restriction profiling of 23s microheterogenic ribosomal repeats for
TRANSCRIPT

Research ArticleRestriction Profiling of 23S Microheterogenic RibosomalRepeats for Detection and Characterizing of E coli and TheirClonal Pathogenic and Phylogroups
Parvathi Jayasree Rajagopalan Nair and Sunita Singh
School of Biotechnology amp Bioinformatics D Y Patil University Plot 15 Sector 50 CBD Belapur Navi MumbaiMaharashtra 400614 India
Correspondence should be addressed to Sunita Singh drsunitadsgmailcom
Received 1 August 2015 Revised 31 October 2015 Accepted 2 November 2015
Academic Editor Olimpia Pepe
Copyright copy 2015 P Jayasree Rajagopalan Nair and S SinghThis is an open access article distributed under theCreative CommonsAttribution License which permits unrestricted use distribution and reproduction in any medium provided the original work isproperly cited
Correlating ribosomal microheterogenicity with unique restriction profiles can prove to be an efficacious and cost-effectiveapproach compared with sequencing for microbial identification An attempt to peruse restriction profiling of 23S ribosomalassemblage was ventured digestion patterns with Bfa I discriminated E coli from its colony morphovars while Hae III profilesassisted in establishing distinct clonal groups Among the gene pool of 399 ribosomal sequences extrapolated from 57 E coligenomes varying degree of predominance (I gt III gt IV gt II) ofHae III pattern was observedThis was also corroborated in samplescollected from clinical commensal and environmental origin K-12 and its descendants showed type I pattern whereas E coli-Band its descendants exhibited type IV both of these patterns being exclusively present in E coli A near-possible association betweenphylogroups and Hae III profiles with presumable correlation between the clonal groups and different pathovars was establishedThe generic nature conservation and barcode gap of 23S rRNA gene make it an ideal choice and substitute to 16S rRNA gene themost preferred region for molecular diagnostics in bacteria
1 Introduction
The need of rapid assay with low detection limits hasreverberated genotypic techniques to gain momentum indiagnostic field which not only revolutionized but enhancedthe authentication process Within a short span the evolv-ing concepts and strategies for molecular detection haveresulted in the shift of ldquogold standardrdquo from being DNA-DNA hybridization (DDH) to currently Nucleic Acid Ampli-fication Techniques (NAATs) and gradually towards Aver-age Nucleotide Identity (ANI) [1ndash4] Among the NAATsPolymerase Chain Reaction (PCR) based on amplificationof 16S rRNA region [5 6] followed by the sequencing ofamplicon forms a simple yet efficient rationale for bacte-rial barcoding [7] Basis for targeting this gene segmentis reasoned by the ubiquitousness in bacterial kingdom tomultiple ribosomal RNA copiesnumbers rrn [8 9] provid-ing sensitivity in PCR reaction Another remarkable feature
of ribosomal appendage is the degree of conservation andheterogenicity with the nonvariable slow evolving regionspreferred for the identification and variable regions beingthe choice for phylogenetic studies [10 11] While design-ing universal or specific primers against these stretchesparameters like primer specificity hybridization efficiencyand amplicon incongruity arising due to microheterogeneitywithin rrn copies must be taken into consideration [1213] These precautionary measures can help in limiting thepitfalls of molecular assay and help to winnow the ingrainedinformation regarding the usability and effectiveness ofprimers for identification An alternative to this scenario isto exploit inter- and intragenic ribosomalmicroheterogeneityfor developing Single Nucleotide Polymorphism (SNP) baseddiscriminative methods The logic behind this stratagem isto develop broad range primers against microdiverse regionsof the targeted stretch which encompass distinct restrictionprofile characteristic to each organism(s) under study [14]
Hindawi Publishing CorporationJournal of PathogensVolume 2015 Article ID 562136 11 pageshttpdxdoiorg1011552015562136
2 Journal of Pathogens
This current study is an endeavor to entitle 23S rRNAgeneas substitute to de facto barcode 16S rRNA gene using ampli-fication ensued by restriction digestion instead of sequencingmaking it a cost-effective approach The exemplification ofthe proposed method was carried out to detect and differ-entiate E coli (candidate) from E aerogenes K pneumoniaeandC koseri (noncandidates)The above choice was finalizedafter considering the following (i) all the selected microbesboth candidate and noncandidate are Gram negative lactosepositive microbes of 120574-Enterobacteriaceae family [15] (ii)all are urinary tract infection (UTI) pathogens with E colibeing reported as the major causative factor [16ndash18] andlastly (iii) all exhibit similar colony semblance and any falsepositive and negative biochemical characterization resultingin misinterpretation with candidate strain
In spite of being the model representative of bacterialkingdom E coli is also acknowledged for its omnipresenceand the capability of switching from good to bad microbePathogenic E coli are classified into aggressive invasivepathogenic hemorrhagic and toxigenic which differs in theirvirulence genes acquired and the type of pathogenicity [19]Comparative study of commensal and pathogenic strainsof the same has shown a rather surprising result of fewerpathovar specific sequences evincing E coli as a genera list[12] Sharing of 40 of core genomes among both virulentand nonvirulent strains indicates clonality [20] This con-ceptualizes an open pangenome having genetic reservoirs toallow gene acquisition for evolving into pathogenic ones [21]
Besides isolates of E coli present in nature the routinelyused nonvirulent lab strains are K-12 B W Crooks and Cwith all being fecal in origin except for strain W (soil isolate)[22] Method of rapid identification of K-12 strains usingmutations in O-antigen gene cluster [23] and determinationof lab strains lineage using multiplex primers [24] have beenreported Studies on the lineages of well-established E colilab strains across the globe have shown that most strainsare derived from either K-12 or B [25 26] Genomes ofthese supposed to be parental strains show 99 geneticidentity with exceptions to IS elements and flagellar genes[27] Phylogenetic classification of E coli strains into A B1B2 D and E groups usingMultiloci Enzyme ElectrophoresisMLEE [28] and triplex [29] has given insights into theconcept of clonality in E coli A comparative analysis ofMLEE and randomamplification restriction profiling against16S-23S intertranscribed region and ribotyping have verifiedthat horizontal gene transfer mechanisms do not disruptorganization of clonal population [30]
No study till date has correlated Amplified RibosomalDNA Restriction Analysis (ARDRA) of a core segment in23S rRNA gene against the pangenome and tried to establisha link between phylogroups pathogroups and restrictiongroups
2 Materials and Methods
21 Bacterial Strains and Culture Conditions Authenticationof the molecular assay in screening and identifying the can-didate organism was executed into parts one with standardstrains of E coli and the other with 25 isolates each from fecal
clinical and environmental origin Lyophilized strains wereprocured from Microbial Type Culture Collection (MTCC)Institute of Microbial Technology (IMTECH ChandigarhIndia) for both identification and clonality studies (Table 1)
Five different sets of fecal droppings from rat chickenpigeon rabbit (pet shops Crawford market Mumbai India)and humans (Excel Pathology LabVashi NaviMumbai) wereconsidered for isolation of commensal samples The strainswere revived in nutrient broth at 37∘C For environmentalstrains water sample from ten local water bodies lying in andaround the parent institute was utilized (areas with hospitalsor health care centers in the vicinity were avoided to preventcross-contamination with clinical samples) For isolating Ecoli of clinical origin urine samples from twenty-five patientssuffering from UTI (Excel Pathology Lab) were considered
The samples were processed within 24 hrs of collection(either in zip log bags or in sterile glass bottle) onMacConkeyagar (MAC) The distinct pink colonies were restreaked ontoboth MAC and Eosin Methylene Blue (EMB) agar respec-tively [31] Isolates giving discrete colony morphology withindeterminate greenish sheen were selected and evaluated forthe false positive and negatives by both microbiochemicaland molecular assays Each of the selected colonies waspicked with sterile wooden toothpicks and transferred to fivemL of nutrient broth and incubated overnight at 37∘CThesesamples were further characterized [32] by IMViC 1mL wasutilized for DNA isolation and the remaining was used forpreparing glycerol stock (25 glycerol) for future reference
22 Genome Database and Microheterogenicity Analysis Theaboriginal step involved the aggregation of Genbank genomesequences of all the strains that were considered for this studyfrom the public data repository National Centre for Biotech-nology Information (NCBI) The data pool (until 11 April2013) comprised an assemblage of 61 completed genomes(Supplementary Data A1 in Supplementary Material avail-able online at httpdxdoiorg1011552015562136) with 57of candidate strains (E coli) and noncandidate strains (Kpneumoniae C koseri and E aerogenes) The following stepswere executed to analyze the microheterogenicity (Figure 1)which can directly assist in evaluating the electability of 23SrRNA gene as an ideal barcode
Two strains one from the candidate and one from non-candidate genome (E coli str K-12 MG1655 (NC 0009132)C koseri ATCC BAA-895 (NC 0097921) E aerogenes(NC 0156631) and K pneumoniae subsp pneumoniaeMGH78578 (NC 0096481)) were selected and assigned as refer-ence strain
For studying intergenic variability among the candidateand noncandidate strains the rrsH (16S RNA gene) and rrlH(23S rRNA gene) sequence on the regular strand (referencestrains) were extrapolated and multiple sequence alignment(MSA) using ClustalW [33] was executed Total mismatchwas enumerated by the addition of number of mismatchesand sequence gaps introduced during the ldquoline-uprdquo of twoor more sequences The mismatch percentage was calculatedby dividing the total mismatches seen across the four strainsby the average base pair (bp) for both 16S and 23S rrn generespectively
Journal of Pathogens 3
Table 1 List of standard strains used in this study
Strains MTCC ATCC NatureBarcodingidentification studies
Noncandidate strainsCitrobacter freundii 1658 8090 ClinicalCitrobacter koseri 1657 27028 Blood cultureKlebsiella pneumonia 432 33495 Urinary cultureEnterobacter aerogenes 111 13048 Sputum
Candidate strains433 15228 Commensal
Escherichia coli 4296 19138 Urine448 25922 Clinical sample
Diversityclonality studies
Escherichia coli
1302 mdash K122622 10798 Descendent of K121667 mdash DH11304 mdash C11687 8739 Crooks strain448 9637 W2692 53846 Descendent of W1303 mdash B1678 mdash BL211679 mdash BL21DE31680 mdash BL21DE3Lys5
E coli namely K-12 B C Crooks andW are commonly used as lab strains Whole genomic data of these strains except strain C is available (NCBI Genomes)
Candidate microbe
Noncandidate microbe
Escherichia coli
Klebsiella pneumoniae (2 G)Enterobacter aerogenes (1 G)
Citrobacter koseri (1 G)
sequences from each genome
57 genomes (G) (21 pathogenic
36 nonpathogenic)
Estimation of diversity (multiple sequence alignment MSA)Intergenic diversity
representative genome Intragenic diversity
Restriction profiling for differentiating candidate
from noncandidates (NEB V20)
85 restriction groups(Suppl A2)
Virtual digestion was individually
carried out on each rrlH
Similar restriction patterns(rejected) Dissimilar restriction patterns
(selected)
Restriction patterns were checked in all the 430 ribosomal segments
Unique profile in candidate alone
(barcoding)Bfa I was chosen
Polymorphic within candidate genome
(clonality)Hae III was chosen
A total of 430 sequences considering both candidate and noncandidate genomes
(ii) Regular strands of eachgenome
(i) Complementary strands ofeach genome
rrlH from each
Extraction of 23S rrn
Figure 1 Schematic representation of methodology for detecting barcode and estimating clonality The primers are designed against theregions that flank these restriction sequences which gives unique bands revealing identification or clonality
For estimating the intragenic variability all the riboso-mal segments from 57 E coli genomes were extracted (7copiesgenome) complementary and regular strands withineach strain were segregated and the percentage mismatchesfor each of the genomes were determined
23 Virtual Restriction Profiling Virtual restriction profilesusing NEB V20 [34] with 85 restriction groups (Supplemen-tary Data A2) were performed for a total of 430 ribosomalsegments that included 399 from 57 E coli genomes and7 8 and 16 rrn sequences from C koseri E aerogenes
4 Journal of Pathogens
Table 2 Restriction site of Bfa I (CTAG) within 23S rrlH of candidate and noncandidates and the restriction profiles with 23S P1880
Restriction profile within rrlH Fragments obtained on ARDRAF E Ci K Ea Mic Fragments (amplicon size)11 687 685 685 685 E 665 bp 215 bp (880 bp)12 874 874 874 C 475 bp 214 bp and 189 bp (878 bp)13 1670 1669 1667 1670 K 475 bp 214 bp and 189 bp (877 bp)14 2336 2339 Ea 475 bp 214 bp and 189 bp (876 bp)15 2652 2653 2650 2654Primers were designed against the regions flanking the bold restriction sitesF number of fragments E Escherichia coli Ci Citrobacter koseri K Klebsiella pneumoniae Ea Enterobacter aerogenes Mic microbe23S P1
880-FP(+) 51015840-GGCGAAAAGAACCCCGGCGA-31015840 (20 nt) 23S P1
880-RP(minus) 51015840-AGGGGTCGACTCACCCTGCC-31015840 (20 nt)
FP forward primer RP reverse primer The primer details were obtained from the results of primer blast and restriction profiling pattern was obtained fromNEB V20 Forward and reverse primers of 23S P1
880lie within domains I and III of 23S rRNA secondary structure respectively with Bfa I sites in domain II
Table 3 Clonal groups based on Hae III (GGCC) profiles within the candidate microbe using 23S P2682
Virtual digest pattern Types of Hae III restriction pattern observed amongst 399 rrn sequences of E coli genomesF C Type I Type II Type III Type IV11 785 Cut site within rrlH of candidate rrn segments12 1042 1984 1984 1984 198413 1116 x 2163 X 216314 1217 x X 2205 220515 1361 2415 2415 2415 241516 1445 Cut site within the amplicon (682 bp)17 1907 28 28 28 2818 1984 x 207 X 20719 2161 x X 249 249110 2205 459 459 459 459111 2415 Fragments obtained for each group
112 2839 28 bp 223 bp and 431 bp 28 bp 179 bp 223 bp and252 bp
28 bp 210 bp 221 bp and223 bp
28 bp 42 bp 179 bp210 bp and 223 bp
Primers were designed against the regions flanking the bold restriction sites F number of fragments C cut sites23S P2
682-FP(+) 51015840CCGACCTGCACGAATGGCGT31015840 (20 nt)
23S P2682
-RP(+) 51015840CAGTTCTCCAGCGCCCACGG31015840 (20 nt)Forward and reverse primers of 23S P2
682lie within domain IV and transition of V-VI of 23S rRNA secondary structure respectively First cut sites lie within
domain IV whereas the rest lie within domain V
and K pneumoniae strains respectively Among the 14common cutters groups (Supplementary Data A3) Bfa I(Supplementary Data A4) was selected as the enzyme forbarcoding as it gave conserved and distinct pattern acrossall the 399 rrn sequences (not shown) in differentiating thecandidate from the noncandidate strains The uniquenessand position of the cuts sites for giving unique bands onagarose gel electrophoresis were also a lead reason consideredfor finalizing this enzyme Hae III displaying polymorphismpattern with 23S rDNA segments of candidate microbe wasfinalized for diversity studies (Figure 1)
24 Primers and Restriction Enzymes for Addressing Identityand Clonality Sequence of rrlH of E coli K-12 str MG1655was used as the template for designing broad range primersusing Primer-BLAST [35] In conjunction with parameterslike GC content 119879
119898value and the amplicon size number of
restriction sites within the amplicon and the size of fragmentafter digestion were kept as the benchmark for finalizing
primersThere are twoprimer sets 23S P1880
[Probe 31778738]was selectedwithBfa I restriction digestion (Table 2) demarkingthe candidate and 23S P2
682[Probe 31781046] (Table 3) with
polymorphicHae III restriction siteswas selected for address-ing clonality Possibility of self-priming andprimer-dimer for-mation of each primer was checked using free web tools Oli-gocal (httpwwwsimgenecomOligoCalc) OligoEvaluator(httpwwwsigmaaldrichcomtechnical-documentsarticlesbiologyoligo-evaluatorhtml) and multiple primer analyzertool (httpwwwthermoscientificbiocomwebtoolsmultiple-primer) maintained by SimGene Sigma and Fermentasresp
BLAST genome (Supplementary Data) assisted in veri-fying the conservation and specificity of primers ampliconsize Bfa I recognition sites and region of hybridizationacross the genome of 931 bacterial families After omitting thenonannotated sequences all of the completed whole genomedata aggregated under phyla Actinobacteria BacteroidetesChlamydiae Cyanobacteria Firmicutes Proteobacteria and
Journal of Pathogens 5
Spirochaetae of the bacterial kingdom were considered forthe analysis
25 DNA Isolation Amplification and Restriction DigestionDNA isolation was performed according to Parvathi andSingh [36] The reaction soup comprised 2120583L of supernatant(DNA) 05 120583M of each forward and reverse primer 1x ofassay buffer (10mM Tris-HCl pH 88 50mM KCl) MgCl
2
(15mM) dNTPmix (800 120583M) and 05U120583LTaq polymerase(Kappa Systems KK5004) Standard genomic DNA of Ecoli K-12 (Merck) and nuclease-free water were used aspositive and negative control The reactions were carriedout in a thermal cycler (Bioer) with the program set for25 cycles comprising denaturation (95∘C) annealing (68∘C)and extension (72∘C) for 30 s each 3 120583L of the PCR productwas digested with 2U of respective enzyme (FermentasThermo Scientific) in 5120583L reactionmix at 37∘C for two hoursThe amplicons and their restriction products were analysedby agarose gel electrophoresis (AGE) at 5Vcm in 2 and25 agarose gels respectively each containing 05120583gmL ofEtBr The gels were viewed and photographed in BioImagingSystems (Syngene)
3 Result and Discussion
31 Genetic Diversity within 23S Ribosomal Repeats for Identi-fication of E coli 23S rRNA gene showed 087 intergenicvariations compared with 16S rRNA within the referencegenomes (Supplementary Data A5) The result seemed obvi-ous due to the increased frame size of the latter but thisalso encouraged perusing microheterogenicity analysis ofthe largest ribosomal segment for further approach Thesizes of target segments varied from 2903 to 2907 bp withmaximum gene length observed in ATCC8739 (Crookrsquosstrain) accounting for 2970ndash2985 bp followed by E24377Adisplaying sequences of 2923ndash2925 bp One rrn of UM 146and ETECH10407 were of 1762 bp and 1561 bp respectively(Supplementary Data A9) Absolutely no link between rrnsize and pathogenicity could be defined because the varia-tions were random ATCC8739 being a laboratory cultureof fecal origin and UM146 an EIAC strain E23477A andETECH10407 of ETEC origin
Ribosomal genes were found to be distributed acrosscomplementary and regular strand with majority of thegenomes (46 out of 57) showing segregation in five regularand two complementary strands All the 23S rrn sequencesdistributed within the regular and complementary strandsacross respective genomes were individually sorted for MSAto attain an understanding of intragenic variation Analysisrevealed the highest level of mismatches in complementarystrands compared with their counterparts (SupplementaryData A6) The highest level of mismatches mounting to06 corresponding to 50 of the targeted gene size wasperceived in complementary strands of P12b (commensal)E23489 (EPEC) and ETEC H10407 The greatest dispar-ity among sequences in regular strand was observed inAPEC 01 with 003 (882903) followed by CFT043 (UPEC)and 042 (EAEC) having 0016 (sim42ndash512903) dissimilarity23S ribosomal copies of ED1a (commensal) showed zero
percent mismatches in both complementary and regularstrands making it an ideal cluster with the highest degreeof conservation Similar values of mismatches were seen inSE11 (Commensal) CB9615 (EPEC) and 0104H4 2009El-2011 (EAEC) These results indicate that there is no cor-relation between intragenic diversity of target stretch andpathogenicity of strains These differences substantiated 23SRNA clusters having microdiverse organization with bothconserved and variable stretches within them Sequevarsarising due to this heterogeneity unavoidably lead to biasedresults differential amplification and chimeric moleculesThus the method of screening and targeting restriction pro-file pattern within a region of choice (ribosomal segments)and specific to bacteria of interest demonstrates authentic andsimpler practice than routine molecular strategy involvingamplification and sequencing
32 Barcoding E coli from Its Colony Morphovars ARDRAof 23S P1880 with Bfa I PCR amplification of the stan-dard strains and isolates of different origin (commensalurine and clinical sample) with 23S P1
880gave amplicons
(880 bp) of high intensity with no primer-dimer formation(Figure 2(a)) Restriction digestion of the amplicons withBfa I gave unique fragments of 665 bp and 215 bp in E coliwhereas C koseri E aerogenes and K pneumoniae eachgave fragments of 475 bp 214 bp and 189 bp respectively(Figure 2(b)) The genotypic strategy proved superior tobiochemical characterization taking into consideration all thepossible positive negative and false positive and negativeresults across 25 isolates each from clinical commensal andenvironmental origin (Supplementary Data A7) 18 (72)of clinical 22 (88) of commensal and all the 25 (100)environment isolates showed distinct restriction pattern likethat of E coli BLAST genome search across 931 bacterialfamilies verified the specificity of primers and restrictionpreference for Bfa I The primers showed exclusive bindingonly with Enterobacteriaceae family (Supplementary DataA8) Among the various strains in the family only Shigellaspp Salmonella enterica (Arizona) and Proteus mirabilusexhibited similar restriction preference to that of E coli Inspite of Shigella spp being taxa in disguise of E coli [37] theycan be easily screened using selective medium because of nolactose fermentation [38] whereas Salmonella enterica andProteus mirabilis showed amplicon heterogeneity of 1073 bpand 880 bp during amplification thus making it easy to avoiderroneous conclusions and misinterpretation
33 Categorizing E coli StrainsUsingARDRAof 23S P2682 withHae III A detailed analysis of the Hae III profile across 399sequences [Supplementary Data A9] of ribosomal segmentsof E coli indicated four possible profiles types I II IIIand IV with varying degree of predominance within thesesegments The rest of the candidate strains showed type IIIpatterns but types I and IV were exclusive for E coli strainsMajority of genomes (119899 = 32) as in 15 nonpathogenic and17 pathogenic strains showed type I profiles (57) followedby type III (35) predominating in 15 strains Six strainsshowed type IV pattern (10) of which four were B strainsType I groups represent E coli strains that lacked recognition
6 Journal of Pathogens
1 7
880 bp
65432
(a)
8
665bp
475bp
215 bp 214 bp
178bp
1 765432
(b)
Figure 2 (a) Broad Range PCR of 23S P1880
with genomic DNA isolated from standard strains Lane 1 100 bp ladder Lane 2 blank Lane 3E coli Lane 4 C freundii Lane 5 C koseri Lane 6 K pneumonia Lane 7 E aerogenes PCR product of single representative of E coli wasloaded (b) ARDRA of 23S P1
880
amplicons with Bfa I Lanes 1 2 and 3 E coli (ATCC15223 ATCC25922 and ATCC11775) Lane 4C freundii(ATCC8090) Lane 5 C koseri (ATCC27028) Lane 6 K pneumonia (ATCC33495) Lane 7 E aerogenes (ATCC13048) Lane 8 100 bp ladder
elements due to change at 2205th and 2161st base with respectto 23S rDNA or with reference to amplicons at 207th or 249thbase positions Type IV strains are contrary to type I withintact recognition sequence at both of these sites Strains withtype II profiles lacked 2161st (249th) cut site and type IIIlacked the 2205th (207th) cut positions respectively Type IIwas the rarest with its patterns observed to be ambiguous incertain rrn segments
Heterogenicity in few genomes was noted two rrn amongseven repeats ofW (NC 0176851 each in regular and comple-mentary strand) and KO11FL (NC 0176601 each in regularand complementary strand) and one in KO11FL (NC 0169021 regular) showed type II profile in spite of type I beingtheir primary pattern One among seven of the B strains REL606 (NC 0129671 regular) BL21-Gold (NC 0129471 com-plementary) BL21DE3 (NC 0129712 regular) and BL21DE3(NC 0128922 regular) showed type I profiles apart frombeing prevalent with type IV pattern Similarly two rrnamongNRG 873C (NC 0176341 each in regular and comple-mentary strand) and LF82 (NC 0119931 each in regular andcomplementary strand) and one in CFT073 (NC 0044311regular) showed type I pattern while exhibiting type IIIpattern predominance APEC01 (NC 0085631) being primein type III showed one rrn segment of type I and type IV(both in regular strand) E coli strain 042 (NC 0176261)displayed preponderance in type I pattern but three rrn (tworegular and one complementary) copies exhibited type IIIpattern One of the regular rrn elements in LF82 showed typeIV pattern The fragmentation pattern obtained by ARDRAof 23S P2
682with Hae III in the standard samples showed
that all K-12 C W Crooks and their respective descendantsexhibited type I profileswith type IVprofiles being exclusivelypresented by B strains (Figures 3(a) and 3(b))
Among the isolates 21 out of 22 commensal and 16 of18 clinical samples showed type I pattern heterogenicity ofpatterns was not observed These results reaffirmed predom-inance (93) of type I in nature Multisequence typing hasshown commensal strains to be categorized primarily undergroup A or group B1 whereas groups B2 D and E comprisegenerally pathogenic strains K-12 B and Crooks strain comeunder phylogroup A whereas strain W is a B1 phylogroup
34 Local Alignment of 23S rrn Hae III Profiles versus GlobalAlignment to Establish Clonality in E coli A number ofgenes that are of core dispensable and unique in naturemake up for pangenome the latter two are gained due torecombinational or transpositional events To obtain similarclustering on comparing whole genome would emphasizethe significance of these restriction profiles and help inestablishing the clonal nature and predominance of a singleclone Hence dendogram constructs with 682 bp ampliconfrom a representative of each restriction profile will showdistinct clustering of these profiles due to the fact that a smallsubset of segments within the whole genome was consideredThe global alignment of whole genome sequences (Figure 4)of the entire 57 E coli strains displayed distinct concentrateof type I III and IV profiles An intrinsic analysis of thespot of divergence and respective clade formations showedinteresting and riveting findings The strains at the ancestralroot showed type I pattern and the next divergent node point(D1) displayed formation of two distinct clans of type I andtype IV profiles segregated at the maximum distance fromone another Numbering of nodes was done by consideringthe precedence of branch point at every node A possiblescenario of two additional mutational events that are oftransitional nature (A to G) at 2205th and 2161st bases has
Journal of Pathogens 7
682bp
987654321
9998400
8998400
7998400
6998400
5998400
4998400
3998400
2998400
1998400
(a)
431bp
223 bp
210 bp
179bp
223 + 210 bp
223+
210+
87654321
8998400
7998400
6998400
5998400
4998400
3998400
2998400
1998400
(b)
Figure 3 (a) Broad range PCR of 23S P2682
with genomic DNA isolated from standard strains of E coli The upper row comprises Lane 1K-12 strains Lane 2 derivatives of K-12 strains Lane 3 DHI strain Lane 4 strain C1 Lane 6 Crookrsquos strain Lane 7 Walkerrsquos strain Lane 8descendent of W Lane 9 blank The lower row comprises Lane 11015840 B strain Lane 21015840 BL21 strain Lane 31015840 BL21DE3 Lane 41015840 BL21DE3Lys5(B3) Lane 61015840 E coli (ATCC 15223 commensal isolate) Lane 71015840 E coli (ATCC 11775 urine sample) Lane 81015840 E coli (ATCC 25922 clinical)Lanes 551015840 and Lanes 991015840 in both rows are 100 bp ladder and blank respectively (b) ARDRA of 23S P2
682
amplicons with Hae III Lane 1 ofboth rows comprises 100 bp ladder The digested products are loaded as per the order given in Figure 2(a)
introduced Hae III recognition patterns D6 gave lineages ofB1 phylogroups D9 giving rise to B2 clades (D12) of type Iprofiles and D clades with varied Hae III patterns (D11) D10gave rise to two nodes (with strains being type I groups)with one showing B1 phylogroups (D13) and the other beingpredominant in E phylogroups followed by B1 An almostexemplary corelation between the clonal groups and differentpathovars was found with this study Majority of commensalstrains and EIEC and all EAEC and EPEC were shown tobe having type I profile (Table 4) All UPEC ExPEC APECEIEC and single strain of EPEC and two commensal strainsshowed type III pattern B form strains along with a singlestrain of UPEC and environmental isolate exhibited type IVprofiles
Similarly a near-pristine association between phylo-groups and Hae III profiles was established Strains comingunder phylogroups A were preponderant in type I whereastype IV profiles were exclusively for B forms Type I profileswere observed in the strains (119899) in the order of A gt B1 gt E gtDwith type III dominant in B2 strains Phylogroup D showedequal occurrence of type III and type IV fragmentationpattern followed by a single strain with type I profile Theseresults assisted in enumerating a conjecture which works upa scenario of primordial microbial population where E colistrains had two clonal types (Figure 5(a)) which coexistedtogether (type I and type IV clones) Of these type I strainswould have been the predominant clone in comparison totype IV based on the dendogram data with maximum genepool of root strains showing type I Type III may have beenthe next predominant clone (an intermediary one) that hasevolved either from type I (by addition of a new recognition
Table 4 Corelation between phylogroups Hae III profiles andnature of E coli strains (genome data analysis of 57 E coli strainscollated from Genbank)
Strain Type A B1 B2 D E Total Out ofCommensal I 10 5 mdash mdash mdash 15 21Commensal III mdash mdash 2 mdash mdash 2 21Commensal IV 4 mdash mdash mdash mdash 4 21EPEC I mdash mdash mdash mdash 7 7 8EPEC III mdash mdash 1 mdash mdash 1 8EAEC I mdash 4 mdash 1 mdash 5 5EHEC I mdash 2 mdash mdash mdash 2 3EHEC II mdash 1 mdash mdash mdash 1 3ETEC I 1 1 1 mdash mdash 3 3UPEC III mdash mdash 7 mdash mdash 7 8UPEC IV mdash mdash mdash 1 mdash 1 8EIEC III mdash mdash 3 mdash mdash 3 3ExPEC III mdash mdash 2 2 mdash 4 4APEC III mdash mdash 1 mdash 1 1ENV IV mdash mdash mdash 1 1 1APEC avian pathogenic EAEC enteroaggressive EHEC enterohemorrhag-ic ETEC enterotoxigenic EIEC enteroinvasive EPEC enteropathogenicENV environmental ExPEC extraintestinal pathogenic UPEC uropatho-genic
site) or from type IV (by deletion of one recognition site)strains In a different scenario maybe all three types coexistedor type III would have given rise to type IVwith an additionalmutational event This situation may be hardly convincing
8 Journal of Pathogens
D1
D2
D3
D4
D5
D6
D7
D8
D9
D10
D11
D12
D13
D14
BL21(DE3)_1lowastBL21(DE3)_2lowast
BL21-Gold(DE3)pLysS AGlowast
B str REL606lowastA phylogroup
ETEC H10407lowast (B2)
UMNK88 (A)
(B1 clades) Str 2009EL-2050Str 2009EL-2071
Str 2011-349355989
B1 phylogroup
Str EC4115Str TW14359
Str EDL933Str Sakai
Xuzhou21Str CB9615
Str RM12598
E phylogroup
Str 11128Str 11368 B1 phylogroup
NA114
SE15LF82lowastlowastlowastStr NRG 857Clowastlowastlowast
UM146
UTI89IHE3034
APEC O1lowastlowastS88
Clone D i2Clone D i14
ABU 83972
CFT073536
ED1aStr E234869
B2 phylogroup
UMN026
IAI39Str CE10
SMS-3-5
D phylogroup
O42lowastlowastlowast
(B2 clades)
(D clades)
(B1 clades)
Wlowastlowastlowast
WKO11FLlowastlowast
KO11FLE24377A
SE11
B1 phylogroup
IAI1ATCC 8739
P12blowastHS
BW2952
K-12 substr DH10BDH1
DH1 (ME8569)K-12 substr W3110
K-12 substr MG1655
A phylogroup
1
2
10
3156
Str 12009lowastlowast
Commensal ETEC EHEC EPEC UPEC
ExPECAPECEAECEIECENV
Figure 4 Analysis of distribution of Hae III profiles phylogroups and pathovars within the global alignment dendogram constructcomprising 57 E coli genomes The dendogram of the whole genome sequence of 57 genomes was obtained from the NCBI web site forE coli and was redrawn using Photoshop CS6 for colour coordination for pathogenic and nonpathogenic strains and inputting the clonalgroups and phylogroups (httpwwwncbinlmnihgovgenometerm=escherichia+coli) Among the 57 genomes 32 were of type I (56)18 were of type III (31) and 6 and 1 were of type IV (10) and type II (2) respectively showing that type I is predominant and type IIthe rarest To correlate between the phylogroupsHae III profiles and pathovars the lineages were considered as clades derived from variousdivergence point (D) during the course of evolution
as dendogram segregation shows type III strains evolving orpresent after type IV strains The clustering also shows thatpredominance of type III is more than type IV and maybeevolutionary processesmutation have triggered the increasein type III profiles On similar line type II clones would havebeen evolved from type I (addition) or type IV (deletion)but due to the very limited type II restriction profile patterns(rare) as observed from the gene pool data the chances orapproximation of such event is difficult to prove Based onthese results no new insights into phylogroup distributionacross candidate strains can be provided however withrespect to the Hae III profile an additional angle can beconsidered Phylogroups A and B1 are predominant withtype I Hae III profiles whereas B2 phylogroups exclusivelyrepresented type III profile (Figure 5(b))
An intrinsic metagenomic and pangenomic analysis ofmost bacterial species exhibited mosaicisms of their inherentblueprint [39] In such situations core gene elements sharedacross the species are limited in comparison to dispensable(flexible) genetic contents that concede uniqueness to aparticular strain These flexible contents help in niche adapt-ability and give bacteria the ability to infect and invade Amajor concern is while diagnosing both true and opportunistpathogenic bacteria the former causing disease in virtuallyany susceptible host and the latter invoking diseases inimmunocompromised individuals ldquoMolecular barcodingrdquoan evolved term which implies the method of allele specificamplification of relatively small DNA sequences or barcodes[40] for spotting a particular species is a thrust area formicrobial diagnostics A plethora of functional genes ranging
Journal of Pathogens 9
Type I(28 459)
Type IV(28 207 249 and 459)
Hypothesis
Type III(28 249 and 459)
+ minus
+
Primordial
NonpathogenicA gt B1 gt B2
type I gt IV gt III
PathogenicB2 gt B1 gt E gt D
type I (IAT)III(P)gt type IVII
Predominant clones
Rarest
+minus
Type II(28 207 and 459)
Restriction profilesand
clonesPhylogroups and
clones
2∘ clonesintermediate
(a) (b)
