restriction fragment length polymorphisms and sequence
TRANSCRIPT

Vol. 59, No. 5INFECTION AND IMMUNITY, May 1991, p. 1803-18100019-9567/91/051803-08$02.00/0Copyright C 1991, American Society for Microbiology
Restriction Fragment Length Polymorphisms and SequenceVariation within the spaP Gene of Streptococcus mutans
Serotype c IsolatesL. J. BRADY,' P. J. CROWLEY,' J. K.-C. MA,2 C. KELLY,2 S. F. LEE,' T. LEHNER,2 AND A. S. BLEIWEISl*
Department of Oral Biology, University of Florida, Gainesville, Florida 32610,1 and Department ofImmunology, UnitedMedical and Dental Schools of Guy's and St. Thomas's Hospitals, London Bridge, London SE] 9RT, England2
Received 31 October 1990/Accepted 8 February 1991
A restriction fragment length polymorphism study was undertaken to determine the extent and location ofheterogeneity within spaP encoding the Mr 185,000 cell surface protein P1 (antigen I/II) of Streptococcus mutansserotype c isolates. The gene was found to be highly conserved except for a central variable (V) region predictedto encode less than 150 amino acids. Sequence analysis identified two V-region variants. These differences wereindependent of the geographic source of the isolates. Southern analysis using synthetic oligonucleotide probesindicated that nonretention of P1 (I/II) by some isolates is not due to a deletion of the 3'-terminal DNA necessary
to encode an intact carboxy terminus.
Streptococcus mutans has been implicated as a causativeagent of human dental caries (14). Serotype c organismsexpress a protein of approximate Mr 185,000, first identifiedas antigen I/MI (36, 45) and also referred to as antigen B (37,38), IF (18), PAc (33, 34, 43), MSL-1 (7), and P1 (11).Analogous proteins have also been described for Streptococ-cus sobrinus serotype d and g strains (1, 16, 32), S. mutansserotype f strains (2), and Streptococcus sanguis strains (6).These proteins are designated SpaA (or PAg), SR, andSSP-5, respectively. The cell surface protein P1 (I/II) isassociated with a fuzzy surface layer surrounding the cellwall in strains designated "retainers" or released into theculture supernatant in strains designated "nonretainers" (3).P1 has been shown to mediate the binding of retainer strainsof S. mutans to salivary agglutinin-coated hydroxylapatitebeads (25, 27) and to increase the hydrophobicity of suchstrains (24, 25, 27, 31). At present, P1 (I/II) is believed tofunction as an adhesin enabling S. mutans to bind to thesalivary pellicle of tooth surfaces or to other plaque micro-organisms (8, 12, 27, 29).The gene encoding P1 (I/II) has been cloned (26, 33) and
sequenced independently by two research groups (21, 34),and they designated it spaP and pac, respectively. Compar-ison of the sequences derived from the two strains of S.mutans serotype c, NG5 and MT8148, reveals a high degreeof homology, although some differences can be noted.Amino acid sequences deduced from spaP and pac indicatethat these genes encode polypeptides of 1,561 [P1 (I/II)] and1,565 (PAc) amino acids with predicted Mrs (following thecleavage of signal peptides) of 166,159 and 166,817, respec-tively. Differences between predicted Mrs and those mea-sured by sodium dodecyl sulfate (SDS)-polyacrylamide gelelectrophoresis have been attributed to an abundance ofproline residues. This has been proposed to cause aberrantgel migration of group A streptococcal M6 protein (15) andgroup G streptococcal protein G (13). P1 (1/11) and PAc sharethe following features: a 38-residue N-terminal signal pep-tide, an alanine-rich (A) region in the amino-terminal third ofthe molecule comprising three 82-residue tandem repeats
* Corresponding author.
(residues 219 to 464 of P1, with residues 186 to 218 repre-senting a fourth partial degenerate repeat), a proline-rich (P)region located in the central portion of the molecule com-prising three 39-residue tandem repeats (residues 847 to 963of P1) preceded by a 7-residue partial repeat sequence, aC-terminal region rich in proline residues believed to spanthe cell wall, and a potential transmembrane domain consist-ing primarily of hydrophobic amino acids followed by acharged cytoplasmic tail. The C-terminal molecular architec-ture is similar to that observed for other streptococcal wallproteins, most notably group A streptococcal M protein (9).The alanine-rich repeats of the A region demonstrate a7-residue periodicity predicted to result in an a-helical coiledcoil structure (22), a feature also observed with M protein (9)and pneumococcal PspA (44a). A schematic diagram of spaPis shown in Fig. 1.The spaP and pac genes are highly conserved, with most
nucleotide substitutions resulting in alternative codon usageor conservative amino acid substitutions (21). There are atotal of 36 predicted single amino acid substitutions betweenP1 (I/Il) and PAc; however, 20 of these substitutions areclustered within a variable segment (V region) of less than150 amino acids [residues 679 to 823 of P1 (I/II) and 679 to827 of PAc] immediately upstream of the P-region repeats(Fig. 1). A single residue deletion in PAc [corresponding toresidue 752 of P1 (I/Il)] and the replacement of 9 P1 (I/Il)residues (796 to 804) with 14 nonhomologous PAc residues(795 to 808) occur within the V region as well. However,only two amino acid substitutions occur in the A region andonly one occurs in the P region.To examine the extent and location of heterogeneity
within the spaP gene at the molecular level among variousisolates of serotype c S. mutans, a restriction fragmentlength polymorphism (RFLP) study of the entire gene wasundertaken. Specific probes were generated by using clonedspaP DNA. Of particular interest was the V region, in whichmost of the differences between spaP and pac are clustered.Sequence analysis of this part of the gene from several S.mutans isolates was performed to determine whether theywere spaP-like or pac-like or possessed unique V-regionsequences. In addition, oligonucleotide probes correspond-ing to the 3' terminus of the gene were used to determine
1803
on March 20, 2018 by guest
http://iai.asm.org/
Dow
nloaded from

1804 BRADY ET AL.
5'
(1)HINDIII(1538)
1 2 3 1 1 2 3M rT.:.-}-.-.T-.7-.-3
HINDIII1(4019)
3'
(4782K6mmJ% J % %,%-%L' % %I% % %,%4 r ¶ - i -XXI-I
SIGNAL SEQUENCE
PROMOTERREGION
\A - REGION/(Alanine-rich)(655-1491)
(a.a.186-464)
V-REGIO1 b-REGI N(Variable) (Proline-rich)
(2134-2512) (2617-2988)(a.a.679-823) (a.a.840-963)
WALL MEMBRANESPANNING SPANNINGREGION REGION(Proline-nch) (4705-4766)(4555-4704) (a.a. 1536-1556)(a.a.1486-1535)
CYrOPLASMICTAIL(4767-4782)
(a.a.1557-1561)
FIG. 1. Schematic representation of spaP based on the sequence derived from S. mutans serotype c isolate NG5 (21). Bold numbers inparentheses denote nucleic acid positions, whereas numbers following "a.a." denote amino acid residues.
whether P1 (I/II)-nonretaining isolates of S. mutans lack theDNA necessary to encode an intact carboxy terminus.
