results - shodhgangashodhganga.inflibnet.ac.in/bitstream/10603/62303/7/08_results.pdf · results...
TRANSCRIPT

RESULTS
Hamelia patens Jacq. belong to the family Rubiaceae was collected from
different parts of Kerala. Plant collected from Parassala has remarkable
morphological differences (posses dark green leaves and small flowers) compared to
all other accessions (posses light green leaves and large flowers). In the present study
Acc. No. 1 (from Kariavattom, Fig. 1) and Acc. No. 3 (from Parassala, Fig. 2) were
selected, since they were morphologically dissimilar (Fig. 3).
4.1 Phenology
The flowering phenology showed uniformity throughout the year. The
inflorescence is a terminal cyme of about three inches long bearing flowers in a
helicoid cyme. The flower open around 1-2 am and last for one day. The petals,
stamens and stigma drop off while the calyx is persistent with ovary. Flowers are
reddish orange in colour, pedicellate, bisexual, actinomorphic, complete, regular,
pentamerous, and epigynous. Sepals are five in number and gamosepalous. Corolla
consists of five gamopetalous petals with cylindrical corolla tube. Androecium has
five stamens, in epipetalous condition and anther dehisce around 11-12 pm. Style is
long with linear shaped stigma, in which sticky excudates were seen from very early
stages onwards. Ovary is pentacarpellary, syncarpous, pentalocular with numerous
ovules on axile placentation. Mature fruits (drup) persist for 2-3 weeks with sterile
seeds (Figs. 4-6) and then drop off.
4.2 Structure of reproductive parts
4.2.1 Anther
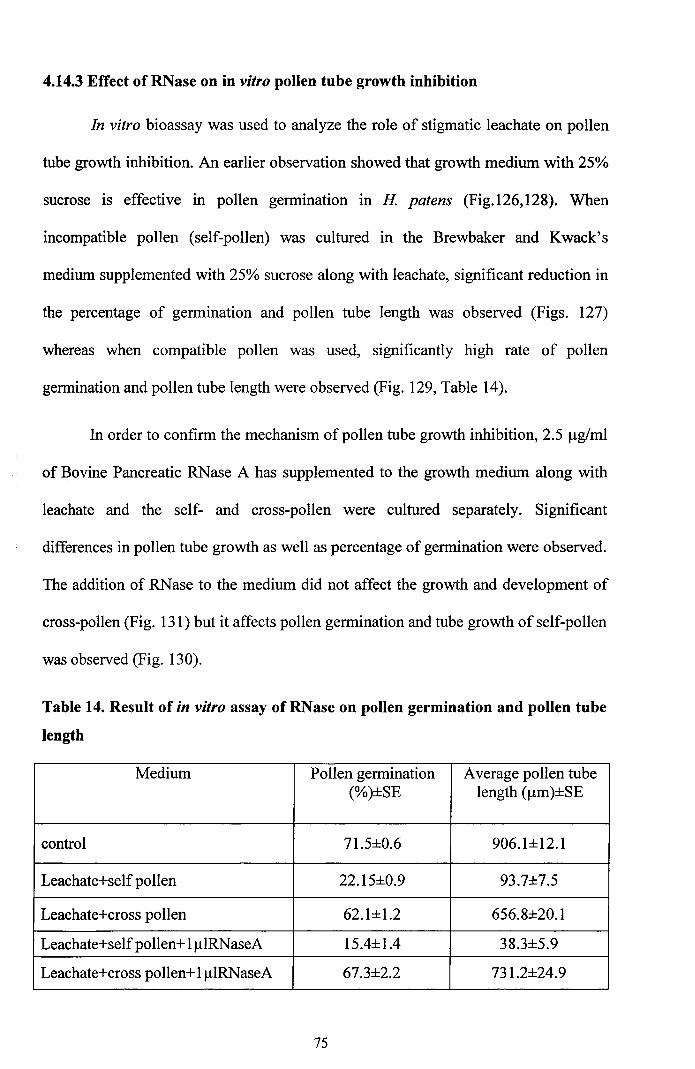
Fully matured anther is bithecous and tetrasporangiate (Fig. 7).
Microsporangium consists of a mass of haploid pollen enclosed within the

microsporangial wall. Outermost layer of the anther is epidermis, followed by a single
layer of endothecium, two or three middle layers and a single layered tapetum
(Figs. 8, 9). When pollen matures, the tapetum disintegrates and the dehiscence of
anther takes place longitudinally at the time ofanthesis.
4.2.2 Pistil
Stigma is wet, papillate and linear in shape (Fig. 10). Unicellular and rod
shaped papillae are arranged uniformly throughout the surface of stigma (Fig. 11).
The cryotome sections of stigma revealed two zones, an outer zone and inner zone.
The outer zone consists of epidermis and 1-3 layers of laterally extended cells
constituting the secretory tissue. The inner zone consists of parenchymatous ground
tissue with central transmitting zone and vascular strands on either side (Fig. 12).
SEM studies revealed that the linear shaped stigma is covered with unicellular
and rod shaped papillae. From the very early stage onwards to the stage of maturity,
the morphology of stigma surface was found to be similar (Figs. 13-20). The surface
of stigma is irregular and posses copious exudate from the very early stage of the bud
which accumulates all around the stigmatic surface. Below the papillar cells, the
subepidermal cells of the stigma converge into a short solid style made up of a central
core of transmitting tissue surrounded by parenchymatous cortex and vascular tissues.
SEM showed numerous sterile ovules on axile placentation (Figs. 21-23).
Ovary wall consists of a single layered parenchymatous epidermis. Vascular tissues
are clearly visible intermittently.
57