Figure 5 (a) Hypothesis of clonality in E coli strains based on 23S rrn restriction profiling The primordial soup of E coli microbes wouldhave been comprised of two clonal groups types I and IV coexisting with each other but predominated by type I strains Over the courseof evolution on accumulating mutation as a means for variation type I or type IV would have given rise to type III by addition (former) ordeletion (latter) of recognition sequence The second scenario could be a rare event because accumulation of two mutations in a conservedregion would be difficult Type II being the rarest would have been evolved in similar basis to type III (b) Hypothesis of linking phylogroupsHae III profiles and clonal types in E coli This diagram illustrates the clonal groups of commensal E coli strains having both A phylogroups(dominant) and B1 phylogroups each being type I profiles Pathogenic counterparts showed dominancy in type III restriction pattern followedby type I pattern With respect to phylogroups pathovars were predominant in B2 and B1 followed by D and E groups
from housekeeping to repair are employed for characterizingbacteria of which rRNA is referred to as the gold stretch[41 42] Ribosomal segments organized as operon clusters of16S 23S and 5S rDNA and these assemblages are distributedfrom single to multiple copies across the bacterial genophore[43] The occurrence of redundancy in maintaining differentcopies of these genes is not associated with virulence but anin-built ecological strategy that contributes to the particularbacterium with a competitive edge and better chance ofsurvival [44] A futuristic approach of coupling the techniqueand results with antibiotic resistance pattern colony char-acteristics and genomic profiles of bacterial strains of bothcommensal and pathogenic nature can help in monitoringand managing infections in a better manner
4 Conclusion
The causatum of postgenomic era genomics and proteomicshas enhanced this progress by improvising the techniquesused for diagnostics The strategy of ARDRA utilized inthis study was rapid and a cost-effective approach to dif-ferentiate E coli from its colony morphovars along withgiving a predicament of nature of origin or clonal groupsThe mutations reported in this work for detection andestablishing clonality namely T874A (Bfa I) G2161A andG2205A (Hae III) have not been mentioned in any researchpaper or within the Ribosomal RNA Mutation Database
[45] making this finding aboriginal This basic methodologycan be adapted for detection of other bacterial organismsby targeting SNPs linked to recognition sequences therebyvalidating rrn sequences as marker for diagnostics A cumu-lative approach of detection of unique restriction pattern of atargeted gene and its cross-reference across bacterial familiescan circumvent changes of false positives
Disclosure
This work was presented (poster) in part at the 5th Interna-tional Barcoding of Life Kunming China 2013
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
Acknowledgments
The authors acknowledge and express their gratitude to Pro-fessor D A Bhiwgade (Ex-Director) and Professor DebjaniDasgupta Director School of Biotechnology amp Bioinformat-ics D Y Patil University as well as the management for theirsupport and for providing the infrastructure to carry out thecurrent work
10 Journal of Pathogens
References
[1] M Richter and R Rossello-Mora ldquoShifting the genomic goldstandard for the prokaryotic species definitionrdquo Proceedings ofthe National Academy of Sciences of the United States of Americavol 106 no 45 pp 19126ndash19131 2009
[2] A F Auch M von Jan H-P Klenk and M Goker ldquoDigitalDNA-DNA hybridization for microbial species delineation bymeans of genome-to-genome sequence comparisonrdquo Standardsin Genomic Sciences vol 2 no 1 pp 117ndash134 2010
[3] D H Martin M Nsuami J Schachter et al ldquoUse of multiplenucleic acid amplification tests to define the infected-patientlsquogold standardrsquo in clinical trials of new diagnostic tests forChlamydia trachomatis infectionsrdquo Journal of Clinical Microbi-ology vol 42 no 10 pp 4749ndash4758 2004
[4] E A Mothershed and A M Whitney ldquoNucleic acid-basedmethods for the detection of bacterial pathogens presentand future considerations for the clinical laboratoryrdquo ClinicaChimica Acta vol 363 no 1-2 pp 206ndash220 2006
[5] J E Clarridge III ldquoImpact of 16S rRNA gene sequence analysisfor identification of bacteria on clinical microbiology andinfectious diseasesrdquo Clinical Microbiology Reviews vol 17 no4 pp 840ndash862 2004
[6] O Mizrahi-Man E R Davenport and Y Gilad ldquoTaxonomicclassification of bacterial 16S rRNA genes using short sequenc-ing reads evaluation of effective study designsrdquo PLoS ONE vol8 no 1 Article ID e53608 2013
[7] D E Lebonah A Dileep K Chandrasekhar S Sreevani BSreedevi and J Pramoda Kumari ldquoDNA barcoding on bacteriaa reviewrdquo Advances in Biology vol 2014 Article ID 541787 9pages 2014
[8] J Rajendhran and P Gunasekaran ldquoMicrobial phylogeny anddiversity small subunit ribosomal RNA sequence analysis andbeyondrdquo Microbiological Research vol 166 no 2 pp 99ndash1102011
[9] R Srinivasan U Karaoz M Volegova et al ldquoUse of 16S rRNAgene for identification of a broad range of clinically relevantbacterial pathogensrdquo PLoS ONE vol 10 no 2 Article IDe0117617 2015
[10] S Chakravorty D Helb M Burday N Connell and D AllandldquoA detailed analysis of 16S ribosomal RNA gene segments forthe diagnosis of pathogenic bacteriardquo Journal of MicrobiologicalMethods vol 69 no 2 pp 330ndash339 2007
[11] C R Woese ldquoReview bacterial evolutionrdquo MicrobiologicalReviews vol 51 pp 221ndash271 1987
[12] M Touchon C Hoede O Tenaillon et al ldquoOrganised genomedynamics in the Escherichia coli species results in highly diverseadaptive pathsrdquo PLoS Genetics vol 5 no 1 Article ID e10003442009
[13] J M Janda and S L Abbott ldquo16S rRNA gene sequencingfor bacterial identification in the diagnostic laboratory plusesperils and pitfallsrdquo Journal of Clinical Microbiology vol 45 no9 pp 2761ndash2764 2007
[14] J-J Lu C-L Perng S-Y Lee and C-CWan ldquoUse of PCRwithuniversal primers and restriction endonuclease digestions fordetection and identification of common bacterial pathogens incerebrospinal fluidrdquo Journal of Clinical Microbiology vol 38 no6 pp 2076ndash2080 2000
[15] N M Guentzel ldquoEscherichia Klebsiella Enterobacter SerratiaCitrobacter andProteusrdquo inMedicalMicrobiology S Baron Edchapter 26 University of Texas Medical Branch at GalvestonGalveston Tex USA 1996
[16] P Behzadip E Behzadi H Yazdanbod R Aghapour A MCheshmeh and S D Omran ldquoA survey on urinary tract infec-tions associated with the three most common uropathogenicbacteriardquoMaedica (Buchar) vol 5 no 2 pp 111ndash115 2010
[17] B CMetri P Jyothi andBV Peerapur ldquoAntibiotic resistance inCitrobacter spp isolated from urinary tract infectionrdquo UrologyAnnals vol 5 no 4 pp 312ndash313 2013
[18] N Taneja S S Chatterjee M Singh S Singh and M SharmaldquoPediatric urinary tract infections in a tertiary care center fromnorth Indiardquo Indian Journal of Medical Research vol 131 no 1pp 101ndash105 2010
[19] J B Kaper J P Nataro and H L T Mobley ldquoPathogenicEscherichia colirdquo Nature Reviews Microbiology vol 2 no 2 pp123ndash140 2004
[20] E J de Muinck K Lagesen J E Afset et al ldquoComparisonsof infant Escherichia coli isolates link genomic profiles withadaptation to the ecological nicherdquo BMC Genomics vol 14 no1 article 81 2013
[21] D A Rasko M J Rosovitz G S A Myers et al ldquoThepangenome structure of Escherichia coli comparative genomicanalysis of E coli commensal and pathogenic isolatesrdquo Journalof Bacteriology vol 190 no 20 pp 6881ndash6893 2008
[22] C T Archer J F Kim H Jeong et al ldquoThe genome sequenceof E coli W (ATCC 9637) comparative genome analysis andan improved genome-scale reconstruction of E colirdquo BMCGenomics vol 12 article no 9 2011
[23] PKuhnert J Nicolet and J Frey ldquoRapid and accurate identifica-tion of Escherichia coliK-12 strainsrdquoApplied and EnvironmentalMicrobiology vol 61 no 11 pp 4135ndash4139 1995
[24] A P Bauer S M Dieckmann W Ludwig and K-H SchleiferldquoRapid identification of Escherichia coli safety and laboratorystrain lineages based on multiplex-PCRrdquo FEMS MicrobiologyLetters vol 269 no 1 pp 36ndash40 2007
[25] B J Bachmann ldquoPedigrees of some mutant strains ofEscherichia coli K-12rdquo Bacteriological Reviews vol 36 no 4 pp525ndash557 1972
[26] P Daegelen F W Studier R E Lenski S Cure and J F KimldquoTracing ancestors and relatives of Escherichia coli B and thederivation of B strains REL606 and BL21(DE3)rdquo Journal ofMolecular Biology vol 394 no 4 pp 634ndash643 2009
[27] F W Studier P Daegelen R E Lenski S Maslov and J F KimldquoUnderstanding the differences between genome sequences ofEscherichia coli B strains REL606 and BL21(DE3) and compar-ison of the E coli B and K-12 genomesrdquo Journal of MolecularBiology vol 394 no 4 pp 653ndash680 2009
[28] R K Selander and B R Levin ldquoGenetic diversity and structurein Escherichia coli populationsrdquo Science vol 210 no 4469 pp545ndash547 1980
[29] D M Gordon O Clermont H Tolley and E DenamurldquoAssigning Escherichia coli strains to phylogenetic groupsmulti-locus sequence typing versus the PCR triplex methodrdquoEnvironmental Microbiology vol 10 no 10 pp 2484ndash24962008
[30] P Desjardins B Picard B Kaltenbock J Elion and EDenamur ldquoSex in Escherichia coli does not disrupt the clonalstructure of the population evidence from random amplifiedpolymorphic DNA and restriction-fragment-length polymor-phismrdquo Journal of Molecular Evolution vol 41 no 4 pp 440ndash448 1995
Journal of Pathogens 11
[31] N D Jasuja R Saxena S Chandra and S C Joshi ldquoIsolationand identification of microorganism from polyhouse agricul-ture soil of RajasthanrdquoAfrican Journal ofMicrobiology Researchvol 7 no 41 pp 4886ndash4891 2013
[32] V Hemraj S Diksha and G Avneet ldquoA review of commonlyused biochemical test for bacteriardquo Innovare Journal of LifeSciences vol 1 pp 1ndash7 2013
[33] J D Thompson D G Higgins and T J Gibson ldquoCLUSTALWimproving the sensitivity of progressive multiple sequencealignment through sequence weighting position-specific gappenalties and weight matrix choicerdquoNucleic Acids Research vol22 no 22 pp 4673ndash4680 1994
[34] T Vincze J Posfai and R J Roberts ldquoNEBcutter a program tocleave DNA with restriction enzymesrdquo Nucleic Acids Researchvol 31 no 13 pp 3688ndash3691 2003
[35] J Ye G Coulouris I Zaretskaya I Cutcutache S Rozen andT Madden ldquoPrimer-BLAST a tool to design target-specificprimers for polymerase chain reactionrdquo BMC Bioinformaticsvol 13 article 134 2012
[36] J R Parvathi and S Singh ldquoFacile approaches for microbialDNA extraction comparative study with colonies lsquopickedrsquo fromthree different bacteriological mediumrdquo Journal of Pure andApplied Microbiology vol 9 no 3 pp 2373ndash2378 2015
[37] R Lan and P R Reeves ldquoEscherichia coli in disguise molecularorigins of Shigellardquo Microbes and Infection vol 4 no 11 pp1125ndash1132 2002
[38] D J Leininger J R Roberson and F Elvinger ldquoUse ofeosin methylene blue agar to differentiate Escherichia coli fromother gram-negative mastitis pathogensrdquo Journal of VeterinaryDiagnostic Investigation vol 13 no 3 pp 273ndash275 2001
[39] U Dobrindt F Agerer K Michaelis et al ldquoAnalysis of genomeplasticity in pathogenic and commensal Escherichia coli isolatesby use of DNA arraysrdquo Journal of Bacteriology vol 185 no 6pp 1831ndash1840 2003
[40] V Savolainen R S Cowan A P Vogler G K Roderickand R Lane ldquoTowards writing the encyclopaedia of life anintroduction to DNA barcodingrdquo Philosophical Transactions ofthe Royal Society B Biological Sciences vol 360 no 1462 pp1805ndash1811 2005
[41] T A Isenbarger C E Carr S S Johnson et al ldquoThe mostconserved genome segments for life detection on earth andother planetsrdquo Origins of Life and Evolution of Biospheres vol38 no 6 pp 517ndash533 2008
[42] S R Santos and H Ochman ldquoIdentification and phylogeneticsorting of bacterial lineages with universally conserved genesand proteinsrdquoEnvironmentalMicrobiology vol 6 no 7 pp 754ndash759 2004
[43] R Rastogi M Wu I DasGupta and G E Fox ldquoVisualizationof ribosomal RNA operon copy number distributionrdquo BMCMicrobiology vol 9 no 1 article 208 2009
[44] J A Klappenbach J M Dunbar and T M Schmidt ldquorRNAoperon copy number reflects ecological strategies of bacteriardquoApplied and Environmental Microbiology vol 66 no 4 pp1328ndash1333 2000
[45] J A Klappenbach P R Saxman J R Cole and T M SchmidtldquoRrndb the ribosomal RNA operon copy number databaserdquoNucleic Acids Research vol 29 no 1 pp 181ndash184 2001
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom

2 Journal of Pathogens
This current study is an endeavor to entitle 23S rRNAgeneas substitute to de facto barcode 16S rRNA gene using ampli-fication ensued by restriction digestion instead of sequencingmaking it a cost-effective approach The exemplification ofthe proposed method was carried out to detect and differ-entiate E coli (candidate) from E aerogenes K pneumoniaeandC koseri (noncandidates)The above choice was finalizedafter considering the following (i) all the selected microbesboth candidate and noncandidate are Gram negative lactosepositive microbes of 120574-Enterobacteriaceae family [15] (ii)all are urinary tract infection (UTI) pathogens with E colibeing reported as the major causative factor [16ndash18] andlastly (iii) all exhibit similar colony semblance and any falsepositive and negative biochemical characterization resultingin misinterpretation with candidate strain
In spite of being the model representative of bacterialkingdom E coli is also acknowledged for its omnipresenceand the capability of switching from good to bad microbePathogenic E coli are classified into aggressive invasivepathogenic hemorrhagic and toxigenic which differs in theirvirulence genes acquired and the type of pathogenicity [19]Comparative study of commensal and pathogenic strainsof the same has shown a rather surprising result of fewerpathovar specific sequences evincing E coli as a genera list[12] Sharing of 40 of core genomes among both virulentand nonvirulent strains indicates clonality [20] This con-ceptualizes an open pangenome having genetic reservoirs toallow gene acquisition for evolving into pathogenic ones [21]
Besides isolates of E coli present in nature the routinelyused nonvirulent lab strains are K-12 B W Crooks and Cwith all being fecal in origin except for strain W (soil isolate)[22] Method of rapid identification of K-12 strains usingmutations in O-antigen gene cluster [23] and determinationof lab strains lineage using multiplex primers [24] have beenreported Studies on the lineages of well-established E colilab strains across the globe have shown that most strainsare derived from either K-12 or B [25 26] Genomes ofthese supposed to be parental strains show 99 geneticidentity with exceptions to IS elements and flagellar genes[27] Phylogenetic classification of E coli strains into A B1B2 D and E groups usingMultiloci Enzyme ElectrophoresisMLEE [28] and triplex [29] has given insights into theconcept of clonality in E coli A comparative analysis ofMLEE and randomamplification restriction profiling against16S-23S intertranscribed region and ribotyping have verifiedthat horizontal gene transfer mechanisms do not disruptorganization of clonal population [30]
No study till date has correlated Amplified RibosomalDNA Restriction Analysis (ARDRA) of a core segment in23S rRNA gene against the pangenome and tried to establisha link between phylogroups pathogroups and restrictiongroups
2 Materials and Methods
21 Bacterial Strains and Culture Conditions Authenticationof the molecular assay in screening and identifying the can-didate organism was executed into parts one with standardstrains of E coli and the other with 25 isolates each from fecal
clinical and environmental origin Lyophilized strains wereprocured from Microbial Type Culture Collection (MTCC)Institute of Microbial Technology (IMTECH ChandigarhIndia) for both identification and clonality studies (Table 1)
Five different sets of fecal droppings from rat chickenpigeon rabbit (pet shops Crawford market Mumbai India)and humans (Excel Pathology LabVashi NaviMumbai) wereconsidered for isolation of commensal samples The strainswere revived in nutrient broth at 37∘C For environmentalstrains water sample from ten local water bodies lying in andaround the parent institute was utilized (areas with hospitalsor health care centers in the vicinity were avoided to preventcross-contamination with clinical samples) For isolating Ecoli of clinical origin urine samples from twenty-five patientssuffering from UTI (Excel Pathology Lab) were considered
The samples were processed within 24 hrs of collection(either in zip log bags or in sterile glass bottle) onMacConkeyagar (MAC) The distinct pink colonies were restreaked ontoboth MAC and Eosin Methylene Blue (EMB) agar respec-tively [31] Isolates giving discrete colony morphology withindeterminate greenish sheen were selected and evaluated forthe false positive and negatives by both microbiochemicaland molecular assays Each of the selected colonies waspicked with sterile wooden toothpicks and transferred to fivemL of nutrient broth and incubated overnight at 37∘CThesesamples were further characterized [32] by IMViC 1mL wasutilized for DNA isolation and the remaining was used forpreparing glycerol stock (25 glycerol) for future reference
22 Genome Database and Microheterogenicity Analysis Theaboriginal step involved the aggregation of Genbank genomesequences of all the strains that were considered for this studyfrom the public data repository National Centre for Biotech-nology Information (NCBI) The data pool (until 11 April2013) comprised an assemblage of 61 completed genomes(Supplementary Data A1 in Supplementary Material avail-able online at httpdxdoiorg1011552015562136) with 57of candidate strains (E coli) and noncandidate strains (Kpneumoniae C koseri and E aerogenes) The following stepswere executed to analyze the microheterogenicity (Figure 1)which can directly assist in evaluating the electability of 23SrRNA gene as an ideal barcode
Two strains one from the candidate and one from non-candidate genome (E coli str K-12 MG1655 (NC 0009132)C koseri ATCC BAA-895 (NC 0097921) E aerogenes(NC 0156631) and K pneumoniae subsp pneumoniaeMGH78578 (NC 0096481)) were selected and assigned as refer-ence strain
For studying intergenic variability among the candidateand noncandidate strains the rrsH (16S RNA gene) and rrlH(23S rRNA gene) sequence on the regular strand (referencestrains) were extrapolated and multiple sequence alignment(MSA) using ClustalW [33] was executed Total mismatchwas enumerated by the addition of number of mismatchesand sequence gaps introduced during the ldquoline-uprdquo of twoor more sequences The mismatch percentage was calculatedby dividing the total mismatches seen across the four strainsby the average base pair (bp) for both 16S and 23S rrn generespectively
Journal of Pathogens 3
Table 1 List of standard strains used in this study
Strains MTCC ATCC NatureBarcodingidentification studies
Noncandidate strainsCitrobacter freundii 1658 8090 ClinicalCitrobacter koseri 1657 27028 Blood cultureKlebsiella pneumonia 432 33495 Urinary cultureEnterobacter aerogenes 111 13048 Sputum
Candidate strains433 15228 Commensal
Escherichia coli 4296 19138 Urine448 25922 Clinical sample
Diversityclonality studies
Escherichia coli
1302 mdash K122622 10798 Descendent of K121667 mdash DH11304 mdash C11687 8739 Crooks strain448 9637 W2692 53846 Descendent of W1303 mdash B1678 mdash BL211679 mdash BL21DE31680 mdash BL21DE3Lys5
E coli namely K-12 B C Crooks andW are commonly used as lab strains Whole genomic data of these strains except strain C is available (NCBI Genomes)
Candidate microbe
Noncandidate microbe
Escherichia coli
Klebsiella pneumoniae (2 G)Enterobacter aerogenes (1 G)
Citrobacter koseri (1 G)
sequences from each genome
57 genomes (G) (21 pathogenic
36 nonpathogenic)
Estimation of diversity (multiple sequence alignment MSA)Intergenic diversity
representative genome Intragenic diversity
Restriction profiling for differentiating candidate
from noncandidates (NEB V20)
85 restriction groups(Suppl A2)
Virtual digestion was individually
carried out on each rrlH
Similar restriction patterns(rejected) Dissimilar restriction patterns
(selected)
Restriction patterns were checked in all the 430 ribosomal segments
Unique profile in candidate alone
(barcoding)Bfa I was chosen
Polymorphic within candidate genome
(clonality)Hae III was chosen
A total of 430 sequences considering both candidate and noncandidate genomes
(ii) Regular strands of eachgenome
(i) Complementary strands ofeach genome
rrlH from each
Extraction of 23S rrn
Figure 1 Schematic representation of methodology for detecting barcode and estimating clonality The primers are designed against theregions that flank these restriction sequences which gives unique bands revealing identification or clonality
For estimating the intragenic variability all the riboso-mal segments from 57 E coli genomes were extracted (7copiesgenome) complementary and regular strands withineach strain were segregated and the percentage mismatchesfor each of the genomes were determined
23 Virtual Restriction Profiling Virtual restriction profilesusing NEB V20 [34] with 85 restriction groups (Supplemen-tary Data A2) were performed for a total of 430 ribosomalsegments that included 399 from 57 E coli genomes and7 8 and 16 rrn sequences from C koseri E aerogenes
4 Journal of Pathogens
Table 2 Restriction site of Bfa I (CTAG) within 23S rrlH of candidate and noncandidates and the restriction profiles with 23S P1880
Restriction profile within rrlH Fragments obtained on ARDRAF E Ci K Ea Mic Fragments (amplicon size)11 687 685 685 685 E 665 bp 215 bp (880 bp)12 874 874 874 C 475 bp 214 bp and 189 bp (878 bp)13 1670 1669 1667 1670 K 475 bp 214 bp and 189 bp (877 bp)14 2336 2339 Ea 475 bp 214 bp and 189 bp (876 bp)15 2652 2653 2650 2654Primers were designed against the regions flanking the bold restriction sitesF number of fragments E Escherichia coli Ci Citrobacter koseri K Klebsiella pneumoniae Ea Enterobacter aerogenes Mic microbe23S P1
880-FP(+) 51015840-GGCGAAAAGAACCCCGGCGA-31015840 (20 nt) 23S P1
880-RP(minus) 51015840-AGGGGTCGACTCACCCTGCC-31015840 (20 nt)
FP forward primer RP reverse primer The primer details were obtained from the results of primer blast and restriction profiling pattern was obtained fromNEB V20 Forward and reverse primers of 23S P1
880lie within domains I and III of 23S rRNA secondary structure respectively with Bfa I sites in domain II
Table 3 Clonal groups based on Hae III (GGCC) profiles within the candidate microbe using 23S P2682
Virtual digest pattern Types of Hae III restriction pattern observed amongst 399 rrn sequences of E coli genomesF C Type I Type II Type III Type IV11 785 Cut site within rrlH of candidate rrn segments12 1042 1984 1984 1984 198413 1116 x 2163 X 216314 1217 x X 2205 220515 1361 2415 2415 2415 241516 1445 Cut site within the amplicon (682 bp)17 1907 28 28 28 2818 1984 x 207 X 20719 2161 x X 249 249110 2205 459 459 459 459111 2415 Fragments obtained for each group
112 2839 28 bp 223 bp and 431 bp 28 bp 179 bp 223 bp and252 bp
28 bp 210 bp 221 bp and223 bp
28 bp 42 bp 179 bp210 bp and 223 bp
Primers were designed against the regions flanking the bold restriction sites F number of fragments C cut sites23S P2
682-FP(+) 51015840CCGACCTGCACGAATGGCGT31015840 (20 nt)
23S P2682
-RP(+) 51015840CAGTTCTCCAGCGCCCACGG31015840 (20 nt)Forward and reverse primers of 23S P2
682lie within domain IV and transition of V-VI of 23S rRNA secondary structure respectively First cut sites lie within
domain IV whereas the rest lie within domain V
and K pneumoniae strains respectively Among the 14common cutters groups (Supplementary Data A3) Bfa I(Supplementary Data A4) was selected as the enzyme forbarcoding as it gave conserved and distinct pattern acrossall the 399 rrn sequences (not shown) in differentiating thecandidate from the noncandidate strains The uniquenessand position of the cuts sites for giving unique bands onagarose gel electrophoresis were also a lead reason consideredfor finalizing this enzyme Hae III displaying polymorphismpattern with 23S rDNA segments of candidate microbe wasfinalized for diversity studies (Figure 1)
24 Primers and Restriction Enzymes for Addressing Identityand Clonality Sequence of rrlH of E coli K-12 str MG1655was used as the template for designing broad range primersusing Primer-BLAST [35] In conjunction with parameterslike GC content 119879
119898value and the amplicon size number of
restriction sites within the amplicon and the size of fragmentafter digestion were kept as the benchmark for finalizing
primersThere are twoprimer sets 23S P1880
[Probe 31778738]was selectedwithBfa I restriction digestion (Table 2) demarkingthe candidate and 23S P2
682[Probe 31781046] (Table 3) with
polymorphicHae III restriction siteswas selected for address-ing clonality Possibility of self-priming andprimer-dimer for-mation of each primer was checked using free web tools Oli-gocal (httpwwwsimgenecomOligoCalc) OligoEvaluator(httpwwwsigmaaldrichcomtechnical-documentsarticlesbiologyoligo-evaluatorhtml) and multiple primer analyzertool (httpwwwthermoscientificbiocomwebtoolsmultiple-primer) maintained by SimGene Sigma and Fermentasresp
BLAST genome (Supplementary Data) assisted in veri-fying the conservation and specificity of primers ampliconsize Bfa I recognition sites and region of hybridizationacross the genome of 931 bacterial families After omitting thenonannotated sequences all of the completed whole genomedata aggregated under phyla Actinobacteria BacteroidetesChlamydiae Cyanobacteria Firmicutes Proteobacteria and
Journal of Pathogens 5
Spirochaetae of the bacterial kingdom were considered forthe analysis
25 DNA Isolation Amplification and Restriction DigestionDNA isolation was performed according to Parvathi andSingh [36] The reaction soup comprised 2120583L of supernatant(DNA) 05 120583M of each forward and reverse primer 1x ofassay buffer (10mM Tris-HCl pH 88 50mM KCl) MgCl
2
(15mM) dNTPmix (800 120583M) and 05U120583LTaq polymerase(Kappa Systems KK5004) Standard genomic DNA of Ecoli K-12 (Merck) and nuclease-free water were used aspositive and negative control The reactions were carriedout in a thermal cycler (Bioer) with the program set for25 cycles comprising denaturation (95∘C) annealing (68∘C)and extension (72∘C) for 30 s each 3 120583L of the PCR productwas digested with 2U of respective enzyme (FermentasThermo Scientific) in 5120583L reactionmix at 37∘C for two hoursThe amplicons and their restriction products were analysedby agarose gel electrophoresis (AGE) at 5Vcm in 2 and25 agarose gels respectively each containing 05120583gmL ofEtBr The gels were viewed and photographed in BioImagingSystems (Syngene)
3 Result and Discussion
31 Genetic Diversity within 23S Ribosomal Repeats for Identi-fication of E coli 23S rRNA gene showed 087 intergenicvariations compared with 16S rRNA within the referencegenomes (Supplementary Data A5) The result seemed obvi-ous due to the increased frame size of the latter but thisalso encouraged perusing microheterogenicity analysis ofthe largest ribosomal segment for further approach Thesizes of target segments varied from 2903 to 2907 bp withmaximum gene length observed in ATCC8739 (Crookrsquosstrain) accounting for 2970ndash2985 bp followed by E24377Adisplaying sequences of 2923ndash2925 bp One rrn of UM 146and ETECH10407 were of 1762 bp and 1561 bp respectively(Supplementary Data A9) Absolutely no link between rrnsize and pathogenicity could be defined because the varia-tions were random ATCC8739 being a laboratory cultureof fecal origin and UM146 an EIAC strain E23477A andETECH10407 of ETEC origin
Ribosomal genes were found to be distributed acrosscomplementary and regular strand with majority of thegenomes (46 out of 57) showing segregation in five regularand two complementary strands All the 23S rrn sequencesdistributed within the regular and complementary strandsacross respective genomes were individually sorted for MSAto attain an understanding of intragenic variation Analysisrevealed the highest level of mismatches in complementarystrands compared with their counterparts (SupplementaryData A6) The highest level of mismatches mounting to06 corresponding to 50 of the targeted gene size wasperceived in complementary strands of P12b (commensal)E23489 (EPEC) and ETEC H10407 The greatest dispar-ity among sequences in regular strand was observed inAPEC 01 with 003 (882903) followed by CFT043 (UPEC)and 042 (EAEC) having 0016 (sim42ndash512903) dissimilarity23S ribosomal copies of ED1a (commensal) showed zero
percent mismatches in both complementary and regularstrands making it an ideal cluster with the highest degreeof conservation Similar values of mismatches were seen inSE11 (Commensal) CB9615 (EPEC) and 0104H4 2009El-2011 (EAEC) These results indicate that there is no cor-relation between intragenic diversity of target stretch andpathogenicity of strains These differences substantiated 23SRNA clusters having microdiverse organization with bothconserved and variable stretches within them Sequevarsarising due to this heterogeneity unavoidably lead to biasedresults differential amplification and chimeric moleculesThus the method of screening and targeting restriction pro-file pattern within a region of choice (ribosomal segments)and specific to bacteria of interest demonstrates authentic andsimpler practice than routine molecular strategy involvingamplification and sequencing
32 Barcoding E coli from Its Colony Morphovars ARDRAof 23S P1880 with Bfa I PCR amplification of the stan-dard strains and isolates of different origin (commensalurine and clinical sample) with 23S P1
880gave amplicons
(880 bp) of high intensity with no primer-dimer formation(Figure 2(a)) Restriction digestion of the amplicons withBfa I gave unique fragments of 665 bp and 215 bp in E coliwhereas C koseri E aerogenes and K pneumoniae eachgave fragments of 475 bp 214 bp and 189 bp respectively(Figure 2(b)) The genotypic strategy proved superior tobiochemical characterization taking into consideration all thepossible positive negative and false positive and negativeresults across 25 isolates each from clinical commensal andenvironmental origin (Supplementary Data A7) 18 (72)of clinical 22 (88) of commensal and all the 25 (100)environment isolates showed distinct restriction pattern likethat of E coli BLAST genome search across 931 bacterialfamilies verified the specificity of primers and restrictionpreference for Bfa I The primers showed exclusive bindingonly with Enterobacteriaceae family (Supplementary DataA8) Among the various strains in the family only Shigellaspp Salmonella enterica (Arizona) and Proteus mirabilusexhibited similar restriction preference to that of E coli Inspite of Shigella spp being taxa in disguise of E coli [37] theycan be easily screened using selective medium because of nolactose fermentation [38] whereas Salmonella enterica andProteus mirabilis showed amplicon heterogeneity of 1073 bpand 880 bp during amplification thus making it easy to avoiderroneous conclusions and misinterpretation
33 Categorizing E coli StrainsUsingARDRAof 23S P2682 withHae III A detailed analysis of the Hae III profile across 399sequences [Supplementary Data A9] of ribosomal segmentsof E coli indicated four possible profiles types I II IIIand IV with varying degree of predominance within thesesegments The rest of the candidate strains showed type IIIpatterns but types I and IV were exclusive for E coli strainsMajority of genomes (119899 = 32) as in 15 nonpathogenic and17 pathogenic strains showed type I profiles (57) followedby type III (35) predominating in 15 strains Six strainsshowed type IV pattern (10) of which four were B strainsType I groups represent E coli strains that lacked recognition
6 Journal of Pathogens
1 7
880 bp
65432
(a)
8
665bp
475bp
215 bp 214 bp
178bp
1 765432
(b)
Figure 2 (a) Broad Range PCR of 23S P1880
with genomic DNA isolated from standard strains Lane 1 100 bp ladder Lane 2 blank Lane 3E coli Lane 4 C freundii Lane 5 C koseri Lane 6 K pneumonia Lane 7 E aerogenes PCR product of single representative of E coli wasloaded (b) ARDRA of 23S P1
880
amplicons with Bfa I Lanes 1 2 and 3 E coli (ATCC15223 ATCC25922 and ATCC11775) Lane 4C freundii(ATCC8090) Lane 5 C koseri (ATCC27028) Lane 6 K pneumonia (ATCC33495) Lane 7 E aerogenes (ATCC13048) Lane 8 100 bp ladder
elements due to change at 2205th and 2161st base with respectto 23S rDNA or with reference to amplicons at 207th or 249thbase positions Type IV strains are contrary to type I withintact recognition sequence at both of these sites Strains withtype II profiles lacked 2161st (249th) cut site and type IIIlacked the 2205th (207th) cut positions respectively Type IIwas the rarest with its patterns observed to be ambiguous incertain rrn segments
Heterogenicity in few genomes was noted two rrn amongseven repeats ofW (NC 0176851 each in regular and comple-mentary strand) and KO11FL (NC 0176601 each in regularand complementary strand) and one in KO11FL (NC 0169021 regular) showed type II profile in spite of type I beingtheir primary pattern One among seven of the B strains REL606 (NC 0129671 regular) BL21-Gold (NC 0129471 com-plementary) BL21DE3 (NC 0129712 regular) and BL21DE3(NC 0128922 regular) showed type I profiles apart frombeing prevalent with type IV pattern Similarly two rrnamongNRG 873C (NC 0176341 each in regular and comple-mentary strand) and LF82 (NC 0119931 each in regular andcomplementary strand) and one in CFT073 (NC 0044311regular) showed type I pattern while exhibiting type IIIpattern predominance APEC01 (NC 0085631) being primein type III showed one rrn segment of type I and type IV(both in regular strand) E coli strain 042 (NC 0176261)displayed preponderance in type I pattern but three rrn (tworegular and one complementary) copies exhibited type IIIpattern One of the regular rrn elements in LF82 showed typeIV pattern The fragmentation pattern obtained by ARDRAof 23S P2
682with Hae III in the standard samples showed
that all K-12 C W Crooks and their respective descendantsexhibited type I profileswith type IVprofiles being exclusivelypresented by B strains (Figures 3(a) and 3(b))
Among the isolates 21 out of 22 commensal and 16 of18 clinical samples showed type I pattern heterogenicity ofpatterns was not observed These results reaffirmed predom-inance (93) of type I in nature Multisequence typing hasshown commensal strains to be categorized primarily undergroup A or group B1 whereas groups B2 D and E comprisegenerally pathogenic strains K-12 B and Crooks strain comeunder phylogroup A whereas strain W is a B1 phylogroup