MATERIALS AND METHODSBacterial strains and culture conditions. S. mutans sero-
type c isolates used in this study were as follows: Ingbritt175, Ingbritt 162, NG8, NG7, and NG5 from K. Knox,Institute of Dental Research, Sydney, Australia; CCY1,CCY2, Ingbritt, Guy's c, and K2 from T. Lehner, UnitedMedical and Dental Schools of Guy's and St. Thomas'sHospitals, London, England; GS5 from R. Gibbons, ForsythDental Center, Boston, Massachusetts; and DP5 and DP6,fresh isolates from our laboratory. All streptococcal isolateswere grown aerobically for 16 h at 37°C in Todd-Hewittbroth (Difco Laboratories, Detroit, Mich.) supplementedwith 0.3% yeast extract. Escherichia coli JM109 (Interna-tional Biotechnologies, Inc., New Haven, Conn.) andrecombinant strains from our laboratory were grown aero-bically at 37°C with vigorous shaking in L13 medium (1%tryptone, 0.5% yeast extract, and 1%o [wt/vol] NaCl, pH 7.0).Medium for the growth of recombinant strains was supple-mented with 20 ,ug of ampicillin per ml (wt/vol) (SigmaChemical Co., St. Louis, Mo.).
Isolation of chromosomal DNA. Streptococcal cells fromstationary-phase cultures (1 liter) were harvested by centrif-ugation at 10,000 x g (10 min, 4°C) and washed twice with 30ml of 10 mM Tris-HCl-1 mM EDTA buffer, pH 7.5 (TE).Cell pellets (-1.5 g wet weight) were suspended in 5 ml ofTE and incubated at 65°C for 20 min. Five milliliters of TEcontaining 40 mg of lysozyme per ml (wt/vol) and 200 U ofmutanolysin (Sigma) were added to each suspension (1 h,37°C), followed by the addition of 0.5 ml ofTE containing 10mg of predigested pronase E per ml (wt/vol) (Sigma) (1 h,37°C). Suspensions were diluted to 15 ml with TE, and 5 mlof 20% (wt/vol) SDS (Swartz/Mann Biotech, Cleveland,Ohio) was added to lyse the cells. Cesium chloride (Sigma)was added to each sample to a density of 1.55 ± 0.03 g/ml(-30 g), and samples were incubated (15 min, 65°C) todissolve precipitated SDS. Samples were centrifuged in38.5-ml Beckman ultraclear Quick Seal tubes in a VTi 50vertical tube rotor (175,000 x g, 24 h, room temperature).The DNA band collected from each gradient was dialyzedonce against 200 volumes of sterile 10 mM Tris-HCl-5 mMEDTA buffer, pH 8.0 (16 h, room temperature), and twiceagainst 10 mM Tris-HCl, pH 8.0 (4 h, 4°C and 16 h, 4°C),treated with 100 jtg of boiled RNase per ml (wt/vol) (Sigma)
for 1 h at 37°C and then with 100 p.g of proteinase K per ml(Sigma) for 2 h at 37°C, brought to 15 ml with TE, extractedonce with an equal volume of phenol-chloroform and oncewith an equal volume of chloroform, and precipitated with 2volumes of ice-cold 95% ethanol containing 2.5% (wt/vol)potassium acetate (30 min, -85°C). Precipitated DNA wasspooled onto sterile glass rods, rinsed with ice-cold 70%ethanol, and air dried. The DNA from each isolate wasdissolved in 1 ml of TE and quantitated by reading absorb-ance at 260 and 280 nm.
Isolation of plasmid DNA. Plasmids were isolated by themethod of Ish-Horowitz and Burke (19).
Subcloning. The gene encoding P1, spaP, was cloned fromHindIII partially digested NG5 chromosomal DNA andligated into pUC18 as previously described (26), resulting inthe generation of the recombinant plasmid pSM2949. Asubclone containing recombinant plasmid p26L10 was gen-erated by partially digesting pSM2949 with HindIII. A 5.3-kbDNA fragment containing the 2.7-kb pUC18 vector and the2.6-kb 3' half of the original spaP insert was isolated from anagarose gel by electroelution (28) and purified by using anElutip-d column (Schleicher and Schuell, Inc., Keene,N.H.). The DNA was ligated with T4 ligase (InternationalBiotechnologies, Inc., New Haven, Conn.) and transformedinto E. coli JM109 as described previously (26). A secondsubclone containing the recombinant plasmid p26R3, corre-sponding to the 5' half of the original spaP insert, wasgenerated by digesting pSM2949 to completion with HindIl,PstI, and Scal. The 2.6-kb fragment was isolated and puri-fied as described above and cloned into E. coli JM109 byusing pUC18.
Oligonucleotide synthesis. Oligonucleotides were synthe-sized with a DNA synthesizer (model 380B; Applied Biosys-tems, Foster City, Calif.).
Restriction enzymes. The following restriction enzymeswere used in this study: AlwNI, BspHI, HgiAI and DraIII(New England Biolabs, Beverly, Mass.); NheI, DraI, andEcoRV (Promega Corp., Madison, Wis.); HaeII, Ncil, ScaI,and PvuII (Bethesda Research Laboratories [I3RL], Gaith-ersburg, Md.); EaeI, BsmI, XMnI, and HaeII (Stratagene,La Jolla, Calif.); Eco47III (United States Biochemical Corp.,Cleveland, Ohio); XbaI (Pharmacia-LKB Corp., Piscat-away, N.J.); HindIII and PstI (International Biotechnolo-gies, Inc.); and Nsp(7524)I (Amersham Corp., ArlingtonHeights, Ill.). Each enzyme was used according to themanufacturer's instructions.
INFECT. IMMUN.