4.3 Cytology
4.3.1 Meiosis
Meiotic chromosome studies showed the presence of 12 bivalents at
metaphase (Fig. 24) and subsequent stages ofmeiosis were normal. The chromosomes
were small in size.
4.3.2 Pollen cytology
Pollen grains were kept in lactopropionic orcein and observed after 24 hrs.
The protoplast completely extruded out of the pollen exine and the presence of two
intensely stained nuclei was distinctly seen (Fig. 25).
4.4 Pollen viability studies
4.4.1 Fluorochromatic reaction test.
When pollen grains were mounted in FDA solution and observed under UV
light, about 90% of pollen grains showed a bright green or yellowish green
fluorescence which revealed their viability status (Fig. 26).
4.4.2 In vitro germination
In order to assess the percentage of viable pollen grains, in vitro test was
conducted. Pollen germination commenced after 3 hrs in the medium supplemented
with 5% sucrose and attained a maximum germination of 72.5% was observed the
medium with 25% sucrose (Fig. 27, Table 4, Fig. I). Above 25%, there was a gradual
decrease in the rate of pollen germination, the content of the pollen cytoplasm
extruded out through the germ pore.
58
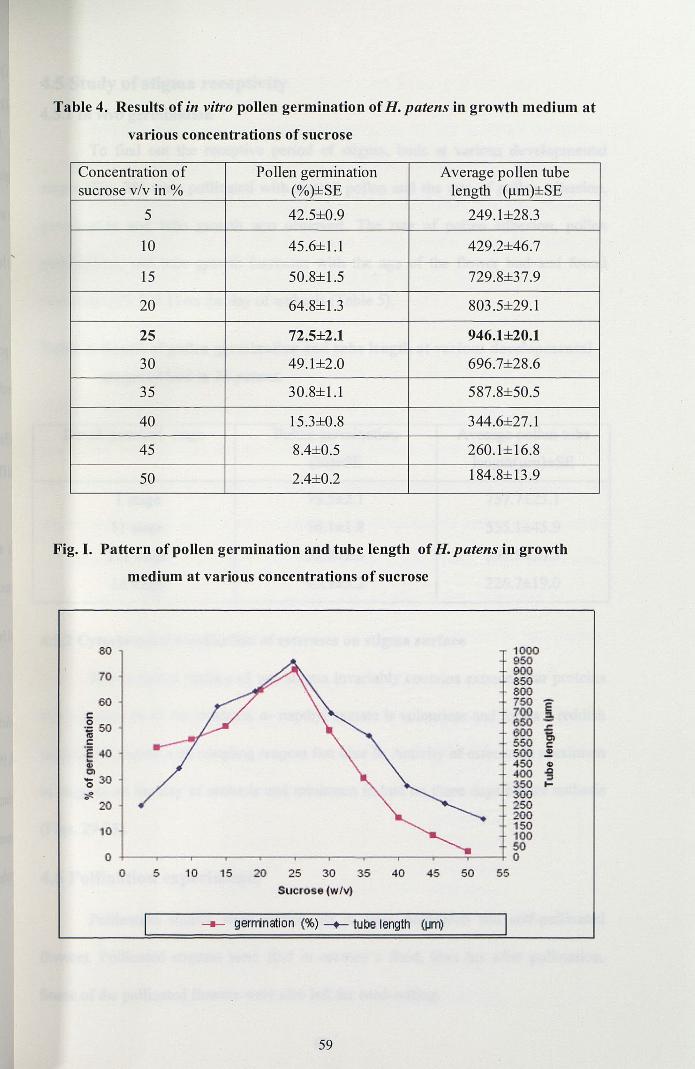
Table 4. Results of in vitro pollen germination ofH. patens in growth medium at
various concentrations of sucrose
Concentration of Pollen germination Average pollen tubesucrose v/v in % (%)±SE length (~m)±SE
5 42.5±0.9 249.1±28.3
10 45.6±1.1 429.2±46.7
15 50.8±1.5 729.8±37.9
20 64.8±13 803.5±29.1
25 72.5±2.1 946.1±20.1
30 49.1±2.0 696.7±28.6
35 30.8±1.1 587.8±50.5
40 15.3±0.8 344.6±27.1
45 8.4±0.5 260.1±16.8
50 2.4±0.2 184.8±13.9
Fig. I. Pattern of pollen germination and tube length ofH. patens in growth
medium at various concentrations of sucrose
SO
70
60l:.g 50IIIC
's 40~~
Q 30~
20
10
00 5 10 15 20 25 30 35 40
Sucrose {w/v)
1000950900850800750 E700 a650WO
,t;"6l550 c:
500 .!450 CI)
400 .til:::I
350 ...3()O250200150100500
45 50 55
-II- gerrrination (%) -+- tube length (IJm)
59
4.5 Study of stigma receptivity
4.5.1 In vivo germination
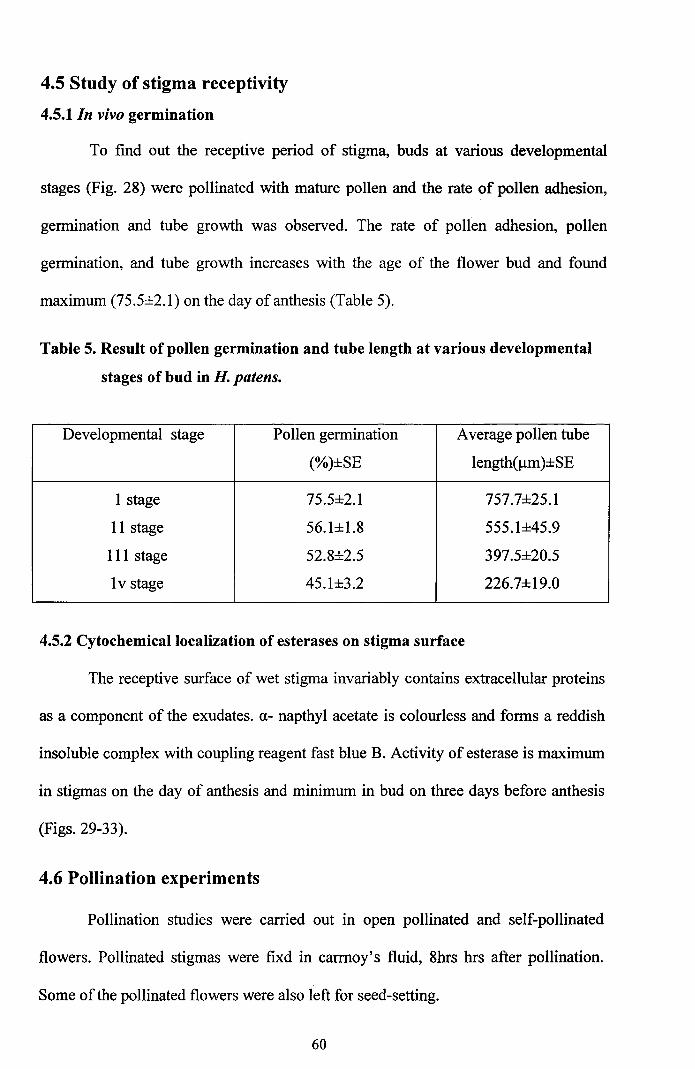
To find out the receptive period of stigma, buds at various developmental
stages (Fig. 28) were pollinated with mature pollen and the rate of pollen adhesion,
germination and tube growth was observed. The rate of pollen adhesion, pollen
germination, and tube growth increases with the age of the flower bud and found
maximum (75.5±2.l) on the day ofanthesis (Table 5).
Table 5. Result of pollen germination and tube length at various developmental
stages of bud in H. patens.
Developmental stage Pollen germination Average pollen tube
(%)±SE length(llm)±SE
I stage 75.5±2.1 757.7±25.1
II stage 56.1±1.8 555.1±45.9
III stage 52.8±2.5 397.5±20.5
Iv stage 45.1±3.2 226.7±19.0
4.5.2 Cytochemical localization of esterases on stigma surface
The receptive surface of wet stigma invariably contains extracellular proteins
as a component of the exudates. 0.- napthyl acetate is colourless and forms a reddish
insoluble complex with coupling reagent fast blue B. Activity of esterase is maximum
in stigmas on the day of anthesis and minimum in bud on three days before anthesis
(Figs. 29-33).
4.6 Pollination experiments
Pollination studies were carried out in open pollinated and self-pollinated
flowers. Pollinated stigmas were fixd in carrnoy's fluid, 8hrs hrs after pollination.
Some of the pollinated flowers were also left for seed-setting.
60
4.6.1 Open pollination
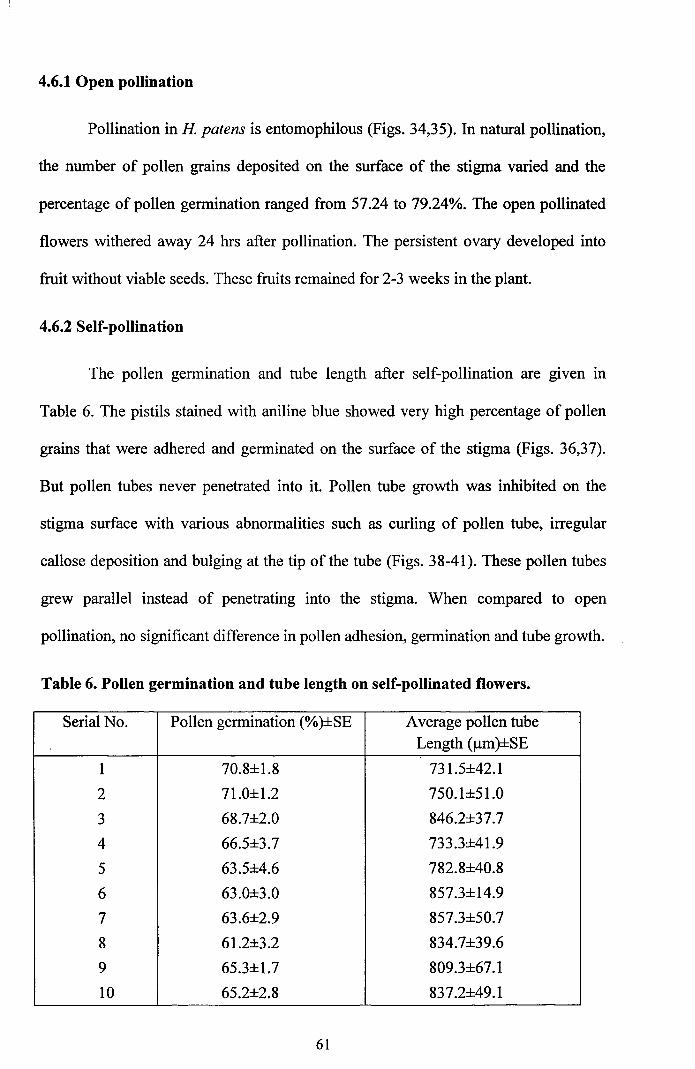
Pollination in H patens is entomophilous (Figs. 34,35). In natural pollination,
the number of pollen grains deposited on the surface of the stigma varied and the
percentage of pollen germination ranged from 57.24 to 79.24%. The open pollinated
flowers withered away 24 hrs after pollination. The persistent ovary developed into
fruit without viable seeds. These fruits remained for 2-3 weeks in the plant.
4.6.2 Self-pollination
The pollen germination and tube length after self-pollination are given in
Table 6. The pistils stained with aniline blue showed very high percentage of pollen
grains that were adhered and germinated on the surface of the stigma (Figs. 36,37).
But pollen tubes never penetrated into it. Pollen tube growth was inhibited on the
stigma surface with various abnormalities such as curling of pollen tube, irregular
callose deposition and bulging at the tip of the tube (Figs. 38-41). These pollen tubes
grew parallel instead of penetrating into the stigma. When compared to open
pollination, no significant difference in pollen adhesion, germination and tube growth.
Table 6. Pollen germination and tube length on self-pollinated flowers.
Serial No. Pollen germination (%)±SE Average pollen tubeLength (j.lm)±SE
1 70.8±1.8 731.5±42.1
2 71.0±1.2 750.1±51.0
3 68.7±2.0 846.2±37.7
4 66.5±3.7 733.3±41.9
5 63.5±4.6 782.8±40.8
6 63.0±3.0 857.3±14.9
7 63.6±2.9 857.3±50.7
8 61.2±3.2 834.7±39.6
9 65.3±1.7 809.3±67.1
10 65.2±2.8 837.2±49.1
61
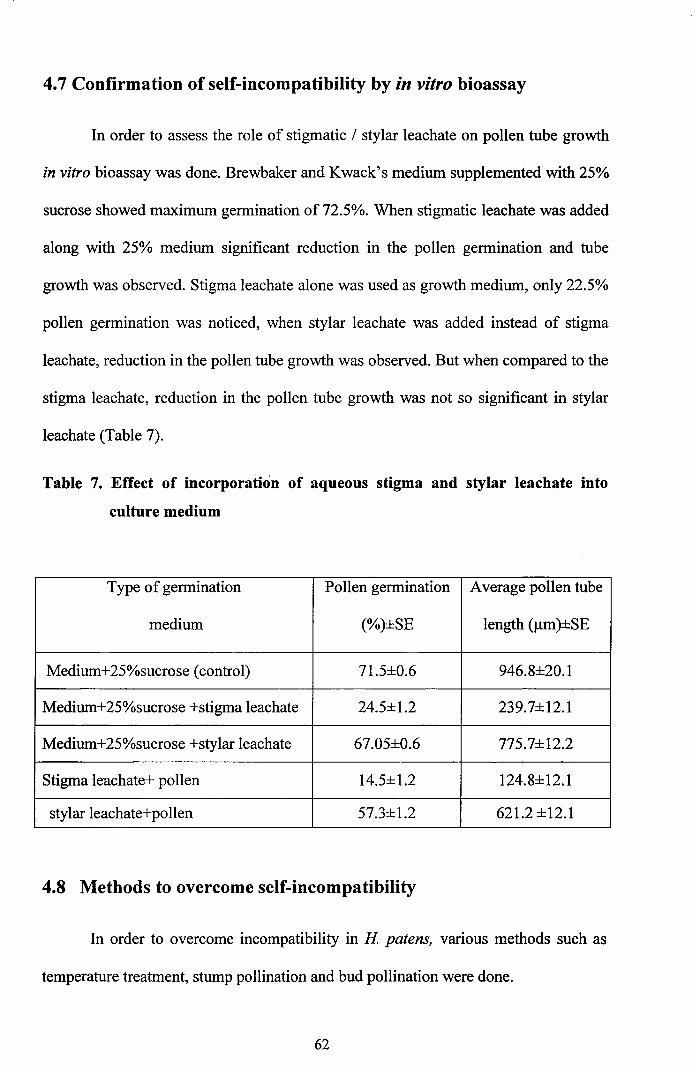
4.7 Confirmation of self-incompatibility by in vitro bioassay
In order to assess the role of stigmatic / stylar leachate on pollen tube growth
in vitro bioassay was done. Brewbaker and Kwack's medium supplemented with 25%
sucrose showed maximum germination of 72.5%. When stigmatic leachate was added
along with 25% medium significant reduction in the pollen germination and tube
growth was observed. Stigma leachate alone was used as growth medium, only 22.5%
pollen germination was noticed, when stylar leachate was added instead of stigma
leachate, reduction in the pollen tube growth was observed. But when compared to the
stigma leachate, reduction in the pollen tube growth was not so significant in stylar
leachate (Table 7).
Table 7. Effect of incorporation of aqueous stigma and stylar leachate into
culture medium
Type of germination Pollen germination Average pollen tube
medium (%)±SE length (Jlm)±SE
Medium+25%sucrose (control) 71.5±O.6 946.8±20.1
Medium+25%sucrose +stigma leachate 24.5±1.2 239.7±12.1
Medium+25%sucrose +stylar leachate 67.05±O.6 775.7±12.2
Stigma leachate+ pollen 14.5±1.2 124.8±12.1
stylar leachate+pollen 57.3±1.2 621.2 ±12.1
4.8 Methods to overcome self-incompatibility
In order to overcome incompatibility in H patens, various methods such as
temperature treatment, stump pollination and bud pollination were done.
62
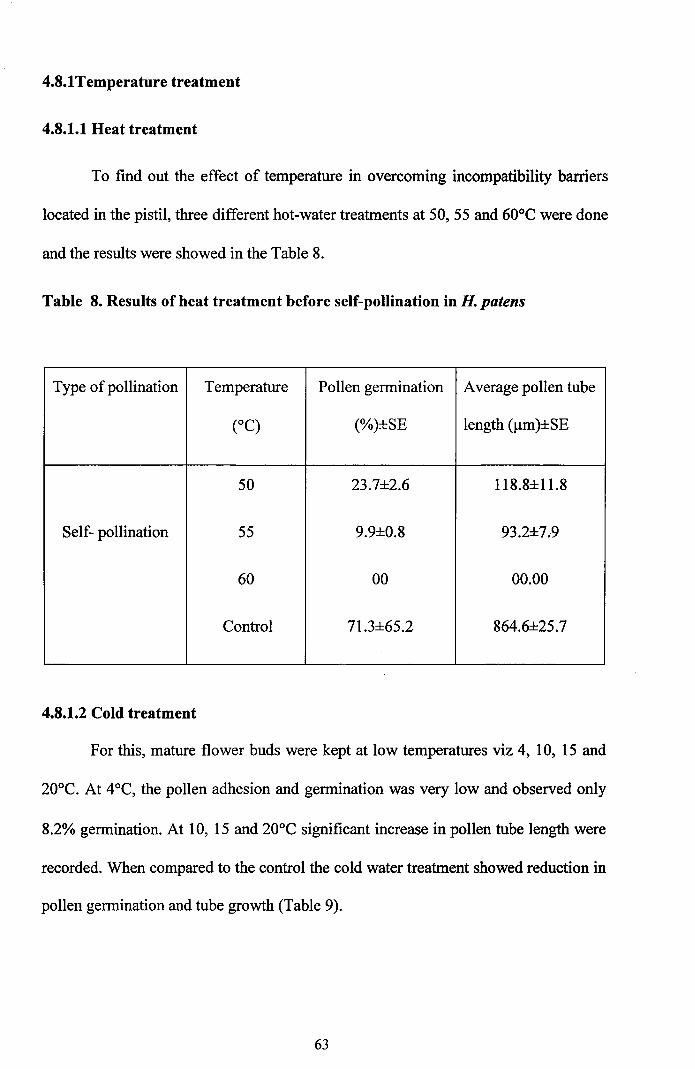
4.8.1Temperature treatment
4.8.1.1 Heat treatment
To find out the effect of temperature in overcoming incompatibility barriers
located in the pistil, three different hot-water treatments at 50,55 and 60°C were done
and the results were showed in the Table 8.
Table 8. Results of heat treatment before self-pollination in H. patens
Type ofpollination Temperature Pollen germination Average pollen tube
(OC) (%)±SE length (J..Lm)±SE
50 23.7±2.6 118.8±11.8
Self- pollination 55 9.9±0.8 93.2±7.9
60 00 00.00
Control 71.3±65.2 864.6±25.7
4.8.1.2 Cold treatment
For this, mature flower buds were kept at low temperatures viz 4, 10, 15 and
20°C. At 4°C, the pollen adhesion and germination was very low and observed only
8.2% germination. At 10, 15 and 20°C significant increase in pollen tube length were
recorded. When compared to the control the cold water treatment showed reduction in
pollen germination and tube growth (Table 9).
63
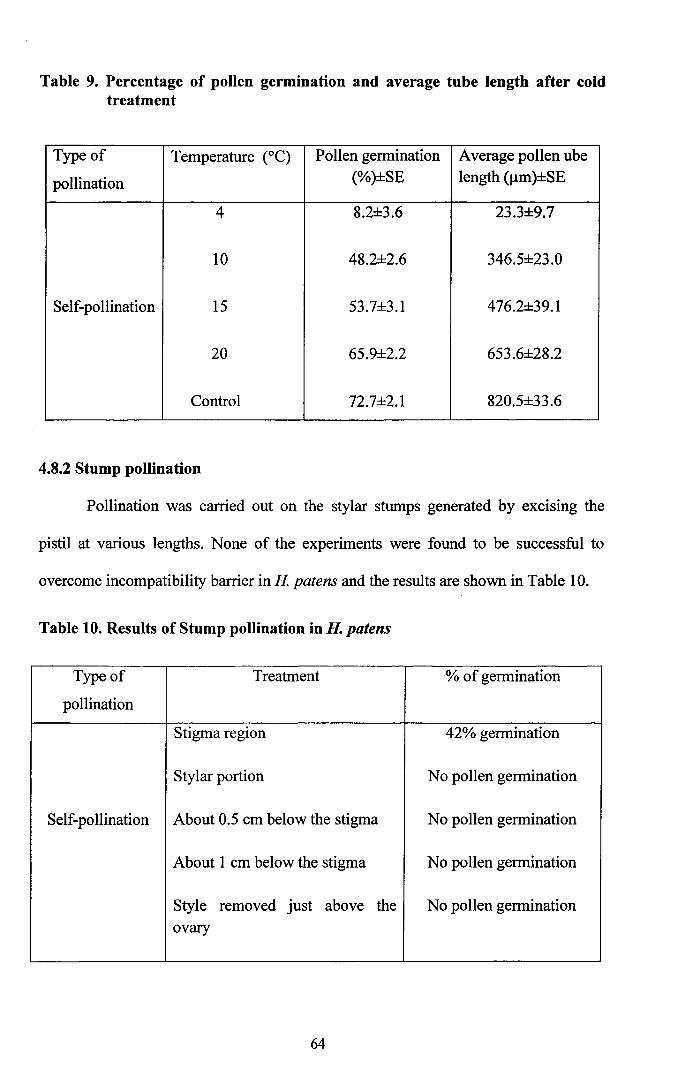
Table 9. Percentage of pollen germination and average tube length after coldtreatment
Type of Temperature (OC) Pollen germination Average pollen ube
pollination (%)±SE length (Jlm)±SE
4 8.2±3.6 23.3±9.7
10 48.2±2.6 346.5±23.0
Self-pollination 15 53.7±3.l 476.2±39.1
20 65.9±2.2 653.6±28.2
Control 72.7±2.1 820.5±33.6
4.8.2 Stump pollination
Pollination was carried out on the stylar stumps generated by excising the
pistil at various lengths. None of the experiments were found to be successful to
overcome incompatibility barrier in H patens and the results are shown in Table 10.
Table 10. Results of Stump pollination in H. patens
Type of Treatment % of germination
pollination
Stigma region 42% germination
Stylar portion No pollen germination
Self-pollination About 0.5 cm below the stigma No pollen germination
About 1 cm below the stigma No pollen germination
Style removed just above the No pollen germinationovary
64
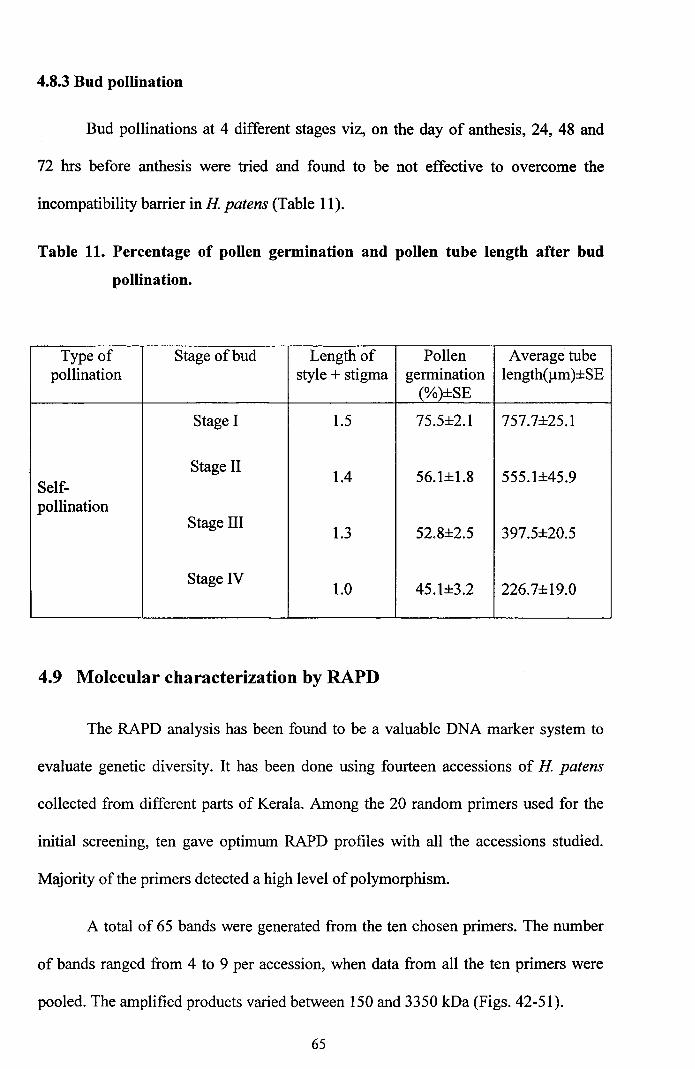
4.8.3 Bud pollination
Bud pollinations at 4 different stages viz, on the day of anthesis, 24, 48 and
72 hrs before anthesis were tried and found to be not effective to overcome the
incompatibility barrier in H patens (Table 11).
Table 11. Percentage of pollen germination and pollen tube length after bud
pollination.
Type of Stage of bud Length of Pollen Average tubepollination style + stigma germination length(llm)±SE
(%)±SE
Stage I 1.5 75.5±2.1 757.7±25.1
Stage II1.4 56.1±1.8 555.1±45.9
Self-pollination
Stage III1.3 52.8±2.5 397.5±20.5
Stage IV1.0 45.1±3.2 226.7±19.0
4.9 Molecular characterization by RAPD
The RAPD analysis has been found to be a valuable DNA marker system to
evaluate genetic diversity. It has been done using fourteen accessions of H patens
collected from different parts of Kerala. Among the 20 random primers used for the
initial screening, ten gave optimum RAPD profiles with all the accessions studied.
Majority of the primers detected a high level ofpolymorphism.
A total of 65 bands were generated from the ten chosen primers. The number
of bands ranged from 4 to 9 per accession, when data from all the ten primers were
pooled. The amplified products varied between 150 and 3350 kDa (Figs. 42-51).
65
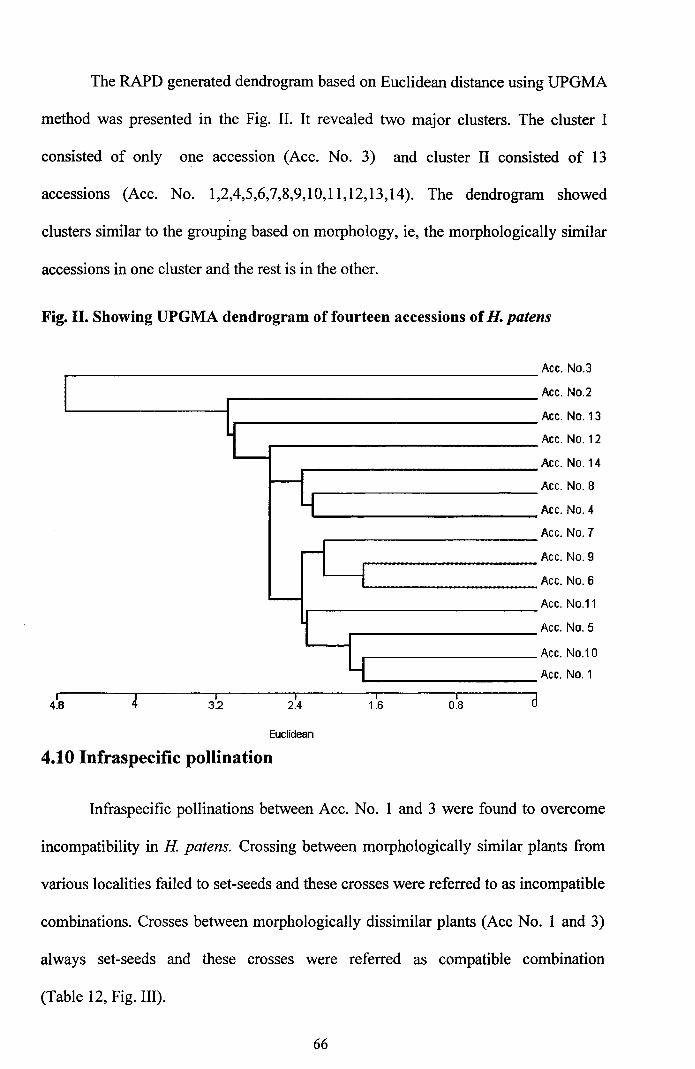
The RAPD generated dendrogram based on Euclidean distance using UPGMA
method was presented in the Fig. II. It revealed two major clusters. The cluster I
consisted of only one accession (Ace. No.3) and cluster II consisted of 13
accessions (Ace. No. 1,2,4,5,6,7,8,9,10,11,12,13,14). The dendrogram showed
clusters similar to the grouping based on morphology, ie, the morphologically similar
accessions in one cluster and the rest is in the other.
Fig. II. Showing UPGMA dendrogram of fourteen accessions ofH. patens
I4.8
I3.2
Euclidean
Ace. NO.3
Ace. NO.2
Ace. No. 13
Ace. No. 12
Ace. No. 14
Ace. NO.8
Ace. NO.4
Ace. NO.7
Ace. NO.9
Ace. NO.6
Ace. NO.11
Ace. NO.5
Ace. NO.10
Ace. NO.1
d
4.10 Infraspecific pollination
Infraspecific pollinations between Ace. No. 1 and 3 were found to overcome
incompatibility in H patens. Crossing between morphologically similar plants from
various localities failed to set-seeds and these crosses were referred to as incompatible
combinations. Crosses between morphologically dissimilar plants (Ace No.1 and 3)
always set-seeds and these crosses were referred as compatible combination
(Table 12, Fig. III).
66
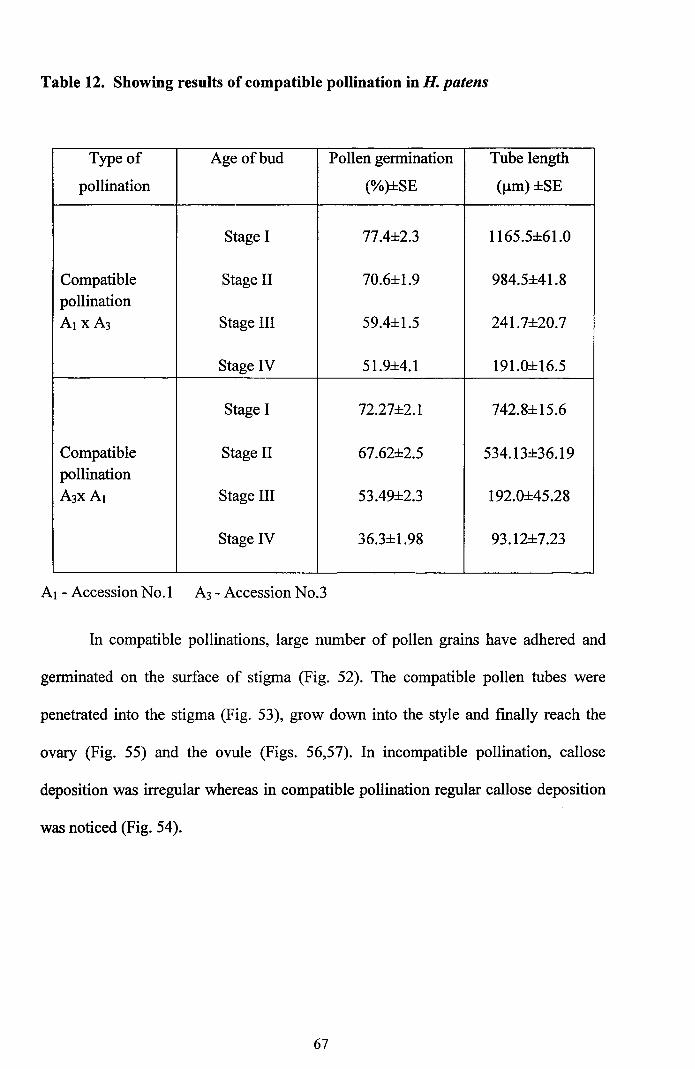
Table 12. Showing results of compatible pollination in H. patens
Type of Age of bud Pollen gennination Tube length
pollination (%)±SE ()lm) ±SE
Stage I 77.4±2.3 1165.5±61.0
Compatible Stage II 70.6±1.9 984.5±41.8pollinationAl xA3 Stage III 59.4±1.5 241.7±20.7
Stage IV 51.9±4.1 191.0±16.5
Stage I 72.27±2.1 742.8±15.6
Compatible Stage II 67.62±2.5 534.13±36.19pollination
A3XA I Stage III 53.49±2.3 192.0±45.28
Stage IV 36.3±1.98 93.12±7.23
Al - Accession No.1 A3 - Accession No.3
In compatible pollinations, large number of pollen grains have adhered and
genninated on the surface of stigma (Fig. 52). The compatible pollen tubes were
penetrated into the stigma (Fig. 53), grow down into the style and finally reach the
ovary (Fig. 55) and the ovule (Figs. 56,57). In incompatible pollination, callose
deposition was irregular whereas in compatible pollination regular callose deposition
was noticed (Fig. 54).
67
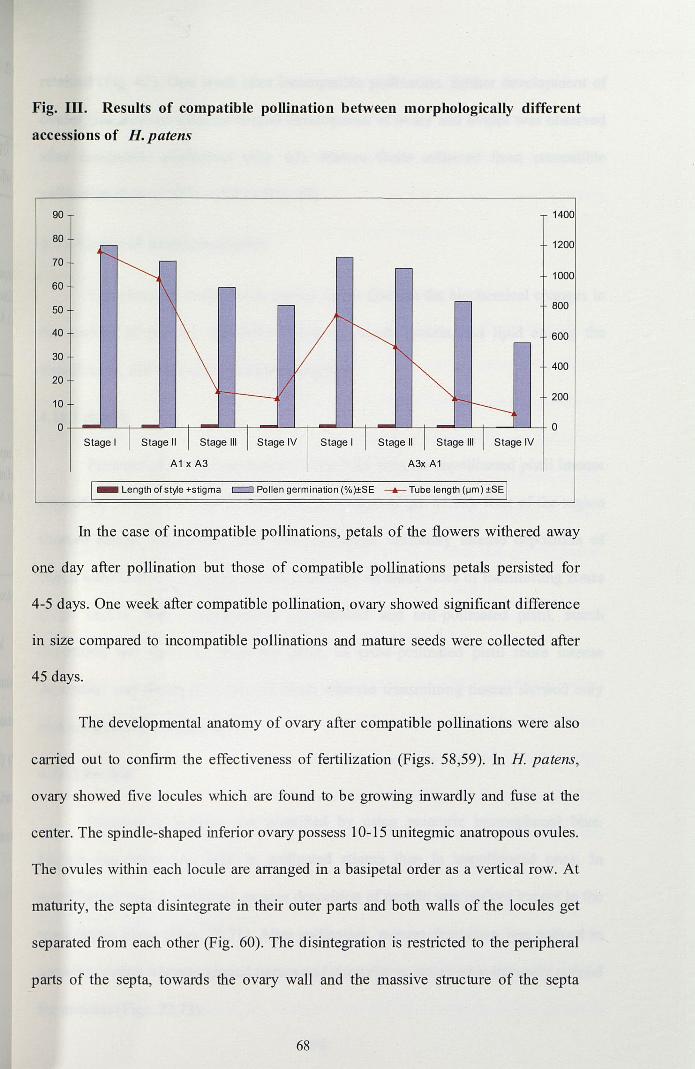
Fig. Ill. Results of compatible pollination between morphologically different
accessions of H. patens
90
80
70
60
50
40
30
20
10
O-j-----J-L--L.+-----'-+---"--'-t--------L-t-------'--;~-----L_t_-----'-+_----'----'_+
Stage I
A1 x A3
Stage IV Stage I
A3x A1
_ Length of style +stigma c::J Pollen germination (%)±SE -.- Tube length (IJm) ±SE
In the case of incompatible pollinations, petals of the flowers withered away
one day after pollination but those of compatible pollinations petals persisted for
4-5 days. One week after compatible pollination, ovary showed significant difference
in size compared to incompatible pollinations and mature seeds were collected after
45 days.
The developmental anatomy of ovary after compatible pollinations were also
carried out to confirm the effectiveness of fertilization (Figs. 58,59). In H patens,
ovary showed five locules which are found to be growing inwardly and fuse at the
center. The spindle-shaped inferior ovary possess 10-15 unitegmic anatropous ovules.
The ovules within each locule are arranged in a basipetal order as a vertical row. At
maturity, the septa disintegrate in their outer parts and both walls of the locules get
separated from each other (Fig. 60). The disintegration is restricted to the peripheral
parts of the septa, towards the ovary wall and the massive structure of the septa
68