34 Local Alignment of 23S rrn Hae III Profiles versus GlobalAlignment to Establish Clonality in E coli A number ofgenes that are of core dispensable and unique in naturemake up for pangenome the latter two are gained due torecombinational or transpositional events To obtain similarclustering on comparing whole genome would emphasizethe significance of these restriction profiles and help inestablishing the clonal nature and predominance of a singleclone Hence dendogram constructs with 682 bp ampliconfrom a representative of each restriction profile will showdistinct clustering of these profiles due to the fact that a smallsubset of segments within the whole genome was consideredThe global alignment of whole genome sequences (Figure 4)of the entire 57 E coli strains displayed distinct concentrateof type I III and IV profiles An intrinsic analysis of thespot of divergence and respective clade formations showedinteresting and riveting findings The strains at the ancestralroot showed type I pattern and the next divergent node point(D1) displayed formation of two distinct clans of type I andtype IV profiles segregated at the maximum distance fromone another Numbering of nodes was done by consideringthe precedence of branch point at every node A possiblescenario of two additional mutational events that are oftransitional nature (A to G) at 2205th and 2161st bases has
Journal of Pathogens 7
682bp
987654321
9998400
8998400
7998400
6998400
5998400
4998400
3998400
2998400
1998400
(a)
431bp
223 bp
210 bp
179bp
223 + 210 bp
223+
210+
87654321
8998400
7998400
6998400
5998400
4998400
3998400
2998400
1998400
(b)
Figure 3 (a) Broad range PCR of 23S P2682
with genomic DNA isolated from standard strains of E coli The upper row comprises Lane 1K-12 strains Lane 2 derivatives of K-12 strains Lane 3 DHI strain Lane 4 strain C1 Lane 6 Crookrsquos strain Lane 7 Walkerrsquos strain Lane 8descendent of W Lane 9 blank The lower row comprises Lane 11015840 B strain Lane 21015840 BL21 strain Lane 31015840 BL21DE3 Lane 41015840 BL21DE3Lys5(B3) Lane 61015840 E coli (ATCC 15223 commensal isolate) Lane 71015840 E coli (ATCC 11775 urine sample) Lane 81015840 E coli (ATCC 25922 clinical)Lanes 551015840 and Lanes 991015840 in both rows are 100 bp ladder and blank respectively (b) ARDRA of 23S P2
682
amplicons with Hae III Lane 1 ofboth rows comprises 100 bp ladder The digested products are loaded as per the order given in Figure 2(a)
introduced Hae III recognition patterns D6 gave lineages ofB1 phylogroups D9 giving rise to B2 clades (D12) of type Iprofiles and D clades with varied Hae III patterns (D11) D10gave rise to two nodes (with strains being type I groups)with one showing B1 phylogroups (D13) and the other beingpredominant in E phylogroups followed by B1 An almostexemplary corelation between the clonal groups and differentpathovars was found with this study Majority of commensalstrains and EIEC and all EAEC and EPEC were shown tobe having type I profile (Table 4) All UPEC ExPEC APECEIEC and single strain of EPEC and two commensal strainsshowed type III pattern B form strains along with a singlestrain of UPEC and environmental isolate exhibited type IVprofiles
Similarly a near-pristine association between phylo-groups and Hae III profiles was established Strains comingunder phylogroups A were preponderant in type I whereastype IV profiles were exclusively for B forms Type I profileswere observed in the strains (119899) in the order of A gt B1 gt E gtDwith type III dominant in B2 strains Phylogroup D showedequal occurrence of type III and type IV fragmentationpattern followed by a single strain with type I profile Theseresults assisted in enumerating a conjecture which works upa scenario of primordial microbial population where E colistrains had two clonal types (Figure 5(a)) which coexistedtogether (type I and type IV clones) Of these type I strainswould have been the predominant clone in comparison totype IV based on the dendogram data with maximum genepool of root strains showing type I Type III may have beenthe next predominant clone (an intermediary one) that hasevolved either from type I (by addition of a new recognition
Table 4 Corelation between phylogroups Hae III profiles andnature of E coli strains (genome data analysis of 57 E coli strainscollated from Genbank)
Strain Type A B1 B2 D E Total Out ofCommensal I 10 5 mdash mdash mdash 15 21Commensal III mdash mdash 2 mdash mdash 2 21Commensal IV 4 mdash mdash mdash mdash 4 21EPEC I mdash mdash mdash mdash 7 7 8EPEC III mdash mdash 1 mdash mdash 1 8EAEC I mdash 4 mdash 1 mdash 5 5EHEC I mdash 2 mdash mdash mdash 2 3EHEC II mdash 1 mdash mdash mdash 1 3ETEC I 1 1 1 mdash mdash 3 3UPEC III mdash mdash 7 mdash mdash 7 8UPEC IV mdash mdash mdash 1 mdash 1 8EIEC III mdash mdash 3 mdash mdash 3 3ExPEC III mdash mdash 2 2 mdash 4 4APEC III mdash mdash 1 mdash 1 1ENV IV mdash mdash mdash 1 1 1APEC avian pathogenic EAEC enteroaggressive EHEC enterohemorrhag-ic ETEC enterotoxigenic EIEC enteroinvasive EPEC enteropathogenicENV environmental ExPEC extraintestinal pathogenic UPEC uropatho-genic
site) or from type IV (by deletion of one recognition site)strains In a different scenario maybe all three types coexistedor type III would have given rise to type IVwith an additionalmutational event This situation may be hardly convincing
8 Journal of Pathogens
D1
D2
D3
D4
D5
D6
D7
D8
D9
D10
D11
D12
D13
D14
BL21(DE3)_1lowastBL21(DE3)_2lowast
BL21-Gold(DE3)pLysS AGlowast
B str REL606lowastA phylogroup
ETEC H10407lowast (B2)
UMNK88 (A)
(B1 clades) Str 2009EL-2050Str 2009EL-2071
Str 2011-349355989
B1 phylogroup
Str EC4115Str TW14359
Str EDL933Str Sakai
Xuzhou21Str CB9615
Str RM12598
E phylogroup
Str 11128Str 11368 B1 phylogroup
NA114
SE15LF82lowastlowastlowastStr NRG 857Clowastlowastlowast
UM146
UTI89IHE3034
APEC O1lowastlowastS88
Clone D i2Clone D i14
ABU 83972
CFT073536
ED1aStr E234869
B2 phylogroup
UMN026
IAI39Str CE10
SMS-3-5
D phylogroup
O42lowastlowastlowast
(B2 clades)
(D clades)
(B1 clades)
Wlowastlowastlowast
WKO11FLlowastlowast
KO11FLE24377A
SE11
B1 phylogroup
IAI1ATCC 8739
P12blowastHS
BW2952
K-12 substr DH10BDH1
DH1 (ME8569)K-12 substr W3110
K-12 substr MG1655
A phylogroup
1
2
10
3156
Str 12009lowastlowast
Commensal ETEC EHEC EPEC UPEC
ExPECAPECEAECEIECENV
Figure 4 Analysis of distribution of Hae III profiles phylogroups and pathovars within the global alignment dendogram constructcomprising 57 E coli genomes The dendogram of the whole genome sequence of 57 genomes was obtained from the NCBI web site forE coli and was redrawn using Photoshop CS6 for colour coordination for pathogenic and nonpathogenic strains and inputting the clonalgroups and phylogroups (httpwwwncbinlmnihgovgenometerm=escherichia+coli) Among the 57 genomes 32 were of type I (56)18 were of type III (31) and 6 and 1 were of type IV (10) and type II (2) respectively showing that type I is predominant and type IIthe rarest To correlate between the phylogroupsHae III profiles and pathovars the lineages were considered as clades derived from variousdivergence point (D) during the course of evolution
as dendogram segregation shows type III strains evolving orpresent after type IV strains The clustering also shows thatpredominance of type III is more than type IV and maybeevolutionary processesmutation have triggered the increasein type III profiles On similar line type II clones would havebeen evolved from type I (addition) or type IV (deletion)but due to the very limited type II restriction profile patterns(rare) as observed from the gene pool data the chances orapproximation of such event is difficult to prove Based onthese results no new insights into phylogroup distributionacross candidate strains can be provided however withrespect to the Hae III profile an additional angle can beconsidered Phylogroups A and B1 are predominant withtype I Hae III profiles whereas B2 phylogroups exclusivelyrepresented type III profile (Figure 5(b))
An intrinsic metagenomic and pangenomic analysis ofmost bacterial species exhibited mosaicisms of their inherentblueprint [39] In such situations core gene elements sharedacross the species are limited in comparison to dispensable(flexible) genetic contents that concede uniqueness to aparticular strain These flexible contents help in niche adapt-ability and give bacteria the ability to infect and invade Amajor concern is while diagnosing both true and opportunistpathogenic bacteria the former causing disease in virtuallyany susceptible host and the latter invoking diseases inimmunocompromised individuals ldquoMolecular barcodingrdquoan evolved term which implies the method of allele specificamplification of relatively small DNA sequences or barcodes[40] for spotting a particular species is a thrust area formicrobial diagnostics A plethora of functional genes ranging
Journal of Pathogens 9
Type I(28 459)
Type IV(28 207 249 and 459)
Hypothesis
Type III(28 249 and 459)
+ minus
+
Primordial
NonpathogenicA gt B1 gt B2
type I gt IV gt III
PathogenicB2 gt B1 gt E gt D
type I (IAT)III(P)gt type IVII
Predominant clones
Rarest
+minus
Type II(28 207 and 459)
Restriction profilesand
clonesPhylogroups and
clones
2∘ clonesintermediate
(a) (b)
Figure 5 (a) Hypothesis of clonality in E coli strains based on 23S rrn restriction profiling The primordial soup of E coli microbes wouldhave been comprised of two clonal groups types I and IV coexisting with each other but predominated by type I strains Over the courseof evolution on accumulating mutation as a means for variation type I or type IV would have given rise to type III by addition (former) ordeletion (latter) of recognition sequence The second scenario could be a rare event because accumulation of two mutations in a conservedregion would be difficult Type II being the rarest would have been evolved in similar basis to type III (b) Hypothesis of linking phylogroupsHae III profiles and clonal types in E coli This diagram illustrates the clonal groups of commensal E coli strains having both A phylogroups(dominant) and B1 phylogroups each being type I profiles Pathogenic counterparts showed dominancy in type III restriction pattern followedby type I pattern With respect to phylogroups pathovars were predominant in B2 and B1 followed by D and E groups
from housekeeping to repair are employed for characterizingbacteria of which rRNA is referred to as the gold stretch[41 42] Ribosomal segments organized as operon clusters of16S 23S and 5S rDNA and these assemblages are distributedfrom single to multiple copies across the bacterial genophore[43] The occurrence of redundancy in maintaining differentcopies of these genes is not associated with virulence but anin-built ecological strategy that contributes to the particularbacterium with a competitive edge and better chance ofsurvival [44] A futuristic approach of coupling the techniqueand results with antibiotic resistance pattern colony char-acteristics and genomic profiles of bacterial strains of bothcommensal and pathogenic nature can help in monitoringand managing infections in a better manner
4 Conclusion
The causatum of postgenomic era genomics and proteomicshas enhanced this progress by improvising the techniquesused for diagnostics The strategy of ARDRA utilized inthis study was rapid and a cost-effective approach to dif-ferentiate E coli from its colony morphovars along withgiving a predicament of nature of origin or clonal groupsThe mutations reported in this work for detection andestablishing clonality namely T874A (Bfa I) G2161A andG2205A (Hae III) have not been mentioned in any researchpaper or within the Ribosomal RNA Mutation Database
[45] making this finding aboriginal This basic methodologycan be adapted for detection of other bacterial organismsby targeting SNPs linked to recognition sequences therebyvalidating rrn sequences as marker for diagnostics A cumu-lative approach of detection of unique restriction pattern of atargeted gene and its cross-reference across bacterial familiescan circumvent changes of false positives
Disclosure
This work was presented (poster) in part at the 5th Interna-tional Barcoding of Life Kunming China 2013
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
Acknowledgments
The authors acknowledge and express their gratitude to Pro-fessor D A Bhiwgade (Ex-Director) and Professor DebjaniDasgupta Director School of Biotechnology amp Bioinformat-ics D Y Patil University as well as the management for theirsupport and for providing the infrastructure to carry out thecurrent work
10 Journal of Pathogens
References
[1] M Richter and R Rossello-Mora ldquoShifting the genomic goldstandard for the prokaryotic species definitionrdquo Proceedings ofthe National Academy of Sciences of the United States of Americavol 106 no 45 pp 19126ndash19131 2009
[2] A F Auch M von Jan H-P Klenk and M Goker ldquoDigitalDNA-DNA hybridization for microbial species delineation bymeans of genome-to-genome sequence comparisonrdquo Standardsin Genomic Sciences vol 2 no 1 pp 117ndash134 2010
[3] D H Martin M Nsuami J Schachter et al ldquoUse of multiplenucleic acid amplification tests to define the infected-patientlsquogold standardrsquo in clinical trials of new diagnostic tests forChlamydia trachomatis infectionsrdquo Journal of Clinical Microbi-ology vol 42 no 10 pp 4749ndash4758 2004
[4] E A Mothershed and A M Whitney ldquoNucleic acid-basedmethods for the detection of bacterial pathogens presentand future considerations for the clinical laboratoryrdquo ClinicaChimica Acta vol 363 no 1-2 pp 206ndash220 2006
[5] J E Clarridge III ldquoImpact of 16S rRNA gene sequence analysisfor identification of bacteria on clinical microbiology andinfectious diseasesrdquo Clinical Microbiology Reviews vol 17 no4 pp 840ndash862 2004
[6] O Mizrahi-Man E R Davenport and Y Gilad ldquoTaxonomicclassification of bacterial 16S rRNA genes using short sequenc-ing reads evaluation of effective study designsrdquo PLoS ONE vol8 no 1 Article ID e53608 2013
[7] D E Lebonah A Dileep K Chandrasekhar S Sreevani BSreedevi and J Pramoda Kumari ldquoDNA barcoding on bacteriaa reviewrdquo Advances in Biology vol 2014 Article ID 541787 9pages 2014
[8] J Rajendhran and P Gunasekaran ldquoMicrobial phylogeny anddiversity small subunit ribosomal RNA sequence analysis andbeyondrdquo Microbiological Research vol 166 no 2 pp 99ndash1102011
[9] R Srinivasan U Karaoz M Volegova et al ldquoUse of 16S rRNAgene for identification of a broad range of clinically relevantbacterial pathogensrdquo PLoS ONE vol 10 no 2 Article IDe0117617 2015
[10] S Chakravorty D Helb M Burday N Connell and D AllandldquoA detailed analysis of 16S ribosomal RNA gene segments forthe diagnosis of pathogenic bacteriardquo Journal of MicrobiologicalMethods vol 69 no 2 pp 330ndash339 2007
[11] C R Woese ldquoReview bacterial evolutionrdquo MicrobiologicalReviews vol 51 pp 221ndash271 1987
[12] M Touchon C Hoede O Tenaillon et al ldquoOrganised genomedynamics in the Escherichia coli species results in highly diverseadaptive pathsrdquo PLoS Genetics vol 5 no 1 Article ID e10003442009
[13] J M Janda and S L Abbott ldquo16S rRNA gene sequencingfor bacterial identification in the diagnostic laboratory plusesperils and pitfallsrdquo Journal of Clinical Microbiology vol 45 no9 pp 2761ndash2764 2007
[14] J-J Lu C-L Perng S-Y Lee and C-CWan ldquoUse of PCRwithuniversal primers and restriction endonuclease digestions fordetection and identification of common bacterial pathogens incerebrospinal fluidrdquo Journal of Clinical Microbiology vol 38 no6 pp 2076ndash2080 2000
[15] N M Guentzel ldquoEscherichia Klebsiella Enterobacter SerratiaCitrobacter andProteusrdquo inMedicalMicrobiology S Baron Edchapter 26 University of Texas Medical Branch at GalvestonGalveston Tex USA 1996
[16] P Behzadip E Behzadi H Yazdanbod R Aghapour A MCheshmeh and S D Omran ldquoA survey on urinary tract infec-tions associated with the three most common uropathogenicbacteriardquoMaedica (Buchar) vol 5 no 2 pp 111ndash115 2010
[17] B CMetri P Jyothi andBV Peerapur ldquoAntibiotic resistance inCitrobacter spp isolated from urinary tract infectionrdquo UrologyAnnals vol 5 no 4 pp 312ndash313 2013
[18] N Taneja S S Chatterjee M Singh S Singh and M SharmaldquoPediatric urinary tract infections in a tertiary care center fromnorth Indiardquo Indian Journal of Medical Research vol 131 no 1pp 101ndash105 2010
[19] J B Kaper J P Nataro and H L T Mobley ldquoPathogenicEscherichia colirdquo Nature Reviews Microbiology vol 2 no 2 pp123ndash140 2004
[20] E J de Muinck K Lagesen J E Afset et al ldquoComparisonsof infant Escherichia coli isolates link genomic profiles withadaptation to the ecological nicherdquo BMC Genomics vol 14 no1 article 81 2013
[21] D A Rasko M J Rosovitz G S A Myers et al ldquoThepangenome structure of Escherichia coli comparative genomicanalysis of E coli commensal and pathogenic isolatesrdquo Journalof Bacteriology vol 190 no 20 pp 6881ndash6893 2008
[22] C T Archer J F Kim H Jeong et al ldquoThe genome sequenceof E coli W (ATCC 9637) comparative genome analysis andan improved genome-scale reconstruction of E colirdquo BMCGenomics vol 12 article no 9 2011
[23] PKuhnert J Nicolet and J Frey ldquoRapid and accurate identifica-tion of Escherichia coliK-12 strainsrdquoApplied and EnvironmentalMicrobiology vol 61 no 11 pp 4135ndash4139 1995
[24] A P Bauer S M Dieckmann W Ludwig and K-H SchleiferldquoRapid identification of Escherichia coli safety and laboratorystrain lineages based on multiplex-PCRrdquo FEMS MicrobiologyLetters vol 269 no 1 pp 36ndash40 2007
[25] B J Bachmann ldquoPedigrees of some mutant strains ofEscherichia coli K-12rdquo Bacteriological Reviews vol 36 no 4 pp525ndash557 1972
[26] P Daegelen F W Studier R E Lenski S Cure and J F KimldquoTracing ancestors and relatives of Escherichia coli B and thederivation of B strains REL606 and BL21(DE3)rdquo Journal ofMolecular Biology vol 394 no 4 pp 634ndash643 2009
[27] F W Studier P Daegelen R E Lenski S Maslov and J F KimldquoUnderstanding the differences between genome sequences ofEscherichia coli B strains REL606 and BL21(DE3) and compar-ison of the E coli B and K-12 genomesrdquo Journal of MolecularBiology vol 394 no 4 pp 653ndash680 2009
[28] R K Selander and B R Levin ldquoGenetic diversity and structurein Escherichia coli populationsrdquo Science vol 210 no 4469 pp545ndash547 1980
[29] D M Gordon O Clermont H Tolley and E DenamurldquoAssigning Escherichia coli strains to phylogenetic groupsmulti-locus sequence typing versus the PCR triplex methodrdquoEnvironmental Microbiology vol 10 no 10 pp 2484ndash24962008
[30] P Desjardins B Picard B Kaltenbock J Elion and EDenamur ldquoSex in Escherichia coli does not disrupt the clonalstructure of the population evidence from random amplifiedpolymorphic DNA and restriction-fragment-length polymor-phismrdquo Journal of Molecular Evolution vol 41 no 4 pp 440ndash448 1995
Journal of Pathogens 11
[31] N D Jasuja R Saxena S Chandra and S C Joshi ldquoIsolationand identification of microorganism from polyhouse agricul-ture soil of RajasthanrdquoAfrican Journal ofMicrobiology Researchvol 7 no 41 pp 4886ndash4891 2013
[32] V Hemraj S Diksha and G Avneet ldquoA review of commonlyused biochemical test for bacteriardquo Innovare Journal of LifeSciences vol 1 pp 1ndash7 2013
[33] J D Thompson D G Higgins and T J Gibson ldquoCLUSTALWimproving the sensitivity of progressive multiple sequencealignment through sequence weighting position-specific gappenalties and weight matrix choicerdquoNucleic Acids Research vol22 no 22 pp 4673ndash4680 1994
[34] T Vincze J Posfai and R J Roberts ldquoNEBcutter a program tocleave DNA with restriction enzymesrdquo Nucleic Acids Researchvol 31 no 13 pp 3688ndash3691 2003
[35] J Ye G Coulouris I Zaretskaya I Cutcutache S Rozen andT Madden ldquoPrimer-BLAST a tool to design target-specificprimers for polymerase chain reactionrdquo BMC Bioinformaticsvol 13 article 134 2012
[36] J R Parvathi and S Singh ldquoFacile approaches for microbialDNA extraction comparative study with colonies lsquopickedrsquo fromthree different bacteriological mediumrdquo Journal of Pure andApplied Microbiology vol 9 no 3 pp 2373ndash2378 2015
[37] R Lan and P R Reeves ldquoEscherichia coli in disguise molecularorigins of Shigellardquo Microbes and Infection vol 4 no 11 pp1125ndash1132 2002
[38] D J Leininger J R Roberson and F Elvinger ldquoUse ofeosin methylene blue agar to differentiate Escherichia coli fromother gram-negative mastitis pathogensrdquo Journal of VeterinaryDiagnostic Investigation vol 13 no 3 pp 273ndash275 2001
[39] U Dobrindt F Agerer K Michaelis et al ldquoAnalysis of genomeplasticity in pathogenic and commensal Escherichia coli isolatesby use of DNA arraysrdquo Journal of Bacteriology vol 185 no 6pp 1831ndash1840 2003
[40] V Savolainen R S Cowan A P Vogler G K Roderickand R Lane ldquoTowards writing the encyclopaedia of life anintroduction to DNA barcodingrdquo Philosophical Transactions ofthe Royal Society B Biological Sciences vol 360 no 1462 pp1805ndash1811 2005
[41] T A Isenbarger C E Carr S S Johnson et al ldquoThe mostconserved genome segments for life detection on earth andother planetsrdquo Origins of Life and Evolution of Biospheres vol38 no 6 pp 517ndash533 2008
[42] S R Santos and H Ochman ldquoIdentification and phylogeneticsorting of bacterial lineages with universally conserved genesand proteinsrdquoEnvironmentalMicrobiology vol 6 no 7 pp 754ndash759 2004
[43] R Rastogi M Wu I DasGupta and G E Fox ldquoVisualizationof ribosomal RNA operon copy number distributionrdquo BMCMicrobiology vol 9 no 1 article 208 2009
[44] J A Klappenbach J M Dunbar and T M Schmidt ldquorRNAoperon copy number reflects ecological strategies of bacteriardquoApplied and Environmental Microbiology vol 66 no 4 pp1328ndash1333 2000
[45] J A Klappenbach P R Saxman J R Cole and T M SchmidtldquoRrndb the ribosomal RNA operon copy number databaserdquoNucleic Acids Research vol 29 no 1 pp 181ndash184 2001
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom

Journal of Pathogens 3
Table 1 List of standard strains used in this study
Strains MTCC ATCC NatureBarcodingidentification studies
Noncandidate strainsCitrobacter freundii 1658 8090 ClinicalCitrobacter koseri 1657 27028 Blood cultureKlebsiella pneumonia 432 33495 Urinary cultureEnterobacter aerogenes 111 13048 Sputum
Candidate strains433 15228 Commensal
Escherichia coli 4296 19138 Urine448 25922 Clinical sample
Diversityclonality studies
Escherichia coli
1302 mdash K122622 10798 Descendent of K121667 mdash DH11304 mdash C11687 8739 Crooks strain448 9637 W2692 53846 Descendent of W1303 mdash B1678 mdash BL211679 mdash BL21DE31680 mdash BL21DE3Lys5
E coli namely K-12 B C Crooks andW are commonly used as lab strains Whole genomic data of these strains except strain C is available (NCBI Genomes)
Candidate microbe
Noncandidate microbe
Escherichia coli
Klebsiella pneumoniae (2 G)Enterobacter aerogenes (1 G)
Citrobacter koseri (1 G)
sequences from each genome
57 genomes (G) (21 pathogenic
36 nonpathogenic)
Estimation of diversity (multiple sequence alignment MSA)Intergenic diversity
representative genome Intragenic diversity
Restriction profiling for differentiating candidate
from noncandidates (NEB V20)
85 restriction groups(Suppl A2)
Virtual digestion was individually
carried out on each rrlH
Similar restriction patterns(rejected) Dissimilar restriction patterns
(selected)
Restriction patterns were checked in all the 430 ribosomal segments
Unique profile in candidate alone
(barcoding)Bfa I was chosen
Polymorphic within candidate genome
(clonality)Hae III was chosen
A total of 430 sequences considering both candidate and noncandidate genomes
(ii) Regular strands of eachgenome
(i) Complementary strands ofeach genome
rrlH from each
Extraction of 23S rrn
Figure 1 Schematic representation of methodology for detecting barcode and estimating clonality The primers are designed against theregions that flank these restriction sequences which gives unique bands revealing identification or clonality
For estimating the intragenic variability all the riboso-mal segments from 57 E coli genomes were extracted (7copiesgenome) complementary and regular strands withineach strain were segregated and the percentage mismatchesfor each of the genomes were determined
23 Virtual Restriction Profiling Virtual restriction profilesusing NEB V20 [34] with 85 restriction groups (Supplemen-tary Data A2) were performed for a total of 430 ribosomalsegments that included 399 from 57 E coli genomes and7 8 and 16 rrn sequences from C koseri E aerogenes
4 Journal of Pathogens
Table 2 Restriction site of Bfa I (CTAG) within 23S rrlH of candidate and noncandidates and the restriction profiles with 23S P1880
Restriction profile within rrlH Fragments obtained on ARDRAF E Ci K Ea Mic Fragments (amplicon size)11 687 685 685 685 E 665 bp 215 bp (880 bp)12 874 874 874 C 475 bp 214 bp and 189 bp (878 bp)13 1670 1669 1667 1670 K 475 bp 214 bp and 189 bp (877 bp)14 2336 2339 Ea 475 bp 214 bp and 189 bp (876 bp)15 2652 2653 2650 2654Primers were designed against the regions flanking the bold restriction sitesF number of fragments E Escherichia coli Ci Citrobacter koseri K Klebsiella pneumoniae Ea Enterobacter aerogenes Mic microbe23S P1
880-FP(+) 51015840-GGCGAAAAGAACCCCGGCGA-31015840 (20 nt) 23S P1
880-RP(minus) 51015840-AGGGGTCGACTCACCCTGCC-31015840 (20 nt)
FP forward primer RP reverse primer The primer details were obtained from the results of primer blast and restriction profiling pattern was obtained fromNEB V20 Forward and reverse primers of 23S P1
880lie within domains I and III of 23S rRNA secondary structure respectively with Bfa I sites in domain II
Table 3 Clonal groups based on Hae III (GGCC) profiles within the candidate microbe using 23S P2682
Virtual digest pattern Types of Hae III restriction pattern observed amongst 399 rrn sequences of E coli genomesF C Type I Type II Type III Type IV11 785 Cut site within rrlH of candidate rrn segments12 1042 1984 1984 1984 198413 1116 x 2163 X 216314 1217 x X 2205 220515 1361 2415 2415 2415 241516 1445 Cut site within the amplicon (682 bp)17 1907 28 28 28 2818 1984 x 207 X 20719 2161 x X 249 249110 2205 459 459 459 459111 2415 Fragments obtained for each group
112 2839 28 bp 223 bp and 431 bp 28 bp 179 bp 223 bp and252 bp
28 bp 210 bp 221 bp and223 bp
28 bp 42 bp 179 bp210 bp and 223 bp
Primers were designed against the regions flanking the bold restriction sites F number of fragments C cut sites23S P2
682-FP(+) 51015840CCGACCTGCACGAATGGCGT31015840 (20 nt)
23S P2682
-RP(+) 51015840CAGTTCTCCAGCGCCCACGG31015840 (20 nt)Forward and reverse primers of 23S P2
682lie within domain IV and transition of V-VI of 23S rRNA secondary structure respectively First cut sites lie within
domain IV whereas the rest lie within domain V
and K pneumoniae strains respectively Among the 14common cutters groups (Supplementary Data A3) Bfa I(Supplementary Data A4) was selected as the enzyme forbarcoding as it gave conserved and distinct pattern acrossall the 399 rrn sequences (not shown) in differentiating thecandidate from the noncandidate strains The uniquenessand position of the cuts sites for giving unique bands onagarose gel electrophoresis were also a lead reason consideredfor finalizing this enzyme Hae III displaying polymorphismpattern with 23S rDNA segments of candidate microbe wasfinalized for diversity studies (Figure 1)
24 Primers and Restriction Enzymes for Addressing Identityand Clonality Sequence of rrlH of E coli K-12 str MG1655was used as the template for designing broad range primersusing Primer-BLAST [35] In conjunction with parameterslike GC content 119879
119898value and the amplicon size number of
restriction sites within the amplicon and the size of fragmentafter digestion were kept as the benchmark for finalizing
primersThere are twoprimer sets 23S P1880
[Probe 31778738]was selectedwithBfa I restriction digestion (Table 2) demarkingthe candidate and 23S P2
682[Probe 31781046] (Table 3) with
polymorphicHae III restriction siteswas selected for address-ing clonality Possibility of self-priming andprimer-dimer for-mation of each primer was checked using free web tools Oli-gocal (httpwwwsimgenecomOligoCalc) OligoEvaluator(httpwwwsigmaaldrichcomtechnical-documentsarticlesbiologyoligo-evaluatorhtml) and multiple primer analyzertool (httpwwwthermoscientificbiocomwebtoolsmultiple-primer) maintained by SimGene Sigma and Fermentasresp
BLAST genome (Supplementary Data) assisted in veri-fying the conservation and specificity of primers ampliconsize Bfa I recognition sites and region of hybridizationacross the genome of 931 bacterial families After omitting thenonannotated sequences all of the completed whole genomedata aggregated under phyla Actinobacteria BacteroidetesChlamydiae Cyanobacteria Firmicutes Proteobacteria and
Journal of Pathogens 5
Spirochaetae of the bacterial kingdom were considered forthe analysis
25 DNA Isolation Amplification and Restriction DigestionDNA isolation was performed according to Parvathi andSingh [36] The reaction soup comprised 2120583L of supernatant(DNA) 05 120583M of each forward and reverse primer 1x ofassay buffer (10mM Tris-HCl pH 88 50mM KCl) MgCl
2
(15mM) dNTPmix (800 120583M) and 05U120583LTaq polymerase(Kappa Systems KK5004) Standard genomic DNA of Ecoli K-12 (Merck) and nuclease-free water were used aspositive and negative control The reactions were carriedout in a thermal cycler (Bioer) with the program set for25 cycles comprising denaturation (95∘C) annealing (68∘C)and extension (72∘C) for 30 s each 3 120583L of the PCR productwas digested with 2U of respective enzyme (FermentasThermo Scientific) in 5120583L reactionmix at 37∘C for two hoursThe amplicons and their restriction products were analysedby agarose gel electrophoresis (AGE) at 5Vcm in 2 and25 agarose gels respectively each containing 05120583gmL ofEtBr The gels were viewed and photographed in BioImagingSystems (Syngene)
3 Result and Discussion
31 Genetic Diversity within 23S Ribosomal Repeats for Identi-fication of E coli 23S rRNA gene showed 087 intergenicvariations compared with 16S rRNA within the referencegenomes (Supplementary Data A5) The result seemed obvi-ous due to the increased frame size of the latter but thisalso encouraged perusing microheterogenicity analysis ofthe largest ribosomal segment for further approach Thesizes of target segments varied from 2903 to 2907 bp withmaximum gene length observed in ATCC8739 (Crookrsquosstrain) accounting for 2970ndash2985 bp followed by E24377Adisplaying sequences of 2923ndash2925 bp One rrn of UM 146and ETECH10407 were of 1762 bp and 1561 bp respectively(Supplementary Data A9) Absolutely no link between rrnsize and pathogenicity could be defined because the varia-tions were random ATCC8739 being a laboratory cultureof fecal origin and UM146 an EIAC strain E23477A andETECH10407 of ETEC origin
Ribosomal genes were found to be distributed acrosscomplementary and regular strand with majority of thegenomes (46 out of 57) showing segregation in five regularand two complementary strands All the 23S rrn sequencesdistributed within the regular and complementary strandsacross respective genomes were individually sorted for MSAto attain an understanding of intragenic variation Analysisrevealed the highest level of mismatches in complementarystrands compared with their counterparts (SupplementaryData A6) The highest level of mismatches mounting to06 corresponding to 50 of the targeted gene size wasperceived in complementary strands of P12b (commensal)E23489 (EPEC) and ETEC H10407 The greatest dispar-ity among sequences in regular strand was observed inAPEC 01 with 003 (882903) followed by CFT043 (UPEC)and 042 (EAEC) having 0016 (sim42ndash512903) dissimilarity23S ribosomal copies of ED1a (commensal) showed zero
percent mismatches in both complementary and regularstrands making it an ideal cluster with the highest degreeof conservation Similar values of mismatches were seen inSE11 (Commensal) CB9615 (EPEC) and 0104H4 2009El-2011 (EAEC) These results indicate that there is no cor-relation between intragenic diversity of target stretch andpathogenicity of strains These differences substantiated 23SRNA clusters having microdiverse organization with bothconserved and variable stretches within them Sequevarsarising due to this heterogeneity unavoidably lead to biasedresults differential amplification and chimeric moleculesThus the method of screening and targeting restriction pro-file pattern within a region of choice (ribosomal segments)and specific to bacteria of interest demonstrates authentic andsimpler practice than routine molecular strategy involvingamplification and sequencing
32 Barcoding E coli from Its Colony Morphovars ARDRAof 23S P1880 with Bfa I PCR amplification of the stan-dard strains and isolates of different origin (commensalurine and clinical sample) with 23S P1
880gave amplicons
(880 bp) of high intensity with no primer-dimer formation(Figure 2(a)) Restriction digestion of the amplicons withBfa I gave unique fragments of 665 bp and 215 bp in E coliwhereas C koseri E aerogenes and K pneumoniae eachgave fragments of 475 bp 214 bp and 189 bp respectively(Figure 2(b)) The genotypic strategy proved superior tobiochemical characterization taking into consideration all thepossible positive negative and false positive and negativeresults across 25 isolates each from clinical commensal andenvironmental origin (Supplementary Data A7) 18 (72)of clinical 22 (88) of commensal and all the 25 (100)environment isolates showed distinct restriction pattern likethat of E coli BLAST genome search across 931 bacterialfamilies verified the specificity of primers and restrictionpreference for Bfa I The primers showed exclusive bindingonly with Enterobacteriaceae family (Supplementary DataA8) Among the various strains in the family only Shigellaspp Salmonella enterica (Arizona) and Proteus mirabilusexhibited similar restriction preference to that of E coli Inspite of Shigella spp being taxa in disguise of E coli [37] theycan be easily screened using selective medium because of nolactose fermentation [38] whereas Salmonella enterica andProteus mirabilis showed amplicon heterogeneity of 1073 bpand 880 bp during amplification thus making it easy to avoiderroneous conclusions and misinterpretation
33 Categorizing E coli StrainsUsingARDRAof 23S P2682 withHae III A detailed analysis of the Hae III profile across 399sequences [Supplementary Data A9] of ribosomal segmentsof E coli indicated four possible profiles types I II IIIand IV with varying degree of predominance within thesesegments The rest of the candidate strains showed type IIIpatterns but types I and IV were exclusive for E coli strainsMajority of genomes (119899 = 32) as in 15 nonpathogenic and17 pathogenic strains showed type I profiles (57) followedby type III (35) predominating in 15 strains Six strainsshowed type IV pattern (10) of which four were B strainsType I groups represent E coli strains that lacked recognition
6 Journal of Pathogens
1 7
880 bp
65432
(a)
8
665bp
475bp
215 bp 214 bp
178bp
1 765432
(b)
Figure 2 (a) Broad Range PCR of 23S P1880
with genomic DNA isolated from standard strains Lane 1 100 bp ladder Lane 2 blank Lane 3E coli Lane 4 C freundii Lane 5 C koseri Lane 6 K pneumonia Lane 7 E aerogenes PCR product of single representative of E coli wasloaded (b) ARDRA of 23S P1
880
amplicons with Bfa I Lanes 1 2 and 3 E coli (ATCC15223 ATCC25922 and ATCC11775) Lane 4C freundii(ATCC8090) Lane 5 C koseri (ATCC27028) Lane 6 K pneumonia (ATCC33495) Lane 7 E aerogenes (ATCC13048) Lane 8 100 bp ladder
elements due to change at 2205th and 2161st base with respectto 23S rDNA or with reference to amplicons at 207th or 249thbase positions Type IV strains are contrary to type I withintact recognition sequence at both of these sites Strains withtype II profiles lacked 2161st (249th) cut site and type IIIlacked the 2205th (207th) cut positions respectively Type IIwas the rarest with its patterns observed to be ambiguous incertain rrn segments