%1Fo/Rw .%/o.
on March 20, 2018 by guest
http://iai.asm.org/
Dow
nloaded from

spaP GENE OF S. MUTANS SEROTYPE c ISOLATES 1805
Biotinylation. A 1.4-kb DraIII-HindIll fragment of p26R3and the entire p26L10 plasmid were labeled with biotin-7-dATP (BRL) by using a nick translation kit (BRL) accordingto the manufacturer's suggested protocol.Four 18-base oligonucleotides (spanning bases 3961 to
3979, 4029 to 4047, 4321 to 4339, and 4731 to 4749 of the 3'terminus of spaP) were end labeled with biotin-7-dATP asdescribed previously (20).Southern hybridization. Complete DNA digests were elec-
trophoresed at 15 V/cm of gel on 0.7% agarose in lx TAEbuffer (0.04 M Tris-acetate-0.002 M EDTA, pH 8.6), depu-rinated in 0.25 M HCl (15 min), denatured in 1.5 M NaCl-0.5M NaOH (twice for 20 min), and neutralized in 1.5 MNaCI-1.0 M Tris-HCl, pH 7.5 (20 min). Gels were washed in0.1 x TAE, and the DNA was transferred onto PhotoGenenylon membranes (BRL) by electroblotting in 0.1 x TAE (1h, 200 mA). Southern blots were prehybridized, hybridizedwith biotinylated probe DNA, and developed with the Pho-toGene nonradioactive detection system (BRL) according tothe manufacturer's suggested protocol.For Southern blots in which biotinylated oligonucleotides
were used as probes, digested DNA was electroblotted ontonitrocellulose (Bio-Rad Laboratories, Richmond, Calif.) in0.1 x TAE (1 h, 200 mA). Prehybridization was performedwith 2x SSC (0.3 M NaCl-0.03 M sodium citrate, pH 7.0)containing 0.5% (wt/vol) Ficoll, 0.5% (wt/vol) polyvinylpyr-rolidone, 0.5% (wt/vol) bovine serum albumin, 1% (wt/vol)SDS, and 0.5 mg of sheared denatured salmon sperm DNAper ml (2 h, 37°C). Hybridization was performed with thesame solution containing approximately 2 ,ug of biotinylatedoligonucleotide probe DNA per ml (16 h, 37°C). Posthybrid-ization washes and development were performed with theBluGene nonradioactive detection system (BRL) accordingto the manufacturer's suggested protocol.PCR. Chromosomal DNA was prepared from S. mutans
isolates as described above. Using the primers 5'-GCTTCCGCTTATACA-3' (nucleotides 2065 to 2079 of spaP) and3'-TAGGAAGATTAACCGCACG-5' (complementary tonucleotides 2554 to 2572 of spaP), a 0.5-kb fragment whichspanned most of the V region of spaP was amplified by thepolymerase chain reaction (PCR) (39). The reaction was 30cycles (94°C for 2 min, 55°C for 1 min, 72°C for 2 min) witha thermal cycler (Bioexcellence, Techne Ltd., Cambridge,England).
Nucleotide sequencing. Double-stranded DNA from thePCR was sequenced directly by using a modified procedure(44) based on the dideoxy chain termination method (40).The oligonucleotide primers used for the PCR were also usedto sequence the fragments. The sequences were assembledand analyzed by using the Staden Plus program (Amersham,England).
RESULTS
A complete restriction map of spaP based on the sequencedetermined for S. mutans NG5 was generated (21). Thoserestriction endonucleases that demonstrated a single recog-nition site within the region of interest (e.g., the A or Vregion) or a single recognition site within the entire genewere chosen for use in this study (see Fig. 2). To ensure thatany RFLPs detected were the result of alteration of therestriction site within the region of interest and not ofchanges elsewhere in the coding or flanking sequences,chromosomal DNA was predigested with "boundary" en-zymes, which demonstrated conserved sites among theisolates tested. These sites corresponded as closely as pos-
sible to the 5' and 3' ends of each probe. Analysis of strainNG5 served as a positive control.
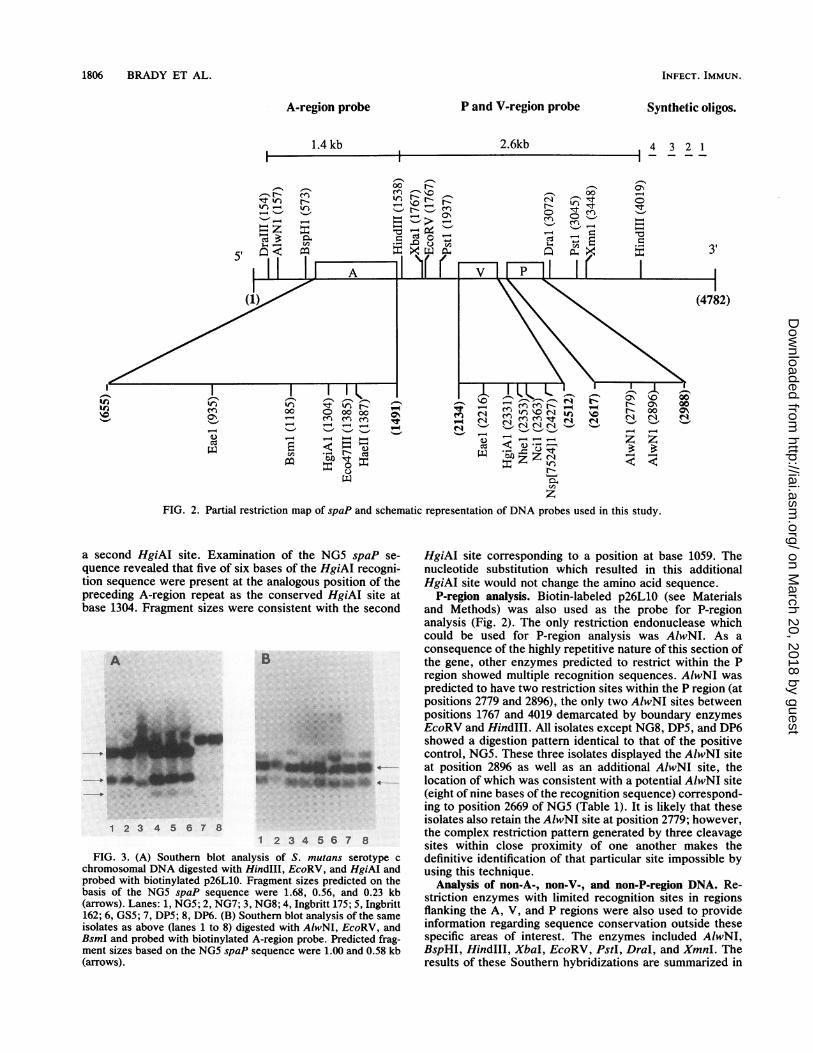
V-region analysis. Biotin-labeled p26L10 (see Materialsand Methods) was used as the probe for V-region analysis(Fig. 2). The insert in this recombinant plasmid includedthat portion of spaP spanning the HindlIl sites from posi-tions 1538 to 4019. The 5' and 3' boundary enzymes forthe V-region experiments were EcoRV (position 1767) andHindIII (position 4019), respectively. The restriction endo-nucleases EaeI, HgiAI, NheI, NciI, and Nsp(7524)I wereused to analyze polymorphisms within the V region. TheV-region Southern analysis using HgiAI as the third enzymeis shown in Fig. 3A. Fragment sizes predicted on the basis ofthe spaP sequence were 1.68, 0.56, and 0.23 kb and wereobserved for isolates NG5, NG7, Ingbritt 175, Ingbritt 162,and GS5. Isolates NG8, DP5, and DP6 displayed a differentpattern, indicating that they lacked the HgiAI site at position2331. The summary of the results of all V-region experimentsis shown in Table 1. RFLPs observed in this region paral-leled the differences observed by comparison of spaP andpac gene sequences. On the basis of RFLPs within the Vregion, there appeared to be two families of isolates. IsolatesNG8, DP5, and DP6 lacked those restriction sites present inthe spaP sequence which were also absent from the pacsequence (34). These three isolates demonstrated differentEaeI, HgiAI, NheI, Ncl, and Nsp(7524)I sites than the onespredicted for NG5; however, the exact locations of thesesites could not be determined by RFLP analysis sincecorresponding partial recognition sequences were not lo-cated in the NG5 sequence. All other isolates possessed thespaP restriction sites predicted from the NG5 sequence withone exception; GS5 lacked the EaeI site at position 2216.A fragment of the gene (nucleotides 2065 to 2572) encoding
