and production can go on unabated despite external control measures. Thus, our experience is that high levels of internal P loading may keep lakes in a eutrophic state for years after the reduction of external P loading.
Preventing P l oading from l ake s ediments
The cycling of P between the sediments and water is complex and poorly understood. The P release from sediment depends on both redox and pH and involves
much longer than, and differs from, the initial route of degradation and eutrophication. This discrepancy is called hysteresis (Scheffer 1998 ). For example, in Lake Veluwe (the Netherlands) macrophytes disappeared due to ongoing eutrophication at total phosphorus (TP) levels above 200 μ g L − 1 , whereas the return of macro-phytes during the lake ’ s restoration or re - oligotrophicationoccurred only at a TP concentration below 100 μ g L − 1
(Ibelings et al . 2007 ). In this case, the presence of bottom - dwelling fi sh and their interaction with wind promoted sediment resuspension causing high turbid-ity of water, which reduced the light intensity and thus prevented the return of macrophytes. Most lakes expe-riencing ongoing re - oligotrophication approach a new equilibrium (macrophyte presence, changes in algal composition and decrease in their abundance) in 10 – 15 years (Anderson et al . 2005 ; van Donk et al . 2008 ). Some researchers (Mooij et al . 2005 ) expect global climate changes will counteract the effects of nutrient reduction in lakes and delay the recovery process. Indeed, heavier rainfall may lead to higher runoff of nutrients from farmland into surface waters and higher temperatures to earlier and more release of nutrients from the sediments into the water column.
Figure 18.5 Long - term monitoring of the effects of various P reduction measures in the shallow Dutch Lake Veluwe almost immediately after the lake ’ s formation. Note the marked increase in Secchi - disc transparency in the mid - 1990s. The fl uctuations in transparency, especially the decrease during 2001, are caused by wet winter periods. (Courtesy of M.L. Meijer et al ., unpublished results.)
0
150
200
100
Chl
orop
hyll
(mg
m-3
)
2000
50
250
1960 1970 1980 19900.0
0.8
1.0
0.6
Tran
spar
ency
(m
)2000
0.4
1.2
1960 1970 1980 1990
0.0
1.5
2.0
1.0P
hosp
horu
s (g
m-2
yr-
1 )
0.5
3.0
0.0
0.3
0.4
0.2
Tota
l pho
spho
rus
(mg
l-1 )
0.1
0.5
0.2
2.5
(c)
(a)
(d)
(b)

240 Restoration ecology
removal by dredging are rare, however, because they are relatively expensive. Moreover, the sediment dred-ging eliminates the toxic and hazardous compounds and rooted aquatic plants, in addition to P removal. Nevertheless, sediment removal has three disadvan-tages: (1) during dredging, P from the sediments is released into the water column, (2) fi nding disposal sites for the extracted sediments is a major dilemma for the lake restorers and managers and (3) new underly-ing sediment layers exposed to the water could even enhance P release.
Dilution and fl ushing of the l ake with n utrient - p oor w ater
Lakes can be replenished with water from extraneous sources or from lakes with lower nutrient levels but preferably rich in Ca 2++ and HCO 3−− . Dilution as a res-toration tool, therefore, implies reducing per se the nutrient levels in lake water to limiting concentrations (i.e. fl ushing rate with diluted water exceeds the algal growth rate). Cooke et al . ( 2005 , 149 – 64) cite several examples where this technique was successfully applied in lakes in the United States and in Europe. The winter period is the best for fl ushing since both the dilution water and the lake to be diluted have less suspended material and algae than in other seasons. Because good - quality dilution water is generally scarce and expensive, due to the logistics of its transport, the tech-nique has not gained great popularity. Flushing may also be done with bicarbonate - rich water. In two lakes in the Netherlands, winter fl ushing with bicarbonate-rich (and P - poor) dilution water since 1979 resulted in a pronounced drop in P - release. The increased bicar-bonate is supposed to cause a better binding of P to the sediments through buffering and lowering of the pH (Hosper & Meijer 1986 ; Hosper 1997 ).