Heterogenicity in few genomes was noted two rrn amongseven repeats ofW (NC 0176851 each in regular and comple-mentary strand) and KO11FL (NC 0176601 each in regularand complementary strand) and one in KO11FL (NC 0169021 regular) showed type II profile in spite of type I beingtheir primary pattern One among seven of the B strains REL606 (NC 0129671 regular) BL21-Gold (NC 0129471 com-plementary) BL21DE3 (NC 0129712 regular) and BL21DE3(NC 0128922 regular) showed type I profiles apart frombeing prevalent with type IV pattern Similarly two rrnamongNRG 873C (NC 0176341 each in regular and comple-mentary strand) and LF82 (NC 0119931 each in regular andcomplementary strand) and one in CFT073 (NC 0044311regular) showed type I pattern while exhibiting type IIIpattern predominance APEC01 (NC 0085631) being primein type III showed one rrn segment of type I and type IV(both in regular strand) E coli strain 042 (NC 0176261)displayed preponderance in type I pattern but three rrn (tworegular and one complementary) copies exhibited type IIIpattern One of the regular rrn elements in LF82 showed typeIV pattern The fragmentation pattern obtained by ARDRAof 23S P2
682with Hae III in the standard samples showed
that all K-12 C W Crooks and their respective descendantsexhibited type I profileswith type IVprofiles being exclusivelypresented by B strains (Figures 3(a) and 3(b))
Among the isolates 21 out of 22 commensal and 16 of18 clinical samples showed type I pattern heterogenicity ofpatterns was not observed These results reaffirmed predom-inance (93) of type I in nature Multisequence typing hasshown commensal strains to be categorized primarily undergroup A or group B1 whereas groups B2 D and E comprisegenerally pathogenic strains K-12 B and Crooks strain comeunder phylogroup A whereas strain W is a B1 phylogroup
34 Local Alignment of 23S rrn Hae III Profiles versus GlobalAlignment to Establish Clonality in E coli A number ofgenes that are of core dispensable and unique in naturemake up for pangenome the latter two are gained due torecombinational or transpositional events To obtain similarclustering on comparing whole genome would emphasizethe significance of these restriction profiles and help inestablishing the clonal nature and predominance of a singleclone Hence dendogram constructs with 682 bp ampliconfrom a representative of each restriction profile will showdistinct clustering of these profiles due to the fact that a smallsubset of segments within the whole genome was consideredThe global alignment of whole genome sequences (Figure 4)of the entire 57 E coli strains displayed distinct concentrateof type I III and IV profiles An intrinsic analysis of thespot of divergence and respective clade formations showedinteresting and riveting findings The strains at the ancestralroot showed type I pattern and the next divergent node point(D1) displayed formation of two distinct clans of type I andtype IV profiles segregated at the maximum distance fromone another Numbering of nodes was done by consideringthe precedence of branch point at every node A possiblescenario of two additional mutational events that are oftransitional nature (A to G) at 2205th and 2161st bases has
Journal of Pathogens 7
682bp
987654321
9998400
8998400
7998400
6998400
5998400
4998400
3998400
2998400
1998400
(a)
431bp
223 bp
210 bp
179bp
223 + 210 bp
223+
210+
87654321
8998400
7998400
6998400
5998400
4998400
3998400
2998400
1998400
(b)
Figure 3 (a) Broad range PCR of 23S P2682
with genomic DNA isolated from standard strains of E coli The upper row comprises Lane 1K-12 strains Lane 2 derivatives of K-12 strains Lane 3 DHI strain Lane 4 strain C1 Lane 6 Crookrsquos strain Lane 7 Walkerrsquos strain Lane 8descendent of W Lane 9 blank The lower row comprises Lane 11015840 B strain Lane 21015840 BL21 strain Lane 31015840 BL21DE3 Lane 41015840 BL21DE3Lys5(B3) Lane 61015840 E coli (ATCC 15223 commensal isolate) Lane 71015840 E coli (ATCC 11775 urine sample) Lane 81015840 E coli (ATCC 25922 clinical)Lanes 551015840 and Lanes 991015840 in both rows are 100 bp ladder and blank respectively (b) ARDRA of 23S P2
682
amplicons with Hae III Lane 1 ofboth rows comprises 100 bp ladder The digested products are loaded as per the order given in Figure 2(a)
introduced Hae III recognition patterns D6 gave lineages ofB1 phylogroups D9 giving rise to B2 clades (D12) of type Iprofiles and D clades with varied Hae III patterns (D11) D10gave rise to two nodes (with strains being type I groups)with one showing B1 phylogroups (D13) and the other beingpredominant in E phylogroups followed by B1 An almostexemplary corelation between the clonal groups and differentpathovars was found with this study Majority of commensalstrains and EIEC and all EAEC and EPEC were shown tobe having type I profile (Table 4) All UPEC ExPEC APECEIEC and single strain of EPEC and two commensal strainsshowed type III pattern B form strains along with a singlestrain of UPEC and environmental isolate exhibited type IVprofiles
Similarly a near-pristine association between phylo-groups and Hae III profiles was established Strains comingunder phylogroups A were preponderant in type I whereastype IV profiles were exclusively for B forms Type I profileswere observed in the strains (119899) in the order of A gt B1 gt E gtDwith type III dominant in B2 strains Phylogroup D showedequal occurrence of type III and type IV fragmentationpattern followed by a single strain with type I profile Theseresults assisted in enumerating a conjecture which works upa scenario of primordial microbial population where E colistrains had two clonal types (Figure 5(a)) which coexistedtogether (type I and type IV clones) Of these type I strainswould have been the predominant clone in comparison totype IV based on the dendogram data with maximum genepool of root strains showing type I Type III may have beenthe next predominant clone (an intermediary one) that hasevolved either from type I (by addition of a new recognition
Table 4 Corelation between phylogroups Hae III profiles andnature of E coli strains (genome data analysis of 57 E coli strainscollated from Genbank)
Strain Type A B1 B2 D E Total Out ofCommensal I 10 5 mdash mdash mdash 15 21Commensal III mdash mdash 2 mdash mdash 2 21Commensal IV 4 mdash mdash mdash mdash 4 21EPEC I mdash mdash mdash mdash 7 7 8EPEC III mdash mdash 1 mdash mdash 1 8EAEC I mdash 4 mdash 1 mdash 5 5EHEC I mdash 2 mdash mdash mdash 2 3EHEC II mdash 1 mdash mdash mdash 1 3ETEC I 1 1 1 mdash mdash 3 3UPEC III mdash mdash 7 mdash mdash 7 8UPEC IV mdash mdash mdash 1 mdash 1 8EIEC III mdash mdash 3 mdash mdash 3 3ExPEC III mdash mdash 2 2 mdash 4 4APEC III mdash mdash 1 mdash 1 1ENV IV mdash mdash mdash 1 1 1APEC avian pathogenic EAEC enteroaggressive EHEC enterohemorrhag-ic ETEC enterotoxigenic EIEC enteroinvasive EPEC enteropathogenicENV environmental ExPEC extraintestinal pathogenic UPEC uropatho-genic
site) or from type IV (by deletion of one recognition site)strains In a different scenario maybe all three types coexistedor type III would have given rise to type IVwith an additionalmutational event This situation may be hardly convincing
8 Journal of Pathogens
D1
D2
D3
D4
D5
D6
D7
D8
D9
D10
D11
D12
D13
D14
BL21(DE3)_1lowastBL21(DE3)_2lowast
BL21-Gold(DE3)pLysS AGlowast
B str REL606lowastA phylogroup
ETEC H10407lowast (B2)
UMNK88 (A)
(B1 clades) Str 2009EL-2050Str 2009EL-2071
Str 2011-349355989
B1 phylogroup
Str EC4115Str TW14359
Str EDL933Str Sakai
Xuzhou21Str CB9615
Str RM12598
E phylogroup
Str 11128Str 11368 B1 phylogroup
NA114
SE15LF82lowastlowastlowastStr NRG 857Clowastlowastlowast
UM146
UTI89IHE3034
APEC O1lowastlowastS88
Clone D i2Clone D i14
ABU 83972
CFT073536
ED1aStr E234869
B2 phylogroup
UMN026
IAI39Str CE10
SMS-3-5
D phylogroup
O42lowastlowastlowast
(B2 clades)
(D clades)
(B1 clades)
Wlowastlowastlowast
WKO11FLlowastlowast
KO11FLE24377A
SE11
B1 phylogroup
IAI1ATCC 8739
P12blowastHS
BW2952
K-12 substr DH10BDH1
DH1 (ME8569)K-12 substr W3110
K-12 substr MG1655
A phylogroup
1
2
10
3156
Str 12009lowastlowast
Commensal ETEC EHEC EPEC UPEC
ExPECAPECEAECEIECENV
Figure 4 Analysis of distribution of Hae III profiles phylogroups and pathovars within the global alignment dendogram constructcomprising 57 E coli genomes The dendogram of the whole genome sequence of 57 genomes was obtained from the NCBI web site forE coli and was redrawn using Photoshop CS6 for colour coordination for pathogenic and nonpathogenic strains and inputting the clonalgroups and phylogroups (httpwwwncbinlmnihgovgenometerm=escherichia+coli) Among the 57 genomes 32 were of type I (56)18 were of type III (31) and 6 and 1 were of type IV (10) and type II (2) respectively showing that type I is predominant and type IIthe rarest To correlate between the phylogroupsHae III profiles and pathovars the lineages were considered as clades derived from variousdivergence point (D) during the course of evolution
as dendogram segregation shows type III strains evolving orpresent after type IV strains The clustering also shows thatpredominance of type III is more than type IV and maybeevolutionary processesmutation have triggered the increasein type III profiles On similar line type II clones would havebeen evolved from type I (addition) or type IV (deletion)but due to the very limited type II restriction profile patterns(rare) as observed from the gene pool data the chances orapproximation of such event is difficult to prove Based onthese results no new insights into phylogroup distributionacross candidate strains can be provided however withrespect to the Hae III profile an additional angle can beconsidered Phylogroups A and B1 are predominant withtype I Hae III profiles whereas B2 phylogroups exclusivelyrepresented type III profile (Figure 5(b))
An intrinsic metagenomic and pangenomic analysis ofmost bacterial species exhibited mosaicisms of their inherentblueprint [39] In such situations core gene elements sharedacross the species are limited in comparison to dispensable(flexible) genetic contents that concede uniqueness to aparticular strain These flexible contents help in niche adapt-ability and give bacteria the ability to infect and invade Amajor concern is while diagnosing both true and opportunistpathogenic bacteria the former causing disease in virtuallyany susceptible host and the latter invoking diseases inimmunocompromised individuals ldquoMolecular barcodingrdquoan evolved term which implies the method of allele specificamplification of relatively small DNA sequences or barcodes[40] for spotting a particular species is a thrust area formicrobial diagnostics A plethora of functional genes ranging
Journal of Pathogens 9
Type I(28 459)
Type IV(28 207 249 and 459)
Hypothesis
Type III(28 249 and 459)
+ minus
+
Primordial
NonpathogenicA gt B1 gt B2
type I gt IV gt III
PathogenicB2 gt B1 gt E gt D
type I (IAT)III(P)gt type IVII
Predominant clones
Rarest
+minus
Type II(28 207 and 459)
Restriction profilesand
clonesPhylogroups and
clones
2∘ clonesintermediate
(a) (b)
Figure 5 (a) Hypothesis of clonality in E coli strains based on 23S rrn restriction profiling The primordial soup of E coli microbes wouldhave been comprised of two clonal groups types I and IV coexisting with each other but predominated by type I strains Over the courseof evolution on accumulating mutation as a means for variation type I or type IV would have given rise to type III by addition (former) ordeletion (latter) of recognition sequence The second scenario could be a rare event because accumulation of two mutations in a conservedregion would be difficult Type II being the rarest would have been evolved in similar basis to type III (b) Hypothesis of linking phylogroupsHae III profiles and clonal types in E coli This diagram illustrates the clonal groups of commensal E coli strains having both A phylogroups(dominant) and B1 phylogroups each being type I profiles Pathogenic counterparts showed dominancy in type III restriction pattern followedby type I pattern With respect to phylogroups pathovars were predominant in B2 and B1 followed by D and E groups
from housekeeping to repair are employed for characterizingbacteria of which rRNA is referred to as the gold stretch[41 42] Ribosomal segments organized as operon clusters of16S 23S and 5S rDNA and these assemblages are distributedfrom single to multiple copies across the bacterial genophore[43] The occurrence of redundancy in maintaining differentcopies of these genes is not associated with virulence but anin-built ecological strategy that contributes to the particularbacterium with a competitive edge and better chance ofsurvival [44] A futuristic approach of coupling the techniqueand results with antibiotic resistance pattern colony char-acteristics and genomic profiles of bacterial strains of bothcommensal and pathogenic nature can help in monitoringand managing infections in a better manner
4 Conclusion
The causatum of postgenomic era genomics and proteomicshas enhanced this progress by improvising the techniquesused for diagnostics The strategy of ARDRA utilized inthis study was rapid and a cost-effective approach to dif-ferentiate E coli from its colony morphovars along withgiving a predicament of nature of origin or clonal groupsThe mutations reported in this work for detection andestablishing clonality namely T874A (Bfa I) G2161A andG2205A (Hae III) have not been mentioned in any researchpaper or within the Ribosomal RNA Mutation Database
[45] making this finding aboriginal This basic methodologycan be adapted for detection of other bacterial organismsby targeting SNPs linked to recognition sequences therebyvalidating rrn sequences as marker for diagnostics A cumu-lative approach of detection of unique restriction pattern of atargeted gene and its cross-reference across bacterial familiescan circumvent changes of false positives
Disclosure
This work was presented (poster) in part at the 5th Interna-tional Barcoding of Life Kunming China 2013
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
Acknowledgments
The authors acknowledge and express their gratitude to Pro-fessor D A Bhiwgade (Ex-Director) and Professor DebjaniDasgupta Director School of Biotechnology amp Bioinformat-ics D Y Patil University as well as the management for theirsupport and for providing the infrastructure to carry out thecurrent work
10 Journal of Pathogens
References
[1] M Richter and R Rossello-Mora ldquoShifting the genomic goldstandard for the prokaryotic species definitionrdquo Proceedings ofthe National Academy of Sciences of the United States of Americavol 106 no 45 pp 19126ndash19131 2009
[2] A F Auch M von Jan H-P Klenk and M Goker ldquoDigitalDNA-DNA hybridization for microbial species delineation bymeans of genome-to-genome sequence comparisonrdquo Standardsin Genomic Sciences vol 2 no 1 pp 117ndash134 2010
[3] D H Martin M Nsuami J Schachter et al ldquoUse of multiplenucleic acid amplification tests to define the infected-patientlsquogold standardrsquo in clinical trials of new diagnostic tests forChlamydia trachomatis infectionsrdquo Journal of Clinical Microbi-ology vol 42 no 10 pp 4749ndash4758 2004
[4] E A Mothershed and A M Whitney ldquoNucleic acid-basedmethods for the detection of bacterial pathogens presentand future considerations for the clinical laboratoryrdquo ClinicaChimica Acta vol 363 no 1-2 pp 206ndash220 2006
[5] J E Clarridge III ldquoImpact of 16S rRNA gene sequence analysisfor identification of bacteria on clinical microbiology andinfectious diseasesrdquo Clinical Microbiology Reviews vol 17 no4 pp 840ndash862 2004
[6] O Mizrahi-Man E R Davenport and Y Gilad ldquoTaxonomicclassification of bacterial 16S rRNA genes using short sequenc-ing reads evaluation of effective study designsrdquo PLoS ONE vol8 no 1 Article ID e53608 2013
[7] D E Lebonah A Dileep K Chandrasekhar S Sreevani BSreedevi and J Pramoda Kumari ldquoDNA barcoding on bacteriaa reviewrdquo Advances in Biology vol 2014 Article ID 541787 9pages 2014
[8] J Rajendhran and P Gunasekaran ldquoMicrobial phylogeny anddiversity small subunit ribosomal RNA sequence analysis andbeyondrdquo Microbiological Research vol 166 no 2 pp 99ndash1102011
[9] R Srinivasan U Karaoz M Volegova et al ldquoUse of 16S rRNAgene for identification of a broad range of clinically relevantbacterial pathogensrdquo PLoS ONE vol 10 no 2 Article IDe0117617 2015
[10] S Chakravorty D Helb M Burday N Connell and D AllandldquoA detailed analysis of 16S ribosomal RNA gene segments forthe diagnosis of pathogenic bacteriardquo Journal of MicrobiologicalMethods vol 69 no 2 pp 330ndash339 2007
[11] C R Woese ldquoReview bacterial evolutionrdquo MicrobiologicalReviews vol 51 pp 221ndash271 1987
[12] M Touchon C Hoede O Tenaillon et al ldquoOrganised genomedynamics in the Escherichia coli species results in highly diverseadaptive pathsrdquo PLoS Genetics vol 5 no 1 Article ID e10003442009
[13] J M Janda and S L Abbott ldquo16S rRNA gene sequencingfor bacterial identification in the diagnostic laboratory plusesperils and pitfallsrdquo Journal of Clinical Microbiology vol 45 no9 pp 2761ndash2764 2007
[14] J-J Lu C-L Perng S-Y Lee and C-CWan ldquoUse of PCRwithuniversal primers and restriction endonuclease digestions fordetection and identification of common bacterial pathogens incerebrospinal fluidrdquo Journal of Clinical Microbiology vol 38 no6 pp 2076ndash2080 2000
[15] N M Guentzel ldquoEscherichia Klebsiella Enterobacter SerratiaCitrobacter andProteusrdquo inMedicalMicrobiology S Baron Edchapter 26 University of Texas Medical Branch at GalvestonGalveston Tex USA 1996
[16] P Behzadip E Behzadi H Yazdanbod R Aghapour A MCheshmeh and S D Omran ldquoA survey on urinary tract infec-tions associated with the three most common uropathogenicbacteriardquoMaedica (Buchar) vol 5 no 2 pp 111ndash115 2010
[17] B CMetri P Jyothi andBV Peerapur ldquoAntibiotic resistance inCitrobacter spp isolated from urinary tract infectionrdquo UrologyAnnals vol 5 no 4 pp 312ndash313 2013
[18] N Taneja S S Chatterjee M Singh S Singh and M SharmaldquoPediatric urinary tract infections in a tertiary care center fromnorth Indiardquo Indian Journal of Medical Research vol 131 no 1pp 101ndash105 2010
[19] J B Kaper J P Nataro and H L T Mobley ldquoPathogenicEscherichia colirdquo Nature Reviews Microbiology vol 2 no 2 pp123ndash140 2004
[20] E J de Muinck K Lagesen J E Afset et al ldquoComparisonsof infant Escherichia coli isolates link genomic profiles withadaptation to the ecological nicherdquo BMC Genomics vol 14 no1 article 81 2013
[21] D A Rasko M J Rosovitz G S A Myers et al ldquoThepangenome structure of Escherichia coli comparative genomicanalysis of E coli commensal and pathogenic isolatesrdquo Journalof Bacteriology vol 190 no 20 pp 6881ndash6893 2008
[22] C T Archer J F Kim H Jeong et al ldquoThe genome sequenceof E coli W (ATCC 9637) comparative genome analysis andan improved genome-scale reconstruction of E colirdquo BMCGenomics vol 12 article no 9 2011
[23] PKuhnert J Nicolet and J Frey ldquoRapid and accurate identifica-tion of Escherichia coliK-12 strainsrdquoApplied and EnvironmentalMicrobiology vol 61 no 11 pp 4135ndash4139 1995
[24] A P Bauer S M Dieckmann W Ludwig and K-H SchleiferldquoRapid identification of Escherichia coli safety and laboratorystrain lineages based on multiplex-PCRrdquo FEMS MicrobiologyLetters vol 269 no 1 pp 36ndash40 2007
[25] B J Bachmann ldquoPedigrees of some mutant strains ofEscherichia coli K-12rdquo Bacteriological Reviews vol 36 no 4 pp525ndash557 1972
[26] P Daegelen F W Studier R E Lenski S Cure and J F KimldquoTracing ancestors and relatives of Escherichia coli B and thederivation of B strains REL606 and BL21(DE3)rdquo Journal ofMolecular Biology vol 394 no 4 pp 634ndash643 2009
[27] F W Studier P Daegelen R E Lenski S Maslov and J F KimldquoUnderstanding the differences between genome sequences ofEscherichia coli B strains REL606 and BL21(DE3) and compar-ison of the E coli B and K-12 genomesrdquo Journal of MolecularBiology vol 394 no 4 pp 653ndash680 2009
[28] R K Selander and B R Levin ldquoGenetic diversity and structurein Escherichia coli populationsrdquo Science vol 210 no 4469 pp545ndash547 1980
[29] D M Gordon O Clermont H Tolley and E DenamurldquoAssigning Escherichia coli strains to phylogenetic groupsmulti-locus sequence typing versus the PCR triplex methodrdquoEnvironmental Microbiology vol 10 no 10 pp 2484ndash24962008
[30] P Desjardins B Picard B Kaltenbock J Elion and EDenamur ldquoSex in Escherichia coli does not disrupt the clonalstructure of the population evidence from random amplifiedpolymorphic DNA and restriction-fragment-length polymor-phismrdquo Journal of Molecular Evolution vol 41 no 4 pp 440ndash448 1995
Journal of Pathogens 11
[31] N D Jasuja R Saxena S Chandra and S C Joshi ldquoIsolationand identification of microorganism from polyhouse agricul-ture soil of RajasthanrdquoAfrican Journal ofMicrobiology Researchvol 7 no 41 pp 4886ndash4891 2013
[32] V Hemraj S Diksha and G Avneet ldquoA review of commonlyused biochemical test for bacteriardquo Innovare Journal of LifeSciences vol 1 pp 1ndash7 2013
[33] J D Thompson D G Higgins and T J Gibson ldquoCLUSTALWimproving the sensitivity of progressive multiple sequencealignment through sequence weighting position-specific gappenalties and weight matrix choicerdquoNucleic Acids Research vol22 no 22 pp 4673ndash4680 1994
[34] T Vincze J Posfai and R J Roberts ldquoNEBcutter a program tocleave DNA with restriction enzymesrdquo Nucleic Acids Researchvol 31 no 13 pp 3688ndash3691 2003
[35] J Ye G Coulouris I Zaretskaya I Cutcutache S Rozen andT Madden ldquoPrimer-BLAST a tool to design target-specificprimers for polymerase chain reactionrdquo BMC Bioinformaticsvol 13 article 134 2012
[36] J R Parvathi and S Singh ldquoFacile approaches for microbialDNA extraction comparative study with colonies lsquopickedrsquo fromthree different bacteriological mediumrdquo Journal of Pure andApplied Microbiology vol 9 no 3 pp 2373ndash2378 2015
[37] R Lan and P R Reeves ldquoEscherichia coli in disguise molecularorigins of Shigellardquo Microbes and Infection vol 4 no 11 pp1125ndash1132 2002
[38] D J Leininger J R Roberson and F Elvinger ldquoUse ofeosin methylene blue agar to differentiate Escherichia coli fromother gram-negative mastitis pathogensrdquo Journal of VeterinaryDiagnostic Investigation vol 13 no 3 pp 273ndash275 2001
[39] U Dobrindt F Agerer K Michaelis et al ldquoAnalysis of genomeplasticity in pathogenic and commensal Escherichia coli isolatesby use of DNA arraysrdquo Journal of Bacteriology vol 185 no 6pp 1831ndash1840 2003
[40] V Savolainen R S Cowan A P Vogler G K Roderickand R Lane ldquoTowards writing the encyclopaedia of life anintroduction to DNA barcodingrdquo Philosophical Transactions ofthe Royal Society B Biological Sciences vol 360 no 1462 pp1805ndash1811 2005
[41] T A Isenbarger C E Carr S S Johnson et al ldquoThe mostconserved genome segments for life detection on earth andother planetsrdquo Origins of Life and Evolution of Biospheres vol38 no 6 pp 517ndash533 2008
[42] S R Santos and H Ochman ldquoIdentification and phylogeneticsorting of bacterial lineages with universally conserved genesand proteinsrdquoEnvironmentalMicrobiology vol 6 no 7 pp 754ndash759 2004
[43] R Rastogi M Wu I DasGupta and G E Fox ldquoVisualizationof ribosomal RNA operon copy number distributionrdquo BMCMicrobiology vol 9 no 1 article 208 2009
[44] J A Klappenbach J M Dunbar and T M Schmidt ldquorRNAoperon copy number reflects ecological strategies of bacteriardquoApplied and Environmental Microbiology vol 66 no 4 pp1328ndash1333 2000
[45] J A Klappenbach P R Saxman J R Cole and T M SchmidtldquoRrndb the ribosomal RNA operon copy number databaserdquoNucleic Acids Research vol 29 no 1 pp 181ndash184 2001
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom
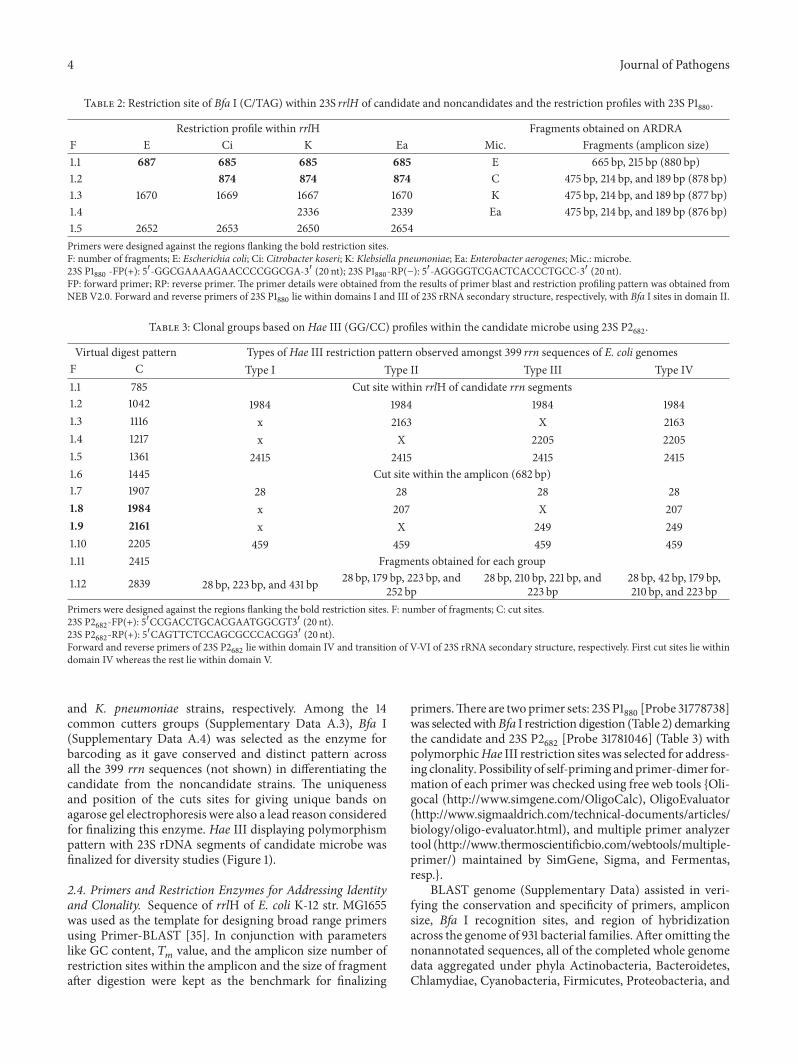
4 Journal of Pathogens
Table 2 Restriction site of Bfa I (CTAG) within 23S rrlH of candidate and noncandidates and the restriction profiles with 23S P1880
Restriction profile within rrlH Fragments obtained on ARDRAF E Ci K Ea Mic Fragments (amplicon size)11 687 685 685 685 E 665 bp 215 bp (880 bp)12 874 874 874 C 475 bp 214 bp and 189 bp (878 bp)13 1670 1669 1667 1670 K 475 bp 214 bp and 189 bp (877 bp)14 2336 2339 Ea 475 bp 214 bp and 189 bp (876 bp)15 2652 2653 2650 2654Primers were designed against the regions flanking the bold restriction sitesF number of fragments E Escherichia coli Ci Citrobacter koseri K Klebsiella pneumoniae Ea Enterobacter aerogenes Mic microbe23S P1
880-FP(+) 51015840-GGCGAAAAGAACCCCGGCGA-31015840 (20 nt) 23S P1
880-RP(minus) 51015840-AGGGGTCGACTCACCCTGCC-31015840 (20 nt)
FP forward primer RP reverse primer The primer details were obtained from the results of primer blast and restriction profiling pattern was obtained fromNEB V20 Forward and reverse primers of 23S P1
880lie within domains I and III of 23S rRNA secondary structure respectively with Bfa I sites in domain II
Table 3 Clonal groups based on Hae III (GGCC) profiles within the candidate microbe using 23S P2682
Virtual digest pattern Types of Hae III restriction pattern observed amongst 399 rrn sequences of E coli genomesF C Type I Type II Type III Type IV11 785 Cut site within rrlH of candidate rrn segments12 1042 1984 1984 1984 198413 1116 x 2163 X 216314 1217 x X 2205 220515 1361 2415 2415 2415 241516 1445 Cut site within the amplicon (682 bp)17 1907 28 28 28 2818 1984 x 207 X 20719 2161 x X 249 249110 2205 459 459 459 459111 2415 Fragments obtained for each group
112 2839 28 bp 223 bp and 431 bp 28 bp 179 bp 223 bp and252 bp
28 bp 210 bp 221 bp and223 bp
28 bp 42 bp 179 bp210 bp and 223 bp
Primers were designed against the regions flanking the bold restriction sites F number of fragments C cut sites23S P2
682-FP(+) 51015840CCGACCTGCACGAATGGCGT31015840 (20 nt)
23S P2682
-RP(+) 51015840CAGTTCTCCAGCGCCCACGG31015840 (20 nt)Forward and reverse primers of 23S P2
682lie within domain IV and transition of V-VI of 23S rRNA secondary structure respectively First cut sites lie within
domain IV whereas the rest lie within domain V
and K pneumoniae strains respectively Among the 14common cutters groups (Supplementary Data A3) Bfa I(Supplementary Data A4) was selected as the enzyme forbarcoding as it gave conserved and distinct pattern acrossall the 399 rrn sequences (not shown) in differentiating thecandidate from the noncandidate strains The uniquenessand position of the cuts sites for giving unique bands onagarose gel electrophoresis were also a lead reason consideredfor finalizing this enzyme Hae III displaying polymorphismpattern with 23S rDNA segments of candidate microbe wasfinalized for diversity studies (Figure 1)
24 Primers and Restriction Enzymes for Addressing Identityand Clonality Sequence of rrlH of E coli K-12 str MG1655was used as the template for designing broad range primersusing Primer-BLAST [35] In conjunction with parameterslike GC content 119879
119898value and the amplicon size number of
restriction sites within the amplicon and the size of fragmentafter digestion were kept as the benchmark for finalizing
primersThere are twoprimer sets 23S P1880
[Probe 31778738]was selectedwithBfa I restriction digestion (Table 2) demarkingthe candidate and 23S P2
682[Probe 31781046] (Table 3) with
polymorphicHae III restriction siteswas selected for address-ing clonality Possibility of self-priming andprimer-dimer for-mation of each primer was checked using free web tools Oli-gocal (httpwwwsimgenecomOligoCalc) OligoEvaluator(httpwwwsigmaaldrichcomtechnical-documentsarticlesbiologyoligo-evaluatorhtml) and multiple primer analyzertool (httpwwwthermoscientificbiocomwebtoolsmultiple-primer) maintained by SimGene Sigma and Fermentasresp
BLAST genome (Supplementary Data) assisted in veri-fying the conservation and specificity of primers ampliconsize Bfa I recognition sites and region of hybridizationacross the genome of 931 bacterial families After omitting thenonannotated sequences all of the completed whole genomedata aggregated under phyla Actinobacteria BacteroidetesChlamydiae Cyanobacteria Firmicutes Proteobacteria and
Journal of Pathogens 5
Spirochaetae of the bacterial kingdom were considered forthe analysis
25 DNA Isolation Amplification and Restriction DigestionDNA isolation was performed according to Parvathi andSingh [36] The reaction soup comprised 2120583L of supernatant(DNA) 05 120583M of each forward and reverse primer 1x ofassay buffer (10mM Tris-HCl pH 88 50mM KCl) MgCl
2
(15mM) dNTPmix (800 120583M) and 05U120583LTaq polymerase(Kappa Systems KK5004) Standard genomic DNA of Ecoli K-12 (Merck) and nuclease-free water were used aspositive and negative control The reactions were carriedout in a thermal cycler (Bioer) with the program set for25 cycles comprising denaturation (95∘C) annealing (68∘C)and extension (72∘C) for 30 s each 3 120583L of the PCR productwas digested with 2U of respective enzyme (FermentasThermo Scientific) in 5120583L reactionmix at 37∘C for two hoursThe amplicons and their restriction products were analysedby agarose gel electrophoresis (AGE) at 5Vcm in 2 and25 agarose gels respectively each containing 05120583gmL ofEtBr The gels were viewed and photographed in BioImagingSystems (Syngene)
3 Result and Discussion
31 Genetic Diversity within 23S Ribosomal Repeats for Identi-fication of E coli 23S rRNA gene showed 087 intergenicvariations compared with 16S rRNA within the referencegenomes (Supplementary Data A5) The result seemed obvi-ous due to the increased frame size of the latter but thisalso encouraged perusing microheterogenicity analysis ofthe largest ribosomal segment for further approach Thesizes of target segments varied from 2903 to 2907 bp withmaximum gene length observed in ATCC8739 (Crookrsquosstrain) accounting for 2970ndash2985 bp followed by E24377Adisplaying sequences of 2923ndash2925 bp One rrn of UM 146and ETECH10407 were of 1762 bp and 1561 bp respectively(Supplementary Data A9) Absolutely no link between rrnsize and pathogenicity could be defined because the varia-tions were random ATCC8739 being a laboratory cultureof fecal origin and UM146 an EIAC strain E23477A andETECH10407 of ETEC origin
Ribosomal genes were found to be distributed acrosscomplementary and regular strand with majority of thegenomes (46 out of 57) showing segregation in five regularand two complementary strands All the 23S rrn sequencesdistributed within the regular and complementary strandsacross respective genomes were individually sorted for MSAto attain an understanding of intragenic variation Analysisrevealed the highest level of mismatches in complementarystrands compared with their counterparts (SupplementaryData A6) The highest level of mismatches mounting to06 corresponding to 50 of the targeted gene size wasperceived in complementary strands of P12b (commensal)E23489 (EPEC) and ETEC H10407 The greatest dispar-ity among sequences in regular strand was observed inAPEC 01 with 003 (882903) followed by CFT043 (UPEC)and 042 (EAEC) having 0016 (sim42ndash512903) dissimilarity23S ribosomal copies of ED1a (commensal) showed zero
percent mismatches in both complementary and regularstrands making it an ideal cluster with the highest degreeof conservation Similar values of mismatches were seen inSE11 (Commensal) CB9615 (EPEC) and 0104H4 2009El-2011 (EAEC) These results indicate that there is no cor-relation between intragenic diversity of target stretch andpathogenicity of strains These differences substantiated 23SRNA clusters having microdiverse organization with bothconserved and variable stretches within them Sequevarsarising due to this heterogeneity unavoidably lead to biasedresults differential amplification and chimeric moleculesThus the method of screening and targeting restriction pro-file pattern within a region of choice (ribosomal segments)and specific to bacteria of interest demonstrates authentic andsimpler practice than routine molecular strategy involvingamplification and sequencing
32 Barcoding E coli from Its Colony Morphovars ARDRAof 23S P1880 with Bfa I PCR amplification of the stan-dard strains and isolates of different origin (commensalurine and clinical sample) with 23S P1
880gave amplicons
(880 bp) of high intensity with no primer-dimer formation(Figure 2(a)) Restriction digestion of the amplicons withBfa I gave unique fragments of 665 bp and 215 bp in E coliwhereas C koseri E aerogenes and K pneumoniae eachgave fragments of 475 bp 214 bp and 189 bp respectively(Figure 2(b)) The genotypic strategy proved superior tobiochemical characterization taking into consideration all thepossible positive negative and false positive and negativeresults across 25 isolates each from clinical commensal andenvironmental origin (Supplementary Data A7) 18 (72)of clinical 22 (88) of commensal and all the 25 (100)environment isolates showed distinct restriction pattern likethat of E coli BLAST genome search across 931 bacterialfamilies verified the specificity of primers and restrictionpreference for Bfa I The primers showed exclusive bindingonly with Enterobacteriaceae family (Supplementary DataA8) Among the various strains in the family only Shigellaspp Salmonella enterica (Arizona) and Proteus mirabilusexhibited similar restriction preference to that of E coli Inspite of Shigella spp being taxa in disguise of E coli [37] theycan be easily screened using selective medium because of nolactose fermentation [38] whereas Salmonella enterica andProteus mirabilis showed amplicon heterogeneity of 1073 bpand 880 bp during amplification thus making it easy to avoiderroneous conclusions and misinterpretation
33 Categorizing E coli StrainsUsingARDRAof 23S P2682 withHae III A detailed analysis of the Hae III profile across 399sequences [Supplementary Data A9] of ribosomal segmentsof E coli indicated four possible profiles types I II IIIand IV with varying degree of predominance within thesesegments The rest of the candidate strains showed type IIIpatterns but types I and IV were exclusive for E coli strainsMajority of genomes (119899 = 32) as in 15 nonpathogenic and17 pathogenic strains showed type I profiles (57) followedby type III (35) predominating in 15 strains Six strainsshowed type IV pattern (10) of which four were B strainsType I groups represent E coli strains that lacked recognition
6 Journal of Pathogens
1 7
880 bp
65432
(a)
8
665bp
475bp
215 bp 214 bp
178bp
1 765432
(b)
Figure 2 (a) Broad Range PCR of 23S P1880
with genomic DNA isolated from standard strains Lane 1 100 bp ladder Lane 2 blank Lane 3E coli Lane 4 C freundii Lane 5 C koseri Lane 6 K pneumonia Lane 7 E aerogenes PCR product of single representative of E coli wasloaded (b) ARDRA of 23S P1
880
amplicons with Bfa I Lanes 1 2 and 3 E coli (ATCC15223 ATCC25922 and ATCC11775) Lane 4C freundii(ATCC8090) Lane 5 C koseri (ATCC27028) Lane 6 K pneumonia (ATCC33495) Lane 7 E aerogenes (ATCC13048) Lane 8 100 bp ladder
elements due to change at 2205th and 2161st base with respectto 23S rDNA or with reference to amplicons at 207th or 249thbase positions Type IV strains are contrary to type I withintact recognition sequence at both of these sites Strains withtype II profiles lacked 2161st (249th) cut site and type IIIlacked the 2205th (207th) cut positions respectively Type IIwas the rarest with its patterns observed to be ambiguous incertain rrn segments