the V region was amplified with the PCR from isolates GS5,NG8, DP5, and DP6. Five additional isolates, CCY1, CCW1,Ingbritt, Guy's c, and K2, were also included in this part ofthe study. Nucleotide sequencing of the PCR productsdemonstrated that there were only two major variant formsof this region, since all the isolates showed virtual identitywith either spaP or pac. Occasional nucleotide substitutionswere detected that were indicative of strain variation (Table2).
A-region analysis. The DNA probe encompassing the Aregion (Fig. 2) was generated by cleavage of p26R3 (seeMaterials and Methods) at the DraIllI site at position 154, toremove non-spaP 5'-flanking DNA, and at the HindIll site atposition 1538. AlwNI was used as the 5' boundary enzyme inthis series of Southern hybridizations since its recognitionsite at position 187 was conserved among all isolates tested.EcoRV, which demonstrated a conserved site at position1767, was used as the 3' boundary enzyme (Fig. 2) since theHindIII site at position 1538 was not present in isolates NG8,DP5, and DP6. Chromosomal DNA from eight isolates of S.mutans serotype c was digested with AlwNI, EcoRV, and athird enzyme with a single recognition site within the spaP Aregion. The Southern analysis using BsmI as the thirdenzyme is shown in Fig. 3B. Fragment sizes predicted on thebasis of the spaP sequence were 1.00 and 0.58 kb. All strainstested demonstrated these fragments. Fainter higher-molec-ular-weight bands represent hybridization to partially di-gested DNA. The restriction endonucleases EaeI, HgiAI,Eco47III, and HaeII also were used to investigate RFLPswithin the A region (Table 1). Only one polymorphism wasdetected within the alanine-rich repeats of the A region.Isolates NG7, Ingbritt 175, and Ingbritt 162 demonstrated aSouthern hybridization pattern that indicated the presence of
VOL. 59, 1991
on March 20, 2018 by guest
http://iai.asm.org/
Dow
nloaded from
1806 BRADY ET AL.
A-region probe
1.4kb
P and V-region probe
2.6kb
Synthetic oligos.
-4 3 2 1
00N
I I I km' "t mn r00 0 0o oo
4,T- -4P- -4 -._/- / _, _/"
¢--4
m =z o
I--,
V-
clir
ci
V 11 P 1z
ON
_p'2
\Cl
CZ
0011OC)~
- 0
1-4
ON0-
._
l(4782)
CON- ~~~00 Q%C"~ ~ C
W-)
_
C,)
z-
FIG. 2. Partial restriction map of spaP and schematic representation of DNA probes used in this study.
a second HgiAI site. Examination of the NG5 spaP se-quence revealed that five of six bases of the HgiAI recogni-tion sequence were present at the analogous position of thepreceding A-region repeat as the conserved HgiAI site atbase 1304. Fragment sizes were consistent with the second
A B
1 2 3 4 5 6 7 8
..
12 345678FIG. 3. (A) Southern blot analysis of S. mutans serotype c
chromosomal DNA digested with HindIII, EcoRV, and HgiAI andprobed with biotinylated p26L10. Fragment sizes predicted on thebasis of the NG5 spaP sequence were 1.68, 0.56, and 0.23 kb(arrows). Lanes: 1, NG5; 2, NG7; 3, NG8; 4, Ingbritt 175; 5, Ingbritt162; 6, GS5; 7, DP5; 8, DP6. (B) Southern blot analysis of the sameisolates as above (lanes 1 to 8) digested with AIwNI, EcoRV, andBsmI and probed with biotinylated A-region probe. Predicted frag-ment sizes based on the NG5 spaP sequence were 1.00 and 0.58 kb(arrows).
HgiAI site corresponding to a position at base 1059. Thenucleotide substitution which resulted in this additionalHgiAI site would not change the amino acid sequence.
P-region analysis. Biotin-labeled p26L10 (see Materialsand Methods) was also used as the probe for P-regionanalysis (Fig. 2). The only restriction endonuclease whichcould be used for P-region analysis was AlwNI. As aconsequence of the highly repetitive nature of this section ofthe gene, other enzymes predicted to restrict within the Pregion showed multiple recognition sequences. AlwNI waspredicted to have two restriction sites within the P region (atpositions 2779 and 2896), the only two AlwNI sites betweenpositions 1767 and 4019 demarcated by boundary enzymesEcoRV and Hindlll. All isolates except NG8, DP5, and DP6showed a digestion pattern identical to that of the positivecontrol, NG5. These three isolates displayed the AlwNI siteat position 2896 as well as an additional AlwNI site, thelocation of which was consistent with a potential AlwNI site(eight of nine bases of the recognition sequence) correspond-ing to position 2669 of NG5 (Table 1). It is likely that theseisolates also retain the AlwNI site at position 2779; however,the complex restriction pattern generated by three cleavagesites within close proximity of one another makes thedefinitive identification of that particular site impossible byusing this technique.