retained (Fig. 61). One week after incompatible pollination, further development of
ovules was arrested whereas normal development of ovary and ovules was observed
after compatible pollination (Fig. 62). Mature fruits collected from compatible
pollination showed 95% viability (Fig. 63).
4.11 General histochemistry
Cytochemical studies were carried out to find out the biochemical changes in
the amount of primary metabolites such as starch, protein and lipid among the
unpollinated, self- and cross-pollinated stigmas.
4.11.1 Starch
Presence of starch was detected using I-KI stain. In unpollinated pistil intense
deposition of starch was observed in secretory zone (Figs. 64,65). Rest of the region
showed meager deposition. After self-pollination, uniformly intense deposition of
starch was observed in ground tissues especially on either sides of transmitting zones
(Figs. 66,67). When compared to unpollinated and self-pollinated pistil, starch
deposition was more in pollinated pistil. In cross-pollinated pistil more intense
deposition was observed in ground tissue whereas transmitting tissues showed only
meager deposition (Figs. 68,69).
4.11.2 Protein
Presence of protein was identified by usmg mercurIC bromophenol blue.
Protein deposition was high in pollinated stigma than in unpollinated ones. In
unpollinated stigma uniformly meager deposition of protein was noticed except in the
transmitting tissue (Figs. 70,71). After pollination, meagre deposition was noticed in
secretory region whereas ground tissues and transmitting zones were intensely stained
for proteins (Figs. 72,73).
69