Heterogenicity in few genomes was noted two rrn amongseven repeats ofW (NC 0176851 each in regular and comple-mentary strand) and KO11FL (NC 0176601 each in regularand complementary strand) and one in KO11FL (NC 0169021 regular) showed type II profile in spite of type I beingtheir primary pattern One among seven of the B strains REL606 (NC 0129671 regular) BL21-Gold (NC 0129471 com-plementary) BL21DE3 (NC 0129712 regular) and BL21DE3(NC 0128922 regular) showed type I profiles apart frombeing prevalent with type IV pattern Similarly two rrnamongNRG 873C (NC 0176341 each in regular and comple-mentary strand) and LF82 (NC 0119931 each in regular andcomplementary strand) and one in CFT073 (NC 0044311regular) showed type I pattern while exhibiting type IIIpattern predominance APEC01 (NC 0085631) being primein type III showed one rrn segment of type I and type IV(both in regular strand) E coli strain 042 (NC 0176261)displayed preponderance in type I pattern but three rrn (tworegular and one complementary) copies exhibited type IIIpattern One of the regular rrn elements in LF82 showed typeIV pattern The fragmentation pattern obtained by ARDRAof 23S P2
682with Hae III in the standard samples showed
that all K-12 C W Crooks and their respective descendantsexhibited type I profileswith type IVprofiles being exclusivelypresented by B strains (Figures 3(a) and 3(b))
Among the isolates 21 out of 22 commensal and 16 of18 clinical samples showed type I pattern heterogenicity ofpatterns was not observed These results reaffirmed predom-inance (93) of type I in nature Multisequence typing hasshown commensal strains to be categorized primarily undergroup A or group B1 whereas groups B2 D and E comprisegenerally pathogenic strains K-12 B and Crooks strain comeunder phylogroup A whereas strain W is a B1 phylogroup
34 Local Alignment of 23S rrn Hae III Profiles versus GlobalAlignment to Establish Clonality in E coli A number ofgenes that are of core dispensable and unique in naturemake up for pangenome the latter two are gained due torecombinational or transpositional events To obtain similarclustering on comparing whole genome would emphasizethe significance of these restriction profiles and help inestablishing the clonal nature and predominance of a singleclone Hence dendogram constructs with 682 bp ampliconfrom a representative of each restriction profile will showdistinct clustering of these profiles due to the fact that a smallsubset of segments within the whole genome was consideredThe global alignment of whole genome sequences (Figure 4)of the entire 57 E coli strains displayed distinct concentrateof type I III and IV profiles An intrinsic analysis of thespot of divergence and respective clade formations showedinteresting and riveting findings The strains at the ancestralroot showed type I pattern and the next divergent node point(D1) displayed formation of two distinct clans of type I andtype IV profiles segregated at the maximum distance fromone another Numbering of nodes was done by consideringthe precedence of branch point at every node A possiblescenario of two additional mutational events that are oftransitional nature (A to G) at 2205th and 2161st bases has
Journal of Pathogens 7
682bp
987654321
9998400
8998400
7998400
6998400
5998400
4998400
3998400
2998400
1998400
(a)
431bp
223 bp
210 bp
179bp
223 + 210 bp
223+
210+
87654321
8998400
7998400
6998400
5998400
4998400
3998400
2998400
1998400
(b)
Figure 3 (a) Broad range PCR of 23S P2682
with genomic DNA isolated from standard strains of E coli The upper row comprises Lane 1K-12 strains Lane 2 derivatives of K-12 strains Lane 3 DHI strain Lane 4 strain C1 Lane 6 Crookrsquos strain Lane 7 Walkerrsquos strain Lane 8descendent of W Lane 9 blank The lower row comprises Lane 11015840 B strain Lane 21015840 BL21 strain Lane 31015840 BL21DE3 Lane 41015840 BL21DE3Lys5(B3) Lane 61015840 E coli (ATCC 15223 commensal isolate) Lane 71015840 E coli (ATCC 11775 urine sample) Lane 81015840 E coli (ATCC 25922 clinical)Lanes 551015840 and Lanes 991015840 in both rows are 100 bp ladder and blank respectively (b) ARDRA of 23S P2
682
amplicons with Hae III Lane 1 ofboth rows comprises 100 bp ladder The digested products are loaded as per the order given in Figure 2(a)
introduced Hae III recognition patterns D6 gave lineages ofB1 phylogroups D9 giving rise to B2 clades (D12) of type Iprofiles and D clades with varied Hae III patterns (D11) D10gave rise to two nodes (with strains being type I groups)with one showing B1 phylogroups (D13) and the other beingpredominant in E phylogroups followed by B1 An almostexemplary corelation between the clonal groups and differentpathovars was found with this study Majority of commensalstrains and EIEC and all EAEC and EPEC were shown tobe having type I profile (Table 4) All UPEC ExPEC APECEIEC and single strain of EPEC and two commensal strainsshowed type III pattern B form strains along with a singlestrain of UPEC and environmental isolate exhibited type IVprofiles
Similarly a near-pristine association between phylo-groups and Hae III profiles was established Strains comingunder phylogroups A were preponderant in type I whereastype IV profiles were exclusively for B forms Type I profileswere observed in the strains (119899) in the order of A gt B1 gt E gtDwith type III dominant in B2 strains Phylogroup D showedequal occurrence of type III and type IV fragmentationpattern followed by a single strain with type I profile Theseresults assisted in enumerating a conjecture which works upa scenario of primordial microbial population where E colistrains had two clonal types (Figure 5(a)) which coexistedtogether (type I and type IV clones) Of these type I strainswould have been the predominant clone in comparison totype IV based on the dendogram data with maximum genepool of root strains showing type I Type III may have beenthe next predominant clone (an intermediary one) that hasevolved either from type I (by addition of a new recognition
Table 4 Corelation between phylogroups Hae III profiles andnature of E coli strains (genome data analysis of 57 E coli strainscollated from Genbank)
Strain Type A B1 B2 D E Total Out ofCommensal I 10 5 mdash mdash mdash 15 21Commensal III mdash mdash 2 mdash mdash 2 21Commensal IV 4 mdash mdash mdash mdash 4 21EPEC I mdash mdash mdash mdash 7 7 8EPEC III mdash mdash 1 mdash mdash 1 8EAEC I mdash 4 mdash 1 mdash 5 5EHEC I mdash 2 mdash mdash mdash 2 3EHEC II mdash 1 mdash mdash mdash 1 3ETEC I 1 1 1 mdash mdash 3 3UPEC III mdash mdash 7 mdash mdash 7 8UPEC IV mdash mdash mdash 1 mdash 1 8EIEC III mdash mdash 3 mdash mdash 3 3ExPEC III mdash mdash 2 2 mdash 4 4APEC III mdash mdash 1 mdash 1 1ENV IV mdash mdash mdash 1 1 1APEC avian pathogenic EAEC enteroaggressive EHEC enterohemorrhag-ic ETEC enterotoxigenic EIEC enteroinvasive EPEC enteropathogenicENV environmental ExPEC extraintestinal pathogenic UPEC uropatho-genic
site) or from type IV (by deletion of one recognition site)strains In a different scenario maybe all three types coexistedor type III would have given rise to type IVwith an additionalmutational event This situation may be hardly convincing
8 Journal of Pathogens
D1
D2
D3
D4
D5
D6
D7
D8
D9
D10
D11
D12
D13
D14
BL21(DE3)_1lowastBL21(DE3)_2lowast
BL21-Gold(DE3)pLysS AGlowast
B str REL606lowastA phylogroup
ETEC H10407lowast (B2)
UMNK88 (A)
(B1 clades) Str 2009EL-2050Str 2009EL-2071
Str 2011-349355989
B1 phylogroup
Str EC4115Str TW14359
Str EDL933Str Sakai
Xuzhou21Str CB9615
Str RM12598
E phylogroup
Str 11128Str 11368 B1 phylogroup
NA114
SE15LF82lowastlowastlowastStr NRG 857Clowastlowastlowast
UM146
UTI89IHE3034
APEC O1lowastlowastS88
Clone D i2Clone D i14
ABU 83972
CFT073536
ED1aStr E234869
B2 phylogroup
UMN026
IAI39Str CE10
SMS-3-5
D phylogroup
O42lowastlowastlowast
(B2 clades)
(D clades)
(B1 clades)
Wlowastlowastlowast
WKO11FLlowastlowast
KO11FLE24377A
SE11
B1 phylogroup
IAI1ATCC 8739
P12blowastHS
BW2952
K-12 substr DH10BDH1
DH1 (ME8569)K-12 substr W3110
K-12 substr MG1655
A phylogroup
1
2
10
3156
Str 12009lowastlowast
Commensal ETEC EHEC EPEC UPEC
ExPECAPECEAECEIECENV
Figure 4 Analysis of distribution of Hae III profiles phylogroups and pathovars within the global alignment dendogram constructcomprising 57 E coli genomes The dendogram of the whole genome sequence of 57 genomes was obtained from the NCBI web site forE coli and was redrawn using Photoshop CS6 for colour coordination for pathogenic and nonpathogenic strains and inputting the clonalgroups and phylogroups (httpwwwncbinlmnihgovgenometerm=escherichia+coli) Among the 57 genomes 32 were of type I (56)18 were of type III (31) and 6 and 1 were of type IV (10) and type II (2) respectively showing that type I is predominant and type IIthe rarest To correlate between the phylogroupsHae III profiles and pathovars the lineages were considered as clades derived from variousdivergence point (D) during the course of evolution
as dendogram segregation shows type III strains evolving orpresent after type IV strains The clustering also shows thatpredominance of type III is more than type IV and maybeevolutionary processesmutation have triggered the increasein type III profiles On similar line type II clones would havebeen evolved from type I (addition) or type IV (deletion)but due to the very limited type II restriction profile patterns(rare) as observed from the gene pool data the chances orapproximation of such event is difficult to prove Based onthese results no new insights into phylogroup distributionacross candidate strains can be provided however withrespect to the Hae III profile an additional angle can beconsidered Phylogroups A and B1 are predominant withtype I Hae III profiles whereas B2 phylogroups exclusivelyrepresented type III profile (Figure 5(b))
An intrinsic metagenomic and pangenomic analysis ofmost bacterial species exhibited mosaicisms of their inherentblueprint [39] In such situations core gene elements sharedacross the species are limited in comparison to dispensable(flexible) genetic contents that concede uniqueness to aparticular strain These flexible contents help in niche adapt-ability and give bacteria the ability to infect and invade Amajor concern is while diagnosing both true and opportunistpathogenic bacteria the former causing disease in virtuallyany susceptible host and the latter invoking diseases inimmunocompromised individuals ldquoMolecular barcodingrdquoan evolved term which implies the method of allele specificamplification of relatively small DNA sequences or barcodes[40] for spotting a particular species is a thrust area formicrobial diagnostics A plethora of functional genes ranging
Journal of Pathogens 9
Type I(28 459)
Type IV(28 207 249 and 459)
Hypothesis
Type III(28 249 and 459)
+ minus
+
Primordial
NonpathogenicA gt B1 gt B2
type I gt IV gt III
PathogenicB2 gt B1 gt E gt D
type I (IAT)III(P)gt type IVII
Predominant clones
Rarest
+minus
Type II(28 207 and 459)
Restriction profilesand
clonesPhylogroups and
clones
2∘ clonesintermediate
(a) (b)
Figure 5 (a) Hypothesis of clonality in E coli strains based on 23S rrn restriction profiling The primordial soup of E coli microbes wouldhave been comprised of two clonal groups types I and IV coexisting with each other but predominated by type I strains Over the courseof evolution on accumulating mutation as a means for variation type I or type IV would have given rise to type III by addition (former) ordeletion (latter) of recognition sequence The second scenario could be a rare event because accumulation of two mutations in a conservedregion would be difficult Type II being the rarest would have been evolved in similar basis to type III (b) Hypothesis of linking phylogroupsHae III profiles and clonal types in E coli This diagram illustrates the clonal groups of commensal E coli strains having both A phylogroups(dominant) and B1 phylogroups each being type I profiles Pathogenic counterparts showed dominancy in type III restriction pattern followedby type I pattern With respect to phylogroups pathovars were predominant in B2 and B1 followed by D and E groups
from housekeeping to repair are employed for characterizingbacteria of which rRNA is referred to as the gold stretch[41 42] Ribosomal segments organized as operon clusters of16S 23S and 5S rDNA and these assemblages are distributedfrom single to multiple copies across the bacterial genophore[43] The occurrence of redundancy in maintaining differentcopies of these genes is not associated with virulence but anin-built ecological strategy that contributes to the particularbacterium with a competitive edge and better chance ofsurvival [44] A futuristic approach of coupling the techniqueand results with antibiotic resistance pattern colony char-acteristics and genomic profiles of bacterial strains of bothcommensal and pathogenic nature can help in monitoringand managing infections in a better manner
4 Conclusion
The causatum of postgenomic era genomics and proteomicshas enhanced this progress by improvising the techniquesused for diagnostics The strategy of ARDRA utilized inthis study was rapid and a cost-effective approach to dif-ferentiate E coli from its colony morphovars along withgiving a predicament of nature of origin or clonal groupsThe mutations reported in this work for detection andestablishing clonality namely T874A (Bfa I) G2161A andG2205A (Hae III) have not been mentioned in any researchpaper or within the Ribosomal RNA Mutation Database
[45] making this finding aboriginal This basic methodologycan be adapted for detection of other bacterial organismsby targeting SNPs linked to recognition sequences therebyvalidating rrn sequences as marker for diagnostics A cumu-lative approach of detection of unique restriction pattern of atargeted gene and its cross-reference across bacterial familiescan circumvent changes of false positives
Disclosure
This work was presented (poster) in part at the 5th Interna-tional Barcoding of Life Kunming China 2013
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
Acknowledgments
The authors acknowledge and express their gratitude to Pro-fessor D A Bhiwgade (Ex-Director) and Professor DebjaniDasgupta Director School of Biotechnology amp Bioinformat-ics D Y Patil University as well as the management for theirsupport and for providing the infrastructure to carry out thecurrent work
10 Journal of Pathogens
References
[1] M Richter and R Rossello-Mora ldquoShifting the genomic goldstandard for the prokaryotic species definitionrdquo Proceedings ofthe National Academy of Sciences of the United States of Americavol 106 no 45 pp 19126ndash19131 2009
[2] A F Auch M von Jan H-P Klenk and M Goker ldquoDigitalDNA-DNA hybridization for microbial species delineation bymeans of genome-to-genome sequence comparisonrdquo Standardsin Genomic Sciences vol 2 no 1 pp 117ndash134 2010
[3] D H Martin M Nsuami J Schachter et al ldquoUse of multiplenucleic acid amplification tests to define the infected-patientlsquogold standardrsquo in clinical trials of new diagnostic tests forChlamydia trachomatis infectionsrdquo Journal of Clinical Microbi-ology vol 42 no 10 pp 4749ndash4758 2004
[4] E A Mothershed and A M Whitney ldquoNucleic acid-basedmethods for the detection of bacterial pathogens presentand future considerations for the clinical laboratoryrdquo ClinicaChimica Acta vol 363 no 1-2 pp 206ndash220 2006
[5] J E Clarridge III ldquoImpact of 16S rRNA gene sequence analysisfor identification of bacteria on clinical microbiology andinfectious diseasesrdquo Clinical Microbiology Reviews vol 17 no4 pp 840ndash862 2004
[6] O Mizrahi-Man E R Davenport and Y Gilad ldquoTaxonomicclassification of bacterial 16S rRNA genes using short sequenc-ing reads evaluation of effective study designsrdquo PLoS ONE vol8 no 1 Article ID e53608 2013
[7] D E Lebonah A Dileep K Chandrasekhar S Sreevani BSreedevi and J Pramoda Kumari ldquoDNA barcoding on bacteriaa reviewrdquo Advances in Biology vol 2014 Article ID 541787 9pages 2014
[8] J Rajendhran and P Gunasekaran ldquoMicrobial phylogeny anddiversity small subunit ribosomal RNA sequence analysis andbeyondrdquo Microbiological Research vol 166 no 2 pp 99ndash1102011
[9] R Srinivasan U Karaoz M Volegova et al ldquoUse of 16S rRNAgene for identification of a broad range of clinically relevantbacterial pathogensrdquo PLoS ONE vol 10 no 2 Article IDe0117617 2015
[10] S Chakravorty D Helb M Burday N Connell and D AllandldquoA detailed analysis of 16S ribosomal RNA gene segments forthe diagnosis of pathogenic bacteriardquo Journal of MicrobiologicalMethods vol 69 no 2 pp 330ndash339 2007
[11] C R Woese ldquoReview bacterial evolutionrdquo MicrobiologicalReviews vol 51 pp 221ndash271 1987
[12] M Touchon C Hoede O Tenaillon et al ldquoOrganised genomedynamics in the Escherichia coli species results in highly diverseadaptive pathsrdquo PLoS Genetics vol 5 no 1 Article ID e10003442009
[13] J M Janda and S L Abbott ldquo16S rRNA gene sequencingfor bacterial identification in the diagnostic laboratory plusesperils and pitfallsrdquo Journal of Clinical Microbiology vol 45 no9 pp 2761ndash2764 2007
[14] J-J Lu C-L Perng S-Y Lee and C-CWan ldquoUse of PCRwithuniversal primers and restriction endonuclease digestions fordetection and identification of common bacterial pathogens incerebrospinal fluidrdquo Journal of Clinical Microbiology vol 38 no6 pp 2076ndash2080 2000
[15] N M Guentzel ldquoEscherichia Klebsiella Enterobacter SerratiaCitrobacter andProteusrdquo inMedicalMicrobiology S Baron Edchapter 26 University of Texas Medical Branch at GalvestonGalveston Tex USA 1996
[16] P Behzadip E Behzadi H Yazdanbod R Aghapour A MCheshmeh and S D Omran ldquoA survey on urinary tract infec-tions associated with the three most common uropathogenicbacteriardquoMaedica (Buchar) vol 5 no 2 pp 111ndash115 2010
[17] B CMetri P Jyothi andBV Peerapur ldquoAntibiotic resistance inCitrobacter spp isolated from urinary tract infectionrdquo UrologyAnnals vol 5 no 4 pp 312ndash313 2013
[18] N Taneja S S Chatterjee M Singh S Singh and M SharmaldquoPediatric urinary tract infections in a tertiary care center fromnorth Indiardquo Indian Journal of Medical Research vol 131 no 1pp 101ndash105 2010
[19] J B Kaper J P Nataro and H L T Mobley ldquoPathogenicEscherichia colirdquo Nature Reviews Microbiology vol 2 no 2 pp123ndash140 2004
[20] E J de Muinck K Lagesen J E Afset et al ldquoComparisonsof infant Escherichia coli isolates link genomic profiles withadaptation to the ecological nicherdquo BMC Genomics vol 14 no1 article 81 2013
[21] D A Rasko M J Rosovitz G S A Myers et al ldquoThepangenome structure of Escherichia coli comparative genomicanalysis of E coli commensal and pathogenic isolatesrdquo Journalof Bacteriology vol 190 no 20 pp 6881ndash6893 2008
[22] C T Archer J F Kim H Jeong et al ldquoThe genome sequenceof E coli W (ATCC 9637) comparative genome analysis andan improved genome-scale reconstruction of E colirdquo BMCGenomics vol 12 article no 9 2011
[23] PKuhnert J Nicolet and J Frey ldquoRapid and accurate identifica-tion of Escherichia coliK-12 strainsrdquoApplied and EnvironmentalMicrobiology vol 61 no 11 pp 4135ndash4139 1995
[24] A P Bauer S M Dieckmann W Ludwig and K-H SchleiferldquoRapid identification of Escherichia coli safety and laboratorystrain lineages based on multiplex-PCRrdquo FEMS MicrobiologyLetters vol 269 no 1 pp 36ndash40 2007
[25] B J Bachmann ldquoPedigrees of some mutant strains ofEscherichia coli K-12rdquo Bacteriological Reviews vol 36 no 4 pp525ndash557 1972
[26] P Daegelen F W Studier R E Lenski S Cure and J F KimldquoTracing ancestors and relatives of Escherichia coli B and thederivation of B strains REL606 and BL21(DE3)rdquo Journal ofMolecular Biology vol 394 no 4 pp 634ndash643 2009
[27] F W Studier P Daegelen R E Lenski S Maslov and J F KimldquoUnderstanding the differences between genome sequences ofEscherichia coli B strains REL606 and BL21(DE3) and compar-ison of the E coli B and K-12 genomesrdquo Journal of MolecularBiology vol 394 no 4 pp 653ndash680 2009
[28] R K Selander and B R Levin ldquoGenetic diversity and structurein Escherichia coli populationsrdquo Science vol 210 no 4469 pp545ndash547 1980
[29] D M Gordon O Clermont H Tolley and E DenamurldquoAssigning Escherichia coli strains to phylogenetic groupsmulti-locus sequence typing versus the PCR triplex methodrdquoEnvironmental Microbiology vol 10 no 10 pp 2484ndash24962008
[30] P Desjardins B Picard B Kaltenbock J Elion and EDenamur ldquoSex in Escherichia coli does not disrupt the clonalstructure of the population evidence from random amplifiedpolymorphic DNA and restriction-fragment-length polymor-phismrdquo Journal of Molecular Evolution vol 41 no 4 pp 440ndash448 1995
Journal of Pathogens 11
[31] N D Jasuja R Saxena S Chandra and S C Joshi ldquoIsolationand identification of microorganism from polyhouse agricul-ture soil of RajasthanrdquoAfrican Journal ofMicrobiology Researchvol 7 no 41 pp 4886ndash4891 2013
[32] V Hemraj S Diksha and G Avneet ldquoA review of commonlyused biochemical test for bacteriardquo Innovare Journal of LifeSciences vol 1 pp 1ndash7 2013
[33] J D Thompson D G Higgins and T J Gibson ldquoCLUSTALWimproving the sensitivity of progressive multiple sequencealignment through sequence weighting position-specific gappenalties and weight matrix choicerdquoNucleic Acids Research vol22 no 22 pp 4673ndash4680 1994
[34] T Vincze J Posfai and R J Roberts ldquoNEBcutter a program tocleave DNA with restriction enzymesrdquo Nucleic Acids Researchvol 31 no 13 pp 3688ndash3691 2003
[35] J Ye G Coulouris I Zaretskaya I Cutcutache S Rozen andT Madden ldquoPrimer-BLAST a tool to design target-specificprimers for polymerase chain reactionrdquo BMC Bioinformaticsvol 13 article 134 2012
[36] J R Parvathi and S Singh ldquoFacile approaches for microbialDNA extraction comparative study with colonies lsquopickedrsquo fromthree different bacteriological mediumrdquo Journal of Pure andApplied Microbiology vol 9 no 3 pp 2373ndash2378 2015
[37] R Lan and P R Reeves ldquoEscherichia coli in disguise molecularorigins of Shigellardquo Microbes and Infection vol 4 no 11 pp1125ndash1132 2002
[38] D J Leininger J R Roberson and F Elvinger ldquoUse ofeosin methylene blue agar to differentiate Escherichia coli fromother gram-negative mastitis pathogensrdquo Journal of VeterinaryDiagnostic Investigation vol 13 no 3 pp 273ndash275 2001
[39] U Dobrindt F Agerer K Michaelis et al ldquoAnalysis of genomeplasticity in pathogenic and commensal Escherichia coli isolatesby use of DNA arraysrdquo Journal of Bacteriology vol 185 no 6pp 1831ndash1840 2003
[40] V Savolainen R S Cowan A P Vogler G K Roderickand R Lane ldquoTowards writing the encyclopaedia of life anintroduction to DNA barcodingrdquo Philosophical Transactions ofthe Royal Society B Biological Sciences vol 360 no 1462 pp1805ndash1811 2005
[41] T A Isenbarger C E Carr S S Johnson et al ldquoThe mostconserved genome segments for life detection on earth andother planetsrdquo Origins of Life and Evolution of Biospheres vol38 no 6 pp 517ndash533 2008
[42] S R Santos and H Ochman ldquoIdentification and phylogeneticsorting of bacterial lineages with universally conserved genesand proteinsrdquoEnvironmentalMicrobiology vol 6 no 7 pp 754ndash759 2004
[43] R Rastogi M Wu I DasGupta and G E Fox ldquoVisualizationof ribosomal RNA operon copy number distributionrdquo BMCMicrobiology vol 9 no 1 article 208 2009
[44] J A Klappenbach J M Dunbar and T M Schmidt ldquorRNAoperon copy number reflects ecological strategies of bacteriardquoApplied and Environmental Microbiology vol 66 no 4 pp1328ndash1333 2000
[45] J A Klappenbach P R Saxman J R Cole and T M SchmidtldquoRrndb the ribosomal RNA operon copy number databaserdquoNucleic Acids Research vol 29 no 1 pp 181ndash184 2001
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom

Journal of Pathogens 5
Spirochaetae of the bacterial kingdom were considered forthe analysis
25 DNA Isolation Amplification and Restriction DigestionDNA isolation was performed according to Parvathi andSingh [36] The reaction soup comprised 2120583L of supernatant(DNA) 05 120583M of each forward and reverse primer 1x ofassay buffer (10mM Tris-HCl pH 88 50mM KCl) MgCl
2
(15mM) dNTPmix (800 120583M) and 05U120583LTaq polymerase(Kappa Systems KK5004) Standard genomic DNA of Ecoli K-12 (Merck) and nuclease-free water were used aspositive and negative control The reactions were carriedout in a thermal cycler (Bioer) with the program set for25 cycles comprising denaturation (95∘C) annealing (68∘C)and extension (72∘C) for 30 s each 3 120583L of the PCR productwas digested with 2U of respective enzyme (FermentasThermo Scientific) in 5120583L reactionmix at 37∘C for two hoursThe amplicons and their restriction products were analysedby agarose gel electrophoresis (AGE) at 5Vcm in 2 and25 agarose gels respectively each containing 05120583gmL ofEtBr The gels were viewed and photographed in BioImagingSystems (Syngene)
3 Result and Discussion
31 Genetic Diversity within 23S Ribosomal Repeats for Identi-fication of E coli 23S rRNA gene showed 087 intergenicvariations compared with 16S rRNA within the referencegenomes (Supplementary Data A5) The result seemed obvi-ous due to the increased frame size of the latter but thisalso encouraged perusing microheterogenicity analysis ofthe largest ribosomal segment for further approach Thesizes of target segments varied from 2903 to 2907 bp withmaximum gene length observed in ATCC8739 (Crookrsquosstrain) accounting for 2970ndash2985 bp followed by E24377Adisplaying sequences of 2923ndash2925 bp One rrn of UM 146and ETECH10407 were of 1762 bp and 1561 bp respectively(Supplementary Data A9) Absolutely no link between rrnsize and pathogenicity could be defined because the varia-tions were random ATCC8739 being a laboratory cultureof fecal origin and UM146 an EIAC strain E23477A andETECH10407 of ETEC origin
Ribosomal genes were found to be distributed acrosscomplementary and regular strand with majority of thegenomes (46 out of 57) showing segregation in five regularand two complementary strands All the 23S rrn sequencesdistributed within the regular and complementary strandsacross respective genomes were individually sorted for MSAto attain an understanding of intragenic variation Analysisrevealed the highest level of mismatches in complementarystrands compared with their counterparts (SupplementaryData A6) The highest level of mismatches mounting to06 corresponding to 50 of the targeted gene size wasperceived in complementary strands of P12b (commensal)E23489 (EPEC) and ETEC H10407 The greatest dispar-ity among sequences in regular strand was observed inAPEC 01 with 003 (882903) followed by CFT043 (UPEC)and 042 (EAEC) having 0016 (sim42ndash512903) dissimilarity23S ribosomal copies of ED1a (commensal) showed zero
percent mismatches in both complementary and regularstrands making it an ideal cluster with the highest degreeof conservation Similar values of mismatches were seen inSE11 (Commensal) CB9615 (EPEC) and 0104H4 2009El-2011 (EAEC) These results indicate that there is no cor-relation between intragenic diversity of target stretch andpathogenicity of strains These differences substantiated 23SRNA clusters having microdiverse organization with bothconserved and variable stretches within them Sequevarsarising due to this heterogeneity unavoidably lead to biasedresults differential amplification and chimeric moleculesThus the method of screening and targeting restriction pro-file pattern within a region of choice (ribosomal segments)and specific to bacteria of interest demonstrates authentic andsimpler practice than routine molecular strategy involvingamplification and sequencing
32 Barcoding E coli from Its Colony Morphovars ARDRAof 23S P1880 with Bfa I PCR amplification of the stan-dard strains and isolates of different origin (commensalurine and clinical sample) with 23S P1
880gave amplicons
(880 bp) of high intensity with no primer-dimer formation(Figure 2(a)) Restriction digestion of the amplicons withBfa I gave unique fragments of 665 bp and 215 bp in E coliwhereas C koseri E aerogenes and K pneumoniae eachgave fragments of 475 bp 214 bp and 189 bp respectively(Figure 2(b)) The genotypic strategy proved superior tobiochemical characterization taking into consideration all thepossible positive negative and false positive and negativeresults across 25 isolates each from clinical commensal andenvironmental origin (Supplementary Data A7) 18 (72)of clinical 22 (88) of commensal and all the 25 (100)environment isolates showed distinct restriction pattern likethat of E coli BLAST genome search across 931 bacterialfamilies verified the specificity of primers and restrictionpreference for Bfa I The primers showed exclusive bindingonly with Enterobacteriaceae family (Supplementary DataA8) Among the various strains in the family only Shigellaspp Salmonella enterica (Arizona) and Proteus mirabilusexhibited similar restriction preference to that of E coli Inspite of Shigella spp being taxa in disguise of E coli [37] theycan be easily screened using selective medium because of nolactose fermentation [38] whereas Salmonella enterica andProteus mirabilis showed amplicon heterogeneity of 1073 bpand 880 bp during amplification thus making it easy to avoiderroneous conclusions and misinterpretation
33 Categorizing E coli StrainsUsingARDRAof 23S P2682 withHae III A detailed analysis of the Hae III profile across 399sequences [Supplementary Data A9] of ribosomal segmentsof E coli indicated four possible profiles types I II IIIand IV with varying degree of predominance within thesesegments The rest of the candidate strains showed type IIIpatterns but types I and IV were exclusive for E coli strainsMajority of genomes (119899 = 32) as in 15 nonpathogenic and17 pathogenic strains showed type I profiles (57) followedby type III (35) predominating in 15 strains Six strainsshowed type IV pattern (10) of which four were B strainsType I groups represent E coli strains that lacked recognition
6 Journal of Pathogens
1 7
880 bp
65432
(a)
8
665bp
475bp
215 bp 214 bp
178bp
1 765432
(b)
Figure 2 (a) Broad Range PCR of 23S P1880
with genomic DNA isolated from standard strains Lane 1 100 bp ladder Lane 2 blank Lane 3E coli Lane 4 C freundii Lane 5 C koseri Lane 6 K pneumonia Lane 7 E aerogenes PCR product of single representative of E coli wasloaded (b) ARDRA of 23S P1
880
amplicons with Bfa I Lanes 1 2 and 3 E coli (ATCC15223 ATCC25922 and ATCC11775) Lane 4C freundii(ATCC8090) Lane 5 C koseri (ATCC27028) Lane 6 K pneumonia (ATCC33495) Lane 7 E aerogenes (ATCC13048) Lane 8 100 bp ladder
elements due to change at 2205th and 2161st base with respectto 23S rDNA or with reference to amplicons at 207th or 249thbase positions Type IV strains are contrary to type I withintact recognition sequence at both of these sites Strains withtype II profiles lacked 2161st (249th) cut site and type IIIlacked the 2205th (207th) cut positions respectively Type IIwas the rarest with its patterns observed to be ambiguous incertain rrn segments
Heterogenicity in few genomes was noted two rrn amongseven repeats ofW (NC 0176851 each in regular and comple-mentary strand) and KO11FL (NC 0176601 each in regularand complementary strand) and one in KO11FL (NC 0169021 regular) showed type II profile in spite of type I beingtheir primary pattern One among seven of the B strains REL606 (NC 0129671 regular) BL21-Gold (NC 0129471 com-plementary) BL21DE3 (NC 0129712 regular) and BL21DE3(NC 0128922 regular) showed type I profiles apart frombeing prevalent with type IV pattern Similarly two rrnamongNRG 873C (NC 0176341 each in regular and comple-mentary strand) and LF82 (NC 0119931 each in regular andcomplementary strand) and one in CFT073 (NC 0044311regular) showed type I pattern while exhibiting type IIIpattern predominance APEC01 (NC 0085631) being primein type III showed one rrn segment of type I and type IV(both in regular strand) E coli strain 042 (NC 0176261)displayed preponderance in type I pattern but three rrn (tworegular and one complementary) copies exhibited type IIIpattern One of the regular rrn elements in LF82 showed typeIV pattern The fragmentation pattern obtained by ARDRAof 23S P2
682with Hae III in the standard samples showed
that all K-12 C W Crooks and their respective descendantsexhibited type I profileswith type IVprofiles being exclusivelypresented by B strains (Figures 3(a) and 3(b))
Among the isolates 21 out of 22 commensal and 16 of18 clinical samples showed type I pattern heterogenicity ofpatterns was not observed These results reaffirmed predom-inance (93) of type I in nature Multisequence typing hasshown commensal strains to be categorized primarily undergroup A or group B1 whereas groups B2 D and E comprisegenerally pathogenic strains K-12 B and Crooks strain comeunder phylogroup A whereas strain W is a B1 phylogroup
34 Local Alignment of 23S rrn Hae III Profiles versus GlobalAlignment to Establish Clonality in E coli A number ofgenes that are of core dispensable and unique in naturemake up for pangenome the latter two are gained due torecombinational or transpositional events To obtain similarclustering on comparing whole genome would emphasizethe significance of these restriction profiles and help inestablishing the clonal nature and predominance of a singleclone Hence dendogram constructs with 682 bp ampliconfrom a representative of each restriction profile will showdistinct clustering of these profiles due to the fact that a smallsubset of segments within the whole genome was consideredThe global alignment of whole genome sequences (Figure 4)of the entire 57 E coli strains displayed distinct concentrateof type I III and IV profiles An intrinsic analysis of thespot of divergence and respective clade formations showedinteresting and riveting findings The strains at the ancestralroot showed type I pattern and the next divergent node point(D1) displayed formation of two distinct clans of type I andtype IV profiles segregated at the maximum distance fromone another Numbering of nodes was done by consideringthe precedence of branch point at every node A possiblescenario of two additional mutational events that are oftransitional nature (A to G) at 2205th and 2161st bases has
Journal of Pathogens 7
682bp
987654321
9998400
8998400
7998400
6998400
5998400
4998400
3998400
2998400
1998400
(a)
431bp
223 bp
210 bp
179bp
223 + 210 bp
223+
210+
87654321
8998400
7998400
6998400
5998400
4998400
3998400
2998400
1998400
(b)
Figure 3 (a) Broad range PCR of 23S P2682
with genomic DNA isolated from standard strains of E coli The upper row comprises Lane 1K-12 strains Lane 2 derivatives of K-12 strains Lane 3 DHI strain Lane 4 strain C1 Lane 6 Crookrsquos strain Lane 7 Walkerrsquos strain Lane 8descendent of W Lane 9 blank The lower row comprises Lane 11015840 B strain Lane 21015840 BL21 strain Lane 31015840 BL21DE3 Lane 41015840 BL21DE3Lys5(B3) Lane 61015840 E coli (ATCC 15223 commensal isolate) Lane 71015840 E coli (ATCC 11775 urine sample) Lane 81015840 E coli (ATCC 25922 clinical)Lanes 551015840 and Lanes 991015840 in both rows are 100 bp ladder and blank respectively (b) ARDRA of 23S P2
682
amplicons with Hae III Lane 1 ofboth rows comprises 100 bp ladder The digested products are loaded as per the order given in Figure 2(a)
introduced Hae III recognition patterns D6 gave lineages ofB1 phylogroups D9 giving rise to B2 clades (D12) of type Iprofiles and D clades with varied Hae III patterns (D11) D10gave rise to two nodes (with strains being type I groups)with one showing B1 phylogroups (D13) and the other beingpredominant in E phylogroups followed by B1 An almostexemplary corelation between the clonal groups and differentpathovars was found with this study Majority of commensalstrains and EIEC and all EAEC and EPEC were shown tobe having type I profile (Table 4) All UPEC ExPEC APECEIEC and single strain of EPEC and two commensal strainsshowed type III pattern B form strains along with a singlestrain of UPEC and environmental isolate exhibited type IVprofiles