Analysis of non-A-, non-V-, and non-P-region DNA. Re-striction enzymes with limited recognition sites in regionsflanking the A, V, and P regions were also used to provideinformation regarding sequence conservation outside thesespecific areas of interest. The enzymes included AlwNI,BspHI, HindIII, XbaI, EcoRV, PstI, DraI, and XmnI. Theresults of these Southern hybridizations are summarized in
5S
(1)
3'
_/1
inON
a)Ce
INFECT. IMMUN.
I-
~-
on March 20, 2018 by guest
http://iai.asm.org/
Dow
nloaded from

spaP GENE OF S. MUTANS SEROTYPE c ISOLATES 1807
I.300A03t C)
00 ONto ,019.g.- _ . _.
P..P.-9b6 r+F,
o+
++++
I + + + + +
I
+ x xx
0
0),Oct
00
_ _
0
C,00
t2
00 2
00~
-JO
00~
Vo
00
Ss
000
08f
%
0sn
'0t
CDo0
0
a
0F'
Table 1. With the exception of the HindIII site at position1538 (missing from isolates NG8, DP5, and DP6), there wereno RFLPs detected with these enzymes.
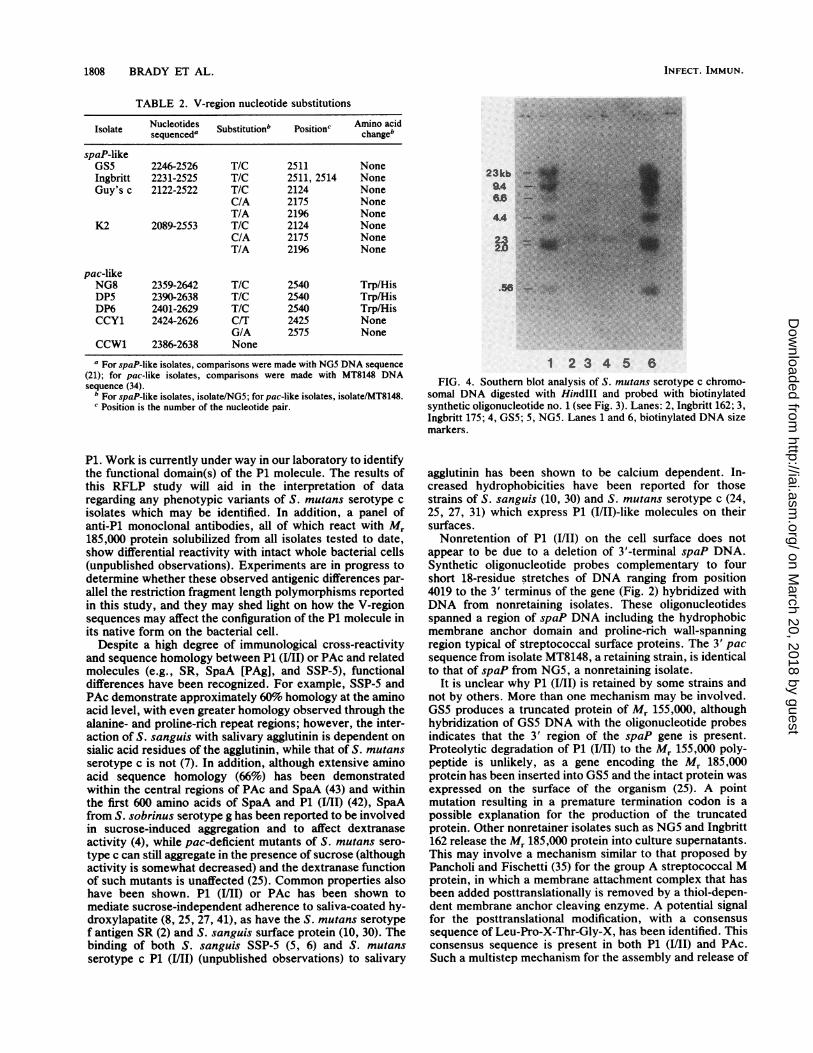
Southern analysis of nonretaining isolates with 3' oligonu-cleotides. To determine whether S. mutans isolates which donot retain P1 (I/II) on their cell surfaces but which release themolecule into culture supernatants have a deletion at the 3'end of the spaP gene (where the C-terminal membrane andwall anchor sequences are located), Southern hybridizationswere performed with four end-labeled 3' oligonucleotideprobes. The positions of these 18-residue oligonucleotideswithin the spaP sequence derived from NG5 are shown inFig. 2. All four oligonucleotide probes hybridized withHindIII-restricted DNA from all Pl-nonretaining isolates,including GS5, which produces a truncated Mr 155,000protein instead of a full-length Mr 185,000 product (23). Arepresentative Southern blot is shown in Fig. 4. Since allisolates were shown to have a conserved HindIII site atposition 4019, differences in fragment sizes reflect polymor-phisms in flanking DNA downstream of the 3' terminus ofspaP.
tT
COT0)
w
CO)
I10
0
C,,Z
0u)-CD
DISCUSSION
Analysis of S. mutans serotype c isolates indicated thatthe spaP gene is highly conserved, with the exception ofvariable region differences first identified by comparing spaPand pac sequences. Analysis of the A region demonstratedcomplete conservation of five restriction sites, with anadditional HgiAI site in three of eight isolates tested. Anal-ysis of the P region was limited to the use ofAlwNI. Threeof eight isolates demonstrated an additional AlwNI siteupstream from those predicted for NG5 spaP. Given therepetitive nature of the A and P regions, the identification ofadditional restriction sites in some isolates is not surprising.RFLP analysis of spaP outside the A, P, and V regionsdemonstrated no polymorphisms among eight restrictionsites, with the exception of three of eight isolates whichlacked the HindIII site at position 1538.RFLP analysis of the V regions of eight isolates revealed
two patterns. One is spaP-like (21), and such isolates retainthose V-region restriction sites predicted for spaP from theNG5 sequence. The second is pac-like (34), and theseisolates lack the restriction sites for the five enzymes tested.Furthermore, the sequences encoding the V region from anumber of S. mutans isolates were shown to be either spaP-orpac-like. No other variants were found. Whether isolateswere spaP- orpac-like was independent of their geographicsource. Both spaP- and pac-like isolates were identifiedamong isolates from New Guinea, the United States, andEngland. Thus, at least two types of S. mutans serotype corganisms appear to exist worldwide.