When compared, protein deposition was found to be higher in self-pollinated
stigma than in cross-pollinated one. Uniformly stained ground tissues and meagrely
stained transmitting tissues were seen in cross-pollinated stigma (Figs. 74,75). Cross
pollinated pistil showed slight reduction in the localization of protein in transmitting
tissues and ground tissues when compared to self-pollinated pistil.
4.11.3 Lipids
Presence of lipid was studied using Sudan Black-B. In unpollinated stigma,
localized intense deposition of lipid was observed in the secretory zone of the stigma,
where exudates are seen (Figs. 76,77). Ground tissues below the secretory zone are
free from deposition of lipids but the intense deposition was noticed in ground tissues
on either side of the vascular tissues. After self-pollination, intense deposition was
noticed at the secretory zone of the stigma and vascular region (Figs. 78,79). In cross
pollinated pistil, meagre deposition was observed in transmitting and ground tissues
whereas intense deposition was seen in secretory region (Figs. 80,81).
4.11.4 Periodic acid Schiff (PAS) reaction for polysaccharides
PAS reaction showed uniform distribution of PAS positive bodies.
Polysaccharides appeared as dark pink coloured bodies. Uniformly less intense
deposition of PAS positive bodies were observed in unpollinated stigma (Figs. 82,83)
where as localized intense deposition was noticed in pollinated one (Figs. 84,85).
When compared to self-pollinated pistil, cross-pollinated pistils showed less
deposition (Figs. 86,87).
4.12 Enzyme cytochemistry
Since the enzymes control biochemical reactions, and their synthesis is under
the control of specific gene (s), any change in the activity of enzyme would reflect in
70

the pattern of gene expression and corresponding metabolic events in the cell. Hence,
the enzymes can be used as a tool to study the problem of self-incompatibility at the
biochemical level. In the present investigation, cytochemical changes in the
localization of different enzymes such as succinic dehydrogenase, acid phosphatase,
ATPase, peroxidase and esterase in unpollinated, self-pollinated and cross-pollinated
stigmas were studied.
4.12.1 Succinic dehydrogenase
SDH activity was localized as blue coloured bodies. In unpollinated pistil, the
activity was uniformly high in the entire region except transmitting zone, where
intense SDH activity was observed (Figs. 88,89). After self-pollination localized
intense activity was observed on either side of the transmitting region and secretory
zone (Figs. 90,91). When compared to self-pollinated pistil, cross-pollinated pistil
showed meagre activity except in the tissue on either side of transmitting tissue
(Figs. 92,93).
4.12.2 Acid phosphatase
Acid phosphatase activity appeared as blackish brown deposits and was very
intense in papillar, epidermal and sub epidermal regions (Fig. 94). The central core of
transmitting tissue showed uniformly less enzyme activity (Fig. 95). Maximum
activity was observed in the unpollinated stigma. Self- and cross-pollinated pistil
showed slight decrease in the activity of APase (Figs. 96-99). Secretory zone and
vascular regions showed some localized intense activity.
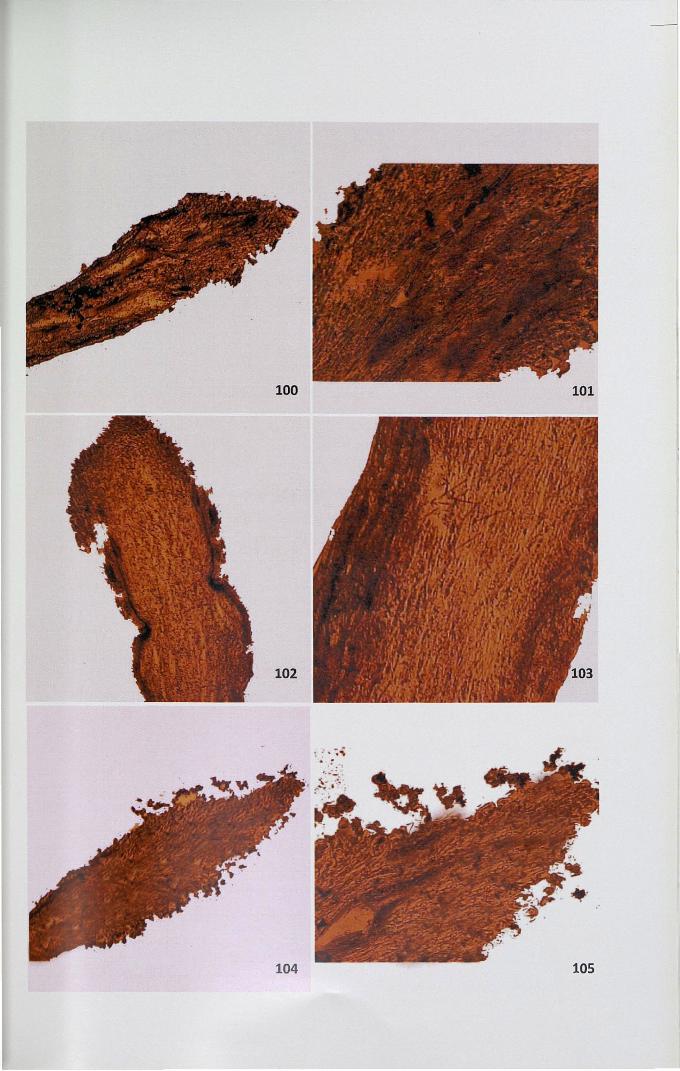
4.12.3 Adenosine triphosphatase
The activity site of ATPase was observed as brown black deposits of lead
sulphide distributed as granules. In unpollinated stigma, papillar region showed
71