Similarly a near-pristine association between phylo-groups and Hae III profiles was established Strains comingunder phylogroups A were preponderant in type I whereastype IV profiles were exclusively for B forms Type I profileswere observed in the strains (119899) in the order of A gt B1 gt E gtDwith type III dominant in B2 strains Phylogroup D showedequal occurrence of type III and type IV fragmentationpattern followed by a single strain with type I profile Theseresults assisted in enumerating a conjecture which works upa scenario of primordial microbial population where E colistrains had two clonal types (Figure 5(a)) which coexistedtogether (type I and type IV clones) Of these type I strainswould have been the predominant clone in comparison totype IV based on the dendogram data with maximum genepool of root strains showing type I Type III may have beenthe next predominant clone (an intermediary one) that hasevolved either from type I (by addition of a new recognition
Table 4 Corelation between phylogroups Hae III profiles andnature of E coli strains (genome data analysis of 57 E coli strainscollated from Genbank)
Strain Type A B1 B2 D E Total Out ofCommensal I 10 5 mdash mdash mdash 15 21Commensal III mdash mdash 2 mdash mdash 2 21Commensal IV 4 mdash mdash mdash mdash 4 21EPEC I mdash mdash mdash mdash 7 7 8EPEC III mdash mdash 1 mdash mdash 1 8EAEC I mdash 4 mdash 1 mdash 5 5EHEC I mdash 2 mdash mdash mdash 2 3EHEC II mdash 1 mdash mdash mdash 1 3ETEC I 1 1 1 mdash mdash 3 3UPEC III mdash mdash 7 mdash mdash 7 8UPEC IV mdash mdash mdash 1 mdash 1 8EIEC III mdash mdash 3 mdash mdash 3 3ExPEC III mdash mdash 2 2 mdash 4 4APEC III mdash mdash 1 mdash 1 1ENV IV mdash mdash mdash 1 1 1APEC avian pathogenic EAEC enteroaggressive EHEC enterohemorrhag-ic ETEC enterotoxigenic EIEC enteroinvasive EPEC enteropathogenicENV environmental ExPEC extraintestinal pathogenic UPEC uropatho-genic
site) or from type IV (by deletion of one recognition site)strains In a different scenario maybe all three types coexistedor type III would have given rise to type IVwith an additionalmutational event This situation may be hardly convincing
8 Journal of Pathogens
D1
D2
D3
D4
D5
D6
D7
D8
D9
D10
D11
D12
D13
D14
BL21(DE3)_1lowastBL21(DE3)_2lowast
BL21-Gold(DE3)pLysS AGlowast
B str REL606lowastA phylogroup
ETEC H10407lowast (B2)
UMNK88 (A)
(B1 clades) Str 2009EL-2050Str 2009EL-2071
Str 2011-349355989
B1 phylogroup
Str EC4115Str TW14359
Str EDL933Str Sakai
Xuzhou21Str CB9615
Str RM12598
E phylogroup
Str 11128Str 11368 B1 phylogroup
NA114
SE15LF82lowastlowastlowastStr NRG 857Clowastlowastlowast
UM146
UTI89IHE3034
APEC O1lowastlowastS88
Clone D i2Clone D i14
ABU 83972
CFT073536
ED1aStr E234869
B2 phylogroup
UMN026
IAI39Str CE10
SMS-3-5
D phylogroup
O42lowastlowastlowast
(B2 clades)
(D clades)
(B1 clades)
Wlowastlowastlowast
WKO11FLlowastlowast
KO11FLE24377A
SE11
B1 phylogroup
IAI1ATCC 8739
P12blowastHS
BW2952
K-12 substr DH10BDH1
DH1 (ME8569)K-12 substr W3110
K-12 substr MG1655
A phylogroup
1
2
10
3156
Str 12009lowastlowast
Commensal ETEC EHEC EPEC UPEC
ExPECAPECEAECEIECENV
Figure 4 Analysis of distribution of Hae III profiles phylogroups and pathovars within the global alignment dendogram constructcomprising 57 E coli genomes The dendogram of the whole genome sequence of 57 genomes was obtained from the NCBI web site forE coli and was redrawn using Photoshop CS6 for colour coordination for pathogenic and nonpathogenic strains and inputting the clonalgroups and phylogroups (httpwwwncbinlmnihgovgenometerm=escherichia+coli) Among the 57 genomes 32 were of type I (56)18 were of type III (31) and 6 and 1 were of type IV (10) and type II (2) respectively showing that type I is predominant and type IIthe rarest To correlate between the phylogroupsHae III profiles and pathovars the lineages were considered as clades derived from variousdivergence point (D) during the course of evolution
as dendogram segregation shows type III strains evolving orpresent after type IV strains The clustering also shows thatpredominance of type III is more than type IV and maybeevolutionary processesmutation have triggered the increasein type III profiles On similar line type II clones would havebeen evolved from type I (addition) or type IV (deletion)but due to the very limited type II restriction profile patterns(rare) as observed from the gene pool data the chances orapproximation of such event is difficult to prove Based onthese results no new insights into phylogroup distributionacross candidate strains can be provided however withrespect to the Hae III profile an additional angle can beconsidered Phylogroups A and B1 are predominant withtype I Hae III profiles whereas B2 phylogroups exclusivelyrepresented type III profile (Figure 5(b))
An intrinsic metagenomic and pangenomic analysis ofmost bacterial species exhibited mosaicisms of their inherentblueprint [39] In such situations core gene elements sharedacross the species are limited in comparison to dispensable(flexible) genetic contents that concede uniqueness to aparticular strain These flexible contents help in niche adapt-ability and give bacteria the ability to infect and invade Amajor concern is while diagnosing both true and opportunistpathogenic bacteria the former causing disease in virtuallyany susceptible host and the latter invoking diseases inimmunocompromised individuals ldquoMolecular barcodingrdquoan evolved term which implies the method of allele specificamplification of relatively small DNA sequences or barcodes[40] for spotting a particular species is a thrust area formicrobial diagnostics A plethora of functional genes ranging
Journal of Pathogens 9
Type I(28 459)
Type IV(28 207 249 and 459)
Hypothesis
Type III(28 249 and 459)
+ minus
+
Primordial
NonpathogenicA gt B1 gt B2
type I gt IV gt III
PathogenicB2 gt B1 gt E gt D
type I (IAT)III(P)gt type IVII
Predominant clones
Rarest
+minus
Type II(28 207 and 459)
Restriction profilesand
clonesPhylogroups and
clones
2∘ clonesintermediate
(a) (b)
Figure 5 (a) Hypothesis of clonality in E coli strains based on 23S rrn restriction profiling The primordial soup of E coli microbes wouldhave been comprised of two clonal groups types I and IV coexisting with each other but predominated by type I strains Over the courseof evolution on accumulating mutation as a means for variation type I or type IV would have given rise to type III by addition (former) ordeletion (latter) of recognition sequence The second scenario could be a rare event because accumulation of two mutations in a conservedregion would be difficult Type II being the rarest would have been evolved in similar basis to type III (b) Hypothesis of linking phylogroupsHae III profiles and clonal types in E coli This diagram illustrates the clonal groups of commensal E coli strains having both A phylogroups(dominant) and B1 phylogroups each being type I profiles Pathogenic counterparts showed dominancy in type III restriction pattern followedby type I pattern With respect to phylogroups pathovars were predominant in B2 and B1 followed by D and E groups
from housekeeping to repair are employed for characterizingbacteria of which rRNA is referred to as the gold stretch[41 42] Ribosomal segments organized as operon clusters of16S 23S and 5S rDNA and these assemblages are distributedfrom single to multiple copies across the bacterial genophore[43] The occurrence of redundancy in maintaining differentcopies of these genes is not associated with virulence but anin-built ecological strategy that contributes to the particularbacterium with a competitive edge and better chance ofsurvival [44] A futuristic approach of coupling the techniqueand results with antibiotic resistance pattern colony char-acteristics and genomic profiles of bacterial strains of bothcommensal and pathogenic nature can help in monitoringand managing infections in a better manner
4 Conclusion
The causatum of postgenomic era genomics and proteomicshas enhanced this progress by improvising the techniquesused for diagnostics The strategy of ARDRA utilized inthis study was rapid and a cost-effective approach to dif-ferentiate E coli from its colony morphovars along withgiving a predicament of nature of origin or clonal groupsThe mutations reported in this work for detection andestablishing clonality namely T874A (Bfa I) G2161A andG2205A (Hae III) have not been mentioned in any researchpaper or within the Ribosomal RNA Mutation Database
[45] making this finding aboriginal This basic methodologycan be adapted for detection of other bacterial organismsby targeting SNPs linked to recognition sequences therebyvalidating rrn sequences as marker for diagnostics A cumu-lative approach of detection of unique restriction pattern of atargeted gene and its cross-reference across bacterial familiescan circumvent changes of false positives
Disclosure
This work was presented (poster) in part at the 5th Interna-tional Barcoding of Life Kunming China 2013
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
Acknowledgments
The authors acknowledge and express their gratitude to Pro-fessor D A Bhiwgade (Ex-Director) and Professor DebjaniDasgupta Director School of Biotechnology amp Bioinformat-ics D Y Patil University as well as the management for theirsupport and for providing the infrastructure to carry out thecurrent work
10 Journal of Pathogens
References
[1] M Richter and R Rossello-Mora ldquoShifting the genomic goldstandard for the prokaryotic species definitionrdquo Proceedings ofthe National Academy of Sciences of the United States of Americavol 106 no 45 pp 19126ndash19131 2009
[2] A F Auch M von Jan H-P Klenk and M Goker ldquoDigitalDNA-DNA hybridization for microbial species delineation bymeans of genome-to-genome sequence comparisonrdquo Standardsin Genomic Sciences vol 2 no 1 pp 117ndash134 2010
[3] D H Martin M Nsuami J Schachter et al ldquoUse of multiplenucleic acid amplification tests to define the infected-patientlsquogold standardrsquo in clinical trials of new diagnostic tests forChlamydia trachomatis infectionsrdquo Journal of Clinical Microbi-ology vol 42 no 10 pp 4749ndash4758 2004
[4] E A Mothershed and A M Whitney ldquoNucleic acid-basedmethods for the detection of bacterial pathogens presentand future considerations for the clinical laboratoryrdquo ClinicaChimica Acta vol 363 no 1-2 pp 206ndash220 2006
[5] J E Clarridge III ldquoImpact of 16S rRNA gene sequence analysisfor identification of bacteria on clinical microbiology andinfectious diseasesrdquo Clinical Microbiology Reviews vol 17 no4 pp 840ndash862 2004
[6] O Mizrahi-Man E R Davenport and Y Gilad ldquoTaxonomicclassification of bacterial 16S rRNA genes using short sequenc-ing reads evaluation of effective study designsrdquo PLoS ONE vol8 no 1 Article ID e53608 2013
[7] D E Lebonah A Dileep K Chandrasekhar S Sreevani BSreedevi and J Pramoda Kumari ldquoDNA barcoding on bacteriaa reviewrdquo Advances in Biology vol 2014 Article ID 541787 9pages 2014
[8] J Rajendhran and P Gunasekaran ldquoMicrobial phylogeny anddiversity small subunit ribosomal RNA sequence analysis andbeyondrdquo Microbiological Research vol 166 no 2 pp 99ndash1102011
[9] R Srinivasan U Karaoz M Volegova et al ldquoUse of 16S rRNAgene for identification of a broad range of clinically relevantbacterial pathogensrdquo PLoS ONE vol 10 no 2 Article IDe0117617 2015
[10] S Chakravorty D Helb M Burday N Connell and D AllandldquoA detailed analysis of 16S ribosomal RNA gene segments forthe diagnosis of pathogenic bacteriardquo Journal of MicrobiologicalMethods vol 69 no 2 pp 330ndash339 2007
[11] C R Woese ldquoReview bacterial evolutionrdquo MicrobiologicalReviews vol 51 pp 221ndash271 1987
[12] M Touchon C Hoede O Tenaillon et al ldquoOrganised genomedynamics in the Escherichia coli species results in highly diverseadaptive pathsrdquo PLoS Genetics vol 5 no 1 Article ID e10003442009
[13] J M Janda and S L Abbott ldquo16S rRNA gene sequencingfor bacterial identification in the diagnostic laboratory plusesperils and pitfallsrdquo Journal of Clinical Microbiology vol 45 no9 pp 2761ndash2764 2007
[14] J-J Lu C-L Perng S-Y Lee and C-CWan ldquoUse of PCRwithuniversal primers and restriction endonuclease digestions fordetection and identification of common bacterial pathogens incerebrospinal fluidrdquo Journal of Clinical Microbiology vol 38 no6 pp 2076ndash2080 2000
[15] N M Guentzel ldquoEscherichia Klebsiella Enterobacter SerratiaCitrobacter andProteusrdquo inMedicalMicrobiology S Baron Edchapter 26 University of Texas Medical Branch at GalvestonGalveston Tex USA 1996
[16] P Behzadip E Behzadi H Yazdanbod R Aghapour A MCheshmeh and S D Omran ldquoA survey on urinary tract infec-tions associated with the three most common uropathogenicbacteriardquoMaedica (Buchar) vol 5 no 2 pp 111ndash115 2010
[17] B CMetri P Jyothi andBV Peerapur ldquoAntibiotic resistance inCitrobacter spp isolated from urinary tract infectionrdquo UrologyAnnals vol 5 no 4 pp 312ndash313 2013
[18] N Taneja S S Chatterjee M Singh S Singh and M SharmaldquoPediatric urinary tract infections in a tertiary care center fromnorth Indiardquo Indian Journal of Medical Research vol 131 no 1pp 101ndash105 2010
[19] J B Kaper J P Nataro and H L T Mobley ldquoPathogenicEscherichia colirdquo Nature Reviews Microbiology vol 2 no 2 pp123ndash140 2004
[20] E J de Muinck K Lagesen J E Afset et al ldquoComparisonsof infant Escherichia coli isolates link genomic profiles withadaptation to the ecological nicherdquo BMC Genomics vol 14 no1 article 81 2013
[21] D A Rasko M J Rosovitz G S A Myers et al ldquoThepangenome structure of Escherichia coli comparative genomicanalysis of E coli commensal and pathogenic isolatesrdquo Journalof Bacteriology vol 190 no 20 pp 6881ndash6893 2008
[22] C T Archer J F Kim H Jeong et al ldquoThe genome sequenceof E coli W (ATCC 9637) comparative genome analysis andan improved genome-scale reconstruction of E colirdquo BMCGenomics vol 12 article no 9 2011
[23] PKuhnert J Nicolet and J Frey ldquoRapid and accurate identifica-tion of Escherichia coliK-12 strainsrdquoApplied and EnvironmentalMicrobiology vol 61 no 11 pp 4135ndash4139 1995
[24] A P Bauer S M Dieckmann W Ludwig and K-H SchleiferldquoRapid identification of Escherichia coli safety and laboratorystrain lineages based on multiplex-PCRrdquo FEMS MicrobiologyLetters vol 269 no 1 pp 36ndash40 2007
[25] B J Bachmann ldquoPedigrees of some mutant strains ofEscherichia coli K-12rdquo Bacteriological Reviews vol 36 no 4 pp525ndash557 1972
[26] P Daegelen F W Studier R E Lenski S Cure and J F KimldquoTracing ancestors and relatives of Escherichia coli B and thederivation of B strains REL606 and BL21(DE3)rdquo Journal ofMolecular Biology vol 394 no 4 pp 634ndash643 2009
[27] F W Studier P Daegelen R E Lenski S Maslov and J F KimldquoUnderstanding the differences between genome sequences ofEscherichia coli B strains REL606 and BL21(DE3) and compar-ison of the E coli B and K-12 genomesrdquo Journal of MolecularBiology vol 394 no 4 pp 653ndash680 2009
[28] R K Selander and B R Levin ldquoGenetic diversity and structurein Escherichia coli populationsrdquo Science vol 210 no 4469 pp545ndash547 1980
[29] D M Gordon O Clermont H Tolley and E DenamurldquoAssigning Escherichia coli strains to phylogenetic groupsmulti-locus sequence typing versus the PCR triplex methodrdquoEnvironmental Microbiology vol 10 no 10 pp 2484ndash24962008
[30] P Desjardins B Picard B Kaltenbock J Elion and EDenamur ldquoSex in Escherichia coli does not disrupt the clonalstructure of the population evidence from random amplifiedpolymorphic DNA and restriction-fragment-length polymor-phismrdquo Journal of Molecular Evolution vol 41 no 4 pp 440ndash448 1995
Journal of Pathogens 11
[31] N D Jasuja R Saxena S Chandra and S C Joshi ldquoIsolationand identification of microorganism from polyhouse agricul-ture soil of RajasthanrdquoAfrican Journal ofMicrobiology Researchvol 7 no 41 pp 4886ndash4891 2013
[32] V Hemraj S Diksha and G Avneet ldquoA review of commonlyused biochemical test for bacteriardquo Innovare Journal of LifeSciences vol 1 pp 1ndash7 2013
[33] J D Thompson D G Higgins and T J Gibson ldquoCLUSTALWimproving the sensitivity of progressive multiple sequencealignment through sequence weighting position-specific gappenalties and weight matrix choicerdquoNucleic Acids Research vol22 no 22 pp 4673ndash4680 1994
[34] T Vincze J Posfai and R J Roberts ldquoNEBcutter a program tocleave DNA with restriction enzymesrdquo Nucleic Acids Researchvol 31 no 13 pp 3688ndash3691 2003
[35] J Ye G Coulouris I Zaretskaya I Cutcutache S Rozen andT Madden ldquoPrimer-BLAST a tool to design target-specificprimers for polymerase chain reactionrdquo BMC Bioinformaticsvol 13 article 134 2012
[36] J R Parvathi and S Singh ldquoFacile approaches for microbialDNA extraction comparative study with colonies lsquopickedrsquo fromthree different bacteriological mediumrdquo Journal of Pure andApplied Microbiology vol 9 no 3 pp 2373ndash2378 2015
[37] R Lan and P R Reeves ldquoEscherichia coli in disguise molecularorigins of Shigellardquo Microbes and Infection vol 4 no 11 pp1125ndash1132 2002
[38] D J Leininger J R Roberson and F Elvinger ldquoUse ofeosin methylene blue agar to differentiate Escherichia coli fromother gram-negative mastitis pathogensrdquo Journal of VeterinaryDiagnostic Investigation vol 13 no 3 pp 273ndash275 2001
[39] U Dobrindt F Agerer K Michaelis et al ldquoAnalysis of genomeplasticity in pathogenic and commensal Escherichia coli isolatesby use of DNA arraysrdquo Journal of Bacteriology vol 185 no 6pp 1831ndash1840 2003
[40] V Savolainen R S Cowan A P Vogler G K Roderickand R Lane ldquoTowards writing the encyclopaedia of life anintroduction to DNA barcodingrdquo Philosophical Transactions ofthe Royal Society B Biological Sciences vol 360 no 1462 pp1805ndash1811 2005
[41] T A Isenbarger C E Carr S S Johnson et al ldquoThe mostconserved genome segments for life detection on earth andother planetsrdquo Origins of Life and Evolution of Biospheres vol38 no 6 pp 517ndash533 2008
[42] S R Santos and H Ochman ldquoIdentification and phylogeneticsorting of bacterial lineages with universally conserved genesand proteinsrdquoEnvironmentalMicrobiology vol 6 no 7 pp 754ndash759 2004
[43] R Rastogi M Wu I DasGupta and G E Fox ldquoVisualizationof ribosomal RNA operon copy number distributionrdquo BMCMicrobiology vol 9 no 1 article 208 2009
[44] J A Klappenbach J M Dunbar and T M Schmidt ldquorRNAoperon copy number reflects ecological strategies of bacteriardquoApplied and Environmental Microbiology vol 66 no 4 pp1328ndash1333 2000
[45] J A Klappenbach P R Saxman J R Cole and T M SchmidtldquoRrndb the ribosomal RNA operon copy number databaserdquoNucleic Acids Research vol 29 no 1 pp 181ndash184 2001
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom

6 Journal of Pathogens
1 7
880 bp
65432
(a)
8
665bp
475bp
215 bp 214 bp
178bp
1 765432
(b)
Figure 2 (a) Broad Range PCR of 23S P1880
with genomic DNA isolated from standard strains Lane 1 100 bp ladder Lane 2 blank Lane 3E coli Lane 4 C freundii Lane 5 C koseri Lane 6 K pneumonia Lane 7 E aerogenes PCR product of single representative of E coli wasloaded (b) ARDRA of 23S P1
880
amplicons with Bfa I Lanes 1 2 and 3 E coli (ATCC15223 ATCC25922 and ATCC11775) Lane 4C freundii(ATCC8090) Lane 5 C koseri (ATCC27028) Lane 6 K pneumonia (ATCC33495) Lane 7 E aerogenes (ATCC13048) Lane 8 100 bp ladder
elements due to change at 2205th and 2161st base with respectto 23S rDNA or with reference to amplicons at 207th or 249thbase positions Type IV strains are contrary to type I withintact recognition sequence at both of these sites Strains withtype II profiles lacked 2161st (249th) cut site and type IIIlacked the 2205th (207th) cut positions respectively Type IIwas the rarest with its patterns observed to be ambiguous incertain rrn segments
Heterogenicity in few genomes was noted two rrn amongseven repeats ofW (NC 0176851 each in regular and comple-mentary strand) and KO11FL (NC 0176601 each in regularand complementary strand) and one in KO11FL (NC 0169021 regular) showed type II profile in spite of type I beingtheir primary pattern One among seven of the B strains REL606 (NC 0129671 regular) BL21-Gold (NC 0129471 com-plementary) BL21DE3 (NC 0129712 regular) and BL21DE3(NC 0128922 regular) showed type I profiles apart frombeing prevalent with type IV pattern Similarly two rrnamongNRG 873C (NC 0176341 each in regular and comple-mentary strand) and LF82 (NC 0119931 each in regular andcomplementary strand) and one in CFT073 (NC 0044311regular) showed type I pattern while exhibiting type IIIpattern predominance APEC01 (NC 0085631) being primein type III showed one rrn segment of type I and type IV(both in regular strand) E coli strain 042 (NC 0176261)displayed preponderance in type I pattern but three rrn (tworegular and one complementary) copies exhibited type IIIpattern One of the regular rrn elements in LF82 showed typeIV pattern The fragmentation pattern obtained by ARDRAof 23S P2
682with Hae III in the standard samples showed
that all K-12 C W Crooks and their respective descendantsexhibited type I profileswith type IVprofiles being exclusivelypresented by B strains (Figures 3(a) and 3(b))
Among the isolates 21 out of 22 commensal and 16 of18 clinical samples showed type I pattern heterogenicity ofpatterns was not observed These results reaffirmed predom-inance (93) of type I in nature Multisequence typing hasshown commensal strains to be categorized primarily undergroup A or group B1 whereas groups B2 D and E comprisegenerally pathogenic strains K-12 B and Crooks strain comeunder phylogroup A whereas strain W is a B1 phylogroup
34 Local Alignment of 23S rrn Hae III Profiles versus GlobalAlignment to Establish Clonality in E coli A number ofgenes that are of core dispensable and unique in naturemake up for pangenome the latter two are gained due torecombinational or transpositional events To obtain similarclustering on comparing whole genome would emphasizethe significance of these restriction profiles and help inestablishing the clonal nature and predominance of a singleclone Hence dendogram constructs with 682 bp ampliconfrom a representative of each restriction profile will showdistinct clustering of these profiles due to the fact that a smallsubset of segments within the whole genome was consideredThe global alignment of whole genome sequences (Figure 4)of the entire 57 E coli strains displayed distinct concentrateof type I III and IV profiles An intrinsic analysis of thespot of divergence and respective clade formations showedinteresting and riveting findings The strains at the ancestralroot showed type I pattern and the next divergent node point(D1) displayed formation of two distinct clans of type I andtype IV profiles segregated at the maximum distance fromone another Numbering of nodes was done by consideringthe precedence of branch point at every node A possiblescenario of two additional mutational events that are oftransitional nature (A to G) at 2205th and 2161st bases has
Journal of Pathogens 7
682bp
987654321
9998400
8998400
7998400
6998400
5998400
4998400
3998400
2998400
1998400
(a)
431bp
223 bp
210 bp
179bp
223 + 210 bp
223+
210+
87654321
8998400
7998400
6998400
5998400
4998400
3998400
2998400
1998400
(b)
Figure 3 (a) Broad range PCR of 23S P2682
with genomic DNA isolated from standard strains of E coli The upper row comprises Lane 1K-12 strains Lane 2 derivatives of K-12 strains Lane 3 DHI strain Lane 4 strain C1 Lane 6 Crookrsquos strain Lane 7 Walkerrsquos strain Lane 8descendent of W Lane 9 blank The lower row comprises Lane 11015840 B strain Lane 21015840 BL21 strain Lane 31015840 BL21DE3 Lane 41015840 BL21DE3Lys5(B3) Lane 61015840 E coli (ATCC 15223 commensal isolate) Lane 71015840 E coli (ATCC 11775 urine sample) Lane 81015840 E coli (ATCC 25922 clinical)Lanes 551015840 and Lanes 991015840 in both rows are 100 bp ladder and blank respectively (b) ARDRA of 23S P2
682
amplicons with Hae III Lane 1 ofboth rows comprises 100 bp ladder The digested products are loaded as per the order given in Figure 2(a)
introduced Hae III recognition patterns D6 gave lineages ofB1 phylogroups D9 giving rise to B2 clades (D12) of type Iprofiles and D clades with varied Hae III patterns (D11) D10gave rise to two nodes (with strains being type I groups)with one showing B1 phylogroups (D13) and the other beingpredominant in E phylogroups followed by B1 An almostexemplary corelation between the clonal groups and differentpathovars was found with this study Majority of commensalstrains and EIEC and all EAEC and EPEC were shown tobe having type I profile (Table 4) All UPEC ExPEC APECEIEC and single strain of EPEC and two commensal strainsshowed type III pattern B form strains along with a singlestrain of UPEC and environmental isolate exhibited type IVprofiles
Similarly a near-pristine association between phylo-groups and Hae III profiles was established Strains comingunder phylogroups A were preponderant in type I whereastype IV profiles were exclusively for B forms Type I profileswere observed in the strains (119899) in the order of A gt B1 gt E gtDwith type III dominant in B2 strains Phylogroup D showedequal occurrence of type III and type IV fragmentationpattern followed by a single strain with type I profile Theseresults assisted in enumerating a conjecture which works upa scenario of primordial microbial population where E colistrains had two clonal types (Figure 5(a)) which coexistedtogether (type I and type IV clones) Of these type I strainswould have been the predominant clone in comparison totype IV based on the dendogram data with maximum genepool of root strains showing type I Type III may have beenthe next predominant clone (an intermediary one) that hasevolved either from type I (by addition of a new recognition
Table 4 Corelation between phylogroups Hae III profiles andnature of E coli strains (genome data analysis of 57 E coli strainscollated from Genbank)
Strain Type A B1 B2 D E Total Out ofCommensal I 10 5 mdash mdash mdash 15 21Commensal III mdash mdash 2 mdash mdash 2 21Commensal IV 4 mdash mdash mdash mdash 4 21EPEC I mdash mdash mdash mdash 7 7 8EPEC III mdash mdash 1 mdash mdash 1 8EAEC I mdash 4 mdash 1 mdash 5 5EHEC I mdash 2 mdash mdash mdash 2 3EHEC II mdash 1 mdash mdash mdash 1 3ETEC I 1 1 1 mdash mdash 3 3UPEC III mdash mdash 7 mdash mdash 7 8UPEC IV mdash mdash mdash 1 mdash 1 8EIEC III mdash mdash 3 mdash mdash 3 3ExPEC III mdash mdash 2 2 mdash 4 4APEC III mdash mdash 1 mdash 1 1ENV IV mdash mdash mdash 1 1 1APEC avian pathogenic EAEC enteroaggressive EHEC enterohemorrhag-ic ETEC enterotoxigenic EIEC enteroinvasive EPEC enteropathogenicENV environmental ExPEC extraintestinal pathogenic UPEC uropatho-genic
site) or from type IV (by deletion of one recognition site)strains In a different scenario maybe all three types coexistedor type III would have given rise to type IVwith an additionalmutational event This situation may be hardly convincing
8 Journal of Pathogens
D1
D2
D3
D4
D5
D6
D7
D8
D9
D10
D11
D12
D13
D14
BL21(DE3)_1lowastBL21(DE3)_2lowast
BL21-Gold(DE3)pLysS AGlowast
B str REL606lowastA phylogroup
ETEC H10407lowast (B2)
UMNK88 (A)
(B1 clades) Str 2009EL-2050Str 2009EL-2071
Str 2011-349355989
B1 phylogroup
Str EC4115Str TW14359
Str EDL933Str Sakai
Xuzhou21Str CB9615
Str RM12598
E phylogroup
Str 11128Str 11368 B1 phylogroup
NA114
SE15LF82lowastlowastlowastStr NRG 857Clowastlowastlowast
UM146
UTI89IHE3034
APEC O1lowastlowastS88
Clone D i2Clone D i14
ABU 83972
CFT073536
ED1aStr E234869
B2 phylogroup
UMN026
IAI39Str CE10
SMS-3-5
D phylogroup
O42lowastlowastlowast
(B2 clades)
(D clades)
(B1 clades)
Wlowastlowastlowast
WKO11FLlowastlowast
KO11FLE24377A
SE11
B1 phylogroup
IAI1ATCC 8739
P12blowastHS
BW2952
K-12 substr DH10BDH1
DH1 (ME8569)K-12 substr W3110
K-12 substr MG1655
A phylogroup
1
2
10
3156
Str 12009lowastlowast
Commensal ETEC EHEC EPEC UPEC
ExPECAPECEAECEIECENV
Figure 4 Analysis of distribution of Hae III profiles phylogroups and pathovars within the global alignment dendogram constructcomprising 57 E coli genomes The dendogram of the whole genome sequence of 57 genomes was obtained from the NCBI web site forE coli and was redrawn using Photoshop CS6 for colour coordination for pathogenic and nonpathogenic strains and inputting the clonalgroups and phylogroups (httpwwwncbinlmnihgovgenometerm=escherichia+coli) Among the 57 genomes 32 were of type I (56)18 were of type III (31) and 6 and 1 were of type IV (10) and type II (2) respectively showing that type I is predominant and type IIthe rarest To correlate between the phylogroupsHae III profiles and pathovars the lineages were considered as clades derived from variousdivergence point (D) during the course of evolution
as dendogram segregation shows type III strains evolving orpresent after type IV strains The clustering also shows thatpredominance of type III is more than type IV and maybeevolutionary processesmutation have triggered the increasein type III profiles On similar line type II clones would havebeen evolved from type I (addition) or type IV (deletion)but due to the very limited type II restriction profile patterns(rare) as observed from the gene pool data the chances orapproximation of such event is difficult to prove Based onthese results no new insights into phylogroup distributionacross candidate strains can be provided however withrespect to the Hae III profile an additional angle can beconsidered Phylogroups A and B1 are predominant withtype I Hae III profiles whereas B2 phylogroups exclusivelyrepresented type III profile (Figure 5(b))
An intrinsic metagenomic and pangenomic analysis ofmost bacterial species exhibited mosaicisms of their inherentblueprint [39] In such situations core gene elements sharedacross the species are limited in comparison to dispensable(flexible) genetic contents that concede uniqueness to aparticular strain These flexible contents help in niche adapt-ability and give bacteria the ability to infect and invade Amajor concern is while diagnosing both true and opportunistpathogenic bacteria the former causing disease in virtuallyany susceptible host and the latter invoking diseases inimmunocompromised individuals ldquoMolecular barcodingrdquoan evolved term which implies the method of allele specificamplification of relatively small DNA sequences or barcodes[40] for spotting a particular species is a thrust area formicrobial diagnostics A plethora of functional genes ranging
Journal of Pathogens 9
Type I(28 459)
Type IV(28 207 249 and 459)
Hypothesis
Type III(28 249 and 459)
+ minus
+
Primordial
NonpathogenicA gt B1 gt B2
type I gt IV gt III
PathogenicB2 gt B1 gt E gt D
type I (IAT)III(P)gt type IVII
Predominant clones
Rarest
+minus
Type II(28 207 and 459)
Restriction profilesand
clonesPhylogroups and
clones
2∘ clonesintermediate
(a) (b)
Figure 5 (a) Hypothesis of clonality in E coli strains based on 23S rrn restriction profiling The primordial soup of E coli microbes wouldhave been comprised of two clonal groups types I and IV coexisting with each other but predominated by type I strains Over the courseof evolution on accumulating mutation as a means for variation type I or type IV would have given rise to type III by addition (former) ordeletion (latter) of recognition sequence The second scenario could be a rare event because accumulation of two mutations in a conservedregion would be difficult Type II being the rarest would have been evolved in similar basis to type III (b) Hypothesis of linking phylogroupsHae III profiles and clonal types in E coli This diagram illustrates the clonal groups of commensal E coli strains having both A phylogroups(dominant) and B1 phylogroups each being type I profiles Pathogenic counterparts showed dominancy in type III restriction pattern followedby type I pattern With respect to phylogroups pathovars were predominant in B2 and B1 followed by D and E groups
from housekeeping to repair are employed for characterizingbacteria of which rRNA is referred to as the gold stretch[41 42] Ribosomal segments organized as operon clusters of16S 23S and 5S rDNA and these assemblages are distributedfrom single to multiple copies across the bacterial genophore[43] The occurrence of redundancy in maintaining differentcopies of these genes is not associated with virulence but anin-built ecological strategy that contributes to the particularbacterium with a competitive edge and better chance ofsurvival [44] A futuristic approach of coupling the techniqueand results with antibiotic resistance pattern colony char-acteristics and genomic profiles of bacterial strains of bothcommensal and pathogenic nature can help in monitoringand managing infections in a better manner
4 Conclusion
The causatum of postgenomic era genomics and proteomicshas enhanced this progress by improvising the techniquesused for diagnostics The strategy of ARDRA utilized inthis study was rapid and a cost-effective approach to dif-ferentiate E coli from its colony morphovars along withgiving a predicament of nature of origin or clonal groupsThe mutations reported in this work for detection andestablishing clonality namely T874A (Bfa I) G2161A andG2205A (Hae III) have not been mentioned in any researchpaper or within the Ribosomal RNA Mutation Database
[45] making this finding aboriginal This basic methodologycan be adapted for detection of other bacterial organismsby targeting SNPs linked to recognition sequences therebyvalidating rrn sequences as marker for diagnostics A cumu-lative approach of detection of unique restriction pattern of atargeted gene and its cross-reference across bacterial familiescan circumvent changes of false positives
Disclosure
This work was presented (poster) in part at the 5th Interna-tional Barcoding of Life Kunming China 2013
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
Acknowledgments
The authors acknowledge and express their gratitude to Pro-fessor D A Bhiwgade (Ex-Director) and Professor DebjaniDasgupta Director School of Biotechnology amp Bioinformat-ics D Y Patil University as well as the management for theirsupport and for providing the infrastructure to carry out thecurrent work
10 Journal of Pathogens
References
[1] M Richter and R Rossello-Mora ldquoShifting the genomic goldstandard for the prokaryotic species definitionrdquo Proceedings ofthe National Academy of Sciences of the United States of Americavol 106 no 45 pp 19126ndash19131 2009
[2] A F Auch M von Jan H-P Klenk and M Goker ldquoDigitalDNA-DNA hybridization for microbial species delineation bymeans of genome-to-genome sequence comparisonrdquo Standardsin Genomic Sciences vol 2 no 1 pp 117ndash134 2010
[3] D H Martin M Nsuami J Schachter et al ldquoUse of multiplenucleic acid amplification tests to define the infected-patientlsquogold standardrsquo in clinical trials of new diagnostic tests forChlamydia trachomatis infectionsrdquo Journal of Clinical Microbi-ology vol 42 no 10 pp 4749ndash4758 2004
[4] E A Mothershed and A M Whitney ldquoNucleic acid-basedmethods for the detection of bacterial pathogens presentand future considerations for the clinical laboratoryrdquo ClinicaChimica Acta vol 363 no 1-2 pp 206ndash220 2006
[5] J E Clarridge III ldquoImpact of 16S rRNA gene sequence analysisfor identification of bacteria on clinical microbiology andinfectious diseasesrdquo Clinical Microbiology Reviews vol 17 no4 pp 840ndash862 2004
[6] O Mizrahi-Man E R Davenport and Y Gilad ldquoTaxonomicclassification of bacterial 16S rRNA genes using short sequenc-ing reads evaluation of effective study designsrdquo PLoS ONE vol8 no 1 Article ID e53608 2013