Molecular heterogeneity among streptococcal proteins hasbeen demonstrated. Huang et al. (17) reported RFLPs withinthe streptokinase gene of group A streptococci in strains ofboth the same and different serotypes. More recently,Johnston et al. (19a) used the PCR and sequence analysis todefine two variable regions, Vl and V2, also within thestreptokinase gene of group A streptococci. Their resultssuggested the existence of at least four different Vl se-quences, two of which appear to be associated with strainsimplicated in the pathogenesis of poststreptococcal glomer-ulonephritis. The significance of the V-region polymor-phisms within the spaP genes of serotype c S. mutansorganisms is not yet known since it is still unclear how thevariable region may impact on the structure and function of
VOL. 59, 1991
cz z
CCC0 co"
0.
.~ Q. rA
* r
Oo.00
CC
DO
0
0
Pco
rA 0
CD
-, CL,
coo
0 >0. PZcD
Co-
ZC)0
C,_4.
0o_
(Du*
D CD
Ci20nCo
0
CD
0 .
ox
C'C
0
CD
Co
PIP
. _
CD0*co5-
0
CD
+ + + + +
on March 20, 2018 by guest
http://iai.asm.org/
Dow
nloaded from
1808 BRADY ET AL.
TABLE 2. V-region nucleotide substitutions
Nucleotides Amino acidIsolate sequenceda Substitution' Position' changeb
spaP-likeGS5 2246-2526 T/C 2511 NoneIngbritt 2231-2525 T/C 2511, 2514 NoneGuy's c 2122-2522 T/C 2124 None
C/A 2175 NoneT/A 21% None
K2 2089-2553 T/C 2124 NoneC/A 2175 NoneT/A 21% None
pac-likeNG8 2359-2642 T/C 2540 Trp/HisDP5 2390-2638 T/C 2540 Trp/HisDP6 2401-2629 T/C 2540 Trp/HisCCY1 2424-2626 C/T 2425 None
G/A 2575 NoneCCW1 2386-2638 None
a For spaP-like isolates, comparisons were made with NG5 DNA sequence(21); for pac-like isolates, comparisons were made with MT8148 DNAsequence (34).
b For spaP-like isolates, isolate/NG5; forpac-like isolates, isolate/MT8148.c Position is the number of the nucleotide pair.
P1. Work is currently under way in our laboratory to identifythe functional domain(s) of the P1 molecule. The results ofthis RFLP study will aid in the interpretation of dataregarding any phenotypic variants of S. mutans serotype cisolates which may be identified. In addition, a panel ofanti-Pl monoclonal antibodies, all of which react with M,185,000 protein solubilized from all isolates tested to date,show differential reactivity with intact whole bacterial cells(unpublished observations). Experiments are in progress todetermine whether these observed antigenic differences par-allel the restriction fragment length polymorphisms reportedin this study, and they may shed light on how the V-regionsequences may affect the configuration of the P1 molecule inits native form on the bacterial cell.
Despite a high degree of immunological cross-reactivityand sequence homology between P1 (I/II) or PAc and relatedmolecules (e.g., SR, SpaA [PAg], and SSP-5), functionaldifferences have been recognized. For example, SSP-5 andPAc demonstrate approximately 60% homology at the aminoacid level, with even greater homology observed through thealanine- and proline-rich repeat regions; however, the inter-action of S. sanguis with salivary agglutinin is dependent onsialic acid residues of the agglutinin, while that of S. mutansserotype c is not (7). In addition, although extensive aminoacid sequence homology (66%) has been demonstratedwithin the central regions of PAc and SpaA (43) and withinthe first 600 amino acids of SpaA and P1 (I/II) (42), SpaAfrom S. sobrinus serotype g has been reported to be involvedin sucrose-induced aggregation and to affect dextranaseactivity (4), while pac-deficient mutants of S. mutans sero-type c can still aggregate in the presence of sucrose (althoughactivity is somewhat decreased) and the dextranase functionof such mutants is unaffected (25). Common properties alsohave been shown. P1 (I/II) or PAc has been shown tomediate sucrose-independent adherence to saliva-coated hy-droxylapatite (8, 25, 27, 41), as have the S. mutans serotypef antigen SR (2) and S. sanguis surface protein (10, 30). Thebinding of both S. sanguis SSP-5 (5, 6) and S. mutansserotype c P1 (I/II) (unpublished observations) to salivary
23kb '*Ei9A
4A
1 2334 5 6FIG. 4. Southern blot analysis of S. mutans serotype c chromo-
somal DNA digested with Hindlll and probed with biotinylatedsynthetic oligonucleotide no. 1 (see Fig. 3). Lanes: 2, Ingbritt 162; 3,Ingbritt 175; 4, GS5; 5, NG5. Lanes 1 and 6, biotinylated DNA sizemarkers.
agglutinin has been shown to be calcium dependent. In-creased hydrophobicities have been reported for thosestrains of S. sanguis (10, 30) and S. mutans serotype c (24,25, 27, 31) which express P1 (1/11)-like molecules on theirsurfaces.Nonretention of P1 (I/Il) on the cell surface does not
appear to be due to a deletion of 3'-terminal spaP DNA.Synthetic oligonucleotide probes complementary to fourshort 18-residue stretches of DNA ranging from position4019 to the 3' terminus of the gene (Fig. 2) hybridized withDNA from nonretaining isolates. These oligonucleotidesspanned a region of spaP DNA including the hydrophobicmembrane anchor domain and proline-rich wall-spanningregion typical of streptococcal surface proteins. The 3' pacsequence from isolate MT8148, a retaining strain, is identicalto that of spaP from NG5, a nonretaining isolate.
It is unclear why P1 (I/Il) is retained by some strains andnot by others. More than one mechanism may be involved.GSS produces a truncated protein of Mr 155,000, althoughhybridization of GSS DNA with the oligonucleotide probesindicates that the 3' region of the spaP gene is present.Proteolytic degradation of P1 (I/II) to the Mr 155,000 poly-peptide is unlikely, as a gene encoding the Mr 185,000protein has been inserted into 055 and the intact protein wasexpressed on the surface of the organism (25). A pointmutation resulting in a premature termination codon is apossible explanation for the production of the truncatedprotein. Other nonretainer isolates such as NG5 and Ingbritt162 release the Mr 185,000 protein into culture supernatants.This may involve a mechanism similar to that proposed byPancholi and Fischetti (35) for the group A streptococcal Mprotein, in which a membrane attachment complex that hasbeen added posttranslationally is removed by a thiol-depen-dent membrane anchor cleaving enzyme. A potential signalfor the posttranslational modification, with a consensussequence of Leu-Pro-X-Thr-Gly-X, has been identified. Thisconsensus sequence is present in both P1 (I/Il) and PAc.Such a multistep mechanism for the assembly and release of
INFECT. IMMUN.
on March 20, 2018 by guest
http://iai.asm.org/
Dow
nloaded from

spaP GENE OF S. MUTANS SEROTYPE c ISOLATES 1809
streptococcal surface proteins provides an explanation forthe nonretention of P1 (MI/I) by isolates which appear toencode all the information necessary for cell-surface attach-ment. Nonretention of P1 may result from differences out-side spaP, i.e., in the gene(s) encoding the membrane anchorcleaving enzyme activity or the putative membrane anchorcomplex.