intense activity compared to the ground tissue (Figs. 100,101). The activity of this
enzyme was high in unpollinated pistil. When compared to self-pollinated pistil
(Figs. 102,103), cross-pollinated pistil showed less intense activity (Figs. 104,105).
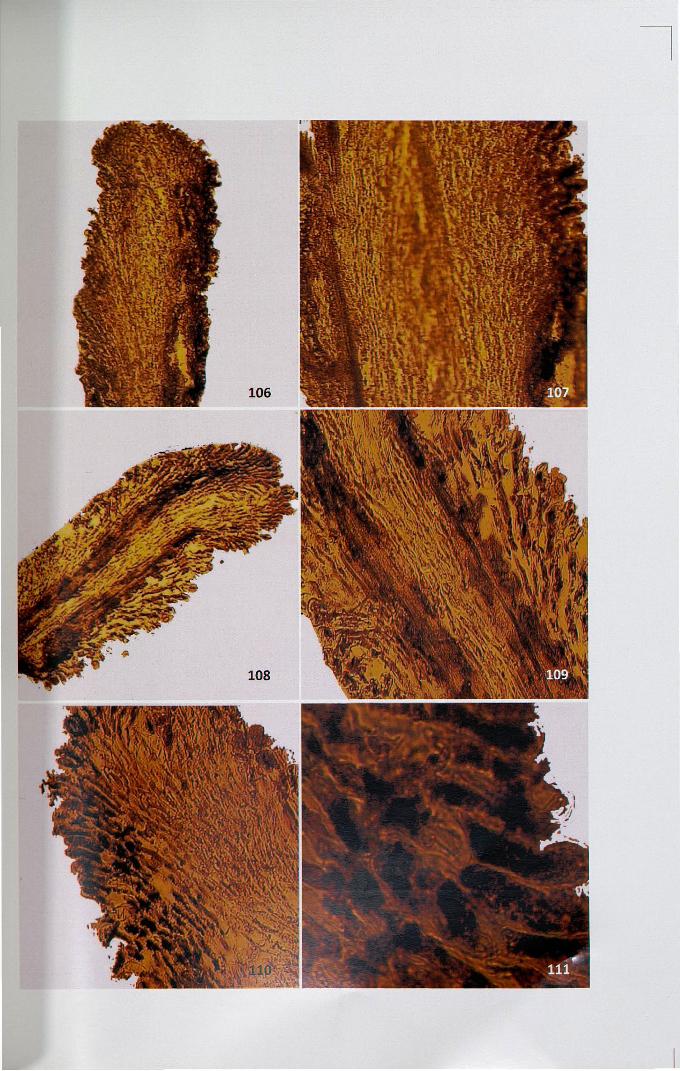
4.12.4 Peroxidase
Activity site appeared as reddish brown reaction products. Intense activity was
observed in papillar and secretory zone and the rest of the region showed
comparatively less uniform distribution (Figs. 106,107). In H patens, pollination
stimulated the activity of peroxidase in the pistil (Figs. 108,109). In the papilla region,
uniformly intense deposition was noticed. In cross-pollinated pistil, less intense
deposition was observed (Figs. 110,111).
4.12.5 Esterase
Esterase activity was detected as brick red coloured granules. In unpollinated
stigma, uniformly less activity (Fig. 112) was observed except in the region of
secondary tissues (Fig. 113) when compared to unpollinated stigma self- and cross
pollinated pistils showed higher activity (Figs. 114-117).
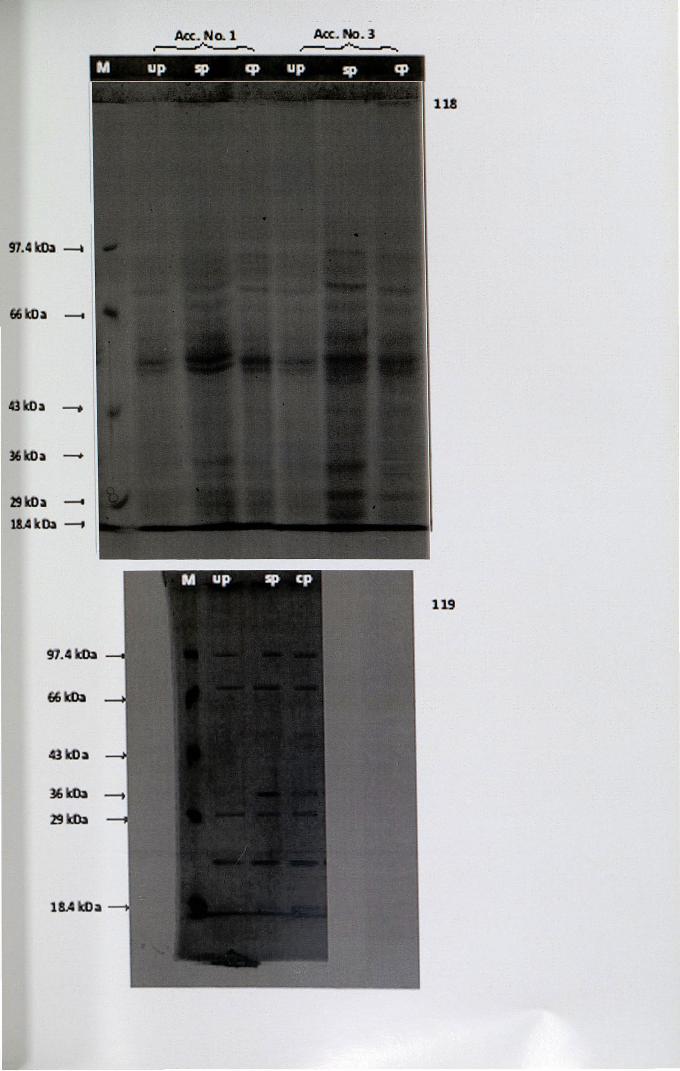
4.13 Electrophoresis
Since infraspecific pollination was found to be successful in H patens, the
protein profiles of stigma homogenate and leachate of unpollinated, self- and cross
pollinated pistils were analysed using the technique of SDS-PAGE. Stigma
homogenate revealed 15 protein bands with molecular weight ranging from
20-100 kDa (Fig. 118). The protein bands of self- and cross-pollinated pistils were
common except in 36 kDa region. When compared the band with molecular weight
36 kDa was found to be absent in unpollinated pistil and also the intensity of the
bands were less intense.
72

However, the electrophoretic studies of stigma leachate of unpollinated, self
and cross-pollinated pistil enabled the detection of five common bands with molecular
weights 23, 30, 36, 70 and 99 kDa (Fig. 119), but showed quantitative difference. It is
noteworthy that protein band at 36 kDa region was common in stigma leachate as well
as homogenate of self- and cross-pollinated pistil, while the band is totally absent in
unpollinated pistil.
4.14 RNase activity
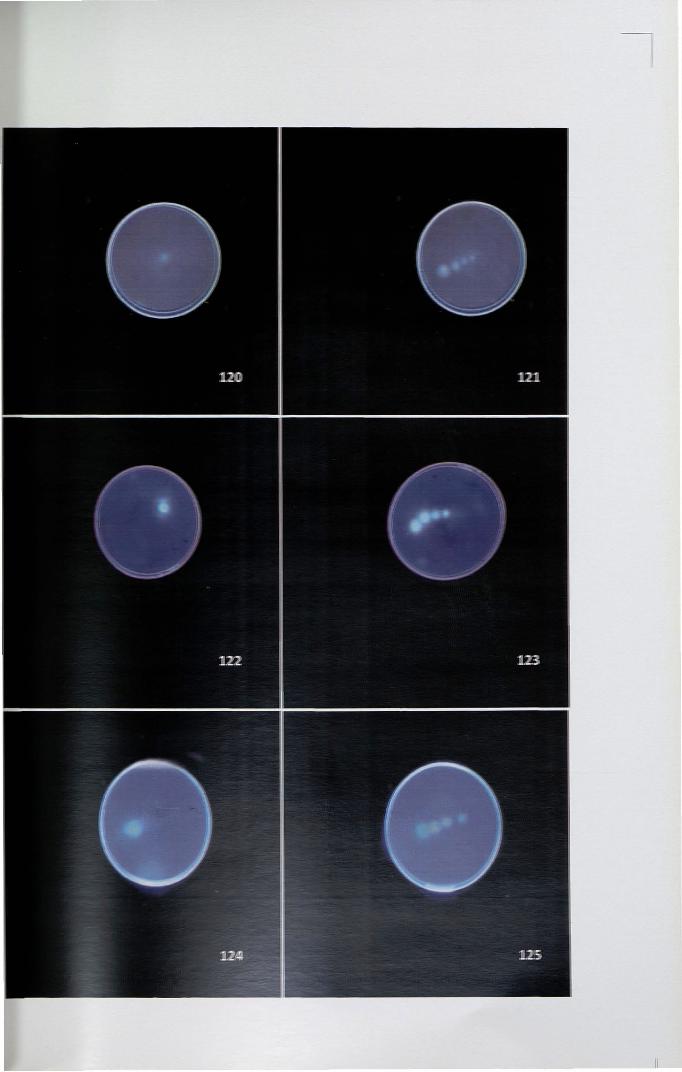
4.14.1 Diffusion plate assay
In H patens, the diffusion plate assay for RNase activity appeared as white
halos in a blue background around the stigmatic surface. In vivo germination studies
using decolourized aniline blue confirmed that the inhibition is only on the stigmatic
surface. From the analyses, it was also observed that the RNase activity in the
stigmatic diffusate is less in early developmental stages and reaches maximum at
anthesis. When compared, the activity was very high in self-pollinated pistil followed
by cross-pollinated and less in unpollinated pistil (Figs. 120-125).
4.14.2 Quantitative spectrophotometric assay of RNase activity
Spectrophotometric assay was carried out to check the RNase activity and
found that the activity was high in self-pollinated than unpollinated and cross
pollinated stigma. Further analysis was done to find out RNase activity in stigma or
style which confirmed the activity high on the stigma than style (Table 13, Fig. IV).
73
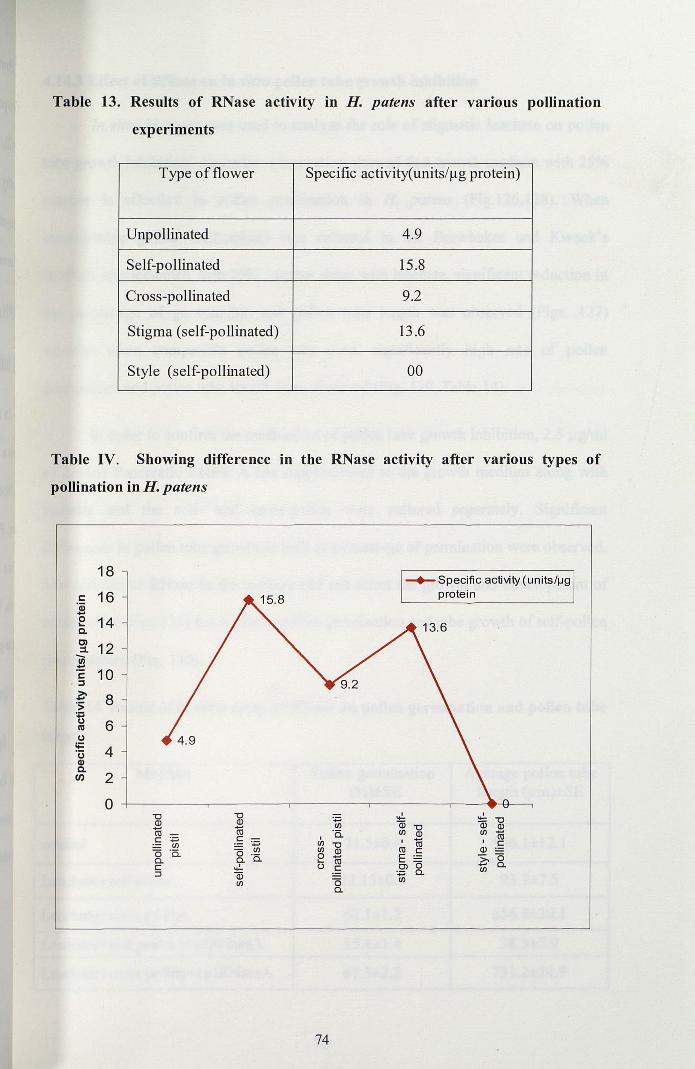
Table 13. Results of RNase activity in H. patens after various pollination
experiments
Type of flower Specific activity(units/~g protein)
Unpollinated 4.9
Self-pollinated 15.8
Cross-pollinated 9.2
Stigma (self-pollinated) 13.6
Style (self-pollinated) 00
Table IV, Showing difference in the RNase activity after various types of
pollination in H. patens
18
s:: 16'(jj-E 14c.Cl 12:::l.-III:!::
10s::,j
~ 8'S;..CJ 6I1lCJ
li= 4'uQ)C.In 2
0"C "CQ) Q)- -m mc :;:::; c
'';:::;IJl0 '5. 0 IJl
Cl. Cl. '5.c .,!.j (ii
IJl
-+-Specific activity (units/1J9protein
.,!. I:;:::; lii "CIJl (ii "C Q)'5. IJl Q) IJl coI
I - IIJl "C m cIJl Q) m c Q)0 co E Z, 0... 0() c OJ Cl.
:;:::; Cl. IJl
0 IJl
Cl.
74
4.14.3 Effect of RNase on in vitro pollen tube growth inhibition
In vitro bioassay was used to analyze the role of stigmatic leachate on pollen
tube growth inhibition. An earlier observation showed that growth medium with 25%
sucrose is effective in pollen germination in H patens (Fig.126,128). When
incompatible pollen (self-pollen) was cultured in the Brewbaker and Kwack's
medium supplemented with 25% sucrose along with leachate, significant reduction in
the percentage of germination and pollen tube length was observed (Figs. 127)
whereas when compatible pollen was used, significantly high rate of pollen
germination and pollen tube length were observed (Fig. 129, Table 14).
In order to confirm the mechanism of pollen tube growth inhibition, 2.5 Ilg/ml
of Bovine Pancreatic RNase A has supplemented to the growth medium along with
leachate and the self- and cross-pollen were cultured separately. Significant
differences in pollen tube growth as well as percentage of germination were observed.
The addition of RNase to the medium did not affect the growth and development of
cross-pollen (Fig. 131) but it affects pollen germination and tube growth of self-pollen
was observed (Fig. 130).
Table 14. Result of in vitro assay of RNase on pollen germination and pollen tube
length
Medium Pollen germination Average pollen tube(%)±SE length (Ilm)±SE
control 71.5±0.6 906.1±12.1
Leachate+selfpollen 22.15±0.9 93.7±7.5
Leachate+cross pollen 62.1±1.2 656.8±20.1
Leachate+self pollen+I1llRNaseA 15.4±1.4 38.3±5.9
Leachate+cross pollen+I1llRNaseA 67.3±2.2 731.2±24.9
75