[7] D E Lebonah A Dileep K Chandrasekhar S Sreevani BSreedevi and J Pramoda Kumari ldquoDNA barcoding on bacteriaa reviewrdquo Advances in Biology vol 2014 Article ID 541787 9pages 2014
[8] J Rajendhran and P Gunasekaran ldquoMicrobial phylogeny anddiversity small subunit ribosomal RNA sequence analysis andbeyondrdquo Microbiological Research vol 166 no 2 pp 99ndash1102011
[9] R Srinivasan U Karaoz M Volegova et al ldquoUse of 16S rRNAgene for identification of a broad range of clinically relevantbacterial pathogensrdquo PLoS ONE vol 10 no 2 Article IDe0117617 2015
[10] S Chakravorty D Helb M Burday N Connell and D AllandldquoA detailed analysis of 16S ribosomal RNA gene segments forthe diagnosis of pathogenic bacteriardquo Journal of MicrobiologicalMethods vol 69 no 2 pp 330ndash339 2007
[11] C R Woese ldquoReview bacterial evolutionrdquo MicrobiologicalReviews vol 51 pp 221ndash271 1987
[12] M Touchon C Hoede O Tenaillon et al ldquoOrganised genomedynamics in the Escherichia coli species results in highly diverseadaptive pathsrdquo PLoS Genetics vol 5 no 1 Article ID e10003442009
[13] J M Janda and S L Abbott ldquo16S rRNA gene sequencingfor bacterial identification in the diagnostic laboratory plusesperils and pitfallsrdquo Journal of Clinical Microbiology vol 45 no9 pp 2761ndash2764 2007
[14] J-J Lu C-L Perng S-Y Lee and C-CWan ldquoUse of PCRwithuniversal primers and restriction endonuclease digestions fordetection and identification of common bacterial pathogens incerebrospinal fluidrdquo Journal of Clinical Microbiology vol 38 no6 pp 2076ndash2080 2000
[15] N M Guentzel ldquoEscherichia Klebsiella Enterobacter SerratiaCitrobacter andProteusrdquo inMedicalMicrobiology S Baron Edchapter 26 University of Texas Medical Branch at GalvestonGalveston Tex USA 1996
[16] P Behzadip E Behzadi H Yazdanbod R Aghapour A MCheshmeh and S D Omran ldquoA survey on urinary tract infec-tions associated with the three most common uropathogenicbacteriardquoMaedica (Buchar) vol 5 no 2 pp 111ndash115 2010
[17] B CMetri P Jyothi andBV Peerapur ldquoAntibiotic resistance inCitrobacter spp isolated from urinary tract infectionrdquo UrologyAnnals vol 5 no 4 pp 312ndash313 2013
[18] N Taneja S S Chatterjee M Singh S Singh and M SharmaldquoPediatric urinary tract infections in a tertiary care center fromnorth Indiardquo Indian Journal of Medical Research vol 131 no 1pp 101ndash105 2010
[19] J B Kaper J P Nataro and H L T Mobley ldquoPathogenicEscherichia colirdquo Nature Reviews Microbiology vol 2 no 2 pp123ndash140 2004
[20] E J de Muinck K Lagesen J E Afset et al ldquoComparisonsof infant Escherichia coli isolates link genomic profiles withadaptation to the ecological nicherdquo BMC Genomics vol 14 no1 article 81 2013
[21] D A Rasko M J Rosovitz G S A Myers et al ldquoThepangenome structure of Escherichia coli comparative genomicanalysis of E coli commensal and pathogenic isolatesrdquo Journalof Bacteriology vol 190 no 20 pp 6881ndash6893 2008
[22] C T Archer J F Kim H Jeong et al ldquoThe genome sequenceof E coli W (ATCC 9637) comparative genome analysis andan improved genome-scale reconstruction of E colirdquo BMCGenomics vol 12 article no 9 2011
[23] PKuhnert J Nicolet and J Frey ldquoRapid and accurate identifica-tion of Escherichia coliK-12 strainsrdquoApplied and EnvironmentalMicrobiology vol 61 no 11 pp 4135ndash4139 1995
[24] A P Bauer S M Dieckmann W Ludwig and K-H SchleiferldquoRapid identification of Escherichia coli safety and laboratorystrain lineages based on multiplex-PCRrdquo FEMS MicrobiologyLetters vol 269 no 1 pp 36ndash40 2007
[25] B J Bachmann ldquoPedigrees of some mutant strains ofEscherichia coli K-12rdquo Bacteriological Reviews vol 36 no 4 pp525ndash557 1972
[26] P Daegelen F W Studier R E Lenski S Cure and J F KimldquoTracing ancestors and relatives of Escherichia coli B and thederivation of B strains REL606 and BL21(DE3)rdquo Journal ofMolecular Biology vol 394 no 4 pp 634ndash643 2009
[27] F W Studier P Daegelen R E Lenski S Maslov and J F KimldquoUnderstanding the differences between genome sequences ofEscherichia coli B strains REL606 and BL21(DE3) and compar-ison of the E coli B and K-12 genomesrdquo Journal of MolecularBiology vol 394 no 4 pp 653ndash680 2009
[28] R K Selander and B R Levin ldquoGenetic diversity and structurein Escherichia coli populationsrdquo Science vol 210 no 4469 pp545ndash547 1980
[29] D M Gordon O Clermont H Tolley and E DenamurldquoAssigning Escherichia coli strains to phylogenetic groupsmulti-locus sequence typing versus the PCR triplex methodrdquoEnvironmental Microbiology vol 10 no 10 pp 2484ndash24962008
[30] P Desjardins B Picard B Kaltenbock J Elion and EDenamur ldquoSex in Escherichia coli does not disrupt the clonalstructure of the population evidence from random amplifiedpolymorphic DNA and restriction-fragment-length polymor-phismrdquo Journal of Molecular Evolution vol 41 no 4 pp 440ndash448 1995
Journal of Pathogens 11
[31] N D Jasuja R Saxena S Chandra and S C Joshi ldquoIsolationand identification of microorganism from polyhouse agricul-ture soil of RajasthanrdquoAfrican Journal ofMicrobiology Researchvol 7 no 41 pp 4886ndash4891 2013
[32] V Hemraj S Diksha and G Avneet ldquoA review of commonlyused biochemical test for bacteriardquo Innovare Journal of LifeSciences vol 1 pp 1ndash7 2013
[33] J D Thompson D G Higgins and T J Gibson ldquoCLUSTALWimproving the sensitivity of progressive multiple sequencealignment through sequence weighting position-specific gappenalties and weight matrix choicerdquoNucleic Acids Research vol22 no 22 pp 4673ndash4680 1994
[34] T Vincze J Posfai and R J Roberts ldquoNEBcutter a program tocleave DNA with restriction enzymesrdquo Nucleic Acids Researchvol 31 no 13 pp 3688ndash3691 2003
[35] J Ye G Coulouris I Zaretskaya I Cutcutache S Rozen andT Madden ldquoPrimer-BLAST a tool to design target-specificprimers for polymerase chain reactionrdquo BMC Bioinformaticsvol 13 article 134 2012
[36] J R Parvathi and S Singh ldquoFacile approaches for microbialDNA extraction comparative study with colonies lsquopickedrsquo fromthree different bacteriological mediumrdquo Journal of Pure andApplied Microbiology vol 9 no 3 pp 2373ndash2378 2015
[37] R Lan and P R Reeves ldquoEscherichia coli in disguise molecularorigins of Shigellardquo Microbes and Infection vol 4 no 11 pp1125ndash1132 2002
[38] D J Leininger J R Roberson and F Elvinger ldquoUse ofeosin methylene blue agar to differentiate Escherichia coli fromother gram-negative mastitis pathogensrdquo Journal of VeterinaryDiagnostic Investigation vol 13 no 3 pp 273ndash275 2001
[39] U Dobrindt F Agerer K Michaelis et al ldquoAnalysis of genomeplasticity in pathogenic and commensal Escherichia coli isolatesby use of DNA arraysrdquo Journal of Bacteriology vol 185 no 6pp 1831ndash1840 2003
[40] V Savolainen R S Cowan A P Vogler G K Roderickand R Lane ldquoTowards writing the encyclopaedia of life anintroduction to DNA barcodingrdquo Philosophical Transactions ofthe Royal Society B Biological Sciences vol 360 no 1462 pp1805ndash1811 2005
[41] T A Isenbarger C E Carr S S Johnson et al ldquoThe mostconserved genome segments for life detection on earth andother planetsrdquo Origins of Life and Evolution of Biospheres vol38 no 6 pp 517ndash533 2008
[42] S R Santos and H Ochman ldquoIdentification and phylogeneticsorting of bacterial lineages with universally conserved genesand proteinsrdquoEnvironmentalMicrobiology vol 6 no 7 pp 754ndash759 2004
[43] R Rastogi M Wu I DasGupta and G E Fox ldquoVisualizationof ribosomal RNA operon copy number distributionrdquo BMCMicrobiology vol 9 no 1 article 208 2009
[44] J A Klappenbach J M Dunbar and T M Schmidt ldquorRNAoperon copy number reflects ecological strategies of bacteriardquoApplied and Environmental Microbiology vol 66 no 4 pp1328ndash1333 2000
[45] J A Klappenbach P R Saxman J R Cole and T M SchmidtldquoRrndb the ribosomal RNA operon copy number databaserdquoNucleic Acids Research vol 29 no 1 pp 181ndash184 2001
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom

Journal of Pathogens 7
682bp
987654321
9998400
8998400
7998400
6998400
5998400
4998400
3998400
2998400
1998400
(a)
431bp
223 bp
210 bp
179bp
223 + 210 bp
223+
210+
87654321
8998400
7998400
6998400
5998400
4998400
3998400
2998400
1998400
(b)
Figure 3 (a) Broad range PCR of 23S P2682
with genomic DNA isolated from standard strains of E coli The upper row comprises Lane 1K-12 strains Lane 2 derivatives of K-12 strains Lane 3 DHI strain Lane 4 strain C1 Lane 6 Crookrsquos strain Lane 7 Walkerrsquos strain Lane 8descendent of W Lane 9 blank The lower row comprises Lane 11015840 B strain Lane 21015840 BL21 strain Lane 31015840 BL21DE3 Lane 41015840 BL21DE3Lys5(B3) Lane 61015840 E coli (ATCC 15223 commensal isolate) Lane 71015840 E coli (ATCC 11775 urine sample) Lane 81015840 E coli (ATCC 25922 clinical)Lanes 551015840 and Lanes 991015840 in both rows are 100 bp ladder and blank respectively (b) ARDRA of 23S P2
682
amplicons with Hae III Lane 1 ofboth rows comprises 100 bp ladder The digested products are loaded as per the order given in Figure 2(a)
introduced Hae III recognition patterns D6 gave lineages ofB1 phylogroups D9 giving rise to B2 clades (D12) of type Iprofiles and D clades with varied Hae III patterns (D11) D10gave rise to two nodes (with strains being type I groups)with one showing B1 phylogroups (D13) and the other beingpredominant in E phylogroups followed by B1 An almostexemplary corelation between the clonal groups and differentpathovars was found with this study Majority of commensalstrains and EIEC and all EAEC and EPEC were shown tobe having type I profile (Table 4) All UPEC ExPEC APECEIEC and single strain of EPEC and two commensal strainsshowed type III pattern B form strains along with a singlestrain of UPEC and environmental isolate exhibited type IVprofiles
Similarly a near-pristine association between phylo-groups and Hae III profiles was established Strains comingunder phylogroups A were preponderant in type I whereastype IV profiles were exclusively for B forms Type I profileswere observed in the strains (119899) in the order of A gt B1 gt E gtDwith type III dominant in B2 strains Phylogroup D showedequal occurrence of type III and type IV fragmentationpattern followed by a single strain with type I profile Theseresults assisted in enumerating a conjecture which works upa scenario of primordial microbial population where E colistrains had two clonal types (Figure 5(a)) which coexistedtogether (type I and type IV clones) Of these type I strainswould have been the predominant clone in comparison totype IV based on the dendogram data with maximum genepool of root strains showing type I Type III may have beenthe next predominant clone (an intermediary one) that hasevolved either from type I (by addition of a new recognition
Table 4 Corelation between phylogroups Hae III profiles andnature of E coli strains (genome data analysis of 57 E coli strainscollated from Genbank)
Strain Type A B1 B2 D E Total Out ofCommensal I 10 5 mdash mdash mdash 15 21Commensal III mdash mdash 2 mdash mdash 2 21Commensal IV 4 mdash mdash mdash mdash 4 21EPEC I mdash mdash mdash mdash 7 7 8EPEC III mdash mdash 1 mdash mdash 1 8EAEC I mdash 4 mdash 1 mdash 5 5EHEC I mdash 2 mdash mdash mdash 2 3EHEC II mdash 1 mdash mdash mdash 1 3ETEC I 1 1 1 mdash mdash 3 3UPEC III mdash mdash 7 mdash mdash 7 8UPEC IV mdash mdash mdash 1 mdash 1 8EIEC III mdash mdash 3 mdash mdash 3 3ExPEC III mdash mdash 2 2 mdash 4 4APEC III mdash mdash 1 mdash 1 1ENV IV mdash mdash mdash 1 1 1APEC avian pathogenic EAEC enteroaggressive EHEC enterohemorrhag-ic ETEC enterotoxigenic EIEC enteroinvasive EPEC enteropathogenicENV environmental ExPEC extraintestinal pathogenic UPEC uropatho-genic
site) or from type IV (by deletion of one recognition site)strains In a different scenario maybe all three types coexistedor type III would have given rise to type IVwith an additionalmutational event This situation may be hardly convincing
8 Journal of Pathogens
D1
D2
D3
D4
D5
D6
D7
D8
D9
D10
D11
D12
D13
D14
BL21(DE3)_1lowastBL21(DE3)_2lowast
BL21-Gold(DE3)pLysS AGlowast
B str REL606lowastA phylogroup
ETEC H10407lowast (B2)
UMNK88 (A)
(B1 clades) Str 2009EL-2050Str 2009EL-2071
Str 2011-349355989
B1 phylogroup
Str EC4115Str TW14359
Str EDL933Str Sakai
Xuzhou21Str CB9615
Str RM12598
E phylogroup
Str 11128Str 11368 B1 phylogroup
NA114
SE15LF82lowastlowastlowastStr NRG 857Clowastlowastlowast
UM146
UTI89IHE3034
APEC O1lowastlowastS88
Clone D i2Clone D i14
ABU 83972
CFT073536
ED1aStr E234869
B2 phylogroup
UMN026
IAI39Str CE10
SMS-3-5
D phylogroup
O42lowastlowastlowast
(B2 clades)
(D clades)
(B1 clades)
Wlowastlowastlowast
WKO11FLlowastlowast
KO11FLE24377A
SE11
B1 phylogroup
IAI1ATCC 8739
P12blowastHS
BW2952
K-12 substr DH10BDH1
DH1 (ME8569)K-12 substr W3110
K-12 substr MG1655
A phylogroup
1
2
10
3156
Str 12009lowastlowast
Commensal ETEC EHEC EPEC UPEC
ExPECAPECEAECEIECENV
Figure 4 Analysis of distribution of Hae III profiles phylogroups and pathovars within the global alignment dendogram constructcomprising 57 E coli genomes The dendogram of the whole genome sequence of 57 genomes was obtained from the NCBI web site forE coli and was redrawn using Photoshop CS6 for colour coordination for pathogenic and nonpathogenic strains and inputting the clonalgroups and phylogroups (httpwwwncbinlmnihgovgenometerm=escherichia+coli) Among the 57 genomes 32 were of type I (56)18 were of type III (31) and 6 and 1 were of type IV (10) and type II (2) respectively showing that type I is predominant and type IIthe rarest To correlate between the phylogroupsHae III profiles and pathovars the lineages were considered as clades derived from variousdivergence point (D) during the course of evolution
as dendogram segregation shows type III strains evolving orpresent after type IV strains The clustering also shows thatpredominance of type III is more than type IV and maybeevolutionary processesmutation have triggered the increasein type III profiles On similar line type II clones would havebeen evolved from type I (addition) or type IV (deletion)but due to the very limited type II restriction profile patterns(rare) as observed from the gene pool data the chances orapproximation of such event is difficult to prove Based onthese results no new insights into phylogroup distributionacross candidate strains can be provided however withrespect to the Hae III profile an additional angle can beconsidered Phylogroups A and B1 are predominant withtype I Hae III profiles whereas B2 phylogroups exclusivelyrepresented type III profile (Figure 5(b))
An intrinsic metagenomic and pangenomic analysis ofmost bacterial species exhibited mosaicisms of their inherentblueprint [39] In such situations core gene elements sharedacross the species are limited in comparison to dispensable(flexible) genetic contents that concede uniqueness to aparticular strain These flexible contents help in niche adapt-ability and give bacteria the ability to infect and invade Amajor concern is while diagnosing both true and opportunistpathogenic bacteria the former causing disease in virtuallyany susceptible host and the latter invoking diseases inimmunocompromised individuals ldquoMolecular barcodingrdquoan evolved term which implies the method of allele specificamplification of relatively small DNA sequences or barcodes[40] for spotting a particular species is a thrust area formicrobial diagnostics A plethora of functional genes ranging
Journal of Pathogens 9
Type I(28 459)
Type IV(28 207 249 and 459)
Hypothesis
Type III(28 249 and 459)
+ minus
+
Primordial
NonpathogenicA gt B1 gt B2
type I gt IV gt III
PathogenicB2 gt B1 gt E gt D
type I (IAT)III(P)gt type IVII
Predominant clones
Rarest
+minus
Type II(28 207 and 459)
Restriction profilesand
clonesPhylogroups and
clones
2∘ clonesintermediate
(a) (b)
Figure 5 (a) Hypothesis of clonality in E coli strains based on 23S rrn restriction profiling The primordial soup of E coli microbes wouldhave been comprised of two clonal groups types I and IV coexisting with each other but predominated by type I strains Over the courseof evolution on accumulating mutation as a means for variation type I or type IV would have given rise to type III by addition (former) ordeletion (latter) of recognition sequence The second scenario could be a rare event because accumulation of two mutations in a conservedregion would be difficult Type II being the rarest would have been evolved in similar basis to type III (b) Hypothesis of linking phylogroupsHae III profiles and clonal types in E coli This diagram illustrates the clonal groups of commensal E coli strains having both A phylogroups(dominant) and B1 phylogroups each being type I profiles Pathogenic counterparts showed dominancy in type III restriction pattern followedby type I pattern With respect to phylogroups pathovars were predominant in B2 and B1 followed by D and E groups
from housekeeping to repair are employed for characterizingbacteria of which rRNA is referred to as the gold stretch[41 42] Ribosomal segments organized as operon clusters of16S 23S and 5S rDNA and these assemblages are distributedfrom single to multiple copies across the bacterial genophore[43] The occurrence of redundancy in maintaining differentcopies of these genes is not associated with virulence but anin-built ecological strategy that contributes to the particularbacterium with a competitive edge and better chance ofsurvival [44] A futuristic approach of coupling the techniqueand results with antibiotic resistance pattern colony char-acteristics and genomic profiles of bacterial strains of bothcommensal and pathogenic nature can help in monitoringand managing infections in a better manner
4 Conclusion
The causatum of postgenomic era genomics and proteomicshas enhanced this progress by improvising the techniquesused for diagnostics The strategy of ARDRA utilized inthis study was rapid and a cost-effective approach to dif-ferentiate E coli from its colony morphovars along withgiving a predicament of nature of origin or clonal groupsThe mutations reported in this work for detection andestablishing clonality namely T874A (Bfa I) G2161A andG2205A (Hae III) have not been mentioned in any researchpaper or within the Ribosomal RNA Mutation Database
[45] making this finding aboriginal This basic methodologycan be adapted for detection of other bacterial organismsby targeting SNPs linked to recognition sequences therebyvalidating rrn sequences as marker for diagnostics A cumu-lative approach of detection of unique restriction pattern of atargeted gene and its cross-reference across bacterial familiescan circumvent changes of false positives
Disclosure
This work was presented (poster) in part at the 5th Interna-tional Barcoding of Life Kunming China 2013
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
Acknowledgments
The authors acknowledge and express their gratitude to Pro-fessor D A Bhiwgade (Ex-Director) and Professor DebjaniDasgupta Director School of Biotechnology amp Bioinformat-ics D Y Patil University as well as the management for theirsupport and for providing the infrastructure to carry out thecurrent work
10 Journal of Pathogens
References
[1] M Richter and R Rossello-Mora ldquoShifting the genomic goldstandard for the prokaryotic species definitionrdquo Proceedings ofthe National Academy of Sciences of the United States of Americavol 106 no 45 pp 19126ndash19131 2009
[2] A F Auch M von Jan H-P Klenk and M Goker ldquoDigitalDNA-DNA hybridization for microbial species delineation bymeans of genome-to-genome sequence comparisonrdquo Standardsin Genomic Sciences vol 2 no 1 pp 117ndash134 2010
[3] D H Martin M Nsuami J Schachter et al ldquoUse of multiplenucleic acid amplification tests to define the infected-patientlsquogold standardrsquo in clinical trials of new diagnostic tests forChlamydia trachomatis infectionsrdquo Journal of Clinical Microbi-ology vol 42 no 10 pp 4749ndash4758 2004
[4] E A Mothershed and A M Whitney ldquoNucleic acid-basedmethods for the detection of bacterial pathogens presentand future considerations for the clinical laboratoryrdquo ClinicaChimica Acta vol 363 no 1-2 pp 206ndash220 2006
[5] J E Clarridge III ldquoImpact of 16S rRNA gene sequence analysisfor identification of bacteria on clinical microbiology andinfectious diseasesrdquo Clinical Microbiology Reviews vol 17 no4 pp 840ndash862 2004
[6] O Mizrahi-Man E R Davenport and Y Gilad ldquoTaxonomicclassification of bacterial 16S rRNA genes using short sequenc-ing reads evaluation of effective study designsrdquo PLoS ONE vol8 no 1 Article ID e53608 2013
[7] D E Lebonah A Dileep K Chandrasekhar S Sreevani BSreedevi and J Pramoda Kumari ldquoDNA barcoding on bacteriaa reviewrdquo Advances in Biology vol 2014 Article ID 541787 9pages 2014
[8] J Rajendhran and P Gunasekaran ldquoMicrobial phylogeny anddiversity small subunit ribosomal RNA sequence analysis andbeyondrdquo Microbiological Research vol 166 no 2 pp 99ndash1102011
[9] R Srinivasan U Karaoz M Volegova et al ldquoUse of 16S rRNAgene for identification of a broad range of clinically relevantbacterial pathogensrdquo PLoS ONE vol 10 no 2 Article IDe0117617 2015
[10] S Chakravorty D Helb M Burday N Connell and D AllandldquoA detailed analysis of 16S ribosomal RNA gene segments forthe diagnosis of pathogenic bacteriardquo Journal of MicrobiologicalMethods vol 69 no 2 pp 330ndash339 2007
[11] C R Woese ldquoReview bacterial evolutionrdquo MicrobiologicalReviews vol 51 pp 221ndash271 1987
[12] M Touchon C Hoede O Tenaillon et al ldquoOrganised genomedynamics in the Escherichia coli species results in highly diverseadaptive pathsrdquo PLoS Genetics vol 5 no 1 Article ID e10003442009
[13] J M Janda and S L Abbott ldquo16S rRNA gene sequencingfor bacterial identification in the diagnostic laboratory plusesperils and pitfallsrdquo Journal of Clinical Microbiology vol 45 no9 pp 2761ndash2764 2007
[14] J-J Lu C-L Perng S-Y Lee and C-CWan ldquoUse of PCRwithuniversal primers and restriction endonuclease digestions fordetection and identification of common bacterial pathogens incerebrospinal fluidrdquo Journal of Clinical Microbiology vol 38 no6 pp 2076ndash2080 2000
[15] N M Guentzel ldquoEscherichia Klebsiella Enterobacter SerratiaCitrobacter andProteusrdquo inMedicalMicrobiology S Baron Edchapter 26 University of Texas Medical Branch at GalvestonGalveston Tex USA 1996
[16] P Behzadip E Behzadi H Yazdanbod R Aghapour A MCheshmeh and S D Omran ldquoA survey on urinary tract infec-tions associated with the three most common uropathogenicbacteriardquoMaedica (Buchar) vol 5 no 2 pp 111ndash115 2010
[17] B CMetri P Jyothi andBV Peerapur ldquoAntibiotic resistance inCitrobacter spp isolated from urinary tract infectionrdquo UrologyAnnals vol 5 no 4 pp 312ndash313 2013
[18] N Taneja S S Chatterjee M Singh S Singh and M SharmaldquoPediatric urinary tract infections in a tertiary care center fromnorth Indiardquo Indian Journal of Medical Research vol 131 no 1pp 101ndash105 2010
[19] J B Kaper J P Nataro and H L T Mobley ldquoPathogenicEscherichia colirdquo Nature Reviews Microbiology vol 2 no 2 pp123ndash140 2004
[20] E J de Muinck K Lagesen J E Afset et al ldquoComparisonsof infant Escherichia coli isolates link genomic profiles withadaptation to the ecological nicherdquo BMC Genomics vol 14 no1 article 81 2013
[21] D A Rasko M J Rosovitz G S A Myers et al ldquoThepangenome structure of Escherichia coli comparative genomicanalysis of E coli commensal and pathogenic isolatesrdquo Journalof Bacteriology vol 190 no 20 pp 6881ndash6893 2008
[22] C T Archer J F Kim H Jeong et al ldquoThe genome sequenceof E coli W (ATCC 9637) comparative genome analysis andan improved genome-scale reconstruction of E colirdquo BMCGenomics vol 12 article no 9 2011
[23] PKuhnert J Nicolet and J Frey ldquoRapid and accurate identifica-tion of Escherichia coliK-12 strainsrdquoApplied and EnvironmentalMicrobiology vol 61 no 11 pp 4135ndash4139 1995
[24] A P Bauer S M Dieckmann W Ludwig and K-H SchleiferldquoRapid identification of Escherichia coli safety and laboratorystrain lineages based on multiplex-PCRrdquo FEMS MicrobiologyLetters vol 269 no 1 pp 36ndash40 2007
[25] B J Bachmann ldquoPedigrees of some mutant strains ofEscherichia coli K-12rdquo Bacteriological Reviews vol 36 no 4 pp525ndash557 1972
[26] P Daegelen F W Studier R E Lenski S Cure and J F KimldquoTracing ancestors and relatives of Escherichia coli B and thederivation of B strains REL606 and BL21(DE3)rdquo Journal ofMolecular Biology vol 394 no 4 pp 634ndash643 2009
[27] F W Studier P Daegelen R E Lenski S Maslov and J F KimldquoUnderstanding the differences between genome sequences ofEscherichia coli B strains REL606 and BL21(DE3) and compar-ison of the E coli B and K-12 genomesrdquo Journal of MolecularBiology vol 394 no 4 pp 653ndash680 2009
[28] R K Selander and B R Levin ldquoGenetic diversity and structurein Escherichia coli populationsrdquo Science vol 210 no 4469 pp545ndash547 1980
[29] D M Gordon O Clermont H Tolley and E DenamurldquoAssigning Escherichia coli strains to phylogenetic groupsmulti-locus sequence typing versus the PCR triplex methodrdquoEnvironmental Microbiology vol 10 no 10 pp 2484ndash24962008
[30] P Desjardins B Picard B Kaltenbock J Elion and EDenamur ldquoSex in Escherichia coli does not disrupt the clonalstructure of the population evidence from random amplifiedpolymorphic DNA and restriction-fragment-length polymor-phismrdquo Journal of Molecular Evolution vol 41 no 4 pp 440ndash448 1995
Journal of Pathogens 11
[31] N D Jasuja R Saxena S Chandra and S C Joshi ldquoIsolationand identification of microorganism from polyhouse agricul-ture soil of RajasthanrdquoAfrican Journal ofMicrobiology Researchvol 7 no 41 pp 4886ndash4891 2013
[32] V Hemraj S Diksha and G Avneet ldquoA review of commonlyused biochemical test for bacteriardquo Innovare Journal of LifeSciences vol 1 pp 1ndash7 2013
[33] J D Thompson D G Higgins and T J Gibson ldquoCLUSTALWimproving the sensitivity of progressive multiple sequencealignment through sequence weighting position-specific gappenalties and weight matrix choicerdquoNucleic Acids Research vol22 no 22 pp 4673ndash4680 1994
[34] T Vincze J Posfai and R J Roberts ldquoNEBcutter a program tocleave DNA with restriction enzymesrdquo Nucleic Acids Researchvol 31 no 13 pp 3688ndash3691 2003
[35] J Ye G Coulouris I Zaretskaya I Cutcutache S Rozen andT Madden ldquoPrimer-BLAST a tool to design target-specificprimers for polymerase chain reactionrdquo BMC Bioinformaticsvol 13 article 134 2012
[36] J R Parvathi and S Singh ldquoFacile approaches for microbialDNA extraction comparative study with colonies lsquopickedrsquo fromthree different bacteriological mediumrdquo Journal of Pure andApplied Microbiology vol 9 no 3 pp 2373ndash2378 2015
[37] R Lan and P R Reeves ldquoEscherichia coli in disguise molecularorigins of Shigellardquo Microbes and Infection vol 4 no 11 pp1125ndash1132 2002
[38] D J Leininger J R Roberson and F Elvinger ldquoUse ofeosin methylene blue agar to differentiate Escherichia coli fromother gram-negative mastitis pathogensrdquo Journal of VeterinaryDiagnostic Investigation vol 13 no 3 pp 273ndash275 2001
[39] U Dobrindt F Agerer K Michaelis et al ldquoAnalysis of genomeplasticity in pathogenic and commensal Escherichia coli isolatesby use of DNA arraysrdquo Journal of Bacteriology vol 185 no 6pp 1831ndash1840 2003
[40] V Savolainen R S Cowan A P Vogler G K Roderickand R Lane ldquoTowards writing the encyclopaedia of life anintroduction to DNA barcodingrdquo Philosophical Transactions ofthe Royal Society B Biological Sciences vol 360 no 1462 pp1805ndash1811 2005
[41] T A Isenbarger C E Carr S S Johnson et al ldquoThe mostconserved genome segments for life detection on earth andother planetsrdquo Origins of Life and Evolution of Biospheres vol38 no 6 pp 517ndash533 2008
[42] S R Santos and H Ochman ldquoIdentification and phylogeneticsorting of bacterial lineages with universally conserved genesand proteinsrdquoEnvironmentalMicrobiology vol 6 no 7 pp 754ndash759 2004
[43] R Rastogi M Wu I DasGupta and G E Fox ldquoVisualizationof ribosomal RNA operon copy number distributionrdquo BMCMicrobiology vol 9 no 1 article 208 2009
[44] J A Klappenbach J M Dunbar and T M Schmidt ldquorRNAoperon copy number reflects ecological strategies of bacteriardquoApplied and Environmental Microbiology vol 66 no 4 pp1328ndash1333 2000
[45] J A Klappenbach P R Saxman J R Cole and T M SchmidtldquoRrndb the ribosomal RNA operon copy number databaserdquoNucleic Acids Research vol 29 no 1 pp 181ndash184 2001
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom
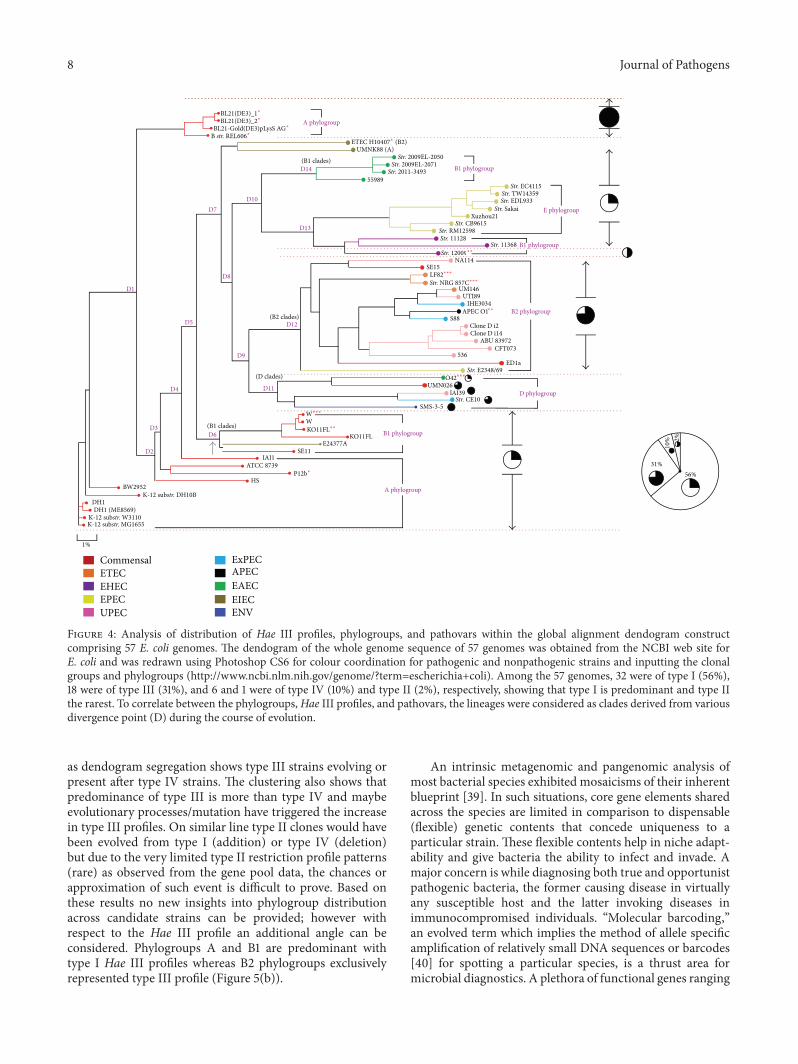
8 Journal of Pathogens
D1
D2
D3
D4
D5
D6
D7
D8
D9
D10
D11
D12
D13
D14
BL21(DE3)_1lowastBL21(DE3)_2lowast
BL21-Gold(DE3)pLysS AGlowast
B str REL606lowastA phylogroup
ETEC H10407lowast (B2)
UMNK88 (A)
(B1 clades) Str 2009EL-2050Str 2009EL-2071
Str 2011-349355989
B1 phylogroup
Str EC4115Str TW14359
Str EDL933Str Sakai
Xuzhou21Str CB9615
Str RM12598
E phylogroup
Str 11128Str 11368 B1 phylogroup
NA114
SE15LF82lowastlowastlowastStr NRG 857Clowastlowastlowast
UM146
UTI89IHE3034
APEC O1lowastlowastS88
Clone D i2Clone D i14
ABU 83972
CFT073536
ED1aStr E234869
B2 phylogroup
UMN026
IAI39Str CE10
SMS-3-5
D phylogroup
O42lowastlowastlowast
(B2 clades)
(D clades)
(B1 clades)
Wlowastlowastlowast
WKO11FLlowastlowast
KO11FLE24377A
SE11
B1 phylogroup
IAI1ATCC 8739
P12blowastHS
BW2952
K-12 substr DH10BDH1
DH1 (ME8569)K-12 substr W3110
K-12 substr MG1655
A phylogroup
1
2
10
3156
Str 12009lowastlowast
Commensal ETEC EHEC EPEC UPEC
ExPECAPECEAECEIECENV
Figure 4 Analysis of distribution of Hae III profiles phylogroups and pathovars within the global alignment dendogram constructcomprising 57 E coli genomes The dendogram of the whole genome sequence of 57 genomes was obtained from the NCBI web site forE coli and was redrawn using Photoshop CS6 for colour coordination for pathogenic and nonpathogenic strains and inputting the clonalgroups and phylogroups (httpwwwncbinlmnihgovgenometerm=escherichia+coli) Among the 57 genomes 32 were of type I (56)18 were of type III (31) and 6 and 1 were of type IV (10) and type II (2) respectively showing that type I is predominant and type IIthe rarest To correlate between the phylogroupsHae III profiles and pathovars the lineages were considered as clades derived from variousdivergence point (D) during the course of evolution
as dendogram segregation shows type III strains evolving orpresent after type IV strains The clustering also shows thatpredominance of type III is more than type IV and maybeevolutionary processesmutation have triggered the increasein type III profiles On similar line type II clones would havebeen evolved from type I (addition) or type IV (deletion)but due to the very limited type II restriction profile patterns(rare) as observed from the gene pool data the chances orapproximation of such event is difficult to prove Based onthese results no new insights into phylogroup distributionacross candidate strains can be provided however withrespect to the Hae III profile an additional angle can beconsidered Phylogroups A and B1 are predominant withtype I Hae III profiles whereas B2 phylogroups exclusivelyrepresented type III profile (Figure 5(b))
An intrinsic metagenomic and pangenomic analysis ofmost bacterial species exhibited mosaicisms of their inherentblueprint [39] In such situations core gene elements sharedacross the species are limited in comparison to dispensable(flexible) genetic contents that concede uniqueness to aparticular strain These flexible contents help in niche adapt-ability and give bacteria the ability to infect and invade Amajor concern is while diagnosing both true and opportunistpathogenic bacteria the former causing disease in virtuallyany susceptible host and the latter invoking diseases inimmunocompromised individuals ldquoMolecular barcodingrdquoan evolved term which implies the method of allele specificamplification of relatively small DNA sequences or barcodes[40] for spotting a particular species is a thrust area formicrobial diagnostics A plethora of functional genes ranging
Journal of Pathogens 9
Type I(28 459)
Type IV(28 207 249 and 459)
Hypothesis
Type III(28 249 and 459)
+ minus
+
Primordial
NonpathogenicA gt B1 gt B2
type I gt IV gt III
PathogenicB2 gt B1 gt E gt D
type I (IAT)III(P)gt type IVII
Predominant clones
Rarest
+minus
Type II(28 207 and 459)
Restriction profilesand
clonesPhylogroups and
clones
2∘ clonesintermediate
(a) (b)
Figure 5 (a) Hypothesis of clonality in E coli strains based on 23S rrn restriction profiling The primordial soup of E coli microbes wouldhave been comprised of two clonal groups types I and IV coexisting with each other but predominated by type I strains Over the courseof evolution on accumulating mutation as a means for variation type I or type IV would have given rise to type III by addition (former) ordeletion (latter) of recognition sequence The second scenario could be a rare event because accumulation of two mutations in a conservedregion would be difficult Type II being the rarest would have been evolved in similar basis to type III (b) Hypothesis of linking phylogroupsHae III profiles and clonal types in E coli This diagram illustrates the clonal groups of commensal E coli strains having both A phylogroups(dominant) and B1 phylogroups each being type I profiles Pathogenic counterparts showed dominancy in type III restriction pattern followedby type I pattern With respect to phylogroups pathovars were predominant in B2 and B1 followed by D and E groups
from housekeeping to repair are employed for characterizingbacteria of which rRNA is referred to as the gold stretch[41 42] Ribosomal segments organized as operon clusters of16S 23S and 5S rDNA and these assemblages are distributedfrom single to multiple copies across the bacterial genophore[43] The occurrence of redundancy in maintaining differentcopies of these genes is not associated with virulence but anin-built ecological strategy that contributes to the particularbacterium with a competitive edge and better chance ofsurvival [44] A futuristic approach of coupling the techniqueand results with antibiotic resistance pattern colony char-acteristics and genomic profiles of bacterial strains of bothcommensal and pathogenic nature can help in monitoringand managing infections in a better manner
4 Conclusion
The causatum of postgenomic era genomics and proteomicshas enhanced this progress by improvising the techniquesused for diagnostics The strategy of ARDRA utilized inthis study was rapid and a cost-effective approach to dif-ferentiate E coli from its colony morphovars along withgiving a predicament of nature of origin or clonal groupsThe mutations reported in this work for detection andestablishing clonality namely T874A (Bfa I) G2161A andG2205A (Hae III) have not been mentioned in any researchpaper or within the Ribosomal RNA Mutation Database
[45] making this finding aboriginal This basic methodologycan be adapted for detection of other bacterial organismsby targeting SNPs linked to recognition sequences therebyvalidating rrn sequences as marker for diagnostics A cumu-lative approach of detection of unique restriction pattern of atargeted gene and its cross-reference across bacterial familiescan circumvent changes of false positives
Disclosure
This work was presented (poster) in part at the 5th Interna-tional Barcoding of Life Kunming China 2013
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
Acknowledgments