ACKNOWLEDGMENTS
This work was supported by U.S. Public Health Service grantR37-DE-08007 to A.S.B. from the National Institute of DentalResearch, NIH, Bethesda, Md., and by the Dickinson Trust (toT.L.).
REFERENCES1. Abiko, Y., M. Hayakawa, H. Aoki, S. Saito, and H. Takiguchi.
1989. Cloning of the gene for cell-surface protein antigen A fromStreptococcus sobrinus (serotype d). Arch. Oral Biol. 34:571-575.
2. Ackermans, F., J. P. Klein, J. A. Ogier, H. Bazin, F. Cormont,and R. M. Frank. 1985. Purification and characterization of asaliva-interacting cell wall protein from Streptococcus mutansserotype f by using monoclonal antibody immunoaffinity chro-matography. Biochem. J. 228:211-217.
3. Ayakawa, G. Y., L. W. Boushell, P. J. Crowley, G. W. Erdos,W. P. McArthur, and A. S. Bleiweis. 1987. Isolation andcharacterization of monoclonal antibodies specific for antigenP1, a major surface protein of mutans streptococci. Infect.Immun. 55:2759-2767.
4. Curtiss, R., III, S. A. Larrimore, R. G. Holt, J. F. Barrett, R.Barletta, H. H. Murchison, S. M. Michalek, and S. Saito. 1983.Analysis of Streptococcus mutans virulence attributes usingrecombinant DNA and immunological techniques, p. 95-104. InR. J. Doyle and J. E. Ciardi (ed.), Glucosyltransferases, glu-cans, sucrose and dental caries. IRL Press, Washington, D.C.
5. Demuth, D. R., P. Berthold, P. S. Leboy, E. E. Golub, C. A.Davis, and D. Malamud. 1989. Saliva-mediated aggregation ofEnterococcus faecalis transformed with a Streptococcus san-guis gene encoding the SSP-5 surface antigen. Infect. Immun.57:1470-1475.
6. Demuth, D. R., C. A. Davis, A. M. Corner, R. J. Lamont, P. S.Leboy, and D. Malamud. 1988. Cloning and expression of aStreptococcus sanguis surface antigen that interacts with ahuman salivary agglutinin. Infect. Immun. 56:2484-2490.
7. Demuth, D. R., E. E. Golub, and D. Malamud. 1990. Strepto-coccal-host interactions: structural and functional analysis of aStreptococcus sanguis receptor for a human salivary glycopro-tein. J. Biol. Chem. 265:7120-7126.
8. Douglas, C. W. I., and R. R. B. Russell. 1984. Effect of specificantisera upon Streptococcus mutans adherence to saliva-coatedhydroxylapatite. FEMS Microbiol. Lett. 25:211-214.
9. Fischetti, V. A. 1989. Streptococcal M protein: molecular designand biological behavior. Clin. Microbiol. Rev. 2:285-314.
10. Fives-Taylor, P. M., and D. W. Thompson. 1985. Surfaceproperties of Streptococcus sanguis FW213 mutants nonadher-ent to saliva-coated hydroxyapatite. Infeet. Immun. 47:752-759.
11. Forester, H., N. Hunter, and K. W. Knox. 1983. Characteristicsof a high molecular weight extracellular protein of Streptococ-cus mutans. J. Gen. Microbiol. 129:2779-2788.
12. Gibbons, R. J. 1984. Adherent interactions which may affectmicrobial ecology in the mouth. J. Dent. Res. 63:378-385.
13. Guss, B., M. Eliasson, A. Olsson, M. Uhlen, A.-K. Frej, H.Jornvall, J.-I. Flock, and M. Lindberg. 1986. Structure of theIgG-binding regions of streptococcal protein G. EMBO J.5:1567-1575.
14. Hamada, S., and H. D. Slade. 1980. Biology, immunology, andcariogenicity of Streptococcus mutans. Microbiol. Rev. 44:331-384.
15. Hollingshead, S. K., V. A. Fischetti, and J. R. Scott. 1986.Complete nucleotide sequence of type 6 M protein of the groupA streptococcus. J. Biol. Chem. 261:1677-1686.
16. Holt, R. G., Y. Abiko, S. Saito, J. Smorawinska, J. B. Hansen,and R. Curtiss III. 1982. Streptococcus mutans genes that codefor extracellular proteins in Escherichia coli K-12. Infect. Im-mun. 38:147-156.
17. Huang, T.-T., H. Malke, and J. J. Ferretti. 1989. Heterogeneityof the streptokinase gene in group A streptococci. Infect.Immun. 57:502-506.
18. Hughes, M., S. M. MacHardy, A. J. Sheppard, and N. C. Woods.1980. Evidence for an immunological relationship betweenStreptococcus mutans and human cardiac tissue. Infect. Im-mun. 27:576-588.
19. Ish-Horowitz, D., and J. F. Burke. 1981. Rapid and efficientcosmid vector cloning. Nucleic Acids Res. 9:2989-2998.
19a.Johnston, K. H., J. D. Chaiban, and P. C. Wheeler. 1990.Analysis of the variable domain of the streptokinase gene fromstreptococci associated with post-streptococcal glomerulone-phritis, abstr. 27. Am. Soc. Microbiol. 3rd Int. Conf. Strepto-coccal Genetics.
20. Karger, B. 1989. Preparation and use of biotinylated oligonucle-otide probes. Focus 11:57-58.
21. Kelly, C., P. Evans, L. Bergmeier, S. F. Lee, A. Progulske-Fox,A. C. Harris, A. Aitken, A. S. Bleiweis, and T. Lehner. 1989.Sequence analysis of the cloned streptococcal surface antigenI/II. FEBS Lett. 258:127-132.