Lake b iomanipulation
1. The t heoretical a spects
Lake biomanipulation or food web manipulation has become routine for improving water quality of shallow lakes and reservoirs. It both has an ecological basis (Reynolds 1994 ) and complements the restoration methods involving nutrient reduction. In conjunction, these two measures can speed up lake rehabilitation. Two important hypotheses have enhanced our insights into food web relationships: (1) the size – effi ciency
bacterial decomposition of organic matter, including algal blooms. Several techniques have been applied to inactivate sediment P. Oxidation of organic matter en-hances P binding in the sediment complexes. ‘ Sealing ’ or stripping inactivates the sediment P, using the salts of Ca, Fe or Al, to precipitate all P species to sediment as a fl ock (see Cooke et al . 2005 ). The process removes up to 90 – 95% of P. In the Netherlands, Boers et al . (1992) applied FeCl 3 solution (about 5 mg Fe 3+ l − 1 d − 1 ) for P stripping by mixing the solution with lake water and surface sediments using a water jet. However, high external loading of P nullifi ed the positive effects. Although Al immobilizes P more effi ciently than Fe, it is potentially toxic and its use for P reduction in public water supplies is generally avoided. Recently some researchers have attempted to treat lake water with chemical compounds including Phoslock ® , a commer-cial bentonite product coated with lanthanum, to reduce P concentration in lake water. For example, Egemose et al . (2010) treated water from three Danish lakes with Phoslock ® and evaluated the effects on P concentration. Although this compound took several days to reduce P levels, the sediment stability increased by 265%. Moreover, this compound improved sediment consolidation and removed colouration from the water, thereby improving light penetration, which in turn improved conditions for macrophyte colonization.
Wind - induced resuspension of organic matter and nutrients and mixing with the water column can also be reduced by creating deeper pits in shallow lakes. Penning et al . (2010) carried out a model study in Lake Loosdrecht, a shallow (Z mean 1.8 m) and peaty lake in the Netherlands. They calculate the effect of creating three deep pits ( c . 12 m), covering 10% (120 ha) of the lake ’ s total surface area ( ∼ 1200 ha). The study predicts a decrease in the concentration of organic detritus by 25%, and in the light attenuation coeffi cient ( α ) from 2.5 m − 1 to 2.2 m − 1 . Together with the P load reduction measures and decrease in chlorophyll - a level, α is anticipated to decrease to 2.0 m − 1 , and to 1.6 m − 1 if combined with the deepening of the lake. Such improvements of the underwater light climate are the fi rst steps for the recovery of a submerged macrophyte community. Such studies are extremely useful in shallow, eutrophic lakes where wind - induced mixing (and mixing reinforced by benthivorous fi sh) and con-stant resuspension of nutrient - rich sediments delay the lake recovery processes (Gulati & van Donk 2002 ).
Sediment dredging leads to a reduction of the inter-nal P loading. Field studies concerning sediment

Restoration of freshwater lakes 241
clarity and promote diverse biological communities (Jeppesen & Sammalkorpi 2002 ). In shallow lakes, the development of macrophytes is the most striking side effect of these changes in the food chain. The macro-phytes compete for nutrients with phytoplankton and further limit the latter ’ s growth during spring and early summer, leading to clear water. Biomanipulation is applied as an ecological restoration method in which the strategy is to induce a drastic reduction of the fi sh stock.
For shallow lakes, application of biomanipulation is based essentially on the knowledge that top - down manipulations in the food web, especially reduction of planktivorous fi sh biomass, can lead to desirable changes at lower trophic levels. The theory relies on the hypothesis of alternative stable states : a turbid state dominated by phytoplankton blooms, and a clear state dominated by macrophytes (Scheffer et al . 1993 ; Moss 1998 ; see also Chapter 6 ). There is ample evidence to support the existence of alternative stable states in lakes (Gulati & van Donk 2002 ), but it is important to recognize that the hysteresis effect (see section 18.3.1 ) retards recovery or restoration from a turbid to a clear state. For a lake to shift from a turbid state to a clear - water state, substantial reductions of the plank-tivore and bottom - dwelling benthivore fi sh stocks are needed before macrophytes can establish and multiply.