4.14.4 Effect of RNase regulator on in vitro pollen tube growth
Generally two chemicals namely zinc sulfate and cupric sulfate inhibit the
activity of stigmatic RNase and promote pollen germination. In this study also
different concentrations of these chemicals were applied in the medium having
stigmatic leachate. Zinc sulfate (l mM) was found to be more effective in reducing
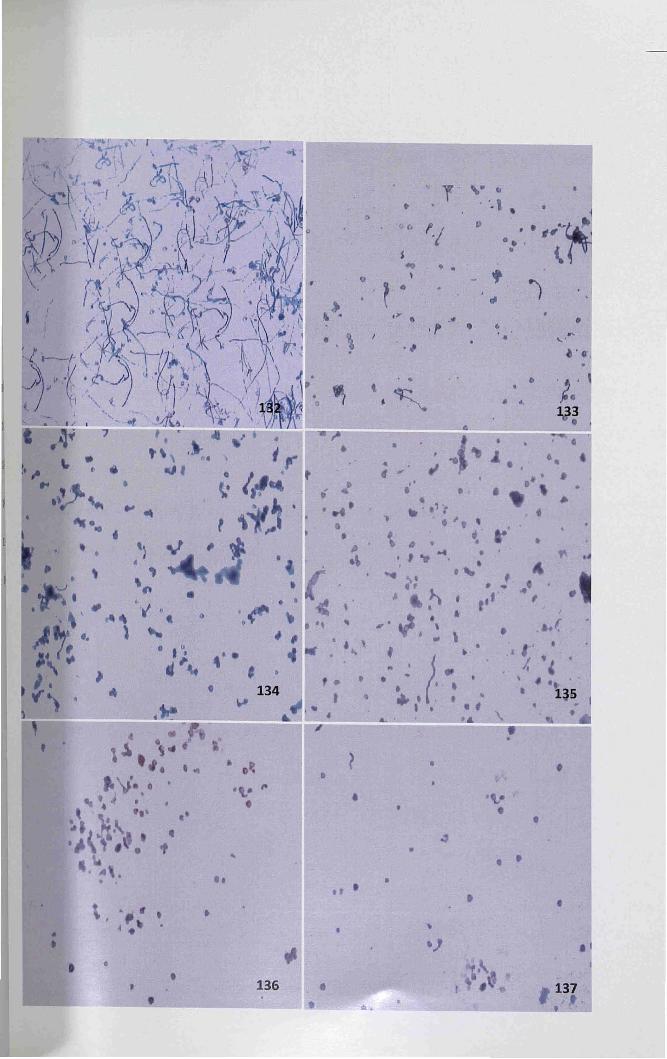
the inhibitory activity of RNase on pollen tube growth (Fig. 132). After the addition
of ImM zinc sulfate, the inhibition of pollen tube growth was undetected. But when
the concentration exceeds ImM, the pollen tube exhibited disturbed growth
(Fig. 134,136). Additionally, zinc sulfate was superior to cupric sulfate as a chemical
ofpollen tube growth in vitro by decreasing the RNase activity. The cupric sulfate did
bind to the outer surface of pollen the tissue. This disturbed the pollen tube
germination (Figs. 133,135,137).
76

Figures 1- 6
I. H patens Accession No.l- Habit
2. H patens Accession No.2- Habit
3. Morphological variation between the two accessions
4. Fruit and L.S offruit
5. C.S of the ovary with sterile ovules x 4
6. Stereomicrograph ofC.S ofthe fruit


Figures 7-12
7. Stamen from a single flower
8. C.S of anther - Tetrasporangiate showing fibrous endothecium and stomiumx4
9. C.S of anther- A portion enlarged showing fibrous endothecium and pollengrains x 40
10. Stereomicrograph of mature stigma and style
11. L.S ofthe stigma showing papillae x 10
12. L.S of the stigma showing vascular tissues and central transmitting tissue x4.(t-transmitting tissue, v- vascular tissue)
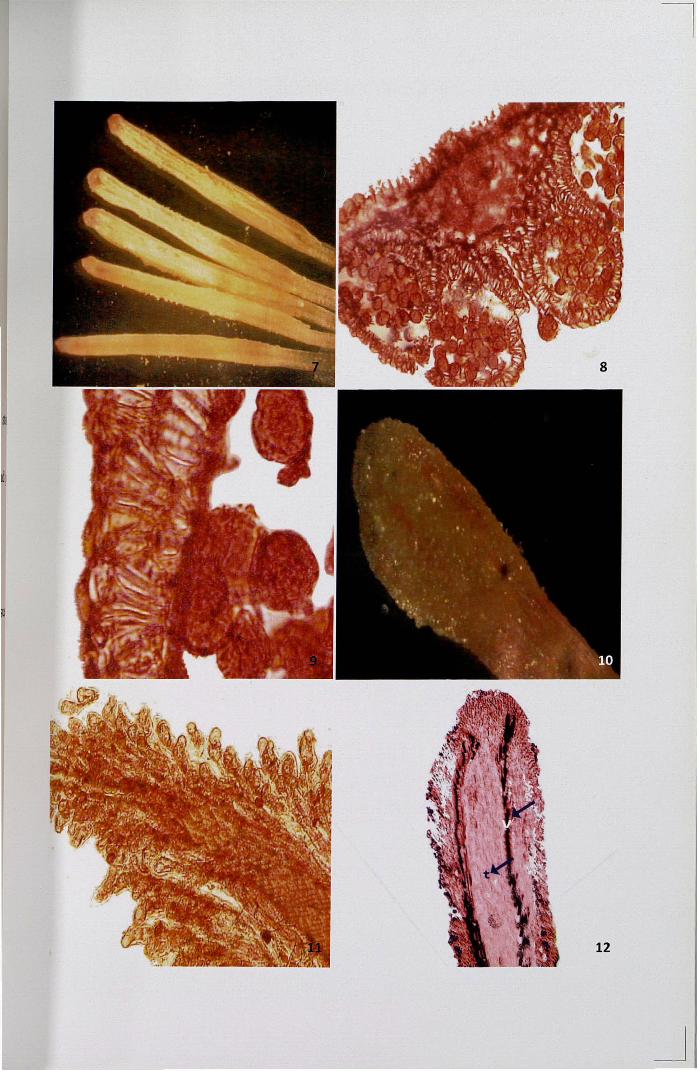

Figures 13- 20. Scanning Electron Micrographs of stigma at different developmental
stages
13. Four days before anthesis
14. Irregular stigmatic surface four days before anthesis - a portion enlarged
15. Three days before anthesis
16. Stigma surface three days before anthesis - a portion enlarged
17. Two days before anthesis
18. Fully developed papillae covering the stigmatic surface - a portionenlarged
19. One day before anthesis
20. Stigma surface with pollen grains one day before anthesis
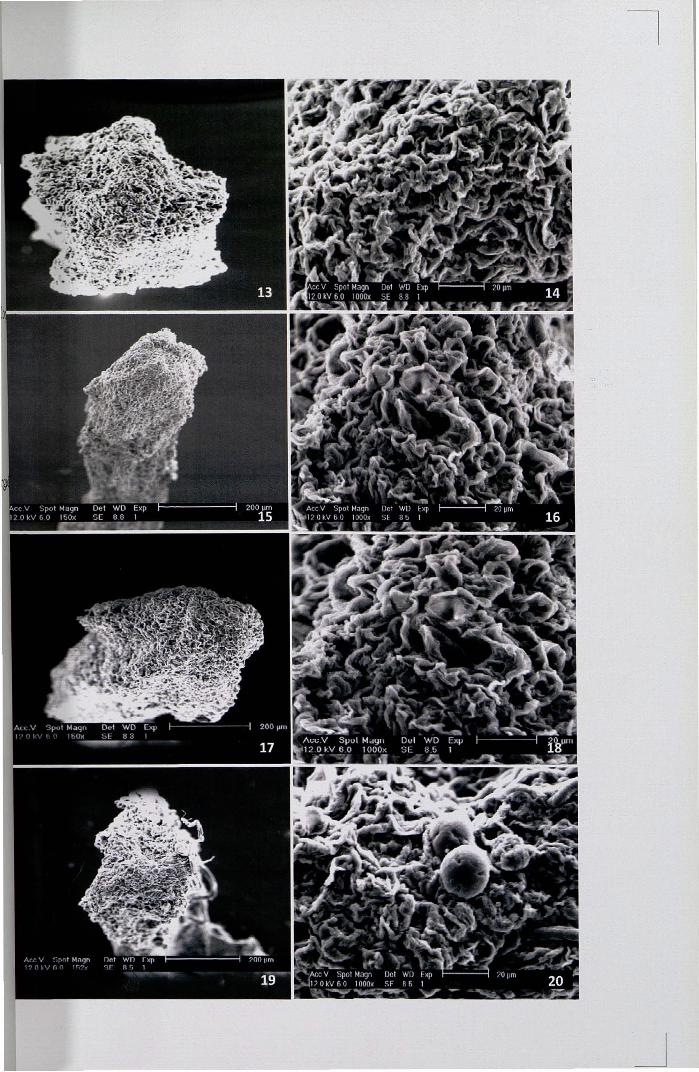

Figures 21-26
21. SEM of L.S of the ovary showing sterile ovules (ov-ovule)
22,23. SEM ofL.S ovary - a portion enlarged (ov-ovule)
24. Meiotic metaphase n = 12 x 40
25. Pollen grain showing two nuclei at the time of shedding x 40
26. Pollen grains showing intense fluorescence by FeR test x 10
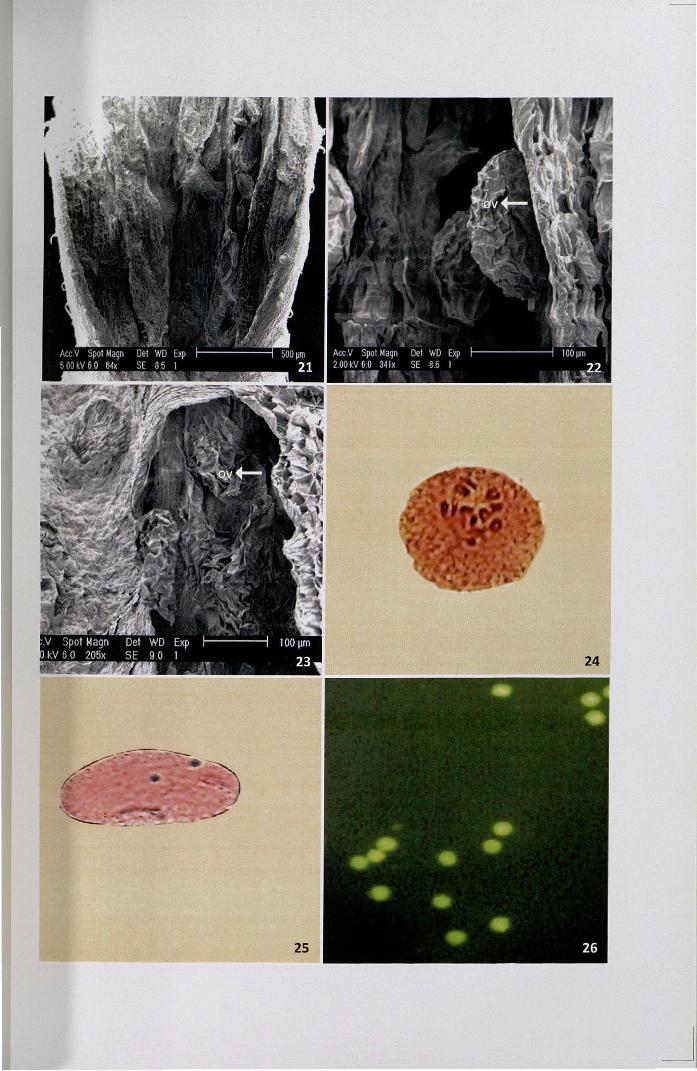

Figures 27-35
27. In vitro pollen germination in Brewbaker and Kwack's medium supplementedwith 25% sucrose x 10
28. Buds at various developmental stages
29-32. Stereomicrograph of localization of esterase on the stigma surface atdifferent developmental stages
29. Bud at 3 days before anthesis
30. Bud at 2 days before anthesis
31. Bud at one day before anthesis
32. Bud on the day of anthesis
33. L.S of the stigma and style showing the activity of esterase x 40
34-35. Various floral visitors
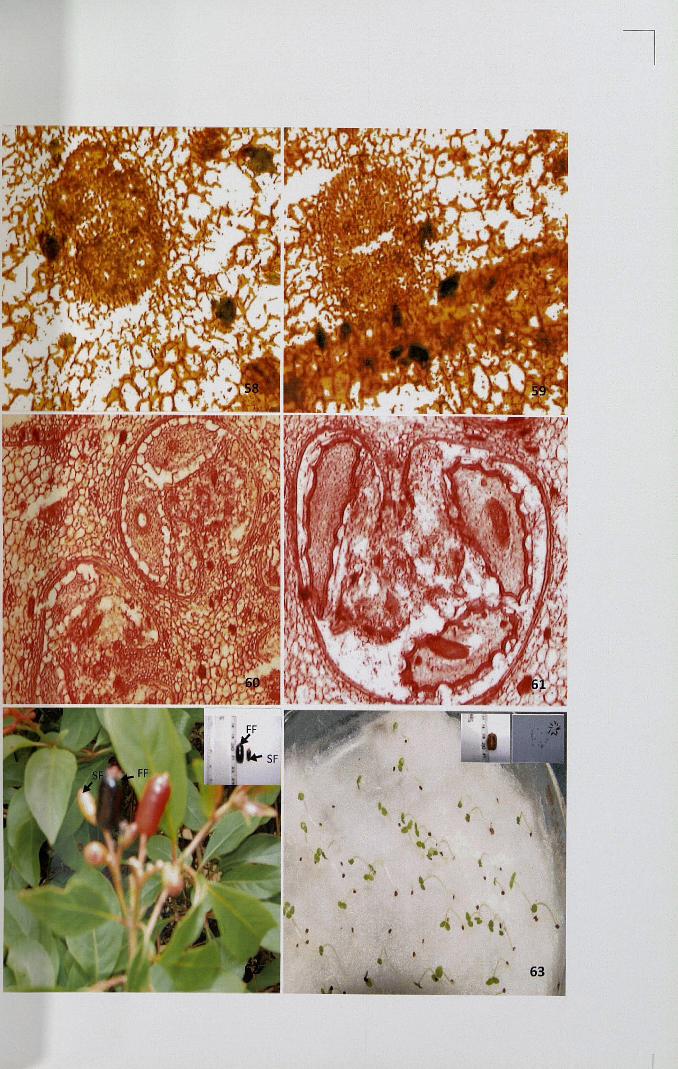

Figures 64-69
64. Unpollinated pistil showing localized intense deposition of starch x 4
65. Unpollinated pistil showing intense deposition of starch in the secretoryregion x 10
66,67. Self-pollinated pistil showing uniformly intense deposition in the vascularregion x 10
68,69. Cross-pollinated pistil showing intense deposition of starch x 4, x 10
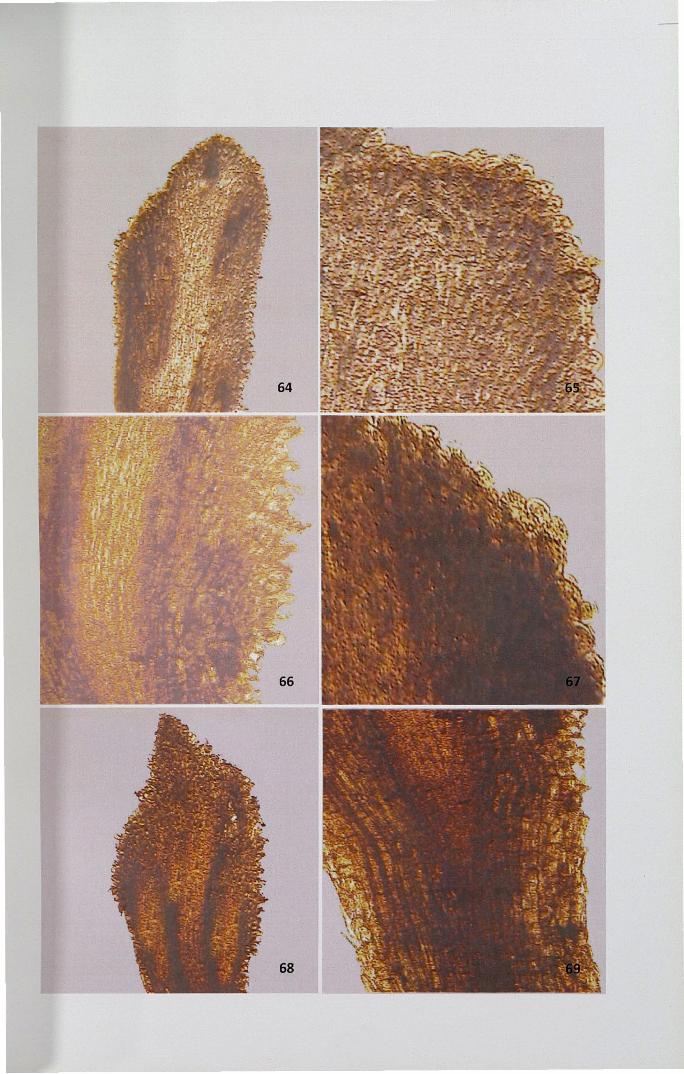

Figures 70-75
70. Meagre deposition ofprotein before pollination x 4
71. Uniformly meagre deposition ofprotein in the secretory zone x 40
72,73. Meagre deposition ofprotein in secretory region and intensely stainedtransmitting zones in self-pollinated stigma x 4, x 10
74,75. Uniformly stained ground and transmitting tissue and intenselystained vascular tissues in cross-pollinated stigma x 4, x 10
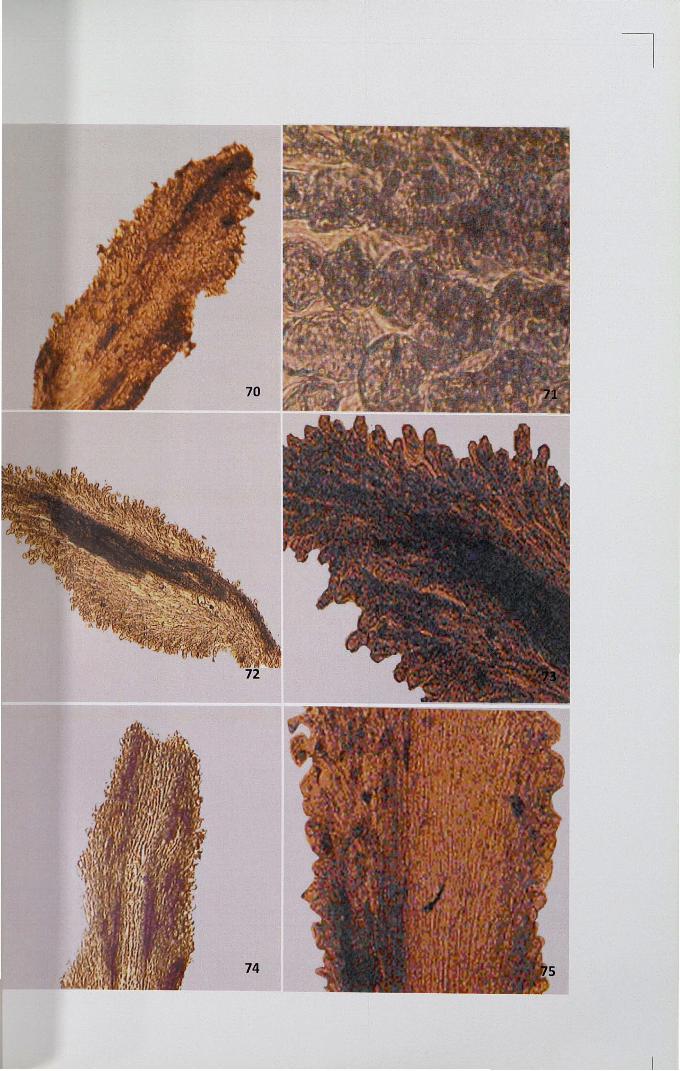

Figures 76-81
76.
77.
78,79.
80.
81.
Localized intense deposition of lipid in the secretory zone ofunpollinated stigma x 4
Intense deposition of lipids in ground tissues on either side of thevascular tissues in unpollinated stigma x 10
Intense deposition of lipids in stigma after self-pollination x 4, x 10
Meagre deposition of lipid in the transmitting zone of cross-pollinatedstigma x 10
Intense deposition of lipid in the secretory region after cross-pollination x 40


Figure 82-87
82,83. Uniformly less intense deposition ofPAS positive bodies in maturestigma x 10
84. Localized intense deposition ofPAS positive bodies in self-pollinatedpistil x 4
85. Deposition of PAS bodies in papillar region x 40
86. Cross-pollinated pistils showed less PAS deposition x 4
87. Uniformly meagre deposition of PAS positive bodies x 40


Figures 88-93
88. Unpollinated pistil showing intense activity ofSDH at transmittingregion and vascular tissues x 10
89. Intense activity of SDH at transmitting region x 40
90. Localized intense activity of SDH on either side of the transmittingregion after self-pollination x 10
91. Intense activity ofSDH in secretory zone and vascular tissue afterself-pollination x 10
92. Cross-pollinated pistil showing meagre activity of SDH x 4
93. Localised intense activity of SDH after cross-pollination x 10


Figures 94-99
194. Intense activity of acid phosphatase in papillar, epidermal and subepidermalregions ofunpollinated stigma x 4
95. The central core oftransmitting tissue showing uniformly less enzymeactivity x 10
96,97. Activity ofAPase in self-pollinated stigma x 4, x 10
98,99. Activity ofAPase in cross-pollinated stigma x 40
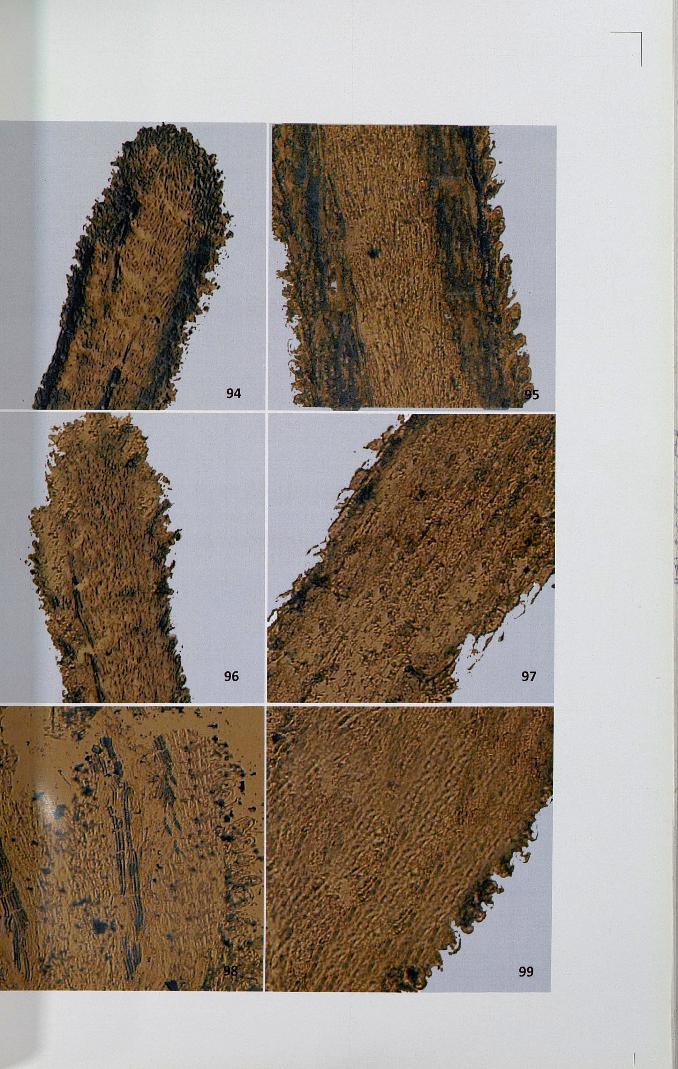

Figures 100-105
100,101. High activity of ATPase in unpollinated stigma x 4, x 10
102,103. Intense activity ofATPase in self-pollinated stigma x 4, x 10
104,105. Activity ofATPase in cross-pollinated stigma x 10, x 10
100
102
101

Figures 76-81
76.
77.
78,79.
80.
81.
Localized intense deposition of lipid in the secretory zone ofunpollinated stigma x 4
Intense deposition of lipids in ground tissues on either side of thevascular tissues in unpollinated stigma x 10
Intense deposition of lipids in stigma after self-pollination x 4, x 10
Meagre deposition of lipid in the transmitting zone of cross-pollinatedstigma x 10
Intense deposition of lipid in the secretory region after cross-pollination x 40


Figure 82-87
82,83. Uniformly less intense deposition ofPAS positive bodies in maturestigma x 10
84. Localized intense deposition ofPAS positive bodies in self-pollinatedpistil x 4
85. Deposition of PAS bodies in papillar region x 40
86. Cross-pollinated pistils showed less PAS deposition x 4
87. Uniformly meagre deposition of PAS positive bodies x 40


Figures 88-93
88. Unpollinated pistil showing intense activity ofSDH at transmittingregion and vascular tissues x 10
89. Intense activity of SDH at transmitting region x 40
90. Localized intense activity of SDH on either side of the transmittingregion after self-pollination x 10
91. Intense activity ofSDH in secretory zone and vascular tissue afterself-pollination x 10
92. Cross-pollinated pistil showing meagre activity of SDH x 4
93. Localised intense activity of SDH after cross-pollination x 10


Figures 94-99
94. Intense activity of acid phosphatase in papillar, epidermal and subepidermalregions ofunpollinated stigma x 4
95. The central core oftransmitting tissue showing uniformly less enzymeactivity x 10
96,97. Activity ofAPase in self-pollinated stigma x 4, x 10
98,99. Activity ofAPase in cross-pollinated stigma x 40
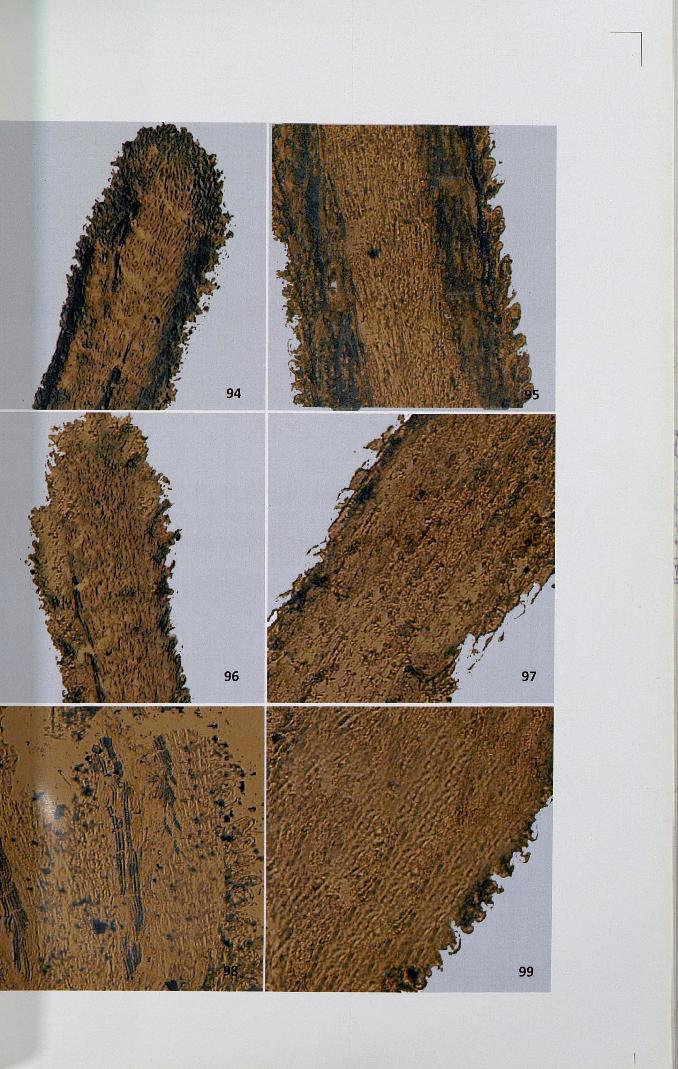

Figures 100-105
100,101. High activity of ATPase in unpollinated stigma x 4, x 10
102,103. Intense activity ofATPase in self-pollinated stigma x 4, x 10
104,105. Activity ofATPase in cross-pollinated stigma x 10, x 10
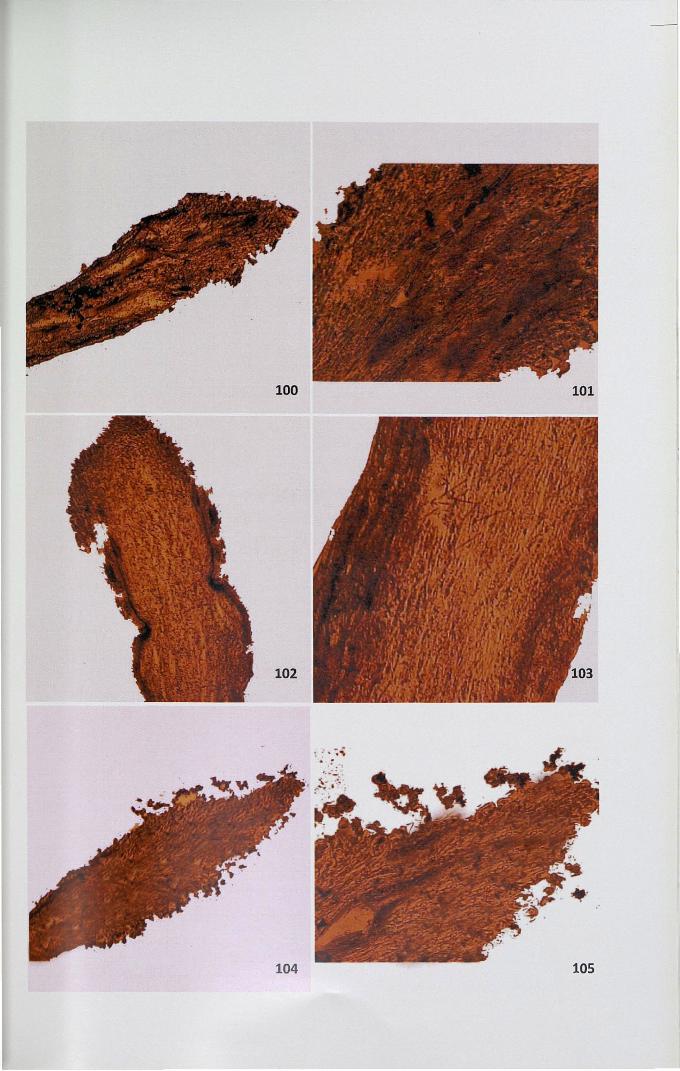
100
102
101

Figures 106-111
106. Activity ofperoxidase in unpollinated stigma x 4
107. Intense activity ofperoxidase in papillar and secretory region ofunpollinated stigma x 10
108. Activity ofperoxidase on stigma after self-pollination x 4
109. Intense activity ofperoxidase after cross-pollination x 10
110. Intense activity ofperoxidase after cross-pollination x 10
111. Intense activity ofperoxidase at papillar region after cross-pollination x 40

Figures 112 -120.
112. Meagre activity of esterase in unpollinated stigma x 10
113. Intense activity ofesterase in secondary tissues x 10
114,115. Self-pollinated stigma showing increase in the activity of esterase x 10
116,117. Intense activity of esterase on the surface ofcross-pollinated stigma x 10
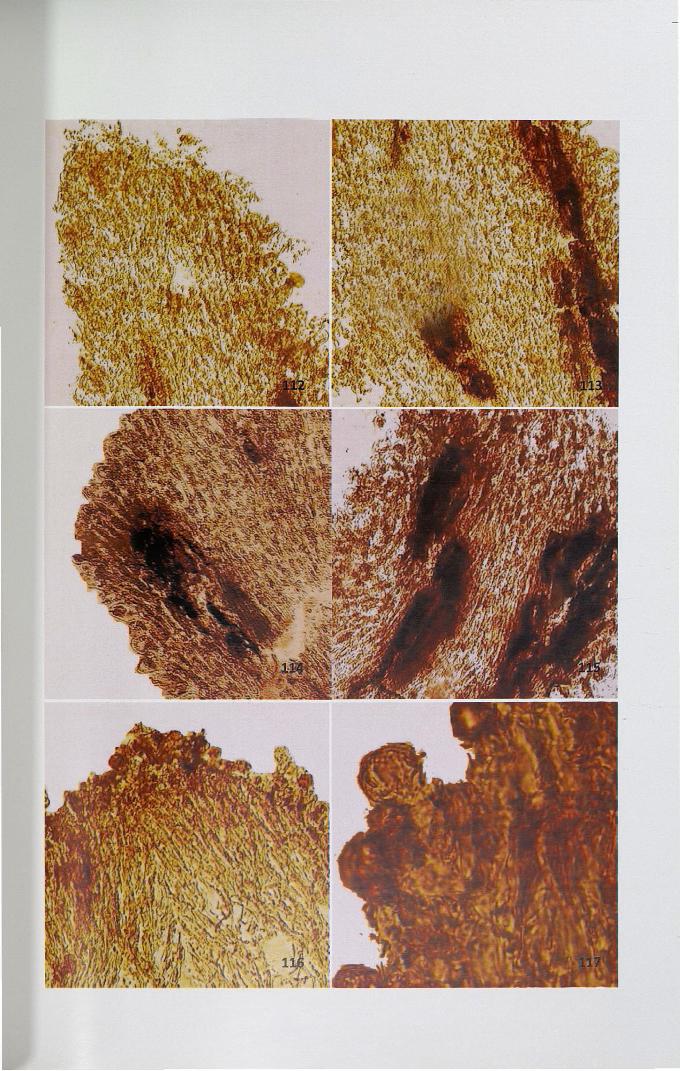

Figure 118. Protein profile of stigma homogenate ofunpollinated, self- and crosspollinated stigma
Figure 119. Protein profile of stigma leachate ofunpollinated, self- and crosspollinated stigma
M - Molecular marker, up - unpollinated, sp - self-pollinated,
cp - cross- pollinated pistil
U.8

Figure 120-125. In H patens, diffusion plate assay showing RNase activity appeared
as white halos on blue background
120. Mature unpollinated stigma showing less RNase activity x 4
121. RNase activity ofunpollinated stigma at different developmental stages x4
122. Mature self-pollinated stigma showing high RNase activity x 4
123. RNase activity of self-pollinated stigma at different developmental stages x4
124. Cross-pollinated stigma with less RNase activity x 4
125. RNase activity on stigma at different developmental stages aftercross-pollination x 4
____ll

Figure 126-131. In vitro pollen tube growth inhibition by self stigma leachate of
Hpatens
126. Pollen germination at 25% sucrose supplemented in Brewbacker'smedium (control) x 4
127. Pollen germination in 25% sucrose medium with self stigma leachate x 4
128. Self-pollen cultured with cross stigma leachate containing sucrosemedium x 4
130. Self-pollen cultured with self stigma leachate containing medium withRNaseA x4
129,131. Self-pollen cultured with cross stigma leachate containing mediumwith RNaseA x 4
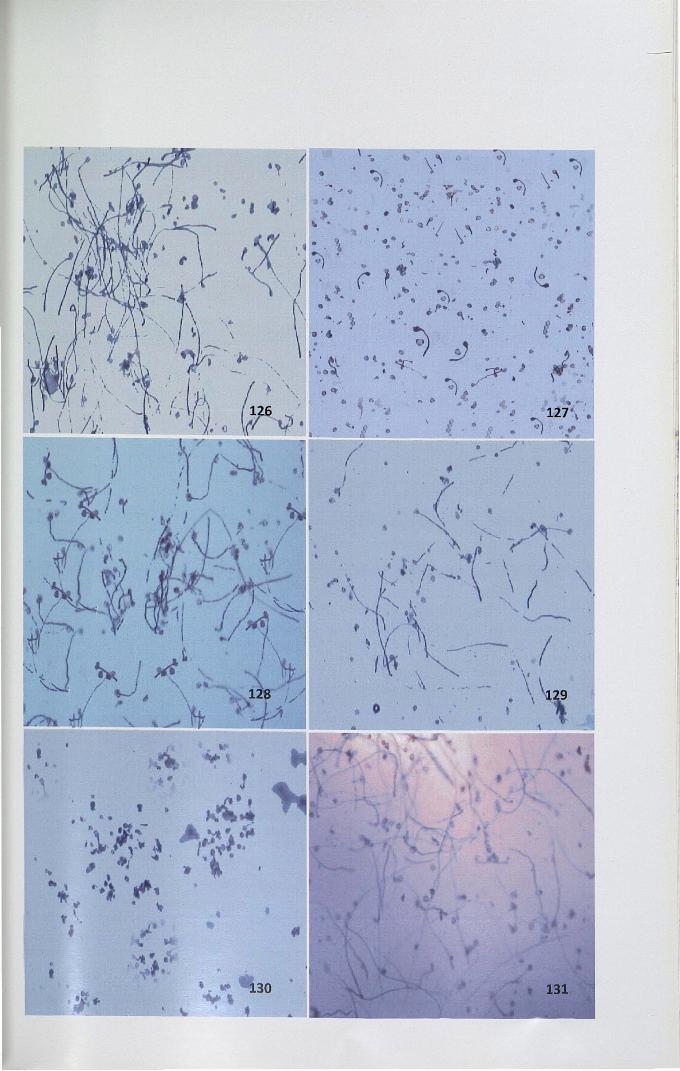
• •
. '..... I
" '
. .."..
/I
~- .
~
\ . ~~, ... , ~
'. ,.. ' . .
•
..1,..e -
, 1'/.'. . \.'.. $ . .
(~" • ,r~. 0.. .;.~,
~,. .. .! I. ~ .. .. '. .'
',I
'. • .~.:J ....."'~~~ ~ 7/
• ct VJ. "
•
\ ... \-.~ . ~, ..
',I •
I
\
•..•
,--, ~
"\,,
, l,.

Figure 132-137. In vitro pollen tube growth by self stigma leachate ofH patens
containing RNase inhibitor (ZnS04 and CUS04)
132. Self-pollen cultured with self stigma leachate containing 1mM ZnS04 x 4
133. Self-pollen cultured with self stigma leachate containing 1mM CUS04 x 4
134. Self-pollen cultured with self stigma leachate containing 2mM ZnS04 x 4
135. Self-pollen cultured with self stigma leachate containing 2mM CUS04 x 4
136. Self-pollen cultured with self stigma leachate containing 5mM ZnS04 x 4
137. Self-pollen cultured with self stigma leachate containing 5mM CUS04 x 4