22. Kelly, C., P. Evans, J. K.-C. Ma, L. A. Bergmeier, W. Taylor,L. J. Brady, S. F. Lee, A. S. Bleiweis, and T. Lehner. 1990.Sequencing and characterization of the 185 kDa cell surfaceantigen of Streptococcus mutans. Arch. Oral Biol. 35(Suppl.):33S-38S.
23. Knox, K. W., L. Hardy, and A. D. Wicken. 1986. Comparativestudies on the protein profiles and hydrophobicity of strains ofStreptococcus mutans serotype c. J. Gen. Microbiol. 132:2541-2548.
24. Koga, T., H. Asakawa, N. Okahashi, and I. Takahashi. 1989.Effect of subculturing on expression of a cell-surface proteinantigen by Streptococcus mutans. J. Gen. Microbiol. 135:3199-3207.
25. Koga, T., N. Okahashi, I. Takahashi, T. Kanamoto, H. Asakawa,and M. Iwaki. 1990. Surface hydrophobicity, adherence, andaggregation of cell surface protein antigen mutants of Strepto-coccus mutans serotype c. Infect. Immun. 58:289-296.
26. Lee, S. F., A. Progulske-Fox, and A. S. Bleiweis. 1988. Molecularcloning and expression of a Streptococcus mutans major surfaceprotein antigen, P1 (I/II), in Escherichia coli. Infect. Immun.56:2114-2119.
27. Lee, S. F., A. Progulske-Fox, G. W. Erdos, D. A. Piacentini,G. Y. Ayakawa, P. J. Crowley, and A. S. Bleiweis. 1989.Construction and characterization of isogenic mutants of Strep-tococcus mutans deficient in major surface protein antigen P1(HII). Infect. Immun. 57:3306-3313.
28. Maniatis, T., E. F. Fritsch, and J. Sambrook. 1982. Molecularcloning: a laboratory manual. Cold Spring Harbor Laboratory,Cold Spring Harbor, N.Y.
29. McBride, B. C., M. Song, B. Krasse, and J. Olson. 1984.Biochemical and immunological differences between hydropho-bic and hydrophilic strains of Streptococcus mutans. Infect.Imnmun. 44:68-75.
30. Morris, E. J., N. Ganeshkumar, and B. C. McBride. 1985. Cellsurface components of Streptococcus sanguis: relationship toaggregation, adherence, and hydrophobicity. J. Bacteriol. 164:255-262.
31. Ohta, H., H. Kato, N. Okahashi, I. Takahashi, S. Hamada, andT. Koga. 1989. Characterization of a cell-surface protein antigenof hydrophilic Streptococcus mutans strain GS-5. J. Gen. Mi-crobiol. 135:981-988.
32. Okahashi, N., T. Koga, and S. Hamada. 1986. Purification andimmunochemical properties of a protein antigen from serotype gStreptococcus mutans. Microbiol. Immunol. 30:34-47.
33. Okahashi, N., C. Sasakawa, M. Yoshikawa, S. Hamada, and T.Koga. 1989. Cloning of a surface protein antigen gene fromserotype c Streptococcus mutans. Mol. Microbiol. 3:221-228.
34. Okahashi, N., C. Sasakawa, M. Yoshikawa, S. Hamada, and T.Koga. 1989. Molecular characterization of a surface protein
VOL. 59, 1991
on March 20, 2018 by guest
http://iai.asm.org/
Dow
nloaded from

1810 BRADY ET AL. INFECT. IMMUN.
antigen gene from serotype c Streptococcus mutans, implicatedin dental caries. Mol. Microbiol. 3:673-678.
35. Pancholi, V., and V. A. Fischetti. 1989. Identification of an
endogenous membrane anchor-cleaving enzyme for group Astreptococcal M protein. J. Exp. Med. 170:2119-2133.
36. Russell, M. W., E. D. Bergmeier, E. D. Zanders, and T. Lehner.1980. Protein antigens of Streptococcus mutans: purificationand properties of a double antigen and its protease-resistantcomponent. Infect. Immun. 28:486-493.
37. Russell, R. R. B. 1979. Wall associated antigens of Streptococ-cus mutans. J. Gen. Microbiol. 114:109-115.
38. Russell, R. R. B. 1980. Distribution of cross-reactive antigens Aand B in Streptococcus mutans and other oral streptococci. J.Gen. Microbiol. 118:383-388.
39. Saiki, R. K., D. H. Gelfand, S. Stoffel, S. J. Scharf, R. Higuchi,G. T. Horn, K. B. Mullis, and H. A. Erlich. 1988. Primerdirected enzymatic amplification of DNA with a thermostableDNA polymerase. Science 239:487-491.
40. Sanger, F., S. Nicklen, and A. R. Coulson. 1977. DNA sequenc-
ing with chain-terminating inhibitors. Proc. Natl. Acad. Sci.USA 74:5463-5467.
41. Staat, R. H., S. D. Langley, and R. J. Doyle. 1980. Streptococcusmutans adherence: presumptive evidence for protein-mediatedattachment followed by glucan-dependent cellular accumula-
tion. Infect. Immun. 27:675-681.42. Staffileno, L. K., M. Hendricks, R. LaPolla, P. Van Hook, J. I.
Rosen, J. Warner, K. Hoey, D. Wegemer, R. B. Naso, R. D.Sublett, B. Waldschmidt, M. Leong, G. G. Thornton, T. Lehner,and J. A. Haron. 1990. Cloning and sequencing of the aminoterminal nucleotides of the antigen I/II of Streptococcus sobri-nus and the immune response to the corresponding syntheticpeptides. Arch. Oral Biol. 35(Suppl.):47S-52S.
43. Takahashi, I., N. Okahashi, C. Sasakawa, M. Yoshikawa, S.Hamada, and T. Koga. 1989. Homology between surface proteinantigen genes of Streptococcus sobrinus and Streptococcusmutans. FEBS Lett. 249:383-388.
44. Winship, P. R. 1989. An improved method for directly sequenc-ing PCR amplified material using dimethyl sulphoxide. NucleicAcids Res. 17:1266.
44a.Yother, J., and D. E. Briles. 1990. Generation of truncated formsof pneumococcal surface protein A (PspA) and DNA sequenc-ing of pspA, abstr. D-106, p. 98. Abstr. 90th Annu. Meet. Am.Soc. Microbiol. 1990. American Society for Microbiology,Washington, D.C.
45. Zanders, E. D., and T. Lehner. 1981. Separation and character-ization of a protein antigen from cells of Streptococcus mutans.J. Gen. Microbiol. 122:217-225.
on March 20, 2018 by guest
http://iai.asm.org/
Dow
nloaded from