hypothesis (Hall et al . 1976 ) and (2) the cascading trophic interactions hypothesis (Carpenter & Kitchell 1992 ). These two hypotheses, but especially the latter, eluci-date ecosystem functioning and structure, and inter-trophic feedback effects. Whereas the fi rst hypothesis enhances our insight into the fi sh – zooplankton dynamics (i.e. fi sh tend to prey on large - bodied zoo-plankton), the trophic cascade hypothesis shows how intertrophic effects cascade down the food chain. In other words:
Piscivorous fi sh ↓ Planktivorous fi sh ↓ Zooplankton ↓ Algae
In short, the lake food web is infl uenced by nutrient inputs (bottom - up control) on the one hand, and zoo-plankton grazers and predatory fi sh (top - down control) on the other (Figure 18.6 ). Intense predation by pisci-vorous fi sh will lead to a decrease in the biomass of planktivorous fi sh and an increase in that of larger zoo-plankters, especially Daphnia spp. These changes cause a reduction of phytoplankton biomass, improve water
Figure 18.6 A simplifi ed diagram of top - down and bottom - up control measures in the food web of shallow lakes. Nutrient reductions in the infl ows and major biomanipulation measures are indicated with arrows. (From Gulati and van Donk 2002 .)
PLANKTIVORES:FISH
LIGHT
ALGAE
Zooplankton
Bioturbation
Macrophytes
Piscivores
Shading
Grazing CompetitionNutrients
N, P
Uptake
Uptake
PredationPredation
S
S
Reduction,removal
Refuges
Reductionin inflows
Restocking
Inoculation?
Interference,toxicity
Limitations

242 Restoration ecology
cacy of restoration measures are also likely to vary with lake depth (see Moss 1998 ). In addition, because shallow lakes are more likely to be colonized by macro-phytes, a shift to a clear - water state in these lakes will occur earlier than in the deeper ones.
3. Lessons l earned
Gulati et al . (2008) attribute the failure of lake bioma-nipulation measures to one or more bottlenecks, including (1) ineffective biomass reduction of plank-tivorous and benthivorous fi sh and the inability to sustain fi sh at low levels, (2) inadequate reduction of external P and a concomitant increase of P release from sediments, (3) poor edibility of cyanobacteria to zooplankton, (4) reduction of macrophytes by fi sh and waterfowl and (5) recurrent failure of introduced northern pike (piscivorous fi sh). Among these, bottle-necks (1), (2) and (4) are crucial to overcome for lake biomanipulation to be successful (see also Gliwicz 2005 ). Below, we highlight the importance of selected biotic factors for applying biomanipulation as a lake restoration measure.
Importance of fi sh
In shallow lakes, it is relatively easy to manipulate fi sh (Lammens 1999 ) and produce rapid effects through fi shing on the planktivores and benthivores. After fi shing, the standing stock usually ranges from 150 kg FW ha − 1 to 20 kg FW ha − 1 (Lammens et al . 2002 ). Maintenance of a fi sh stock at low densities is desirable but generally hard to achieve. This is because there is less competition between fi sh individuals for food leading to higher growth. Furthermore, because the amount of P per unit body weight in fi sh is quite high, P recycling by fi sh seems to retard lake restoration (Sereda et al . 2008 ). Studies on Loosdrecht lakes showed that the P regenerated from fi sh was about 140% of the external P loading (van Liere & Janse 1992 ). Thus, nutrients and algal concentrations are expected to decline upon removal of planktivores. For fi sh removal to be successful, a substantial reduction of the fi sh stock must take place.
Role of m acrophytes
Macrophytes stabilize lakes after biomanipulation (Coops & Hosper 2002 ). They infl uence various proc-esses in shallow lakes (Figure 18.7 ). Firstly, macro-phytes can accumulate relatively large amounts of N
Measures to reduce these fi sh stocks need to be repeated to prevent re - establishment of their populations. An improvement in the underwater light climate (i.e. more visibility) indicates that the reduction of planktivore and benthivore fi sh is succesful and that the top - down cascading effects are re - established (Hosper 1997 ; Meijer 2000 ): phytoplankton biomass is controlled by zooplankton which in turn are not heavily predated by planktivores because piscivores control them.
2. Historical o verview
Lake biomanipulation as a restoration measure started in the early 1970s. State - of - the - art reviews are given by Gulati and van Donk (2002) and Jeppesen et al . (2007a) . From these studies, it becomes clear that in almost all biomanipulated lakes in Europe, the stand-ing stock of planktivorous fi sh was effectively reduced but Daphnia populations did not increase in the years thereafter. Both poor food quality (high cyanobacterial densities) and/or predation by remaining planktivores probably contributed to the failure of the daphnids to develop high population densities. Northern pike ( Esox lucius ), which was introduced to control the planktivo-rous fi sh, mainly bream, was rarely a success. These studies showed that the size - selective predation on larger zooplankton by the planktivorous fi sh led to a decrease of daphnids and to their grazing on phyto-plankton, so that water quality did not improve (Gulati & van Donk 2002 ). In other words, the trophic cascadebetween phytoplankton and zooplankton was largely decoupled (Kasprzak et al . 2007 ). In both Denmark and the Netherlands, the effect of fi sh removal was less obvious because lakes tended to return to the turbid state unless fi sh removal was repeated annually. Other factors that caused biomanipulation to fail were (1) insuffi cient reduction in external P loading (e.g. Benn-dorf et al . 2002 ), (2) continuation of internal P loading and (3) the failure of macrophytes to appear.
There are more cases of long - term failure than success for lake biomanipulation, mainly because the nutrient input effects on the pelagic food web tend to persist even after the top - down manipulation measures (McQueen et al . 1986 ). Thus, reduction of nutrients from the lake ’ s catchment, rather than in - lake only, is an important prerequisite for success of biomanipula-tion. In deeper lakes the failure of biomanipulation measures is also caused by high P loading (Benndorf et al . 2002 ). Because sediment – water interactions differ with depth, the nutrient dynamics and the effi -

Restoration of freshwater lakes 243
large - bodied zooplankton decreases. The decreased pre-dation effects cascade in the food web and lead to a marked increase in larger - bodied zooplankton, espe-cially daphnids, as well as their grazing pressure on algae (Potthof et al . 2008 ) leading to a clear - water phase. Clear - water conditions have been reported for several lakes in western Europe following reductions of planktivorous fi sh (e.g. Gulati 1990 ; Jeppesen et al . 1999 ) and in the United States (Schrage & Downing 2004 ). It can thus be concluded that reduction of the planktivorous fi sh stock helps to restore the large - bodied zooplankters, particularly those comprising Daphniaspecies. Such a change will lead to a reduction in algae biomass and improvement of water transparency.
Role of z ebra m ussels
The zebra mussel, Dreissena polymorpha , a bio - invader in many lakes in temperate regions, is a potential tool for lake management (Dionisio Pires et al . 2010 ). Because zebra mussels are not found everywhere in the world, the use of these bivalves is less general than other techniques, like external P reduction, which apply universally. Use of zebra mussel in lake mana-gement therefore only applies to temperate regions. Dionisio Pires et al . (2004) demonstrated that adult mussels can act as biofi lters: the mussels clear the lake from cyanobacteria at much higher rates than other particles in the water (Figure 18.8 ). In some shallow
and P from both water and sediments (Barko & James 1998 ), reducing bioavailability of these elements for algae (van Donk & van de Bund 2002 ). Moreover, the algae (periphyton) adhering to macrophytes act as major nutrient sinks. Secondly, the macrophytes provide refuge for larger bodied zooplankton and young fi sh against fi sh predation (Moss 1998 ), and thus promote zooplankton grazing. Thirdly, macro-phytes consolidate bottom sediments and reduce their resuspension by fi sh and wind. Finally, both denitrifi ca-tion in the macrophyte beds and the release of allelo-pathic substances by macrophytes are reported to adversely affect phytoplankton (Figure 18.7 ), although the mechanisms for such negative feedbacks are as yet unclear (van Donk & van de Bund 2002 ). Thus, all these characteristics related to macrophyte develop-ment and growth lead to improved underwater light climate in lakes. For macrophytes to be able to stabilize lakes after biomanipulation, it is important that they can develop into a stable population. This depends on the amount of light reaching the bottom, the presence of propagules (plant material) in the bottom, predation by waterfowl and the softness of the sediment (Scheffer 1998 ).
Role of z ooplankton
In lake restoration studies, the standing stock of plank-tivorous fi sh is always reduced so that predation on
Figure 18.7 A diagrammatic representation of the mechanisms and factors causing sediment resuspension and turbidity in shallow lakes in relation to macrophytes (submerged plants). Different feedback mechanisms and their relative intensities are indicated with arrows (Allelop. subs. = allelopathic substances). (From Gulati and van Donk 2002 .)
SEDIMENTRESUSPENSION
Feeding
Turbidity PHYTOPLANKTON
PLANKTIVORE andBENTHIVORE FISH
ZOOPLANKTON
SUBMERGEDPLANTS
Herbivory(fish, birds)
Allelop. subs.
Refuge
Waves (wind)
Nutrients

244 Restoration ecology
when emissions of SO 2 and NO x peaked. By the mid - 1980s, there was a perceptible decrease in emissions (Mason 2002 ). By the year 2000, two thirds of surface waters in Europe, where long - term monitoring (n = 56) had been carried out, showed reductions in acid deposition. Additionally, modelling studies of acid-ifi cation in Norwegian surface waters indicate that these waters will continue to recover slowly (Larssen et al . 2010 ). By 1983, almost all countries had com-mitted to reduce their SO 2 emissions by 30% within a decade, and to decrease by 70 – 80% by 2010 (United Nations 1994 ). However, contrary to these emission reductions, the NO x emissions have tended to increase, for example in the United Kingdom. Therefore, acidifi -cation related to N is still a grave problem.
Liming is the most common technique for restoring acidifi ed lakes, for example in Scandinavian countries, the United Kingdom (Scotland and Wales) and eastern Canada. Limestone, which includes calcite (CaCO 3 ) and powdered dolomite, is the most commonly used compound. Also, quicklime (CaO) and slaked lime (Ca(OH)2 ) have been used in addition to alum ((Al2 SO 4 ) 3 · 14H 2 O). Liming produces longer lasting buffering effects and pH generally does not rise above 7.0. The liming of Loch Fleet, Galloway, southwest Scotland, facilitated the introduction and sustenance of a trout population.
Both in Sweden and in Norway, large - scale liming is used as a national strategy for preserving species threatened by acidifi cation (Henrikson & Brodin 1995 ). In Sweden alone, between 7500 and 11 000 km of streams were limed annually to raise the pH to > 6.0 in order to help indigenous fauna and fl ora to survive. Nearly 90% of the acidifi ed bodies of surface water have thus been restored.
Adjusting w ater l evel fl uctuations
Shallow lakes are particularly sensitive to rapid changes in water level. Therefore, water level fl uctuations (WLF) may have an overriding effect on shallow lake ecosys-tem functioning and management (Coops & Hosper 2002 ). Water levels in shallow lakes naturally fl uctu-ate both seasonally and annually depending largely on regional climatic conditions and past and current human activities (Beklioglu et al . 2001 ). WLF can also be disastrous for submersed plant communities. Extremely high water levels in the vegetation period reduce light availability, and low water levels may damage plants due to ice and wave action during
Dutch lakes the areas colonized by zebra mussels are clearer, even to the naked eye, than the other lake parts.
In contrast to the practice in some European coun-tries, in North America, zebra mussels, and their congeners the quagga mussels ( Dreissena rostriformis bugensis ), are eradicated from the lakes they have invaded, instead of being used as biofi ltrators (Strayer 2009 ). Both the zebra mussel and the quagga mussel have caused much economic and ecological damage by settling on hard substrates where they clog pipes of the cooling systems. These mussels strongly compete with other fi lter feeders and disrupt lake food webs. Eradi-cation programmes in North America will have diffi -culties in achieving success because these mussels reproduce readily and fast.
18.3.3 Other measures
There are several other measures available for lake res-toration. In this section, we will highlight only three that are science - based, aimed at counteracting acidifi -cation, excessive water level fl uctuations and oxygen - poor conditions, respectively.
Restoration of a cidifi ed s ystems
Actions to reduce acidifying emissions in Europe, the United States and Canada began in the mid - 1970s,
Figure 18.8 Clearance rates of adult zebra mussels (Dreissena ) on different food particles from Lake Zwemlust, the Netherlands. The clearance rates on cyanobacteria are signifi cantly higher than those on other phytoplankton and detritus as indicated by the letters a and b above the bars. (From Dionisio Pires et al . 2004 . Reproduced by permission of Blackwell Publishing.)
0
25
Cle
aran
ce r
ate
(ml m
g of
dry
wei
ght o
f D
reis
sena
-1 h
-1)
Cyanobacteria
20
10
5
15
Other phyto-plankton
Detritus
a
bb

Restoration of freshwater lakes 245
tions in the upper mixed layers, so that the dominant phytoplankton shifts to noncyanobacterial forms. In the Netherlands, artifi cial mixing is used to disrupt stratifi cation of the water column (i.e. a water column that is divided into an upper and a lower layer due to the increase in spring and summer temperature) to prevent bloom formation by cyanobacteria (Huisman et al . 2004 ). Thus, mixing or circulation, unlike nutri-ent reduction, produces instantaneous improvements in water quality, and without decreasing the nutrient concentrations or loading. However, the technique seems to be more effective for lakes in which stratifi ca-tion of the water column occurs.
18.4 PERSPECTIVES
Although in many European countries freshwater lakes and reservoirs constitute only a small fraction of the land area, their importance for human health, recreation and national economies is considerable. There is obviously an urgent need to further curtail, divert and treat the unwarranted inputs into the lakes and reservoirs of nutrients, organics, silt and contami-nants from the watershed. Despite more than a quarter century of nutrient reduction measures in runoff waters, many lakes still have poor water quality. The principal cause for the delay of responses to corrective measures is the large stockpiles of P in lake sediments. The slow release of this element allows eutrophication symptoms to persist and even proliferate. Secondly, inadequate nutrient reductions in the runoff waters and the virtually unabated and diffuse inputs from the agricultural grounds in the lake catchment areas act as a major bottleneck to lake restoration. It is, there-fore, diffi cult to predict the response of an ecosystem under restoration. Thus, further manipulations of both chemical and biological processes are needed to sustain the positive effects of the corrective measures. However, monitoring of water quality and restoration of the in - lake processes alone will be a futile exercise if not carried out in a watershed context. This latter involves documenting the entire landscape setting, including habitat type, hydrological regime, soil properties, topography and invasive species, which all can hamper restoration measures. Moreover, chances of improving a lake are often better if the chosen measures comple-ment each other.
It is diffi cult to draw any conclusions on the effects of climate change on lake restoration. At present, we
winter and to desiccation during summer. The plans by water managers in the last 20 years, to allow more dynamic water levels in set - aside nature areas, espe-cially, entail extending the upper and lower limits of the permissible annual water level fl uctuations (Coops & Hosper 2002 ). Near - natural water levels that allow wider fl uctuations than the current ‘ fi xed ’ levels are considered the best option by most water managers.
A good case study on WLF management is that of Lake Okeechobee (Florida, United States, headwaters of the Everglades). As a subtropical lake ecosystem, it exhibits large seasonal and interannual variation in rainfall and high evapotranspiration. This can result in large and unpredictable variations in water level that can affect ecosystem dynamics. In response to this, as well as to other environmental concerns, a 30 - year plan was initiated to construct structures to restore the quantity, quality, timing and distribution of water (Havens 2002 ). This overall programme is called Com-prehensive Everglades Restoration Program (CERP). CERP is expected to reduce the occurrence of damaging high and low WLF and to increase the occurrence of spring water level recessions that benefi t native biota. Johnson et al . (2007) studied the response to hydrologi-cal variations in Lake Okeechobee of both vegetation and fi sheries. Their fi ndings indicate that lakes with a high water level and prolonged fl ooding are likely to show substantial losses of aquatic and wetland plants as well as a negative impact on fi sh populations, as is the case at Lake Okeechobee. In contrast, shallower lakes with moderate water level variations were reported to have widespread stands of emergent and submerged aquatic vegetation in most years. Conse-quently, habitat structure was improved and food resources increased for largemouth bass ( Micropterus salmoides ) and other fi sh species, alligators, wading birds and snail kites ( Rostrhamus sociabilis ).
Mixing of the w ater c olumn
Oxygen - poor water conditions can be counteracted by artifi cial mixing, which involves aerating or oxygenat-ing lakes using pumps, jets and bubbling air at the lake bottom. The technique has been used in the United States to prevent fi sh dying during ice cover (Cooke et al . 2005 ). Aeration oxidizes substances in the water column, allows removal of Fe and Mn from the water into the sediments more rapidly and reduces internal P loading from sediment. Aeration seems to prevent cyanobacteria from exploiting the optimal light condi-

246 Restoration ecology
with increasing temperature, these changes will lead to greater predation pressure of fi sh on zooplankton and macro - invertebrates. Such a scenario seems to be especially valid for temperate lakes where temperature rises are perhaps relatively easier to predict. Successful biomanipulation will therefore probably be more diffi -cult under a changing climate. Finally, the predicted climate changes expected to intensify the negative impact of eutrophication on water quality in temper-ate waters (Mooij et al . 2005 ). Figure 18.9 illustrates the different restoration measures as discussed in the text, and shows which of these measures are negatively impacted by climate change. Regarding tropical waters, we do not have at present the basic information, espe-cially on biological aspects of lake restoration, let alone predicting the climate change effects for these waters.
Lake restoration plans nowadays typically envisage near - natural restoration , that is, allowing nature to take its course, and emphasizing that a lake is an integral part of a landscape comprising other aquatic, semi - aquatic and terrestrial ecosystems (see papers in Nien-huis & Gulati 2002 ) and taking climate change into
know little about long - term climate change and its consequences for hydrology and water management practices (e.g. the impact of fl ooding or droughts, or both, as well as of human water use on ecosystems). However, several model studies predict that warming will act as a catalyst for higher growth of cyanobacte-ria in lakes at higher temperatures than other phyto-plankters (Paerl & Huisman 2008 ). Accor ding to Jeppesen et al . (2007b) , global warming is expected to intensify eutrophication and its symptoms, via nutri-ent loading, increased rates of evaporation losses and changes in water levels. For example, the fi sh commu-nity structure could be affected by increases in tem-perature, eutrophication and salinity. Both the life history traits and the feeding behaviour of many organisms are likely to be affected: changes in life history of many fi sh species (like a shorter body size and life span) will lead to changes in fi sh community compositions in lakes. As a result of this, less predators will be present which will benefi t herbivores, plankti-vores and omnivores and, hence, a disruption of the food chain may take place (see section 18.3.2 ). Together
Figure 18.9 Diagrammatic depiction of different restoration methods applied to lakes, illustrating also the predicted effects of climate change on lake restoration measures. The possible restoration measures (as discussed in the text) are shown in boxes and numbered. The circle on the right side of the fi gure shows how biomanipulation operates: it leads to reduction of planktivorous fi sh (PL) and increase of piscivorous fi sh (PI). See text for its effects on the food web. Climate changes are likely to negate effects of some of the restoration measures, especially nutrient reduction (increase in both nutrient release from thebottom and external nutrient loading) and on the food web (positive effects on planktivore fi sh and inedible phytoplankton and negative effects on piscivore fi sh in temperate waters), hence hampering biomanipulation efforts. These negative effects of the climate change are portrayed as the three unhappy smileys.

Restoration of freshwater lakes 247
Sustainability of the positive effects on water quality is central to the remedial measures. The experi-ence gained from the failures and occasional successes of the last two decades should make it possible to develop more enduring strategies for a more sustaina-ble restoration of our lake ecosystems. Lastly, the long - term aims of water management should be to create and maintain sustainable and healthy aquatic envi-ronments that possess optimal properties for their assigned functions.
ACKNOWLEDGEMENTS
The authors thank Professor Emeritus Rob Hart from the University of KwaZulu - Natal, South Africa, and Dr Harry Hosper from the Centre for Water Management (RWS Waterdienst, the Netherlands), for their com-ments on a previous version of this chapter.
account. First, such a measure includes reinforcing the shoreline vegetation of lakes to prevent wind - and wave - induced erosion and improving the propensity of the land – water transition to develop a natural biodi-versity. Secondly, excavating deeper pits within the shallower lake parts offers a viable control measure to isolate and bury the organic matter shifted by wind to these pits. The burial of the nutrient - rich lake sedi-ments will retard in - lake nutrient release rates from the sediments. Thirdly, more exploratory work is needed for creating artifi cial islands in small and shallow lakes. Such an island within a lake, facing the commonly prevailing winds, will reduce the wind fetch factor and slow down shore - line erosion on the leeward side of the island. Fourthly, feasibility plans are also under way to deploy water level management, to encourage the shoreline macrovegetation and develop greater natural evolution of the aquatic and semi - aquatic ecosystems.