The authors acknowledge and express their gratitude to Pro-fessor D A Bhiwgade (Ex-Director) and Professor DebjaniDasgupta Director School of Biotechnology amp Bioinformat-ics D Y Patil University as well as the management for theirsupport and for providing the infrastructure to carry out thecurrent work
10 Journal of Pathogens
References
[1] M Richter and R Rossello-Mora ldquoShifting the genomic goldstandard for the prokaryotic species definitionrdquo Proceedings ofthe National Academy of Sciences of the United States of Americavol 106 no 45 pp 19126ndash19131 2009
[2] A F Auch M von Jan H-P Klenk and M Goker ldquoDigitalDNA-DNA hybridization for microbial species delineation bymeans of genome-to-genome sequence comparisonrdquo Standardsin Genomic Sciences vol 2 no 1 pp 117ndash134 2010
[3] D H Martin M Nsuami J Schachter et al ldquoUse of multiplenucleic acid amplification tests to define the infected-patientlsquogold standardrsquo in clinical trials of new diagnostic tests forChlamydia trachomatis infectionsrdquo Journal of Clinical Microbi-ology vol 42 no 10 pp 4749ndash4758 2004
[4] E A Mothershed and A M Whitney ldquoNucleic acid-basedmethods for the detection of bacterial pathogens presentand future considerations for the clinical laboratoryrdquo ClinicaChimica Acta vol 363 no 1-2 pp 206ndash220 2006
[5] J E Clarridge III ldquoImpact of 16S rRNA gene sequence analysisfor identification of bacteria on clinical microbiology andinfectious diseasesrdquo Clinical Microbiology Reviews vol 17 no4 pp 840ndash862 2004
[6] O Mizrahi-Man E R Davenport and Y Gilad ldquoTaxonomicclassification of bacterial 16S rRNA genes using short sequenc-ing reads evaluation of effective study designsrdquo PLoS ONE vol8 no 1 Article ID e53608 2013
[7] D E Lebonah A Dileep K Chandrasekhar S Sreevani BSreedevi and J Pramoda Kumari ldquoDNA barcoding on bacteriaa reviewrdquo Advances in Biology vol 2014 Article ID 541787 9pages 2014
[8] J Rajendhran and P Gunasekaran ldquoMicrobial phylogeny anddiversity small subunit ribosomal RNA sequence analysis andbeyondrdquo Microbiological Research vol 166 no 2 pp 99ndash1102011
[9] R Srinivasan U Karaoz M Volegova et al ldquoUse of 16S rRNAgene for identification of a broad range of clinically relevantbacterial pathogensrdquo PLoS ONE vol 10 no 2 Article IDe0117617 2015
[10] S Chakravorty D Helb M Burday N Connell and D AllandldquoA detailed analysis of 16S ribosomal RNA gene segments forthe diagnosis of pathogenic bacteriardquo Journal of MicrobiologicalMethods vol 69 no 2 pp 330ndash339 2007
[11] C R Woese ldquoReview bacterial evolutionrdquo MicrobiologicalReviews vol 51 pp 221ndash271 1987
[12] M Touchon C Hoede O Tenaillon et al ldquoOrganised genomedynamics in the Escherichia coli species results in highly diverseadaptive pathsrdquo PLoS Genetics vol 5 no 1 Article ID e10003442009
[13] J M Janda and S L Abbott ldquo16S rRNA gene sequencingfor bacterial identification in the diagnostic laboratory plusesperils and pitfallsrdquo Journal of Clinical Microbiology vol 45 no9 pp 2761ndash2764 2007
[14] J-J Lu C-L Perng S-Y Lee and C-CWan ldquoUse of PCRwithuniversal primers and restriction endonuclease digestions fordetection and identification of common bacterial pathogens incerebrospinal fluidrdquo Journal of Clinical Microbiology vol 38 no6 pp 2076ndash2080 2000
[15] N M Guentzel ldquoEscherichia Klebsiella Enterobacter SerratiaCitrobacter andProteusrdquo inMedicalMicrobiology S Baron Edchapter 26 University of Texas Medical Branch at GalvestonGalveston Tex USA 1996
[16] P Behzadip E Behzadi H Yazdanbod R Aghapour A MCheshmeh and S D Omran ldquoA survey on urinary tract infec-tions associated with the three most common uropathogenicbacteriardquoMaedica (Buchar) vol 5 no 2 pp 111ndash115 2010
[17] B CMetri P Jyothi andBV Peerapur ldquoAntibiotic resistance inCitrobacter spp isolated from urinary tract infectionrdquo UrologyAnnals vol 5 no 4 pp 312ndash313 2013
[18] N Taneja S S Chatterjee M Singh S Singh and M SharmaldquoPediatric urinary tract infections in a tertiary care center fromnorth Indiardquo Indian Journal of Medical Research vol 131 no 1pp 101ndash105 2010
[19] J B Kaper J P Nataro and H L T Mobley ldquoPathogenicEscherichia colirdquo Nature Reviews Microbiology vol 2 no 2 pp123ndash140 2004
[20] E J de Muinck K Lagesen J E Afset et al ldquoComparisonsof infant Escherichia coli isolates link genomic profiles withadaptation to the ecological nicherdquo BMC Genomics vol 14 no1 article 81 2013
[21] D A Rasko M J Rosovitz G S A Myers et al ldquoThepangenome structure of Escherichia coli comparative genomicanalysis of E coli commensal and pathogenic isolatesrdquo Journalof Bacteriology vol 190 no 20 pp 6881ndash6893 2008
[22] C T Archer J F Kim H Jeong et al ldquoThe genome sequenceof E coli W (ATCC 9637) comparative genome analysis andan improved genome-scale reconstruction of E colirdquo BMCGenomics vol 12 article no 9 2011
[23] PKuhnert J Nicolet and J Frey ldquoRapid and accurate identifica-tion of Escherichia coliK-12 strainsrdquoApplied and EnvironmentalMicrobiology vol 61 no 11 pp 4135ndash4139 1995
[24] A P Bauer S M Dieckmann W Ludwig and K-H SchleiferldquoRapid identification of Escherichia coli safety and laboratorystrain lineages based on multiplex-PCRrdquo FEMS MicrobiologyLetters vol 269 no 1 pp 36ndash40 2007
[25] B J Bachmann ldquoPedigrees of some mutant strains ofEscherichia coli K-12rdquo Bacteriological Reviews vol 36 no 4 pp525ndash557 1972
[26] P Daegelen F W Studier R E Lenski S Cure and J F KimldquoTracing ancestors and relatives of Escherichia coli B and thederivation of B strains REL606 and BL21(DE3)rdquo Journal ofMolecular Biology vol 394 no 4 pp 634ndash643 2009
[27] F W Studier P Daegelen R E Lenski S Maslov and J F KimldquoUnderstanding the differences between genome sequences ofEscherichia coli B strains REL606 and BL21(DE3) and compar-ison of the E coli B and K-12 genomesrdquo Journal of MolecularBiology vol 394 no 4 pp 653ndash680 2009
[28] R K Selander and B R Levin ldquoGenetic diversity and structurein Escherichia coli populationsrdquo Science vol 210 no 4469 pp545ndash547 1980
[29] D M Gordon O Clermont H Tolley and E DenamurldquoAssigning Escherichia coli strains to phylogenetic groupsmulti-locus sequence typing versus the PCR triplex methodrdquoEnvironmental Microbiology vol 10 no 10 pp 2484ndash24962008
[30] P Desjardins B Picard B Kaltenbock J Elion and EDenamur ldquoSex in Escherichia coli does not disrupt the clonalstructure of the population evidence from random amplifiedpolymorphic DNA and restriction-fragment-length polymor-phismrdquo Journal of Molecular Evolution vol 41 no 4 pp 440ndash448 1995
Journal of Pathogens 11
[31] N D Jasuja R Saxena S Chandra and S C Joshi ldquoIsolationand identification of microorganism from polyhouse agricul-ture soil of RajasthanrdquoAfrican Journal ofMicrobiology Researchvol 7 no 41 pp 4886ndash4891 2013
[32] V Hemraj S Diksha and G Avneet ldquoA review of commonlyused biochemical test for bacteriardquo Innovare Journal of LifeSciences vol 1 pp 1ndash7 2013
[33] J D Thompson D G Higgins and T J Gibson ldquoCLUSTALWimproving the sensitivity of progressive multiple sequencealignment through sequence weighting position-specific gappenalties and weight matrix choicerdquoNucleic Acids Research vol22 no 22 pp 4673ndash4680 1994
[34] T Vincze J Posfai and R J Roberts ldquoNEBcutter a program tocleave DNA with restriction enzymesrdquo Nucleic Acids Researchvol 31 no 13 pp 3688ndash3691 2003
[35] J Ye G Coulouris I Zaretskaya I Cutcutache S Rozen andT Madden ldquoPrimer-BLAST a tool to design target-specificprimers for polymerase chain reactionrdquo BMC Bioinformaticsvol 13 article 134 2012
[36] J R Parvathi and S Singh ldquoFacile approaches for microbialDNA extraction comparative study with colonies lsquopickedrsquo fromthree different bacteriological mediumrdquo Journal of Pure andApplied Microbiology vol 9 no 3 pp 2373ndash2378 2015
[37] R Lan and P R Reeves ldquoEscherichia coli in disguise molecularorigins of Shigellardquo Microbes and Infection vol 4 no 11 pp1125ndash1132 2002
[38] D J Leininger J R Roberson and F Elvinger ldquoUse ofeosin methylene blue agar to differentiate Escherichia coli fromother gram-negative mastitis pathogensrdquo Journal of VeterinaryDiagnostic Investigation vol 13 no 3 pp 273ndash275 2001
[39] U Dobrindt F Agerer K Michaelis et al ldquoAnalysis of genomeplasticity in pathogenic and commensal Escherichia coli isolatesby use of DNA arraysrdquo Journal of Bacteriology vol 185 no 6pp 1831ndash1840 2003
[40] V Savolainen R S Cowan A P Vogler G K Roderickand R Lane ldquoTowards writing the encyclopaedia of life anintroduction to DNA barcodingrdquo Philosophical Transactions ofthe Royal Society B Biological Sciences vol 360 no 1462 pp1805ndash1811 2005
[41] T A Isenbarger C E Carr S S Johnson et al ldquoThe mostconserved genome segments for life detection on earth andother planetsrdquo Origins of Life and Evolution of Biospheres vol38 no 6 pp 517ndash533 2008
[42] S R Santos and H Ochman ldquoIdentification and phylogeneticsorting of bacterial lineages with universally conserved genesand proteinsrdquoEnvironmentalMicrobiology vol 6 no 7 pp 754ndash759 2004
[43] R Rastogi M Wu I DasGupta and G E Fox ldquoVisualizationof ribosomal RNA operon copy number distributionrdquo BMCMicrobiology vol 9 no 1 article 208 2009
[44] J A Klappenbach J M Dunbar and T M Schmidt ldquorRNAoperon copy number reflects ecological strategies of bacteriardquoApplied and Environmental Microbiology vol 66 no 4 pp1328ndash1333 2000
[45] J A Klappenbach P R Saxman J R Cole and T M SchmidtldquoRrndb the ribosomal RNA operon copy number databaserdquoNucleic Acids Research vol 29 no 1 pp 181ndash184 2001
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom

Journal of Pathogens 9
Type I(28 459)
Type IV(28 207 249 and 459)
Hypothesis
Type III(28 249 and 459)
+ minus
+
Primordial
NonpathogenicA gt B1 gt B2
type I gt IV gt III
PathogenicB2 gt B1 gt E gt D
type I (IAT)III(P)gt type IVII
Predominant clones
Rarest
+minus
Type II(28 207 and 459)
Restriction profilesand
clonesPhylogroups and
clones
2∘ clonesintermediate
(a) (b)
Figure 5 (a) Hypothesis of clonality in E coli strains based on 23S rrn restriction profiling The primordial soup of E coli microbes wouldhave been comprised of two clonal groups types I and IV coexisting with each other but predominated by type I strains Over the courseof evolution on accumulating mutation as a means for variation type I or type IV would have given rise to type III by addition (former) ordeletion (latter) of recognition sequence The second scenario could be a rare event because accumulation of two mutations in a conservedregion would be difficult Type II being the rarest would have been evolved in similar basis to type III (b) Hypothesis of linking phylogroupsHae III profiles and clonal types in E coli This diagram illustrates the clonal groups of commensal E coli strains having both A phylogroups(dominant) and B1 phylogroups each being type I profiles Pathogenic counterparts showed dominancy in type III restriction pattern followedby type I pattern With respect to phylogroups pathovars were predominant in B2 and B1 followed by D and E groups
from housekeeping to repair are employed for characterizingbacteria of which rRNA is referred to as the gold stretch[41 42] Ribosomal segments organized as operon clusters of16S 23S and 5S rDNA and these assemblages are distributedfrom single to multiple copies across the bacterial genophore[43] The occurrence of redundancy in maintaining differentcopies of these genes is not associated with virulence but anin-built ecological strategy that contributes to the particularbacterium with a competitive edge and better chance ofsurvival [44] A futuristic approach of coupling the techniqueand results with antibiotic resistance pattern colony char-acteristics and genomic profiles of bacterial strains of bothcommensal and pathogenic nature can help in monitoringand managing infections in a better manner
4 Conclusion
The causatum of postgenomic era genomics and proteomicshas enhanced this progress by improvising the techniquesused for diagnostics The strategy of ARDRA utilized inthis study was rapid and a cost-effective approach to dif-ferentiate E coli from its colony morphovars along withgiving a predicament of nature of origin or clonal groupsThe mutations reported in this work for detection andestablishing clonality namely T874A (Bfa I) G2161A andG2205A (Hae III) have not been mentioned in any researchpaper or within the Ribosomal RNA Mutation Database
[45] making this finding aboriginal This basic methodologycan be adapted for detection of other bacterial organismsby targeting SNPs linked to recognition sequences therebyvalidating rrn sequences as marker for diagnostics A cumu-lative approach of detection of unique restriction pattern of atargeted gene and its cross-reference across bacterial familiescan circumvent changes of false positives
Disclosure
This work was presented (poster) in part at the 5th Interna-tional Barcoding of Life Kunming China 2013
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
Acknowledgments
The authors acknowledge and express their gratitude to Pro-fessor D A Bhiwgade (Ex-Director) and Professor DebjaniDasgupta Director School of Biotechnology amp Bioinformat-ics D Y Patil University as well as the management for theirsupport and for providing the infrastructure to carry out thecurrent work
10 Journal of Pathogens
References
[1] M Richter and R Rossello-Mora ldquoShifting the genomic goldstandard for the prokaryotic species definitionrdquo Proceedings ofthe National Academy of Sciences of the United States of Americavol 106 no 45 pp 19126ndash19131 2009
[2] A F Auch M von Jan H-P Klenk and M Goker ldquoDigitalDNA-DNA hybridization for microbial species delineation bymeans of genome-to-genome sequence comparisonrdquo Standardsin Genomic Sciences vol 2 no 1 pp 117ndash134 2010
[3] D H Martin M Nsuami J Schachter et al ldquoUse of multiplenucleic acid amplification tests to define the infected-patientlsquogold standardrsquo in clinical trials of new diagnostic tests forChlamydia trachomatis infectionsrdquo Journal of Clinical Microbi-ology vol 42 no 10 pp 4749ndash4758 2004
[4] E A Mothershed and A M Whitney ldquoNucleic acid-basedmethods for the detection of bacterial pathogens presentand future considerations for the clinical laboratoryrdquo ClinicaChimica Acta vol 363 no 1-2 pp 206ndash220 2006
[5] J E Clarridge III ldquoImpact of 16S rRNA gene sequence analysisfor identification of bacteria on clinical microbiology andinfectious diseasesrdquo Clinical Microbiology Reviews vol 17 no4 pp 840ndash862 2004
[6] O Mizrahi-Man E R Davenport and Y Gilad ldquoTaxonomicclassification of bacterial 16S rRNA genes using short sequenc-ing reads evaluation of effective study designsrdquo PLoS ONE vol8 no 1 Article ID e53608 2013
[7] D E Lebonah A Dileep K Chandrasekhar S Sreevani BSreedevi and J Pramoda Kumari ldquoDNA barcoding on bacteriaa reviewrdquo Advances in Biology vol 2014 Article ID 541787 9pages 2014
[8] J Rajendhran and P Gunasekaran ldquoMicrobial phylogeny anddiversity small subunit ribosomal RNA sequence analysis andbeyondrdquo Microbiological Research vol 166 no 2 pp 99ndash1102011
[9] R Srinivasan U Karaoz M Volegova et al ldquoUse of 16S rRNAgene for identification of a broad range of clinically relevantbacterial pathogensrdquo PLoS ONE vol 10 no 2 Article IDe0117617 2015
[10] S Chakravorty D Helb M Burday N Connell and D AllandldquoA detailed analysis of 16S ribosomal RNA gene segments forthe diagnosis of pathogenic bacteriardquo Journal of MicrobiologicalMethods vol 69 no 2 pp 330ndash339 2007
[11] C R Woese ldquoReview bacterial evolutionrdquo MicrobiologicalReviews vol 51 pp 221ndash271 1987
[12] M Touchon C Hoede O Tenaillon et al ldquoOrganised genomedynamics in the Escherichia coli species results in highly diverseadaptive pathsrdquo PLoS Genetics vol 5 no 1 Article ID e10003442009
[13] J M Janda and S L Abbott ldquo16S rRNA gene sequencingfor bacterial identification in the diagnostic laboratory plusesperils and pitfallsrdquo Journal of Clinical Microbiology vol 45 no9 pp 2761ndash2764 2007
[14] J-J Lu C-L Perng S-Y Lee and C-CWan ldquoUse of PCRwithuniversal primers and restriction endonuclease digestions fordetection and identification of common bacterial pathogens incerebrospinal fluidrdquo Journal of Clinical Microbiology vol 38 no6 pp 2076ndash2080 2000
[15] N M Guentzel ldquoEscherichia Klebsiella Enterobacter SerratiaCitrobacter andProteusrdquo inMedicalMicrobiology S Baron Edchapter 26 University of Texas Medical Branch at GalvestonGalveston Tex USA 1996
[16] P Behzadip E Behzadi H Yazdanbod R Aghapour A MCheshmeh and S D Omran ldquoA survey on urinary tract infec-tions associated with the three most common uropathogenicbacteriardquoMaedica (Buchar) vol 5 no 2 pp 111ndash115 2010
[17] B CMetri P Jyothi andBV Peerapur ldquoAntibiotic resistance inCitrobacter spp isolated from urinary tract infectionrdquo UrologyAnnals vol 5 no 4 pp 312ndash313 2013
[18] N Taneja S S Chatterjee M Singh S Singh and M SharmaldquoPediatric urinary tract infections in a tertiary care center fromnorth Indiardquo Indian Journal of Medical Research vol 131 no 1pp 101ndash105 2010
[19] J B Kaper J P Nataro and H L T Mobley ldquoPathogenicEscherichia colirdquo Nature Reviews Microbiology vol 2 no 2 pp123ndash140 2004
[20] E J de Muinck K Lagesen J E Afset et al ldquoComparisonsof infant Escherichia coli isolates link genomic profiles withadaptation to the ecological nicherdquo BMC Genomics vol 14 no1 article 81 2013
[21] D A Rasko M J Rosovitz G S A Myers et al ldquoThepangenome structure of Escherichia coli comparative genomicanalysis of E coli commensal and pathogenic isolatesrdquo Journalof Bacteriology vol 190 no 20 pp 6881ndash6893 2008
[22] C T Archer J F Kim H Jeong et al ldquoThe genome sequenceof E coli W (ATCC 9637) comparative genome analysis andan improved genome-scale reconstruction of E colirdquo BMCGenomics vol 12 article no 9 2011
[23] PKuhnert J Nicolet and J Frey ldquoRapid and accurate identifica-tion of Escherichia coliK-12 strainsrdquoApplied and EnvironmentalMicrobiology vol 61 no 11 pp 4135ndash4139 1995
[24] A P Bauer S M Dieckmann W Ludwig and K-H SchleiferldquoRapid identification of Escherichia coli safety and laboratorystrain lineages based on multiplex-PCRrdquo FEMS MicrobiologyLetters vol 269 no 1 pp 36ndash40 2007
[25] B J Bachmann ldquoPedigrees of some mutant strains ofEscherichia coli K-12rdquo Bacteriological Reviews vol 36 no 4 pp525ndash557 1972
[26] P Daegelen F W Studier R E Lenski S Cure and J F KimldquoTracing ancestors and relatives of Escherichia coli B and thederivation of B strains REL606 and BL21(DE3)rdquo Journal ofMolecular Biology vol 394 no 4 pp 634ndash643 2009
[27] F W Studier P Daegelen R E Lenski S Maslov and J F KimldquoUnderstanding the differences between genome sequences ofEscherichia coli B strains REL606 and BL21(DE3) and compar-ison of the E coli B and K-12 genomesrdquo Journal of MolecularBiology vol 394 no 4 pp 653ndash680 2009
[28] R K Selander and B R Levin ldquoGenetic diversity and structurein Escherichia coli populationsrdquo Science vol 210 no 4469 pp545ndash547 1980
[29] D M Gordon O Clermont H Tolley and E DenamurldquoAssigning Escherichia coli strains to phylogenetic groupsmulti-locus sequence typing versus the PCR triplex methodrdquoEnvironmental Microbiology vol 10 no 10 pp 2484ndash24962008
[30] P Desjardins B Picard B Kaltenbock J Elion and EDenamur ldquoSex in Escherichia coli does not disrupt the clonalstructure of the population evidence from random amplifiedpolymorphic DNA and restriction-fragment-length polymor-phismrdquo Journal of Molecular Evolution vol 41 no 4 pp 440ndash448 1995
Journal of Pathogens 11
[31] N D Jasuja R Saxena S Chandra and S C Joshi ldquoIsolationand identification of microorganism from polyhouse agricul-ture soil of RajasthanrdquoAfrican Journal ofMicrobiology Researchvol 7 no 41 pp 4886ndash4891 2013
[32] V Hemraj S Diksha and G Avneet ldquoA review of commonlyused biochemical test for bacteriardquo Innovare Journal of LifeSciences vol 1 pp 1ndash7 2013
[33] J D Thompson D G Higgins and T J Gibson ldquoCLUSTALWimproving the sensitivity of progressive multiple sequencealignment through sequence weighting position-specific gappenalties and weight matrix choicerdquoNucleic Acids Research vol22 no 22 pp 4673ndash4680 1994
[34] T Vincze J Posfai and R J Roberts ldquoNEBcutter a program tocleave DNA with restriction enzymesrdquo Nucleic Acids Researchvol 31 no 13 pp 3688ndash3691 2003
[35] J Ye G Coulouris I Zaretskaya I Cutcutache S Rozen andT Madden ldquoPrimer-BLAST a tool to design target-specificprimers for polymerase chain reactionrdquo BMC Bioinformaticsvol 13 article 134 2012
[36] J R Parvathi and S Singh ldquoFacile approaches for microbialDNA extraction comparative study with colonies lsquopickedrsquo fromthree different bacteriological mediumrdquo Journal of Pure andApplied Microbiology vol 9 no 3 pp 2373ndash2378 2015
[37] R Lan and P R Reeves ldquoEscherichia coli in disguise molecularorigins of Shigellardquo Microbes and Infection vol 4 no 11 pp1125ndash1132 2002
[38] D J Leininger J R Roberson and F Elvinger ldquoUse ofeosin methylene blue agar to differentiate Escherichia coli fromother gram-negative mastitis pathogensrdquo Journal of VeterinaryDiagnostic Investigation vol 13 no 3 pp 273ndash275 2001
[39] U Dobrindt F Agerer K Michaelis et al ldquoAnalysis of genomeplasticity in pathogenic and commensal Escherichia coli isolatesby use of DNA arraysrdquo Journal of Bacteriology vol 185 no 6pp 1831ndash1840 2003
[40] V Savolainen R S Cowan A P Vogler G K Roderickand R Lane ldquoTowards writing the encyclopaedia of life anintroduction to DNA barcodingrdquo Philosophical Transactions ofthe Royal Society B Biological Sciences vol 360 no 1462 pp1805ndash1811 2005
[41] T A Isenbarger C E Carr S S Johnson et al ldquoThe mostconserved genome segments for life detection on earth andother planetsrdquo Origins of Life and Evolution of Biospheres vol38 no 6 pp 517ndash533 2008
[42] S R Santos and H Ochman ldquoIdentification and phylogeneticsorting of bacterial lineages with universally conserved genesand proteinsrdquoEnvironmentalMicrobiology vol 6 no 7 pp 754ndash759 2004
[43] R Rastogi M Wu I DasGupta and G E Fox ldquoVisualizationof ribosomal RNA operon copy number distributionrdquo BMCMicrobiology vol 9 no 1 article 208 2009
[44] J A Klappenbach J M Dunbar and T M Schmidt ldquorRNAoperon copy number reflects ecological strategies of bacteriardquoApplied and Environmental Microbiology vol 66 no 4 pp1328ndash1333 2000
[45] J A Klappenbach P R Saxman J R Cole and T M SchmidtldquoRrndb the ribosomal RNA operon copy number databaserdquoNucleic Acids Research vol 29 no 1 pp 181ndash184 2001
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom

10 Journal of Pathogens
References
[1] M Richter and R Rossello-Mora ldquoShifting the genomic goldstandard for the prokaryotic species definitionrdquo Proceedings ofthe National Academy of Sciences of the United States of Americavol 106 no 45 pp 19126ndash19131 2009
[2] A F Auch M von Jan H-P Klenk and M Goker ldquoDigitalDNA-DNA hybridization for microbial species delineation bymeans of genome-to-genome sequence comparisonrdquo Standardsin Genomic Sciences vol 2 no 1 pp 117ndash134 2010
[3] D H Martin M Nsuami J Schachter et al ldquoUse of multiplenucleic acid amplification tests to define the infected-patientlsquogold standardrsquo in clinical trials of new diagnostic tests forChlamydia trachomatis infectionsrdquo Journal of Clinical Microbi-ology vol 42 no 10 pp 4749ndash4758 2004
[4] E A Mothershed and A M Whitney ldquoNucleic acid-basedmethods for the detection of bacterial pathogens presentand future considerations for the clinical laboratoryrdquo ClinicaChimica Acta vol 363 no 1-2 pp 206ndash220 2006
[5] J E Clarridge III ldquoImpact of 16S rRNA gene sequence analysisfor identification of bacteria on clinical microbiology andinfectious diseasesrdquo Clinical Microbiology Reviews vol 17 no4 pp 840ndash862 2004
[6] O Mizrahi-Man E R Davenport and Y Gilad ldquoTaxonomicclassification of bacterial 16S rRNA genes using short sequenc-ing reads evaluation of effective study designsrdquo PLoS ONE vol8 no 1 Article ID e53608 2013
[7] D E Lebonah A Dileep K Chandrasekhar S Sreevani BSreedevi and J Pramoda Kumari ldquoDNA barcoding on bacteriaa reviewrdquo Advances in Biology vol 2014 Article ID 541787 9pages 2014
[8] J Rajendhran and P Gunasekaran ldquoMicrobial phylogeny anddiversity small subunit ribosomal RNA sequence analysis andbeyondrdquo Microbiological Research vol 166 no 2 pp 99ndash1102011
[9] R Srinivasan U Karaoz M Volegova et al ldquoUse of 16S rRNAgene for identification of a broad range of clinically relevantbacterial pathogensrdquo PLoS ONE vol 10 no 2 Article IDe0117617 2015
[10] S Chakravorty D Helb M Burday N Connell and D AllandldquoA detailed analysis of 16S ribosomal RNA gene segments forthe diagnosis of pathogenic bacteriardquo Journal of MicrobiologicalMethods vol 69 no 2 pp 330ndash339 2007
[11] C R Woese ldquoReview bacterial evolutionrdquo MicrobiologicalReviews vol 51 pp 221ndash271 1987
[12] M Touchon C Hoede O Tenaillon et al ldquoOrganised genomedynamics in the Escherichia coli species results in highly diverseadaptive pathsrdquo PLoS Genetics vol 5 no 1 Article ID e10003442009
[13] J M Janda and S L Abbott ldquo16S rRNA gene sequencingfor bacterial identification in the diagnostic laboratory plusesperils and pitfallsrdquo Journal of Clinical Microbiology vol 45 no9 pp 2761ndash2764 2007
[14] J-J Lu C-L Perng S-Y Lee and C-CWan ldquoUse of PCRwithuniversal primers and restriction endonuclease digestions fordetection and identification of common bacterial pathogens incerebrospinal fluidrdquo Journal of Clinical Microbiology vol 38 no6 pp 2076ndash2080 2000
[15] N M Guentzel ldquoEscherichia Klebsiella Enterobacter SerratiaCitrobacter andProteusrdquo inMedicalMicrobiology S Baron Edchapter 26 University of Texas Medical Branch at GalvestonGalveston Tex USA 1996
[16] P Behzadip E Behzadi H Yazdanbod R Aghapour A MCheshmeh and S D Omran ldquoA survey on urinary tract infec-tions associated with the three most common uropathogenicbacteriardquoMaedica (Buchar) vol 5 no 2 pp 111ndash115 2010
[17] B CMetri P Jyothi andBV Peerapur ldquoAntibiotic resistance inCitrobacter spp isolated from urinary tract infectionrdquo UrologyAnnals vol 5 no 4 pp 312ndash313 2013
[18] N Taneja S S Chatterjee M Singh S Singh and M SharmaldquoPediatric urinary tract infections in a tertiary care center fromnorth Indiardquo Indian Journal of Medical Research vol 131 no 1pp 101ndash105 2010
[19] J B Kaper J P Nataro and H L T Mobley ldquoPathogenicEscherichia colirdquo Nature Reviews Microbiology vol 2 no 2 pp123ndash140 2004
[20] E J de Muinck K Lagesen J E Afset et al ldquoComparisonsof infant Escherichia coli isolates link genomic profiles withadaptation to the ecological nicherdquo BMC Genomics vol 14 no1 article 81 2013
[21] D A Rasko M J Rosovitz G S A Myers et al ldquoThepangenome structure of Escherichia coli comparative genomicanalysis of E coli commensal and pathogenic isolatesrdquo Journalof Bacteriology vol 190 no 20 pp 6881ndash6893 2008
[22] C T Archer J F Kim H Jeong et al ldquoThe genome sequenceof E coli W (ATCC 9637) comparative genome analysis andan improved genome-scale reconstruction of E colirdquo BMCGenomics vol 12 article no 9 2011
[23] PKuhnert J Nicolet and J Frey ldquoRapid and accurate identifica-tion of Escherichia coliK-12 strainsrdquoApplied and EnvironmentalMicrobiology vol 61 no 11 pp 4135ndash4139 1995
[24] A P Bauer S M Dieckmann W Ludwig and K-H SchleiferldquoRapid identification of Escherichia coli safety and laboratorystrain lineages based on multiplex-PCRrdquo FEMS MicrobiologyLetters vol 269 no 1 pp 36ndash40 2007
[25] B J Bachmann ldquoPedigrees of some mutant strains ofEscherichia coli K-12rdquo Bacteriological Reviews vol 36 no 4 pp525ndash557 1972
[26] P Daegelen F W Studier R E Lenski S Cure and J F KimldquoTracing ancestors and relatives of Escherichia coli B and thederivation of B strains REL606 and BL21(DE3)rdquo Journal ofMolecular Biology vol 394 no 4 pp 634ndash643 2009
[27] F W Studier P Daegelen R E Lenski S Maslov and J F KimldquoUnderstanding the differences between genome sequences ofEscherichia coli B strains REL606 and BL21(DE3) and compar-ison of the E coli B and K-12 genomesrdquo Journal of MolecularBiology vol 394 no 4 pp 653ndash680 2009
[28] R K Selander and B R Levin ldquoGenetic diversity and structurein Escherichia coli populationsrdquo Science vol 210 no 4469 pp545ndash547 1980
[29] D M Gordon O Clermont H Tolley and E DenamurldquoAssigning Escherichia coli strains to phylogenetic groupsmulti-locus sequence typing versus the PCR triplex methodrdquoEnvironmental Microbiology vol 10 no 10 pp 2484ndash24962008
[30] P Desjardins B Picard B Kaltenbock J Elion and EDenamur ldquoSex in Escherichia coli does not disrupt the clonalstructure of the population evidence from random amplifiedpolymorphic DNA and restriction-fragment-length polymor-phismrdquo Journal of Molecular Evolution vol 41 no 4 pp 440ndash448 1995
Journal of Pathogens 11
[31] N D Jasuja R Saxena S Chandra and S C Joshi ldquoIsolationand identification of microorganism from polyhouse agricul-ture soil of RajasthanrdquoAfrican Journal ofMicrobiology Researchvol 7 no 41 pp 4886ndash4891 2013
[32] V Hemraj S Diksha and G Avneet ldquoA review of commonlyused biochemical test for bacteriardquo Innovare Journal of LifeSciences vol 1 pp 1ndash7 2013
[33] J D Thompson D G Higgins and T J Gibson ldquoCLUSTALWimproving the sensitivity of progressive multiple sequencealignment through sequence weighting position-specific gappenalties and weight matrix choicerdquoNucleic Acids Research vol22 no 22 pp 4673ndash4680 1994
[34] T Vincze J Posfai and R J Roberts ldquoNEBcutter a program tocleave DNA with restriction enzymesrdquo Nucleic Acids Researchvol 31 no 13 pp 3688ndash3691 2003
[35] J Ye G Coulouris I Zaretskaya I Cutcutache S Rozen andT Madden ldquoPrimer-BLAST a tool to design target-specificprimers for polymerase chain reactionrdquo BMC Bioinformaticsvol 13 article 134 2012
[36] J R Parvathi and S Singh ldquoFacile approaches for microbialDNA extraction comparative study with colonies lsquopickedrsquo fromthree different bacteriological mediumrdquo Journal of Pure andApplied Microbiology vol 9 no 3 pp 2373ndash2378 2015
[37] R Lan and P R Reeves ldquoEscherichia coli in disguise molecularorigins of Shigellardquo Microbes and Infection vol 4 no 11 pp1125ndash1132 2002
[38] D J Leininger J R Roberson and F Elvinger ldquoUse ofeosin methylene blue agar to differentiate Escherichia coli fromother gram-negative mastitis pathogensrdquo Journal of VeterinaryDiagnostic Investigation vol 13 no 3 pp 273ndash275 2001
[39] U Dobrindt F Agerer K Michaelis et al ldquoAnalysis of genomeplasticity in pathogenic and commensal Escherichia coli isolatesby use of DNA arraysrdquo Journal of Bacteriology vol 185 no 6pp 1831ndash1840 2003
[40] V Savolainen R S Cowan A P Vogler G K Roderickand R Lane ldquoTowards writing the encyclopaedia of life anintroduction to DNA barcodingrdquo Philosophical Transactions ofthe Royal Society B Biological Sciences vol 360 no 1462 pp1805ndash1811 2005
[41] T A Isenbarger C E Carr S S Johnson et al ldquoThe mostconserved genome segments for life detection on earth andother planetsrdquo Origins of Life and Evolution of Biospheres vol38 no 6 pp 517ndash533 2008
[42] S R Santos and H Ochman ldquoIdentification and phylogeneticsorting of bacterial lineages with universally conserved genesand proteinsrdquoEnvironmentalMicrobiology vol 6 no 7 pp 754ndash759 2004
[43] R Rastogi M Wu I DasGupta and G E Fox ldquoVisualizationof ribosomal RNA operon copy number distributionrdquo BMCMicrobiology vol 9 no 1 article 208 2009
[44] J A Klappenbach J M Dunbar and T M Schmidt ldquorRNAoperon copy number reflects ecological strategies of bacteriardquoApplied and Environmental Microbiology vol 66 no 4 pp1328ndash1333 2000
[45] J A Klappenbach P R Saxman J R Cole and T M SchmidtldquoRrndb the ribosomal RNA operon copy number databaserdquoNucleic Acids Research vol 29 no 1 pp 181ndash184 2001
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom

Journal of Pathogens 11
[31] N D Jasuja R Saxena S Chandra and S C Joshi ldquoIsolationand identification of microorganism from polyhouse agricul-ture soil of RajasthanrdquoAfrican Journal ofMicrobiology Researchvol 7 no 41 pp 4886ndash4891 2013
[32] V Hemraj S Diksha and G Avneet ldquoA review of commonlyused biochemical test for bacteriardquo Innovare Journal of LifeSciences vol 1 pp 1ndash7 2013
[33] J D Thompson D G Higgins and T J Gibson ldquoCLUSTALWimproving the sensitivity of progressive multiple sequencealignment through sequence weighting position-specific gappenalties and weight matrix choicerdquoNucleic Acids Research vol22 no 22 pp 4673ndash4680 1994
[34] T Vincze J Posfai and R J Roberts ldquoNEBcutter a program tocleave DNA with restriction enzymesrdquo Nucleic Acids Researchvol 31 no 13 pp 3688ndash3691 2003
[35] J Ye G Coulouris I Zaretskaya I Cutcutache S Rozen andT Madden ldquoPrimer-BLAST a tool to design target-specificprimers for polymerase chain reactionrdquo BMC Bioinformaticsvol 13 article 134 2012
[36] J R Parvathi and S Singh ldquoFacile approaches for microbialDNA extraction comparative study with colonies lsquopickedrsquo fromthree different bacteriological mediumrdquo Journal of Pure andApplied Microbiology vol 9 no 3 pp 2373ndash2378 2015
[37] R Lan and P R Reeves ldquoEscherichia coli in disguise molecularorigins of Shigellardquo Microbes and Infection vol 4 no 11 pp1125ndash1132 2002
[38] D J Leininger J R Roberson and F Elvinger ldquoUse ofeosin methylene blue agar to differentiate Escherichia coli fromother gram-negative mastitis pathogensrdquo Journal of VeterinaryDiagnostic Investigation vol 13 no 3 pp 273ndash275 2001
[39] U Dobrindt F Agerer K Michaelis et al ldquoAnalysis of genomeplasticity in pathogenic and commensal Escherichia coli isolatesby use of DNA arraysrdquo Journal of Bacteriology vol 185 no 6pp 1831ndash1840 2003
[40] V Savolainen R S Cowan A P Vogler G K Roderickand R Lane ldquoTowards writing the encyclopaedia of life anintroduction to DNA barcodingrdquo Philosophical Transactions ofthe Royal Society B Biological Sciences vol 360 no 1462 pp1805ndash1811 2005
[41] T A Isenbarger C E Carr S S Johnson et al ldquoThe mostconserved genome segments for life detection on earth andother planetsrdquo Origins of Life and Evolution of Biospheres vol38 no 6 pp 517ndash533 2008
[42] S R Santos and H Ochman ldquoIdentification and phylogeneticsorting of bacterial lineages with universally conserved genesand proteinsrdquoEnvironmentalMicrobiology vol 6 no 7 pp 754ndash759 2004
[43] R Rastogi M Wu I DasGupta and G E Fox ldquoVisualizationof ribosomal RNA operon copy number distributionrdquo BMCMicrobiology vol 9 no 1 article 208 2009
[44] J A Klappenbach J M Dunbar and T M Schmidt ldquorRNAoperon copy number reflects ecological strategies of bacteriardquoApplied and Environmental Microbiology vol 66 no 4 pp1328ndash1333 2000
[45] J A Klappenbach P R Saxman J R Cole and T M SchmidtldquoRrndb the ribosomal RNA operon copy number databaserdquoNucleic Acids Research vol 29 no 1 pp 181ndash184 2001
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom