review article possible future monoclonal antibody (mab)-based therapy against arbovirus
TRANSCRIPT

Hindawi Publishing CorporationBioMed Research InternationalVolume 2013 Article ID 838491 21 pageshttpdxdoiorg1011552013838491
Review ArticlePossible Future Monoclonal Antibody (mAb)-Based Therapyagainst Arbovirus Infections
Giuseppe Sautto Nicasio Mancini Giacomo GoriniMassimo Clementi and Roberto Burioni
Laboratorio di Microbiologia e Virologia Universita Vita-Salute San Raffaele 20132 Milano Italy
Correspondence should be addressed to Giuseppe Sautto sauttogiuseppehsrit
Received 10 May 2013 Revised 5 July 2013 Accepted 11 July 2013
Academic Editor Aldo Manzin
Copyright copy 2013 Giuseppe Sautto et alThis is an open access article distributed under theCreativeCommonsAttribution Licensewhich permits unrestricted use distribution and reproduction in any medium provided the original work is properly cited
More than 150 arboviruses belonging to different families are known to infect humans causing endemic infections as well asepidemic outbreaks Effective vaccines to limit the occurrence of some of these infections have been licensed while for the othersseveral new immunogens are under development mostly for their improvements concerning safety and effectiveness profiles Onthe other hand specific and effective antiviral drugs are not yet available posing an urgentmedical need in particular for emergencycases Neutralizing monoclonal antibodies (mAbs) have been demonstrated to be effective in the treatment of several infectiousdiseases as well as in preliminary in vitro and in vivo models of arbovirus-related infections Given their specific antiviral activityas well-tolerated molecules with limited side effects mAbs could represent a new therapeutic approach for the development ofan effective treatment as well as useful tools in the study of the host-virus interplay and in the development of more effectiveimmunogens However before their use as candidate therapeutics possible hurdles (eg Ab-dependent enhancement of infectionoccurrence of viral escape variants) must be carefully evaluated In this review are described themain arboviruses infecting humansand candidate mAbs to be possibly used in a future passive immunotherapy
1 Introduction
Arthropod-borne virus (arbovirus) infections are increas-ingly becoming an emerging medical problem mostly affect-ing endemic areas such as developing countries or upcomingeconomies (like China and India) In particular the majoroutbreak source of arbovirus-related diseases in endemicareas is mostly related to the presence of the viruses inan animal reservoir and a following expansion in humansMoreover epidemic episodes which occur mainly duringseasons with increased disease activity or outbreaks (egbecause of climate variations) have also been described Inaddition increasing traveling to exotic and medically high-risk locations has enlarged this problem also to previouslynon-endemic areas due to the global rise of travelers andmovement of large populations [1]
Of the over 545 suspected arbovirus species the mostknown virus-transmitting arthropods (vectors) are mosqui-toes (mostly femaleAedes aegypti andAedes albopictus) ticksmidges and sandflies Humans are usually dead-end hosts
as they do not develop the high viremia required to infectarthropods that is sustained by vertebrate animal reservoirs[1]
Although several arboviruses of clinical significance inhumans are known (more than 150) only a restricted group ofthem is globally diffused the majority of which are zoonoticand belong to the Flaviviridae Bunyaviridae or Togaviridaefamilies with a small number belonging to Reoviridae andOrthomyxoviridae [1] Highly effective vaccines for several ofthem are available including tick-borne encephalitis (TBEV)[2] yellow fever (YFV) [3] and Japanese encephalitis (JEV)viruses [4] but for no one of them a specific antiviral drugis currently approved for clinical use In the course of viralinfections neutralizing monoclonal antibody (mAb)-basedtherapy represents a promising and safe alternative strategyin particular when a specific and efficacious treatment is notyet available [5ndash11] At present human mAb-based passiveimmunotherapies for arbovirosis are at very early stage ofdevelopment However previous studies in mice have shownthat passive transfer of either monoclonal or polyclonal Abs
2 BioMed Research International
can be protective against homologous or heterologous denguevirus (DENV) challenge as well as against other flavivirusesand human arboviruses Moreover engineering renderingmAbs capable of crossing the blood-brain barrier in order tolimit viral dissemination within CNS may be considered
Finally a possible administration of mAbs in those sub-jects that could be at risk of exposure to arbovirus infectionssuch as travelers in endemic areas could reduce the possi-ble incidence and consequent augmented risk of epidemicepisodes
In this review we describe the major clinical relevant andworldwide diffused arboviruses infecting humans and therecentlymajor describedmAbs to be possibly used in a futurepassive immunotherapy
2 Flaviviruses
The Flavivirus genus including more than 70 viruses is theonly onewithin the Flaviviridae family which holds arboviru-ses that are responsible for significant morbidity and mortal-ity worldwide [12]
About 25 billion people are at risk of infection in tropicaland subtropical countries mainly South-East and SouthAsiaCentral and South America and the Caribbean In additionmultiple Flavivirus infections have been reported in the sameareas complicating early diagnosis and identification [13]
Flavivirus infections can cause fever encephalitis hem-orrhagic disease flaccid paralysis and death in humansHowever the immunopathogenesis of these viruses is notfully understood In the last decade the flaviviruses havereemerged as aggressive human pathogens [13]
The human flaviviruses includes 53 recognized speciesHowever five of them are considered clinically important likeDENV YFV JEV TBEV and West Nile virus (WNV)
The genome of all the members of the Flaviviridae familyconsists of a 95ndash125 kb positive-sense single-stranded RNAThey are enveloped small virions (40ndash60 nm in diameter)with two or more species of envelope glycoproteins (eg Mand E proteins) which are involved in the binding and fusionprocesses In particular the precursor of the mature M pro-tein (prM) interacts with E glycoproteins acting as a chaper-one and preventing the fusion of the viruswith themembranein the cell during egress through acidic compartments ofthe secretory pathway Then cleavage of prM by the cellularprotease furin during transit through the Golgi network is arequired step in the viral lifecycle that defines the transitionfrom an immature non-infectious virus particle into an infec-tious form However immature infectious virions retainingsome uncleaved prM molecules could be released [14]
The M and E glycoproteins constitute an icosahedralscaffold surrounding a nucleocapsid which consists of theviral genome complexed with a core of approximately 30 nmcomposed of multiple copies of a small basic capsid (C) pro-tein Binding uptake and fusion by target cells are believed toinvolve clathrin-mediated and low-pH-induced endocytosis[14]
Flaviviruses can utilize multiple receptors for differentcell types and host species They are thought to firstly inter-act with dendritic cells through DC-SIGN and L-SIGN
binding of glycans on E glycoprotein dimers In additionhighly sulfated glycosaminoglycans (eg heparan sulfate)have been demonstrated to play an important role in theinitial attachment of several flaviviruses to their target cellsOther molecules identified as possible receptors are inte-grins mannose-binding receptor on macrophages laminin-binding protein GRP78 (BiP) and CD14 [15]
Uncoating and replication of the viral genome througha minus-strand RNA intermediate occurs in the cyto-plasmic replication complexes associated with perinuclearmembranes where viral proteins are produced as part of asingle long polyprotein of more than 3000 amino acidsgenerating three structural (C prM and E) and sevennon-structural (NS1 NS2A NS2B the viral protease NS3NS4A NS4B and the RNA-dependent RNA polymeraseNS5) proteins by cleavage of host and viral proteases respec-tively Progeny virions are thought to assemble by bud-ding into an intracellular membrane compartment proba-bly the endoplasmic reticulum then transited through thehost secretory pathway and released at the cell surface[16]
Efforts to develop effective prophylactic approaches forseveral clinically important flaviviruses are underway [17]The crucial role of the humoral immune response againstFlavivirus infections is well established as infection withone serotype provides life-long protective immunity to thehomologous infecting serotype and cross-protection in thefirst few months against the other serotypes Converselyindividuals experiencing a secondary infectionwith a distinctserotype are at greater risk of severe complications asdiscussed later [18]
Only a limited number of Flavivirus vaccines are availabletoday however no approved antiviral drugs are yet availablefor their clinical use Thus giving the lack of vaccines aswell as specific antiviral drugs broadly cross-neutralizingmAbs could be helpful in the development of an effectivetherapeutic strategy against these infections as well as in theprogress towards effective immunogens
In the next paragraphs we describe the molecular targetsof flaviviruses including DENV JEV TBEV WNV YFV andSt Louis encephalitis virus (SLEV) followed by a descriptionof the therapeutic candidate mAbs directed against them
21 Domains and Functions of the Surface Envelope (E) Glyco-protein of Flaviviruses Themajor target of the host humoralimmune response and of neutralizing Abs against flavivirusesis represented by the envelope (E) glycoprotein which is a56-kDa protein and the major represented antigen on thesurface of virions [19] However Ab response towards otherstructural and non-structural proteins such as prM and NS1respectively has also been described [20 21] However prM-specific Abs display limited neutralizing activity while anti-NS1 Abs have been detected only during the convalescentperiod of a primary DENV infection but were stronglyidentified during the acute phase of a secondary DENVinfection suggesting that both Abs could contribute to thepathogenesis of the life-threatening dengue hemorrhagicfeverdengue shock syndrome (DHFDSS) [22]
BioMed Research International 3
The crystallographic structure of the E protein of TBEVhas been proposed as a model for the envelope protein of theflaviviruses [23]
Further studies performed throughX-ray crystallographyhave revealed that the E protein is a type II viral fusionprotein with three 120573-barrel domains (D) DI DII and DIII(corresponding to the antigenic domains C A and B resp)connected to the viral membrane by a helical structurecalled the stem anchor [24] In its native form the E proteinfolds as an homodimer with an antiparallel structure andan unusual herringbone pseudoicosaedral symmetry patternwith the M protein located centrally within the symmetryconsistent with a head-to-tail configuration lying parallel tothe envelope lipid bilayer The DIII of the E glycoprotein(amino acid residues 295ndash395) has an immunoglobulin-like fold contains seven 120573-strands as well as type- andsubcomplex-specific neutralizing B-cell epitopes and is theproposed receptor-binding domain through four peptideloops on the solvent exposed face Directly linked to DIIIthere is the stemtransmembrane region spanning aminoacid residues 401ndash495 and containing regions important foroligomerization with prM protein (amino acid residues 431ndash449 and 450ndash472) [25] DII has a long finger-like structureand contains two extended loops that project from DI andthe highly conserved and hydrophobic glycine-rich fusionloop at its tip (amino acid residues 98ndash110) interacting withthe stem region and the endosomal membrane of the cellduring the fusion process [26] In the trimeric conformationthe hydrophobic fusion peptide is exposed in DII to mediatethe fusion with the membranes of the cell DI is a sim120amino acid central domain formed by a discontinuous hingeregion consisting in a central 120573-barrel of eight strands thatconnectsDII andDIII andplays a crucial role in the structuralrearrangement of E from a homodimer to a trimer whichoccurs on exposure to low pH of endosomes that is requiredfor the membrane fusion process In some flaviviruses anN-linked glycosylation site is present at amino acid position154ndash156 of the E protein and does not appear to be necessaryfor E function but it has been associated with the increasedneuroinvasiveness of WNV lineage I outbreak strains fromthe USA and a pH-sensitive decrease in stability of the non-glycosylated strains [27]
22 Flavivirus Neutralizing B-Cell Epitopes As previouslydescribed Abs are a significant component of the hostrsquosprotective response against Flavivirus infections Howevervirus-specific Abs have been implicated in the pathogenesisof severe clinicalmanifestations following a secondaryDENVas well as YFV andWNV infections
Moreover cross-reactivity between Flavivirus serogroupscould complicate the interpretation of diagnostic assays butcould be of interest in isolating cross-reactive and cross-neutralizing mAbs However Ab-dependent enhancement(ADE) of infection has been described for this genus as itrepresents the major risk of complications when reinfectionoccursThus great caremust be taken in evaluating neutraliz-ing activity and possibleAb-mediated infection enhancementin the characterization of future possible therapeutic mAbsagainst these viruses [28]
However prophylactic and therapeutic use of neutral-izing mAbs for Flavivirus infections has been shown to beeffective in animal models as reported later
Neutralizing epitopes were found to be located in each ofthe three domains of the E protein and have been confirmedto be surface exposed in the high-resolution X-ray crystalstructure of the pre-fusion dimer of DENV2 E glycoprotein[29]
Most potentmAbs against flaviviruses are directed towardepitopes on DIII For this reason DIII-based immunogensare under evaluation as promising subunit Flavivirus candi-date vaccines [30 31] However several cross-reactive mAbsthat bind to residues from the AB loop were found to bepoorly neutralizing as this loop projects toward the lipidbilayer in the mature viral particle Conversely lateral ridgeregion of DIII (eg BC DE and FG loops) is targeted bystrong serotype-specific neutralizing mAbs [32ndash35] Cross-reactive mAbs specific for DII have also been described butwith less neutralizing and variable profiles However few ofthem whose epitope encompasses the fusion loop are cross-reactive and neutralizing [36]
23 Ab-Dependent Enhancement (ADE) of Infection In vitroand in vivo studies have demonstrated that increased diseaseseverity causing DHFDSS upon reinfection with a differentDENV serotype is mostly due to the phenomenon of ADE ofinfection determined by cross-reacting but poorly or non-neutralizing Abs generated during the primary infectionthat facilitate virus entry through Fc-120574 or complementreceptors present on cells such as monocytesmacrophages(Figure 1(a)) Additionally in 6 to 9 month-old children ofDENV immune mothers severe disease is associated withprimary infection possibly because of declining levels ofneutralizing maternal Abs [64]
These cross-reactive Abs are mainly directed against theE and prM glycoproteins In particular experimental lines ofevidence have suggested that cross-reactiveAbs againstDII ofE protein can under certain conditions enhance infectivityof WNV in vitro [65] However the phenomenon of ADEcould reflect the presence of non-neutralizing concentrationsof virus-reactive Abs and has been observed for severalflaviviruses in vitro and in vivo such as WNV TBEV JEVand YFV as well as other Flaviviridae members [28 66ndash68]In particular it has been described that ADE of infectionoccurs at virus-boundedAb concentrations in the upper limitby the stoichiometric threshold for neutralization and atlower concentrations by the minimal number of Abs thatallow attachment of the virion to cells Furthermore Abs thatrecognize infrequently displayed epitopes that do not supportneutralizationmay enhance infection even at saturation [69]
Thus the ADE of infection mechanismmay pose a threatfor the development of a safe and efficacious vaccine as wellas a possible immunotherapy against Flaviviridae
Indeed cross-neutralizing mAbs as candidate therapeu-tics for these infections may be considered after having wellascertained their broadly cross-neutralizing activity and theabsence of possible mAb-mediated ADE of infection mech-anisms Alternatively the risk of ADE of infection could beovertaken by the removal of the Ab heavy chain (ie the CH2
4 BioMed Research International
receptorFc-120574
Cellular receptor
(a)
Cellular receptor
(b) (c)
Figure 1 Mechanisms of antibody-dependent enhancement (ADE) of infection (a) After binding to the viral epitope the Ab is recognizedby a cellular Fc-120574 receptor bringing the viral particle in proximity of the target cell (b) the binding of the Ab induces conformational changeswithin the structure of the viral target protein These changes improve the affinity for the cellular receptor (c) molecular mimicry by a viralmotif of cellular membrane components leads an autoreactive Ab to bind both the viral and the cellular target bringing the virus in proximityof the target cell
and CH3 portions or direct expression of the mAb as aFab fragment) andor deletion of the N-linked sugars onIgG molecules that are both required for interactions withFc-120574 receptors (Fc-120574R) or eventually by the blocking ofFc-120574R engagement with anti-Fc-120574R Abs [70] Similarly theidentification and detailed examination of cross-neutralizingepitopes that do not promote ADE of infection may definenovel targets for vaccine development
However in some instances ADE of infection mayoccur when Ab molecules cross-react with both viral andcellular antigens (Figure 1(c)) In this regard Fc-120574 receptor-independentmechanismof infection in particular forDENVprM-specific Abs able to bind simultaneously to the virus andtarget cells has been described Indeed othermembers of theFlaviviridae belonging to another genus like hepatitis C virus(HCV) have been shown to elicit polyreactive Abs that areprobably responsible for an ADE of infection mechanism aswell as of secondary clinical manifestations In connectionwith this several studies employing both polyclonal andmonoclonal Abs suggested the presence of common epitopescontaining particular short motifs on NS1 and the threedomains of the E protein as well as human proteins thatmay play a role in ADE of infection mechanisms and thusin DHFDSS pathogenesis [70] Interestingly these commonmotifs have been found to be additionally represented inDENV2 strains which have greater human pathogenic capac-ities [22]
Moreover another molecular mechanism of ADE ofinfection could consist in the facilitation of conformationalchanges of the targeted surface protein that are required forvirus entry as described and hypothesized for other viruses(eg HIV HCV and HSV) (Figure 1(b)) [71 72]
Finally Fc-120574R engagement could result also in ldquointrinsicrdquoADE of infection mechanisms involving signaling eventssuch as inhibition of antiviral response and increased viralreplication Disrupted signaling events include the RIG-IMDA5 cascade and type I interferon (IFN) response aswell as induction and suppression of anti-inflammatory(IL-10) and proinflammatory (IL-12 and TNF-120572) cytokinesrespectively [73]
24 Dengue Virus (DENV) Dengue virus (DENV) is respon-sible for 50ndash100 million symptomatic infections each yearresulting in 500000 hospitalizations and over 20000 deathswhich occurs mostly in tropical and subtropical regions ofthe world Although malaria remains the most importantcause for systemic febrile disease in travelers chikungunyavirus (CHIKV) and DENV are increasingly diagnosed withdengue currently being the second most important cause forfebrile disease in travelers
Decreases in mosquito control efforts during the endof the 90s coupled with societal factors (eg globalizationmigrations and dense urbanization) have contributed to thereemergence of flaviviruses such as DENV in South and Cen-tral America Development of effective DENV vaccines thatexhibit cross-protection thought to be important for prevent-ing subsequent dengue-associated immunopathogenesis isproving to be particularly challenging [1]
A vaccine for this infection is not yet available despiteconsiderable public and private efforts This difficulty ismainly due to the perceived need to simultaneously protectthe four known serotypes of DENV (DENV1ndash4) which shareabout 70 of sequence homology while genotypes can varyup to 3 Moreover as anticipated and discussed previously
BioMed Research International 5
non-protective Abs may contribute to more severe clinicaloutcomes in vaccinated people [74]
241 Murine versus Human Anti-DENV Humoral ImmuneResponse It is well ascertained that anti-E DIII-directed Absare virtually absent from the naıve human repertoire as theyare directed away from this domain probably against weaklyneutralizing and immunodominant regions Moreover it hasbeen demonstrated that over than 90 of the human Abresponse of primary DENV infected patients is able to bindonly native DENV virus particles instead of a recombinantform of the E protein posing difficulties in the cloning strate-gies of neutralizing mAbs [39] Thus it can be concludedthat in vivo only a small fraction of DENV-specific Abs areresponsible for neutralization
Conversely murine mAbs that recognize all the threedomains of E have been identified Epitope-mapping studiesusingmurine serotype-specificmAbs have demonstrated thatloops located in the lateral ridge region of DIII consti-tute the strongest neutralizing B-cell target In this regionsequence diversity between serotype is high while cross-reactive murine mAbs that recognize another loop showedgenerally a weaker neutralization profile In particular type-specific mAbs with neutralizing activity against DENV2localized to the BC DE and FG loops on the lateral ridgeof DIII whereas subcomplex-specific mAbs recognized anadjacent epitope centered on the connecting A strand of DIIIat residues K305 K307 and K310 [32ndash35]
Cross-reactive mAbs directed against prM and NS1 havealso been described but as anticipated they feature weaklyneutralizing or absent as well as ADE of infection activityIn the following paragraphs we report the better character-ized neutralizing mouse and human mAbs describing theirmolecular features further summarized in Table 1
242 Anti-DENV Murine mAbs Murine mAbs directedagainst all the three domains of the E protein and endowedof different neutralizing and binding characteristics havebeen described However only DIII and DII are classicallyrecognized by neutralizing Abs In this regard Sukupolvi-Petty et al isolated twenty-four anti-DENV2 mouse mAbswith moderate or strong neutralizing activity against thehomologous virus in cell culture assays Binding sites weremapped for the majority of these in distinct epitopes inregions located in DI (lateral ridge) DII (dimer interfacelateral ridge and fusion loop) and DIII (lateral ridge CC1015840loop and A strand) Moreover 16 of the neutralizing mAbswere tested in mice with most of them being protectivewhen given as prophylaxis Seven of these had post-exposuretherapeutic activity when administered as a single dose byintraperitoneal route even 3 days after intracranial infectionFor the mAbs with the greatest therapeutic potential protec-tion was confirmed with an Ab-enhanced vascular leakagemouse model of DENV2 infection [35]
Recently Deng et al described a cross-reactive murinemAb named 2A10G6 that is able to recognize DENV1ndash4 YFV WNV JEV and TBEV to potently cross-neutralizeDENV1ndash4 YFV and to a lesser extent WNV This mAbrecognizes a highly conserved motif (amino acid residues
98ndash101) located within the fusion loop of the FlavivirusDII of the E glycoprotein Moreover this mAb exerts itsneutralizing activity in a post-attachment step during thevirus entry process as demonstrated by kinetic neutralizationtests performed in vitro Additionally protection experimentsperformed in mouse models showed that treatment with100 120583gmL of 2A10G6 conferred full protection against lethalDENV2 challenge and 20 120583gmL and 4120583gmL of 2A10G6protected 89 and 40 of infected mice from lethal chal-lenge respectively For infection with DENV1 3 and 4 use of100 120583gmL of 2A10G6 conferred partial protection and 5377 and 73 of the infected mice survived after challengerespectively Finally the protection profile of 2A10G6 againstWNV showed that prophylactic administration with a singledose of 200mg of 2A10G6 conferred 80 protection in miceMost importantly 3 of 8 (375) mice survived when 2A10G6was administered one day after WNV challenge SimilarlyanothermousemAb 4G2 is able to recognize the fusion loopat the extremity of DII from all four DENV serotypes and toprevent syncytia formation [36]
Rajamanonmani et al described a mouse mAb 9F12raised against theDIII of DENV2E protein that is able to rec-ognize DENV1ndash4 serotypes as well asWNV and to neutralizefive DENV strains representative of all DENV serotypes [41]Similarly Cockburn et al reported a comparative high-resolution crystallographic analysis of an A-strand DIIImurinemAb 4E11 in complex with its target domain of the Eprotein from the four DENV serotypes MAb 4E11 is capableof recognizing and neutralizing all four serotypes with IC50values varying between 1 and 100 nMThe structures reportedalso highlight the mechanism by which anti-A-strand mAbsdisrupt the architecture of the mature virion inducing dimerdissociation premature fusion-loop exposure and concomi-tant particle inactivation [40] Also Midgley et al function-ally characterized another murine mAb 2H12 raised in miceagainstDIII of E protein Similarly to the previously describedmAbs 2H12 is able to bind all the four DENV serotypes ina epitope encompassing the conserved 314ETQH317 motifHowever the neutralizing potential of 2H12 is lower than anumber of other anti-DIII mAbs with IC50 values rangingfrom 056 to 145 nM for DENV1 2 and 4 On the other handit showed no ADE of infection activity [38]
Austin et al isolated a murine mAb named E111 whichrecognize a novel CC1015840-loop epitope on DIII of the E proteinfrom two different DENV1 genotypes Docking of the mAbstructure onto the available cryoelectron microscopy modelsof DENV virions revealed that the E111 epitope was inaccessi-ble suggesting that this mAb recognizes an uncharacterizedvirus conformation While the affinity of binding betweenE111 and DIII varied by genotype a limited correlation withneutralizing activity was observed These data support theconclusion that potent neutralization depends on genotype-dependent exposure of the CC1015840 loop epitope [49] In fact aspreviously described elsewhere binding of some E reactiveAbs depends on the dynamicmovement of proteinmoleculesldquobreathingrdquo in the virion particle leading to transient expo-sure of hidden epitopes For instance optimal binding ofmousemAb 1A1D-2 to EDIII requires incubation at 37∘C [75]
6 BioMed Research InternationalTa
ble1Schematicsummaryof
theb
estcharacterized
mAb
sagainstarbo
viralp
atho
gens
mAb
Orig
inRe
activ
ityTargetprotein
Epito
peClon
ingstr
ategy
Form
atisotype
Neutralizingactiv
ityRe
ference
48A
Hum
anDEN
V1ndash4
EDII
EBVtransfo
rmation
IgG
IC50
(120583gmL)
DEN
V115
DEN
V2gt40
DEN
V324
DEN
V4gt40
[37]
D11C
IC50
(120583gmL)
DEN
V115
DEN
V210
DEN
V3102
DEN
V416
16D
IC50
(120583gmL)
DEN
V115
DEN
V202
DEN
V305
DEN
V427
2H12
Murine
DEN
V1ndash4
EDIII(ABloop
aa
314ndash
317)
Hybrid
omafrom
BALB
cmou
seim
mun
ized
with
DEN
V2EDIII
IgG2b
IC50
nM
DEN
V1056ndash54
DEN
V329
DEN
V4145
[38]
C9Murinechim
eric
DEN
V2
EDIII
Phaged
isplayof
achim
ericmurine
hybridom
alibrary
VH1V1205811
PRNT5
0850120583
gmL
[39]
4E11
Murine
DEN
V1ndash4
EDIII(str
andA308
312andstrand
G387
389391)
Hybrid
oma
IgG2a120581
IC50
(120583gmL)
DEN
V1016
DEN
V2013
DEN
V-38
DEN
V-415
[40]
4G2
Murine
DEN
V1ndash4
EDII(fu
sionloop
)Hybrid
omafrom
DEN
V2Eim
mun
ized
mice
IgG
PRNT5
0(120583gmL)
DEN
V215
[39]
9F12
Murine
DEN
V1ndash4WNV
EDIII(aa
305307310
330Astr
andandBC
loop
)
Hybrid
omafrom
BALB
cmou
seim
mun
ized
with
DEN
V-2EDIII
IgG1k
DEN
V1ndash4P
RNT5
02sdot10minus8ndash2sdot10minus7M
[41]
2A10G6
Murine
DEN
V1ndash4YF
VWNV
JEV
TBEV
EDII(Fusionloop
aa
98ndash101)
Hybrid
omafrom
BALB
cmou
seim
mun
ized
with
inactiv
ated
DEN
V2
IgG1
PRNT5
0(120583gmL)
DEN
V12
DEN
V215
DEN
V321
DEN
V418
YNF36
WNV
46
[36]
mAb
11Hum
anDEN
V1ndash4WNV
SLEV
YFV
JEV
MVEV
EDII(fu
sionloop
)Ph
aged
isplayof
human
scFv
Fusio
nprotein
scFv-Fc
PRNT8
0(120583gmL)
WNV
125
DEN
V2625
[4243]
BioMed Research International 7Ta
ble1Con
tinued
mAb
Orig
inRe
activ
ityTargetprotein
Epito
peClon
ingstr
ategy
Form
atisotype
Neutralizingactiv
ityRe
ference
E16
(MGAW
N1)
Murine
WNV
EDIII(LR
aa3
02ndash309)
Hybrid
omafrom
immun
ized
micew
ithWNVE
IgG2bhum
anized
PRNT5
04ndash
18ng
PRNT9
053ndash297
ng[4445]
1A1D
-2Murine
DEN
V1ndash3
EDIII(Astr
andaa
307
310and312)
Hybrid
omafrom
immun
ized
micew
ithdifferent
pH-tr
eated
virus
IgG2a
DEN
V2PR
NT5
021n
M[4647]
1F4
Hum
an
DEN
V1
E(virion
)
DI-DII
Electro
fusio
nof
infected
mem
oryBcells
from
DEN
V-im
mun
esub
jects
with
EBV
IgG
DEN
V1IC5
0011120583gmL
[48]
2D22
DEN
V2
DIII
DEN
V2IC50
008120583gmL
5J7
DEN
V3
DI-DII
DEN
V3IC50
010120583gmL
E105
Murine
DEN
V1
E
DIII(BC
loop
G328
T329
andD330DE
loop
K361E
and
E362K
FGloop
K3
85)
Hybrid
omas
deriv
edfro
mC5
7BL6
IFN-120572120573Rminus
minusmice
infected
with
DEN
V1
IgG
PRNT5
0DEN
V-1
05ndash592ng
mL
[32]
E106
DIII(BC
loop
G328
T329
andD330DE
loop
K361E
and
E362K
FGloop
K3
85A
strand
S305
K307E
309K3
10and
E311)
PRNT5
0DEN
V-1
06ndash
592ng
mL
E111
DIII(CC1015840loop
)PR
NT5
038ndash25120583gmL
[49]
CR4374
Hum
anWNV
EDIII
Phaged
isplayof
scFv
IgGlib
rary
VH2-05V
L1e
PRNT5
0018120583gmL
PRNT9
0095120583gmL
[50]
CR4353
VH3-30V
k3-A27
PRNT5
00026120583
gmL
PRNT9
0364120583gmL
1A5
Chim
panzee
DEN
V1ndash4WNV
JEV
LGTV
EDII(aaG
106)
Phaged
isplayof
Fab
library
from
DEN
V1ndash4
infected
chim
panzees
Hum
anized
IgG1
PRNT5
0(120583gmL)
DEN
V1048
DEN
V2095
DEN
V332
DEN
V443
WNVD
ENV438
JEV
21LG
TV28
[51]
8 BioMed Research International
Table1Con
tinued
mAb
Orig
inRe
activ
ityTargetprotein
Epito
peClon
ingstr
ategy
Form
atisotype
Neutralizingactiv
ityRe
ference
mAb
11Hum
anDEN
V1ndash4WNV
EDII(fu
sionloop
W101G104G106)
scFv
library
scFv-Fc
PRNT8
0(120583gmL)
DEN
V2625
WNV
125
[4243]
3B4C
-4
Murine
SLEV
E
1a
Hybrid
oma
IgG
SLEV
PRNTlt17
[5253]
1B2C
-5SLEV
1bSLEV
PRNTlt17
6B5A
-2SLEV
1cSLEV
PRNT
48
4A4C
-4SLEV
1dSLEV
PRNT
29
1B5D
-1SLEV
JEV
2SLEV
PRNTlt17
2B5B
-3SL
EVJEV
MVEV
WNV
YFV
3SLEV
PRNT
23
2B6B
-2AllFlaviviru
s4a
SLEV
PRNTlt17
6B6C
-1AllFlaviviru
s4b
(inDII)
SLEV
PRNT
23
A3
Chim
panzee
JEV
E2
DIa
aK1
79Ph
aged
isplay
Hum
anized
PRNT5
0004ndash0
2nM
[54]
B2DIIaaI126
Phaged
isplay
PRNT5
0002ndash2
nM
E3DIIIa
aG132
Phaged
isplay
PRNT5
0014ndash0
93n
M
FabT
JE12B0
2Hum
anJEV
ENA
Phaged
isplay
Fab
FRNT5
0502120583gmL
[55]
5F10
Hum
anCH
IKV
E2Dom
ainB
EBVtransfo
rmation
IgG11205822
IC50lt100n
gmL
[56]
8B10
Hum
anCH
IKV
E1-E2
E2Dom
ainA
EBVtransfo
rmation
IgG1k
IC50lt100n
gmL
CHK-
152
Murine
CHIK
VE2
aa59
Hybrid
oma
IgG2c
IC50
1ndash3n
gmL
[57]
11E7
Murine
CCHFV
Gn
C-ter
Hybrid
oma
IgG
PRNT8
0diluted
12560
[58]
4-39-C
CMurine
RVFV
G2
Dom
ainIV
Hybrid
oma
IgG
PRNT8
0diluted
1204
80ndash181920
[59]
2C9
Murine
YFV
EHybrid
oma
IgG2a
PRNT9
011
0417D
1105
2Asib
i[60]
5A 7A R3(27)
Hum
anYF
VE
DI-DII
Phaged
isplay
ScFv
PRNT9
001ndash03120583
gmL
[61]
13D6
Murine
TBEV
EDIII
Hybrid
oma
IgGchimeric
PRNT5
019120583gmL
FRNT5
045120583
gmL
IC50
19ndash167120583
gmL
[62]
3B4C
-4(H
y4-26C
)Murine
VEE
VE2
aa182ndash209
Hybrid
oma
IgGhum
anized
PRNT7
0(120583gmL)
394ndash125
[63]
BioMed Research International 9
The structure of the 1A1D-2 bound to EDIII indicates that themAb binds to sites that are transiently exposed during viralldquobreathingrdquo at 37∘C and block infection during attachmentof the virion to the cell In particular this mAb neutralizesDENV1 2 and 3 serotypes and residues K307 and K310 arethemost critical residues for binding of 1A1D-2mAb [46 75]
Finally Shrestha et al immunized mice with a genotype2 strain of DENV1 virus and generated 79 new mAbs 16of which strongly inhibited infection by the homologousvirus and localized to DIII of E protein Surprisingly onlytwo mAbs E105 and E106 retained strong binding andneutralizing activity against all five DENV1 genotypes as wellas being protective in immunocompromised infected miceMoreover E105 and E106 exhibited therapeutic activity evenwhen administered as a single dose four days after inoculationwith a heterologous genotype 4 strain of DENV1 in the samemouse model [32]
243 Other Non-HumanmAbs Other non-human and non-murine as well as humanized mAbs directed against the Eglycoprotein have been described In this regard Goncalvezet al isolated a large panel of anti-E Fab fragments from chim-panzees infected with all four DENV serotypes Howeveronly a limited number of them displayed a cross-neutralizingactivity against DENV1 and 2 and to a lesser extent againstDENV3 and 4 In particular the authors calculated thatamong them the 1A5 mAb that was further humanizedneutralized DENV1ndash4 at a PRNT50 titer of 048 095 32and 43 120583gmL respectively Interestingly the humanized 1A5was also tested for binding and neutralization against theWNVDENV4 chimera JEV strain SA14-14-2 and Langatvirus (LGTV) strain TP 21 giving a PRNT50 titer of 38 21and 28120583gmL respectively Moreover the authors calculatedthat when administered in a dose of 2mg per kg of bodyweight 1A5 would give a serum titer of approximately 40and 20 50 reduction in plaque reduction neutralization test(PRNT50) against DENV1 and DENV2 respectively [76]Furthermore epitope mapping of 1A5 mAb localized to G106within the Flavivirus-conserved fusion loop inDII of DENV2E protein [51]
244 Anti-DENV Human mAbs As anticipated attemptsto isolate neutralizing human mAbs have been morechallenging due to the restricted elicited Ab-repertoire whichrecognizes DIII of E glycoprotein in naıve as well as ininfected patients [77 78] However several groups reportedthe isolation of neutralizing mAbs from infected subjects Inthis regard Setthapramote et al isolated a total of 136 humanhybridoma clones producing specific mAbs against DENVobtained using PBMCs from nine blood samples from fouracute-phase patients secondarily infected with DENV2 andfive convalescent-phase patients Interestingly the authorsfound that most of the acute-phase mAb clones were cross-reactive with all four DENV serotypes with most of themrecognizing the E protein and endowing of neutralizingactivity against all DENV serotypes compared to thosederived from convalescent-phase patients In particularfrom the acute-phase PBMCs 818 were anti-E 66 wereanti-prM and 33 were anti-NS1 while 133 anti-E 133
anti-prM and 533 anti-NS1 mAbs clones were obtainedfrom convalescent-phase PBMCs [79] Previous studiesconfirmed the data obtained by Setthapramote et al onconvalescent-phase patients but this group firstly reportedthe efficient preparation of human mAbs with strongneutralizing activity titers against all four DENV serotypesusing PBMCs from acute-phase patients secondarily infectedwith DENV [80 81]
The group of de Alwis through immunoglobulin deple-tion studies reported that a substantial fraction of DENV-reactive Abs in human immune sera including type-specificneutralizing Abs bound to the intact virion but not torecombinant E proteinThe authors confirmed these observa-tions also isolating human neutralizing mAbs and proposedthat humans produce Abs that neutralize DENV infectionby binding a complex quaternary structure epitope that isexpressed only when E proteins are assembled on a virusparticle [48] Similar findings have been described for severalother viruses including WNV and HIV [82 83] Mappingstudies indicate that this epitope has a footprint that spansadjacent E protein dimers and includes residues at the hingebetween domains I and II of E protein [48]
Recently Costin et al isolated three broadly neutralizinganti-DENVhumanmAbs named 48AD11C and 16DThesemAbswere isolated from three different convalescent patientswith distinct histories of DENV infection All three mAbsrecognized the E glycoprotein with high affinity neutralizedall four serotypes of DENV but mediated ADE of infection inFc receptor-bearing cells at subneutralizing concentrationsMapping studies revealed that all three mAbs bind discontin-uous epitopes within the highly conserved fusion-loop regionof DII (contacting residues W101 L107 andor G109) [37]
25 Yellow Fever Virus (YFV) TheWHO estimates that thereare 200000 cases of yellow fever virus (YFV) infections and30000 related deaths every year especially in Sub-SaharanAfrica In fact different to themajority of human arbovirusesand similarly to DENV and CHIKV YFV has expanded itshost range to include humans as an amplifying host [1]Symptoms occurring from 3 to 6 days after infection withYFV consist mostly in fever chills anorexia lumbosacralpain nausea and vomiting This syndrome lasts about 3days in most cases and is sometimes followed by a one-daylasting period of remission Fever will reoccur together withdark hematemesismelena petechiae and other hemorrhagicsymptoms Convalescence is characterized by deep asthenialasting up to two weeks Rarely prognosis can be fatal Aneffective live-attenuated vaccine (17D) derived from the Asibistrain by serial passages in chicken embryos is available [84]
251 mAbs against YFV In 1983 Schlesinger et al produceda battery of mAbs after immunization of BALBc mice byinjection of 17D YFV vaccine strain [84] Among them thespecificity towards the E and NS1 proteins was describedeven though anti-NS1 Abs were not able to show neutralizingactivity The 13 IgG and 1 IgM anti-E protein mAbs wereclassified in five groups according to their specificity GroupA consisting of the only IgM produced in this experiment
10 BioMed Research International
could neutralize 17D YFV virus only group B 17D and Asibistrains only group C Asibi strain only group D could onlyneutralize Asibi but not 17D strain showing also reactivityagainst DENV2 Zika or Banzi viruses All of the four mAbsbelonging to this group were able to cross-react with Zikaandor Banzi viruses and two (4E11 and 5H3) neutralizedDENV2 virus with 4E11 neutralizing also Banzi virus The3E9 mAb the only component of the group E could notneutralize 17D nor Asibi strains
All of the IgG mAbs resulted able to protect bothprophylactically and therapeutically BALBc or CD-1 micefrom lethal intracerebral challenge with 17D-204 strain [85]The chimeric form of the group B mAb 2C9 IgG (2C9-cIgG) was able to provide AG129 mice a 72 survival whenadministered 24 hours before infection with 17D-204 strainIts murine form could provide the survival of the 95 ofthe mice Appreciable results were also obtained when bothmurine 2C9 and 2C9-cIgG were administered 24 hours post-infection with the survival of 70 and 20 of mice respec-tively Viral presence was not detected in survivingmice [60]
In 2005 Daffis and colleagues constructed two Ab-phage libraries by cloning the repertoire of YFV-infectedpatients Panning was then performed with YFV-17D virionsThe scFv-5A 7A and R3(27) showed a neutralizing activityspanning from50 to 100 inPRNTassays against bothYFV17D-204-WHO and Asibi strains In further tests reactivitywas observed against wild-type YFV strains of West Africagenotype I and II (Nigeria 1987 and Asibi strains resp)and EastCentral Africa (CAR 1986 Ethiopia 1961 strains) Aconcentration from 01 to 03120583gmL could yield a 90 plaquereduction Reactivity was also observed against the strainSenegal 1990 in a lesser extent Production of escape variantscould demonstrate that scFv-5A 7A and R3(27) epitopes arebuilt up by residues extensively separated in themonomers ofE glycoprotein that however result in closed proximity whenthe homodimeric form of E is constituted [61]
26 West Nile Virus (WNV) The West Nile virus (WNV) isan epidemic neurotropic virus estimated to be responsibleof about 36000 cases and 1500 deaths registered in theUnited States between 1992 and 2012 [86] WNV antigenicityallows its classification into the Japanese encephalitis virus(JEV) serocomplex Genomic analysis has revealed two maingenetic lineages of WNV lineage I viruses circulating in theUSA Europe the Middle East Africa India and Australiaand lineage II viruses isolated from Sub-Saharan Africa andMadagascar [1]
WNV was firstly identified in 1937 and is endemicin many countries of Africa Middle East and West AsiaHowever after 1990 frequent outbreaks of WNV infectionswere reported in Romania Israel and later in North Americaand across the USA in 1999 and recently in 2012 [87]
Most of WNV infected individuals do not develop symp-toms while about 20 develop a self-limiting illness calledWest Nile fever Acute symptoms include fever tirednessand swollen lymph glands but in a minority of cases alsoencephalitis with long-term deficits in cognitive functionand motor skills have been reported However in less than
1 of infected patients WNV having crossed the blood-brain barrier is responsible of neuroinvasive and potentiallylethal form of the disease In these cases degeneration andapoptosis upon infection of neurons and the consequentinflammatory response can occur Related symptoms includehigh fever coma muscle weakness and paralysis Immun-odepression and advanced age have been correlated with anaugment risk to develop a severe disease [87] There are nospecific treatment options or licensed vaccines for humans[88]
WNV is transmitted by infected mosquitos and initialreplication is thought to occur primarily in dendritic cells inthe skin which migrate to secondary lymphoid tissues wherethe replicating virus enters the circulation To date onlyefficacious WNV veterinarian vaccines have been licensedwhile there are no licensed vaccines for protection againstWNV in humans [88]
261 mAbs against WNV Broad-spectrum antivirals suchas type I IFN-120572 ribavirin mycophenolic acid in WNVinfection showed ineffective results in vivo despite in vitrothey showed some activity [89]
However experiments in murine models extrapolationof clinical data as well as passive administration of pooledimmune-120574-globulins (OmriGam) containing a significanttiter of neutralizing Abs before and after infection showedan important role of the humoral immune response incontrolling viremia and prevent viral dissemination Indeedthe development of a neutralizing mAb-based therapy seemsto be encouraging for a possible treatment of infected patients[90ndash92]
Similarly to what has been observed for DENV only anti-E Abs have been identified as neutralizing and protectiveAlso anti-NS1 Abs have shown a protective role howevertheir mechanism of protection has not yet been elucidated asthe NS1 protein is secreted from infected cells and not presenton the virions [93ndash95]
In this regard similarly to what has been describedfor DENV studies performed using a naıve human scFvlibrary for panning with purified WNV E protein supportthe hypothesis that no Abs against the neutralizing DIII canbe isolated However DIII specific Abs were isolated in asubsequent study using immunoglobulin libraries obtainedfrom threeWNV infected patients for biopanning on purifiedinactivated virus virus-like particles consisting of prM andE proteins or recombinant E glycoprotein Although theproportion of DIII-specific mAbs was low (8) comparedwith anti-DII mAbs (47) Two out of the four anti-DIIImAbs were potently neutralizing and protective in vivowhereas only three out of the 24 anti-DII were weaklyneutralizing in vitro and non-protective in vivo In particularresidues that are critical for neutralization lies on the regionsspanning amino acids 305ndash312 330ndash333 and 365 that arelocated on adjacent exposed loops ofDIIIHowever sequencealignment of E protein of different flaviviruses such as DENVand JEV revealed a considerable variation compared tothe whole E protein This observation further suggests thatdifferently from broadly cross-reactive anti-DII Abs anti-DIII neutralizing Abs are virus-type specific However of
BioMed Research International 11
these mAbs two (CR4374 and CR4353) protected mice fromlethal WNV challenge at 50 protective doses of 129 and357 120583gkg of body weight respectively [50]
Sanchez et al using three different immunization strate-gies (ie inactivated virus naked DNA and recombinantprotein) isolated ninemurinemAbsmost of which bound toconformation-dependent epitopes in DIII of the E protein Inparticular neutralizing mAbs named 8B10 11C2 10C5 and17C8 were obtained from mice immunized with inactivatedvirus alone or in combination with a DNA plasmid andbound to the same region of DIII with high affinity Incontrast mAbs obtained by immunization with a solubleversion of the E glycoprotein did not exhibit neutralizingactivity These non-neutralizing mAbs were cross-reactivewith several other flaviviruses including SLEV JEV YFVand Powassan virus confirming the conserved nature ofFlavivirus non-neutralizing epitopes [96]
Gould et al isolated 11 unique human single-chain vari-able region Ab fragments (scFvs) that bind the E protein ofWNVAmong them a humanmAb namedmAb11 expressedas a scFv-Fc fusion protein was further characterized Itrecognizes the fusion loop at the distal end of DII ofthe WNV E protein and cross-reacts with all four DENVserotypes and provides protection against DENV2 and 4as well as WNV [42] Moreover therapeutic studies of thismAb in WNV-infection model mice provided substantialprotection when administered after 5 days post-infectionInterestingly a neutralization escape variant of this mAbfailed to cause lethal encephalitis (at higher infectious doses)or induce the inflammatory responses associated with blood-brain barrier permeability in mice compared to the parentalWNV suggesting an important role for the fusion loop inviral pathogenesis [43]
Oliphant et al isolated an anti-DIII E protein mAbnamed E16 fromhybridomas obtained after immunization ofmice with recombinant WNV E protein which neutralizedall WNV strains with PRNT50 values of 4 to 18 ng andPRNT90 values of 53 to 297 ng One hundred microgramsof mAb protected greater than 90 of mice from lethalinfection and even a single 4120583g treatment of E16 on day 2 afterinfection prevented mortality Moreover humanization ofthis mAb confirmed as therapeutically effective in mice [44]Subsequent studies revealed that mAb E16 neutralization ismediated by engagement of four discontinuous segmentsof DIII including the amino-terminal region (amino acidresidues 302ndash309) and the three connecting loops BC (aminoacid residues 330ndash333) DE (amino acid residues 365ndash368)and FG (amino acid residues 389ndash391) Moreover no ADE ofinfection was detected when E16 mAb was used at saturatingconcentrations [45 47]
Furthermore results of a Phase I safety study of thehumanized E16 mAb (designated MGAWN1) have beenreported and suggested that doses of up to 30mgkgwerewelltolerated with few mild adverse events and would providean excess of virus neutralizing activity for 3-4 weeks aftertreatment However a case of anti-MGAWN1 Ab elicitationoccurred with the consequent increased rate of clearance andindeed impacting efficacy A Phase II safety and efficacy studyof MGAWN1 is ongoing [97] Furthermore the possibility of
preventing or treating WNV-induced memory deficits wasrecently investigated In this study hamsters were treatedintraperitoneally with 32mgkg ofMGAWN1mAb at 45 daysafter subcutaneously challenging with WNV InterestinglyMGAWN1 prevented mortality weight loss and improvedfood consumption of WNV-infected hamsters compared tocontrols [98]
Recently Lelli et al isolated six anti-E mAbs frominactivated-WNV immunized mice In particular three ofthem (3B2 3D6 and 4D3) neutralized lineage I and II WNVwith the first two recognizing the same epitopes located onthe distal lateral surface of DIII (critical amino acid residueK307) Conversely 4D3mAb recognized a novel neutralizingepitope on DII (critical residues S276 and T278) Indeedfurther protective and therapeutic studies are needed toascertain their neutralizing activity in vivo [99]
Finally to conclude like DENV infection Abs directedagainst the M protein are not protective and neutralizingMoreover in humans a skewed humoral immune responseagainst DII has been frequently observed and confirmedalso with the hybridoma technology [50] The isolation andelicitation of neutralizingAbs directed against the fusion loopand DIII of the E protein represent thus the most challengingand promising goal for the development of new effectivetherapeutic inhibitors and immunogens respectively
27 Tick-Borne Encephalitis Virus (TBEV) Tick-borne ence-phalitis virus (TBEV) is one of the most dangerous agentscausing human neuroinfections occurring mostly in Europeand Asia and with a potential fatal prognosis [100] TBEV isbelieved to cause 3500ndash10000 human cases of encephalitisin Europe per year with a high morbidity in Russia CzechRepublic Austria and Germany In particular between 1990and 2007 an average of 8755 cases of TBE was reported peryear in Europe and Russia Despite the fact that Russia isthe country with most infections registered annually CzechRepublic incidence is among the highest in Europe with 400ndash1000 clinical cases reported every year [101]
Three subtypes of TBEV are classified namely EuropeanSiberian and Far-Eastern sharing most of the genetic andantigenic features [102] In fact a high degree of antigenichomogeneity between different strains of TBEV has beendescribed [103]
Clinical manifestations vary among the subtypes butusually they start with a short febrile period of 7ndash14 daysafter the tick bite Fatigue headache and pain in the neckshoulders and lower back together with high fever andvomitingmay be present [100]Thesemanifestations are oftenfollowed by an asymptomatic phase lasting from 2 to 10days after remission from the fever with possible progres-sion to neurological disease Neurological symptoms includemeningitis encephalitis myelitis and radiculitis Mortalityoccurs in 1-2 of the European subtype-infected patientsbut fatal prognosis can occur in up to 20ndash40 of the Far-Eastern subtype-infected patients [104] Mortality rates ofthe Siberian subtype are similar to those observed for theEuropean [105]
Effective and safe vaccines against TBEV produced frominactivated virus have been developed and licensed for their
12 BioMed Research International
use in humans However an emergency therapy in theabsence of a mass immunization is needed as no effectivetreatments are yet available [106]
271 mAbs against TBEV Levanov et al described the chim-erization of two murine mAbs (13D6 and 10C2) directedagainst the DIII and DII respectively of the TBEV E glyco-protein The chimeric mAbs present binding characteristicssimilar to the parentalmAbsMoreover as the parentalmAbsonly the chimeric mAb 13D6 was able to neutralize TBEVinfectivity in vitro In particular neutralization studies withthe murine chimeric and scFv forms of mAb 13D6 were per-formed In particular murine 13D6 showed an IC50 titer of115 120583gmL in Focus Reduction Neutralization Tests (FRNT)against TBEV strain 205 and of 29 120583gmL in PRNT againstTBEV strain Softjin Chimeric 13D6 showed an IC50 titer of45 120583gmL in FRNT against TBEV strain 205 and of 19 120583gmLin PRNT against TBEV strain Softjin ScFv 13D6 showed anIC50 titer of 167 120583gmL in FRNT against TBEV strain 205and of 112 120583gmL in PRNT against TBEV strain Softjin [62]
28 Japanese Encephalitis Virus (JEV) Japanese encephalitisvirus (JEV) is a mosquito-borne flavivirus that over thepast few decades has caused several outbreaks throughoutChina Southeast Asia Australia and Papua New Guineawith a prevalence recently estimated to be of about 70000casesyear [107ndash109] A 40 mortality was recorded in someof the JEV-affected areas Moreover many survivors facesome neurological problems and complications [108] Since1995 the disease has also emerged in Non-Asian regions suchas Northern Australia [110 111] The situation in SoutheastAsia however is further complicated by the overlappingepidemics of JEV and DENV as well as sporadic casesof WNV infections detected in some of the affected areasparticularly in India [112]
Effective vaccines both live-attenuated and inactivatedstrains of JEV have been developed and licensed in majorcountries However as for the other arthropod-borne Fla-vivirus members no specific antiviral drugs are currentlyavailable [4]
281mAbs against JEV Aspreviously described for the otherFlavivirusmembers many groups isolated and characterizedanti-JEVmAbs showing different neutralizing and protectiveproperties In this regard Gupta et al used combinations ofanti-E JEV mAbs (Hs1-4) in mice protection experimentsIn particular they found that the singularly mAb protectionranged from 45 to 65 when 100120583g of mAb were adminis-tered while equimolar combinations of two or three mAbsgave 85ndash90 or 100 protection respectively [113] In similarexperiments Lee et al demonstrated that a Flavivirus anti-NS1 mAb named 16NS1 cross-reacted with JEV as well asWNV and exhibited protective activity against WNV as wellas a lethal JEV infection However no neutralizing activitywas observed using this mAb against both WNV and JEV inin vitro experiments suggesting the participation of otherAb-mediatedmechanisms in vivo In particular 95ofmicewere
protected when 500120583g of mAb were administered intraperi-toneally and concomitantly to intramuscular injection of JEV
Overlapping peptide mapping analysis combined withsite-specific mutations identified the 116KAWGKSILFA125region and critical amino acid residues W118 and I122 as16NS1 mAb epitope highly conserved in WNV and JEVstrains [114]
Arakawa et al isolated from a combinatorial human Fablibrary constructed from peripheral blood lymphocytesobtained from JEV hyperimmune volunteers Among 188randomly selected clones FabTJE12B02 showed the best 50focus reduction endpoint at the concentration of 502 120583gmLagainst the JEV strain Nakayama [55]
Goncalvez et al isolated three mAbs named Fabs A3B2 and E3 by repertoire cloning from chimpanzees initiallyimmunized with inactivated JE-VAX and then boosted withattenuated JEV SA14-14-2 In particular these mAbs reactedwith epitopes in three different E domains in DI (amino acidresidue K179) in DII (I126) and in DIII (G132) for Fabs A3B2 and E3 respectively Moreover these Fabs as well as thederived humanized counterpart mAbs exhibited high neu-tralizing activities against a broad spectrum of JEV genotypestrains Moreover these mAbs exhibited a 50 protectivedose of 084 120583g (B2) 58120583g (A3) and 247120583g (E3) in mousemodels Finally administration of 200 120583gmouse of mAb B2one day after otherwise lethal JEV infection protected 50of mice and significantly prolonged the average survival timecompared to that of mice in the unprotected group [54]
29 St Louis Encephalitis Virus (SLEV) St Louis encephalitisvirus (SLEV) was first discovered as the mosquito-borneagent responsible for over 1000 cases of encephalitis duringa 1933 summer outbreak in St Louis (Missouri) and nowis a reemerging human pathogen widely distributed in theAmerican continent causing several human encephalitisoutbreaks over the last 80 years Additional epidemics haveindeed occurred from 1964 to 2006 in the Americas rangingfrom the US to Argentina and Brazil As a member of theFlavivirus genus the SLEVE glycoprotein ectodomain is 68identical to serocomplex-related JEVEbut only 46 and 40identical to those ofDENV2 andTBEV viruses fromdifferentserocomplexes [115]
291 mAbs against SLEV In 1983 Roehrig et al isolatedtwenty-one hybridomas producing murine mAbs specificfor the E glycoprotein of SLEV strain MSI-7 Serologicreactivities were initially determined by cross-reactivity indi-rect immunofluorescence assays using 22 strains of SLEVand 8 other related flaviviruses Four groups demonstratingtype- subcomplex- supercomplex- and group-specific reac-tivity patterns were identified Analysis of hemagglutination-inhibition and virus neutralization subdivided the cross-reactivity groups into eight epitopes (E-1a b c d E-2 E-3 and E-4a b) one of them following localized in DII (4b)[52 53]
Moreover among the previously described isolated anti-WNV mAbs by Gould et al the chimeric scFv-Fc 79 effec-tively neutralized also SLEV resulting in gt80 of PRNT80when used at 5 120583gmL [42]
BioMed Research International 13
3 Alphaviruses
Major human arboviruses in the Togaviridae family Chikun-gunya (CHIKV) and Venezuelan equine encephalomyelitis(VEEV) viruses belong to the Alphavirus genus which iscomposed of viruses with an icosahedral nucleocapsid sur-rounded by a lipid envelope and glycoprotein spikes Thestructural proteins of alphaviruses arise through co- andpost-translational processing of a polyprotein encoded by asingle positive-stranded RNAproducing the capsid (C) PE26 K and E1 The PE2 glycoprotein is a precursor containingthe E3 glycoprotein fused to the amino terminus of the E2envelope glycoprotein The PE2 glycoprotein is followed by6K a small membrane-associated protein and E1 the secondpolypeptide component of glycoprotein spikes
Trimerized heterodimers of the E1 and E2 viral glyco-proteins form the surface spikes and contain determinantsof viral tropism and virulence The E3 glycoprotein acts as asignal for transport of PE2 across themembranes of the roughendoplasmic reticulum and may promote the formation andintracellular transport of E1-PE2 heterodimers to the cellsurface During transport to the cell surface PE2 undergoesa maturational cleavage event by a furin-like protease toproduce E2 and E3 [116]
The E2 glycoprotein promotes specificity of virus bindingto the host cell surface and is a target of the humoral immuneresponse The E1 glycoprotein mediates fusion of the virionenvelope with the membranes of acidified endosomes allow-ing release of the nucleocapsid into the cytoplasm and theonset of viral replicationAlphavirus E1 shares no appreciablesequence identity to Flavivirus E protein Despite differencesin their amino acid sequences and arrangements on the viralparticle the structures of E and E1 are remarkably similarIndeed the conservation of three domains has been welldocumented in crystal structures of flavivirus E ectodomainsfrom TBEV DENV JEV and WNV as well as Alphavirus E1from Semliki Forest virus (SFV) and CHIKV Abs to the E1glycoprotein do not typically neutralize virus infectivity invitro but can protect against lethal challenge in animals
By analogy with the prM protein of dengue Flavivirusfurin cleavage of PE2 may begin after transit of an acidiclate component of the Golgi body where E3 is thought tosuppress the acid pH-triggered activation of glycoprotein E1fusion capability Protective anti-E3 mouse mAbs have alsobeen described [117]
31 Chikungunya Virus (CHIKV) The chikungunya virus(CHIKV) belongs to the Semliki Forest clade and was firstlyisolated in 1953 in Tanzania during an epidemic outbreakfollowing occurred also in Asia and Africa In the last fivedecades several CHIKV outbreaks have been described bothin Africa and Asia separated by gaps lasting from two totwenty years In 2005-2006 about 300000 cases out of785000 inhabitants were reported in La Reunion island witha fatal prognosis for 237 of the patients [117] Neither Europenor the Americas have had outbreaks of CHIKV so far exceptfor imported isolated cases Three genotypes of CHIKV aredescribed Asian EastCentralSouth African (ECSA) andWest African with an amino acid identity spanning from
952 to 998 Recent epidemics in Africa and Indiansubcontinent were caused by strains belonging to the ECSAgenotype Transmitted by Aedes mosquitoes CHIKV ismaintained in the human population by human-mosquito-human transmission [1] The disease is characterized bydengue-like symptoms such as chills high fever headacheand persistent myalgia and a further incapacitating arthralgia(from which the name chikungunya) which affects 40 ofinfected subjects However prognosis is rarely fatal [116]
There is currently no commercial vaccine and antiviraltreatment forCHIKV although some candidate vaccines havebeen tested in humans In this regard in 2000 US Armyperformed a Phase II clinical trial testing a live-attenuatedCHIKV vaccine (TSI-GSD-218) derived from a 1962 strain(15561) of an outbreak in Thailand Out of 58 every patientdeveloped neutralizingAbs and 5 lamentedmild tomoderatejoint pain [117] A phase III trial of this candidate vaccineis ongoing Furthermore a new formulation using virus-likeparticles was able to induce neutralizing Abs in macaquesagainst different CHIKV strains [117 118] Indeed it has beendescribed that infection seems to elicit long-lasting protectiveimmunity and cross-protection among CHIKV and otheralphaviruses
CHIKV entry into host cell is demonstrated to be medi-ated by envelope glycoproteins E1 and E2 which allowvirus fusion to cell membrane in low pH conditions andrecognition of an unknown cellular receptor respectively[116 119ndash121] Even though the genome replication relies onan error-prone RNA-dependent RNA-polymerasis CHIKVstrains S27 and 05115 Reunion showed in E1 and E2 alow amino acid variation of 068 and 33 respectivelyand such a low variability may be needed for an effectivereplication in two phylogenetically distant hosts [116 122]Recently the crystal structure of E1 and E2 heterodimerhas been resolved both at low pH and natural conditionsshedding light on E1 and E2 ectodomainsrsquo structure Inparticular similarly to flaviviruses E protein E1 ectodomainis made of the N-terminal DI the DII containing the fusionloop and the DIII at the C-terminal On the other hand E2ectodomain comprises theN-terminal domainA the domainB supposed to interact with the host cellrsquos unknown receptorand domain C at the C-terminal E1 DIII and E2 domain Care located close to the viral membrane [123 124]
311 mAbs against CHIKV Warter et al described 5F10 and8B10 two human mAbs which strongly neutralized severalCHIKV isolates in vitro without cross-reactivity againstother alphaviruses but to Onyong-nyong virus [56] Mixedpreparation of 5F10 and 8F10 did not show neither synergisticnor addictive effect in vitro and studies upon escape mutantsdemonstrated that 5F10 mAb binds at the tip of the E2domain B while 8B10 recognizes residues close to E1 fusionloop and amino acids within E2 domain A which form atransitional epitope under low pH conditions The authorsspeculated that 5F10 and 8B10 may inhibit CHIKV entry andfusion to the cell membrane respectively It is worth notingthat the previously mentioned escape mutants displayedmutations associated with reduced viral fitness in vitro evenafter 13 neutralizationamplification rounds In the same
14 BioMed Research International
study cell-to-cell transmission was firstly demonstrated as anescapemechanism enhanced by the E2mutation R82G [122]
In further in vivo studies 5F10 and 8B10 significantlydelayed CHIKV-caused death of AGR129 mice both in pro-phylactic and therapeutic tests Interestingly in therapeutictreatment these mAbs showed a synergistic effect whenadministered in combination with the total amount of mAbsinjected being the same of the single-mAbs Possibly thesetreatment did not allow themice to survive CHIKV challengedue to Ig clearing and 6ndash10 days half-life usually described forhuman mAbs [125]
Recently Pal et al cloned thirty-six murine mAbs ableto neutralize the ECSA La Reunion 2006 OPY-1 strain ofCHIKV (CHIKV-LR) the majority of which also neutralizeinfection of other strains corresponding to the Asian andWest African genotypes In particularmAbCHK-152 showedthe highest and broadest neutralizing activity with FRNTvalues indicating an IC50 value of 1 to 3 ngmL dependingon the viral strain used Among the thirty-six mAbs four(CHK-102 CHK-152 and CHK-166 CHK-263) could pre-vent lethality of immunodeficient Ifnarminusminus C57BL6 micewhen administered one day before exposition with CHK-152 and CHK-263 showing the ability to protect mice at thelowest dose (10 120583g) Therapeutic studies were performed onthese mAbs administering a single dose of 100 120583g 24 hoursafter infection The highest activity was described for CHK-166 able to protect 63 of the mice Testing the combinedactivity of the mAbs in therapeutic studies administration ofCHK-166 plus CHK-152 in a dose of 250120583g each turned out tobe themost effective protecting 71 of the IfnarminusminusC57BL6mice 60 hours after infection [57]
32 Venezuelan Equine Encephalomyelitis Virus (VEEV)Venezuelan equine encephalomyelitis virus (VEEV) is main-tained in a natural transmission cycle between mosquitoesand small rodents The first documented outbreaks occurredin the 1930s and several epidemics have been reported so farin Latin-American countries such as Venezuela ColombiaPeru Ecuador Bolivia Honduras Mexico and Panama withhundreds of thousands of human cases being reported anda case-fatality rate up to 1 during the 1969 Ecuador out-break Clinical manifestations of VEE are indistinguishablefrom Dengue [126] Six serogroups (IndashVI) are currentlyrecognized within the VEEV complex VEEV caused humanand equine outbreaks in the Americas for nearly a centuryEquine epizootics have high mortality (38ndash83) leading toa high viremia followed by a lethal encephalitis and often tohuman epidemics involving thousands of cases and hundredsof deaths Indeed VEEV is infectious for humans by theairborne route and has been responsible for a number oflaboratory infections In humans the disease is usually self-limiting with a febrile illness with 1ndash4 of cases progressingto severe encephalitis It is a hazard to laboratory workershas been developed as a biological weapon and is a potentialbioterrorist agent [127] There are no antiviral drugs andvaccines licensed for the treatment of VEEV infection inhumans Indeed although experimental live-attenuated vac-cines have been developed (eg TC-83) with good levels ofprotection in equine and mice in humans they may fail to
give protection in the majority of cases [128] Consideringthe protective role exerted by humoral immune response inanimal models it could be concluded that treatment withspecific IgG may have a beneficial antiviral effect in humanairborne infections with pathogenic strains of VEEV
321 mAbs against VEEV Several works support that mAbsmay protect against airborne VEEV as well as after airborneexposure toVEEVAb reactivity to both surface glycoproteins(E1 and E2) is associated with neutralization of the virusin vitro and passive protection against virus challenge Aseries of anti-E1 and anti-E2 mouse and humanized mAbshave been described recently Among them Phillpotts et alexamined two anti-E2 murine mAbs named 1A4A-1 and1A3A-9 both having potent protective activity against subcu-taneous VEEV challenge in mice Both mAbs had a similarhalf-life (58 and 100 days) in mouse serum after a singleintraperitoneal dose suggesting that mAbs delivered by thisroute at or around the time of VEEV infection persists athigh levels in the blood as well as secretions in respiratorytransudation throughout the clinical course of the disease Inparticular both mAbs administered 24 h prior to airbornechallenges had a substantial protective effect (90ndash100)Treatment of mice 2 or 24 h after airborne infection with asingle intraperitoneal dose of 100 120583g of 1A3A-9 mAb led toapproximately 50 survival There was no beneficial effectwhen mAb treatment was delayed to 72 h Moreover therewas evidence of synergy in vitro in PRNT between 1A3A-9and 1A4A-1 as has been demonstrated for other virusesand mAb pairs However no synergy was found in mouseprotection when mAbs were delivered intraperitoneally as amixture in equal parts to animals challenged 24 h previouslywith VEEV [129]
In a subsequent study Phillpotts et al reported two otherprotective mAbs The 3B2A-9 mAb protected against allthe serogroup I strains while 1A3B-7 mAb protected wellagainst challenge with all of the viruses tested Both 3B2A-9 and 1A3B-7 protected against airborne exposure to theIAB serogroup virus strainTrinidad donkey (TrD)Howeverthere was no evidence of synergistic protection when thesemAbs were combined in equal proportions An intraperi-toneal dose of 10 120583g was sufficient to protect 50 of themice with either mAb [130] According to other data themechanism of protection did not appear to depend uponneutralization [131] Similar results were obtained by OrsquoBrienet al which described a non-neutralizing mAb (IgG2a)named CUF37-2a able to protect 50 of mice from asubcutaneous VEEV challenge when a dose of 915 120583g of mAbwas administered 24 h prior to challenge [132]
In a following study the 1A3B-7 mAb was humanized(and following reported as Hu1A3B-7) maintaining the samefeatures of its murine counterpart In particular evaluation ofin vitro studies indicated that Hu1A3B-7 retained both broadspecificity and neutralizing activity Furthermore in vivoexperiments showed that Hu1A3B-7 successfully protectedmice against lethal subcutaneous and aerosol challenges withVEEV strain TrD Moreover the effectiveness of the human-ization process was determined by assessing proliferationresponses in human T-cells to peptides derived from the
BioMed Research International 15
murine and humanized versions of the VH and VL domainsThis analysis showed that the number of human T-cellepitopes within the humanized Ab had been substantiallyreduced indicating that Hu1A3B-7 may have low immuno-genicity in vivo [133]
Similarly Hunt et al described a humanizedmurinemAbderived from the 3B4C-4 mAb recognizing the E2c epitope(amino acid residues 182ndash209) and following named Hy4-26C showing neutralizing activity similar to the murinemAb Moreover it was protective (70ndash100 of survival) at100 ng dose in mice animal model intraperitoneal inoculatedand at 500120583g for intranasal challenge (80) Therapeuticstudies in mice revealed that Hy4-26C was able to cure miceup to 24 h following infection at 10 120583g on the contrary themouse mAb when given 1 hour after virus challenge [63]
In another work Hunt et al isolated two neutralizinghumanized Fabs F5 and L1A7 employing a blocking strategy[134 135] The anti-E2 specific F5 IgG had a 70 PRNTendpoint of 10 ngmL equivalent to that described for themost effective neutralizing anti-VEEVE2mAbs In particularthe E1-specific hFab L1A7 had a PRNT endpoint of 3120583gmL300-fold lower than F5
Further studies revealed that F5 epitope is located inthe 115ndash199 amino acid region Moreover F5 IgG had potentability to protect mice from infection by either route whenadministered 24 h before exposure however mice treated24 h and 48 h after aerosol exposure developed central ner-vous system infections but exhibited no clinical signs ofdisease [135]
Hu et al reported that passive immunization with thehumanized chimericmousemAbhu1A4A1IgG1-2A inmice at50 120583g 24 h before or after virulent VEEV challenge providedcomplete protection indicating that hu1A4A1IgG1-2A haspotent prophylactic and therapeutic effects against VEEVinfection [136]
Anti-VEEV mAbs isolated from a non-human primategene library has also been reported In particular thehumanized chimeric scFv-Fc ToR67-3B4 recognized viableas well as formalin and 120573-propiolactone inactivated virusparticles It detected specifically the viral E1 envelope proteinof VEEV but did not react with reduced viral glycopro-tein preparations suggesting that recognition depends uponconformational epitopes The recombinant Ab was able todetect multiple VEEV subtypes and displayed only marginalcross-reactivity to otherAlphavirus species except for Easternequine encephalitis virus (EEEV) In addition the scFv-Fcfusion described here might be of therapeutic use since itsuccessfully inactivated VEEV in a murine disease modelIn particular when the recombinant Ab was administered 6hours after challenge 80 to 100 of mice survived lethalVEEV IAB or IE infection Forty to sixty percent of micesurvived when scFv-Fc ToR67-3B4 was applied 6 hours afterchallenge with VEEV subtypes II and former IIIA [137]
4 Bunyaviridae
TheBunyaviridae family contains human arboviruses belong-ing to the Orthobunyavirus Phlebovirus and Nairovirus
genera Bunyaviridae includes enveloped viruses with a frag-mented single-stranded RNA genome of negative polarityTheir tripartite genome consists of a small (S) a medium(M) and a large (L) fragment and the envelope glycoproteinsGn and Gc that are cleaved out of a polyprotein synthesizedby the M fragment These glycoproteins have been provedto mediate the formation of the virus particle to play a rolein the interaction with cell surface receptors to mediate theentry of the virus into cells and to serve as targets for themajority of neutralizing Abs described so far
Orthobunyavirus transmitted through mosquitoes ormidges vectors are divided in 18 serogroups based on cross-titrations in haemagglutination inhibition assays and neutral-ization assays and correlating with main vector preferencesHowever the most clinically important viruses belong onlyto two serogroups the California encephalitis and the Simbuserogroups
Crimean-Congo hemorrhagic fever virus (CCHFV) isconsidered the only one of clinical relevance within theNairovirus genus which uses tick as main vector and is divid-ed in seven serogroups while Toscana virus and Rift Valleyfever virus (RVFV) which are transmitted by sandflies andmosquitoes respectively belong to the Phlebovirus genus[138]
MAbs against theNairovirusCCHFV and the PhlebovirusRVFV have been described
41 Crimean-Congo Hemorrhagic Fever Virus (CCHFV)Crimean-Congo Hemorrhagic Fever Virus (CCHFV) dis-tribution covers the greatest geographic range of any tick-borne virus known as viral isolation andor disease hasbeen reported from more than 30 countries in SoutheasternEurope Africa Asia and Middle East [139 140] Namelyfrom 1953 to 2010 about 6000 human cases were reportedin Southeastern Europe CCHFV causes sporadic outbreakswith mortality rates ranging from 10 to 80 [141 142] Asignificant variation in the time of incubation has beendescribed [139] Prehemorrhagic symptoms include highfever chills headache photophobia and back and abdominalpains Among other symptoms neuropsychiatric changeshave been reported in some patients In severe cases 3ndash6 daysafter the onset of the disease hemorrhagic symptoms occurwith petechiae ecchymosis bleeding in the form of melenahematemesis epistaxis [139]
In comparisonwith the nucleocapsid proteins inCCHFVGn and Gc show a higher degree of antigenic variabilityprobably due to their exposition to the host immune systemhypothesis corroborated in the description by Hewson et alin 2004 of four M segment phylogenic groups namely M1M2 M3 and M4 [141 142]
411 mAbs against CCHFV Blackburn et al firstly describedin 1987 murine anti-CCHFV mAbs able to recognize nucle-ocapsid proteins but no neutralizing activity has beendescribed thus far [143]
On the other hand Bertolotti-Ciarlet et al produced apanel of murine mAbs recognizing conformational epitopes
16 BioMed Research International
within theGn andGc glycoproteins expressed by the CCHFVstrain IbAr10200 The effectiveness of the mAbs was studiedby performing PRNT80 and none of the anti-Gn mAbsshowed neutralizing activity although many of the anti-GcmAbs neutralized IbAr10200 strain in vitro with clones 8A15E3 12A9 6C2 and 9H3 showing activity at a gt5120 dilutionMAbs 11E7 and 30F7 exhibited neutralizing activity at 2560dilution A suckling mice protection test was thereforeperformed for these mAbs The previously mentionedanti-Gc Abs were capable to protect mice to an appreciabledegree when applied 24 hours before and in a weakermanner 24 hours after virus challenge Many of the anti-GnmAbs were able to confer significant protection to IbAr10200both 24 hours before and after virus administration Therelevant effectiveness of the anti-Gn mAbs 6B12 10E1113G8 and 10G4 suggests that these mAbs may possess someneutralizing activity due to Ab-based effector mechanisms(eg ADCC) [58]
In further studies the same group characterized thebroadly cross-reactive neutralizing activity of the previouslydescribed murine mAb 11E7 able to bind conformationalepitopes within the C-terminal region of the Gn glycoproteinof all CCHFVM groups [58 141]
42 Rift Valley Fever Virus (RVFV) Similarly to CCHFV RiftValley Fever Virus (RVFV) causes sporadic outbreaks Thelargest epidemic occurred in Egypt in 1977 with an estimated200000 infections 18000 patients manifesting symptomsand 600 fatal prognoses [144] Symptoms include hemor-rhagic episodes fever encephalitis and blindness [145]
The major target of the humoral immune response is theRVFV glycoproteins Gc (G1) and Gn (G2) ranging fromnucleotides 480ndash2090 and 2091ndash3614 respectively [144]
421 mAbs against RVFV Within G2 glycoprotein are des-cribed three neutralizing antigenic sites namely epitope Inucleotides 792ndash893 (amino acid residues 258ndash291) epitopeII nucleotides 1164ndash1196 (amino acid residues 382ndash392) andepitope IV nucleotides 858ndash917 (amino acid residues 280ndash299) [59 145]
Despite the high degree of variability observed inCCHFVglycoproteins RVFV G2 presented an unexpected high con-servation among 22 isolates [59] In the same study onlythe LUNYO and 900060 isolates were resistant to in vitroneutralization by murine mAb 4D4 (recognizing epitope II)All other isolates were neutralized with PRNT80 titers from10240 togt81920On the other hand only SNS isolate showedreduced sensitivity to 4ndash39-CC (epitope IV) All other isolateswere neutralized with PRNT80 titers from 20480 to gt81920Protection studies on mice would be of interest for thedevelopment of a passive immunization prophylaxis [59]
Protection studies from lethal South African RVFVAN1830 strain infection were successfully performed on abattery of anti-G1 and anti-G2 mAbs produced by Besselaarand Blackburn with the strongly neutralizing anti-G1 mAb3E5 and anti-G2mAb 9C4 being the most effective [146]The9C4 antigenic area maps within G2 epitope I and a broadlyneutralizing activity may be investigated
5 Conclusions
Arbovirus infections have acquired increasing interest giventhe augmented globalization and tourism movement all overthe world that could be at the basis of epidemic eventsFurthermore the same vectors can sometimes transmit sev-eral arboviruses concomitantly complicating the diagnosis aswell as the therapy For example confusing mixed epidemicshave occasionally been described such as YFV plus CHIKVDENVplus CHIKV ormore recentlyPlasmodium falciparummalaria plus DENV1 and CHIKV on Madagascarrsquos east coast[1]
In the last decade several efforts have been employed inthe development of effective immunogens and therapeuticsgiving the current absence of specific antiviral drugs as wellas effective vaccines for themost diffused arbovirus infections(ie DENV andWNV)
However at the moment mosquito control is the bestavailable method for preventing arbovirus infections Thusto control the emerging public health problem of arbovirusinfections new antiviral therapeutic strategies that providepotent and broadly cross-protective immunity (especially forflavivirus infections) are an urgent globally medical need
As described in this review the majority of highlyneutralizing mAbs are of murine origin and indeed theirutility in the treatment of these infections is limited In facttheir employment can elicit an immunogenic response (iehuman anti-mouse Abs HAMA) the therapeutic efficacycould be reduced by a relatively faster clearance in humans(compared to human Abs) potentially exacerbated by theHAMA response Finally murine Abs exhibit relatively weakeffector functions (eg ADCC) compared to human AbsIndeed chimerization humanization or better the isolationand characterization of fully human broadly neutralizingmAbs would be the best choice for their possible use in afuture mAb-based therapy against arbovirus infections [147]
Finally possible ADE of infection mechanisms should beevaluated before considering a mAb as a possible candidatetherapeutic in a post-exposure setting as well as in the devel-opment of new Ab-eliciting immunogens
Conflict of Interests
The authors declare no conflict of interests in particular withOmriGam and any other cited Ab-based preparation men-tioned in the paper
References
[1] N Cleton M Koopmans J Reimerink G J Godeke and CReusken ldquoCome fly with me review of clinically importantarboviruses for global travelersrdquo Journal of Clinical Virology vol55 no 3 pp 191ndash203 2012
[2] F X Heinz H Holzmann A Essl and M Kundi ldquoField effec-tiveness of vaccination against tick-borne encephalitisrdquoVaccinevol 25 no 43 pp 7559ndash7567 2007
[3] A D Barrett and D E Teuwen ldquoYellow fever vaccinemdashhowdoes it work and why do rare cases of serious adverse events
BioMed Research International 17
take placerdquo Current Opinion in Immunology vol 21 no 3 pp308ndash313 2009
[4] S B Halstead and S J Thomas ldquoNew vaccines for Japaneseencephalitisrdquo Current Infectious Disease Reports vol 12 no 3pp 174ndash180 2010
[5] R Burioni M Perotti N Mancini and M Clementi ldquoPer-spectives for the utilization of neutralizing human monoclonalantibodies as anti-HCV drugsrdquo Journal of Hepatology vol 49no 2 pp 299ndash300 2008
[6] R A Diotti G A Sautto L Solforosi NManciniM Clementiand R Burioni ldquoNeutralization activity and kinetics of twobroad-range human monoclonal IgG1 derived from recombi-nant Fab fragments and directed against Hepatitis C virus E2glycoproteinrdquo New Microbiologica vol 35 no 4 pp 475ndash4792012
[7] R Burioni F Canducci N Mancini et al ldquoMolecular cloningof the first humanmonoclona antibodies neutralizing with highpotency Swine-origin Influenza A pandemic virus (S-OIV)rdquoNew Microbiologica vol 32 no 4 pp 319ndash324 2009
[8] P K Chandra S Hazari B Poat et al ldquoIntracytoplasmic stableexpression of IgG1 antibody targeting NS3 helicase inhibitsreplication of highly efficient hepatitis C Virus 2a clonerdquoVirology Journal vol 7 article 118 2010
[9] N Clementi D de Marco N Mancini et al ldquoA human mono-clonal antibody with neutralizing activity against highly diver-gent influenza subtypesrdquo PLoS ONE vol 6 no 12 Article IDe28001 2011
[10] D de Marco N Clementi N Mancini et al ldquoA non-VH1-69 heterosubtypic neutralizing human monoclonal antibodyprotects mice against H1N1 and H5N1 virusesrdquo PLoS ONE vol7 no 4 Article ID e34415 2012
[11] L Solforosi N Mancini F Canducci et al ldquoA phage displayvector optimized for the generation of human antibody com-binatorial libraries and the molecular cloning of monoclonalantibody fragmentsrdquoNewMicrobiologica vol 35 no 3 pp 289ndash294 2012
[12] E V Koonin and V V Dolja ldquoEvolution and taxonomy ofpositive-strand RNA viruses implications of comparative anal-ysis of amino acid sequencesrdquo Critical Reviews in Biochemistryand Molecular Biology vol 28 no 5 pp 375ndash430 1993
[13] J S Mackenzie D J Gubler and L R Petersen ldquoEmergingflaviviruses the spread and resurgence of Japanese encephalitisWest Nile and dengue virusesrdquo Nature Medicine vol 10 no 12pp S98ndashS109 2004
[14] T J SmithW E Brandt J L Swanson J MMcCown and E LBuescher ldquoPhysical and biological properties of dengue-2 virusand associated antigensrdquo Journal of Virology vol 5 no 4 pp524ndash532 1970
[15] J M Smit B Moesker I Rodenhuis-Zybert and J WilschutldquoFlavivirus cell entry and membrane fusionrdquo Viruses vol 3 no2 pp 160ndash171 2011
[16] R J Kuhn W Zhang M G Rossmann et al ldquoStructure ofdengue virus implications for flavivirus organization matura-tion and fusionrdquo Cell vol 108 no 5 pp 717ndash725 2002
[17] K Stiasny J H Aberle M Keller B Grubeck-Loebenstein andF X Heinz ldquoAge affects quantity but not quality of antibodyresponses after vaccination with an inactivated flavivirus vac-cine against tick-borne encephalitisrdquo PLoS ONE vol 7 no 3Article ID e34145 2012
[18] LThomasOVerlaetenACabie et al ldquoInfluence of the dengueserotype previous dengue infection and plasma viral load
on clinical presentation and outcome during a dengue-2 anddengue-4 co-epidemicrdquo American Journal of Tropical Medicineand Hygiene vol 78 no 6 pp 990ndash998 2008
[19] Y Zhang J Corver P R Chipman et al ldquoStructures of imma-ture flavivirus particlesrdquo EMBO Journal vol 22 no 11 pp2604ndash2613 2003
[20] A K I Falconar ldquoIdentification of an epitope on the denguevirus membrane (M) protein defined by cross-protective mon-oclonal antibodies design of an improved epitope sequencebased on common determinants present in both envelope (EandM) proteinsrdquo Archives of Virology vol 144 no 12 pp 2313ndash2330 1999
[21] K-J Huang Y-C Yang Y-S Lin et al ldquoThe dual-specific bind-ing of dengue virus and target cells for the antibody-dependentenhancement of dengue virus infectionrdquo Journal of Immunol-ogy vol 176 no 5 pp 2825ndash2832 2006
[22] A K I Falconar ldquoAntibody responses are generated to immun-odominant ELKKLE-typemotifs on the nonstructural-1 glyco-protein during live dengue virus infections inmice and humansimplications for diagnosis pathogenesis and vaccine designrdquoClinical and Vaccine Immunology vol 14 no 5 pp 493ndash5042007
[23] C W Mandl F Guirakhoo H Holzmann F X Heinz and CKunz ldquoAntigenic structure of the flavivirus envelope protein Eat the molecular level using the tick-borne encephalitis virus asa modelrdquo Journal of Virology vol 63 no 2 pp 564ndash571 1989
[24] F A Rey F X Heinz C Mandl C Kunz and S C HarrisonldquoThe envelope glycoprotein from tick-borne encephalitis virusat 2 A resolutionrdquo Nature vol 375 no 6529 pp 291ndash298 1995
[25] S L Allison K Stiasny K Stadler CWMandl and F XHeinzldquoMapping of functional elements in the stem-anchor regionof tick-borne encephalitis virus envelope protein Erdquo Journal ofVirology vol 73 no 7 pp 5605ndash5612 1999
[26] D E Klein J L Choi and S C Harrison ldquoStructure of a denguevirus envelope protein late-stage fusion intermediaterdquo Journal ofVirology vol 87 no 4 pp 2287ndash2293 2013
[27] S L Hanna T C Pierson M D Sanchez A A Ahmed M MMurtadha and R W Doms ldquoN-linked glycosylation of WestNile virus envelope proteins influences particle assembly andinfectivityrdquo Journal of Virology vol 79 no 21 pp 13262ndash132742005
[28] N Mancini G Sautto N Clementi et al ldquoNeutralization inter-fering antibodies a ldquonovelrdquo example of humoral immune dys-function facilitating viral escaperdquoViruses vol 4 no 9 pp 1731ndash1752 2012
[29] Y Modis S Ogata D Clements and S C Harrison ldquoVariablesurface epitopes in the crystal structure of dengue virus type3 envelope glycoproteinrdquo Journal of Virology vol 79 no 2 pp1223ndash1231 2005
[30] N Miller ldquoRecent progress in dengue vaccine research anddevelopmentrdquo Current Opinion in Molecular Therapeutics vol12 no 1 pp 31ndash38 2010
[31] J H Chavez J R Silva A A Amarilla and L T Moraes Figu-eiredo ldquoDomain III peptides from flavivirus envelope proteinare useful antigens for serologic diagnosis and targets forimmunizationrdquo Biologicals vol 38 no 6 pp 613ndash618 2010
[32] B Shrestha J D Brien S Sukupolvi-Petty et al ldquoThe develop-ment of therapeutic antibodies that neutralize homologous andheterologous genotypes of dengue virus type 1rdquo PLoS Pathogensvol 6 no 4 Article ID e1000823 2010
18 BioMed Research International
[33] G D Gromowski N D Barrett and A D T Barrett ldquoChar-acterization of dengue virus complex-specific neutralizing epi-topes on envelope protein domain III of dengue 2 virusrdquo Journalof Virology vol 82 no 17 pp 8828ndash8837 2008
[34] S Sukupolvi-Petty S K Austin W E Purtha et al ldquoType- andsubcomplex-specific neutralizing antibodies against domain IIIof dengue virus type 2 envelope protein recognize adjacentepitopesrdquo Journal of Virology vol 81 no 23 pp 12816ndash128262007
[35] S Sukupolvi-Petty S K Austin M Engle et al ldquoStructure andfunction analysis of therapeutic monoclonal antibodies againstdengue virus type 2rdquo Journal of Virology vol 84 no 18 pp9227ndash9239 2010
[36] Y-Q Deng J-X Dai G-H Ji et al ldquoA broadly flaviviruscross-neutralizingmonoclonal antibody that recognizes a novelepitope within the fusion loop of e proteinrdquo PLoS ONE vol 6no 1 Article ID e16059 2011
[37] J M Costin E Zaitseva K M Kahle et al ldquoMechanistic studyof broadly neutralizing human monoclonal antibodies againstdengue virus that target the fusion looprdquo Journal of Virologyvol 87 no 1 pp 52ndash66 2013
[38] CMMidgley A FlanaganH B Tran et al ldquoStructural analysisof a dengue cross-reactive antibody complexed with envelopedomain III reveals the molecular basis of cross-reactivityrdquoJournal of Immunology vol 188 no 10 pp 4971ndash4979 2012
[39] N J Moreland P Susanto E Lim et al ldquoPhage display appro-aches for the isolation of monoclonal antibodies against denguevirus envelope domain III from human and mouse derivedlibrariesrdquo International Journal ofMolecular Sciences vol 13 no3 pp 2618ndash2635 2012
[40] J J B Cockburn M E Navarro Sanchez N Fretes et alldquoMechanism of dengue virus broad cross-neutralization by amonoclonal antibodyrdquo Structure vol 20 no 2 pp 303ndash3142012
[41] R Rajamanonmani C Nkenfou P Clancy et al ldquoOn a mousemonoclonal antibody that neutralizes all four dengue virusserotypesrdquo Journal of General Virology vol 90 no 4 pp 799ndash809 2009
[42] L H Gould J Sui H Foellmer et al ldquoProtective and herapeuticcapacity of human single-chain Fv-Fc fusion proteins againstWest Nile virusrdquo Journal of Virology vol 79 no 23 pp 14606ndash14613 2005
[43] H Sultana H G Foellmer G Neelakanta et al ldquoFusion LoopPeptide of the West Nile virus envelope protein is essential forpathogenesis and is recognized by a therapeutic cross-reactivehuman monoclonal antibodyrdquo Journal of Immunology vol 183no 1 pp 650ndash660 2009
[44] T Oliphant M Engle G E Nybakken et al ldquoDevelopment ofa humanized monoclonal antibody with therapeutic potentialagainstWest Nile virusrdquoNature Medicine vol 11 no 5 pp 522ndash530 2005
[45] G E Nybakken T Oliphant S Johnson S Burke M S Dia-mond and D H Fremont ldquoStructural basis of West Nile virusneutralization by a therapeutic antibodyrdquo Nature vol 437 no7059 pp 764ndash769 2005
[46] G D Gromowski J T Roehrig M S Diamond J C LeeT J Pitcher and A D T Barrett ldquoMutations of an antibodybinding energy hot spot on domain III of the dengue 2 envelopeglycoprotein exploited for neutralization escaperdquo Virology vol407 no 2 pp 237ndash246 2010
[47] J T Roehrig R A Bolin andR G Kelly ldquoMonoclonal antibodymapping of the envelope glycoprotein of the dengue 2 virusJamaicardquo Virology vol 246 no 2 pp 317ndash328 1998
[48] R de Alwis S A Smith N P Olivarez et al ldquoIdentification ofhumanneutralizing antibodies that bind to complex epitopes ondengue virionsrdquo Proceedings of the National Academy of Sciencesof the United States of America vol 109 no 19 pp 7439ndash74442012
[49] S K Austin K A Dowd B Shrestha et al ldquoStructural basis ofdifferential neutralization of DENV-1 genotypes by an antibodythat recognizes a cryptic epitoperdquo PLoS Pathogens vol 8 no 10Article ID e1002930 2012
[50] M Throsby C Geuijen J Goudsmit et al ldquoIsolation and char-acterization of human monoclonal antibodies from individualsinfected with West Nile Virusrdquo Journal of Virology vol 80 no14 pp 6982ndash6992 2006
[51] A P Goncalvez R H Purcell and C-J Lai ldquoEpitope deter-minants of a chimpanzee Fab antibody that efficiently cross-neutralizes dengue type 1 and type 2 viruses map to inside andin close proximity to fusion loop of the dengue type 2 virusenvelope glycoproteinrdquo Journal of Virology vol 78 no 23 pp12919ndash12928 2004
[52] J T Roehrig J H Mathews and D W Trent ldquoIdentification ofepitopes on the E glycoprotein of Saint Louis encephalitis virususing monoclonal antibodiesrdquo Virology vol 128 no 1 pp 118ndash126 1983
[53] J T Roehrig L A Staudinger A R Hunt J H Mathews andC D Blair ldquoAntibody prophylaxis and therapy for flavivirusencephalitis infectionsrdquo Annals of the New York Academy ofSciences vol 951 pp 286ndash297 2001
[54] A P Goncalvez C-H Chien K Tubthong et al ldquoHumanizedmonoclonal antibodies derived from chimpanzee fabs protectagainst Japanese encephalitis virus in vitro and in vivordquo Journalof Virology vol 82 no 14 pp 7009ndash7021 2008
[55] M Arakawa T Yamashiro G-I Uechi M Tadano and ANishizono ldquoConstruction of humanFab (1205741120581) library and iden-tification of human monoclonal Fab possessing neutralizingpotency against Japanese encephalitis virusrdquo Microbiology andImmunology vol 51 no 6 pp 617ndash625 2007
[56] L Warter C Y Lee R Thiagarajan et al ldquoChikungunya virusenvelope-specific human monoclonal antibodies with broadneutralization potencyrdquo Journal of Immunology vol 186 no 5pp 3258ndash3264 2011
[57] P Pal K A Dowd J D Brien et al ldquoDevelopment of a highlyprotective combination monoclonal antibody therapy againstchikungunya virusrdquo PLoS Pathogens vol 9 no 4 Article IDe1003312 2013
[58] A Bertolotti-Ciarlet J Smith K Strecker et al ldquoCellular local-ization and antigenic characterization of Crimean-Congo hem-orrhagic fever virus glycoproteinsrdquo Journal of Virology vol 79no 10 pp 6152ndash6161 2005
[59] J K Battles and J M Dalrymple ldquoGenetic variation amonggeographic isolates of Rift Valley fever virusrdquo American Journalof Tropical Medicine and Hygiene vol 39 no 6 pp 617ndash6311988
[60] B AThibodeaux N C Garbino NM Liss et al ldquoA humanizedIgG but not IgM antibody is effective in prophylaxis and therapyof yellow fever infection in an AG12917D-204 peripheralchallenge mouse modelrdquo Antiviral Research vol 94 no 1 pp1ndash8 2012
BioMed Research International 19
[61] S Daffis R E Kontermann J Korimbocus H Zeller H-DKlenk and J Ter Meulen ldquoAntibody responses against wild-type yellow fever virus and the 17D vaccine strain charac-terization with human monoclonal antibody fragments andneutralization escape variantsrdquoVirology vol 337 no 2 pp 262ndash272 2005
[62] L N Levanov L E Matveev E P Goncharova et al ldquoChimericantibodies against tick-borne encephalitis virusrdquo Vaccine vol28 no 32 pp 5265ndash5271 2010
[63] A R Hunt S Frederickson C Hinkel K S Bowdish and J TRoehrig ldquoA humanized murine monoclonal antibody protectsmice either before or after challenge with virulent Venezuelanequine encephalomyelitis virusrdquo Journal of General Virologyvol 87 no 9 pp 2467ndash2476 2006
[64] J M da Silva Voorham I A Rodenhuis-Zybert N V A Nunezet al ldquoAntibodies against the envelope glycoprotein promoteinfectivity of immature dengue virus serotype 2rdquo PLoS ONEvol 7 no 3 Article ID e29957 2012
[65] T C Pierson Q Xu S Nelson et al ldquoThe stoichiometry ofantibody-mediated neutralization and enhancement of westnile virus infectionrdquoCell Host andMicrobe vol 1 no 2 pp 135ndash145 2007
[66] G Sautto N Mancini R A Diotti L Solforosi M Clementiand R Burioni ldquoAnti-hepatitis C virus E2 (HCVE2) glyco-proteinmonoclonal antibodies andneutralization interferencerdquoAntiviral Research vol 96 no 1 pp 82ndash89 2012
[67] R Burioni Y Matsuura N Mancini et al ldquoDiverging effectsof human recombinant anti-hepatitis C virus (HCV) antibodyfragments derived from a single patient on the infectivityof a vesicular stomatitis virusHCV pseudotyperdquo Journal ofVirology vol 76 no 22 pp 11775ndash11779 2002
[68] R J Phillpotts J R Stephenson and J S Porterfield ldquoAntibody-dependent enhancement of tick-borne encephalitus virus infec-tivityrdquo Journal of General Virology vol 66 no 8 pp 1831ndash18371985
[69] K A Dowd and T C Pierson ldquoAntibody-mediated neutraliza-tion of flaviviruses a reductionist viewrdquo Virology vol 411 no 2pp 306ndash315 2011
[70] A K I Falconar ldquoMonoclonal antibodies that bind to commonepitopes on the dengue virus type 2 nonstructural-1 andenvelope glycoproteins display weak neutralizing activity anddifferentiated responses to virulent strains implications forpathogenesis and vaccinesrdquo Clinical and Vaccine Immunologyvol 15 no 3 pp 549ndash561 2008
[71] K Meyer A Banerjee S E Frey R B Belshe and R Ray ldquoAweak neutralizing antibody response to hepatitis C virus enve-lope glycoprotein enhances virus infectionrdquo PLoS ONE vol 6no 8 Article ID e23699 2011
[72] R Burioni R AWilliamson P P Sanna F E Bloom and D RBurton ldquoRecombinant human Fab to glycoprotein D neutral-izes infectivity and prevents cell-to-cell transmission of herpessimplex viruses 1 and 2 in vitrordquo Proceedings of the NationalAcademy of Sciences of the United States of America vol 91 no1 pp 355ndash359 1994
[73] M S Rolph A Zaid N E Rulli and S Mahalingam ldquoDown-regulation of interferon-120573 in antibody-dependent enhancementof dengue viral infections of human macrophages is dependenton interleukin-6rdquo Journal of Infectious Diseases vol 204 no 3pp 489ndash491 2011
[74] S S Whitehead J E Blaney A P Durbin and B R MurphyldquoProspects for a dengue virus vaccinerdquoNature ReviewsMicrobi-ology vol 5 no 7 pp 518ndash528 2007
[75] S-M Lok V Kostyuchenko G ENybakken et al ldquoBinding of aneutralizing antibody to dengue virus alters the arrangement ofsurface glycoproteinsrdquoNature Structural andMolecular Biologyvol 15 no 3 pp 312ndash317 2008
[76] A P Goncalvez R Men C Wernly R H Purcell and C-J Lai ldquoChimpanzee Fab fragments and a derived humanizedimmunogiobulin G1 antibody that efficiently cross-neutralizedengue type 1 and type 2 virusesrdquo Journal of Virology vol 78no 23 pp 12910ndash12918 2004
[77] WM P BWahala A A Kraus L B HaymoreM A Accavitti-Loper and A M de Silva ldquoDengue virus neutralization byhuman immune sera role of envelope protein domain III-reactive antibodyrdquo Virology vol 392 no 1 pp 103ndash113 2009
[78] W D Crill H R Hughes M J Delorey and G-J J ChangldquoHumoral immune responses of dengue fever patients usingepitope-specific serotype-2 virus-like particle antigensrdquo PLoSONE vol 4 no 4 Article ID e4991 2009
[79] C Setthapramote T Sasaki O Puiprom et al ldquoHuman mono-clonal antibodies to neutralize all dengue virus serotypes usinglymphocytes from patients at acute phase of the secondaryinfectionrdquo Biochemical and Biophysical Research Communica-tions vol 423 no 4 pp 867ndash872 2012
[80] M Beltramello K L Williams C P Simmons et al ldquoThehuman immune response to dengue virus is dominated byhighly cross-reactive antibodies endowed with neutralizing andenhancing activityrdquo Cell Host andMicrobe vol 8 no 3 pp 271ndash283 2010
[81] C M Midgley M Bajwa-Joseph S Vasanawathana et al ldquoAnin-depth analysis of original antigenic sin in dengue virusinfectionrdquo Journal of Virology vol 85 no 22 pp 410ndash421 2011
[82] B Kaufmann M R Vogt J Goudsmit et al ldquoNeutralizationof West Nile virus by cross-linking of its surface proteins withFab fragments of the human monoclonal antibody CR4354rdquoProceedings of the National Academy of Sciences of the UnitedStates of America vol 107 no 44 pp 18950ndash18955 2010
[83] B Spurrier J M Sampson M Totrov et al ldquoStructural analysisof human and macaque mAbs 2909 and 25B implicationsfor the configuration of the quaternary neutralizing epitope ofHIV-1 gp120rdquo Structure vol 19 no 5 pp 691ndash699 2011
[84] J J SchlesingerMWBrandriss andT PMonath ldquoMonoclonalantibodies distinguish between wild and vaccine strains ofyellow fever virus by neutralization hemagglutination inhibi-tion and immune precipitation of the virus envelope proteinrdquoVirology vol 125 no 1 pp 8ndash17 1983
[85] M W Brandriss J J Schlesinger E E Walsh and M BrisellildquoLethal 17D yellow fever encephalitis in mice I Passive protec-tion by monoclonal antibodies to the envelope proteins of 17Dyellow fever and dengue 2 virusesrdquo Journal of General Virologyvol 67 no 2 pp 229ndash234 1986
[86] H Cho and M S Diamond ldquoImmune responses to West Nilevirus infection in the central nervous systemrdquo Viruses vol 4no 12 pp 3812ndash3830 2012
[87] D W Beasley A D Barrett and R B Tesh ldquoResurgence ofWest Nile neurologic disease in the United States in 2012 whathappened What needs to be donerdquo Antiviral Research vol 99no 1 pp 1ndash5 2013
[88] M Throsby J Ter Meulen C Geuijen J Goudsmit and J DeKruif ldquoMapping and analysis of West Nile virus-specific mon-oclonal antibodies prospects for vaccine developmentrdquo ExpertReview of Vaccines vol 6 no 2 pp 183ndash191 2007
[89] M S Diamond T G Roberts D Edgil B Lu J Ernst and EHarris ldquoModulation of dengue virus infection in human cells
20 BioMed Research International
by alpha beta and gamma interferonsrdquo Journal of Virology vol74 no 11 pp 4957ndash4966 2000
[90] Z Shimoni M J Niven S Pitlick and S Bulvik ldquoTreatmentof West Nile virus encephalitis with intravenous immunoglob-ulinrdquo Emerging Infectious Diseases vol 7 no 4 article 759 2001
[91] AHamdan PGreen EMendelsonM R Kramer S Pitlik andM Weinberger ldquoPossible benefit of intravenous immunoglob-ulin therapy in a lung transplant recipient with West Nile virusencephalitisrdquoTransplant InfectiousDisease vol 4 no 3 pp 160ndash162 2002
[92] M Haley A S Retter D Fowler J Gea-Banacloche and NP OrsquoGrady ldquoThe role for intravenous immunoglobulin in thetreatment of West Nile virus encephalitisrdquo Clinical InfectiousDiseases vol 37 no 6 pp e88ndashe90 2003
[93] K M Chung G E Nybakken B S Thompson et al ldquoAnti-bodies against West Nile virus nonstructural protein NS1prevent lethal infection through Fc 120574 receptor-dependent and-independent mechanismsrdquo Journal of Virology vol 80 no 3pp 1340ndash1351 2006
[94] E A Henchal L S Henchal and J J Schlesinger ldquoSynergisticinteractions of anti-NS1 monoclonal antibodies protect pas-sively immunized mice from lethal challenge with dengue 2virusrdquo Journal of General Virology vol 69 no 8 pp 2101ndash21071988
[95] J T Roehrig ldquoAntigenic structure of flavivirus proteinsrdquoAdvances in Virus Research vol 59 pp 141ndash175 2003
[96] M D Sanchez T C Pierson D McAllister et al ldquoCharacteri-zation of neutralizing antibodies to West Nile virusrdquo Virologyvol 336 no 1 pp 70ndash82 2005
[97] J H Beigel J L Nordstrom S R Pillemer et al ldquoSafety andpharmacokinetics of single intravenous dose of MGAWN1 anovel monoclonal antibody to West Nile virusrdquo AntimicrobialAgents and Chemotherapy vol 54 no 6 pp 2431ndash2436 2010
[98] C A Smeraski V Siddharthan and J D Morrey ldquoTreatmentof spatial memory impairment in hamsters infected with WestNile virus using a humanizedmonoclonal antibodyMGAWN1rdquoAntiviral Research vol 91 no 1 pp 43ndash49 2011
[99] D Lelli AMoreno E Brocchi et al ldquoWestNile virus character-ization and diagnostic applications of monoclonal antibodiesrdquoVirology Journal vol 9 article 81 2012
[100] T S Gritsun V A Lashkevich and E A Gould ldquoTick-borneencephalitisrdquo Antiviral Research vol 57 no 1-2 pp 129ndash1462003
[101] D Ruzek T S Gritsun N L Forrester et al ldquoMutations inthe NS2B and NS3 genes affect mouse neuroinvasiveness of aWestern European field strain of tick-borne encephalitis virusrdquoVirology vol 374 no 2 pp 249ndash255 2008
[102] M Ecker S L Allison T Meixner and F X Heinz ldquoSequenceanalysis and genetic classification of tick-borne encephalitisviruses from Europe and Asiardquo Journal of General Virology vol80 no 1 pp 179ndash185 1999
[103] H Holzmann M S Vorobyova I P Ladyzhenskaya et alldquoMolecular epidemiology of tick-borne encephalitis viruscross-protection between European and Far Eastern subtypesrdquoVaccine vol 10 no 5 pp 345ndash349 1992
[104] C W Mandl ldquoSteps of the tick-borne encephalitis virus repli-cation cycle that affect neuropathogenesisrdquo Virus Research vol111 no 2 pp 161ndash174 2005
[105] A V Atrasheuskaya T M Fredeking and G M IgnatyevldquoChanges in immune parameters and their correction in humancases of tick-borne encephalitisrdquo Clinical and ExperimentalImmunology vol 131 no 1 pp 148ndash154 2003
[106] K L Mansfield N Johnson L P Phipps J R Stephenson AR Fooks and T Solomon ldquoTick-borne encephalitis virusmdashareview of an emerging zoonosisrdquo Journal of General Virologyvol 90 no 8 pp 1781ndash1794 2009
[107] DWVaughn andCHHoke Jr ldquoThe epidemiology of Japaneseencephalitis prospects for preventionrdquo Epidemiologic Reviewsvol 14 pp 197ndash221 1992
[108] T Solomon H Ni D W C Beasley M Ekkelenkamp M JCardosa and A D T Barrett ldquoOrigin and evolution of Japaneseencephalitis virus in Southeast Asiardquo Journal of Virology vol 77no 5 pp 3091ndash3098 2003
[109] M R Wilson ldquoEmerging viral infectionsrdquo Current Opinion inNeurology vol 26 no 3 pp 301ndash306 2013
[110] J N Hanna S A Ritchie D A Phillips et al ldquoAn outbreakof Japanese encephalitis in the Torres Strait Australia 1995rdquoMedical Journal of Australia vol 165 no 5 pp 256ndash260 1996
[111] J N Hanna S A Ritchie D A Phillips et al ldquoJapanese ence-phalitis in north Queensland Australia 1998rdquo Medical Journalof Australia vol 170 no 11 pp 533ndash536 1999
[112] SGeorgeMGourie-Devi and J A Rao ldquoIsolation ofWestNilevirus from the brains of children who had died of encephalitisrdquoBulletin of theWorldHealthOrganization vol 62 no 6 pp 879ndash882 1984
[113] A K Gupta A A Koshy and V J Lad ldquoEnhanced protec-tion of mice against Japanese encephalitis virus infection bycombinations ofmonoclonal antibodies to glycoprotein ErdquoActaVirologica vol 55 no 2 pp 165ndash168 2011
[114] T H Lee B-H Song S-I Yun et al ldquoA cross-protective mAbrecognizes a novel epitope within the flavivirus NS1 proteinrdquoJournal of General Virology vol 93 no 1 pp 20ndash26 2012
[115] S C Weaver ldquoHost range amplification and arboviral diseaseemergencerdquo Archives of Virology Supplementum no 19 pp 33ndash44 2005
[116] M Solignat B Gay S Higgs L Briant and C Devaux ldquoRepli-cation cycle of chikungunya a re-emerging arbovirusrdquoVirologyvol 393 no 2 pp 183ndash197 2009
[117] O Schwartz and M L Albert ldquoBiology and pathogenesis ofchikungunya virusrdquo Nature Reviews Microbiology vol 8 no 7pp 491ndash500 2010
[118] W Akahata Z-Y Yang H Andersen et al ldquoA virus-like particlevaccine for epidemic Chikungunya virus protects nonhumanprimates against infectionrdquo Nature Medicine vol 16 no 3 pp334ndash338 2010
[119] M Kielian and A Helenius ldquopH-Iinduced alterations in thefusogenic spike protein of Semliki Forest virusrdquo Journal of CellBiology vol 101 no 6 pp 2284ndash2291 1985
[120] J White and A Helenius ldquopH-Dependent fusion between theSemliki Forest virus membrane and liposomesrdquo Proceedings ofthe National Academy of Sciences of the United States of Americavol 77 no 6 pp 3273ndash3277 1980
[121] J Dubuisson and C M Rice ldquoSindbis virus attachment isola-tion and characterization of mutants with impaired binding tovertebrate cellsrdquo Journal of Virology vol 67 no 6 pp 3363ndash33741993
[122] C Y Lee Y-W Kam J Fric et al ldquoChikungunya virus neutral-ization antigens and direct cell-to-cell transmission are revealedby human antibody-escapemutantsrdquo PLoS Pathogens vol 7 no12 Article ID e1002390 2011
[123] J E Voss M-C Vaney S Duquerroy et al ldquoGlycoproteinorganization of Chikungunya virus particles revealed by X-raycrystallographyrdquo Nature vol 468 no 7324 pp 709ndash712 2010
BioMed Research International 21
[124] L Li J Jose Y Xiang R J Kuhn and M G Rossmann ldquoStruc-tural changes of envelope proteins during alphavirus fusionrdquoNature vol 468 no 7324 pp 705ndash708 2010
[125] J Fric S Bertin-Maghit C I Wang A Nardin and L WarterldquoUse of human monoclonal antibodies to treat Chikungunyavirus infectionrdquo Journal of Infectious Diseases vol 207 no 2 pp319ndash322 2013
[126] P V Aguilar J G Estrada-Franco R Navarro-Lopez C FerroA D Haddow and S C Weaver ldquoEndemic Venezuelanequine encephalitis in the Americas hidden under the dengueumbrellardquo Future Virology vol 6 no 6 pp 721ndash740 2011
[127] S C Weaver R Winegar I D Manger and N L ForresterldquoAlphaviruses population genetics and determinants of emer-gencerdquo Antiviral Research vol 94 no 3 pp 242ndash257 2012
[128] S Paessler and S C Weaver ldquoVaccines for Venezuelan equineencephalitisrdquo Vaccine vol 27 no 4 pp D80ndashD85 2009
[129] R J Phillpotts ldquoVenezuelan equine encephalitis virus complex-specific monoclonal antibody provides broad protection inmurine models against airborne challenge with viruses fromserogroups I II and IIIrdquoVirus Research vol 120 no 1-2 pp 107ndash112 2006
[130] R J Phillpotts L D Jones and S C Howard ldquoMonoclonalantibody protectsmice against infection anddiseasewhen giveneither before or up to 24 h after airborne challenge with virulentVenezuelan equine encephalitis virusrdquoVaccine vol 20 no 11-12pp 1497ndash1504 2002
[131] J H Mathews J T Roehrig and D W Trent ldquoRole of com-plement and the Fc portion of immunoglobulin G in immunityto Venezuelan equine encephalomyelitis virus infection withglycoprotein-specific monoclonal antibodiesrdquo Journal of Virol-ogy vol 55 no 3 pp 594ndash600 1985
[132] L M OrsquoBrien C D Underwood-Fowler S A Goodchild AL Phelps and R J Phillpotts ldquoDevelopment of a novel mon-oclonal antibody with reactivity to a wide range of Venezuelanequine encephalitis virus strainsrdquoVirology Journal vol 6 article206 2009
[133] L M OrsquoBrien S A Goodchild R J Phillpotts and S D Per-kins ldquoA humanised murine monoclonal antibody protectsmice from Venezuelan equine encephalitis virus Evergladesvirus and Mucambo virus when administered up to 48h afterairborne challengerdquo Virology vol 426 no 2 pp 100ndash105 2012
[134] N Clementi N Mancini L Solforosi M Castelli M Clementiand R Burioni ldquoPhage display-based strategies for cloning andoptimization of monoclonal antibodies directed against humanpathogensrdquo International Journal of Molecular Sciences vol 13no 7 pp 8273ndash8292 2012
[135] A R Hunt R A Bowen S Frederickson T Maruyama JT Roehrig and C D Blair ldquoTreatment of mice with humanmonoclonal antibody 24h after lethal aerosol challenge withvirulent Venezuelan equine encephalitis virus prevents diseasebut not infectionrdquo Virology vol 414 no 2 pp 146ndash152 2011
[136] W-G Hu A L Phelps S Jager et al ldquoA recombinant human-ized monoclonal antibody completely protects mice againstlethal challenge with Venezuelan equine encephalitis virusrdquoVaccine vol 28 no 34 pp 5558ndash5564 2010
[137] T Rulker L Voss P Thullier et al ldquoIsolation and characterisa-tion of a human-like antibody fragment (scFv) that inactivatesVEEV in vitro and in vivordquo PLoS ONE vol 7 no 5 Article IDe37242 2012
[138] C T Walter and J N Barr ldquoRecent advances in the molecularand cellular biology of bunyavirusesrdquo Journal of General Virol-ogy vol 92 no 11 pp 2467ndash2484 2011
[139] C A Whitehouse ldquoCrimean-Congo hemorrhagic feverrdquo Anti-viral Research vol 64 no 3 pp 145ndash160 2004
[140] M Mertens K Schmidt A Ozkul and M H Groschup ldquoTheimpact of Crimean-Congo hemorrhagic fever virus on publichealthrdquo Antiviral Research vol 98 no 2 pp 248ndash260 2013
[141] A A Ahmed J M McFalls C Hoffmann et al ldquoPresence ofbroadly reactive and group-specific neutralizing epitopes onnewly described isolates of Crimean-Congo hemorrhagic fevervirusrdquo Journal of General Virology vol 86 no 12 pp 3327ndash33362005
[142] R Hewson A Gmyl L Gmyl et al ldquoEvidence of segment reas-sortment in Crimean-Congo haemorrhagic fever virusrdquo Journalof General Virology vol 85 no 10 pp 3059ndash3070 2004
[143] N K Blackburn T G Besselaar A J Shepherd and R Swane-poel ldquoPreparation and use of monoclonal antibodies for iden-tifying Crimean-Congo hemorrhagic fever virusrdquo AmericanJournal of Tropical Medicine and Hygiene vol 37 no 2 pp 392ndash397 1987
[144] T Ikegami ldquoMolecular biology and genetic diversity of RiftValley fever virusrdquoAntiviral Research vol 95 no 3 pp 293ndash3102012
[145] KKeegan andM S Collett ldquoUse of bacterial expression cloningto define the amino acid sequences of antigenic determinantson the G2 glycoprotein of Rift Valley fever virusrdquo Journal ofVirology vol 58 no 2 pp 263ndash270 1986
[146] T G Besselaar and N K Blackburn ldquoTopological mapping ofantigenic sites on the Rift Valley fever virus envelope glycopro-teins using monoclonal antibodiesrdquo Archives of Virology vol121 no 1ndash4 pp 111ndash124 1991
[147] L G Presta ldquoEngineering of therapeutic antibodies to mini-mize immunogenicity and optimize functionrdquo Advanced DrugDelivery Reviews vol 58 no 5-6 pp 640ndash656 2006
Submit your manuscripts athttpwwwhindawicom
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anatomy Research International
PeptidesInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporation httpwwwhindawicom
International Journal of
Volume 2014
Zoology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Molecular Biology International
GenomicsInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioinformaticsAdvances in
Marine BiologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Signal TransductionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Evolutionary BiologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Biochemistry Research International
ArchaeaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Genetics Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in
Virolog y
Hindawi Publishing Corporationhttpwwwhindawicom
Nucleic AcidsJournal of
Volume 2014
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Enzyme Research
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Microbiology

2 BioMed Research International
can be protective against homologous or heterologous denguevirus (DENV) challenge as well as against other flavivirusesand human arboviruses Moreover engineering renderingmAbs capable of crossing the blood-brain barrier in order tolimit viral dissemination within CNS may be considered
Finally a possible administration of mAbs in those sub-jects that could be at risk of exposure to arbovirus infectionssuch as travelers in endemic areas could reduce the possi-ble incidence and consequent augmented risk of epidemicepisodes
In this review we describe the major clinical relevant andworldwide diffused arboviruses infecting humans and therecentlymajor describedmAbs to be possibly used in a futurepassive immunotherapy
2 Flaviviruses
The Flavivirus genus including more than 70 viruses is theonly onewithin the Flaviviridae family which holds arboviru-ses that are responsible for significant morbidity and mortal-ity worldwide [12]
About 25 billion people are at risk of infection in tropicaland subtropical countries mainly South-East and SouthAsiaCentral and South America and the Caribbean In additionmultiple Flavivirus infections have been reported in the sameareas complicating early diagnosis and identification [13]
Flavivirus infections can cause fever encephalitis hem-orrhagic disease flaccid paralysis and death in humansHowever the immunopathogenesis of these viruses is notfully understood In the last decade the flaviviruses havereemerged as aggressive human pathogens [13]
The human flaviviruses includes 53 recognized speciesHowever five of them are considered clinically important likeDENV YFV JEV TBEV and West Nile virus (WNV)
The genome of all the members of the Flaviviridae familyconsists of a 95ndash125 kb positive-sense single-stranded RNAThey are enveloped small virions (40ndash60 nm in diameter)with two or more species of envelope glycoproteins (eg Mand E proteins) which are involved in the binding and fusionprocesses In particular the precursor of the mature M pro-tein (prM) interacts with E glycoproteins acting as a chaper-one and preventing the fusion of the viruswith themembranein the cell during egress through acidic compartments ofthe secretory pathway Then cleavage of prM by the cellularprotease furin during transit through the Golgi network is arequired step in the viral lifecycle that defines the transitionfrom an immature non-infectious virus particle into an infec-tious form However immature infectious virions retainingsome uncleaved prM molecules could be released [14]
The M and E glycoproteins constitute an icosahedralscaffold surrounding a nucleocapsid which consists of theviral genome complexed with a core of approximately 30 nmcomposed of multiple copies of a small basic capsid (C) pro-tein Binding uptake and fusion by target cells are believed toinvolve clathrin-mediated and low-pH-induced endocytosis[14]
Flaviviruses can utilize multiple receptors for differentcell types and host species They are thought to firstly inter-act with dendritic cells through DC-SIGN and L-SIGN
binding of glycans on E glycoprotein dimers In additionhighly sulfated glycosaminoglycans (eg heparan sulfate)have been demonstrated to play an important role in theinitial attachment of several flaviviruses to their target cellsOther molecules identified as possible receptors are inte-grins mannose-binding receptor on macrophages laminin-binding protein GRP78 (BiP) and CD14 [15]
Uncoating and replication of the viral genome througha minus-strand RNA intermediate occurs in the cyto-plasmic replication complexes associated with perinuclearmembranes where viral proteins are produced as part of asingle long polyprotein of more than 3000 amino acidsgenerating three structural (C prM and E) and sevennon-structural (NS1 NS2A NS2B the viral protease NS3NS4A NS4B and the RNA-dependent RNA polymeraseNS5) proteins by cleavage of host and viral proteases respec-tively Progeny virions are thought to assemble by bud-ding into an intracellular membrane compartment proba-bly the endoplasmic reticulum then transited through thehost secretory pathway and released at the cell surface[16]
Efforts to develop effective prophylactic approaches forseveral clinically important flaviviruses are underway [17]The crucial role of the humoral immune response againstFlavivirus infections is well established as infection withone serotype provides life-long protective immunity to thehomologous infecting serotype and cross-protection in thefirst few months against the other serotypes Converselyindividuals experiencing a secondary infectionwith a distinctserotype are at greater risk of severe complications asdiscussed later [18]
Only a limited number of Flavivirus vaccines are availabletoday however no approved antiviral drugs are yet availablefor their clinical use Thus giving the lack of vaccines aswell as specific antiviral drugs broadly cross-neutralizingmAbs could be helpful in the development of an effectivetherapeutic strategy against these infections as well as in theprogress towards effective immunogens
In the next paragraphs we describe the molecular targetsof flaviviruses including DENV JEV TBEV WNV YFV andSt Louis encephalitis virus (SLEV) followed by a descriptionof the therapeutic candidate mAbs directed against them
21 Domains and Functions of the Surface Envelope (E) Glyco-protein of Flaviviruses Themajor target of the host humoralimmune response and of neutralizing Abs against flavivirusesis represented by the envelope (E) glycoprotein which is a56-kDa protein and the major represented antigen on thesurface of virions [19] However Ab response towards otherstructural and non-structural proteins such as prM and NS1respectively has also been described [20 21] However prM-specific Abs display limited neutralizing activity while anti-NS1 Abs have been detected only during the convalescentperiod of a primary DENV infection but were stronglyidentified during the acute phase of a secondary DENVinfection suggesting that both Abs could contribute to thepathogenesis of the life-threatening dengue hemorrhagicfeverdengue shock syndrome (DHFDSS) [22]
BioMed Research International 3
The crystallographic structure of the E protein of TBEVhas been proposed as a model for the envelope protein of theflaviviruses [23]
Further studies performed throughX-ray crystallographyhave revealed that the E protein is a type II viral fusionprotein with three 120573-barrel domains (D) DI DII and DIII(corresponding to the antigenic domains C A and B resp)connected to the viral membrane by a helical structurecalled the stem anchor [24] In its native form the E proteinfolds as an homodimer with an antiparallel structure andan unusual herringbone pseudoicosaedral symmetry patternwith the M protein located centrally within the symmetryconsistent with a head-to-tail configuration lying parallel tothe envelope lipid bilayer The DIII of the E glycoprotein(amino acid residues 295ndash395) has an immunoglobulin-like fold contains seven 120573-strands as well as type- andsubcomplex-specific neutralizing B-cell epitopes and is theproposed receptor-binding domain through four peptideloops on the solvent exposed face Directly linked to DIIIthere is the stemtransmembrane region spanning aminoacid residues 401ndash495 and containing regions important foroligomerization with prM protein (amino acid residues 431ndash449 and 450ndash472) [25] DII has a long finger-like structureand contains two extended loops that project from DI andthe highly conserved and hydrophobic glycine-rich fusionloop at its tip (amino acid residues 98ndash110) interacting withthe stem region and the endosomal membrane of the cellduring the fusion process [26] In the trimeric conformationthe hydrophobic fusion peptide is exposed in DII to mediatethe fusion with the membranes of the cell DI is a sim120amino acid central domain formed by a discontinuous hingeregion consisting in a central 120573-barrel of eight strands thatconnectsDII andDIII andplays a crucial role in the structuralrearrangement of E from a homodimer to a trimer whichoccurs on exposure to low pH of endosomes that is requiredfor the membrane fusion process In some flaviviruses anN-linked glycosylation site is present at amino acid position154ndash156 of the E protein and does not appear to be necessaryfor E function but it has been associated with the increasedneuroinvasiveness of WNV lineage I outbreak strains fromthe USA and a pH-sensitive decrease in stability of the non-glycosylated strains [27]
22 Flavivirus Neutralizing B-Cell Epitopes As previouslydescribed Abs are a significant component of the hostrsquosprotective response against Flavivirus infections Howevervirus-specific Abs have been implicated in the pathogenesisof severe clinicalmanifestations following a secondaryDENVas well as YFV andWNV infections
Moreover cross-reactivity between Flavivirus serogroupscould complicate the interpretation of diagnostic assays butcould be of interest in isolating cross-reactive and cross-neutralizing mAbs However Ab-dependent enhancement(ADE) of infection has been described for this genus as itrepresents the major risk of complications when reinfectionoccursThus great caremust be taken in evaluating neutraliz-ing activity and possibleAb-mediated infection enhancementin the characterization of future possible therapeutic mAbsagainst these viruses [28]
However prophylactic and therapeutic use of neutral-izing mAbs for Flavivirus infections has been shown to beeffective in animal models as reported later
Neutralizing epitopes were found to be located in each ofthe three domains of the E protein and have been confirmedto be surface exposed in the high-resolution X-ray crystalstructure of the pre-fusion dimer of DENV2 E glycoprotein[29]
Most potentmAbs against flaviviruses are directed towardepitopes on DIII For this reason DIII-based immunogensare under evaluation as promising subunit Flavivirus candi-date vaccines [30 31] However several cross-reactive mAbsthat bind to residues from the AB loop were found to bepoorly neutralizing as this loop projects toward the lipidbilayer in the mature viral particle Conversely lateral ridgeregion of DIII (eg BC DE and FG loops) is targeted bystrong serotype-specific neutralizing mAbs [32ndash35] Cross-reactive mAbs specific for DII have also been described butwith less neutralizing and variable profiles However few ofthem whose epitope encompasses the fusion loop are cross-reactive and neutralizing [36]
23 Ab-Dependent Enhancement (ADE) of Infection In vitroand in vivo studies have demonstrated that increased diseaseseverity causing DHFDSS upon reinfection with a differentDENV serotype is mostly due to the phenomenon of ADE ofinfection determined by cross-reacting but poorly or non-neutralizing Abs generated during the primary infectionthat facilitate virus entry through Fc-120574 or complementreceptors present on cells such as monocytesmacrophages(Figure 1(a)) Additionally in 6 to 9 month-old children ofDENV immune mothers severe disease is associated withprimary infection possibly because of declining levels ofneutralizing maternal Abs [64]
These cross-reactive Abs are mainly directed against theE and prM glycoproteins In particular experimental lines ofevidence have suggested that cross-reactiveAbs againstDII ofE protein can under certain conditions enhance infectivityof WNV in vitro [65] However the phenomenon of ADEcould reflect the presence of non-neutralizing concentrationsof virus-reactive Abs and has been observed for severalflaviviruses in vitro and in vivo such as WNV TBEV JEVand YFV as well as other Flaviviridae members [28 66ndash68]In particular it has been described that ADE of infectionoccurs at virus-boundedAb concentrations in the upper limitby the stoichiometric threshold for neutralization and atlower concentrations by the minimal number of Abs thatallow attachment of the virion to cells Furthermore Abs thatrecognize infrequently displayed epitopes that do not supportneutralizationmay enhance infection even at saturation [69]
Thus the ADE of infection mechanismmay pose a threatfor the development of a safe and efficacious vaccine as wellas a possible immunotherapy against Flaviviridae
Indeed cross-neutralizing mAbs as candidate therapeu-tics for these infections may be considered after having wellascertained their broadly cross-neutralizing activity and theabsence of possible mAb-mediated ADE of infection mech-anisms Alternatively the risk of ADE of infection could beovertaken by the removal of the Ab heavy chain (ie the CH2
4 BioMed Research International
receptorFc-120574
Cellular receptor
(a)
Cellular receptor
(b) (c)
Figure 1 Mechanisms of antibody-dependent enhancement (ADE) of infection (a) After binding to the viral epitope the Ab is recognizedby a cellular Fc-120574 receptor bringing the viral particle in proximity of the target cell (b) the binding of the Ab induces conformational changeswithin the structure of the viral target protein These changes improve the affinity for the cellular receptor (c) molecular mimicry by a viralmotif of cellular membrane components leads an autoreactive Ab to bind both the viral and the cellular target bringing the virus in proximityof the target cell
and CH3 portions or direct expression of the mAb as aFab fragment) andor deletion of the N-linked sugars onIgG molecules that are both required for interactions withFc-120574 receptors (Fc-120574R) or eventually by the blocking ofFc-120574R engagement with anti-Fc-120574R Abs [70] Similarly theidentification and detailed examination of cross-neutralizingepitopes that do not promote ADE of infection may definenovel targets for vaccine development
However in some instances ADE of infection mayoccur when Ab molecules cross-react with both viral andcellular antigens (Figure 1(c)) In this regard Fc-120574 receptor-independentmechanismof infection in particular forDENVprM-specific Abs able to bind simultaneously to the virus andtarget cells has been described Indeed othermembers of theFlaviviridae belonging to another genus like hepatitis C virus(HCV) have been shown to elicit polyreactive Abs that areprobably responsible for an ADE of infection mechanism aswell as of secondary clinical manifestations In connectionwith this several studies employing both polyclonal andmonoclonal Abs suggested the presence of common epitopescontaining particular short motifs on NS1 and the threedomains of the E protein as well as human proteins thatmay play a role in ADE of infection mechanisms and thusin DHFDSS pathogenesis [70] Interestingly these commonmotifs have been found to be additionally represented inDENV2 strains which have greater human pathogenic capac-ities [22]
Moreover another molecular mechanism of ADE ofinfection could consist in the facilitation of conformationalchanges of the targeted surface protein that are required forvirus entry as described and hypothesized for other viruses(eg HIV HCV and HSV) (Figure 1(b)) [71 72]
Finally Fc-120574R engagement could result also in ldquointrinsicrdquoADE of infection mechanisms involving signaling eventssuch as inhibition of antiviral response and increased viralreplication Disrupted signaling events include the RIG-IMDA5 cascade and type I interferon (IFN) response aswell as induction and suppression of anti-inflammatory(IL-10) and proinflammatory (IL-12 and TNF-120572) cytokinesrespectively [73]
24 Dengue Virus (DENV) Dengue virus (DENV) is respon-sible for 50ndash100 million symptomatic infections each yearresulting in 500000 hospitalizations and over 20000 deathswhich occurs mostly in tropical and subtropical regions ofthe world Although malaria remains the most importantcause for systemic febrile disease in travelers chikungunyavirus (CHIKV) and DENV are increasingly diagnosed withdengue currently being the second most important cause forfebrile disease in travelers
Decreases in mosquito control efforts during the endof the 90s coupled with societal factors (eg globalizationmigrations and dense urbanization) have contributed to thereemergence of flaviviruses such as DENV in South and Cen-tral America Development of effective DENV vaccines thatexhibit cross-protection thought to be important for prevent-ing subsequent dengue-associated immunopathogenesis isproving to be particularly challenging [1]
A vaccine for this infection is not yet available despiteconsiderable public and private efforts This difficulty ismainly due to the perceived need to simultaneously protectthe four known serotypes of DENV (DENV1ndash4) which shareabout 70 of sequence homology while genotypes can varyup to 3 Moreover as anticipated and discussed previously
BioMed Research International 5
non-protective Abs may contribute to more severe clinicaloutcomes in vaccinated people [74]
241 Murine versus Human Anti-DENV Humoral ImmuneResponse It is well ascertained that anti-E DIII-directed Absare virtually absent from the naıve human repertoire as theyare directed away from this domain probably against weaklyneutralizing and immunodominant regions Moreover it hasbeen demonstrated that over than 90 of the human Abresponse of primary DENV infected patients is able to bindonly native DENV virus particles instead of a recombinantform of the E protein posing difficulties in the cloning strate-gies of neutralizing mAbs [39] Thus it can be concludedthat in vivo only a small fraction of DENV-specific Abs areresponsible for neutralization
Conversely murine mAbs that recognize all the threedomains of E have been identified Epitope-mapping studiesusingmurine serotype-specificmAbs have demonstrated thatloops located in the lateral ridge region of DIII consti-tute the strongest neutralizing B-cell target In this regionsequence diversity between serotype is high while cross-reactive murine mAbs that recognize another loop showedgenerally a weaker neutralization profile In particular type-specific mAbs with neutralizing activity against DENV2localized to the BC DE and FG loops on the lateral ridgeof DIII whereas subcomplex-specific mAbs recognized anadjacent epitope centered on the connecting A strand of DIIIat residues K305 K307 and K310 [32ndash35]
Cross-reactive mAbs directed against prM and NS1 havealso been described but as anticipated they feature weaklyneutralizing or absent as well as ADE of infection activityIn the following paragraphs we report the better character-ized neutralizing mouse and human mAbs describing theirmolecular features further summarized in Table 1
242 Anti-DENV Murine mAbs Murine mAbs directedagainst all the three domains of the E protein and endowedof different neutralizing and binding characteristics havebeen described However only DIII and DII are classicallyrecognized by neutralizing Abs In this regard Sukupolvi-Petty et al isolated twenty-four anti-DENV2 mouse mAbswith moderate or strong neutralizing activity against thehomologous virus in cell culture assays Binding sites weremapped for the majority of these in distinct epitopes inregions located in DI (lateral ridge) DII (dimer interfacelateral ridge and fusion loop) and DIII (lateral ridge CC1015840loop and A strand) Moreover 16 of the neutralizing mAbswere tested in mice with most of them being protectivewhen given as prophylaxis Seven of these had post-exposuretherapeutic activity when administered as a single dose byintraperitoneal route even 3 days after intracranial infectionFor the mAbs with the greatest therapeutic potential protec-tion was confirmed with an Ab-enhanced vascular leakagemouse model of DENV2 infection [35]
Recently Deng et al described a cross-reactive murinemAb named 2A10G6 that is able to recognize DENV1ndash4 YFV WNV JEV and TBEV to potently cross-neutralizeDENV1ndash4 YFV and to a lesser extent WNV This mAbrecognizes a highly conserved motif (amino acid residues
98ndash101) located within the fusion loop of the FlavivirusDII of the E glycoprotein Moreover this mAb exerts itsneutralizing activity in a post-attachment step during thevirus entry process as demonstrated by kinetic neutralizationtests performed in vitro Additionally protection experimentsperformed in mouse models showed that treatment with100 120583gmL of 2A10G6 conferred full protection against lethalDENV2 challenge and 20 120583gmL and 4120583gmL of 2A10G6protected 89 and 40 of infected mice from lethal chal-lenge respectively For infection with DENV1 3 and 4 use of100 120583gmL of 2A10G6 conferred partial protection and 5377 and 73 of the infected mice survived after challengerespectively Finally the protection profile of 2A10G6 againstWNV showed that prophylactic administration with a singledose of 200mg of 2A10G6 conferred 80 protection in miceMost importantly 3 of 8 (375) mice survived when 2A10G6was administered one day after WNV challenge SimilarlyanothermousemAb 4G2 is able to recognize the fusion loopat the extremity of DII from all four DENV serotypes and toprevent syncytia formation [36]
Rajamanonmani et al described a mouse mAb 9F12raised against theDIII of DENV2E protein that is able to rec-ognize DENV1ndash4 serotypes as well asWNV and to neutralizefive DENV strains representative of all DENV serotypes [41]Similarly Cockburn et al reported a comparative high-resolution crystallographic analysis of an A-strand DIIImurinemAb 4E11 in complex with its target domain of the Eprotein from the four DENV serotypes MAb 4E11 is capableof recognizing and neutralizing all four serotypes with IC50values varying between 1 and 100 nMThe structures reportedalso highlight the mechanism by which anti-A-strand mAbsdisrupt the architecture of the mature virion inducing dimerdissociation premature fusion-loop exposure and concomi-tant particle inactivation [40] Also Midgley et al function-ally characterized another murine mAb 2H12 raised in miceagainstDIII of E protein Similarly to the previously describedmAbs 2H12 is able to bind all the four DENV serotypes ina epitope encompassing the conserved 314ETQH317 motifHowever the neutralizing potential of 2H12 is lower than anumber of other anti-DIII mAbs with IC50 values rangingfrom 056 to 145 nM for DENV1 2 and 4 On the other handit showed no ADE of infection activity [38]
Austin et al isolated a murine mAb named E111 whichrecognize a novel CC1015840-loop epitope on DIII of the E proteinfrom two different DENV1 genotypes Docking of the mAbstructure onto the available cryoelectron microscopy modelsof DENV virions revealed that the E111 epitope was inaccessi-ble suggesting that this mAb recognizes an uncharacterizedvirus conformation While the affinity of binding betweenE111 and DIII varied by genotype a limited correlation withneutralizing activity was observed These data support theconclusion that potent neutralization depends on genotype-dependent exposure of the CC1015840 loop epitope [49] In fact aspreviously described elsewhere binding of some E reactiveAbs depends on the dynamicmovement of proteinmoleculesldquobreathingrdquo in the virion particle leading to transient expo-sure of hidden epitopes For instance optimal binding ofmousemAb 1A1D-2 to EDIII requires incubation at 37∘C [75]
6 BioMed Research InternationalTa
ble1Schematicsummaryof
theb
estcharacterized
mAb
sagainstarbo
viralp
atho
gens
mAb
Orig
inRe
activ
ityTargetprotein
Epito
peClon
ingstr
ategy
Form
atisotype
Neutralizingactiv
ityRe
ference
48A
Hum
anDEN
V1ndash4
EDII
EBVtransfo
rmation
IgG
IC50
(120583gmL)
DEN
V115
DEN
V2gt40
DEN
V324
DEN
V4gt40
[37]
D11C
IC50
(120583gmL)
DEN
V115
DEN
V210
DEN
V3102
DEN
V416
16D
IC50
(120583gmL)
DEN
V115
DEN
V202
DEN
V305
DEN
V427
2H12
Murine
DEN
V1ndash4
EDIII(ABloop
aa
314ndash
317)
Hybrid
omafrom
BALB
cmou
seim
mun
ized
with
DEN
V2EDIII
IgG2b
IC50
nM
DEN
V1056ndash54
DEN
V329
DEN
V4145
[38]
C9Murinechim
eric
DEN
V2
EDIII
Phaged
isplayof
achim
ericmurine
hybridom
alibrary
VH1V1205811
PRNT5
0850120583
gmL
[39]
4E11
Murine
DEN
V1ndash4
EDIII(str
andA308
312andstrand
G387
389391)
Hybrid
oma
IgG2a120581
IC50
(120583gmL)
DEN
V1016
DEN
V2013
DEN
V-38
DEN
V-415
[40]
4G2
Murine
DEN
V1ndash4
EDII(fu
sionloop
)Hybrid
omafrom
DEN
V2Eim
mun
ized
mice
IgG
PRNT5
0(120583gmL)
DEN
V215
[39]
9F12
Murine
DEN
V1ndash4WNV
EDIII(aa
305307310
330Astr
andandBC
loop
)
Hybrid
omafrom
BALB
cmou
seim
mun
ized
with
DEN
V-2EDIII
IgG1k
DEN
V1ndash4P
RNT5
02sdot10minus8ndash2sdot10minus7M
[41]
2A10G6
Murine
DEN
V1ndash4YF
VWNV
JEV
TBEV
EDII(Fusionloop
aa
98ndash101)
Hybrid
omafrom
BALB
cmou
seim
mun
ized
with
inactiv
ated
DEN
V2
IgG1
PRNT5
0(120583gmL)
DEN
V12
DEN
V215
DEN
V321
DEN
V418
YNF36
WNV
46
[36]
mAb
11Hum
anDEN
V1ndash4WNV
SLEV
YFV
JEV
MVEV
EDII(fu
sionloop
)Ph
aged
isplayof
human
scFv
Fusio
nprotein
scFv-Fc
PRNT8
0(120583gmL)
WNV
125
DEN
V2625
[4243]
BioMed Research International 7Ta
ble1Con
tinued
mAb
Orig
inRe
activ
ityTargetprotein
Epito
peClon
ingstr
ategy
Form
atisotype
Neutralizingactiv
ityRe
ference
E16
(MGAW
N1)
Murine
WNV
EDIII(LR
aa3
02ndash309)
Hybrid
omafrom
immun
ized
micew
ithWNVE
IgG2bhum
anized
PRNT5
04ndash
18ng
PRNT9
053ndash297
ng[4445]
1A1D
-2Murine
DEN
V1ndash3
EDIII(Astr
andaa
307
310and312)
Hybrid
omafrom
immun
ized
micew
ithdifferent
pH-tr
eated
virus
IgG2a
DEN
V2PR
NT5
021n
M[4647]
1F4
Hum
an
DEN
V1
E(virion
)
DI-DII
Electro
fusio
nof
infected
mem
oryBcells
from
DEN
V-im
mun
esub
jects
with
EBV
IgG
DEN
V1IC5
0011120583gmL
[48]
2D22
DEN
V2
DIII
DEN
V2IC50
008120583gmL
5J7
DEN
V3
DI-DII
DEN
V3IC50
010120583gmL
E105
Murine
DEN
V1
E
DIII(BC
loop
G328
T329
andD330DE
loop
K361E
and
E362K
FGloop
K3
85)
Hybrid
omas
deriv
edfro
mC5
7BL6
IFN-120572120573Rminus
minusmice
infected
with
DEN
V1
IgG
PRNT5
0DEN
V-1
05ndash592ng
mL
[32]
E106
DIII(BC
loop
G328
T329
andD330DE
loop
K361E
and
E362K
FGloop
K3
85A
strand
S305
K307E
309K3
10and
E311)
PRNT5
0DEN
V-1
06ndash
592ng
mL
E111
DIII(CC1015840loop
)PR
NT5
038ndash25120583gmL
[49]
CR4374
Hum
anWNV
EDIII
Phaged
isplayof
scFv
IgGlib
rary
VH2-05V
L1e
PRNT5
0018120583gmL
PRNT9
0095120583gmL
[50]
CR4353
VH3-30V
k3-A27
PRNT5
00026120583
gmL
PRNT9
0364120583gmL
1A5
Chim
panzee
DEN
V1ndash4WNV
JEV
LGTV
EDII(aaG
106)
Phaged
isplayof
Fab
library
from
DEN
V1ndash4
infected
chim
panzees
Hum
anized
IgG1
PRNT5
0(120583gmL)
DEN
V1048
DEN
V2095
DEN
V332
DEN
V443
WNVD
ENV438
JEV
21LG
TV28
[51]
8 BioMed Research International
Table1Con
tinued
mAb
Orig
inRe
activ
ityTargetprotein
Epito
peClon
ingstr
ategy
Form
atisotype
Neutralizingactiv
ityRe
ference
mAb
11Hum
anDEN
V1ndash4WNV
EDII(fu
sionloop
W101G104G106)
scFv
library
scFv-Fc
PRNT8
0(120583gmL)
DEN
V2625
WNV
125
[4243]
3B4C
-4
Murine
SLEV
E
1a
Hybrid
oma
IgG
SLEV
PRNTlt17
[5253]
1B2C
-5SLEV
1bSLEV
PRNTlt17
6B5A
-2SLEV
1cSLEV
PRNT
48
4A4C
-4SLEV
1dSLEV
PRNT
29
1B5D
-1SLEV
JEV
2SLEV
PRNTlt17
2B5B
-3SL
EVJEV
MVEV
WNV
YFV
3SLEV
PRNT
23
2B6B
-2AllFlaviviru
s4a
SLEV
PRNTlt17
6B6C
-1AllFlaviviru
s4b
(inDII)
SLEV
PRNT
23
A3
Chim
panzee
JEV
E2
DIa
aK1
79Ph
aged
isplay
Hum
anized
PRNT5
0004ndash0
2nM
[54]
B2DIIaaI126
Phaged
isplay
PRNT5
0002ndash2
nM
E3DIIIa
aG132
Phaged
isplay
PRNT5
0014ndash0
93n
M
FabT
JE12B0
2Hum
anJEV
ENA
Phaged
isplay
Fab
FRNT5
0502120583gmL
[55]
5F10
Hum
anCH
IKV
E2Dom
ainB
EBVtransfo
rmation
IgG11205822
IC50lt100n
gmL
[56]
8B10
Hum
anCH
IKV
E1-E2
E2Dom
ainA
EBVtransfo
rmation
IgG1k
IC50lt100n
gmL
CHK-
152
Murine
CHIK
VE2
aa59
Hybrid
oma
IgG2c
IC50
1ndash3n
gmL
[57]
11E7
Murine
CCHFV
Gn
C-ter
Hybrid
oma
IgG
PRNT8
0diluted
12560
[58]
4-39-C
CMurine
RVFV
G2
Dom
ainIV
Hybrid
oma
IgG
PRNT8
0diluted
1204
80ndash181920
[59]
2C9
Murine
YFV
EHybrid
oma
IgG2a
PRNT9
011
0417D
1105
2Asib
i[60]
5A 7A R3(27)
Hum
anYF
VE
DI-DII
Phaged
isplay
ScFv
PRNT9
001ndash03120583
gmL
[61]
13D6
Murine
TBEV
EDIII
Hybrid
oma
IgGchimeric
PRNT5
019120583gmL
FRNT5
045120583
gmL
IC50
19ndash167120583
gmL
[62]
3B4C
-4(H
y4-26C
)Murine
VEE
VE2
aa182ndash209
Hybrid
oma
IgGhum
anized
PRNT7
0(120583gmL)
394ndash125
[63]
BioMed Research International 9
The structure of the 1A1D-2 bound to EDIII indicates that themAb binds to sites that are transiently exposed during viralldquobreathingrdquo at 37∘C and block infection during attachmentof the virion to the cell In particular this mAb neutralizesDENV1 2 and 3 serotypes and residues K307 and K310 arethemost critical residues for binding of 1A1D-2mAb [46 75]
Finally Shrestha et al immunized mice with a genotype2 strain of DENV1 virus and generated 79 new mAbs 16of which strongly inhibited infection by the homologousvirus and localized to DIII of E protein Surprisingly onlytwo mAbs E105 and E106 retained strong binding andneutralizing activity against all five DENV1 genotypes as wellas being protective in immunocompromised infected miceMoreover E105 and E106 exhibited therapeutic activity evenwhen administered as a single dose four days after inoculationwith a heterologous genotype 4 strain of DENV1 in the samemouse model [32]
243 Other Non-HumanmAbs Other non-human and non-murine as well as humanized mAbs directed against the Eglycoprotein have been described In this regard Goncalvezet al isolated a large panel of anti-E Fab fragments from chim-panzees infected with all four DENV serotypes Howeveronly a limited number of them displayed a cross-neutralizingactivity against DENV1 and 2 and to a lesser extent againstDENV3 and 4 In particular the authors calculated thatamong them the 1A5 mAb that was further humanizedneutralized DENV1ndash4 at a PRNT50 titer of 048 095 32and 43 120583gmL respectively Interestingly the humanized 1A5was also tested for binding and neutralization against theWNVDENV4 chimera JEV strain SA14-14-2 and Langatvirus (LGTV) strain TP 21 giving a PRNT50 titer of 38 21and 28120583gmL respectively Moreover the authors calculatedthat when administered in a dose of 2mg per kg of bodyweight 1A5 would give a serum titer of approximately 40and 20 50 reduction in plaque reduction neutralization test(PRNT50) against DENV1 and DENV2 respectively [76]Furthermore epitope mapping of 1A5 mAb localized to G106within the Flavivirus-conserved fusion loop inDII of DENV2E protein [51]
244 Anti-DENV Human mAbs As anticipated attemptsto isolate neutralizing human mAbs have been morechallenging due to the restricted elicited Ab-repertoire whichrecognizes DIII of E glycoprotein in naıve as well as ininfected patients [77 78] However several groups reportedthe isolation of neutralizing mAbs from infected subjects Inthis regard Setthapramote et al isolated a total of 136 humanhybridoma clones producing specific mAbs against DENVobtained using PBMCs from nine blood samples from fouracute-phase patients secondarily infected with DENV2 andfive convalescent-phase patients Interestingly the authorsfound that most of the acute-phase mAb clones were cross-reactive with all four DENV serotypes with most of themrecognizing the E protein and endowing of neutralizingactivity against all DENV serotypes compared to thosederived from convalescent-phase patients In particularfrom the acute-phase PBMCs 818 were anti-E 66 wereanti-prM and 33 were anti-NS1 while 133 anti-E 133
anti-prM and 533 anti-NS1 mAbs clones were obtainedfrom convalescent-phase PBMCs [79] Previous studiesconfirmed the data obtained by Setthapramote et al onconvalescent-phase patients but this group firstly reportedthe efficient preparation of human mAbs with strongneutralizing activity titers against all four DENV serotypesusing PBMCs from acute-phase patients secondarily infectedwith DENV [80 81]
The group of de Alwis through immunoglobulin deple-tion studies reported that a substantial fraction of DENV-reactive Abs in human immune sera including type-specificneutralizing Abs bound to the intact virion but not torecombinant E proteinThe authors confirmed these observa-tions also isolating human neutralizing mAbs and proposedthat humans produce Abs that neutralize DENV infectionby binding a complex quaternary structure epitope that isexpressed only when E proteins are assembled on a virusparticle [48] Similar findings have been described for severalother viruses including WNV and HIV [82 83] Mappingstudies indicate that this epitope has a footprint that spansadjacent E protein dimers and includes residues at the hingebetween domains I and II of E protein [48]
Recently Costin et al isolated three broadly neutralizinganti-DENVhumanmAbs named 48AD11C and 16DThesemAbswere isolated from three different convalescent patientswith distinct histories of DENV infection All three mAbsrecognized the E glycoprotein with high affinity neutralizedall four serotypes of DENV but mediated ADE of infection inFc receptor-bearing cells at subneutralizing concentrationsMapping studies revealed that all three mAbs bind discontin-uous epitopes within the highly conserved fusion-loop regionof DII (contacting residues W101 L107 andor G109) [37]
25 Yellow Fever Virus (YFV) TheWHO estimates that thereare 200000 cases of yellow fever virus (YFV) infections and30000 related deaths every year especially in Sub-SaharanAfrica In fact different to themajority of human arbovirusesand similarly to DENV and CHIKV YFV has expanded itshost range to include humans as an amplifying host [1]Symptoms occurring from 3 to 6 days after infection withYFV consist mostly in fever chills anorexia lumbosacralpain nausea and vomiting This syndrome lasts about 3days in most cases and is sometimes followed by a one-daylasting period of remission Fever will reoccur together withdark hematemesismelena petechiae and other hemorrhagicsymptoms Convalescence is characterized by deep asthenialasting up to two weeks Rarely prognosis can be fatal Aneffective live-attenuated vaccine (17D) derived from the Asibistrain by serial passages in chicken embryos is available [84]
251 mAbs against YFV In 1983 Schlesinger et al produceda battery of mAbs after immunization of BALBc mice byinjection of 17D YFV vaccine strain [84] Among them thespecificity towards the E and NS1 proteins was describedeven though anti-NS1 Abs were not able to show neutralizingactivity The 13 IgG and 1 IgM anti-E protein mAbs wereclassified in five groups according to their specificity GroupA consisting of the only IgM produced in this experiment
10 BioMed Research International
could neutralize 17D YFV virus only group B 17D and Asibistrains only group C Asibi strain only group D could onlyneutralize Asibi but not 17D strain showing also reactivityagainst DENV2 Zika or Banzi viruses All of the four mAbsbelonging to this group were able to cross-react with Zikaandor Banzi viruses and two (4E11 and 5H3) neutralizedDENV2 virus with 4E11 neutralizing also Banzi virus The3E9 mAb the only component of the group E could notneutralize 17D nor Asibi strains
All of the IgG mAbs resulted able to protect bothprophylactically and therapeutically BALBc or CD-1 micefrom lethal intracerebral challenge with 17D-204 strain [85]The chimeric form of the group B mAb 2C9 IgG (2C9-cIgG) was able to provide AG129 mice a 72 survival whenadministered 24 hours before infection with 17D-204 strainIts murine form could provide the survival of the 95 ofthe mice Appreciable results were also obtained when bothmurine 2C9 and 2C9-cIgG were administered 24 hours post-infection with the survival of 70 and 20 of mice respec-tively Viral presence was not detected in survivingmice [60]
In 2005 Daffis and colleagues constructed two Ab-phage libraries by cloning the repertoire of YFV-infectedpatients Panning was then performed with YFV-17D virionsThe scFv-5A 7A and R3(27) showed a neutralizing activityspanning from50 to 100 inPRNTassays against bothYFV17D-204-WHO and Asibi strains In further tests reactivitywas observed against wild-type YFV strains of West Africagenotype I and II (Nigeria 1987 and Asibi strains resp)and EastCentral Africa (CAR 1986 Ethiopia 1961 strains) Aconcentration from 01 to 03120583gmL could yield a 90 plaquereduction Reactivity was also observed against the strainSenegal 1990 in a lesser extent Production of escape variantscould demonstrate that scFv-5A 7A and R3(27) epitopes arebuilt up by residues extensively separated in themonomers ofE glycoprotein that however result in closed proximity whenthe homodimeric form of E is constituted [61]
26 West Nile Virus (WNV) The West Nile virus (WNV) isan epidemic neurotropic virus estimated to be responsibleof about 36000 cases and 1500 deaths registered in theUnited States between 1992 and 2012 [86] WNV antigenicityallows its classification into the Japanese encephalitis virus(JEV) serocomplex Genomic analysis has revealed two maingenetic lineages of WNV lineage I viruses circulating in theUSA Europe the Middle East Africa India and Australiaand lineage II viruses isolated from Sub-Saharan Africa andMadagascar [1]
WNV was firstly identified in 1937 and is endemicin many countries of Africa Middle East and West AsiaHowever after 1990 frequent outbreaks of WNV infectionswere reported in Romania Israel and later in North Americaand across the USA in 1999 and recently in 2012 [87]
Most of WNV infected individuals do not develop symp-toms while about 20 develop a self-limiting illness calledWest Nile fever Acute symptoms include fever tirednessand swollen lymph glands but in a minority of cases alsoencephalitis with long-term deficits in cognitive functionand motor skills have been reported However in less than
1 of infected patients WNV having crossed the blood-brain barrier is responsible of neuroinvasive and potentiallylethal form of the disease In these cases degeneration andapoptosis upon infection of neurons and the consequentinflammatory response can occur Related symptoms includehigh fever coma muscle weakness and paralysis Immun-odepression and advanced age have been correlated with anaugment risk to develop a severe disease [87] There are nospecific treatment options or licensed vaccines for humans[88]
WNV is transmitted by infected mosquitos and initialreplication is thought to occur primarily in dendritic cells inthe skin which migrate to secondary lymphoid tissues wherethe replicating virus enters the circulation To date onlyefficacious WNV veterinarian vaccines have been licensedwhile there are no licensed vaccines for protection againstWNV in humans [88]
261 mAbs against WNV Broad-spectrum antivirals suchas type I IFN-120572 ribavirin mycophenolic acid in WNVinfection showed ineffective results in vivo despite in vitrothey showed some activity [89]
However experiments in murine models extrapolationof clinical data as well as passive administration of pooledimmune-120574-globulins (OmriGam) containing a significanttiter of neutralizing Abs before and after infection showedan important role of the humoral immune response incontrolling viremia and prevent viral dissemination Indeedthe development of a neutralizing mAb-based therapy seemsto be encouraging for a possible treatment of infected patients[90ndash92]
Similarly to what has been observed for DENV only anti-E Abs have been identified as neutralizing and protectiveAlso anti-NS1 Abs have shown a protective role howevertheir mechanism of protection has not yet been elucidated asthe NS1 protein is secreted from infected cells and not presenton the virions [93ndash95]
In this regard similarly to what has been describedfor DENV studies performed using a naıve human scFvlibrary for panning with purified WNV E protein supportthe hypothesis that no Abs against the neutralizing DIII canbe isolated However DIII specific Abs were isolated in asubsequent study using immunoglobulin libraries obtainedfrom threeWNV infected patients for biopanning on purifiedinactivated virus virus-like particles consisting of prM andE proteins or recombinant E glycoprotein Although theproportion of DIII-specific mAbs was low (8) comparedwith anti-DII mAbs (47) Two out of the four anti-DIIImAbs were potently neutralizing and protective in vivowhereas only three out of the 24 anti-DII were weaklyneutralizing in vitro and non-protective in vivo In particularresidues that are critical for neutralization lies on the regionsspanning amino acids 305ndash312 330ndash333 and 365 that arelocated on adjacent exposed loops ofDIIIHowever sequencealignment of E protein of different flaviviruses such as DENVand JEV revealed a considerable variation compared tothe whole E protein This observation further suggests thatdifferently from broadly cross-reactive anti-DII Abs anti-DIII neutralizing Abs are virus-type specific However of
BioMed Research International 11
these mAbs two (CR4374 and CR4353) protected mice fromlethal WNV challenge at 50 protective doses of 129 and357 120583gkg of body weight respectively [50]
Sanchez et al using three different immunization strate-gies (ie inactivated virus naked DNA and recombinantprotein) isolated ninemurinemAbsmost of which bound toconformation-dependent epitopes in DIII of the E protein Inparticular neutralizing mAbs named 8B10 11C2 10C5 and17C8 were obtained from mice immunized with inactivatedvirus alone or in combination with a DNA plasmid andbound to the same region of DIII with high affinity Incontrast mAbs obtained by immunization with a solubleversion of the E glycoprotein did not exhibit neutralizingactivity These non-neutralizing mAbs were cross-reactivewith several other flaviviruses including SLEV JEV YFVand Powassan virus confirming the conserved nature ofFlavivirus non-neutralizing epitopes [96]
Gould et al isolated 11 unique human single-chain vari-able region Ab fragments (scFvs) that bind the E protein ofWNVAmong them a humanmAb namedmAb11 expressedas a scFv-Fc fusion protein was further characterized Itrecognizes the fusion loop at the distal end of DII ofthe WNV E protein and cross-reacts with all four DENVserotypes and provides protection against DENV2 and 4as well as WNV [42] Moreover therapeutic studies of thismAb in WNV-infection model mice provided substantialprotection when administered after 5 days post-infectionInterestingly a neutralization escape variant of this mAbfailed to cause lethal encephalitis (at higher infectious doses)or induce the inflammatory responses associated with blood-brain barrier permeability in mice compared to the parentalWNV suggesting an important role for the fusion loop inviral pathogenesis [43]
Oliphant et al isolated an anti-DIII E protein mAbnamed E16 fromhybridomas obtained after immunization ofmice with recombinant WNV E protein which neutralizedall WNV strains with PRNT50 values of 4 to 18 ng andPRNT90 values of 53 to 297 ng One hundred microgramsof mAb protected greater than 90 of mice from lethalinfection and even a single 4120583g treatment of E16 on day 2 afterinfection prevented mortality Moreover humanization ofthis mAb confirmed as therapeutically effective in mice [44]Subsequent studies revealed that mAb E16 neutralization ismediated by engagement of four discontinuous segmentsof DIII including the amino-terminal region (amino acidresidues 302ndash309) and the three connecting loops BC (aminoacid residues 330ndash333) DE (amino acid residues 365ndash368)and FG (amino acid residues 389ndash391) Moreover no ADE ofinfection was detected when E16 mAb was used at saturatingconcentrations [45 47]
Furthermore results of a Phase I safety study of thehumanized E16 mAb (designated MGAWN1) have beenreported and suggested that doses of up to 30mgkgwerewelltolerated with few mild adverse events and would providean excess of virus neutralizing activity for 3-4 weeks aftertreatment However a case of anti-MGAWN1 Ab elicitationoccurred with the consequent increased rate of clearance andindeed impacting efficacy A Phase II safety and efficacy studyof MGAWN1 is ongoing [97] Furthermore the possibility of
preventing or treating WNV-induced memory deficits wasrecently investigated In this study hamsters were treatedintraperitoneally with 32mgkg ofMGAWN1mAb at 45 daysafter subcutaneously challenging with WNV InterestinglyMGAWN1 prevented mortality weight loss and improvedfood consumption of WNV-infected hamsters compared tocontrols [98]
Recently Lelli et al isolated six anti-E mAbs frominactivated-WNV immunized mice In particular three ofthem (3B2 3D6 and 4D3) neutralized lineage I and II WNVwith the first two recognizing the same epitopes located onthe distal lateral surface of DIII (critical amino acid residueK307) Conversely 4D3mAb recognized a novel neutralizingepitope on DII (critical residues S276 and T278) Indeedfurther protective and therapeutic studies are needed toascertain their neutralizing activity in vivo [99]
Finally to conclude like DENV infection Abs directedagainst the M protein are not protective and neutralizingMoreover in humans a skewed humoral immune responseagainst DII has been frequently observed and confirmedalso with the hybridoma technology [50] The isolation andelicitation of neutralizingAbs directed against the fusion loopand DIII of the E protein represent thus the most challengingand promising goal for the development of new effectivetherapeutic inhibitors and immunogens respectively
27 Tick-Borne Encephalitis Virus (TBEV) Tick-borne ence-phalitis virus (TBEV) is one of the most dangerous agentscausing human neuroinfections occurring mostly in Europeand Asia and with a potential fatal prognosis [100] TBEV isbelieved to cause 3500ndash10000 human cases of encephalitisin Europe per year with a high morbidity in Russia CzechRepublic Austria and Germany In particular between 1990and 2007 an average of 8755 cases of TBE was reported peryear in Europe and Russia Despite the fact that Russia isthe country with most infections registered annually CzechRepublic incidence is among the highest in Europe with 400ndash1000 clinical cases reported every year [101]
Three subtypes of TBEV are classified namely EuropeanSiberian and Far-Eastern sharing most of the genetic andantigenic features [102] In fact a high degree of antigenichomogeneity between different strains of TBEV has beendescribed [103]
Clinical manifestations vary among the subtypes butusually they start with a short febrile period of 7ndash14 daysafter the tick bite Fatigue headache and pain in the neckshoulders and lower back together with high fever andvomitingmay be present [100]Thesemanifestations are oftenfollowed by an asymptomatic phase lasting from 2 to 10days after remission from the fever with possible progres-sion to neurological disease Neurological symptoms includemeningitis encephalitis myelitis and radiculitis Mortalityoccurs in 1-2 of the European subtype-infected patientsbut fatal prognosis can occur in up to 20ndash40 of the Far-Eastern subtype-infected patients [104] Mortality rates ofthe Siberian subtype are similar to those observed for theEuropean [105]
Effective and safe vaccines against TBEV produced frominactivated virus have been developed and licensed for their
12 BioMed Research International
use in humans However an emergency therapy in theabsence of a mass immunization is needed as no effectivetreatments are yet available [106]
271 mAbs against TBEV Levanov et al described the chim-erization of two murine mAbs (13D6 and 10C2) directedagainst the DIII and DII respectively of the TBEV E glyco-protein The chimeric mAbs present binding characteristicssimilar to the parentalmAbsMoreover as the parentalmAbsonly the chimeric mAb 13D6 was able to neutralize TBEVinfectivity in vitro In particular neutralization studies withthe murine chimeric and scFv forms of mAb 13D6 were per-formed In particular murine 13D6 showed an IC50 titer of115 120583gmL in Focus Reduction Neutralization Tests (FRNT)against TBEV strain 205 and of 29 120583gmL in PRNT againstTBEV strain Softjin Chimeric 13D6 showed an IC50 titer of45 120583gmL in FRNT against TBEV strain 205 and of 19 120583gmLin PRNT against TBEV strain Softjin ScFv 13D6 showed anIC50 titer of 167 120583gmL in FRNT against TBEV strain 205and of 112 120583gmL in PRNT against TBEV strain Softjin [62]
28 Japanese Encephalitis Virus (JEV) Japanese encephalitisvirus (JEV) is a mosquito-borne flavivirus that over thepast few decades has caused several outbreaks throughoutChina Southeast Asia Australia and Papua New Guineawith a prevalence recently estimated to be of about 70000casesyear [107ndash109] A 40 mortality was recorded in someof the JEV-affected areas Moreover many survivors facesome neurological problems and complications [108] Since1995 the disease has also emerged in Non-Asian regions suchas Northern Australia [110 111] The situation in SoutheastAsia however is further complicated by the overlappingepidemics of JEV and DENV as well as sporadic casesof WNV infections detected in some of the affected areasparticularly in India [112]
Effective vaccines both live-attenuated and inactivatedstrains of JEV have been developed and licensed in majorcountries However as for the other arthropod-borne Fla-vivirus members no specific antiviral drugs are currentlyavailable [4]
281mAbs against JEV Aspreviously described for the otherFlavivirusmembers many groups isolated and characterizedanti-JEVmAbs showing different neutralizing and protectiveproperties In this regard Gupta et al used combinations ofanti-E JEV mAbs (Hs1-4) in mice protection experimentsIn particular they found that the singularly mAb protectionranged from 45 to 65 when 100120583g of mAb were adminis-tered while equimolar combinations of two or three mAbsgave 85ndash90 or 100 protection respectively [113] In similarexperiments Lee et al demonstrated that a Flavivirus anti-NS1 mAb named 16NS1 cross-reacted with JEV as well asWNV and exhibited protective activity against WNV as wellas a lethal JEV infection However no neutralizing activitywas observed using this mAb against both WNV and JEV inin vitro experiments suggesting the participation of otherAb-mediatedmechanisms in vivo In particular 95ofmicewere
protected when 500120583g of mAb were administered intraperi-toneally and concomitantly to intramuscular injection of JEV
Overlapping peptide mapping analysis combined withsite-specific mutations identified the 116KAWGKSILFA125region and critical amino acid residues W118 and I122 as16NS1 mAb epitope highly conserved in WNV and JEVstrains [114]
Arakawa et al isolated from a combinatorial human Fablibrary constructed from peripheral blood lymphocytesobtained from JEV hyperimmune volunteers Among 188randomly selected clones FabTJE12B02 showed the best 50focus reduction endpoint at the concentration of 502 120583gmLagainst the JEV strain Nakayama [55]
Goncalvez et al isolated three mAbs named Fabs A3B2 and E3 by repertoire cloning from chimpanzees initiallyimmunized with inactivated JE-VAX and then boosted withattenuated JEV SA14-14-2 In particular these mAbs reactedwith epitopes in three different E domains in DI (amino acidresidue K179) in DII (I126) and in DIII (G132) for Fabs A3B2 and E3 respectively Moreover these Fabs as well as thederived humanized counterpart mAbs exhibited high neu-tralizing activities against a broad spectrum of JEV genotypestrains Moreover these mAbs exhibited a 50 protectivedose of 084 120583g (B2) 58120583g (A3) and 247120583g (E3) in mousemodels Finally administration of 200 120583gmouse of mAb B2one day after otherwise lethal JEV infection protected 50of mice and significantly prolonged the average survival timecompared to that of mice in the unprotected group [54]
29 St Louis Encephalitis Virus (SLEV) St Louis encephalitisvirus (SLEV) was first discovered as the mosquito-borneagent responsible for over 1000 cases of encephalitis duringa 1933 summer outbreak in St Louis (Missouri) and nowis a reemerging human pathogen widely distributed in theAmerican continent causing several human encephalitisoutbreaks over the last 80 years Additional epidemics haveindeed occurred from 1964 to 2006 in the Americas rangingfrom the US to Argentina and Brazil As a member of theFlavivirus genus the SLEVE glycoprotein ectodomain is 68identical to serocomplex-related JEVEbut only 46 and 40identical to those ofDENV2 andTBEV viruses fromdifferentserocomplexes [115]
291 mAbs against SLEV In 1983 Roehrig et al isolatedtwenty-one hybridomas producing murine mAbs specificfor the E glycoprotein of SLEV strain MSI-7 Serologicreactivities were initially determined by cross-reactivity indi-rect immunofluorescence assays using 22 strains of SLEVand 8 other related flaviviruses Four groups demonstratingtype- subcomplex- supercomplex- and group-specific reac-tivity patterns were identified Analysis of hemagglutination-inhibition and virus neutralization subdivided the cross-reactivity groups into eight epitopes (E-1a b c d E-2 E-3 and E-4a b) one of them following localized in DII (4b)[52 53]
Moreover among the previously described isolated anti-WNV mAbs by Gould et al the chimeric scFv-Fc 79 effec-tively neutralized also SLEV resulting in gt80 of PRNT80when used at 5 120583gmL [42]
BioMed Research International 13
3 Alphaviruses
Major human arboviruses in the Togaviridae family Chikun-gunya (CHIKV) and Venezuelan equine encephalomyelitis(VEEV) viruses belong to the Alphavirus genus which iscomposed of viruses with an icosahedral nucleocapsid sur-rounded by a lipid envelope and glycoprotein spikes Thestructural proteins of alphaviruses arise through co- andpost-translational processing of a polyprotein encoded by asingle positive-stranded RNAproducing the capsid (C) PE26 K and E1 The PE2 glycoprotein is a precursor containingthe E3 glycoprotein fused to the amino terminus of the E2envelope glycoprotein The PE2 glycoprotein is followed by6K a small membrane-associated protein and E1 the secondpolypeptide component of glycoprotein spikes
Trimerized heterodimers of the E1 and E2 viral glyco-proteins form the surface spikes and contain determinantsof viral tropism and virulence The E3 glycoprotein acts as asignal for transport of PE2 across themembranes of the roughendoplasmic reticulum and may promote the formation andintracellular transport of E1-PE2 heterodimers to the cellsurface During transport to the cell surface PE2 undergoesa maturational cleavage event by a furin-like protease toproduce E2 and E3 [116]
The E2 glycoprotein promotes specificity of virus bindingto the host cell surface and is a target of the humoral immuneresponse The E1 glycoprotein mediates fusion of the virionenvelope with the membranes of acidified endosomes allow-ing release of the nucleocapsid into the cytoplasm and theonset of viral replicationAlphavirus E1 shares no appreciablesequence identity to Flavivirus E protein Despite differencesin their amino acid sequences and arrangements on the viralparticle the structures of E and E1 are remarkably similarIndeed the conservation of three domains has been welldocumented in crystal structures of flavivirus E ectodomainsfrom TBEV DENV JEV and WNV as well as Alphavirus E1from Semliki Forest virus (SFV) and CHIKV Abs to the E1glycoprotein do not typically neutralize virus infectivity invitro but can protect against lethal challenge in animals
By analogy with the prM protein of dengue Flavivirusfurin cleavage of PE2 may begin after transit of an acidiclate component of the Golgi body where E3 is thought tosuppress the acid pH-triggered activation of glycoprotein E1fusion capability Protective anti-E3 mouse mAbs have alsobeen described [117]
31 Chikungunya Virus (CHIKV) The chikungunya virus(CHIKV) belongs to the Semliki Forest clade and was firstlyisolated in 1953 in Tanzania during an epidemic outbreakfollowing occurred also in Asia and Africa In the last fivedecades several CHIKV outbreaks have been described bothin Africa and Asia separated by gaps lasting from two totwenty years In 2005-2006 about 300000 cases out of785000 inhabitants were reported in La Reunion island witha fatal prognosis for 237 of the patients [117] Neither Europenor the Americas have had outbreaks of CHIKV so far exceptfor imported isolated cases Three genotypes of CHIKV aredescribed Asian EastCentralSouth African (ECSA) andWest African with an amino acid identity spanning from
952 to 998 Recent epidemics in Africa and Indiansubcontinent were caused by strains belonging to the ECSAgenotype Transmitted by Aedes mosquitoes CHIKV ismaintained in the human population by human-mosquito-human transmission [1] The disease is characterized bydengue-like symptoms such as chills high fever headacheand persistent myalgia and a further incapacitating arthralgia(from which the name chikungunya) which affects 40 ofinfected subjects However prognosis is rarely fatal [116]
There is currently no commercial vaccine and antiviraltreatment forCHIKV although some candidate vaccines havebeen tested in humans In this regard in 2000 US Armyperformed a Phase II clinical trial testing a live-attenuatedCHIKV vaccine (TSI-GSD-218) derived from a 1962 strain(15561) of an outbreak in Thailand Out of 58 every patientdeveloped neutralizingAbs and 5 lamentedmild tomoderatejoint pain [117] A phase III trial of this candidate vaccineis ongoing Furthermore a new formulation using virus-likeparticles was able to induce neutralizing Abs in macaquesagainst different CHIKV strains [117 118] Indeed it has beendescribed that infection seems to elicit long-lasting protectiveimmunity and cross-protection among CHIKV and otheralphaviruses
CHIKV entry into host cell is demonstrated to be medi-ated by envelope glycoproteins E1 and E2 which allowvirus fusion to cell membrane in low pH conditions andrecognition of an unknown cellular receptor respectively[116 119ndash121] Even though the genome replication relies onan error-prone RNA-dependent RNA-polymerasis CHIKVstrains S27 and 05115 Reunion showed in E1 and E2 alow amino acid variation of 068 and 33 respectivelyand such a low variability may be needed for an effectivereplication in two phylogenetically distant hosts [116 122]Recently the crystal structure of E1 and E2 heterodimerhas been resolved both at low pH and natural conditionsshedding light on E1 and E2 ectodomainsrsquo structure Inparticular similarly to flaviviruses E protein E1 ectodomainis made of the N-terminal DI the DII containing the fusionloop and the DIII at the C-terminal On the other hand E2ectodomain comprises theN-terminal domainA the domainB supposed to interact with the host cellrsquos unknown receptorand domain C at the C-terminal E1 DIII and E2 domain Care located close to the viral membrane [123 124]
311 mAbs against CHIKV Warter et al described 5F10 and8B10 two human mAbs which strongly neutralized severalCHIKV isolates in vitro without cross-reactivity againstother alphaviruses but to Onyong-nyong virus [56] Mixedpreparation of 5F10 and 8F10 did not show neither synergisticnor addictive effect in vitro and studies upon escape mutantsdemonstrated that 5F10 mAb binds at the tip of the E2domain B while 8B10 recognizes residues close to E1 fusionloop and amino acids within E2 domain A which form atransitional epitope under low pH conditions The authorsspeculated that 5F10 and 8B10 may inhibit CHIKV entry andfusion to the cell membrane respectively It is worth notingthat the previously mentioned escape mutants displayedmutations associated with reduced viral fitness in vitro evenafter 13 neutralizationamplification rounds In the same
14 BioMed Research International
study cell-to-cell transmission was firstly demonstrated as anescapemechanism enhanced by the E2mutation R82G [122]
In further in vivo studies 5F10 and 8B10 significantlydelayed CHIKV-caused death of AGR129 mice both in pro-phylactic and therapeutic tests Interestingly in therapeutictreatment these mAbs showed a synergistic effect whenadministered in combination with the total amount of mAbsinjected being the same of the single-mAbs Possibly thesetreatment did not allow themice to survive CHIKV challengedue to Ig clearing and 6ndash10 days half-life usually described forhuman mAbs [125]
Recently Pal et al cloned thirty-six murine mAbs ableto neutralize the ECSA La Reunion 2006 OPY-1 strain ofCHIKV (CHIKV-LR) the majority of which also neutralizeinfection of other strains corresponding to the Asian andWest African genotypes In particularmAbCHK-152 showedthe highest and broadest neutralizing activity with FRNTvalues indicating an IC50 value of 1 to 3 ngmL dependingon the viral strain used Among the thirty-six mAbs four(CHK-102 CHK-152 and CHK-166 CHK-263) could pre-vent lethality of immunodeficient Ifnarminusminus C57BL6 micewhen administered one day before exposition with CHK-152 and CHK-263 showing the ability to protect mice at thelowest dose (10 120583g) Therapeutic studies were performed onthese mAbs administering a single dose of 100 120583g 24 hoursafter infection The highest activity was described for CHK-166 able to protect 63 of the mice Testing the combinedactivity of the mAbs in therapeutic studies administration ofCHK-166 plus CHK-152 in a dose of 250120583g each turned out tobe themost effective protecting 71 of the IfnarminusminusC57BL6mice 60 hours after infection [57]
32 Venezuelan Equine Encephalomyelitis Virus (VEEV)Venezuelan equine encephalomyelitis virus (VEEV) is main-tained in a natural transmission cycle between mosquitoesand small rodents The first documented outbreaks occurredin the 1930s and several epidemics have been reported so farin Latin-American countries such as Venezuela ColombiaPeru Ecuador Bolivia Honduras Mexico and Panama withhundreds of thousands of human cases being reported anda case-fatality rate up to 1 during the 1969 Ecuador out-break Clinical manifestations of VEE are indistinguishablefrom Dengue [126] Six serogroups (IndashVI) are currentlyrecognized within the VEEV complex VEEV caused humanand equine outbreaks in the Americas for nearly a centuryEquine epizootics have high mortality (38ndash83) leading toa high viremia followed by a lethal encephalitis and often tohuman epidemics involving thousands of cases and hundredsof deaths Indeed VEEV is infectious for humans by theairborne route and has been responsible for a number oflaboratory infections In humans the disease is usually self-limiting with a febrile illness with 1ndash4 of cases progressingto severe encephalitis It is a hazard to laboratory workershas been developed as a biological weapon and is a potentialbioterrorist agent [127] There are no antiviral drugs andvaccines licensed for the treatment of VEEV infection inhumans Indeed although experimental live-attenuated vac-cines have been developed (eg TC-83) with good levels ofprotection in equine and mice in humans they may fail to
give protection in the majority of cases [128] Consideringthe protective role exerted by humoral immune response inanimal models it could be concluded that treatment withspecific IgG may have a beneficial antiviral effect in humanairborne infections with pathogenic strains of VEEV
321 mAbs against VEEV Several works support that mAbsmay protect against airborne VEEV as well as after airborneexposure toVEEVAb reactivity to both surface glycoproteins(E1 and E2) is associated with neutralization of the virusin vitro and passive protection against virus challenge Aseries of anti-E1 and anti-E2 mouse and humanized mAbshave been described recently Among them Phillpotts et alexamined two anti-E2 murine mAbs named 1A4A-1 and1A3A-9 both having potent protective activity against subcu-taneous VEEV challenge in mice Both mAbs had a similarhalf-life (58 and 100 days) in mouse serum after a singleintraperitoneal dose suggesting that mAbs delivered by thisroute at or around the time of VEEV infection persists athigh levels in the blood as well as secretions in respiratorytransudation throughout the clinical course of the disease Inparticular both mAbs administered 24 h prior to airbornechallenges had a substantial protective effect (90ndash100)Treatment of mice 2 or 24 h after airborne infection with asingle intraperitoneal dose of 100 120583g of 1A3A-9 mAb led toapproximately 50 survival There was no beneficial effectwhen mAb treatment was delayed to 72 h Moreover therewas evidence of synergy in vitro in PRNT between 1A3A-9and 1A4A-1 as has been demonstrated for other virusesand mAb pairs However no synergy was found in mouseprotection when mAbs were delivered intraperitoneally as amixture in equal parts to animals challenged 24 h previouslywith VEEV [129]
In a subsequent study Phillpotts et al reported two otherprotective mAbs The 3B2A-9 mAb protected against allthe serogroup I strains while 1A3B-7 mAb protected wellagainst challenge with all of the viruses tested Both 3B2A-9 and 1A3B-7 protected against airborne exposure to theIAB serogroup virus strainTrinidad donkey (TrD)Howeverthere was no evidence of synergistic protection when thesemAbs were combined in equal proportions An intraperi-toneal dose of 10 120583g was sufficient to protect 50 of themice with either mAb [130] According to other data themechanism of protection did not appear to depend uponneutralization [131] Similar results were obtained by OrsquoBrienet al which described a non-neutralizing mAb (IgG2a)named CUF37-2a able to protect 50 of mice from asubcutaneous VEEV challenge when a dose of 915 120583g of mAbwas administered 24 h prior to challenge [132]
In a following study the 1A3B-7 mAb was humanized(and following reported as Hu1A3B-7) maintaining the samefeatures of its murine counterpart In particular evaluation ofin vitro studies indicated that Hu1A3B-7 retained both broadspecificity and neutralizing activity Furthermore in vivoexperiments showed that Hu1A3B-7 successfully protectedmice against lethal subcutaneous and aerosol challenges withVEEV strain TrD Moreover the effectiveness of the human-ization process was determined by assessing proliferationresponses in human T-cells to peptides derived from the
BioMed Research International 15
murine and humanized versions of the VH and VL domainsThis analysis showed that the number of human T-cellepitopes within the humanized Ab had been substantiallyreduced indicating that Hu1A3B-7 may have low immuno-genicity in vivo [133]
Similarly Hunt et al described a humanizedmurinemAbderived from the 3B4C-4 mAb recognizing the E2c epitope(amino acid residues 182ndash209) and following named Hy4-26C showing neutralizing activity similar to the murinemAb Moreover it was protective (70ndash100 of survival) at100 ng dose in mice animal model intraperitoneal inoculatedand at 500120583g for intranasal challenge (80) Therapeuticstudies in mice revealed that Hy4-26C was able to cure miceup to 24 h following infection at 10 120583g on the contrary themouse mAb when given 1 hour after virus challenge [63]
In another work Hunt et al isolated two neutralizinghumanized Fabs F5 and L1A7 employing a blocking strategy[134 135] The anti-E2 specific F5 IgG had a 70 PRNTendpoint of 10 ngmL equivalent to that described for themost effective neutralizing anti-VEEVE2mAbs In particularthe E1-specific hFab L1A7 had a PRNT endpoint of 3120583gmL300-fold lower than F5
Further studies revealed that F5 epitope is located inthe 115ndash199 amino acid region Moreover F5 IgG had potentability to protect mice from infection by either route whenadministered 24 h before exposure however mice treated24 h and 48 h after aerosol exposure developed central ner-vous system infections but exhibited no clinical signs ofdisease [135]
Hu et al reported that passive immunization with thehumanized chimericmousemAbhu1A4A1IgG1-2A inmice at50 120583g 24 h before or after virulent VEEV challenge providedcomplete protection indicating that hu1A4A1IgG1-2A haspotent prophylactic and therapeutic effects against VEEVinfection [136]
Anti-VEEV mAbs isolated from a non-human primategene library has also been reported In particular thehumanized chimeric scFv-Fc ToR67-3B4 recognized viableas well as formalin and 120573-propiolactone inactivated virusparticles It detected specifically the viral E1 envelope proteinof VEEV but did not react with reduced viral glycopro-tein preparations suggesting that recognition depends uponconformational epitopes The recombinant Ab was able todetect multiple VEEV subtypes and displayed only marginalcross-reactivity to otherAlphavirus species except for Easternequine encephalitis virus (EEEV) In addition the scFv-Fcfusion described here might be of therapeutic use since itsuccessfully inactivated VEEV in a murine disease modelIn particular when the recombinant Ab was administered 6hours after challenge 80 to 100 of mice survived lethalVEEV IAB or IE infection Forty to sixty percent of micesurvived when scFv-Fc ToR67-3B4 was applied 6 hours afterchallenge with VEEV subtypes II and former IIIA [137]
4 Bunyaviridae
TheBunyaviridae family contains human arboviruses belong-ing to the Orthobunyavirus Phlebovirus and Nairovirus
genera Bunyaviridae includes enveloped viruses with a frag-mented single-stranded RNA genome of negative polarityTheir tripartite genome consists of a small (S) a medium(M) and a large (L) fragment and the envelope glycoproteinsGn and Gc that are cleaved out of a polyprotein synthesizedby the M fragment These glycoproteins have been provedto mediate the formation of the virus particle to play a rolein the interaction with cell surface receptors to mediate theentry of the virus into cells and to serve as targets for themajority of neutralizing Abs described so far
Orthobunyavirus transmitted through mosquitoes ormidges vectors are divided in 18 serogroups based on cross-titrations in haemagglutination inhibition assays and neutral-ization assays and correlating with main vector preferencesHowever the most clinically important viruses belong onlyto two serogroups the California encephalitis and the Simbuserogroups
Crimean-Congo hemorrhagic fever virus (CCHFV) isconsidered the only one of clinical relevance within theNairovirus genus which uses tick as main vector and is divid-ed in seven serogroups while Toscana virus and Rift Valleyfever virus (RVFV) which are transmitted by sandflies andmosquitoes respectively belong to the Phlebovirus genus[138]
MAbs against theNairovirusCCHFV and the PhlebovirusRVFV have been described
41 Crimean-Congo Hemorrhagic Fever Virus (CCHFV)Crimean-Congo Hemorrhagic Fever Virus (CCHFV) dis-tribution covers the greatest geographic range of any tick-borne virus known as viral isolation andor disease hasbeen reported from more than 30 countries in SoutheasternEurope Africa Asia and Middle East [139 140] Namelyfrom 1953 to 2010 about 6000 human cases were reportedin Southeastern Europe CCHFV causes sporadic outbreakswith mortality rates ranging from 10 to 80 [141 142] Asignificant variation in the time of incubation has beendescribed [139] Prehemorrhagic symptoms include highfever chills headache photophobia and back and abdominalpains Among other symptoms neuropsychiatric changeshave been reported in some patients In severe cases 3ndash6 daysafter the onset of the disease hemorrhagic symptoms occurwith petechiae ecchymosis bleeding in the form of melenahematemesis epistaxis [139]
In comparisonwith the nucleocapsid proteins inCCHFVGn and Gc show a higher degree of antigenic variabilityprobably due to their exposition to the host immune systemhypothesis corroborated in the description by Hewson et alin 2004 of four M segment phylogenic groups namely M1M2 M3 and M4 [141 142]
411 mAbs against CCHFV Blackburn et al firstly describedin 1987 murine anti-CCHFV mAbs able to recognize nucle-ocapsid proteins but no neutralizing activity has beendescribed thus far [143]
On the other hand Bertolotti-Ciarlet et al produced apanel of murine mAbs recognizing conformational epitopes
16 BioMed Research International
within theGn andGc glycoproteins expressed by the CCHFVstrain IbAr10200 The effectiveness of the mAbs was studiedby performing PRNT80 and none of the anti-Gn mAbsshowed neutralizing activity although many of the anti-GcmAbs neutralized IbAr10200 strain in vitro with clones 8A15E3 12A9 6C2 and 9H3 showing activity at a gt5120 dilutionMAbs 11E7 and 30F7 exhibited neutralizing activity at 2560dilution A suckling mice protection test was thereforeperformed for these mAbs The previously mentionedanti-Gc Abs were capable to protect mice to an appreciabledegree when applied 24 hours before and in a weakermanner 24 hours after virus challenge Many of the anti-GnmAbs were able to confer significant protection to IbAr10200both 24 hours before and after virus administration Therelevant effectiveness of the anti-Gn mAbs 6B12 10E1113G8 and 10G4 suggests that these mAbs may possess someneutralizing activity due to Ab-based effector mechanisms(eg ADCC) [58]
In further studies the same group characterized thebroadly cross-reactive neutralizing activity of the previouslydescribed murine mAb 11E7 able to bind conformationalepitopes within the C-terminal region of the Gn glycoproteinof all CCHFVM groups [58 141]
42 Rift Valley Fever Virus (RVFV) Similarly to CCHFV RiftValley Fever Virus (RVFV) causes sporadic outbreaks Thelargest epidemic occurred in Egypt in 1977 with an estimated200000 infections 18000 patients manifesting symptomsand 600 fatal prognoses [144] Symptoms include hemor-rhagic episodes fever encephalitis and blindness [145]
The major target of the humoral immune response is theRVFV glycoproteins Gc (G1) and Gn (G2) ranging fromnucleotides 480ndash2090 and 2091ndash3614 respectively [144]
421 mAbs against RVFV Within G2 glycoprotein are des-cribed three neutralizing antigenic sites namely epitope Inucleotides 792ndash893 (amino acid residues 258ndash291) epitopeII nucleotides 1164ndash1196 (amino acid residues 382ndash392) andepitope IV nucleotides 858ndash917 (amino acid residues 280ndash299) [59 145]
Despite the high degree of variability observed inCCHFVglycoproteins RVFV G2 presented an unexpected high con-servation among 22 isolates [59] In the same study onlythe LUNYO and 900060 isolates were resistant to in vitroneutralization by murine mAb 4D4 (recognizing epitope II)All other isolates were neutralized with PRNT80 titers from10240 togt81920On the other hand only SNS isolate showedreduced sensitivity to 4ndash39-CC (epitope IV) All other isolateswere neutralized with PRNT80 titers from 20480 to gt81920Protection studies on mice would be of interest for thedevelopment of a passive immunization prophylaxis [59]
Protection studies from lethal South African RVFVAN1830 strain infection were successfully performed on abattery of anti-G1 and anti-G2 mAbs produced by Besselaarand Blackburn with the strongly neutralizing anti-G1 mAb3E5 and anti-G2mAb 9C4 being the most effective [146]The9C4 antigenic area maps within G2 epitope I and a broadlyneutralizing activity may be investigated
5 Conclusions
Arbovirus infections have acquired increasing interest giventhe augmented globalization and tourism movement all overthe world that could be at the basis of epidemic eventsFurthermore the same vectors can sometimes transmit sev-eral arboviruses concomitantly complicating the diagnosis aswell as the therapy For example confusing mixed epidemicshave occasionally been described such as YFV plus CHIKVDENVplus CHIKV ormore recentlyPlasmodium falciparummalaria plus DENV1 and CHIKV on Madagascarrsquos east coast[1]
In the last decade several efforts have been employed inthe development of effective immunogens and therapeuticsgiving the current absence of specific antiviral drugs as wellas effective vaccines for themost diffused arbovirus infections(ie DENV andWNV)
However at the moment mosquito control is the bestavailable method for preventing arbovirus infections Thusto control the emerging public health problem of arbovirusinfections new antiviral therapeutic strategies that providepotent and broadly cross-protective immunity (especially forflavivirus infections) are an urgent globally medical need
As described in this review the majority of highlyneutralizing mAbs are of murine origin and indeed theirutility in the treatment of these infections is limited In facttheir employment can elicit an immunogenic response (iehuman anti-mouse Abs HAMA) the therapeutic efficacycould be reduced by a relatively faster clearance in humans(compared to human Abs) potentially exacerbated by theHAMA response Finally murine Abs exhibit relatively weakeffector functions (eg ADCC) compared to human AbsIndeed chimerization humanization or better the isolationand characterization of fully human broadly neutralizingmAbs would be the best choice for their possible use in afuture mAb-based therapy against arbovirus infections [147]
Finally possible ADE of infection mechanisms should beevaluated before considering a mAb as a possible candidatetherapeutic in a post-exposure setting as well as in the devel-opment of new Ab-eliciting immunogens
Conflict of Interests
The authors declare no conflict of interests in particular withOmriGam and any other cited Ab-based preparation men-tioned in the paper
References
[1] N Cleton M Koopmans J Reimerink G J Godeke and CReusken ldquoCome fly with me review of clinically importantarboviruses for global travelersrdquo Journal of Clinical Virology vol55 no 3 pp 191ndash203 2012
[2] F X Heinz H Holzmann A Essl and M Kundi ldquoField effec-tiveness of vaccination against tick-borne encephalitisrdquoVaccinevol 25 no 43 pp 7559ndash7567 2007
[3] A D Barrett and D E Teuwen ldquoYellow fever vaccinemdashhowdoes it work and why do rare cases of serious adverse events
BioMed Research International 17
take placerdquo Current Opinion in Immunology vol 21 no 3 pp308ndash313 2009
[4] S B Halstead and S J Thomas ldquoNew vaccines for Japaneseencephalitisrdquo Current Infectious Disease Reports vol 12 no 3pp 174ndash180 2010
[5] R Burioni M Perotti N Mancini and M Clementi ldquoPer-spectives for the utilization of neutralizing human monoclonalantibodies as anti-HCV drugsrdquo Journal of Hepatology vol 49no 2 pp 299ndash300 2008
[6] R A Diotti G A Sautto L Solforosi NManciniM Clementiand R Burioni ldquoNeutralization activity and kinetics of twobroad-range human monoclonal IgG1 derived from recombi-nant Fab fragments and directed against Hepatitis C virus E2glycoproteinrdquo New Microbiologica vol 35 no 4 pp 475ndash4792012
[7] R Burioni F Canducci N Mancini et al ldquoMolecular cloningof the first humanmonoclona antibodies neutralizing with highpotency Swine-origin Influenza A pandemic virus (S-OIV)rdquoNew Microbiologica vol 32 no 4 pp 319ndash324 2009
[8] P K Chandra S Hazari B Poat et al ldquoIntracytoplasmic stableexpression of IgG1 antibody targeting NS3 helicase inhibitsreplication of highly efficient hepatitis C Virus 2a clonerdquoVirology Journal vol 7 article 118 2010
[9] N Clementi D de Marco N Mancini et al ldquoA human mono-clonal antibody with neutralizing activity against highly diver-gent influenza subtypesrdquo PLoS ONE vol 6 no 12 Article IDe28001 2011
[10] D de Marco N Clementi N Mancini et al ldquoA non-VH1-69 heterosubtypic neutralizing human monoclonal antibodyprotects mice against H1N1 and H5N1 virusesrdquo PLoS ONE vol7 no 4 Article ID e34415 2012
[11] L Solforosi N Mancini F Canducci et al ldquoA phage displayvector optimized for the generation of human antibody com-binatorial libraries and the molecular cloning of monoclonalantibody fragmentsrdquoNewMicrobiologica vol 35 no 3 pp 289ndash294 2012
[12] E V Koonin and V V Dolja ldquoEvolution and taxonomy ofpositive-strand RNA viruses implications of comparative anal-ysis of amino acid sequencesrdquo Critical Reviews in Biochemistryand Molecular Biology vol 28 no 5 pp 375ndash430 1993
[13] J S Mackenzie D J Gubler and L R Petersen ldquoEmergingflaviviruses the spread and resurgence of Japanese encephalitisWest Nile and dengue virusesrdquo Nature Medicine vol 10 no 12pp S98ndashS109 2004
[14] T J SmithW E Brandt J L Swanson J MMcCown and E LBuescher ldquoPhysical and biological properties of dengue-2 virusand associated antigensrdquo Journal of Virology vol 5 no 4 pp524ndash532 1970
[15] J M Smit B Moesker I Rodenhuis-Zybert and J WilschutldquoFlavivirus cell entry and membrane fusionrdquo Viruses vol 3 no2 pp 160ndash171 2011
[16] R J Kuhn W Zhang M G Rossmann et al ldquoStructure ofdengue virus implications for flavivirus organization matura-tion and fusionrdquo Cell vol 108 no 5 pp 717ndash725 2002
[17] K Stiasny J H Aberle M Keller B Grubeck-Loebenstein andF X Heinz ldquoAge affects quantity but not quality of antibodyresponses after vaccination with an inactivated flavivirus vac-cine against tick-borne encephalitisrdquo PLoS ONE vol 7 no 3Article ID e34145 2012
[18] LThomasOVerlaetenACabie et al ldquoInfluence of the dengueserotype previous dengue infection and plasma viral load
on clinical presentation and outcome during a dengue-2 anddengue-4 co-epidemicrdquo American Journal of Tropical Medicineand Hygiene vol 78 no 6 pp 990ndash998 2008
[19] Y Zhang J Corver P R Chipman et al ldquoStructures of imma-ture flavivirus particlesrdquo EMBO Journal vol 22 no 11 pp2604ndash2613 2003
[20] A K I Falconar ldquoIdentification of an epitope on the denguevirus membrane (M) protein defined by cross-protective mon-oclonal antibodies design of an improved epitope sequencebased on common determinants present in both envelope (EandM) proteinsrdquo Archives of Virology vol 144 no 12 pp 2313ndash2330 1999
[21] K-J Huang Y-C Yang Y-S Lin et al ldquoThe dual-specific bind-ing of dengue virus and target cells for the antibody-dependentenhancement of dengue virus infectionrdquo Journal of Immunol-ogy vol 176 no 5 pp 2825ndash2832 2006
[22] A K I Falconar ldquoAntibody responses are generated to immun-odominant ELKKLE-typemotifs on the nonstructural-1 glyco-protein during live dengue virus infections inmice and humansimplications for diagnosis pathogenesis and vaccine designrdquoClinical and Vaccine Immunology vol 14 no 5 pp 493ndash5042007
[23] C W Mandl F Guirakhoo H Holzmann F X Heinz and CKunz ldquoAntigenic structure of the flavivirus envelope protein Eat the molecular level using the tick-borne encephalitis virus asa modelrdquo Journal of Virology vol 63 no 2 pp 564ndash571 1989
[24] F A Rey F X Heinz C Mandl C Kunz and S C HarrisonldquoThe envelope glycoprotein from tick-borne encephalitis virusat 2 A resolutionrdquo Nature vol 375 no 6529 pp 291ndash298 1995
[25] S L Allison K Stiasny K Stadler CWMandl and F XHeinzldquoMapping of functional elements in the stem-anchor regionof tick-borne encephalitis virus envelope protein Erdquo Journal ofVirology vol 73 no 7 pp 5605ndash5612 1999
[26] D E Klein J L Choi and S C Harrison ldquoStructure of a denguevirus envelope protein late-stage fusion intermediaterdquo Journal ofVirology vol 87 no 4 pp 2287ndash2293 2013
[27] S L Hanna T C Pierson M D Sanchez A A Ahmed M MMurtadha and R W Doms ldquoN-linked glycosylation of WestNile virus envelope proteins influences particle assembly andinfectivityrdquo Journal of Virology vol 79 no 21 pp 13262ndash132742005
[28] N Mancini G Sautto N Clementi et al ldquoNeutralization inter-fering antibodies a ldquonovelrdquo example of humoral immune dys-function facilitating viral escaperdquoViruses vol 4 no 9 pp 1731ndash1752 2012
[29] Y Modis S Ogata D Clements and S C Harrison ldquoVariablesurface epitopes in the crystal structure of dengue virus type3 envelope glycoproteinrdquo Journal of Virology vol 79 no 2 pp1223ndash1231 2005
[30] N Miller ldquoRecent progress in dengue vaccine research anddevelopmentrdquo Current Opinion in Molecular Therapeutics vol12 no 1 pp 31ndash38 2010
[31] J H Chavez J R Silva A A Amarilla and L T Moraes Figu-eiredo ldquoDomain III peptides from flavivirus envelope proteinare useful antigens for serologic diagnosis and targets forimmunizationrdquo Biologicals vol 38 no 6 pp 613ndash618 2010
[32] B Shrestha J D Brien S Sukupolvi-Petty et al ldquoThe develop-ment of therapeutic antibodies that neutralize homologous andheterologous genotypes of dengue virus type 1rdquo PLoS Pathogensvol 6 no 4 Article ID e1000823 2010
18 BioMed Research International
[33] G D Gromowski N D Barrett and A D T Barrett ldquoChar-acterization of dengue virus complex-specific neutralizing epi-topes on envelope protein domain III of dengue 2 virusrdquo Journalof Virology vol 82 no 17 pp 8828ndash8837 2008
[34] S Sukupolvi-Petty S K Austin W E Purtha et al ldquoType- andsubcomplex-specific neutralizing antibodies against domain IIIof dengue virus type 2 envelope protein recognize adjacentepitopesrdquo Journal of Virology vol 81 no 23 pp 12816ndash128262007
[35] S Sukupolvi-Petty S K Austin M Engle et al ldquoStructure andfunction analysis of therapeutic monoclonal antibodies againstdengue virus type 2rdquo Journal of Virology vol 84 no 18 pp9227ndash9239 2010
[36] Y-Q Deng J-X Dai G-H Ji et al ldquoA broadly flaviviruscross-neutralizingmonoclonal antibody that recognizes a novelepitope within the fusion loop of e proteinrdquo PLoS ONE vol 6no 1 Article ID e16059 2011
[37] J M Costin E Zaitseva K M Kahle et al ldquoMechanistic studyof broadly neutralizing human monoclonal antibodies againstdengue virus that target the fusion looprdquo Journal of Virologyvol 87 no 1 pp 52ndash66 2013
[38] CMMidgley A FlanaganH B Tran et al ldquoStructural analysisof a dengue cross-reactive antibody complexed with envelopedomain III reveals the molecular basis of cross-reactivityrdquoJournal of Immunology vol 188 no 10 pp 4971ndash4979 2012
[39] N J Moreland P Susanto E Lim et al ldquoPhage display appro-aches for the isolation of monoclonal antibodies against denguevirus envelope domain III from human and mouse derivedlibrariesrdquo International Journal ofMolecular Sciences vol 13 no3 pp 2618ndash2635 2012
[40] J J B Cockburn M E Navarro Sanchez N Fretes et alldquoMechanism of dengue virus broad cross-neutralization by amonoclonal antibodyrdquo Structure vol 20 no 2 pp 303ndash3142012
[41] R Rajamanonmani C Nkenfou P Clancy et al ldquoOn a mousemonoclonal antibody that neutralizes all four dengue virusserotypesrdquo Journal of General Virology vol 90 no 4 pp 799ndash809 2009
[42] L H Gould J Sui H Foellmer et al ldquoProtective and herapeuticcapacity of human single-chain Fv-Fc fusion proteins againstWest Nile virusrdquo Journal of Virology vol 79 no 23 pp 14606ndash14613 2005
[43] H Sultana H G Foellmer G Neelakanta et al ldquoFusion LoopPeptide of the West Nile virus envelope protein is essential forpathogenesis and is recognized by a therapeutic cross-reactivehuman monoclonal antibodyrdquo Journal of Immunology vol 183no 1 pp 650ndash660 2009
[44] T Oliphant M Engle G E Nybakken et al ldquoDevelopment ofa humanized monoclonal antibody with therapeutic potentialagainstWest Nile virusrdquoNature Medicine vol 11 no 5 pp 522ndash530 2005
[45] G E Nybakken T Oliphant S Johnson S Burke M S Dia-mond and D H Fremont ldquoStructural basis of West Nile virusneutralization by a therapeutic antibodyrdquo Nature vol 437 no7059 pp 764ndash769 2005
[46] G D Gromowski J T Roehrig M S Diamond J C LeeT J Pitcher and A D T Barrett ldquoMutations of an antibodybinding energy hot spot on domain III of the dengue 2 envelopeglycoprotein exploited for neutralization escaperdquo Virology vol407 no 2 pp 237ndash246 2010
[47] J T Roehrig R A Bolin andR G Kelly ldquoMonoclonal antibodymapping of the envelope glycoprotein of the dengue 2 virusJamaicardquo Virology vol 246 no 2 pp 317ndash328 1998
[48] R de Alwis S A Smith N P Olivarez et al ldquoIdentification ofhumanneutralizing antibodies that bind to complex epitopes ondengue virionsrdquo Proceedings of the National Academy of Sciencesof the United States of America vol 109 no 19 pp 7439ndash74442012
[49] S K Austin K A Dowd B Shrestha et al ldquoStructural basis ofdifferential neutralization of DENV-1 genotypes by an antibodythat recognizes a cryptic epitoperdquo PLoS Pathogens vol 8 no 10Article ID e1002930 2012
[50] M Throsby C Geuijen J Goudsmit et al ldquoIsolation and char-acterization of human monoclonal antibodies from individualsinfected with West Nile Virusrdquo Journal of Virology vol 80 no14 pp 6982ndash6992 2006
[51] A P Goncalvez R H Purcell and C-J Lai ldquoEpitope deter-minants of a chimpanzee Fab antibody that efficiently cross-neutralizes dengue type 1 and type 2 viruses map to inside andin close proximity to fusion loop of the dengue type 2 virusenvelope glycoproteinrdquo Journal of Virology vol 78 no 23 pp12919ndash12928 2004
[52] J T Roehrig J H Mathews and D W Trent ldquoIdentification ofepitopes on the E glycoprotein of Saint Louis encephalitis virususing monoclonal antibodiesrdquo Virology vol 128 no 1 pp 118ndash126 1983
[53] J T Roehrig L A Staudinger A R Hunt J H Mathews andC D Blair ldquoAntibody prophylaxis and therapy for flavivirusencephalitis infectionsrdquo Annals of the New York Academy ofSciences vol 951 pp 286ndash297 2001
[54] A P Goncalvez C-H Chien K Tubthong et al ldquoHumanizedmonoclonal antibodies derived from chimpanzee fabs protectagainst Japanese encephalitis virus in vitro and in vivordquo Journalof Virology vol 82 no 14 pp 7009ndash7021 2008
[55] M Arakawa T Yamashiro G-I Uechi M Tadano and ANishizono ldquoConstruction of humanFab (1205741120581) library and iden-tification of human monoclonal Fab possessing neutralizingpotency against Japanese encephalitis virusrdquo Microbiology andImmunology vol 51 no 6 pp 617ndash625 2007
[56] L Warter C Y Lee R Thiagarajan et al ldquoChikungunya virusenvelope-specific human monoclonal antibodies with broadneutralization potencyrdquo Journal of Immunology vol 186 no 5pp 3258ndash3264 2011
[57] P Pal K A Dowd J D Brien et al ldquoDevelopment of a highlyprotective combination monoclonal antibody therapy againstchikungunya virusrdquo PLoS Pathogens vol 9 no 4 Article IDe1003312 2013
[58] A Bertolotti-Ciarlet J Smith K Strecker et al ldquoCellular local-ization and antigenic characterization of Crimean-Congo hem-orrhagic fever virus glycoproteinsrdquo Journal of Virology vol 79no 10 pp 6152ndash6161 2005
[59] J K Battles and J M Dalrymple ldquoGenetic variation amonggeographic isolates of Rift Valley fever virusrdquo American Journalof Tropical Medicine and Hygiene vol 39 no 6 pp 617ndash6311988
[60] B AThibodeaux N C Garbino NM Liss et al ldquoA humanizedIgG but not IgM antibody is effective in prophylaxis and therapyof yellow fever infection in an AG12917D-204 peripheralchallenge mouse modelrdquo Antiviral Research vol 94 no 1 pp1ndash8 2012
BioMed Research International 19
[61] S Daffis R E Kontermann J Korimbocus H Zeller H-DKlenk and J Ter Meulen ldquoAntibody responses against wild-type yellow fever virus and the 17D vaccine strain charac-terization with human monoclonal antibody fragments andneutralization escape variantsrdquoVirology vol 337 no 2 pp 262ndash272 2005
[62] L N Levanov L E Matveev E P Goncharova et al ldquoChimericantibodies against tick-borne encephalitis virusrdquo Vaccine vol28 no 32 pp 5265ndash5271 2010
[63] A R Hunt S Frederickson C Hinkel K S Bowdish and J TRoehrig ldquoA humanized murine monoclonal antibody protectsmice either before or after challenge with virulent Venezuelanequine encephalomyelitis virusrdquo Journal of General Virologyvol 87 no 9 pp 2467ndash2476 2006
[64] J M da Silva Voorham I A Rodenhuis-Zybert N V A Nunezet al ldquoAntibodies against the envelope glycoprotein promoteinfectivity of immature dengue virus serotype 2rdquo PLoS ONEvol 7 no 3 Article ID e29957 2012
[65] T C Pierson Q Xu S Nelson et al ldquoThe stoichiometry ofantibody-mediated neutralization and enhancement of westnile virus infectionrdquoCell Host andMicrobe vol 1 no 2 pp 135ndash145 2007
[66] G Sautto N Mancini R A Diotti L Solforosi M Clementiand R Burioni ldquoAnti-hepatitis C virus E2 (HCVE2) glyco-proteinmonoclonal antibodies andneutralization interferencerdquoAntiviral Research vol 96 no 1 pp 82ndash89 2012
[67] R Burioni Y Matsuura N Mancini et al ldquoDiverging effectsof human recombinant anti-hepatitis C virus (HCV) antibodyfragments derived from a single patient on the infectivityof a vesicular stomatitis virusHCV pseudotyperdquo Journal ofVirology vol 76 no 22 pp 11775ndash11779 2002
[68] R J Phillpotts J R Stephenson and J S Porterfield ldquoAntibody-dependent enhancement of tick-borne encephalitus virus infec-tivityrdquo Journal of General Virology vol 66 no 8 pp 1831ndash18371985
[69] K A Dowd and T C Pierson ldquoAntibody-mediated neutraliza-tion of flaviviruses a reductionist viewrdquo Virology vol 411 no 2pp 306ndash315 2011
[70] A K I Falconar ldquoMonoclonal antibodies that bind to commonepitopes on the dengue virus type 2 nonstructural-1 andenvelope glycoproteins display weak neutralizing activity anddifferentiated responses to virulent strains implications forpathogenesis and vaccinesrdquo Clinical and Vaccine Immunologyvol 15 no 3 pp 549ndash561 2008
[71] K Meyer A Banerjee S E Frey R B Belshe and R Ray ldquoAweak neutralizing antibody response to hepatitis C virus enve-lope glycoprotein enhances virus infectionrdquo PLoS ONE vol 6no 8 Article ID e23699 2011
[72] R Burioni R AWilliamson P P Sanna F E Bloom and D RBurton ldquoRecombinant human Fab to glycoprotein D neutral-izes infectivity and prevents cell-to-cell transmission of herpessimplex viruses 1 and 2 in vitrordquo Proceedings of the NationalAcademy of Sciences of the United States of America vol 91 no1 pp 355ndash359 1994
[73] M S Rolph A Zaid N E Rulli and S Mahalingam ldquoDown-regulation of interferon-120573 in antibody-dependent enhancementof dengue viral infections of human macrophages is dependenton interleukin-6rdquo Journal of Infectious Diseases vol 204 no 3pp 489ndash491 2011
[74] S S Whitehead J E Blaney A P Durbin and B R MurphyldquoProspects for a dengue virus vaccinerdquoNature ReviewsMicrobi-ology vol 5 no 7 pp 518ndash528 2007
[75] S-M Lok V Kostyuchenko G ENybakken et al ldquoBinding of aneutralizing antibody to dengue virus alters the arrangement ofsurface glycoproteinsrdquoNature Structural andMolecular Biologyvol 15 no 3 pp 312ndash317 2008
[76] A P Goncalvez R Men C Wernly R H Purcell and C-J Lai ldquoChimpanzee Fab fragments and a derived humanizedimmunogiobulin G1 antibody that efficiently cross-neutralizedengue type 1 and type 2 virusesrdquo Journal of Virology vol 78no 23 pp 12910ndash12918 2004
[77] WM P BWahala A A Kraus L B HaymoreM A Accavitti-Loper and A M de Silva ldquoDengue virus neutralization byhuman immune sera role of envelope protein domain III-reactive antibodyrdquo Virology vol 392 no 1 pp 103ndash113 2009
[78] W D Crill H R Hughes M J Delorey and G-J J ChangldquoHumoral immune responses of dengue fever patients usingepitope-specific serotype-2 virus-like particle antigensrdquo PLoSONE vol 4 no 4 Article ID e4991 2009
[79] C Setthapramote T Sasaki O Puiprom et al ldquoHuman mono-clonal antibodies to neutralize all dengue virus serotypes usinglymphocytes from patients at acute phase of the secondaryinfectionrdquo Biochemical and Biophysical Research Communica-tions vol 423 no 4 pp 867ndash872 2012
[80] M Beltramello K L Williams C P Simmons et al ldquoThehuman immune response to dengue virus is dominated byhighly cross-reactive antibodies endowed with neutralizing andenhancing activityrdquo Cell Host andMicrobe vol 8 no 3 pp 271ndash283 2010
[81] C M Midgley M Bajwa-Joseph S Vasanawathana et al ldquoAnin-depth analysis of original antigenic sin in dengue virusinfectionrdquo Journal of Virology vol 85 no 22 pp 410ndash421 2011
[82] B Kaufmann M R Vogt J Goudsmit et al ldquoNeutralizationof West Nile virus by cross-linking of its surface proteins withFab fragments of the human monoclonal antibody CR4354rdquoProceedings of the National Academy of Sciences of the UnitedStates of America vol 107 no 44 pp 18950ndash18955 2010
[83] B Spurrier J M Sampson M Totrov et al ldquoStructural analysisof human and macaque mAbs 2909 and 25B implicationsfor the configuration of the quaternary neutralizing epitope ofHIV-1 gp120rdquo Structure vol 19 no 5 pp 691ndash699 2011
[84] J J SchlesingerMWBrandriss andT PMonath ldquoMonoclonalantibodies distinguish between wild and vaccine strains ofyellow fever virus by neutralization hemagglutination inhibi-tion and immune precipitation of the virus envelope proteinrdquoVirology vol 125 no 1 pp 8ndash17 1983
[85] M W Brandriss J J Schlesinger E E Walsh and M BrisellildquoLethal 17D yellow fever encephalitis in mice I Passive protec-tion by monoclonal antibodies to the envelope proteins of 17Dyellow fever and dengue 2 virusesrdquo Journal of General Virologyvol 67 no 2 pp 229ndash234 1986
[86] H Cho and M S Diamond ldquoImmune responses to West Nilevirus infection in the central nervous systemrdquo Viruses vol 4no 12 pp 3812ndash3830 2012
[87] D W Beasley A D Barrett and R B Tesh ldquoResurgence ofWest Nile neurologic disease in the United States in 2012 whathappened What needs to be donerdquo Antiviral Research vol 99no 1 pp 1ndash5 2013
[88] M Throsby J Ter Meulen C Geuijen J Goudsmit and J DeKruif ldquoMapping and analysis of West Nile virus-specific mon-oclonal antibodies prospects for vaccine developmentrdquo ExpertReview of Vaccines vol 6 no 2 pp 183ndash191 2007
[89] M S Diamond T G Roberts D Edgil B Lu J Ernst and EHarris ldquoModulation of dengue virus infection in human cells
20 BioMed Research International
by alpha beta and gamma interferonsrdquo Journal of Virology vol74 no 11 pp 4957ndash4966 2000
[90] Z Shimoni M J Niven S Pitlick and S Bulvik ldquoTreatmentof West Nile virus encephalitis with intravenous immunoglob-ulinrdquo Emerging Infectious Diseases vol 7 no 4 article 759 2001
[91] AHamdan PGreen EMendelsonM R Kramer S Pitlik andM Weinberger ldquoPossible benefit of intravenous immunoglob-ulin therapy in a lung transplant recipient with West Nile virusencephalitisrdquoTransplant InfectiousDisease vol 4 no 3 pp 160ndash162 2002
[92] M Haley A S Retter D Fowler J Gea-Banacloche and NP OrsquoGrady ldquoThe role for intravenous immunoglobulin in thetreatment of West Nile virus encephalitisrdquo Clinical InfectiousDiseases vol 37 no 6 pp e88ndashe90 2003
[93] K M Chung G E Nybakken B S Thompson et al ldquoAnti-bodies against West Nile virus nonstructural protein NS1prevent lethal infection through Fc 120574 receptor-dependent and-independent mechanismsrdquo Journal of Virology vol 80 no 3pp 1340ndash1351 2006
[94] E A Henchal L S Henchal and J J Schlesinger ldquoSynergisticinteractions of anti-NS1 monoclonal antibodies protect pas-sively immunized mice from lethal challenge with dengue 2virusrdquo Journal of General Virology vol 69 no 8 pp 2101ndash21071988
[95] J T Roehrig ldquoAntigenic structure of flavivirus proteinsrdquoAdvances in Virus Research vol 59 pp 141ndash175 2003
[96] M D Sanchez T C Pierson D McAllister et al ldquoCharacteri-zation of neutralizing antibodies to West Nile virusrdquo Virologyvol 336 no 1 pp 70ndash82 2005
[97] J H Beigel J L Nordstrom S R Pillemer et al ldquoSafety andpharmacokinetics of single intravenous dose of MGAWN1 anovel monoclonal antibody to West Nile virusrdquo AntimicrobialAgents and Chemotherapy vol 54 no 6 pp 2431ndash2436 2010
[98] C A Smeraski V Siddharthan and J D Morrey ldquoTreatmentof spatial memory impairment in hamsters infected with WestNile virus using a humanizedmonoclonal antibodyMGAWN1rdquoAntiviral Research vol 91 no 1 pp 43ndash49 2011
[99] D Lelli AMoreno E Brocchi et al ldquoWestNile virus character-ization and diagnostic applications of monoclonal antibodiesrdquoVirology Journal vol 9 article 81 2012
[100] T S Gritsun V A Lashkevich and E A Gould ldquoTick-borneencephalitisrdquo Antiviral Research vol 57 no 1-2 pp 129ndash1462003
[101] D Ruzek T S Gritsun N L Forrester et al ldquoMutations inthe NS2B and NS3 genes affect mouse neuroinvasiveness of aWestern European field strain of tick-borne encephalitis virusrdquoVirology vol 374 no 2 pp 249ndash255 2008
[102] M Ecker S L Allison T Meixner and F X Heinz ldquoSequenceanalysis and genetic classification of tick-borne encephalitisviruses from Europe and Asiardquo Journal of General Virology vol80 no 1 pp 179ndash185 1999
[103] H Holzmann M S Vorobyova I P Ladyzhenskaya et alldquoMolecular epidemiology of tick-borne encephalitis viruscross-protection between European and Far Eastern subtypesrdquoVaccine vol 10 no 5 pp 345ndash349 1992
[104] C W Mandl ldquoSteps of the tick-borne encephalitis virus repli-cation cycle that affect neuropathogenesisrdquo Virus Research vol111 no 2 pp 161ndash174 2005
[105] A V Atrasheuskaya T M Fredeking and G M IgnatyevldquoChanges in immune parameters and their correction in humancases of tick-borne encephalitisrdquo Clinical and ExperimentalImmunology vol 131 no 1 pp 148ndash154 2003
[106] K L Mansfield N Johnson L P Phipps J R Stephenson AR Fooks and T Solomon ldquoTick-borne encephalitis virusmdashareview of an emerging zoonosisrdquo Journal of General Virologyvol 90 no 8 pp 1781ndash1794 2009
[107] DWVaughn andCHHoke Jr ldquoThe epidemiology of Japaneseencephalitis prospects for preventionrdquo Epidemiologic Reviewsvol 14 pp 197ndash221 1992
[108] T Solomon H Ni D W C Beasley M Ekkelenkamp M JCardosa and A D T Barrett ldquoOrigin and evolution of Japaneseencephalitis virus in Southeast Asiardquo Journal of Virology vol 77no 5 pp 3091ndash3098 2003
[109] M R Wilson ldquoEmerging viral infectionsrdquo Current Opinion inNeurology vol 26 no 3 pp 301ndash306 2013
[110] J N Hanna S A Ritchie D A Phillips et al ldquoAn outbreakof Japanese encephalitis in the Torres Strait Australia 1995rdquoMedical Journal of Australia vol 165 no 5 pp 256ndash260 1996
[111] J N Hanna S A Ritchie D A Phillips et al ldquoJapanese ence-phalitis in north Queensland Australia 1998rdquo Medical Journalof Australia vol 170 no 11 pp 533ndash536 1999
[112] SGeorgeMGourie-Devi and J A Rao ldquoIsolation ofWestNilevirus from the brains of children who had died of encephalitisrdquoBulletin of theWorldHealthOrganization vol 62 no 6 pp 879ndash882 1984
[113] A K Gupta A A Koshy and V J Lad ldquoEnhanced protec-tion of mice against Japanese encephalitis virus infection bycombinations ofmonoclonal antibodies to glycoprotein ErdquoActaVirologica vol 55 no 2 pp 165ndash168 2011
[114] T H Lee B-H Song S-I Yun et al ldquoA cross-protective mAbrecognizes a novel epitope within the flavivirus NS1 proteinrdquoJournal of General Virology vol 93 no 1 pp 20ndash26 2012
[115] S C Weaver ldquoHost range amplification and arboviral diseaseemergencerdquo Archives of Virology Supplementum no 19 pp 33ndash44 2005
[116] M Solignat B Gay S Higgs L Briant and C Devaux ldquoRepli-cation cycle of chikungunya a re-emerging arbovirusrdquoVirologyvol 393 no 2 pp 183ndash197 2009
[117] O Schwartz and M L Albert ldquoBiology and pathogenesis ofchikungunya virusrdquo Nature Reviews Microbiology vol 8 no 7pp 491ndash500 2010
[118] W Akahata Z-Y Yang H Andersen et al ldquoA virus-like particlevaccine for epidemic Chikungunya virus protects nonhumanprimates against infectionrdquo Nature Medicine vol 16 no 3 pp334ndash338 2010
[119] M Kielian and A Helenius ldquopH-Iinduced alterations in thefusogenic spike protein of Semliki Forest virusrdquo Journal of CellBiology vol 101 no 6 pp 2284ndash2291 1985
[120] J White and A Helenius ldquopH-Dependent fusion between theSemliki Forest virus membrane and liposomesrdquo Proceedings ofthe National Academy of Sciences of the United States of Americavol 77 no 6 pp 3273ndash3277 1980
[121] J Dubuisson and C M Rice ldquoSindbis virus attachment isola-tion and characterization of mutants with impaired binding tovertebrate cellsrdquo Journal of Virology vol 67 no 6 pp 3363ndash33741993
[122] C Y Lee Y-W Kam J Fric et al ldquoChikungunya virus neutral-ization antigens and direct cell-to-cell transmission are revealedby human antibody-escapemutantsrdquo PLoS Pathogens vol 7 no12 Article ID e1002390 2011
[123] J E Voss M-C Vaney S Duquerroy et al ldquoGlycoproteinorganization of Chikungunya virus particles revealed by X-raycrystallographyrdquo Nature vol 468 no 7324 pp 709ndash712 2010
BioMed Research International 21
[124] L Li J Jose Y Xiang R J Kuhn and M G Rossmann ldquoStruc-tural changes of envelope proteins during alphavirus fusionrdquoNature vol 468 no 7324 pp 705ndash708 2010
[125] J Fric S Bertin-Maghit C I Wang A Nardin and L WarterldquoUse of human monoclonal antibodies to treat Chikungunyavirus infectionrdquo Journal of Infectious Diseases vol 207 no 2 pp319ndash322 2013
[126] P V Aguilar J G Estrada-Franco R Navarro-Lopez C FerroA D Haddow and S C Weaver ldquoEndemic Venezuelanequine encephalitis in the Americas hidden under the dengueumbrellardquo Future Virology vol 6 no 6 pp 721ndash740 2011
[127] S C Weaver R Winegar I D Manger and N L ForresterldquoAlphaviruses population genetics and determinants of emer-gencerdquo Antiviral Research vol 94 no 3 pp 242ndash257 2012
[128] S Paessler and S C Weaver ldquoVaccines for Venezuelan equineencephalitisrdquo Vaccine vol 27 no 4 pp D80ndashD85 2009
[129] R J Phillpotts ldquoVenezuelan equine encephalitis virus complex-specific monoclonal antibody provides broad protection inmurine models against airborne challenge with viruses fromserogroups I II and IIIrdquoVirus Research vol 120 no 1-2 pp 107ndash112 2006
[130] R J Phillpotts L D Jones and S C Howard ldquoMonoclonalantibody protectsmice against infection anddiseasewhen giveneither before or up to 24 h after airborne challenge with virulentVenezuelan equine encephalitis virusrdquoVaccine vol 20 no 11-12pp 1497ndash1504 2002
[131] J H Mathews J T Roehrig and D W Trent ldquoRole of com-plement and the Fc portion of immunoglobulin G in immunityto Venezuelan equine encephalomyelitis virus infection withglycoprotein-specific monoclonal antibodiesrdquo Journal of Virol-ogy vol 55 no 3 pp 594ndash600 1985
[132] L M OrsquoBrien C D Underwood-Fowler S A Goodchild AL Phelps and R J Phillpotts ldquoDevelopment of a novel mon-oclonal antibody with reactivity to a wide range of Venezuelanequine encephalitis virus strainsrdquoVirology Journal vol 6 article206 2009
[133] L M OrsquoBrien S A Goodchild R J Phillpotts and S D Per-kins ldquoA humanised murine monoclonal antibody protectsmice from Venezuelan equine encephalitis virus Evergladesvirus and Mucambo virus when administered up to 48h afterairborne challengerdquo Virology vol 426 no 2 pp 100ndash105 2012
[134] N Clementi N Mancini L Solforosi M Castelli M Clementiand R Burioni ldquoPhage display-based strategies for cloning andoptimization of monoclonal antibodies directed against humanpathogensrdquo International Journal of Molecular Sciences vol 13no 7 pp 8273ndash8292 2012
[135] A R Hunt R A Bowen S Frederickson T Maruyama JT Roehrig and C D Blair ldquoTreatment of mice with humanmonoclonal antibody 24h after lethal aerosol challenge withvirulent Venezuelan equine encephalitis virus prevents diseasebut not infectionrdquo Virology vol 414 no 2 pp 146ndash152 2011
[136] W-G Hu A L Phelps S Jager et al ldquoA recombinant human-ized monoclonal antibody completely protects mice againstlethal challenge with Venezuelan equine encephalitis virusrdquoVaccine vol 28 no 34 pp 5558ndash5564 2010
[137] T Rulker L Voss P Thullier et al ldquoIsolation and characterisa-tion of a human-like antibody fragment (scFv) that inactivatesVEEV in vitro and in vivordquo PLoS ONE vol 7 no 5 Article IDe37242 2012
[138] C T Walter and J N Barr ldquoRecent advances in the molecularand cellular biology of bunyavirusesrdquo Journal of General Virol-ogy vol 92 no 11 pp 2467ndash2484 2011
[139] C A Whitehouse ldquoCrimean-Congo hemorrhagic feverrdquo Anti-viral Research vol 64 no 3 pp 145ndash160 2004
[140] M Mertens K Schmidt A Ozkul and M H Groschup ldquoTheimpact of Crimean-Congo hemorrhagic fever virus on publichealthrdquo Antiviral Research vol 98 no 2 pp 248ndash260 2013
[141] A A Ahmed J M McFalls C Hoffmann et al ldquoPresence ofbroadly reactive and group-specific neutralizing epitopes onnewly described isolates of Crimean-Congo hemorrhagic fevervirusrdquo Journal of General Virology vol 86 no 12 pp 3327ndash33362005
[142] R Hewson A Gmyl L Gmyl et al ldquoEvidence of segment reas-sortment in Crimean-Congo haemorrhagic fever virusrdquo Journalof General Virology vol 85 no 10 pp 3059ndash3070 2004
[143] N K Blackburn T G Besselaar A J Shepherd and R Swane-poel ldquoPreparation and use of monoclonal antibodies for iden-tifying Crimean-Congo hemorrhagic fever virusrdquo AmericanJournal of Tropical Medicine and Hygiene vol 37 no 2 pp 392ndash397 1987
[144] T Ikegami ldquoMolecular biology and genetic diversity of RiftValley fever virusrdquoAntiviral Research vol 95 no 3 pp 293ndash3102012
[145] KKeegan andM S Collett ldquoUse of bacterial expression cloningto define the amino acid sequences of antigenic determinantson the G2 glycoprotein of Rift Valley fever virusrdquo Journal ofVirology vol 58 no 2 pp 263ndash270 1986
[146] T G Besselaar and N K Blackburn ldquoTopological mapping ofantigenic sites on the Rift Valley fever virus envelope glycopro-teins using monoclonal antibodiesrdquo Archives of Virology vol121 no 1ndash4 pp 111ndash124 1991
[147] L G Presta ldquoEngineering of therapeutic antibodies to mini-mize immunogenicity and optimize functionrdquo Advanced DrugDelivery Reviews vol 58 no 5-6 pp 640ndash656 2006
Submit your manuscripts athttpwwwhindawicom
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anatomy Research International
PeptidesInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporation httpwwwhindawicom
International Journal of
Volume 2014
Zoology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Molecular Biology International
GenomicsInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioinformaticsAdvances in
Marine BiologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Signal TransductionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Evolutionary BiologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Biochemistry Research International
ArchaeaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Genetics Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in
Virolog y
Hindawi Publishing Corporationhttpwwwhindawicom
Nucleic AcidsJournal of
Volume 2014
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Enzyme Research
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Microbiology

BioMed Research International 3
The crystallographic structure of the E protein of TBEVhas been proposed as a model for the envelope protein of theflaviviruses [23]
Further studies performed throughX-ray crystallographyhave revealed that the E protein is a type II viral fusionprotein with three 120573-barrel domains (D) DI DII and DIII(corresponding to the antigenic domains C A and B resp)connected to the viral membrane by a helical structurecalled the stem anchor [24] In its native form the E proteinfolds as an homodimer with an antiparallel structure andan unusual herringbone pseudoicosaedral symmetry patternwith the M protein located centrally within the symmetryconsistent with a head-to-tail configuration lying parallel tothe envelope lipid bilayer The DIII of the E glycoprotein(amino acid residues 295ndash395) has an immunoglobulin-like fold contains seven 120573-strands as well as type- andsubcomplex-specific neutralizing B-cell epitopes and is theproposed receptor-binding domain through four peptideloops on the solvent exposed face Directly linked to DIIIthere is the stemtransmembrane region spanning aminoacid residues 401ndash495 and containing regions important foroligomerization with prM protein (amino acid residues 431ndash449 and 450ndash472) [25] DII has a long finger-like structureand contains two extended loops that project from DI andthe highly conserved and hydrophobic glycine-rich fusionloop at its tip (amino acid residues 98ndash110) interacting withthe stem region and the endosomal membrane of the cellduring the fusion process [26] In the trimeric conformationthe hydrophobic fusion peptide is exposed in DII to mediatethe fusion with the membranes of the cell DI is a sim120amino acid central domain formed by a discontinuous hingeregion consisting in a central 120573-barrel of eight strands thatconnectsDII andDIII andplays a crucial role in the structuralrearrangement of E from a homodimer to a trimer whichoccurs on exposure to low pH of endosomes that is requiredfor the membrane fusion process In some flaviviruses anN-linked glycosylation site is present at amino acid position154ndash156 of the E protein and does not appear to be necessaryfor E function but it has been associated with the increasedneuroinvasiveness of WNV lineage I outbreak strains fromthe USA and a pH-sensitive decrease in stability of the non-glycosylated strains [27]
22 Flavivirus Neutralizing B-Cell Epitopes As previouslydescribed Abs are a significant component of the hostrsquosprotective response against Flavivirus infections Howevervirus-specific Abs have been implicated in the pathogenesisof severe clinicalmanifestations following a secondaryDENVas well as YFV andWNV infections
Moreover cross-reactivity between Flavivirus serogroupscould complicate the interpretation of diagnostic assays butcould be of interest in isolating cross-reactive and cross-neutralizing mAbs However Ab-dependent enhancement(ADE) of infection has been described for this genus as itrepresents the major risk of complications when reinfectionoccursThus great caremust be taken in evaluating neutraliz-ing activity and possibleAb-mediated infection enhancementin the characterization of future possible therapeutic mAbsagainst these viruses [28]
However prophylactic and therapeutic use of neutral-izing mAbs for Flavivirus infections has been shown to beeffective in animal models as reported later
Neutralizing epitopes were found to be located in each ofthe three domains of the E protein and have been confirmedto be surface exposed in the high-resolution X-ray crystalstructure of the pre-fusion dimer of DENV2 E glycoprotein[29]
Most potentmAbs against flaviviruses are directed towardepitopes on DIII For this reason DIII-based immunogensare under evaluation as promising subunit Flavivirus candi-date vaccines [30 31] However several cross-reactive mAbsthat bind to residues from the AB loop were found to bepoorly neutralizing as this loop projects toward the lipidbilayer in the mature viral particle Conversely lateral ridgeregion of DIII (eg BC DE and FG loops) is targeted bystrong serotype-specific neutralizing mAbs [32ndash35] Cross-reactive mAbs specific for DII have also been described butwith less neutralizing and variable profiles However few ofthem whose epitope encompasses the fusion loop are cross-reactive and neutralizing [36]
23 Ab-Dependent Enhancement (ADE) of Infection In vitroand in vivo studies have demonstrated that increased diseaseseverity causing DHFDSS upon reinfection with a differentDENV serotype is mostly due to the phenomenon of ADE ofinfection determined by cross-reacting but poorly or non-neutralizing Abs generated during the primary infectionthat facilitate virus entry through Fc-120574 or complementreceptors present on cells such as monocytesmacrophages(Figure 1(a)) Additionally in 6 to 9 month-old children ofDENV immune mothers severe disease is associated withprimary infection possibly because of declining levels ofneutralizing maternal Abs [64]
These cross-reactive Abs are mainly directed against theE and prM glycoproteins In particular experimental lines ofevidence have suggested that cross-reactiveAbs againstDII ofE protein can under certain conditions enhance infectivityof WNV in vitro [65] However the phenomenon of ADEcould reflect the presence of non-neutralizing concentrationsof virus-reactive Abs and has been observed for severalflaviviruses in vitro and in vivo such as WNV TBEV JEVand YFV as well as other Flaviviridae members [28 66ndash68]In particular it has been described that ADE of infectionoccurs at virus-boundedAb concentrations in the upper limitby the stoichiometric threshold for neutralization and atlower concentrations by the minimal number of Abs thatallow attachment of the virion to cells Furthermore Abs thatrecognize infrequently displayed epitopes that do not supportneutralizationmay enhance infection even at saturation [69]
Thus the ADE of infection mechanismmay pose a threatfor the development of a safe and efficacious vaccine as wellas a possible immunotherapy against Flaviviridae
Indeed cross-neutralizing mAbs as candidate therapeu-tics for these infections may be considered after having wellascertained their broadly cross-neutralizing activity and theabsence of possible mAb-mediated ADE of infection mech-anisms Alternatively the risk of ADE of infection could beovertaken by the removal of the Ab heavy chain (ie the CH2
4 BioMed Research International
receptorFc-120574
Cellular receptor
(a)
Cellular receptor
(b) (c)
Figure 1 Mechanisms of antibody-dependent enhancement (ADE) of infection (a) After binding to the viral epitope the Ab is recognizedby a cellular Fc-120574 receptor bringing the viral particle in proximity of the target cell (b) the binding of the Ab induces conformational changeswithin the structure of the viral target protein These changes improve the affinity for the cellular receptor (c) molecular mimicry by a viralmotif of cellular membrane components leads an autoreactive Ab to bind both the viral and the cellular target bringing the virus in proximityof the target cell
and CH3 portions or direct expression of the mAb as aFab fragment) andor deletion of the N-linked sugars onIgG molecules that are both required for interactions withFc-120574 receptors (Fc-120574R) or eventually by the blocking ofFc-120574R engagement with anti-Fc-120574R Abs [70] Similarly theidentification and detailed examination of cross-neutralizingepitopes that do not promote ADE of infection may definenovel targets for vaccine development
However in some instances ADE of infection mayoccur when Ab molecules cross-react with both viral andcellular antigens (Figure 1(c)) In this regard Fc-120574 receptor-independentmechanismof infection in particular forDENVprM-specific Abs able to bind simultaneously to the virus andtarget cells has been described Indeed othermembers of theFlaviviridae belonging to another genus like hepatitis C virus(HCV) have been shown to elicit polyreactive Abs that areprobably responsible for an ADE of infection mechanism aswell as of secondary clinical manifestations In connectionwith this several studies employing both polyclonal andmonoclonal Abs suggested the presence of common epitopescontaining particular short motifs on NS1 and the threedomains of the E protein as well as human proteins thatmay play a role in ADE of infection mechanisms and thusin DHFDSS pathogenesis [70] Interestingly these commonmotifs have been found to be additionally represented inDENV2 strains which have greater human pathogenic capac-ities [22]
Moreover another molecular mechanism of ADE ofinfection could consist in the facilitation of conformationalchanges of the targeted surface protein that are required forvirus entry as described and hypothesized for other viruses(eg HIV HCV and HSV) (Figure 1(b)) [71 72]
Finally Fc-120574R engagement could result also in ldquointrinsicrdquoADE of infection mechanisms involving signaling eventssuch as inhibition of antiviral response and increased viralreplication Disrupted signaling events include the RIG-IMDA5 cascade and type I interferon (IFN) response aswell as induction and suppression of anti-inflammatory(IL-10) and proinflammatory (IL-12 and TNF-120572) cytokinesrespectively [73]
24 Dengue Virus (DENV) Dengue virus (DENV) is respon-sible for 50ndash100 million symptomatic infections each yearresulting in 500000 hospitalizations and over 20000 deathswhich occurs mostly in tropical and subtropical regions ofthe world Although malaria remains the most importantcause for systemic febrile disease in travelers chikungunyavirus (CHIKV) and DENV are increasingly diagnosed withdengue currently being the second most important cause forfebrile disease in travelers
Decreases in mosquito control efforts during the endof the 90s coupled with societal factors (eg globalizationmigrations and dense urbanization) have contributed to thereemergence of flaviviruses such as DENV in South and Cen-tral America Development of effective DENV vaccines thatexhibit cross-protection thought to be important for prevent-ing subsequent dengue-associated immunopathogenesis isproving to be particularly challenging [1]
A vaccine for this infection is not yet available despiteconsiderable public and private efforts This difficulty ismainly due to the perceived need to simultaneously protectthe four known serotypes of DENV (DENV1ndash4) which shareabout 70 of sequence homology while genotypes can varyup to 3 Moreover as anticipated and discussed previously
BioMed Research International 5
non-protective Abs may contribute to more severe clinicaloutcomes in vaccinated people [74]
241 Murine versus Human Anti-DENV Humoral ImmuneResponse It is well ascertained that anti-E DIII-directed Absare virtually absent from the naıve human repertoire as theyare directed away from this domain probably against weaklyneutralizing and immunodominant regions Moreover it hasbeen demonstrated that over than 90 of the human Abresponse of primary DENV infected patients is able to bindonly native DENV virus particles instead of a recombinantform of the E protein posing difficulties in the cloning strate-gies of neutralizing mAbs [39] Thus it can be concludedthat in vivo only a small fraction of DENV-specific Abs areresponsible for neutralization
Conversely murine mAbs that recognize all the threedomains of E have been identified Epitope-mapping studiesusingmurine serotype-specificmAbs have demonstrated thatloops located in the lateral ridge region of DIII consti-tute the strongest neutralizing B-cell target In this regionsequence diversity between serotype is high while cross-reactive murine mAbs that recognize another loop showedgenerally a weaker neutralization profile In particular type-specific mAbs with neutralizing activity against DENV2localized to the BC DE and FG loops on the lateral ridgeof DIII whereas subcomplex-specific mAbs recognized anadjacent epitope centered on the connecting A strand of DIIIat residues K305 K307 and K310 [32ndash35]
Cross-reactive mAbs directed against prM and NS1 havealso been described but as anticipated they feature weaklyneutralizing or absent as well as ADE of infection activityIn the following paragraphs we report the better character-ized neutralizing mouse and human mAbs describing theirmolecular features further summarized in Table 1
242 Anti-DENV Murine mAbs Murine mAbs directedagainst all the three domains of the E protein and endowedof different neutralizing and binding characteristics havebeen described However only DIII and DII are classicallyrecognized by neutralizing Abs In this regard Sukupolvi-Petty et al isolated twenty-four anti-DENV2 mouse mAbswith moderate or strong neutralizing activity against thehomologous virus in cell culture assays Binding sites weremapped for the majority of these in distinct epitopes inregions located in DI (lateral ridge) DII (dimer interfacelateral ridge and fusion loop) and DIII (lateral ridge CC1015840loop and A strand) Moreover 16 of the neutralizing mAbswere tested in mice with most of them being protectivewhen given as prophylaxis Seven of these had post-exposuretherapeutic activity when administered as a single dose byintraperitoneal route even 3 days after intracranial infectionFor the mAbs with the greatest therapeutic potential protec-tion was confirmed with an Ab-enhanced vascular leakagemouse model of DENV2 infection [35]
Recently Deng et al described a cross-reactive murinemAb named 2A10G6 that is able to recognize DENV1ndash4 YFV WNV JEV and TBEV to potently cross-neutralizeDENV1ndash4 YFV and to a lesser extent WNV This mAbrecognizes a highly conserved motif (amino acid residues
98ndash101) located within the fusion loop of the FlavivirusDII of the E glycoprotein Moreover this mAb exerts itsneutralizing activity in a post-attachment step during thevirus entry process as demonstrated by kinetic neutralizationtests performed in vitro Additionally protection experimentsperformed in mouse models showed that treatment with100 120583gmL of 2A10G6 conferred full protection against lethalDENV2 challenge and 20 120583gmL and 4120583gmL of 2A10G6protected 89 and 40 of infected mice from lethal chal-lenge respectively For infection with DENV1 3 and 4 use of100 120583gmL of 2A10G6 conferred partial protection and 5377 and 73 of the infected mice survived after challengerespectively Finally the protection profile of 2A10G6 againstWNV showed that prophylactic administration with a singledose of 200mg of 2A10G6 conferred 80 protection in miceMost importantly 3 of 8 (375) mice survived when 2A10G6was administered one day after WNV challenge SimilarlyanothermousemAb 4G2 is able to recognize the fusion loopat the extremity of DII from all four DENV serotypes and toprevent syncytia formation [36]
Rajamanonmani et al described a mouse mAb 9F12raised against theDIII of DENV2E protein that is able to rec-ognize DENV1ndash4 serotypes as well asWNV and to neutralizefive DENV strains representative of all DENV serotypes [41]Similarly Cockburn et al reported a comparative high-resolution crystallographic analysis of an A-strand DIIImurinemAb 4E11 in complex with its target domain of the Eprotein from the four DENV serotypes MAb 4E11 is capableof recognizing and neutralizing all four serotypes with IC50values varying between 1 and 100 nMThe structures reportedalso highlight the mechanism by which anti-A-strand mAbsdisrupt the architecture of the mature virion inducing dimerdissociation premature fusion-loop exposure and concomi-tant particle inactivation [40] Also Midgley et al function-ally characterized another murine mAb 2H12 raised in miceagainstDIII of E protein Similarly to the previously describedmAbs 2H12 is able to bind all the four DENV serotypes ina epitope encompassing the conserved 314ETQH317 motifHowever the neutralizing potential of 2H12 is lower than anumber of other anti-DIII mAbs with IC50 values rangingfrom 056 to 145 nM for DENV1 2 and 4 On the other handit showed no ADE of infection activity [38]
Austin et al isolated a murine mAb named E111 whichrecognize a novel CC1015840-loop epitope on DIII of the E proteinfrom two different DENV1 genotypes Docking of the mAbstructure onto the available cryoelectron microscopy modelsof DENV virions revealed that the E111 epitope was inaccessi-ble suggesting that this mAb recognizes an uncharacterizedvirus conformation While the affinity of binding betweenE111 and DIII varied by genotype a limited correlation withneutralizing activity was observed These data support theconclusion that potent neutralization depends on genotype-dependent exposure of the CC1015840 loop epitope [49] In fact aspreviously described elsewhere binding of some E reactiveAbs depends on the dynamicmovement of proteinmoleculesldquobreathingrdquo in the virion particle leading to transient expo-sure of hidden epitopes For instance optimal binding ofmousemAb 1A1D-2 to EDIII requires incubation at 37∘C [75]
6 BioMed Research InternationalTa
ble1Schematicsummaryof
theb
estcharacterized
mAb
sagainstarbo
viralp
atho
gens
mAb
Orig
inRe
activ
ityTargetprotein
Epito
peClon
ingstr
ategy
Form
atisotype
Neutralizingactiv
ityRe
ference
48A
Hum
anDEN
V1ndash4
EDII
EBVtransfo
rmation
IgG
IC50
(120583gmL)
DEN
V115
DEN
V2gt40
DEN
V324
DEN
V4gt40
[37]
D11C
IC50
(120583gmL)
DEN
V115
DEN
V210
DEN
V3102
DEN
V416
16D
IC50
(120583gmL)
DEN
V115
DEN
V202
DEN
V305
DEN
V427
2H12
Murine
DEN
V1ndash4
EDIII(ABloop
aa
314ndash
317)
Hybrid
omafrom
BALB
cmou
seim
mun
ized
with
DEN
V2EDIII
IgG2b
IC50
nM
DEN
V1056ndash54
DEN
V329
DEN
V4145
[38]
C9Murinechim
eric
DEN
V2
EDIII
Phaged
isplayof
achim
ericmurine
hybridom
alibrary
VH1V1205811
PRNT5
0850120583
gmL
[39]
4E11
Murine
DEN
V1ndash4
EDIII(str
andA308
312andstrand
G387
389391)
Hybrid
oma
IgG2a120581
IC50
(120583gmL)
DEN
V1016
DEN
V2013
DEN
V-38
DEN
V-415
[40]
4G2
Murine
DEN
V1ndash4
EDII(fu
sionloop
)Hybrid
omafrom
DEN
V2Eim
mun
ized
mice
IgG
PRNT5
0(120583gmL)
DEN
V215
[39]
9F12
Murine
DEN
V1ndash4WNV
EDIII(aa
305307310
330Astr
andandBC
loop
)
Hybrid
omafrom
BALB
cmou
seim
mun
ized
with
DEN
V-2EDIII
IgG1k
DEN
V1ndash4P
RNT5
02sdot10minus8ndash2sdot10minus7M
[41]
2A10G6
Murine
DEN
V1ndash4YF
VWNV
JEV
TBEV
EDII(Fusionloop
aa
98ndash101)
Hybrid
omafrom
BALB
cmou
seim
mun
ized
with
inactiv
ated
DEN
V2
IgG1
PRNT5
0(120583gmL)
DEN
V12
DEN
V215
DEN
V321
DEN
V418
YNF36
WNV
46
[36]
mAb
11Hum
anDEN
V1ndash4WNV
SLEV
YFV
JEV
MVEV
EDII(fu
sionloop
)Ph
aged
isplayof
human
scFv
Fusio
nprotein
scFv-Fc
PRNT8
0(120583gmL)
WNV
125
DEN
V2625
[4243]
BioMed Research International 7Ta
ble1Con
tinued
mAb
Orig
inRe
activ
ityTargetprotein
Epito
peClon
ingstr
ategy
Form
atisotype
Neutralizingactiv
ityRe
ference
E16
(MGAW
N1)
Murine
WNV
EDIII(LR
aa3
02ndash309)
Hybrid
omafrom
immun
ized
micew
ithWNVE
IgG2bhum
anized
PRNT5
04ndash
18ng
PRNT9
053ndash297
ng[4445]
1A1D
-2Murine
DEN
V1ndash3
EDIII(Astr
andaa
307
310and312)
Hybrid
omafrom
immun
ized
micew
ithdifferent
pH-tr
eated
virus
IgG2a
DEN
V2PR
NT5
021n
M[4647]
1F4
Hum
an
DEN
V1
E(virion
)
DI-DII
Electro
fusio
nof
infected
mem
oryBcells
from
DEN
V-im
mun
esub
jects
with
EBV
IgG
DEN
V1IC5
0011120583gmL
[48]
2D22
DEN
V2
DIII
DEN
V2IC50
008120583gmL
5J7
DEN
V3
DI-DII
DEN
V3IC50
010120583gmL
E105
Murine
DEN
V1
E
DIII(BC
loop
G328
T329
andD330DE
loop
K361E
and
E362K
FGloop
K3
85)
Hybrid
omas
deriv
edfro
mC5
7BL6
IFN-120572120573Rminus
minusmice
infected
with
DEN
V1
IgG
PRNT5
0DEN
V-1
05ndash592ng
mL
[32]
E106
DIII(BC
loop
G328
T329
andD330DE
loop
K361E
and
E362K
FGloop
K3
85A
strand
S305
K307E
309K3
10and
E311)
PRNT5
0DEN
V-1
06ndash
592ng
mL
E111
DIII(CC1015840loop
)PR
NT5
038ndash25120583gmL
[49]
CR4374
Hum
anWNV
EDIII
Phaged
isplayof
scFv
IgGlib
rary
VH2-05V
L1e
PRNT5
0018120583gmL
PRNT9
0095120583gmL
[50]
CR4353
VH3-30V
k3-A27
PRNT5
00026120583
gmL
PRNT9
0364120583gmL
1A5
Chim
panzee
DEN
V1ndash4WNV
JEV
LGTV
EDII(aaG
106)
Phaged
isplayof
Fab
library
from
DEN
V1ndash4
infected
chim
panzees
Hum
anized
IgG1
PRNT5
0(120583gmL)
DEN
V1048
DEN
V2095
DEN
V332
DEN
V443
WNVD
ENV438
JEV
21LG
TV28
[51]
8 BioMed Research International
Table1Con
tinued
mAb
Orig
inRe
activ
ityTargetprotein
Epito
peClon
ingstr
ategy
Form
atisotype
Neutralizingactiv
ityRe
ference
mAb
11Hum
anDEN
V1ndash4WNV
EDII(fu
sionloop
W101G104G106)
scFv
library
scFv-Fc
PRNT8
0(120583gmL)
DEN
V2625
WNV
125
[4243]
3B4C
-4
Murine
SLEV
E
1a
Hybrid
oma
IgG
SLEV
PRNTlt17
[5253]
1B2C
-5SLEV
1bSLEV
PRNTlt17
6B5A
-2SLEV
1cSLEV
PRNT
48
4A4C
-4SLEV
1dSLEV
PRNT
29
1B5D
-1SLEV
JEV
2SLEV
PRNTlt17
2B5B
-3SL
EVJEV
MVEV
WNV
YFV
3SLEV
PRNT
23
2B6B
-2AllFlaviviru
s4a
SLEV
PRNTlt17
6B6C
-1AllFlaviviru
s4b
(inDII)
SLEV
PRNT
23
A3
Chim
panzee
JEV
E2
DIa
aK1
79Ph
aged
isplay
Hum
anized
PRNT5
0004ndash0
2nM
[54]
B2DIIaaI126
Phaged
isplay
PRNT5
0002ndash2
nM
E3DIIIa
aG132
Phaged
isplay
PRNT5
0014ndash0
93n
M
FabT
JE12B0
2Hum
anJEV
ENA
Phaged
isplay
Fab
FRNT5
0502120583gmL
[55]
5F10
Hum
anCH
IKV
E2Dom
ainB
EBVtransfo
rmation
IgG11205822
IC50lt100n
gmL
[56]
8B10
Hum
anCH
IKV
E1-E2
E2Dom
ainA
EBVtransfo
rmation
IgG1k
IC50lt100n
gmL
CHK-
152
Murine
CHIK
VE2
aa59
Hybrid
oma
IgG2c
IC50
1ndash3n
gmL
[57]
11E7
Murine
CCHFV
Gn
C-ter
Hybrid
oma
IgG
PRNT8
0diluted
12560
[58]
4-39-C
CMurine
RVFV
G2
Dom
ainIV
Hybrid
oma
IgG
PRNT8
0diluted
1204
80ndash181920
[59]
2C9
Murine
YFV
EHybrid
oma
IgG2a
PRNT9
011
0417D
1105
2Asib
i[60]
5A 7A R3(27)
Hum
anYF
VE
DI-DII
Phaged
isplay
ScFv
PRNT9
001ndash03120583
gmL
[61]
13D6
Murine
TBEV
EDIII
Hybrid
oma
IgGchimeric
PRNT5
019120583gmL
FRNT5
045120583
gmL
IC50
19ndash167120583
gmL
[62]
3B4C
-4(H
y4-26C
)Murine
VEE
VE2
aa182ndash209
Hybrid
oma
IgGhum
anized
PRNT7
0(120583gmL)
394ndash125
[63]
BioMed Research International 9
The structure of the 1A1D-2 bound to EDIII indicates that themAb binds to sites that are transiently exposed during viralldquobreathingrdquo at 37∘C and block infection during attachmentof the virion to the cell In particular this mAb neutralizesDENV1 2 and 3 serotypes and residues K307 and K310 arethemost critical residues for binding of 1A1D-2mAb [46 75]
Finally Shrestha et al immunized mice with a genotype2 strain of DENV1 virus and generated 79 new mAbs 16of which strongly inhibited infection by the homologousvirus and localized to DIII of E protein Surprisingly onlytwo mAbs E105 and E106 retained strong binding andneutralizing activity against all five DENV1 genotypes as wellas being protective in immunocompromised infected miceMoreover E105 and E106 exhibited therapeutic activity evenwhen administered as a single dose four days after inoculationwith a heterologous genotype 4 strain of DENV1 in the samemouse model [32]
243 Other Non-HumanmAbs Other non-human and non-murine as well as humanized mAbs directed against the Eglycoprotein have been described In this regard Goncalvezet al isolated a large panel of anti-E Fab fragments from chim-panzees infected with all four DENV serotypes Howeveronly a limited number of them displayed a cross-neutralizingactivity against DENV1 and 2 and to a lesser extent againstDENV3 and 4 In particular the authors calculated thatamong them the 1A5 mAb that was further humanizedneutralized DENV1ndash4 at a PRNT50 titer of 048 095 32and 43 120583gmL respectively Interestingly the humanized 1A5was also tested for binding and neutralization against theWNVDENV4 chimera JEV strain SA14-14-2 and Langatvirus (LGTV) strain TP 21 giving a PRNT50 titer of 38 21and 28120583gmL respectively Moreover the authors calculatedthat when administered in a dose of 2mg per kg of bodyweight 1A5 would give a serum titer of approximately 40and 20 50 reduction in plaque reduction neutralization test(PRNT50) against DENV1 and DENV2 respectively [76]Furthermore epitope mapping of 1A5 mAb localized to G106within the Flavivirus-conserved fusion loop inDII of DENV2E protein [51]
244 Anti-DENV Human mAbs As anticipated attemptsto isolate neutralizing human mAbs have been morechallenging due to the restricted elicited Ab-repertoire whichrecognizes DIII of E glycoprotein in naıve as well as ininfected patients [77 78] However several groups reportedthe isolation of neutralizing mAbs from infected subjects Inthis regard Setthapramote et al isolated a total of 136 humanhybridoma clones producing specific mAbs against DENVobtained using PBMCs from nine blood samples from fouracute-phase patients secondarily infected with DENV2 andfive convalescent-phase patients Interestingly the authorsfound that most of the acute-phase mAb clones were cross-reactive with all four DENV serotypes with most of themrecognizing the E protein and endowing of neutralizingactivity against all DENV serotypes compared to thosederived from convalescent-phase patients In particularfrom the acute-phase PBMCs 818 were anti-E 66 wereanti-prM and 33 were anti-NS1 while 133 anti-E 133
anti-prM and 533 anti-NS1 mAbs clones were obtainedfrom convalescent-phase PBMCs [79] Previous studiesconfirmed the data obtained by Setthapramote et al onconvalescent-phase patients but this group firstly reportedthe efficient preparation of human mAbs with strongneutralizing activity titers against all four DENV serotypesusing PBMCs from acute-phase patients secondarily infectedwith DENV [80 81]
The group of de Alwis through immunoglobulin deple-tion studies reported that a substantial fraction of DENV-reactive Abs in human immune sera including type-specificneutralizing Abs bound to the intact virion but not torecombinant E proteinThe authors confirmed these observa-tions also isolating human neutralizing mAbs and proposedthat humans produce Abs that neutralize DENV infectionby binding a complex quaternary structure epitope that isexpressed only when E proteins are assembled on a virusparticle [48] Similar findings have been described for severalother viruses including WNV and HIV [82 83] Mappingstudies indicate that this epitope has a footprint that spansadjacent E protein dimers and includes residues at the hingebetween domains I and II of E protein [48]
Recently Costin et al isolated three broadly neutralizinganti-DENVhumanmAbs named 48AD11C and 16DThesemAbswere isolated from three different convalescent patientswith distinct histories of DENV infection All three mAbsrecognized the E glycoprotein with high affinity neutralizedall four serotypes of DENV but mediated ADE of infection inFc receptor-bearing cells at subneutralizing concentrationsMapping studies revealed that all three mAbs bind discontin-uous epitopes within the highly conserved fusion-loop regionof DII (contacting residues W101 L107 andor G109) [37]
25 Yellow Fever Virus (YFV) TheWHO estimates that thereare 200000 cases of yellow fever virus (YFV) infections and30000 related deaths every year especially in Sub-SaharanAfrica In fact different to themajority of human arbovirusesand similarly to DENV and CHIKV YFV has expanded itshost range to include humans as an amplifying host [1]Symptoms occurring from 3 to 6 days after infection withYFV consist mostly in fever chills anorexia lumbosacralpain nausea and vomiting This syndrome lasts about 3days in most cases and is sometimes followed by a one-daylasting period of remission Fever will reoccur together withdark hematemesismelena petechiae and other hemorrhagicsymptoms Convalescence is characterized by deep asthenialasting up to two weeks Rarely prognosis can be fatal Aneffective live-attenuated vaccine (17D) derived from the Asibistrain by serial passages in chicken embryos is available [84]
251 mAbs against YFV In 1983 Schlesinger et al produceda battery of mAbs after immunization of BALBc mice byinjection of 17D YFV vaccine strain [84] Among them thespecificity towards the E and NS1 proteins was describedeven though anti-NS1 Abs were not able to show neutralizingactivity The 13 IgG and 1 IgM anti-E protein mAbs wereclassified in five groups according to their specificity GroupA consisting of the only IgM produced in this experiment
10 BioMed Research International
could neutralize 17D YFV virus only group B 17D and Asibistrains only group C Asibi strain only group D could onlyneutralize Asibi but not 17D strain showing also reactivityagainst DENV2 Zika or Banzi viruses All of the four mAbsbelonging to this group were able to cross-react with Zikaandor Banzi viruses and two (4E11 and 5H3) neutralizedDENV2 virus with 4E11 neutralizing also Banzi virus The3E9 mAb the only component of the group E could notneutralize 17D nor Asibi strains
All of the IgG mAbs resulted able to protect bothprophylactically and therapeutically BALBc or CD-1 micefrom lethal intracerebral challenge with 17D-204 strain [85]The chimeric form of the group B mAb 2C9 IgG (2C9-cIgG) was able to provide AG129 mice a 72 survival whenadministered 24 hours before infection with 17D-204 strainIts murine form could provide the survival of the 95 ofthe mice Appreciable results were also obtained when bothmurine 2C9 and 2C9-cIgG were administered 24 hours post-infection with the survival of 70 and 20 of mice respec-tively Viral presence was not detected in survivingmice [60]
In 2005 Daffis and colleagues constructed two Ab-phage libraries by cloning the repertoire of YFV-infectedpatients Panning was then performed with YFV-17D virionsThe scFv-5A 7A and R3(27) showed a neutralizing activityspanning from50 to 100 inPRNTassays against bothYFV17D-204-WHO and Asibi strains In further tests reactivitywas observed against wild-type YFV strains of West Africagenotype I and II (Nigeria 1987 and Asibi strains resp)and EastCentral Africa (CAR 1986 Ethiopia 1961 strains) Aconcentration from 01 to 03120583gmL could yield a 90 plaquereduction Reactivity was also observed against the strainSenegal 1990 in a lesser extent Production of escape variantscould demonstrate that scFv-5A 7A and R3(27) epitopes arebuilt up by residues extensively separated in themonomers ofE glycoprotein that however result in closed proximity whenthe homodimeric form of E is constituted [61]
26 West Nile Virus (WNV) The West Nile virus (WNV) isan epidemic neurotropic virus estimated to be responsibleof about 36000 cases and 1500 deaths registered in theUnited States between 1992 and 2012 [86] WNV antigenicityallows its classification into the Japanese encephalitis virus(JEV) serocomplex Genomic analysis has revealed two maingenetic lineages of WNV lineage I viruses circulating in theUSA Europe the Middle East Africa India and Australiaand lineage II viruses isolated from Sub-Saharan Africa andMadagascar [1]
WNV was firstly identified in 1937 and is endemicin many countries of Africa Middle East and West AsiaHowever after 1990 frequent outbreaks of WNV infectionswere reported in Romania Israel and later in North Americaand across the USA in 1999 and recently in 2012 [87]
Most of WNV infected individuals do not develop symp-toms while about 20 develop a self-limiting illness calledWest Nile fever Acute symptoms include fever tirednessand swollen lymph glands but in a minority of cases alsoencephalitis with long-term deficits in cognitive functionand motor skills have been reported However in less than
1 of infected patients WNV having crossed the blood-brain barrier is responsible of neuroinvasive and potentiallylethal form of the disease In these cases degeneration andapoptosis upon infection of neurons and the consequentinflammatory response can occur Related symptoms includehigh fever coma muscle weakness and paralysis Immun-odepression and advanced age have been correlated with anaugment risk to develop a severe disease [87] There are nospecific treatment options or licensed vaccines for humans[88]
WNV is transmitted by infected mosquitos and initialreplication is thought to occur primarily in dendritic cells inthe skin which migrate to secondary lymphoid tissues wherethe replicating virus enters the circulation To date onlyefficacious WNV veterinarian vaccines have been licensedwhile there are no licensed vaccines for protection againstWNV in humans [88]
261 mAbs against WNV Broad-spectrum antivirals suchas type I IFN-120572 ribavirin mycophenolic acid in WNVinfection showed ineffective results in vivo despite in vitrothey showed some activity [89]
However experiments in murine models extrapolationof clinical data as well as passive administration of pooledimmune-120574-globulins (OmriGam) containing a significanttiter of neutralizing Abs before and after infection showedan important role of the humoral immune response incontrolling viremia and prevent viral dissemination Indeedthe development of a neutralizing mAb-based therapy seemsto be encouraging for a possible treatment of infected patients[90ndash92]
Similarly to what has been observed for DENV only anti-E Abs have been identified as neutralizing and protectiveAlso anti-NS1 Abs have shown a protective role howevertheir mechanism of protection has not yet been elucidated asthe NS1 protein is secreted from infected cells and not presenton the virions [93ndash95]
In this regard similarly to what has been describedfor DENV studies performed using a naıve human scFvlibrary for panning with purified WNV E protein supportthe hypothesis that no Abs against the neutralizing DIII canbe isolated However DIII specific Abs were isolated in asubsequent study using immunoglobulin libraries obtainedfrom threeWNV infected patients for biopanning on purifiedinactivated virus virus-like particles consisting of prM andE proteins or recombinant E glycoprotein Although theproportion of DIII-specific mAbs was low (8) comparedwith anti-DII mAbs (47) Two out of the four anti-DIIImAbs were potently neutralizing and protective in vivowhereas only three out of the 24 anti-DII were weaklyneutralizing in vitro and non-protective in vivo In particularresidues that are critical for neutralization lies on the regionsspanning amino acids 305ndash312 330ndash333 and 365 that arelocated on adjacent exposed loops ofDIIIHowever sequencealignment of E protein of different flaviviruses such as DENVand JEV revealed a considerable variation compared tothe whole E protein This observation further suggests thatdifferently from broadly cross-reactive anti-DII Abs anti-DIII neutralizing Abs are virus-type specific However of
BioMed Research International 11
these mAbs two (CR4374 and CR4353) protected mice fromlethal WNV challenge at 50 protective doses of 129 and357 120583gkg of body weight respectively [50]
Sanchez et al using three different immunization strate-gies (ie inactivated virus naked DNA and recombinantprotein) isolated ninemurinemAbsmost of which bound toconformation-dependent epitopes in DIII of the E protein Inparticular neutralizing mAbs named 8B10 11C2 10C5 and17C8 were obtained from mice immunized with inactivatedvirus alone or in combination with a DNA plasmid andbound to the same region of DIII with high affinity Incontrast mAbs obtained by immunization with a solubleversion of the E glycoprotein did not exhibit neutralizingactivity These non-neutralizing mAbs were cross-reactivewith several other flaviviruses including SLEV JEV YFVand Powassan virus confirming the conserved nature ofFlavivirus non-neutralizing epitopes [96]
Gould et al isolated 11 unique human single-chain vari-able region Ab fragments (scFvs) that bind the E protein ofWNVAmong them a humanmAb namedmAb11 expressedas a scFv-Fc fusion protein was further characterized Itrecognizes the fusion loop at the distal end of DII ofthe WNV E protein and cross-reacts with all four DENVserotypes and provides protection against DENV2 and 4as well as WNV [42] Moreover therapeutic studies of thismAb in WNV-infection model mice provided substantialprotection when administered after 5 days post-infectionInterestingly a neutralization escape variant of this mAbfailed to cause lethal encephalitis (at higher infectious doses)or induce the inflammatory responses associated with blood-brain barrier permeability in mice compared to the parentalWNV suggesting an important role for the fusion loop inviral pathogenesis [43]
Oliphant et al isolated an anti-DIII E protein mAbnamed E16 fromhybridomas obtained after immunization ofmice with recombinant WNV E protein which neutralizedall WNV strains with PRNT50 values of 4 to 18 ng andPRNT90 values of 53 to 297 ng One hundred microgramsof mAb protected greater than 90 of mice from lethalinfection and even a single 4120583g treatment of E16 on day 2 afterinfection prevented mortality Moreover humanization ofthis mAb confirmed as therapeutically effective in mice [44]Subsequent studies revealed that mAb E16 neutralization ismediated by engagement of four discontinuous segmentsof DIII including the amino-terminal region (amino acidresidues 302ndash309) and the three connecting loops BC (aminoacid residues 330ndash333) DE (amino acid residues 365ndash368)and FG (amino acid residues 389ndash391) Moreover no ADE ofinfection was detected when E16 mAb was used at saturatingconcentrations [45 47]
Furthermore results of a Phase I safety study of thehumanized E16 mAb (designated MGAWN1) have beenreported and suggested that doses of up to 30mgkgwerewelltolerated with few mild adverse events and would providean excess of virus neutralizing activity for 3-4 weeks aftertreatment However a case of anti-MGAWN1 Ab elicitationoccurred with the consequent increased rate of clearance andindeed impacting efficacy A Phase II safety and efficacy studyof MGAWN1 is ongoing [97] Furthermore the possibility of
preventing or treating WNV-induced memory deficits wasrecently investigated In this study hamsters were treatedintraperitoneally with 32mgkg ofMGAWN1mAb at 45 daysafter subcutaneously challenging with WNV InterestinglyMGAWN1 prevented mortality weight loss and improvedfood consumption of WNV-infected hamsters compared tocontrols [98]
Recently Lelli et al isolated six anti-E mAbs frominactivated-WNV immunized mice In particular three ofthem (3B2 3D6 and 4D3) neutralized lineage I and II WNVwith the first two recognizing the same epitopes located onthe distal lateral surface of DIII (critical amino acid residueK307) Conversely 4D3mAb recognized a novel neutralizingepitope on DII (critical residues S276 and T278) Indeedfurther protective and therapeutic studies are needed toascertain their neutralizing activity in vivo [99]
Finally to conclude like DENV infection Abs directedagainst the M protein are not protective and neutralizingMoreover in humans a skewed humoral immune responseagainst DII has been frequently observed and confirmedalso with the hybridoma technology [50] The isolation andelicitation of neutralizingAbs directed against the fusion loopand DIII of the E protein represent thus the most challengingand promising goal for the development of new effectivetherapeutic inhibitors and immunogens respectively
27 Tick-Borne Encephalitis Virus (TBEV) Tick-borne ence-phalitis virus (TBEV) is one of the most dangerous agentscausing human neuroinfections occurring mostly in Europeand Asia and with a potential fatal prognosis [100] TBEV isbelieved to cause 3500ndash10000 human cases of encephalitisin Europe per year with a high morbidity in Russia CzechRepublic Austria and Germany In particular between 1990and 2007 an average of 8755 cases of TBE was reported peryear in Europe and Russia Despite the fact that Russia isthe country with most infections registered annually CzechRepublic incidence is among the highest in Europe with 400ndash1000 clinical cases reported every year [101]
Three subtypes of TBEV are classified namely EuropeanSiberian and Far-Eastern sharing most of the genetic andantigenic features [102] In fact a high degree of antigenichomogeneity between different strains of TBEV has beendescribed [103]
Clinical manifestations vary among the subtypes butusually they start with a short febrile period of 7ndash14 daysafter the tick bite Fatigue headache and pain in the neckshoulders and lower back together with high fever andvomitingmay be present [100]Thesemanifestations are oftenfollowed by an asymptomatic phase lasting from 2 to 10days after remission from the fever with possible progres-sion to neurological disease Neurological symptoms includemeningitis encephalitis myelitis and radiculitis Mortalityoccurs in 1-2 of the European subtype-infected patientsbut fatal prognosis can occur in up to 20ndash40 of the Far-Eastern subtype-infected patients [104] Mortality rates ofthe Siberian subtype are similar to those observed for theEuropean [105]
Effective and safe vaccines against TBEV produced frominactivated virus have been developed and licensed for their
12 BioMed Research International
use in humans However an emergency therapy in theabsence of a mass immunization is needed as no effectivetreatments are yet available [106]
271 mAbs against TBEV Levanov et al described the chim-erization of two murine mAbs (13D6 and 10C2) directedagainst the DIII and DII respectively of the TBEV E glyco-protein The chimeric mAbs present binding characteristicssimilar to the parentalmAbsMoreover as the parentalmAbsonly the chimeric mAb 13D6 was able to neutralize TBEVinfectivity in vitro In particular neutralization studies withthe murine chimeric and scFv forms of mAb 13D6 were per-formed In particular murine 13D6 showed an IC50 titer of115 120583gmL in Focus Reduction Neutralization Tests (FRNT)against TBEV strain 205 and of 29 120583gmL in PRNT againstTBEV strain Softjin Chimeric 13D6 showed an IC50 titer of45 120583gmL in FRNT against TBEV strain 205 and of 19 120583gmLin PRNT against TBEV strain Softjin ScFv 13D6 showed anIC50 titer of 167 120583gmL in FRNT against TBEV strain 205and of 112 120583gmL in PRNT against TBEV strain Softjin [62]
28 Japanese Encephalitis Virus (JEV) Japanese encephalitisvirus (JEV) is a mosquito-borne flavivirus that over thepast few decades has caused several outbreaks throughoutChina Southeast Asia Australia and Papua New Guineawith a prevalence recently estimated to be of about 70000casesyear [107ndash109] A 40 mortality was recorded in someof the JEV-affected areas Moreover many survivors facesome neurological problems and complications [108] Since1995 the disease has also emerged in Non-Asian regions suchas Northern Australia [110 111] The situation in SoutheastAsia however is further complicated by the overlappingepidemics of JEV and DENV as well as sporadic casesof WNV infections detected in some of the affected areasparticularly in India [112]
Effective vaccines both live-attenuated and inactivatedstrains of JEV have been developed and licensed in majorcountries However as for the other arthropod-borne Fla-vivirus members no specific antiviral drugs are currentlyavailable [4]
281mAbs against JEV Aspreviously described for the otherFlavivirusmembers many groups isolated and characterizedanti-JEVmAbs showing different neutralizing and protectiveproperties In this regard Gupta et al used combinations ofanti-E JEV mAbs (Hs1-4) in mice protection experimentsIn particular they found that the singularly mAb protectionranged from 45 to 65 when 100120583g of mAb were adminis-tered while equimolar combinations of two or three mAbsgave 85ndash90 or 100 protection respectively [113] In similarexperiments Lee et al demonstrated that a Flavivirus anti-NS1 mAb named 16NS1 cross-reacted with JEV as well asWNV and exhibited protective activity against WNV as wellas a lethal JEV infection However no neutralizing activitywas observed using this mAb against both WNV and JEV inin vitro experiments suggesting the participation of otherAb-mediatedmechanisms in vivo In particular 95ofmicewere
protected when 500120583g of mAb were administered intraperi-toneally and concomitantly to intramuscular injection of JEV
Overlapping peptide mapping analysis combined withsite-specific mutations identified the 116KAWGKSILFA125region and critical amino acid residues W118 and I122 as16NS1 mAb epitope highly conserved in WNV and JEVstrains [114]
Arakawa et al isolated from a combinatorial human Fablibrary constructed from peripheral blood lymphocytesobtained from JEV hyperimmune volunteers Among 188randomly selected clones FabTJE12B02 showed the best 50focus reduction endpoint at the concentration of 502 120583gmLagainst the JEV strain Nakayama [55]
Goncalvez et al isolated three mAbs named Fabs A3B2 and E3 by repertoire cloning from chimpanzees initiallyimmunized with inactivated JE-VAX and then boosted withattenuated JEV SA14-14-2 In particular these mAbs reactedwith epitopes in three different E domains in DI (amino acidresidue K179) in DII (I126) and in DIII (G132) for Fabs A3B2 and E3 respectively Moreover these Fabs as well as thederived humanized counterpart mAbs exhibited high neu-tralizing activities against a broad spectrum of JEV genotypestrains Moreover these mAbs exhibited a 50 protectivedose of 084 120583g (B2) 58120583g (A3) and 247120583g (E3) in mousemodels Finally administration of 200 120583gmouse of mAb B2one day after otherwise lethal JEV infection protected 50of mice and significantly prolonged the average survival timecompared to that of mice in the unprotected group [54]
29 St Louis Encephalitis Virus (SLEV) St Louis encephalitisvirus (SLEV) was first discovered as the mosquito-borneagent responsible for over 1000 cases of encephalitis duringa 1933 summer outbreak in St Louis (Missouri) and nowis a reemerging human pathogen widely distributed in theAmerican continent causing several human encephalitisoutbreaks over the last 80 years Additional epidemics haveindeed occurred from 1964 to 2006 in the Americas rangingfrom the US to Argentina and Brazil As a member of theFlavivirus genus the SLEVE glycoprotein ectodomain is 68identical to serocomplex-related JEVEbut only 46 and 40identical to those ofDENV2 andTBEV viruses fromdifferentserocomplexes [115]
291 mAbs against SLEV In 1983 Roehrig et al isolatedtwenty-one hybridomas producing murine mAbs specificfor the E glycoprotein of SLEV strain MSI-7 Serologicreactivities were initially determined by cross-reactivity indi-rect immunofluorescence assays using 22 strains of SLEVand 8 other related flaviviruses Four groups demonstratingtype- subcomplex- supercomplex- and group-specific reac-tivity patterns were identified Analysis of hemagglutination-inhibition and virus neutralization subdivided the cross-reactivity groups into eight epitopes (E-1a b c d E-2 E-3 and E-4a b) one of them following localized in DII (4b)[52 53]
Moreover among the previously described isolated anti-WNV mAbs by Gould et al the chimeric scFv-Fc 79 effec-tively neutralized also SLEV resulting in gt80 of PRNT80when used at 5 120583gmL [42]
BioMed Research International 13
3 Alphaviruses
Major human arboviruses in the Togaviridae family Chikun-gunya (CHIKV) and Venezuelan equine encephalomyelitis(VEEV) viruses belong to the Alphavirus genus which iscomposed of viruses with an icosahedral nucleocapsid sur-rounded by a lipid envelope and glycoprotein spikes Thestructural proteins of alphaviruses arise through co- andpost-translational processing of a polyprotein encoded by asingle positive-stranded RNAproducing the capsid (C) PE26 K and E1 The PE2 glycoprotein is a precursor containingthe E3 glycoprotein fused to the amino terminus of the E2envelope glycoprotein The PE2 glycoprotein is followed by6K a small membrane-associated protein and E1 the secondpolypeptide component of glycoprotein spikes
Trimerized heterodimers of the E1 and E2 viral glyco-proteins form the surface spikes and contain determinantsof viral tropism and virulence The E3 glycoprotein acts as asignal for transport of PE2 across themembranes of the roughendoplasmic reticulum and may promote the formation andintracellular transport of E1-PE2 heterodimers to the cellsurface During transport to the cell surface PE2 undergoesa maturational cleavage event by a furin-like protease toproduce E2 and E3 [116]
The E2 glycoprotein promotes specificity of virus bindingto the host cell surface and is a target of the humoral immuneresponse The E1 glycoprotein mediates fusion of the virionenvelope with the membranes of acidified endosomes allow-ing release of the nucleocapsid into the cytoplasm and theonset of viral replicationAlphavirus E1 shares no appreciablesequence identity to Flavivirus E protein Despite differencesin their amino acid sequences and arrangements on the viralparticle the structures of E and E1 are remarkably similarIndeed the conservation of three domains has been welldocumented in crystal structures of flavivirus E ectodomainsfrom TBEV DENV JEV and WNV as well as Alphavirus E1from Semliki Forest virus (SFV) and CHIKV Abs to the E1glycoprotein do not typically neutralize virus infectivity invitro but can protect against lethal challenge in animals
By analogy with the prM protein of dengue Flavivirusfurin cleavage of PE2 may begin after transit of an acidiclate component of the Golgi body where E3 is thought tosuppress the acid pH-triggered activation of glycoprotein E1fusion capability Protective anti-E3 mouse mAbs have alsobeen described [117]
31 Chikungunya Virus (CHIKV) The chikungunya virus(CHIKV) belongs to the Semliki Forest clade and was firstlyisolated in 1953 in Tanzania during an epidemic outbreakfollowing occurred also in Asia and Africa In the last fivedecades several CHIKV outbreaks have been described bothin Africa and Asia separated by gaps lasting from two totwenty years In 2005-2006 about 300000 cases out of785000 inhabitants were reported in La Reunion island witha fatal prognosis for 237 of the patients [117] Neither Europenor the Americas have had outbreaks of CHIKV so far exceptfor imported isolated cases Three genotypes of CHIKV aredescribed Asian EastCentralSouth African (ECSA) andWest African with an amino acid identity spanning from
952 to 998 Recent epidemics in Africa and Indiansubcontinent were caused by strains belonging to the ECSAgenotype Transmitted by Aedes mosquitoes CHIKV ismaintained in the human population by human-mosquito-human transmission [1] The disease is characterized bydengue-like symptoms such as chills high fever headacheand persistent myalgia and a further incapacitating arthralgia(from which the name chikungunya) which affects 40 ofinfected subjects However prognosis is rarely fatal [116]
There is currently no commercial vaccine and antiviraltreatment forCHIKV although some candidate vaccines havebeen tested in humans In this regard in 2000 US Armyperformed a Phase II clinical trial testing a live-attenuatedCHIKV vaccine (TSI-GSD-218) derived from a 1962 strain(15561) of an outbreak in Thailand Out of 58 every patientdeveloped neutralizingAbs and 5 lamentedmild tomoderatejoint pain [117] A phase III trial of this candidate vaccineis ongoing Furthermore a new formulation using virus-likeparticles was able to induce neutralizing Abs in macaquesagainst different CHIKV strains [117 118] Indeed it has beendescribed that infection seems to elicit long-lasting protectiveimmunity and cross-protection among CHIKV and otheralphaviruses
CHIKV entry into host cell is demonstrated to be medi-ated by envelope glycoproteins E1 and E2 which allowvirus fusion to cell membrane in low pH conditions andrecognition of an unknown cellular receptor respectively[116 119ndash121] Even though the genome replication relies onan error-prone RNA-dependent RNA-polymerasis CHIKVstrains S27 and 05115 Reunion showed in E1 and E2 alow amino acid variation of 068 and 33 respectivelyand such a low variability may be needed for an effectivereplication in two phylogenetically distant hosts [116 122]Recently the crystal structure of E1 and E2 heterodimerhas been resolved both at low pH and natural conditionsshedding light on E1 and E2 ectodomainsrsquo structure Inparticular similarly to flaviviruses E protein E1 ectodomainis made of the N-terminal DI the DII containing the fusionloop and the DIII at the C-terminal On the other hand E2ectodomain comprises theN-terminal domainA the domainB supposed to interact with the host cellrsquos unknown receptorand domain C at the C-terminal E1 DIII and E2 domain Care located close to the viral membrane [123 124]
311 mAbs against CHIKV Warter et al described 5F10 and8B10 two human mAbs which strongly neutralized severalCHIKV isolates in vitro without cross-reactivity againstother alphaviruses but to Onyong-nyong virus [56] Mixedpreparation of 5F10 and 8F10 did not show neither synergisticnor addictive effect in vitro and studies upon escape mutantsdemonstrated that 5F10 mAb binds at the tip of the E2domain B while 8B10 recognizes residues close to E1 fusionloop and amino acids within E2 domain A which form atransitional epitope under low pH conditions The authorsspeculated that 5F10 and 8B10 may inhibit CHIKV entry andfusion to the cell membrane respectively It is worth notingthat the previously mentioned escape mutants displayedmutations associated with reduced viral fitness in vitro evenafter 13 neutralizationamplification rounds In the same
14 BioMed Research International
study cell-to-cell transmission was firstly demonstrated as anescapemechanism enhanced by the E2mutation R82G [122]
In further in vivo studies 5F10 and 8B10 significantlydelayed CHIKV-caused death of AGR129 mice both in pro-phylactic and therapeutic tests Interestingly in therapeutictreatment these mAbs showed a synergistic effect whenadministered in combination with the total amount of mAbsinjected being the same of the single-mAbs Possibly thesetreatment did not allow themice to survive CHIKV challengedue to Ig clearing and 6ndash10 days half-life usually described forhuman mAbs [125]
Recently Pal et al cloned thirty-six murine mAbs ableto neutralize the ECSA La Reunion 2006 OPY-1 strain ofCHIKV (CHIKV-LR) the majority of which also neutralizeinfection of other strains corresponding to the Asian andWest African genotypes In particularmAbCHK-152 showedthe highest and broadest neutralizing activity with FRNTvalues indicating an IC50 value of 1 to 3 ngmL dependingon the viral strain used Among the thirty-six mAbs four(CHK-102 CHK-152 and CHK-166 CHK-263) could pre-vent lethality of immunodeficient Ifnarminusminus C57BL6 micewhen administered one day before exposition with CHK-152 and CHK-263 showing the ability to protect mice at thelowest dose (10 120583g) Therapeutic studies were performed onthese mAbs administering a single dose of 100 120583g 24 hoursafter infection The highest activity was described for CHK-166 able to protect 63 of the mice Testing the combinedactivity of the mAbs in therapeutic studies administration ofCHK-166 plus CHK-152 in a dose of 250120583g each turned out tobe themost effective protecting 71 of the IfnarminusminusC57BL6mice 60 hours after infection [57]
32 Venezuelan Equine Encephalomyelitis Virus (VEEV)Venezuelan equine encephalomyelitis virus (VEEV) is main-tained in a natural transmission cycle between mosquitoesand small rodents The first documented outbreaks occurredin the 1930s and several epidemics have been reported so farin Latin-American countries such as Venezuela ColombiaPeru Ecuador Bolivia Honduras Mexico and Panama withhundreds of thousands of human cases being reported anda case-fatality rate up to 1 during the 1969 Ecuador out-break Clinical manifestations of VEE are indistinguishablefrom Dengue [126] Six serogroups (IndashVI) are currentlyrecognized within the VEEV complex VEEV caused humanand equine outbreaks in the Americas for nearly a centuryEquine epizootics have high mortality (38ndash83) leading toa high viremia followed by a lethal encephalitis and often tohuman epidemics involving thousands of cases and hundredsof deaths Indeed VEEV is infectious for humans by theairborne route and has been responsible for a number oflaboratory infections In humans the disease is usually self-limiting with a febrile illness with 1ndash4 of cases progressingto severe encephalitis It is a hazard to laboratory workershas been developed as a biological weapon and is a potentialbioterrorist agent [127] There are no antiviral drugs andvaccines licensed for the treatment of VEEV infection inhumans Indeed although experimental live-attenuated vac-cines have been developed (eg TC-83) with good levels ofprotection in equine and mice in humans they may fail to
give protection in the majority of cases [128] Consideringthe protective role exerted by humoral immune response inanimal models it could be concluded that treatment withspecific IgG may have a beneficial antiviral effect in humanairborne infections with pathogenic strains of VEEV
321 mAbs against VEEV Several works support that mAbsmay protect against airborne VEEV as well as after airborneexposure toVEEVAb reactivity to both surface glycoproteins(E1 and E2) is associated with neutralization of the virusin vitro and passive protection against virus challenge Aseries of anti-E1 and anti-E2 mouse and humanized mAbshave been described recently Among them Phillpotts et alexamined two anti-E2 murine mAbs named 1A4A-1 and1A3A-9 both having potent protective activity against subcu-taneous VEEV challenge in mice Both mAbs had a similarhalf-life (58 and 100 days) in mouse serum after a singleintraperitoneal dose suggesting that mAbs delivered by thisroute at or around the time of VEEV infection persists athigh levels in the blood as well as secretions in respiratorytransudation throughout the clinical course of the disease Inparticular both mAbs administered 24 h prior to airbornechallenges had a substantial protective effect (90ndash100)Treatment of mice 2 or 24 h after airborne infection with asingle intraperitoneal dose of 100 120583g of 1A3A-9 mAb led toapproximately 50 survival There was no beneficial effectwhen mAb treatment was delayed to 72 h Moreover therewas evidence of synergy in vitro in PRNT between 1A3A-9and 1A4A-1 as has been demonstrated for other virusesand mAb pairs However no synergy was found in mouseprotection when mAbs were delivered intraperitoneally as amixture in equal parts to animals challenged 24 h previouslywith VEEV [129]
In a subsequent study Phillpotts et al reported two otherprotective mAbs The 3B2A-9 mAb protected against allthe serogroup I strains while 1A3B-7 mAb protected wellagainst challenge with all of the viruses tested Both 3B2A-9 and 1A3B-7 protected against airborne exposure to theIAB serogroup virus strainTrinidad donkey (TrD)Howeverthere was no evidence of synergistic protection when thesemAbs were combined in equal proportions An intraperi-toneal dose of 10 120583g was sufficient to protect 50 of themice with either mAb [130] According to other data themechanism of protection did not appear to depend uponneutralization [131] Similar results were obtained by OrsquoBrienet al which described a non-neutralizing mAb (IgG2a)named CUF37-2a able to protect 50 of mice from asubcutaneous VEEV challenge when a dose of 915 120583g of mAbwas administered 24 h prior to challenge [132]
In a following study the 1A3B-7 mAb was humanized(and following reported as Hu1A3B-7) maintaining the samefeatures of its murine counterpart In particular evaluation ofin vitro studies indicated that Hu1A3B-7 retained both broadspecificity and neutralizing activity Furthermore in vivoexperiments showed that Hu1A3B-7 successfully protectedmice against lethal subcutaneous and aerosol challenges withVEEV strain TrD Moreover the effectiveness of the human-ization process was determined by assessing proliferationresponses in human T-cells to peptides derived from the
BioMed Research International 15
murine and humanized versions of the VH and VL domainsThis analysis showed that the number of human T-cellepitopes within the humanized Ab had been substantiallyreduced indicating that Hu1A3B-7 may have low immuno-genicity in vivo [133]
Similarly Hunt et al described a humanizedmurinemAbderived from the 3B4C-4 mAb recognizing the E2c epitope(amino acid residues 182ndash209) and following named Hy4-26C showing neutralizing activity similar to the murinemAb Moreover it was protective (70ndash100 of survival) at100 ng dose in mice animal model intraperitoneal inoculatedand at 500120583g for intranasal challenge (80) Therapeuticstudies in mice revealed that Hy4-26C was able to cure miceup to 24 h following infection at 10 120583g on the contrary themouse mAb when given 1 hour after virus challenge [63]
In another work Hunt et al isolated two neutralizinghumanized Fabs F5 and L1A7 employing a blocking strategy[134 135] The anti-E2 specific F5 IgG had a 70 PRNTendpoint of 10 ngmL equivalent to that described for themost effective neutralizing anti-VEEVE2mAbs In particularthe E1-specific hFab L1A7 had a PRNT endpoint of 3120583gmL300-fold lower than F5
Further studies revealed that F5 epitope is located inthe 115ndash199 amino acid region Moreover F5 IgG had potentability to protect mice from infection by either route whenadministered 24 h before exposure however mice treated24 h and 48 h after aerosol exposure developed central ner-vous system infections but exhibited no clinical signs ofdisease [135]
Hu et al reported that passive immunization with thehumanized chimericmousemAbhu1A4A1IgG1-2A inmice at50 120583g 24 h before or after virulent VEEV challenge providedcomplete protection indicating that hu1A4A1IgG1-2A haspotent prophylactic and therapeutic effects against VEEVinfection [136]
Anti-VEEV mAbs isolated from a non-human primategene library has also been reported In particular thehumanized chimeric scFv-Fc ToR67-3B4 recognized viableas well as formalin and 120573-propiolactone inactivated virusparticles It detected specifically the viral E1 envelope proteinof VEEV but did not react with reduced viral glycopro-tein preparations suggesting that recognition depends uponconformational epitopes The recombinant Ab was able todetect multiple VEEV subtypes and displayed only marginalcross-reactivity to otherAlphavirus species except for Easternequine encephalitis virus (EEEV) In addition the scFv-Fcfusion described here might be of therapeutic use since itsuccessfully inactivated VEEV in a murine disease modelIn particular when the recombinant Ab was administered 6hours after challenge 80 to 100 of mice survived lethalVEEV IAB or IE infection Forty to sixty percent of micesurvived when scFv-Fc ToR67-3B4 was applied 6 hours afterchallenge with VEEV subtypes II and former IIIA [137]
4 Bunyaviridae
TheBunyaviridae family contains human arboviruses belong-ing to the Orthobunyavirus Phlebovirus and Nairovirus
genera Bunyaviridae includes enveloped viruses with a frag-mented single-stranded RNA genome of negative polarityTheir tripartite genome consists of a small (S) a medium(M) and a large (L) fragment and the envelope glycoproteinsGn and Gc that are cleaved out of a polyprotein synthesizedby the M fragment These glycoproteins have been provedto mediate the formation of the virus particle to play a rolein the interaction with cell surface receptors to mediate theentry of the virus into cells and to serve as targets for themajority of neutralizing Abs described so far
Orthobunyavirus transmitted through mosquitoes ormidges vectors are divided in 18 serogroups based on cross-titrations in haemagglutination inhibition assays and neutral-ization assays and correlating with main vector preferencesHowever the most clinically important viruses belong onlyto two serogroups the California encephalitis and the Simbuserogroups
Crimean-Congo hemorrhagic fever virus (CCHFV) isconsidered the only one of clinical relevance within theNairovirus genus which uses tick as main vector and is divid-ed in seven serogroups while Toscana virus and Rift Valleyfever virus (RVFV) which are transmitted by sandflies andmosquitoes respectively belong to the Phlebovirus genus[138]
MAbs against theNairovirusCCHFV and the PhlebovirusRVFV have been described
41 Crimean-Congo Hemorrhagic Fever Virus (CCHFV)Crimean-Congo Hemorrhagic Fever Virus (CCHFV) dis-tribution covers the greatest geographic range of any tick-borne virus known as viral isolation andor disease hasbeen reported from more than 30 countries in SoutheasternEurope Africa Asia and Middle East [139 140] Namelyfrom 1953 to 2010 about 6000 human cases were reportedin Southeastern Europe CCHFV causes sporadic outbreakswith mortality rates ranging from 10 to 80 [141 142] Asignificant variation in the time of incubation has beendescribed [139] Prehemorrhagic symptoms include highfever chills headache photophobia and back and abdominalpains Among other symptoms neuropsychiatric changeshave been reported in some patients In severe cases 3ndash6 daysafter the onset of the disease hemorrhagic symptoms occurwith petechiae ecchymosis bleeding in the form of melenahematemesis epistaxis [139]
In comparisonwith the nucleocapsid proteins inCCHFVGn and Gc show a higher degree of antigenic variabilityprobably due to their exposition to the host immune systemhypothesis corroborated in the description by Hewson et alin 2004 of four M segment phylogenic groups namely M1M2 M3 and M4 [141 142]
411 mAbs against CCHFV Blackburn et al firstly describedin 1987 murine anti-CCHFV mAbs able to recognize nucle-ocapsid proteins but no neutralizing activity has beendescribed thus far [143]
On the other hand Bertolotti-Ciarlet et al produced apanel of murine mAbs recognizing conformational epitopes
16 BioMed Research International
within theGn andGc glycoproteins expressed by the CCHFVstrain IbAr10200 The effectiveness of the mAbs was studiedby performing PRNT80 and none of the anti-Gn mAbsshowed neutralizing activity although many of the anti-GcmAbs neutralized IbAr10200 strain in vitro with clones 8A15E3 12A9 6C2 and 9H3 showing activity at a gt5120 dilutionMAbs 11E7 and 30F7 exhibited neutralizing activity at 2560dilution A suckling mice protection test was thereforeperformed for these mAbs The previously mentionedanti-Gc Abs were capable to protect mice to an appreciabledegree when applied 24 hours before and in a weakermanner 24 hours after virus challenge Many of the anti-GnmAbs were able to confer significant protection to IbAr10200both 24 hours before and after virus administration Therelevant effectiveness of the anti-Gn mAbs 6B12 10E1113G8 and 10G4 suggests that these mAbs may possess someneutralizing activity due to Ab-based effector mechanisms(eg ADCC) [58]
In further studies the same group characterized thebroadly cross-reactive neutralizing activity of the previouslydescribed murine mAb 11E7 able to bind conformationalepitopes within the C-terminal region of the Gn glycoproteinof all CCHFVM groups [58 141]
42 Rift Valley Fever Virus (RVFV) Similarly to CCHFV RiftValley Fever Virus (RVFV) causes sporadic outbreaks Thelargest epidemic occurred in Egypt in 1977 with an estimated200000 infections 18000 patients manifesting symptomsand 600 fatal prognoses [144] Symptoms include hemor-rhagic episodes fever encephalitis and blindness [145]
The major target of the humoral immune response is theRVFV glycoproteins Gc (G1) and Gn (G2) ranging fromnucleotides 480ndash2090 and 2091ndash3614 respectively [144]
421 mAbs against RVFV Within G2 glycoprotein are des-cribed three neutralizing antigenic sites namely epitope Inucleotides 792ndash893 (amino acid residues 258ndash291) epitopeII nucleotides 1164ndash1196 (amino acid residues 382ndash392) andepitope IV nucleotides 858ndash917 (amino acid residues 280ndash299) [59 145]
Despite the high degree of variability observed inCCHFVglycoproteins RVFV G2 presented an unexpected high con-servation among 22 isolates [59] In the same study onlythe LUNYO and 900060 isolates were resistant to in vitroneutralization by murine mAb 4D4 (recognizing epitope II)All other isolates were neutralized with PRNT80 titers from10240 togt81920On the other hand only SNS isolate showedreduced sensitivity to 4ndash39-CC (epitope IV) All other isolateswere neutralized with PRNT80 titers from 20480 to gt81920Protection studies on mice would be of interest for thedevelopment of a passive immunization prophylaxis [59]
Protection studies from lethal South African RVFVAN1830 strain infection were successfully performed on abattery of anti-G1 and anti-G2 mAbs produced by Besselaarand Blackburn with the strongly neutralizing anti-G1 mAb3E5 and anti-G2mAb 9C4 being the most effective [146]The9C4 antigenic area maps within G2 epitope I and a broadlyneutralizing activity may be investigated
5 Conclusions
Arbovirus infections have acquired increasing interest giventhe augmented globalization and tourism movement all overthe world that could be at the basis of epidemic eventsFurthermore the same vectors can sometimes transmit sev-eral arboviruses concomitantly complicating the diagnosis aswell as the therapy For example confusing mixed epidemicshave occasionally been described such as YFV plus CHIKVDENVplus CHIKV ormore recentlyPlasmodium falciparummalaria plus DENV1 and CHIKV on Madagascarrsquos east coast[1]
In the last decade several efforts have been employed inthe development of effective immunogens and therapeuticsgiving the current absence of specific antiviral drugs as wellas effective vaccines for themost diffused arbovirus infections(ie DENV andWNV)
However at the moment mosquito control is the bestavailable method for preventing arbovirus infections Thusto control the emerging public health problem of arbovirusinfections new antiviral therapeutic strategies that providepotent and broadly cross-protective immunity (especially forflavivirus infections) are an urgent globally medical need
As described in this review the majority of highlyneutralizing mAbs are of murine origin and indeed theirutility in the treatment of these infections is limited In facttheir employment can elicit an immunogenic response (iehuman anti-mouse Abs HAMA) the therapeutic efficacycould be reduced by a relatively faster clearance in humans(compared to human Abs) potentially exacerbated by theHAMA response Finally murine Abs exhibit relatively weakeffector functions (eg ADCC) compared to human AbsIndeed chimerization humanization or better the isolationand characterization of fully human broadly neutralizingmAbs would be the best choice for their possible use in afuture mAb-based therapy against arbovirus infections [147]
Finally possible ADE of infection mechanisms should beevaluated before considering a mAb as a possible candidatetherapeutic in a post-exposure setting as well as in the devel-opment of new Ab-eliciting immunogens
Conflict of Interests
The authors declare no conflict of interests in particular withOmriGam and any other cited Ab-based preparation men-tioned in the paper
References
[1] N Cleton M Koopmans J Reimerink G J Godeke and CReusken ldquoCome fly with me review of clinically importantarboviruses for global travelersrdquo Journal of Clinical Virology vol55 no 3 pp 191ndash203 2012
[2] F X Heinz H Holzmann A Essl and M Kundi ldquoField effec-tiveness of vaccination against tick-borne encephalitisrdquoVaccinevol 25 no 43 pp 7559ndash7567 2007
[3] A D Barrett and D E Teuwen ldquoYellow fever vaccinemdashhowdoes it work and why do rare cases of serious adverse events
BioMed Research International 17
take placerdquo Current Opinion in Immunology vol 21 no 3 pp308ndash313 2009
[4] S B Halstead and S J Thomas ldquoNew vaccines for Japaneseencephalitisrdquo Current Infectious Disease Reports vol 12 no 3pp 174ndash180 2010
[5] R Burioni M Perotti N Mancini and M Clementi ldquoPer-spectives for the utilization of neutralizing human monoclonalantibodies as anti-HCV drugsrdquo Journal of Hepatology vol 49no 2 pp 299ndash300 2008
[6] R A Diotti G A Sautto L Solforosi NManciniM Clementiand R Burioni ldquoNeutralization activity and kinetics of twobroad-range human monoclonal IgG1 derived from recombi-nant Fab fragments and directed against Hepatitis C virus E2glycoproteinrdquo New Microbiologica vol 35 no 4 pp 475ndash4792012
[7] R Burioni F Canducci N Mancini et al ldquoMolecular cloningof the first humanmonoclona antibodies neutralizing with highpotency Swine-origin Influenza A pandemic virus (S-OIV)rdquoNew Microbiologica vol 32 no 4 pp 319ndash324 2009
[8] P K Chandra S Hazari B Poat et al ldquoIntracytoplasmic stableexpression of IgG1 antibody targeting NS3 helicase inhibitsreplication of highly efficient hepatitis C Virus 2a clonerdquoVirology Journal vol 7 article 118 2010
[9] N Clementi D de Marco N Mancini et al ldquoA human mono-clonal antibody with neutralizing activity against highly diver-gent influenza subtypesrdquo PLoS ONE vol 6 no 12 Article IDe28001 2011
[10] D de Marco N Clementi N Mancini et al ldquoA non-VH1-69 heterosubtypic neutralizing human monoclonal antibodyprotects mice against H1N1 and H5N1 virusesrdquo PLoS ONE vol7 no 4 Article ID e34415 2012
[11] L Solforosi N Mancini F Canducci et al ldquoA phage displayvector optimized for the generation of human antibody com-binatorial libraries and the molecular cloning of monoclonalantibody fragmentsrdquoNewMicrobiologica vol 35 no 3 pp 289ndash294 2012
[12] E V Koonin and V V Dolja ldquoEvolution and taxonomy ofpositive-strand RNA viruses implications of comparative anal-ysis of amino acid sequencesrdquo Critical Reviews in Biochemistryand Molecular Biology vol 28 no 5 pp 375ndash430 1993
[13] J S Mackenzie D J Gubler and L R Petersen ldquoEmergingflaviviruses the spread and resurgence of Japanese encephalitisWest Nile and dengue virusesrdquo Nature Medicine vol 10 no 12pp S98ndashS109 2004
[14] T J SmithW E Brandt J L Swanson J MMcCown and E LBuescher ldquoPhysical and biological properties of dengue-2 virusand associated antigensrdquo Journal of Virology vol 5 no 4 pp524ndash532 1970
[15] J M Smit B Moesker I Rodenhuis-Zybert and J WilschutldquoFlavivirus cell entry and membrane fusionrdquo Viruses vol 3 no2 pp 160ndash171 2011
[16] R J Kuhn W Zhang M G Rossmann et al ldquoStructure ofdengue virus implications for flavivirus organization matura-tion and fusionrdquo Cell vol 108 no 5 pp 717ndash725 2002
[17] K Stiasny J H Aberle M Keller B Grubeck-Loebenstein andF X Heinz ldquoAge affects quantity but not quality of antibodyresponses after vaccination with an inactivated flavivirus vac-cine against tick-borne encephalitisrdquo PLoS ONE vol 7 no 3Article ID e34145 2012
[18] LThomasOVerlaetenACabie et al ldquoInfluence of the dengueserotype previous dengue infection and plasma viral load
on clinical presentation and outcome during a dengue-2 anddengue-4 co-epidemicrdquo American Journal of Tropical Medicineand Hygiene vol 78 no 6 pp 990ndash998 2008
[19] Y Zhang J Corver P R Chipman et al ldquoStructures of imma-ture flavivirus particlesrdquo EMBO Journal vol 22 no 11 pp2604ndash2613 2003
[20] A K I Falconar ldquoIdentification of an epitope on the denguevirus membrane (M) protein defined by cross-protective mon-oclonal antibodies design of an improved epitope sequencebased on common determinants present in both envelope (EandM) proteinsrdquo Archives of Virology vol 144 no 12 pp 2313ndash2330 1999
[21] K-J Huang Y-C Yang Y-S Lin et al ldquoThe dual-specific bind-ing of dengue virus and target cells for the antibody-dependentenhancement of dengue virus infectionrdquo Journal of Immunol-ogy vol 176 no 5 pp 2825ndash2832 2006
[22] A K I Falconar ldquoAntibody responses are generated to immun-odominant ELKKLE-typemotifs on the nonstructural-1 glyco-protein during live dengue virus infections inmice and humansimplications for diagnosis pathogenesis and vaccine designrdquoClinical and Vaccine Immunology vol 14 no 5 pp 493ndash5042007
[23] C W Mandl F Guirakhoo H Holzmann F X Heinz and CKunz ldquoAntigenic structure of the flavivirus envelope protein Eat the molecular level using the tick-borne encephalitis virus asa modelrdquo Journal of Virology vol 63 no 2 pp 564ndash571 1989
[24] F A Rey F X Heinz C Mandl C Kunz and S C HarrisonldquoThe envelope glycoprotein from tick-borne encephalitis virusat 2 A resolutionrdquo Nature vol 375 no 6529 pp 291ndash298 1995
[25] S L Allison K Stiasny K Stadler CWMandl and F XHeinzldquoMapping of functional elements in the stem-anchor regionof tick-borne encephalitis virus envelope protein Erdquo Journal ofVirology vol 73 no 7 pp 5605ndash5612 1999
[26] D E Klein J L Choi and S C Harrison ldquoStructure of a denguevirus envelope protein late-stage fusion intermediaterdquo Journal ofVirology vol 87 no 4 pp 2287ndash2293 2013
[27] S L Hanna T C Pierson M D Sanchez A A Ahmed M MMurtadha and R W Doms ldquoN-linked glycosylation of WestNile virus envelope proteins influences particle assembly andinfectivityrdquo Journal of Virology vol 79 no 21 pp 13262ndash132742005
[28] N Mancini G Sautto N Clementi et al ldquoNeutralization inter-fering antibodies a ldquonovelrdquo example of humoral immune dys-function facilitating viral escaperdquoViruses vol 4 no 9 pp 1731ndash1752 2012
[29] Y Modis S Ogata D Clements and S C Harrison ldquoVariablesurface epitopes in the crystal structure of dengue virus type3 envelope glycoproteinrdquo Journal of Virology vol 79 no 2 pp1223ndash1231 2005
[30] N Miller ldquoRecent progress in dengue vaccine research anddevelopmentrdquo Current Opinion in Molecular Therapeutics vol12 no 1 pp 31ndash38 2010
[31] J H Chavez J R Silva A A Amarilla and L T Moraes Figu-eiredo ldquoDomain III peptides from flavivirus envelope proteinare useful antigens for serologic diagnosis and targets forimmunizationrdquo Biologicals vol 38 no 6 pp 613ndash618 2010
[32] B Shrestha J D Brien S Sukupolvi-Petty et al ldquoThe develop-ment of therapeutic antibodies that neutralize homologous andheterologous genotypes of dengue virus type 1rdquo PLoS Pathogensvol 6 no 4 Article ID e1000823 2010
18 BioMed Research International
[33] G D Gromowski N D Barrett and A D T Barrett ldquoChar-acterization of dengue virus complex-specific neutralizing epi-topes on envelope protein domain III of dengue 2 virusrdquo Journalof Virology vol 82 no 17 pp 8828ndash8837 2008
[34] S Sukupolvi-Petty S K Austin W E Purtha et al ldquoType- andsubcomplex-specific neutralizing antibodies against domain IIIof dengue virus type 2 envelope protein recognize adjacentepitopesrdquo Journal of Virology vol 81 no 23 pp 12816ndash128262007
[35] S Sukupolvi-Petty S K Austin M Engle et al ldquoStructure andfunction analysis of therapeutic monoclonal antibodies againstdengue virus type 2rdquo Journal of Virology vol 84 no 18 pp9227ndash9239 2010
[36] Y-Q Deng J-X Dai G-H Ji et al ldquoA broadly flaviviruscross-neutralizingmonoclonal antibody that recognizes a novelepitope within the fusion loop of e proteinrdquo PLoS ONE vol 6no 1 Article ID e16059 2011
[37] J M Costin E Zaitseva K M Kahle et al ldquoMechanistic studyof broadly neutralizing human monoclonal antibodies againstdengue virus that target the fusion looprdquo Journal of Virologyvol 87 no 1 pp 52ndash66 2013
[38] CMMidgley A FlanaganH B Tran et al ldquoStructural analysisof a dengue cross-reactive antibody complexed with envelopedomain III reveals the molecular basis of cross-reactivityrdquoJournal of Immunology vol 188 no 10 pp 4971ndash4979 2012
[39] N J Moreland P Susanto E Lim et al ldquoPhage display appro-aches for the isolation of monoclonal antibodies against denguevirus envelope domain III from human and mouse derivedlibrariesrdquo International Journal ofMolecular Sciences vol 13 no3 pp 2618ndash2635 2012
[40] J J B Cockburn M E Navarro Sanchez N Fretes et alldquoMechanism of dengue virus broad cross-neutralization by amonoclonal antibodyrdquo Structure vol 20 no 2 pp 303ndash3142012
[41] R Rajamanonmani C Nkenfou P Clancy et al ldquoOn a mousemonoclonal antibody that neutralizes all four dengue virusserotypesrdquo Journal of General Virology vol 90 no 4 pp 799ndash809 2009
[42] L H Gould J Sui H Foellmer et al ldquoProtective and herapeuticcapacity of human single-chain Fv-Fc fusion proteins againstWest Nile virusrdquo Journal of Virology vol 79 no 23 pp 14606ndash14613 2005
[43] H Sultana H G Foellmer G Neelakanta et al ldquoFusion LoopPeptide of the West Nile virus envelope protein is essential forpathogenesis and is recognized by a therapeutic cross-reactivehuman monoclonal antibodyrdquo Journal of Immunology vol 183no 1 pp 650ndash660 2009
[44] T Oliphant M Engle G E Nybakken et al ldquoDevelopment ofa humanized monoclonal antibody with therapeutic potentialagainstWest Nile virusrdquoNature Medicine vol 11 no 5 pp 522ndash530 2005
[45] G E Nybakken T Oliphant S Johnson S Burke M S Dia-mond and D H Fremont ldquoStructural basis of West Nile virusneutralization by a therapeutic antibodyrdquo Nature vol 437 no7059 pp 764ndash769 2005
[46] G D Gromowski J T Roehrig M S Diamond J C LeeT J Pitcher and A D T Barrett ldquoMutations of an antibodybinding energy hot spot on domain III of the dengue 2 envelopeglycoprotein exploited for neutralization escaperdquo Virology vol407 no 2 pp 237ndash246 2010
[47] J T Roehrig R A Bolin andR G Kelly ldquoMonoclonal antibodymapping of the envelope glycoprotein of the dengue 2 virusJamaicardquo Virology vol 246 no 2 pp 317ndash328 1998
[48] R de Alwis S A Smith N P Olivarez et al ldquoIdentification ofhumanneutralizing antibodies that bind to complex epitopes ondengue virionsrdquo Proceedings of the National Academy of Sciencesof the United States of America vol 109 no 19 pp 7439ndash74442012
[49] S K Austin K A Dowd B Shrestha et al ldquoStructural basis ofdifferential neutralization of DENV-1 genotypes by an antibodythat recognizes a cryptic epitoperdquo PLoS Pathogens vol 8 no 10Article ID e1002930 2012
[50] M Throsby C Geuijen J Goudsmit et al ldquoIsolation and char-acterization of human monoclonal antibodies from individualsinfected with West Nile Virusrdquo Journal of Virology vol 80 no14 pp 6982ndash6992 2006
[51] A P Goncalvez R H Purcell and C-J Lai ldquoEpitope deter-minants of a chimpanzee Fab antibody that efficiently cross-neutralizes dengue type 1 and type 2 viruses map to inside andin close proximity to fusion loop of the dengue type 2 virusenvelope glycoproteinrdquo Journal of Virology vol 78 no 23 pp12919ndash12928 2004
[52] J T Roehrig J H Mathews and D W Trent ldquoIdentification ofepitopes on the E glycoprotein of Saint Louis encephalitis virususing monoclonal antibodiesrdquo Virology vol 128 no 1 pp 118ndash126 1983
[53] J T Roehrig L A Staudinger A R Hunt J H Mathews andC D Blair ldquoAntibody prophylaxis and therapy for flavivirusencephalitis infectionsrdquo Annals of the New York Academy ofSciences vol 951 pp 286ndash297 2001
[54] A P Goncalvez C-H Chien K Tubthong et al ldquoHumanizedmonoclonal antibodies derived from chimpanzee fabs protectagainst Japanese encephalitis virus in vitro and in vivordquo Journalof Virology vol 82 no 14 pp 7009ndash7021 2008
[55] M Arakawa T Yamashiro G-I Uechi M Tadano and ANishizono ldquoConstruction of humanFab (1205741120581) library and iden-tification of human monoclonal Fab possessing neutralizingpotency against Japanese encephalitis virusrdquo Microbiology andImmunology vol 51 no 6 pp 617ndash625 2007
[56] L Warter C Y Lee R Thiagarajan et al ldquoChikungunya virusenvelope-specific human monoclonal antibodies with broadneutralization potencyrdquo Journal of Immunology vol 186 no 5pp 3258ndash3264 2011
[57] P Pal K A Dowd J D Brien et al ldquoDevelopment of a highlyprotective combination monoclonal antibody therapy againstchikungunya virusrdquo PLoS Pathogens vol 9 no 4 Article IDe1003312 2013
[58] A Bertolotti-Ciarlet J Smith K Strecker et al ldquoCellular local-ization and antigenic characterization of Crimean-Congo hem-orrhagic fever virus glycoproteinsrdquo Journal of Virology vol 79no 10 pp 6152ndash6161 2005
[59] J K Battles and J M Dalrymple ldquoGenetic variation amonggeographic isolates of Rift Valley fever virusrdquo American Journalof Tropical Medicine and Hygiene vol 39 no 6 pp 617ndash6311988
[60] B AThibodeaux N C Garbino NM Liss et al ldquoA humanizedIgG but not IgM antibody is effective in prophylaxis and therapyof yellow fever infection in an AG12917D-204 peripheralchallenge mouse modelrdquo Antiviral Research vol 94 no 1 pp1ndash8 2012
BioMed Research International 19
[61] S Daffis R E Kontermann J Korimbocus H Zeller H-DKlenk and J Ter Meulen ldquoAntibody responses against wild-type yellow fever virus and the 17D vaccine strain charac-terization with human monoclonal antibody fragments andneutralization escape variantsrdquoVirology vol 337 no 2 pp 262ndash272 2005
[62] L N Levanov L E Matveev E P Goncharova et al ldquoChimericantibodies against tick-borne encephalitis virusrdquo Vaccine vol28 no 32 pp 5265ndash5271 2010
[63] A R Hunt S Frederickson C Hinkel K S Bowdish and J TRoehrig ldquoA humanized murine monoclonal antibody protectsmice either before or after challenge with virulent Venezuelanequine encephalomyelitis virusrdquo Journal of General Virologyvol 87 no 9 pp 2467ndash2476 2006
[64] J M da Silva Voorham I A Rodenhuis-Zybert N V A Nunezet al ldquoAntibodies against the envelope glycoprotein promoteinfectivity of immature dengue virus serotype 2rdquo PLoS ONEvol 7 no 3 Article ID e29957 2012
[65] T C Pierson Q Xu S Nelson et al ldquoThe stoichiometry ofantibody-mediated neutralization and enhancement of westnile virus infectionrdquoCell Host andMicrobe vol 1 no 2 pp 135ndash145 2007
[66] G Sautto N Mancini R A Diotti L Solforosi M Clementiand R Burioni ldquoAnti-hepatitis C virus E2 (HCVE2) glyco-proteinmonoclonal antibodies andneutralization interferencerdquoAntiviral Research vol 96 no 1 pp 82ndash89 2012
[67] R Burioni Y Matsuura N Mancini et al ldquoDiverging effectsof human recombinant anti-hepatitis C virus (HCV) antibodyfragments derived from a single patient on the infectivityof a vesicular stomatitis virusHCV pseudotyperdquo Journal ofVirology vol 76 no 22 pp 11775ndash11779 2002
[68] R J Phillpotts J R Stephenson and J S Porterfield ldquoAntibody-dependent enhancement of tick-borne encephalitus virus infec-tivityrdquo Journal of General Virology vol 66 no 8 pp 1831ndash18371985
[69] K A Dowd and T C Pierson ldquoAntibody-mediated neutraliza-tion of flaviviruses a reductionist viewrdquo Virology vol 411 no 2pp 306ndash315 2011
[70] A K I Falconar ldquoMonoclonal antibodies that bind to commonepitopes on the dengue virus type 2 nonstructural-1 andenvelope glycoproteins display weak neutralizing activity anddifferentiated responses to virulent strains implications forpathogenesis and vaccinesrdquo Clinical and Vaccine Immunologyvol 15 no 3 pp 549ndash561 2008
[71] K Meyer A Banerjee S E Frey R B Belshe and R Ray ldquoAweak neutralizing antibody response to hepatitis C virus enve-lope glycoprotein enhances virus infectionrdquo PLoS ONE vol 6no 8 Article ID e23699 2011
[72] R Burioni R AWilliamson P P Sanna F E Bloom and D RBurton ldquoRecombinant human Fab to glycoprotein D neutral-izes infectivity and prevents cell-to-cell transmission of herpessimplex viruses 1 and 2 in vitrordquo Proceedings of the NationalAcademy of Sciences of the United States of America vol 91 no1 pp 355ndash359 1994
[73] M S Rolph A Zaid N E Rulli and S Mahalingam ldquoDown-regulation of interferon-120573 in antibody-dependent enhancementof dengue viral infections of human macrophages is dependenton interleukin-6rdquo Journal of Infectious Diseases vol 204 no 3pp 489ndash491 2011
[74] S S Whitehead J E Blaney A P Durbin and B R MurphyldquoProspects for a dengue virus vaccinerdquoNature ReviewsMicrobi-ology vol 5 no 7 pp 518ndash528 2007
[75] S-M Lok V Kostyuchenko G ENybakken et al ldquoBinding of aneutralizing antibody to dengue virus alters the arrangement ofsurface glycoproteinsrdquoNature Structural andMolecular Biologyvol 15 no 3 pp 312ndash317 2008
[76] A P Goncalvez R Men C Wernly R H Purcell and C-J Lai ldquoChimpanzee Fab fragments and a derived humanizedimmunogiobulin G1 antibody that efficiently cross-neutralizedengue type 1 and type 2 virusesrdquo Journal of Virology vol 78no 23 pp 12910ndash12918 2004
[77] WM P BWahala A A Kraus L B HaymoreM A Accavitti-Loper and A M de Silva ldquoDengue virus neutralization byhuman immune sera role of envelope protein domain III-reactive antibodyrdquo Virology vol 392 no 1 pp 103ndash113 2009
[78] W D Crill H R Hughes M J Delorey and G-J J ChangldquoHumoral immune responses of dengue fever patients usingepitope-specific serotype-2 virus-like particle antigensrdquo PLoSONE vol 4 no 4 Article ID e4991 2009
[79] C Setthapramote T Sasaki O Puiprom et al ldquoHuman mono-clonal antibodies to neutralize all dengue virus serotypes usinglymphocytes from patients at acute phase of the secondaryinfectionrdquo Biochemical and Biophysical Research Communica-tions vol 423 no 4 pp 867ndash872 2012
[80] M Beltramello K L Williams C P Simmons et al ldquoThehuman immune response to dengue virus is dominated byhighly cross-reactive antibodies endowed with neutralizing andenhancing activityrdquo Cell Host andMicrobe vol 8 no 3 pp 271ndash283 2010
[81] C M Midgley M Bajwa-Joseph S Vasanawathana et al ldquoAnin-depth analysis of original antigenic sin in dengue virusinfectionrdquo Journal of Virology vol 85 no 22 pp 410ndash421 2011
[82] B Kaufmann M R Vogt J Goudsmit et al ldquoNeutralizationof West Nile virus by cross-linking of its surface proteins withFab fragments of the human monoclonal antibody CR4354rdquoProceedings of the National Academy of Sciences of the UnitedStates of America vol 107 no 44 pp 18950ndash18955 2010
[83] B Spurrier J M Sampson M Totrov et al ldquoStructural analysisof human and macaque mAbs 2909 and 25B implicationsfor the configuration of the quaternary neutralizing epitope ofHIV-1 gp120rdquo Structure vol 19 no 5 pp 691ndash699 2011
[84] J J SchlesingerMWBrandriss andT PMonath ldquoMonoclonalantibodies distinguish between wild and vaccine strains ofyellow fever virus by neutralization hemagglutination inhibi-tion and immune precipitation of the virus envelope proteinrdquoVirology vol 125 no 1 pp 8ndash17 1983
[85] M W Brandriss J J Schlesinger E E Walsh and M BrisellildquoLethal 17D yellow fever encephalitis in mice I Passive protec-tion by monoclonal antibodies to the envelope proteins of 17Dyellow fever and dengue 2 virusesrdquo Journal of General Virologyvol 67 no 2 pp 229ndash234 1986
[86] H Cho and M S Diamond ldquoImmune responses to West Nilevirus infection in the central nervous systemrdquo Viruses vol 4no 12 pp 3812ndash3830 2012
[87] D W Beasley A D Barrett and R B Tesh ldquoResurgence ofWest Nile neurologic disease in the United States in 2012 whathappened What needs to be donerdquo Antiviral Research vol 99no 1 pp 1ndash5 2013
[88] M Throsby J Ter Meulen C Geuijen J Goudsmit and J DeKruif ldquoMapping and analysis of West Nile virus-specific mon-oclonal antibodies prospects for vaccine developmentrdquo ExpertReview of Vaccines vol 6 no 2 pp 183ndash191 2007
[89] M S Diamond T G Roberts D Edgil B Lu J Ernst and EHarris ldquoModulation of dengue virus infection in human cells
20 BioMed Research International
by alpha beta and gamma interferonsrdquo Journal of Virology vol74 no 11 pp 4957ndash4966 2000
[90] Z Shimoni M J Niven S Pitlick and S Bulvik ldquoTreatmentof West Nile virus encephalitis with intravenous immunoglob-ulinrdquo Emerging Infectious Diseases vol 7 no 4 article 759 2001
[91] AHamdan PGreen EMendelsonM R Kramer S Pitlik andM Weinberger ldquoPossible benefit of intravenous immunoglob-ulin therapy in a lung transplant recipient with West Nile virusencephalitisrdquoTransplant InfectiousDisease vol 4 no 3 pp 160ndash162 2002
[92] M Haley A S Retter D Fowler J Gea-Banacloche and NP OrsquoGrady ldquoThe role for intravenous immunoglobulin in thetreatment of West Nile virus encephalitisrdquo Clinical InfectiousDiseases vol 37 no 6 pp e88ndashe90 2003
[93] K M Chung G E Nybakken B S Thompson et al ldquoAnti-bodies against West Nile virus nonstructural protein NS1prevent lethal infection through Fc 120574 receptor-dependent and-independent mechanismsrdquo Journal of Virology vol 80 no 3pp 1340ndash1351 2006
[94] E A Henchal L S Henchal and J J Schlesinger ldquoSynergisticinteractions of anti-NS1 monoclonal antibodies protect pas-sively immunized mice from lethal challenge with dengue 2virusrdquo Journal of General Virology vol 69 no 8 pp 2101ndash21071988
[95] J T Roehrig ldquoAntigenic structure of flavivirus proteinsrdquoAdvances in Virus Research vol 59 pp 141ndash175 2003
[96] M D Sanchez T C Pierson D McAllister et al ldquoCharacteri-zation of neutralizing antibodies to West Nile virusrdquo Virologyvol 336 no 1 pp 70ndash82 2005
[97] J H Beigel J L Nordstrom S R Pillemer et al ldquoSafety andpharmacokinetics of single intravenous dose of MGAWN1 anovel monoclonal antibody to West Nile virusrdquo AntimicrobialAgents and Chemotherapy vol 54 no 6 pp 2431ndash2436 2010
[98] C A Smeraski V Siddharthan and J D Morrey ldquoTreatmentof spatial memory impairment in hamsters infected with WestNile virus using a humanizedmonoclonal antibodyMGAWN1rdquoAntiviral Research vol 91 no 1 pp 43ndash49 2011
[99] D Lelli AMoreno E Brocchi et al ldquoWestNile virus character-ization and diagnostic applications of monoclonal antibodiesrdquoVirology Journal vol 9 article 81 2012
[100] T S Gritsun V A Lashkevich and E A Gould ldquoTick-borneencephalitisrdquo Antiviral Research vol 57 no 1-2 pp 129ndash1462003
[101] D Ruzek T S Gritsun N L Forrester et al ldquoMutations inthe NS2B and NS3 genes affect mouse neuroinvasiveness of aWestern European field strain of tick-borne encephalitis virusrdquoVirology vol 374 no 2 pp 249ndash255 2008
[102] M Ecker S L Allison T Meixner and F X Heinz ldquoSequenceanalysis and genetic classification of tick-borne encephalitisviruses from Europe and Asiardquo Journal of General Virology vol80 no 1 pp 179ndash185 1999
[103] H Holzmann M S Vorobyova I P Ladyzhenskaya et alldquoMolecular epidemiology of tick-borne encephalitis viruscross-protection between European and Far Eastern subtypesrdquoVaccine vol 10 no 5 pp 345ndash349 1992
[104] C W Mandl ldquoSteps of the tick-borne encephalitis virus repli-cation cycle that affect neuropathogenesisrdquo Virus Research vol111 no 2 pp 161ndash174 2005
[105] A V Atrasheuskaya T M Fredeking and G M IgnatyevldquoChanges in immune parameters and their correction in humancases of tick-borne encephalitisrdquo Clinical and ExperimentalImmunology vol 131 no 1 pp 148ndash154 2003
[106] K L Mansfield N Johnson L P Phipps J R Stephenson AR Fooks and T Solomon ldquoTick-borne encephalitis virusmdashareview of an emerging zoonosisrdquo Journal of General Virologyvol 90 no 8 pp 1781ndash1794 2009
[107] DWVaughn andCHHoke Jr ldquoThe epidemiology of Japaneseencephalitis prospects for preventionrdquo Epidemiologic Reviewsvol 14 pp 197ndash221 1992
[108] T Solomon H Ni D W C Beasley M Ekkelenkamp M JCardosa and A D T Barrett ldquoOrigin and evolution of Japaneseencephalitis virus in Southeast Asiardquo Journal of Virology vol 77no 5 pp 3091ndash3098 2003
[109] M R Wilson ldquoEmerging viral infectionsrdquo Current Opinion inNeurology vol 26 no 3 pp 301ndash306 2013
[110] J N Hanna S A Ritchie D A Phillips et al ldquoAn outbreakof Japanese encephalitis in the Torres Strait Australia 1995rdquoMedical Journal of Australia vol 165 no 5 pp 256ndash260 1996
[111] J N Hanna S A Ritchie D A Phillips et al ldquoJapanese ence-phalitis in north Queensland Australia 1998rdquo Medical Journalof Australia vol 170 no 11 pp 533ndash536 1999
[112] SGeorgeMGourie-Devi and J A Rao ldquoIsolation ofWestNilevirus from the brains of children who had died of encephalitisrdquoBulletin of theWorldHealthOrganization vol 62 no 6 pp 879ndash882 1984
[113] A K Gupta A A Koshy and V J Lad ldquoEnhanced protec-tion of mice against Japanese encephalitis virus infection bycombinations ofmonoclonal antibodies to glycoprotein ErdquoActaVirologica vol 55 no 2 pp 165ndash168 2011
[114] T H Lee B-H Song S-I Yun et al ldquoA cross-protective mAbrecognizes a novel epitope within the flavivirus NS1 proteinrdquoJournal of General Virology vol 93 no 1 pp 20ndash26 2012
[115] S C Weaver ldquoHost range amplification and arboviral diseaseemergencerdquo Archives of Virology Supplementum no 19 pp 33ndash44 2005
[116] M Solignat B Gay S Higgs L Briant and C Devaux ldquoRepli-cation cycle of chikungunya a re-emerging arbovirusrdquoVirologyvol 393 no 2 pp 183ndash197 2009
[117] O Schwartz and M L Albert ldquoBiology and pathogenesis ofchikungunya virusrdquo Nature Reviews Microbiology vol 8 no 7pp 491ndash500 2010
[118] W Akahata Z-Y Yang H Andersen et al ldquoA virus-like particlevaccine for epidemic Chikungunya virus protects nonhumanprimates against infectionrdquo Nature Medicine vol 16 no 3 pp334ndash338 2010
[119] M Kielian and A Helenius ldquopH-Iinduced alterations in thefusogenic spike protein of Semliki Forest virusrdquo Journal of CellBiology vol 101 no 6 pp 2284ndash2291 1985
[120] J White and A Helenius ldquopH-Dependent fusion between theSemliki Forest virus membrane and liposomesrdquo Proceedings ofthe National Academy of Sciences of the United States of Americavol 77 no 6 pp 3273ndash3277 1980
[121] J Dubuisson and C M Rice ldquoSindbis virus attachment isola-tion and characterization of mutants with impaired binding tovertebrate cellsrdquo Journal of Virology vol 67 no 6 pp 3363ndash33741993
[122] C Y Lee Y-W Kam J Fric et al ldquoChikungunya virus neutral-ization antigens and direct cell-to-cell transmission are revealedby human antibody-escapemutantsrdquo PLoS Pathogens vol 7 no12 Article ID e1002390 2011
[123] J E Voss M-C Vaney S Duquerroy et al ldquoGlycoproteinorganization of Chikungunya virus particles revealed by X-raycrystallographyrdquo Nature vol 468 no 7324 pp 709ndash712 2010
BioMed Research International 21
[124] L Li J Jose Y Xiang R J Kuhn and M G Rossmann ldquoStruc-tural changes of envelope proteins during alphavirus fusionrdquoNature vol 468 no 7324 pp 705ndash708 2010
[125] J Fric S Bertin-Maghit C I Wang A Nardin and L WarterldquoUse of human monoclonal antibodies to treat Chikungunyavirus infectionrdquo Journal of Infectious Diseases vol 207 no 2 pp319ndash322 2013
[126] P V Aguilar J G Estrada-Franco R Navarro-Lopez C FerroA D Haddow and S C Weaver ldquoEndemic Venezuelanequine encephalitis in the Americas hidden under the dengueumbrellardquo Future Virology vol 6 no 6 pp 721ndash740 2011
[127] S C Weaver R Winegar I D Manger and N L ForresterldquoAlphaviruses population genetics and determinants of emer-gencerdquo Antiviral Research vol 94 no 3 pp 242ndash257 2012
[128] S Paessler and S C Weaver ldquoVaccines for Venezuelan equineencephalitisrdquo Vaccine vol 27 no 4 pp D80ndashD85 2009
[129] R J Phillpotts ldquoVenezuelan equine encephalitis virus complex-specific monoclonal antibody provides broad protection inmurine models against airborne challenge with viruses fromserogroups I II and IIIrdquoVirus Research vol 120 no 1-2 pp 107ndash112 2006
[130] R J Phillpotts L D Jones and S C Howard ldquoMonoclonalantibody protectsmice against infection anddiseasewhen giveneither before or up to 24 h after airborne challenge with virulentVenezuelan equine encephalitis virusrdquoVaccine vol 20 no 11-12pp 1497ndash1504 2002
[131] J H Mathews J T Roehrig and D W Trent ldquoRole of com-plement and the Fc portion of immunoglobulin G in immunityto Venezuelan equine encephalomyelitis virus infection withglycoprotein-specific monoclonal antibodiesrdquo Journal of Virol-ogy vol 55 no 3 pp 594ndash600 1985
[132] L M OrsquoBrien C D Underwood-Fowler S A Goodchild AL Phelps and R J Phillpotts ldquoDevelopment of a novel mon-oclonal antibody with reactivity to a wide range of Venezuelanequine encephalitis virus strainsrdquoVirology Journal vol 6 article206 2009
[133] L M OrsquoBrien S A Goodchild R J Phillpotts and S D Per-kins ldquoA humanised murine monoclonal antibody protectsmice from Venezuelan equine encephalitis virus Evergladesvirus and Mucambo virus when administered up to 48h afterairborne challengerdquo Virology vol 426 no 2 pp 100ndash105 2012
[134] N Clementi N Mancini L Solforosi M Castelli M Clementiand R Burioni ldquoPhage display-based strategies for cloning andoptimization of monoclonal antibodies directed against humanpathogensrdquo International Journal of Molecular Sciences vol 13no 7 pp 8273ndash8292 2012
[135] A R Hunt R A Bowen S Frederickson T Maruyama JT Roehrig and C D Blair ldquoTreatment of mice with humanmonoclonal antibody 24h after lethal aerosol challenge withvirulent Venezuelan equine encephalitis virus prevents diseasebut not infectionrdquo Virology vol 414 no 2 pp 146ndash152 2011
[136] W-G Hu A L Phelps S Jager et al ldquoA recombinant human-ized monoclonal antibody completely protects mice againstlethal challenge with Venezuelan equine encephalitis virusrdquoVaccine vol 28 no 34 pp 5558ndash5564 2010
[137] T Rulker L Voss P Thullier et al ldquoIsolation and characterisa-tion of a human-like antibody fragment (scFv) that inactivatesVEEV in vitro and in vivordquo PLoS ONE vol 7 no 5 Article IDe37242 2012
[138] C T Walter and J N Barr ldquoRecent advances in the molecularand cellular biology of bunyavirusesrdquo Journal of General Virol-ogy vol 92 no 11 pp 2467ndash2484 2011
[139] C A Whitehouse ldquoCrimean-Congo hemorrhagic feverrdquo Anti-viral Research vol 64 no 3 pp 145ndash160 2004
[140] M Mertens K Schmidt A Ozkul and M H Groschup ldquoTheimpact of Crimean-Congo hemorrhagic fever virus on publichealthrdquo Antiviral Research vol 98 no 2 pp 248ndash260 2013
[141] A A Ahmed J M McFalls C Hoffmann et al ldquoPresence ofbroadly reactive and group-specific neutralizing epitopes onnewly described isolates of Crimean-Congo hemorrhagic fevervirusrdquo Journal of General Virology vol 86 no 12 pp 3327ndash33362005
[142] R Hewson A Gmyl L Gmyl et al ldquoEvidence of segment reas-sortment in Crimean-Congo haemorrhagic fever virusrdquo Journalof General Virology vol 85 no 10 pp 3059ndash3070 2004
[143] N K Blackburn T G Besselaar A J Shepherd and R Swane-poel ldquoPreparation and use of monoclonal antibodies for iden-tifying Crimean-Congo hemorrhagic fever virusrdquo AmericanJournal of Tropical Medicine and Hygiene vol 37 no 2 pp 392ndash397 1987
[144] T Ikegami ldquoMolecular biology and genetic diversity of RiftValley fever virusrdquoAntiviral Research vol 95 no 3 pp 293ndash3102012
[145] KKeegan andM S Collett ldquoUse of bacterial expression cloningto define the amino acid sequences of antigenic determinantson the G2 glycoprotein of Rift Valley fever virusrdquo Journal ofVirology vol 58 no 2 pp 263ndash270 1986
[146] T G Besselaar and N K Blackburn ldquoTopological mapping ofantigenic sites on the Rift Valley fever virus envelope glycopro-teins using monoclonal antibodiesrdquo Archives of Virology vol121 no 1ndash4 pp 111ndash124 1991
[147] L G Presta ldquoEngineering of therapeutic antibodies to mini-mize immunogenicity and optimize functionrdquo Advanced DrugDelivery Reviews vol 58 no 5-6 pp 640ndash656 2006
Submit your manuscripts athttpwwwhindawicom
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anatomy Research International
PeptidesInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporation httpwwwhindawicom
International Journal of
Volume 2014
Zoology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Molecular Biology International
GenomicsInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioinformaticsAdvances in
Marine BiologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Signal TransductionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Evolutionary BiologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Biochemistry Research International
ArchaeaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Genetics Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in
Virolog y
Hindawi Publishing Corporationhttpwwwhindawicom
Nucleic AcidsJournal of
Volume 2014
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Enzyme Research
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Microbiology

4 BioMed Research International
receptorFc-120574
Cellular receptor
(a)
Cellular receptor
(b) (c)
Figure 1 Mechanisms of antibody-dependent enhancement (ADE) of infection (a) After binding to the viral epitope the Ab is recognizedby a cellular Fc-120574 receptor bringing the viral particle in proximity of the target cell (b) the binding of the Ab induces conformational changeswithin the structure of the viral target protein These changes improve the affinity for the cellular receptor (c) molecular mimicry by a viralmotif of cellular membrane components leads an autoreactive Ab to bind both the viral and the cellular target bringing the virus in proximityof the target cell
and CH3 portions or direct expression of the mAb as aFab fragment) andor deletion of the N-linked sugars onIgG molecules that are both required for interactions withFc-120574 receptors (Fc-120574R) or eventually by the blocking ofFc-120574R engagement with anti-Fc-120574R Abs [70] Similarly theidentification and detailed examination of cross-neutralizingepitopes that do not promote ADE of infection may definenovel targets for vaccine development
However in some instances ADE of infection mayoccur when Ab molecules cross-react with both viral andcellular antigens (Figure 1(c)) In this regard Fc-120574 receptor-independentmechanismof infection in particular forDENVprM-specific Abs able to bind simultaneously to the virus andtarget cells has been described Indeed othermembers of theFlaviviridae belonging to another genus like hepatitis C virus(HCV) have been shown to elicit polyreactive Abs that areprobably responsible for an ADE of infection mechanism aswell as of secondary clinical manifestations In connectionwith this several studies employing both polyclonal andmonoclonal Abs suggested the presence of common epitopescontaining particular short motifs on NS1 and the threedomains of the E protein as well as human proteins thatmay play a role in ADE of infection mechanisms and thusin DHFDSS pathogenesis [70] Interestingly these commonmotifs have been found to be additionally represented inDENV2 strains which have greater human pathogenic capac-ities [22]
Moreover another molecular mechanism of ADE ofinfection could consist in the facilitation of conformationalchanges of the targeted surface protein that are required forvirus entry as described and hypothesized for other viruses(eg HIV HCV and HSV) (Figure 1(b)) [71 72]
Finally Fc-120574R engagement could result also in ldquointrinsicrdquoADE of infection mechanisms involving signaling eventssuch as inhibition of antiviral response and increased viralreplication Disrupted signaling events include the RIG-IMDA5 cascade and type I interferon (IFN) response aswell as induction and suppression of anti-inflammatory(IL-10) and proinflammatory (IL-12 and TNF-120572) cytokinesrespectively [73]
24 Dengue Virus (DENV) Dengue virus (DENV) is respon-sible for 50ndash100 million symptomatic infections each yearresulting in 500000 hospitalizations and over 20000 deathswhich occurs mostly in tropical and subtropical regions ofthe world Although malaria remains the most importantcause for systemic febrile disease in travelers chikungunyavirus (CHIKV) and DENV are increasingly diagnosed withdengue currently being the second most important cause forfebrile disease in travelers
Decreases in mosquito control efforts during the endof the 90s coupled with societal factors (eg globalizationmigrations and dense urbanization) have contributed to thereemergence of flaviviruses such as DENV in South and Cen-tral America Development of effective DENV vaccines thatexhibit cross-protection thought to be important for prevent-ing subsequent dengue-associated immunopathogenesis isproving to be particularly challenging [1]
A vaccine for this infection is not yet available despiteconsiderable public and private efforts This difficulty ismainly due to the perceived need to simultaneously protectthe four known serotypes of DENV (DENV1ndash4) which shareabout 70 of sequence homology while genotypes can varyup to 3 Moreover as anticipated and discussed previously
BioMed Research International 5
non-protective Abs may contribute to more severe clinicaloutcomes in vaccinated people [74]
241 Murine versus Human Anti-DENV Humoral ImmuneResponse It is well ascertained that anti-E DIII-directed Absare virtually absent from the naıve human repertoire as theyare directed away from this domain probably against weaklyneutralizing and immunodominant regions Moreover it hasbeen demonstrated that over than 90 of the human Abresponse of primary DENV infected patients is able to bindonly native DENV virus particles instead of a recombinantform of the E protein posing difficulties in the cloning strate-gies of neutralizing mAbs [39] Thus it can be concludedthat in vivo only a small fraction of DENV-specific Abs areresponsible for neutralization
Conversely murine mAbs that recognize all the threedomains of E have been identified Epitope-mapping studiesusingmurine serotype-specificmAbs have demonstrated thatloops located in the lateral ridge region of DIII consti-tute the strongest neutralizing B-cell target In this regionsequence diversity between serotype is high while cross-reactive murine mAbs that recognize another loop showedgenerally a weaker neutralization profile In particular type-specific mAbs with neutralizing activity against DENV2localized to the BC DE and FG loops on the lateral ridgeof DIII whereas subcomplex-specific mAbs recognized anadjacent epitope centered on the connecting A strand of DIIIat residues K305 K307 and K310 [32ndash35]
Cross-reactive mAbs directed against prM and NS1 havealso been described but as anticipated they feature weaklyneutralizing or absent as well as ADE of infection activityIn the following paragraphs we report the better character-ized neutralizing mouse and human mAbs describing theirmolecular features further summarized in Table 1
242 Anti-DENV Murine mAbs Murine mAbs directedagainst all the three domains of the E protein and endowedof different neutralizing and binding characteristics havebeen described However only DIII and DII are classicallyrecognized by neutralizing Abs In this regard Sukupolvi-Petty et al isolated twenty-four anti-DENV2 mouse mAbswith moderate or strong neutralizing activity against thehomologous virus in cell culture assays Binding sites weremapped for the majority of these in distinct epitopes inregions located in DI (lateral ridge) DII (dimer interfacelateral ridge and fusion loop) and DIII (lateral ridge CC1015840loop and A strand) Moreover 16 of the neutralizing mAbswere tested in mice with most of them being protectivewhen given as prophylaxis Seven of these had post-exposuretherapeutic activity when administered as a single dose byintraperitoneal route even 3 days after intracranial infectionFor the mAbs with the greatest therapeutic potential protec-tion was confirmed with an Ab-enhanced vascular leakagemouse model of DENV2 infection [35]
Recently Deng et al described a cross-reactive murinemAb named 2A10G6 that is able to recognize DENV1ndash4 YFV WNV JEV and TBEV to potently cross-neutralizeDENV1ndash4 YFV and to a lesser extent WNV This mAbrecognizes a highly conserved motif (amino acid residues
98ndash101) located within the fusion loop of the FlavivirusDII of the E glycoprotein Moreover this mAb exerts itsneutralizing activity in a post-attachment step during thevirus entry process as demonstrated by kinetic neutralizationtests performed in vitro Additionally protection experimentsperformed in mouse models showed that treatment with100 120583gmL of 2A10G6 conferred full protection against lethalDENV2 challenge and 20 120583gmL and 4120583gmL of 2A10G6protected 89 and 40 of infected mice from lethal chal-lenge respectively For infection with DENV1 3 and 4 use of100 120583gmL of 2A10G6 conferred partial protection and 5377 and 73 of the infected mice survived after challengerespectively Finally the protection profile of 2A10G6 againstWNV showed that prophylactic administration with a singledose of 200mg of 2A10G6 conferred 80 protection in miceMost importantly 3 of 8 (375) mice survived when 2A10G6was administered one day after WNV challenge SimilarlyanothermousemAb 4G2 is able to recognize the fusion loopat the extremity of DII from all four DENV serotypes and toprevent syncytia formation [36]
Rajamanonmani et al described a mouse mAb 9F12raised against theDIII of DENV2E protein that is able to rec-ognize DENV1ndash4 serotypes as well asWNV and to neutralizefive DENV strains representative of all DENV serotypes [41]Similarly Cockburn et al reported a comparative high-resolution crystallographic analysis of an A-strand DIIImurinemAb 4E11 in complex with its target domain of the Eprotein from the four DENV serotypes MAb 4E11 is capableof recognizing and neutralizing all four serotypes with IC50values varying between 1 and 100 nMThe structures reportedalso highlight the mechanism by which anti-A-strand mAbsdisrupt the architecture of the mature virion inducing dimerdissociation premature fusion-loop exposure and concomi-tant particle inactivation [40] Also Midgley et al function-ally characterized another murine mAb 2H12 raised in miceagainstDIII of E protein Similarly to the previously describedmAbs 2H12 is able to bind all the four DENV serotypes ina epitope encompassing the conserved 314ETQH317 motifHowever the neutralizing potential of 2H12 is lower than anumber of other anti-DIII mAbs with IC50 values rangingfrom 056 to 145 nM for DENV1 2 and 4 On the other handit showed no ADE of infection activity [38]
Austin et al isolated a murine mAb named E111 whichrecognize a novel CC1015840-loop epitope on DIII of the E proteinfrom two different DENV1 genotypes Docking of the mAbstructure onto the available cryoelectron microscopy modelsof DENV virions revealed that the E111 epitope was inaccessi-ble suggesting that this mAb recognizes an uncharacterizedvirus conformation While the affinity of binding betweenE111 and DIII varied by genotype a limited correlation withneutralizing activity was observed These data support theconclusion that potent neutralization depends on genotype-dependent exposure of the CC1015840 loop epitope [49] In fact aspreviously described elsewhere binding of some E reactiveAbs depends on the dynamicmovement of proteinmoleculesldquobreathingrdquo in the virion particle leading to transient expo-sure of hidden epitopes For instance optimal binding ofmousemAb 1A1D-2 to EDIII requires incubation at 37∘C [75]
6 BioMed Research InternationalTa
ble1Schematicsummaryof
theb
estcharacterized
mAb
sagainstarbo
viralp
atho
gens
mAb
Orig
inRe
activ
ityTargetprotein
Epito
peClon
ingstr
ategy
Form
atisotype
Neutralizingactiv
ityRe
ference
48A
Hum
anDEN
V1ndash4
EDII
EBVtransfo
rmation
IgG
IC50
(120583gmL)
DEN
V115
DEN
V2gt40
DEN
V324
DEN
V4gt40
[37]
D11C
IC50
(120583gmL)
DEN
V115
DEN
V210
DEN
V3102
DEN
V416
16D
IC50
(120583gmL)
DEN
V115
DEN
V202
DEN
V305
DEN
V427
2H12
Murine
DEN
V1ndash4
EDIII(ABloop
aa
314ndash
317)
Hybrid
omafrom
BALB
cmou
seim
mun
ized
with
DEN
V2EDIII
IgG2b
IC50
nM
DEN
V1056ndash54
DEN
V329
DEN
V4145
[38]
C9Murinechim
eric
DEN
V2
EDIII
Phaged
isplayof
achim
ericmurine
hybridom
alibrary
VH1V1205811
PRNT5
0850120583
gmL
[39]
4E11
Murine
DEN
V1ndash4
EDIII(str
andA308
312andstrand
G387
389391)
Hybrid
oma
IgG2a120581
IC50
(120583gmL)
DEN
V1016
DEN
V2013
DEN
V-38
DEN
V-415
[40]
4G2
Murine
DEN
V1ndash4
EDII(fu
sionloop
)Hybrid
omafrom
DEN
V2Eim
mun
ized
mice
IgG
PRNT5
0(120583gmL)
DEN
V215
[39]
9F12
Murine
DEN
V1ndash4WNV
EDIII(aa
305307310
330Astr
andandBC
loop
)
Hybrid
omafrom
BALB
cmou
seim
mun
ized
with
DEN
V-2EDIII
IgG1k
DEN
V1ndash4P
RNT5
02sdot10minus8ndash2sdot10minus7M
[41]
2A10G6
Murine
DEN
V1ndash4YF
VWNV
JEV
TBEV
EDII(Fusionloop
aa
98ndash101)
Hybrid
omafrom
BALB
cmou
seim
mun
ized
with
inactiv
ated
DEN
V2
IgG1
PRNT5
0(120583gmL)
DEN
V12
DEN
V215
DEN
V321
DEN
V418
YNF36
WNV
46
[36]
mAb
11Hum
anDEN
V1ndash4WNV
SLEV
YFV
JEV
MVEV
EDII(fu
sionloop
)Ph
aged
isplayof
human
scFv
Fusio
nprotein
scFv-Fc
PRNT8
0(120583gmL)
WNV
125
DEN
V2625
[4243]
BioMed Research International 7Ta
ble1Con
tinued
mAb
Orig
inRe
activ
ityTargetprotein
Epito
peClon
ingstr
ategy
Form
atisotype
Neutralizingactiv
ityRe
ference
E16
(MGAW
N1)
Murine
WNV
EDIII(LR
aa3
02ndash309)
Hybrid
omafrom
immun
ized
micew
ithWNVE
IgG2bhum
anized
PRNT5
04ndash
18ng
PRNT9
053ndash297
ng[4445]
1A1D
-2Murine
DEN
V1ndash3
EDIII(Astr
andaa
307
310and312)
Hybrid
omafrom
immun
ized
micew
ithdifferent
pH-tr
eated
virus
IgG2a
DEN
V2PR
NT5
021n
M[4647]
1F4
Hum
an
DEN
V1
E(virion
)
DI-DII
Electro
fusio
nof
infected
mem
oryBcells
from
DEN
V-im
mun
esub
jects
with
EBV
IgG
DEN
V1IC5
0011120583gmL
[48]
2D22
DEN
V2
DIII
DEN
V2IC50
008120583gmL
5J7
DEN
V3
DI-DII
DEN
V3IC50
010120583gmL
E105
Murine
DEN
V1
E
DIII(BC
loop
G328
T329
andD330DE
loop
K361E
and
E362K
FGloop
K3
85)
Hybrid
omas
deriv
edfro
mC5
7BL6
IFN-120572120573Rminus
minusmice
infected
with
DEN
V1
IgG
PRNT5
0DEN
V-1
05ndash592ng
mL
[32]
E106
DIII(BC
loop
G328
T329
andD330DE
loop
K361E
and
E362K
FGloop
K3
85A
strand
S305
K307E
309K3
10and
E311)
PRNT5
0DEN
V-1
06ndash
592ng
mL
E111
DIII(CC1015840loop
)PR
NT5
038ndash25120583gmL
[49]
CR4374
Hum
anWNV
EDIII
Phaged
isplayof
scFv
IgGlib
rary
VH2-05V
L1e
PRNT5
0018120583gmL
PRNT9
0095120583gmL
[50]
CR4353
VH3-30V
k3-A27
PRNT5
00026120583
gmL
PRNT9
0364120583gmL
1A5
Chim
panzee
DEN
V1ndash4WNV
JEV
LGTV
EDII(aaG
106)
Phaged
isplayof
Fab
library
from
DEN
V1ndash4
infected
chim
panzees
Hum
anized
IgG1
PRNT5
0(120583gmL)
DEN
V1048
DEN
V2095
DEN
V332
DEN
V443
WNVD
ENV438
JEV
21LG
TV28
[51]
8 BioMed Research International
Table1Con
tinued
mAb
Orig
inRe
activ
ityTargetprotein
Epito
peClon
ingstr
ategy
Form
atisotype
Neutralizingactiv
ityRe
ference
mAb
11Hum
anDEN
V1ndash4WNV
EDII(fu
sionloop
W101G104G106)
scFv
library
scFv-Fc
PRNT8
0(120583gmL)
DEN
V2625
WNV
125
[4243]
3B4C
-4
Murine
SLEV
E
1a
Hybrid
oma
IgG
SLEV
PRNTlt17
[5253]
1B2C
-5SLEV
1bSLEV
PRNTlt17
6B5A
-2SLEV
1cSLEV
PRNT
48
4A4C
-4SLEV
1dSLEV
PRNT
29
1B5D
-1SLEV
JEV
2SLEV
PRNTlt17
2B5B
-3SL
EVJEV
MVEV
WNV
YFV
3SLEV
PRNT
23
2B6B
-2AllFlaviviru
s4a
SLEV
PRNTlt17
6B6C
-1AllFlaviviru
s4b
(inDII)
SLEV
PRNT
23
A3
Chim
panzee
JEV
E2
DIa
aK1
79Ph
aged
isplay
Hum
anized
PRNT5
0004ndash0
2nM
[54]
B2DIIaaI126
Phaged
isplay
PRNT5
0002ndash2
nM
E3DIIIa
aG132
Phaged
isplay
PRNT5
0014ndash0
93n
M
FabT
JE12B0
2Hum
anJEV
ENA
Phaged
isplay
Fab
FRNT5
0502120583gmL
[55]
5F10
Hum
anCH
IKV
E2Dom
ainB
EBVtransfo
rmation
IgG11205822
IC50lt100n
gmL
[56]
8B10
Hum
anCH
IKV
E1-E2
E2Dom
ainA
EBVtransfo
rmation
IgG1k
IC50lt100n
gmL
CHK-
152
Murine
CHIK
VE2
aa59
Hybrid
oma
IgG2c
IC50
1ndash3n
gmL
[57]
11E7
Murine
CCHFV
Gn
C-ter
Hybrid
oma
IgG
PRNT8
0diluted
12560
[58]
4-39-C
CMurine
RVFV
G2
Dom
ainIV
Hybrid
oma
IgG
PRNT8
0diluted
1204
80ndash181920
[59]
2C9
Murine
YFV
EHybrid
oma
IgG2a
PRNT9
011
0417D
1105
2Asib
i[60]
5A 7A R3(27)
Hum
anYF
VE
DI-DII
Phaged
isplay
ScFv
PRNT9
001ndash03120583
gmL
[61]
13D6
Murine
TBEV
EDIII
Hybrid
oma
IgGchimeric
PRNT5
019120583gmL
FRNT5
045120583
gmL
IC50
19ndash167120583
gmL
[62]
3B4C
-4(H
y4-26C
)Murine
VEE
VE2
aa182ndash209
Hybrid
oma
IgGhum
anized
PRNT7
0(120583gmL)
394ndash125
[63]
BioMed Research International 9
The structure of the 1A1D-2 bound to EDIII indicates that themAb binds to sites that are transiently exposed during viralldquobreathingrdquo at 37∘C and block infection during attachmentof the virion to the cell In particular this mAb neutralizesDENV1 2 and 3 serotypes and residues K307 and K310 arethemost critical residues for binding of 1A1D-2mAb [46 75]
Finally Shrestha et al immunized mice with a genotype2 strain of DENV1 virus and generated 79 new mAbs 16of which strongly inhibited infection by the homologousvirus and localized to DIII of E protein Surprisingly onlytwo mAbs E105 and E106 retained strong binding andneutralizing activity against all five DENV1 genotypes as wellas being protective in immunocompromised infected miceMoreover E105 and E106 exhibited therapeutic activity evenwhen administered as a single dose four days after inoculationwith a heterologous genotype 4 strain of DENV1 in the samemouse model [32]
243 Other Non-HumanmAbs Other non-human and non-murine as well as humanized mAbs directed against the Eglycoprotein have been described In this regard Goncalvezet al isolated a large panel of anti-E Fab fragments from chim-panzees infected with all four DENV serotypes Howeveronly a limited number of them displayed a cross-neutralizingactivity against DENV1 and 2 and to a lesser extent againstDENV3 and 4 In particular the authors calculated thatamong them the 1A5 mAb that was further humanizedneutralized DENV1ndash4 at a PRNT50 titer of 048 095 32and 43 120583gmL respectively Interestingly the humanized 1A5was also tested for binding and neutralization against theWNVDENV4 chimera JEV strain SA14-14-2 and Langatvirus (LGTV) strain TP 21 giving a PRNT50 titer of 38 21and 28120583gmL respectively Moreover the authors calculatedthat when administered in a dose of 2mg per kg of bodyweight 1A5 would give a serum titer of approximately 40and 20 50 reduction in plaque reduction neutralization test(PRNT50) against DENV1 and DENV2 respectively [76]Furthermore epitope mapping of 1A5 mAb localized to G106within the Flavivirus-conserved fusion loop inDII of DENV2E protein [51]
244 Anti-DENV Human mAbs As anticipated attemptsto isolate neutralizing human mAbs have been morechallenging due to the restricted elicited Ab-repertoire whichrecognizes DIII of E glycoprotein in naıve as well as ininfected patients [77 78] However several groups reportedthe isolation of neutralizing mAbs from infected subjects Inthis regard Setthapramote et al isolated a total of 136 humanhybridoma clones producing specific mAbs against DENVobtained using PBMCs from nine blood samples from fouracute-phase patients secondarily infected with DENV2 andfive convalescent-phase patients Interestingly the authorsfound that most of the acute-phase mAb clones were cross-reactive with all four DENV serotypes with most of themrecognizing the E protein and endowing of neutralizingactivity against all DENV serotypes compared to thosederived from convalescent-phase patients In particularfrom the acute-phase PBMCs 818 were anti-E 66 wereanti-prM and 33 were anti-NS1 while 133 anti-E 133
anti-prM and 533 anti-NS1 mAbs clones were obtainedfrom convalescent-phase PBMCs [79] Previous studiesconfirmed the data obtained by Setthapramote et al onconvalescent-phase patients but this group firstly reportedthe efficient preparation of human mAbs with strongneutralizing activity titers against all four DENV serotypesusing PBMCs from acute-phase patients secondarily infectedwith DENV [80 81]
The group of de Alwis through immunoglobulin deple-tion studies reported that a substantial fraction of DENV-reactive Abs in human immune sera including type-specificneutralizing Abs bound to the intact virion but not torecombinant E proteinThe authors confirmed these observa-tions also isolating human neutralizing mAbs and proposedthat humans produce Abs that neutralize DENV infectionby binding a complex quaternary structure epitope that isexpressed only when E proteins are assembled on a virusparticle [48] Similar findings have been described for severalother viruses including WNV and HIV [82 83] Mappingstudies indicate that this epitope has a footprint that spansadjacent E protein dimers and includes residues at the hingebetween domains I and II of E protein [48]
Recently Costin et al isolated three broadly neutralizinganti-DENVhumanmAbs named 48AD11C and 16DThesemAbswere isolated from three different convalescent patientswith distinct histories of DENV infection All three mAbsrecognized the E glycoprotein with high affinity neutralizedall four serotypes of DENV but mediated ADE of infection inFc receptor-bearing cells at subneutralizing concentrationsMapping studies revealed that all three mAbs bind discontin-uous epitopes within the highly conserved fusion-loop regionof DII (contacting residues W101 L107 andor G109) [37]
25 Yellow Fever Virus (YFV) TheWHO estimates that thereare 200000 cases of yellow fever virus (YFV) infections and30000 related deaths every year especially in Sub-SaharanAfrica In fact different to themajority of human arbovirusesand similarly to DENV and CHIKV YFV has expanded itshost range to include humans as an amplifying host [1]Symptoms occurring from 3 to 6 days after infection withYFV consist mostly in fever chills anorexia lumbosacralpain nausea and vomiting This syndrome lasts about 3days in most cases and is sometimes followed by a one-daylasting period of remission Fever will reoccur together withdark hematemesismelena petechiae and other hemorrhagicsymptoms Convalescence is characterized by deep asthenialasting up to two weeks Rarely prognosis can be fatal Aneffective live-attenuated vaccine (17D) derived from the Asibistrain by serial passages in chicken embryos is available [84]
251 mAbs against YFV In 1983 Schlesinger et al produceda battery of mAbs after immunization of BALBc mice byinjection of 17D YFV vaccine strain [84] Among them thespecificity towards the E and NS1 proteins was describedeven though anti-NS1 Abs were not able to show neutralizingactivity The 13 IgG and 1 IgM anti-E protein mAbs wereclassified in five groups according to their specificity GroupA consisting of the only IgM produced in this experiment
10 BioMed Research International
could neutralize 17D YFV virus only group B 17D and Asibistrains only group C Asibi strain only group D could onlyneutralize Asibi but not 17D strain showing also reactivityagainst DENV2 Zika or Banzi viruses All of the four mAbsbelonging to this group were able to cross-react with Zikaandor Banzi viruses and two (4E11 and 5H3) neutralizedDENV2 virus with 4E11 neutralizing also Banzi virus The3E9 mAb the only component of the group E could notneutralize 17D nor Asibi strains
All of the IgG mAbs resulted able to protect bothprophylactically and therapeutically BALBc or CD-1 micefrom lethal intracerebral challenge with 17D-204 strain [85]The chimeric form of the group B mAb 2C9 IgG (2C9-cIgG) was able to provide AG129 mice a 72 survival whenadministered 24 hours before infection with 17D-204 strainIts murine form could provide the survival of the 95 ofthe mice Appreciable results were also obtained when bothmurine 2C9 and 2C9-cIgG were administered 24 hours post-infection with the survival of 70 and 20 of mice respec-tively Viral presence was not detected in survivingmice [60]
In 2005 Daffis and colleagues constructed two Ab-phage libraries by cloning the repertoire of YFV-infectedpatients Panning was then performed with YFV-17D virionsThe scFv-5A 7A and R3(27) showed a neutralizing activityspanning from50 to 100 inPRNTassays against bothYFV17D-204-WHO and Asibi strains In further tests reactivitywas observed against wild-type YFV strains of West Africagenotype I and II (Nigeria 1987 and Asibi strains resp)and EastCentral Africa (CAR 1986 Ethiopia 1961 strains) Aconcentration from 01 to 03120583gmL could yield a 90 plaquereduction Reactivity was also observed against the strainSenegal 1990 in a lesser extent Production of escape variantscould demonstrate that scFv-5A 7A and R3(27) epitopes arebuilt up by residues extensively separated in themonomers ofE glycoprotein that however result in closed proximity whenthe homodimeric form of E is constituted [61]
26 West Nile Virus (WNV) The West Nile virus (WNV) isan epidemic neurotropic virus estimated to be responsibleof about 36000 cases and 1500 deaths registered in theUnited States between 1992 and 2012 [86] WNV antigenicityallows its classification into the Japanese encephalitis virus(JEV) serocomplex Genomic analysis has revealed two maingenetic lineages of WNV lineage I viruses circulating in theUSA Europe the Middle East Africa India and Australiaand lineage II viruses isolated from Sub-Saharan Africa andMadagascar [1]
WNV was firstly identified in 1937 and is endemicin many countries of Africa Middle East and West AsiaHowever after 1990 frequent outbreaks of WNV infectionswere reported in Romania Israel and later in North Americaand across the USA in 1999 and recently in 2012 [87]
Most of WNV infected individuals do not develop symp-toms while about 20 develop a self-limiting illness calledWest Nile fever Acute symptoms include fever tirednessand swollen lymph glands but in a minority of cases alsoencephalitis with long-term deficits in cognitive functionand motor skills have been reported However in less than
1 of infected patients WNV having crossed the blood-brain barrier is responsible of neuroinvasive and potentiallylethal form of the disease In these cases degeneration andapoptosis upon infection of neurons and the consequentinflammatory response can occur Related symptoms includehigh fever coma muscle weakness and paralysis Immun-odepression and advanced age have been correlated with anaugment risk to develop a severe disease [87] There are nospecific treatment options or licensed vaccines for humans[88]
WNV is transmitted by infected mosquitos and initialreplication is thought to occur primarily in dendritic cells inthe skin which migrate to secondary lymphoid tissues wherethe replicating virus enters the circulation To date onlyefficacious WNV veterinarian vaccines have been licensedwhile there are no licensed vaccines for protection againstWNV in humans [88]
261 mAbs against WNV Broad-spectrum antivirals suchas type I IFN-120572 ribavirin mycophenolic acid in WNVinfection showed ineffective results in vivo despite in vitrothey showed some activity [89]
However experiments in murine models extrapolationof clinical data as well as passive administration of pooledimmune-120574-globulins (OmriGam) containing a significanttiter of neutralizing Abs before and after infection showedan important role of the humoral immune response incontrolling viremia and prevent viral dissemination Indeedthe development of a neutralizing mAb-based therapy seemsto be encouraging for a possible treatment of infected patients[90ndash92]
Similarly to what has been observed for DENV only anti-E Abs have been identified as neutralizing and protectiveAlso anti-NS1 Abs have shown a protective role howevertheir mechanism of protection has not yet been elucidated asthe NS1 protein is secreted from infected cells and not presenton the virions [93ndash95]
In this regard similarly to what has been describedfor DENV studies performed using a naıve human scFvlibrary for panning with purified WNV E protein supportthe hypothesis that no Abs against the neutralizing DIII canbe isolated However DIII specific Abs were isolated in asubsequent study using immunoglobulin libraries obtainedfrom threeWNV infected patients for biopanning on purifiedinactivated virus virus-like particles consisting of prM andE proteins or recombinant E glycoprotein Although theproportion of DIII-specific mAbs was low (8) comparedwith anti-DII mAbs (47) Two out of the four anti-DIIImAbs were potently neutralizing and protective in vivowhereas only three out of the 24 anti-DII were weaklyneutralizing in vitro and non-protective in vivo In particularresidues that are critical for neutralization lies on the regionsspanning amino acids 305ndash312 330ndash333 and 365 that arelocated on adjacent exposed loops ofDIIIHowever sequencealignment of E protein of different flaviviruses such as DENVand JEV revealed a considerable variation compared tothe whole E protein This observation further suggests thatdifferently from broadly cross-reactive anti-DII Abs anti-DIII neutralizing Abs are virus-type specific However of
BioMed Research International 11
these mAbs two (CR4374 and CR4353) protected mice fromlethal WNV challenge at 50 protective doses of 129 and357 120583gkg of body weight respectively [50]
Sanchez et al using three different immunization strate-gies (ie inactivated virus naked DNA and recombinantprotein) isolated ninemurinemAbsmost of which bound toconformation-dependent epitopes in DIII of the E protein Inparticular neutralizing mAbs named 8B10 11C2 10C5 and17C8 were obtained from mice immunized with inactivatedvirus alone or in combination with a DNA plasmid andbound to the same region of DIII with high affinity Incontrast mAbs obtained by immunization with a solubleversion of the E glycoprotein did not exhibit neutralizingactivity These non-neutralizing mAbs were cross-reactivewith several other flaviviruses including SLEV JEV YFVand Powassan virus confirming the conserved nature ofFlavivirus non-neutralizing epitopes [96]
Gould et al isolated 11 unique human single-chain vari-able region Ab fragments (scFvs) that bind the E protein ofWNVAmong them a humanmAb namedmAb11 expressedas a scFv-Fc fusion protein was further characterized Itrecognizes the fusion loop at the distal end of DII ofthe WNV E protein and cross-reacts with all four DENVserotypes and provides protection against DENV2 and 4as well as WNV [42] Moreover therapeutic studies of thismAb in WNV-infection model mice provided substantialprotection when administered after 5 days post-infectionInterestingly a neutralization escape variant of this mAbfailed to cause lethal encephalitis (at higher infectious doses)or induce the inflammatory responses associated with blood-brain barrier permeability in mice compared to the parentalWNV suggesting an important role for the fusion loop inviral pathogenesis [43]
Oliphant et al isolated an anti-DIII E protein mAbnamed E16 fromhybridomas obtained after immunization ofmice with recombinant WNV E protein which neutralizedall WNV strains with PRNT50 values of 4 to 18 ng andPRNT90 values of 53 to 297 ng One hundred microgramsof mAb protected greater than 90 of mice from lethalinfection and even a single 4120583g treatment of E16 on day 2 afterinfection prevented mortality Moreover humanization ofthis mAb confirmed as therapeutically effective in mice [44]Subsequent studies revealed that mAb E16 neutralization ismediated by engagement of four discontinuous segmentsof DIII including the amino-terminal region (amino acidresidues 302ndash309) and the three connecting loops BC (aminoacid residues 330ndash333) DE (amino acid residues 365ndash368)and FG (amino acid residues 389ndash391) Moreover no ADE ofinfection was detected when E16 mAb was used at saturatingconcentrations [45 47]
Furthermore results of a Phase I safety study of thehumanized E16 mAb (designated MGAWN1) have beenreported and suggested that doses of up to 30mgkgwerewelltolerated with few mild adverse events and would providean excess of virus neutralizing activity for 3-4 weeks aftertreatment However a case of anti-MGAWN1 Ab elicitationoccurred with the consequent increased rate of clearance andindeed impacting efficacy A Phase II safety and efficacy studyof MGAWN1 is ongoing [97] Furthermore the possibility of
preventing or treating WNV-induced memory deficits wasrecently investigated In this study hamsters were treatedintraperitoneally with 32mgkg ofMGAWN1mAb at 45 daysafter subcutaneously challenging with WNV InterestinglyMGAWN1 prevented mortality weight loss and improvedfood consumption of WNV-infected hamsters compared tocontrols [98]
Recently Lelli et al isolated six anti-E mAbs frominactivated-WNV immunized mice In particular three ofthem (3B2 3D6 and 4D3) neutralized lineage I and II WNVwith the first two recognizing the same epitopes located onthe distal lateral surface of DIII (critical amino acid residueK307) Conversely 4D3mAb recognized a novel neutralizingepitope on DII (critical residues S276 and T278) Indeedfurther protective and therapeutic studies are needed toascertain their neutralizing activity in vivo [99]
Finally to conclude like DENV infection Abs directedagainst the M protein are not protective and neutralizingMoreover in humans a skewed humoral immune responseagainst DII has been frequently observed and confirmedalso with the hybridoma technology [50] The isolation andelicitation of neutralizingAbs directed against the fusion loopand DIII of the E protein represent thus the most challengingand promising goal for the development of new effectivetherapeutic inhibitors and immunogens respectively
27 Tick-Borne Encephalitis Virus (TBEV) Tick-borne ence-phalitis virus (TBEV) is one of the most dangerous agentscausing human neuroinfections occurring mostly in Europeand Asia and with a potential fatal prognosis [100] TBEV isbelieved to cause 3500ndash10000 human cases of encephalitisin Europe per year with a high morbidity in Russia CzechRepublic Austria and Germany In particular between 1990and 2007 an average of 8755 cases of TBE was reported peryear in Europe and Russia Despite the fact that Russia isthe country with most infections registered annually CzechRepublic incidence is among the highest in Europe with 400ndash1000 clinical cases reported every year [101]
Three subtypes of TBEV are classified namely EuropeanSiberian and Far-Eastern sharing most of the genetic andantigenic features [102] In fact a high degree of antigenichomogeneity between different strains of TBEV has beendescribed [103]
Clinical manifestations vary among the subtypes butusually they start with a short febrile period of 7ndash14 daysafter the tick bite Fatigue headache and pain in the neckshoulders and lower back together with high fever andvomitingmay be present [100]Thesemanifestations are oftenfollowed by an asymptomatic phase lasting from 2 to 10days after remission from the fever with possible progres-sion to neurological disease Neurological symptoms includemeningitis encephalitis myelitis and radiculitis Mortalityoccurs in 1-2 of the European subtype-infected patientsbut fatal prognosis can occur in up to 20ndash40 of the Far-Eastern subtype-infected patients [104] Mortality rates ofthe Siberian subtype are similar to those observed for theEuropean [105]
Effective and safe vaccines against TBEV produced frominactivated virus have been developed and licensed for their
12 BioMed Research International
use in humans However an emergency therapy in theabsence of a mass immunization is needed as no effectivetreatments are yet available [106]
271 mAbs against TBEV Levanov et al described the chim-erization of two murine mAbs (13D6 and 10C2) directedagainst the DIII and DII respectively of the TBEV E glyco-protein The chimeric mAbs present binding characteristicssimilar to the parentalmAbsMoreover as the parentalmAbsonly the chimeric mAb 13D6 was able to neutralize TBEVinfectivity in vitro In particular neutralization studies withthe murine chimeric and scFv forms of mAb 13D6 were per-formed In particular murine 13D6 showed an IC50 titer of115 120583gmL in Focus Reduction Neutralization Tests (FRNT)against TBEV strain 205 and of 29 120583gmL in PRNT againstTBEV strain Softjin Chimeric 13D6 showed an IC50 titer of45 120583gmL in FRNT against TBEV strain 205 and of 19 120583gmLin PRNT against TBEV strain Softjin ScFv 13D6 showed anIC50 titer of 167 120583gmL in FRNT against TBEV strain 205and of 112 120583gmL in PRNT against TBEV strain Softjin [62]
28 Japanese Encephalitis Virus (JEV) Japanese encephalitisvirus (JEV) is a mosquito-borne flavivirus that over thepast few decades has caused several outbreaks throughoutChina Southeast Asia Australia and Papua New Guineawith a prevalence recently estimated to be of about 70000casesyear [107ndash109] A 40 mortality was recorded in someof the JEV-affected areas Moreover many survivors facesome neurological problems and complications [108] Since1995 the disease has also emerged in Non-Asian regions suchas Northern Australia [110 111] The situation in SoutheastAsia however is further complicated by the overlappingepidemics of JEV and DENV as well as sporadic casesof WNV infections detected in some of the affected areasparticularly in India [112]
Effective vaccines both live-attenuated and inactivatedstrains of JEV have been developed and licensed in majorcountries However as for the other arthropod-borne Fla-vivirus members no specific antiviral drugs are currentlyavailable [4]
281mAbs against JEV Aspreviously described for the otherFlavivirusmembers many groups isolated and characterizedanti-JEVmAbs showing different neutralizing and protectiveproperties In this regard Gupta et al used combinations ofanti-E JEV mAbs (Hs1-4) in mice protection experimentsIn particular they found that the singularly mAb protectionranged from 45 to 65 when 100120583g of mAb were adminis-tered while equimolar combinations of two or three mAbsgave 85ndash90 or 100 protection respectively [113] In similarexperiments Lee et al demonstrated that a Flavivirus anti-NS1 mAb named 16NS1 cross-reacted with JEV as well asWNV and exhibited protective activity against WNV as wellas a lethal JEV infection However no neutralizing activitywas observed using this mAb against both WNV and JEV inin vitro experiments suggesting the participation of otherAb-mediatedmechanisms in vivo In particular 95ofmicewere
protected when 500120583g of mAb were administered intraperi-toneally and concomitantly to intramuscular injection of JEV
Overlapping peptide mapping analysis combined withsite-specific mutations identified the 116KAWGKSILFA125region and critical amino acid residues W118 and I122 as16NS1 mAb epitope highly conserved in WNV and JEVstrains [114]
Arakawa et al isolated from a combinatorial human Fablibrary constructed from peripheral blood lymphocytesobtained from JEV hyperimmune volunteers Among 188randomly selected clones FabTJE12B02 showed the best 50focus reduction endpoint at the concentration of 502 120583gmLagainst the JEV strain Nakayama [55]
Goncalvez et al isolated three mAbs named Fabs A3B2 and E3 by repertoire cloning from chimpanzees initiallyimmunized with inactivated JE-VAX and then boosted withattenuated JEV SA14-14-2 In particular these mAbs reactedwith epitopes in three different E domains in DI (amino acidresidue K179) in DII (I126) and in DIII (G132) for Fabs A3B2 and E3 respectively Moreover these Fabs as well as thederived humanized counterpart mAbs exhibited high neu-tralizing activities against a broad spectrum of JEV genotypestrains Moreover these mAbs exhibited a 50 protectivedose of 084 120583g (B2) 58120583g (A3) and 247120583g (E3) in mousemodels Finally administration of 200 120583gmouse of mAb B2one day after otherwise lethal JEV infection protected 50of mice and significantly prolonged the average survival timecompared to that of mice in the unprotected group [54]
29 St Louis Encephalitis Virus (SLEV) St Louis encephalitisvirus (SLEV) was first discovered as the mosquito-borneagent responsible for over 1000 cases of encephalitis duringa 1933 summer outbreak in St Louis (Missouri) and nowis a reemerging human pathogen widely distributed in theAmerican continent causing several human encephalitisoutbreaks over the last 80 years Additional epidemics haveindeed occurred from 1964 to 2006 in the Americas rangingfrom the US to Argentina and Brazil As a member of theFlavivirus genus the SLEVE glycoprotein ectodomain is 68identical to serocomplex-related JEVEbut only 46 and 40identical to those ofDENV2 andTBEV viruses fromdifferentserocomplexes [115]
291 mAbs against SLEV In 1983 Roehrig et al isolatedtwenty-one hybridomas producing murine mAbs specificfor the E glycoprotein of SLEV strain MSI-7 Serologicreactivities were initially determined by cross-reactivity indi-rect immunofluorescence assays using 22 strains of SLEVand 8 other related flaviviruses Four groups demonstratingtype- subcomplex- supercomplex- and group-specific reac-tivity patterns were identified Analysis of hemagglutination-inhibition and virus neutralization subdivided the cross-reactivity groups into eight epitopes (E-1a b c d E-2 E-3 and E-4a b) one of them following localized in DII (4b)[52 53]
Moreover among the previously described isolated anti-WNV mAbs by Gould et al the chimeric scFv-Fc 79 effec-tively neutralized also SLEV resulting in gt80 of PRNT80when used at 5 120583gmL [42]
BioMed Research International 13
3 Alphaviruses
Major human arboviruses in the Togaviridae family Chikun-gunya (CHIKV) and Venezuelan equine encephalomyelitis(VEEV) viruses belong to the Alphavirus genus which iscomposed of viruses with an icosahedral nucleocapsid sur-rounded by a lipid envelope and glycoprotein spikes Thestructural proteins of alphaviruses arise through co- andpost-translational processing of a polyprotein encoded by asingle positive-stranded RNAproducing the capsid (C) PE26 K and E1 The PE2 glycoprotein is a precursor containingthe E3 glycoprotein fused to the amino terminus of the E2envelope glycoprotein The PE2 glycoprotein is followed by6K a small membrane-associated protein and E1 the secondpolypeptide component of glycoprotein spikes
Trimerized heterodimers of the E1 and E2 viral glyco-proteins form the surface spikes and contain determinantsof viral tropism and virulence The E3 glycoprotein acts as asignal for transport of PE2 across themembranes of the roughendoplasmic reticulum and may promote the formation andintracellular transport of E1-PE2 heterodimers to the cellsurface During transport to the cell surface PE2 undergoesa maturational cleavage event by a furin-like protease toproduce E2 and E3 [116]
The E2 glycoprotein promotes specificity of virus bindingto the host cell surface and is a target of the humoral immuneresponse The E1 glycoprotein mediates fusion of the virionenvelope with the membranes of acidified endosomes allow-ing release of the nucleocapsid into the cytoplasm and theonset of viral replicationAlphavirus E1 shares no appreciablesequence identity to Flavivirus E protein Despite differencesin their amino acid sequences and arrangements on the viralparticle the structures of E and E1 are remarkably similarIndeed the conservation of three domains has been welldocumented in crystal structures of flavivirus E ectodomainsfrom TBEV DENV JEV and WNV as well as Alphavirus E1from Semliki Forest virus (SFV) and CHIKV Abs to the E1glycoprotein do not typically neutralize virus infectivity invitro but can protect against lethal challenge in animals
By analogy with the prM protein of dengue Flavivirusfurin cleavage of PE2 may begin after transit of an acidiclate component of the Golgi body where E3 is thought tosuppress the acid pH-triggered activation of glycoprotein E1fusion capability Protective anti-E3 mouse mAbs have alsobeen described [117]
31 Chikungunya Virus (CHIKV) The chikungunya virus(CHIKV) belongs to the Semliki Forest clade and was firstlyisolated in 1953 in Tanzania during an epidemic outbreakfollowing occurred also in Asia and Africa In the last fivedecades several CHIKV outbreaks have been described bothin Africa and Asia separated by gaps lasting from two totwenty years In 2005-2006 about 300000 cases out of785000 inhabitants were reported in La Reunion island witha fatal prognosis for 237 of the patients [117] Neither Europenor the Americas have had outbreaks of CHIKV so far exceptfor imported isolated cases Three genotypes of CHIKV aredescribed Asian EastCentralSouth African (ECSA) andWest African with an amino acid identity spanning from
952 to 998 Recent epidemics in Africa and Indiansubcontinent were caused by strains belonging to the ECSAgenotype Transmitted by Aedes mosquitoes CHIKV ismaintained in the human population by human-mosquito-human transmission [1] The disease is characterized bydengue-like symptoms such as chills high fever headacheand persistent myalgia and a further incapacitating arthralgia(from which the name chikungunya) which affects 40 ofinfected subjects However prognosis is rarely fatal [116]
There is currently no commercial vaccine and antiviraltreatment forCHIKV although some candidate vaccines havebeen tested in humans In this regard in 2000 US Armyperformed a Phase II clinical trial testing a live-attenuatedCHIKV vaccine (TSI-GSD-218) derived from a 1962 strain(15561) of an outbreak in Thailand Out of 58 every patientdeveloped neutralizingAbs and 5 lamentedmild tomoderatejoint pain [117] A phase III trial of this candidate vaccineis ongoing Furthermore a new formulation using virus-likeparticles was able to induce neutralizing Abs in macaquesagainst different CHIKV strains [117 118] Indeed it has beendescribed that infection seems to elicit long-lasting protectiveimmunity and cross-protection among CHIKV and otheralphaviruses
CHIKV entry into host cell is demonstrated to be medi-ated by envelope glycoproteins E1 and E2 which allowvirus fusion to cell membrane in low pH conditions andrecognition of an unknown cellular receptor respectively[116 119ndash121] Even though the genome replication relies onan error-prone RNA-dependent RNA-polymerasis CHIKVstrains S27 and 05115 Reunion showed in E1 and E2 alow amino acid variation of 068 and 33 respectivelyand such a low variability may be needed for an effectivereplication in two phylogenetically distant hosts [116 122]Recently the crystal structure of E1 and E2 heterodimerhas been resolved both at low pH and natural conditionsshedding light on E1 and E2 ectodomainsrsquo structure Inparticular similarly to flaviviruses E protein E1 ectodomainis made of the N-terminal DI the DII containing the fusionloop and the DIII at the C-terminal On the other hand E2ectodomain comprises theN-terminal domainA the domainB supposed to interact with the host cellrsquos unknown receptorand domain C at the C-terminal E1 DIII and E2 domain Care located close to the viral membrane [123 124]
311 mAbs against CHIKV Warter et al described 5F10 and8B10 two human mAbs which strongly neutralized severalCHIKV isolates in vitro without cross-reactivity againstother alphaviruses but to Onyong-nyong virus [56] Mixedpreparation of 5F10 and 8F10 did not show neither synergisticnor addictive effect in vitro and studies upon escape mutantsdemonstrated that 5F10 mAb binds at the tip of the E2domain B while 8B10 recognizes residues close to E1 fusionloop and amino acids within E2 domain A which form atransitional epitope under low pH conditions The authorsspeculated that 5F10 and 8B10 may inhibit CHIKV entry andfusion to the cell membrane respectively It is worth notingthat the previously mentioned escape mutants displayedmutations associated with reduced viral fitness in vitro evenafter 13 neutralizationamplification rounds In the same
14 BioMed Research International
study cell-to-cell transmission was firstly demonstrated as anescapemechanism enhanced by the E2mutation R82G [122]
In further in vivo studies 5F10 and 8B10 significantlydelayed CHIKV-caused death of AGR129 mice both in pro-phylactic and therapeutic tests Interestingly in therapeutictreatment these mAbs showed a synergistic effect whenadministered in combination with the total amount of mAbsinjected being the same of the single-mAbs Possibly thesetreatment did not allow themice to survive CHIKV challengedue to Ig clearing and 6ndash10 days half-life usually described forhuman mAbs [125]
Recently Pal et al cloned thirty-six murine mAbs ableto neutralize the ECSA La Reunion 2006 OPY-1 strain ofCHIKV (CHIKV-LR) the majority of which also neutralizeinfection of other strains corresponding to the Asian andWest African genotypes In particularmAbCHK-152 showedthe highest and broadest neutralizing activity with FRNTvalues indicating an IC50 value of 1 to 3 ngmL dependingon the viral strain used Among the thirty-six mAbs four(CHK-102 CHK-152 and CHK-166 CHK-263) could pre-vent lethality of immunodeficient Ifnarminusminus C57BL6 micewhen administered one day before exposition with CHK-152 and CHK-263 showing the ability to protect mice at thelowest dose (10 120583g) Therapeutic studies were performed onthese mAbs administering a single dose of 100 120583g 24 hoursafter infection The highest activity was described for CHK-166 able to protect 63 of the mice Testing the combinedactivity of the mAbs in therapeutic studies administration ofCHK-166 plus CHK-152 in a dose of 250120583g each turned out tobe themost effective protecting 71 of the IfnarminusminusC57BL6mice 60 hours after infection [57]
32 Venezuelan Equine Encephalomyelitis Virus (VEEV)Venezuelan equine encephalomyelitis virus (VEEV) is main-tained in a natural transmission cycle between mosquitoesand small rodents The first documented outbreaks occurredin the 1930s and several epidemics have been reported so farin Latin-American countries such as Venezuela ColombiaPeru Ecuador Bolivia Honduras Mexico and Panama withhundreds of thousands of human cases being reported anda case-fatality rate up to 1 during the 1969 Ecuador out-break Clinical manifestations of VEE are indistinguishablefrom Dengue [126] Six serogroups (IndashVI) are currentlyrecognized within the VEEV complex VEEV caused humanand equine outbreaks in the Americas for nearly a centuryEquine epizootics have high mortality (38ndash83) leading toa high viremia followed by a lethal encephalitis and often tohuman epidemics involving thousands of cases and hundredsof deaths Indeed VEEV is infectious for humans by theairborne route and has been responsible for a number oflaboratory infections In humans the disease is usually self-limiting with a febrile illness with 1ndash4 of cases progressingto severe encephalitis It is a hazard to laboratory workershas been developed as a biological weapon and is a potentialbioterrorist agent [127] There are no antiviral drugs andvaccines licensed for the treatment of VEEV infection inhumans Indeed although experimental live-attenuated vac-cines have been developed (eg TC-83) with good levels ofprotection in equine and mice in humans they may fail to
give protection in the majority of cases [128] Consideringthe protective role exerted by humoral immune response inanimal models it could be concluded that treatment withspecific IgG may have a beneficial antiviral effect in humanairborne infections with pathogenic strains of VEEV
321 mAbs against VEEV Several works support that mAbsmay protect against airborne VEEV as well as after airborneexposure toVEEVAb reactivity to both surface glycoproteins(E1 and E2) is associated with neutralization of the virusin vitro and passive protection against virus challenge Aseries of anti-E1 and anti-E2 mouse and humanized mAbshave been described recently Among them Phillpotts et alexamined two anti-E2 murine mAbs named 1A4A-1 and1A3A-9 both having potent protective activity against subcu-taneous VEEV challenge in mice Both mAbs had a similarhalf-life (58 and 100 days) in mouse serum after a singleintraperitoneal dose suggesting that mAbs delivered by thisroute at or around the time of VEEV infection persists athigh levels in the blood as well as secretions in respiratorytransudation throughout the clinical course of the disease Inparticular both mAbs administered 24 h prior to airbornechallenges had a substantial protective effect (90ndash100)Treatment of mice 2 or 24 h after airborne infection with asingle intraperitoneal dose of 100 120583g of 1A3A-9 mAb led toapproximately 50 survival There was no beneficial effectwhen mAb treatment was delayed to 72 h Moreover therewas evidence of synergy in vitro in PRNT between 1A3A-9and 1A4A-1 as has been demonstrated for other virusesand mAb pairs However no synergy was found in mouseprotection when mAbs were delivered intraperitoneally as amixture in equal parts to animals challenged 24 h previouslywith VEEV [129]
In a subsequent study Phillpotts et al reported two otherprotective mAbs The 3B2A-9 mAb protected against allthe serogroup I strains while 1A3B-7 mAb protected wellagainst challenge with all of the viruses tested Both 3B2A-9 and 1A3B-7 protected against airborne exposure to theIAB serogroup virus strainTrinidad donkey (TrD)Howeverthere was no evidence of synergistic protection when thesemAbs were combined in equal proportions An intraperi-toneal dose of 10 120583g was sufficient to protect 50 of themice with either mAb [130] According to other data themechanism of protection did not appear to depend uponneutralization [131] Similar results were obtained by OrsquoBrienet al which described a non-neutralizing mAb (IgG2a)named CUF37-2a able to protect 50 of mice from asubcutaneous VEEV challenge when a dose of 915 120583g of mAbwas administered 24 h prior to challenge [132]
In a following study the 1A3B-7 mAb was humanized(and following reported as Hu1A3B-7) maintaining the samefeatures of its murine counterpart In particular evaluation ofin vitro studies indicated that Hu1A3B-7 retained both broadspecificity and neutralizing activity Furthermore in vivoexperiments showed that Hu1A3B-7 successfully protectedmice against lethal subcutaneous and aerosol challenges withVEEV strain TrD Moreover the effectiveness of the human-ization process was determined by assessing proliferationresponses in human T-cells to peptides derived from the
BioMed Research International 15
murine and humanized versions of the VH and VL domainsThis analysis showed that the number of human T-cellepitopes within the humanized Ab had been substantiallyreduced indicating that Hu1A3B-7 may have low immuno-genicity in vivo [133]
Similarly Hunt et al described a humanizedmurinemAbderived from the 3B4C-4 mAb recognizing the E2c epitope(amino acid residues 182ndash209) and following named Hy4-26C showing neutralizing activity similar to the murinemAb Moreover it was protective (70ndash100 of survival) at100 ng dose in mice animal model intraperitoneal inoculatedand at 500120583g for intranasal challenge (80) Therapeuticstudies in mice revealed that Hy4-26C was able to cure miceup to 24 h following infection at 10 120583g on the contrary themouse mAb when given 1 hour after virus challenge [63]
In another work Hunt et al isolated two neutralizinghumanized Fabs F5 and L1A7 employing a blocking strategy[134 135] The anti-E2 specific F5 IgG had a 70 PRNTendpoint of 10 ngmL equivalent to that described for themost effective neutralizing anti-VEEVE2mAbs In particularthe E1-specific hFab L1A7 had a PRNT endpoint of 3120583gmL300-fold lower than F5
Further studies revealed that F5 epitope is located inthe 115ndash199 amino acid region Moreover F5 IgG had potentability to protect mice from infection by either route whenadministered 24 h before exposure however mice treated24 h and 48 h after aerosol exposure developed central ner-vous system infections but exhibited no clinical signs ofdisease [135]
Hu et al reported that passive immunization with thehumanized chimericmousemAbhu1A4A1IgG1-2A inmice at50 120583g 24 h before or after virulent VEEV challenge providedcomplete protection indicating that hu1A4A1IgG1-2A haspotent prophylactic and therapeutic effects against VEEVinfection [136]
Anti-VEEV mAbs isolated from a non-human primategene library has also been reported In particular thehumanized chimeric scFv-Fc ToR67-3B4 recognized viableas well as formalin and 120573-propiolactone inactivated virusparticles It detected specifically the viral E1 envelope proteinof VEEV but did not react with reduced viral glycopro-tein preparations suggesting that recognition depends uponconformational epitopes The recombinant Ab was able todetect multiple VEEV subtypes and displayed only marginalcross-reactivity to otherAlphavirus species except for Easternequine encephalitis virus (EEEV) In addition the scFv-Fcfusion described here might be of therapeutic use since itsuccessfully inactivated VEEV in a murine disease modelIn particular when the recombinant Ab was administered 6hours after challenge 80 to 100 of mice survived lethalVEEV IAB or IE infection Forty to sixty percent of micesurvived when scFv-Fc ToR67-3B4 was applied 6 hours afterchallenge with VEEV subtypes II and former IIIA [137]
4 Bunyaviridae
TheBunyaviridae family contains human arboviruses belong-ing to the Orthobunyavirus Phlebovirus and Nairovirus
genera Bunyaviridae includes enveloped viruses with a frag-mented single-stranded RNA genome of negative polarityTheir tripartite genome consists of a small (S) a medium(M) and a large (L) fragment and the envelope glycoproteinsGn and Gc that are cleaved out of a polyprotein synthesizedby the M fragment These glycoproteins have been provedto mediate the formation of the virus particle to play a rolein the interaction with cell surface receptors to mediate theentry of the virus into cells and to serve as targets for themajority of neutralizing Abs described so far
Orthobunyavirus transmitted through mosquitoes ormidges vectors are divided in 18 serogroups based on cross-titrations in haemagglutination inhibition assays and neutral-ization assays and correlating with main vector preferencesHowever the most clinically important viruses belong onlyto two serogroups the California encephalitis and the Simbuserogroups
Crimean-Congo hemorrhagic fever virus (CCHFV) isconsidered the only one of clinical relevance within theNairovirus genus which uses tick as main vector and is divid-ed in seven serogroups while Toscana virus and Rift Valleyfever virus (RVFV) which are transmitted by sandflies andmosquitoes respectively belong to the Phlebovirus genus[138]
MAbs against theNairovirusCCHFV and the PhlebovirusRVFV have been described
41 Crimean-Congo Hemorrhagic Fever Virus (CCHFV)Crimean-Congo Hemorrhagic Fever Virus (CCHFV) dis-tribution covers the greatest geographic range of any tick-borne virus known as viral isolation andor disease hasbeen reported from more than 30 countries in SoutheasternEurope Africa Asia and Middle East [139 140] Namelyfrom 1953 to 2010 about 6000 human cases were reportedin Southeastern Europe CCHFV causes sporadic outbreakswith mortality rates ranging from 10 to 80 [141 142] Asignificant variation in the time of incubation has beendescribed [139] Prehemorrhagic symptoms include highfever chills headache photophobia and back and abdominalpains Among other symptoms neuropsychiatric changeshave been reported in some patients In severe cases 3ndash6 daysafter the onset of the disease hemorrhagic symptoms occurwith petechiae ecchymosis bleeding in the form of melenahematemesis epistaxis [139]
In comparisonwith the nucleocapsid proteins inCCHFVGn and Gc show a higher degree of antigenic variabilityprobably due to their exposition to the host immune systemhypothesis corroborated in the description by Hewson et alin 2004 of four M segment phylogenic groups namely M1M2 M3 and M4 [141 142]
411 mAbs against CCHFV Blackburn et al firstly describedin 1987 murine anti-CCHFV mAbs able to recognize nucle-ocapsid proteins but no neutralizing activity has beendescribed thus far [143]
On the other hand Bertolotti-Ciarlet et al produced apanel of murine mAbs recognizing conformational epitopes
16 BioMed Research International
within theGn andGc glycoproteins expressed by the CCHFVstrain IbAr10200 The effectiveness of the mAbs was studiedby performing PRNT80 and none of the anti-Gn mAbsshowed neutralizing activity although many of the anti-GcmAbs neutralized IbAr10200 strain in vitro with clones 8A15E3 12A9 6C2 and 9H3 showing activity at a gt5120 dilutionMAbs 11E7 and 30F7 exhibited neutralizing activity at 2560dilution A suckling mice protection test was thereforeperformed for these mAbs The previously mentionedanti-Gc Abs were capable to protect mice to an appreciabledegree when applied 24 hours before and in a weakermanner 24 hours after virus challenge Many of the anti-GnmAbs were able to confer significant protection to IbAr10200both 24 hours before and after virus administration Therelevant effectiveness of the anti-Gn mAbs 6B12 10E1113G8 and 10G4 suggests that these mAbs may possess someneutralizing activity due to Ab-based effector mechanisms(eg ADCC) [58]
In further studies the same group characterized thebroadly cross-reactive neutralizing activity of the previouslydescribed murine mAb 11E7 able to bind conformationalepitopes within the C-terminal region of the Gn glycoproteinof all CCHFVM groups [58 141]
42 Rift Valley Fever Virus (RVFV) Similarly to CCHFV RiftValley Fever Virus (RVFV) causes sporadic outbreaks Thelargest epidemic occurred in Egypt in 1977 with an estimated200000 infections 18000 patients manifesting symptomsand 600 fatal prognoses [144] Symptoms include hemor-rhagic episodes fever encephalitis and blindness [145]
The major target of the humoral immune response is theRVFV glycoproteins Gc (G1) and Gn (G2) ranging fromnucleotides 480ndash2090 and 2091ndash3614 respectively [144]
421 mAbs against RVFV Within G2 glycoprotein are des-cribed three neutralizing antigenic sites namely epitope Inucleotides 792ndash893 (amino acid residues 258ndash291) epitopeII nucleotides 1164ndash1196 (amino acid residues 382ndash392) andepitope IV nucleotides 858ndash917 (amino acid residues 280ndash299) [59 145]
Despite the high degree of variability observed inCCHFVglycoproteins RVFV G2 presented an unexpected high con-servation among 22 isolates [59] In the same study onlythe LUNYO and 900060 isolates were resistant to in vitroneutralization by murine mAb 4D4 (recognizing epitope II)All other isolates were neutralized with PRNT80 titers from10240 togt81920On the other hand only SNS isolate showedreduced sensitivity to 4ndash39-CC (epitope IV) All other isolateswere neutralized with PRNT80 titers from 20480 to gt81920Protection studies on mice would be of interest for thedevelopment of a passive immunization prophylaxis [59]
Protection studies from lethal South African RVFVAN1830 strain infection were successfully performed on abattery of anti-G1 and anti-G2 mAbs produced by Besselaarand Blackburn with the strongly neutralizing anti-G1 mAb3E5 and anti-G2mAb 9C4 being the most effective [146]The9C4 antigenic area maps within G2 epitope I and a broadlyneutralizing activity may be investigated
5 Conclusions
Arbovirus infections have acquired increasing interest giventhe augmented globalization and tourism movement all overthe world that could be at the basis of epidemic eventsFurthermore the same vectors can sometimes transmit sev-eral arboviruses concomitantly complicating the diagnosis aswell as the therapy For example confusing mixed epidemicshave occasionally been described such as YFV plus CHIKVDENVplus CHIKV ormore recentlyPlasmodium falciparummalaria plus DENV1 and CHIKV on Madagascarrsquos east coast[1]
In the last decade several efforts have been employed inthe development of effective immunogens and therapeuticsgiving the current absence of specific antiviral drugs as wellas effective vaccines for themost diffused arbovirus infections(ie DENV andWNV)
However at the moment mosquito control is the bestavailable method for preventing arbovirus infections Thusto control the emerging public health problem of arbovirusinfections new antiviral therapeutic strategies that providepotent and broadly cross-protective immunity (especially forflavivirus infections) are an urgent globally medical need
As described in this review the majority of highlyneutralizing mAbs are of murine origin and indeed theirutility in the treatment of these infections is limited In facttheir employment can elicit an immunogenic response (iehuman anti-mouse Abs HAMA) the therapeutic efficacycould be reduced by a relatively faster clearance in humans(compared to human Abs) potentially exacerbated by theHAMA response Finally murine Abs exhibit relatively weakeffector functions (eg ADCC) compared to human AbsIndeed chimerization humanization or better the isolationand characterization of fully human broadly neutralizingmAbs would be the best choice for their possible use in afuture mAb-based therapy against arbovirus infections [147]
Finally possible ADE of infection mechanisms should beevaluated before considering a mAb as a possible candidatetherapeutic in a post-exposure setting as well as in the devel-opment of new Ab-eliciting immunogens
Conflict of Interests
The authors declare no conflict of interests in particular withOmriGam and any other cited Ab-based preparation men-tioned in the paper
References
[1] N Cleton M Koopmans J Reimerink G J Godeke and CReusken ldquoCome fly with me review of clinically importantarboviruses for global travelersrdquo Journal of Clinical Virology vol55 no 3 pp 191ndash203 2012
[2] F X Heinz H Holzmann A Essl and M Kundi ldquoField effec-tiveness of vaccination against tick-borne encephalitisrdquoVaccinevol 25 no 43 pp 7559ndash7567 2007
[3] A D Barrett and D E Teuwen ldquoYellow fever vaccinemdashhowdoes it work and why do rare cases of serious adverse events
BioMed Research International 17
take placerdquo Current Opinion in Immunology vol 21 no 3 pp308ndash313 2009
[4] S B Halstead and S J Thomas ldquoNew vaccines for Japaneseencephalitisrdquo Current Infectious Disease Reports vol 12 no 3pp 174ndash180 2010
[5] R Burioni M Perotti N Mancini and M Clementi ldquoPer-spectives for the utilization of neutralizing human monoclonalantibodies as anti-HCV drugsrdquo Journal of Hepatology vol 49no 2 pp 299ndash300 2008
[6] R A Diotti G A Sautto L Solforosi NManciniM Clementiand R Burioni ldquoNeutralization activity and kinetics of twobroad-range human monoclonal IgG1 derived from recombi-nant Fab fragments and directed against Hepatitis C virus E2glycoproteinrdquo New Microbiologica vol 35 no 4 pp 475ndash4792012
[7] R Burioni F Canducci N Mancini et al ldquoMolecular cloningof the first humanmonoclona antibodies neutralizing with highpotency Swine-origin Influenza A pandemic virus (S-OIV)rdquoNew Microbiologica vol 32 no 4 pp 319ndash324 2009
[8] P K Chandra S Hazari B Poat et al ldquoIntracytoplasmic stableexpression of IgG1 antibody targeting NS3 helicase inhibitsreplication of highly efficient hepatitis C Virus 2a clonerdquoVirology Journal vol 7 article 118 2010
[9] N Clementi D de Marco N Mancini et al ldquoA human mono-clonal antibody with neutralizing activity against highly diver-gent influenza subtypesrdquo PLoS ONE vol 6 no 12 Article IDe28001 2011
[10] D de Marco N Clementi N Mancini et al ldquoA non-VH1-69 heterosubtypic neutralizing human monoclonal antibodyprotects mice against H1N1 and H5N1 virusesrdquo PLoS ONE vol7 no 4 Article ID e34415 2012
[11] L Solforosi N Mancini F Canducci et al ldquoA phage displayvector optimized for the generation of human antibody com-binatorial libraries and the molecular cloning of monoclonalantibody fragmentsrdquoNewMicrobiologica vol 35 no 3 pp 289ndash294 2012
[12] E V Koonin and V V Dolja ldquoEvolution and taxonomy ofpositive-strand RNA viruses implications of comparative anal-ysis of amino acid sequencesrdquo Critical Reviews in Biochemistryand Molecular Biology vol 28 no 5 pp 375ndash430 1993
[13] J S Mackenzie D J Gubler and L R Petersen ldquoEmergingflaviviruses the spread and resurgence of Japanese encephalitisWest Nile and dengue virusesrdquo Nature Medicine vol 10 no 12pp S98ndashS109 2004
[14] T J SmithW E Brandt J L Swanson J MMcCown and E LBuescher ldquoPhysical and biological properties of dengue-2 virusand associated antigensrdquo Journal of Virology vol 5 no 4 pp524ndash532 1970
[15] J M Smit B Moesker I Rodenhuis-Zybert and J WilschutldquoFlavivirus cell entry and membrane fusionrdquo Viruses vol 3 no2 pp 160ndash171 2011
[16] R J Kuhn W Zhang M G Rossmann et al ldquoStructure ofdengue virus implications for flavivirus organization matura-tion and fusionrdquo Cell vol 108 no 5 pp 717ndash725 2002
[17] K Stiasny J H Aberle M Keller B Grubeck-Loebenstein andF X Heinz ldquoAge affects quantity but not quality of antibodyresponses after vaccination with an inactivated flavivirus vac-cine against tick-borne encephalitisrdquo PLoS ONE vol 7 no 3Article ID e34145 2012
[18] LThomasOVerlaetenACabie et al ldquoInfluence of the dengueserotype previous dengue infection and plasma viral load
on clinical presentation and outcome during a dengue-2 anddengue-4 co-epidemicrdquo American Journal of Tropical Medicineand Hygiene vol 78 no 6 pp 990ndash998 2008
[19] Y Zhang J Corver P R Chipman et al ldquoStructures of imma-ture flavivirus particlesrdquo EMBO Journal vol 22 no 11 pp2604ndash2613 2003
[20] A K I Falconar ldquoIdentification of an epitope on the denguevirus membrane (M) protein defined by cross-protective mon-oclonal antibodies design of an improved epitope sequencebased on common determinants present in both envelope (EandM) proteinsrdquo Archives of Virology vol 144 no 12 pp 2313ndash2330 1999
[21] K-J Huang Y-C Yang Y-S Lin et al ldquoThe dual-specific bind-ing of dengue virus and target cells for the antibody-dependentenhancement of dengue virus infectionrdquo Journal of Immunol-ogy vol 176 no 5 pp 2825ndash2832 2006
[22] A K I Falconar ldquoAntibody responses are generated to immun-odominant ELKKLE-typemotifs on the nonstructural-1 glyco-protein during live dengue virus infections inmice and humansimplications for diagnosis pathogenesis and vaccine designrdquoClinical and Vaccine Immunology vol 14 no 5 pp 493ndash5042007
[23] C W Mandl F Guirakhoo H Holzmann F X Heinz and CKunz ldquoAntigenic structure of the flavivirus envelope protein Eat the molecular level using the tick-borne encephalitis virus asa modelrdquo Journal of Virology vol 63 no 2 pp 564ndash571 1989
[24] F A Rey F X Heinz C Mandl C Kunz and S C HarrisonldquoThe envelope glycoprotein from tick-borne encephalitis virusat 2 A resolutionrdquo Nature vol 375 no 6529 pp 291ndash298 1995
[25] S L Allison K Stiasny K Stadler CWMandl and F XHeinzldquoMapping of functional elements in the stem-anchor regionof tick-borne encephalitis virus envelope protein Erdquo Journal ofVirology vol 73 no 7 pp 5605ndash5612 1999
[26] D E Klein J L Choi and S C Harrison ldquoStructure of a denguevirus envelope protein late-stage fusion intermediaterdquo Journal ofVirology vol 87 no 4 pp 2287ndash2293 2013
[27] S L Hanna T C Pierson M D Sanchez A A Ahmed M MMurtadha and R W Doms ldquoN-linked glycosylation of WestNile virus envelope proteins influences particle assembly andinfectivityrdquo Journal of Virology vol 79 no 21 pp 13262ndash132742005
[28] N Mancini G Sautto N Clementi et al ldquoNeutralization inter-fering antibodies a ldquonovelrdquo example of humoral immune dys-function facilitating viral escaperdquoViruses vol 4 no 9 pp 1731ndash1752 2012
[29] Y Modis S Ogata D Clements and S C Harrison ldquoVariablesurface epitopes in the crystal structure of dengue virus type3 envelope glycoproteinrdquo Journal of Virology vol 79 no 2 pp1223ndash1231 2005
[30] N Miller ldquoRecent progress in dengue vaccine research anddevelopmentrdquo Current Opinion in Molecular Therapeutics vol12 no 1 pp 31ndash38 2010
[31] J H Chavez J R Silva A A Amarilla and L T Moraes Figu-eiredo ldquoDomain III peptides from flavivirus envelope proteinare useful antigens for serologic diagnosis and targets forimmunizationrdquo Biologicals vol 38 no 6 pp 613ndash618 2010
[32] B Shrestha J D Brien S Sukupolvi-Petty et al ldquoThe develop-ment of therapeutic antibodies that neutralize homologous andheterologous genotypes of dengue virus type 1rdquo PLoS Pathogensvol 6 no 4 Article ID e1000823 2010
18 BioMed Research International
[33] G D Gromowski N D Barrett and A D T Barrett ldquoChar-acterization of dengue virus complex-specific neutralizing epi-topes on envelope protein domain III of dengue 2 virusrdquo Journalof Virology vol 82 no 17 pp 8828ndash8837 2008
[34] S Sukupolvi-Petty S K Austin W E Purtha et al ldquoType- andsubcomplex-specific neutralizing antibodies against domain IIIof dengue virus type 2 envelope protein recognize adjacentepitopesrdquo Journal of Virology vol 81 no 23 pp 12816ndash128262007
[35] S Sukupolvi-Petty S K Austin M Engle et al ldquoStructure andfunction analysis of therapeutic monoclonal antibodies againstdengue virus type 2rdquo Journal of Virology vol 84 no 18 pp9227ndash9239 2010
[36] Y-Q Deng J-X Dai G-H Ji et al ldquoA broadly flaviviruscross-neutralizingmonoclonal antibody that recognizes a novelepitope within the fusion loop of e proteinrdquo PLoS ONE vol 6no 1 Article ID e16059 2011
[37] J M Costin E Zaitseva K M Kahle et al ldquoMechanistic studyof broadly neutralizing human monoclonal antibodies againstdengue virus that target the fusion looprdquo Journal of Virologyvol 87 no 1 pp 52ndash66 2013
[38] CMMidgley A FlanaganH B Tran et al ldquoStructural analysisof a dengue cross-reactive antibody complexed with envelopedomain III reveals the molecular basis of cross-reactivityrdquoJournal of Immunology vol 188 no 10 pp 4971ndash4979 2012
[39] N J Moreland P Susanto E Lim et al ldquoPhage display appro-aches for the isolation of monoclonal antibodies against denguevirus envelope domain III from human and mouse derivedlibrariesrdquo International Journal ofMolecular Sciences vol 13 no3 pp 2618ndash2635 2012
[40] J J B Cockburn M E Navarro Sanchez N Fretes et alldquoMechanism of dengue virus broad cross-neutralization by amonoclonal antibodyrdquo Structure vol 20 no 2 pp 303ndash3142012
[41] R Rajamanonmani C Nkenfou P Clancy et al ldquoOn a mousemonoclonal antibody that neutralizes all four dengue virusserotypesrdquo Journal of General Virology vol 90 no 4 pp 799ndash809 2009
[42] L H Gould J Sui H Foellmer et al ldquoProtective and herapeuticcapacity of human single-chain Fv-Fc fusion proteins againstWest Nile virusrdquo Journal of Virology vol 79 no 23 pp 14606ndash14613 2005
[43] H Sultana H G Foellmer G Neelakanta et al ldquoFusion LoopPeptide of the West Nile virus envelope protein is essential forpathogenesis and is recognized by a therapeutic cross-reactivehuman monoclonal antibodyrdquo Journal of Immunology vol 183no 1 pp 650ndash660 2009
[44] T Oliphant M Engle G E Nybakken et al ldquoDevelopment ofa humanized monoclonal antibody with therapeutic potentialagainstWest Nile virusrdquoNature Medicine vol 11 no 5 pp 522ndash530 2005
[45] G E Nybakken T Oliphant S Johnson S Burke M S Dia-mond and D H Fremont ldquoStructural basis of West Nile virusneutralization by a therapeutic antibodyrdquo Nature vol 437 no7059 pp 764ndash769 2005
[46] G D Gromowski J T Roehrig M S Diamond J C LeeT J Pitcher and A D T Barrett ldquoMutations of an antibodybinding energy hot spot on domain III of the dengue 2 envelopeglycoprotein exploited for neutralization escaperdquo Virology vol407 no 2 pp 237ndash246 2010
[47] J T Roehrig R A Bolin andR G Kelly ldquoMonoclonal antibodymapping of the envelope glycoprotein of the dengue 2 virusJamaicardquo Virology vol 246 no 2 pp 317ndash328 1998
[48] R de Alwis S A Smith N P Olivarez et al ldquoIdentification ofhumanneutralizing antibodies that bind to complex epitopes ondengue virionsrdquo Proceedings of the National Academy of Sciencesof the United States of America vol 109 no 19 pp 7439ndash74442012
[49] S K Austin K A Dowd B Shrestha et al ldquoStructural basis ofdifferential neutralization of DENV-1 genotypes by an antibodythat recognizes a cryptic epitoperdquo PLoS Pathogens vol 8 no 10Article ID e1002930 2012
[50] M Throsby C Geuijen J Goudsmit et al ldquoIsolation and char-acterization of human monoclonal antibodies from individualsinfected with West Nile Virusrdquo Journal of Virology vol 80 no14 pp 6982ndash6992 2006
[51] A P Goncalvez R H Purcell and C-J Lai ldquoEpitope deter-minants of a chimpanzee Fab antibody that efficiently cross-neutralizes dengue type 1 and type 2 viruses map to inside andin close proximity to fusion loop of the dengue type 2 virusenvelope glycoproteinrdquo Journal of Virology vol 78 no 23 pp12919ndash12928 2004
[52] J T Roehrig J H Mathews and D W Trent ldquoIdentification ofepitopes on the E glycoprotein of Saint Louis encephalitis virususing monoclonal antibodiesrdquo Virology vol 128 no 1 pp 118ndash126 1983
[53] J T Roehrig L A Staudinger A R Hunt J H Mathews andC D Blair ldquoAntibody prophylaxis and therapy for flavivirusencephalitis infectionsrdquo Annals of the New York Academy ofSciences vol 951 pp 286ndash297 2001
[54] A P Goncalvez C-H Chien K Tubthong et al ldquoHumanizedmonoclonal antibodies derived from chimpanzee fabs protectagainst Japanese encephalitis virus in vitro and in vivordquo Journalof Virology vol 82 no 14 pp 7009ndash7021 2008
[55] M Arakawa T Yamashiro G-I Uechi M Tadano and ANishizono ldquoConstruction of humanFab (1205741120581) library and iden-tification of human monoclonal Fab possessing neutralizingpotency against Japanese encephalitis virusrdquo Microbiology andImmunology vol 51 no 6 pp 617ndash625 2007
[56] L Warter C Y Lee R Thiagarajan et al ldquoChikungunya virusenvelope-specific human monoclonal antibodies with broadneutralization potencyrdquo Journal of Immunology vol 186 no 5pp 3258ndash3264 2011
[57] P Pal K A Dowd J D Brien et al ldquoDevelopment of a highlyprotective combination monoclonal antibody therapy againstchikungunya virusrdquo PLoS Pathogens vol 9 no 4 Article IDe1003312 2013
[58] A Bertolotti-Ciarlet J Smith K Strecker et al ldquoCellular local-ization and antigenic characterization of Crimean-Congo hem-orrhagic fever virus glycoproteinsrdquo Journal of Virology vol 79no 10 pp 6152ndash6161 2005
[59] J K Battles and J M Dalrymple ldquoGenetic variation amonggeographic isolates of Rift Valley fever virusrdquo American Journalof Tropical Medicine and Hygiene vol 39 no 6 pp 617ndash6311988
[60] B AThibodeaux N C Garbino NM Liss et al ldquoA humanizedIgG but not IgM antibody is effective in prophylaxis and therapyof yellow fever infection in an AG12917D-204 peripheralchallenge mouse modelrdquo Antiviral Research vol 94 no 1 pp1ndash8 2012
BioMed Research International 19
[61] S Daffis R E Kontermann J Korimbocus H Zeller H-DKlenk and J Ter Meulen ldquoAntibody responses against wild-type yellow fever virus and the 17D vaccine strain charac-terization with human monoclonal antibody fragments andneutralization escape variantsrdquoVirology vol 337 no 2 pp 262ndash272 2005
[62] L N Levanov L E Matveev E P Goncharova et al ldquoChimericantibodies against tick-borne encephalitis virusrdquo Vaccine vol28 no 32 pp 5265ndash5271 2010
[63] A R Hunt S Frederickson C Hinkel K S Bowdish and J TRoehrig ldquoA humanized murine monoclonal antibody protectsmice either before or after challenge with virulent Venezuelanequine encephalomyelitis virusrdquo Journal of General Virologyvol 87 no 9 pp 2467ndash2476 2006
[64] J M da Silva Voorham I A Rodenhuis-Zybert N V A Nunezet al ldquoAntibodies against the envelope glycoprotein promoteinfectivity of immature dengue virus serotype 2rdquo PLoS ONEvol 7 no 3 Article ID e29957 2012
[65] T C Pierson Q Xu S Nelson et al ldquoThe stoichiometry ofantibody-mediated neutralization and enhancement of westnile virus infectionrdquoCell Host andMicrobe vol 1 no 2 pp 135ndash145 2007
[66] G Sautto N Mancini R A Diotti L Solforosi M Clementiand R Burioni ldquoAnti-hepatitis C virus E2 (HCVE2) glyco-proteinmonoclonal antibodies andneutralization interferencerdquoAntiviral Research vol 96 no 1 pp 82ndash89 2012
[67] R Burioni Y Matsuura N Mancini et al ldquoDiverging effectsof human recombinant anti-hepatitis C virus (HCV) antibodyfragments derived from a single patient on the infectivityof a vesicular stomatitis virusHCV pseudotyperdquo Journal ofVirology vol 76 no 22 pp 11775ndash11779 2002
[68] R J Phillpotts J R Stephenson and J S Porterfield ldquoAntibody-dependent enhancement of tick-borne encephalitus virus infec-tivityrdquo Journal of General Virology vol 66 no 8 pp 1831ndash18371985
[69] K A Dowd and T C Pierson ldquoAntibody-mediated neutraliza-tion of flaviviruses a reductionist viewrdquo Virology vol 411 no 2pp 306ndash315 2011
[70] A K I Falconar ldquoMonoclonal antibodies that bind to commonepitopes on the dengue virus type 2 nonstructural-1 andenvelope glycoproteins display weak neutralizing activity anddifferentiated responses to virulent strains implications forpathogenesis and vaccinesrdquo Clinical and Vaccine Immunologyvol 15 no 3 pp 549ndash561 2008
[71] K Meyer A Banerjee S E Frey R B Belshe and R Ray ldquoAweak neutralizing antibody response to hepatitis C virus enve-lope glycoprotein enhances virus infectionrdquo PLoS ONE vol 6no 8 Article ID e23699 2011
[72] R Burioni R AWilliamson P P Sanna F E Bloom and D RBurton ldquoRecombinant human Fab to glycoprotein D neutral-izes infectivity and prevents cell-to-cell transmission of herpessimplex viruses 1 and 2 in vitrordquo Proceedings of the NationalAcademy of Sciences of the United States of America vol 91 no1 pp 355ndash359 1994
[73] M S Rolph A Zaid N E Rulli and S Mahalingam ldquoDown-regulation of interferon-120573 in antibody-dependent enhancementof dengue viral infections of human macrophages is dependenton interleukin-6rdquo Journal of Infectious Diseases vol 204 no 3pp 489ndash491 2011
[74] S S Whitehead J E Blaney A P Durbin and B R MurphyldquoProspects for a dengue virus vaccinerdquoNature ReviewsMicrobi-ology vol 5 no 7 pp 518ndash528 2007
[75] S-M Lok V Kostyuchenko G ENybakken et al ldquoBinding of aneutralizing antibody to dengue virus alters the arrangement ofsurface glycoproteinsrdquoNature Structural andMolecular Biologyvol 15 no 3 pp 312ndash317 2008
[76] A P Goncalvez R Men C Wernly R H Purcell and C-J Lai ldquoChimpanzee Fab fragments and a derived humanizedimmunogiobulin G1 antibody that efficiently cross-neutralizedengue type 1 and type 2 virusesrdquo Journal of Virology vol 78no 23 pp 12910ndash12918 2004
[77] WM P BWahala A A Kraus L B HaymoreM A Accavitti-Loper and A M de Silva ldquoDengue virus neutralization byhuman immune sera role of envelope protein domain III-reactive antibodyrdquo Virology vol 392 no 1 pp 103ndash113 2009
[78] W D Crill H R Hughes M J Delorey and G-J J ChangldquoHumoral immune responses of dengue fever patients usingepitope-specific serotype-2 virus-like particle antigensrdquo PLoSONE vol 4 no 4 Article ID e4991 2009
[79] C Setthapramote T Sasaki O Puiprom et al ldquoHuman mono-clonal antibodies to neutralize all dengue virus serotypes usinglymphocytes from patients at acute phase of the secondaryinfectionrdquo Biochemical and Biophysical Research Communica-tions vol 423 no 4 pp 867ndash872 2012
[80] M Beltramello K L Williams C P Simmons et al ldquoThehuman immune response to dengue virus is dominated byhighly cross-reactive antibodies endowed with neutralizing andenhancing activityrdquo Cell Host andMicrobe vol 8 no 3 pp 271ndash283 2010
[81] C M Midgley M Bajwa-Joseph S Vasanawathana et al ldquoAnin-depth analysis of original antigenic sin in dengue virusinfectionrdquo Journal of Virology vol 85 no 22 pp 410ndash421 2011
[82] B Kaufmann M R Vogt J Goudsmit et al ldquoNeutralizationof West Nile virus by cross-linking of its surface proteins withFab fragments of the human monoclonal antibody CR4354rdquoProceedings of the National Academy of Sciences of the UnitedStates of America vol 107 no 44 pp 18950ndash18955 2010
[83] B Spurrier J M Sampson M Totrov et al ldquoStructural analysisof human and macaque mAbs 2909 and 25B implicationsfor the configuration of the quaternary neutralizing epitope ofHIV-1 gp120rdquo Structure vol 19 no 5 pp 691ndash699 2011
[84] J J SchlesingerMWBrandriss andT PMonath ldquoMonoclonalantibodies distinguish between wild and vaccine strains ofyellow fever virus by neutralization hemagglutination inhibi-tion and immune precipitation of the virus envelope proteinrdquoVirology vol 125 no 1 pp 8ndash17 1983
[85] M W Brandriss J J Schlesinger E E Walsh and M BrisellildquoLethal 17D yellow fever encephalitis in mice I Passive protec-tion by monoclonal antibodies to the envelope proteins of 17Dyellow fever and dengue 2 virusesrdquo Journal of General Virologyvol 67 no 2 pp 229ndash234 1986
[86] H Cho and M S Diamond ldquoImmune responses to West Nilevirus infection in the central nervous systemrdquo Viruses vol 4no 12 pp 3812ndash3830 2012
[87] D W Beasley A D Barrett and R B Tesh ldquoResurgence ofWest Nile neurologic disease in the United States in 2012 whathappened What needs to be donerdquo Antiviral Research vol 99no 1 pp 1ndash5 2013
[88] M Throsby J Ter Meulen C Geuijen J Goudsmit and J DeKruif ldquoMapping and analysis of West Nile virus-specific mon-oclonal antibodies prospects for vaccine developmentrdquo ExpertReview of Vaccines vol 6 no 2 pp 183ndash191 2007
[89] M S Diamond T G Roberts D Edgil B Lu J Ernst and EHarris ldquoModulation of dengue virus infection in human cells
20 BioMed Research International
by alpha beta and gamma interferonsrdquo Journal of Virology vol74 no 11 pp 4957ndash4966 2000
[90] Z Shimoni M J Niven S Pitlick and S Bulvik ldquoTreatmentof West Nile virus encephalitis with intravenous immunoglob-ulinrdquo Emerging Infectious Diseases vol 7 no 4 article 759 2001
[91] AHamdan PGreen EMendelsonM R Kramer S Pitlik andM Weinberger ldquoPossible benefit of intravenous immunoglob-ulin therapy in a lung transplant recipient with West Nile virusencephalitisrdquoTransplant InfectiousDisease vol 4 no 3 pp 160ndash162 2002
[92] M Haley A S Retter D Fowler J Gea-Banacloche and NP OrsquoGrady ldquoThe role for intravenous immunoglobulin in thetreatment of West Nile virus encephalitisrdquo Clinical InfectiousDiseases vol 37 no 6 pp e88ndashe90 2003
[93] K M Chung G E Nybakken B S Thompson et al ldquoAnti-bodies against West Nile virus nonstructural protein NS1prevent lethal infection through Fc 120574 receptor-dependent and-independent mechanismsrdquo Journal of Virology vol 80 no 3pp 1340ndash1351 2006
[94] E A Henchal L S Henchal and J J Schlesinger ldquoSynergisticinteractions of anti-NS1 monoclonal antibodies protect pas-sively immunized mice from lethal challenge with dengue 2virusrdquo Journal of General Virology vol 69 no 8 pp 2101ndash21071988
[95] J T Roehrig ldquoAntigenic structure of flavivirus proteinsrdquoAdvances in Virus Research vol 59 pp 141ndash175 2003
[96] M D Sanchez T C Pierson D McAllister et al ldquoCharacteri-zation of neutralizing antibodies to West Nile virusrdquo Virologyvol 336 no 1 pp 70ndash82 2005
[97] J H Beigel J L Nordstrom S R Pillemer et al ldquoSafety andpharmacokinetics of single intravenous dose of MGAWN1 anovel monoclonal antibody to West Nile virusrdquo AntimicrobialAgents and Chemotherapy vol 54 no 6 pp 2431ndash2436 2010
[98] C A Smeraski V Siddharthan and J D Morrey ldquoTreatmentof spatial memory impairment in hamsters infected with WestNile virus using a humanizedmonoclonal antibodyMGAWN1rdquoAntiviral Research vol 91 no 1 pp 43ndash49 2011
[99] D Lelli AMoreno E Brocchi et al ldquoWestNile virus character-ization and diagnostic applications of monoclonal antibodiesrdquoVirology Journal vol 9 article 81 2012
[100] T S Gritsun V A Lashkevich and E A Gould ldquoTick-borneencephalitisrdquo Antiviral Research vol 57 no 1-2 pp 129ndash1462003
[101] D Ruzek T S Gritsun N L Forrester et al ldquoMutations inthe NS2B and NS3 genes affect mouse neuroinvasiveness of aWestern European field strain of tick-borne encephalitis virusrdquoVirology vol 374 no 2 pp 249ndash255 2008
[102] M Ecker S L Allison T Meixner and F X Heinz ldquoSequenceanalysis and genetic classification of tick-borne encephalitisviruses from Europe and Asiardquo Journal of General Virology vol80 no 1 pp 179ndash185 1999
[103] H Holzmann M S Vorobyova I P Ladyzhenskaya et alldquoMolecular epidemiology of tick-borne encephalitis viruscross-protection between European and Far Eastern subtypesrdquoVaccine vol 10 no 5 pp 345ndash349 1992
[104] C W Mandl ldquoSteps of the tick-borne encephalitis virus repli-cation cycle that affect neuropathogenesisrdquo Virus Research vol111 no 2 pp 161ndash174 2005
[105] A V Atrasheuskaya T M Fredeking and G M IgnatyevldquoChanges in immune parameters and their correction in humancases of tick-borne encephalitisrdquo Clinical and ExperimentalImmunology vol 131 no 1 pp 148ndash154 2003
[106] K L Mansfield N Johnson L P Phipps J R Stephenson AR Fooks and T Solomon ldquoTick-borne encephalitis virusmdashareview of an emerging zoonosisrdquo Journal of General Virologyvol 90 no 8 pp 1781ndash1794 2009
[107] DWVaughn andCHHoke Jr ldquoThe epidemiology of Japaneseencephalitis prospects for preventionrdquo Epidemiologic Reviewsvol 14 pp 197ndash221 1992
[108] T Solomon H Ni D W C Beasley M Ekkelenkamp M JCardosa and A D T Barrett ldquoOrigin and evolution of Japaneseencephalitis virus in Southeast Asiardquo Journal of Virology vol 77no 5 pp 3091ndash3098 2003
[109] M R Wilson ldquoEmerging viral infectionsrdquo Current Opinion inNeurology vol 26 no 3 pp 301ndash306 2013
[110] J N Hanna S A Ritchie D A Phillips et al ldquoAn outbreakof Japanese encephalitis in the Torres Strait Australia 1995rdquoMedical Journal of Australia vol 165 no 5 pp 256ndash260 1996
[111] J N Hanna S A Ritchie D A Phillips et al ldquoJapanese ence-phalitis in north Queensland Australia 1998rdquo Medical Journalof Australia vol 170 no 11 pp 533ndash536 1999
[112] SGeorgeMGourie-Devi and J A Rao ldquoIsolation ofWestNilevirus from the brains of children who had died of encephalitisrdquoBulletin of theWorldHealthOrganization vol 62 no 6 pp 879ndash882 1984
[113] A K Gupta A A Koshy and V J Lad ldquoEnhanced protec-tion of mice against Japanese encephalitis virus infection bycombinations ofmonoclonal antibodies to glycoprotein ErdquoActaVirologica vol 55 no 2 pp 165ndash168 2011
[114] T H Lee B-H Song S-I Yun et al ldquoA cross-protective mAbrecognizes a novel epitope within the flavivirus NS1 proteinrdquoJournal of General Virology vol 93 no 1 pp 20ndash26 2012
[115] S C Weaver ldquoHost range amplification and arboviral diseaseemergencerdquo Archives of Virology Supplementum no 19 pp 33ndash44 2005
[116] M Solignat B Gay S Higgs L Briant and C Devaux ldquoRepli-cation cycle of chikungunya a re-emerging arbovirusrdquoVirologyvol 393 no 2 pp 183ndash197 2009
[117] O Schwartz and M L Albert ldquoBiology and pathogenesis ofchikungunya virusrdquo Nature Reviews Microbiology vol 8 no 7pp 491ndash500 2010
[118] W Akahata Z-Y Yang H Andersen et al ldquoA virus-like particlevaccine for epidemic Chikungunya virus protects nonhumanprimates against infectionrdquo Nature Medicine vol 16 no 3 pp334ndash338 2010
[119] M Kielian and A Helenius ldquopH-Iinduced alterations in thefusogenic spike protein of Semliki Forest virusrdquo Journal of CellBiology vol 101 no 6 pp 2284ndash2291 1985
[120] J White and A Helenius ldquopH-Dependent fusion between theSemliki Forest virus membrane and liposomesrdquo Proceedings ofthe National Academy of Sciences of the United States of Americavol 77 no 6 pp 3273ndash3277 1980
[121] J Dubuisson and C M Rice ldquoSindbis virus attachment isola-tion and characterization of mutants with impaired binding tovertebrate cellsrdquo Journal of Virology vol 67 no 6 pp 3363ndash33741993
[122] C Y Lee Y-W Kam J Fric et al ldquoChikungunya virus neutral-ization antigens and direct cell-to-cell transmission are revealedby human antibody-escapemutantsrdquo PLoS Pathogens vol 7 no12 Article ID e1002390 2011
[123] J E Voss M-C Vaney S Duquerroy et al ldquoGlycoproteinorganization of Chikungunya virus particles revealed by X-raycrystallographyrdquo Nature vol 468 no 7324 pp 709ndash712 2010
BioMed Research International 21
[124] L Li J Jose Y Xiang R J Kuhn and M G Rossmann ldquoStruc-tural changes of envelope proteins during alphavirus fusionrdquoNature vol 468 no 7324 pp 705ndash708 2010
[125] J Fric S Bertin-Maghit C I Wang A Nardin and L WarterldquoUse of human monoclonal antibodies to treat Chikungunyavirus infectionrdquo Journal of Infectious Diseases vol 207 no 2 pp319ndash322 2013
[126] P V Aguilar J G Estrada-Franco R Navarro-Lopez C FerroA D Haddow and S C Weaver ldquoEndemic Venezuelanequine encephalitis in the Americas hidden under the dengueumbrellardquo Future Virology vol 6 no 6 pp 721ndash740 2011
[127] S C Weaver R Winegar I D Manger and N L ForresterldquoAlphaviruses population genetics and determinants of emer-gencerdquo Antiviral Research vol 94 no 3 pp 242ndash257 2012
[128] S Paessler and S C Weaver ldquoVaccines for Venezuelan equineencephalitisrdquo Vaccine vol 27 no 4 pp D80ndashD85 2009
[129] R J Phillpotts ldquoVenezuelan equine encephalitis virus complex-specific monoclonal antibody provides broad protection inmurine models against airborne challenge with viruses fromserogroups I II and IIIrdquoVirus Research vol 120 no 1-2 pp 107ndash112 2006
[130] R J Phillpotts L D Jones and S C Howard ldquoMonoclonalantibody protectsmice against infection anddiseasewhen giveneither before or up to 24 h after airborne challenge with virulentVenezuelan equine encephalitis virusrdquoVaccine vol 20 no 11-12pp 1497ndash1504 2002
[131] J H Mathews J T Roehrig and D W Trent ldquoRole of com-plement and the Fc portion of immunoglobulin G in immunityto Venezuelan equine encephalomyelitis virus infection withglycoprotein-specific monoclonal antibodiesrdquo Journal of Virol-ogy vol 55 no 3 pp 594ndash600 1985
[132] L M OrsquoBrien C D Underwood-Fowler S A Goodchild AL Phelps and R J Phillpotts ldquoDevelopment of a novel mon-oclonal antibody with reactivity to a wide range of Venezuelanequine encephalitis virus strainsrdquoVirology Journal vol 6 article206 2009
[133] L M OrsquoBrien S A Goodchild R J Phillpotts and S D Per-kins ldquoA humanised murine monoclonal antibody protectsmice from Venezuelan equine encephalitis virus Evergladesvirus and Mucambo virus when administered up to 48h afterairborne challengerdquo Virology vol 426 no 2 pp 100ndash105 2012
[134] N Clementi N Mancini L Solforosi M Castelli M Clementiand R Burioni ldquoPhage display-based strategies for cloning andoptimization of monoclonal antibodies directed against humanpathogensrdquo International Journal of Molecular Sciences vol 13no 7 pp 8273ndash8292 2012
[135] A R Hunt R A Bowen S Frederickson T Maruyama JT Roehrig and C D Blair ldquoTreatment of mice with humanmonoclonal antibody 24h after lethal aerosol challenge withvirulent Venezuelan equine encephalitis virus prevents diseasebut not infectionrdquo Virology vol 414 no 2 pp 146ndash152 2011
[136] W-G Hu A L Phelps S Jager et al ldquoA recombinant human-ized monoclonal antibody completely protects mice againstlethal challenge with Venezuelan equine encephalitis virusrdquoVaccine vol 28 no 34 pp 5558ndash5564 2010
[137] T Rulker L Voss P Thullier et al ldquoIsolation and characterisa-tion of a human-like antibody fragment (scFv) that inactivatesVEEV in vitro and in vivordquo PLoS ONE vol 7 no 5 Article IDe37242 2012
[138] C T Walter and J N Barr ldquoRecent advances in the molecularand cellular biology of bunyavirusesrdquo Journal of General Virol-ogy vol 92 no 11 pp 2467ndash2484 2011
[139] C A Whitehouse ldquoCrimean-Congo hemorrhagic feverrdquo Anti-viral Research vol 64 no 3 pp 145ndash160 2004
[140] M Mertens K Schmidt A Ozkul and M H Groschup ldquoTheimpact of Crimean-Congo hemorrhagic fever virus on publichealthrdquo Antiviral Research vol 98 no 2 pp 248ndash260 2013
[141] A A Ahmed J M McFalls C Hoffmann et al ldquoPresence ofbroadly reactive and group-specific neutralizing epitopes onnewly described isolates of Crimean-Congo hemorrhagic fevervirusrdquo Journal of General Virology vol 86 no 12 pp 3327ndash33362005
[142] R Hewson A Gmyl L Gmyl et al ldquoEvidence of segment reas-sortment in Crimean-Congo haemorrhagic fever virusrdquo Journalof General Virology vol 85 no 10 pp 3059ndash3070 2004
[143] N K Blackburn T G Besselaar A J Shepherd and R Swane-poel ldquoPreparation and use of monoclonal antibodies for iden-tifying Crimean-Congo hemorrhagic fever virusrdquo AmericanJournal of Tropical Medicine and Hygiene vol 37 no 2 pp 392ndash397 1987
[144] T Ikegami ldquoMolecular biology and genetic diversity of RiftValley fever virusrdquoAntiviral Research vol 95 no 3 pp 293ndash3102012
[145] KKeegan andM S Collett ldquoUse of bacterial expression cloningto define the amino acid sequences of antigenic determinantson the G2 glycoprotein of Rift Valley fever virusrdquo Journal ofVirology vol 58 no 2 pp 263ndash270 1986
[146] T G Besselaar and N K Blackburn ldquoTopological mapping ofantigenic sites on the Rift Valley fever virus envelope glycopro-teins using monoclonal antibodiesrdquo Archives of Virology vol121 no 1ndash4 pp 111ndash124 1991
[147] L G Presta ldquoEngineering of therapeutic antibodies to mini-mize immunogenicity and optimize functionrdquo Advanced DrugDelivery Reviews vol 58 no 5-6 pp 640ndash656 2006
Submit your manuscripts athttpwwwhindawicom
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anatomy Research International
PeptidesInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporation httpwwwhindawicom
International Journal of
Volume 2014
Zoology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Molecular Biology International
GenomicsInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioinformaticsAdvances in
Marine BiologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Signal TransductionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Evolutionary BiologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Biochemistry Research International
ArchaeaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Genetics Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in
Virolog y
Hindawi Publishing Corporationhttpwwwhindawicom
Nucleic AcidsJournal of
Volume 2014
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Enzyme Research
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Microbiology

BioMed Research International 5
non-protective Abs may contribute to more severe clinicaloutcomes in vaccinated people [74]
241 Murine versus Human Anti-DENV Humoral ImmuneResponse It is well ascertained that anti-E DIII-directed Absare virtually absent from the naıve human repertoire as theyare directed away from this domain probably against weaklyneutralizing and immunodominant regions Moreover it hasbeen demonstrated that over than 90 of the human Abresponse of primary DENV infected patients is able to bindonly native DENV virus particles instead of a recombinantform of the E protein posing difficulties in the cloning strate-gies of neutralizing mAbs [39] Thus it can be concludedthat in vivo only a small fraction of DENV-specific Abs areresponsible for neutralization
Conversely murine mAbs that recognize all the threedomains of E have been identified Epitope-mapping studiesusingmurine serotype-specificmAbs have demonstrated thatloops located in the lateral ridge region of DIII consti-tute the strongest neutralizing B-cell target In this regionsequence diversity between serotype is high while cross-reactive murine mAbs that recognize another loop showedgenerally a weaker neutralization profile In particular type-specific mAbs with neutralizing activity against DENV2localized to the BC DE and FG loops on the lateral ridgeof DIII whereas subcomplex-specific mAbs recognized anadjacent epitope centered on the connecting A strand of DIIIat residues K305 K307 and K310 [32ndash35]
Cross-reactive mAbs directed against prM and NS1 havealso been described but as anticipated they feature weaklyneutralizing or absent as well as ADE of infection activityIn the following paragraphs we report the better character-ized neutralizing mouse and human mAbs describing theirmolecular features further summarized in Table 1
242 Anti-DENV Murine mAbs Murine mAbs directedagainst all the three domains of the E protein and endowedof different neutralizing and binding characteristics havebeen described However only DIII and DII are classicallyrecognized by neutralizing Abs In this regard Sukupolvi-Petty et al isolated twenty-four anti-DENV2 mouse mAbswith moderate or strong neutralizing activity against thehomologous virus in cell culture assays Binding sites weremapped for the majority of these in distinct epitopes inregions located in DI (lateral ridge) DII (dimer interfacelateral ridge and fusion loop) and DIII (lateral ridge CC1015840loop and A strand) Moreover 16 of the neutralizing mAbswere tested in mice with most of them being protectivewhen given as prophylaxis Seven of these had post-exposuretherapeutic activity when administered as a single dose byintraperitoneal route even 3 days after intracranial infectionFor the mAbs with the greatest therapeutic potential protec-tion was confirmed with an Ab-enhanced vascular leakagemouse model of DENV2 infection [35]
Recently Deng et al described a cross-reactive murinemAb named 2A10G6 that is able to recognize DENV1ndash4 YFV WNV JEV and TBEV to potently cross-neutralizeDENV1ndash4 YFV and to a lesser extent WNV This mAbrecognizes a highly conserved motif (amino acid residues
98ndash101) located within the fusion loop of the FlavivirusDII of the E glycoprotein Moreover this mAb exerts itsneutralizing activity in a post-attachment step during thevirus entry process as demonstrated by kinetic neutralizationtests performed in vitro Additionally protection experimentsperformed in mouse models showed that treatment with100 120583gmL of 2A10G6 conferred full protection against lethalDENV2 challenge and 20 120583gmL and 4120583gmL of 2A10G6protected 89 and 40 of infected mice from lethal chal-lenge respectively For infection with DENV1 3 and 4 use of100 120583gmL of 2A10G6 conferred partial protection and 5377 and 73 of the infected mice survived after challengerespectively Finally the protection profile of 2A10G6 againstWNV showed that prophylactic administration with a singledose of 200mg of 2A10G6 conferred 80 protection in miceMost importantly 3 of 8 (375) mice survived when 2A10G6was administered one day after WNV challenge SimilarlyanothermousemAb 4G2 is able to recognize the fusion loopat the extremity of DII from all four DENV serotypes and toprevent syncytia formation [36]
Rajamanonmani et al described a mouse mAb 9F12raised against theDIII of DENV2E protein that is able to rec-ognize DENV1ndash4 serotypes as well asWNV and to neutralizefive DENV strains representative of all DENV serotypes [41]Similarly Cockburn et al reported a comparative high-resolution crystallographic analysis of an A-strand DIIImurinemAb 4E11 in complex with its target domain of the Eprotein from the four DENV serotypes MAb 4E11 is capableof recognizing and neutralizing all four serotypes with IC50values varying between 1 and 100 nMThe structures reportedalso highlight the mechanism by which anti-A-strand mAbsdisrupt the architecture of the mature virion inducing dimerdissociation premature fusion-loop exposure and concomi-tant particle inactivation [40] Also Midgley et al function-ally characterized another murine mAb 2H12 raised in miceagainstDIII of E protein Similarly to the previously describedmAbs 2H12 is able to bind all the four DENV serotypes ina epitope encompassing the conserved 314ETQH317 motifHowever the neutralizing potential of 2H12 is lower than anumber of other anti-DIII mAbs with IC50 values rangingfrom 056 to 145 nM for DENV1 2 and 4 On the other handit showed no ADE of infection activity [38]
Austin et al isolated a murine mAb named E111 whichrecognize a novel CC1015840-loop epitope on DIII of the E proteinfrom two different DENV1 genotypes Docking of the mAbstructure onto the available cryoelectron microscopy modelsof DENV virions revealed that the E111 epitope was inaccessi-ble suggesting that this mAb recognizes an uncharacterizedvirus conformation While the affinity of binding betweenE111 and DIII varied by genotype a limited correlation withneutralizing activity was observed These data support theconclusion that potent neutralization depends on genotype-dependent exposure of the CC1015840 loop epitope [49] In fact aspreviously described elsewhere binding of some E reactiveAbs depends on the dynamicmovement of proteinmoleculesldquobreathingrdquo in the virion particle leading to transient expo-sure of hidden epitopes For instance optimal binding ofmousemAb 1A1D-2 to EDIII requires incubation at 37∘C [75]
6 BioMed Research InternationalTa
ble1Schematicsummaryof
theb
estcharacterized
mAb
sagainstarbo
viralp
atho
gens
mAb
Orig
inRe
activ
ityTargetprotein
Epito
peClon
ingstr
ategy
Form
atisotype
Neutralizingactiv
ityRe
ference
48A
Hum
anDEN
V1ndash4
EDII
EBVtransfo
rmation
IgG
IC50
(120583gmL)
DEN
V115
DEN
V2gt40
DEN
V324
DEN
V4gt40
[37]
D11C
IC50
(120583gmL)
DEN
V115
DEN
V210
DEN
V3102
DEN
V416
16D
IC50
(120583gmL)
DEN
V115
DEN
V202
DEN
V305
DEN
V427
2H12
Murine
DEN
V1ndash4
EDIII(ABloop
aa
314ndash
317)
Hybrid
omafrom
BALB
cmou
seim
mun
ized
with
DEN
V2EDIII
IgG2b
IC50
nM
DEN
V1056ndash54
DEN
V329
DEN
V4145
[38]
C9Murinechim
eric
DEN
V2
EDIII
Phaged
isplayof
achim
ericmurine
hybridom
alibrary
VH1V1205811
PRNT5
0850120583
gmL
[39]
4E11
Murine
DEN
V1ndash4
EDIII(str
andA308
312andstrand
G387
389391)
Hybrid
oma
IgG2a120581
IC50
(120583gmL)
DEN
V1016
DEN
V2013
DEN
V-38
DEN
V-415
[40]
4G2
Murine
DEN
V1ndash4
EDII(fu
sionloop
)Hybrid
omafrom
DEN
V2Eim
mun
ized
mice
IgG
PRNT5
0(120583gmL)
DEN
V215
[39]
9F12
Murine
DEN
V1ndash4WNV
EDIII(aa
305307310
330Astr
andandBC
loop
)
Hybrid
omafrom
BALB
cmou
seim
mun
ized
with
DEN
V-2EDIII
IgG1k
DEN
V1ndash4P
RNT5
02sdot10minus8ndash2sdot10minus7M
[41]
2A10G6
Murine
DEN
V1ndash4YF
VWNV
JEV
TBEV
EDII(Fusionloop
aa
98ndash101)
Hybrid
omafrom
BALB
cmou
seim
mun
ized
with
inactiv
ated
DEN
V2
IgG1
PRNT5
0(120583gmL)
DEN
V12
DEN
V215
DEN
V321
DEN
V418
YNF36
WNV
46
[36]
mAb
11Hum
anDEN
V1ndash4WNV
SLEV
YFV
JEV
MVEV
EDII(fu
sionloop
)Ph
aged
isplayof
human
scFv
Fusio
nprotein
scFv-Fc
PRNT8
0(120583gmL)
WNV
125
DEN
V2625
[4243]
BioMed Research International 7Ta
ble1Con
tinued
mAb
Orig
inRe
activ
ityTargetprotein
Epito
peClon
ingstr
ategy
Form
atisotype
Neutralizingactiv
ityRe
ference
E16
(MGAW
N1)
Murine
WNV
EDIII(LR
aa3
02ndash309)
Hybrid
omafrom
immun
ized
micew
ithWNVE
IgG2bhum
anized
PRNT5
04ndash
18ng
PRNT9
053ndash297
ng[4445]
1A1D
-2Murine
DEN
V1ndash3
EDIII(Astr
andaa
307
310and312)
Hybrid
omafrom
immun
ized
micew
ithdifferent
pH-tr
eated
virus
IgG2a
DEN
V2PR
NT5
021n
M[4647]
1F4
Hum
an
DEN
V1
E(virion
)
DI-DII
Electro
fusio
nof
infected
mem
oryBcells
from
DEN
V-im
mun
esub
jects
with
EBV
IgG
DEN
V1IC5
0011120583gmL
[48]
2D22
DEN
V2
DIII
DEN
V2IC50
008120583gmL
5J7
DEN
V3
DI-DII
DEN
V3IC50
010120583gmL
E105
Murine
DEN
V1
E
DIII(BC
loop
G328
T329
andD330DE
loop
K361E
and
E362K
FGloop
K3
85)
Hybrid
omas
deriv
edfro
mC5
7BL6
IFN-120572120573Rminus
minusmice
infected
with
DEN
V1
IgG
PRNT5
0DEN
V-1
05ndash592ng
mL
[32]
E106
DIII(BC
loop
G328
T329
andD330DE
loop
K361E
and
E362K
FGloop
K3
85A
strand
S305
K307E
309K3
10and
E311)
PRNT5
0DEN
V-1
06ndash
592ng
mL
E111
DIII(CC1015840loop
)PR
NT5
038ndash25120583gmL
[49]
CR4374
Hum
anWNV
EDIII
Phaged
isplayof
scFv
IgGlib
rary
VH2-05V
L1e
PRNT5
0018120583gmL
PRNT9
0095120583gmL
[50]
CR4353
VH3-30V
k3-A27
PRNT5
00026120583
gmL
PRNT9
0364120583gmL
1A5
Chim
panzee
DEN
V1ndash4WNV
JEV
LGTV
EDII(aaG
106)
Phaged
isplayof
Fab
library
from
DEN
V1ndash4
infected
chim
panzees
Hum
anized
IgG1
PRNT5
0(120583gmL)
DEN
V1048
DEN
V2095
DEN
V332
DEN
V443
WNVD
ENV438
JEV
21LG
TV28
[51]
8 BioMed Research International
Table1Con
tinued
mAb
Orig
inRe
activ
ityTargetprotein
Epito
peClon
ingstr
ategy
Form
atisotype
Neutralizingactiv
ityRe
ference
mAb
11Hum
anDEN
V1ndash4WNV
EDII(fu
sionloop
W101G104G106)
scFv
library
scFv-Fc
PRNT8
0(120583gmL)
DEN
V2625
WNV
125
[4243]
3B4C
-4
Murine
SLEV
E
1a
Hybrid
oma
IgG
SLEV
PRNTlt17
[5253]
1B2C
-5SLEV
1bSLEV
PRNTlt17
6B5A
-2SLEV
1cSLEV
PRNT
48
4A4C
-4SLEV
1dSLEV
PRNT
29
1B5D
-1SLEV
JEV
2SLEV
PRNTlt17
2B5B
-3SL
EVJEV
MVEV
WNV
YFV
3SLEV
PRNT
23
2B6B
-2AllFlaviviru
s4a
SLEV
PRNTlt17
6B6C
-1AllFlaviviru
s4b
(inDII)
SLEV
PRNT
23
A3
Chim
panzee
JEV
E2
DIa
aK1
79Ph
aged
isplay
Hum
anized
PRNT5
0004ndash0
2nM
[54]
B2DIIaaI126
Phaged
isplay
PRNT5
0002ndash2
nM
E3DIIIa
aG132
Phaged
isplay
PRNT5
0014ndash0
93n
M
FabT
JE12B0
2Hum
anJEV
ENA
Phaged
isplay
Fab
FRNT5
0502120583gmL
[55]
5F10
Hum
anCH
IKV
E2Dom
ainB
EBVtransfo
rmation
IgG11205822
IC50lt100n
gmL
[56]
8B10
Hum
anCH
IKV
E1-E2
E2Dom
ainA
EBVtransfo
rmation
IgG1k
IC50lt100n
gmL
CHK-
152
Murine
CHIK
VE2
aa59
Hybrid
oma
IgG2c
IC50
1ndash3n
gmL
[57]
11E7
Murine
CCHFV
Gn
C-ter
Hybrid
oma
IgG
PRNT8
0diluted
12560
[58]
4-39-C
CMurine
RVFV
G2
Dom
ainIV
Hybrid
oma
IgG
PRNT8
0diluted
1204
80ndash181920
[59]
2C9
Murine
YFV
EHybrid
oma
IgG2a
PRNT9
011
0417D
1105
2Asib
i[60]
5A 7A R3(27)
Hum
anYF
VE
DI-DII
Phaged
isplay
ScFv
PRNT9
001ndash03120583
gmL
[61]
13D6
Murine
TBEV
EDIII
Hybrid
oma
IgGchimeric
PRNT5
019120583gmL
FRNT5
045120583
gmL
IC50
19ndash167120583
gmL
[62]
3B4C
-4(H
y4-26C
)Murine
VEE
VE2
aa182ndash209
Hybrid
oma
IgGhum
anized
PRNT7
0(120583gmL)
394ndash125
[63]
BioMed Research International 9
The structure of the 1A1D-2 bound to EDIII indicates that themAb binds to sites that are transiently exposed during viralldquobreathingrdquo at 37∘C and block infection during attachmentof the virion to the cell In particular this mAb neutralizesDENV1 2 and 3 serotypes and residues K307 and K310 arethemost critical residues for binding of 1A1D-2mAb [46 75]
Finally Shrestha et al immunized mice with a genotype2 strain of DENV1 virus and generated 79 new mAbs 16of which strongly inhibited infection by the homologousvirus and localized to DIII of E protein Surprisingly onlytwo mAbs E105 and E106 retained strong binding andneutralizing activity against all five DENV1 genotypes as wellas being protective in immunocompromised infected miceMoreover E105 and E106 exhibited therapeutic activity evenwhen administered as a single dose four days after inoculationwith a heterologous genotype 4 strain of DENV1 in the samemouse model [32]
243 Other Non-HumanmAbs Other non-human and non-murine as well as humanized mAbs directed against the Eglycoprotein have been described In this regard Goncalvezet al isolated a large panel of anti-E Fab fragments from chim-panzees infected with all four DENV serotypes Howeveronly a limited number of them displayed a cross-neutralizingactivity against DENV1 and 2 and to a lesser extent againstDENV3 and 4 In particular the authors calculated thatamong them the 1A5 mAb that was further humanizedneutralized DENV1ndash4 at a PRNT50 titer of 048 095 32and 43 120583gmL respectively Interestingly the humanized 1A5was also tested for binding and neutralization against theWNVDENV4 chimera JEV strain SA14-14-2 and Langatvirus (LGTV) strain TP 21 giving a PRNT50 titer of 38 21and 28120583gmL respectively Moreover the authors calculatedthat when administered in a dose of 2mg per kg of bodyweight 1A5 would give a serum titer of approximately 40and 20 50 reduction in plaque reduction neutralization test(PRNT50) against DENV1 and DENV2 respectively [76]Furthermore epitope mapping of 1A5 mAb localized to G106within the Flavivirus-conserved fusion loop inDII of DENV2E protein [51]
244 Anti-DENV Human mAbs As anticipated attemptsto isolate neutralizing human mAbs have been morechallenging due to the restricted elicited Ab-repertoire whichrecognizes DIII of E glycoprotein in naıve as well as ininfected patients [77 78] However several groups reportedthe isolation of neutralizing mAbs from infected subjects Inthis regard Setthapramote et al isolated a total of 136 humanhybridoma clones producing specific mAbs against DENVobtained using PBMCs from nine blood samples from fouracute-phase patients secondarily infected with DENV2 andfive convalescent-phase patients Interestingly the authorsfound that most of the acute-phase mAb clones were cross-reactive with all four DENV serotypes with most of themrecognizing the E protein and endowing of neutralizingactivity against all DENV serotypes compared to thosederived from convalescent-phase patients In particularfrom the acute-phase PBMCs 818 were anti-E 66 wereanti-prM and 33 were anti-NS1 while 133 anti-E 133
anti-prM and 533 anti-NS1 mAbs clones were obtainedfrom convalescent-phase PBMCs [79] Previous studiesconfirmed the data obtained by Setthapramote et al onconvalescent-phase patients but this group firstly reportedthe efficient preparation of human mAbs with strongneutralizing activity titers against all four DENV serotypesusing PBMCs from acute-phase patients secondarily infectedwith DENV [80 81]
The group of de Alwis through immunoglobulin deple-tion studies reported that a substantial fraction of DENV-reactive Abs in human immune sera including type-specificneutralizing Abs bound to the intact virion but not torecombinant E proteinThe authors confirmed these observa-tions also isolating human neutralizing mAbs and proposedthat humans produce Abs that neutralize DENV infectionby binding a complex quaternary structure epitope that isexpressed only when E proteins are assembled on a virusparticle [48] Similar findings have been described for severalother viruses including WNV and HIV [82 83] Mappingstudies indicate that this epitope has a footprint that spansadjacent E protein dimers and includes residues at the hingebetween domains I and II of E protein [48]
Recently Costin et al isolated three broadly neutralizinganti-DENVhumanmAbs named 48AD11C and 16DThesemAbswere isolated from three different convalescent patientswith distinct histories of DENV infection All three mAbsrecognized the E glycoprotein with high affinity neutralizedall four serotypes of DENV but mediated ADE of infection inFc receptor-bearing cells at subneutralizing concentrationsMapping studies revealed that all three mAbs bind discontin-uous epitopes within the highly conserved fusion-loop regionof DII (contacting residues W101 L107 andor G109) [37]
25 Yellow Fever Virus (YFV) TheWHO estimates that thereare 200000 cases of yellow fever virus (YFV) infections and30000 related deaths every year especially in Sub-SaharanAfrica In fact different to themajority of human arbovirusesand similarly to DENV and CHIKV YFV has expanded itshost range to include humans as an amplifying host [1]Symptoms occurring from 3 to 6 days after infection withYFV consist mostly in fever chills anorexia lumbosacralpain nausea and vomiting This syndrome lasts about 3days in most cases and is sometimes followed by a one-daylasting period of remission Fever will reoccur together withdark hematemesismelena petechiae and other hemorrhagicsymptoms Convalescence is characterized by deep asthenialasting up to two weeks Rarely prognosis can be fatal Aneffective live-attenuated vaccine (17D) derived from the Asibistrain by serial passages in chicken embryos is available [84]
251 mAbs against YFV In 1983 Schlesinger et al produceda battery of mAbs after immunization of BALBc mice byinjection of 17D YFV vaccine strain [84] Among them thespecificity towards the E and NS1 proteins was describedeven though anti-NS1 Abs were not able to show neutralizingactivity The 13 IgG and 1 IgM anti-E protein mAbs wereclassified in five groups according to their specificity GroupA consisting of the only IgM produced in this experiment
10 BioMed Research International
could neutralize 17D YFV virus only group B 17D and Asibistrains only group C Asibi strain only group D could onlyneutralize Asibi but not 17D strain showing also reactivityagainst DENV2 Zika or Banzi viruses All of the four mAbsbelonging to this group were able to cross-react with Zikaandor Banzi viruses and two (4E11 and 5H3) neutralizedDENV2 virus with 4E11 neutralizing also Banzi virus The3E9 mAb the only component of the group E could notneutralize 17D nor Asibi strains
All of the IgG mAbs resulted able to protect bothprophylactically and therapeutically BALBc or CD-1 micefrom lethal intracerebral challenge with 17D-204 strain [85]The chimeric form of the group B mAb 2C9 IgG (2C9-cIgG) was able to provide AG129 mice a 72 survival whenadministered 24 hours before infection with 17D-204 strainIts murine form could provide the survival of the 95 ofthe mice Appreciable results were also obtained when bothmurine 2C9 and 2C9-cIgG were administered 24 hours post-infection with the survival of 70 and 20 of mice respec-tively Viral presence was not detected in survivingmice [60]
In 2005 Daffis and colleagues constructed two Ab-phage libraries by cloning the repertoire of YFV-infectedpatients Panning was then performed with YFV-17D virionsThe scFv-5A 7A and R3(27) showed a neutralizing activityspanning from50 to 100 inPRNTassays against bothYFV17D-204-WHO and Asibi strains In further tests reactivitywas observed against wild-type YFV strains of West Africagenotype I and II (Nigeria 1987 and Asibi strains resp)and EastCentral Africa (CAR 1986 Ethiopia 1961 strains) Aconcentration from 01 to 03120583gmL could yield a 90 plaquereduction Reactivity was also observed against the strainSenegal 1990 in a lesser extent Production of escape variantscould demonstrate that scFv-5A 7A and R3(27) epitopes arebuilt up by residues extensively separated in themonomers ofE glycoprotein that however result in closed proximity whenthe homodimeric form of E is constituted [61]
26 West Nile Virus (WNV) The West Nile virus (WNV) isan epidemic neurotropic virus estimated to be responsibleof about 36000 cases and 1500 deaths registered in theUnited States between 1992 and 2012 [86] WNV antigenicityallows its classification into the Japanese encephalitis virus(JEV) serocomplex Genomic analysis has revealed two maingenetic lineages of WNV lineage I viruses circulating in theUSA Europe the Middle East Africa India and Australiaand lineage II viruses isolated from Sub-Saharan Africa andMadagascar [1]
WNV was firstly identified in 1937 and is endemicin many countries of Africa Middle East and West AsiaHowever after 1990 frequent outbreaks of WNV infectionswere reported in Romania Israel and later in North Americaand across the USA in 1999 and recently in 2012 [87]
Most of WNV infected individuals do not develop symp-toms while about 20 develop a self-limiting illness calledWest Nile fever Acute symptoms include fever tirednessand swollen lymph glands but in a minority of cases alsoencephalitis with long-term deficits in cognitive functionand motor skills have been reported However in less than
1 of infected patients WNV having crossed the blood-brain barrier is responsible of neuroinvasive and potentiallylethal form of the disease In these cases degeneration andapoptosis upon infection of neurons and the consequentinflammatory response can occur Related symptoms includehigh fever coma muscle weakness and paralysis Immun-odepression and advanced age have been correlated with anaugment risk to develop a severe disease [87] There are nospecific treatment options or licensed vaccines for humans[88]
WNV is transmitted by infected mosquitos and initialreplication is thought to occur primarily in dendritic cells inthe skin which migrate to secondary lymphoid tissues wherethe replicating virus enters the circulation To date onlyefficacious WNV veterinarian vaccines have been licensedwhile there are no licensed vaccines for protection againstWNV in humans [88]
261 mAbs against WNV Broad-spectrum antivirals suchas type I IFN-120572 ribavirin mycophenolic acid in WNVinfection showed ineffective results in vivo despite in vitrothey showed some activity [89]
However experiments in murine models extrapolationof clinical data as well as passive administration of pooledimmune-120574-globulins (OmriGam) containing a significanttiter of neutralizing Abs before and after infection showedan important role of the humoral immune response incontrolling viremia and prevent viral dissemination Indeedthe development of a neutralizing mAb-based therapy seemsto be encouraging for a possible treatment of infected patients[90ndash92]
Similarly to what has been observed for DENV only anti-E Abs have been identified as neutralizing and protectiveAlso anti-NS1 Abs have shown a protective role howevertheir mechanism of protection has not yet been elucidated asthe NS1 protein is secreted from infected cells and not presenton the virions [93ndash95]
In this regard similarly to what has been describedfor DENV studies performed using a naıve human scFvlibrary for panning with purified WNV E protein supportthe hypothesis that no Abs against the neutralizing DIII canbe isolated However DIII specific Abs were isolated in asubsequent study using immunoglobulin libraries obtainedfrom threeWNV infected patients for biopanning on purifiedinactivated virus virus-like particles consisting of prM andE proteins or recombinant E glycoprotein Although theproportion of DIII-specific mAbs was low (8) comparedwith anti-DII mAbs (47) Two out of the four anti-DIIImAbs were potently neutralizing and protective in vivowhereas only three out of the 24 anti-DII were weaklyneutralizing in vitro and non-protective in vivo In particularresidues that are critical for neutralization lies on the regionsspanning amino acids 305ndash312 330ndash333 and 365 that arelocated on adjacent exposed loops ofDIIIHowever sequencealignment of E protein of different flaviviruses such as DENVand JEV revealed a considerable variation compared tothe whole E protein This observation further suggests thatdifferently from broadly cross-reactive anti-DII Abs anti-DIII neutralizing Abs are virus-type specific However of
BioMed Research International 11
these mAbs two (CR4374 and CR4353) protected mice fromlethal WNV challenge at 50 protective doses of 129 and357 120583gkg of body weight respectively [50]
Sanchez et al using three different immunization strate-gies (ie inactivated virus naked DNA and recombinantprotein) isolated ninemurinemAbsmost of which bound toconformation-dependent epitopes in DIII of the E protein Inparticular neutralizing mAbs named 8B10 11C2 10C5 and17C8 were obtained from mice immunized with inactivatedvirus alone or in combination with a DNA plasmid andbound to the same region of DIII with high affinity Incontrast mAbs obtained by immunization with a solubleversion of the E glycoprotein did not exhibit neutralizingactivity These non-neutralizing mAbs were cross-reactivewith several other flaviviruses including SLEV JEV YFVand Powassan virus confirming the conserved nature ofFlavivirus non-neutralizing epitopes [96]
Gould et al isolated 11 unique human single-chain vari-able region Ab fragments (scFvs) that bind the E protein ofWNVAmong them a humanmAb namedmAb11 expressedas a scFv-Fc fusion protein was further characterized Itrecognizes the fusion loop at the distal end of DII ofthe WNV E protein and cross-reacts with all four DENVserotypes and provides protection against DENV2 and 4as well as WNV [42] Moreover therapeutic studies of thismAb in WNV-infection model mice provided substantialprotection when administered after 5 days post-infectionInterestingly a neutralization escape variant of this mAbfailed to cause lethal encephalitis (at higher infectious doses)or induce the inflammatory responses associated with blood-brain barrier permeability in mice compared to the parentalWNV suggesting an important role for the fusion loop inviral pathogenesis [43]
Oliphant et al isolated an anti-DIII E protein mAbnamed E16 fromhybridomas obtained after immunization ofmice with recombinant WNV E protein which neutralizedall WNV strains with PRNT50 values of 4 to 18 ng andPRNT90 values of 53 to 297 ng One hundred microgramsof mAb protected greater than 90 of mice from lethalinfection and even a single 4120583g treatment of E16 on day 2 afterinfection prevented mortality Moreover humanization ofthis mAb confirmed as therapeutically effective in mice [44]Subsequent studies revealed that mAb E16 neutralization ismediated by engagement of four discontinuous segmentsof DIII including the amino-terminal region (amino acidresidues 302ndash309) and the three connecting loops BC (aminoacid residues 330ndash333) DE (amino acid residues 365ndash368)and FG (amino acid residues 389ndash391) Moreover no ADE ofinfection was detected when E16 mAb was used at saturatingconcentrations [45 47]
Furthermore results of a Phase I safety study of thehumanized E16 mAb (designated MGAWN1) have beenreported and suggested that doses of up to 30mgkgwerewelltolerated with few mild adverse events and would providean excess of virus neutralizing activity for 3-4 weeks aftertreatment However a case of anti-MGAWN1 Ab elicitationoccurred with the consequent increased rate of clearance andindeed impacting efficacy A Phase II safety and efficacy studyof MGAWN1 is ongoing [97] Furthermore the possibility of
preventing or treating WNV-induced memory deficits wasrecently investigated In this study hamsters were treatedintraperitoneally with 32mgkg ofMGAWN1mAb at 45 daysafter subcutaneously challenging with WNV InterestinglyMGAWN1 prevented mortality weight loss and improvedfood consumption of WNV-infected hamsters compared tocontrols [98]
Recently Lelli et al isolated six anti-E mAbs frominactivated-WNV immunized mice In particular three ofthem (3B2 3D6 and 4D3) neutralized lineage I and II WNVwith the first two recognizing the same epitopes located onthe distal lateral surface of DIII (critical amino acid residueK307) Conversely 4D3mAb recognized a novel neutralizingepitope on DII (critical residues S276 and T278) Indeedfurther protective and therapeutic studies are needed toascertain their neutralizing activity in vivo [99]
Finally to conclude like DENV infection Abs directedagainst the M protein are not protective and neutralizingMoreover in humans a skewed humoral immune responseagainst DII has been frequently observed and confirmedalso with the hybridoma technology [50] The isolation andelicitation of neutralizingAbs directed against the fusion loopand DIII of the E protein represent thus the most challengingand promising goal for the development of new effectivetherapeutic inhibitors and immunogens respectively
27 Tick-Borne Encephalitis Virus (TBEV) Tick-borne ence-phalitis virus (TBEV) is one of the most dangerous agentscausing human neuroinfections occurring mostly in Europeand Asia and with a potential fatal prognosis [100] TBEV isbelieved to cause 3500ndash10000 human cases of encephalitisin Europe per year with a high morbidity in Russia CzechRepublic Austria and Germany In particular between 1990and 2007 an average of 8755 cases of TBE was reported peryear in Europe and Russia Despite the fact that Russia isthe country with most infections registered annually CzechRepublic incidence is among the highest in Europe with 400ndash1000 clinical cases reported every year [101]
Three subtypes of TBEV are classified namely EuropeanSiberian and Far-Eastern sharing most of the genetic andantigenic features [102] In fact a high degree of antigenichomogeneity between different strains of TBEV has beendescribed [103]
Clinical manifestations vary among the subtypes butusually they start with a short febrile period of 7ndash14 daysafter the tick bite Fatigue headache and pain in the neckshoulders and lower back together with high fever andvomitingmay be present [100]Thesemanifestations are oftenfollowed by an asymptomatic phase lasting from 2 to 10days after remission from the fever with possible progres-sion to neurological disease Neurological symptoms includemeningitis encephalitis myelitis and radiculitis Mortalityoccurs in 1-2 of the European subtype-infected patientsbut fatal prognosis can occur in up to 20ndash40 of the Far-Eastern subtype-infected patients [104] Mortality rates ofthe Siberian subtype are similar to those observed for theEuropean [105]
Effective and safe vaccines against TBEV produced frominactivated virus have been developed and licensed for their
12 BioMed Research International
use in humans However an emergency therapy in theabsence of a mass immunization is needed as no effectivetreatments are yet available [106]
271 mAbs against TBEV Levanov et al described the chim-erization of two murine mAbs (13D6 and 10C2) directedagainst the DIII and DII respectively of the TBEV E glyco-protein The chimeric mAbs present binding characteristicssimilar to the parentalmAbsMoreover as the parentalmAbsonly the chimeric mAb 13D6 was able to neutralize TBEVinfectivity in vitro In particular neutralization studies withthe murine chimeric and scFv forms of mAb 13D6 were per-formed In particular murine 13D6 showed an IC50 titer of115 120583gmL in Focus Reduction Neutralization Tests (FRNT)against TBEV strain 205 and of 29 120583gmL in PRNT againstTBEV strain Softjin Chimeric 13D6 showed an IC50 titer of45 120583gmL in FRNT against TBEV strain 205 and of 19 120583gmLin PRNT against TBEV strain Softjin ScFv 13D6 showed anIC50 titer of 167 120583gmL in FRNT against TBEV strain 205and of 112 120583gmL in PRNT against TBEV strain Softjin [62]
28 Japanese Encephalitis Virus (JEV) Japanese encephalitisvirus (JEV) is a mosquito-borne flavivirus that over thepast few decades has caused several outbreaks throughoutChina Southeast Asia Australia and Papua New Guineawith a prevalence recently estimated to be of about 70000casesyear [107ndash109] A 40 mortality was recorded in someof the JEV-affected areas Moreover many survivors facesome neurological problems and complications [108] Since1995 the disease has also emerged in Non-Asian regions suchas Northern Australia [110 111] The situation in SoutheastAsia however is further complicated by the overlappingepidemics of JEV and DENV as well as sporadic casesof WNV infections detected in some of the affected areasparticularly in India [112]
Effective vaccines both live-attenuated and inactivatedstrains of JEV have been developed and licensed in majorcountries However as for the other arthropod-borne Fla-vivirus members no specific antiviral drugs are currentlyavailable [4]
281mAbs against JEV Aspreviously described for the otherFlavivirusmembers many groups isolated and characterizedanti-JEVmAbs showing different neutralizing and protectiveproperties In this regard Gupta et al used combinations ofanti-E JEV mAbs (Hs1-4) in mice protection experimentsIn particular they found that the singularly mAb protectionranged from 45 to 65 when 100120583g of mAb were adminis-tered while equimolar combinations of two or three mAbsgave 85ndash90 or 100 protection respectively [113] In similarexperiments Lee et al demonstrated that a Flavivirus anti-NS1 mAb named 16NS1 cross-reacted with JEV as well asWNV and exhibited protective activity against WNV as wellas a lethal JEV infection However no neutralizing activitywas observed using this mAb against both WNV and JEV inin vitro experiments suggesting the participation of otherAb-mediatedmechanisms in vivo In particular 95ofmicewere
protected when 500120583g of mAb were administered intraperi-toneally and concomitantly to intramuscular injection of JEV
Overlapping peptide mapping analysis combined withsite-specific mutations identified the 116KAWGKSILFA125region and critical amino acid residues W118 and I122 as16NS1 mAb epitope highly conserved in WNV and JEVstrains [114]
Arakawa et al isolated from a combinatorial human Fablibrary constructed from peripheral blood lymphocytesobtained from JEV hyperimmune volunteers Among 188randomly selected clones FabTJE12B02 showed the best 50focus reduction endpoint at the concentration of 502 120583gmLagainst the JEV strain Nakayama [55]
Goncalvez et al isolated three mAbs named Fabs A3B2 and E3 by repertoire cloning from chimpanzees initiallyimmunized with inactivated JE-VAX and then boosted withattenuated JEV SA14-14-2 In particular these mAbs reactedwith epitopes in three different E domains in DI (amino acidresidue K179) in DII (I126) and in DIII (G132) for Fabs A3B2 and E3 respectively Moreover these Fabs as well as thederived humanized counterpart mAbs exhibited high neu-tralizing activities against a broad spectrum of JEV genotypestrains Moreover these mAbs exhibited a 50 protectivedose of 084 120583g (B2) 58120583g (A3) and 247120583g (E3) in mousemodels Finally administration of 200 120583gmouse of mAb B2one day after otherwise lethal JEV infection protected 50of mice and significantly prolonged the average survival timecompared to that of mice in the unprotected group [54]
29 St Louis Encephalitis Virus (SLEV) St Louis encephalitisvirus (SLEV) was first discovered as the mosquito-borneagent responsible for over 1000 cases of encephalitis duringa 1933 summer outbreak in St Louis (Missouri) and nowis a reemerging human pathogen widely distributed in theAmerican continent causing several human encephalitisoutbreaks over the last 80 years Additional epidemics haveindeed occurred from 1964 to 2006 in the Americas rangingfrom the US to Argentina and Brazil As a member of theFlavivirus genus the SLEVE glycoprotein ectodomain is 68identical to serocomplex-related JEVEbut only 46 and 40identical to those ofDENV2 andTBEV viruses fromdifferentserocomplexes [115]
291 mAbs against SLEV In 1983 Roehrig et al isolatedtwenty-one hybridomas producing murine mAbs specificfor the E glycoprotein of SLEV strain MSI-7 Serologicreactivities were initially determined by cross-reactivity indi-rect immunofluorescence assays using 22 strains of SLEVand 8 other related flaviviruses Four groups demonstratingtype- subcomplex- supercomplex- and group-specific reac-tivity patterns were identified Analysis of hemagglutination-inhibition and virus neutralization subdivided the cross-reactivity groups into eight epitopes (E-1a b c d E-2 E-3 and E-4a b) one of them following localized in DII (4b)[52 53]
Moreover among the previously described isolated anti-WNV mAbs by Gould et al the chimeric scFv-Fc 79 effec-tively neutralized also SLEV resulting in gt80 of PRNT80when used at 5 120583gmL [42]
BioMed Research International 13
3 Alphaviruses
Major human arboviruses in the Togaviridae family Chikun-gunya (CHIKV) and Venezuelan equine encephalomyelitis(VEEV) viruses belong to the Alphavirus genus which iscomposed of viruses with an icosahedral nucleocapsid sur-rounded by a lipid envelope and glycoprotein spikes Thestructural proteins of alphaviruses arise through co- andpost-translational processing of a polyprotein encoded by asingle positive-stranded RNAproducing the capsid (C) PE26 K and E1 The PE2 glycoprotein is a precursor containingthe E3 glycoprotein fused to the amino terminus of the E2envelope glycoprotein The PE2 glycoprotein is followed by6K a small membrane-associated protein and E1 the secondpolypeptide component of glycoprotein spikes
Trimerized heterodimers of the E1 and E2 viral glyco-proteins form the surface spikes and contain determinantsof viral tropism and virulence The E3 glycoprotein acts as asignal for transport of PE2 across themembranes of the roughendoplasmic reticulum and may promote the formation andintracellular transport of E1-PE2 heterodimers to the cellsurface During transport to the cell surface PE2 undergoesa maturational cleavage event by a furin-like protease toproduce E2 and E3 [116]
The E2 glycoprotein promotes specificity of virus bindingto the host cell surface and is a target of the humoral immuneresponse The E1 glycoprotein mediates fusion of the virionenvelope with the membranes of acidified endosomes allow-ing release of the nucleocapsid into the cytoplasm and theonset of viral replicationAlphavirus E1 shares no appreciablesequence identity to Flavivirus E protein Despite differencesin their amino acid sequences and arrangements on the viralparticle the structures of E and E1 are remarkably similarIndeed the conservation of three domains has been welldocumented in crystal structures of flavivirus E ectodomainsfrom TBEV DENV JEV and WNV as well as Alphavirus E1from Semliki Forest virus (SFV) and CHIKV Abs to the E1glycoprotein do not typically neutralize virus infectivity invitro but can protect against lethal challenge in animals
By analogy with the prM protein of dengue Flavivirusfurin cleavage of PE2 may begin after transit of an acidiclate component of the Golgi body where E3 is thought tosuppress the acid pH-triggered activation of glycoprotein E1fusion capability Protective anti-E3 mouse mAbs have alsobeen described [117]
31 Chikungunya Virus (CHIKV) The chikungunya virus(CHIKV) belongs to the Semliki Forest clade and was firstlyisolated in 1953 in Tanzania during an epidemic outbreakfollowing occurred also in Asia and Africa In the last fivedecades several CHIKV outbreaks have been described bothin Africa and Asia separated by gaps lasting from two totwenty years In 2005-2006 about 300000 cases out of785000 inhabitants were reported in La Reunion island witha fatal prognosis for 237 of the patients [117] Neither Europenor the Americas have had outbreaks of CHIKV so far exceptfor imported isolated cases Three genotypes of CHIKV aredescribed Asian EastCentralSouth African (ECSA) andWest African with an amino acid identity spanning from
952 to 998 Recent epidemics in Africa and Indiansubcontinent were caused by strains belonging to the ECSAgenotype Transmitted by Aedes mosquitoes CHIKV ismaintained in the human population by human-mosquito-human transmission [1] The disease is characterized bydengue-like symptoms such as chills high fever headacheand persistent myalgia and a further incapacitating arthralgia(from which the name chikungunya) which affects 40 ofinfected subjects However prognosis is rarely fatal [116]
There is currently no commercial vaccine and antiviraltreatment forCHIKV although some candidate vaccines havebeen tested in humans In this regard in 2000 US Armyperformed a Phase II clinical trial testing a live-attenuatedCHIKV vaccine (TSI-GSD-218) derived from a 1962 strain(15561) of an outbreak in Thailand Out of 58 every patientdeveloped neutralizingAbs and 5 lamentedmild tomoderatejoint pain [117] A phase III trial of this candidate vaccineis ongoing Furthermore a new formulation using virus-likeparticles was able to induce neutralizing Abs in macaquesagainst different CHIKV strains [117 118] Indeed it has beendescribed that infection seems to elicit long-lasting protectiveimmunity and cross-protection among CHIKV and otheralphaviruses
CHIKV entry into host cell is demonstrated to be medi-ated by envelope glycoproteins E1 and E2 which allowvirus fusion to cell membrane in low pH conditions andrecognition of an unknown cellular receptor respectively[116 119ndash121] Even though the genome replication relies onan error-prone RNA-dependent RNA-polymerasis CHIKVstrains S27 and 05115 Reunion showed in E1 and E2 alow amino acid variation of 068 and 33 respectivelyand such a low variability may be needed for an effectivereplication in two phylogenetically distant hosts [116 122]Recently the crystal structure of E1 and E2 heterodimerhas been resolved both at low pH and natural conditionsshedding light on E1 and E2 ectodomainsrsquo structure Inparticular similarly to flaviviruses E protein E1 ectodomainis made of the N-terminal DI the DII containing the fusionloop and the DIII at the C-terminal On the other hand E2ectodomain comprises theN-terminal domainA the domainB supposed to interact with the host cellrsquos unknown receptorand domain C at the C-terminal E1 DIII and E2 domain Care located close to the viral membrane [123 124]
311 mAbs against CHIKV Warter et al described 5F10 and8B10 two human mAbs which strongly neutralized severalCHIKV isolates in vitro without cross-reactivity againstother alphaviruses but to Onyong-nyong virus [56] Mixedpreparation of 5F10 and 8F10 did not show neither synergisticnor addictive effect in vitro and studies upon escape mutantsdemonstrated that 5F10 mAb binds at the tip of the E2domain B while 8B10 recognizes residues close to E1 fusionloop and amino acids within E2 domain A which form atransitional epitope under low pH conditions The authorsspeculated that 5F10 and 8B10 may inhibit CHIKV entry andfusion to the cell membrane respectively It is worth notingthat the previously mentioned escape mutants displayedmutations associated with reduced viral fitness in vitro evenafter 13 neutralizationamplification rounds In the same
14 BioMed Research International
study cell-to-cell transmission was firstly demonstrated as anescapemechanism enhanced by the E2mutation R82G [122]
In further in vivo studies 5F10 and 8B10 significantlydelayed CHIKV-caused death of AGR129 mice both in pro-phylactic and therapeutic tests Interestingly in therapeutictreatment these mAbs showed a synergistic effect whenadministered in combination with the total amount of mAbsinjected being the same of the single-mAbs Possibly thesetreatment did not allow themice to survive CHIKV challengedue to Ig clearing and 6ndash10 days half-life usually described forhuman mAbs [125]
Recently Pal et al cloned thirty-six murine mAbs ableto neutralize the ECSA La Reunion 2006 OPY-1 strain ofCHIKV (CHIKV-LR) the majority of which also neutralizeinfection of other strains corresponding to the Asian andWest African genotypes In particularmAbCHK-152 showedthe highest and broadest neutralizing activity with FRNTvalues indicating an IC50 value of 1 to 3 ngmL dependingon the viral strain used Among the thirty-six mAbs four(CHK-102 CHK-152 and CHK-166 CHK-263) could pre-vent lethality of immunodeficient Ifnarminusminus C57BL6 micewhen administered one day before exposition with CHK-152 and CHK-263 showing the ability to protect mice at thelowest dose (10 120583g) Therapeutic studies were performed onthese mAbs administering a single dose of 100 120583g 24 hoursafter infection The highest activity was described for CHK-166 able to protect 63 of the mice Testing the combinedactivity of the mAbs in therapeutic studies administration ofCHK-166 plus CHK-152 in a dose of 250120583g each turned out tobe themost effective protecting 71 of the IfnarminusminusC57BL6mice 60 hours after infection [57]
32 Venezuelan Equine Encephalomyelitis Virus (VEEV)Venezuelan equine encephalomyelitis virus (VEEV) is main-tained in a natural transmission cycle between mosquitoesand small rodents The first documented outbreaks occurredin the 1930s and several epidemics have been reported so farin Latin-American countries such as Venezuela ColombiaPeru Ecuador Bolivia Honduras Mexico and Panama withhundreds of thousands of human cases being reported anda case-fatality rate up to 1 during the 1969 Ecuador out-break Clinical manifestations of VEE are indistinguishablefrom Dengue [126] Six serogroups (IndashVI) are currentlyrecognized within the VEEV complex VEEV caused humanand equine outbreaks in the Americas for nearly a centuryEquine epizootics have high mortality (38ndash83) leading toa high viremia followed by a lethal encephalitis and often tohuman epidemics involving thousands of cases and hundredsof deaths Indeed VEEV is infectious for humans by theairborne route and has been responsible for a number oflaboratory infections In humans the disease is usually self-limiting with a febrile illness with 1ndash4 of cases progressingto severe encephalitis It is a hazard to laboratory workershas been developed as a biological weapon and is a potentialbioterrorist agent [127] There are no antiviral drugs andvaccines licensed for the treatment of VEEV infection inhumans Indeed although experimental live-attenuated vac-cines have been developed (eg TC-83) with good levels ofprotection in equine and mice in humans they may fail to
give protection in the majority of cases [128] Consideringthe protective role exerted by humoral immune response inanimal models it could be concluded that treatment withspecific IgG may have a beneficial antiviral effect in humanairborne infections with pathogenic strains of VEEV
321 mAbs against VEEV Several works support that mAbsmay protect against airborne VEEV as well as after airborneexposure toVEEVAb reactivity to both surface glycoproteins(E1 and E2) is associated with neutralization of the virusin vitro and passive protection against virus challenge Aseries of anti-E1 and anti-E2 mouse and humanized mAbshave been described recently Among them Phillpotts et alexamined two anti-E2 murine mAbs named 1A4A-1 and1A3A-9 both having potent protective activity against subcu-taneous VEEV challenge in mice Both mAbs had a similarhalf-life (58 and 100 days) in mouse serum after a singleintraperitoneal dose suggesting that mAbs delivered by thisroute at or around the time of VEEV infection persists athigh levels in the blood as well as secretions in respiratorytransudation throughout the clinical course of the disease Inparticular both mAbs administered 24 h prior to airbornechallenges had a substantial protective effect (90ndash100)Treatment of mice 2 or 24 h after airborne infection with asingle intraperitoneal dose of 100 120583g of 1A3A-9 mAb led toapproximately 50 survival There was no beneficial effectwhen mAb treatment was delayed to 72 h Moreover therewas evidence of synergy in vitro in PRNT between 1A3A-9and 1A4A-1 as has been demonstrated for other virusesand mAb pairs However no synergy was found in mouseprotection when mAbs were delivered intraperitoneally as amixture in equal parts to animals challenged 24 h previouslywith VEEV [129]
In a subsequent study Phillpotts et al reported two otherprotective mAbs The 3B2A-9 mAb protected against allthe serogroup I strains while 1A3B-7 mAb protected wellagainst challenge with all of the viruses tested Both 3B2A-9 and 1A3B-7 protected against airborne exposure to theIAB serogroup virus strainTrinidad donkey (TrD)Howeverthere was no evidence of synergistic protection when thesemAbs were combined in equal proportions An intraperi-toneal dose of 10 120583g was sufficient to protect 50 of themice with either mAb [130] According to other data themechanism of protection did not appear to depend uponneutralization [131] Similar results were obtained by OrsquoBrienet al which described a non-neutralizing mAb (IgG2a)named CUF37-2a able to protect 50 of mice from asubcutaneous VEEV challenge when a dose of 915 120583g of mAbwas administered 24 h prior to challenge [132]
In a following study the 1A3B-7 mAb was humanized(and following reported as Hu1A3B-7) maintaining the samefeatures of its murine counterpart In particular evaluation ofin vitro studies indicated that Hu1A3B-7 retained both broadspecificity and neutralizing activity Furthermore in vivoexperiments showed that Hu1A3B-7 successfully protectedmice against lethal subcutaneous and aerosol challenges withVEEV strain TrD Moreover the effectiveness of the human-ization process was determined by assessing proliferationresponses in human T-cells to peptides derived from the
BioMed Research International 15
murine and humanized versions of the VH and VL domainsThis analysis showed that the number of human T-cellepitopes within the humanized Ab had been substantiallyreduced indicating that Hu1A3B-7 may have low immuno-genicity in vivo [133]
Similarly Hunt et al described a humanizedmurinemAbderived from the 3B4C-4 mAb recognizing the E2c epitope(amino acid residues 182ndash209) and following named Hy4-26C showing neutralizing activity similar to the murinemAb Moreover it was protective (70ndash100 of survival) at100 ng dose in mice animal model intraperitoneal inoculatedand at 500120583g for intranasal challenge (80) Therapeuticstudies in mice revealed that Hy4-26C was able to cure miceup to 24 h following infection at 10 120583g on the contrary themouse mAb when given 1 hour after virus challenge [63]
In another work Hunt et al isolated two neutralizinghumanized Fabs F5 and L1A7 employing a blocking strategy[134 135] The anti-E2 specific F5 IgG had a 70 PRNTendpoint of 10 ngmL equivalent to that described for themost effective neutralizing anti-VEEVE2mAbs In particularthe E1-specific hFab L1A7 had a PRNT endpoint of 3120583gmL300-fold lower than F5
Further studies revealed that F5 epitope is located inthe 115ndash199 amino acid region Moreover F5 IgG had potentability to protect mice from infection by either route whenadministered 24 h before exposure however mice treated24 h and 48 h after aerosol exposure developed central ner-vous system infections but exhibited no clinical signs ofdisease [135]
Hu et al reported that passive immunization with thehumanized chimericmousemAbhu1A4A1IgG1-2A inmice at50 120583g 24 h before or after virulent VEEV challenge providedcomplete protection indicating that hu1A4A1IgG1-2A haspotent prophylactic and therapeutic effects against VEEVinfection [136]
Anti-VEEV mAbs isolated from a non-human primategene library has also been reported In particular thehumanized chimeric scFv-Fc ToR67-3B4 recognized viableas well as formalin and 120573-propiolactone inactivated virusparticles It detected specifically the viral E1 envelope proteinof VEEV but did not react with reduced viral glycopro-tein preparations suggesting that recognition depends uponconformational epitopes The recombinant Ab was able todetect multiple VEEV subtypes and displayed only marginalcross-reactivity to otherAlphavirus species except for Easternequine encephalitis virus (EEEV) In addition the scFv-Fcfusion described here might be of therapeutic use since itsuccessfully inactivated VEEV in a murine disease modelIn particular when the recombinant Ab was administered 6hours after challenge 80 to 100 of mice survived lethalVEEV IAB or IE infection Forty to sixty percent of micesurvived when scFv-Fc ToR67-3B4 was applied 6 hours afterchallenge with VEEV subtypes II and former IIIA [137]
4 Bunyaviridae
TheBunyaviridae family contains human arboviruses belong-ing to the Orthobunyavirus Phlebovirus and Nairovirus
genera Bunyaviridae includes enveloped viruses with a frag-mented single-stranded RNA genome of negative polarityTheir tripartite genome consists of a small (S) a medium(M) and a large (L) fragment and the envelope glycoproteinsGn and Gc that are cleaved out of a polyprotein synthesizedby the M fragment These glycoproteins have been provedto mediate the formation of the virus particle to play a rolein the interaction with cell surface receptors to mediate theentry of the virus into cells and to serve as targets for themajority of neutralizing Abs described so far
Orthobunyavirus transmitted through mosquitoes ormidges vectors are divided in 18 serogroups based on cross-titrations in haemagglutination inhibition assays and neutral-ization assays and correlating with main vector preferencesHowever the most clinically important viruses belong onlyto two serogroups the California encephalitis and the Simbuserogroups
Crimean-Congo hemorrhagic fever virus (CCHFV) isconsidered the only one of clinical relevance within theNairovirus genus which uses tick as main vector and is divid-ed in seven serogroups while Toscana virus and Rift Valleyfever virus (RVFV) which are transmitted by sandflies andmosquitoes respectively belong to the Phlebovirus genus[138]
MAbs against theNairovirusCCHFV and the PhlebovirusRVFV have been described
41 Crimean-Congo Hemorrhagic Fever Virus (CCHFV)Crimean-Congo Hemorrhagic Fever Virus (CCHFV) dis-tribution covers the greatest geographic range of any tick-borne virus known as viral isolation andor disease hasbeen reported from more than 30 countries in SoutheasternEurope Africa Asia and Middle East [139 140] Namelyfrom 1953 to 2010 about 6000 human cases were reportedin Southeastern Europe CCHFV causes sporadic outbreakswith mortality rates ranging from 10 to 80 [141 142] Asignificant variation in the time of incubation has beendescribed [139] Prehemorrhagic symptoms include highfever chills headache photophobia and back and abdominalpains Among other symptoms neuropsychiatric changeshave been reported in some patients In severe cases 3ndash6 daysafter the onset of the disease hemorrhagic symptoms occurwith petechiae ecchymosis bleeding in the form of melenahematemesis epistaxis [139]
In comparisonwith the nucleocapsid proteins inCCHFVGn and Gc show a higher degree of antigenic variabilityprobably due to their exposition to the host immune systemhypothesis corroborated in the description by Hewson et alin 2004 of four M segment phylogenic groups namely M1M2 M3 and M4 [141 142]
411 mAbs against CCHFV Blackburn et al firstly describedin 1987 murine anti-CCHFV mAbs able to recognize nucle-ocapsid proteins but no neutralizing activity has beendescribed thus far [143]
On the other hand Bertolotti-Ciarlet et al produced apanel of murine mAbs recognizing conformational epitopes
16 BioMed Research International
within theGn andGc glycoproteins expressed by the CCHFVstrain IbAr10200 The effectiveness of the mAbs was studiedby performing PRNT80 and none of the anti-Gn mAbsshowed neutralizing activity although many of the anti-GcmAbs neutralized IbAr10200 strain in vitro with clones 8A15E3 12A9 6C2 and 9H3 showing activity at a gt5120 dilutionMAbs 11E7 and 30F7 exhibited neutralizing activity at 2560dilution A suckling mice protection test was thereforeperformed for these mAbs The previously mentionedanti-Gc Abs were capable to protect mice to an appreciabledegree when applied 24 hours before and in a weakermanner 24 hours after virus challenge Many of the anti-GnmAbs were able to confer significant protection to IbAr10200both 24 hours before and after virus administration Therelevant effectiveness of the anti-Gn mAbs 6B12 10E1113G8 and 10G4 suggests that these mAbs may possess someneutralizing activity due to Ab-based effector mechanisms(eg ADCC) [58]
In further studies the same group characterized thebroadly cross-reactive neutralizing activity of the previouslydescribed murine mAb 11E7 able to bind conformationalepitopes within the C-terminal region of the Gn glycoproteinof all CCHFVM groups [58 141]
42 Rift Valley Fever Virus (RVFV) Similarly to CCHFV RiftValley Fever Virus (RVFV) causes sporadic outbreaks Thelargest epidemic occurred in Egypt in 1977 with an estimated200000 infections 18000 patients manifesting symptomsand 600 fatal prognoses [144] Symptoms include hemor-rhagic episodes fever encephalitis and blindness [145]
The major target of the humoral immune response is theRVFV glycoproteins Gc (G1) and Gn (G2) ranging fromnucleotides 480ndash2090 and 2091ndash3614 respectively [144]
421 mAbs against RVFV Within G2 glycoprotein are des-cribed three neutralizing antigenic sites namely epitope Inucleotides 792ndash893 (amino acid residues 258ndash291) epitopeII nucleotides 1164ndash1196 (amino acid residues 382ndash392) andepitope IV nucleotides 858ndash917 (amino acid residues 280ndash299) [59 145]
Despite the high degree of variability observed inCCHFVglycoproteins RVFV G2 presented an unexpected high con-servation among 22 isolates [59] In the same study onlythe LUNYO and 900060 isolates were resistant to in vitroneutralization by murine mAb 4D4 (recognizing epitope II)All other isolates were neutralized with PRNT80 titers from10240 togt81920On the other hand only SNS isolate showedreduced sensitivity to 4ndash39-CC (epitope IV) All other isolateswere neutralized with PRNT80 titers from 20480 to gt81920Protection studies on mice would be of interest for thedevelopment of a passive immunization prophylaxis [59]
Protection studies from lethal South African RVFVAN1830 strain infection were successfully performed on abattery of anti-G1 and anti-G2 mAbs produced by Besselaarand Blackburn with the strongly neutralizing anti-G1 mAb3E5 and anti-G2mAb 9C4 being the most effective [146]The9C4 antigenic area maps within G2 epitope I and a broadlyneutralizing activity may be investigated
5 Conclusions
Arbovirus infections have acquired increasing interest giventhe augmented globalization and tourism movement all overthe world that could be at the basis of epidemic eventsFurthermore the same vectors can sometimes transmit sev-eral arboviruses concomitantly complicating the diagnosis aswell as the therapy For example confusing mixed epidemicshave occasionally been described such as YFV plus CHIKVDENVplus CHIKV ormore recentlyPlasmodium falciparummalaria plus DENV1 and CHIKV on Madagascarrsquos east coast[1]
In the last decade several efforts have been employed inthe development of effective immunogens and therapeuticsgiving the current absence of specific antiviral drugs as wellas effective vaccines for themost diffused arbovirus infections(ie DENV andWNV)
However at the moment mosquito control is the bestavailable method for preventing arbovirus infections Thusto control the emerging public health problem of arbovirusinfections new antiviral therapeutic strategies that providepotent and broadly cross-protective immunity (especially forflavivirus infections) are an urgent globally medical need
As described in this review the majority of highlyneutralizing mAbs are of murine origin and indeed theirutility in the treatment of these infections is limited In facttheir employment can elicit an immunogenic response (iehuman anti-mouse Abs HAMA) the therapeutic efficacycould be reduced by a relatively faster clearance in humans(compared to human Abs) potentially exacerbated by theHAMA response Finally murine Abs exhibit relatively weakeffector functions (eg ADCC) compared to human AbsIndeed chimerization humanization or better the isolationand characterization of fully human broadly neutralizingmAbs would be the best choice for their possible use in afuture mAb-based therapy against arbovirus infections [147]
Finally possible ADE of infection mechanisms should beevaluated before considering a mAb as a possible candidatetherapeutic in a post-exposure setting as well as in the devel-opment of new Ab-eliciting immunogens
Conflict of Interests
The authors declare no conflict of interests in particular withOmriGam and any other cited Ab-based preparation men-tioned in the paper
References
[1] N Cleton M Koopmans J Reimerink G J Godeke and CReusken ldquoCome fly with me review of clinically importantarboviruses for global travelersrdquo Journal of Clinical Virology vol55 no 3 pp 191ndash203 2012
[2] F X Heinz H Holzmann A Essl and M Kundi ldquoField effec-tiveness of vaccination against tick-borne encephalitisrdquoVaccinevol 25 no 43 pp 7559ndash7567 2007
[3] A D Barrett and D E Teuwen ldquoYellow fever vaccinemdashhowdoes it work and why do rare cases of serious adverse events
BioMed Research International 17
take placerdquo Current Opinion in Immunology vol 21 no 3 pp308ndash313 2009
[4] S B Halstead and S J Thomas ldquoNew vaccines for Japaneseencephalitisrdquo Current Infectious Disease Reports vol 12 no 3pp 174ndash180 2010
[5] R Burioni M Perotti N Mancini and M Clementi ldquoPer-spectives for the utilization of neutralizing human monoclonalantibodies as anti-HCV drugsrdquo Journal of Hepatology vol 49no 2 pp 299ndash300 2008
[6] R A Diotti G A Sautto L Solforosi NManciniM Clementiand R Burioni ldquoNeutralization activity and kinetics of twobroad-range human monoclonal IgG1 derived from recombi-nant Fab fragments and directed against Hepatitis C virus E2glycoproteinrdquo New Microbiologica vol 35 no 4 pp 475ndash4792012
[7] R Burioni F Canducci N Mancini et al ldquoMolecular cloningof the first humanmonoclona antibodies neutralizing with highpotency Swine-origin Influenza A pandemic virus (S-OIV)rdquoNew Microbiologica vol 32 no 4 pp 319ndash324 2009
[8] P K Chandra S Hazari B Poat et al ldquoIntracytoplasmic stableexpression of IgG1 antibody targeting NS3 helicase inhibitsreplication of highly efficient hepatitis C Virus 2a clonerdquoVirology Journal vol 7 article 118 2010
[9] N Clementi D de Marco N Mancini et al ldquoA human mono-clonal antibody with neutralizing activity against highly diver-gent influenza subtypesrdquo PLoS ONE vol 6 no 12 Article IDe28001 2011
[10] D de Marco N Clementi N Mancini et al ldquoA non-VH1-69 heterosubtypic neutralizing human monoclonal antibodyprotects mice against H1N1 and H5N1 virusesrdquo PLoS ONE vol7 no 4 Article ID e34415 2012
[11] L Solforosi N Mancini F Canducci et al ldquoA phage displayvector optimized for the generation of human antibody com-binatorial libraries and the molecular cloning of monoclonalantibody fragmentsrdquoNewMicrobiologica vol 35 no 3 pp 289ndash294 2012
[12] E V Koonin and V V Dolja ldquoEvolution and taxonomy ofpositive-strand RNA viruses implications of comparative anal-ysis of amino acid sequencesrdquo Critical Reviews in Biochemistryand Molecular Biology vol 28 no 5 pp 375ndash430 1993
[13] J S Mackenzie D J Gubler and L R Petersen ldquoEmergingflaviviruses the spread and resurgence of Japanese encephalitisWest Nile and dengue virusesrdquo Nature Medicine vol 10 no 12pp S98ndashS109 2004
[14] T J SmithW E Brandt J L Swanson J MMcCown and E LBuescher ldquoPhysical and biological properties of dengue-2 virusand associated antigensrdquo Journal of Virology vol 5 no 4 pp524ndash532 1970
[15] J M Smit B Moesker I Rodenhuis-Zybert and J WilschutldquoFlavivirus cell entry and membrane fusionrdquo Viruses vol 3 no2 pp 160ndash171 2011
[16] R J Kuhn W Zhang M G Rossmann et al ldquoStructure ofdengue virus implications for flavivirus organization matura-tion and fusionrdquo Cell vol 108 no 5 pp 717ndash725 2002
[17] K Stiasny J H Aberle M Keller B Grubeck-Loebenstein andF X Heinz ldquoAge affects quantity but not quality of antibodyresponses after vaccination with an inactivated flavivirus vac-cine against tick-borne encephalitisrdquo PLoS ONE vol 7 no 3Article ID e34145 2012
[18] LThomasOVerlaetenACabie et al ldquoInfluence of the dengueserotype previous dengue infection and plasma viral load
on clinical presentation and outcome during a dengue-2 anddengue-4 co-epidemicrdquo American Journal of Tropical Medicineand Hygiene vol 78 no 6 pp 990ndash998 2008
[19] Y Zhang J Corver P R Chipman et al ldquoStructures of imma-ture flavivirus particlesrdquo EMBO Journal vol 22 no 11 pp2604ndash2613 2003
[20] A K I Falconar ldquoIdentification of an epitope on the denguevirus membrane (M) protein defined by cross-protective mon-oclonal antibodies design of an improved epitope sequencebased on common determinants present in both envelope (EandM) proteinsrdquo Archives of Virology vol 144 no 12 pp 2313ndash2330 1999
[21] K-J Huang Y-C Yang Y-S Lin et al ldquoThe dual-specific bind-ing of dengue virus and target cells for the antibody-dependentenhancement of dengue virus infectionrdquo Journal of Immunol-ogy vol 176 no 5 pp 2825ndash2832 2006
[22] A K I Falconar ldquoAntibody responses are generated to immun-odominant ELKKLE-typemotifs on the nonstructural-1 glyco-protein during live dengue virus infections inmice and humansimplications for diagnosis pathogenesis and vaccine designrdquoClinical and Vaccine Immunology vol 14 no 5 pp 493ndash5042007
[23] C W Mandl F Guirakhoo H Holzmann F X Heinz and CKunz ldquoAntigenic structure of the flavivirus envelope protein Eat the molecular level using the tick-borne encephalitis virus asa modelrdquo Journal of Virology vol 63 no 2 pp 564ndash571 1989
[24] F A Rey F X Heinz C Mandl C Kunz and S C HarrisonldquoThe envelope glycoprotein from tick-borne encephalitis virusat 2 A resolutionrdquo Nature vol 375 no 6529 pp 291ndash298 1995
[25] S L Allison K Stiasny K Stadler CWMandl and F XHeinzldquoMapping of functional elements in the stem-anchor regionof tick-borne encephalitis virus envelope protein Erdquo Journal ofVirology vol 73 no 7 pp 5605ndash5612 1999
[26] D E Klein J L Choi and S C Harrison ldquoStructure of a denguevirus envelope protein late-stage fusion intermediaterdquo Journal ofVirology vol 87 no 4 pp 2287ndash2293 2013
[27] S L Hanna T C Pierson M D Sanchez A A Ahmed M MMurtadha and R W Doms ldquoN-linked glycosylation of WestNile virus envelope proteins influences particle assembly andinfectivityrdquo Journal of Virology vol 79 no 21 pp 13262ndash132742005
[28] N Mancini G Sautto N Clementi et al ldquoNeutralization inter-fering antibodies a ldquonovelrdquo example of humoral immune dys-function facilitating viral escaperdquoViruses vol 4 no 9 pp 1731ndash1752 2012
[29] Y Modis S Ogata D Clements and S C Harrison ldquoVariablesurface epitopes in the crystal structure of dengue virus type3 envelope glycoproteinrdquo Journal of Virology vol 79 no 2 pp1223ndash1231 2005
[30] N Miller ldquoRecent progress in dengue vaccine research anddevelopmentrdquo Current Opinion in Molecular Therapeutics vol12 no 1 pp 31ndash38 2010
[31] J H Chavez J R Silva A A Amarilla and L T Moraes Figu-eiredo ldquoDomain III peptides from flavivirus envelope proteinare useful antigens for serologic diagnosis and targets forimmunizationrdquo Biologicals vol 38 no 6 pp 613ndash618 2010
[32] B Shrestha J D Brien S Sukupolvi-Petty et al ldquoThe develop-ment of therapeutic antibodies that neutralize homologous andheterologous genotypes of dengue virus type 1rdquo PLoS Pathogensvol 6 no 4 Article ID e1000823 2010
18 BioMed Research International
[33] G D Gromowski N D Barrett and A D T Barrett ldquoChar-acterization of dengue virus complex-specific neutralizing epi-topes on envelope protein domain III of dengue 2 virusrdquo Journalof Virology vol 82 no 17 pp 8828ndash8837 2008
[34] S Sukupolvi-Petty S K Austin W E Purtha et al ldquoType- andsubcomplex-specific neutralizing antibodies against domain IIIof dengue virus type 2 envelope protein recognize adjacentepitopesrdquo Journal of Virology vol 81 no 23 pp 12816ndash128262007
[35] S Sukupolvi-Petty S K Austin M Engle et al ldquoStructure andfunction analysis of therapeutic monoclonal antibodies againstdengue virus type 2rdquo Journal of Virology vol 84 no 18 pp9227ndash9239 2010
[36] Y-Q Deng J-X Dai G-H Ji et al ldquoA broadly flaviviruscross-neutralizingmonoclonal antibody that recognizes a novelepitope within the fusion loop of e proteinrdquo PLoS ONE vol 6no 1 Article ID e16059 2011
[37] J M Costin E Zaitseva K M Kahle et al ldquoMechanistic studyof broadly neutralizing human monoclonal antibodies againstdengue virus that target the fusion looprdquo Journal of Virologyvol 87 no 1 pp 52ndash66 2013
[38] CMMidgley A FlanaganH B Tran et al ldquoStructural analysisof a dengue cross-reactive antibody complexed with envelopedomain III reveals the molecular basis of cross-reactivityrdquoJournal of Immunology vol 188 no 10 pp 4971ndash4979 2012
[39] N J Moreland P Susanto E Lim et al ldquoPhage display appro-aches for the isolation of monoclonal antibodies against denguevirus envelope domain III from human and mouse derivedlibrariesrdquo International Journal ofMolecular Sciences vol 13 no3 pp 2618ndash2635 2012
[40] J J B Cockburn M E Navarro Sanchez N Fretes et alldquoMechanism of dengue virus broad cross-neutralization by amonoclonal antibodyrdquo Structure vol 20 no 2 pp 303ndash3142012
[41] R Rajamanonmani C Nkenfou P Clancy et al ldquoOn a mousemonoclonal antibody that neutralizes all four dengue virusserotypesrdquo Journal of General Virology vol 90 no 4 pp 799ndash809 2009
[42] L H Gould J Sui H Foellmer et al ldquoProtective and herapeuticcapacity of human single-chain Fv-Fc fusion proteins againstWest Nile virusrdquo Journal of Virology vol 79 no 23 pp 14606ndash14613 2005
[43] H Sultana H G Foellmer G Neelakanta et al ldquoFusion LoopPeptide of the West Nile virus envelope protein is essential forpathogenesis and is recognized by a therapeutic cross-reactivehuman monoclonal antibodyrdquo Journal of Immunology vol 183no 1 pp 650ndash660 2009
[44] T Oliphant M Engle G E Nybakken et al ldquoDevelopment ofa humanized monoclonal antibody with therapeutic potentialagainstWest Nile virusrdquoNature Medicine vol 11 no 5 pp 522ndash530 2005
[45] G E Nybakken T Oliphant S Johnson S Burke M S Dia-mond and D H Fremont ldquoStructural basis of West Nile virusneutralization by a therapeutic antibodyrdquo Nature vol 437 no7059 pp 764ndash769 2005
[46] G D Gromowski J T Roehrig M S Diamond J C LeeT J Pitcher and A D T Barrett ldquoMutations of an antibodybinding energy hot spot on domain III of the dengue 2 envelopeglycoprotein exploited for neutralization escaperdquo Virology vol407 no 2 pp 237ndash246 2010
[47] J T Roehrig R A Bolin andR G Kelly ldquoMonoclonal antibodymapping of the envelope glycoprotein of the dengue 2 virusJamaicardquo Virology vol 246 no 2 pp 317ndash328 1998
[48] R de Alwis S A Smith N P Olivarez et al ldquoIdentification ofhumanneutralizing antibodies that bind to complex epitopes ondengue virionsrdquo Proceedings of the National Academy of Sciencesof the United States of America vol 109 no 19 pp 7439ndash74442012
[49] S K Austin K A Dowd B Shrestha et al ldquoStructural basis ofdifferential neutralization of DENV-1 genotypes by an antibodythat recognizes a cryptic epitoperdquo PLoS Pathogens vol 8 no 10Article ID e1002930 2012
[50] M Throsby C Geuijen J Goudsmit et al ldquoIsolation and char-acterization of human monoclonal antibodies from individualsinfected with West Nile Virusrdquo Journal of Virology vol 80 no14 pp 6982ndash6992 2006
[51] A P Goncalvez R H Purcell and C-J Lai ldquoEpitope deter-minants of a chimpanzee Fab antibody that efficiently cross-neutralizes dengue type 1 and type 2 viruses map to inside andin close proximity to fusion loop of the dengue type 2 virusenvelope glycoproteinrdquo Journal of Virology vol 78 no 23 pp12919ndash12928 2004
[52] J T Roehrig J H Mathews and D W Trent ldquoIdentification ofepitopes on the E glycoprotein of Saint Louis encephalitis virususing monoclonal antibodiesrdquo Virology vol 128 no 1 pp 118ndash126 1983
[53] J T Roehrig L A Staudinger A R Hunt J H Mathews andC D Blair ldquoAntibody prophylaxis and therapy for flavivirusencephalitis infectionsrdquo Annals of the New York Academy ofSciences vol 951 pp 286ndash297 2001
[54] A P Goncalvez C-H Chien K Tubthong et al ldquoHumanizedmonoclonal antibodies derived from chimpanzee fabs protectagainst Japanese encephalitis virus in vitro and in vivordquo Journalof Virology vol 82 no 14 pp 7009ndash7021 2008
[55] M Arakawa T Yamashiro G-I Uechi M Tadano and ANishizono ldquoConstruction of humanFab (1205741120581) library and iden-tification of human monoclonal Fab possessing neutralizingpotency against Japanese encephalitis virusrdquo Microbiology andImmunology vol 51 no 6 pp 617ndash625 2007
[56] L Warter C Y Lee R Thiagarajan et al ldquoChikungunya virusenvelope-specific human monoclonal antibodies with broadneutralization potencyrdquo Journal of Immunology vol 186 no 5pp 3258ndash3264 2011
[57] P Pal K A Dowd J D Brien et al ldquoDevelopment of a highlyprotective combination monoclonal antibody therapy againstchikungunya virusrdquo PLoS Pathogens vol 9 no 4 Article IDe1003312 2013
[58] A Bertolotti-Ciarlet J Smith K Strecker et al ldquoCellular local-ization and antigenic characterization of Crimean-Congo hem-orrhagic fever virus glycoproteinsrdquo Journal of Virology vol 79no 10 pp 6152ndash6161 2005
[59] J K Battles and J M Dalrymple ldquoGenetic variation amonggeographic isolates of Rift Valley fever virusrdquo American Journalof Tropical Medicine and Hygiene vol 39 no 6 pp 617ndash6311988
[60] B AThibodeaux N C Garbino NM Liss et al ldquoA humanizedIgG but not IgM antibody is effective in prophylaxis and therapyof yellow fever infection in an AG12917D-204 peripheralchallenge mouse modelrdquo Antiviral Research vol 94 no 1 pp1ndash8 2012
BioMed Research International 19
[61] S Daffis R E Kontermann J Korimbocus H Zeller H-DKlenk and J Ter Meulen ldquoAntibody responses against wild-type yellow fever virus and the 17D vaccine strain charac-terization with human monoclonal antibody fragments andneutralization escape variantsrdquoVirology vol 337 no 2 pp 262ndash272 2005
[62] L N Levanov L E Matveev E P Goncharova et al ldquoChimericantibodies against tick-borne encephalitis virusrdquo Vaccine vol28 no 32 pp 5265ndash5271 2010
[63] A R Hunt S Frederickson C Hinkel K S Bowdish and J TRoehrig ldquoA humanized murine monoclonal antibody protectsmice either before or after challenge with virulent Venezuelanequine encephalomyelitis virusrdquo Journal of General Virologyvol 87 no 9 pp 2467ndash2476 2006
[64] J M da Silva Voorham I A Rodenhuis-Zybert N V A Nunezet al ldquoAntibodies against the envelope glycoprotein promoteinfectivity of immature dengue virus serotype 2rdquo PLoS ONEvol 7 no 3 Article ID e29957 2012
[65] T C Pierson Q Xu S Nelson et al ldquoThe stoichiometry ofantibody-mediated neutralization and enhancement of westnile virus infectionrdquoCell Host andMicrobe vol 1 no 2 pp 135ndash145 2007
[66] G Sautto N Mancini R A Diotti L Solforosi M Clementiand R Burioni ldquoAnti-hepatitis C virus E2 (HCVE2) glyco-proteinmonoclonal antibodies andneutralization interferencerdquoAntiviral Research vol 96 no 1 pp 82ndash89 2012
[67] R Burioni Y Matsuura N Mancini et al ldquoDiverging effectsof human recombinant anti-hepatitis C virus (HCV) antibodyfragments derived from a single patient on the infectivityof a vesicular stomatitis virusHCV pseudotyperdquo Journal ofVirology vol 76 no 22 pp 11775ndash11779 2002
[68] R J Phillpotts J R Stephenson and J S Porterfield ldquoAntibody-dependent enhancement of tick-borne encephalitus virus infec-tivityrdquo Journal of General Virology vol 66 no 8 pp 1831ndash18371985
[69] K A Dowd and T C Pierson ldquoAntibody-mediated neutraliza-tion of flaviviruses a reductionist viewrdquo Virology vol 411 no 2pp 306ndash315 2011
[70] A K I Falconar ldquoMonoclonal antibodies that bind to commonepitopes on the dengue virus type 2 nonstructural-1 andenvelope glycoproteins display weak neutralizing activity anddifferentiated responses to virulent strains implications forpathogenesis and vaccinesrdquo Clinical and Vaccine Immunologyvol 15 no 3 pp 549ndash561 2008
[71] K Meyer A Banerjee S E Frey R B Belshe and R Ray ldquoAweak neutralizing antibody response to hepatitis C virus enve-lope glycoprotein enhances virus infectionrdquo PLoS ONE vol 6no 8 Article ID e23699 2011
[72] R Burioni R AWilliamson P P Sanna F E Bloom and D RBurton ldquoRecombinant human Fab to glycoprotein D neutral-izes infectivity and prevents cell-to-cell transmission of herpessimplex viruses 1 and 2 in vitrordquo Proceedings of the NationalAcademy of Sciences of the United States of America vol 91 no1 pp 355ndash359 1994
[73] M S Rolph A Zaid N E Rulli and S Mahalingam ldquoDown-regulation of interferon-120573 in antibody-dependent enhancementof dengue viral infections of human macrophages is dependenton interleukin-6rdquo Journal of Infectious Diseases vol 204 no 3pp 489ndash491 2011
[74] S S Whitehead J E Blaney A P Durbin and B R MurphyldquoProspects for a dengue virus vaccinerdquoNature ReviewsMicrobi-ology vol 5 no 7 pp 518ndash528 2007
[75] S-M Lok V Kostyuchenko G ENybakken et al ldquoBinding of aneutralizing antibody to dengue virus alters the arrangement ofsurface glycoproteinsrdquoNature Structural andMolecular Biologyvol 15 no 3 pp 312ndash317 2008
[76] A P Goncalvez R Men C Wernly R H Purcell and C-J Lai ldquoChimpanzee Fab fragments and a derived humanizedimmunogiobulin G1 antibody that efficiently cross-neutralizedengue type 1 and type 2 virusesrdquo Journal of Virology vol 78no 23 pp 12910ndash12918 2004
[77] WM P BWahala A A Kraus L B HaymoreM A Accavitti-Loper and A M de Silva ldquoDengue virus neutralization byhuman immune sera role of envelope protein domain III-reactive antibodyrdquo Virology vol 392 no 1 pp 103ndash113 2009
[78] W D Crill H R Hughes M J Delorey and G-J J ChangldquoHumoral immune responses of dengue fever patients usingepitope-specific serotype-2 virus-like particle antigensrdquo PLoSONE vol 4 no 4 Article ID e4991 2009
[79] C Setthapramote T Sasaki O Puiprom et al ldquoHuman mono-clonal antibodies to neutralize all dengue virus serotypes usinglymphocytes from patients at acute phase of the secondaryinfectionrdquo Biochemical and Biophysical Research Communica-tions vol 423 no 4 pp 867ndash872 2012
[80] M Beltramello K L Williams C P Simmons et al ldquoThehuman immune response to dengue virus is dominated byhighly cross-reactive antibodies endowed with neutralizing andenhancing activityrdquo Cell Host andMicrobe vol 8 no 3 pp 271ndash283 2010
[81] C M Midgley M Bajwa-Joseph S Vasanawathana et al ldquoAnin-depth analysis of original antigenic sin in dengue virusinfectionrdquo Journal of Virology vol 85 no 22 pp 410ndash421 2011
[82] B Kaufmann M R Vogt J Goudsmit et al ldquoNeutralizationof West Nile virus by cross-linking of its surface proteins withFab fragments of the human monoclonal antibody CR4354rdquoProceedings of the National Academy of Sciences of the UnitedStates of America vol 107 no 44 pp 18950ndash18955 2010
[83] B Spurrier J M Sampson M Totrov et al ldquoStructural analysisof human and macaque mAbs 2909 and 25B implicationsfor the configuration of the quaternary neutralizing epitope ofHIV-1 gp120rdquo Structure vol 19 no 5 pp 691ndash699 2011
[84] J J SchlesingerMWBrandriss andT PMonath ldquoMonoclonalantibodies distinguish between wild and vaccine strains ofyellow fever virus by neutralization hemagglutination inhibi-tion and immune precipitation of the virus envelope proteinrdquoVirology vol 125 no 1 pp 8ndash17 1983
[85] M W Brandriss J J Schlesinger E E Walsh and M BrisellildquoLethal 17D yellow fever encephalitis in mice I Passive protec-tion by monoclonal antibodies to the envelope proteins of 17Dyellow fever and dengue 2 virusesrdquo Journal of General Virologyvol 67 no 2 pp 229ndash234 1986
[86] H Cho and M S Diamond ldquoImmune responses to West Nilevirus infection in the central nervous systemrdquo Viruses vol 4no 12 pp 3812ndash3830 2012
[87] D W Beasley A D Barrett and R B Tesh ldquoResurgence ofWest Nile neurologic disease in the United States in 2012 whathappened What needs to be donerdquo Antiviral Research vol 99no 1 pp 1ndash5 2013
[88] M Throsby J Ter Meulen C Geuijen J Goudsmit and J DeKruif ldquoMapping and analysis of West Nile virus-specific mon-oclonal antibodies prospects for vaccine developmentrdquo ExpertReview of Vaccines vol 6 no 2 pp 183ndash191 2007
[89] M S Diamond T G Roberts D Edgil B Lu J Ernst and EHarris ldquoModulation of dengue virus infection in human cells
20 BioMed Research International
by alpha beta and gamma interferonsrdquo Journal of Virology vol74 no 11 pp 4957ndash4966 2000
[90] Z Shimoni M J Niven S Pitlick and S Bulvik ldquoTreatmentof West Nile virus encephalitis with intravenous immunoglob-ulinrdquo Emerging Infectious Diseases vol 7 no 4 article 759 2001
[91] AHamdan PGreen EMendelsonM R Kramer S Pitlik andM Weinberger ldquoPossible benefit of intravenous immunoglob-ulin therapy in a lung transplant recipient with West Nile virusencephalitisrdquoTransplant InfectiousDisease vol 4 no 3 pp 160ndash162 2002
[92] M Haley A S Retter D Fowler J Gea-Banacloche and NP OrsquoGrady ldquoThe role for intravenous immunoglobulin in thetreatment of West Nile virus encephalitisrdquo Clinical InfectiousDiseases vol 37 no 6 pp e88ndashe90 2003
[93] K M Chung G E Nybakken B S Thompson et al ldquoAnti-bodies against West Nile virus nonstructural protein NS1prevent lethal infection through Fc 120574 receptor-dependent and-independent mechanismsrdquo Journal of Virology vol 80 no 3pp 1340ndash1351 2006
[94] E A Henchal L S Henchal and J J Schlesinger ldquoSynergisticinteractions of anti-NS1 monoclonal antibodies protect pas-sively immunized mice from lethal challenge with dengue 2virusrdquo Journal of General Virology vol 69 no 8 pp 2101ndash21071988
[95] J T Roehrig ldquoAntigenic structure of flavivirus proteinsrdquoAdvances in Virus Research vol 59 pp 141ndash175 2003
[96] M D Sanchez T C Pierson D McAllister et al ldquoCharacteri-zation of neutralizing antibodies to West Nile virusrdquo Virologyvol 336 no 1 pp 70ndash82 2005
[97] J H Beigel J L Nordstrom S R Pillemer et al ldquoSafety andpharmacokinetics of single intravenous dose of MGAWN1 anovel monoclonal antibody to West Nile virusrdquo AntimicrobialAgents and Chemotherapy vol 54 no 6 pp 2431ndash2436 2010
[98] C A Smeraski V Siddharthan and J D Morrey ldquoTreatmentof spatial memory impairment in hamsters infected with WestNile virus using a humanizedmonoclonal antibodyMGAWN1rdquoAntiviral Research vol 91 no 1 pp 43ndash49 2011
[99] D Lelli AMoreno E Brocchi et al ldquoWestNile virus character-ization and diagnostic applications of monoclonal antibodiesrdquoVirology Journal vol 9 article 81 2012
[100] T S Gritsun V A Lashkevich and E A Gould ldquoTick-borneencephalitisrdquo Antiviral Research vol 57 no 1-2 pp 129ndash1462003
[101] D Ruzek T S Gritsun N L Forrester et al ldquoMutations inthe NS2B and NS3 genes affect mouse neuroinvasiveness of aWestern European field strain of tick-borne encephalitis virusrdquoVirology vol 374 no 2 pp 249ndash255 2008
[102] M Ecker S L Allison T Meixner and F X Heinz ldquoSequenceanalysis and genetic classification of tick-borne encephalitisviruses from Europe and Asiardquo Journal of General Virology vol80 no 1 pp 179ndash185 1999
[103] H Holzmann M S Vorobyova I P Ladyzhenskaya et alldquoMolecular epidemiology of tick-borne encephalitis viruscross-protection between European and Far Eastern subtypesrdquoVaccine vol 10 no 5 pp 345ndash349 1992
[104] C W Mandl ldquoSteps of the tick-borne encephalitis virus repli-cation cycle that affect neuropathogenesisrdquo Virus Research vol111 no 2 pp 161ndash174 2005
[105] A V Atrasheuskaya T M Fredeking and G M IgnatyevldquoChanges in immune parameters and their correction in humancases of tick-borne encephalitisrdquo Clinical and ExperimentalImmunology vol 131 no 1 pp 148ndash154 2003
[106] K L Mansfield N Johnson L P Phipps J R Stephenson AR Fooks and T Solomon ldquoTick-borne encephalitis virusmdashareview of an emerging zoonosisrdquo Journal of General Virologyvol 90 no 8 pp 1781ndash1794 2009
[107] DWVaughn andCHHoke Jr ldquoThe epidemiology of Japaneseencephalitis prospects for preventionrdquo Epidemiologic Reviewsvol 14 pp 197ndash221 1992
[108] T Solomon H Ni D W C Beasley M Ekkelenkamp M JCardosa and A D T Barrett ldquoOrigin and evolution of Japaneseencephalitis virus in Southeast Asiardquo Journal of Virology vol 77no 5 pp 3091ndash3098 2003
[109] M R Wilson ldquoEmerging viral infectionsrdquo Current Opinion inNeurology vol 26 no 3 pp 301ndash306 2013
[110] J N Hanna S A Ritchie D A Phillips et al ldquoAn outbreakof Japanese encephalitis in the Torres Strait Australia 1995rdquoMedical Journal of Australia vol 165 no 5 pp 256ndash260 1996
[111] J N Hanna S A Ritchie D A Phillips et al ldquoJapanese ence-phalitis in north Queensland Australia 1998rdquo Medical Journalof Australia vol 170 no 11 pp 533ndash536 1999
[112] SGeorgeMGourie-Devi and J A Rao ldquoIsolation ofWestNilevirus from the brains of children who had died of encephalitisrdquoBulletin of theWorldHealthOrganization vol 62 no 6 pp 879ndash882 1984
[113] A K Gupta A A Koshy and V J Lad ldquoEnhanced protec-tion of mice against Japanese encephalitis virus infection bycombinations ofmonoclonal antibodies to glycoprotein ErdquoActaVirologica vol 55 no 2 pp 165ndash168 2011
[114] T H Lee B-H Song S-I Yun et al ldquoA cross-protective mAbrecognizes a novel epitope within the flavivirus NS1 proteinrdquoJournal of General Virology vol 93 no 1 pp 20ndash26 2012
[115] S C Weaver ldquoHost range amplification and arboviral diseaseemergencerdquo Archives of Virology Supplementum no 19 pp 33ndash44 2005
[116] M Solignat B Gay S Higgs L Briant and C Devaux ldquoRepli-cation cycle of chikungunya a re-emerging arbovirusrdquoVirologyvol 393 no 2 pp 183ndash197 2009
[117] O Schwartz and M L Albert ldquoBiology and pathogenesis ofchikungunya virusrdquo Nature Reviews Microbiology vol 8 no 7pp 491ndash500 2010
[118] W Akahata Z-Y Yang H Andersen et al ldquoA virus-like particlevaccine for epidemic Chikungunya virus protects nonhumanprimates against infectionrdquo Nature Medicine vol 16 no 3 pp334ndash338 2010
[119] M Kielian and A Helenius ldquopH-Iinduced alterations in thefusogenic spike protein of Semliki Forest virusrdquo Journal of CellBiology vol 101 no 6 pp 2284ndash2291 1985
[120] J White and A Helenius ldquopH-Dependent fusion between theSemliki Forest virus membrane and liposomesrdquo Proceedings ofthe National Academy of Sciences of the United States of Americavol 77 no 6 pp 3273ndash3277 1980
[121] J Dubuisson and C M Rice ldquoSindbis virus attachment isola-tion and characterization of mutants with impaired binding tovertebrate cellsrdquo Journal of Virology vol 67 no 6 pp 3363ndash33741993
[122] C Y Lee Y-W Kam J Fric et al ldquoChikungunya virus neutral-ization antigens and direct cell-to-cell transmission are revealedby human antibody-escapemutantsrdquo PLoS Pathogens vol 7 no12 Article ID e1002390 2011
[123] J E Voss M-C Vaney S Duquerroy et al ldquoGlycoproteinorganization of Chikungunya virus particles revealed by X-raycrystallographyrdquo Nature vol 468 no 7324 pp 709ndash712 2010
BioMed Research International 21
[124] L Li J Jose Y Xiang R J Kuhn and M G Rossmann ldquoStruc-tural changes of envelope proteins during alphavirus fusionrdquoNature vol 468 no 7324 pp 705ndash708 2010
[125] J Fric S Bertin-Maghit C I Wang A Nardin and L WarterldquoUse of human monoclonal antibodies to treat Chikungunyavirus infectionrdquo Journal of Infectious Diseases vol 207 no 2 pp319ndash322 2013
[126] P V Aguilar J G Estrada-Franco R Navarro-Lopez C FerroA D Haddow and S C Weaver ldquoEndemic Venezuelanequine encephalitis in the Americas hidden under the dengueumbrellardquo Future Virology vol 6 no 6 pp 721ndash740 2011
[127] S C Weaver R Winegar I D Manger and N L ForresterldquoAlphaviruses population genetics and determinants of emer-gencerdquo Antiviral Research vol 94 no 3 pp 242ndash257 2012
[128] S Paessler and S C Weaver ldquoVaccines for Venezuelan equineencephalitisrdquo Vaccine vol 27 no 4 pp D80ndashD85 2009
[129] R J Phillpotts ldquoVenezuelan equine encephalitis virus complex-specific monoclonal antibody provides broad protection inmurine models against airborne challenge with viruses fromserogroups I II and IIIrdquoVirus Research vol 120 no 1-2 pp 107ndash112 2006
[130] R J Phillpotts L D Jones and S C Howard ldquoMonoclonalantibody protectsmice against infection anddiseasewhen giveneither before or up to 24 h after airborne challenge with virulentVenezuelan equine encephalitis virusrdquoVaccine vol 20 no 11-12pp 1497ndash1504 2002
[131] J H Mathews J T Roehrig and D W Trent ldquoRole of com-plement and the Fc portion of immunoglobulin G in immunityto Venezuelan equine encephalomyelitis virus infection withglycoprotein-specific monoclonal antibodiesrdquo Journal of Virol-ogy vol 55 no 3 pp 594ndash600 1985
[132] L M OrsquoBrien C D Underwood-Fowler S A Goodchild AL Phelps and R J Phillpotts ldquoDevelopment of a novel mon-oclonal antibody with reactivity to a wide range of Venezuelanequine encephalitis virus strainsrdquoVirology Journal vol 6 article206 2009
[133] L M OrsquoBrien S A Goodchild R J Phillpotts and S D Per-kins ldquoA humanised murine monoclonal antibody protectsmice from Venezuelan equine encephalitis virus Evergladesvirus and Mucambo virus when administered up to 48h afterairborne challengerdquo Virology vol 426 no 2 pp 100ndash105 2012
[134] N Clementi N Mancini L Solforosi M Castelli M Clementiand R Burioni ldquoPhage display-based strategies for cloning andoptimization of monoclonal antibodies directed against humanpathogensrdquo International Journal of Molecular Sciences vol 13no 7 pp 8273ndash8292 2012
[135] A R Hunt R A Bowen S Frederickson T Maruyama JT Roehrig and C D Blair ldquoTreatment of mice with humanmonoclonal antibody 24h after lethal aerosol challenge withvirulent Venezuelan equine encephalitis virus prevents diseasebut not infectionrdquo Virology vol 414 no 2 pp 146ndash152 2011
[136] W-G Hu A L Phelps S Jager et al ldquoA recombinant human-ized monoclonal antibody completely protects mice againstlethal challenge with Venezuelan equine encephalitis virusrdquoVaccine vol 28 no 34 pp 5558ndash5564 2010
[137] T Rulker L Voss P Thullier et al ldquoIsolation and characterisa-tion of a human-like antibody fragment (scFv) that inactivatesVEEV in vitro and in vivordquo PLoS ONE vol 7 no 5 Article IDe37242 2012
[138] C T Walter and J N Barr ldquoRecent advances in the molecularand cellular biology of bunyavirusesrdquo Journal of General Virol-ogy vol 92 no 11 pp 2467ndash2484 2011
[139] C A Whitehouse ldquoCrimean-Congo hemorrhagic feverrdquo Anti-viral Research vol 64 no 3 pp 145ndash160 2004
[140] M Mertens K Schmidt A Ozkul and M H Groschup ldquoTheimpact of Crimean-Congo hemorrhagic fever virus on publichealthrdquo Antiviral Research vol 98 no 2 pp 248ndash260 2013
[141] A A Ahmed J M McFalls C Hoffmann et al ldquoPresence ofbroadly reactive and group-specific neutralizing epitopes onnewly described isolates of Crimean-Congo hemorrhagic fevervirusrdquo Journal of General Virology vol 86 no 12 pp 3327ndash33362005
[142] R Hewson A Gmyl L Gmyl et al ldquoEvidence of segment reas-sortment in Crimean-Congo haemorrhagic fever virusrdquo Journalof General Virology vol 85 no 10 pp 3059ndash3070 2004
[143] N K Blackburn T G Besselaar A J Shepherd and R Swane-poel ldquoPreparation and use of monoclonal antibodies for iden-tifying Crimean-Congo hemorrhagic fever virusrdquo AmericanJournal of Tropical Medicine and Hygiene vol 37 no 2 pp 392ndash397 1987
[144] T Ikegami ldquoMolecular biology and genetic diversity of RiftValley fever virusrdquoAntiviral Research vol 95 no 3 pp 293ndash3102012
[145] KKeegan andM S Collett ldquoUse of bacterial expression cloningto define the amino acid sequences of antigenic determinantson the G2 glycoprotein of Rift Valley fever virusrdquo Journal ofVirology vol 58 no 2 pp 263ndash270 1986
[146] T G Besselaar and N K Blackburn ldquoTopological mapping ofantigenic sites on the Rift Valley fever virus envelope glycopro-teins using monoclonal antibodiesrdquo Archives of Virology vol121 no 1ndash4 pp 111ndash124 1991
[147] L G Presta ldquoEngineering of therapeutic antibodies to mini-mize immunogenicity and optimize functionrdquo Advanced DrugDelivery Reviews vol 58 no 5-6 pp 640ndash656 2006
Submit your manuscripts athttpwwwhindawicom
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anatomy Research International
PeptidesInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporation httpwwwhindawicom
International Journal of
Volume 2014
Zoology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Molecular Biology International
GenomicsInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioinformaticsAdvances in
Marine BiologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Signal TransductionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Evolutionary BiologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Biochemistry Research International
ArchaeaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Genetics Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in
Virolog y
Hindawi Publishing Corporationhttpwwwhindawicom
Nucleic AcidsJournal of
Volume 2014
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Enzyme Research
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Microbiology

6 BioMed Research InternationalTa
ble1Schematicsummaryof
theb
estcharacterized
mAb
sagainstarbo
viralp
atho
gens
mAb
Orig
inRe
activ
ityTargetprotein
Epito
peClon
ingstr
ategy
Form
atisotype
Neutralizingactiv
ityRe
ference
48A
Hum
anDEN
V1ndash4
EDII
EBVtransfo
rmation
IgG
IC50
(120583gmL)
DEN
V115
DEN
V2gt40
DEN
V324
DEN
V4gt40
[37]
D11C
IC50
(120583gmL)
DEN
V115
DEN
V210
DEN
V3102
DEN
V416
16D
IC50
(120583gmL)
DEN
V115
DEN
V202
DEN
V305
DEN
V427
2H12
Murine
DEN
V1ndash4
EDIII(ABloop
aa
314ndash
317)
Hybrid
omafrom
BALB
cmou
seim
mun
ized
with
DEN
V2EDIII
IgG2b
IC50
nM
DEN
V1056ndash54
DEN
V329
DEN
V4145
[38]
C9Murinechim
eric
DEN
V2
EDIII
Phaged
isplayof
achim
ericmurine
hybridom
alibrary
VH1V1205811
PRNT5
0850120583
gmL
[39]
4E11
Murine
DEN
V1ndash4
EDIII(str
andA308
312andstrand
G387
389391)
Hybrid
oma
IgG2a120581
IC50
(120583gmL)
DEN
V1016
DEN
V2013
DEN
V-38
DEN
V-415
[40]
4G2
Murine
DEN
V1ndash4
EDII(fu
sionloop
)Hybrid
omafrom
DEN
V2Eim
mun
ized
mice
IgG
PRNT5
0(120583gmL)
DEN
V215
[39]
9F12
Murine
DEN
V1ndash4WNV
EDIII(aa
305307310
330Astr
andandBC
loop
)
Hybrid
omafrom
BALB
cmou
seim
mun
ized
with
DEN
V-2EDIII
IgG1k
DEN
V1ndash4P
RNT5
02sdot10minus8ndash2sdot10minus7M
[41]
2A10G6
Murine
DEN
V1ndash4YF
VWNV
JEV
TBEV
EDII(Fusionloop
aa
98ndash101)
Hybrid
omafrom
BALB
cmou
seim
mun
ized
with
inactiv
ated
DEN
V2
IgG1
PRNT5
0(120583gmL)
DEN
V12
DEN
V215
DEN
V321
DEN
V418
YNF36
WNV
46
[36]
mAb
11Hum
anDEN
V1ndash4WNV
SLEV
YFV
JEV
MVEV
EDII(fu
sionloop
)Ph
aged
isplayof
human
scFv
Fusio
nprotein
scFv-Fc
PRNT8
0(120583gmL)
WNV
125
DEN
V2625
[4243]
BioMed Research International 7Ta
ble1Con
tinued
mAb
Orig
inRe
activ
ityTargetprotein
Epito
peClon
ingstr
ategy
Form
atisotype
Neutralizingactiv
ityRe
ference
E16
(MGAW
N1)
Murine
WNV
EDIII(LR
aa3
02ndash309)
Hybrid
omafrom
immun
ized
micew
ithWNVE
IgG2bhum
anized
PRNT5
04ndash
18ng
PRNT9
053ndash297
ng[4445]
1A1D
-2Murine
DEN
V1ndash3
EDIII(Astr
andaa
307
310and312)
Hybrid
omafrom
immun
ized
micew
ithdifferent
pH-tr
eated
virus
IgG2a
DEN
V2PR
NT5
021n
M[4647]
1F4
Hum
an
DEN
V1
E(virion
)
DI-DII
Electro
fusio
nof
infected
mem
oryBcells
from
DEN
V-im
mun
esub
jects
with
EBV
IgG
DEN
V1IC5
0011120583gmL
[48]
2D22
DEN
V2
DIII
DEN
V2IC50
008120583gmL
5J7
DEN
V3
DI-DII
DEN
V3IC50
010120583gmL
E105
Murine
DEN
V1
E
DIII(BC
loop
G328
T329
andD330DE
loop
K361E
and
E362K
FGloop
K3
85)
Hybrid
omas
deriv
edfro
mC5
7BL6
IFN-120572120573Rminus
minusmice
infected
with
DEN
V1
IgG
PRNT5
0DEN
V-1
05ndash592ng
mL
[32]
E106
DIII(BC
loop
G328
T329
andD330DE
loop
K361E
and
E362K
FGloop
K3
85A
strand
S305
K307E
309K3
10and
E311)
PRNT5
0DEN
V-1
06ndash
592ng
mL
E111
DIII(CC1015840loop
)PR
NT5
038ndash25120583gmL
[49]
CR4374
Hum
anWNV
EDIII
Phaged
isplayof
scFv
IgGlib
rary
VH2-05V
L1e
PRNT5
0018120583gmL
PRNT9
0095120583gmL
[50]
CR4353
VH3-30V
k3-A27
PRNT5
00026120583
gmL
PRNT9
0364120583gmL
1A5
Chim
panzee
DEN
V1ndash4WNV
JEV
LGTV
EDII(aaG
106)
Phaged
isplayof
Fab
library
from
DEN
V1ndash4
infected
chim
panzees
Hum
anized
IgG1
PRNT5
0(120583gmL)
DEN
V1048
DEN
V2095
DEN
V332
DEN
V443
WNVD
ENV438
JEV
21LG
TV28
[51]
8 BioMed Research International
Table1Con
tinued
mAb
Orig
inRe
activ
ityTargetprotein
Epito
peClon
ingstr
ategy
Form
atisotype
Neutralizingactiv
ityRe
ference
mAb
11Hum
anDEN
V1ndash4WNV
EDII(fu
sionloop
W101G104G106)
scFv
library
scFv-Fc
PRNT8
0(120583gmL)
DEN
V2625
WNV
125
[4243]
3B4C
-4
Murine
SLEV
E
1a
Hybrid
oma
IgG
SLEV
PRNTlt17
[5253]
1B2C
-5SLEV
1bSLEV
PRNTlt17
6B5A
-2SLEV
1cSLEV
PRNT
48
4A4C
-4SLEV
1dSLEV
PRNT
29
1B5D
-1SLEV
JEV
2SLEV
PRNTlt17
2B5B
-3SL
EVJEV
MVEV
WNV
YFV
3SLEV
PRNT
23
2B6B
-2AllFlaviviru
s4a
SLEV
PRNTlt17
6B6C
-1AllFlaviviru
s4b
(inDII)
SLEV
PRNT
23
A3
Chim
panzee
JEV
E2
DIa
aK1
79Ph
aged
isplay
Hum
anized
PRNT5
0004ndash0
2nM
[54]
B2DIIaaI126
Phaged
isplay
PRNT5
0002ndash2
nM
E3DIIIa
aG132
Phaged
isplay
PRNT5
0014ndash0
93n
M
FabT
JE12B0
2Hum
anJEV
ENA
Phaged
isplay
Fab
FRNT5
0502120583gmL
[55]
5F10
Hum
anCH
IKV
E2Dom
ainB
EBVtransfo
rmation
IgG11205822
IC50lt100n
gmL
[56]
8B10
Hum
anCH
IKV
E1-E2
E2Dom
ainA
EBVtransfo
rmation
IgG1k
IC50lt100n
gmL
CHK-
152
Murine
CHIK
VE2
aa59
Hybrid
oma
IgG2c
IC50
1ndash3n
gmL
[57]
11E7
Murine
CCHFV
Gn
C-ter
Hybrid
oma
IgG
PRNT8
0diluted
12560
[58]
4-39-C
CMurine
RVFV
G2
Dom
ainIV
Hybrid
oma
IgG
PRNT8
0diluted
1204
80ndash181920
[59]
2C9
Murine
YFV
EHybrid
oma
IgG2a
PRNT9
011
0417D
1105
2Asib
i[60]
5A 7A R3(27)
Hum
anYF
VE
DI-DII
Phaged
isplay
ScFv
PRNT9
001ndash03120583
gmL
[61]
13D6
Murine
TBEV
EDIII
Hybrid
oma
IgGchimeric
PRNT5
019120583gmL
FRNT5
045120583
gmL
IC50
19ndash167120583
gmL
[62]
3B4C
-4(H
y4-26C
)Murine
VEE
VE2
aa182ndash209
Hybrid
oma
IgGhum
anized
PRNT7
0(120583gmL)
394ndash125
[63]
BioMed Research International 9
The structure of the 1A1D-2 bound to EDIII indicates that themAb binds to sites that are transiently exposed during viralldquobreathingrdquo at 37∘C and block infection during attachmentof the virion to the cell In particular this mAb neutralizesDENV1 2 and 3 serotypes and residues K307 and K310 arethemost critical residues for binding of 1A1D-2mAb [46 75]
Finally Shrestha et al immunized mice with a genotype2 strain of DENV1 virus and generated 79 new mAbs 16of which strongly inhibited infection by the homologousvirus and localized to DIII of E protein Surprisingly onlytwo mAbs E105 and E106 retained strong binding andneutralizing activity against all five DENV1 genotypes as wellas being protective in immunocompromised infected miceMoreover E105 and E106 exhibited therapeutic activity evenwhen administered as a single dose four days after inoculationwith a heterologous genotype 4 strain of DENV1 in the samemouse model [32]
243 Other Non-HumanmAbs Other non-human and non-murine as well as humanized mAbs directed against the Eglycoprotein have been described In this regard Goncalvezet al isolated a large panel of anti-E Fab fragments from chim-panzees infected with all four DENV serotypes Howeveronly a limited number of them displayed a cross-neutralizingactivity against DENV1 and 2 and to a lesser extent againstDENV3 and 4 In particular the authors calculated thatamong them the 1A5 mAb that was further humanizedneutralized DENV1ndash4 at a PRNT50 titer of 048 095 32and 43 120583gmL respectively Interestingly the humanized 1A5was also tested for binding and neutralization against theWNVDENV4 chimera JEV strain SA14-14-2 and Langatvirus (LGTV) strain TP 21 giving a PRNT50 titer of 38 21and 28120583gmL respectively Moreover the authors calculatedthat when administered in a dose of 2mg per kg of bodyweight 1A5 would give a serum titer of approximately 40and 20 50 reduction in plaque reduction neutralization test(PRNT50) against DENV1 and DENV2 respectively [76]Furthermore epitope mapping of 1A5 mAb localized to G106within the Flavivirus-conserved fusion loop inDII of DENV2E protein [51]
244 Anti-DENV Human mAbs As anticipated attemptsto isolate neutralizing human mAbs have been morechallenging due to the restricted elicited Ab-repertoire whichrecognizes DIII of E glycoprotein in naıve as well as ininfected patients [77 78] However several groups reportedthe isolation of neutralizing mAbs from infected subjects Inthis regard Setthapramote et al isolated a total of 136 humanhybridoma clones producing specific mAbs against DENVobtained using PBMCs from nine blood samples from fouracute-phase patients secondarily infected with DENV2 andfive convalescent-phase patients Interestingly the authorsfound that most of the acute-phase mAb clones were cross-reactive with all four DENV serotypes with most of themrecognizing the E protein and endowing of neutralizingactivity against all DENV serotypes compared to thosederived from convalescent-phase patients In particularfrom the acute-phase PBMCs 818 were anti-E 66 wereanti-prM and 33 were anti-NS1 while 133 anti-E 133
anti-prM and 533 anti-NS1 mAbs clones were obtainedfrom convalescent-phase PBMCs [79] Previous studiesconfirmed the data obtained by Setthapramote et al onconvalescent-phase patients but this group firstly reportedthe efficient preparation of human mAbs with strongneutralizing activity titers against all four DENV serotypesusing PBMCs from acute-phase patients secondarily infectedwith DENV [80 81]
The group of de Alwis through immunoglobulin deple-tion studies reported that a substantial fraction of DENV-reactive Abs in human immune sera including type-specificneutralizing Abs bound to the intact virion but not torecombinant E proteinThe authors confirmed these observa-tions also isolating human neutralizing mAbs and proposedthat humans produce Abs that neutralize DENV infectionby binding a complex quaternary structure epitope that isexpressed only when E proteins are assembled on a virusparticle [48] Similar findings have been described for severalother viruses including WNV and HIV [82 83] Mappingstudies indicate that this epitope has a footprint that spansadjacent E protein dimers and includes residues at the hingebetween domains I and II of E protein [48]
Recently Costin et al isolated three broadly neutralizinganti-DENVhumanmAbs named 48AD11C and 16DThesemAbswere isolated from three different convalescent patientswith distinct histories of DENV infection All three mAbsrecognized the E glycoprotein with high affinity neutralizedall four serotypes of DENV but mediated ADE of infection inFc receptor-bearing cells at subneutralizing concentrationsMapping studies revealed that all three mAbs bind discontin-uous epitopes within the highly conserved fusion-loop regionof DII (contacting residues W101 L107 andor G109) [37]
25 Yellow Fever Virus (YFV) TheWHO estimates that thereare 200000 cases of yellow fever virus (YFV) infections and30000 related deaths every year especially in Sub-SaharanAfrica In fact different to themajority of human arbovirusesand similarly to DENV and CHIKV YFV has expanded itshost range to include humans as an amplifying host [1]Symptoms occurring from 3 to 6 days after infection withYFV consist mostly in fever chills anorexia lumbosacralpain nausea and vomiting This syndrome lasts about 3days in most cases and is sometimes followed by a one-daylasting period of remission Fever will reoccur together withdark hematemesismelena petechiae and other hemorrhagicsymptoms Convalescence is characterized by deep asthenialasting up to two weeks Rarely prognosis can be fatal Aneffective live-attenuated vaccine (17D) derived from the Asibistrain by serial passages in chicken embryos is available [84]
251 mAbs against YFV In 1983 Schlesinger et al produceda battery of mAbs after immunization of BALBc mice byinjection of 17D YFV vaccine strain [84] Among them thespecificity towards the E and NS1 proteins was describedeven though anti-NS1 Abs were not able to show neutralizingactivity The 13 IgG and 1 IgM anti-E protein mAbs wereclassified in five groups according to their specificity GroupA consisting of the only IgM produced in this experiment
10 BioMed Research International
could neutralize 17D YFV virus only group B 17D and Asibistrains only group C Asibi strain only group D could onlyneutralize Asibi but not 17D strain showing also reactivityagainst DENV2 Zika or Banzi viruses All of the four mAbsbelonging to this group were able to cross-react with Zikaandor Banzi viruses and two (4E11 and 5H3) neutralizedDENV2 virus with 4E11 neutralizing also Banzi virus The3E9 mAb the only component of the group E could notneutralize 17D nor Asibi strains
All of the IgG mAbs resulted able to protect bothprophylactically and therapeutically BALBc or CD-1 micefrom lethal intracerebral challenge with 17D-204 strain [85]The chimeric form of the group B mAb 2C9 IgG (2C9-cIgG) was able to provide AG129 mice a 72 survival whenadministered 24 hours before infection with 17D-204 strainIts murine form could provide the survival of the 95 ofthe mice Appreciable results were also obtained when bothmurine 2C9 and 2C9-cIgG were administered 24 hours post-infection with the survival of 70 and 20 of mice respec-tively Viral presence was not detected in survivingmice [60]
In 2005 Daffis and colleagues constructed two Ab-phage libraries by cloning the repertoire of YFV-infectedpatients Panning was then performed with YFV-17D virionsThe scFv-5A 7A and R3(27) showed a neutralizing activityspanning from50 to 100 inPRNTassays against bothYFV17D-204-WHO and Asibi strains In further tests reactivitywas observed against wild-type YFV strains of West Africagenotype I and II (Nigeria 1987 and Asibi strains resp)and EastCentral Africa (CAR 1986 Ethiopia 1961 strains) Aconcentration from 01 to 03120583gmL could yield a 90 plaquereduction Reactivity was also observed against the strainSenegal 1990 in a lesser extent Production of escape variantscould demonstrate that scFv-5A 7A and R3(27) epitopes arebuilt up by residues extensively separated in themonomers ofE glycoprotein that however result in closed proximity whenthe homodimeric form of E is constituted [61]
26 West Nile Virus (WNV) The West Nile virus (WNV) isan epidemic neurotropic virus estimated to be responsibleof about 36000 cases and 1500 deaths registered in theUnited States between 1992 and 2012 [86] WNV antigenicityallows its classification into the Japanese encephalitis virus(JEV) serocomplex Genomic analysis has revealed two maingenetic lineages of WNV lineage I viruses circulating in theUSA Europe the Middle East Africa India and Australiaand lineage II viruses isolated from Sub-Saharan Africa andMadagascar [1]
WNV was firstly identified in 1937 and is endemicin many countries of Africa Middle East and West AsiaHowever after 1990 frequent outbreaks of WNV infectionswere reported in Romania Israel and later in North Americaand across the USA in 1999 and recently in 2012 [87]
Most of WNV infected individuals do not develop symp-toms while about 20 develop a self-limiting illness calledWest Nile fever Acute symptoms include fever tirednessand swollen lymph glands but in a minority of cases alsoencephalitis with long-term deficits in cognitive functionand motor skills have been reported However in less than
1 of infected patients WNV having crossed the blood-brain barrier is responsible of neuroinvasive and potentiallylethal form of the disease In these cases degeneration andapoptosis upon infection of neurons and the consequentinflammatory response can occur Related symptoms includehigh fever coma muscle weakness and paralysis Immun-odepression and advanced age have been correlated with anaugment risk to develop a severe disease [87] There are nospecific treatment options or licensed vaccines for humans[88]
WNV is transmitted by infected mosquitos and initialreplication is thought to occur primarily in dendritic cells inthe skin which migrate to secondary lymphoid tissues wherethe replicating virus enters the circulation To date onlyefficacious WNV veterinarian vaccines have been licensedwhile there are no licensed vaccines for protection againstWNV in humans [88]
261 mAbs against WNV Broad-spectrum antivirals suchas type I IFN-120572 ribavirin mycophenolic acid in WNVinfection showed ineffective results in vivo despite in vitrothey showed some activity [89]
However experiments in murine models extrapolationof clinical data as well as passive administration of pooledimmune-120574-globulins (OmriGam) containing a significanttiter of neutralizing Abs before and after infection showedan important role of the humoral immune response incontrolling viremia and prevent viral dissemination Indeedthe development of a neutralizing mAb-based therapy seemsto be encouraging for a possible treatment of infected patients[90ndash92]
Similarly to what has been observed for DENV only anti-E Abs have been identified as neutralizing and protectiveAlso anti-NS1 Abs have shown a protective role howevertheir mechanism of protection has not yet been elucidated asthe NS1 protein is secreted from infected cells and not presenton the virions [93ndash95]
In this regard similarly to what has been describedfor DENV studies performed using a naıve human scFvlibrary for panning with purified WNV E protein supportthe hypothesis that no Abs against the neutralizing DIII canbe isolated However DIII specific Abs were isolated in asubsequent study using immunoglobulin libraries obtainedfrom threeWNV infected patients for biopanning on purifiedinactivated virus virus-like particles consisting of prM andE proteins or recombinant E glycoprotein Although theproportion of DIII-specific mAbs was low (8) comparedwith anti-DII mAbs (47) Two out of the four anti-DIIImAbs were potently neutralizing and protective in vivowhereas only three out of the 24 anti-DII were weaklyneutralizing in vitro and non-protective in vivo In particularresidues that are critical for neutralization lies on the regionsspanning amino acids 305ndash312 330ndash333 and 365 that arelocated on adjacent exposed loops ofDIIIHowever sequencealignment of E protein of different flaviviruses such as DENVand JEV revealed a considerable variation compared tothe whole E protein This observation further suggests thatdifferently from broadly cross-reactive anti-DII Abs anti-DIII neutralizing Abs are virus-type specific However of
BioMed Research International 11
these mAbs two (CR4374 and CR4353) protected mice fromlethal WNV challenge at 50 protective doses of 129 and357 120583gkg of body weight respectively [50]
Sanchez et al using three different immunization strate-gies (ie inactivated virus naked DNA and recombinantprotein) isolated ninemurinemAbsmost of which bound toconformation-dependent epitopes in DIII of the E protein Inparticular neutralizing mAbs named 8B10 11C2 10C5 and17C8 were obtained from mice immunized with inactivatedvirus alone or in combination with a DNA plasmid andbound to the same region of DIII with high affinity Incontrast mAbs obtained by immunization with a solubleversion of the E glycoprotein did not exhibit neutralizingactivity These non-neutralizing mAbs were cross-reactivewith several other flaviviruses including SLEV JEV YFVand Powassan virus confirming the conserved nature ofFlavivirus non-neutralizing epitopes [96]
Gould et al isolated 11 unique human single-chain vari-able region Ab fragments (scFvs) that bind the E protein ofWNVAmong them a humanmAb namedmAb11 expressedas a scFv-Fc fusion protein was further characterized Itrecognizes the fusion loop at the distal end of DII ofthe WNV E protein and cross-reacts with all four DENVserotypes and provides protection against DENV2 and 4as well as WNV [42] Moreover therapeutic studies of thismAb in WNV-infection model mice provided substantialprotection when administered after 5 days post-infectionInterestingly a neutralization escape variant of this mAbfailed to cause lethal encephalitis (at higher infectious doses)or induce the inflammatory responses associated with blood-brain barrier permeability in mice compared to the parentalWNV suggesting an important role for the fusion loop inviral pathogenesis [43]
Oliphant et al isolated an anti-DIII E protein mAbnamed E16 fromhybridomas obtained after immunization ofmice with recombinant WNV E protein which neutralizedall WNV strains with PRNT50 values of 4 to 18 ng andPRNT90 values of 53 to 297 ng One hundred microgramsof mAb protected greater than 90 of mice from lethalinfection and even a single 4120583g treatment of E16 on day 2 afterinfection prevented mortality Moreover humanization ofthis mAb confirmed as therapeutically effective in mice [44]Subsequent studies revealed that mAb E16 neutralization ismediated by engagement of four discontinuous segmentsof DIII including the amino-terminal region (amino acidresidues 302ndash309) and the three connecting loops BC (aminoacid residues 330ndash333) DE (amino acid residues 365ndash368)and FG (amino acid residues 389ndash391) Moreover no ADE ofinfection was detected when E16 mAb was used at saturatingconcentrations [45 47]
Furthermore results of a Phase I safety study of thehumanized E16 mAb (designated MGAWN1) have beenreported and suggested that doses of up to 30mgkgwerewelltolerated with few mild adverse events and would providean excess of virus neutralizing activity for 3-4 weeks aftertreatment However a case of anti-MGAWN1 Ab elicitationoccurred with the consequent increased rate of clearance andindeed impacting efficacy A Phase II safety and efficacy studyof MGAWN1 is ongoing [97] Furthermore the possibility of
preventing or treating WNV-induced memory deficits wasrecently investigated In this study hamsters were treatedintraperitoneally with 32mgkg ofMGAWN1mAb at 45 daysafter subcutaneously challenging with WNV InterestinglyMGAWN1 prevented mortality weight loss and improvedfood consumption of WNV-infected hamsters compared tocontrols [98]
Recently Lelli et al isolated six anti-E mAbs frominactivated-WNV immunized mice In particular three ofthem (3B2 3D6 and 4D3) neutralized lineage I and II WNVwith the first two recognizing the same epitopes located onthe distal lateral surface of DIII (critical amino acid residueK307) Conversely 4D3mAb recognized a novel neutralizingepitope on DII (critical residues S276 and T278) Indeedfurther protective and therapeutic studies are needed toascertain their neutralizing activity in vivo [99]
Finally to conclude like DENV infection Abs directedagainst the M protein are not protective and neutralizingMoreover in humans a skewed humoral immune responseagainst DII has been frequently observed and confirmedalso with the hybridoma technology [50] The isolation andelicitation of neutralizingAbs directed against the fusion loopand DIII of the E protein represent thus the most challengingand promising goal for the development of new effectivetherapeutic inhibitors and immunogens respectively
27 Tick-Borne Encephalitis Virus (TBEV) Tick-borne ence-phalitis virus (TBEV) is one of the most dangerous agentscausing human neuroinfections occurring mostly in Europeand Asia and with a potential fatal prognosis [100] TBEV isbelieved to cause 3500ndash10000 human cases of encephalitisin Europe per year with a high morbidity in Russia CzechRepublic Austria and Germany In particular between 1990and 2007 an average of 8755 cases of TBE was reported peryear in Europe and Russia Despite the fact that Russia isthe country with most infections registered annually CzechRepublic incidence is among the highest in Europe with 400ndash1000 clinical cases reported every year [101]
Three subtypes of TBEV are classified namely EuropeanSiberian and Far-Eastern sharing most of the genetic andantigenic features [102] In fact a high degree of antigenichomogeneity between different strains of TBEV has beendescribed [103]
Clinical manifestations vary among the subtypes butusually they start with a short febrile period of 7ndash14 daysafter the tick bite Fatigue headache and pain in the neckshoulders and lower back together with high fever andvomitingmay be present [100]Thesemanifestations are oftenfollowed by an asymptomatic phase lasting from 2 to 10days after remission from the fever with possible progres-sion to neurological disease Neurological symptoms includemeningitis encephalitis myelitis and radiculitis Mortalityoccurs in 1-2 of the European subtype-infected patientsbut fatal prognosis can occur in up to 20ndash40 of the Far-Eastern subtype-infected patients [104] Mortality rates ofthe Siberian subtype are similar to those observed for theEuropean [105]
Effective and safe vaccines against TBEV produced frominactivated virus have been developed and licensed for their
12 BioMed Research International
use in humans However an emergency therapy in theabsence of a mass immunization is needed as no effectivetreatments are yet available [106]
271 mAbs against TBEV Levanov et al described the chim-erization of two murine mAbs (13D6 and 10C2) directedagainst the DIII and DII respectively of the TBEV E glyco-protein The chimeric mAbs present binding characteristicssimilar to the parentalmAbsMoreover as the parentalmAbsonly the chimeric mAb 13D6 was able to neutralize TBEVinfectivity in vitro In particular neutralization studies withthe murine chimeric and scFv forms of mAb 13D6 were per-formed In particular murine 13D6 showed an IC50 titer of115 120583gmL in Focus Reduction Neutralization Tests (FRNT)against TBEV strain 205 and of 29 120583gmL in PRNT againstTBEV strain Softjin Chimeric 13D6 showed an IC50 titer of45 120583gmL in FRNT against TBEV strain 205 and of 19 120583gmLin PRNT against TBEV strain Softjin ScFv 13D6 showed anIC50 titer of 167 120583gmL in FRNT against TBEV strain 205and of 112 120583gmL in PRNT against TBEV strain Softjin [62]
28 Japanese Encephalitis Virus (JEV) Japanese encephalitisvirus (JEV) is a mosquito-borne flavivirus that over thepast few decades has caused several outbreaks throughoutChina Southeast Asia Australia and Papua New Guineawith a prevalence recently estimated to be of about 70000casesyear [107ndash109] A 40 mortality was recorded in someof the JEV-affected areas Moreover many survivors facesome neurological problems and complications [108] Since1995 the disease has also emerged in Non-Asian regions suchas Northern Australia [110 111] The situation in SoutheastAsia however is further complicated by the overlappingepidemics of JEV and DENV as well as sporadic casesof WNV infections detected in some of the affected areasparticularly in India [112]
Effective vaccines both live-attenuated and inactivatedstrains of JEV have been developed and licensed in majorcountries However as for the other arthropod-borne Fla-vivirus members no specific antiviral drugs are currentlyavailable [4]
281mAbs against JEV Aspreviously described for the otherFlavivirusmembers many groups isolated and characterizedanti-JEVmAbs showing different neutralizing and protectiveproperties In this regard Gupta et al used combinations ofanti-E JEV mAbs (Hs1-4) in mice protection experimentsIn particular they found that the singularly mAb protectionranged from 45 to 65 when 100120583g of mAb were adminis-tered while equimolar combinations of two or three mAbsgave 85ndash90 or 100 protection respectively [113] In similarexperiments Lee et al demonstrated that a Flavivirus anti-NS1 mAb named 16NS1 cross-reacted with JEV as well asWNV and exhibited protective activity against WNV as wellas a lethal JEV infection However no neutralizing activitywas observed using this mAb against both WNV and JEV inin vitro experiments suggesting the participation of otherAb-mediatedmechanisms in vivo In particular 95ofmicewere
protected when 500120583g of mAb were administered intraperi-toneally and concomitantly to intramuscular injection of JEV
Overlapping peptide mapping analysis combined withsite-specific mutations identified the 116KAWGKSILFA125region and critical amino acid residues W118 and I122 as16NS1 mAb epitope highly conserved in WNV and JEVstrains [114]
Arakawa et al isolated from a combinatorial human Fablibrary constructed from peripheral blood lymphocytesobtained from JEV hyperimmune volunteers Among 188randomly selected clones FabTJE12B02 showed the best 50focus reduction endpoint at the concentration of 502 120583gmLagainst the JEV strain Nakayama [55]
Goncalvez et al isolated three mAbs named Fabs A3B2 and E3 by repertoire cloning from chimpanzees initiallyimmunized with inactivated JE-VAX and then boosted withattenuated JEV SA14-14-2 In particular these mAbs reactedwith epitopes in three different E domains in DI (amino acidresidue K179) in DII (I126) and in DIII (G132) for Fabs A3B2 and E3 respectively Moreover these Fabs as well as thederived humanized counterpart mAbs exhibited high neu-tralizing activities against a broad spectrum of JEV genotypestrains Moreover these mAbs exhibited a 50 protectivedose of 084 120583g (B2) 58120583g (A3) and 247120583g (E3) in mousemodels Finally administration of 200 120583gmouse of mAb B2one day after otherwise lethal JEV infection protected 50of mice and significantly prolonged the average survival timecompared to that of mice in the unprotected group [54]
29 St Louis Encephalitis Virus (SLEV) St Louis encephalitisvirus (SLEV) was first discovered as the mosquito-borneagent responsible for over 1000 cases of encephalitis duringa 1933 summer outbreak in St Louis (Missouri) and nowis a reemerging human pathogen widely distributed in theAmerican continent causing several human encephalitisoutbreaks over the last 80 years Additional epidemics haveindeed occurred from 1964 to 2006 in the Americas rangingfrom the US to Argentina and Brazil As a member of theFlavivirus genus the SLEVE glycoprotein ectodomain is 68identical to serocomplex-related JEVEbut only 46 and 40identical to those ofDENV2 andTBEV viruses fromdifferentserocomplexes [115]
291 mAbs against SLEV In 1983 Roehrig et al isolatedtwenty-one hybridomas producing murine mAbs specificfor the E glycoprotein of SLEV strain MSI-7 Serologicreactivities were initially determined by cross-reactivity indi-rect immunofluorescence assays using 22 strains of SLEVand 8 other related flaviviruses Four groups demonstratingtype- subcomplex- supercomplex- and group-specific reac-tivity patterns were identified Analysis of hemagglutination-inhibition and virus neutralization subdivided the cross-reactivity groups into eight epitopes (E-1a b c d E-2 E-3 and E-4a b) one of them following localized in DII (4b)[52 53]
Moreover among the previously described isolated anti-WNV mAbs by Gould et al the chimeric scFv-Fc 79 effec-tively neutralized also SLEV resulting in gt80 of PRNT80when used at 5 120583gmL [42]
BioMed Research International 13
3 Alphaviruses
Major human arboviruses in the Togaviridae family Chikun-gunya (CHIKV) and Venezuelan equine encephalomyelitis(VEEV) viruses belong to the Alphavirus genus which iscomposed of viruses with an icosahedral nucleocapsid sur-rounded by a lipid envelope and glycoprotein spikes Thestructural proteins of alphaviruses arise through co- andpost-translational processing of a polyprotein encoded by asingle positive-stranded RNAproducing the capsid (C) PE26 K and E1 The PE2 glycoprotein is a precursor containingthe E3 glycoprotein fused to the amino terminus of the E2envelope glycoprotein The PE2 glycoprotein is followed by6K a small membrane-associated protein and E1 the secondpolypeptide component of glycoprotein spikes
Trimerized heterodimers of the E1 and E2 viral glyco-proteins form the surface spikes and contain determinantsof viral tropism and virulence The E3 glycoprotein acts as asignal for transport of PE2 across themembranes of the roughendoplasmic reticulum and may promote the formation andintracellular transport of E1-PE2 heterodimers to the cellsurface During transport to the cell surface PE2 undergoesa maturational cleavage event by a furin-like protease toproduce E2 and E3 [116]
The E2 glycoprotein promotes specificity of virus bindingto the host cell surface and is a target of the humoral immuneresponse The E1 glycoprotein mediates fusion of the virionenvelope with the membranes of acidified endosomes allow-ing release of the nucleocapsid into the cytoplasm and theonset of viral replicationAlphavirus E1 shares no appreciablesequence identity to Flavivirus E protein Despite differencesin their amino acid sequences and arrangements on the viralparticle the structures of E and E1 are remarkably similarIndeed the conservation of three domains has been welldocumented in crystal structures of flavivirus E ectodomainsfrom TBEV DENV JEV and WNV as well as Alphavirus E1from Semliki Forest virus (SFV) and CHIKV Abs to the E1glycoprotein do not typically neutralize virus infectivity invitro but can protect against lethal challenge in animals
By analogy with the prM protein of dengue Flavivirusfurin cleavage of PE2 may begin after transit of an acidiclate component of the Golgi body where E3 is thought tosuppress the acid pH-triggered activation of glycoprotein E1fusion capability Protective anti-E3 mouse mAbs have alsobeen described [117]
31 Chikungunya Virus (CHIKV) The chikungunya virus(CHIKV) belongs to the Semliki Forest clade and was firstlyisolated in 1953 in Tanzania during an epidemic outbreakfollowing occurred also in Asia and Africa In the last fivedecades several CHIKV outbreaks have been described bothin Africa and Asia separated by gaps lasting from two totwenty years In 2005-2006 about 300000 cases out of785000 inhabitants were reported in La Reunion island witha fatal prognosis for 237 of the patients [117] Neither Europenor the Americas have had outbreaks of CHIKV so far exceptfor imported isolated cases Three genotypes of CHIKV aredescribed Asian EastCentralSouth African (ECSA) andWest African with an amino acid identity spanning from
952 to 998 Recent epidemics in Africa and Indiansubcontinent were caused by strains belonging to the ECSAgenotype Transmitted by Aedes mosquitoes CHIKV ismaintained in the human population by human-mosquito-human transmission [1] The disease is characterized bydengue-like symptoms such as chills high fever headacheand persistent myalgia and a further incapacitating arthralgia(from which the name chikungunya) which affects 40 ofinfected subjects However prognosis is rarely fatal [116]
There is currently no commercial vaccine and antiviraltreatment forCHIKV although some candidate vaccines havebeen tested in humans In this regard in 2000 US Armyperformed a Phase II clinical trial testing a live-attenuatedCHIKV vaccine (TSI-GSD-218) derived from a 1962 strain(15561) of an outbreak in Thailand Out of 58 every patientdeveloped neutralizingAbs and 5 lamentedmild tomoderatejoint pain [117] A phase III trial of this candidate vaccineis ongoing Furthermore a new formulation using virus-likeparticles was able to induce neutralizing Abs in macaquesagainst different CHIKV strains [117 118] Indeed it has beendescribed that infection seems to elicit long-lasting protectiveimmunity and cross-protection among CHIKV and otheralphaviruses
CHIKV entry into host cell is demonstrated to be medi-ated by envelope glycoproteins E1 and E2 which allowvirus fusion to cell membrane in low pH conditions andrecognition of an unknown cellular receptor respectively[116 119ndash121] Even though the genome replication relies onan error-prone RNA-dependent RNA-polymerasis CHIKVstrains S27 and 05115 Reunion showed in E1 and E2 alow amino acid variation of 068 and 33 respectivelyand such a low variability may be needed for an effectivereplication in two phylogenetically distant hosts [116 122]Recently the crystal structure of E1 and E2 heterodimerhas been resolved both at low pH and natural conditionsshedding light on E1 and E2 ectodomainsrsquo structure Inparticular similarly to flaviviruses E protein E1 ectodomainis made of the N-terminal DI the DII containing the fusionloop and the DIII at the C-terminal On the other hand E2ectodomain comprises theN-terminal domainA the domainB supposed to interact with the host cellrsquos unknown receptorand domain C at the C-terminal E1 DIII and E2 domain Care located close to the viral membrane [123 124]
311 mAbs against CHIKV Warter et al described 5F10 and8B10 two human mAbs which strongly neutralized severalCHIKV isolates in vitro without cross-reactivity againstother alphaviruses but to Onyong-nyong virus [56] Mixedpreparation of 5F10 and 8F10 did not show neither synergisticnor addictive effect in vitro and studies upon escape mutantsdemonstrated that 5F10 mAb binds at the tip of the E2domain B while 8B10 recognizes residues close to E1 fusionloop and amino acids within E2 domain A which form atransitional epitope under low pH conditions The authorsspeculated that 5F10 and 8B10 may inhibit CHIKV entry andfusion to the cell membrane respectively It is worth notingthat the previously mentioned escape mutants displayedmutations associated with reduced viral fitness in vitro evenafter 13 neutralizationamplification rounds In the same
14 BioMed Research International
study cell-to-cell transmission was firstly demonstrated as anescapemechanism enhanced by the E2mutation R82G [122]
In further in vivo studies 5F10 and 8B10 significantlydelayed CHIKV-caused death of AGR129 mice both in pro-phylactic and therapeutic tests Interestingly in therapeutictreatment these mAbs showed a synergistic effect whenadministered in combination with the total amount of mAbsinjected being the same of the single-mAbs Possibly thesetreatment did not allow themice to survive CHIKV challengedue to Ig clearing and 6ndash10 days half-life usually described forhuman mAbs [125]
Recently Pal et al cloned thirty-six murine mAbs ableto neutralize the ECSA La Reunion 2006 OPY-1 strain ofCHIKV (CHIKV-LR) the majority of which also neutralizeinfection of other strains corresponding to the Asian andWest African genotypes In particularmAbCHK-152 showedthe highest and broadest neutralizing activity with FRNTvalues indicating an IC50 value of 1 to 3 ngmL dependingon the viral strain used Among the thirty-six mAbs four(CHK-102 CHK-152 and CHK-166 CHK-263) could pre-vent lethality of immunodeficient Ifnarminusminus C57BL6 micewhen administered one day before exposition with CHK-152 and CHK-263 showing the ability to protect mice at thelowest dose (10 120583g) Therapeutic studies were performed onthese mAbs administering a single dose of 100 120583g 24 hoursafter infection The highest activity was described for CHK-166 able to protect 63 of the mice Testing the combinedactivity of the mAbs in therapeutic studies administration ofCHK-166 plus CHK-152 in a dose of 250120583g each turned out tobe themost effective protecting 71 of the IfnarminusminusC57BL6mice 60 hours after infection [57]
32 Venezuelan Equine Encephalomyelitis Virus (VEEV)Venezuelan equine encephalomyelitis virus (VEEV) is main-tained in a natural transmission cycle between mosquitoesand small rodents The first documented outbreaks occurredin the 1930s and several epidemics have been reported so farin Latin-American countries such as Venezuela ColombiaPeru Ecuador Bolivia Honduras Mexico and Panama withhundreds of thousands of human cases being reported anda case-fatality rate up to 1 during the 1969 Ecuador out-break Clinical manifestations of VEE are indistinguishablefrom Dengue [126] Six serogroups (IndashVI) are currentlyrecognized within the VEEV complex VEEV caused humanand equine outbreaks in the Americas for nearly a centuryEquine epizootics have high mortality (38ndash83) leading toa high viremia followed by a lethal encephalitis and often tohuman epidemics involving thousands of cases and hundredsof deaths Indeed VEEV is infectious for humans by theairborne route and has been responsible for a number oflaboratory infections In humans the disease is usually self-limiting with a febrile illness with 1ndash4 of cases progressingto severe encephalitis It is a hazard to laboratory workershas been developed as a biological weapon and is a potentialbioterrorist agent [127] There are no antiviral drugs andvaccines licensed for the treatment of VEEV infection inhumans Indeed although experimental live-attenuated vac-cines have been developed (eg TC-83) with good levels ofprotection in equine and mice in humans they may fail to
give protection in the majority of cases [128] Consideringthe protective role exerted by humoral immune response inanimal models it could be concluded that treatment withspecific IgG may have a beneficial antiviral effect in humanairborne infections with pathogenic strains of VEEV
321 mAbs against VEEV Several works support that mAbsmay protect against airborne VEEV as well as after airborneexposure toVEEVAb reactivity to both surface glycoproteins(E1 and E2) is associated with neutralization of the virusin vitro and passive protection against virus challenge Aseries of anti-E1 and anti-E2 mouse and humanized mAbshave been described recently Among them Phillpotts et alexamined two anti-E2 murine mAbs named 1A4A-1 and1A3A-9 both having potent protective activity against subcu-taneous VEEV challenge in mice Both mAbs had a similarhalf-life (58 and 100 days) in mouse serum after a singleintraperitoneal dose suggesting that mAbs delivered by thisroute at or around the time of VEEV infection persists athigh levels in the blood as well as secretions in respiratorytransudation throughout the clinical course of the disease Inparticular both mAbs administered 24 h prior to airbornechallenges had a substantial protective effect (90ndash100)Treatment of mice 2 or 24 h after airborne infection with asingle intraperitoneal dose of 100 120583g of 1A3A-9 mAb led toapproximately 50 survival There was no beneficial effectwhen mAb treatment was delayed to 72 h Moreover therewas evidence of synergy in vitro in PRNT between 1A3A-9and 1A4A-1 as has been demonstrated for other virusesand mAb pairs However no synergy was found in mouseprotection when mAbs were delivered intraperitoneally as amixture in equal parts to animals challenged 24 h previouslywith VEEV [129]
In a subsequent study Phillpotts et al reported two otherprotective mAbs The 3B2A-9 mAb protected against allthe serogroup I strains while 1A3B-7 mAb protected wellagainst challenge with all of the viruses tested Both 3B2A-9 and 1A3B-7 protected against airborne exposure to theIAB serogroup virus strainTrinidad donkey (TrD)Howeverthere was no evidence of synergistic protection when thesemAbs were combined in equal proportions An intraperi-toneal dose of 10 120583g was sufficient to protect 50 of themice with either mAb [130] According to other data themechanism of protection did not appear to depend uponneutralization [131] Similar results were obtained by OrsquoBrienet al which described a non-neutralizing mAb (IgG2a)named CUF37-2a able to protect 50 of mice from asubcutaneous VEEV challenge when a dose of 915 120583g of mAbwas administered 24 h prior to challenge [132]
In a following study the 1A3B-7 mAb was humanized(and following reported as Hu1A3B-7) maintaining the samefeatures of its murine counterpart In particular evaluation ofin vitro studies indicated that Hu1A3B-7 retained both broadspecificity and neutralizing activity Furthermore in vivoexperiments showed that Hu1A3B-7 successfully protectedmice against lethal subcutaneous and aerosol challenges withVEEV strain TrD Moreover the effectiveness of the human-ization process was determined by assessing proliferationresponses in human T-cells to peptides derived from the
BioMed Research International 15
murine and humanized versions of the VH and VL domainsThis analysis showed that the number of human T-cellepitopes within the humanized Ab had been substantiallyreduced indicating that Hu1A3B-7 may have low immuno-genicity in vivo [133]
Similarly Hunt et al described a humanizedmurinemAbderived from the 3B4C-4 mAb recognizing the E2c epitope(amino acid residues 182ndash209) and following named Hy4-26C showing neutralizing activity similar to the murinemAb Moreover it was protective (70ndash100 of survival) at100 ng dose in mice animal model intraperitoneal inoculatedand at 500120583g for intranasal challenge (80) Therapeuticstudies in mice revealed that Hy4-26C was able to cure miceup to 24 h following infection at 10 120583g on the contrary themouse mAb when given 1 hour after virus challenge [63]
In another work Hunt et al isolated two neutralizinghumanized Fabs F5 and L1A7 employing a blocking strategy[134 135] The anti-E2 specific F5 IgG had a 70 PRNTendpoint of 10 ngmL equivalent to that described for themost effective neutralizing anti-VEEVE2mAbs In particularthe E1-specific hFab L1A7 had a PRNT endpoint of 3120583gmL300-fold lower than F5
Further studies revealed that F5 epitope is located inthe 115ndash199 amino acid region Moreover F5 IgG had potentability to protect mice from infection by either route whenadministered 24 h before exposure however mice treated24 h and 48 h after aerosol exposure developed central ner-vous system infections but exhibited no clinical signs ofdisease [135]
Hu et al reported that passive immunization with thehumanized chimericmousemAbhu1A4A1IgG1-2A inmice at50 120583g 24 h before or after virulent VEEV challenge providedcomplete protection indicating that hu1A4A1IgG1-2A haspotent prophylactic and therapeutic effects against VEEVinfection [136]
Anti-VEEV mAbs isolated from a non-human primategene library has also been reported In particular thehumanized chimeric scFv-Fc ToR67-3B4 recognized viableas well as formalin and 120573-propiolactone inactivated virusparticles It detected specifically the viral E1 envelope proteinof VEEV but did not react with reduced viral glycopro-tein preparations suggesting that recognition depends uponconformational epitopes The recombinant Ab was able todetect multiple VEEV subtypes and displayed only marginalcross-reactivity to otherAlphavirus species except for Easternequine encephalitis virus (EEEV) In addition the scFv-Fcfusion described here might be of therapeutic use since itsuccessfully inactivated VEEV in a murine disease modelIn particular when the recombinant Ab was administered 6hours after challenge 80 to 100 of mice survived lethalVEEV IAB or IE infection Forty to sixty percent of micesurvived when scFv-Fc ToR67-3B4 was applied 6 hours afterchallenge with VEEV subtypes II and former IIIA [137]
4 Bunyaviridae
TheBunyaviridae family contains human arboviruses belong-ing to the Orthobunyavirus Phlebovirus and Nairovirus
genera Bunyaviridae includes enveloped viruses with a frag-mented single-stranded RNA genome of negative polarityTheir tripartite genome consists of a small (S) a medium(M) and a large (L) fragment and the envelope glycoproteinsGn and Gc that are cleaved out of a polyprotein synthesizedby the M fragment These glycoproteins have been provedto mediate the formation of the virus particle to play a rolein the interaction with cell surface receptors to mediate theentry of the virus into cells and to serve as targets for themajority of neutralizing Abs described so far
Orthobunyavirus transmitted through mosquitoes ormidges vectors are divided in 18 serogroups based on cross-titrations in haemagglutination inhibition assays and neutral-ization assays and correlating with main vector preferencesHowever the most clinically important viruses belong onlyto two serogroups the California encephalitis and the Simbuserogroups
Crimean-Congo hemorrhagic fever virus (CCHFV) isconsidered the only one of clinical relevance within theNairovirus genus which uses tick as main vector and is divid-ed in seven serogroups while Toscana virus and Rift Valleyfever virus (RVFV) which are transmitted by sandflies andmosquitoes respectively belong to the Phlebovirus genus[138]
MAbs against theNairovirusCCHFV and the PhlebovirusRVFV have been described
41 Crimean-Congo Hemorrhagic Fever Virus (CCHFV)Crimean-Congo Hemorrhagic Fever Virus (CCHFV) dis-tribution covers the greatest geographic range of any tick-borne virus known as viral isolation andor disease hasbeen reported from more than 30 countries in SoutheasternEurope Africa Asia and Middle East [139 140] Namelyfrom 1953 to 2010 about 6000 human cases were reportedin Southeastern Europe CCHFV causes sporadic outbreakswith mortality rates ranging from 10 to 80 [141 142] Asignificant variation in the time of incubation has beendescribed [139] Prehemorrhagic symptoms include highfever chills headache photophobia and back and abdominalpains Among other symptoms neuropsychiatric changeshave been reported in some patients In severe cases 3ndash6 daysafter the onset of the disease hemorrhagic symptoms occurwith petechiae ecchymosis bleeding in the form of melenahematemesis epistaxis [139]
In comparisonwith the nucleocapsid proteins inCCHFVGn and Gc show a higher degree of antigenic variabilityprobably due to their exposition to the host immune systemhypothesis corroborated in the description by Hewson et alin 2004 of four M segment phylogenic groups namely M1M2 M3 and M4 [141 142]
411 mAbs against CCHFV Blackburn et al firstly describedin 1987 murine anti-CCHFV mAbs able to recognize nucle-ocapsid proteins but no neutralizing activity has beendescribed thus far [143]
On the other hand Bertolotti-Ciarlet et al produced apanel of murine mAbs recognizing conformational epitopes
16 BioMed Research International
within theGn andGc glycoproteins expressed by the CCHFVstrain IbAr10200 The effectiveness of the mAbs was studiedby performing PRNT80 and none of the anti-Gn mAbsshowed neutralizing activity although many of the anti-GcmAbs neutralized IbAr10200 strain in vitro with clones 8A15E3 12A9 6C2 and 9H3 showing activity at a gt5120 dilutionMAbs 11E7 and 30F7 exhibited neutralizing activity at 2560dilution A suckling mice protection test was thereforeperformed for these mAbs The previously mentionedanti-Gc Abs were capable to protect mice to an appreciabledegree when applied 24 hours before and in a weakermanner 24 hours after virus challenge Many of the anti-GnmAbs were able to confer significant protection to IbAr10200both 24 hours before and after virus administration Therelevant effectiveness of the anti-Gn mAbs 6B12 10E1113G8 and 10G4 suggests that these mAbs may possess someneutralizing activity due to Ab-based effector mechanisms(eg ADCC) [58]
In further studies the same group characterized thebroadly cross-reactive neutralizing activity of the previouslydescribed murine mAb 11E7 able to bind conformationalepitopes within the C-terminal region of the Gn glycoproteinof all CCHFVM groups [58 141]
42 Rift Valley Fever Virus (RVFV) Similarly to CCHFV RiftValley Fever Virus (RVFV) causes sporadic outbreaks Thelargest epidemic occurred in Egypt in 1977 with an estimated200000 infections 18000 patients manifesting symptomsand 600 fatal prognoses [144] Symptoms include hemor-rhagic episodes fever encephalitis and blindness [145]
The major target of the humoral immune response is theRVFV glycoproteins Gc (G1) and Gn (G2) ranging fromnucleotides 480ndash2090 and 2091ndash3614 respectively [144]
421 mAbs against RVFV Within G2 glycoprotein are des-cribed three neutralizing antigenic sites namely epitope Inucleotides 792ndash893 (amino acid residues 258ndash291) epitopeII nucleotides 1164ndash1196 (amino acid residues 382ndash392) andepitope IV nucleotides 858ndash917 (amino acid residues 280ndash299) [59 145]
Despite the high degree of variability observed inCCHFVglycoproteins RVFV G2 presented an unexpected high con-servation among 22 isolates [59] In the same study onlythe LUNYO and 900060 isolates were resistant to in vitroneutralization by murine mAb 4D4 (recognizing epitope II)All other isolates were neutralized with PRNT80 titers from10240 togt81920On the other hand only SNS isolate showedreduced sensitivity to 4ndash39-CC (epitope IV) All other isolateswere neutralized with PRNT80 titers from 20480 to gt81920Protection studies on mice would be of interest for thedevelopment of a passive immunization prophylaxis [59]
Protection studies from lethal South African RVFVAN1830 strain infection were successfully performed on abattery of anti-G1 and anti-G2 mAbs produced by Besselaarand Blackburn with the strongly neutralizing anti-G1 mAb3E5 and anti-G2mAb 9C4 being the most effective [146]The9C4 antigenic area maps within G2 epitope I and a broadlyneutralizing activity may be investigated
5 Conclusions
Arbovirus infections have acquired increasing interest giventhe augmented globalization and tourism movement all overthe world that could be at the basis of epidemic eventsFurthermore the same vectors can sometimes transmit sev-eral arboviruses concomitantly complicating the diagnosis aswell as the therapy For example confusing mixed epidemicshave occasionally been described such as YFV plus CHIKVDENVplus CHIKV ormore recentlyPlasmodium falciparummalaria plus DENV1 and CHIKV on Madagascarrsquos east coast[1]
In the last decade several efforts have been employed inthe development of effective immunogens and therapeuticsgiving the current absence of specific antiviral drugs as wellas effective vaccines for themost diffused arbovirus infections(ie DENV andWNV)
However at the moment mosquito control is the bestavailable method for preventing arbovirus infections Thusto control the emerging public health problem of arbovirusinfections new antiviral therapeutic strategies that providepotent and broadly cross-protective immunity (especially forflavivirus infections) are an urgent globally medical need
As described in this review the majority of highlyneutralizing mAbs are of murine origin and indeed theirutility in the treatment of these infections is limited In facttheir employment can elicit an immunogenic response (iehuman anti-mouse Abs HAMA) the therapeutic efficacycould be reduced by a relatively faster clearance in humans(compared to human Abs) potentially exacerbated by theHAMA response Finally murine Abs exhibit relatively weakeffector functions (eg ADCC) compared to human AbsIndeed chimerization humanization or better the isolationand characterization of fully human broadly neutralizingmAbs would be the best choice for their possible use in afuture mAb-based therapy against arbovirus infections [147]
Finally possible ADE of infection mechanisms should beevaluated before considering a mAb as a possible candidatetherapeutic in a post-exposure setting as well as in the devel-opment of new Ab-eliciting immunogens
Conflict of Interests
The authors declare no conflict of interests in particular withOmriGam and any other cited Ab-based preparation men-tioned in the paper
References
[1] N Cleton M Koopmans J Reimerink G J Godeke and CReusken ldquoCome fly with me review of clinically importantarboviruses for global travelersrdquo Journal of Clinical Virology vol55 no 3 pp 191ndash203 2012
[2] F X Heinz H Holzmann A Essl and M Kundi ldquoField effec-tiveness of vaccination against tick-borne encephalitisrdquoVaccinevol 25 no 43 pp 7559ndash7567 2007
[3] A D Barrett and D E Teuwen ldquoYellow fever vaccinemdashhowdoes it work and why do rare cases of serious adverse events
BioMed Research International 17
take placerdquo Current Opinion in Immunology vol 21 no 3 pp308ndash313 2009
[4] S B Halstead and S J Thomas ldquoNew vaccines for Japaneseencephalitisrdquo Current Infectious Disease Reports vol 12 no 3pp 174ndash180 2010
[5] R Burioni M Perotti N Mancini and M Clementi ldquoPer-spectives for the utilization of neutralizing human monoclonalantibodies as anti-HCV drugsrdquo Journal of Hepatology vol 49no 2 pp 299ndash300 2008
[6] R A Diotti G A Sautto L Solforosi NManciniM Clementiand R Burioni ldquoNeutralization activity and kinetics of twobroad-range human monoclonal IgG1 derived from recombi-nant Fab fragments and directed against Hepatitis C virus E2glycoproteinrdquo New Microbiologica vol 35 no 4 pp 475ndash4792012
[7] R Burioni F Canducci N Mancini et al ldquoMolecular cloningof the first humanmonoclona antibodies neutralizing with highpotency Swine-origin Influenza A pandemic virus (S-OIV)rdquoNew Microbiologica vol 32 no 4 pp 319ndash324 2009
[8] P K Chandra S Hazari B Poat et al ldquoIntracytoplasmic stableexpression of IgG1 antibody targeting NS3 helicase inhibitsreplication of highly efficient hepatitis C Virus 2a clonerdquoVirology Journal vol 7 article 118 2010
[9] N Clementi D de Marco N Mancini et al ldquoA human mono-clonal antibody with neutralizing activity against highly diver-gent influenza subtypesrdquo PLoS ONE vol 6 no 12 Article IDe28001 2011
[10] D de Marco N Clementi N Mancini et al ldquoA non-VH1-69 heterosubtypic neutralizing human monoclonal antibodyprotects mice against H1N1 and H5N1 virusesrdquo PLoS ONE vol7 no 4 Article ID e34415 2012
[11] L Solforosi N Mancini F Canducci et al ldquoA phage displayvector optimized for the generation of human antibody com-binatorial libraries and the molecular cloning of monoclonalantibody fragmentsrdquoNewMicrobiologica vol 35 no 3 pp 289ndash294 2012
[12] E V Koonin and V V Dolja ldquoEvolution and taxonomy ofpositive-strand RNA viruses implications of comparative anal-ysis of amino acid sequencesrdquo Critical Reviews in Biochemistryand Molecular Biology vol 28 no 5 pp 375ndash430 1993
[13] J S Mackenzie D J Gubler and L R Petersen ldquoEmergingflaviviruses the spread and resurgence of Japanese encephalitisWest Nile and dengue virusesrdquo Nature Medicine vol 10 no 12pp S98ndashS109 2004
[14] T J SmithW E Brandt J L Swanson J MMcCown and E LBuescher ldquoPhysical and biological properties of dengue-2 virusand associated antigensrdquo Journal of Virology vol 5 no 4 pp524ndash532 1970
[15] J M Smit B Moesker I Rodenhuis-Zybert and J WilschutldquoFlavivirus cell entry and membrane fusionrdquo Viruses vol 3 no2 pp 160ndash171 2011
[16] R J Kuhn W Zhang M G Rossmann et al ldquoStructure ofdengue virus implications for flavivirus organization matura-tion and fusionrdquo Cell vol 108 no 5 pp 717ndash725 2002
[17] K Stiasny J H Aberle M Keller B Grubeck-Loebenstein andF X Heinz ldquoAge affects quantity but not quality of antibodyresponses after vaccination with an inactivated flavivirus vac-cine against tick-borne encephalitisrdquo PLoS ONE vol 7 no 3Article ID e34145 2012
[18] LThomasOVerlaetenACabie et al ldquoInfluence of the dengueserotype previous dengue infection and plasma viral load
on clinical presentation and outcome during a dengue-2 anddengue-4 co-epidemicrdquo American Journal of Tropical Medicineand Hygiene vol 78 no 6 pp 990ndash998 2008
[19] Y Zhang J Corver P R Chipman et al ldquoStructures of imma-ture flavivirus particlesrdquo EMBO Journal vol 22 no 11 pp2604ndash2613 2003
[20] A K I Falconar ldquoIdentification of an epitope on the denguevirus membrane (M) protein defined by cross-protective mon-oclonal antibodies design of an improved epitope sequencebased on common determinants present in both envelope (EandM) proteinsrdquo Archives of Virology vol 144 no 12 pp 2313ndash2330 1999
[21] K-J Huang Y-C Yang Y-S Lin et al ldquoThe dual-specific bind-ing of dengue virus and target cells for the antibody-dependentenhancement of dengue virus infectionrdquo Journal of Immunol-ogy vol 176 no 5 pp 2825ndash2832 2006
[22] A K I Falconar ldquoAntibody responses are generated to immun-odominant ELKKLE-typemotifs on the nonstructural-1 glyco-protein during live dengue virus infections inmice and humansimplications for diagnosis pathogenesis and vaccine designrdquoClinical and Vaccine Immunology vol 14 no 5 pp 493ndash5042007
[23] C W Mandl F Guirakhoo H Holzmann F X Heinz and CKunz ldquoAntigenic structure of the flavivirus envelope protein Eat the molecular level using the tick-borne encephalitis virus asa modelrdquo Journal of Virology vol 63 no 2 pp 564ndash571 1989
[24] F A Rey F X Heinz C Mandl C Kunz and S C HarrisonldquoThe envelope glycoprotein from tick-borne encephalitis virusat 2 A resolutionrdquo Nature vol 375 no 6529 pp 291ndash298 1995
[25] S L Allison K Stiasny K Stadler CWMandl and F XHeinzldquoMapping of functional elements in the stem-anchor regionof tick-borne encephalitis virus envelope protein Erdquo Journal ofVirology vol 73 no 7 pp 5605ndash5612 1999
[26] D E Klein J L Choi and S C Harrison ldquoStructure of a denguevirus envelope protein late-stage fusion intermediaterdquo Journal ofVirology vol 87 no 4 pp 2287ndash2293 2013
[27] S L Hanna T C Pierson M D Sanchez A A Ahmed M MMurtadha and R W Doms ldquoN-linked glycosylation of WestNile virus envelope proteins influences particle assembly andinfectivityrdquo Journal of Virology vol 79 no 21 pp 13262ndash132742005
[28] N Mancini G Sautto N Clementi et al ldquoNeutralization inter-fering antibodies a ldquonovelrdquo example of humoral immune dys-function facilitating viral escaperdquoViruses vol 4 no 9 pp 1731ndash1752 2012
[29] Y Modis S Ogata D Clements and S C Harrison ldquoVariablesurface epitopes in the crystal structure of dengue virus type3 envelope glycoproteinrdquo Journal of Virology vol 79 no 2 pp1223ndash1231 2005
[30] N Miller ldquoRecent progress in dengue vaccine research anddevelopmentrdquo Current Opinion in Molecular Therapeutics vol12 no 1 pp 31ndash38 2010
[31] J H Chavez J R Silva A A Amarilla and L T Moraes Figu-eiredo ldquoDomain III peptides from flavivirus envelope proteinare useful antigens for serologic diagnosis and targets forimmunizationrdquo Biologicals vol 38 no 6 pp 613ndash618 2010
[32] B Shrestha J D Brien S Sukupolvi-Petty et al ldquoThe develop-ment of therapeutic antibodies that neutralize homologous andheterologous genotypes of dengue virus type 1rdquo PLoS Pathogensvol 6 no 4 Article ID e1000823 2010
18 BioMed Research International
[33] G D Gromowski N D Barrett and A D T Barrett ldquoChar-acterization of dengue virus complex-specific neutralizing epi-topes on envelope protein domain III of dengue 2 virusrdquo Journalof Virology vol 82 no 17 pp 8828ndash8837 2008
[34] S Sukupolvi-Petty S K Austin W E Purtha et al ldquoType- andsubcomplex-specific neutralizing antibodies against domain IIIof dengue virus type 2 envelope protein recognize adjacentepitopesrdquo Journal of Virology vol 81 no 23 pp 12816ndash128262007
[35] S Sukupolvi-Petty S K Austin M Engle et al ldquoStructure andfunction analysis of therapeutic monoclonal antibodies againstdengue virus type 2rdquo Journal of Virology vol 84 no 18 pp9227ndash9239 2010
[36] Y-Q Deng J-X Dai G-H Ji et al ldquoA broadly flaviviruscross-neutralizingmonoclonal antibody that recognizes a novelepitope within the fusion loop of e proteinrdquo PLoS ONE vol 6no 1 Article ID e16059 2011
[37] J M Costin E Zaitseva K M Kahle et al ldquoMechanistic studyof broadly neutralizing human monoclonal antibodies againstdengue virus that target the fusion looprdquo Journal of Virologyvol 87 no 1 pp 52ndash66 2013
[38] CMMidgley A FlanaganH B Tran et al ldquoStructural analysisof a dengue cross-reactive antibody complexed with envelopedomain III reveals the molecular basis of cross-reactivityrdquoJournal of Immunology vol 188 no 10 pp 4971ndash4979 2012
[39] N J Moreland P Susanto E Lim et al ldquoPhage display appro-aches for the isolation of monoclonal antibodies against denguevirus envelope domain III from human and mouse derivedlibrariesrdquo International Journal ofMolecular Sciences vol 13 no3 pp 2618ndash2635 2012
[40] J J B Cockburn M E Navarro Sanchez N Fretes et alldquoMechanism of dengue virus broad cross-neutralization by amonoclonal antibodyrdquo Structure vol 20 no 2 pp 303ndash3142012
[41] R Rajamanonmani C Nkenfou P Clancy et al ldquoOn a mousemonoclonal antibody that neutralizes all four dengue virusserotypesrdquo Journal of General Virology vol 90 no 4 pp 799ndash809 2009
[42] L H Gould J Sui H Foellmer et al ldquoProtective and herapeuticcapacity of human single-chain Fv-Fc fusion proteins againstWest Nile virusrdquo Journal of Virology vol 79 no 23 pp 14606ndash14613 2005
[43] H Sultana H G Foellmer G Neelakanta et al ldquoFusion LoopPeptide of the West Nile virus envelope protein is essential forpathogenesis and is recognized by a therapeutic cross-reactivehuman monoclonal antibodyrdquo Journal of Immunology vol 183no 1 pp 650ndash660 2009
[44] T Oliphant M Engle G E Nybakken et al ldquoDevelopment ofa humanized monoclonal antibody with therapeutic potentialagainstWest Nile virusrdquoNature Medicine vol 11 no 5 pp 522ndash530 2005
[45] G E Nybakken T Oliphant S Johnson S Burke M S Dia-mond and D H Fremont ldquoStructural basis of West Nile virusneutralization by a therapeutic antibodyrdquo Nature vol 437 no7059 pp 764ndash769 2005
[46] G D Gromowski J T Roehrig M S Diamond J C LeeT J Pitcher and A D T Barrett ldquoMutations of an antibodybinding energy hot spot on domain III of the dengue 2 envelopeglycoprotein exploited for neutralization escaperdquo Virology vol407 no 2 pp 237ndash246 2010
[47] J T Roehrig R A Bolin andR G Kelly ldquoMonoclonal antibodymapping of the envelope glycoprotein of the dengue 2 virusJamaicardquo Virology vol 246 no 2 pp 317ndash328 1998
[48] R de Alwis S A Smith N P Olivarez et al ldquoIdentification ofhumanneutralizing antibodies that bind to complex epitopes ondengue virionsrdquo Proceedings of the National Academy of Sciencesof the United States of America vol 109 no 19 pp 7439ndash74442012
[49] S K Austin K A Dowd B Shrestha et al ldquoStructural basis ofdifferential neutralization of DENV-1 genotypes by an antibodythat recognizes a cryptic epitoperdquo PLoS Pathogens vol 8 no 10Article ID e1002930 2012
[50] M Throsby C Geuijen J Goudsmit et al ldquoIsolation and char-acterization of human monoclonal antibodies from individualsinfected with West Nile Virusrdquo Journal of Virology vol 80 no14 pp 6982ndash6992 2006
[51] A P Goncalvez R H Purcell and C-J Lai ldquoEpitope deter-minants of a chimpanzee Fab antibody that efficiently cross-neutralizes dengue type 1 and type 2 viruses map to inside andin close proximity to fusion loop of the dengue type 2 virusenvelope glycoproteinrdquo Journal of Virology vol 78 no 23 pp12919ndash12928 2004
[52] J T Roehrig J H Mathews and D W Trent ldquoIdentification ofepitopes on the E glycoprotein of Saint Louis encephalitis virususing monoclonal antibodiesrdquo Virology vol 128 no 1 pp 118ndash126 1983
[53] J T Roehrig L A Staudinger A R Hunt J H Mathews andC D Blair ldquoAntibody prophylaxis and therapy for flavivirusencephalitis infectionsrdquo Annals of the New York Academy ofSciences vol 951 pp 286ndash297 2001
[54] A P Goncalvez C-H Chien K Tubthong et al ldquoHumanizedmonoclonal antibodies derived from chimpanzee fabs protectagainst Japanese encephalitis virus in vitro and in vivordquo Journalof Virology vol 82 no 14 pp 7009ndash7021 2008
[55] M Arakawa T Yamashiro G-I Uechi M Tadano and ANishizono ldquoConstruction of humanFab (1205741120581) library and iden-tification of human monoclonal Fab possessing neutralizingpotency against Japanese encephalitis virusrdquo Microbiology andImmunology vol 51 no 6 pp 617ndash625 2007
[56] L Warter C Y Lee R Thiagarajan et al ldquoChikungunya virusenvelope-specific human monoclonal antibodies with broadneutralization potencyrdquo Journal of Immunology vol 186 no 5pp 3258ndash3264 2011
[57] P Pal K A Dowd J D Brien et al ldquoDevelopment of a highlyprotective combination monoclonal antibody therapy againstchikungunya virusrdquo PLoS Pathogens vol 9 no 4 Article IDe1003312 2013
[58] A Bertolotti-Ciarlet J Smith K Strecker et al ldquoCellular local-ization and antigenic characterization of Crimean-Congo hem-orrhagic fever virus glycoproteinsrdquo Journal of Virology vol 79no 10 pp 6152ndash6161 2005
[59] J K Battles and J M Dalrymple ldquoGenetic variation amonggeographic isolates of Rift Valley fever virusrdquo American Journalof Tropical Medicine and Hygiene vol 39 no 6 pp 617ndash6311988
[60] B AThibodeaux N C Garbino NM Liss et al ldquoA humanizedIgG but not IgM antibody is effective in prophylaxis and therapyof yellow fever infection in an AG12917D-204 peripheralchallenge mouse modelrdquo Antiviral Research vol 94 no 1 pp1ndash8 2012
BioMed Research International 19
[61] S Daffis R E Kontermann J Korimbocus H Zeller H-DKlenk and J Ter Meulen ldquoAntibody responses against wild-type yellow fever virus and the 17D vaccine strain charac-terization with human monoclonal antibody fragments andneutralization escape variantsrdquoVirology vol 337 no 2 pp 262ndash272 2005
[62] L N Levanov L E Matveev E P Goncharova et al ldquoChimericantibodies against tick-borne encephalitis virusrdquo Vaccine vol28 no 32 pp 5265ndash5271 2010
[63] A R Hunt S Frederickson C Hinkel K S Bowdish and J TRoehrig ldquoA humanized murine monoclonal antibody protectsmice either before or after challenge with virulent Venezuelanequine encephalomyelitis virusrdquo Journal of General Virologyvol 87 no 9 pp 2467ndash2476 2006
[64] J M da Silva Voorham I A Rodenhuis-Zybert N V A Nunezet al ldquoAntibodies against the envelope glycoprotein promoteinfectivity of immature dengue virus serotype 2rdquo PLoS ONEvol 7 no 3 Article ID e29957 2012
[65] T C Pierson Q Xu S Nelson et al ldquoThe stoichiometry ofantibody-mediated neutralization and enhancement of westnile virus infectionrdquoCell Host andMicrobe vol 1 no 2 pp 135ndash145 2007
[66] G Sautto N Mancini R A Diotti L Solforosi M Clementiand R Burioni ldquoAnti-hepatitis C virus E2 (HCVE2) glyco-proteinmonoclonal antibodies andneutralization interferencerdquoAntiviral Research vol 96 no 1 pp 82ndash89 2012
[67] R Burioni Y Matsuura N Mancini et al ldquoDiverging effectsof human recombinant anti-hepatitis C virus (HCV) antibodyfragments derived from a single patient on the infectivityof a vesicular stomatitis virusHCV pseudotyperdquo Journal ofVirology vol 76 no 22 pp 11775ndash11779 2002
[68] R J Phillpotts J R Stephenson and J S Porterfield ldquoAntibody-dependent enhancement of tick-borne encephalitus virus infec-tivityrdquo Journal of General Virology vol 66 no 8 pp 1831ndash18371985
[69] K A Dowd and T C Pierson ldquoAntibody-mediated neutraliza-tion of flaviviruses a reductionist viewrdquo Virology vol 411 no 2pp 306ndash315 2011
[70] A K I Falconar ldquoMonoclonal antibodies that bind to commonepitopes on the dengue virus type 2 nonstructural-1 andenvelope glycoproteins display weak neutralizing activity anddifferentiated responses to virulent strains implications forpathogenesis and vaccinesrdquo Clinical and Vaccine Immunologyvol 15 no 3 pp 549ndash561 2008
[71] K Meyer A Banerjee S E Frey R B Belshe and R Ray ldquoAweak neutralizing antibody response to hepatitis C virus enve-lope glycoprotein enhances virus infectionrdquo PLoS ONE vol 6no 8 Article ID e23699 2011
[72] R Burioni R AWilliamson P P Sanna F E Bloom and D RBurton ldquoRecombinant human Fab to glycoprotein D neutral-izes infectivity and prevents cell-to-cell transmission of herpessimplex viruses 1 and 2 in vitrordquo Proceedings of the NationalAcademy of Sciences of the United States of America vol 91 no1 pp 355ndash359 1994
[73] M S Rolph A Zaid N E Rulli and S Mahalingam ldquoDown-regulation of interferon-120573 in antibody-dependent enhancementof dengue viral infections of human macrophages is dependenton interleukin-6rdquo Journal of Infectious Diseases vol 204 no 3pp 489ndash491 2011
[74] S S Whitehead J E Blaney A P Durbin and B R MurphyldquoProspects for a dengue virus vaccinerdquoNature ReviewsMicrobi-ology vol 5 no 7 pp 518ndash528 2007
[75] S-M Lok V Kostyuchenko G ENybakken et al ldquoBinding of aneutralizing antibody to dengue virus alters the arrangement ofsurface glycoproteinsrdquoNature Structural andMolecular Biologyvol 15 no 3 pp 312ndash317 2008
[76] A P Goncalvez R Men C Wernly R H Purcell and C-J Lai ldquoChimpanzee Fab fragments and a derived humanizedimmunogiobulin G1 antibody that efficiently cross-neutralizedengue type 1 and type 2 virusesrdquo Journal of Virology vol 78no 23 pp 12910ndash12918 2004
[77] WM P BWahala A A Kraus L B HaymoreM A Accavitti-Loper and A M de Silva ldquoDengue virus neutralization byhuman immune sera role of envelope protein domain III-reactive antibodyrdquo Virology vol 392 no 1 pp 103ndash113 2009
[78] W D Crill H R Hughes M J Delorey and G-J J ChangldquoHumoral immune responses of dengue fever patients usingepitope-specific serotype-2 virus-like particle antigensrdquo PLoSONE vol 4 no 4 Article ID e4991 2009
[79] C Setthapramote T Sasaki O Puiprom et al ldquoHuman mono-clonal antibodies to neutralize all dengue virus serotypes usinglymphocytes from patients at acute phase of the secondaryinfectionrdquo Biochemical and Biophysical Research Communica-tions vol 423 no 4 pp 867ndash872 2012
[80] M Beltramello K L Williams C P Simmons et al ldquoThehuman immune response to dengue virus is dominated byhighly cross-reactive antibodies endowed with neutralizing andenhancing activityrdquo Cell Host andMicrobe vol 8 no 3 pp 271ndash283 2010
[81] C M Midgley M Bajwa-Joseph S Vasanawathana et al ldquoAnin-depth analysis of original antigenic sin in dengue virusinfectionrdquo Journal of Virology vol 85 no 22 pp 410ndash421 2011
[82] B Kaufmann M R Vogt J Goudsmit et al ldquoNeutralizationof West Nile virus by cross-linking of its surface proteins withFab fragments of the human monoclonal antibody CR4354rdquoProceedings of the National Academy of Sciences of the UnitedStates of America vol 107 no 44 pp 18950ndash18955 2010
[83] B Spurrier J M Sampson M Totrov et al ldquoStructural analysisof human and macaque mAbs 2909 and 25B implicationsfor the configuration of the quaternary neutralizing epitope ofHIV-1 gp120rdquo Structure vol 19 no 5 pp 691ndash699 2011
[84] J J SchlesingerMWBrandriss andT PMonath ldquoMonoclonalantibodies distinguish between wild and vaccine strains ofyellow fever virus by neutralization hemagglutination inhibi-tion and immune precipitation of the virus envelope proteinrdquoVirology vol 125 no 1 pp 8ndash17 1983
[85] M W Brandriss J J Schlesinger E E Walsh and M BrisellildquoLethal 17D yellow fever encephalitis in mice I Passive protec-tion by monoclonal antibodies to the envelope proteins of 17Dyellow fever and dengue 2 virusesrdquo Journal of General Virologyvol 67 no 2 pp 229ndash234 1986
[86] H Cho and M S Diamond ldquoImmune responses to West Nilevirus infection in the central nervous systemrdquo Viruses vol 4no 12 pp 3812ndash3830 2012
[87] D W Beasley A D Barrett and R B Tesh ldquoResurgence ofWest Nile neurologic disease in the United States in 2012 whathappened What needs to be donerdquo Antiviral Research vol 99no 1 pp 1ndash5 2013
[88] M Throsby J Ter Meulen C Geuijen J Goudsmit and J DeKruif ldquoMapping and analysis of West Nile virus-specific mon-oclonal antibodies prospects for vaccine developmentrdquo ExpertReview of Vaccines vol 6 no 2 pp 183ndash191 2007
[89] M S Diamond T G Roberts D Edgil B Lu J Ernst and EHarris ldquoModulation of dengue virus infection in human cells
20 BioMed Research International
by alpha beta and gamma interferonsrdquo Journal of Virology vol74 no 11 pp 4957ndash4966 2000
[90] Z Shimoni M J Niven S Pitlick and S Bulvik ldquoTreatmentof West Nile virus encephalitis with intravenous immunoglob-ulinrdquo Emerging Infectious Diseases vol 7 no 4 article 759 2001
[91] AHamdan PGreen EMendelsonM R Kramer S Pitlik andM Weinberger ldquoPossible benefit of intravenous immunoglob-ulin therapy in a lung transplant recipient with West Nile virusencephalitisrdquoTransplant InfectiousDisease vol 4 no 3 pp 160ndash162 2002
[92] M Haley A S Retter D Fowler J Gea-Banacloche and NP OrsquoGrady ldquoThe role for intravenous immunoglobulin in thetreatment of West Nile virus encephalitisrdquo Clinical InfectiousDiseases vol 37 no 6 pp e88ndashe90 2003
[93] K M Chung G E Nybakken B S Thompson et al ldquoAnti-bodies against West Nile virus nonstructural protein NS1prevent lethal infection through Fc 120574 receptor-dependent and-independent mechanismsrdquo Journal of Virology vol 80 no 3pp 1340ndash1351 2006
[94] E A Henchal L S Henchal and J J Schlesinger ldquoSynergisticinteractions of anti-NS1 monoclonal antibodies protect pas-sively immunized mice from lethal challenge with dengue 2virusrdquo Journal of General Virology vol 69 no 8 pp 2101ndash21071988
[95] J T Roehrig ldquoAntigenic structure of flavivirus proteinsrdquoAdvances in Virus Research vol 59 pp 141ndash175 2003
[96] M D Sanchez T C Pierson D McAllister et al ldquoCharacteri-zation of neutralizing antibodies to West Nile virusrdquo Virologyvol 336 no 1 pp 70ndash82 2005
[97] J H Beigel J L Nordstrom S R Pillemer et al ldquoSafety andpharmacokinetics of single intravenous dose of MGAWN1 anovel monoclonal antibody to West Nile virusrdquo AntimicrobialAgents and Chemotherapy vol 54 no 6 pp 2431ndash2436 2010
[98] C A Smeraski V Siddharthan and J D Morrey ldquoTreatmentof spatial memory impairment in hamsters infected with WestNile virus using a humanizedmonoclonal antibodyMGAWN1rdquoAntiviral Research vol 91 no 1 pp 43ndash49 2011
[99] D Lelli AMoreno E Brocchi et al ldquoWestNile virus character-ization and diagnostic applications of monoclonal antibodiesrdquoVirology Journal vol 9 article 81 2012
[100] T S Gritsun V A Lashkevich and E A Gould ldquoTick-borneencephalitisrdquo Antiviral Research vol 57 no 1-2 pp 129ndash1462003
[101] D Ruzek T S Gritsun N L Forrester et al ldquoMutations inthe NS2B and NS3 genes affect mouse neuroinvasiveness of aWestern European field strain of tick-borne encephalitis virusrdquoVirology vol 374 no 2 pp 249ndash255 2008
[102] M Ecker S L Allison T Meixner and F X Heinz ldquoSequenceanalysis and genetic classification of tick-borne encephalitisviruses from Europe and Asiardquo Journal of General Virology vol80 no 1 pp 179ndash185 1999
[103] H Holzmann M S Vorobyova I P Ladyzhenskaya et alldquoMolecular epidemiology of tick-borne encephalitis viruscross-protection between European and Far Eastern subtypesrdquoVaccine vol 10 no 5 pp 345ndash349 1992
[104] C W Mandl ldquoSteps of the tick-borne encephalitis virus repli-cation cycle that affect neuropathogenesisrdquo Virus Research vol111 no 2 pp 161ndash174 2005
[105] A V Atrasheuskaya T M Fredeking and G M IgnatyevldquoChanges in immune parameters and their correction in humancases of tick-borne encephalitisrdquo Clinical and ExperimentalImmunology vol 131 no 1 pp 148ndash154 2003
[106] K L Mansfield N Johnson L P Phipps J R Stephenson AR Fooks and T Solomon ldquoTick-borne encephalitis virusmdashareview of an emerging zoonosisrdquo Journal of General Virologyvol 90 no 8 pp 1781ndash1794 2009
[107] DWVaughn andCHHoke Jr ldquoThe epidemiology of Japaneseencephalitis prospects for preventionrdquo Epidemiologic Reviewsvol 14 pp 197ndash221 1992
[108] T Solomon H Ni D W C Beasley M Ekkelenkamp M JCardosa and A D T Barrett ldquoOrigin and evolution of Japaneseencephalitis virus in Southeast Asiardquo Journal of Virology vol 77no 5 pp 3091ndash3098 2003
[109] M R Wilson ldquoEmerging viral infectionsrdquo Current Opinion inNeurology vol 26 no 3 pp 301ndash306 2013
[110] J N Hanna S A Ritchie D A Phillips et al ldquoAn outbreakof Japanese encephalitis in the Torres Strait Australia 1995rdquoMedical Journal of Australia vol 165 no 5 pp 256ndash260 1996
[111] J N Hanna S A Ritchie D A Phillips et al ldquoJapanese ence-phalitis in north Queensland Australia 1998rdquo Medical Journalof Australia vol 170 no 11 pp 533ndash536 1999
[112] SGeorgeMGourie-Devi and J A Rao ldquoIsolation ofWestNilevirus from the brains of children who had died of encephalitisrdquoBulletin of theWorldHealthOrganization vol 62 no 6 pp 879ndash882 1984
[113] A K Gupta A A Koshy and V J Lad ldquoEnhanced protec-tion of mice against Japanese encephalitis virus infection bycombinations ofmonoclonal antibodies to glycoprotein ErdquoActaVirologica vol 55 no 2 pp 165ndash168 2011
[114] T H Lee B-H Song S-I Yun et al ldquoA cross-protective mAbrecognizes a novel epitope within the flavivirus NS1 proteinrdquoJournal of General Virology vol 93 no 1 pp 20ndash26 2012
[115] S C Weaver ldquoHost range amplification and arboviral diseaseemergencerdquo Archives of Virology Supplementum no 19 pp 33ndash44 2005
[116] M Solignat B Gay S Higgs L Briant and C Devaux ldquoRepli-cation cycle of chikungunya a re-emerging arbovirusrdquoVirologyvol 393 no 2 pp 183ndash197 2009
[117] O Schwartz and M L Albert ldquoBiology and pathogenesis ofchikungunya virusrdquo Nature Reviews Microbiology vol 8 no 7pp 491ndash500 2010
[118] W Akahata Z-Y Yang H Andersen et al ldquoA virus-like particlevaccine for epidemic Chikungunya virus protects nonhumanprimates against infectionrdquo Nature Medicine vol 16 no 3 pp334ndash338 2010
[119] M Kielian and A Helenius ldquopH-Iinduced alterations in thefusogenic spike protein of Semliki Forest virusrdquo Journal of CellBiology vol 101 no 6 pp 2284ndash2291 1985
[120] J White and A Helenius ldquopH-Dependent fusion between theSemliki Forest virus membrane and liposomesrdquo Proceedings ofthe National Academy of Sciences of the United States of Americavol 77 no 6 pp 3273ndash3277 1980
[121] J Dubuisson and C M Rice ldquoSindbis virus attachment isola-tion and characterization of mutants with impaired binding tovertebrate cellsrdquo Journal of Virology vol 67 no 6 pp 3363ndash33741993
[122] C Y Lee Y-W Kam J Fric et al ldquoChikungunya virus neutral-ization antigens and direct cell-to-cell transmission are revealedby human antibody-escapemutantsrdquo PLoS Pathogens vol 7 no12 Article ID e1002390 2011
[123] J E Voss M-C Vaney S Duquerroy et al ldquoGlycoproteinorganization of Chikungunya virus particles revealed by X-raycrystallographyrdquo Nature vol 468 no 7324 pp 709ndash712 2010
BioMed Research International 21
[124] L Li J Jose Y Xiang R J Kuhn and M G Rossmann ldquoStruc-tural changes of envelope proteins during alphavirus fusionrdquoNature vol 468 no 7324 pp 705ndash708 2010
[125] J Fric S Bertin-Maghit C I Wang A Nardin and L WarterldquoUse of human monoclonal antibodies to treat Chikungunyavirus infectionrdquo Journal of Infectious Diseases vol 207 no 2 pp319ndash322 2013
[126] P V Aguilar J G Estrada-Franco R Navarro-Lopez C FerroA D Haddow and S C Weaver ldquoEndemic Venezuelanequine encephalitis in the Americas hidden under the dengueumbrellardquo Future Virology vol 6 no 6 pp 721ndash740 2011
[127] S C Weaver R Winegar I D Manger and N L ForresterldquoAlphaviruses population genetics and determinants of emer-gencerdquo Antiviral Research vol 94 no 3 pp 242ndash257 2012
[128] S Paessler and S C Weaver ldquoVaccines for Venezuelan equineencephalitisrdquo Vaccine vol 27 no 4 pp D80ndashD85 2009
[129] R J Phillpotts ldquoVenezuelan equine encephalitis virus complex-specific monoclonal antibody provides broad protection inmurine models against airborne challenge with viruses fromserogroups I II and IIIrdquoVirus Research vol 120 no 1-2 pp 107ndash112 2006
[130] R J Phillpotts L D Jones and S C Howard ldquoMonoclonalantibody protectsmice against infection anddiseasewhen giveneither before or up to 24 h after airborne challenge with virulentVenezuelan equine encephalitis virusrdquoVaccine vol 20 no 11-12pp 1497ndash1504 2002
[131] J H Mathews J T Roehrig and D W Trent ldquoRole of com-plement and the Fc portion of immunoglobulin G in immunityto Venezuelan equine encephalomyelitis virus infection withglycoprotein-specific monoclonal antibodiesrdquo Journal of Virol-ogy vol 55 no 3 pp 594ndash600 1985
[132] L M OrsquoBrien C D Underwood-Fowler S A Goodchild AL Phelps and R J Phillpotts ldquoDevelopment of a novel mon-oclonal antibody with reactivity to a wide range of Venezuelanequine encephalitis virus strainsrdquoVirology Journal vol 6 article206 2009
[133] L M OrsquoBrien S A Goodchild R J Phillpotts and S D Per-kins ldquoA humanised murine monoclonal antibody protectsmice from Venezuelan equine encephalitis virus Evergladesvirus and Mucambo virus when administered up to 48h afterairborne challengerdquo Virology vol 426 no 2 pp 100ndash105 2012
[134] N Clementi N Mancini L Solforosi M Castelli M Clementiand R Burioni ldquoPhage display-based strategies for cloning andoptimization of monoclonal antibodies directed against humanpathogensrdquo International Journal of Molecular Sciences vol 13no 7 pp 8273ndash8292 2012
[135] A R Hunt R A Bowen S Frederickson T Maruyama JT Roehrig and C D Blair ldquoTreatment of mice with humanmonoclonal antibody 24h after lethal aerosol challenge withvirulent Venezuelan equine encephalitis virus prevents diseasebut not infectionrdquo Virology vol 414 no 2 pp 146ndash152 2011
[136] W-G Hu A L Phelps S Jager et al ldquoA recombinant human-ized monoclonal antibody completely protects mice againstlethal challenge with Venezuelan equine encephalitis virusrdquoVaccine vol 28 no 34 pp 5558ndash5564 2010
[137] T Rulker L Voss P Thullier et al ldquoIsolation and characterisa-tion of a human-like antibody fragment (scFv) that inactivatesVEEV in vitro and in vivordquo PLoS ONE vol 7 no 5 Article IDe37242 2012
[138] C T Walter and J N Barr ldquoRecent advances in the molecularand cellular biology of bunyavirusesrdquo Journal of General Virol-ogy vol 92 no 11 pp 2467ndash2484 2011
[139] C A Whitehouse ldquoCrimean-Congo hemorrhagic feverrdquo Anti-viral Research vol 64 no 3 pp 145ndash160 2004
[140] M Mertens K Schmidt A Ozkul and M H Groschup ldquoTheimpact of Crimean-Congo hemorrhagic fever virus on publichealthrdquo Antiviral Research vol 98 no 2 pp 248ndash260 2013
[141] A A Ahmed J M McFalls C Hoffmann et al ldquoPresence ofbroadly reactive and group-specific neutralizing epitopes onnewly described isolates of Crimean-Congo hemorrhagic fevervirusrdquo Journal of General Virology vol 86 no 12 pp 3327ndash33362005
[142] R Hewson A Gmyl L Gmyl et al ldquoEvidence of segment reas-sortment in Crimean-Congo haemorrhagic fever virusrdquo Journalof General Virology vol 85 no 10 pp 3059ndash3070 2004
[143] N K Blackburn T G Besselaar A J Shepherd and R Swane-poel ldquoPreparation and use of monoclonal antibodies for iden-tifying Crimean-Congo hemorrhagic fever virusrdquo AmericanJournal of Tropical Medicine and Hygiene vol 37 no 2 pp 392ndash397 1987
[144] T Ikegami ldquoMolecular biology and genetic diversity of RiftValley fever virusrdquoAntiviral Research vol 95 no 3 pp 293ndash3102012
[145] KKeegan andM S Collett ldquoUse of bacterial expression cloningto define the amino acid sequences of antigenic determinantson the G2 glycoprotein of Rift Valley fever virusrdquo Journal ofVirology vol 58 no 2 pp 263ndash270 1986
[146] T G Besselaar and N K Blackburn ldquoTopological mapping ofantigenic sites on the Rift Valley fever virus envelope glycopro-teins using monoclonal antibodiesrdquo Archives of Virology vol121 no 1ndash4 pp 111ndash124 1991
[147] L G Presta ldquoEngineering of therapeutic antibodies to mini-mize immunogenicity and optimize functionrdquo Advanced DrugDelivery Reviews vol 58 no 5-6 pp 640ndash656 2006
Submit your manuscripts athttpwwwhindawicom
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anatomy Research International
PeptidesInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporation httpwwwhindawicom
International Journal of
Volume 2014
Zoology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Molecular Biology International
GenomicsInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioinformaticsAdvances in
Marine BiologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Signal TransductionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Evolutionary BiologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Biochemistry Research International
ArchaeaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Genetics Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in
Virolog y
Hindawi Publishing Corporationhttpwwwhindawicom
Nucleic AcidsJournal of
Volume 2014
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Enzyme Research
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Microbiology

BioMed Research International 7Ta
ble1Con
tinued
mAb
Orig
inRe
activ
ityTargetprotein
Epito
peClon
ingstr
ategy
Form
atisotype
Neutralizingactiv
ityRe
ference
E16
(MGAW
N1)
Murine
WNV
EDIII(LR
aa3
02ndash309)
Hybrid
omafrom
immun
ized
micew
ithWNVE
IgG2bhum
anized
PRNT5
04ndash
18ng
PRNT9
053ndash297
ng[4445]
1A1D
-2Murine
DEN
V1ndash3
EDIII(Astr
andaa
307
310and312)
Hybrid
omafrom
immun
ized
micew
ithdifferent
pH-tr
eated
virus
IgG2a
DEN
V2PR
NT5
021n
M[4647]
1F4
Hum
an
DEN
V1
E(virion
)
DI-DII
Electro
fusio
nof
infected
mem
oryBcells
from
DEN
V-im
mun
esub
jects
with
EBV
IgG
DEN
V1IC5
0011120583gmL
[48]
2D22
DEN
V2
DIII
DEN
V2IC50
008120583gmL
5J7
DEN
V3
DI-DII
DEN
V3IC50
010120583gmL
E105
Murine
DEN
V1
E
DIII(BC
loop
G328
T329
andD330DE
loop
K361E
and
E362K
FGloop
K3
85)
Hybrid
omas
deriv
edfro
mC5
7BL6
IFN-120572120573Rminus
minusmice
infected
with
DEN
V1
IgG
PRNT5
0DEN
V-1
05ndash592ng
mL
[32]
E106
DIII(BC
loop
G328
T329
andD330DE
loop
K361E
and
E362K
FGloop
K3
85A
strand
S305
K307E
309K3
10and
E311)
PRNT5
0DEN
V-1
06ndash
592ng
mL
E111
DIII(CC1015840loop
)PR
NT5
038ndash25120583gmL
[49]
CR4374
Hum
anWNV
EDIII
Phaged
isplayof
scFv
IgGlib
rary
VH2-05V
L1e
PRNT5
0018120583gmL
PRNT9
0095120583gmL
[50]
CR4353
VH3-30V
k3-A27
PRNT5
00026120583
gmL
PRNT9
0364120583gmL
1A5
Chim
panzee
DEN
V1ndash4WNV
JEV
LGTV
EDII(aaG
106)
Phaged
isplayof
Fab
library
from
DEN
V1ndash4
infected
chim
panzees
Hum
anized
IgG1
PRNT5
0(120583gmL)
DEN
V1048
DEN
V2095
DEN
V332
DEN
V443
WNVD
ENV438
JEV
21LG
TV28
[51]
8 BioMed Research International
Table1Con
tinued
mAb
Orig
inRe
activ
ityTargetprotein
Epito
peClon
ingstr
ategy
Form
atisotype
Neutralizingactiv
ityRe
ference
mAb
11Hum
anDEN
V1ndash4WNV
EDII(fu
sionloop
W101G104G106)
scFv
library
scFv-Fc
PRNT8
0(120583gmL)
DEN
V2625
WNV
125
[4243]
3B4C
-4
Murine
SLEV
E
1a
Hybrid
oma
IgG
SLEV
PRNTlt17
[5253]
1B2C
-5SLEV
1bSLEV
PRNTlt17
6B5A
-2SLEV
1cSLEV
PRNT
48
4A4C
-4SLEV
1dSLEV
PRNT
29
1B5D
-1SLEV
JEV
2SLEV
PRNTlt17
2B5B
-3SL
EVJEV
MVEV
WNV
YFV
3SLEV
PRNT
23
2B6B
-2AllFlaviviru
s4a
SLEV
PRNTlt17
6B6C
-1AllFlaviviru
s4b
(inDII)
SLEV
PRNT
23
A3
Chim
panzee
JEV
E2
DIa
aK1
79Ph
aged
isplay
Hum
anized
PRNT5
0004ndash0
2nM
[54]
B2DIIaaI126
Phaged
isplay
PRNT5
0002ndash2
nM
E3DIIIa
aG132
Phaged
isplay
PRNT5
0014ndash0
93n
M
FabT
JE12B0
2Hum
anJEV
ENA
Phaged
isplay
Fab
FRNT5
0502120583gmL
[55]
5F10
Hum
anCH
IKV
E2Dom
ainB
EBVtransfo
rmation
IgG11205822
IC50lt100n
gmL
[56]
8B10
Hum
anCH
IKV
E1-E2
E2Dom
ainA
EBVtransfo
rmation
IgG1k
IC50lt100n
gmL
CHK-
152
Murine
CHIK
VE2
aa59
Hybrid
oma
IgG2c
IC50
1ndash3n
gmL
[57]
11E7
Murine
CCHFV
Gn
C-ter
Hybrid
oma
IgG
PRNT8
0diluted
12560
[58]
4-39-C
CMurine
RVFV
G2
Dom
ainIV
Hybrid
oma
IgG
PRNT8
0diluted
1204
80ndash181920
[59]
2C9
Murine
YFV
EHybrid
oma
IgG2a
PRNT9
011
0417D
1105
2Asib
i[60]
5A 7A R3(27)
Hum
anYF
VE
DI-DII
Phaged
isplay
ScFv
PRNT9
001ndash03120583
gmL
[61]
13D6
Murine
TBEV
EDIII
Hybrid
oma
IgGchimeric
PRNT5
019120583gmL
FRNT5
045120583
gmL
IC50
19ndash167120583
gmL
[62]
3B4C
-4(H
y4-26C
)Murine
VEE
VE2
aa182ndash209
Hybrid
oma
IgGhum
anized
PRNT7
0(120583gmL)
394ndash125
[63]
BioMed Research International 9
The structure of the 1A1D-2 bound to EDIII indicates that themAb binds to sites that are transiently exposed during viralldquobreathingrdquo at 37∘C and block infection during attachmentof the virion to the cell In particular this mAb neutralizesDENV1 2 and 3 serotypes and residues K307 and K310 arethemost critical residues for binding of 1A1D-2mAb [46 75]
Finally Shrestha et al immunized mice with a genotype2 strain of DENV1 virus and generated 79 new mAbs 16of which strongly inhibited infection by the homologousvirus and localized to DIII of E protein Surprisingly onlytwo mAbs E105 and E106 retained strong binding andneutralizing activity against all five DENV1 genotypes as wellas being protective in immunocompromised infected miceMoreover E105 and E106 exhibited therapeutic activity evenwhen administered as a single dose four days after inoculationwith a heterologous genotype 4 strain of DENV1 in the samemouse model [32]
243 Other Non-HumanmAbs Other non-human and non-murine as well as humanized mAbs directed against the Eglycoprotein have been described In this regard Goncalvezet al isolated a large panel of anti-E Fab fragments from chim-panzees infected with all four DENV serotypes Howeveronly a limited number of them displayed a cross-neutralizingactivity against DENV1 and 2 and to a lesser extent againstDENV3 and 4 In particular the authors calculated thatamong them the 1A5 mAb that was further humanizedneutralized DENV1ndash4 at a PRNT50 titer of 048 095 32and 43 120583gmL respectively Interestingly the humanized 1A5was also tested for binding and neutralization against theWNVDENV4 chimera JEV strain SA14-14-2 and Langatvirus (LGTV) strain TP 21 giving a PRNT50 titer of 38 21and 28120583gmL respectively Moreover the authors calculatedthat when administered in a dose of 2mg per kg of bodyweight 1A5 would give a serum titer of approximately 40and 20 50 reduction in plaque reduction neutralization test(PRNT50) against DENV1 and DENV2 respectively [76]Furthermore epitope mapping of 1A5 mAb localized to G106within the Flavivirus-conserved fusion loop inDII of DENV2E protein [51]
244 Anti-DENV Human mAbs As anticipated attemptsto isolate neutralizing human mAbs have been morechallenging due to the restricted elicited Ab-repertoire whichrecognizes DIII of E glycoprotein in naıve as well as ininfected patients [77 78] However several groups reportedthe isolation of neutralizing mAbs from infected subjects Inthis regard Setthapramote et al isolated a total of 136 humanhybridoma clones producing specific mAbs against DENVobtained using PBMCs from nine blood samples from fouracute-phase patients secondarily infected with DENV2 andfive convalescent-phase patients Interestingly the authorsfound that most of the acute-phase mAb clones were cross-reactive with all four DENV serotypes with most of themrecognizing the E protein and endowing of neutralizingactivity against all DENV serotypes compared to thosederived from convalescent-phase patients In particularfrom the acute-phase PBMCs 818 were anti-E 66 wereanti-prM and 33 were anti-NS1 while 133 anti-E 133
anti-prM and 533 anti-NS1 mAbs clones were obtainedfrom convalescent-phase PBMCs [79] Previous studiesconfirmed the data obtained by Setthapramote et al onconvalescent-phase patients but this group firstly reportedthe efficient preparation of human mAbs with strongneutralizing activity titers against all four DENV serotypesusing PBMCs from acute-phase patients secondarily infectedwith DENV [80 81]
The group of de Alwis through immunoglobulin deple-tion studies reported that a substantial fraction of DENV-reactive Abs in human immune sera including type-specificneutralizing Abs bound to the intact virion but not torecombinant E proteinThe authors confirmed these observa-tions also isolating human neutralizing mAbs and proposedthat humans produce Abs that neutralize DENV infectionby binding a complex quaternary structure epitope that isexpressed only when E proteins are assembled on a virusparticle [48] Similar findings have been described for severalother viruses including WNV and HIV [82 83] Mappingstudies indicate that this epitope has a footprint that spansadjacent E protein dimers and includes residues at the hingebetween domains I and II of E protein [48]
Recently Costin et al isolated three broadly neutralizinganti-DENVhumanmAbs named 48AD11C and 16DThesemAbswere isolated from three different convalescent patientswith distinct histories of DENV infection All three mAbsrecognized the E glycoprotein with high affinity neutralizedall four serotypes of DENV but mediated ADE of infection inFc receptor-bearing cells at subneutralizing concentrationsMapping studies revealed that all three mAbs bind discontin-uous epitopes within the highly conserved fusion-loop regionof DII (contacting residues W101 L107 andor G109) [37]
25 Yellow Fever Virus (YFV) TheWHO estimates that thereare 200000 cases of yellow fever virus (YFV) infections and30000 related deaths every year especially in Sub-SaharanAfrica In fact different to themajority of human arbovirusesand similarly to DENV and CHIKV YFV has expanded itshost range to include humans as an amplifying host [1]Symptoms occurring from 3 to 6 days after infection withYFV consist mostly in fever chills anorexia lumbosacralpain nausea and vomiting This syndrome lasts about 3days in most cases and is sometimes followed by a one-daylasting period of remission Fever will reoccur together withdark hematemesismelena petechiae and other hemorrhagicsymptoms Convalescence is characterized by deep asthenialasting up to two weeks Rarely prognosis can be fatal Aneffective live-attenuated vaccine (17D) derived from the Asibistrain by serial passages in chicken embryos is available [84]
251 mAbs against YFV In 1983 Schlesinger et al produceda battery of mAbs after immunization of BALBc mice byinjection of 17D YFV vaccine strain [84] Among them thespecificity towards the E and NS1 proteins was describedeven though anti-NS1 Abs were not able to show neutralizingactivity The 13 IgG and 1 IgM anti-E protein mAbs wereclassified in five groups according to their specificity GroupA consisting of the only IgM produced in this experiment
10 BioMed Research International
could neutralize 17D YFV virus only group B 17D and Asibistrains only group C Asibi strain only group D could onlyneutralize Asibi but not 17D strain showing also reactivityagainst DENV2 Zika or Banzi viruses All of the four mAbsbelonging to this group were able to cross-react with Zikaandor Banzi viruses and two (4E11 and 5H3) neutralizedDENV2 virus with 4E11 neutralizing also Banzi virus The3E9 mAb the only component of the group E could notneutralize 17D nor Asibi strains
All of the IgG mAbs resulted able to protect bothprophylactically and therapeutically BALBc or CD-1 micefrom lethal intracerebral challenge with 17D-204 strain [85]The chimeric form of the group B mAb 2C9 IgG (2C9-cIgG) was able to provide AG129 mice a 72 survival whenadministered 24 hours before infection with 17D-204 strainIts murine form could provide the survival of the 95 ofthe mice Appreciable results were also obtained when bothmurine 2C9 and 2C9-cIgG were administered 24 hours post-infection with the survival of 70 and 20 of mice respec-tively Viral presence was not detected in survivingmice [60]
In 2005 Daffis and colleagues constructed two Ab-phage libraries by cloning the repertoire of YFV-infectedpatients Panning was then performed with YFV-17D virionsThe scFv-5A 7A and R3(27) showed a neutralizing activityspanning from50 to 100 inPRNTassays against bothYFV17D-204-WHO and Asibi strains In further tests reactivitywas observed against wild-type YFV strains of West Africagenotype I and II (Nigeria 1987 and Asibi strains resp)and EastCentral Africa (CAR 1986 Ethiopia 1961 strains) Aconcentration from 01 to 03120583gmL could yield a 90 plaquereduction Reactivity was also observed against the strainSenegal 1990 in a lesser extent Production of escape variantscould demonstrate that scFv-5A 7A and R3(27) epitopes arebuilt up by residues extensively separated in themonomers ofE glycoprotein that however result in closed proximity whenthe homodimeric form of E is constituted [61]
26 West Nile Virus (WNV) The West Nile virus (WNV) isan epidemic neurotropic virus estimated to be responsibleof about 36000 cases and 1500 deaths registered in theUnited States between 1992 and 2012 [86] WNV antigenicityallows its classification into the Japanese encephalitis virus(JEV) serocomplex Genomic analysis has revealed two maingenetic lineages of WNV lineage I viruses circulating in theUSA Europe the Middle East Africa India and Australiaand lineage II viruses isolated from Sub-Saharan Africa andMadagascar [1]
WNV was firstly identified in 1937 and is endemicin many countries of Africa Middle East and West AsiaHowever after 1990 frequent outbreaks of WNV infectionswere reported in Romania Israel and later in North Americaand across the USA in 1999 and recently in 2012 [87]
Most of WNV infected individuals do not develop symp-toms while about 20 develop a self-limiting illness calledWest Nile fever Acute symptoms include fever tirednessand swollen lymph glands but in a minority of cases alsoencephalitis with long-term deficits in cognitive functionand motor skills have been reported However in less than
1 of infected patients WNV having crossed the blood-brain barrier is responsible of neuroinvasive and potentiallylethal form of the disease In these cases degeneration andapoptosis upon infection of neurons and the consequentinflammatory response can occur Related symptoms includehigh fever coma muscle weakness and paralysis Immun-odepression and advanced age have been correlated with anaugment risk to develop a severe disease [87] There are nospecific treatment options or licensed vaccines for humans[88]
WNV is transmitted by infected mosquitos and initialreplication is thought to occur primarily in dendritic cells inthe skin which migrate to secondary lymphoid tissues wherethe replicating virus enters the circulation To date onlyefficacious WNV veterinarian vaccines have been licensedwhile there are no licensed vaccines for protection againstWNV in humans [88]
261 mAbs against WNV Broad-spectrum antivirals suchas type I IFN-120572 ribavirin mycophenolic acid in WNVinfection showed ineffective results in vivo despite in vitrothey showed some activity [89]
However experiments in murine models extrapolationof clinical data as well as passive administration of pooledimmune-120574-globulins (OmriGam) containing a significanttiter of neutralizing Abs before and after infection showedan important role of the humoral immune response incontrolling viremia and prevent viral dissemination Indeedthe development of a neutralizing mAb-based therapy seemsto be encouraging for a possible treatment of infected patients[90ndash92]
Similarly to what has been observed for DENV only anti-E Abs have been identified as neutralizing and protectiveAlso anti-NS1 Abs have shown a protective role howevertheir mechanism of protection has not yet been elucidated asthe NS1 protein is secreted from infected cells and not presenton the virions [93ndash95]
In this regard similarly to what has been describedfor DENV studies performed using a naıve human scFvlibrary for panning with purified WNV E protein supportthe hypothesis that no Abs against the neutralizing DIII canbe isolated However DIII specific Abs were isolated in asubsequent study using immunoglobulin libraries obtainedfrom threeWNV infected patients for biopanning on purifiedinactivated virus virus-like particles consisting of prM andE proteins or recombinant E glycoprotein Although theproportion of DIII-specific mAbs was low (8) comparedwith anti-DII mAbs (47) Two out of the four anti-DIIImAbs were potently neutralizing and protective in vivowhereas only three out of the 24 anti-DII were weaklyneutralizing in vitro and non-protective in vivo In particularresidues that are critical for neutralization lies on the regionsspanning amino acids 305ndash312 330ndash333 and 365 that arelocated on adjacent exposed loops ofDIIIHowever sequencealignment of E protein of different flaviviruses such as DENVand JEV revealed a considerable variation compared tothe whole E protein This observation further suggests thatdifferently from broadly cross-reactive anti-DII Abs anti-DIII neutralizing Abs are virus-type specific However of
BioMed Research International 11
these mAbs two (CR4374 and CR4353) protected mice fromlethal WNV challenge at 50 protective doses of 129 and357 120583gkg of body weight respectively [50]
Sanchez et al using three different immunization strate-gies (ie inactivated virus naked DNA and recombinantprotein) isolated ninemurinemAbsmost of which bound toconformation-dependent epitopes in DIII of the E protein Inparticular neutralizing mAbs named 8B10 11C2 10C5 and17C8 were obtained from mice immunized with inactivatedvirus alone or in combination with a DNA plasmid andbound to the same region of DIII with high affinity Incontrast mAbs obtained by immunization with a solubleversion of the E glycoprotein did not exhibit neutralizingactivity These non-neutralizing mAbs were cross-reactivewith several other flaviviruses including SLEV JEV YFVand Powassan virus confirming the conserved nature ofFlavivirus non-neutralizing epitopes [96]
Gould et al isolated 11 unique human single-chain vari-able region Ab fragments (scFvs) that bind the E protein ofWNVAmong them a humanmAb namedmAb11 expressedas a scFv-Fc fusion protein was further characterized Itrecognizes the fusion loop at the distal end of DII ofthe WNV E protein and cross-reacts with all four DENVserotypes and provides protection against DENV2 and 4as well as WNV [42] Moreover therapeutic studies of thismAb in WNV-infection model mice provided substantialprotection when administered after 5 days post-infectionInterestingly a neutralization escape variant of this mAbfailed to cause lethal encephalitis (at higher infectious doses)or induce the inflammatory responses associated with blood-brain barrier permeability in mice compared to the parentalWNV suggesting an important role for the fusion loop inviral pathogenesis [43]
Oliphant et al isolated an anti-DIII E protein mAbnamed E16 fromhybridomas obtained after immunization ofmice with recombinant WNV E protein which neutralizedall WNV strains with PRNT50 values of 4 to 18 ng andPRNT90 values of 53 to 297 ng One hundred microgramsof mAb protected greater than 90 of mice from lethalinfection and even a single 4120583g treatment of E16 on day 2 afterinfection prevented mortality Moreover humanization ofthis mAb confirmed as therapeutically effective in mice [44]Subsequent studies revealed that mAb E16 neutralization ismediated by engagement of four discontinuous segmentsof DIII including the amino-terminal region (amino acidresidues 302ndash309) and the three connecting loops BC (aminoacid residues 330ndash333) DE (amino acid residues 365ndash368)and FG (amino acid residues 389ndash391) Moreover no ADE ofinfection was detected when E16 mAb was used at saturatingconcentrations [45 47]
Furthermore results of a Phase I safety study of thehumanized E16 mAb (designated MGAWN1) have beenreported and suggested that doses of up to 30mgkgwerewelltolerated with few mild adverse events and would providean excess of virus neutralizing activity for 3-4 weeks aftertreatment However a case of anti-MGAWN1 Ab elicitationoccurred with the consequent increased rate of clearance andindeed impacting efficacy A Phase II safety and efficacy studyof MGAWN1 is ongoing [97] Furthermore the possibility of
preventing or treating WNV-induced memory deficits wasrecently investigated In this study hamsters were treatedintraperitoneally with 32mgkg ofMGAWN1mAb at 45 daysafter subcutaneously challenging with WNV InterestinglyMGAWN1 prevented mortality weight loss and improvedfood consumption of WNV-infected hamsters compared tocontrols [98]
Recently Lelli et al isolated six anti-E mAbs frominactivated-WNV immunized mice In particular three ofthem (3B2 3D6 and 4D3) neutralized lineage I and II WNVwith the first two recognizing the same epitopes located onthe distal lateral surface of DIII (critical amino acid residueK307) Conversely 4D3mAb recognized a novel neutralizingepitope on DII (critical residues S276 and T278) Indeedfurther protective and therapeutic studies are needed toascertain their neutralizing activity in vivo [99]
Finally to conclude like DENV infection Abs directedagainst the M protein are not protective and neutralizingMoreover in humans a skewed humoral immune responseagainst DII has been frequently observed and confirmedalso with the hybridoma technology [50] The isolation andelicitation of neutralizingAbs directed against the fusion loopand DIII of the E protein represent thus the most challengingand promising goal for the development of new effectivetherapeutic inhibitors and immunogens respectively
27 Tick-Borne Encephalitis Virus (TBEV) Tick-borne ence-phalitis virus (TBEV) is one of the most dangerous agentscausing human neuroinfections occurring mostly in Europeand Asia and with a potential fatal prognosis [100] TBEV isbelieved to cause 3500ndash10000 human cases of encephalitisin Europe per year with a high morbidity in Russia CzechRepublic Austria and Germany In particular between 1990and 2007 an average of 8755 cases of TBE was reported peryear in Europe and Russia Despite the fact that Russia isthe country with most infections registered annually CzechRepublic incidence is among the highest in Europe with 400ndash1000 clinical cases reported every year [101]
Three subtypes of TBEV are classified namely EuropeanSiberian and Far-Eastern sharing most of the genetic andantigenic features [102] In fact a high degree of antigenichomogeneity between different strains of TBEV has beendescribed [103]
Clinical manifestations vary among the subtypes butusually they start with a short febrile period of 7ndash14 daysafter the tick bite Fatigue headache and pain in the neckshoulders and lower back together with high fever andvomitingmay be present [100]Thesemanifestations are oftenfollowed by an asymptomatic phase lasting from 2 to 10days after remission from the fever with possible progres-sion to neurological disease Neurological symptoms includemeningitis encephalitis myelitis and radiculitis Mortalityoccurs in 1-2 of the European subtype-infected patientsbut fatal prognosis can occur in up to 20ndash40 of the Far-Eastern subtype-infected patients [104] Mortality rates ofthe Siberian subtype are similar to those observed for theEuropean [105]
Effective and safe vaccines against TBEV produced frominactivated virus have been developed and licensed for their
12 BioMed Research International
use in humans However an emergency therapy in theabsence of a mass immunization is needed as no effectivetreatments are yet available [106]
271 mAbs against TBEV Levanov et al described the chim-erization of two murine mAbs (13D6 and 10C2) directedagainst the DIII and DII respectively of the TBEV E glyco-protein The chimeric mAbs present binding characteristicssimilar to the parentalmAbsMoreover as the parentalmAbsonly the chimeric mAb 13D6 was able to neutralize TBEVinfectivity in vitro In particular neutralization studies withthe murine chimeric and scFv forms of mAb 13D6 were per-formed In particular murine 13D6 showed an IC50 titer of115 120583gmL in Focus Reduction Neutralization Tests (FRNT)against TBEV strain 205 and of 29 120583gmL in PRNT againstTBEV strain Softjin Chimeric 13D6 showed an IC50 titer of45 120583gmL in FRNT against TBEV strain 205 and of 19 120583gmLin PRNT against TBEV strain Softjin ScFv 13D6 showed anIC50 titer of 167 120583gmL in FRNT against TBEV strain 205and of 112 120583gmL in PRNT against TBEV strain Softjin [62]
28 Japanese Encephalitis Virus (JEV) Japanese encephalitisvirus (JEV) is a mosquito-borne flavivirus that over thepast few decades has caused several outbreaks throughoutChina Southeast Asia Australia and Papua New Guineawith a prevalence recently estimated to be of about 70000casesyear [107ndash109] A 40 mortality was recorded in someof the JEV-affected areas Moreover many survivors facesome neurological problems and complications [108] Since1995 the disease has also emerged in Non-Asian regions suchas Northern Australia [110 111] The situation in SoutheastAsia however is further complicated by the overlappingepidemics of JEV and DENV as well as sporadic casesof WNV infections detected in some of the affected areasparticularly in India [112]
Effective vaccines both live-attenuated and inactivatedstrains of JEV have been developed and licensed in majorcountries However as for the other arthropod-borne Fla-vivirus members no specific antiviral drugs are currentlyavailable [4]
281mAbs against JEV Aspreviously described for the otherFlavivirusmembers many groups isolated and characterizedanti-JEVmAbs showing different neutralizing and protectiveproperties In this regard Gupta et al used combinations ofanti-E JEV mAbs (Hs1-4) in mice protection experimentsIn particular they found that the singularly mAb protectionranged from 45 to 65 when 100120583g of mAb were adminis-tered while equimolar combinations of two or three mAbsgave 85ndash90 or 100 protection respectively [113] In similarexperiments Lee et al demonstrated that a Flavivirus anti-NS1 mAb named 16NS1 cross-reacted with JEV as well asWNV and exhibited protective activity against WNV as wellas a lethal JEV infection However no neutralizing activitywas observed using this mAb against both WNV and JEV inin vitro experiments suggesting the participation of otherAb-mediatedmechanisms in vivo In particular 95ofmicewere
protected when 500120583g of mAb were administered intraperi-toneally and concomitantly to intramuscular injection of JEV
Overlapping peptide mapping analysis combined withsite-specific mutations identified the 116KAWGKSILFA125region and critical amino acid residues W118 and I122 as16NS1 mAb epitope highly conserved in WNV and JEVstrains [114]
Arakawa et al isolated from a combinatorial human Fablibrary constructed from peripheral blood lymphocytesobtained from JEV hyperimmune volunteers Among 188randomly selected clones FabTJE12B02 showed the best 50focus reduction endpoint at the concentration of 502 120583gmLagainst the JEV strain Nakayama [55]
Goncalvez et al isolated three mAbs named Fabs A3B2 and E3 by repertoire cloning from chimpanzees initiallyimmunized with inactivated JE-VAX and then boosted withattenuated JEV SA14-14-2 In particular these mAbs reactedwith epitopes in three different E domains in DI (amino acidresidue K179) in DII (I126) and in DIII (G132) for Fabs A3B2 and E3 respectively Moreover these Fabs as well as thederived humanized counterpart mAbs exhibited high neu-tralizing activities against a broad spectrum of JEV genotypestrains Moreover these mAbs exhibited a 50 protectivedose of 084 120583g (B2) 58120583g (A3) and 247120583g (E3) in mousemodels Finally administration of 200 120583gmouse of mAb B2one day after otherwise lethal JEV infection protected 50of mice and significantly prolonged the average survival timecompared to that of mice in the unprotected group [54]
29 St Louis Encephalitis Virus (SLEV) St Louis encephalitisvirus (SLEV) was first discovered as the mosquito-borneagent responsible for over 1000 cases of encephalitis duringa 1933 summer outbreak in St Louis (Missouri) and nowis a reemerging human pathogen widely distributed in theAmerican continent causing several human encephalitisoutbreaks over the last 80 years Additional epidemics haveindeed occurred from 1964 to 2006 in the Americas rangingfrom the US to Argentina and Brazil As a member of theFlavivirus genus the SLEVE glycoprotein ectodomain is 68identical to serocomplex-related JEVEbut only 46 and 40identical to those ofDENV2 andTBEV viruses fromdifferentserocomplexes [115]
291 mAbs against SLEV In 1983 Roehrig et al isolatedtwenty-one hybridomas producing murine mAbs specificfor the E glycoprotein of SLEV strain MSI-7 Serologicreactivities were initially determined by cross-reactivity indi-rect immunofluorescence assays using 22 strains of SLEVand 8 other related flaviviruses Four groups demonstratingtype- subcomplex- supercomplex- and group-specific reac-tivity patterns were identified Analysis of hemagglutination-inhibition and virus neutralization subdivided the cross-reactivity groups into eight epitopes (E-1a b c d E-2 E-3 and E-4a b) one of them following localized in DII (4b)[52 53]
Moreover among the previously described isolated anti-WNV mAbs by Gould et al the chimeric scFv-Fc 79 effec-tively neutralized also SLEV resulting in gt80 of PRNT80when used at 5 120583gmL [42]
BioMed Research International 13
3 Alphaviruses
Major human arboviruses in the Togaviridae family Chikun-gunya (CHIKV) and Venezuelan equine encephalomyelitis(VEEV) viruses belong to the Alphavirus genus which iscomposed of viruses with an icosahedral nucleocapsid sur-rounded by a lipid envelope and glycoprotein spikes Thestructural proteins of alphaviruses arise through co- andpost-translational processing of a polyprotein encoded by asingle positive-stranded RNAproducing the capsid (C) PE26 K and E1 The PE2 glycoprotein is a precursor containingthe E3 glycoprotein fused to the amino terminus of the E2envelope glycoprotein The PE2 glycoprotein is followed by6K a small membrane-associated protein and E1 the secondpolypeptide component of glycoprotein spikes
Trimerized heterodimers of the E1 and E2 viral glyco-proteins form the surface spikes and contain determinantsof viral tropism and virulence The E3 glycoprotein acts as asignal for transport of PE2 across themembranes of the roughendoplasmic reticulum and may promote the formation andintracellular transport of E1-PE2 heterodimers to the cellsurface During transport to the cell surface PE2 undergoesa maturational cleavage event by a furin-like protease toproduce E2 and E3 [116]
The E2 glycoprotein promotes specificity of virus bindingto the host cell surface and is a target of the humoral immuneresponse The E1 glycoprotein mediates fusion of the virionenvelope with the membranes of acidified endosomes allow-ing release of the nucleocapsid into the cytoplasm and theonset of viral replicationAlphavirus E1 shares no appreciablesequence identity to Flavivirus E protein Despite differencesin their amino acid sequences and arrangements on the viralparticle the structures of E and E1 are remarkably similarIndeed the conservation of three domains has been welldocumented in crystal structures of flavivirus E ectodomainsfrom TBEV DENV JEV and WNV as well as Alphavirus E1from Semliki Forest virus (SFV) and CHIKV Abs to the E1glycoprotein do not typically neutralize virus infectivity invitro but can protect against lethal challenge in animals
By analogy with the prM protein of dengue Flavivirusfurin cleavage of PE2 may begin after transit of an acidiclate component of the Golgi body where E3 is thought tosuppress the acid pH-triggered activation of glycoprotein E1fusion capability Protective anti-E3 mouse mAbs have alsobeen described [117]
31 Chikungunya Virus (CHIKV) The chikungunya virus(CHIKV) belongs to the Semliki Forest clade and was firstlyisolated in 1953 in Tanzania during an epidemic outbreakfollowing occurred also in Asia and Africa In the last fivedecades several CHIKV outbreaks have been described bothin Africa and Asia separated by gaps lasting from two totwenty years In 2005-2006 about 300000 cases out of785000 inhabitants were reported in La Reunion island witha fatal prognosis for 237 of the patients [117] Neither Europenor the Americas have had outbreaks of CHIKV so far exceptfor imported isolated cases Three genotypes of CHIKV aredescribed Asian EastCentralSouth African (ECSA) andWest African with an amino acid identity spanning from
952 to 998 Recent epidemics in Africa and Indiansubcontinent were caused by strains belonging to the ECSAgenotype Transmitted by Aedes mosquitoes CHIKV ismaintained in the human population by human-mosquito-human transmission [1] The disease is characterized bydengue-like symptoms such as chills high fever headacheand persistent myalgia and a further incapacitating arthralgia(from which the name chikungunya) which affects 40 ofinfected subjects However prognosis is rarely fatal [116]
There is currently no commercial vaccine and antiviraltreatment forCHIKV although some candidate vaccines havebeen tested in humans In this regard in 2000 US Armyperformed a Phase II clinical trial testing a live-attenuatedCHIKV vaccine (TSI-GSD-218) derived from a 1962 strain(15561) of an outbreak in Thailand Out of 58 every patientdeveloped neutralizingAbs and 5 lamentedmild tomoderatejoint pain [117] A phase III trial of this candidate vaccineis ongoing Furthermore a new formulation using virus-likeparticles was able to induce neutralizing Abs in macaquesagainst different CHIKV strains [117 118] Indeed it has beendescribed that infection seems to elicit long-lasting protectiveimmunity and cross-protection among CHIKV and otheralphaviruses
CHIKV entry into host cell is demonstrated to be medi-ated by envelope glycoproteins E1 and E2 which allowvirus fusion to cell membrane in low pH conditions andrecognition of an unknown cellular receptor respectively[116 119ndash121] Even though the genome replication relies onan error-prone RNA-dependent RNA-polymerasis CHIKVstrains S27 and 05115 Reunion showed in E1 and E2 alow amino acid variation of 068 and 33 respectivelyand such a low variability may be needed for an effectivereplication in two phylogenetically distant hosts [116 122]Recently the crystal structure of E1 and E2 heterodimerhas been resolved both at low pH and natural conditionsshedding light on E1 and E2 ectodomainsrsquo structure Inparticular similarly to flaviviruses E protein E1 ectodomainis made of the N-terminal DI the DII containing the fusionloop and the DIII at the C-terminal On the other hand E2ectodomain comprises theN-terminal domainA the domainB supposed to interact with the host cellrsquos unknown receptorand domain C at the C-terminal E1 DIII and E2 domain Care located close to the viral membrane [123 124]
311 mAbs against CHIKV Warter et al described 5F10 and8B10 two human mAbs which strongly neutralized severalCHIKV isolates in vitro without cross-reactivity againstother alphaviruses but to Onyong-nyong virus [56] Mixedpreparation of 5F10 and 8F10 did not show neither synergisticnor addictive effect in vitro and studies upon escape mutantsdemonstrated that 5F10 mAb binds at the tip of the E2domain B while 8B10 recognizes residues close to E1 fusionloop and amino acids within E2 domain A which form atransitional epitope under low pH conditions The authorsspeculated that 5F10 and 8B10 may inhibit CHIKV entry andfusion to the cell membrane respectively It is worth notingthat the previously mentioned escape mutants displayedmutations associated with reduced viral fitness in vitro evenafter 13 neutralizationamplification rounds In the same
14 BioMed Research International
study cell-to-cell transmission was firstly demonstrated as anescapemechanism enhanced by the E2mutation R82G [122]
In further in vivo studies 5F10 and 8B10 significantlydelayed CHIKV-caused death of AGR129 mice both in pro-phylactic and therapeutic tests Interestingly in therapeutictreatment these mAbs showed a synergistic effect whenadministered in combination with the total amount of mAbsinjected being the same of the single-mAbs Possibly thesetreatment did not allow themice to survive CHIKV challengedue to Ig clearing and 6ndash10 days half-life usually described forhuman mAbs [125]
Recently Pal et al cloned thirty-six murine mAbs ableto neutralize the ECSA La Reunion 2006 OPY-1 strain ofCHIKV (CHIKV-LR) the majority of which also neutralizeinfection of other strains corresponding to the Asian andWest African genotypes In particularmAbCHK-152 showedthe highest and broadest neutralizing activity with FRNTvalues indicating an IC50 value of 1 to 3 ngmL dependingon the viral strain used Among the thirty-six mAbs four(CHK-102 CHK-152 and CHK-166 CHK-263) could pre-vent lethality of immunodeficient Ifnarminusminus C57BL6 micewhen administered one day before exposition with CHK-152 and CHK-263 showing the ability to protect mice at thelowest dose (10 120583g) Therapeutic studies were performed onthese mAbs administering a single dose of 100 120583g 24 hoursafter infection The highest activity was described for CHK-166 able to protect 63 of the mice Testing the combinedactivity of the mAbs in therapeutic studies administration ofCHK-166 plus CHK-152 in a dose of 250120583g each turned out tobe themost effective protecting 71 of the IfnarminusminusC57BL6mice 60 hours after infection [57]
32 Venezuelan Equine Encephalomyelitis Virus (VEEV)Venezuelan equine encephalomyelitis virus (VEEV) is main-tained in a natural transmission cycle between mosquitoesand small rodents The first documented outbreaks occurredin the 1930s and several epidemics have been reported so farin Latin-American countries such as Venezuela ColombiaPeru Ecuador Bolivia Honduras Mexico and Panama withhundreds of thousands of human cases being reported anda case-fatality rate up to 1 during the 1969 Ecuador out-break Clinical manifestations of VEE are indistinguishablefrom Dengue [126] Six serogroups (IndashVI) are currentlyrecognized within the VEEV complex VEEV caused humanand equine outbreaks in the Americas for nearly a centuryEquine epizootics have high mortality (38ndash83) leading toa high viremia followed by a lethal encephalitis and often tohuman epidemics involving thousands of cases and hundredsof deaths Indeed VEEV is infectious for humans by theairborne route and has been responsible for a number oflaboratory infections In humans the disease is usually self-limiting with a febrile illness with 1ndash4 of cases progressingto severe encephalitis It is a hazard to laboratory workershas been developed as a biological weapon and is a potentialbioterrorist agent [127] There are no antiviral drugs andvaccines licensed for the treatment of VEEV infection inhumans Indeed although experimental live-attenuated vac-cines have been developed (eg TC-83) with good levels ofprotection in equine and mice in humans they may fail to
give protection in the majority of cases [128] Consideringthe protective role exerted by humoral immune response inanimal models it could be concluded that treatment withspecific IgG may have a beneficial antiviral effect in humanairborne infections with pathogenic strains of VEEV
321 mAbs against VEEV Several works support that mAbsmay protect against airborne VEEV as well as after airborneexposure toVEEVAb reactivity to both surface glycoproteins(E1 and E2) is associated with neutralization of the virusin vitro and passive protection against virus challenge Aseries of anti-E1 and anti-E2 mouse and humanized mAbshave been described recently Among them Phillpotts et alexamined two anti-E2 murine mAbs named 1A4A-1 and1A3A-9 both having potent protective activity against subcu-taneous VEEV challenge in mice Both mAbs had a similarhalf-life (58 and 100 days) in mouse serum after a singleintraperitoneal dose suggesting that mAbs delivered by thisroute at or around the time of VEEV infection persists athigh levels in the blood as well as secretions in respiratorytransudation throughout the clinical course of the disease Inparticular both mAbs administered 24 h prior to airbornechallenges had a substantial protective effect (90ndash100)Treatment of mice 2 or 24 h after airborne infection with asingle intraperitoneal dose of 100 120583g of 1A3A-9 mAb led toapproximately 50 survival There was no beneficial effectwhen mAb treatment was delayed to 72 h Moreover therewas evidence of synergy in vitro in PRNT between 1A3A-9and 1A4A-1 as has been demonstrated for other virusesand mAb pairs However no synergy was found in mouseprotection when mAbs were delivered intraperitoneally as amixture in equal parts to animals challenged 24 h previouslywith VEEV [129]
In a subsequent study Phillpotts et al reported two otherprotective mAbs The 3B2A-9 mAb protected against allthe serogroup I strains while 1A3B-7 mAb protected wellagainst challenge with all of the viruses tested Both 3B2A-9 and 1A3B-7 protected against airborne exposure to theIAB serogroup virus strainTrinidad donkey (TrD)Howeverthere was no evidence of synergistic protection when thesemAbs were combined in equal proportions An intraperi-toneal dose of 10 120583g was sufficient to protect 50 of themice with either mAb [130] According to other data themechanism of protection did not appear to depend uponneutralization [131] Similar results were obtained by OrsquoBrienet al which described a non-neutralizing mAb (IgG2a)named CUF37-2a able to protect 50 of mice from asubcutaneous VEEV challenge when a dose of 915 120583g of mAbwas administered 24 h prior to challenge [132]
In a following study the 1A3B-7 mAb was humanized(and following reported as Hu1A3B-7) maintaining the samefeatures of its murine counterpart In particular evaluation ofin vitro studies indicated that Hu1A3B-7 retained both broadspecificity and neutralizing activity Furthermore in vivoexperiments showed that Hu1A3B-7 successfully protectedmice against lethal subcutaneous and aerosol challenges withVEEV strain TrD Moreover the effectiveness of the human-ization process was determined by assessing proliferationresponses in human T-cells to peptides derived from the
BioMed Research International 15
murine and humanized versions of the VH and VL domainsThis analysis showed that the number of human T-cellepitopes within the humanized Ab had been substantiallyreduced indicating that Hu1A3B-7 may have low immuno-genicity in vivo [133]
Similarly Hunt et al described a humanizedmurinemAbderived from the 3B4C-4 mAb recognizing the E2c epitope(amino acid residues 182ndash209) and following named Hy4-26C showing neutralizing activity similar to the murinemAb Moreover it was protective (70ndash100 of survival) at100 ng dose in mice animal model intraperitoneal inoculatedand at 500120583g for intranasal challenge (80) Therapeuticstudies in mice revealed that Hy4-26C was able to cure miceup to 24 h following infection at 10 120583g on the contrary themouse mAb when given 1 hour after virus challenge [63]
In another work Hunt et al isolated two neutralizinghumanized Fabs F5 and L1A7 employing a blocking strategy[134 135] The anti-E2 specific F5 IgG had a 70 PRNTendpoint of 10 ngmL equivalent to that described for themost effective neutralizing anti-VEEVE2mAbs In particularthe E1-specific hFab L1A7 had a PRNT endpoint of 3120583gmL300-fold lower than F5
Further studies revealed that F5 epitope is located inthe 115ndash199 amino acid region Moreover F5 IgG had potentability to protect mice from infection by either route whenadministered 24 h before exposure however mice treated24 h and 48 h after aerosol exposure developed central ner-vous system infections but exhibited no clinical signs ofdisease [135]
Hu et al reported that passive immunization with thehumanized chimericmousemAbhu1A4A1IgG1-2A inmice at50 120583g 24 h before or after virulent VEEV challenge providedcomplete protection indicating that hu1A4A1IgG1-2A haspotent prophylactic and therapeutic effects against VEEVinfection [136]
Anti-VEEV mAbs isolated from a non-human primategene library has also been reported In particular thehumanized chimeric scFv-Fc ToR67-3B4 recognized viableas well as formalin and 120573-propiolactone inactivated virusparticles It detected specifically the viral E1 envelope proteinof VEEV but did not react with reduced viral glycopro-tein preparations suggesting that recognition depends uponconformational epitopes The recombinant Ab was able todetect multiple VEEV subtypes and displayed only marginalcross-reactivity to otherAlphavirus species except for Easternequine encephalitis virus (EEEV) In addition the scFv-Fcfusion described here might be of therapeutic use since itsuccessfully inactivated VEEV in a murine disease modelIn particular when the recombinant Ab was administered 6hours after challenge 80 to 100 of mice survived lethalVEEV IAB or IE infection Forty to sixty percent of micesurvived when scFv-Fc ToR67-3B4 was applied 6 hours afterchallenge with VEEV subtypes II and former IIIA [137]
4 Bunyaviridae
TheBunyaviridae family contains human arboviruses belong-ing to the Orthobunyavirus Phlebovirus and Nairovirus
genera Bunyaviridae includes enveloped viruses with a frag-mented single-stranded RNA genome of negative polarityTheir tripartite genome consists of a small (S) a medium(M) and a large (L) fragment and the envelope glycoproteinsGn and Gc that are cleaved out of a polyprotein synthesizedby the M fragment These glycoproteins have been provedto mediate the formation of the virus particle to play a rolein the interaction with cell surface receptors to mediate theentry of the virus into cells and to serve as targets for themajority of neutralizing Abs described so far
Orthobunyavirus transmitted through mosquitoes ormidges vectors are divided in 18 serogroups based on cross-titrations in haemagglutination inhibition assays and neutral-ization assays and correlating with main vector preferencesHowever the most clinically important viruses belong onlyto two serogroups the California encephalitis and the Simbuserogroups
Crimean-Congo hemorrhagic fever virus (CCHFV) isconsidered the only one of clinical relevance within theNairovirus genus which uses tick as main vector and is divid-ed in seven serogroups while Toscana virus and Rift Valleyfever virus (RVFV) which are transmitted by sandflies andmosquitoes respectively belong to the Phlebovirus genus[138]
MAbs against theNairovirusCCHFV and the PhlebovirusRVFV have been described
41 Crimean-Congo Hemorrhagic Fever Virus (CCHFV)Crimean-Congo Hemorrhagic Fever Virus (CCHFV) dis-tribution covers the greatest geographic range of any tick-borne virus known as viral isolation andor disease hasbeen reported from more than 30 countries in SoutheasternEurope Africa Asia and Middle East [139 140] Namelyfrom 1953 to 2010 about 6000 human cases were reportedin Southeastern Europe CCHFV causes sporadic outbreakswith mortality rates ranging from 10 to 80 [141 142] Asignificant variation in the time of incubation has beendescribed [139] Prehemorrhagic symptoms include highfever chills headache photophobia and back and abdominalpains Among other symptoms neuropsychiatric changeshave been reported in some patients In severe cases 3ndash6 daysafter the onset of the disease hemorrhagic symptoms occurwith petechiae ecchymosis bleeding in the form of melenahematemesis epistaxis [139]
In comparisonwith the nucleocapsid proteins inCCHFVGn and Gc show a higher degree of antigenic variabilityprobably due to their exposition to the host immune systemhypothesis corroborated in the description by Hewson et alin 2004 of four M segment phylogenic groups namely M1M2 M3 and M4 [141 142]
411 mAbs against CCHFV Blackburn et al firstly describedin 1987 murine anti-CCHFV mAbs able to recognize nucle-ocapsid proteins but no neutralizing activity has beendescribed thus far [143]
On the other hand Bertolotti-Ciarlet et al produced apanel of murine mAbs recognizing conformational epitopes
16 BioMed Research International
within theGn andGc glycoproteins expressed by the CCHFVstrain IbAr10200 The effectiveness of the mAbs was studiedby performing PRNT80 and none of the anti-Gn mAbsshowed neutralizing activity although many of the anti-GcmAbs neutralized IbAr10200 strain in vitro with clones 8A15E3 12A9 6C2 and 9H3 showing activity at a gt5120 dilutionMAbs 11E7 and 30F7 exhibited neutralizing activity at 2560dilution A suckling mice protection test was thereforeperformed for these mAbs The previously mentionedanti-Gc Abs were capable to protect mice to an appreciabledegree when applied 24 hours before and in a weakermanner 24 hours after virus challenge Many of the anti-GnmAbs were able to confer significant protection to IbAr10200both 24 hours before and after virus administration Therelevant effectiveness of the anti-Gn mAbs 6B12 10E1113G8 and 10G4 suggests that these mAbs may possess someneutralizing activity due to Ab-based effector mechanisms(eg ADCC) [58]
In further studies the same group characterized thebroadly cross-reactive neutralizing activity of the previouslydescribed murine mAb 11E7 able to bind conformationalepitopes within the C-terminal region of the Gn glycoproteinof all CCHFVM groups [58 141]
42 Rift Valley Fever Virus (RVFV) Similarly to CCHFV RiftValley Fever Virus (RVFV) causes sporadic outbreaks Thelargest epidemic occurred in Egypt in 1977 with an estimated200000 infections 18000 patients manifesting symptomsand 600 fatal prognoses [144] Symptoms include hemor-rhagic episodes fever encephalitis and blindness [145]
The major target of the humoral immune response is theRVFV glycoproteins Gc (G1) and Gn (G2) ranging fromnucleotides 480ndash2090 and 2091ndash3614 respectively [144]
421 mAbs against RVFV Within G2 glycoprotein are des-cribed three neutralizing antigenic sites namely epitope Inucleotides 792ndash893 (amino acid residues 258ndash291) epitopeII nucleotides 1164ndash1196 (amino acid residues 382ndash392) andepitope IV nucleotides 858ndash917 (amino acid residues 280ndash299) [59 145]
Despite the high degree of variability observed inCCHFVglycoproteins RVFV G2 presented an unexpected high con-servation among 22 isolates [59] In the same study onlythe LUNYO and 900060 isolates were resistant to in vitroneutralization by murine mAb 4D4 (recognizing epitope II)All other isolates were neutralized with PRNT80 titers from10240 togt81920On the other hand only SNS isolate showedreduced sensitivity to 4ndash39-CC (epitope IV) All other isolateswere neutralized with PRNT80 titers from 20480 to gt81920Protection studies on mice would be of interest for thedevelopment of a passive immunization prophylaxis [59]
Protection studies from lethal South African RVFVAN1830 strain infection were successfully performed on abattery of anti-G1 and anti-G2 mAbs produced by Besselaarand Blackburn with the strongly neutralizing anti-G1 mAb3E5 and anti-G2mAb 9C4 being the most effective [146]The9C4 antigenic area maps within G2 epitope I and a broadlyneutralizing activity may be investigated
5 Conclusions
Arbovirus infections have acquired increasing interest giventhe augmented globalization and tourism movement all overthe world that could be at the basis of epidemic eventsFurthermore the same vectors can sometimes transmit sev-eral arboviruses concomitantly complicating the diagnosis aswell as the therapy For example confusing mixed epidemicshave occasionally been described such as YFV plus CHIKVDENVplus CHIKV ormore recentlyPlasmodium falciparummalaria plus DENV1 and CHIKV on Madagascarrsquos east coast[1]
In the last decade several efforts have been employed inthe development of effective immunogens and therapeuticsgiving the current absence of specific antiviral drugs as wellas effective vaccines for themost diffused arbovirus infections(ie DENV andWNV)
However at the moment mosquito control is the bestavailable method for preventing arbovirus infections Thusto control the emerging public health problem of arbovirusinfections new antiviral therapeutic strategies that providepotent and broadly cross-protective immunity (especially forflavivirus infections) are an urgent globally medical need
As described in this review the majority of highlyneutralizing mAbs are of murine origin and indeed theirutility in the treatment of these infections is limited In facttheir employment can elicit an immunogenic response (iehuman anti-mouse Abs HAMA) the therapeutic efficacycould be reduced by a relatively faster clearance in humans(compared to human Abs) potentially exacerbated by theHAMA response Finally murine Abs exhibit relatively weakeffector functions (eg ADCC) compared to human AbsIndeed chimerization humanization or better the isolationand characterization of fully human broadly neutralizingmAbs would be the best choice for their possible use in afuture mAb-based therapy against arbovirus infections [147]
Finally possible ADE of infection mechanisms should beevaluated before considering a mAb as a possible candidatetherapeutic in a post-exposure setting as well as in the devel-opment of new Ab-eliciting immunogens
Conflict of Interests
The authors declare no conflict of interests in particular withOmriGam and any other cited Ab-based preparation men-tioned in the paper
References
[1] N Cleton M Koopmans J Reimerink G J Godeke and CReusken ldquoCome fly with me review of clinically importantarboviruses for global travelersrdquo Journal of Clinical Virology vol55 no 3 pp 191ndash203 2012
[2] F X Heinz H Holzmann A Essl and M Kundi ldquoField effec-tiveness of vaccination against tick-borne encephalitisrdquoVaccinevol 25 no 43 pp 7559ndash7567 2007
[3] A D Barrett and D E Teuwen ldquoYellow fever vaccinemdashhowdoes it work and why do rare cases of serious adverse events
BioMed Research International 17
take placerdquo Current Opinion in Immunology vol 21 no 3 pp308ndash313 2009
[4] S B Halstead and S J Thomas ldquoNew vaccines for Japaneseencephalitisrdquo Current Infectious Disease Reports vol 12 no 3pp 174ndash180 2010
[5] R Burioni M Perotti N Mancini and M Clementi ldquoPer-spectives for the utilization of neutralizing human monoclonalantibodies as anti-HCV drugsrdquo Journal of Hepatology vol 49no 2 pp 299ndash300 2008
[6] R A Diotti G A Sautto L Solforosi NManciniM Clementiand R Burioni ldquoNeutralization activity and kinetics of twobroad-range human monoclonal IgG1 derived from recombi-nant Fab fragments and directed against Hepatitis C virus E2glycoproteinrdquo New Microbiologica vol 35 no 4 pp 475ndash4792012
[7] R Burioni F Canducci N Mancini et al ldquoMolecular cloningof the first humanmonoclona antibodies neutralizing with highpotency Swine-origin Influenza A pandemic virus (S-OIV)rdquoNew Microbiologica vol 32 no 4 pp 319ndash324 2009
[8] P K Chandra S Hazari B Poat et al ldquoIntracytoplasmic stableexpression of IgG1 antibody targeting NS3 helicase inhibitsreplication of highly efficient hepatitis C Virus 2a clonerdquoVirology Journal vol 7 article 118 2010
[9] N Clementi D de Marco N Mancini et al ldquoA human mono-clonal antibody with neutralizing activity against highly diver-gent influenza subtypesrdquo PLoS ONE vol 6 no 12 Article IDe28001 2011
[10] D de Marco N Clementi N Mancini et al ldquoA non-VH1-69 heterosubtypic neutralizing human monoclonal antibodyprotects mice against H1N1 and H5N1 virusesrdquo PLoS ONE vol7 no 4 Article ID e34415 2012
[11] L Solforosi N Mancini F Canducci et al ldquoA phage displayvector optimized for the generation of human antibody com-binatorial libraries and the molecular cloning of monoclonalantibody fragmentsrdquoNewMicrobiologica vol 35 no 3 pp 289ndash294 2012
[12] E V Koonin and V V Dolja ldquoEvolution and taxonomy ofpositive-strand RNA viruses implications of comparative anal-ysis of amino acid sequencesrdquo Critical Reviews in Biochemistryand Molecular Biology vol 28 no 5 pp 375ndash430 1993
[13] J S Mackenzie D J Gubler and L R Petersen ldquoEmergingflaviviruses the spread and resurgence of Japanese encephalitisWest Nile and dengue virusesrdquo Nature Medicine vol 10 no 12pp S98ndashS109 2004
[14] T J SmithW E Brandt J L Swanson J MMcCown and E LBuescher ldquoPhysical and biological properties of dengue-2 virusand associated antigensrdquo Journal of Virology vol 5 no 4 pp524ndash532 1970
[15] J M Smit B Moesker I Rodenhuis-Zybert and J WilschutldquoFlavivirus cell entry and membrane fusionrdquo Viruses vol 3 no2 pp 160ndash171 2011
[16] R J Kuhn W Zhang M G Rossmann et al ldquoStructure ofdengue virus implications for flavivirus organization matura-tion and fusionrdquo Cell vol 108 no 5 pp 717ndash725 2002
[17] K Stiasny J H Aberle M Keller B Grubeck-Loebenstein andF X Heinz ldquoAge affects quantity but not quality of antibodyresponses after vaccination with an inactivated flavivirus vac-cine against tick-borne encephalitisrdquo PLoS ONE vol 7 no 3Article ID e34145 2012
[18] LThomasOVerlaetenACabie et al ldquoInfluence of the dengueserotype previous dengue infection and plasma viral load
on clinical presentation and outcome during a dengue-2 anddengue-4 co-epidemicrdquo American Journal of Tropical Medicineand Hygiene vol 78 no 6 pp 990ndash998 2008
[19] Y Zhang J Corver P R Chipman et al ldquoStructures of imma-ture flavivirus particlesrdquo EMBO Journal vol 22 no 11 pp2604ndash2613 2003
[20] A K I Falconar ldquoIdentification of an epitope on the denguevirus membrane (M) protein defined by cross-protective mon-oclonal antibodies design of an improved epitope sequencebased on common determinants present in both envelope (EandM) proteinsrdquo Archives of Virology vol 144 no 12 pp 2313ndash2330 1999
[21] K-J Huang Y-C Yang Y-S Lin et al ldquoThe dual-specific bind-ing of dengue virus and target cells for the antibody-dependentenhancement of dengue virus infectionrdquo Journal of Immunol-ogy vol 176 no 5 pp 2825ndash2832 2006
[22] A K I Falconar ldquoAntibody responses are generated to immun-odominant ELKKLE-typemotifs on the nonstructural-1 glyco-protein during live dengue virus infections inmice and humansimplications for diagnosis pathogenesis and vaccine designrdquoClinical and Vaccine Immunology vol 14 no 5 pp 493ndash5042007
[23] C W Mandl F Guirakhoo H Holzmann F X Heinz and CKunz ldquoAntigenic structure of the flavivirus envelope protein Eat the molecular level using the tick-borne encephalitis virus asa modelrdquo Journal of Virology vol 63 no 2 pp 564ndash571 1989
[24] F A Rey F X Heinz C Mandl C Kunz and S C HarrisonldquoThe envelope glycoprotein from tick-borne encephalitis virusat 2 A resolutionrdquo Nature vol 375 no 6529 pp 291ndash298 1995
[25] S L Allison K Stiasny K Stadler CWMandl and F XHeinzldquoMapping of functional elements in the stem-anchor regionof tick-borne encephalitis virus envelope protein Erdquo Journal ofVirology vol 73 no 7 pp 5605ndash5612 1999
[26] D E Klein J L Choi and S C Harrison ldquoStructure of a denguevirus envelope protein late-stage fusion intermediaterdquo Journal ofVirology vol 87 no 4 pp 2287ndash2293 2013
[27] S L Hanna T C Pierson M D Sanchez A A Ahmed M MMurtadha and R W Doms ldquoN-linked glycosylation of WestNile virus envelope proteins influences particle assembly andinfectivityrdquo Journal of Virology vol 79 no 21 pp 13262ndash132742005
[28] N Mancini G Sautto N Clementi et al ldquoNeutralization inter-fering antibodies a ldquonovelrdquo example of humoral immune dys-function facilitating viral escaperdquoViruses vol 4 no 9 pp 1731ndash1752 2012
[29] Y Modis S Ogata D Clements and S C Harrison ldquoVariablesurface epitopes in the crystal structure of dengue virus type3 envelope glycoproteinrdquo Journal of Virology vol 79 no 2 pp1223ndash1231 2005
[30] N Miller ldquoRecent progress in dengue vaccine research anddevelopmentrdquo Current Opinion in Molecular Therapeutics vol12 no 1 pp 31ndash38 2010
[31] J H Chavez J R Silva A A Amarilla and L T Moraes Figu-eiredo ldquoDomain III peptides from flavivirus envelope proteinare useful antigens for serologic diagnosis and targets forimmunizationrdquo Biologicals vol 38 no 6 pp 613ndash618 2010
[32] B Shrestha J D Brien S Sukupolvi-Petty et al ldquoThe develop-ment of therapeutic antibodies that neutralize homologous andheterologous genotypes of dengue virus type 1rdquo PLoS Pathogensvol 6 no 4 Article ID e1000823 2010
18 BioMed Research International
[33] G D Gromowski N D Barrett and A D T Barrett ldquoChar-acterization of dengue virus complex-specific neutralizing epi-topes on envelope protein domain III of dengue 2 virusrdquo Journalof Virology vol 82 no 17 pp 8828ndash8837 2008
[34] S Sukupolvi-Petty S K Austin W E Purtha et al ldquoType- andsubcomplex-specific neutralizing antibodies against domain IIIof dengue virus type 2 envelope protein recognize adjacentepitopesrdquo Journal of Virology vol 81 no 23 pp 12816ndash128262007
[35] S Sukupolvi-Petty S K Austin M Engle et al ldquoStructure andfunction analysis of therapeutic monoclonal antibodies againstdengue virus type 2rdquo Journal of Virology vol 84 no 18 pp9227ndash9239 2010
[36] Y-Q Deng J-X Dai G-H Ji et al ldquoA broadly flaviviruscross-neutralizingmonoclonal antibody that recognizes a novelepitope within the fusion loop of e proteinrdquo PLoS ONE vol 6no 1 Article ID e16059 2011
[37] J M Costin E Zaitseva K M Kahle et al ldquoMechanistic studyof broadly neutralizing human monoclonal antibodies againstdengue virus that target the fusion looprdquo Journal of Virologyvol 87 no 1 pp 52ndash66 2013
[38] CMMidgley A FlanaganH B Tran et al ldquoStructural analysisof a dengue cross-reactive antibody complexed with envelopedomain III reveals the molecular basis of cross-reactivityrdquoJournal of Immunology vol 188 no 10 pp 4971ndash4979 2012
[39] N J Moreland P Susanto E Lim et al ldquoPhage display appro-aches for the isolation of monoclonal antibodies against denguevirus envelope domain III from human and mouse derivedlibrariesrdquo International Journal ofMolecular Sciences vol 13 no3 pp 2618ndash2635 2012
[40] J J B Cockburn M E Navarro Sanchez N Fretes et alldquoMechanism of dengue virus broad cross-neutralization by amonoclonal antibodyrdquo Structure vol 20 no 2 pp 303ndash3142012
[41] R Rajamanonmani C Nkenfou P Clancy et al ldquoOn a mousemonoclonal antibody that neutralizes all four dengue virusserotypesrdquo Journal of General Virology vol 90 no 4 pp 799ndash809 2009
[42] L H Gould J Sui H Foellmer et al ldquoProtective and herapeuticcapacity of human single-chain Fv-Fc fusion proteins againstWest Nile virusrdquo Journal of Virology vol 79 no 23 pp 14606ndash14613 2005
[43] H Sultana H G Foellmer G Neelakanta et al ldquoFusion LoopPeptide of the West Nile virus envelope protein is essential forpathogenesis and is recognized by a therapeutic cross-reactivehuman monoclonal antibodyrdquo Journal of Immunology vol 183no 1 pp 650ndash660 2009
[44] T Oliphant M Engle G E Nybakken et al ldquoDevelopment ofa humanized monoclonal antibody with therapeutic potentialagainstWest Nile virusrdquoNature Medicine vol 11 no 5 pp 522ndash530 2005
[45] G E Nybakken T Oliphant S Johnson S Burke M S Dia-mond and D H Fremont ldquoStructural basis of West Nile virusneutralization by a therapeutic antibodyrdquo Nature vol 437 no7059 pp 764ndash769 2005
[46] G D Gromowski J T Roehrig M S Diamond J C LeeT J Pitcher and A D T Barrett ldquoMutations of an antibodybinding energy hot spot on domain III of the dengue 2 envelopeglycoprotein exploited for neutralization escaperdquo Virology vol407 no 2 pp 237ndash246 2010
[47] J T Roehrig R A Bolin andR G Kelly ldquoMonoclonal antibodymapping of the envelope glycoprotein of the dengue 2 virusJamaicardquo Virology vol 246 no 2 pp 317ndash328 1998
[48] R de Alwis S A Smith N P Olivarez et al ldquoIdentification ofhumanneutralizing antibodies that bind to complex epitopes ondengue virionsrdquo Proceedings of the National Academy of Sciencesof the United States of America vol 109 no 19 pp 7439ndash74442012
[49] S K Austin K A Dowd B Shrestha et al ldquoStructural basis ofdifferential neutralization of DENV-1 genotypes by an antibodythat recognizes a cryptic epitoperdquo PLoS Pathogens vol 8 no 10Article ID e1002930 2012
[50] M Throsby C Geuijen J Goudsmit et al ldquoIsolation and char-acterization of human monoclonal antibodies from individualsinfected with West Nile Virusrdquo Journal of Virology vol 80 no14 pp 6982ndash6992 2006
[51] A P Goncalvez R H Purcell and C-J Lai ldquoEpitope deter-minants of a chimpanzee Fab antibody that efficiently cross-neutralizes dengue type 1 and type 2 viruses map to inside andin close proximity to fusion loop of the dengue type 2 virusenvelope glycoproteinrdquo Journal of Virology vol 78 no 23 pp12919ndash12928 2004
[52] J T Roehrig J H Mathews and D W Trent ldquoIdentification ofepitopes on the E glycoprotein of Saint Louis encephalitis virususing monoclonal antibodiesrdquo Virology vol 128 no 1 pp 118ndash126 1983
[53] J T Roehrig L A Staudinger A R Hunt J H Mathews andC D Blair ldquoAntibody prophylaxis and therapy for flavivirusencephalitis infectionsrdquo Annals of the New York Academy ofSciences vol 951 pp 286ndash297 2001
[54] A P Goncalvez C-H Chien K Tubthong et al ldquoHumanizedmonoclonal antibodies derived from chimpanzee fabs protectagainst Japanese encephalitis virus in vitro and in vivordquo Journalof Virology vol 82 no 14 pp 7009ndash7021 2008
[55] M Arakawa T Yamashiro G-I Uechi M Tadano and ANishizono ldquoConstruction of humanFab (1205741120581) library and iden-tification of human monoclonal Fab possessing neutralizingpotency against Japanese encephalitis virusrdquo Microbiology andImmunology vol 51 no 6 pp 617ndash625 2007
[56] L Warter C Y Lee R Thiagarajan et al ldquoChikungunya virusenvelope-specific human monoclonal antibodies with broadneutralization potencyrdquo Journal of Immunology vol 186 no 5pp 3258ndash3264 2011
[57] P Pal K A Dowd J D Brien et al ldquoDevelopment of a highlyprotective combination monoclonal antibody therapy againstchikungunya virusrdquo PLoS Pathogens vol 9 no 4 Article IDe1003312 2013
[58] A Bertolotti-Ciarlet J Smith K Strecker et al ldquoCellular local-ization and antigenic characterization of Crimean-Congo hem-orrhagic fever virus glycoproteinsrdquo Journal of Virology vol 79no 10 pp 6152ndash6161 2005
[59] J K Battles and J M Dalrymple ldquoGenetic variation amonggeographic isolates of Rift Valley fever virusrdquo American Journalof Tropical Medicine and Hygiene vol 39 no 6 pp 617ndash6311988
[60] B AThibodeaux N C Garbino NM Liss et al ldquoA humanizedIgG but not IgM antibody is effective in prophylaxis and therapyof yellow fever infection in an AG12917D-204 peripheralchallenge mouse modelrdquo Antiviral Research vol 94 no 1 pp1ndash8 2012
BioMed Research International 19
[61] S Daffis R E Kontermann J Korimbocus H Zeller H-DKlenk and J Ter Meulen ldquoAntibody responses against wild-type yellow fever virus and the 17D vaccine strain charac-terization with human monoclonal antibody fragments andneutralization escape variantsrdquoVirology vol 337 no 2 pp 262ndash272 2005
[62] L N Levanov L E Matveev E P Goncharova et al ldquoChimericantibodies against tick-borne encephalitis virusrdquo Vaccine vol28 no 32 pp 5265ndash5271 2010
[63] A R Hunt S Frederickson C Hinkel K S Bowdish and J TRoehrig ldquoA humanized murine monoclonal antibody protectsmice either before or after challenge with virulent Venezuelanequine encephalomyelitis virusrdquo Journal of General Virologyvol 87 no 9 pp 2467ndash2476 2006
[64] J M da Silva Voorham I A Rodenhuis-Zybert N V A Nunezet al ldquoAntibodies against the envelope glycoprotein promoteinfectivity of immature dengue virus serotype 2rdquo PLoS ONEvol 7 no 3 Article ID e29957 2012
[65] T C Pierson Q Xu S Nelson et al ldquoThe stoichiometry ofantibody-mediated neutralization and enhancement of westnile virus infectionrdquoCell Host andMicrobe vol 1 no 2 pp 135ndash145 2007
[66] G Sautto N Mancini R A Diotti L Solforosi M Clementiand R Burioni ldquoAnti-hepatitis C virus E2 (HCVE2) glyco-proteinmonoclonal antibodies andneutralization interferencerdquoAntiviral Research vol 96 no 1 pp 82ndash89 2012
[67] R Burioni Y Matsuura N Mancini et al ldquoDiverging effectsof human recombinant anti-hepatitis C virus (HCV) antibodyfragments derived from a single patient on the infectivityof a vesicular stomatitis virusHCV pseudotyperdquo Journal ofVirology vol 76 no 22 pp 11775ndash11779 2002
[68] R J Phillpotts J R Stephenson and J S Porterfield ldquoAntibody-dependent enhancement of tick-borne encephalitus virus infec-tivityrdquo Journal of General Virology vol 66 no 8 pp 1831ndash18371985
[69] K A Dowd and T C Pierson ldquoAntibody-mediated neutraliza-tion of flaviviruses a reductionist viewrdquo Virology vol 411 no 2pp 306ndash315 2011
[70] A K I Falconar ldquoMonoclonal antibodies that bind to commonepitopes on the dengue virus type 2 nonstructural-1 andenvelope glycoproteins display weak neutralizing activity anddifferentiated responses to virulent strains implications forpathogenesis and vaccinesrdquo Clinical and Vaccine Immunologyvol 15 no 3 pp 549ndash561 2008
[71] K Meyer A Banerjee S E Frey R B Belshe and R Ray ldquoAweak neutralizing antibody response to hepatitis C virus enve-lope glycoprotein enhances virus infectionrdquo PLoS ONE vol 6no 8 Article ID e23699 2011
[72] R Burioni R AWilliamson P P Sanna F E Bloom and D RBurton ldquoRecombinant human Fab to glycoprotein D neutral-izes infectivity and prevents cell-to-cell transmission of herpessimplex viruses 1 and 2 in vitrordquo Proceedings of the NationalAcademy of Sciences of the United States of America vol 91 no1 pp 355ndash359 1994
[73] M S Rolph A Zaid N E Rulli and S Mahalingam ldquoDown-regulation of interferon-120573 in antibody-dependent enhancementof dengue viral infections of human macrophages is dependenton interleukin-6rdquo Journal of Infectious Diseases vol 204 no 3pp 489ndash491 2011
[74] S S Whitehead J E Blaney A P Durbin and B R MurphyldquoProspects for a dengue virus vaccinerdquoNature ReviewsMicrobi-ology vol 5 no 7 pp 518ndash528 2007
[75] S-M Lok V Kostyuchenko G ENybakken et al ldquoBinding of aneutralizing antibody to dengue virus alters the arrangement ofsurface glycoproteinsrdquoNature Structural andMolecular Biologyvol 15 no 3 pp 312ndash317 2008
[76] A P Goncalvez R Men C Wernly R H Purcell and C-J Lai ldquoChimpanzee Fab fragments and a derived humanizedimmunogiobulin G1 antibody that efficiently cross-neutralizedengue type 1 and type 2 virusesrdquo Journal of Virology vol 78no 23 pp 12910ndash12918 2004
[77] WM P BWahala A A Kraus L B HaymoreM A Accavitti-Loper and A M de Silva ldquoDengue virus neutralization byhuman immune sera role of envelope protein domain III-reactive antibodyrdquo Virology vol 392 no 1 pp 103ndash113 2009
[78] W D Crill H R Hughes M J Delorey and G-J J ChangldquoHumoral immune responses of dengue fever patients usingepitope-specific serotype-2 virus-like particle antigensrdquo PLoSONE vol 4 no 4 Article ID e4991 2009
[79] C Setthapramote T Sasaki O Puiprom et al ldquoHuman mono-clonal antibodies to neutralize all dengue virus serotypes usinglymphocytes from patients at acute phase of the secondaryinfectionrdquo Biochemical and Biophysical Research Communica-tions vol 423 no 4 pp 867ndash872 2012
[80] M Beltramello K L Williams C P Simmons et al ldquoThehuman immune response to dengue virus is dominated byhighly cross-reactive antibodies endowed with neutralizing andenhancing activityrdquo Cell Host andMicrobe vol 8 no 3 pp 271ndash283 2010
[81] C M Midgley M Bajwa-Joseph S Vasanawathana et al ldquoAnin-depth analysis of original antigenic sin in dengue virusinfectionrdquo Journal of Virology vol 85 no 22 pp 410ndash421 2011
[82] B Kaufmann M R Vogt J Goudsmit et al ldquoNeutralizationof West Nile virus by cross-linking of its surface proteins withFab fragments of the human monoclonal antibody CR4354rdquoProceedings of the National Academy of Sciences of the UnitedStates of America vol 107 no 44 pp 18950ndash18955 2010
[83] B Spurrier J M Sampson M Totrov et al ldquoStructural analysisof human and macaque mAbs 2909 and 25B implicationsfor the configuration of the quaternary neutralizing epitope ofHIV-1 gp120rdquo Structure vol 19 no 5 pp 691ndash699 2011
[84] J J SchlesingerMWBrandriss andT PMonath ldquoMonoclonalantibodies distinguish between wild and vaccine strains ofyellow fever virus by neutralization hemagglutination inhibi-tion and immune precipitation of the virus envelope proteinrdquoVirology vol 125 no 1 pp 8ndash17 1983
[85] M W Brandriss J J Schlesinger E E Walsh and M BrisellildquoLethal 17D yellow fever encephalitis in mice I Passive protec-tion by monoclonal antibodies to the envelope proteins of 17Dyellow fever and dengue 2 virusesrdquo Journal of General Virologyvol 67 no 2 pp 229ndash234 1986
[86] H Cho and M S Diamond ldquoImmune responses to West Nilevirus infection in the central nervous systemrdquo Viruses vol 4no 12 pp 3812ndash3830 2012
[87] D W Beasley A D Barrett and R B Tesh ldquoResurgence ofWest Nile neurologic disease in the United States in 2012 whathappened What needs to be donerdquo Antiviral Research vol 99no 1 pp 1ndash5 2013
[88] M Throsby J Ter Meulen C Geuijen J Goudsmit and J DeKruif ldquoMapping and analysis of West Nile virus-specific mon-oclonal antibodies prospects for vaccine developmentrdquo ExpertReview of Vaccines vol 6 no 2 pp 183ndash191 2007
[89] M S Diamond T G Roberts D Edgil B Lu J Ernst and EHarris ldquoModulation of dengue virus infection in human cells
20 BioMed Research International
by alpha beta and gamma interferonsrdquo Journal of Virology vol74 no 11 pp 4957ndash4966 2000
[90] Z Shimoni M J Niven S Pitlick and S Bulvik ldquoTreatmentof West Nile virus encephalitis with intravenous immunoglob-ulinrdquo Emerging Infectious Diseases vol 7 no 4 article 759 2001
[91] AHamdan PGreen EMendelsonM R Kramer S Pitlik andM Weinberger ldquoPossible benefit of intravenous immunoglob-ulin therapy in a lung transplant recipient with West Nile virusencephalitisrdquoTransplant InfectiousDisease vol 4 no 3 pp 160ndash162 2002
[92] M Haley A S Retter D Fowler J Gea-Banacloche and NP OrsquoGrady ldquoThe role for intravenous immunoglobulin in thetreatment of West Nile virus encephalitisrdquo Clinical InfectiousDiseases vol 37 no 6 pp e88ndashe90 2003
[93] K M Chung G E Nybakken B S Thompson et al ldquoAnti-bodies against West Nile virus nonstructural protein NS1prevent lethal infection through Fc 120574 receptor-dependent and-independent mechanismsrdquo Journal of Virology vol 80 no 3pp 1340ndash1351 2006
[94] E A Henchal L S Henchal and J J Schlesinger ldquoSynergisticinteractions of anti-NS1 monoclonal antibodies protect pas-sively immunized mice from lethal challenge with dengue 2virusrdquo Journal of General Virology vol 69 no 8 pp 2101ndash21071988
[95] J T Roehrig ldquoAntigenic structure of flavivirus proteinsrdquoAdvances in Virus Research vol 59 pp 141ndash175 2003
[96] M D Sanchez T C Pierson D McAllister et al ldquoCharacteri-zation of neutralizing antibodies to West Nile virusrdquo Virologyvol 336 no 1 pp 70ndash82 2005
[97] J H Beigel J L Nordstrom S R Pillemer et al ldquoSafety andpharmacokinetics of single intravenous dose of MGAWN1 anovel monoclonal antibody to West Nile virusrdquo AntimicrobialAgents and Chemotherapy vol 54 no 6 pp 2431ndash2436 2010
[98] C A Smeraski V Siddharthan and J D Morrey ldquoTreatmentof spatial memory impairment in hamsters infected with WestNile virus using a humanizedmonoclonal antibodyMGAWN1rdquoAntiviral Research vol 91 no 1 pp 43ndash49 2011
[99] D Lelli AMoreno E Brocchi et al ldquoWestNile virus character-ization and diagnostic applications of monoclonal antibodiesrdquoVirology Journal vol 9 article 81 2012
[100] T S Gritsun V A Lashkevich and E A Gould ldquoTick-borneencephalitisrdquo Antiviral Research vol 57 no 1-2 pp 129ndash1462003
[101] D Ruzek T S Gritsun N L Forrester et al ldquoMutations inthe NS2B and NS3 genes affect mouse neuroinvasiveness of aWestern European field strain of tick-borne encephalitis virusrdquoVirology vol 374 no 2 pp 249ndash255 2008
[102] M Ecker S L Allison T Meixner and F X Heinz ldquoSequenceanalysis and genetic classification of tick-borne encephalitisviruses from Europe and Asiardquo Journal of General Virology vol80 no 1 pp 179ndash185 1999
[103] H Holzmann M S Vorobyova I P Ladyzhenskaya et alldquoMolecular epidemiology of tick-borne encephalitis viruscross-protection between European and Far Eastern subtypesrdquoVaccine vol 10 no 5 pp 345ndash349 1992
[104] C W Mandl ldquoSteps of the tick-borne encephalitis virus repli-cation cycle that affect neuropathogenesisrdquo Virus Research vol111 no 2 pp 161ndash174 2005
[105] A V Atrasheuskaya T M Fredeking and G M IgnatyevldquoChanges in immune parameters and their correction in humancases of tick-borne encephalitisrdquo Clinical and ExperimentalImmunology vol 131 no 1 pp 148ndash154 2003
[106] K L Mansfield N Johnson L P Phipps J R Stephenson AR Fooks and T Solomon ldquoTick-borne encephalitis virusmdashareview of an emerging zoonosisrdquo Journal of General Virologyvol 90 no 8 pp 1781ndash1794 2009
[107] DWVaughn andCHHoke Jr ldquoThe epidemiology of Japaneseencephalitis prospects for preventionrdquo Epidemiologic Reviewsvol 14 pp 197ndash221 1992
[108] T Solomon H Ni D W C Beasley M Ekkelenkamp M JCardosa and A D T Barrett ldquoOrigin and evolution of Japaneseencephalitis virus in Southeast Asiardquo Journal of Virology vol 77no 5 pp 3091ndash3098 2003
[109] M R Wilson ldquoEmerging viral infectionsrdquo Current Opinion inNeurology vol 26 no 3 pp 301ndash306 2013
[110] J N Hanna S A Ritchie D A Phillips et al ldquoAn outbreakof Japanese encephalitis in the Torres Strait Australia 1995rdquoMedical Journal of Australia vol 165 no 5 pp 256ndash260 1996
[111] J N Hanna S A Ritchie D A Phillips et al ldquoJapanese ence-phalitis in north Queensland Australia 1998rdquo Medical Journalof Australia vol 170 no 11 pp 533ndash536 1999
[112] SGeorgeMGourie-Devi and J A Rao ldquoIsolation ofWestNilevirus from the brains of children who had died of encephalitisrdquoBulletin of theWorldHealthOrganization vol 62 no 6 pp 879ndash882 1984
[113] A K Gupta A A Koshy and V J Lad ldquoEnhanced protec-tion of mice against Japanese encephalitis virus infection bycombinations ofmonoclonal antibodies to glycoprotein ErdquoActaVirologica vol 55 no 2 pp 165ndash168 2011
[114] T H Lee B-H Song S-I Yun et al ldquoA cross-protective mAbrecognizes a novel epitope within the flavivirus NS1 proteinrdquoJournal of General Virology vol 93 no 1 pp 20ndash26 2012
[115] S C Weaver ldquoHost range amplification and arboviral diseaseemergencerdquo Archives of Virology Supplementum no 19 pp 33ndash44 2005
[116] M Solignat B Gay S Higgs L Briant and C Devaux ldquoRepli-cation cycle of chikungunya a re-emerging arbovirusrdquoVirologyvol 393 no 2 pp 183ndash197 2009
[117] O Schwartz and M L Albert ldquoBiology and pathogenesis ofchikungunya virusrdquo Nature Reviews Microbiology vol 8 no 7pp 491ndash500 2010
[118] W Akahata Z-Y Yang H Andersen et al ldquoA virus-like particlevaccine for epidemic Chikungunya virus protects nonhumanprimates against infectionrdquo Nature Medicine vol 16 no 3 pp334ndash338 2010
[119] M Kielian and A Helenius ldquopH-Iinduced alterations in thefusogenic spike protein of Semliki Forest virusrdquo Journal of CellBiology vol 101 no 6 pp 2284ndash2291 1985
[120] J White and A Helenius ldquopH-Dependent fusion between theSemliki Forest virus membrane and liposomesrdquo Proceedings ofthe National Academy of Sciences of the United States of Americavol 77 no 6 pp 3273ndash3277 1980
[121] J Dubuisson and C M Rice ldquoSindbis virus attachment isola-tion and characterization of mutants with impaired binding tovertebrate cellsrdquo Journal of Virology vol 67 no 6 pp 3363ndash33741993
[122] C Y Lee Y-W Kam J Fric et al ldquoChikungunya virus neutral-ization antigens and direct cell-to-cell transmission are revealedby human antibody-escapemutantsrdquo PLoS Pathogens vol 7 no12 Article ID e1002390 2011
[123] J E Voss M-C Vaney S Duquerroy et al ldquoGlycoproteinorganization of Chikungunya virus particles revealed by X-raycrystallographyrdquo Nature vol 468 no 7324 pp 709ndash712 2010
BioMed Research International 21
[124] L Li J Jose Y Xiang R J Kuhn and M G Rossmann ldquoStruc-tural changes of envelope proteins during alphavirus fusionrdquoNature vol 468 no 7324 pp 705ndash708 2010
[125] J Fric S Bertin-Maghit C I Wang A Nardin and L WarterldquoUse of human monoclonal antibodies to treat Chikungunyavirus infectionrdquo Journal of Infectious Diseases vol 207 no 2 pp319ndash322 2013
[126] P V Aguilar J G Estrada-Franco R Navarro-Lopez C FerroA D Haddow and S C Weaver ldquoEndemic Venezuelanequine encephalitis in the Americas hidden under the dengueumbrellardquo Future Virology vol 6 no 6 pp 721ndash740 2011
[127] S C Weaver R Winegar I D Manger and N L ForresterldquoAlphaviruses population genetics and determinants of emer-gencerdquo Antiviral Research vol 94 no 3 pp 242ndash257 2012
[128] S Paessler and S C Weaver ldquoVaccines for Venezuelan equineencephalitisrdquo Vaccine vol 27 no 4 pp D80ndashD85 2009
[129] R J Phillpotts ldquoVenezuelan equine encephalitis virus complex-specific monoclonal antibody provides broad protection inmurine models against airborne challenge with viruses fromserogroups I II and IIIrdquoVirus Research vol 120 no 1-2 pp 107ndash112 2006
[130] R J Phillpotts L D Jones and S C Howard ldquoMonoclonalantibody protectsmice against infection anddiseasewhen giveneither before or up to 24 h after airborne challenge with virulentVenezuelan equine encephalitis virusrdquoVaccine vol 20 no 11-12pp 1497ndash1504 2002
[131] J H Mathews J T Roehrig and D W Trent ldquoRole of com-plement and the Fc portion of immunoglobulin G in immunityto Venezuelan equine encephalomyelitis virus infection withglycoprotein-specific monoclonal antibodiesrdquo Journal of Virol-ogy vol 55 no 3 pp 594ndash600 1985
[132] L M OrsquoBrien C D Underwood-Fowler S A Goodchild AL Phelps and R J Phillpotts ldquoDevelopment of a novel mon-oclonal antibody with reactivity to a wide range of Venezuelanequine encephalitis virus strainsrdquoVirology Journal vol 6 article206 2009
[133] L M OrsquoBrien S A Goodchild R J Phillpotts and S D Per-kins ldquoA humanised murine monoclonal antibody protectsmice from Venezuelan equine encephalitis virus Evergladesvirus and Mucambo virus when administered up to 48h afterairborne challengerdquo Virology vol 426 no 2 pp 100ndash105 2012
[134] N Clementi N Mancini L Solforosi M Castelli M Clementiand R Burioni ldquoPhage display-based strategies for cloning andoptimization of monoclonal antibodies directed against humanpathogensrdquo International Journal of Molecular Sciences vol 13no 7 pp 8273ndash8292 2012
[135] A R Hunt R A Bowen S Frederickson T Maruyama JT Roehrig and C D Blair ldquoTreatment of mice with humanmonoclonal antibody 24h after lethal aerosol challenge withvirulent Venezuelan equine encephalitis virus prevents diseasebut not infectionrdquo Virology vol 414 no 2 pp 146ndash152 2011
[136] W-G Hu A L Phelps S Jager et al ldquoA recombinant human-ized monoclonal antibody completely protects mice againstlethal challenge with Venezuelan equine encephalitis virusrdquoVaccine vol 28 no 34 pp 5558ndash5564 2010
[137] T Rulker L Voss P Thullier et al ldquoIsolation and characterisa-tion of a human-like antibody fragment (scFv) that inactivatesVEEV in vitro and in vivordquo PLoS ONE vol 7 no 5 Article IDe37242 2012
[138] C T Walter and J N Barr ldquoRecent advances in the molecularand cellular biology of bunyavirusesrdquo Journal of General Virol-ogy vol 92 no 11 pp 2467ndash2484 2011
[139] C A Whitehouse ldquoCrimean-Congo hemorrhagic feverrdquo Anti-viral Research vol 64 no 3 pp 145ndash160 2004
[140] M Mertens K Schmidt A Ozkul and M H Groschup ldquoTheimpact of Crimean-Congo hemorrhagic fever virus on publichealthrdquo Antiviral Research vol 98 no 2 pp 248ndash260 2013
[141] A A Ahmed J M McFalls C Hoffmann et al ldquoPresence ofbroadly reactive and group-specific neutralizing epitopes onnewly described isolates of Crimean-Congo hemorrhagic fevervirusrdquo Journal of General Virology vol 86 no 12 pp 3327ndash33362005
[142] R Hewson A Gmyl L Gmyl et al ldquoEvidence of segment reas-sortment in Crimean-Congo haemorrhagic fever virusrdquo Journalof General Virology vol 85 no 10 pp 3059ndash3070 2004
[143] N K Blackburn T G Besselaar A J Shepherd and R Swane-poel ldquoPreparation and use of monoclonal antibodies for iden-tifying Crimean-Congo hemorrhagic fever virusrdquo AmericanJournal of Tropical Medicine and Hygiene vol 37 no 2 pp 392ndash397 1987
[144] T Ikegami ldquoMolecular biology and genetic diversity of RiftValley fever virusrdquoAntiviral Research vol 95 no 3 pp 293ndash3102012
[145] KKeegan andM S Collett ldquoUse of bacterial expression cloningto define the amino acid sequences of antigenic determinantson the G2 glycoprotein of Rift Valley fever virusrdquo Journal ofVirology vol 58 no 2 pp 263ndash270 1986
[146] T G Besselaar and N K Blackburn ldquoTopological mapping ofantigenic sites on the Rift Valley fever virus envelope glycopro-teins using monoclonal antibodiesrdquo Archives of Virology vol121 no 1ndash4 pp 111ndash124 1991
[147] L G Presta ldquoEngineering of therapeutic antibodies to mini-mize immunogenicity and optimize functionrdquo Advanced DrugDelivery Reviews vol 58 no 5-6 pp 640ndash656 2006
Submit your manuscripts athttpwwwhindawicom
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anatomy Research International
PeptidesInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporation httpwwwhindawicom
International Journal of
Volume 2014
Zoology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Molecular Biology International
GenomicsInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioinformaticsAdvances in
Marine BiologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Signal TransductionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Evolutionary BiologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Biochemistry Research International
ArchaeaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Genetics Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in
Virolog y
Hindawi Publishing Corporationhttpwwwhindawicom
Nucleic AcidsJournal of
Volume 2014
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Enzyme Research
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Microbiology
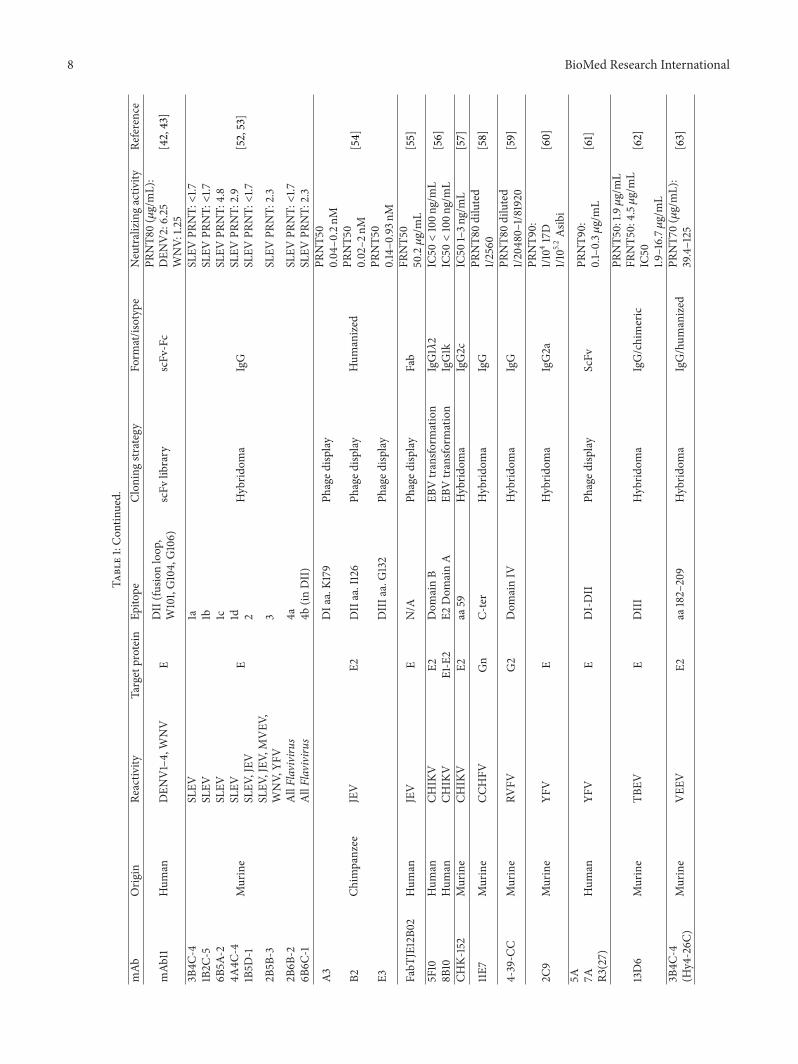
8 BioMed Research International
Table1Con
tinued
mAb
Orig
inRe
activ
ityTargetprotein
Epito
peClon
ingstr
ategy
Form
atisotype
Neutralizingactiv
ityRe
ference
mAb
11Hum
anDEN
V1ndash4WNV
EDII(fu
sionloop
W101G104G106)
scFv
library
scFv-Fc
PRNT8
0(120583gmL)
DEN
V2625
WNV
125
[4243]
3B4C
-4
Murine
SLEV
E
1a
Hybrid
oma
IgG
SLEV
PRNTlt17
[5253]
1B2C
-5SLEV
1bSLEV
PRNTlt17
6B5A
-2SLEV
1cSLEV
PRNT
48
4A4C
-4SLEV
1dSLEV
PRNT
29
1B5D
-1SLEV
JEV
2SLEV
PRNTlt17
2B5B
-3SL
EVJEV
MVEV
WNV
YFV
3SLEV
PRNT
23
2B6B
-2AllFlaviviru
s4a
SLEV
PRNTlt17
6B6C
-1AllFlaviviru
s4b
(inDII)
SLEV
PRNT
23
A3
Chim
panzee
JEV
E2
DIa
aK1
79Ph
aged
isplay
Hum
anized
PRNT5
0004ndash0
2nM
[54]
B2DIIaaI126
Phaged
isplay
PRNT5
0002ndash2
nM
E3DIIIa
aG132
Phaged
isplay
PRNT5
0014ndash0
93n
M
FabT
JE12B0
2Hum
anJEV
ENA
Phaged
isplay
Fab
FRNT5
0502120583gmL
[55]
5F10
Hum
anCH
IKV
E2Dom
ainB
EBVtransfo
rmation
IgG11205822
IC50lt100n
gmL
[56]
8B10
Hum
anCH
IKV
E1-E2
E2Dom
ainA
EBVtransfo
rmation
IgG1k
IC50lt100n
gmL
CHK-
152
Murine
CHIK
VE2
aa59
Hybrid
oma
IgG2c
IC50
1ndash3n
gmL
[57]
11E7
Murine
CCHFV
Gn
C-ter
Hybrid
oma
IgG
PRNT8
0diluted
12560
[58]
4-39-C
CMurine
RVFV
G2
Dom
ainIV
Hybrid
oma
IgG
PRNT8
0diluted
1204
80ndash181920
[59]
2C9
Murine
YFV
EHybrid
oma
IgG2a
PRNT9
011
0417D
1105
2Asib
i[60]
5A 7A R3(27)
Hum
anYF
VE
DI-DII
Phaged
isplay
ScFv
PRNT9
001ndash03120583
gmL
[61]
13D6
Murine
TBEV
EDIII
Hybrid
oma
IgGchimeric
PRNT5
019120583gmL
FRNT5
045120583
gmL
IC50
19ndash167120583
gmL
[62]
3B4C
-4(H
y4-26C
)Murine
VEE
VE2
aa182ndash209
Hybrid
oma
IgGhum
anized
PRNT7
0(120583gmL)
394ndash125
[63]
BioMed Research International 9
The structure of the 1A1D-2 bound to EDIII indicates that themAb binds to sites that are transiently exposed during viralldquobreathingrdquo at 37∘C and block infection during attachmentof the virion to the cell In particular this mAb neutralizesDENV1 2 and 3 serotypes and residues K307 and K310 arethemost critical residues for binding of 1A1D-2mAb [46 75]
Finally Shrestha et al immunized mice with a genotype2 strain of DENV1 virus and generated 79 new mAbs 16of which strongly inhibited infection by the homologousvirus and localized to DIII of E protein Surprisingly onlytwo mAbs E105 and E106 retained strong binding andneutralizing activity against all five DENV1 genotypes as wellas being protective in immunocompromised infected miceMoreover E105 and E106 exhibited therapeutic activity evenwhen administered as a single dose four days after inoculationwith a heterologous genotype 4 strain of DENV1 in the samemouse model [32]
243 Other Non-HumanmAbs Other non-human and non-murine as well as humanized mAbs directed against the Eglycoprotein have been described In this regard Goncalvezet al isolated a large panel of anti-E Fab fragments from chim-panzees infected with all four DENV serotypes Howeveronly a limited number of them displayed a cross-neutralizingactivity against DENV1 and 2 and to a lesser extent againstDENV3 and 4 In particular the authors calculated thatamong them the 1A5 mAb that was further humanizedneutralized DENV1ndash4 at a PRNT50 titer of 048 095 32and 43 120583gmL respectively Interestingly the humanized 1A5was also tested for binding and neutralization against theWNVDENV4 chimera JEV strain SA14-14-2 and Langatvirus (LGTV) strain TP 21 giving a PRNT50 titer of 38 21and 28120583gmL respectively Moreover the authors calculatedthat when administered in a dose of 2mg per kg of bodyweight 1A5 would give a serum titer of approximately 40and 20 50 reduction in plaque reduction neutralization test(PRNT50) against DENV1 and DENV2 respectively [76]Furthermore epitope mapping of 1A5 mAb localized to G106within the Flavivirus-conserved fusion loop inDII of DENV2E protein [51]
244 Anti-DENV Human mAbs As anticipated attemptsto isolate neutralizing human mAbs have been morechallenging due to the restricted elicited Ab-repertoire whichrecognizes DIII of E glycoprotein in naıve as well as ininfected patients [77 78] However several groups reportedthe isolation of neutralizing mAbs from infected subjects Inthis regard Setthapramote et al isolated a total of 136 humanhybridoma clones producing specific mAbs against DENVobtained using PBMCs from nine blood samples from fouracute-phase patients secondarily infected with DENV2 andfive convalescent-phase patients Interestingly the authorsfound that most of the acute-phase mAb clones were cross-reactive with all four DENV serotypes with most of themrecognizing the E protein and endowing of neutralizingactivity against all DENV serotypes compared to thosederived from convalescent-phase patients In particularfrom the acute-phase PBMCs 818 were anti-E 66 wereanti-prM and 33 were anti-NS1 while 133 anti-E 133
anti-prM and 533 anti-NS1 mAbs clones were obtainedfrom convalescent-phase PBMCs [79] Previous studiesconfirmed the data obtained by Setthapramote et al onconvalescent-phase patients but this group firstly reportedthe efficient preparation of human mAbs with strongneutralizing activity titers against all four DENV serotypesusing PBMCs from acute-phase patients secondarily infectedwith DENV [80 81]
The group of de Alwis through immunoglobulin deple-tion studies reported that a substantial fraction of DENV-reactive Abs in human immune sera including type-specificneutralizing Abs bound to the intact virion but not torecombinant E proteinThe authors confirmed these observa-tions also isolating human neutralizing mAbs and proposedthat humans produce Abs that neutralize DENV infectionby binding a complex quaternary structure epitope that isexpressed only when E proteins are assembled on a virusparticle [48] Similar findings have been described for severalother viruses including WNV and HIV [82 83] Mappingstudies indicate that this epitope has a footprint that spansadjacent E protein dimers and includes residues at the hingebetween domains I and II of E protein [48]
Recently Costin et al isolated three broadly neutralizinganti-DENVhumanmAbs named 48AD11C and 16DThesemAbswere isolated from three different convalescent patientswith distinct histories of DENV infection All three mAbsrecognized the E glycoprotein with high affinity neutralizedall four serotypes of DENV but mediated ADE of infection inFc receptor-bearing cells at subneutralizing concentrationsMapping studies revealed that all three mAbs bind discontin-uous epitopes within the highly conserved fusion-loop regionof DII (contacting residues W101 L107 andor G109) [37]
25 Yellow Fever Virus (YFV) TheWHO estimates that thereare 200000 cases of yellow fever virus (YFV) infections and30000 related deaths every year especially in Sub-SaharanAfrica In fact different to themajority of human arbovirusesand similarly to DENV and CHIKV YFV has expanded itshost range to include humans as an amplifying host [1]Symptoms occurring from 3 to 6 days after infection withYFV consist mostly in fever chills anorexia lumbosacralpain nausea and vomiting This syndrome lasts about 3days in most cases and is sometimes followed by a one-daylasting period of remission Fever will reoccur together withdark hematemesismelena petechiae and other hemorrhagicsymptoms Convalescence is characterized by deep asthenialasting up to two weeks Rarely prognosis can be fatal Aneffective live-attenuated vaccine (17D) derived from the Asibistrain by serial passages in chicken embryos is available [84]
251 mAbs against YFV In 1983 Schlesinger et al produceda battery of mAbs after immunization of BALBc mice byinjection of 17D YFV vaccine strain [84] Among them thespecificity towards the E and NS1 proteins was describedeven though anti-NS1 Abs were not able to show neutralizingactivity The 13 IgG and 1 IgM anti-E protein mAbs wereclassified in five groups according to their specificity GroupA consisting of the only IgM produced in this experiment
10 BioMed Research International
could neutralize 17D YFV virus only group B 17D and Asibistrains only group C Asibi strain only group D could onlyneutralize Asibi but not 17D strain showing also reactivityagainst DENV2 Zika or Banzi viruses All of the four mAbsbelonging to this group were able to cross-react with Zikaandor Banzi viruses and two (4E11 and 5H3) neutralizedDENV2 virus with 4E11 neutralizing also Banzi virus The3E9 mAb the only component of the group E could notneutralize 17D nor Asibi strains
All of the IgG mAbs resulted able to protect bothprophylactically and therapeutically BALBc or CD-1 micefrom lethal intracerebral challenge with 17D-204 strain [85]The chimeric form of the group B mAb 2C9 IgG (2C9-cIgG) was able to provide AG129 mice a 72 survival whenadministered 24 hours before infection with 17D-204 strainIts murine form could provide the survival of the 95 ofthe mice Appreciable results were also obtained when bothmurine 2C9 and 2C9-cIgG were administered 24 hours post-infection with the survival of 70 and 20 of mice respec-tively Viral presence was not detected in survivingmice [60]
In 2005 Daffis and colleagues constructed two Ab-phage libraries by cloning the repertoire of YFV-infectedpatients Panning was then performed with YFV-17D virionsThe scFv-5A 7A and R3(27) showed a neutralizing activityspanning from50 to 100 inPRNTassays against bothYFV17D-204-WHO and Asibi strains In further tests reactivitywas observed against wild-type YFV strains of West Africagenotype I and II (Nigeria 1987 and Asibi strains resp)and EastCentral Africa (CAR 1986 Ethiopia 1961 strains) Aconcentration from 01 to 03120583gmL could yield a 90 plaquereduction Reactivity was also observed against the strainSenegal 1990 in a lesser extent Production of escape variantscould demonstrate that scFv-5A 7A and R3(27) epitopes arebuilt up by residues extensively separated in themonomers ofE glycoprotein that however result in closed proximity whenthe homodimeric form of E is constituted [61]
26 West Nile Virus (WNV) The West Nile virus (WNV) isan epidemic neurotropic virus estimated to be responsibleof about 36000 cases and 1500 deaths registered in theUnited States between 1992 and 2012 [86] WNV antigenicityallows its classification into the Japanese encephalitis virus(JEV) serocomplex Genomic analysis has revealed two maingenetic lineages of WNV lineage I viruses circulating in theUSA Europe the Middle East Africa India and Australiaand lineage II viruses isolated from Sub-Saharan Africa andMadagascar [1]
WNV was firstly identified in 1937 and is endemicin many countries of Africa Middle East and West AsiaHowever after 1990 frequent outbreaks of WNV infectionswere reported in Romania Israel and later in North Americaand across the USA in 1999 and recently in 2012 [87]
Most of WNV infected individuals do not develop symp-toms while about 20 develop a self-limiting illness calledWest Nile fever Acute symptoms include fever tirednessand swollen lymph glands but in a minority of cases alsoencephalitis with long-term deficits in cognitive functionand motor skills have been reported However in less than
1 of infected patients WNV having crossed the blood-brain barrier is responsible of neuroinvasive and potentiallylethal form of the disease In these cases degeneration andapoptosis upon infection of neurons and the consequentinflammatory response can occur Related symptoms includehigh fever coma muscle weakness and paralysis Immun-odepression and advanced age have been correlated with anaugment risk to develop a severe disease [87] There are nospecific treatment options or licensed vaccines for humans[88]
WNV is transmitted by infected mosquitos and initialreplication is thought to occur primarily in dendritic cells inthe skin which migrate to secondary lymphoid tissues wherethe replicating virus enters the circulation To date onlyefficacious WNV veterinarian vaccines have been licensedwhile there are no licensed vaccines for protection againstWNV in humans [88]
261 mAbs against WNV Broad-spectrum antivirals suchas type I IFN-120572 ribavirin mycophenolic acid in WNVinfection showed ineffective results in vivo despite in vitrothey showed some activity [89]
However experiments in murine models extrapolationof clinical data as well as passive administration of pooledimmune-120574-globulins (OmriGam) containing a significanttiter of neutralizing Abs before and after infection showedan important role of the humoral immune response incontrolling viremia and prevent viral dissemination Indeedthe development of a neutralizing mAb-based therapy seemsto be encouraging for a possible treatment of infected patients[90ndash92]
Similarly to what has been observed for DENV only anti-E Abs have been identified as neutralizing and protectiveAlso anti-NS1 Abs have shown a protective role howevertheir mechanism of protection has not yet been elucidated asthe NS1 protein is secreted from infected cells and not presenton the virions [93ndash95]
In this regard similarly to what has been describedfor DENV studies performed using a naıve human scFvlibrary for panning with purified WNV E protein supportthe hypothesis that no Abs against the neutralizing DIII canbe isolated However DIII specific Abs were isolated in asubsequent study using immunoglobulin libraries obtainedfrom threeWNV infected patients for biopanning on purifiedinactivated virus virus-like particles consisting of prM andE proteins or recombinant E glycoprotein Although theproportion of DIII-specific mAbs was low (8) comparedwith anti-DII mAbs (47) Two out of the four anti-DIIImAbs were potently neutralizing and protective in vivowhereas only three out of the 24 anti-DII were weaklyneutralizing in vitro and non-protective in vivo In particularresidues that are critical for neutralization lies on the regionsspanning amino acids 305ndash312 330ndash333 and 365 that arelocated on adjacent exposed loops ofDIIIHowever sequencealignment of E protein of different flaviviruses such as DENVand JEV revealed a considerable variation compared tothe whole E protein This observation further suggests thatdifferently from broadly cross-reactive anti-DII Abs anti-DIII neutralizing Abs are virus-type specific However of
BioMed Research International 11
these mAbs two (CR4374 and CR4353) protected mice fromlethal WNV challenge at 50 protective doses of 129 and357 120583gkg of body weight respectively [50]
Sanchez et al using three different immunization strate-gies (ie inactivated virus naked DNA and recombinantprotein) isolated ninemurinemAbsmost of which bound toconformation-dependent epitopes in DIII of the E protein Inparticular neutralizing mAbs named 8B10 11C2 10C5 and17C8 were obtained from mice immunized with inactivatedvirus alone or in combination with a DNA plasmid andbound to the same region of DIII with high affinity Incontrast mAbs obtained by immunization with a solubleversion of the E glycoprotein did not exhibit neutralizingactivity These non-neutralizing mAbs were cross-reactivewith several other flaviviruses including SLEV JEV YFVand Powassan virus confirming the conserved nature ofFlavivirus non-neutralizing epitopes [96]
Gould et al isolated 11 unique human single-chain vari-able region Ab fragments (scFvs) that bind the E protein ofWNVAmong them a humanmAb namedmAb11 expressedas a scFv-Fc fusion protein was further characterized Itrecognizes the fusion loop at the distal end of DII ofthe WNV E protein and cross-reacts with all four DENVserotypes and provides protection against DENV2 and 4as well as WNV [42] Moreover therapeutic studies of thismAb in WNV-infection model mice provided substantialprotection when administered after 5 days post-infectionInterestingly a neutralization escape variant of this mAbfailed to cause lethal encephalitis (at higher infectious doses)or induce the inflammatory responses associated with blood-brain barrier permeability in mice compared to the parentalWNV suggesting an important role for the fusion loop inviral pathogenesis [43]
Oliphant et al isolated an anti-DIII E protein mAbnamed E16 fromhybridomas obtained after immunization ofmice with recombinant WNV E protein which neutralizedall WNV strains with PRNT50 values of 4 to 18 ng andPRNT90 values of 53 to 297 ng One hundred microgramsof mAb protected greater than 90 of mice from lethalinfection and even a single 4120583g treatment of E16 on day 2 afterinfection prevented mortality Moreover humanization ofthis mAb confirmed as therapeutically effective in mice [44]Subsequent studies revealed that mAb E16 neutralization ismediated by engagement of four discontinuous segmentsof DIII including the amino-terminal region (amino acidresidues 302ndash309) and the three connecting loops BC (aminoacid residues 330ndash333) DE (amino acid residues 365ndash368)and FG (amino acid residues 389ndash391) Moreover no ADE ofinfection was detected when E16 mAb was used at saturatingconcentrations [45 47]
Furthermore results of a Phase I safety study of thehumanized E16 mAb (designated MGAWN1) have beenreported and suggested that doses of up to 30mgkgwerewelltolerated with few mild adverse events and would providean excess of virus neutralizing activity for 3-4 weeks aftertreatment However a case of anti-MGAWN1 Ab elicitationoccurred with the consequent increased rate of clearance andindeed impacting efficacy A Phase II safety and efficacy studyof MGAWN1 is ongoing [97] Furthermore the possibility of
preventing or treating WNV-induced memory deficits wasrecently investigated In this study hamsters were treatedintraperitoneally with 32mgkg ofMGAWN1mAb at 45 daysafter subcutaneously challenging with WNV InterestinglyMGAWN1 prevented mortality weight loss and improvedfood consumption of WNV-infected hamsters compared tocontrols [98]
Recently Lelli et al isolated six anti-E mAbs frominactivated-WNV immunized mice In particular three ofthem (3B2 3D6 and 4D3) neutralized lineage I and II WNVwith the first two recognizing the same epitopes located onthe distal lateral surface of DIII (critical amino acid residueK307) Conversely 4D3mAb recognized a novel neutralizingepitope on DII (critical residues S276 and T278) Indeedfurther protective and therapeutic studies are needed toascertain their neutralizing activity in vivo [99]
Finally to conclude like DENV infection Abs directedagainst the M protein are not protective and neutralizingMoreover in humans a skewed humoral immune responseagainst DII has been frequently observed and confirmedalso with the hybridoma technology [50] The isolation andelicitation of neutralizingAbs directed against the fusion loopand DIII of the E protein represent thus the most challengingand promising goal for the development of new effectivetherapeutic inhibitors and immunogens respectively
27 Tick-Borne Encephalitis Virus (TBEV) Tick-borne ence-phalitis virus (TBEV) is one of the most dangerous agentscausing human neuroinfections occurring mostly in Europeand Asia and with a potential fatal prognosis [100] TBEV isbelieved to cause 3500ndash10000 human cases of encephalitisin Europe per year with a high morbidity in Russia CzechRepublic Austria and Germany In particular between 1990and 2007 an average of 8755 cases of TBE was reported peryear in Europe and Russia Despite the fact that Russia isthe country with most infections registered annually CzechRepublic incidence is among the highest in Europe with 400ndash1000 clinical cases reported every year [101]
Three subtypes of TBEV are classified namely EuropeanSiberian and Far-Eastern sharing most of the genetic andantigenic features [102] In fact a high degree of antigenichomogeneity between different strains of TBEV has beendescribed [103]
Clinical manifestations vary among the subtypes butusually they start with a short febrile period of 7ndash14 daysafter the tick bite Fatigue headache and pain in the neckshoulders and lower back together with high fever andvomitingmay be present [100]Thesemanifestations are oftenfollowed by an asymptomatic phase lasting from 2 to 10days after remission from the fever with possible progres-sion to neurological disease Neurological symptoms includemeningitis encephalitis myelitis and radiculitis Mortalityoccurs in 1-2 of the European subtype-infected patientsbut fatal prognosis can occur in up to 20ndash40 of the Far-Eastern subtype-infected patients [104] Mortality rates ofthe Siberian subtype are similar to those observed for theEuropean [105]
Effective and safe vaccines against TBEV produced frominactivated virus have been developed and licensed for their
12 BioMed Research International
use in humans However an emergency therapy in theabsence of a mass immunization is needed as no effectivetreatments are yet available [106]
271 mAbs against TBEV Levanov et al described the chim-erization of two murine mAbs (13D6 and 10C2) directedagainst the DIII and DII respectively of the TBEV E glyco-protein The chimeric mAbs present binding characteristicssimilar to the parentalmAbsMoreover as the parentalmAbsonly the chimeric mAb 13D6 was able to neutralize TBEVinfectivity in vitro In particular neutralization studies withthe murine chimeric and scFv forms of mAb 13D6 were per-formed In particular murine 13D6 showed an IC50 titer of115 120583gmL in Focus Reduction Neutralization Tests (FRNT)against TBEV strain 205 and of 29 120583gmL in PRNT againstTBEV strain Softjin Chimeric 13D6 showed an IC50 titer of45 120583gmL in FRNT against TBEV strain 205 and of 19 120583gmLin PRNT against TBEV strain Softjin ScFv 13D6 showed anIC50 titer of 167 120583gmL in FRNT against TBEV strain 205and of 112 120583gmL in PRNT against TBEV strain Softjin [62]
28 Japanese Encephalitis Virus (JEV) Japanese encephalitisvirus (JEV) is a mosquito-borne flavivirus that over thepast few decades has caused several outbreaks throughoutChina Southeast Asia Australia and Papua New Guineawith a prevalence recently estimated to be of about 70000casesyear [107ndash109] A 40 mortality was recorded in someof the JEV-affected areas Moreover many survivors facesome neurological problems and complications [108] Since1995 the disease has also emerged in Non-Asian regions suchas Northern Australia [110 111] The situation in SoutheastAsia however is further complicated by the overlappingepidemics of JEV and DENV as well as sporadic casesof WNV infections detected in some of the affected areasparticularly in India [112]
Effective vaccines both live-attenuated and inactivatedstrains of JEV have been developed and licensed in majorcountries However as for the other arthropod-borne Fla-vivirus members no specific antiviral drugs are currentlyavailable [4]
281mAbs against JEV Aspreviously described for the otherFlavivirusmembers many groups isolated and characterizedanti-JEVmAbs showing different neutralizing and protectiveproperties In this regard Gupta et al used combinations ofanti-E JEV mAbs (Hs1-4) in mice protection experimentsIn particular they found that the singularly mAb protectionranged from 45 to 65 when 100120583g of mAb were adminis-tered while equimolar combinations of two or three mAbsgave 85ndash90 or 100 protection respectively [113] In similarexperiments Lee et al demonstrated that a Flavivirus anti-NS1 mAb named 16NS1 cross-reacted with JEV as well asWNV and exhibited protective activity against WNV as wellas a lethal JEV infection However no neutralizing activitywas observed using this mAb against both WNV and JEV inin vitro experiments suggesting the participation of otherAb-mediatedmechanisms in vivo In particular 95ofmicewere
protected when 500120583g of mAb were administered intraperi-toneally and concomitantly to intramuscular injection of JEV
Overlapping peptide mapping analysis combined withsite-specific mutations identified the 116KAWGKSILFA125region and critical amino acid residues W118 and I122 as16NS1 mAb epitope highly conserved in WNV and JEVstrains [114]
Arakawa et al isolated from a combinatorial human Fablibrary constructed from peripheral blood lymphocytesobtained from JEV hyperimmune volunteers Among 188randomly selected clones FabTJE12B02 showed the best 50focus reduction endpoint at the concentration of 502 120583gmLagainst the JEV strain Nakayama [55]
Goncalvez et al isolated three mAbs named Fabs A3B2 and E3 by repertoire cloning from chimpanzees initiallyimmunized with inactivated JE-VAX and then boosted withattenuated JEV SA14-14-2 In particular these mAbs reactedwith epitopes in three different E domains in DI (amino acidresidue K179) in DII (I126) and in DIII (G132) for Fabs A3B2 and E3 respectively Moreover these Fabs as well as thederived humanized counterpart mAbs exhibited high neu-tralizing activities against a broad spectrum of JEV genotypestrains Moreover these mAbs exhibited a 50 protectivedose of 084 120583g (B2) 58120583g (A3) and 247120583g (E3) in mousemodels Finally administration of 200 120583gmouse of mAb B2one day after otherwise lethal JEV infection protected 50of mice and significantly prolonged the average survival timecompared to that of mice in the unprotected group [54]
29 St Louis Encephalitis Virus (SLEV) St Louis encephalitisvirus (SLEV) was first discovered as the mosquito-borneagent responsible for over 1000 cases of encephalitis duringa 1933 summer outbreak in St Louis (Missouri) and nowis a reemerging human pathogen widely distributed in theAmerican continent causing several human encephalitisoutbreaks over the last 80 years Additional epidemics haveindeed occurred from 1964 to 2006 in the Americas rangingfrom the US to Argentina and Brazil As a member of theFlavivirus genus the SLEVE glycoprotein ectodomain is 68identical to serocomplex-related JEVEbut only 46 and 40identical to those ofDENV2 andTBEV viruses fromdifferentserocomplexes [115]
291 mAbs against SLEV In 1983 Roehrig et al isolatedtwenty-one hybridomas producing murine mAbs specificfor the E glycoprotein of SLEV strain MSI-7 Serologicreactivities were initially determined by cross-reactivity indi-rect immunofluorescence assays using 22 strains of SLEVand 8 other related flaviviruses Four groups demonstratingtype- subcomplex- supercomplex- and group-specific reac-tivity patterns were identified Analysis of hemagglutination-inhibition and virus neutralization subdivided the cross-reactivity groups into eight epitopes (E-1a b c d E-2 E-3 and E-4a b) one of them following localized in DII (4b)[52 53]
Moreover among the previously described isolated anti-WNV mAbs by Gould et al the chimeric scFv-Fc 79 effec-tively neutralized also SLEV resulting in gt80 of PRNT80when used at 5 120583gmL [42]
BioMed Research International 13
3 Alphaviruses
Major human arboviruses in the Togaviridae family Chikun-gunya (CHIKV) and Venezuelan equine encephalomyelitis(VEEV) viruses belong to the Alphavirus genus which iscomposed of viruses with an icosahedral nucleocapsid sur-rounded by a lipid envelope and glycoprotein spikes Thestructural proteins of alphaviruses arise through co- andpost-translational processing of a polyprotein encoded by asingle positive-stranded RNAproducing the capsid (C) PE26 K and E1 The PE2 glycoprotein is a precursor containingthe E3 glycoprotein fused to the amino terminus of the E2envelope glycoprotein The PE2 glycoprotein is followed by6K a small membrane-associated protein and E1 the secondpolypeptide component of glycoprotein spikes
Trimerized heterodimers of the E1 and E2 viral glyco-proteins form the surface spikes and contain determinantsof viral tropism and virulence The E3 glycoprotein acts as asignal for transport of PE2 across themembranes of the roughendoplasmic reticulum and may promote the formation andintracellular transport of E1-PE2 heterodimers to the cellsurface During transport to the cell surface PE2 undergoesa maturational cleavage event by a furin-like protease toproduce E2 and E3 [116]
The E2 glycoprotein promotes specificity of virus bindingto the host cell surface and is a target of the humoral immuneresponse The E1 glycoprotein mediates fusion of the virionenvelope with the membranes of acidified endosomes allow-ing release of the nucleocapsid into the cytoplasm and theonset of viral replicationAlphavirus E1 shares no appreciablesequence identity to Flavivirus E protein Despite differencesin their amino acid sequences and arrangements on the viralparticle the structures of E and E1 are remarkably similarIndeed the conservation of three domains has been welldocumented in crystal structures of flavivirus E ectodomainsfrom TBEV DENV JEV and WNV as well as Alphavirus E1from Semliki Forest virus (SFV) and CHIKV Abs to the E1glycoprotein do not typically neutralize virus infectivity invitro but can protect against lethal challenge in animals
By analogy with the prM protein of dengue Flavivirusfurin cleavage of PE2 may begin after transit of an acidiclate component of the Golgi body where E3 is thought tosuppress the acid pH-triggered activation of glycoprotein E1fusion capability Protective anti-E3 mouse mAbs have alsobeen described [117]
31 Chikungunya Virus (CHIKV) The chikungunya virus(CHIKV) belongs to the Semliki Forest clade and was firstlyisolated in 1953 in Tanzania during an epidemic outbreakfollowing occurred also in Asia and Africa In the last fivedecades several CHIKV outbreaks have been described bothin Africa and Asia separated by gaps lasting from two totwenty years In 2005-2006 about 300000 cases out of785000 inhabitants were reported in La Reunion island witha fatal prognosis for 237 of the patients [117] Neither Europenor the Americas have had outbreaks of CHIKV so far exceptfor imported isolated cases Three genotypes of CHIKV aredescribed Asian EastCentralSouth African (ECSA) andWest African with an amino acid identity spanning from
952 to 998 Recent epidemics in Africa and Indiansubcontinent were caused by strains belonging to the ECSAgenotype Transmitted by Aedes mosquitoes CHIKV ismaintained in the human population by human-mosquito-human transmission [1] The disease is characterized bydengue-like symptoms such as chills high fever headacheand persistent myalgia and a further incapacitating arthralgia(from which the name chikungunya) which affects 40 ofinfected subjects However prognosis is rarely fatal [116]
There is currently no commercial vaccine and antiviraltreatment forCHIKV although some candidate vaccines havebeen tested in humans In this regard in 2000 US Armyperformed a Phase II clinical trial testing a live-attenuatedCHIKV vaccine (TSI-GSD-218) derived from a 1962 strain(15561) of an outbreak in Thailand Out of 58 every patientdeveloped neutralizingAbs and 5 lamentedmild tomoderatejoint pain [117] A phase III trial of this candidate vaccineis ongoing Furthermore a new formulation using virus-likeparticles was able to induce neutralizing Abs in macaquesagainst different CHIKV strains [117 118] Indeed it has beendescribed that infection seems to elicit long-lasting protectiveimmunity and cross-protection among CHIKV and otheralphaviruses
CHIKV entry into host cell is demonstrated to be medi-ated by envelope glycoproteins E1 and E2 which allowvirus fusion to cell membrane in low pH conditions andrecognition of an unknown cellular receptor respectively[116 119ndash121] Even though the genome replication relies onan error-prone RNA-dependent RNA-polymerasis CHIKVstrains S27 and 05115 Reunion showed in E1 and E2 alow amino acid variation of 068 and 33 respectivelyand such a low variability may be needed for an effectivereplication in two phylogenetically distant hosts [116 122]Recently the crystal structure of E1 and E2 heterodimerhas been resolved both at low pH and natural conditionsshedding light on E1 and E2 ectodomainsrsquo structure Inparticular similarly to flaviviruses E protein E1 ectodomainis made of the N-terminal DI the DII containing the fusionloop and the DIII at the C-terminal On the other hand E2ectodomain comprises theN-terminal domainA the domainB supposed to interact with the host cellrsquos unknown receptorand domain C at the C-terminal E1 DIII and E2 domain Care located close to the viral membrane [123 124]
311 mAbs against CHIKV Warter et al described 5F10 and8B10 two human mAbs which strongly neutralized severalCHIKV isolates in vitro without cross-reactivity againstother alphaviruses but to Onyong-nyong virus [56] Mixedpreparation of 5F10 and 8F10 did not show neither synergisticnor addictive effect in vitro and studies upon escape mutantsdemonstrated that 5F10 mAb binds at the tip of the E2domain B while 8B10 recognizes residues close to E1 fusionloop and amino acids within E2 domain A which form atransitional epitope under low pH conditions The authorsspeculated that 5F10 and 8B10 may inhibit CHIKV entry andfusion to the cell membrane respectively It is worth notingthat the previously mentioned escape mutants displayedmutations associated with reduced viral fitness in vitro evenafter 13 neutralizationamplification rounds In the same
14 BioMed Research International
study cell-to-cell transmission was firstly demonstrated as anescapemechanism enhanced by the E2mutation R82G [122]
In further in vivo studies 5F10 and 8B10 significantlydelayed CHIKV-caused death of AGR129 mice both in pro-phylactic and therapeutic tests Interestingly in therapeutictreatment these mAbs showed a synergistic effect whenadministered in combination with the total amount of mAbsinjected being the same of the single-mAbs Possibly thesetreatment did not allow themice to survive CHIKV challengedue to Ig clearing and 6ndash10 days half-life usually described forhuman mAbs [125]
Recently Pal et al cloned thirty-six murine mAbs ableto neutralize the ECSA La Reunion 2006 OPY-1 strain ofCHIKV (CHIKV-LR) the majority of which also neutralizeinfection of other strains corresponding to the Asian andWest African genotypes In particularmAbCHK-152 showedthe highest and broadest neutralizing activity with FRNTvalues indicating an IC50 value of 1 to 3 ngmL dependingon the viral strain used Among the thirty-six mAbs four(CHK-102 CHK-152 and CHK-166 CHK-263) could pre-vent lethality of immunodeficient Ifnarminusminus C57BL6 micewhen administered one day before exposition with CHK-152 and CHK-263 showing the ability to protect mice at thelowest dose (10 120583g) Therapeutic studies were performed onthese mAbs administering a single dose of 100 120583g 24 hoursafter infection The highest activity was described for CHK-166 able to protect 63 of the mice Testing the combinedactivity of the mAbs in therapeutic studies administration ofCHK-166 plus CHK-152 in a dose of 250120583g each turned out tobe themost effective protecting 71 of the IfnarminusminusC57BL6mice 60 hours after infection [57]
32 Venezuelan Equine Encephalomyelitis Virus (VEEV)Venezuelan equine encephalomyelitis virus (VEEV) is main-tained in a natural transmission cycle between mosquitoesand small rodents The first documented outbreaks occurredin the 1930s and several epidemics have been reported so farin Latin-American countries such as Venezuela ColombiaPeru Ecuador Bolivia Honduras Mexico and Panama withhundreds of thousands of human cases being reported anda case-fatality rate up to 1 during the 1969 Ecuador out-break Clinical manifestations of VEE are indistinguishablefrom Dengue [126] Six serogroups (IndashVI) are currentlyrecognized within the VEEV complex VEEV caused humanand equine outbreaks in the Americas for nearly a centuryEquine epizootics have high mortality (38ndash83) leading toa high viremia followed by a lethal encephalitis and often tohuman epidemics involving thousands of cases and hundredsof deaths Indeed VEEV is infectious for humans by theairborne route and has been responsible for a number oflaboratory infections In humans the disease is usually self-limiting with a febrile illness with 1ndash4 of cases progressingto severe encephalitis It is a hazard to laboratory workershas been developed as a biological weapon and is a potentialbioterrorist agent [127] There are no antiviral drugs andvaccines licensed for the treatment of VEEV infection inhumans Indeed although experimental live-attenuated vac-cines have been developed (eg TC-83) with good levels ofprotection in equine and mice in humans they may fail to
give protection in the majority of cases [128] Consideringthe protective role exerted by humoral immune response inanimal models it could be concluded that treatment withspecific IgG may have a beneficial antiviral effect in humanairborne infections with pathogenic strains of VEEV
321 mAbs against VEEV Several works support that mAbsmay protect against airborne VEEV as well as after airborneexposure toVEEVAb reactivity to both surface glycoproteins(E1 and E2) is associated with neutralization of the virusin vitro and passive protection against virus challenge Aseries of anti-E1 and anti-E2 mouse and humanized mAbshave been described recently Among them Phillpotts et alexamined two anti-E2 murine mAbs named 1A4A-1 and1A3A-9 both having potent protective activity against subcu-taneous VEEV challenge in mice Both mAbs had a similarhalf-life (58 and 100 days) in mouse serum after a singleintraperitoneal dose suggesting that mAbs delivered by thisroute at or around the time of VEEV infection persists athigh levels in the blood as well as secretions in respiratorytransudation throughout the clinical course of the disease Inparticular both mAbs administered 24 h prior to airbornechallenges had a substantial protective effect (90ndash100)Treatment of mice 2 or 24 h after airborne infection with asingle intraperitoneal dose of 100 120583g of 1A3A-9 mAb led toapproximately 50 survival There was no beneficial effectwhen mAb treatment was delayed to 72 h Moreover therewas evidence of synergy in vitro in PRNT between 1A3A-9and 1A4A-1 as has been demonstrated for other virusesand mAb pairs However no synergy was found in mouseprotection when mAbs were delivered intraperitoneally as amixture in equal parts to animals challenged 24 h previouslywith VEEV [129]
In a subsequent study Phillpotts et al reported two otherprotective mAbs The 3B2A-9 mAb protected against allthe serogroup I strains while 1A3B-7 mAb protected wellagainst challenge with all of the viruses tested Both 3B2A-9 and 1A3B-7 protected against airborne exposure to theIAB serogroup virus strainTrinidad donkey (TrD)Howeverthere was no evidence of synergistic protection when thesemAbs were combined in equal proportions An intraperi-toneal dose of 10 120583g was sufficient to protect 50 of themice with either mAb [130] According to other data themechanism of protection did not appear to depend uponneutralization [131] Similar results were obtained by OrsquoBrienet al which described a non-neutralizing mAb (IgG2a)named CUF37-2a able to protect 50 of mice from asubcutaneous VEEV challenge when a dose of 915 120583g of mAbwas administered 24 h prior to challenge [132]
In a following study the 1A3B-7 mAb was humanized(and following reported as Hu1A3B-7) maintaining the samefeatures of its murine counterpart In particular evaluation ofin vitro studies indicated that Hu1A3B-7 retained both broadspecificity and neutralizing activity Furthermore in vivoexperiments showed that Hu1A3B-7 successfully protectedmice against lethal subcutaneous and aerosol challenges withVEEV strain TrD Moreover the effectiveness of the human-ization process was determined by assessing proliferationresponses in human T-cells to peptides derived from the
BioMed Research International 15
murine and humanized versions of the VH and VL domainsThis analysis showed that the number of human T-cellepitopes within the humanized Ab had been substantiallyreduced indicating that Hu1A3B-7 may have low immuno-genicity in vivo [133]
Similarly Hunt et al described a humanizedmurinemAbderived from the 3B4C-4 mAb recognizing the E2c epitope(amino acid residues 182ndash209) and following named Hy4-26C showing neutralizing activity similar to the murinemAb Moreover it was protective (70ndash100 of survival) at100 ng dose in mice animal model intraperitoneal inoculatedand at 500120583g for intranasal challenge (80) Therapeuticstudies in mice revealed that Hy4-26C was able to cure miceup to 24 h following infection at 10 120583g on the contrary themouse mAb when given 1 hour after virus challenge [63]
In another work Hunt et al isolated two neutralizinghumanized Fabs F5 and L1A7 employing a blocking strategy[134 135] The anti-E2 specific F5 IgG had a 70 PRNTendpoint of 10 ngmL equivalent to that described for themost effective neutralizing anti-VEEVE2mAbs In particularthe E1-specific hFab L1A7 had a PRNT endpoint of 3120583gmL300-fold lower than F5
Further studies revealed that F5 epitope is located inthe 115ndash199 amino acid region Moreover F5 IgG had potentability to protect mice from infection by either route whenadministered 24 h before exposure however mice treated24 h and 48 h after aerosol exposure developed central ner-vous system infections but exhibited no clinical signs ofdisease [135]
Hu et al reported that passive immunization with thehumanized chimericmousemAbhu1A4A1IgG1-2A inmice at50 120583g 24 h before or after virulent VEEV challenge providedcomplete protection indicating that hu1A4A1IgG1-2A haspotent prophylactic and therapeutic effects against VEEVinfection [136]
Anti-VEEV mAbs isolated from a non-human primategene library has also been reported In particular thehumanized chimeric scFv-Fc ToR67-3B4 recognized viableas well as formalin and 120573-propiolactone inactivated virusparticles It detected specifically the viral E1 envelope proteinof VEEV but did not react with reduced viral glycopro-tein preparations suggesting that recognition depends uponconformational epitopes The recombinant Ab was able todetect multiple VEEV subtypes and displayed only marginalcross-reactivity to otherAlphavirus species except for Easternequine encephalitis virus (EEEV) In addition the scFv-Fcfusion described here might be of therapeutic use since itsuccessfully inactivated VEEV in a murine disease modelIn particular when the recombinant Ab was administered 6hours after challenge 80 to 100 of mice survived lethalVEEV IAB or IE infection Forty to sixty percent of micesurvived when scFv-Fc ToR67-3B4 was applied 6 hours afterchallenge with VEEV subtypes II and former IIIA [137]
4 Bunyaviridae
TheBunyaviridae family contains human arboviruses belong-ing to the Orthobunyavirus Phlebovirus and Nairovirus
genera Bunyaviridae includes enveloped viruses with a frag-mented single-stranded RNA genome of negative polarityTheir tripartite genome consists of a small (S) a medium(M) and a large (L) fragment and the envelope glycoproteinsGn and Gc that are cleaved out of a polyprotein synthesizedby the M fragment These glycoproteins have been provedto mediate the formation of the virus particle to play a rolein the interaction with cell surface receptors to mediate theentry of the virus into cells and to serve as targets for themajority of neutralizing Abs described so far
Orthobunyavirus transmitted through mosquitoes ormidges vectors are divided in 18 serogroups based on cross-titrations in haemagglutination inhibition assays and neutral-ization assays and correlating with main vector preferencesHowever the most clinically important viruses belong onlyto two serogroups the California encephalitis and the Simbuserogroups
Crimean-Congo hemorrhagic fever virus (CCHFV) isconsidered the only one of clinical relevance within theNairovirus genus which uses tick as main vector and is divid-ed in seven serogroups while Toscana virus and Rift Valleyfever virus (RVFV) which are transmitted by sandflies andmosquitoes respectively belong to the Phlebovirus genus[138]
MAbs against theNairovirusCCHFV and the PhlebovirusRVFV have been described
41 Crimean-Congo Hemorrhagic Fever Virus (CCHFV)Crimean-Congo Hemorrhagic Fever Virus (CCHFV) dis-tribution covers the greatest geographic range of any tick-borne virus known as viral isolation andor disease hasbeen reported from more than 30 countries in SoutheasternEurope Africa Asia and Middle East [139 140] Namelyfrom 1953 to 2010 about 6000 human cases were reportedin Southeastern Europe CCHFV causes sporadic outbreakswith mortality rates ranging from 10 to 80 [141 142] Asignificant variation in the time of incubation has beendescribed [139] Prehemorrhagic symptoms include highfever chills headache photophobia and back and abdominalpains Among other symptoms neuropsychiatric changeshave been reported in some patients In severe cases 3ndash6 daysafter the onset of the disease hemorrhagic symptoms occurwith petechiae ecchymosis bleeding in the form of melenahematemesis epistaxis [139]
In comparisonwith the nucleocapsid proteins inCCHFVGn and Gc show a higher degree of antigenic variabilityprobably due to their exposition to the host immune systemhypothesis corroborated in the description by Hewson et alin 2004 of four M segment phylogenic groups namely M1M2 M3 and M4 [141 142]
411 mAbs against CCHFV Blackburn et al firstly describedin 1987 murine anti-CCHFV mAbs able to recognize nucle-ocapsid proteins but no neutralizing activity has beendescribed thus far [143]
On the other hand Bertolotti-Ciarlet et al produced apanel of murine mAbs recognizing conformational epitopes
16 BioMed Research International
within theGn andGc glycoproteins expressed by the CCHFVstrain IbAr10200 The effectiveness of the mAbs was studiedby performing PRNT80 and none of the anti-Gn mAbsshowed neutralizing activity although many of the anti-GcmAbs neutralized IbAr10200 strain in vitro with clones 8A15E3 12A9 6C2 and 9H3 showing activity at a gt5120 dilutionMAbs 11E7 and 30F7 exhibited neutralizing activity at 2560dilution A suckling mice protection test was thereforeperformed for these mAbs The previously mentionedanti-Gc Abs were capable to protect mice to an appreciabledegree when applied 24 hours before and in a weakermanner 24 hours after virus challenge Many of the anti-GnmAbs were able to confer significant protection to IbAr10200both 24 hours before and after virus administration Therelevant effectiveness of the anti-Gn mAbs 6B12 10E1113G8 and 10G4 suggests that these mAbs may possess someneutralizing activity due to Ab-based effector mechanisms(eg ADCC) [58]
In further studies the same group characterized thebroadly cross-reactive neutralizing activity of the previouslydescribed murine mAb 11E7 able to bind conformationalepitopes within the C-terminal region of the Gn glycoproteinof all CCHFVM groups [58 141]
42 Rift Valley Fever Virus (RVFV) Similarly to CCHFV RiftValley Fever Virus (RVFV) causes sporadic outbreaks Thelargest epidemic occurred in Egypt in 1977 with an estimated200000 infections 18000 patients manifesting symptomsand 600 fatal prognoses [144] Symptoms include hemor-rhagic episodes fever encephalitis and blindness [145]
The major target of the humoral immune response is theRVFV glycoproteins Gc (G1) and Gn (G2) ranging fromnucleotides 480ndash2090 and 2091ndash3614 respectively [144]
421 mAbs against RVFV Within G2 glycoprotein are des-cribed three neutralizing antigenic sites namely epitope Inucleotides 792ndash893 (amino acid residues 258ndash291) epitopeII nucleotides 1164ndash1196 (amino acid residues 382ndash392) andepitope IV nucleotides 858ndash917 (amino acid residues 280ndash299) [59 145]
Despite the high degree of variability observed inCCHFVglycoproteins RVFV G2 presented an unexpected high con-servation among 22 isolates [59] In the same study onlythe LUNYO and 900060 isolates were resistant to in vitroneutralization by murine mAb 4D4 (recognizing epitope II)All other isolates were neutralized with PRNT80 titers from10240 togt81920On the other hand only SNS isolate showedreduced sensitivity to 4ndash39-CC (epitope IV) All other isolateswere neutralized with PRNT80 titers from 20480 to gt81920Protection studies on mice would be of interest for thedevelopment of a passive immunization prophylaxis [59]
Protection studies from lethal South African RVFVAN1830 strain infection were successfully performed on abattery of anti-G1 and anti-G2 mAbs produced by Besselaarand Blackburn with the strongly neutralizing anti-G1 mAb3E5 and anti-G2mAb 9C4 being the most effective [146]The9C4 antigenic area maps within G2 epitope I and a broadlyneutralizing activity may be investigated
5 Conclusions
Arbovirus infections have acquired increasing interest giventhe augmented globalization and tourism movement all overthe world that could be at the basis of epidemic eventsFurthermore the same vectors can sometimes transmit sev-eral arboviruses concomitantly complicating the diagnosis aswell as the therapy For example confusing mixed epidemicshave occasionally been described such as YFV plus CHIKVDENVplus CHIKV ormore recentlyPlasmodium falciparummalaria plus DENV1 and CHIKV on Madagascarrsquos east coast[1]
In the last decade several efforts have been employed inthe development of effective immunogens and therapeuticsgiving the current absence of specific antiviral drugs as wellas effective vaccines for themost diffused arbovirus infections(ie DENV andWNV)
However at the moment mosquito control is the bestavailable method for preventing arbovirus infections Thusto control the emerging public health problem of arbovirusinfections new antiviral therapeutic strategies that providepotent and broadly cross-protective immunity (especially forflavivirus infections) are an urgent globally medical need
As described in this review the majority of highlyneutralizing mAbs are of murine origin and indeed theirutility in the treatment of these infections is limited In facttheir employment can elicit an immunogenic response (iehuman anti-mouse Abs HAMA) the therapeutic efficacycould be reduced by a relatively faster clearance in humans(compared to human Abs) potentially exacerbated by theHAMA response Finally murine Abs exhibit relatively weakeffector functions (eg ADCC) compared to human AbsIndeed chimerization humanization or better the isolationand characterization of fully human broadly neutralizingmAbs would be the best choice for their possible use in afuture mAb-based therapy against arbovirus infections [147]
Finally possible ADE of infection mechanisms should beevaluated before considering a mAb as a possible candidatetherapeutic in a post-exposure setting as well as in the devel-opment of new Ab-eliciting immunogens
Conflict of Interests
The authors declare no conflict of interests in particular withOmriGam and any other cited Ab-based preparation men-tioned in the paper
References
[1] N Cleton M Koopmans J Reimerink G J Godeke and CReusken ldquoCome fly with me review of clinically importantarboviruses for global travelersrdquo Journal of Clinical Virology vol55 no 3 pp 191ndash203 2012
[2] F X Heinz H Holzmann A Essl and M Kundi ldquoField effec-tiveness of vaccination against tick-borne encephalitisrdquoVaccinevol 25 no 43 pp 7559ndash7567 2007
[3] A D Barrett and D E Teuwen ldquoYellow fever vaccinemdashhowdoes it work and why do rare cases of serious adverse events
BioMed Research International 17
take placerdquo Current Opinion in Immunology vol 21 no 3 pp308ndash313 2009
[4] S B Halstead and S J Thomas ldquoNew vaccines for Japaneseencephalitisrdquo Current Infectious Disease Reports vol 12 no 3pp 174ndash180 2010
[5] R Burioni M Perotti N Mancini and M Clementi ldquoPer-spectives for the utilization of neutralizing human monoclonalantibodies as anti-HCV drugsrdquo Journal of Hepatology vol 49no 2 pp 299ndash300 2008
[6] R A Diotti G A Sautto L Solforosi NManciniM Clementiand R Burioni ldquoNeutralization activity and kinetics of twobroad-range human monoclonal IgG1 derived from recombi-nant Fab fragments and directed against Hepatitis C virus E2glycoproteinrdquo New Microbiologica vol 35 no 4 pp 475ndash4792012
[7] R Burioni F Canducci N Mancini et al ldquoMolecular cloningof the first humanmonoclona antibodies neutralizing with highpotency Swine-origin Influenza A pandemic virus (S-OIV)rdquoNew Microbiologica vol 32 no 4 pp 319ndash324 2009
[8] P K Chandra S Hazari B Poat et al ldquoIntracytoplasmic stableexpression of IgG1 antibody targeting NS3 helicase inhibitsreplication of highly efficient hepatitis C Virus 2a clonerdquoVirology Journal vol 7 article 118 2010
[9] N Clementi D de Marco N Mancini et al ldquoA human mono-clonal antibody with neutralizing activity against highly diver-gent influenza subtypesrdquo PLoS ONE vol 6 no 12 Article IDe28001 2011
[10] D de Marco N Clementi N Mancini et al ldquoA non-VH1-69 heterosubtypic neutralizing human monoclonal antibodyprotects mice against H1N1 and H5N1 virusesrdquo PLoS ONE vol7 no 4 Article ID e34415 2012
[11] L Solforosi N Mancini F Canducci et al ldquoA phage displayvector optimized for the generation of human antibody com-binatorial libraries and the molecular cloning of monoclonalantibody fragmentsrdquoNewMicrobiologica vol 35 no 3 pp 289ndash294 2012
[12] E V Koonin and V V Dolja ldquoEvolution and taxonomy ofpositive-strand RNA viruses implications of comparative anal-ysis of amino acid sequencesrdquo Critical Reviews in Biochemistryand Molecular Biology vol 28 no 5 pp 375ndash430 1993
[13] J S Mackenzie D J Gubler and L R Petersen ldquoEmergingflaviviruses the spread and resurgence of Japanese encephalitisWest Nile and dengue virusesrdquo Nature Medicine vol 10 no 12pp S98ndashS109 2004
[14] T J SmithW E Brandt J L Swanson J MMcCown and E LBuescher ldquoPhysical and biological properties of dengue-2 virusand associated antigensrdquo Journal of Virology vol 5 no 4 pp524ndash532 1970
[15] J M Smit B Moesker I Rodenhuis-Zybert and J WilschutldquoFlavivirus cell entry and membrane fusionrdquo Viruses vol 3 no2 pp 160ndash171 2011
[16] R J Kuhn W Zhang M G Rossmann et al ldquoStructure ofdengue virus implications for flavivirus organization matura-tion and fusionrdquo Cell vol 108 no 5 pp 717ndash725 2002
[17] K Stiasny J H Aberle M Keller B Grubeck-Loebenstein andF X Heinz ldquoAge affects quantity but not quality of antibodyresponses after vaccination with an inactivated flavivirus vac-cine against tick-borne encephalitisrdquo PLoS ONE vol 7 no 3Article ID e34145 2012
[18] LThomasOVerlaetenACabie et al ldquoInfluence of the dengueserotype previous dengue infection and plasma viral load
on clinical presentation and outcome during a dengue-2 anddengue-4 co-epidemicrdquo American Journal of Tropical Medicineand Hygiene vol 78 no 6 pp 990ndash998 2008
[19] Y Zhang J Corver P R Chipman et al ldquoStructures of imma-ture flavivirus particlesrdquo EMBO Journal vol 22 no 11 pp2604ndash2613 2003
[20] A K I Falconar ldquoIdentification of an epitope on the denguevirus membrane (M) protein defined by cross-protective mon-oclonal antibodies design of an improved epitope sequencebased on common determinants present in both envelope (EandM) proteinsrdquo Archives of Virology vol 144 no 12 pp 2313ndash2330 1999
[21] K-J Huang Y-C Yang Y-S Lin et al ldquoThe dual-specific bind-ing of dengue virus and target cells for the antibody-dependentenhancement of dengue virus infectionrdquo Journal of Immunol-ogy vol 176 no 5 pp 2825ndash2832 2006
[22] A K I Falconar ldquoAntibody responses are generated to immun-odominant ELKKLE-typemotifs on the nonstructural-1 glyco-protein during live dengue virus infections inmice and humansimplications for diagnosis pathogenesis and vaccine designrdquoClinical and Vaccine Immunology vol 14 no 5 pp 493ndash5042007
[23] C W Mandl F Guirakhoo H Holzmann F X Heinz and CKunz ldquoAntigenic structure of the flavivirus envelope protein Eat the molecular level using the tick-borne encephalitis virus asa modelrdquo Journal of Virology vol 63 no 2 pp 564ndash571 1989
[24] F A Rey F X Heinz C Mandl C Kunz and S C HarrisonldquoThe envelope glycoprotein from tick-borne encephalitis virusat 2 A resolutionrdquo Nature vol 375 no 6529 pp 291ndash298 1995
[25] S L Allison K Stiasny K Stadler CWMandl and F XHeinzldquoMapping of functional elements in the stem-anchor regionof tick-borne encephalitis virus envelope protein Erdquo Journal ofVirology vol 73 no 7 pp 5605ndash5612 1999
[26] D E Klein J L Choi and S C Harrison ldquoStructure of a denguevirus envelope protein late-stage fusion intermediaterdquo Journal ofVirology vol 87 no 4 pp 2287ndash2293 2013
[27] S L Hanna T C Pierson M D Sanchez A A Ahmed M MMurtadha and R W Doms ldquoN-linked glycosylation of WestNile virus envelope proteins influences particle assembly andinfectivityrdquo Journal of Virology vol 79 no 21 pp 13262ndash132742005
[28] N Mancini G Sautto N Clementi et al ldquoNeutralization inter-fering antibodies a ldquonovelrdquo example of humoral immune dys-function facilitating viral escaperdquoViruses vol 4 no 9 pp 1731ndash1752 2012
[29] Y Modis S Ogata D Clements and S C Harrison ldquoVariablesurface epitopes in the crystal structure of dengue virus type3 envelope glycoproteinrdquo Journal of Virology vol 79 no 2 pp1223ndash1231 2005
[30] N Miller ldquoRecent progress in dengue vaccine research anddevelopmentrdquo Current Opinion in Molecular Therapeutics vol12 no 1 pp 31ndash38 2010
[31] J H Chavez J R Silva A A Amarilla and L T Moraes Figu-eiredo ldquoDomain III peptides from flavivirus envelope proteinare useful antigens for serologic diagnosis and targets forimmunizationrdquo Biologicals vol 38 no 6 pp 613ndash618 2010
[32] B Shrestha J D Brien S Sukupolvi-Petty et al ldquoThe develop-ment of therapeutic antibodies that neutralize homologous andheterologous genotypes of dengue virus type 1rdquo PLoS Pathogensvol 6 no 4 Article ID e1000823 2010
18 BioMed Research International
[33] G D Gromowski N D Barrett and A D T Barrett ldquoChar-acterization of dengue virus complex-specific neutralizing epi-topes on envelope protein domain III of dengue 2 virusrdquo Journalof Virology vol 82 no 17 pp 8828ndash8837 2008
[34] S Sukupolvi-Petty S K Austin W E Purtha et al ldquoType- andsubcomplex-specific neutralizing antibodies against domain IIIof dengue virus type 2 envelope protein recognize adjacentepitopesrdquo Journal of Virology vol 81 no 23 pp 12816ndash128262007
[35] S Sukupolvi-Petty S K Austin M Engle et al ldquoStructure andfunction analysis of therapeutic monoclonal antibodies againstdengue virus type 2rdquo Journal of Virology vol 84 no 18 pp9227ndash9239 2010
[36] Y-Q Deng J-X Dai G-H Ji et al ldquoA broadly flaviviruscross-neutralizingmonoclonal antibody that recognizes a novelepitope within the fusion loop of e proteinrdquo PLoS ONE vol 6no 1 Article ID e16059 2011
[37] J M Costin E Zaitseva K M Kahle et al ldquoMechanistic studyof broadly neutralizing human monoclonal antibodies againstdengue virus that target the fusion looprdquo Journal of Virologyvol 87 no 1 pp 52ndash66 2013
[38] CMMidgley A FlanaganH B Tran et al ldquoStructural analysisof a dengue cross-reactive antibody complexed with envelopedomain III reveals the molecular basis of cross-reactivityrdquoJournal of Immunology vol 188 no 10 pp 4971ndash4979 2012
[39] N J Moreland P Susanto E Lim et al ldquoPhage display appro-aches for the isolation of monoclonal antibodies against denguevirus envelope domain III from human and mouse derivedlibrariesrdquo International Journal ofMolecular Sciences vol 13 no3 pp 2618ndash2635 2012
[40] J J B Cockburn M E Navarro Sanchez N Fretes et alldquoMechanism of dengue virus broad cross-neutralization by amonoclonal antibodyrdquo Structure vol 20 no 2 pp 303ndash3142012
[41] R Rajamanonmani C Nkenfou P Clancy et al ldquoOn a mousemonoclonal antibody that neutralizes all four dengue virusserotypesrdquo Journal of General Virology vol 90 no 4 pp 799ndash809 2009
[42] L H Gould J Sui H Foellmer et al ldquoProtective and herapeuticcapacity of human single-chain Fv-Fc fusion proteins againstWest Nile virusrdquo Journal of Virology vol 79 no 23 pp 14606ndash14613 2005
[43] H Sultana H G Foellmer G Neelakanta et al ldquoFusion LoopPeptide of the West Nile virus envelope protein is essential forpathogenesis and is recognized by a therapeutic cross-reactivehuman monoclonal antibodyrdquo Journal of Immunology vol 183no 1 pp 650ndash660 2009
[44] T Oliphant M Engle G E Nybakken et al ldquoDevelopment ofa humanized monoclonal antibody with therapeutic potentialagainstWest Nile virusrdquoNature Medicine vol 11 no 5 pp 522ndash530 2005
[45] G E Nybakken T Oliphant S Johnson S Burke M S Dia-mond and D H Fremont ldquoStructural basis of West Nile virusneutralization by a therapeutic antibodyrdquo Nature vol 437 no7059 pp 764ndash769 2005
[46] G D Gromowski J T Roehrig M S Diamond J C LeeT J Pitcher and A D T Barrett ldquoMutations of an antibodybinding energy hot spot on domain III of the dengue 2 envelopeglycoprotein exploited for neutralization escaperdquo Virology vol407 no 2 pp 237ndash246 2010
[47] J T Roehrig R A Bolin andR G Kelly ldquoMonoclonal antibodymapping of the envelope glycoprotein of the dengue 2 virusJamaicardquo Virology vol 246 no 2 pp 317ndash328 1998
[48] R de Alwis S A Smith N P Olivarez et al ldquoIdentification ofhumanneutralizing antibodies that bind to complex epitopes ondengue virionsrdquo Proceedings of the National Academy of Sciencesof the United States of America vol 109 no 19 pp 7439ndash74442012
[49] S K Austin K A Dowd B Shrestha et al ldquoStructural basis ofdifferential neutralization of DENV-1 genotypes by an antibodythat recognizes a cryptic epitoperdquo PLoS Pathogens vol 8 no 10Article ID e1002930 2012
[50] M Throsby C Geuijen J Goudsmit et al ldquoIsolation and char-acterization of human monoclonal antibodies from individualsinfected with West Nile Virusrdquo Journal of Virology vol 80 no14 pp 6982ndash6992 2006
[51] A P Goncalvez R H Purcell and C-J Lai ldquoEpitope deter-minants of a chimpanzee Fab antibody that efficiently cross-neutralizes dengue type 1 and type 2 viruses map to inside andin close proximity to fusion loop of the dengue type 2 virusenvelope glycoproteinrdquo Journal of Virology vol 78 no 23 pp12919ndash12928 2004
[52] J T Roehrig J H Mathews and D W Trent ldquoIdentification ofepitopes on the E glycoprotein of Saint Louis encephalitis virususing monoclonal antibodiesrdquo Virology vol 128 no 1 pp 118ndash126 1983
[53] J T Roehrig L A Staudinger A R Hunt J H Mathews andC D Blair ldquoAntibody prophylaxis and therapy for flavivirusencephalitis infectionsrdquo Annals of the New York Academy ofSciences vol 951 pp 286ndash297 2001
[54] A P Goncalvez C-H Chien K Tubthong et al ldquoHumanizedmonoclonal antibodies derived from chimpanzee fabs protectagainst Japanese encephalitis virus in vitro and in vivordquo Journalof Virology vol 82 no 14 pp 7009ndash7021 2008
[55] M Arakawa T Yamashiro G-I Uechi M Tadano and ANishizono ldquoConstruction of humanFab (1205741120581) library and iden-tification of human monoclonal Fab possessing neutralizingpotency against Japanese encephalitis virusrdquo Microbiology andImmunology vol 51 no 6 pp 617ndash625 2007
[56] L Warter C Y Lee R Thiagarajan et al ldquoChikungunya virusenvelope-specific human monoclonal antibodies with broadneutralization potencyrdquo Journal of Immunology vol 186 no 5pp 3258ndash3264 2011
[57] P Pal K A Dowd J D Brien et al ldquoDevelopment of a highlyprotective combination monoclonal antibody therapy againstchikungunya virusrdquo PLoS Pathogens vol 9 no 4 Article IDe1003312 2013
[58] A Bertolotti-Ciarlet J Smith K Strecker et al ldquoCellular local-ization and antigenic characterization of Crimean-Congo hem-orrhagic fever virus glycoproteinsrdquo Journal of Virology vol 79no 10 pp 6152ndash6161 2005
[59] J K Battles and J M Dalrymple ldquoGenetic variation amonggeographic isolates of Rift Valley fever virusrdquo American Journalof Tropical Medicine and Hygiene vol 39 no 6 pp 617ndash6311988
[60] B AThibodeaux N C Garbino NM Liss et al ldquoA humanizedIgG but not IgM antibody is effective in prophylaxis and therapyof yellow fever infection in an AG12917D-204 peripheralchallenge mouse modelrdquo Antiviral Research vol 94 no 1 pp1ndash8 2012
BioMed Research International 19
[61] S Daffis R E Kontermann J Korimbocus H Zeller H-DKlenk and J Ter Meulen ldquoAntibody responses against wild-type yellow fever virus and the 17D vaccine strain charac-terization with human monoclonal antibody fragments andneutralization escape variantsrdquoVirology vol 337 no 2 pp 262ndash272 2005
[62] L N Levanov L E Matveev E P Goncharova et al ldquoChimericantibodies against tick-borne encephalitis virusrdquo Vaccine vol28 no 32 pp 5265ndash5271 2010
[63] A R Hunt S Frederickson C Hinkel K S Bowdish and J TRoehrig ldquoA humanized murine monoclonal antibody protectsmice either before or after challenge with virulent Venezuelanequine encephalomyelitis virusrdquo Journal of General Virologyvol 87 no 9 pp 2467ndash2476 2006
[64] J M da Silva Voorham I A Rodenhuis-Zybert N V A Nunezet al ldquoAntibodies against the envelope glycoprotein promoteinfectivity of immature dengue virus serotype 2rdquo PLoS ONEvol 7 no 3 Article ID e29957 2012
[65] T C Pierson Q Xu S Nelson et al ldquoThe stoichiometry ofantibody-mediated neutralization and enhancement of westnile virus infectionrdquoCell Host andMicrobe vol 1 no 2 pp 135ndash145 2007
[66] G Sautto N Mancini R A Diotti L Solforosi M Clementiand R Burioni ldquoAnti-hepatitis C virus E2 (HCVE2) glyco-proteinmonoclonal antibodies andneutralization interferencerdquoAntiviral Research vol 96 no 1 pp 82ndash89 2012
[67] R Burioni Y Matsuura N Mancini et al ldquoDiverging effectsof human recombinant anti-hepatitis C virus (HCV) antibodyfragments derived from a single patient on the infectivityof a vesicular stomatitis virusHCV pseudotyperdquo Journal ofVirology vol 76 no 22 pp 11775ndash11779 2002
[68] R J Phillpotts J R Stephenson and J S Porterfield ldquoAntibody-dependent enhancement of tick-borne encephalitus virus infec-tivityrdquo Journal of General Virology vol 66 no 8 pp 1831ndash18371985
[69] K A Dowd and T C Pierson ldquoAntibody-mediated neutraliza-tion of flaviviruses a reductionist viewrdquo Virology vol 411 no 2pp 306ndash315 2011
[70] A K I Falconar ldquoMonoclonal antibodies that bind to commonepitopes on the dengue virus type 2 nonstructural-1 andenvelope glycoproteins display weak neutralizing activity anddifferentiated responses to virulent strains implications forpathogenesis and vaccinesrdquo Clinical and Vaccine Immunologyvol 15 no 3 pp 549ndash561 2008
[71] K Meyer A Banerjee S E Frey R B Belshe and R Ray ldquoAweak neutralizing antibody response to hepatitis C virus enve-lope glycoprotein enhances virus infectionrdquo PLoS ONE vol 6no 8 Article ID e23699 2011
[72] R Burioni R AWilliamson P P Sanna F E Bloom and D RBurton ldquoRecombinant human Fab to glycoprotein D neutral-izes infectivity and prevents cell-to-cell transmission of herpessimplex viruses 1 and 2 in vitrordquo Proceedings of the NationalAcademy of Sciences of the United States of America vol 91 no1 pp 355ndash359 1994
[73] M S Rolph A Zaid N E Rulli and S Mahalingam ldquoDown-regulation of interferon-120573 in antibody-dependent enhancementof dengue viral infections of human macrophages is dependenton interleukin-6rdquo Journal of Infectious Diseases vol 204 no 3pp 489ndash491 2011
[74] S S Whitehead J E Blaney A P Durbin and B R MurphyldquoProspects for a dengue virus vaccinerdquoNature ReviewsMicrobi-ology vol 5 no 7 pp 518ndash528 2007
[75] S-M Lok V Kostyuchenko G ENybakken et al ldquoBinding of aneutralizing antibody to dengue virus alters the arrangement ofsurface glycoproteinsrdquoNature Structural andMolecular Biologyvol 15 no 3 pp 312ndash317 2008
[76] A P Goncalvez R Men C Wernly R H Purcell and C-J Lai ldquoChimpanzee Fab fragments and a derived humanizedimmunogiobulin G1 antibody that efficiently cross-neutralizedengue type 1 and type 2 virusesrdquo Journal of Virology vol 78no 23 pp 12910ndash12918 2004
[77] WM P BWahala A A Kraus L B HaymoreM A Accavitti-Loper and A M de Silva ldquoDengue virus neutralization byhuman immune sera role of envelope protein domain III-reactive antibodyrdquo Virology vol 392 no 1 pp 103ndash113 2009
[78] W D Crill H R Hughes M J Delorey and G-J J ChangldquoHumoral immune responses of dengue fever patients usingepitope-specific serotype-2 virus-like particle antigensrdquo PLoSONE vol 4 no 4 Article ID e4991 2009
[79] C Setthapramote T Sasaki O Puiprom et al ldquoHuman mono-clonal antibodies to neutralize all dengue virus serotypes usinglymphocytes from patients at acute phase of the secondaryinfectionrdquo Biochemical and Biophysical Research Communica-tions vol 423 no 4 pp 867ndash872 2012
[80] M Beltramello K L Williams C P Simmons et al ldquoThehuman immune response to dengue virus is dominated byhighly cross-reactive antibodies endowed with neutralizing andenhancing activityrdquo Cell Host andMicrobe vol 8 no 3 pp 271ndash283 2010
[81] C M Midgley M Bajwa-Joseph S Vasanawathana et al ldquoAnin-depth analysis of original antigenic sin in dengue virusinfectionrdquo Journal of Virology vol 85 no 22 pp 410ndash421 2011
[82] B Kaufmann M R Vogt J Goudsmit et al ldquoNeutralizationof West Nile virus by cross-linking of its surface proteins withFab fragments of the human monoclonal antibody CR4354rdquoProceedings of the National Academy of Sciences of the UnitedStates of America vol 107 no 44 pp 18950ndash18955 2010
[83] B Spurrier J M Sampson M Totrov et al ldquoStructural analysisof human and macaque mAbs 2909 and 25B implicationsfor the configuration of the quaternary neutralizing epitope ofHIV-1 gp120rdquo Structure vol 19 no 5 pp 691ndash699 2011
[84] J J SchlesingerMWBrandriss andT PMonath ldquoMonoclonalantibodies distinguish between wild and vaccine strains ofyellow fever virus by neutralization hemagglutination inhibi-tion and immune precipitation of the virus envelope proteinrdquoVirology vol 125 no 1 pp 8ndash17 1983
[85] M W Brandriss J J Schlesinger E E Walsh and M BrisellildquoLethal 17D yellow fever encephalitis in mice I Passive protec-tion by monoclonal antibodies to the envelope proteins of 17Dyellow fever and dengue 2 virusesrdquo Journal of General Virologyvol 67 no 2 pp 229ndash234 1986
[86] H Cho and M S Diamond ldquoImmune responses to West Nilevirus infection in the central nervous systemrdquo Viruses vol 4no 12 pp 3812ndash3830 2012
[87] D W Beasley A D Barrett and R B Tesh ldquoResurgence ofWest Nile neurologic disease in the United States in 2012 whathappened What needs to be donerdquo Antiviral Research vol 99no 1 pp 1ndash5 2013
[88] M Throsby J Ter Meulen C Geuijen J Goudsmit and J DeKruif ldquoMapping and analysis of West Nile virus-specific mon-oclonal antibodies prospects for vaccine developmentrdquo ExpertReview of Vaccines vol 6 no 2 pp 183ndash191 2007
[89] M S Diamond T G Roberts D Edgil B Lu J Ernst and EHarris ldquoModulation of dengue virus infection in human cells
20 BioMed Research International
by alpha beta and gamma interferonsrdquo Journal of Virology vol74 no 11 pp 4957ndash4966 2000
[90] Z Shimoni M J Niven S Pitlick and S Bulvik ldquoTreatmentof West Nile virus encephalitis with intravenous immunoglob-ulinrdquo Emerging Infectious Diseases vol 7 no 4 article 759 2001
[91] AHamdan PGreen EMendelsonM R Kramer S Pitlik andM Weinberger ldquoPossible benefit of intravenous immunoglob-ulin therapy in a lung transplant recipient with West Nile virusencephalitisrdquoTransplant InfectiousDisease vol 4 no 3 pp 160ndash162 2002
[92] M Haley A S Retter D Fowler J Gea-Banacloche and NP OrsquoGrady ldquoThe role for intravenous immunoglobulin in thetreatment of West Nile virus encephalitisrdquo Clinical InfectiousDiseases vol 37 no 6 pp e88ndashe90 2003
[93] K M Chung G E Nybakken B S Thompson et al ldquoAnti-bodies against West Nile virus nonstructural protein NS1prevent lethal infection through Fc 120574 receptor-dependent and-independent mechanismsrdquo Journal of Virology vol 80 no 3pp 1340ndash1351 2006
[94] E A Henchal L S Henchal and J J Schlesinger ldquoSynergisticinteractions of anti-NS1 monoclonal antibodies protect pas-sively immunized mice from lethal challenge with dengue 2virusrdquo Journal of General Virology vol 69 no 8 pp 2101ndash21071988
[95] J T Roehrig ldquoAntigenic structure of flavivirus proteinsrdquoAdvances in Virus Research vol 59 pp 141ndash175 2003
[96] M D Sanchez T C Pierson D McAllister et al ldquoCharacteri-zation of neutralizing antibodies to West Nile virusrdquo Virologyvol 336 no 1 pp 70ndash82 2005
[97] J H Beigel J L Nordstrom S R Pillemer et al ldquoSafety andpharmacokinetics of single intravenous dose of MGAWN1 anovel monoclonal antibody to West Nile virusrdquo AntimicrobialAgents and Chemotherapy vol 54 no 6 pp 2431ndash2436 2010
[98] C A Smeraski V Siddharthan and J D Morrey ldquoTreatmentof spatial memory impairment in hamsters infected with WestNile virus using a humanizedmonoclonal antibodyMGAWN1rdquoAntiviral Research vol 91 no 1 pp 43ndash49 2011
[99] D Lelli AMoreno E Brocchi et al ldquoWestNile virus character-ization and diagnostic applications of monoclonal antibodiesrdquoVirology Journal vol 9 article 81 2012
[100] T S Gritsun V A Lashkevich and E A Gould ldquoTick-borneencephalitisrdquo Antiviral Research vol 57 no 1-2 pp 129ndash1462003
[101] D Ruzek T S Gritsun N L Forrester et al ldquoMutations inthe NS2B and NS3 genes affect mouse neuroinvasiveness of aWestern European field strain of tick-borne encephalitis virusrdquoVirology vol 374 no 2 pp 249ndash255 2008
[102] M Ecker S L Allison T Meixner and F X Heinz ldquoSequenceanalysis and genetic classification of tick-borne encephalitisviruses from Europe and Asiardquo Journal of General Virology vol80 no 1 pp 179ndash185 1999
[103] H Holzmann M S Vorobyova I P Ladyzhenskaya et alldquoMolecular epidemiology of tick-borne encephalitis viruscross-protection between European and Far Eastern subtypesrdquoVaccine vol 10 no 5 pp 345ndash349 1992
[104] C W Mandl ldquoSteps of the tick-borne encephalitis virus repli-cation cycle that affect neuropathogenesisrdquo Virus Research vol111 no 2 pp 161ndash174 2005
[105] A V Atrasheuskaya T M Fredeking and G M IgnatyevldquoChanges in immune parameters and their correction in humancases of tick-borne encephalitisrdquo Clinical and ExperimentalImmunology vol 131 no 1 pp 148ndash154 2003
[106] K L Mansfield N Johnson L P Phipps J R Stephenson AR Fooks and T Solomon ldquoTick-borne encephalitis virusmdashareview of an emerging zoonosisrdquo Journal of General Virologyvol 90 no 8 pp 1781ndash1794 2009
[107] DWVaughn andCHHoke Jr ldquoThe epidemiology of Japaneseencephalitis prospects for preventionrdquo Epidemiologic Reviewsvol 14 pp 197ndash221 1992
[108] T Solomon H Ni D W C Beasley M Ekkelenkamp M JCardosa and A D T Barrett ldquoOrigin and evolution of Japaneseencephalitis virus in Southeast Asiardquo Journal of Virology vol 77no 5 pp 3091ndash3098 2003
[109] M R Wilson ldquoEmerging viral infectionsrdquo Current Opinion inNeurology vol 26 no 3 pp 301ndash306 2013
[110] J N Hanna S A Ritchie D A Phillips et al ldquoAn outbreakof Japanese encephalitis in the Torres Strait Australia 1995rdquoMedical Journal of Australia vol 165 no 5 pp 256ndash260 1996
[111] J N Hanna S A Ritchie D A Phillips et al ldquoJapanese ence-phalitis in north Queensland Australia 1998rdquo Medical Journalof Australia vol 170 no 11 pp 533ndash536 1999
[112] SGeorgeMGourie-Devi and J A Rao ldquoIsolation ofWestNilevirus from the brains of children who had died of encephalitisrdquoBulletin of theWorldHealthOrganization vol 62 no 6 pp 879ndash882 1984
[113] A K Gupta A A Koshy and V J Lad ldquoEnhanced protec-tion of mice against Japanese encephalitis virus infection bycombinations ofmonoclonal antibodies to glycoprotein ErdquoActaVirologica vol 55 no 2 pp 165ndash168 2011
[114] T H Lee B-H Song S-I Yun et al ldquoA cross-protective mAbrecognizes a novel epitope within the flavivirus NS1 proteinrdquoJournal of General Virology vol 93 no 1 pp 20ndash26 2012
[115] S C Weaver ldquoHost range amplification and arboviral diseaseemergencerdquo Archives of Virology Supplementum no 19 pp 33ndash44 2005
[116] M Solignat B Gay S Higgs L Briant and C Devaux ldquoRepli-cation cycle of chikungunya a re-emerging arbovirusrdquoVirologyvol 393 no 2 pp 183ndash197 2009
[117] O Schwartz and M L Albert ldquoBiology and pathogenesis ofchikungunya virusrdquo Nature Reviews Microbiology vol 8 no 7pp 491ndash500 2010
[118] W Akahata Z-Y Yang H Andersen et al ldquoA virus-like particlevaccine for epidemic Chikungunya virus protects nonhumanprimates against infectionrdquo Nature Medicine vol 16 no 3 pp334ndash338 2010
[119] M Kielian and A Helenius ldquopH-Iinduced alterations in thefusogenic spike protein of Semliki Forest virusrdquo Journal of CellBiology vol 101 no 6 pp 2284ndash2291 1985
[120] J White and A Helenius ldquopH-Dependent fusion between theSemliki Forest virus membrane and liposomesrdquo Proceedings ofthe National Academy of Sciences of the United States of Americavol 77 no 6 pp 3273ndash3277 1980
[121] J Dubuisson and C M Rice ldquoSindbis virus attachment isola-tion and characterization of mutants with impaired binding tovertebrate cellsrdquo Journal of Virology vol 67 no 6 pp 3363ndash33741993
[122] C Y Lee Y-W Kam J Fric et al ldquoChikungunya virus neutral-ization antigens and direct cell-to-cell transmission are revealedby human antibody-escapemutantsrdquo PLoS Pathogens vol 7 no12 Article ID e1002390 2011
[123] J E Voss M-C Vaney S Duquerroy et al ldquoGlycoproteinorganization of Chikungunya virus particles revealed by X-raycrystallographyrdquo Nature vol 468 no 7324 pp 709ndash712 2010
BioMed Research International 21
[124] L Li J Jose Y Xiang R J Kuhn and M G Rossmann ldquoStruc-tural changes of envelope proteins during alphavirus fusionrdquoNature vol 468 no 7324 pp 705ndash708 2010
[125] J Fric S Bertin-Maghit C I Wang A Nardin and L WarterldquoUse of human monoclonal antibodies to treat Chikungunyavirus infectionrdquo Journal of Infectious Diseases vol 207 no 2 pp319ndash322 2013
[126] P V Aguilar J G Estrada-Franco R Navarro-Lopez C FerroA D Haddow and S C Weaver ldquoEndemic Venezuelanequine encephalitis in the Americas hidden under the dengueumbrellardquo Future Virology vol 6 no 6 pp 721ndash740 2011
[127] S C Weaver R Winegar I D Manger and N L ForresterldquoAlphaviruses population genetics and determinants of emer-gencerdquo Antiviral Research vol 94 no 3 pp 242ndash257 2012
[128] S Paessler and S C Weaver ldquoVaccines for Venezuelan equineencephalitisrdquo Vaccine vol 27 no 4 pp D80ndashD85 2009
[129] R J Phillpotts ldquoVenezuelan equine encephalitis virus complex-specific monoclonal antibody provides broad protection inmurine models against airborne challenge with viruses fromserogroups I II and IIIrdquoVirus Research vol 120 no 1-2 pp 107ndash112 2006
[130] R J Phillpotts L D Jones and S C Howard ldquoMonoclonalantibody protectsmice against infection anddiseasewhen giveneither before or up to 24 h after airborne challenge with virulentVenezuelan equine encephalitis virusrdquoVaccine vol 20 no 11-12pp 1497ndash1504 2002
[131] J H Mathews J T Roehrig and D W Trent ldquoRole of com-plement and the Fc portion of immunoglobulin G in immunityto Venezuelan equine encephalomyelitis virus infection withglycoprotein-specific monoclonal antibodiesrdquo Journal of Virol-ogy vol 55 no 3 pp 594ndash600 1985
[132] L M OrsquoBrien C D Underwood-Fowler S A Goodchild AL Phelps and R J Phillpotts ldquoDevelopment of a novel mon-oclonal antibody with reactivity to a wide range of Venezuelanequine encephalitis virus strainsrdquoVirology Journal vol 6 article206 2009
[133] L M OrsquoBrien S A Goodchild R J Phillpotts and S D Per-kins ldquoA humanised murine monoclonal antibody protectsmice from Venezuelan equine encephalitis virus Evergladesvirus and Mucambo virus when administered up to 48h afterairborne challengerdquo Virology vol 426 no 2 pp 100ndash105 2012
[134] N Clementi N Mancini L Solforosi M Castelli M Clementiand R Burioni ldquoPhage display-based strategies for cloning andoptimization of monoclonal antibodies directed against humanpathogensrdquo International Journal of Molecular Sciences vol 13no 7 pp 8273ndash8292 2012
[135] A R Hunt R A Bowen S Frederickson T Maruyama JT Roehrig and C D Blair ldquoTreatment of mice with humanmonoclonal antibody 24h after lethal aerosol challenge withvirulent Venezuelan equine encephalitis virus prevents diseasebut not infectionrdquo Virology vol 414 no 2 pp 146ndash152 2011
[136] W-G Hu A L Phelps S Jager et al ldquoA recombinant human-ized monoclonal antibody completely protects mice againstlethal challenge with Venezuelan equine encephalitis virusrdquoVaccine vol 28 no 34 pp 5558ndash5564 2010
[137] T Rulker L Voss P Thullier et al ldquoIsolation and characterisa-tion of a human-like antibody fragment (scFv) that inactivatesVEEV in vitro and in vivordquo PLoS ONE vol 7 no 5 Article IDe37242 2012
[138] C T Walter and J N Barr ldquoRecent advances in the molecularand cellular biology of bunyavirusesrdquo Journal of General Virol-ogy vol 92 no 11 pp 2467ndash2484 2011
[139] C A Whitehouse ldquoCrimean-Congo hemorrhagic feverrdquo Anti-viral Research vol 64 no 3 pp 145ndash160 2004
[140] M Mertens K Schmidt A Ozkul and M H Groschup ldquoTheimpact of Crimean-Congo hemorrhagic fever virus on publichealthrdquo Antiviral Research vol 98 no 2 pp 248ndash260 2013
[141] A A Ahmed J M McFalls C Hoffmann et al ldquoPresence ofbroadly reactive and group-specific neutralizing epitopes onnewly described isolates of Crimean-Congo hemorrhagic fevervirusrdquo Journal of General Virology vol 86 no 12 pp 3327ndash33362005
[142] R Hewson A Gmyl L Gmyl et al ldquoEvidence of segment reas-sortment in Crimean-Congo haemorrhagic fever virusrdquo Journalof General Virology vol 85 no 10 pp 3059ndash3070 2004
[143] N K Blackburn T G Besselaar A J Shepherd and R Swane-poel ldquoPreparation and use of monoclonal antibodies for iden-tifying Crimean-Congo hemorrhagic fever virusrdquo AmericanJournal of Tropical Medicine and Hygiene vol 37 no 2 pp 392ndash397 1987
[144] T Ikegami ldquoMolecular biology and genetic diversity of RiftValley fever virusrdquoAntiviral Research vol 95 no 3 pp 293ndash3102012
[145] KKeegan andM S Collett ldquoUse of bacterial expression cloningto define the amino acid sequences of antigenic determinantson the G2 glycoprotein of Rift Valley fever virusrdquo Journal ofVirology vol 58 no 2 pp 263ndash270 1986
[146] T G Besselaar and N K Blackburn ldquoTopological mapping ofantigenic sites on the Rift Valley fever virus envelope glycopro-teins using monoclonal antibodiesrdquo Archives of Virology vol121 no 1ndash4 pp 111ndash124 1991
[147] L G Presta ldquoEngineering of therapeutic antibodies to mini-mize immunogenicity and optimize functionrdquo Advanced DrugDelivery Reviews vol 58 no 5-6 pp 640ndash656 2006
Submit your manuscripts athttpwwwhindawicom
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anatomy Research International
PeptidesInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporation httpwwwhindawicom
International Journal of
Volume 2014
Zoology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Molecular Biology International
GenomicsInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioinformaticsAdvances in
Marine BiologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Signal TransductionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Evolutionary BiologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Biochemistry Research International
ArchaeaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Genetics Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in
Virolog y
Hindawi Publishing Corporationhttpwwwhindawicom
Nucleic AcidsJournal of
Volume 2014
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Enzyme Research
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Microbiology

BioMed Research International 9
The structure of the 1A1D-2 bound to EDIII indicates that themAb binds to sites that are transiently exposed during viralldquobreathingrdquo at 37∘C and block infection during attachmentof the virion to the cell In particular this mAb neutralizesDENV1 2 and 3 serotypes and residues K307 and K310 arethemost critical residues for binding of 1A1D-2mAb [46 75]
Finally Shrestha et al immunized mice with a genotype2 strain of DENV1 virus and generated 79 new mAbs 16of which strongly inhibited infection by the homologousvirus and localized to DIII of E protein Surprisingly onlytwo mAbs E105 and E106 retained strong binding andneutralizing activity against all five DENV1 genotypes as wellas being protective in immunocompromised infected miceMoreover E105 and E106 exhibited therapeutic activity evenwhen administered as a single dose four days after inoculationwith a heterologous genotype 4 strain of DENV1 in the samemouse model [32]
243 Other Non-HumanmAbs Other non-human and non-murine as well as humanized mAbs directed against the Eglycoprotein have been described In this regard Goncalvezet al isolated a large panel of anti-E Fab fragments from chim-panzees infected with all four DENV serotypes Howeveronly a limited number of them displayed a cross-neutralizingactivity against DENV1 and 2 and to a lesser extent againstDENV3 and 4 In particular the authors calculated thatamong them the 1A5 mAb that was further humanizedneutralized DENV1ndash4 at a PRNT50 titer of 048 095 32and 43 120583gmL respectively Interestingly the humanized 1A5was also tested for binding and neutralization against theWNVDENV4 chimera JEV strain SA14-14-2 and Langatvirus (LGTV) strain TP 21 giving a PRNT50 titer of 38 21and 28120583gmL respectively Moreover the authors calculatedthat when administered in a dose of 2mg per kg of bodyweight 1A5 would give a serum titer of approximately 40and 20 50 reduction in plaque reduction neutralization test(PRNT50) against DENV1 and DENV2 respectively [76]Furthermore epitope mapping of 1A5 mAb localized to G106within the Flavivirus-conserved fusion loop inDII of DENV2E protein [51]
244 Anti-DENV Human mAbs As anticipated attemptsto isolate neutralizing human mAbs have been morechallenging due to the restricted elicited Ab-repertoire whichrecognizes DIII of E glycoprotein in naıve as well as ininfected patients [77 78] However several groups reportedthe isolation of neutralizing mAbs from infected subjects Inthis regard Setthapramote et al isolated a total of 136 humanhybridoma clones producing specific mAbs against DENVobtained using PBMCs from nine blood samples from fouracute-phase patients secondarily infected with DENV2 andfive convalescent-phase patients Interestingly the authorsfound that most of the acute-phase mAb clones were cross-reactive with all four DENV serotypes with most of themrecognizing the E protein and endowing of neutralizingactivity against all DENV serotypes compared to thosederived from convalescent-phase patients In particularfrom the acute-phase PBMCs 818 were anti-E 66 wereanti-prM and 33 were anti-NS1 while 133 anti-E 133
anti-prM and 533 anti-NS1 mAbs clones were obtainedfrom convalescent-phase PBMCs [79] Previous studiesconfirmed the data obtained by Setthapramote et al onconvalescent-phase patients but this group firstly reportedthe efficient preparation of human mAbs with strongneutralizing activity titers against all four DENV serotypesusing PBMCs from acute-phase patients secondarily infectedwith DENV [80 81]
The group of de Alwis through immunoglobulin deple-tion studies reported that a substantial fraction of DENV-reactive Abs in human immune sera including type-specificneutralizing Abs bound to the intact virion but not torecombinant E proteinThe authors confirmed these observa-tions also isolating human neutralizing mAbs and proposedthat humans produce Abs that neutralize DENV infectionby binding a complex quaternary structure epitope that isexpressed only when E proteins are assembled on a virusparticle [48] Similar findings have been described for severalother viruses including WNV and HIV [82 83] Mappingstudies indicate that this epitope has a footprint that spansadjacent E protein dimers and includes residues at the hingebetween domains I and II of E protein [48]
Recently Costin et al isolated three broadly neutralizinganti-DENVhumanmAbs named 48AD11C and 16DThesemAbswere isolated from three different convalescent patientswith distinct histories of DENV infection All three mAbsrecognized the E glycoprotein with high affinity neutralizedall four serotypes of DENV but mediated ADE of infection inFc receptor-bearing cells at subneutralizing concentrationsMapping studies revealed that all three mAbs bind discontin-uous epitopes within the highly conserved fusion-loop regionof DII (contacting residues W101 L107 andor G109) [37]
25 Yellow Fever Virus (YFV) TheWHO estimates that thereare 200000 cases of yellow fever virus (YFV) infections and30000 related deaths every year especially in Sub-SaharanAfrica In fact different to themajority of human arbovirusesand similarly to DENV and CHIKV YFV has expanded itshost range to include humans as an amplifying host [1]Symptoms occurring from 3 to 6 days after infection withYFV consist mostly in fever chills anorexia lumbosacralpain nausea and vomiting This syndrome lasts about 3days in most cases and is sometimes followed by a one-daylasting period of remission Fever will reoccur together withdark hematemesismelena petechiae and other hemorrhagicsymptoms Convalescence is characterized by deep asthenialasting up to two weeks Rarely prognosis can be fatal Aneffective live-attenuated vaccine (17D) derived from the Asibistrain by serial passages in chicken embryos is available [84]
251 mAbs against YFV In 1983 Schlesinger et al produceda battery of mAbs after immunization of BALBc mice byinjection of 17D YFV vaccine strain [84] Among them thespecificity towards the E and NS1 proteins was describedeven though anti-NS1 Abs were not able to show neutralizingactivity The 13 IgG and 1 IgM anti-E protein mAbs wereclassified in five groups according to their specificity GroupA consisting of the only IgM produced in this experiment
10 BioMed Research International
could neutralize 17D YFV virus only group B 17D and Asibistrains only group C Asibi strain only group D could onlyneutralize Asibi but not 17D strain showing also reactivityagainst DENV2 Zika or Banzi viruses All of the four mAbsbelonging to this group were able to cross-react with Zikaandor Banzi viruses and two (4E11 and 5H3) neutralizedDENV2 virus with 4E11 neutralizing also Banzi virus The3E9 mAb the only component of the group E could notneutralize 17D nor Asibi strains
All of the IgG mAbs resulted able to protect bothprophylactically and therapeutically BALBc or CD-1 micefrom lethal intracerebral challenge with 17D-204 strain [85]The chimeric form of the group B mAb 2C9 IgG (2C9-cIgG) was able to provide AG129 mice a 72 survival whenadministered 24 hours before infection with 17D-204 strainIts murine form could provide the survival of the 95 ofthe mice Appreciable results were also obtained when bothmurine 2C9 and 2C9-cIgG were administered 24 hours post-infection with the survival of 70 and 20 of mice respec-tively Viral presence was not detected in survivingmice [60]
In 2005 Daffis and colleagues constructed two Ab-phage libraries by cloning the repertoire of YFV-infectedpatients Panning was then performed with YFV-17D virionsThe scFv-5A 7A and R3(27) showed a neutralizing activityspanning from50 to 100 inPRNTassays against bothYFV17D-204-WHO and Asibi strains In further tests reactivitywas observed against wild-type YFV strains of West Africagenotype I and II (Nigeria 1987 and Asibi strains resp)and EastCentral Africa (CAR 1986 Ethiopia 1961 strains) Aconcentration from 01 to 03120583gmL could yield a 90 plaquereduction Reactivity was also observed against the strainSenegal 1990 in a lesser extent Production of escape variantscould demonstrate that scFv-5A 7A and R3(27) epitopes arebuilt up by residues extensively separated in themonomers ofE glycoprotein that however result in closed proximity whenthe homodimeric form of E is constituted [61]
26 West Nile Virus (WNV) The West Nile virus (WNV) isan epidemic neurotropic virus estimated to be responsibleof about 36000 cases and 1500 deaths registered in theUnited States between 1992 and 2012 [86] WNV antigenicityallows its classification into the Japanese encephalitis virus(JEV) serocomplex Genomic analysis has revealed two maingenetic lineages of WNV lineage I viruses circulating in theUSA Europe the Middle East Africa India and Australiaand lineage II viruses isolated from Sub-Saharan Africa andMadagascar [1]
WNV was firstly identified in 1937 and is endemicin many countries of Africa Middle East and West AsiaHowever after 1990 frequent outbreaks of WNV infectionswere reported in Romania Israel and later in North Americaand across the USA in 1999 and recently in 2012 [87]
Most of WNV infected individuals do not develop symp-toms while about 20 develop a self-limiting illness calledWest Nile fever Acute symptoms include fever tirednessand swollen lymph glands but in a minority of cases alsoencephalitis with long-term deficits in cognitive functionand motor skills have been reported However in less than
1 of infected patients WNV having crossed the blood-brain barrier is responsible of neuroinvasive and potentiallylethal form of the disease In these cases degeneration andapoptosis upon infection of neurons and the consequentinflammatory response can occur Related symptoms includehigh fever coma muscle weakness and paralysis Immun-odepression and advanced age have been correlated with anaugment risk to develop a severe disease [87] There are nospecific treatment options or licensed vaccines for humans[88]
WNV is transmitted by infected mosquitos and initialreplication is thought to occur primarily in dendritic cells inthe skin which migrate to secondary lymphoid tissues wherethe replicating virus enters the circulation To date onlyefficacious WNV veterinarian vaccines have been licensedwhile there are no licensed vaccines for protection againstWNV in humans [88]
261 mAbs against WNV Broad-spectrum antivirals suchas type I IFN-120572 ribavirin mycophenolic acid in WNVinfection showed ineffective results in vivo despite in vitrothey showed some activity [89]
However experiments in murine models extrapolationof clinical data as well as passive administration of pooledimmune-120574-globulins (OmriGam) containing a significanttiter of neutralizing Abs before and after infection showedan important role of the humoral immune response incontrolling viremia and prevent viral dissemination Indeedthe development of a neutralizing mAb-based therapy seemsto be encouraging for a possible treatment of infected patients[90ndash92]
Similarly to what has been observed for DENV only anti-E Abs have been identified as neutralizing and protectiveAlso anti-NS1 Abs have shown a protective role howevertheir mechanism of protection has not yet been elucidated asthe NS1 protein is secreted from infected cells and not presenton the virions [93ndash95]
In this regard similarly to what has been describedfor DENV studies performed using a naıve human scFvlibrary for panning with purified WNV E protein supportthe hypothesis that no Abs against the neutralizing DIII canbe isolated However DIII specific Abs were isolated in asubsequent study using immunoglobulin libraries obtainedfrom threeWNV infected patients for biopanning on purifiedinactivated virus virus-like particles consisting of prM andE proteins or recombinant E glycoprotein Although theproportion of DIII-specific mAbs was low (8) comparedwith anti-DII mAbs (47) Two out of the four anti-DIIImAbs were potently neutralizing and protective in vivowhereas only three out of the 24 anti-DII were weaklyneutralizing in vitro and non-protective in vivo In particularresidues that are critical for neutralization lies on the regionsspanning amino acids 305ndash312 330ndash333 and 365 that arelocated on adjacent exposed loops ofDIIIHowever sequencealignment of E protein of different flaviviruses such as DENVand JEV revealed a considerable variation compared tothe whole E protein This observation further suggests thatdifferently from broadly cross-reactive anti-DII Abs anti-DIII neutralizing Abs are virus-type specific However of
BioMed Research International 11
these mAbs two (CR4374 and CR4353) protected mice fromlethal WNV challenge at 50 protective doses of 129 and357 120583gkg of body weight respectively [50]
Sanchez et al using three different immunization strate-gies (ie inactivated virus naked DNA and recombinantprotein) isolated ninemurinemAbsmost of which bound toconformation-dependent epitopes in DIII of the E protein Inparticular neutralizing mAbs named 8B10 11C2 10C5 and17C8 were obtained from mice immunized with inactivatedvirus alone or in combination with a DNA plasmid andbound to the same region of DIII with high affinity Incontrast mAbs obtained by immunization with a solubleversion of the E glycoprotein did not exhibit neutralizingactivity These non-neutralizing mAbs were cross-reactivewith several other flaviviruses including SLEV JEV YFVand Powassan virus confirming the conserved nature ofFlavivirus non-neutralizing epitopes [96]
Gould et al isolated 11 unique human single-chain vari-able region Ab fragments (scFvs) that bind the E protein ofWNVAmong them a humanmAb namedmAb11 expressedas a scFv-Fc fusion protein was further characterized Itrecognizes the fusion loop at the distal end of DII ofthe WNV E protein and cross-reacts with all four DENVserotypes and provides protection against DENV2 and 4as well as WNV [42] Moreover therapeutic studies of thismAb in WNV-infection model mice provided substantialprotection when administered after 5 days post-infectionInterestingly a neutralization escape variant of this mAbfailed to cause lethal encephalitis (at higher infectious doses)or induce the inflammatory responses associated with blood-brain barrier permeability in mice compared to the parentalWNV suggesting an important role for the fusion loop inviral pathogenesis [43]
Oliphant et al isolated an anti-DIII E protein mAbnamed E16 fromhybridomas obtained after immunization ofmice with recombinant WNV E protein which neutralizedall WNV strains with PRNT50 values of 4 to 18 ng andPRNT90 values of 53 to 297 ng One hundred microgramsof mAb protected greater than 90 of mice from lethalinfection and even a single 4120583g treatment of E16 on day 2 afterinfection prevented mortality Moreover humanization ofthis mAb confirmed as therapeutically effective in mice [44]Subsequent studies revealed that mAb E16 neutralization ismediated by engagement of four discontinuous segmentsof DIII including the amino-terminal region (amino acidresidues 302ndash309) and the three connecting loops BC (aminoacid residues 330ndash333) DE (amino acid residues 365ndash368)and FG (amino acid residues 389ndash391) Moreover no ADE ofinfection was detected when E16 mAb was used at saturatingconcentrations [45 47]
Furthermore results of a Phase I safety study of thehumanized E16 mAb (designated MGAWN1) have beenreported and suggested that doses of up to 30mgkgwerewelltolerated with few mild adverse events and would providean excess of virus neutralizing activity for 3-4 weeks aftertreatment However a case of anti-MGAWN1 Ab elicitationoccurred with the consequent increased rate of clearance andindeed impacting efficacy A Phase II safety and efficacy studyof MGAWN1 is ongoing [97] Furthermore the possibility of
preventing or treating WNV-induced memory deficits wasrecently investigated In this study hamsters were treatedintraperitoneally with 32mgkg ofMGAWN1mAb at 45 daysafter subcutaneously challenging with WNV InterestinglyMGAWN1 prevented mortality weight loss and improvedfood consumption of WNV-infected hamsters compared tocontrols [98]
Recently Lelli et al isolated six anti-E mAbs frominactivated-WNV immunized mice In particular three ofthem (3B2 3D6 and 4D3) neutralized lineage I and II WNVwith the first two recognizing the same epitopes located onthe distal lateral surface of DIII (critical amino acid residueK307) Conversely 4D3mAb recognized a novel neutralizingepitope on DII (critical residues S276 and T278) Indeedfurther protective and therapeutic studies are needed toascertain their neutralizing activity in vivo [99]
Finally to conclude like DENV infection Abs directedagainst the M protein are not protective and neutralizingMoreover in humans a skewed humoral immune responseagainst DII has been frequently observed and confirmedalso with the hybridoma technology [50] The isolation andelicitation of neutralizingAbs directed against the fusion loopand DIII of the E protein represent thus the most challengingand promising goal for the development of new effectivetherapeutic inhibitors and immunogens respectively
27 Tick-Borne Encephalitis Virus (TBEV) Tick-borne ence-phalitis virus (TBEV) is one of the most dangerous agentscausing human neuroinfections occurring mostly in Europeand Asia and with a potential fatal prognosis [100] TBEV isbelieved to cause 3500ndash10000 human cases of encephalitisin Europe per year with a high morbidity in Russia CzechRepublic Austria and Germany In particular between 1990and 2007 an average of 8755 cases of TBE was reported peryear in Europe and Russia Despite the fact that Russia isthe country with most infections registered annually CzechRepublic incidence is among the highest in Europe with 400ndash1000 clinical cases reported every year [101]
Three subtypes of TBEV are classified namely EuropeanSiberian and Far-Eastern sharing most of the genetic andantigenic features [102] In fact a high degree of antigenichomogeneity between different strains of TBEV has beendescribed [103]
Clinical manifestations vary among the subtypes butusually they start with a short febrile period of 7ndash14 daysafter the tick bite Fatigue headache and pain in the neckshoulders and lower back together with high fever andvomitingmay be present [100]Thesemanifestations are oftenfollowed by an asymptomatic phase lasting from 2 to 10days after remission from the fever with possible progres-sion to neurological disease Neurological symptoms includemeningitis encephalitis myelitis and radiculitis Mortalityoccurs in 1-2 of the European subtype-infected patientsbut fatal prognosis can occur in up to 20ndash40 of the Far-Eastern subtype-infected patients [104] Mortality rates ofthe Siberian subtype are similar to those observed for theEuropean [105]
Effective and safe vaccines against TBEV produced frominactivated virus have been developed and licensed for their
12 BioMed Research International
use in humans However an emergency therapy in theabsence of a mass immunization is needed as no effectivetreatments are yet available [106]
271 mAbs against TBEV Levanov et al described the chim-erization of two murine mAbs (13D6 and 10C2) directedagainst the DIII and DII respectively of the TBEV E glyco-protein The chimeric mAbs present binding characteristicssimilar to the parentalmAbsMoreover as the parentalmAbsonly the chimeric mAb 13D6 was able to neutralize TBEVinfectivity in vitro In particular neutralization studies withthe murine chimeric and scFv forms of mAb 13D6 were per-formed In particular murine 13D6 showed an IC50 titer of115 120583gmL in Focus Reduction Neutralization Tests (FRNT)against TBEV strain 205 and of 29 120583gmL in PRNT againstTBEV strain Softjin Chimeric 13D6 showed an IC50 titer of45 120583gmL in FRNT against TBEV strain 205 and of 19 120583gmLin PRNT against TBEV strain Softjin ScFv 13D6 showed anIC50 titer of 167 120583gmL in FRNT against TBEV strain 205and of 112 120583gmL in PRNT against TBEV strain Softjin [62]
28 Japanese Encephalitis Virus (JEV) Japanese encephalitisvirus (JEV) is a mosquito-borne flavivirus that over thepast few decades has caused several outbreaks throughoutChina Southeast Asia Australia and Papua New Guineawith a prevalence recently estimated to be of about 70000casesyear [107ndash109] A 40 mortality was recorded in someof the JEV-affected areas Moreover many survivors facesome neurological problems and complications [108] Since1995 the disease has also emerged in Non-Asian regions suchas Northern Australia [110 111] The situation in SoutheastAsia however is further complicated by the overlappingepidemics of JEV and DENV as well as sporadic casesof WNV infections detected in some of the affected areasparticularly in India [112]
Effective vaccines both live-attenuated and inactivatedstrains of JEV have been developed and licensed in majorcountries However as for the other arthropod-borne Fla-vivirus members no specific antiviral drugs are currentlyavailable [4]
281mAbs against JEV Aspreviously described for the otherFlavivirusmembers many groups isolated and characterizedanti-JEVmAbs showing different neutralizing and protectiveproperties In this regard Gupta et al used combinations ofanti-E JEV mAbs (Hs1-4) in mice protection experimentsIn particular they found that the singularly mAb protectionranged from 45 to 65 when 100120583g of mAb were adminis-tered while equimolar combinations of two or three mAbsgave 85ndash90 or 100 protection respectively [113] In similarexperiments Lee et al demonstrated that a Flavivirus anti-NS1 mAb named 16NS1 cross-reacted with JEV as well asWNV and exhibited protective activity against WNV as wellas a lethal JEV infection However no neutralizing activitywas observed using this mAb against both WNV and JEV inin vitro experiments suggesting the participation of otherAb-mediatedmechanisms in vivo In particular 95ofmicewere
protected when 500120583g of mAb were administered intraperi-toneally and concomitantly to intramuscular injection of JEV
Overlapping peptide mapping analysis combined withsite-specific mutations identified the 116KAWGKSILFA125region and critical amino acid residues W118 and I122 as16NS1 mAb epitope highly conserved in WNV and JEVstrains [114]
Arakawa et al isolated from a combinatorial human Fablibrary constructed from peripheral blood lymphocytesobtained from JEV hyperimmune volunteers Among 188randomly selected clones FabTJE12B02 showed the best 50focus reduction endpoint at the concentration of 502 120583gmLagainst the JEV strain Nakayama [55]
Goncalvez et al isolated three mAbs named Fabs A3B2 and E3 by repertoire cloning from chimpanzees initiallyimmunized with inactivated JE-VAX and then boosted withattenuated JEV SA14-14-2 In particular these mAbs reactedwith epitopes in three different E domains in DI (amino acidresidue K179) in DII (I126) and in DIII (G132) for Fabs A3B2 and E3 respectively Moreover these Fabs as well as thederived humanized counterpart mAbs exhibited high neu-tralizing activities against a broad spectrum of JEV genotypestrains Moreover these mAbs exhibited a 50 protectivedose of 084 120583g (B2) 58120583g (A3) and 247120583g (E3) in mousemodels Finally administration of 200 120583gmouse of mAb B2one day after otherwise lethal JEV infection protected 50of mice and significantly prolonged the average survival timecompared to that of mice in the unprotected group [54]
29 St Louis Encephalitis Virus (SLEV) St Louis encephalitisvirus (SLEV) was first discovered as the mosquito-borneagent responsible for over 1000 cases of encephalitis duringa 1933 summer outbreak in St Louis (Missouri) and nowis a reemerging human pathogen widely distributed in theAmerican continent causing several human encephalitisoutbreaks over the last 80 years Additional epidemics haveindeed occurred from 1964 to 2006 in the Americas rangingfrom the US to Argentina and Brazil As a member of theFlavivirus genus the SLEVE glycoprotein ectodomain is 68identical to serocomplex-related JEVEbut only 46 and 40identical to those ofDENV2 andTBEV viruses fromdifferentserocomplexes [115]
291 mAbs against SLEV In 1983 Roehrig et al isolatedtwenty-one hybridomas producing murine mAbs specificfor the E glycoprotein of SLEV strain MSI-7 Serologicreactivities were initially determined by cross-reactivity indi-rect immunofluorescence assays using 22 strains of SLEVand 8 other related flaviviruses Four groups demonstratingtype- subcomplex- supercomplex- and group-specific reac-tivity patterns were identified Analysis of hemagglutination-inhibition and virus neutralization subdivided the cross-reactivity groups into eight epitopes (E-1a b c d E-2 E-3 and E-4a b) one of them following localized in DII (4b)[52 53]
Moreover among the previously described isolated anti-WNV mAbs by Gould et al the chimeric scFv-Fc 79 effec-tively neutralized also SLEV resulting in gt80 of PRNT80when used at 5 120583gmL [42]
BioMed Research International 13
3 Alphaviruses
Major human arboviruses in the Togaviridae family Chikun-gunya (CHIKV) and Venezuelan equine encephalomyelitis(VEEV) viruses belong to the Alphavirus genus which iscomposed of viruses with an icosahedral nucleocapsid sur-rounded by a lipid envelope and glycoprotein spikes Thestructural proteins of alphaviruses arise through co- andpost-translational processing of a polyprotein encoded by asingle positive-stranded RNAproducing the capsid (C) PE26 K and E1 The PE2 glycoprotein is a precursor containingthe E3 glycoprotein fused to the amino terminus of the E2envelope glycoprotein The PE2 glycoprotein is followed by6K a small membrane-associated protein and E1 the secondpolypeptide component of glycoprotein spikes
Trimerized heterodimers of the E1 and E2 viral glyco-proteins form the surface spikes and contain determinantsof viral tropism and virulence The E3 glycoprotein acts as asignal for transport of PE2 across themembranes of the roughendoplasmic reticulum and may promote the formation andintracellular transport of E1-PE2 heterodimers to the cellsurface During transport to the cell surface PE2 undergoesa maturational cleavage event by a furin-like protease toproduce E2 and E3 [116]
The E2 glycoprotein promotes specificity of virus bindingto the host cell surface and is a target of the humoral immuneresponse The E1 glycoprotein mediates fusion of the virionenvelope with the membranes of acidified endosomes allow-ing release of the nucleocapsid into the cytoplasm and theonset of viral replicationAlphavirus E1 shares no appreciablesequence identity to Flavivirus E protein Despite differencesin their amino acid sequences and arrangements on the viralparticle the structures of E and E1 are remarkably similarIndeed the conservation of three domains has been welldocumented in crystal structures of flavivirus E ectodomainsfrom TBEV DENV JEV and WNV as well as Alphavirus E1from Semliki Forest virus (SFV) and CHIKV Abs to the E1glycoprotein do not typically neutralize virus infectivity invitro but can protect against lethal challenge in animals
By analogy with the prM protein of dengue Flavivirusfurin cleavage of PE2 may begin after transit of an acidiclate component of the Golgi body where E3 is thought tosuppress the acid pH-triggered activation of glycoprotein E1fusion capability Protective anti-E3 mouse mAbs have alsobeen described [117]
31 Chikungunya Virus (CHIKV) The chikungunya virus(CHIKV) belongs to the Semliki Forest clade and was firstlyisolated in 1953 in Tanzania during an epidemic outbreakfollowing occurred also in Asia and Africa In the last fivedecades several CHIKV outbreaks have been described bothin Africa and Asia separated by gaps lasting from two totwenty years In 2005-2006 about 300000 cases out of785000 inhabitants were reported in La Reunion island witha fatal prognosis for 237 of the patients [117] Neither Europenor the Americas have had outbreaks of CHIKV so far exceptfor imported isolated cases Three genotypes of CHIKV aredescribed Asian EastCentralSouth African (ECSA) andWest African with an amino acid identity spanning from
952 to 998 Recent epidemics in Africa and Indiansubcontinent were caused by strains belonging to the ECSAgenotype Transmitted by Aedes mosquitoes CHIKV ismaintained in the human population by human-mosquito-human transmission [1] The disease is characterized bydengue-like symptoms such as chills high fever headacheand persistent myalgia and a further incapacitating arthralgia(from which the name chikungunya) which affects 40 ofinfected subjects However prognosis is rarely fatal [116]
There is currently no commercial vaccine and antiviraltreatment forCHIKV although some candidate vaccines havebeen tested in humans In this regard in 2000 US Armyperformed a Phase II clinical trial testing a live-attenuatedCHIKV vaccine (TSI-GSD-218) derived from a 1962 strain(15561) of an outbreak in Thailand Out of 58 every patientdeveloped neutralizingAbs and 5 lamentedmild tomoderatejoint pain [117] A phase III trial of this candidate vaccineis ongoing Furthermore a new formulation using virus-likeparticles was able to induce neutralizing Abs in macaquesagainst different CHIKV strains [117 118] Indeed it has beendescribed that infection seems to elicit long-lasting protectiveimmunity and cross-protection among CHIKV and otheralphaviruses
CHIKV entry into host cell is demonstrated to be medi-ated by envelope glycoproteins E1 and E2 which allowvirus fusion to cell membrane in low pH conditions andrecognition of an unknown cellular receptor respectively[116 119ndash121] Even though the genome replication relies onan error-prone RNA-dependent RNA-polymerasis CHIKVstrains S27 and 05115 Reunion showed in E1 and E2 alow amino acid variation of 068 and 33 respectivelyand such a low variability may be needed for an effectivereplication in two phylogenetically distant hosts [116 122]Recently the crystal structure of E1 and E2 heterodimerhas been resolved both at low pH and natural conditionsshedding light on E1 and E2 ectodomainsrsquo structure Inparticular similarly to flaviviruses E protein E1 ectodomainis made of the N-terminal DI the DII containing the fusionloop and the DIII at the C-terminal On the other hand E2ectodomain comprises theN-terminal domainA the domainB supposed to interact with the host cellrsquos unknown receptorand domain C at the C-terminal E1 DIII and E2 domain Care located close to the viral membrane [123 124]
311 mAbs against CHIKV Warter et al described 5F10 and8B10 two human mAbs which strongly neutralized severalCHIKV isolates in vitro without cross-reactivity againstother alphaviruses but to Onyong-nyong virus [56] Mixedpreparation of 5F10 and 8F10 did not show neither synergisticnor addictive effect in vitro and studies upon escape mutantsdemonstrated that 5F10 mAb binds at the tip of the E2domain B while 8B10 recognizes residues close to E1 fusionloop and amino acids within E2 domain A which form atransitional epitope under low pH conditions The authorsspeculated that 5F10 and 8B10 may inhibit CHIKV entry andfusion to the cell membrane respectively It is worth notingthat the previously mentioned escape mutants displayedmutations associated with reduced viral fitness in vitro evenafter 13 neutralizationamplification rounds In the same
14 BioMed Research International
study cell-to-cell transmission was firstly demonstrated as anescapemechanism enhanced by the E2mutation R82G [122]
In further in vivo studies 5F10 and 8B10 significantlydelayed CHIKV-caused death of AGR129 mice both in pro-phylactic and therapeutic tests Interestingly in therapeutictreatment these mAbs showed a synergistic effect whenadministered in combination with the total amount of mAbsinjected being the same of the single-mAbs Possibly thesetreatment did not allow themice to survive CHIKV challengedue to Ig clearing and 6ndash10 days half-life usually described forhuman mAbs [125]
Recently Pal et al cloned thirty-six murine mAbs ableto neutralize the ECSA La Reunion 2006 OPY-1 strain ofCHIKV (CHIKV-LR) the majority of which also neutralizeinfection of other strains corresponding to the Asian andWest African genotypes In particularmAbCHK-152 showedthe highest and broadest neutralizing activity with FRNTvalues indicating an IC50 value of 1 to 3 ngmL dependingon the viral strain used Among the thirty-six mAbs four(CHK-102 CHK-152 and CHK-166 CHK-263) could pre-vent lethality of immunodeficient Ifnarminusminus C57BL6 micewhen administered one day before exposition with CHK-152 and CHK-263 showing the ability to protect mice at thelowest dose (10 120583g) Therapeutic studies were performed onthese mAbs administering a single dose of 100 120583g 24 hoursafter infection The highest activity was described for CHK-166 able to protect 63 of the mice Testing the combinedactivity of the mAbs in therapeutic studies administration ofCHK-166 plus CHK-152 in a dose of 250120583g each turned out tobe themost effective protecting 71 of the IfnarminusminusC57BL6mice 60 hours after infection [57]
32 Venezuelan Equine Encephalomyelitis Virus (VEEV)Venezuelan equine encephalomyelitis virus (VEEV) is main-tained in a natural transmission cycle between mosquitoesand small rodents The first documented outbreaks occurredin the 1930s and several epidemics have been reported so farin Latin-American countries such as Venezuela ColombiaPeru Ecuador Bolivia Honduras Mexico and Panama withhundreds of thousands of human cases being reported anda case-fatality rate up to 1 during the 1969 Ecuador out-break Clinical manifestations of VEE are indistinguishablefrom Dengue [126] Six serogroups (IndashVI) are currentlyrecognized within the VEEV complex VEEV caused humanand equine outbreaks in the Americas for nearly a centuryEquine epizootics have high mortality (38ndash83) leading toa high viremia followed by a lethal encephalitis and often tohuman epidemics involving thousands of cases and hundredsof deaths Indeed VEEV is infectious for humans by theairborne route and has been responsible for a number oflaboratory infections In humans the disease is usually self-limiting with a febrile illness with 1ndash4 of cases progressingto severe encephalitis It is a hazard to laboratory workershas been developed as a biological weapon and is a potentialbioterrorist agent [127] There are no antiviral drugs andvaccines licensed for the treatment of VEEV infection inhumans Indeed although experimental live-attenuated vac-cines have been developed (eg TC-83) with good levels ofprotection in equine and mice in humans they may fail to
give protection in the majority of cases [128] Consideringthe protective role exerted by humoral immune response inanimal models it could be concluded that treatment withspecific IgG may have a beneficial antiviral effect in humanairborne infections with pathogenic strains of VEEV
321 mAbs against VEEV Several works support that mAbsmay protect against airborne VEEV as well as after airborneexposure toVEEVAb reactivity to both surface glycoproteins(E1 and E2) is associated with neutralization of the virusin vitro and passive protection against virus challenge Aseries of anti-E1 and anti-E2 mouse and humanized mAbshave been described recently Among them Phillpotts et alexamined two anti-E2 murine mAbs named 1A4A-1 and1A3A-9 both having potent protective activity against subcu-taneous VEEV challenge in mice Both mAbs had a similarhalf-life (58 and 100 days) in mouse serum after a singleintraperitoneal dose suggesting that mAbs delivered by thisroute at or around the time of VEEV infection persists athigh levels in the blood as well as secretions in respiratorytransudation throughout the clinical course of the disease Inparticular both mAbs administered 24 h prior to airbornechallenges had a substantial protective effect (90ndash100)Treatment of mice 2 or 24 h after airborne infection with asingle intraperitoneal dose of 100 120583g of 1A3A-9 mAb led toapproximately 50 survival There was no beneficial effectwhen mAb treatment was delayed to 72 h Moreover therewas evidence of synergy in vitro in PRNT between 1A3A-9and 1A4A-1 as has been demonstrated for other virusesand mAb pairs However no synergy was found in mouseprotection when mAbs were delivered intraperitoneally as amixture in equal parts to animals challenged 24 h previouslywith VEEV [129]
In a subsequent study Phillpotts et al reported two otherprotective mAbs The 3B2A-9 mAb protected against allthe serogroup I strains while 1A3B-7 mAb protected wellagainst challenge with all of the viruses tested Both 3B2A-9 and 1A3B-7 protected against airborne exposure to theIAB serogroup virus strainTrinidad donkey (TrD)Howeverthere was no evidence of synergistic protection when thesemAbs were combined in equal proportions An intraperi-toneal dose of 10 120583g was sufficient to protect 50 of themice with either mAb [130] According to other data themechanism of protection did not appear to depend uponneutralization [131] Similar results were obtained by OrsquoBrienet al which described a non-neutralizing mAb (IgG2a)named CUF37-2a able to protect 50 of mice from asubcutaneous VEEV challenge when a dose of 915 120583g of mAbwas administered 24 h prior to challenge [132]
In a following study the 1A3B-7 mAb was humanized(and following reported as Hu1A3B-7) maintaining the samefeatures of its murine counterpart In particular evaluation ofin vitro studies indicated that Hu1A3B-7 retained both broadspecificity and neutralizing activity Furthermore in vivoexperiments showed that Hu1A3B-7 successfully protectedmice against lethal subcutaneous and aerosol challenges withVEEV strain TrD Moreover the effectiveness of the human-ization process was determined by assessing proliferationresponses in human T-cells to peptides derived from the
BioMed Research International 15
murine and humanized versions of the VH and VL domainsThis analysis showed that the number of human T-cellepitopes within the humanized Ab had been substantiallyreduced indicating that Hu1A3B-7 may have low immuno-genicity in vivo [133]
Similarly Hunt et al described a humanizedmurinemAbderived from the 3B4C-4 mAb recognizing the E2c epitope(amino acid residues 182ndash209) and following named Hy4-26C showing neutralizing activity similar to the murinemAb Moreover it was protective (70ndash100 of survival) at100 ng dose in mice animal model intraperitoneal inoculatedand at 500120583g for intranasal challenge (80) Therapeuticstudies in mice revealed that Hy4-26C was able to cure miceup to 24 h following infection at 10 120583g on the contrary themouse mAb when given 1 hour after virus challenge [63]
In another work Hunt et al isolated two neutralizinghumanized Fabs F5 and L1A7 employing a blocking strategy[134 135] The anti-E2 specific F5 IgG had a 70 PRNTendpoint of 10 ngmL equivalent to that described for themost effective neutralizing anti-VEEVE2mAbs In particularthe E1-specific hFab L1A7 had a PRNT endpoint of 3120583gmL300-fold lower than F5
Further studies revealed that F5 epitope is located inthe 115ndash199 amino acid region Moreover F5 IgG had potentability to protect mice from infection by either route whenadministered 24 h before exposure however mice treated24 h and 48 h after aerosol exposure developed central ner-vous system infections but exhibited no clinical signs ofdisease [135]
Hu et al reported that passive immunization with thehumanized chimericmousemAbhu1A4A1IgG1-2A inmice at50 120583g 24 h before or after virulent VEEV challenge providedcomplete protection indicating that hu1A4A1IgG1-2A haspotent prophylactic and therapeutic effects against VEEVinfection [136]
Anti-VEEV mAbs isolated from a non-human primategene library has also been reported In particular thehumanized chimeric scFv-Fc ToR67-3B4 recognized viableas well as formalin and 120573-propiolactone inactivated virusparticles It detected specifically the viral E1 envelope proteinof VEEV but did not react with reduced viral glycopro-tein preparations suggesting that recognition depends uponconformational epitopes The recombinant Ab was able todetect multiple VEEV subtypes and displayed only marginalcross-reactivity to otherAlphavirus species except for Easternequine encephalitis virus (EEEV) In addition the scFv-Fcfusion described here might be of therapeutic use since itsuccessfully inactivated VEEV in a murine disease modelIn particular when the recombinant Ab was administered 6hours after challenge 80 to 100 of mice survived lethalVEEV IAB or IE infection Forty to sixty percent of micesurvived when scFv-Fc ToR67-3B4 was applied 6 hours afterchallenge with VEEV subtypes II and former IIIA [137]
4 Bunyaviridae
TheBunyaviridae family contains human arboviruses belong-ing to the Orthobunyavirus Phlebovirus and Nairovirus
genera Bunyaviridae includes enveloped viruses with a frag-mented single-stranded RNA genome of negative polarityTheir tripartite genome consists of a small (S) a medium(M) and a large (L) fragment and the envelope glycoproteinsGn and Gc that are cleaved out of a polyprotein synthesizedby the M fragment These glycoproteins have been provedto mediate the formation of the virus particle to play a rolein the interaction with cell surface receptors to mediate theentry of the virus into cells and to serve as targets for themajority of neutralizing Abs described so far
Orthobunyavirus transmitted through mosquitoes ormidges vectors are divided in 18 serogroups based on cross-titrations in haemagglutination inhibition assays and neutral-ization assays and correlating with main vector preferencesHowever the most clinically important viruses belong onlyto two serogroups the California encephalitis and the Simbuserogroups
Crimean-Congo hemorrhagic fever virus (CCHFV) isconsidered the only one of clinical relevance within theNairovirus genus which uses tick as main vector and is divid-ed in seven serogroups while Toscana virus and Rift Valleyfever virus (RVFV) which are transmitted by sandflies andmosquitoes respectively belong to the Phlebovirus genus[138]
MAbs against theNairovirusCCHFV and the PhlebovirusRVFV have been described
41 Crimean-Congo Hemorrhagic Fever Virus (CCHFV)Crimean-Congo Hemorrhagic Fever Virus (CCHFV) dis-tribution covers the greatest geographic range of any tick-borne virus known as viral isolation andor disease hasbeen reported from more than 30 countries in SoutheasternEurope Africa Asia and Middle East [139 140] Namelyfrom 1953 to 2010 about 6000 human cases were reportedin Southeastern Europe CCHFV causes sporadic outbreakswith mortality rates ranging from 10 to 80 [141 142] Asignificant variation in the time of incubation has beendescribed [139] Prehemorrhagic symptoms include highfever chills headache photophobia and back and abdominalpains Among other symptoms neuropsychiatric changeshave been reported in some patients In severe cases 3ndash6 daysafter the onset of the disease hemorrhagic symptoms occurwith petechiae ecchymosis bleeding in the form of melenahematemesis epistaxis [139]
In comparisonwith the nucleocapsid proteins inCCHFVGn and Gc show a higher degree of antigenic variabilityprobably due to their exposition to the host immune systemhypothesis corroborated in the description by Hewson et alin 2004 of four M segment phylogenic groups namely M1M2 M3 and M4 [141 142]
411 mAbs against CCHFV Blackburn et al firstly describedin 1987 murine anti-CCHFV mAbs able to recognize nucle-ocapsid proteins but no neutralizing activity has beendescribed thus far [143]
On the other hand Bertolotti-Ciarlet et al produced apanel of murine mAbs recognizing conformational epitopes
16 BioMed Research International
within theGn andGc glycoproteins expressed by the CCHFVstrain IbAr10200 The effectiveness of the mAbs was studiedby performing PRNT80 and none of the anti-Gn mAbsshowed neutralizing activity although many of the anti-GcmAbs neutralized IbAr10200 strain in vitro with clones 8A15E3 12A9 6C2 and 9H3 showing activity at a gt5120 dilutionMAbs 11E7 and 30F7 exhibited neutralizing activity at 2560dilution A suckling mice protection test was thereforeperformed for these mAbs The previously mentionedanti-Gc Abs were capable to protect mice to an appreciabledegree when applied 24 hours before and in a weakermanner 24 hours after virus challenge Many of the anti-GnmAbs were able to confer significant protection to IbAr10200both 24 hours before and after virus administration Therelevant effectiveness of the anti-Gn mAbs 6B12 10E1113G8 and 10G4 suggests that these mAbs may possess someneutralizing activity due to Ab-based effector mechanisms(eg ADCC) [58]
In further studies the same group characterized thebroadly cross-reactive neutralizing activity of the previouslydescribed murine mAb 11E7 able to bind conformationalepitopes within the C-terminal region of the Gn glycoproteinof all CCHFVM groups [58 141]
42 Rift Valley Fever Virus (RVFV) Similarly to CCHFV RiftValley Fever Virus (RVFV) causes sporadic outbreaks Thelargest epidemic occurred in Egypt in 1977 with an estimated200000 infections 18000 patients manifesting symptomsand 600 fatal prognoses [144] Symptoms include hemor-rhagic episodes fever encephalitis and blindness [145]
The major target of the humoral immune response is theRVFV glycoproteins Gc (G1) and Gn (G2) ranging fromnucleotides 480ndash2090 and 2091ndash3614 respectively [144]
421 mAbs against RVFV Within G2 glycoprotein are des-cribed three neutralizing antigenic sites namely epitope Inucleotides 792ndash893 (amino acid residues 258ndash291) epitopeII nucleotides 1164ndash1196 (amino acid residues 382ndash392) andepitope IV nucleotides 858ndash917 (amino acid residues 280ndash299) [59 145]
Despite the high degree of variability observed inCCHFVglycoproteins RVFV G2 presented an unexpected high con-servation among 22 isolates [59] In the same study onlythe LUNYO and 900060 isolates were resistant to in vitroneutralization by murine mAb 4D4 (recognizing epitope II)All other isolates were neutralized with PRNT80 titers from10240 togt81920On the other hand only SNS isolate showedreduced sensitivity to 4ndash39-CC (epitope IV) All other isolateswere neutralized with PRNT80 titers from 20480 to gt81920Protection studies on mice would be of interest for thedevelopment of a passive immunization prophylaxis [59]
Protection studies from lethal South African RVFVAN1830 strain infection were successfully performed on abattery of anti-G1 and anti-G2 mAbs produced by Besselaarand Blackburn with the strongly neutralizing anti-G1 mAb3E5 and anti-G2mAb 9C4 being the most effective [146]The9C4 antigenic area maps within G2 epitope I and a broadlyneutralizing activity may be investigated
5 Conclusions
Arbovirus infections have acquired increasing interest giventhe augmented globalization and tourism movement all overthe world that could be at the basis of epidemic eventsFurthermore the same vectors can sometimes transmit sev-eral arboviruses concomitantly complicating the diagnosis aswell as the therapy For example confusing mixed epidemicshave occasionally been described such as YFV plus CHIKVDENVplus CHIKV ormore recentlyPlasmodium falciparummalaria plus DENV1 and CHIKV on Madagascarrsquos east coast[1]
In the last decade several efforts have been employed inthe development of effective immunogens and therapeuticsgiving the current absence of specific antiviral drugs as wellas effective vaccines for themost diffused arbovirus infections(ie DENV andWNV)
However at the moment mosquito control is the bestavailable method for preventing arbovirus infections Thusto control the emerging public health problem of arbovirusinfections new antiviral therapeutic strategies that providepotent and broadly cross-protective immunity (especially forflavivirus infections) are an urgent globally medical need
As described in this review the majority of highlyneutralizing mAbs are of murine origin and indeed theirutility in the treatment of these infections is limited In facttheir employment can elicit an immunogenic response (iehuman anti-mouse Abs HAMA) the therapeutic efficacycould be reduced by a relatively faster clearance in humans(compared to human Abs) potentially exacerbated by theHAMA response Finally murine Abs exhibit relatively weakeffector functions (eg ADCC) compared to human AbsIndeed chimerization humanization or better the isolationand characterization of fully human broadly neutralizingmAbs would be the best choice for their possible use in afuture mAb-based therapy against arbovirus infections [147]
Finally possible ADE of infection mechanisms should beevaluated before considering a mAb as a possible candidatetherapeutic in a post-exposure setting as well as in the devel-opment of new Ab-eliciting immunogens
Conflict of Interests
The authors declare no conflict of interests in particular withOmriGam and any other cited Ab-based preparation men-tioned in the paper
References
[1] N Cleton M Koopmans J Reimerink G J Godeke and CReusken ldquoCome fly with me review of clinically importantarboviruses for global travelersrdquo Journal of Clinical Virology vol55 no 3 pp 191ndash203 2012
[2] F X Heinz H Holzmann A Essl and M Kundi ldquoField effec-tiveness of vaccination against tick-borne encephalitisrdquoVaccinevol 25 no 43 pp 7559ndash7567 2007
[3] A D Barrett and D E Teuwen ldquoYellow fever vaccinemdashhowdoes it work and why do rare cases of serious adverse events
BioMed Research International 17
take placerdquo Current Opinion in Immunology vol 21 no 3 pp308ndash313 2009
[4] S B Halstead and S J Thomas ldquoNew vaccines for Japaneseencephalitisrdquo Current Infectious Disease Reports vol 12 no 3pp 174ndash180 2010
[5] R Burioni M Perotti N Mancini and M Clementi ldquoPer-spectives for the utilization of neutralizing human monoclonalantibodies as anti-HCV drugsrdquo Journal of Hepatology vol 49no 2 pp 299ndash300 2008
[6] R A Diotti G A Sautto L Solforosi NManciniM Clementiand R Burioni ldquoNeutralization activity and kinetics of twobroad-range human monoclonal IgG1 derived from recombi-nant Fab fragments and directed against Hepatitis C virus E2glycoproteinrdquo New Microbiologica vol 35 no 4 pp 475ndash4792012
[7] R Burioni F Canducci N Mancini et al ldquoMolecular cloningof the first humanmonoclona antibodies neutralizing with highpotency Swine-origin Influenza A pandemic virus (S-OIV)rdquoNew Microbiologica vol 32 no 4 pp 319ndash324 2009
[8] P K Chandra S Hazari B Poat et al ldquoIntracytoplasmic stableexpression of IgG1 antibody targeting NS3 helicase inhibitsreplication of highly efficient hepatitis C Virus 2a clonerdquoVirology Journal vol 7 article 118 2010
[9] N Clementi D de Marco N Mancini et al ldquoA human mono-clonal antibody with neutralizing activity against highly diver-gent influenza subtypesrdquo PLoS ONE vol 6 no 12 Article IDe28001 2011
[10] D de Marco N Clementi N Mancini et al ldquoA non-VH1-69 heterosubtypic neutralizing human monoclonal antibodyprotects mice against H1N1 and H5N1 virusesrdquo PLoS ONE vol7 no 4 Article ID e34415 2012
[11] L Solforosi N Mancini F Canducci et al ldquoA phage displayvector optimized for the generation of human antibody com-binatorial libraries and the molecular cloning of monoclonalantibody fragmentsrdquoNewMicrobiologica vol 35 no 3 pp 289ndash294 2012
[12] E V Koonin and V V Dolja ldquoEvolution and taxonomy ofpositive-strand RNA viruses implications of comparative anal-ysis of amino acid sequencesrdquo Critical Reviews in Biochemistryand Molecular Biology vol 28 no 5 pp 375ndash430 1993
[13] J S Mackenzie D J Gubler and L R Petersen ldquoEmergingflaviviruses the spread and resurgence of Japanese encephalitisWest Nile and dengue virusesrdquo Nature Medicine vol 10 no 12pp S98ndashS109 2004
[14] T J SmithW E Brandt J L Swanson J MMcCown and E LBuescher ldquoPhysical and biological properties of dengue-2 virusand associated antigensrdquo Journal of Virology vol 5 no 4 pp524ndash532 1970
[15] J M Smit B Moesker I Rodenhuis-Zybert and J WilschutldquoFlavivirus cell entry and membrane fusionrdquo Viruses vol 3 no2 pp 160ndash171 2011
[16] R J Kuhn W Zhang M G Rossmann et al ldquoStructure ofdengue virus implications for flavivirus organization matura-tion and fusionrdquo Cell vol 108 no 5 pp 717ndash725 2002
[17] K Stiasny J H Aberle M Keller B Grubeck-Loebenstein andF X Heinz ldquoAge affects quantity but not quality of antibodyresponses after vaccination with an inactivated flavivirus vac-cine against tick-borne encephalitisrdquo PLoS ONE vol 7 no 3Article ID e34145 2012
[18] LThomasOVerlaetenACabie et al ldquoInfluence of the dengueserotype previous dengue infection and plasma viral load
on clinical presentation and outcome during a dengue-2 anddengue-4 co-epidemicrdquo American Journal of Tropical Medicineand Hygiene vol 78 no 6 pp 990ndash998 2008
[19] Y Zhang J Corver P R Chipman et al ldquoStructures of imma-ture flavivirus particlesrdquo EMBO Journal vol 22 no 11 pp2604ndash2613 2003
[20] A K I Falconar ldquoIdentification of an epitope on the denguevirus membrane (M) protein defined by cross-protective mon-oclonal antibodies design of an improved epitope sequencebased on common determinants present in both envelope (EandM) proteinsrdquo Archives of Virology vol 144 no 12 pp 2313ndash2330 1999
[21] K-J Huang Y-C Yang Y-S Lin et al ldquoThe dual-specific bind-ing of dengue virus and target cells for the antibody-dependentenhancement of dengue virus infectionrdquo Journal of Immunol-ogy vol 176 no 5 pp 2825ndash2832 2006
[22] A K I Falconar ldquoAntibody responses are generated to immun-odominant ELKKLE-typemotifs on the nonstructural-1 glyco-protein during live dengue virus infections inmice and humansimplications for diagnosis pathogenesis and vaccine designrdquoClinical and Vaccine Immunology vol 14 no 5 pp 493ndash5042007
[23] C W Mandl F Guirakhoo H Holzmann F X Heinz and CKunz ldquoAntigenic structure of the flavivirus envelope protein Eat the molecular level using the tick-borne encephalitis virus asa modelrdquo Journal of Virology vol 63 no 2 pp 564ndash571 1989
[24] F A Rey F X Heinz C Mandl C Kunz and S C HarrisonldquoThe envelope glycoprotein from tick-borne encephalitis virusat 2 A resolutionrdquo Nature vol 375 no 6529 pp 291ndash298 1995
[25] S L Allison K Stiasny K Stadler CWMandl and F XHeinzldquoMapping of functional elements in the stem-anchor regionof tick-borne encephalitis virus envelope protein Erdquo Journal ofVirology vol 73 no 7 pp 5605ndash5612 1999
[26] D E Klein J L Choi and S C Harrison ldquoStructure of a denguevirus envelope protein late-stage fusion intermediaterdquo Journal ofVirology vol 87 no 4 pp 2287ndash2293 2013
[27] S L Hanna T C Pierson M D Sanchez A A Ahmed M MMurtadha and R W Doms ldquoN-linked glycosylation of WestNile virus envelope proteins influences particle assembly andinfectivityrdquo Journal of Virology vol 79 no 21 pp 13262ndash132742005
[28] N Mancini G Sautto N Clementi et al ldquoNeutralization inter-fering antibodies a ldquonovelrdquo example of humoral immune dys-function facilitating viral escaperdquoViruses vol 4 no 9 pp 1731ndash1752 2012
[29] Y Modis S Ogata D Clements and S C Harrison ldquoVariablesurface epitopes in the crystal structure of dengue virus type3 envelope glycoproteinrdquo Journal of Virology vol 79 no 2 pp1223ndash1231 2005
[30] N Miller ldquoRecent progress in dengue vaccine research anddevelopmentrdquo Current Opinion in Molecular Therapeutics vol12 no 1 pp 31ndash38 2010
[31] J H Chavez J R Silva A A Amarilla and L T Moraes Figu-eiredo ldquoDomain III peptides from flavivirus envelope proteinare useful antigens for serologic diagnosis and targets forimmunizationrdquo Biologicals vol 38 no 6 pp 613ndash618 2010
[32] B Shrestha J D Brien S Sukupolvi-Petty et al ldquoThe develop-ment of therapeutic antibodies that neutralize homologous andheterologous genotypes of dengue virus type 1rdquo PLoS Pathogensvol 6 no 4 Article ID e1000823 2010
18 BioMed Research International
[33] G D Gromowski N D Barrett and A D T Barrett ldquoChar-acterization of dengue virus complex-specific neutralizing epi-topes on envelope protein domain III of dengue 2 virusrdquo Journalof Virology vol 82 no 17 pp 8828ndash8837 2008
[34] S Sukupolvi-Petty S K Austin W E Purtha et al ldquoType- andsubcomplex-specific neutralizing antibodies against domain IIIof dengue virus type 2 envelope protein recognize adjacentepitopesrdquo Journal of Virology vol 81 no 23 pp 12816ndash128262007
[35] S Sukupolvi-Petty S K Austin M Engle et al ldquoStructure andfunction analysis of therapeutic monoclonal antibodies againstdengue virus type 2rdquo Journal of Virology vol 84 no 18 pp9227ndash9239 2010
[36] Y-Q Deng J-X Dai G-H Ji et al ldquoA broadly flaviviruscross-neutralizingmonoclonal antibody that recognizes a novelepitope within the fusion loop of e proteinrdquo PLoS ONE vol 6no 1 Article ID e16059 2011
[37] J M Costin E Zaitseva K M Kahle et al ldquoMechanistic studyof broadly neutralizing human monoclonal antibodies againstdengue virus that target the fusion looprdquo Journal of Virologyvol 87 no 1 pp 52ndash66 2013
[38] CMMidgley A FlanaganH B Tran et al ldquoStructural analysisof a dengue cross-reactive antibody complexed with envelopedomain III reveals the molecular basis of cross-reactivityrdquoJournal of Immunology vol 188 no 10 pp 4971ndash4979 2012
[39] N J Moreland P Susanto E Lim et al ldquoPhage display appro-aches for the isolation of monoclonal antibodies against denguevirus envelope domain III from human and mouse derivedlibrariesrdquo International Journal ofMolecular Sciences vol 13 no3 pp 2618ndash2635 2012
[40] J J B Cockburn M E Navarro Sanchez N Fretes et alldquoMechanism of dengue virus broad cross-neutralization by amonoclonal antibodyrdquo Structure vol 20 no 2 pp 303ndash3142012
[41] R Rajamanonmani C Nkenfou P Clancy et al ldquoOn a mousemonoclonal antibody that neutralizes all four dengue virusserotypesrdquo Journal of General Virology vol 90 no 4 pp 799ndash809 2009
[42] L H Gould J Sui H Foellmer et al ldquoProtective and herapeuticcapacity of human single-chain Fv-Fc fusion proteins againstWest Nile virusrdquo Journal of Virology vol 79 no 23 pp 14606ndash14613 2005
[43] H Sultana H G Foellmer G Neelakanta et al ldquoFusion LoopPeptide of the West Nile virus envelope protein is essential forpathogenesis and is recognized by a therapeutic cross-reactivehuman monoclonal antibodyrdquo Journal of Immunology vol 183no 1 pp 650ndash660 2009
[44] T Oliphant M Engle G E Nybakken et al ldquoDevelopment ofa humanized monoclonal antibody with therapeutic potentialagainstWest Nile virusrdquoNature Medicine vol 11 no 5 pp 522ndash530 2005
[45] G E Nybakken T Oliphant S Johnson S Burke M S Dia-mond and D H Fremont ldquoStructural basis of West Nile virusneutralization by a therapeutic antibodyrdquo Nature vol 437 no7059 pp 764ndash769 2005
[46] G D Gromowski J T Roehrig M S Diamond J C LeeT J Pitcher and A D T Barrett ldquoMutations of an antibodybinding energy hot spot on domain III of the dengue 2 envelopeglycoprotein exploited for neutralization escaperdquo Virology vol407 no 2 pp 237ndash246 2010
[47] J T Roehrig R A Bolin andR G Kelly ldquoMonoclonal antibodymapping of the envelope glycoprotein of the dengue 2 virusJamaicardquo Virology vol 246 no 2 pp 317ndash328 1998
[48] R de Alwis S A Smith N P Olivarez et al ldquoIdentification ofhumanneutralizing antibodies that bind to complex epitopes ondengue virionsrdquo Proceedings of the National Academy of Sciencesof the United States of America vol 109 no 19 pp 7439ndash74442012
[49] S K Austin K A Dowd B Shrestha et al ldquoStructural basis ofdifferential neutralization of DENV-1 genotypes by an antibodythat recognizes a cryptic epitoperdquo PLoS Pathogens vol 8 no 10Article ID e1002930 2012
[50] M Throsby C Geuijen J Goudsmit et al ldquoIsolation and char-acterization of human monoclonal antibodies from individualsinfected with West Nile Virusrdquo Journal of Virology vol 80 no14 pp 6982ndash6992 2006
[51] A P Goncalvez R H Purcell and C-J Lai ldquoEpitope deter-minants of a chimpanzee Fab antibody that efficiently cross-neutralizes dengue type 1 and type 2 viruses map to inside andin close proximity to fusion loop of the dengue type 2 virusenvelope glycoproteinrdquo Journal of Virology vol 78 no 23 pp12919ndash12928 2004
[52] J T Roehrig J H Mathews and D W Trent ldquoIdentification ofepitopes on the E glycoprotein of Saint Louis encephalitis virususing monoclonal antibodiesrdquo Virology vol 128 no 1 pp 118ndash126 1983
[53] J T Roehrig L A Staudinger A R Hunt J H Mathews andC D Blair ldquoAntibody prophylaxis and therapy for flavivirusencephalitis infectionsrdquo Annals of the New York Academy ofSciences vol 951 pp 286ndash297 2001
[54] A P Goncalvez C-H Chien K Tubthong et al ldquoHumanizedmonoclonal antibodies derived from chimpanzee fabs protectagainst Japanese encephalitis virus in vitro and in vivordquo Journalof Virology vol 82 no 14 pp 7009ndash7021 2008
[55] M Arakawa T Yamashiro G-I Uechi M Tadano and ANishizono ldquoConstruction of humanFab (1205741120581) library and iden-tification of human monoclonal Fab possessing neutralizingpotency against Japanese encephalitis virusrdquo Microbiology andImmunology vol 51 no 6 pp 617ndash625 2007
[56] L Warter C Y Lee R Thiagarajan et al ldquoChikungunya virusenvelope-specific human monoclonal antibodies with broadneutralization potencyrdquo Journal of Immunology vol 186 no 5pp 3258ndash3264 2011
[57] P Pal K A Dowd J D Brien et al ldquoDevelopment of a highlyprotective combination monoclonal antibody therapy againstchikungunya virusrdquo PLoS Pathogens vol 9 no 4 Article IDe1003312 2013
[58] A Bertolotti-Ciarlet J Smith K Strecker et al ldquoCellular local-ization and antigenic characterization of Crimean-Congo hem-orrhagic fever virus glycoproteinsrdquo Journal of Virology vol 79no 10 pp 6152ndash6161 2005
[59] J K Battles and J M Dalrymple ldquoGenetic variation amonggeographic isolates of Rift Valley fever virusrdquo American Journalof Tropical Medicine and Hygiene vol 39 no 6 pp 617ndash6311988
[60] B AThibodeaux N C Garbino NM Liss et al ldquoA humanizedIgG but not IgM antibody is effective in prophylaxis and therapyof yellow fever infection in an AG12917D-204 peripheralchallenge mouse modelrdquo Antiviral Research vol 94 no 1 pp1ndash8 2012
BioMed Research International 19
[61] S Daffis R E Kontermann J Korimbocus H Zeller H-DKlenk and J Ter Meulen ldquoAntibody responses against wild-type yellow fever virus and the 17D vaccine strain charac-terization with human monoclonal antibody fragments andneutralization escape variantsrdquoVirology vol 337 no 2 pp 262ndash272 2005
[62] L N Levanov L E Matveev E P Goncharova et al ldquoChimericantibodies against tick-borne encephalitis virusrdquo Vaccine vol28 no 32 pp 5265ndash5271 2010
[63] A R Hunt S Frederickson C Hinkel K S Bowdish and J TRoehrig ldquoA humanized murine monoclonal antibody protectsmice either before or after challenge with virulent Venezuelanequine encephalomyelitis virusrdquo Journal of General Virologyvol 87 no 9 pp 2467ndash2476 2006
[64] J M da Silva Voorham I A Rodenhuis-Zybert N V A Nunezet al ldquoAntibodies against the envelope glycoprotein promoteinfectivity of immature dengue virus serotype 2rdquo PLoS ONEvol 7 no 3 Article ID e29957 2012
[65] T C Pierson Q Xu S Nelson et al ldquoThe stoichiometry ofantibody-mediated neutralization and enhancement of westnile virus infectionrdquoCell Host andMicrobe vol 1 no 2 pp 135ndash145 2007
[66] G Sautto N Mancini R A Diotti L Solforosi M Clementiand R Burioni ldquoAnti-hepatitis C virus E2 (HCVE2) glyco-proteinmonoclonal antibodies andneutralization interferencerdquoAntiviral Research vol 96 no 1 pp 82ndash89 2012
[67] R Burioni Y Matsuura N Mancini et al ldquoDiverging effectsof human recombinant anti-hepatitis C virus (HCV) antibodyfragments derived from a single patient on the infectivityof a vesicular stomatitis virusHCV pseudotyperdquo Journal ofVirology vol 76 no 22 pp 11775ndash11779 2002
[68] R J Phillpotts J R Stephenson and J S Porterfield ldquoAntibody-dependent enhancement of tick-borne encephalitus virus infec-tivityrdquo Journal of General Virology vol 66 no 8 pp 1831ndash18371985
[69] K A Dowd and T C Pierson ldquoAntibody-mediated neutraliza-tion of flaviviruses a reductionist viewrdquo Virology vol 411 no 2pp 306ndash315 2011
[70] A K I Falconar ldquoMonoclonal antibodies that bind to commonepitopes on the dengue virus type 2 nonstructural-1 andenvelope glycoproteins display weak neutralizing activity anddifferentiated responses to virulent strains implications forpathogenesis and vaccinesrdquo Clinical and Vaccine Immunologyvol 15 no 3 pp 549ndash561 2008
[71] K Meyer A Banerjee S E Frey R B Belshe and R Ray ldquoAweak neutralizing antibody response to hepatitis C virus enve-lope glycoprotein enhances virus infectionrdquo PLoS ONE vol 6no 8 Article ID e23699 2011
[72] R Burioni R AWilliamson P P Sanna F E Bloom and D RBurton ldquoRecombinant human Fab to glycoprotein D neutral-izes infectivity and prevents cell-to-cell transmission of herpessimplex viruses 1 and 2 in vitrordquo Proceedings of the NationalAcademy of Sciences of the United States of America vol 91 no1 pp 355ndash359 1994
[73] M S Rolph A Zaid N E Rulli and S Mahalingam ldquoDown-regulation of interferon-120573 in antibody-dependent enhancementof dengue viral infections of human macrophages is dependenton interleukin-6rdquo Journal of Infectious Diseases vol 204 no 3pp 489ndash491 2011
[74] S S Whitehead J E Blaney A P Durbin and B R MurphyldquoProspects for a dengue virus vaccinerdquoNature ReviewsMicrobi-ology vol 5 no 7 pp 518ndash528 2007
[75] S-M Lok V Kostyuchenko G ENybakken et al ldquoBinding of aneutralizing antibody to dengue virus alters the arrangement ofsurface glycoproteinsrdquoNature Structural andMolecular Biologyvol 15 no 3 pp 312ndash317 2008
[76] A P Goncalvez R Men C Wernly R H Purcell and C-J Lai ldquoChimpanzee Fab fragments and a derived humanizedimmunogiobulin G1 antibody that efficiently cross-neutralizedengue type 1 and type 2 virusesrdquo Journal of Virology vol 78no 23 pp 12910ndash12918 2004
[77] WM P BWahala A A Kraus L B HaymoreM A Accavitti-Loper and A M de Silva ldquoDengue virus neutralization byhuman immune sera role of envelope protein domain III-reactive antibodyrdquo Virology vol 392 no 1 pp 103ndash113 2009
[78] W D Crill H R Hughes M J Delorey and G-J J ChangldquoHumoral immune responses of dengue fever patients usingepitope-specific serotype-2 virus-like particle antigensrdquo PLoSONE vol 4 no 4 Article ID e4991 2009
[79] C Setthapramote T Sasaki O Puiprom et al ldquoHuman mono-clonal antibodies to neutralize all dengue virus serotypes usinglymphocytes from patients at acute phase of the secondaryinfectionrdquo Biochemical and Biophysical Research Communica-tions vol 423 no 4 pp 867ndash872 2012
[80] M Beltramello K L Williams C P Simmons et al ldquoThehuman immune response to dengue virus is dominated byhighly cross-reactive antibodies endowed with neutralizing andenhancing activityrdquo Cell Host andMicrobe vol 8 no 3 pp 271ndash283 2010
[81] C M Midgley M Bajwa-Joseph S Vasanawathana et al ldquoAnin-depth analysis of original antigenic sin in dengue virusinfectionrdquo Journal of Virology vol 85 no 22 pp 410ndash421 2011
[82] B Kaufmann M R Vogt J Goudsmit et al ldquoNeutralizationof West Nile virus by cross-linking of its surface proteins withFab fragments of the human monoclonal antibody CR4354rdquoProceedings of the National Academy of Sciences of the UnitedStates of America vol 107 no 44 pp 18950ndash18955 2010
[83] B Spurrier J M Sampson M Totrov et al ldquoStructural analysisof human and macaque mAbs 2909 and 25B implicationsfor the configuration of the quaternary neutralizing epitope ofHIV-1 gp120rdquo Structure vol 19 no 5 pp 691ndash699 2011
[84] J J SchlesingerMWBrandriss andT PMonath ldquoMonoclonalantibodies distinguish between wild and vaccine strains ofyellow fever virus by neutralization hemagglutination inhibi-tion and immune precipitation of the virus envelope proteinrdquoVirology vol 125 no 1 pp 8ndash17 1983
[85] M W Brandriss J J Schlesinger E E Walsh and M BrisellildquoLethal 17D yellow fever encephalitis in mice I Passive protec-tion by monoclonal antibodies to the envelope proteins of 17Dyellow fever and dengue 2 virusesrdquo Journal of General Virologyvol 67 no 2 pp 229ndash234 1986
[86] H Cho and M S Diamond ldquoImmune responses to West Nilevirus infection in the central nervous systemrdquo Viruses vol 4no 12 pp 3812ndash3830 2012
[87] D W Beasley A D Barrett and R B Tesh ldquoResurgence ofWest Nile neurologic disease in the United States in 2012 whathappened What needs to be donerdquo Antiviral Research vol 99no 1 pp 1ndash5 2013
[88] M Throsby J Ter Meulen C Geuijen J Goudsmit and J DeKruif ldquoMapping and analysis of West Nile virus-specific mon-oclonal antibodies prospects for vaccine developmentrdquo ExpertReview of Vaccines vol 6 no 2 pp 183ndash191 2007
[89] M S Diamond T G Roberts D Edgil B Lu J Ernst and EHarris ldquoModulation of dengue virus infection in human cells
20 BioMed Research International
by alpha beta and gamma interferonsrdquo Journal of Virology vol74 no 11 pp 4957ndash4966 2000
[90] Z Shimoni M J Niven S Pitlick and S Bulvik ldquoTreatmentof West Nile virus encephalitis with intravenous immunoglob-ulinrdquo Emerging Infectious Diseases vol 7 no 4 article 759 2001
[91] AHamdan PGreen EMendelsonM R Kramer S Pitlik andM Weinberger ldquoPossible benefit of intravenous immunoglob-ulin therapy in a lung transplant recipient with West Nile virusencephalitisrdquoTransplant InfectiousDisease vol 4 no 3 pp 160ndash162 2002
[92] M Haley A S Retter D Fowler J Gea-Banacloche and NP OrsquoGrady ldquoThe role for intravenous immunoglobulin in thetreatment of West Nile virus encephalitisrdquo Clinical InfectiousDiseases vol 37 no 6 pp e88ndashe90 2003
[93] K M Chung G E Nybakken B S Thompson et al ldquoAnti-bodies against West Nile virus nonstructural protein NS1prevent lethal infection through Fc 120574 receptor-dependent and-independent mechanismsrdquo Journal of Virology vol 80 no 3pp 1340ndash1351 2006
[94] E A Henchal L S Henchal and J J Schlesinger ldquoSynergisticinteractions of anti-NS1 monoclonal antibodies protect pas-sively immunized mice from lethal challenge with dengue 2virusrdquo Journal of General Virology vol 69 no 8 pp 2101ndash21071988
[95] J T Roehrig ldquoAntigenic structure of flavivirus proteinsrdquoAdvances in Virus Research vol 59 pp 141ndash175 2003
[96] M D Sanchez T C Pierson D McAllister et al ldquoCharacteri-zation of neutralizing antibodies to West Nile virusrdquo Virologyvol 336 no 1 pp 70ndash82 2005
[97] J H Beigel J L Nordstrom S R Pillemer et al ldquoSafety andpharmacokinetics of single intravenous dose of MGAWN1 anovel monoclonal antibody to West Nile virusrdquo AntimicrobialAgents and Chemotherapy vol 54 no 6 pp 2431ndash2436 2010
[98] C A Smeraski V Siddharthan and J D Morrey ldquoTreatmentof spatial memory impairment in hamsters infected with WestNile virus using a humanizedmonoclonal antibodyMGAWN1rdquoAntiviral Research vol 91 no 1 pp 43ndash49 2011
[99] D Lelli AMoreno E Brocchi et al ldquoWestNile virus character-ization and diagnostic applications of monoclonal antibodiesrdquoVirology Journal vol 9 article 81 2012
[100] T S Gritsun V A Lashkevich and E A Gould ldquoTick-borneencephalitisrdquo Antiviral Research vol 57 no 1-2 pp 129ndash1462003
[101] D Ruzek T S Gritsun N L Forrester et al ldquoMutations inthe NS2B and NS3 genes affect mouse neuroinvasiveness of aWestern European field strain of tick-borne encephalitis virusrdquoVirology vol 374 no 2 pp 249ndash255 2008
[102] M Ecker S L Allison T Meixner and F X Heinz ldquoSequenceanalysis and genetic classification of tick-borne encephalitisviruses from Europe and Asiardquo Journal of General Virology vol80 no 1 pp 179ndash185 1999
[103] H Holzmann M S Vorobyova I P Ladyzhenskaya et alldquoMolecular epidemiology of tick-borne encephalitis viruscross-protection between European and Far Eastern subtypesrdquoVaccine vol 10 no 5 pp 345ndash349 1992
[104] C W Mandl ldquoSteps of the tick-borne encephalitis virus repli-cation cycle that affect neuropathogenesisrdquo Virus Research vol111 no 2 pp 161ndash174 2005
[105] A V Atrasheuskaya T M Fredeking and G M IgnatyevldquoChanges in immune parameters and their correction in humancases of tick-borne encephalitisrdquo Clinical and ExperimentalImmunology vol 131 no 1 pp 148ndash154 2003
[106] K L Mansfield N Johnson L P Phipps J R Stephenson AR Fooks and T Solomon ldquoTick-borne encephalitis virusmdashareview of an emerging zoonosisrdquo Journal of General Virologyvol 90 no 8 pp 1781ndash1794 2009
[107] DWVaughn andCHHoke Jr ldquoThe epidemiology of Japaneseencephalitis prospects for preventionrdquo Epidemiologic Reviewsvol 14 pp 197ndash221 1992
[108] T Solomon H Ni D W C Beasley M Ekkelenkamp M JCardosa and A D T Barrett ldquoOrigin and evolution of Japaneseencephalitis virus in Southeast Asiardquo Journal of Virology vol 77no 5 pp 3091ndash3098 2003
[109] M R Wilson ldquoEmerging viral infectionsrdquo Current Opinion inNeurology vol 26 no 3 pp 301ndash306 2013
[110] J N Hanna S A Ritchie D A Phillips et al ldquoAn outbreakof Japanese encephalitis in the Torres Strait Australia 1995rdquoMedical Journal of Australia vol 165 no 5 pp 256ndash260 1996
[111] J N Hanna S A Ritchie D A Phillips et al ldquoJapanese ence-phalitis in north Queensland Australia 1998rdquo Medical Journalof Australia vol 170 no 11 pp 533ndash536 1999
[112] SGeorgeMGourie-Devi and J A Rao ldquoIsolation ofWestNilevirus from the brains of children who had died of encephalitisrdquoBulletin of theWorldHealthOrganization vol 62 no 6 pp 879ndash882 1984
[113] A K Gupta A A Koshy and V J Lad ldquoEnhanced protec-tion of mice against Japanese encephalitis virus infection bycombinations ofmonoclonal antibodies to glycoprotein ErdquoActaVirologica vol 55 no 2 pp 165ndash168 2011
[114] T H Lee B-H Song S-I Yun et al ldquoA cross-protective mAbrecognizes a novel epitope within the flavivirus NS1 proteinrdquoJournal of General Virology vol 93 no 1 pp 20ndash26 2012
[115] S C Weaver ldquoHost range amplification and arboviral diseaseemergencerdquo Archives of Virology Supplementum no 19 pp 33ndash44 2005
[116] M Solignat B Gay S Higgs L Briant and C Devaux ldquoRepli-cation cycle of chikungunya a re-emerging arbovirusrdquoVirologyvol 393 no 2 pp 183ndash197 2009
[117] O Schwartz and M L Albert ldquoBiology and pathogenesis ofchikungunya virusrdquo Nature Reviews Microbiology vol 8 no 7pp 491ndash500 2010
[118] W Akahata Z-Y Yang H Andersen et al ldquoA virus-like particlevaccine for epidemic Chikungunya virus protects nonhumanprimates against infectionrdquo Nature Medicine vol 16 no 3 pp334ndash338 2010
[119] M Kielian and A Helenius ldquopH-Iinduced alterations in thefusogenic spike protein of Semliki Forest virusrdquo Journal of CellBiology vol 101 no 6 pp 2284ndash2291 1985
[120] J White and A Helenius ldquopH-Dependent fusion between theSemliki Forest virus membrane and liposomesrdquo Proceedings ofthe National Academy of Sciences of the United States of Americavol 77 no 6 pp 3273ndash3277 1980
[121] J Dubuisson and C M Rice ldquoSindbis virus attachment isola-tion and characterization of mutants with impaired binding tovertebrate cellsrdquo Journal of Virology vol 67 no 6 pp 3363ndash33741993
[122] C Y Lee Y-W Kam J Fric et al ldquoChikungunya virus neutral-ization antigens and direct cell-to-cell transmission are revealedby human antibody-escapemutantsrdquo PLoS Pathogens vol 7 no12 Article ID e1002390 2011
[123] J E Voss M-C Vaney S Duquerroy et al ldquoGlycoproteinorganization of Chikungunya virus particles revealed by X-raycrystallographyrdquo Nature vol 468 no 7324 pp 709ndash712 2010
BioMed Research International 21
[124] L Li J Jose Y Xiang R J Kuhn and M G Rossmann ldquoStruc-tural changes of envelope proteins during alphavirus fusionrdquoNature vol 468 no 7324 pp 705ndash708 2010
[125] J Fric S Bertin-Maghit C I Wang A Nardin and L WarterldquoUse of human monoclonal antibodies to treat Chikungunyavirus infectionrdquo Journal of Infectious Diseases vol 207 no 2 pp319ndash322 2013
[126] P V Aguilar J G Estrada-Franco R Navarro-Lopez C FerroA D Haddow and S C Weaver ldquoEndemic Venezuelanequine encephalitis in the Americas hidden under the dengueumbrellardquo Future Virology vol 6 no 6 pp 721ndash740 2011
[127] S C Weaver R Winegar I D Manger and N L ForresterldquoAlphaviruses population genetics and determinants of emer-gencerdquo Antiviral Research vol 94 no 3 pp 242ndash257 2012
[128] S Paessler and S C Weaver ldquoVaccines for Venezuelan equineencephalitisrdquo Vaccine vol 27 no 4 pp D80ndashD85 2009
[129] R J Phillpotts ldquoVenezuelan equine encephalitis virus complex-specific monoclonal antibody provides broad protection inmurine models against airborne challenge with viruses fromserogroups I II and IIIrdquoVirus Research vol 120 no 1-2 pp 107ndash112 2006
[130] R J Phillpotts L D Jones and S C Howard ldquoMonoclonalantibody protectsmice against infection anddiseasewhen giveneither before or up to 24 h after airborne challenge with virulentVenezuelan equine encephalitis virusrdquoVaccine vol 20 no 11-12pp 1497ndash1504 2002
[131] J H Mathews J T Roehrig and D W Trent ldquoRole of com-plement and the Fc portion of immunoglobulin G in immunityto Venezuelan equine encephalomyelitis virus infection withglycoprotein-specific monoclonal antibodiesrdquo Journal of Virol-ogy vol 55 no 3 pp 594ndash600 1985
[132] L M OrsquoBrien C D Underwood-Fowler S A Goodchild AL Phelps and R J Phillpotts ldquoDevelopment of a novel mon-oclonal antibody with reactivity to a wide range of Venezuelanequine encephalitis virus strainsrdquoVirology Journal vol 6 article206 2009
[133] L M OrsquoBrien S A Goodchild R J Phillpotts and S D Per-kins ldquoA humanised murine monoclonal antibody protectsmice from Venezuelan equine encephalitis virus Evergladesvirus and Mucambo virus when administered up to 48h afterairborne challengerdquo Virology vol 426 no 2 pp 100ndash105 2012
[134] N Clementi N Mancini L Solforosi M Castelli M Clementiand R Burioni ldquoPhage display-based strategies for cloning andoptimization of monoclonal antibodies directed against humanpathogensrdquo International Journal of Molecular Sciences vol 13no 7 pp 8273ndash8292 2012
[135] A R Hunt R A Bowen S Frederickson T Maruyama JT Roehrig and C D Blair ldquoTreatment of mice with humanmonoclonal antibody 24h after lethal aerosol challenge withvirulent Venezuelan equine encephalitis virus prevents diseasebut not infectionrdquo Virology vol 414 no 2 pp 146ndash152 2011
[136] W-G Hu A L Phelps S Jager et al ldquoA recombinant human-ized monoclonal antibody completely protects mice againstlethal challenge with Venezuelan equine encephalitis virusrdquoVaccine vol 28 no 34 pp 5558ndash5564 2010
[137] T Rulker L Voss P Thullier et al ldquoIsolation and characterisa-tion of a human-like antibody fragment (scFv) that inactivatesVEEV in vitro and in vivordquo PLoS ONE vol 7 no 5 Article IDe37242 2012
[138] C T Walter and J N Barr ldquoRecent advances in the molecularand cellular biology of bunyavirusesrdquo Journal of General Virol-ogy vol 92 no 11 pp 2467ndash2484 2011
[139] C A Whitehouse ldquoCrimean-Congo hemorrhagic feverrdquo Anti-viral Research vol 64 no 3 pp 145ndash160 2004
[140] M Mertens K Schmidt A Ozkul and M H Groschup ldquoTheimpact of Crimean-Congo hemorrhagic fever virus on publichealthrdquo Antiviral Research vol 98 no 2 pp 248ndash260 2013
[141] A A Ahmed J M McFalls C Hoffmann et al ldquoPresence ofbroadly reactive and group-specific neutralizing epitopes onnewly described isolates of Crimean-Congo hemorrhagic fevervirusrdquo Journal of General Virology vol 86 no 12 pp 3327ndash33362005
[142] R Hewson A Gmyl L Gmyl et al ldquoEvidence of segment reas-sortment in Crimean-Congo haemorrhagic fever virusrdquo Journalof General Virology vol 85 no 10 pp 3059ndash3070 2004
[143] N K Blackburn T G Besselaar A J Shepherd and R Swane-poel ldquoPreparation and use of monoclonal antibodies for iden-tifying Crimean-Congo hemorrhagic fever virusrdquo AmericanJournal of Tropical Medicine and Hygiene vol 37 no 2 pp 392ndash397 1987
[144] T Ikegami ldquoMolecular biology and genetic diversity of RiftValley fever virusrdquoAntiviral Research vol 95 no 3 pp 293ndash3102012
[145] KKeegan andM S Collett ldquoUse of bacterial expression cloningto define the amino acid sequences of antigenic determinantson the G2 glycoprotein of Rift Valley fever virusrdquo Journal ofVirology vol 58 no 2 pp 263ndash270 1986
[146] T G Besselaar and N K Blackburn ldquoTopological mapping ofantigenic sites on the Rift Valley fever virus envelope glycopro-teins using monoclonal antibodiesrdquo Archives of Virology vol121 no 1ndash4 pp 111ndash124 1991
[147] L G Presta ldquoEngineering of therapeutic antibodies to mini-mize immunogenicity and optimize functionrdquo Advanced DrugDelivery Reviews vol 58 no 5-6 pp 640ndash656 2006
Submit your manuscripts athttpwwwhindawicom
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anatomy Research International
PeptidesInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporation httpwwwhindawicom
International Journal of
Volume 2014
Zoology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Molecular Biology International
GenomicsInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioinformaticsAdvances in
Marine BiologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Signal TransductionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Evolutionary BiologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Biochemistry Research International
ArchaeaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Genetics Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in
Virolog y
Hindawi Publishing Corporationhttpwwwhindawicom
Nucleic AcidsJournal of
Volume 2014
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Enzyme Research
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Microbiology

10 BioMed Research International
could neutralize 17D YFV virus only group B 17D and Asibistrains only group C Asibi strain only group D could onlyneutralize Asibi but not 17D strain showing also reactivityagainst DENV2 Zika or Banzi viruses All of the four mAbsbelonging to this group were able to cross-react with Zikaandor Banzi viruses and two (4E11 and 5H3) neutralizedDENV2 virus with 4E11 neutralizing also Banzi virus The3E9 mAb the only component of the group E could notneutralize 17D nor Asibi strains
All of the IgG mAbs resulted able to protect bothprophylactically and therapeutically BALBc or CD-1 micefrom lethal intracerebral challenge with 17D-204 strain [85]The chimeric form of the group B mAb 2C9 IgG (2C9-cIgG) was able to provide AG129 mice a 72 survival whenadministered 24 hours before infection with 17D-204 strainIts murine form could provide the survival of the 95 ofthe mice Appreciable results were also obtained when bothmurine 2C9 and 2C9-cIgG were administered 24 hours post-infection with the survival of 70 and 20 of mice respec-tively Viral presence was not detected in survivingmice [60]
In 2005 Daffis and colleagues constructed two Ab-phage libraries by cloning the repertoire of YFV-infectedpatients Panning was then performed with YFV-17D virionsThe scFv-5A 7A and R3(27) showed a neutralizing activityspanning from50 to 100 inPRNTassays against bothYFV17D-204-WHO and Asibi strains In further tests reactivitywas observed against wild-type YFV strains of West Africagenotype I and II (Nigeria 1987 and Asibi strains resp)and EastCentral Africa (CAR 1986 Ethiopia 1961 strains) Aconcentration from 01 to 03120583gmL could yield a 90 plaquereduction Reactivity was also observed against the strainSenegal 1990 in a lesser extent Production of escape variantscould demonstrate that scFv-5A 7A and R3(27) epitopes arebuilt up by residues extensively separated in themonomers ofE glycoprotein that however result in closed proximity whenthe homodimeric form of E is constituted [61]
26 West Nile Virus (WNV) The West Nile virus (WNV) isan epidemic neurotropic virus estimated to be responsibleof about 36000 cases and 1500 deaths registered in theUnited States between 1992 and 2012 [86] WNV antigenicityallows its classification into the Japanese encephalitis virus(JEV) serocomplex Genomic analysis has revealed two maingenetic lineages of WNV lineage I viruses circulating in theUSA Europe the Middle East Africa India and Australiaand lineage II viruses isolated from Sub-Saharan Africa andMadagascar [1]
WNV was firstly identified in 1937 and is endemicin many countries of Africa Middle East and West AsiaHowever after 1990 frequent outbreaks of WNV infectionswere reported in Romania Israel and later in North Americaand across the USA in 1999 and recently in 2012 [87]
Most of WNV infected individuals do not develop symp-toms while about 20 develop a self-limiting illness calledWest Nile fever Acute symptoms include fever tirednessand swollen lymph glands but in a minority of cases alsoencephalitis with long-term deficits in cognitive functionand motor skills have been reported However in less than
1 of infected patients WNV having crossed the blood-brain barrier is responsible of neuroinvasive and potentiallylethal form of the disease In these cases degeneration andapoptosis upon infection of neurons and the consequentinflammatory response can occur Related symptoms includehigh fever coma muscle weakness and paralysis Immun-odepression and advanced age have been correlated with anaugment risk to develop a severe disease [87] There are nospecific treatment options or licensed vaccines for humans[88]
WNV is transmitted by infected mosquitos and initialreplication is thought to occur primarily in dendritic cells inthe skin which migrate to secondary lymphoid tissues wherethe replicating virus enters the circulation To date onlyefficacious WNV veterinarian vaccines have been licensedwhile there are no licensed vaccines for protection againstWNV in humans [88]
261 mAbs against WNV Broad-spectrum antivirals suchas type I IFN-120572 ribavirin mycophenolic acid in WNVinfection showed ineffective results in vivo despite in vitrothey showed some activity [89]
However experiments in murine models extrapolationof clinical data as well as passive administration of pooledimmune-120574-globulins (OmriGam) containing a significanttiter of neutralizing Abs before and after infection showedan important role of the humoral immune response incontrolling viremia and prevent viral dissemination Indeedthe development of a neutralizing mAb-based therapy seemsto be encouraging for a possible treatment of infected patients[90ndash92]
Similarly to what has been observed for DENV only anti-E Abs have been identified as neutralizing and protectiveAlso anti-NS1 Abs have shown a protective role howevertheir mechanism of protection has not yet been elucidated asthe NS1 protein is secreted from infected cells and not presenton the virions [93ndash95]
In this regard similarly to what has been describedfor DENV studies performed using a naıve human scFvlibrary for panning with purified WNV E protein supportthe hypothesis that no Abs against the neutralizing DIII canbe isolated However DIII specific Abs were isolated in asubsequent study using immunoglobulin libraries obtainedfrom threeWNV infected patients for biopanning on purifiedinactivated virus virus-like particles consisting of prM andE proteins or recombinant E glycoprotein Although theproportion of DIII-specific mAbs was low (8) comparedwith anti-DII mAbs (47) Two out of the four anti-DIIImAbs were potently neutralizing and protective in vivowhereas only three out of the 24 anti-DII were weaklyneutralizing in vitro and non-protective in vivo In particularresidues that are critical for neutralization lies on the regionsspanning amino acids 305ndash312 330ndash333 and 365 that arelocated on adjacent exposed loops ofDIIIHowever sequencealignment of E protein of different flaviviruses such as DENVand JEV revealed a considerable variation compared tothe whole E protein This observation further suggests thatdifferently from broadly cross-reactive anti-DII Abs anti-DIII neutralizing Abs are virus-type specific However of
BioMed Research International 11
these mAbs two (CR4374 and CR4353) protected mice fromlethal WNV challenge at 50 protective doses of 129 and357 120583gkg of body weight respectively [50]
Sanchez et al using three different immunization strate-gies (ie inactivated virus naked DNA and recombinantprotein) isolated ninemurinemAbsmost of which bound toconformation-dependent epitopes in DIII of the E protein Inparticular neutralizing mAbs named 8B10 11C2 10C5 and17C8 were obtained from mice immunized with inactivatedvirus alone or in combination with a DNA plasmid andbound to the same region of DIII with high affinity Incontrast mAbs obtained by immunization with a solubleversion of the E glycoprotein did not exhibit neutralizingactivity These non-neutralizing mAbs were cross-reactivewith several other flaviviruses including SLEV JEV YFVand Powassan virus confirming the conserved nature ofFlavivirus non-neutralizing epitopes [96]
Gould et al isolated 11 unique human single-chain vari-able region Ab fragments (scFvs) that bind the E protein ofWNVAmong them a humanmAb namedmAb11 expressedas a scFv-Fc fusion protein was further characterized Itrecognizes the fusion loop at the distal end of DII ofthe WNV E protein and cross-reacts with all four DENVserotypes and provides protection against DENV2 and 4as well as WNV [42] Moreover therapeutic studies of thismAb in WNV-infection model mice provided substantialprotection when administered after 5 days post-infectionInterestingly a neutralization escape variant of this mAbfailed to cause lethal encephalitis (at higher infectious doses)or induce the inflammatory responses associated with blood-brain barrier permeability in mice compared to the parentalWNV suggesting an important role for the fusion loop inviral pathogenesis [43]
Oliphant et al isolated an anti-DIII E protein mAbnamed E16 fromhybridomas obtained after immunization ofmice with recombinant WNV E protein which neutralizedall WNV strains with PRNT50 values of 4 to 18 ng andPRNT90 values of 53 to 297 ng One hundred microgramsof mAb protected greater than 90 of mice from lethalinfection and even a single 4120583g treatment of E16 on day 2 afterinfection prevented mortality Moreover humanization ofthis mAb confirmed as therapeutically effective in mice [44]Subsequent studies revealed that mAb E16 neutralization ismediated by engagement of four discontinuous segmentsof DIII including the amino-terminal region (amino acidresidues 302ndash309) and the three connecting loops BC (aminoacid residues 330ndash333) DE (amino acid residues 365ndash368)and FG (amino acid residues 389ndash391) Moreover no ADE ofinfection was detected when E16 mAb was used at saturatingconcentrations [45 47]
Furthermore results of a Phase I safety study of thehumanized E16 mAb (designated MGAWN1) have beenreported and suggested that doses of up to 30mgkgwerewelltolerated with few mild adverse events and would providean excess of virus neutralizing activity for 3-4 weeks aftertreatment However a case of anti-MGAWN1 Ab elicitationoccurred with the consequent increased rate of clearance andindeed impacting efficacy A Phase II safety and efficacy studyof MGAWN1 is ongoing [97] Furthermore the possibility of
preventing or treating WNV-induced memory deficits wasrecently investigated In this study hamsters were treatedintraperitoneally with 32mgkg ofMGAWN1mAb at 45 daysafter subcutaneously challenging with WNV InterestinglyMGAWN1 prevented mortality weight loss and improvedfood consumption of WNV-infected hamsters compared tocontrols [98]
Recently Lelli et al isolated six anti-E mAbs frominactivated-WNV immunized mice In particular three ofthem (3B2 3D6 and 4D3) neutralized lineage I and II WNVwith the first two recognizing the same epitopes located onthe distal lateral surface of DIII (critical amino acid residueK307) Conversely 4D3mAb recognized a novel neutralizingepitope on DII (critical residues S276 and T278) Indeedfurther protective and therapeutic studies are needed toascertain their neutralizing activity in vivo [99]
Finally to conclude like DENV infection Abs directedagainst the M protein are not protective and neutralizingMoreover in humans a skewed humoral immune responseagainst DII has been frequently observed and confirmedalso with the hybridoma technology [50] The isolation andelicitation of neutralizingAbs directed against the fusion loopand DIII of the E protein represent thus the most challengingand promising goal for the development of new effectivetherapeutic inhibitors and immunogens respectively
27 Tick-Borne Encephalitis Virus (TBEV) Tick-borne ence-phalitis virus (TBEV) is one of the most dangerous agentscausing human neuroinfections occurring mostly in Europeand Asia and with a potential fatal prognosis [100] TBEV isbelieved to cause 3500ndash10000 human cases of encephalitisin Europe per year with a high morbidity in Russia CzechRepublic Austria and Germany In particular between 1990and 2007 an average of 8755 cases of TBE was reported peryear in Europe and Russia Despite the fact that Russia isthe country with most infections registered annually CzechRepublic incidence is among the highest in Europe with 400ndash1000 clinical cases reported every year [101]
Three subtypes of TBEV are classified namely EuropeanSiberian and Far-Eastern sharing most of the genetic andantigenic features [102] In fact a high degree of antigenichomogeneity between different strains of TBEV has beendescribed [103]
Clinical manifestations vary among the subtypes butusually they start with a short febrile period of 7ndash14 daysafter the tick bite Fatigue headache and pain in the neckshoulders and lower back together with high fever andvomitingmay be present [100]Thesemanifestations are oftenfollowed by an asymptomatic phase lasting from 2 to 10days after remission from the fever with possible progres-sion to neurological disease Neurological symptoms includemeningitis encephalitis myelitis and radiculitis Mortalityoccurs in 1-2 of the European subtype-infected patientsbut fatal prognosis can occur in up to 20ndash40 of the Far-Eastern subtype-infected patients [104] Mortality rates ofthe Siberian subtype are similar to those observed for theEuropean [105]
Effective and safe vaccines against TBEV produced frominactivated virus have been developed and licensed for their
12 BioMed Research International
use in humans However an emergency therapy in theabsence of a mass immunization is needed as no effectivetreatments are yet available [106]
271 mAbs against TBEV Levanov et al described the chim-erization of two murine mAbs (13D6 and 10C2) directedagainst the DIII and DII respectively of the TBEV E glyco-protein The chimeric mAbs present binding characteristicssimilar to the parentalmAbsMoreover as the parentalmAbsonly the chimeric mAb 13D6 was able to neutralize TBEVinfectivity in vitro In particular neutralization studies withthe murine chimeric and scFv forms of mAb 13D6 were per-formed In particular murine 13D6 showed an IC50 titer of115 120583gmL in Focus Reduction Neutralization Tests (FRNT)against TBEV strain 205 and of 29 120583gmL in PRNT againstTBEV strain Softjin Chimeric 13D6 showed an IC50 titer of45 120583gmL in FRNT against TBEV strain 205 and of 19 120583gmLin PRNT against TBEV strain Softjin ScFv 13D6 showed anIC50 titer of 167 120583gmL in FRNT against TBEV strain 205and of 112 120583gmL in PRNT against TBEV strain Softjin [62]
28 Japanese Encephalitis Virus (JEV) Japanese encephalitisvirus (JEV) is a mosquito-borne flavivirus that over thepast few decades has caused several outbreaks throughoutChina Southeast Asia Australia and Papua New Guineawith a prevalence recently estimated to be of about 70000casesyear [107ndash109] A 40 mortality was recorded in someof the JEV-affected areas Moreover many survivors facesome neurological problems and complications [108] Since1995 the disease has also emerged in Non-Asian regions suchas Northern Australia [110 111] The situation in SoutheastAsia however is further complicated by the overlappingepidemics of JEV and DENV as well as sporadic casesof WNV infections detected in some of the affected areasparticularly in India [112]
Effective vaccines both live-attenuated and inactivatedstrains of JEV have been developed and licensed in majorcountries However as for the other arthropod-borne Fla-vivirus members no specific antiviral drugs are currentlyavailable [4]
281mAbs against JEV Aspreviously described for the otherFlavivirusmembers many groups isolated and characterizedanti-JEVmAbs showing different neutralizing and protectiveproperties In this regard Gupta et al used combinations ofanti-E JEV mAbs (Hs1-4) in mice protection experimentsIn particular they found that the singularly mAb protectionranged from 45 to 65 when 100120583g of mAb were adminis-tered while equimolar combinations of two or three mAbsgave 85ndash90 or 100 protection respectively [113] In similarexperiments Lee et al demonstrated that a Flavivirus anti-NS1 mAb named 16NS1 cross-reacted with JEV as well asWNV and exhibited protective activity against WNV as wellas a lethal JEV infection However no neutralizing activitywas observed using this mAb against both WNV and JEV inin vitro experiments suggesting the participation of otherAb-mediatedmechanisms in vivo In particular 95ofmicewere
protected when 500120583g of mAb were administered intraperi-toneally and concomitantly to intramuscular injection of JEV
Overlapping peptide mapping analysis combined withsite-specific mutations identified the 116KAWGKSILFA125region and critical amino acid residues W118 and I122 as16NS1 mAb epitope highly conserved in WNV and JEVstrains [114]
Arakawa et al isolated from a combinatorial human Fablibrary constructed from peripheral blood lymphocytesobtained from JEV hyperimmune volunteers Among 188randomly selected clones FabTJE12B02 showed the best 50focus reduction endpoint at the concentration of 502 120583gmLagainst the JEV strain Nakayama [55]
Goncalvez et al isolated three mAbs named Fabs A3B2 and E3 by repertoire cloning from chimpanzees initiallyimmunized with inactivated JE-VAX and then boosted withattenuated JEV SA14-14-2 In particular these mAbs reactedwith epitopes in three different E domains in DI (amino acidresidue K179) in DII (I126) and in DIII (G132) for Fabs A3B2 and E3 respectively Moreover these Fabs as well as thederived humanized counterpart mAbs exhibited high neu-tralizing activities against a broad spectrum of JEV genotypestrains Moreover these mAbs exhibited a 50 protectivedose of 084 120583g (B2) 58120583g (A3) and 247120583g (E3) in mousemodels Finally administration of 200 120583gmouse of mAb B2one day after otherwise lethal JEV infection protected 50of mice and significantly prolonged the average survival timecompared to that of mice in the unprotected group [54]
29 St Louis Encephalitis Virus (SLEV) St Louis encephalitisvirus (SLEV) was first discovered as the mosquito-borneagent responsible for over 1000 cases of encephalitis duringa 1933 summer outbreak in St Louis (Missouri) and nowis a reemerging human pathogen widely distributed in theAmerican continent causing several human encephalitisoutbreaks over the last 80 years Additional epidemics haveindeed occurred from 1964 to 2006 in the Americas rangingfrom the US to Argentina and Brazil As a member of theFlavivirus genus the SLEVE glycoprotein ectodomain is 68identical to serocomplex-related JEVEbut only 46 and 40identical to those ofDENV2 andTBEV viruses fromdifferentserocomplexes [115]
291 mAbs against SLEV In 1983 Roehrig et al isolatedtwenty-one hybridomas producing murine mAbs specificfor the E glycoprotein of SLEV strain MSI-7 Serologicreactivities were initially determined by cross-reactivity indi-rect immunofluorescence assays using 22 strains of SLEVand 8 other related flaviviruses Four groups demonstratingtype- subcomplex- supercomplex- and group-specific reac-tivity patterns were identified Analysis of hemagglutination-inhibition and virus neutralization subdivided the cross-reactivity groups into eight epitopes (E-1a b c d E-2 E-3 and E-4a b) one of them following localized in DII (4b)[52 53]
Moreover among the previously described isolated anti-WNV mAbs by Gould et al the chimeric scFv-Fc 79 effec-tively neutralized also SLEV resulting in gt80 of PRNT80when used at 5 120583gmL [42]
BioMed Research International 13
3 Alphaviruses
Major human arboviruses in the Togaviridae family Chikun-gunya (CHIKV) and Venezuelan equine encephalomyelitis(VEEV) viruses belong to the Alphavirus genus which iscomposed of viruses with an icosahedral nucleocapsid sur-rounded by a lipid envelope and glycoprotein spikes Thestructural proteins of alphaviruses arise through co- andpost-translational processing of a polyprotein encoded by asingle positive-stranded RNAproducing the capsid (C) PE26 K and E1 The PE2 glycoprotein is a precursor containingthe E3 glycoprotein fused to the amino terminus of the E2envelope glycoprotein The PE2 glycoprotein is followed by6K a small membrane-associated protein and E1 the secondpolypeptide component of glycoprotein spikes
Trimerized heterodimers of the E1 and E2 viral glyco-proteins form the surface spikes and contain determinantsof viral tropism and virulence The E3 glycoprotein acts as asignal for transport of PE2 across themembranes of the roughendoplasmic reticulum and may promote the formation andintracellular transport of E1-PE2 heterodimers to the cellsurface During transport to the cell surface PE2 undergoesa maturational cleavage event by a furin-like protease toproduce E2 and E3 [116]
The E2 glycoprotein promotes specificity of virus bindingto the host cell surface and is a target of the humoral immuneresponse The E1 glycoprotein mediates fusion of the virionenvelope with the membranes of acidified endosomes allow-ing release of the nucleocapsid into the cytoplasm and theonset of viral replicationAlphavirus E1 shares no appreciablesequence identity to Flavivirus E protein Despite differencesin their amino acid sequences and arrangements on the viralparticle the structures of E and E1 are remarkably similarIndeed the conservation of three domains has been welldocumented in crystal structures of flavivirus E ectodomainsfrom TBEV DENV JEV and WNV as well as Alphavirus E1from Semliki Forest virus (SFV) and CHIKV Abs to the E1glycoprotein do not typically neutralize virus infectivity invitro but can protect against lethal challenge in animals
By analogy with the prM protein of dengue Flavivirusfurin cleavage of PE2 may begin after transit of an acidiclate component of the Golgi body where E3 is thought tosuppress the acid pH-triggered activation of glycoprotein E1fusion capability Protective anti-E3 mouse mAbs have alsobeen described [117]
31 Chikungunya Virus (CHIKV) The chikungunya virus(CHIKV) belongs to the Semliki Forest clade and was firstlyisolated in 1953 in Tanzania during an epidemic outbreakfollowing occurred also in Asia and Africa In the last fivedecades several CHIKV outbreaks have been described bothin Africa and Asia separated by gaps lasting from two totwenty years In 2005-2006 about 300000 cases out of785000 inhabitants were reported in La Reunion island witha fatal prognosis for 237 of the patients [117] Neither Europenor the Americas have had outbreaks of CHIKV so far exceptfor imported isolated cases Three genotypes of CHIKV aredescribed Asian EastCentralSouth African (ECSA) andWest African with an amino acid identity spanning from
952 to 998 Recent epidemics in Africa and Indiansubcontinent were caused by strains belonging to the ECSAgenotype Transmitted by Aedes mosquitoes CHIKV ismaintained in the human population by human-mosquito-human transmission [1] The disease is characterized bydengue-like symptoms such as chills high fever headacheand persistent myalgia and a further incapacitating arthralgia(from which the name chikungunya) which affects 40 ofinfected subjects However prognosis is rarely fatal [116]
There is currently no commercial vaccine and antiviraltreatment forCHIKV although some candidate vaccines havebeen tested in humans In this regard in 2000 US Armyperformed a Phase II clinical trial testing a live-attenuatedCHIKV vaccine (TSI-GSD-218) derived from a 1962 strain(15561) of an outbreak in Thailand Out of 58 every patientdeveloped neutralizingAbs and 5 lamentedmild tomoderatejoint pain [117] A phase III trial of this candidate vaccineis ongoing Furthermore a new formulation using virus-likeparticles was able to induce neutralizing Abs in macaquesagainst different CHIKV strains [117 118] Indeed it has beendescribed that infection seems to elicit long-lasting protectiveimmunity and cross-protection among CHIKV and otheralphaviruses
CHIKV entry into host cell is demonstrated to be medi-ated by envelope glycoproteins E1 and E2 which allowvirus fusion to cell membrane in low pH conditions andrecognition of an unknown cellular receptor respectively[116 119ndash121] Even though the genome replication relies onan error-prone RNA-dependent RNA-polymerasis CHIKVstrains S27 and 05115 Reunion showed in E1 and E2 alow amino acid variation of 068 and 33 respectivelyand such a low variability may be needed for an effectivereplication in two phylogenetically distant hosts [116 122]Recently the crystal structure of E1 and E2 heterodimerhas been resolved both at low pH and natural conditionsshedding light on E1 and E2 ectodomainsrsquo structure Inparticular similarly to flaviviruses E protein E1 ectodomainis made of the N-terminal DI the DII containing the fusionloop and the DIII at the C-terminal On the other hand E2ectodomain comprises theN-terminal domainA the domainB supposed to interact with the host cellrsquos unknown receptorand domain C at the C-terminal E1 DIII and E2 domain Care located close to the viral membrane [123 124]
311 mAbs against CHIKV Warter et al described 5F10 and8B10 two human mAbs which strongly neutralized severalCHIKV isolates in vitro without cross-reactivity againstother alphaviruses but to Onyong-nyong virus [56] Mixedpreparation of 5F10 and 8F10 did not show neither synergisticnor addictive effect in vitro and studies upon escape mutantsdemonstrated that 5F10 mAb binds at the tip of the E2domain B while 8B10 recognizes residues close to E1 fusionloop and amino acids within E2 domain A which form atransitional epitope under low pH conditions The authorsspeculated that 5F10 and 8B10 may inhibit CHIKV entry andfusion to the cell membrane respectively It is worth notingthat the previously mentioned escape mutants displayedmutations associated with reduced viral fitness in vitro evenafter 13 neutralizationamplification rounds In the same
14 BioMed Research International
study cell-to-cell transmission was firstly demonstrated as anescapemechanism enhanced by the E2mutation R82G [122]
In further in vivo studies 5F10 and 8B10 significantlydelayed CHIKV-caused death of AGR129 mice both in pro-phylactic and therapeutic tests Interestingly in therapeutictreatment these mAbs showed a synergistic effect whenadministered in combination with the total amount of mAbsinjected being the same of the single-mAbs Possibly thesetreatment did not allow themice to survive CHIKV challengedue to Ig clearing and 6ndash10 days half-life usually described forhuman mAbs [125]
Recently Pal et al cloned thirty-six murine mAbs ableto neutralize the ECSA La Reunion 2006 OPY-1 strain ofCHIKV (CHIKV-LR) the majority of which also neutralizeinfection of other strains corresponding to the Asian andWest African genotypes In particularmAbCHK-152 showedthe highest and broadest neutralizing activity with FRNTvalues indicating an IC50 value of 1 to 3 ngmL dependingon the viral strain used Among the thirty-six mAbs four(CHK-102 CHK-152 and CHK-166 CHK-263) could pre-vent lethality of immunodeficient Ifnarminusminus C57BL6 micewhen administered one day before exposition with CHK-152 and CHK-263 showing the ability to protect mice at thelowest dose (10 120583g) Therapeutic studies were performed onthese mAbs administering a single dose of 100 120583g 24 hoursafter infection The highest activity was described for CHK-166 able to protect 63 of the mice Testing the combinedactivity of the mAbs in therapeutic studies administration ofCHK-166 plus CHK-152 in a dose of 250120583g each turned out tobe themost effective protecting 71 of the IfnarminusminusC57BL6mice 60 hours after infection [57]
32 Venezuelan Equine Encephalomyelitis Virus (VEEV)Venezuelan equine encephalomyelitis virus (VEEV) is main-tained in a natural transmission cycle between mosquitoesand small rodents The first documented outbreaks occurredin the 1930s and several epidemics have been reported so farin Latin-American countries such as Venezuela ColombiaPeru Ecuador Bolivia Honduras Mexico and Panama withhundreds of thousands of human cases being reported anda case-fatality rate up to 1 during the 1969 Ecuador out-break Clinical manifestations of VEE are indistinguishablefrom Dengue [126] Six serogroups (IndashVI) are currentlyrecognized within the VEEV complex VEEV caused humanand equine outbreaks in the Americas for nearly a centuryEquine epizootics have high mortality (38ndash83) leading toa high viremia followed by a lethal encephalitis and often tohuman epidemics involving thousands of cases and hundredsof deaths Indeed VEEV is infectious for humans by theairborne route and has been responsible for a number oflaboratory infections In humans the disease is usually self-limiting with a febrile illness with 1ndash4 of cases progressingto severe encephalitis It is a hazard to laboratory workershas been developed as a biological weapon and is a potentialbioterrorist agent [127] There are no antiviral drugs andvaccines licensed for the treatment of VEEV infection inhumans Indeed although experimental live-attenuated vac-cines have been developed (eg TC-83) with good levels ofprotection in equine and mice in humans they may fail to
give protection in the majority of cases [128] Consideringthe protective role exerted by humoral immune response inanimal models it could be concluded that treatment withspecific IgG may have a beneficial antiviral effect in humanairborne infections with pathogenic strains of VEEV
321 mAbs against VEEV Several works support that mAbsmay protect against airborne VEEV as well as after airborneexposure toVEEVAb reactivity to both surface glycoproteins(E1 and E2) is associated with neutralization of the virusin vitro and passive protection against virus challenge Aseries of anti-E1 and anti-E2 mouse and humanized mAbshave been described recently Among them Phillpotts et alexamined two anti-E2 murine mAbs named 1A4A-1 and1A3A-9 both having potent protective activity against subcu-taneous VEEV challenge in mice Both mAbs had a similarhalf-life (58 and 100 days) in mouse serum after a singleintraperitoneal dose suggesting that mAbs delivered by thisroute at or around the time of VEEV infection persists athigh levels in the blood as well as secretions in respiratorytransudation throughout the clinical course of the disease Inparticular both mAbs administered 24 h prior to airbornechallenges had a substantial protective effect (90ndash100)Treatment of mice 2 or 24 h after airborne infection with asingle intraperitoneal dose of 100 120583g of 1A3A-9 mAb led toapproximately 50 survival There was no beneficial effectwhen mAb treatment was delayed to 72 h Moreover therewas evidence of synergy in vitro in PRNT between 1A3A-9and 1A4A-1 as has been demonstrated for other virusesand mAb pairs However no synergy was found in mouseprotection when mAbs were delivered intraperitoneally as amixture in equal parts to animals challenged 24 h previouslywith VEEV [129]
In a subsequent study Phillpotts et al reported two otherprotective mAbs The 3B2A-9 mAb protected against allthe serogroup I strains while 1A3B-7 mAb protected wellagainst challenge with all of the viruses tested Both 3B2A-9 and 1A3B-7 protected against airborne exposure to theIAB serogroup virus strainTrinidad donkey (TrD)Howeverthere was no evidence of synergistic protection when thesemAbs were combined in equal proportions An intraperi-toneal dose of 10 120583g was sufficient to protect 50 of themice with either mAb [130] According to other data themechanism of protection did not appear to depend uponneutralization [131] Similar results were obtained by OrsquoBrienet al which described a non-neutralizing mAb (IgG2a)named CUF37-2a able to protect 50 of mice from asubcutaneous VEEV challenge when a dose of 915 120583g of mAbwas administered 24 h prior to challenge [132]
In a following study the 1A3B-7 mAb was humanized(and following reported as Hu1A3B-7) maintaining the samefeatures of its murine counterpart In particular evaluation ofin vitro studies indicated that Hu1A3B-7 retained both broadspecificity and neutralizing activity Furthermore in vivoexperiments showed that Hu1A3B-7 successfully protectedmice against lethal subcutaneous and aerosol challenges withVEEV strain TrD Moreover the effectiveness of the human-ization process was determined by assessing proliferationresponses in human T-cells to peptides derived from the
BioMed Research International 15
murine and humanized versions of the VH and VL domainsThis analysis showed that the number of human T-cellepitopes within the humanized Ab had been substantiallyreduced indicating that Hu1A3B-7 may have low immuno-genicity in vivo [133]
Similarly Hunt et al described a humanizedmurinemAbderived from the 3B4C-4 mAb recognizing the E2c epitope(amino acid residues 182ndash209) and following named Hy4-26C showing neutralizing activity similar to the murinemAb Moreover it was protective (70ndash100 of survival) at100 ng dose in mice animal model intraperitoneal inoculatedand at 500120583g for intranasal challenge (80) Therapeuticstudies in mice revealed that Hy4-26C was able to cure miceup to 24 h following infection at 10 120583g on the contrary themouse mAb when given 1 hour after virus challenge [63]
In another work Hunt et al isolated two neutralizinghumanized Fabs F5 and L1A7 employing a blocking strategy[134 135] The anti-E2 specific F5 IgG had a 70 PRNTendpoint of 10 ngmL equivalent to that described for themost effective neutralizing anti-VEEVE2mAbs In particularthe E1-specific hFab L1A7 had a PRNT endpoint of 3120583gmL300-fold lower than F5
Further studies revealed that F5 epitope is located inthe 115ndash199 amino acid region Moreover F5 IgG had potentability to protect mice from infection by either route whenadministered 24 h before exposure however mice treated24 h and 48 h after aerosol exposure developed central ner-vous system infections but exhibited no clinical signs ofdisease [135]
Hu et al reported that passive immunization with thehumanized chimericmousemAbhu1A4A1IgG1-2A inmice at50 120583g 24 h before or after virulent VEEV challenge providedcomplete protection indicating that hu1A4A1IgG1-2A haspotent prophylactic and therapeutic effects against VEEVinfection [136]
Anti-VEEV mAbs isolated from a non-human primategene library has also been reported In particular thehumanized chimeric scFv-Fc ToR67-3B4 recognized viableas well as formalin and 120573-propiolactone inactivated virusparticles It detected specifically the viral E1 envelope proteinof VEEV but did not react with reduced viral glycopro-tein preparations suggesting that recognition depends uponconformational epitopes The recombinant Ab was able todetect multiple VEEV subtypes and displayed only marginalcross-reactivity to otherAlphavirus species except for Easternequine encephalitis virus (EEEV) In addition the scFv-Fcfusion described here might be of therapeutic use since itsuccessfully inactivated VEEV in a murine disease modelIn particular when the recombinant Ab was administered 6hours after challenge 80 to 100 of mice survived lethalVEEV IAB or IE infection Forty to sixty percent of micesurvived when scFv-Fc ToR67-3B4 was applied 6 hours afterchallenge with VEEV subtypes II and former IIIA [137]
4 Bunyaviridae
TheBunyaviridae family contains human arboviruses belong-ing to the Orthobunyavirus Phlebovirus and Nairovirus
genera Bunyaviridae includes enveloped viruses with a frag-mented single-stranded RNA genome of negative polarityTheir tripartite genome consists of a small (S) a medium(M) and a large (L) fragment and the envelope glycoproteinsGn and Gc that are cleaved out of a polyprotein synthesizedby the M fragment These glycoproteins have been provedto mediate the formation of the virus particle to play a rolein the interaction with cell surface receptors to mediate theentry of the virus into cells and to serve as targets for themajority of neutralizing Abs described so far
Orthobunyavirus transmitted through mosquitoes ormidges vectors are divided in 18 serogroups based on cross-titrations in haemagglutination inhibition assays and neutral-ization assays and correlating with main vector preferencesHowever the most clinically important viruses belong onlyto two serogroups the California encephalitis and the Simbuserogroups
Crimean-Congo hemorrhagic fever virus (CCHFV) isconsidered the only one of clinical relevance within theNairovirus genus which uses tick as main vector and is divid-ed in seven serogroups while Toscana virus and Rift Valleyfever virus (RVFV) which are transmitted by sandflies andmosquitoes respectively belong to the Phlebovirus genus[138]
MAbs against theNairovirusCCHFV and the PhlebovirusRVFV have been described
41 Crimean-Congo Hemorrhagic Fever Virus (CCHFV)Crimean-Congo Hemorrhagic Fever Virus (CCHFV) dis-tribution covers the greatest geographic range of any tick-borne virus known as viral isolation andor disease hasbeen reported from more than 30 countries in SoutheasternEurope Africa Asia and Middle East [139 140] Namelyfrom 1953 to 2010 about 6000 human cases were reportedin Southeastern Europe CCHFV causes sporadic outbreakswith mortality rates ranging from 10 to 80 [141 142] Asignificant variation in the time of incubation has beendescribed [139] Prehemorrhagic symptoms include highfever chills headache photophobia and back and abdominalpains Among other symptoms neuropsychiatric changeshave been reported in some patients In severe cases 3ndash6 daysafter the onset of the disease hemorrhagic symptoms occurwith petechiae ecchymosis bleeding in the form of melenahematemesis epistaxis [139]
In comparisonwith the nucleocapsid proteins inCCHFVGn and Gc show a higher degree of antigenic variabilityprobably due to their exposition to the host immune systemhypothesis corroborated in the description by Hewson et alin 2004 of four M segment phylogenic groups namely M1M2 M3 and M4 [141 142]
411 mAbs against CCHFV Blackburn et al firstly describedin 1987 murine anti-CCHFV mAbs able to recognize nucle-ocapsid proteins but no neutralizing activity has beendescribed thus far [143]
On the other hand Bertolotti-Ciarlet et al produced apanel of murine mAbs recognizing conformational epitopes
16 BioMed Research International
within theGn andGc glycoproteins expressed by the CCHFVstrain IbAr10200 The effectiveness of the mAbs was studiedby performing PRNT80 and none of the anti-Gn mAbsshowed neutralizing activity although many of the anti-GcmAbs neutralized IbAr10200 strain in vitro with clones 8A15E3 12A9 6C2 and 9H3 showing activity at a gt5120 dilutionMAbs 11E7 and 30F7 exhibited neutralizing activity at 2560dilution A suckling mice protection test was thereforeperformed for these mAbs The previously mentionedanti-Gc Abs were capable to protect mice to an appreciabledegree when applied 24 hours before and in a weakermanner 24 hours after virus challenge Many of the anti-GnmAbs were able to confer significant protection to IbAr10200both 24 hours before and after virus administration Therelevant effectiveness of the anti-Gn mAbs 6B12 10E1113G8 and 10G4 suggests that these mAbs may possess someneutralizing activity due to Ab-based effector mechanisms(eg ADCC) [58]
In further studies the same group characterized thebroadly cross-reactive neutralizing activity of the previouslydescribed murine mAb 11E7 able to bind conformationalepitopes within the C-terminal region of the Gn glycoproteinof all CCHFVM groups [58 141]
42 Rift Valley Fever Virus (RVFV) Similarly to CCHFV RiftValley Fever Virus (RVFV) causes sporadic outbreaks Thelargest epidemic occurred in Egypt in 1977 with an estimated200000 infections 18000 patients manifesting symptomsand 600 fatal prognoses [144] Symptoms include hemor-rhagic episodes fever encephalitis and blindness [145]
The major target of the humoral immune response is theRVFV glycoproteins Gc (G1) and Gn (G2) ranging fromnucleotides 480ndash2090 and 2091ndash3614 respectively [144]
421 mAbs against RVFV Within G2 glycoprotein are des-cribed three neutralizing antigenic sites namely epitope Inucleotides 792ndash893 (amino acid residues 258ndash291) epitopeII nucleotides 1164ndash1196 (amino acid residues 382ndash392) andepitope IV nucleotides 858ndash917 (amino acid residues 280ndash299) [59 145]
Despite the high degree of variability observed inCCHFVglycoproteins RVFV G2 presented an unexpected high con-servation among 22 isolates [59] In the same study onlythe LUNYO and 900060 isolates were resistant to in vitroneutralization by murine mAb 4D4 (recognizing epitope II)All other isolates were neutralized with PRNT80 titers from10240 togt81920On the other hand only SNS isolate showedreduced sensitivity to 4ndash39-CC (epitope IV) All other isolateswere neutralized with PRNT80 titers from 20480 to gt81920Protection studies on mice would be of interest for thedevelopment of a passive immunization prophylaxis [59]
Protection studies from lethal South African RVFVAN1830 strain infection were successfully performed on abattery of anti-G1 and anti-G2 mAbs produced by Besselaarand Blackburn with the strongly neutralizing anti-G1 mAb3E5 and anti-G2mAb 9C4 being the most effective [146]The9C4 antigenic area maps within G2 epitope I and a broadlyneutralizing activity may be investigated
5 Conclusions
Arbovirus infections have acquired increasing interest giventhe augmented globalization and tourism movement all overthe world that could be at the basis of epidemic eventsFurthermore the same vectors can sometimes transmit sev-eral arboviruses concomitantly complicating the diagnosis aswell as the therapy For example confusing mixed epidemicshave occasionally been described such as YFV plus CHIKVDENVplus CHIKV ormore recentlyPlasmodium falciparummalaria plus DENV1 and CHIKV on Madagascarrsquos east coast[1]
In the last decade several efforts have been employed inthe development of effective immunogens and therapeuticsgiving the current absence of specific antiviral drugs as wellas effective vaccines for themost diffused arbovirus infections(ie DENV andWNV)
However at the moment mosquito control is the bestavailable method for preventing arbovirus infections Thusto control the emerging public health problem of arbovirusinfections new antiviral therapeutic strategies that providepotent and broadly cross-protective immunity (especially forflavivirus infections) are an urgent globally medical need
As described in this review the majority of highlyneutralizing mAbs are of murine origin and indeed theirutility in the treatment of these infections is limited In facttheir employment can elicit an immunogenic response (iehuman anti-mouse Abs HAMA) the therapeutic efficacycould be reduced by a relatively faster clearance in humans(compared to human Abs) potentially exacerbated by theHAMA response Finally murine Abs exhibit relatively weakeffector functions (eg ADCC) compared to human AbsIndeed chimerization humanization or better the isolationand characterization of fully human broadly neutralizingmAbs would be the best choice for their possible use in afuture mAb-based therapy against arbovirus infections [147]
Finally possible ADE of infection mechanisms should beevaluated before considering a mAb as a possible candidatetherapeutic in a post-exposure setting as well as in the devel-opment of new Ab-eliciting immunogens
Conflict of Interests
The authors declare no conflict of interests in particular withOmriGam and any other cited Ab-based preparation men-tioned in the paper
References
[1] N Cleton M Koopmans J Reimerink G J Godeke and CReusken ldquoCome fly with me review of clinically importantarboviruses for global travelersrdquo Journal of Clinical Virology vol55 no 3 pp 191ndash203 2012
[2] F X Heinz H Holzmann A Essl and M Kundi ldquoField effec-tiveness of vaccination against tick-borne encephalitisrdquoVaccinevol 25 no 43 pp 7559ndash7567 2007
[3] A D Barrett and D E Teuwen ldquoYellow fever vaccinemdashhowdoes it work and why do rare cases of serious adverse events
BioMed Research International 17
take placerdquo Current Opinion in Immunology vol 21 no 3 pp308ndash313 2009
[4] S B Halstead and S J Thomas ldquoNew vaccines for Japaneseencephalitisrdquo Current Infectious Disease Reports vol 12 no 3pp 174ndash180 2010
[5] R Burioni M Perotti N Mancini and M Clementi ldquoPer-spectives for the utilization of neutralizing human monoclonalantibodies as anti-HCV drugsrdquo Journal of Hepatology vol 49no 2 pp 299ndash300 2008
[6] R A Diotti G A Sautto L Solforosi NManciniM Clementiand R Burioni ldquoNeutralization activity and kinetics of twobroad-range human monoclonal IgG1 derived from recombi-nant Fab fragments and directed against Hepatitis C virus E2glycoproteinrdquo New Microbiologica vol 35 no 4 pp 475ndash4792012
[7] R Burioni F Canducci N Mancini et al ldquoMolecular cloningof the first humanmonoclona antibodies neutralizing with highpotency Swine-origin Influenza A pandemic virus (S-OIV)rdquoNew Microbiologica vol 32 no 4 pp 319ndash324 2009
[8] P K Chandra S Hazari B Poat et al ldquoIntracytoplasmic stableexpression of IgG1 antibody targeting NS3 helicase inhibitsreplication of highly efficient hepatitis C Virus 2a clonerdquoVirology Journal vol 7 article 118 2010
[9] N Clementi D de Marco N Mancini et al ldquoA human mono-clonal antibody with neutralizing activity against highly diver-gent influenza subtypesrdquo PLoS ONE vol 6 no 12 Article IDe28001 2011
[10] D de Marco N Clementi N Mancini et al ldquoA non-VH1-69 heterosubtypic neutralizing human monoclonal antibodyprotects mice against H1N1 and H5N1 virusesrdquo PLoS ONE vol7 no 4 Article ID e34415 2012
[11] L Solforosi N Mancini F Canducci et al ldquoA phage displayvector optimized for the generation of human antibody com-binatorial libraries and the molecular cloning of monoclonalantibody fragmentsrdquoNewMicrobiologica vol 35 no 3 pp 289ndash294 2012
[12] E V Koonin and V V Dolja ldquoEvolution and taxonomy ofpositive-strand RNA viruses implications of comparative anal-ysis of amino acid sequencesrdquo Critical Reviews in Biochemistryand Molecular Biology vol 28 no 5 pp 375ndash430 1993
[13] J S Mackenzie D J Gubler and L R Petersen ldquoEmergingflaviviruses the spread and resurgence of Japanese encephalitisWest Nile and dengue virusesrdquo Nature Medicine vol 10 no 12pp S98ndashS109 2004
[14] T J SmithW E Brandt J L Swanson J MMcCown and E LBuescher ldquoPhysical and biological properties of dengue-2 virusand associated antigensrdquo Journal of Virology vol 5 no 4 pp524ndash532 1970
[15] J M Smit B Moesker I Rodenhuis-Zybert and J WilschutldquoFlavivirus cell entry and membrane fusionrdquo Viruses vol 3 no2 pp 160ndash171 2011
[16] R J Kuhn W Zhang M G Rossmann et al ldquoStructure ofdengue virus implications for flavivirus organization matura-tion and fusionrdquo Cell vol 108 no 5 pp 717ndash725 2002
[17] K Stiasny J H Aberle M Keller B Grubeck-Loebenstein andF X Heinz ldquoAge affects quantity but not quality of antibodyresponses after vaccination with an inactivated flavivirus vac-cine against tick-borne encephalitisrdquo PLoS ONE vol 7 no 3Article ID e34145 2012
[18] LThomasOVerlaetenACabie et al ldquoInfluence of the dengueserotype previous dengue infection and plasma viral load
on clinical presentation and outcome during a dengue-2 anddengue-4 co-epidemicrdquo American Journal of Tropical Medicineand Hygiene vol 78 no 6 pp 990ndash998 2008
[19] Y Zhang J Corver P R Chipman et al ldquoStructures of imma-ture flavivirus particlesrdquo EMBO Journal vol 22 no 11 pp2604ndash2613 2003
[20] A K I Falconar ldquoIdentification of an epitope on the denguevirus membrane (M) protein defined by cross-protective mon-oclonal antibodies design of an improved epitope sequencebased on common determinants present in both envelope (EandM) proteinsrdquo Archives of Virology vol 144 no 12 pp 2313ndash2330 1999
[21] K-J Huang Y-C Yang Y-S Lin et al ldquoThe dual-specific bind-ing of dengue virus and target cells for the antibody-dependentenhancement of dengue virus infectionrdquo Journal of Immunol-ogy vol 176 no 5 pp 2825ndash2832 2006
[22] A K I Falconar ldquoAntibody responses are generated to immun-odominant ELKKLE-typemotifs on the nonstructural-1 glyco-protein during live dengue virus infections inmice and humansimplications for diagnosis pathogenesis and vaccine designrdquoClinical and Vaccine Immunology vol 14 no 5 pp 493ndash5042007
[23] C W Mandl F Guirakhoo H Holzmann F X Heinz and CKunz ldquoAntigenic structure of the flavivirus envelope protein Eat the molecular level using the tick-borne encephalitis virus asa modelrdquo Journal of Virology vol 63 no 2 pp 564ndash571 1989
[24] F A Rey F X Heinz C Mandl C Kunz and S C HarrisonldquoThe envelope glycoprotein from tick-borne encephalitis virusat 2 A resolutionrdquo Nature vol 375 no 6529 pp 291ndash298 1995
[25] S L Allison K Stiasny K Stadler CWMandl and F XHeinzldquoMapping of functional elements in the stem-anchor regionof tick-borne encephalitis virus envelope protein Erdquo Journal ofVirology vol 73 no 7 pp 5605ndash5612 1999
[26] D E Klein J L Choi and S C Harrison ldquoStructure of a denguevirus envelope protein late-stage fusion intermediaterdquo Journal ofVirology vol 87 no 4 pp 2287ndash2293 2013
[27] S L Hanna T C Pierson M D Sanchez A A Ahmed M MMurtadha and R W Doms ldquoN-linked glycosylation of WestNile virus envelope proteins influences particle assembly andinfectivityrdquo Journal of Virology vol 79 no 21 pp 13262ndash132742005
[28] N Mancini G Sautto N Clementi et al ldquoNeutralization inter-fering antibodies a ldquonovelrdquo example of humoral immune dys-function facilitating viral escaperdquoViruses vol 4 no 9 pp 1731ndash1752 2012
[29] Y Modis S Ogata D Clements and S C Harrison ldquoVariablesurface epitopes in the crystal structure of dengue virus type3 envelope glycoproteinrdquo Journal of Virology vol 79 no 2 pp1223ndash1231 2005
[30] N Miller ldquoRecent progress in dengue vaccine research anddevelopmentrdquo Current Opinion in Molecular Therapeutics vol12 no 1 pp 31ndash38 2010
[31] J H Chavez J R Silva A A Amarilla and L T Moraes Figu-eiredo ldquoDomain III peptides from flavivirus envelope proteinare useful antigens for serologic diagnosis and targets forimmunizationrdquo Biologicals vol 38 no 6 pp 613ndash618 2010
[32] B Shrestha J D Brien S Sukupolvi-Petty et al ldquoThe develop-ment of therapeutic antibodies that neutralize homologous andheterologous genotypes of dengue virus type 1rdquo PLoS Pathogensvol 6 no 4 Article ID e1000823 2010
18 BioMed Research International
[33] G D Gromowski N D Barrett and A D T Barrett ldquoChar-acterization of dengue virus complex-specific neutralizing epi-topes on envelope protein domain III of dengue 2 virusrdquo Journalof Virology vol 82 no 17 pp 8828ndash8837 2008
[34] S Sukupolvi-Petty S K Austin W E Purtha et al ldquoType- andsubcomplex-specific neutralizing antibodies against domain IIIof dengue virus type 2 envelope protein recognize adjacentepitopesrdquo Journal of Virology vol 81 no 23 pp 12816ndash128262007
[35] S Sukupolvi-Petty S K Austin M Engle et al ldquoStructure andfunction analysis of therapeutic monoclonal antibodies againstdengue virus type 2rdquo Journal of Virology vol 84 no 18 pp9227ndash9239 2010
[36] Y-Q Deng J-X Dai G-H Ji et al ldquoA broadly flaviviruscross-neutralizingmonoclonal antibody that recognizes a novelepitope within the fusion loop of e proteinrdquo PLoS ONE vol 6no 1 Article ID e16059 2011
[37] J M Costin E Zaitseva K M Kahle et al ldquoMechanistic studyof broadly neutralizing human monoclonal antibodies againstdengue virus that target the fusion looprdquo Journal of Virologyvol 87 no 1 pp 52ndash66 2013
[38] CMMidgley A FlanaganH B Tran et al ldquoStructural analysisof a dengue cross-reactive antibody complexed with envelopedomain III reveals the molecular basis of cross-reactivityrdquoJournal of Immunology vol 188 no 10 pp 4971ndash4979 2012
[39] N J Moreland P Susanto E Lim et al ldquoPhage display appro-aches for the isolation of monoclonal antibodies against denguevirus envelope domain III from human and mouse derivedlibrariesrdquo International Journal ofMolecular Sciences vol 13 no3 pp 2618ndash2635 2012
[40] J J B Cockburn M E Navarro Sanchez N Fretes et alldquoMechanism of dengue virus broad cross-neutralization by amonoclonal antibodyrdquo Structure vol 20 no 2 pp 303ndash3142012
[41] R Rajamanonmani C Nkenfou P Clancy et al ldquoOn a mousemonoclonal antibody that neutralizes all four dengue virusserotypesrdquo Journal of General Virology vol 90 no 4 pp 799ndash809 2009
[42] L H Gould J Sui H Foellmer et al ldquoProtective and herapeuticcapacity of human single-chain Fv-Fc fusion proteins againstWest Nile virusrdquo Journal of Virology vol 79 no 23 pp 14606ndash14613 2005
[43] H Sultana H G Foellmer G Neelakanta et al ldquoFusion LoopPeptide of the West Nile virus envelope protein is essential forpathogenesis and is recognized by a therapeutic cross-reactivehuman monoclonal antibodyrdquo Journal of Immunology vol 183no 1 pp 650ndash660 2009
[44] T Oliphant M Engle G E Nybakken et al ldquoDevelopment ofa humanized monoclonal antibody with therapeutic potentialagainstWest Nile virusrdquoNature Medicine vol 11 no 5 pp 522ndash530 2005
[45] G E Nybakken T Oliphant S Johnson S Burke M S Dia-mond and D H Fremont ldquoStructural basis of West Nile virusneutralization by a therapeutic antibodyrdquo Nature vol 437 no7059 pp 764ndash769 2005
[46] G D Gromowski J T Roehrig M S Diamond J C LeeT J Pitcher and A D T Barrett ldquoMutations of an antibodybinding energy hot spot on domain III of the dengue 2 envelopeglycoprotein exploited for neutralization escaperdquo Virology vol407 no 2 pp 237ndash246 2010
[47] J T Roehrig R A Bolin andR G Kelly ldquoMonoclonal antibodymapping of the envelope glycoprotein of the dengue 2 virusJamaicardquo Virology vol 246 no 2 pp 317ndash328 1998
[48] R de Alwis S A Smith N P Olivarez et al ldquoIdentification ofhumanneutralizing antibodies that bind to complex epitopes ondengue virionsrdquo Proceedings of the National Academy of Sciencesof the United States of America vol 109 no 19 pp 7439ndash74442012
[49] S K Austin K A Dowd B Shrestha et al ldquoStructural basis ofdifferential neutralization of DENV-1 genotypes by an antibodythat recognizes a cryptic epitoperdquo PLoS Pathogens vol 8 no 10Article ID e1002930 2012
[50] M Throsby C Geuijen J Goudsmit et al ldquoIsolation and char-acterization of human monoclonal antibodies from individualsinfected with West Nile Virusrdquo Journal of Virology vol 80 no14 pp 6982ndash6992 2006
[51] A P Goncalvez R H Purcell and C-J Lai ldquoEpitope deter-minants of a chimpanzee Fab antibody that efficiently cross-neutralizes dengue type 1 and type 2 viruses map to inside andin close proximity to fusion loop of the dengue type 2 virusenvelope glycoproteinrdquo Journal of Virology vol 78 no 23 pp12919ndash12928 2004
[52] J T Roehrig J H Mathews and D W Trent ldquoIdentification ofepitopes on the E glycoprotein of Saint Louis encephalitis virususing monoclonal antibodiesrdquo Virology vol 128 no 1 pp 118ndash126 1983
[53] J T Roehrig L A Staudinger A R Hunt J H Mathews andC D Blair ldquoAntibody prophylaxis and therapy for flavivirusencephalitis infectionsrdquo Annals of the New York Academy ofSciences vol 951 pp 286ndash297 2001
[54] A P Goncalvez C-H Chien K Tubthong et al ldquoHumanizedmonoclonal antibodies derived from chimpanzee fabs protectagainst Japanese encephalitis virus in vitro and in vivordquo Journalof Virology vol 82 no 14 pp 7009ndash7021 2008
[55] M Arakawa T Yamashiro G-I Uechi M Tadano and ANishizono ldquoConstruction of humanFab (1205741120581) library and iden-tification of human monoclonal Fab possessing neutralizingpotency against Japanese encephalitis virusrdquo Microbiology andImmunology vol 51 no 6 pp 617ndash625 2007
[56] L Warter C Y Lee R Thiagarajan et al ldquoChikungunya virusenvelope-specific human monoclonal antibodies with broadneutralization potencyrdquo Journal of Immunology vol 186 no 5pp 3258ndash3264 2011
[57] P Pal K A Dowd J D Brien et al ldquoDevelopment of a highlyprotective combination monoclonal antibody therapy againstchikungunya virusrdquo PLoS Pathogens vol 9 no 4 Article IDe1003312 2013
[58] A Bertolotti-Ciarlet J Smith K Strecker et al ldquoCellular local-ization and antigenic characterization of Crimean-Congo hem-orrhagic fever virus glycoproteinsrdquo Journal of Virology vol 79no 10 pp 6152ndash6161 2005
[59] J K Battles and J M Dalrymple ldquoGenetic variation amonggeographic isolates of Rift Valley fever virusrdquo American Journalof Tropical Medicine and Hygiene vol 39 no 6 pp 617ndash6311988
[60] B AThibodeaux N C Garbino NM Liss et al ldquoA humanizedIgG but not IgM antibody is effective in prophylaxis and therapyof yellow fever infection in an AG12917D-204 peripheralchallenge mouse modelrdquo Antiviral Research vol 94 no 1 pp1ndash8 2012
BioMed Research International 19
[61] S Daffis R E Kontermann J Korimbocus H Zeller H-DKlenk and J Ter Meulen ldquoAntibody responses against wild-type yellow fever virus and the 17D vaccine strain charac-terization with human monoclonal antibody fragments andneutralization escape variantsrdquoVirology vol 337 no 2 pp 262ndash272 2005
[62] L N Levanov L E Matveev E P Goncharova et al ldquoChimericantibodies against tick-borne encephalitis virusrdquo Vaccine vol28 no 32 pp 5265ndash5271 2010
[63] A R Hunt S Frederickson C Hinkel K S Bowdish and J TRoehrig ldquoA humanized murine monoclonal antibody protectsmice either before or after challenge with virulent Venezuelanequine encephalomyelitis virusrdquo Journal of General Virologyvol 87 no 9 pp 2467ndash2476 2006
[64] J M da Silva Voorham I A Rodenhuis-Zybert N V A Nunezet al ldquoAntibodies against the envelope glycoprotein promoteinfectivity of immature dengue virus serotype 2rdquo PLoS ONEvol 7 no 3 Article ID e29957 2012
[65] T C Pierson Q Xu S Nelson et al ldquoThe stoichiometry ofantibody-mediated neutralization and enhancement of westnile virus infectionrdquoCell Host andMicrobe vol 1 no 2 pp 135ndash145 2007
[66] G Sautto N Mancini R A Diotti L Solforosi M Clementiand R Burioni ldquoAnti-hepatitis C virus E2 (HCVE2) glyco-proteinmonoclonal antibodies andneutralization interferencerdquoAntiviral Research vol 96 no 1 pp 82ndash89 2012
[67] R Burioni Y Matsuura N Mancini et al ldquoDiverging effectsof human recombinant anti-hepatitis C virus (HCV) antibodyfragments derived from a single patient on the infectivityof a vesicular stomatitis virusHCV pseudotyperdquo Journal ofVirology vol 76 no 22 pp 11775ndash11779 2002
[68] R J Phillpotts J R Stephenson and J S Porterfield ldquoAntibody-dependent enhancement of tick-borne encephalitus virus infec-tivityrdquo Journal of General Virology vol 66 no 8 pp 1831ndash18371985
[69] K A Dowd and T C Pierson ldquoAntibody-mediated neutraliza-tion of flaviviruses a reductionist viewrdquo Virology vol 411 no 2pp 306ndash315 2011
[70] A K I Falconar ldquoMonoclonal antibodies that bind to commonepitopes on the dengue virus type 2 nonstructural-1 andenvelope glycoproteins display weak neutralizing activity anddifferentiated responses to virulent strains implications forpathogenesis and vaccinesrdquo Clinical and Vaccine Immunologyvol 15 no 3 pp 549ndash561 2008
[71] K Meyer A Banerjee S E Frey R B Belshe and R Ray ldquoAweak neutralizing antibody response to hepatitis C virus enve-lope glycoprotein enhances virus infectionrdquo PLoS ONE vol 6no 8 Article ID e23699 2011
[72] R Burioni R AWilliamson P P Sanna F E Bloom and D RBurton ldquoRecombinant human Fab to glycoprotein D neutral-izes infectivity and prevents cell-to-cell transmission of herpessimplex viruses 1 and 2 in vitrordquo Proceedings of the NationalAcademy of Sciences of the United States of America vol 91 no1 pp 355ndash359 1994
[73] M S Rolph A Zaid N E Rulli and S Mahalingam ldquoDown-regulation of interferon-120573 in antibody-dependent enhancementof dengue viral infections of human macrophages is dependenton interleukin-6rdquo Journal of Infectious Diseases vol 204 no 3pp 489ndash491 2011
[74] S S Whitehead J E Blaney A P Durbin and B R MurphyldquoProspects for a dengue virus vaccinerdquoNature ReviewsMicrobi-ology vol 5 no 7 pp 518ndash528 2007
[75] S-M Lok V Kostyuchenko G ENybakken et al ldquoBinding of aneutralizing antibody to dengue virus alters the arrangement ofsurface glycoproteinsrdquoNature Structural andMolecular Biologyvol 15 no 3 pp 312ndash317 2008
[76] A P Goncalvez R Men C Wernly R H Purcell and C-J Lai ldquoChimpanzee Fab fragments and a derived humanizedimmunogiobulin G1 antibody that efficiently cross-neutralizedengue type 1 and type 2 virusesrdquo Journal of Virology vol 78no 23 pp 12910ndash12918 2004
[77] WM P BWahala A A Kraus L B HaymoreM A Accavitti-Loper and A M de Silva ldquoDengue virus neutralization byhuman immune sera role of envelope protein domain III-reactive antibodyrdquo Virology vol 392 no 1 pp 103ndash113 2009
[78] W D Crill H R Hughes M J Delorey and G-J J ChangldquoHumoral immune responses of dengue fever patients usingepitope-specific serotype-2 virus-like particle antigensrdquo PLoSONE vol 4 no 4 Article ID e4991 2009
[79] C Setthapramote T Sasaki O Puiprom et al ldquoHuman mono-clonal antibodies to neutralize all dengue virus serotypes usinglymphocytes from patients at acute phase of the secondaryinfectionrdquo Biochemical and Biophysical Research Communica-tions vol 423 no 4 pp 867ndash872 2012
[80] M Beltramello K L Williams C P Simmons et al ldquoThehuman immune response to dengue virus is dominated byhighly cross-reactive antibodies endowed with neutralizing andenhancing activityrdquo Cell Host andMicrobe vol 8 no 3 pp 271ndash283 2010
[81] C M Midgley M Bajwa-Joseph S Vasanawathana et al ldquoAnin-depth analysis of original antigenic sin in dengue virusinfectionrdquo Journal of Virology vol 85 no 22 pp 410ndash421 2011
[82] B Kaufmann M R Vogt J Goudsmit et al ldquoNeutralizationof West Nile virus by cross-linking of its surface proteins withFab fragments of the human monoclonal antibody CR4354rdquoProceedings of the National Academy of Sciences of the UnitedStates of America vol 107 no 44 pp 18950ndash18955 2010
[83] B Spurrier J M Sampson M Totrov et al ldquoStructural analysisof human and macaque mAbs 2909 and 25B implicationsfor the configuration of the quaternary neutralizing epitope ofHIV-1 gp120rdquo Structure vol 19 no 5 pp 691ndash699 2011
[84] J J SchlesingerMWBrandriss andT PMonath ldquoMonoclonalantibodies distinguish between wild and vaccine strains ofyellow fever virus by neutralization hemagglutination inhibi-tion and immune precipitation of the virus envelope proteinrdquoVirology vol 125 no 1 pp 8ndash17 1983
[85] M W Brandriss J J Schlesinger E E Walsh and M BrisellildquoLethal 17D yellow fever encephalitis in mice I Passive protec-tion by monoclonal antibodies to the envelope proteins of 17Dyellow fever and dengue 2 virusesrdquo Journal of General Virologyvol 67 no 2 pp 229ndash234 1986
[86] H Cho and M S Diamond ldquoImmune responses to West Nilevirus infection in the central nervous systemrdquo Viruses vol 4no 12 pp 3812ndash3830 2012
[87] D W Beasley A D Barrett and R B Tesh ldquoResurgence ofWest Nile neurologic disease in the United States in 2012 whathappened What needs to be donerdquo Antiviral Research vol 99no 1 pp 1ndash5 2013
[88] M Throsby J Ter Meulen C Geuijen J Goudsmit and J DeKruif ldquoMapping and analysis of West Nile virus-specific mon-oclonal antibodies prospects for vaccine developmentrdquo ExpertReview of Vaccines vol 6 no 2 pp 183ndash191 2007
[89] M S Diamond T G Roberts D Edgil B Lu J Ernst and EHarris ldquoModulation of dengue virus infection in human cells
20 BioMed Research International
by alpha beta and gamma interferonsrdquo Journal of Virology vol74 no 11 pp 4957ndash4966 2000
[90] Z Shimoni M J Niven S Pitlick and S Bulvik ldquoTreatmentof West Nile virus encephalitis with intravenous immunoglob-ulinrdquo Emerging Infectious Diseases vol 7 no 4 article 759 2001
[91] AHamdan PGreen EMendelsonM R Kramer S Pitlik andM Weinberger ldquoPossible benefit of intravenous immunoglob-ulin therapy in a lung transplant recipient with West Nile virusencephalitisrdquoTransplant InfectiousDisease vol 4 no 3 pp 160ndash162 2002
[92] M Haley A S Retter D Fowler J Gea-Banacloche and NP OrsquoGrady ldquoThe role for intravenous immunoglobulin in thetreatment of West Nile virus encephalitisrdquo Clinical InfectiousDiseases vol 37 no 6 pp e88ndashe90 2003
[93] K M Chung G E Nybakken B S Thompson et al ldquoAnti-bodies against West Nile virus nonstructural protein NS1prevent lethal infection through Fc 120574 receptor-dependent and-independent mechanismsrdquo Journal of Virology vol 80 no 3pp 1340ndash1351 2006
[94] E A Henchal L S Henchal and J J Schlesinger ldquoSynergisticinteractions of anti-NS1 monoclonal antibodies protect pas-sively immunized mice from lethal challenge with dengue 2virusrdquo Journal of General Virology vol 69 no 8 pp 2101ndash21071988
[95] J T Roehrig ldquoAntigenic structure of flavivirus proteinsrdquoAdvances in Virus Research vol 59 pp 141ndash175 2003
[96] M D Sanchez T C Pierson D McAllister et al ldquoCharacteri-zation of neutralizing antibodies to West Nile virusrdquo Virologyvol 336 no 1 pp 70ndash82 2005
[97] J H Beigel J L Nordstrom S R Pillemer et al ldquoSafety andpharmacokinetics of single intravenous dose of MGAWN1 anovel monoclonal antibody to West Nile virusrdquo AntimicrobialAgents and Chemotherapy vol 54 no 6 pp 2431ndash2436 2010
[98] C A Smeraski V Siddharthan and J D Morrey ldquoTreatmentof spatial memory impairment in hamsters infected with WestNile virus using a humanizedmonoclonal antibodyMGAWN1rdquoAntiviral Research vol 91 no 1 pp 43ndash49 2011
[99] D Lelli AMoreno E Brocchi et al ldquoWestNile virus character-ization and diagnostic applications of monoclonal antibodiesrdquoVirology Journal vol 9 article 81 2012
[100] T S Gritsun V A Lashkevich and E A Gould ldquoTick-borneencephalitisrdquo Antiviral Research vol 57 no 1-2 pp 129ndash1462003
[101] D Ruzek T S Gritsun N L Forrester et al ldquoMutations inthe NS2B and NS3 genes affect mouse neuroinvasiveness of aWestern European field strain of tick-borne encephalitis virusrdquoVirology vol 374 no 2 pp 249ndash255 2008
[102] M Ecker S L Allison T Meixner and F X Heinz ldquoSequenceanalysis and genetic classification of tick-borne encephalitisviruses from Europe and Asiardquo Journal of General Virology vol80 no 1 pp 179ndash185 1999
[103] H Holzmann M S Vorobyova I P Ladyzhenskaya et alldquoMolecular epidemiology of tick-borne encephalitis viruscross-protection between European and Far Eastern subtypesrdquoVaccine vol 10 no 5 pp 345ndash349 1992
[104] C W Mandl ldquoSteps of the tick-borne encephalitis virus repli-cation cycle that affect neuropathogenesisrdquo Virus Research vol111 no 2 pp 161ndash174 2005
[105] A V Atrasheuskaya T M Fredeking and G M IgnatyevldquoChanges in immune parameters and their correction in humancases of tick-borne encephalitisrdquo Clinical and ExperimentalImmunology vol 131 no 1 pp 148ndash154 2003
[106] K L Mansfield N Johnson L P Phipps J R Stephenson AR Fooks and T Solomon ldquoTick-borne encephalitis virusmdashareview of an emerging zoonosisrdquo Journal of General Virologyvol 90 no 8 pp 1781ndash1794 2009
[107] DWVaughn andCHHoke Jr ldquoThe epidemiology of Japaneseencephalitis prospects for preventionrdquo Epidemiologic Reviewsvol 14 pp 197ndash221 1992
[108] T Solomon H Ni D W C Beasley M Ekkelenkamp M JCardosa and A D T Barrett ldquoOrigin and evolution of Japaneseencephalitis virus in Southeast Asiardquo Journal of Virology vol 77no 5 pp 3091ndash3098 2003
[109] M R Wilson ldquoEmerging viral infectionsrdquo Current Opinion inNeurology vol 26 no 3 pp 301ndash306 2013
[110] J N Hanna S A Ritchie D A Phillips et al ldquoAn outbreakof Japanese encephalitis in the Torres Strait Australia 1995rdquoMedical Journal of Australia vol 165 no 5 pp 256ndash260 1996
[111] J N Hanna S A Ritchie D A Phillips et al ldquoJapanese ence-phalitis in north Queensland Australia 1998rdquo Medical Journalof Australia vol 170 no 11 pp 533ndash536 1999
[112] SGeorgeMGourie-Devi and J A Rao ldquoIsolation ofWestNilevirus from the brains of children who had died of encephalitisrdquoBulletin of theWorldHealthOrganization vol 62 no 6 pp 879ndash882 1984
[113] A K Gupta A A Koshy and V J Lad ldquoEnhanced protec-tion of mice against Japanese encephalitis virus infection bycombinations ofmonoclonal antibodies to glycoprotein ErdquoActaVirologica vol 55 no 2 pp 165ndash168 2011
[114] T H Lee B-H Song S-I Yun et al ldquoA cross-protective mAbrecognizes a novel epitope within the flavivirus NS1 proteinrdquoJournal of General Virology vol 93 no 1 pp 20ndash26 2012
[115] S C Weaver ldquoHost range amplification and arboviral diseaseemergencerdquo Archives of Virology Supplementum no 19 pp 33ndash44 2005
[116] M Solignat B Gay S Higgs L Briant and C Devaux ldquoRepli-cation cycle of chikungunya a re-emerging arbovirusrdquoVirologyvol 393 no 2 pp 183ndash197 2009
[117] O Schwartz and M L Albert ldquoBiology and pathogenesis ofchikungunya virusrdquo Nature Reviews Microbiology vol 8 no 7pp 491ndash500 2010
[118] W Akahata Z-Y Yang H Andersen et al ldquoA virus-like particlevaccine for epidemic Chikungunya virus protects nonhumanprimates against infectionrdquo Nature Medicine vol 16 no 3 pp334ndash338 2010
[119] M Kielian and A Helenius ldquopH-Iinduced alterations in thefusogenic spike protein of Semliki Forest virusrdquo Journal of CellBiology vol 101 no 6 pp 2284ndash2291 1985
[120] J White and A Helenius ldquopH-Dependent fusion between theSemliki Forest virus membrane and liposomesrdquo Proceedings ofthe National Academy of Sciences of the United States of Americavol 77 no 6 pp 3273ndash3277 1980
[121] J Dubuisson and C M Rice ldquoSindbis virus attachment isola-tion and characterization of mutants with impaired binding tovertebrate cellsrdquo Journal of Virology vol 67 no 6 pp 3363ndash33741993
[122] C Y Lee Y-W Kam J Fric et al ldquoChikungunya virus neutral-ization antigens and direct cell-to-cell transmission are revealedby human antibody-escapemutantsrdquo PLoS Pathogens vol 7 no12 Article ID e1002390 2011
[123] J E Voss M-C Vaney S Duquerroy et al ldquoGlycoproteinorganization of Chikungunya virus particles revealed by X-raycrystallographyrdquo Nature vol 468 no 7324 pp 709ndash712 2010
BioMed Research International 21
[124] L Li J Jose Y Xiang R J Kuhn and M G Rossmann ldquoStruc-tural changes of envelope proteins during alphavirus fusionrdquoNature vol 468 no 7324 pp 705ndash708 2010
[125] J Fric S Bertin-Maghit C I Wang A Nardin and L WarterldquoUse of human monoclonal antibodies to treat Chikungunyavirus infectionrdquo Journal of Infectious Diseases vol 207 no 2 pp319ndash322 2013
[126] P V Aguilar J G Estrada-Franco R Navarro-Lopez C FerroA D Haddow and S C Weaver ldquoEndemic Venezuelanequine encephalitis in the Americas hidden under the dengueumbrellardquo Future Virology vol 6 no 6 pp 721ndash740 2011
[127] S C Weaver R Winegar I D Manger and N L ForresterldquoAlphaviruses population genetics and determinants of emer-gencerdquo Antiviral Research vol 94 no 3 pp 242ndash257 2012
[128] S Paessler and S C Weaver ldquoVaccines for Venezuelan equineencephalitisrdquo Vaccine vol 27 no 4 pp D80ndashD85 2009
[129] R J Phillpotts ldquoVenezuelan equine encephalitis virus complex-specific monoclonal antibody provides broad protection inmurine models against airborne challenge with viruses fromserogroups I II and IIIrdquoVirus Research vol 120 no 1-2 pp 107ndash112 2006
[130] R J Phillpotts L D Jones and S C Howard ldquoMonoclonalantibody protectsmice against infection anddiseasewhen giveneither before or up to 24 h after airborne challenge with virulentVenezuelan equine encephalitis virusrdquoVaccine vol 20 no 11-12pp 1497ndash1504 2002
[131] J H Mathews J T Roehrig and D W Trent ldquoRole of com-plement and the Fc portion of immunoglobulin G in immunityto Venezuelan equine encephalomyelitis virus infection withglycoprotein-specific monoclonal antibodiesrdquo Journal of Virol-ogy vol 55 no 3 pp 594ndash600 1985
[132] L M OrsquoBrien C D Underwood-Fowler S A Goodchild AL Phelps and R J Phillpotts ldquoDevelopment of a novel mon-oclonal antibody with reactivity to a wide range of Venezuelanequine encephalitis virus strainsrdquoVirology Journal vol 6 article206 2009
[133] L M OrsquoBrien S A Goodchild R J Phillpotts and S D Per-kins ldquoA humanised murine monoclonal antibody protectsmice from Venezuelan equine encephalitis virus Evergladesvirus and Mucambo virus when administered up to 48h afterairborne challengerdquo Virology vol 426 no 2 pp 100ndash105 2012
[134] N Clementi N Mancini L Solforosi M Castelli M Clementiand R Burioni ldquoPhage display-based strategies for cloning andoptimization of monoclonal antibodies directed against humanpathogensrdquo International Journal of Molecular Sciences vol 13no 7 pp 8273ndash8292 2012
[135] A R Hunt R A Bowen S Frederickson T Maruyama JT Roehrig and C D Blair ldquoTreatment of mice with humanmonoclonal antibody 24h after lethal aerosol challenge withvirulent Venezuelan equine encephalitis virus prevents diseasebut not infectionrdquo Virology vol 414 no 2 pp 146ndash152 2011
[136] W-G Hu A L Phelps S Jager et al ldquoA recombinant human-ized monoclonal antibody completely protects mice againstlethal challenge with Venezuelan equine encephalitis virusrdquoVaccine vol 28 no 34 pp 5558ndash5564 2010
[137] T Rulker L Voss P Thullier et al ldquoIsolation and characterisa-tion of a human-like antibody fragment (scFv) that inactivatesVEEV in vitro and in vivordquo PLoS ONE vol 7 no 5 Article IDe37242 2012
[138] C T Walter and J N Barr ldquoRecent advances in the molecularand cellular biology of bunyavirusesrdquo Journal of General Virol-ogy vol 92 no 11 pp 2467ndash2484 2011
[139] C A Whitehouse ldquoCrimean-Congo hemorrhagic feverrdquo Anti-viral Research vol 64 no 3 pp 145ndash160 2004
[140] M Mertens K Schmidt A Ozkul and M H Groschup ldquoTheimpact of Crimean-Congo hemorrhagic fever virus on publichealthrdquo Antiviral Research vol 98 no 2 pp 248ndash260 2013
[141] A A Ahmed J M McFalls C Hoffmann et al ldquoPresence ofbroadly reactive and group-specific neutralizing epitopes onnewly described isolates of Crimean-Congo hemorrhagic fevervirusrdquo Journal of General Virology vol 86 no 12 pp 3327ndash33362005
[142] R Hewson A Gmyl L Gmyl et al ldquoEvidence of segment reas-sortment in Crimean-Congo haemorrhagic fever virusrdquo Journalof General Virology vol 85 no 10 pp 3059ndash3070 2004
[143] N K Blackburn T G Besselaar A J Shepherd and R Swane-poel ldquoPreparation and use of monoclonal antibodies for iden-tifying Crimean-Congo hemorrhagic fever virusrdquo AmericanJournal of Tropical Medicine and Hygiene vol 37 no 2 pp 392ndash397 1987
[144] T Ikegami ldquoMolecular biology and genetic diversity of RiftValley fever virusrdquoAntiviral Research vol 95 no 3 pp 293ndash3102012
[145] KKeegan andM S Collett ldquoUse of bacterial expression cloningto define the amino acid sequences of antigenic determinantson the G2 glycoprotein of Rift Valley fever virusrdquo Journal ofVirology vol 58 no 2 pp 263ndash270 1986
[146] T G Besselaar and N K Blackburn ldquoTopological mapping ofantigenic sites on the Rift Valley fever virus envelope glycopro-teins using monoclonal antibodiesrdquo Archives of Virology vol121 no 1ndash4 pp 111ndash124 1991
[147] L G Presta ldquoEngineering of therapeutic antibodies to mini-mize immunogenicity and optimize functionrdquo Advanced DrugDelivery Reviews vol 58 no 5-6 pp 640ndash656 2006
Submit your manuscripts athttpwwwhindawicom
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anatomy Research International
PeptidesInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporation httpwwwhindawicom
International Journal of
Volume 2014
Zoology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Molecular Biology International
GenomicsInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioinformaticsAdvances in
Marine BiologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Signal TransductionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Evolutionary BiologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Biochemistry Research International
ArchaeaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Genetics Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in
Virolog y
Hindawi Publishing Corporationhttpwwwhindawicom
Nucleic AcidsJournal of
Volume 2014
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Enzyme Research
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Microbiology

BioMed Research International 11
these mAbs two (CR4374 and CR4353) protected mice fromlethal WNV challenge at 50 protective doses of 129 and357 120583gkg of body weight respectively [50]
Sanchez et al using three different immunization strate-gies (ie inactivated virus naked DNA and recombinantprotein) isolated ninemurinemAbsmost of which bound toconformation-dependent epitopes in DIII of the E protein Inparticular neutralizing mAbs named 8B10 11C2 10C5 and17C8 were obtained from mice immunized with inactivatedvirus alone or in combination with a DNA plasmid andbound to the same region of DIII with high affinity Incontrast mAbs obtained by immunization with a solubleversion of the E glycoprotein did not exhibit neutralizingactivity These non-neutralizing mAbs were cross-reactivewith several other flaviviruses including SLEV JEV YFVand Powassan virus confirming the conserved nature ofFlavivirus non-neutralizing epitopes [96]
Gould et al isolated 11 unique human single-chain vari-able region Ab fragments (scFvs) that bind the E protein ofWNVAmong them a humanmAb namedmAb11 expressedas a scFv-Fc fusion protein was further characterized Itrecognizes the fusion loop at the distal end of DII ofthe WNV E protein and cross-reacts with all four DENVserotypes and provides protection against DENV2 and 4as well as WNV [42] Moreover therapeutic studies of thismAb in WNV-infection model mice provided substantialprotection when administered after 5 days post-infectionInterestingly a neutralization escape variant of this mAbfailed to cause lethal encephalitis (at higher infectious doses)or induce the inflammatory responses associated with blood-brain barrier permeability in mice compared to the parentalWNV suggesting an important role for the fusion loop inviral pathogenesis [43]
Oliphant et al isolated an anti-DIII E protein mAbnamed E16 fromhybridomas obtained after immunization ofmice with recombinant WNV E protein which neutralizedall WNV strains with PRNT50 values of 4 to 18 ng andPRNT90 values of 53 to 297 ng One hundred microgramsof mAb protected greater than 90 of mice from lethalinfection and even a single 4120583g treatment of E16 on day 2 afterinfection prevented mortality Moreover humanization ofthis mAb confirmed as therapeutically effective in mice [44]Subsequent studies revealed that mAb E16 neutralization ismediated by engagement of four discontinuous segmentsof DIII including the amino-terminal region (amino acidresidues 302ndash309) and the three connecting loops BC (aminoacid residues 330ndash333) DE (amino acid residues 365ndash368)and FG (amino acid residues 389ndash391) Moreover no ADE ofinfection was detected when E16 mAb was used at saturatingconcentrations [45 47]
Furthermore results of a Phase I safety study of thehumanized E16 mAb (designated MGAWN1) have beenreported and suggested that doses of up to 30mgkgwerewelltolerated with few mild adverse events and would providean excess of virus neutralizing activity for 3-4 weeks aftertreatment However a case of anti-MGAWN1 Ab elicitationoccurred with the consequent increased rate of clearance andindeed impacting efficacy A Phase II safety and efficacy studyof MGAWN1 is ongoing [97] Furthermore the possibility of
preventing or treating WNV-induced memory deficits wasrecently investigated In this study hamsters were treatedintraperitoneally with 32mgkg ofMGAWN1mAb at 45 daysafter subcutaneously challenging with WNV InterestinglyMGAWN1 prevented mortality weight loss and improvedfood consumption of WNV-infected hamsters compared tocontrols [98]
Recently Lelli et al isolated six anti-E mAbs frominactivated-WNV immunized mice In particular three ofthem (3B2 3D6 and 4D3) neutralized lineage I and II WNVwith the first two recognizing the same epitopes located onthe distal lateral surface of DIII (critical amino acid residueK307) Conversely 4D3mAb recognized a novel neutralizingepitope on DII (critical residues S276 and T278) Indeedfurther protective and therapeutic studies are needed toascertain their neutralizing activity in vivo [99]
Finally to conclude like DENV infection Abs directedagainst the M protein are not protective and neutralizingMoreover in humans a skewed humoral immune responseagainst DII has been frequently observed and confirmedalso with the hybridoma technology [50] The isolation andelicitation of neutralizingAbs directed against the fusion loopand DIII of the E protein represent thus the most challengingand promising goal for the development of new effectivetherapeutic inhibitors and immunogens respectively
27 Tick-Borne Encephalitis Virus (TBEV) Tick-borne ence-phalitis virus (TBEV) is one of the most dangerous agentscausing human neuroinfections occurring mostly in Europeand Asia and with a potential fatal prognosis [100] TBEV isbelieved to cause 3500ndash10000 human cases of encephalitisin Europe per year with a high morbidity in Russia CzechRepublic Austria and Germany In particular between 1990and 2007 an average of 8755 cases of TBE was reported peryear in Europe and Russia Despite the fact that Russia isthe country with most infections registered annually CzechRepublic incidence is among the highest in Europe with 400ndash1000 clinical cases reported every year [101]
Three subtypes of TBEV are classified namely EuropeanSiberian and Far-Eastern sharing most of the genetic andantigenic features [102] In fact a high degree of antigenichomogeneity between different strains of TBEV has beendescribed [103]
Clinical manifestations vary among the subtypes butusually they start with a short febrile period of 7ndash14 daysafter the tick bite Fatigue headache and pain in the neckshoulders and lower back together with high fever andvomitingmay be present [100]Thesemanifestations are oftenfollowed by an asymptomatic phase lasting from 2 to 10days after remission from the fever with possible progres-sion to neurological disease Neurological symptoms includemeningitis encephalitis myelitis and radiculitis Mortalityoccurs in 1-2 of the European subtype-infected patientsbut fatal prognosis can occur in up to 20ndash40 of the Far-Eastern subtype-infected patients [104] Mortality rates ofthe Siberian subtype are similar to those observed for theEuropean [105]
Effective and safe vaccines against TBEV produced frominactivated virus have been developed and licensed for their
12 BioMed Research International
use in humans However an emergency therapy in theabsence of a mass immunization is needed as no effectivetreatments are yet available [106]
271 mAbs against TBEV Levanov et al described the chim-erization of two murine mAbs (13D6 and 10C2) directedagainst the DIII and DII respectively of the TBEV E glyco-protein The chimeric mAbs present binding characteristicssimilar to the parentalmAbsMoreover as the parentalmAbsonly the chimeric mAb 13D6 was able to neutralize TBEVinfectivity in vitro In particular neutralization studies withthe murine chimeric and scFv forms of mAb 13D6 were per-formed In particular murine 13D6 showed an IC50 titer of115 120583gmL in Focus Reduction Neutralization Tests (FRNT)against TBEV strain 205 and of 29 120583gmL in PRNT againstTBEV strain Softjin Chimeric 13D6 showed an IC50 titer of45 120583gmL in FRNT against TBEV strain 205 and of 19 120583gmLin PRNT against TBEV strain Softjin ScFv 13D6 showed anIC50 titer of 167 120583gmL in FRNT against TBEV strain 205and of 112 120583gmL in PRNT against TBEV strain Softjin [62]
28 Japanese Encephalitis Virus (JEV) Japanese encephalitisvirus (JEV) is a mosquito-borne flavivirus that over thepast few decades has caused several outbreaks throughoutChina Southeast Asia Australia and Papua New Guineawith a prevalence recently estimated to be of about 70000casesyear [107ndash109] A 40 mortality was recorded in someof the JEV-affected areas Moreover many survivors facesome neurological problems and complications [108] Since1995 the disease has also emerged in Non-Asian regions suchas Northern Australia [110 111] The situation in SoutheastAsia however is further complicated by the overlappingepidemics of JEV and DENV as well as sporadic casesof WNV infections detected in some of the affected areasparticularly in India [112]
Effective vaccines both live-attenuated and inactivatedstrains of JEV have been developed and licensed in majorcountries However as for the other arthropod-borne Fla-vivirus members no specific antiviral drugs are currentlyavailable [4]
281mAbs against JEV Aspreviously described for the otherFlavivirusmembers many groups isolated and characterizedanti-JEVmAbs showing different neutralizing and protectiveproperties In this regard Gupta et al used combinations ofanti-E JEV mAbs (Hs1-4) in mice protection experimentsIn particular they found that the singularly mAb protectionranged from 45 to 65 when 100120583g of mAb were adminis-tered while equimolar combinations of two or three mAbsgave 85ndash90 or 100 protection respectively [113] In similarexperiments Lee et al demonstrated that a Flavivirus anti-NS1 mAb named 16NS1 cross-reacted with JEV as well asWNV and exhibited protective activity against WNV as wellas a lethal JEV infection However no neutralizing activitywas observed using this mAb against both WNV and JEV inin vitro experiments suggesting the participation of otherAb-mediatedmechanisms in vivo In particular 95ofmicewere
protected when 500120583g of mAb were administered intraperi-toneally and concomitantly to intramuscular injection of JEV
Overlapping peptide mapping analysis combined withsite-specific mutations identified the 116KAWGKSILFA125region and critical amino acid residues W118 and I122 as16NS1 mAb epitope highly conserved in WNV and JEVstrains [114]
Arakawa et al isolated from a combinatorial human Fablibrary constructed from peripheral blood lymphocytesobtained from JEV hyperimmune volunteers Among 188randomly selected clones FabTJE12B02 showed the best 50focus reduction endpoint at the concentration of 502 120583gmLagainst the JEV strain Nakayama [55]
Goncalvez et al isolated three mAbs named Fabs A3B2 and E3 by repertoire cloning from chimpanzees initiallyimmunized with inactivated JE-VAX and then boosted withattenuated JEV SA14-14-2 In particular these mAbs reactedwith epitopes in three different E domains in DI (amino acidresidue K179) in DII (I126) and in DIII (G132) for Fabs A3B2 and E3 respectively Moreover these Fabs as well as thederived humanized counterpart mAbs exhibited high neu-tralizing activities against a broad spectrum of JEV genotypestrains Moreover these mAbs exhibited a 50 protectivedose of 084 120583g (B2) 58120583g (A3) and 247120583g (E3) in mousemodels Finally administration of 200 120583gmouse of mAb B2one day after otherwise lethal JEV infection protected 50of mice and significantly prolonged the average survival timecompared to that of mice in the unprotected group [54]
29 St Louis Encephalitis Virus (SLEV) St Louis encephalitisvirus (SLEV) was first discovered as the mosquito-borneagent responsible for over 1000 cases of encephalitis duringa 1933 summer outbreak in St Louis (Missouri) and nowis a reemerging human pathogen widely distributed in theAmerican continent causing several human encephalitisoutbreaks over the last 80 years Additional epidemics haveindeed occurred from 1964 to 2006 in the Americas rangingfrom the US to Argentina and Brazil As a member of theFlavivirus genus the SLEVE glycoprotein ectodomain is 68identical to serocomplex-related JEVEbut only 46 and 40identical to those ofDENV2 andTBEV viruses fromdifferentserocomplexes [115]
291 mAbs against SLEV In 1983 Roehrig et al isolatedtwenty-one hybridomas producing murine mAbs specificfor the E glycoprotein of SLEV strain MSI-7 Serologicreactivities were initially determined by cross-reactivity indi-rect immunofluorescence assays using 22 strains of SLEVand 8 other related flaviviruses Four groups demonstratingtype- subcomplex- supercomplex- and group-specific reac-tivity patterns were identified Analysis of hemagglutination-inhibition and virus neutralization subdivided the cross-reactivity groups into eight epitopes (E-1a b c d E-2 E-3 and E-4a b) one of them following localized in DII (4b)[52 53]
Moreover among the previously described isolated anti-WNV mAbs by Gould et al the chimeric scFv-Fc 79 effec-tively neutralized also SLEV resulting in gt80 of PRNT80when used at 5 120583gmL [42]
BioMed Research International 13
3 Alphaviruses
Major human arboviruses in the Togaviridae family Chikun-gunya (CHIKV) and Venezuelan equine encephalomyelitis(VEEV) viruses belong to the Alphavirus genus which iscomposed of viruses with an icosahedral nucleocapsid sur-rounded by a lipid envelope and glycoprotein spikes Thestructural proteins of alphaviruses arise through co- andpost-translational processing of a polyprotein encoded by asingle positive-stranded RNAproducing the capsid (C) PE26 K and E1 The PE2 glycoprotein is a precursor containingthe E3 glycoprotein fused to the amino terminus of the E2envelope glycoprotein The PE2 glycoprotein is followed by6K a small membrane-associated protein and E1 the secondpolypeptide component of glycoprotein spikes
Trimerized heterodimers of the E1 and E2 viral glyco-proteins form the surface spikes and contain determinantsof viral tropism and virulence The E3 glycoprotein acts as asignal for transport of PE2 across themembranes of the roughendoplasmic reticulum and may promote the formation andintracellular transport of E1-PE2 heterodimers to the cellsurface During transport to the cell surface PE2 undergoesa maturational cleavage event by a furin-like protease toproduce E2 and E3 [116]
The E2 glycoprotein promotes specificity of virus bindingto the host cell surface and is a target of the humoral immuneresponse The E1 glycoprotein mediates fusion of the virionenvelope with the membranes of acidified endosomes allow-ing release of the nucleocapsid into the cytoplasm and theonset of viral replicationAlphavirus E1 shares no appreciablesequence identity to Flavivirus E protein Despite differencesin their amino acid sequences and arrangements on the viralparticle the structures of E and E1 are remarkably similarIndeed the conservation of three domains has been welldocumented in crystal structures of flavivirus E ectodomainsfrom TBEV DENV JEV and WNV as well as Alphavirus E1from Semliki Forest virus (SFV) and CHIKV Abs to the E1glycoprotein do not typically neutralize virus infectivity invitro but can protect against lethal challenge in animals
By analogy with the prM protein of dengue Flavivirusfurin cleavage of PE2 may begin after transit of an acidiclate component of the Golgi body where E3 is thought tosuppress the acid pH-triggered activation of glycoprotein E1fusion capability Protective anti-E3 mouse mAbs have alsobeen described [117]
31 Chikungunya Virus (CHIKV) The chikungunya virus(CHIKV) belongs to the Semliki Forest clade and was firstlyisolated in 1953 in Tanzania during an epidemic outbreakfollowing occurred also in Asia and Africa In the last fivedecades several CHIKV outbreaks have been described bothin Africa and Asia separated by gaps lasting from two totwenty years In 2005-2006 about 300000 cases out of785000 inhabitants were reported in La Reunion island witha fatal prognosis for 237 of the patients [117] Neither Europenor the Americas have had outbreaks of CHIKV so far exceptfor imported isolated cases Three genotypes of CHIKV aredescribed Asian EastCentralSouth African (ECSA) andWest African with an amino acid identity spanning from
952 to 998 Recent epidemics in Africa and Indiansubcontinent were caused by strains belonging to the ECSAgenotype Transmitted by Aedes mosquitoes CHIKV ismaintained in the human population by human-mosquito-human transmission [1] The disease is characterized bydengue-like symptoms such as chills high fever headacheand persistent myalgia and a further incapacitating arthralgia(from which the name chikungunya) which affects 40 ofinfected subjects However prognosis is rarely fatal [116]
There is currently no commercial vaccine and antiviraltreatment forCHIKV although some candidate vaccines havebeen tested in humans In this regard in 2000 US Armyperformed a Phase II clinical trial testing a live-attenuatedCHIKV vaccine (TSI-GSD-218) derived from a 1962 strain(15561) of an outbreak in Thailand Out of 58 every patientdeveloped neutralizingAbs and 5 lamentedmild tomoderatejoint pain [117] A phase III trial of this candidate vaccineis ongoing Furthermore a new formulation using virus-likeparticles was able to induce neutralizing Abs in macaquesagainst different CHIKV strains [117 118] Indeed it has beendescribed that infection seems to elicit long-lasting protectiveimmunity and cross-protection among CHIKV and otheralphaviruses
CHIKV entry into host cell is demonstrated to be medi-ated by envelope glycoproteins E1 and E2 which allowvirus fusion to cell membrane in low pH conditions andrecognition of an unknown cellular receptor respectively[116 119ndash121] Even though the genome replication relies onan error-prone RNA-dependent RNA-polymerasis CHIKVstrains S27 and 05115 Reunion showed in E1 and E2 alow amino acid variation of 068 and 33 respectivelyand such a low variability may be needed for an effectivereplication in two phylogenetically distant hosts [116 122]Recently the crystal structure of E1 and E2 heterodimerhas been resolved both at low pH and natural conditionsshedding light on E1 and E2 ectodomainsrsquo structure Inparticular similarly to flaviviruses E protein E1 ectodomainis made of the N-terminal DI the DII containing the fusionloop and the DIII at the C-terminal On the other hand E2ectodomain comprises theN-terminal domainA the domainB supposed to interact with the host cellrsquos unknown receptorand domain C at the C-terminal E1 DIII and E2 domain Care located close to the viral membrane [123 124]
311 mAbs against CHIKV Warter et al described 5F10 and8B10 two human mAbs which strongly neutralized severalCHIKV isolates in vitro without cross-reactivity againstother alphaviruses but to Onyong-nyong virus [56] Mixedpreparation of 5F10 and 8F10 did not show neither synergisticnor addictive effect in vitro and studies upon escape mutantsdemonstrated that 5F10 mAb binds at the tip of the E2domain B while 8B10 recognizes residues close to E1 fusionloop and amino acids within E2 domain A which form atransitional epitope under low pH conditions The authorsspeculated that 5F10 and 8B10 may inhibit CHIKV entry andfusion to the cell membrane respectively It is worth notingthat the previously mentioned escape mutants displayedmutations associated with reduced viral fitness in vitro evenafter 13 neutralizationamplification rounds In the same
14 BioMed Research International
study cell-to-cell transmission was firstly demonstrated as anescapemechanism enhanced by the E2mutation R82G [122]
In further in vivo studies 5F10 and 8B10 significantlydelayed CHIKV-caused death of AGR129 mice both in pro-phylactic and therapeutic tests Interestingly in therapeutictreatment these mAbs showed a synergistic effect whenadministered in combination with the total amount of mAbsinjected being the same of the single-mAbs Possibly thesetreatment did not allow themice to survive CHIKV challengedue to Ig clearing and 6ndash10 days half-life usually described forhuman mAbs [125]
Recently Pal et al cloned thirty-six murine mAbs ableto neutralize the ECSA La Reunion 2006 OPY-1 strain ofCHIKV (CHIKV-LR) the majority of which also neutralizeinfection of other strains corresponding to the Asian andWest African genotypes In particularmAbCHK-152 showedthe highest and broadest neutralizing activity with FRNTvalues indicating an IC50 value of 1 to 3 ngmL dependingon the viral strain used Among the thirty-six mAbs four(CHK-102 CHK-152 and CHK-166 CHK-263) could pre-vent lethality of immunodeficient Ifnarminusminus C57BL6 micewhen administered one day before exposition with CHK-152 and CHK-263 showing the ability to protect mice at thelowest dose (10 120583g) Therapeutic studies were performed onthese mAbs administering a single dose of 100 120583g 24 hoursafter infection The highest activity was described for CHK-166 able to protect 63 of the mice Testing the combinedactivity of the mAbs in therapeutic studies administration ofCHK-166 plus CHK-152 in a dose of 250120583g each turned out tobe themost effective protecting 71 of the IfnarminusminusC57BL6mice 60 hours after infection [57]
32 Venezuelan Equine Encephalomyelitis Virus (VEEV)Venezuelan equine encephalomyelitis virus (VEEV) is main-tained in a natural transmission cycle between mosquitoesand small rodents The first documented outbreaks occurredin the 1930s and several epidemics have been reported so farin Latin-American countries such as Venezuela ColombiaPeru Ecuador Bolivia Honduras Mexico and Panama withhundreds of thousands of human cases being reported anda case-fatality rate up to 1 during the 1969 Ecuador out-break Clinical manifestations of VEE are indistinguishablefrom Dengue [126] Six serogroups (IndashVI) are currentlyrecognized within the VEEV complex VEEV caused humanand equine outbreaks in the Americas for nearly a centuryEquine epizootics have high mortality (38ndash83) leading toa high viremia followed by a lethal encephalitis and often tohuman epidemics involving thousands of cases and hundredsof deaths Indeed VEEV is infectious for humans by theairborne route and has been responsible for a number oflaboratory infections In humans the disease is usually self-limiting with a febrile illness with 1ndash4 of cases progressingto severe encephalitis It is a hazard to laboratory workershas been developed as a biological weapon and is a potentialbioterrorist agent [127] There are no antiviral drugs andvaccines licensed for the treatment of VEEV infection inhumans Indeed although experimental live-attenuated vac-cines have been developed (eg TC-83) with good levels ofprotection in equine and mice in humans they may fail to
give protection in the majority of cases [128] Consideringthe protective role exerted by humoral immune response inanimal models it could be concluded that treatment withspecific IgG may have a beneficial antiviral effect in humanairborne infections with pathogenic strains of VEEV
321 mAbs against VEEV Several works support that mAbsmay protect against airborne VEEV as well as after airborneexposure toVEEVAb reactivity to both surface glycoproteins(E1 and E2) is associated with neutralization of the virusin vitro and passive protection against virus challenge Aseries of anti-E1 and anti-E2 mouse and humanized mAbshave been described recently Among them Phillpotts et alexamined two anti-E2 murine mAbs named 1A4A-1 and1A3A-9 both having potent protective activity against subcu-taneous VEEV challenge in mice Both mAbs had a similarhalf-life (58 and 100 days) in mouse serum after a singleintraperitoneal dose suggesting that mAbs delivered by thisroute at or around the time of VEEV infection persists athigh levels in the blood as well as secretions in respiratorytransudation throughout the clinical course of the disease Inparticular both mAbs administered 24 h prior to airbornechallenges had a substantial protective effect (90ndash100)Treatment of mice 2 or 24 h after airborne infection with asingle intraperitoneal dose of 100 120583g of 1A3A-9 mAb led toapproximately 50 survival There was no beneficial effectwhen mAb treatment was delayed to 72 h Moreover therewas evidence of synergy in vitro in PRNT between 1A3A-9and 1A4A-1 as has been demonstrated for other virusesand mAb pairs However no synergy was found in mouseprotection when mAbs were delivered intraperitoneally as amixture in equal parts to animals challenged 24 h previouslywith VEEV [129]
In a subsequent study Phillpotts et al reported two otherprotective mAbs The 3B2A-9 mAb protected against allthe serogroup I strains while 1A3B-7 mAb protected wellagainst challenge with all of the viruses tested Both 3B2A-9 and 1A3B-7 protected against airborne exposure to theIAB serogroup virus strainTrinidad donkey (TrD)Howeverthere was no evidence of synergistic protection when thesemAbs were combined in equal proportions An intraperi-toneal dose of 10 120583g was sufficient to protect 50 of themice with either mAb [130] According to other data themechanism of protection did not appear to depend uponneutralization [131] Similar results were obtained by OrsquoBrienet al which described a non-neutralizing mAb (IgG2a)named CUF37-2a able to protect 50 of mice from asubcutaneous VEEV challenge when a dose of 915 120583g of mAbwas administered 24 h prior to challenge [132]
In a following study the 1A3B-7 mAb was humanized(and following reported as Hu1A3B-7) maintaining the samefeatures of its murine counterpart In particular evaluation ofin vitro studies indicated that Hu1A3B-7 retained both broadspecificity and neutralizing activity Furthermore in vivoexperiments showed that Hu1A3B-7 successfully protectedmice against lethal subcutaneous and aerosol challenges withVEEV strain TrD Moreover the effectiveness of the human-ization process was determined by assessing proliferationresponses in human T-cells to peptides derived from the
BioMed Research International 15
murine and humanized versions of the VH and VL domainsThis analysis showed that the number of human T-cellepitopes within the humanized Ab had been substantiallyreduced indicating that Hu1A3B-7 may have low immuno-genicity in vivo [133]
Similarly Hunt et al described a humanizedmurinemAbderived from the 3B4C-4 mAb recognizing the E2c epitope(amino acid residues 182ndash209) and following named Hy4-26C showing neutralizing activity similar to the murinemAb Moreover it was protective (70ndash100 of survival) at100 ng dose in mice animal model intraperitoneal inoculatedand at 500120583g for intranasal challenge (80) Therapeuticstudies in mice revealed that Hy4-26C was able to cure miceup to 24 h following infection at 10 120583g on the contrary themouse mAb when given 1 hour after virus challenge [63]
In another work Hunt et al isolated two neutralizinghumanized Fabs F5 and L1A7 employing a blocking strategy[134 135] The anti-E2 specific F5 IgG had a 70 PRNTendpoint of 10 ngmL equivalent to that described for themost effective neutralizing anti-VEEVE2mAbs In particularthe E1-specific hFab L1A7 had a PRNT endpoint of 3120583gmL300-fold lower than F5
Further studies revealed that F5 epitope is located inthe 115ndash199 amino acid region Moreover F5 IgG had potentability to protect mice from infection by either route whenadministered 24 h before exposure however mice treated24 h and 48 h after aerosol exposure developed central ner-vous system infections but exhibited no clinical signs ofdisease [135]
Hu et al reported that passive immunization with thehumanized chimericmousemAbhu1A4A1IgG1-2A inmice at50 120583g 24 h before or after virulent VEEV challenge providedcomplete protection indicating that hu1A4A1IgG1-2A haspotent prophylactic and therapeutic effects against VEEVinfection [136]
Anti-VEEV mAbs isolated from a non-human primategene library has also been reported In particular thehumanized chimeric scFv-Fc ToR67-3B4 recognized viableas well as formalin and 120573-propiolactone inactivated virusparticles It detected specifically the viral E1 envelope proteinof VEEV but did not react with reduced viral glycopro-tein preparations suggesting that recognition depends uponconformational epitopes The recombinant Ab was able todetect multiple VEEV subtypes and displayed only marginalcross-reactivity to otherAlphavirus species except for Easternequine encephalitis virus (EEEV) In addition the scFv-Fcfusion described here might be of therapeutic use since itsuccessfully inactivated VEEV in a murine disease modelIn particular when the recombinant Ab was administered 6hours after challenge 80 to 100 of mice survived lethalVEEV IAB or IE infection Forty to sixty percent of micesurvived when scFv-Fc ToR67-3B4 was applied 6 hours afterchallenge with VEEV subtypes II and former IIIA [137]
4 Bunyaviridae
TheBunyaviridae family contains human arboviruses belong-ing to the Orthobunyavirus Phlebovirus and Nairovirus
genera Bunyaviridae includes enveloped viruses with a frag-mented single-stranded RNA genome of negative polarityTheir tripartite genome consists of a small (S) a medium(M) and a large (L) fragment and the envelope glycoproteinsGn and Gc that are cleaved out of a polyprotein synthesizedby the M fragment These glycoproteins have been provedto mediate the formation of the virus particle to play a rolein the interaction with cell surface receptors to mediate theentry of the virus into cells and to serve as targets for themajority of neutralizing Abs described so far
Orthobunyavirus transmitted through mosquitoes ormidges vectors are divided in 18 serogroups based on cross-titrations in haemagglutination inhibition assays and neutral-ization assays and correlating with main vector preferencesHowever the most clinically important viruses belong onlyto two serogroups the California encephalitis and the Simbuserogroups
Crimean-Congo hemorrhagic fever virus (CCHFV) isconsidered the only one of clinical relevance within theNairovirus genus which uses tick as main vector and is divid-ed in seven serogroups while Toscana virus and Rift Valleyfever virus (RVFV) which are transmitted by sandflies andmosquitoes respectively belong to the Phlebovirus genus[138]
MAbs against theNairovirusCCHFV and the PhlebovirusRVFV have been described
41 Crimean-Congo Hemorrhagic Fever Virus (CCHFV)Crimean-Congo Hemorrhagic Fever Virus (CCHFV) dis-tribution covers the greatest geographic range of any tick-borne virus known as viral isolation andor disease hasbeen reported from more than 30 countries in SoutheasternEurope Africa Asia and Middle East [139 140] Namelyfrom 1953 to 2010 about 6000 human cases were reportedin Southeastern Europe CCHFV causes sporadic outbreakswith mortality rates ranging from 10 to 80 [141 142] Asignificant variation in the time of incubation has beendescribed [139] Prehemorrhagic symptoms include highfever chills headache photophobia and back and abdominalpains Among other symptoms neuropsychiatric changeshave been reported in some patients In severe cases 3ndash6 daysafter the onset of the disease hemorrhagic symptoms occurwith petechiae ecchymosis bleeding in the form of melenahematemesis epistaxis [139]
In comparisonwith the nucleocapsid proteins inCCHFVGn and Gc show a higher degree of antigenic variabilityprobably due to their exposition to the host immune systemhypothesis corroborated in the description by Hewson et alin 2004 of four M segment phylogenic groups namely M1M2 M3 and M4 [141 142]
411 mAbs against CCHFV Blackburn et al firstly describedin 1987 murine anti-CCHFV mAbs able to recognize nucle-ocapsid proteins but no neutralizing activity has beendescribed thus far [143]
On the other hand Bertolotti-Ciarlet et al produced apanel of murine mAbs recognizing conformational epitopes
16 BioMed Research International
within theGn andGc glycoproteins expressed by the CCHFVstrain IbAr10200 The effectiveness of the mAbs was studiedby performing PRNT80 and none of the anti-Gn mAbsshowed neutralizing activity although many of the anti-GcmAbs neutralized IbAr10200 strain in vitro with clones 8A15E3 12A9 6C2 and 9H3 showing activity at a gt5120 dilutionMAbs 11E7 and 30F7 exhibited neutralizing activity at 2560dilution A suckling mice protection test was thereforeperformed for these mAbs The previously mentionedanti-Gc Abs were capable to protect mice to an appreciabledegree when applied 24 hours before and in a weakermanner 24 hours after virus challenge Many of the anti-GnmAbs were able to confer significant protection to IbAr10200both 24 hours before and after virus administration Therelevant effectiveness of the anti-Gn mAbs 6B12 10E1113G8 and 10G4 suggests that these mAbs may possess someneutralizing activity due to Ab-based effector mechanisms(eg ADCC) [58]
In further studies the same group characterized thebroadly cross-reactive neutralizing activity of the previouslydescribed murine mAb 11E7 able to bind conformationalepitopes within the C-terminal region of the Gn glycoproteinof all CCHFVM groups [58 141]
42 Rift Valley Fever Virus (RVFV) Similarly to CCHFV RiftValley Fever Virus (RVFV) causes sporadic outbreaks Thelargest epidemic occurred in Egypt in 1977 with an estimated200000 infections 18000 patients manifesting symptomsand 600 fatal prognoses [144] Symptoms include hemor-rhagic episodes fever encephalitis and blindness [145]
The major target of the humoral immune response is theRVFV glycoproteins Gc (G1) and Gn (G2) ranging fromnucleotides 480ndash2090 and 2091ndash3614 respectively [144]
421 mAbs against RVFV Within G2 glycoprotein are des-cribed three neutralizing antigenic sites namely epitope Inucleotides 792ndash893 (amino acid residues 258ndash291) epitopeII nucleotides 1164ndash1196 (amino acid residues 382ndash392) andepitope IV nucleotides 858ndash917 (amino acid residues 280ndash299) [59 145]
Despite the high degree of variability observed inCCHFVglycoproteins RVFV G2 presented an unexpected high con-servation among 22 isolates [59] In the same study onlythe LUNYO and 900060 isolates were resistant to in vitroneutralization by murine mAb 4D4 (recognizing epitope II)All other isolates were neutralized with PRNT80 titers from10240 togt81920On the other hand only SNS isolate showedreduced sensitivity to 4ndash39-CC (epitope IV) All other isolateswere neutralized with PRNT80 titers from 20480 to gt81920Protection studies on mice would be of interest for thedevelopment of a passive immunization prophylaxis [59]
Protection studies from lethal South African RVFVAN1830 strain infection were successfully performed on abattery of anti-G1 and anti-G2 mAbs produced by Besselaarand Blackburn with the strongly neutralizing anti-G1 mAb3E5 and anti-G2mAb 9C4 being the most effective [146]The9C4 antigenic area maps within G2 epitope I and a broadlyneutralizing activity may be investigated
5 Conclusions
Arbovirus infections have acquired increasing interest giventhe augmented globalization and tourism movement all overthe world that could be at the basis of epidemic eventsFurthermore the same vectors can sometimes transmit sev-eral arboviruses concomitantly complicating the diagnosis aswell as the therapy For example confusing mixed epidemicshave occasionally been described such as YFV plus CHIKVDENVplus CHIKV ormore recentlyPlasmodium falciparummalaria plus DENV1 and CHIKV on Madagascarrsquos east coast[1]
In the last decade several efforts have been employed inthe development of effective immunogens and therapeuticsgiving the current absence of specific antiviral drugs as wellas effective vaccines for themost diffused arbovirus infections(ie DENV andWNV)
However at the moment mosquito control is the bestavailable method for preventing arbovirus infections Thusto control the emerging public health problem of arbovirusinfections new antiviral therapeutic strategies that providepotent and broadly cross-protective immunity (especially forflavivirus infections) are an urgent globally medical need
As described in this review the majority of highlyneutralizing mAbs are of murine origin and indeed theirutility in the treatment of these infections is limited In facttheir employment can elicit an immunogenic response (iehuman anti-mouse Abs HAMA) the therapeutic efficacycould be reduced by a relatively faster clearance in humans(compared to human Abs) potentially exacerbated by theHAMA response Finally murine Abs exhibit relatively weakeffector functions (eg ADCC) compared to human AbsIndeed chimerization humanization or better the isolationand characterization of fully human broadly neutralizingmAbs would be the best choice for their possible use in afuture mAb-based therapy against arbovirus infections [147]
Finally possible ADE of infection mechanisms should beevaluated before considering a mAb as a possible candidatetherapeutic in a post-exposure setting as well as in the devel-opment of new Ab-eliciting immunogens
Conflict of Interests
The authors declare no conflict of interests in particular withOmriGam and any other cited Ab-based preparation men-tioned in the paper
References
[1] N Cleton M Koopmans J Reimerink G J Godeke and CReusken ldquoCome fly with me review of clinically importantarboviruses for global travelersrdquo Journal of Clinical Virology vol55 no 3 pp 191ndash203 2012
[2] F X Heinz H Holzmann A Essl and M Kundi ldquoField effec-tiveness of vaccination against tick-borne encephalitisrdquoVaccinevol 25 no 43 pp 7559ndash7567 2007
[3] A D Barrett and D E Teuwen ldquoYellow fever vaccinemdashhowdoes it work and why do rare cases of serious adverse events
BioMed Research International 17
take placerdquo Current Opinion in Immunology vol 21 no 3 pp308ndash313 2009
[4] S B Halstead and S J Thomas ldquoNew vaccines for Japaneseencephalitisrdquo Current Infectious Disease Reports vol 12 no 3pp 174ndash180 2010
[5] R Burioni M Perotti N Mancini and M Clementi ldquoPer-spectives for the utilization of neutralizing human monoclonalantibodies as anti-HCV drugsrdquo Journal of Hepatology vol 49no 2 pp 299ndash300 2008
[6] R A Diotti G A Sautto L Solforosi NManciniM Clementiand R Burioni ldquoNeutralization activity and kinetics of twobroad-range human monoclonal IgG1 derived from recombi-nant Fab fragments and directed against Hepatitis C virus E2glycoproteinrdquo New Microbiologica vol 35 no 4 pp 475ndash4792012
[7] R Burioni F Canducci N Mancini et al ldquoMolecular cloningof the first humanmonoclona antibodies neutralizing with highpotency Swine-origin Influenza A pandemic virus (S-OIV)rdquoNew Microbiologica vol 32 no 4 pp 319ndash324 2009
[8] P K Chandra S Hazari B Poat et al ldquoIntracytoplasmic stableexpression of IgG1 antibody targeting NS3 helicase inhibitsreplication of highly efficient hepatitis C Virus 2a clonerdquoVirology Journal vol 7 article 118 2010
[9] N Clementi D de Marco N Mancini et al ldquoA human mono-clonal antibody with neutralizing activity against highly diver-gent influenza subtypesrdquo PLoS ONE vol 6 no 12 Article IDe28001 2011
[10] D de Marco N Clementi N Mancini et al ldquoA non-VH1-69 heterosubtypic neutralizing human monoclonal antibodyprotects mice against H1N1 and H5N1 virusesrdquo PLoS ONE vol7 no 4 Article ID e34415 2012
[11] L Solforosi N Mancini F Canducci et al ldquoA phage displayvector optimized for the generation of human antibody com-binatorial libraries and the molecular cloning of monoclonalantibody fragmentsrdquoNewMicrobiologica vol 35 no 3 pp 289ndash294 2012
[12] E V Koonin and V V Dolja ldquoEvolution and taxonomy ofpositive-strand RNA viruses implications of comparative anal-ysis of amino acid sequencesrdquo Critical Reviews in Biochemistryand Molecular Biology vol 28 no 5 pp 375ndash430 1993
[13] J S Mackenzie D J Gubler and L R Petersen ldquoEmergingflaviviruses the spread and resurgence of Japanese encephalitisWest Nile and dengue virusesrdquo Nature Medicine vol 10 no 12pp S98ndashS109 2004
[14] T J SmithW E Brandt J L Swanson J MMcCown and E LBuescher ldquoPhysical and biological properties of dengue-2 virusand associated antigensrdquo Journal of Virology vol 5 no 4 pp524ndash532 1970
[15] J M Smit B Moesker I Rodenhuis-Zybert and J WilschutldquoFlavivirus cell entry and membrane fusionrdquo Viruses vol 3 no2 pp 160ndash171 2011
[16] R J Kuhn W Zhang M G Rossmann et al ldquoStructure ofdengue virus implications for flavivirus organization matura-tion and fusionrdquo Cell vol 108 no 5 pp 717ndash725 2002
[17] K Stiasny J H Aberle M Keller B Grubeck-Loebenstein andF X Heinz ldquoAge affects quantity but not quality of antibodyresponses after vaccination with an inactivated flavivirus vac-cine against tick-borne encephalitisrdquo PLoS ONE vol 7 no 3Article ID e34145 2012
[18] LThomasOVerlaetenACabie et al ldquoInfluence of the dengueserotype previous dengue infection and plasma viral load
on clinical presentation and outcome during a dengue-2 anddengue-4 co-epidemicrdquo American Journal of Tropical Medicineand Hygiene vol 78 no 6 pp 990ndash998 2008
[19] Y Zhang J Corver P R Chipman et al ldquoStructures of imma-ture flavivirus particlesrdquo EMBO Journal vol 22 no 11 pp2604ndash2613 2003
[20] A K I Falconar ldquoIdentification of an epitope on the denguevirus membrane (M) protein defined by cross-protective mon-oclonal antibodies design of an improved epitope sequencebased on common determinants present in both envelope (EandM) proteinsrdquo Archives of Virology vol 144 no 12 pp 2313ndash2330 1999
[21] K-J Huang Y-C Yang Y-S Lin et al ldquoThe dual-specific bind-ing of dengue virus and target cells for the antibody-dependentenhancement of dengue virus infectionrdquo Journal of Immunol-ogy vol 176 no 5 pp 2825ndash2832 2006
[22] A K I Falconar ldquoAntibody responses are generated to immun-odominant ELKKLE-typemotifs on the nonstructural-1 glyco-protein during live dengue virus infections inmice and humansimplications for diagnosis pathogenesis and vaccine designrdquoClinical and Vaccine Immunology vol 14 no 5 pp 493ndash5042007
[23] C W Mandl F Guirakhoo H Holzmann F X Heinz and CKunz ldquoAntigenic structure of the flavivirus envelope protein Eat the molecular level using the tick-borne encephalitis virus asa modelrdquo Journal of Virology vol 63 no 2 pp 564ndash571 1989
[24] F A Rey F X Heinz C Mandl C Kunz and S C HarrisonldquoThe envelope glycoprotein from tick-borne encephalitis virusat 2 A resolutionrdquo Nature vol 375 no 6529 pp 291ndash298 1995
[25] S L Allison K Stiasny K Stadler CWMandl and F XHeinzldquoMapping of functional elements in the stem-anchor regionof tick-borne encephalitis virus envelope protein Erdquo Journal ofVirology vol 73 no 7 pp 5605ndash5612 1999
[26] D E Klein J L Choi and S C Harrison ldquoStructure of a denguevirus envelope protein late-stage fusion intermediaterdquo Journal ofVirology vol 87 no 4 pp 2287ndash2293 2013
[27] S L Hanna T C Pierson M D Sanchez A A Ahmed M MMurtadha and R W Doms ldquoN-linked glycosylation of WestNile virus envelope proteins influences particle assembly andinfectivityrdquo Journal of Virology vol 79 no 21 pp 13262ndash132742005
[28] N Mancini G Sautto N Clementi et al ldquoNeutralization inter-fering antibodies a ldquonovelrdquo example of humoral immune dys-function facilitating viral escaperdquoViruses vol 4 no 9 pp 1731ndash1752 2012
[29] Y Modis S Ogata D Clements and S C Harrison ldquoVariablesurface epitopes in the crystal structure of dengue virus type3 envelope glycoproteinrdquo Journal of Virology vol 79 no 2 pp1223ndash1231 2005
[30] N Miller ldquoRecent progress in dengue vaccine research anddevelopmentrdquo Current Opinion in Molecular Therapeutics vol12 no 1 pp 31ndash38 2010
[31] J H Chavez J R Silva A A Amarilla and L T Moraes Figu-eiredo ldquoDomain III peptides from flavivirus envelope proteinare useful antigens for serologic diagnosis and targets forimmunizationrdquo Biologicals vol 38 no 6 pp 613ndash618 2010
[32] B Shrestha J D Brien S Sukupolvi-Petty et al ldquoThe develop-ment of therapeutic antibodies that neutralize homologous andheterologous genotypes of dengue virus type 1rdquo PLoS Pathogensvol 6 no 4 Article ID e1000823 2010
18 BioMed Research International
[33] G D Gromowski N D Barrett and A D T Barrett ldquoChar-acterization of dengue virus complex-specific neutralizing epi-topes on envelope protein domain III of dengue 2 virusrdquo Journalof Virology vol 82 no 17 pp 8828ndash8837 2008
[34] S Sukupolvi-Petty S K Austin W E Purtha et al ldquoType- andsubcomplex-specific neutralizing antibodies against domain IIIof dengue virus type 2 envelope protein recognize adjacentepitopesrdquo Journal of Virology vol 81 no 23 pp 12816ndash128262007
[35] S Sukupolvi-Petty S K Austin M Engle et al ldquoStructure andfunction analysis of therapeutic monoclonal antibodies againstdengue virus type 2rdquo Journal of Virology vol 84 no 18 pp9227ndash9239 2010
[36] Y-Q Deng J-X Dai G-H Ji et al ldquoA broadly flaviviruscross-neutralizingmonoclonal antibody that recognizes a novelepitope within the fusion loop of e proteinrdquo PLoS ONE vol 6no 1 Article ID e16059 2011
[37] J M Costin E Zaitseva K M Kahle et al ldquoMechanistic studyof broadly neutralizing human monoclonal antibodies againstdengue virus that target the fusion looprdquo Journal of Virologyvol 87 no 1 pp 52ndash66 2013
[38] CMMidgley A FlanaganH B Tran et al ldquoStructural analysisof a dengue cross-reactive antibody complexed with envelopedomain III reveals the molecular basis of cross-reactivityrdquoJournal of Immunology vol 188 no 10 pp 4971ndash4979 2012
[39] N J Moreland P Susanto E Lim et al ldquoPhage display appro-aches for the isolation of monoclonal antibodies against denguevirus envelope domain III from human and mouse derivedlibrariesrdquo International Journal ofMolecular Sciences vol 13 no3 pp 2618ndash2635 2012
[40] J J B Cockburn M E Navarro Sanchez N Fretes et alldquoMechanism of dengue virus broad cross-neutralization by amonoclonal antibodyrdquo Structure vol 20 no 2 pp 303ndash3142012
[41] R Rajamanonmani C Nkenfou P Clancy et al ldquoOn a mousemonoclonal antibody that neutralizes all four dengue virusserotypesrdquo Journal of General Virology vol 90 no 4 pp 799ndash809 2009
[42] L H Gould J Sui H Foellmer et al ldquoProtective and herapeuticcapacity of human single-chain Fv-Fc fusion proteins againstWest Nile virusrdquo Journal of Virology vol 79 no 23 pp 14606ndash14613 2005
[43] H Sultana H G Foellmer G Neelakanta et al ldquoFusion LoopPeptide of the West Nile virus envelope protein is essential forpathogenesis and is recognized by a therapeutic cross-reactivehuman monoclonal antibodyrdquo Journal of Immunology vol 183no 1 pp 650ndash660 2009
[44] T Oliphant M Engle G E Nybakken et al ldquoDevelopment ofa humanized monoclonal antibody with therapeutic potentialagainstWest Nile virusrdquoNature Medicine vol 11 no 5 pp 522ndash530 2005
[45] G E Nybakken T Oliphant S Johnson S Burke M S Dia-mond and D H Fremont ldquoStructural basis of West Nile virusneutralization by a therapeutic antibodyrdquo Nature vol 437 no7059 pp 764ndash769 2005
[46] G D Gromowski J T Roehrig M S Diamond J C LeeT J Pitcher and A D T Barrett ldquoMutations of an antibodybinding energy hot spot on domain III of the dengue 2 envelopeglycoprotein exploited for neutralization escaperdquo Virology vol407 no 2 pp 237ndash246 2010
[47] J T Roehrig R A Bolin andR G Kelly ldquoMonoclonal antibodymapping of the envelope glycoprotein of the dengue 2 virusJamaicardquo Virology vol 246 no 2 pp 317ndash328 1998
[48] R de Alwis S A Smith N P Olivarez et al ldquoIdentification ofhumanneutralizing antibodies that bind to complex epitopes ondengue virionsrdquo Proceedings of the National Academy of Sciencesof the United States of America vol 109 no 19 pp 7439ndash74442012
[49] S K Austin K A Dowd B Shrestha et al ldquoStructural basis ofdifferential neutralization of DENV-1 genotypes by an antibodythat recognizes a cryptic epitoperdquo PLoS Pathogens vol 8 no 10Article ID e1002930 2012
[50] M Throsby C Geuijen J Goudsmit et al ldquoIsolation and char-acterization of human monoclonal antibodies from individualsinfected with West Nile Virusrdquo Journal of Virology vol 80 no14 pp 6982ndash6992 2006
[51] A P Goncalvez R H Purcell and C-J Lai ldquoEpitope deter-minants of a chimpanzee Fab antibody that efficiently cross-neutralizes dengue type 1 and type 2 viruses map to inside andin close proximity to fusion loop of the dengue type 2 virusenvelope glycoproteinrdquo Journal of Virology vol 78 no 23 pp12919ndash12928 2004
[52] J T Roehrig J H Mathews and D W Trent ldquoIdentification ofepitopes on the E glycoprotein of Saint Louis encephalitis virususing monoclonal antibodiesrdquo Virology vol 128 no 1 pp 118ndash126 1983
[53] J T Roehrig L A Staudinger A R Hunt J H Mathews andC D Blair ldquoAntibody prophylaxis and therapy for flavivirusencephalitis infectionsrdquo Annals of the New York Academy ofSciences vol 951 pp 286ndash297 2001
[54] A P Goncalvez C-H Chien K Tubthong et al ldquoHumanizedmonoclonal antibodies derived from chimpanzee fabs protectagainst Japanese encephalitis virus in vitro and in vivordquo Journalof Virology vol 82 no 14 pp 7009ndash7021 2008
[55] M Arakawa T Yamashiro G-I Uechi M Tadano and ANishizono ldquoConstruction of humanFab (1205741120581) library and iden-tification of human monoclonal Fab possessing neutralizingpotency against Japanese encephalitis virusrdquo Microbiology andImmunology vol 51 no 6 pp 617ndash625 2007
[56] L Warter C Y Lee R Thiagarajan et al ldquoChikungunya virusenvelope-specific human monoclonal antibodies with broadneutralization potencyrdquo Journal of Immunology vol 186 no 5pp 3258ndash3264 2011
[57] P Pal K A Dowd J D Brien et al ldquoDevelopment of a highlyprotective combination monoclonal antibody therapy againstchikungunya virusrdquo PLoS Pathogens vol 9 no 4 Article IDe1003312 2013
[58] A Bertolotti-Ciarlet J Smith K Strecker et al ldquoCellular local-ization and antigenic characterization of Crimean-Congo hem-orrhagic fever virus glycoproteinsrdquo Journal of Virology vol 79no 10 pp 6152ndash6161 2005
[59] J K Battles and J M Dalrymple ldquoGenetic variation amonggeographic isolates of Rift Valley fever virusrdquo American Journalof Tropical Medicine and Hygiene vol 39 no 6 pp 617ndash6311988
[60] B AThibodeaux N C Garbino NM Liss et al ldquoA humanizedIgG but not IgM antibody is effective in prophylaxis and therapyof yellow fever infection in an AG12917D-204 peripheralchallenge mouse modelrdquo Antiviral Research vol 94 no 1 pp1ndash8 2012
BioMed Research International 19
[61] S Daffis R E Kontermann J Korimbocus H Zeller H-DKlenk and J Ter Meulen ldquoAntibody responses against wild-type yellow fever virus and the 17D vaccine strain charac-terization with human monoclonal antibody fragments andneutralization escape variantsrdquoVirology vol 337 no 2 pp 262ndash272 2005
[62] L N Levanov L E Matveev E P Goncharova et al ldquoChimericantibodies against tick-borne encephalitis virusrdquo Vaccine vol28 no 32 pp 5265ndash5271 2010
[63] A R Hunt S Frederickson C Hinkel K S Bowdish and J TRoehrig ldquoA humanized murine monoclonal antibody protectsmice either before or after challenge with virulent Venezuelanequine encephalomyelitis virusrdquo Journal of General Virologyvol 87 no 9 pp 2467ndash2476 2006
[64] J M da Silva Voorham I A Rodenhuis-Zybert N V A Nunezet al ldquoAntibodies against the envelope glycoprotein promoteinfectivity of immature dengue virus serotype 2rdquo PLoS ONEvol 7 no 3 Article ID e29957 2012
[65] T C Pierson Q Xu S Nelson et al ldquoThe stoichiometry ofantibody-mediated neutralization and enhancement of westnile virus infectionrdquoCell Host andMicrobe vol 1 no 2 pp 135ndash145 2007
[66] G Sautto N Mancini R A Diotti L Solforosi M Clementiand R Burioni ldquoAnti-hepatitis C virus E2 (HCVE2) glyco-proteinmonoclonal antibodies andneutralization interferencerdquoAntiviral Research vol 96 no 1 pp 82ndash89 2012
[67] R Burioni Y Matsuura N Mancini et al ldquoDiverging effectsof human recombinant anti-hepatitis C virus (HCV) antibodyfragments derived from a single patient on the infectivityof a vesicular stomatitis virusHCV pseudotyperdquo Journal ofVirology vol 76 no 22 pp 11775ndash11779 2002
[68] R J Phillpotts J R Stephenson and J S Porterfield ldquoAntibody-dependent enhancement of tick-borne encephalitus virus infec-tivityrdquo Journal of General Virology vol 66 no 8 pp 1831ndash18371985
[69] K A Dowd and T C Pierson ldquoAntibody-mediated neutraliza-tion of flaviviruses a reductionist viewrdquo Virology vol 411 no 2pp 306ndash315 2011
[70] A K I Falconar ldquoMonoclonal antibodies that bind to commonepitopes on the dengue virus type 2 nonstructural-1 andenvelope glycoproteins display weak neutralizing activity anddifferentiated responses to virulent strains implications forpathogenesis and vaccinesrdquo Clinical and Vaccine Immunologyvol 15 no 3 pp 549ndash561 2008
[71] K Meyer A Banerjee S E Frey R B Belshe and R Ray ldquoAweak neutralizing antibody response to hepatitis C virus enve-lope glycoprotein enhances virus infectionrdquo PLoS ONE vol 6no 8 Article ID e23699 2011
[72] R Burioni R AWilliamson P P Sanna F E Bloom and D RBurton ldquoRecombinant human Fab to glycoprotein D neutral-izes infectivity and prevents cell-to-cell transmission of herpessimplex viruses 1 and 2 in vitrordquo Proceedings of the NationalAcademy of Sciences of the United States of America vol 91 no1 pp 355ndash359 1994
[73] M S Rolph A Zaid N E Rulli and S Mahalingam ldquoDown-regulation of interferon-120573 in antibody-dependent enhancementof dengue viral infections of human macrophages is dependenton interleukin-6rdquo Journal of Infectious Diseases vol 204 no 3pp 489ndash491 2011
[74] S S Whitehead J E Blaney A P Durbin and B R MurphyldquoProspects for a dengue virus vaccinerdquoNature ReviewsMicrobi-ology vol 5 no 7 pp 518ndash528 2007
[75] S-M Lok V Kostyuchenko G ENybakken et al ldquoBinding of aneutralizing antibody to dengue virus alters the arrangement ofsurface glycoproteinsrdquoNature Structural andMolecular Biologyvol 15 no 3 pp 312ndash317 2008
[76] A P Goncalvez R Men C Wernly R H Purcell and C-J Lai ldquoChimpanzee Fab fragments and a derived humanizedimmunogiobulin G1 antibody that efficiently cross-neutralizedengue type 1 and type 2 virusesrdquo Journal of Virology vol 78no 23 pp 12910ndash12918 2004
[77] WM P BWahala A A Kraus L B HaymoreM A Accavitti-Loper and A M de Silva ldquoDengue virus neutralization byhuman immune sera role of envelope protein domain III-reactive antibodyrdquo Virology vol 392 no 1 pp 103ndash113 2009
[78] W D Crill H R Hughes M J Delorey and G-J J ChangldquoHumoral immune responses of dengue fever patients usingepitope-specific serotype-2 virus-like particle antigensrdquo PLoSONE vol 4 no 4 Article ID e4991 2009
[79] C Setthapramote T Sasaki O Puiprom et al ldquoHuman mono-clonal antibodies to neutralize all dengue virus serotypes usinglymphocytes from patients at acute phase of the secondaryinfectionrdquo Biochemical and Biophysical Research Communica-tions vol 423 no 4 pp 867ndash872 2012
[80] M Beltramello K L Williams C P Simmons et al ldquoThehuman immune response to dengue virus is dominated byhighly cross-reactive antibodies endowed with neutralizing andenhancing activityrdquo Cell Host andMicrobe vol 8 no 3 pp 271ndash283 2010
[81] C M Midgley M Bajwa-Joseph S Vasanawathana et al ldquoAnin-depth analysis of original antigenic sin in dengue virusinfectionrdquo Journal of Virology vol 85 no 22 pp 410ndash421 2011
[82] B Kaufmann M R Vogt J Goudsmit et al ldquoNeutralizationof West Nile virus by cross-linking of its surface proteins withFab fragments of the human monoclonal antibody CR4354rdquoProceedings of the National Academy of Sciences of the UnitedStates of America vol 107 no 44 pp 18950ndash18955 2010
[83] B Spurrier J M Sampson M Totrov et al ldquoStructural analysisof human and macaque mAbs 2909 and 25B implicationsfor the configuration of the quaternary neutralizing epitope ofHIV-1 gp120rdquo Structure vol 19 no 5 pp 691ndash699 2011
[84] J J SchlesingerMWBrandriss andT PMonath ldquoMonoclonalantibodies distinguish between wild and vaccine strains ofyellow fever virus by neutralization hemagglutination inhibi-tion and immune precipitation of the virus envelope proteinrdquoVirology vol 125 no 1 pp 8ndash17 1983
[85] M W Brandriss J J Schlesinger E E Walsh and M BrisellildquoLethal 17D yellow fever encephalitis in mice I Passive protec-tion by monoclonal antibodies to the envelope proteins of 17Dyellow fever and dengue 2 virusesrdquo Journal of General Virologyvol 67 no 2 pp 229ndash234 1986
[86] H Cho and M S Diamond ldquoImmune responses to West Nilevirus infection in the central nervous systemrdquo Viruses vol 4no 12 pp 3812ndash3830 2012
[87] D W Beasley A D Barrett and R B Tesh ldquoResurgence ofWest Nile neurologic disease in the United States in 2012 whathappened What needs to be donerdquo Antiviral Research vol 99no 1 pp 1ndash5 2013
[88] M Throsby J Ter Meulen C Geuijen J Goudsmit and J DeKruif ldquoMapping and analysis of West Nile virus-specific mon-oclonal antibodies prospects for vaccine developmentrdquo ExpertReview of Vaccines vol 6 no 2 pp 183ndash191 2007
[89] M S Diamond T G Roberts D Edgil B Lu J Ernst and EHarris ldquoModulation of dengue virus infection in human cells
20 BioMed Research International
by alpha beta and gamma interferonsrdquo Journal of Virology vol74 no 11 pp 4957ndash4966 2000
[90] Z Shimoni M J Niven S Pitlick and S Bulvik ldquoTreatmentof West Nile virus encephalitis with intravenous immunoglob-ulinrdquo Emerging Infectious Diseases vol 7 no 4 article 759 2001
[91] AHamdan PGreen EMendelsonM R Kramer S Pitlik andM Weinberger ldquoPossible benefit of intravenous immunoglob-ulin therapy in a lung transplant recipient with West Nile virusencephalitisrdquoTransplant InfectiousDisease vol 4 no 3 pp 160ndash162 2002
[92] M Haley A S Retter D Fowler J Gea-Banacloche and NP OrsquoGrady ldquoThe role for intravenous immunoglobulin in thetreatment of West Nile virus encephalitisrdquo Clinical InfectiousDiseases vol 37 no 6 pp e88ndashe90 2003
[93] K M Chung G E Nybakken B S Thompson et al ldquoAnti-bodies against West Nile virus nonstructural protein NS1prevent lethal infection through Fc 120574 receptor-dependent and-independent mechanismsrdquo Journal of Virology vol 80 no 3pp 1340ndash1351 2006
[94] E A Henchal L S Henchal and J J Schlesinger ldquoSynergisticinteractions of anti-NS1 monoclonal antibodies protect pas-sively immunized mice from lethal challenge with dengue 2virusrdquo Journal of General Virology vol 69 no 8 pp 2101ndash21071988
[95] J T Roehrig ldquoAntigenic structure of flavivirus proteinsrdquoAdvances in Virus Research vol 59 pp 141ndash175 2003
[96] M D Sanchez T C Pierson D McAllister et al ldquoCharacteri-zation of neutralizing antibodies to West Nile virusrdquo Virologyvol 336 no 1 pp 70ndash82 2005
[97] J H Beigel J L Nordstrom S R Pillemer et al ldquoSafety andpharmacokinetics of single intravenous dose of MGAWN1 anovel monoclonal antibody to West Nile virusrdquo AntimicrobialAgents and Chemotherapy vol 54 no 6 pp 2431ndash2436 2010
[98] C A Smeraski V Siddharthan and J D Morrey ldquoTreatmentof spatial memory impairment in hamsters infected with WestNile virus using a humanizedmonoclonal antibodyMGAWN1rdquoAntiviral Research vol 91 no 1 pp 43ndash49 2011
[99] D Lelli AMoreno E Brocchi et al ldquoWestNile virus character-ization and diagnostic applications of monoclonal antibodiesrdquoVirology Journal vol 9 article 81 2012
[100] T S Gritsun V A Lashkevich and E A Gould ldquoTick-borneencephalitisrdquo Antiviral Research vol 57 no 1-2 pp 129ndash1462003
[101] D Ruzek T S Gritsun N L Forrester et al ldquoMutations inthe NS2B and NS3 genes affect mouse neuroinvasiveness of aWestern European field strain of tick-borne encephalitis virusrdquoVirology vol 374 no 2 pp 249ndash255 2008
[102] M Ecker S L Allison T Meixner and F X Heinz ldquoSequenceanalysis and genetic classification of tick-borne encephalitisviruses from Europe and Asiardquo Journal of General Virology vol80 no 1 pp 179ndash185 1999
[103] H Holzmann M S Vorobyova I P Ladyzhenskaya et alldquoMolecular epidemiology of tick-borne encephalitis viruscross-protection between European and Far Eastern subtypesrdquoVaccine vol 10 no 5 pp 345ndash349 1992
[104] C W Mandl ldquoSteps of the tick-borne encephalitis virus repli-cation cycle that affect neuropathogenesisrdquo Virus Research vol111 no 2 pp 161ndash174 2005
[105] A V Atrasheuskaya T M Fredeking and G M IgnatyevldquoChanges in immune parameters and their correction in humancases of tick-borne encephalitisrdquo Clinical and ExperimentalImmunology vol 131 no 1 pp 148ndash154 2003
[106] K L Mansfield N Johnson L P Phipps J R Stephenson AR Fooks and T Solomon ldquoTick-borne encephalitis virusmdashareview of an emerging zoonosisrdquo Journal of General Virologyvol 90 no 8 pp 1781ndash1794 2009
[107] DWVaughn andCHHoke Jr ldquoThe epidemiology of Japaneseencephalitis prospects for preventionrdquo Epidemiologic Reviewsvol 14 pp 197ndash221 1992
[108] T Solomon H Ni D W C Beasley M Ekkelenkamp M JCardosa and A D T Barrett ldquoOrigin and evolution of Japaneseencephalitis virus in Southeast Asiardquo Journal of Virology vol 77no 5 pp 3091ndash3098 2003
[109] M R Wilson ldquoEmerging viral infectionsrdquo Current Opinion inNeurology vol 26 no 3 pp 301ndash306 2013
[110] J N Hanna S A Ritchie D A Phillips et al ldquoAn outbreakof Japanese encephalitis in the Torres Strait Australia 1995rdquoMedical Journal of Australia vol 165 no 5 pp 256ndash260 1996
[111] J N Hanna S A Ritchie D A Phillips et al ldquoJapanese ence-phalitis in north Queensland Australia 1998rdquo Medical Journalof Australia vol 170 no 11 pp 533ndash536 1999
[112] SGeorgeMGourie-Devi and J A Rao ldquoIsolation ofWestNilevirus from the brains of children who had died of encephalitisrdquoBulletin of theWorldHealthOrganization vol 62 no 6 pp 879ndash882 1984
[113] A K Gupta A A Koshy and V J Lad ldquoEnhanced protec-tion of mice against Japanese encephalitis virus infection bycombinations ofmonoclonal antibodies to glycoprotein ErdquoActaVirologica vol 55 no 2 pp 165ndash168 2011
[114] T H Lee B-H Song S-I Yun et al ldquoA cross-protective mAbrecognizes a novel epitope within the flavivirus NS1 proteinrdquoJournal of General Virology vol 93 no 1 pp 20ndash26 2012
[115] S C Weaver ldquoHost range amplification and arboviral diseaseemergencerdquo Archives of Virology Supplementum no 19 pp 33ndash44 2005
[116] M Solignat B Gay S Higgs L Briant and C Devaux ldquoRepli-cation cycle of chikungunya a re-emerging arbovirusrdquoVirologyvol 393 no 2 pp 183ndash197 2009
[117] O Schwartz and M L Albert ldquoBiology and pathogenesis ofchikungunya virusrdquo Nature Reviews Microbiology vol 8 no 7pp 491ndash500 2010
[118] W Akahata Z-Y Yang H Andersen et al ldquoA virus-like particlevaccine for epidemic Chikungunya virus protects nonhumanprimates against infectionrdquo Nature Medicine vol 16 no 3 pp334ndash338 2010
[119] M Kielian and A Helenius ldquopH-Iinduced alterations in thefusogenic spike protein of Semliki Forest virusrdquo Journal of CellBiology vol 101 no 6 pp 2284ndash2291 1985
[120] J White and A Helenius ldquopH-Dependent fusion between theSemliki Forest virus membrane and liposomesrdquo Proceedings ofthe National Academy of Sciences of the United States of Americavol 77 no 6 pp 3273ndash3277 1980
[121] J Dubuisson and C M Rice ldquoSindbis virus attachment isola-tion and characterization of mutants with impaired binding tovertebrate cellsrdquo Journal of Virology vol 67 no 6 pp 3363ndash33741993
[122] C Y Lee Y-W Kam J Fric et al ldquoChikungunya virus neutral-ization antigens and direct cell-to-cell transmission are revealedby human antibody-escapemutantsrdquo PLoS Pathogens vol 7 no12 Article ID e1002390 2011
[123] J E Voss M-C Vaney S Duquerroy et al ldquoGlycoproteinorganization of Chikungunya virus particles revealed by X-raycrystallographyrdquo Nature vol 468 no 7324 pp 709ndash712 2010
BioMed Research International 21
[124] L Li J Jose Y Xiang R J Kuhn and M G Rossmann ldquoStruc-tural changes of envelope proteins during alphavirus fusionrdquoNature vol 468 no 7324 pp 705ndash708 2010
[125] J Fric S Bertin-Maghit C I Wang A Nardin and L WarterldquoUse of human monoclonal antibodies to treat Chikungunyavirus infectionrdquo Journal of Infectious Diseases vol 207 no 2 pp319ndash322 2013
[126] P V Aguilar J G Estrada-Franco R Navarro-Lopez C FerroA D Haddow and S C Weaver ldquoEndemic Venezuelanequine encephalitis in the Americas hidden under the dengueumbrellardquo Future Virology vol 6 no 6 pp 721ndash740 2011
[127] S C Weaver R Winegar I D Manger and N L ForresterldquoAlphaviruses population genetics and determinants of emer-gencerdquo Antiviral Research vol 94 no 3 pp 242ndash257 2012
[128] S Paessler and S C Weaver ldquoVaccines for Venezuelan equineencephalitisrdquo Vaccine vol 27 no 4 pp D80ndashD85 2009
[129] R J Phillpotts ldquoVenezuelan equine encephalitis virus complex-specific monoclonal antibody provides broad protection inmurine models against airborne challenge with viruses fromserogroups I II and IIIrdquoVirus Research vol 120 no 1-2 pp 107ndash112 2006
[130] R J Phillpotts L D Jones and S C Howard ldquoMonoclonalantibody protectsmice against infection anddiseasewhen giveneither before or up to 24 h after airborne challenge with virulentVenezuelan equine encephalitis virusrdquoVaccine vol 20 no 11-12pp 1497ndash1504 2002
[131] J H Mathews J T Roehrig and D W Trent ldquoRole of com-plement and the Fc portion of immunoglobulin G in immunityto Venezuelan equine encephalomyelitis virus infection withglycoprotein-specific monoclonal antibodiesrdquo Journal of Virol-ogy vol 55 no 3 pp 594ndash600 1985
[132] L M OrsquoBrien C D Underwood-Fowler S A Goodchild AL Phelps and R J Phillpotts ldquoDevelopment of a novel mon-oclonal antibody with reactivity to a wide range of Venezuelanequine encephalitis virus strainsrdquoVirology Journal vol 6 article206 2009
[133] L M OrsquoBrien S A Goodchild R J Phillpotts and S D Per-kins ldquoA humanised murine monoclonal antibody protectsmice from Venezuelan equine encephalitis virus Evergladesvirus and Mucambo virus when administered up to 48h afterairborne challengerdquo Virology vol 426 no 2 pp 100ndash105 2012
[134] N Clementi N Mancini L Solforosi M Castelli M Clementiand R Burioni ldquoPhage display-based strategies for cloning andoptimization of monoclonal antibodies directed against humanpathogensrdquo International Journal of Molecular Sciences vol 13no 7 pp 8273ndash8292 2012
[135] A R Hunt R A Bowen S Frederickson T Maruyama JT Roehrig and C D Blair ldquoTreatment of mice with humanmonoclonal antibody 24h after lethal aerosol challenge withvirulent Venezuelan equine encephalitis virus prevents diseasebut not infectionrdquo Virology vol 414 no 2 pp 146ndash152 2011
[136] W-G Hu A L Phelps S Jager et al ldquoA recombinant human-ized monoclonal antibody completely protects mice againstlethal challenge with Venezuelan equine encephalitis virusrdquoVaccine vol 28 no 34 pp 5558ndash5564 2010
[137] T Rulker L Voss P Thullier et al ldquoIsolation and characterisa-tion of a human-like antibody fragment (scFv) that inactivatesVEEV in vitro and in vivordquo PLoS ONE vol 7 no 5 Article IDe37242 2012
[138] C T Walter and J N Barr ldquoRecent advances in the molecularand cellular biology of bunyavirusesrdquo Journal of General Virol-ogy vol 92 no 11 pp 2467ndash2484 2011
[139] C A Whitehouse ldquoCrimean-Congo hemorrhagic feverrdquo Anti-viral Research vol 64 no 3 pp 145ndash160 2004
[140] M Mertens K Schmidt A Ozkul and M H Groschup ldquoTheimpact of Crimean-Congo hemorrhagic fever virus on publichealthrdquo Antiviral Research vol 98 no 2 pp 248ndash260 2013
[141] A A Ahmed J M McFalls C Hoffmann et al ldquoPresence ofbroadly reactive and group-specific neutralizing epitopes onnewly described isolates of Crimean-Congo hemorrhagic fevervirusrdquo Journal of General Virology vol 86 no 12 pp 3327ndash33362005
[142] R Hewson A Gmyl L Gmyl et al ldquoEvidence of segment reas-sortment in Crimean-Congo haemorrhagic fever virusrdquo Journalof General Virology vol 85 no 10 pp 3059ndash3070 2004
[143] N K Blackburn T G Besselaar A J Shepherd and R Swane-poel ldquoPreparation and use of monoclonal antibodies for iden-tifying Crimean-Congo hemorrhagic fever virusrdquo AmericanJournal of Tropical Medicine and Hygiene vol 37 no 2 pp 392ndash397 1987
[144] T Ikegami ldquoMolecular biology and genetic diversity of RiftValley fever virusrdquoAntiviral Research vol 95 no 3 pp 293ndash3102012
[145] KKeegan andM S Collett ldquoUse of bacterial expression cloningto define the amino acid sequences of antigenic determinantson the G2 glycoprotein of Rift Valley fever virusrdquo Journal ofVirology vol 58 no 2 pp 263ndash270 1986
[146] T G Besselaar and N K Blackburn ldquoTopological mapping ofantigenic sites on the Rift Valley fever virus envelope glycopro-teins using monoclonal antibodiesrdquo Archives of Virology vol121 no 1ndash4 pp 111ndash124 1991
[147] L G Presta ldquoEngineering of therapeutic antibodies to mini-mize immunogenicity and optimize functionrdquo Advanced DrugDelivery Reviews vol 58 no 5-6 pp 640ndash656 2006
Submit your manuscripts athttpwwwhindawicom
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anatomy Research International
PeptidesInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporation httpwwwhindawicom
International Journal of
Volume 2014
Zoology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Molecular Biology International
GenomicsInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioinformaticsAdvances in
Marine BiologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Signal TransductionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Evolutionary BiologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Biochemistry Research International
ArchaeaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Genetics Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in
Virolog y
Hindawi Publishing Corporationhttpwwwhindawicom
Nucleic AcidsJournal of
Volume 2014
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Enzyme Research
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Microbiology

12 BioMed Research International
use in humans However an emergency therapy in theabsence of a mass immunization is needed as no effectivetreatments are yet available [106]
271 mAbs against TBEV Levanov et al described the chim-erization of two murine mAbs (13D6 and 10C2) directedagainst the DIII and DII respectively of the TBEV E glyco-protein The chimeric mAbs present binding characteristicssimilar to the parentalmAbsMoreover as the parentalmAbsonly the chimeric mAb 13D6 was able to neutralize TBEVinfectivity in vitro In particular neutralization studies withthe murine chimeric and scFv forms of mAb 13D6 were per-formed In particular murine 13D6 showed an IC50 titer of115 120583gmL in Focus Reduction Neutralization Tests (FRNT)against TBEV strain 205 and of 29 120583gmL in PRNT againstTBEV strain Softjin Chimeric 13D6 showed an IC50 titer of45 120583gmL in FRNT against TBEV strain 205 and of 19 120583gmLin PRNT against TBEV strain Softjin ScFv 13D6 showed anIC50 titer of 167 120583gmL in FRNT against TBEV strain 205and of 112 120583gmL in PRNT against TBEV strain Softjin [62]
28 Japanese Encephalitis Virus (JEV) Japanese encephalitisvirus (JEV) is a mosquito-borne flavivirus that over thepast few decades has caused several outbreaks throughoutChina Southeast Asia Australia and Papua New Guineawith a prevalence recently estimated to be of about 70000casesyear [107ndash109] A 40 mortality was recorded in someof the JEV-affected areas Moreover many survivors facesome neurological problems and complications [108] Since1995 the disease has also emerged in Non-Asian regions suchas Northern Australia [110 111] The situation in SoutheastAsia however is further complicated by the overlappingepidemics of JEV and DENV as well as sporadic casesof WNV infections detected in some of the affected areasparticularly in India [112]
Effective vaccines both live-attenuated and inactivatedstrains of JEV have been developed and licensed in majorcountries However as for the other arthropod-borne Fla-vivirus members no specific antiviral drugs are currentlyavailable [4]
281mAbs against JEV Aspreviously described for the otherFlavivirusmembers many groups isolated and characterizedanti-JEVmAbs showing different neutralizing and protectiveproperties In this regard Gupta et al used combinations ofanti-E JEV mAbs (Hs1-4) in mice protection experimentsIn particular they found that the singularly mAb protectionranged from 45 to 65 when 100120583g of mAb were adminis-tered while equimolar combinations of two or three mAbsgave 85ndash90 or 100 protection respectively [113] In similarexperiments Lee et al demonstrated that a Flavivirus anti-NS1 mAb named 16NS1 cross-reacted with JEV as well asWNV and exhibited protective activity against WNV as wellas a lethal JEV infection However no neutralizing activitywas observed using this mAb against both WNV and JEV inin vitro experiments suggesting the participation of otherAb-mediatedmechanisms in vivo In particular 95ofmicewere
protected when 500120583g of mAb were administered intraperi-toneally and concomitantly to intramuscular injection of JEV
Overlapping peptide mapping analysis combined withsite-specific mutations identified the 116KAWGKSILFA125region and critical amino acid residues W118 and I122 as16NS1 mAb epitope highly conserved in WNV and JEVstrains [114]
Arakawa et al isolated from a combinatorial human Fablibrary constructed from peripheral blood lymphocytesobtained from JEV hyperimmune volunteers Among 188randomly selected clones FabTJE12B02 showed the best 50focus reduction endpoint at the concentration of 502 120583gmLagainst the JEV strain Nakayama [55]
Goncalvez et al isolated three mAbs named Fabs A3B2 and E3 by repertoire cloning from chimpanzees initiallyimmunized with inactivated JE-VAX and then boosted withattenuated JEV SA14-14-2 In particular these mAbs reactedwith epitopes in three different E domains in DI (amino acidresidue K179) in DII (I126) and in DIII (G132) for Fabs A3B2 and E3 respectively Moreover these Fabs as well as thederived humanized counterpart mAbs exhibited high neu-tralizing activities against a broad spectrum of JEV genotypestrains Moreover these mAbs exhibited a 50 protectivedose of 084 120583g (B2) 58120583g (A3) and 247120583g (E3) in mousemodels Finally administration of 200 120583gmouse of mAb B2one day after otherwise lethal JEV infection protected 50of mice and significantly prolonged the average survival timecompared to that of mice in the unprotected group [54]
29 St Louis Encephalitis Virus (SLEV) St Louis encephalitisvirus (SLEV) was first discovered as the mosquito-borneagent responsible for over 1000 cases of encephalitis duringa 1933 summer outbreak in St Louis (Missouri) and nowis a reemerging human pathogen widely distributed in theAmerican continent causing several human encephalitisoutbreaks over the last 80 years Additional epidemics haveindeed occurred from 1964 to 2006 in the Americas rangingfrom the US to Argentina and Brazil As a member of theFlavivirus genus the SLEVE glycoprotein ectodomain is 68identical to serocomplex-related JEVEbut only 46 and 40identical to those ofDENV2 andTBEV viruses fromdifferentserocomplexes [115]
291 mAbs against SLEV In 1983 Roehrig et al isolatedtwenty-one hybridomas producing murine mAbs specificfor the E glycoprotein of SLEV strain MSI-7 Serologicreactivities were initially determined by cross-reactivity indi-rect immunofluorescence assays using 22 strains of SLEVand 8 other related flaviviruses Four groups demonstratingtype- subcomplex- supercomplex- and group-specific reac-tivity patterns were identified Analysis of hemagglutination-inhibition and virus neutralization subdivided the cross-reactivity groups into eight epitopes (E-1a b c d E-2 E-3 and E-4a b) one of them following localized in DII (4b)[52 53]
Moreover among the previously described isolated anti-WNV mAbs by Gould et al the chimeric scFv-Fc 79 effec-tively neutralized also SLEV resulting in gt80 of PRNT80when used at 5 120583gmL [42]
BioMed Research International 13
3 Alphaviruses
Major human arboviruses in the Togaviridae family Chikun-gunya (CHIKV) and Venezuelan equine encephalomyelitis(VEEV) viruses belong to the Alphavirus genus which iscomposed of viruses with an icosahedral nucleocapsid sur-rounded by a lipid envelope and glycoprotein spikes Thestructural proteins of alphaviruses arise through co- andpost-translational processing of a polyprotein encoded by asingle positive-stranded RNAproducing the capsid (C) PE26 K and E1 The PE2 glycoprotein is a precursor containingthe E3 glycoprotein fused to the amino terminus of the E2envelope glycoprotein The PE2 glycoprotein is followed by6K a small membrane-associated protein and E1 the secondpolypeptide component of glycoprotein spikes
Trimerized heterodimers of the E1 and E2 viral glyco-proteins form the surface spikes and contain determinantsof viral tropism and virulence The E3 glycoprotein acts as asignal for transport of PE2 across themembranes of the roughendoplasmic reticulum and may promote the formation andintracellular transport of E1-PE2 heterodimers to the cellsurface During transport to the cell surface PE2 undergoesa maturational cleavage event by a furin-like protease toproduce E2 and E3 [116]
The E2 glycoprotein promotes specificity of virus bindingto the host cell surface and is a target of the humoral immuneresponse The E1 glycoprotein mediates fusion of the virionenvelope with the membranes of acidified endosomes allow-ing release of the nucleocapsid into the cytoplasm and theonset of viral replicationAlphavirus E1 shares no appreciablesequence identity to Flavivirus E protein Despite differencesin their amino acid sequences and arrangements on the viralparticle the structures of E and E1 are remarkably similarIndeed the conservation of three domains has been welldocumented in crystal structures of flavivirus E ectodomainsfrom TBEV DENV JEV and WNV as well as Alphavirus E1from Semliki Forest virus (SFV) and CHIKV Abs to the E1glycoprotein do not typically neutralize virus infectivity invitro but can protect against lethal challenge in animals
By analogy with the prM protein of dengue Flavivirusfurin cleavage of PE2 may begin after transit of an acidiclate component of the Golgi body where E3 is thought tosuppress the acid pH-triggered activation of glycoprotein E1fusion capability Protective anti-E3 mouse mAbs have alsobeen described [117]
31 Chikungunya Virus (CHIKV) The chikungunya virus(CHIKV) belongs to the Semliki Forest clade and was firstlyisolated in 1953 in Tanzania during an epidemic outbreakfollowing occurred also in Asia and Africa In the last fivedecades several CHIKV outbreaks have been described bothin Africa and Asia separated by gaps lasting from two totwenty years In 2005-2006 about 300000 cases out of785000 inhabitants were reported in La Reunion island witha fatal prognosis for 237 of the patients [117] Neither Europenor the Americas have had outbreaks of CHIKV so far exceptfor imported isolated cases Three genotypes of CHIKV aredescribed Asian EastCentralSouth African (ECSA) andWest African with an amino acid identity spanning from
952 to 998 Recent epidemics in Africa and Indiansubcontinent were caused by strains belonging to the ECSAgenotype Transmitted by Aedes mosquitoes CHIKV ismaintained in the human population by human-mosquito-human transmission [1] The disease is characterized bydengue-like symptoms such as chills high fever headacheand persistent myalgia and a further incapacitating arthralgia(from which the name chikungunya) which affects 40 ofinfected subjects However prognosis is rarely fatal [116]
There is currently no commercial vaccine and antiviraltreatment forCHIKV although some candidate vaccines havebeen tested in humans In this regard in 2000 US Armyperformed a Phase II clinical trial testing a live-attenuatedCHIKV vaccine (TSI-GSD-218) derived from a 1962 strain(15561) of an outbreak in Thailand Out of 58 every patientdeveloped neutralizingAbs and 5 lamentedmild tomoderatejoint pain [117] A phase III trial of this candidate vaccineis ongoing Furthermore a new formulation using virus-likeparticles was able to induce neutralizing Abs in macaquesagainst different CHIKV strains [117 118] Indeed it has beendescribed that infection seems to elicit long-lasting protectiveimmunity and cross-protection among CHIKV and otheralphaviruses
CHIKV entry into host cell is demonstrated to be medi-ated by envelope glycoproteins E1 and E2 which allowvirus fusion to cell membrane in low pH conditions andrecognition of an unknown cellular receptor respectively[116 119ndash121] Even though the genome replication relies onan error-prone RNA-dependent RNA-polymerasis CHIKVstrains S27 and 05115 Reunion showed in E1 and E2 alow amino acid variation of 068 and 33 respectivelyand such a low variability may be needed for an effectivereplication in two phylogenetically distant hosts [116 122]Recently the crystal structure of E1 and E2 heterodimerhas been resolved both at low pH and natural conditionsshedding light on E1 and E2 ectodomainsrsquo structure Inparticular similarly to flaviviruses E protein E1 ectodomainis made of the N-terminal DI the DII containing the fusionloop and the DIII at the C-terminal On the other hand E2ectodomain comprises theN-terminal domainA the domainB supposed to interact with the host cellrsquos unknown receptorand domain C at the C-terminal E1 DIII and E2 domain Care located close to the viral membrane [123 124]
311 mAbs against CHIKV Warter et al described 5F10 and8B10 two human mAbs which strongly neutralized severalCHIKV isolates in vitro without cross-reactivity againstother alphaviruses but to Onyong-nyong virus [56] Mixedpreparation of 5F10 and 8F10 did not show neither synergisticnor addictive effect in vitro and studies upon escape mutantsdemonstrated that 5F10 mAb binds at the tip of the E2domain B while 8B10 recognizes residues close to E1 fusionloop and amino acids within E2 domain A which form atransitional epitope under low pH conditions The authorsspeculated that 5F10 and 8B10 may inhibit CHIKV entry andfusion to the cell membrane respectively It is worth notingthat the previously mentioned escape mutants displayedmutations associated with reduced viral fitness in vitro evenafter 13 neutralizationamplification rounds In the same
14 BioMed Research International
study cell-to-cell transmission was firstly demonstrated as anescapemechanism enhanced by the E2mutation R82G [122]
In further in vivo studies 5F10 and 8B10 significantlydelayed CHIKV-caused death of AGR129 mice both in pro-phylactic and therapeutic tests Interestingly in therapeutictreatment these mAbs showed a synergistic effect whenadministered in combination with the total amount of mAbsinjected being the same of the single-mAbs Possibly thesetreatment did not allow themice to survive CHIKV challengedue to Ig clearing and 6ndash10 days half-life usually described forhuman mAbs [125]
Recently Pal et al cloned thirty-six murine mAbs ableto neutralize the ECSA La Reunion 2006 OPY-1 strain ofCHIKV (CHIKV-LR) the majority of which also neutralizeinfection of other strains corresponding to the Asian andWest African genotypes In particularmAbCHK-152 showedthe highest and broadest neutralizing activity with FRNTvalues indicating an IC50 value of 1 to 3 ngmL dependingon the viral strain used Among the thirty-six mAbs four(CHK-102 CHK-152 and CHK-166 CHK-263) could pre-vent lethality of immunodeficient Ifnarminusminus C57BL6 micewhen administered one day before exposition with CHK-152 and CHK-263 showing the ability to protect mice at thelowest dose (10 120583g) Therapeutic studies were performed onthese mAbs administering a single dose of 100 120583g 24 hoursafter infection The highest activity was described for CHK-166 able to protect 63 of the mice Testing the combinedactivity of the mAbs in therapeutic studies administration ofCHK-166 plus CHK-152 in a dose of 250120583g each turned out tobe themost effective protecting 71 of the IfnarminusminusC57BL6mice 60 hours after infection [57]
32 Venezuelan Equine Encephalomyelitis Virus (VEEV)Venezuelan equine encephalomyelitis virus (VEEV) is main-tained in a natural transmission cycle between mosquitoesand small rodents The first documented outbreaks occurredin the 1930s and several epidemics have been reported so farin Latin-American countries such as Venezuela ColombiaPeru Ecuador Bolivia Honduras Mexico and Panama withhundreds of thousands of human cases being reported anda case-fatality rate up to 1 during the 1969 Ecuador out-break Clinical manifestations of VEE are indistinguishablefrom Dengue [126] Six serogroups (IndashVI) are currentlyrecognized within the VEEV complex VEEV caused humanand equine outbreaks in the Americas for nearly a centuryEquine epizootics have high mortality (38ndash83) leading toa high viremia followed by a lethal encephalitis and often tohuman epidemics involving thousands of cases and hundredsof deaths Indeed VEEV is infectious for humans by theairborne route and has been responsible for a number oflaboratory infections In humans the disease is usually self-limiting with a febrile illness with 1ndash4 of cases progressingto severe encephalitis It is a hazard to laboratory workershas been developed as a biological weapon and is a potentialbioterrorist agent [127] There are no antiviral drugs andvaccines licensed for the treatment of VEEV infection inhumans Indeed although experimental live-attenuated vac-cines have been developed (eg TC-83) with good levels ofprotection in equine and mice in humans they may fail to
give protection in the majority of cases [128] Consideringthe protective role exerted by humoral immune response inanimal models it could be concluded that treatment withspecific IgG may have a beneficial antiviral effect in humanairborne infections with pathogenic strains of VEEV
321 mAbs against VEEV Several works support that mAbsmay protect against airborne VEEV as well as after airborneexposure toVEEVAb reactivity to both surface glycoproteins(E1 and E2) is associated with neutralization of the virusin vitro and passive protection against virus challenge Aseries of anti-E1 and anti-E2 mouse and humanized mAbshave been described recently Among them Phillpotts et alexamined two anti-E2 murine mAbs named 1A4A-1 and1A3A-9 both having potent protective activity against subcu-taneous VEEV challenge in mice Both mAbs had a similarhalf-life (58 and 100 days) in mouse serum after a singleintraperitoneal dose suggesting that mAbs delivered by thisroute at or around the time of VEEV infection persists athigh levels in the blood as well as secretions in respiratorytransudation throughout the clinical course of the disease Inparticular both mAbs administered 24 h prior to airbornechallenges had a substantial protective effect (90ndash100)Treatment of mice 2 or 24 h after airborne infection with asingle intraperitoneal dose of 100 120583g of 1A3A-9 mAb led toapproximately 50 survival There was no beneficial effectwhen mAb treatment was delayed to 72 h Moreover therewas evidence of synergy in vitro in PRNT between 1A3A-9and 1A4A-1 as has been demonstrated for other virusesand mAb pairs However no synergy was found in mouseprotection when mAbs were delivered intraperitoneally as amixture in equal parts to animals challenged 24 h previouslywith VEEV [129]
In a subsequent study Phillpotts et al reported two otherprotective mAbs The 3B2A-9 mAb protected against allthe serogroup I strains while 1A3B-7 mAb protected wellagainst challenge with all of the viruses tested Both 3B2A-9 and 1A3B-7 protected against airborne exposure to theIAB serogroup virus strainTrinidad donkey (TrD)Howeverthere was no evidence of synergistic protection when thesemAbs were combined in equal proportions An intraperi-toneal dose of 10 120583g was sufficient to protect 50 of themice with either mAb [130] According to other data themechanism of protection did not appear to depend uponneutralization [131] Similar results were obtained by OrsquoBrienet al which described a non-neutralizing mAb (IgG2a)named CUF37-2a able to protect 50 of mice from asubcutaneous VEEV challenge when a dose of 915 120583g of mAbwas administered 24 h prior to challenge [132]
In a following study the 1A3B-7 mAb was humanized(and following reported as Hu1A3B-7) maintaining the samefeatures of its murine counterpart In particular evaluation ofin vitro studies indicated that Hu1A3B-7 retained both broadspecificity and neutralizing activity Furthermore in vivoexperiments showed that Hu1A3B-7 successfully protectedmice against lethal subcutaneous and aerosol challenges withVEEV strain TrD Moreover the effectiveness of the human-ization process was determined by assessing proliferationresponses in human T-cells to peptides derived from the
BioMed Research International 15
murine and humanized versions of the VH and VL domainsThis analysis showed that the number of human T-cellepitopes within the humanized Ab had been substantiallyreduced indicating that Hu1A3B-7 may have low immuno-genicity in vivo [133]
Similarly Hunt et al described a humanizedmurinemAbderived from the 3B4C-4 mAb recognizing the E2c epitope(amino acid residues 182ndash209) and following named Hy4-26C showing neutralizing activity similar to the murinemAb Moreover it was protective (70ndash100 of survival) at100 ng dose in mice animal model intraperitoneal inoculatedand at 500120583g for intranasal challenge (80) Therapeuticstudies in mice revealed that Hy4-26C was able to cure miceup to 24 h following infection at 10 120583g on the contrary themouse mAb when given 1 hour after virus challenge [63]
In another work Hunt et al isolated two neutralizinghumanized Fabs F5 and L1A7 employing a blocking strategy[134 135] The anti-E2 specific F5 IgG had a 70 PRNTendpoint of 10 ngmL equivalent to that described for themost effective neutralizing anti-VEEVE2mAbs In particularthe E1-specific hFab L1A7 had a PRNT endpoint of 3120583gmL300-fold lower than F5
Further studies revealed that F5 epitope is located inthe 115ndash199 amino acid region Moreover F5 IgG had potentability to protect mice from infection by either route whenadministered 24 h before exposure however mice treated24 h and 48 h after aerosol exposure developed central ner-vous system infections but exhibited no clinical signs ofdisease [135]
Hu et al reported that passive immunization with thehumanized chimericmousemAbhu1A4A1IgG1-2A inmice at50 120583g 24 h before or after virulent VEEV challenge providedcomplete protection indicating that hu1A4A1IgG1-2A haspotent prophylactic and therapeutic effects against VEEVinfection [136]
Anti-VEEV mAbs isolated from a non-human primategene library has also been reported In particular thehumanized chimeric scFv-Fc ToR67-3B4 recognized viableas well as formalin and 120573-propiolactone inactivated virusparticles It detected specifically the viral E1 envelope proteinof VEEV but did not react with reduced viral glycopro-tein preparations suggesting that recognition depends uponconformational epitopes The recombinant Ab was able todetect multiple VEEV subtypes and displayed only marginalcross-reactivity to otherAlphavirus species except for Easternequine encephalitis virus (EEEV) In addition the scFv-Fcfusion described here might be of therapeutic use since itsuccessfully inactivated VEEV in a murine disease modelIn particular when the recombinant Ab was administered 6hours after challenge 80 to 100 of mice survived lethalVEEV IAB or IE infection Forty to sixty percent of micesurvived when scFv-Fc ToR67-3B4 was applied 6 hours afterchallenge with VEEV subtypes II and former IIIA [137]
4 Bunyaviridae
TheBunyaviridae family contains human arboviruses belong-ing to the Orthobunyavirus Phlebovirus and Nairovirus
genera Bunyaviridae includes enveloped viruses with a frag-mented single-stranded RNA genome of negative polarityTheir tripartite genome consists of a small (S) a medium(M) and a large (L) fragment and the envelope glycoproteinsGn and Gc that are cleaved out of a polyprotein synthesizedby the M fragment These glycoproteins have been provedto mediate the formation of the virus particle to play a rolein the interaction with cell surface receptors to mediate theentry of the virus into cells and to serve as targets for themajority of neutralizing Abs described so far
Orthobunyavirus transmitted through mosquitoes ormidges vectors are divided in 18 serogroups based on cross-titrations in haemagglutination inhibition assays and neutral-ization assays and correlating with main vector preferencesHowever the most clinically important viruses belong onlyto two serogroups the California encephalitis and the Simbuserogroups
Crimean-Congo hemorrhagic fever virus (CCHFV) isconsidered the only one of clinical relevance within theNairovirus genus which uses tick as main vector and is divid-ed in seven serogroups while Toscana virus and Rift Valleyfever virus (RVFV) which are transmitted by sandflies andmosquitoes respectively belong to the Phlebovirus genus[138]
MAbs against theNairovirusCCHFV and the PhlebovirusRVFV have been described
41 Crimean-Congo Hemorrhagic Fever Virus (CCHFV)Crimean-Congo Hemorrhagic Fever Virus (CCHFV) dis-tribution covers the greatest geographic range of any tick-borne virus known as viral isolation andor disease hasbeen reported from more than 30 countries in SoutheasternEurope Africa Asia and Middle East [139 140] Namelyfrom 1953 to 2010 about 6000 human cases were reportedin Southeastern Europe CCHFV causes sporadic outbreakswith mortality rates ranging from 10 to 80 [141 142] Asignificant variation in the time of incubation has beendescribed [139] Prehemorrhagic symptoms include highfever chills headache photophobia and back and abdominalpains Among other symptoms neuropsychiatric changeshave been reported in some patients In severe cases 3ndash6 daysafter the onset of the disease hemorrhagic symptoms occurwith petechiae ecchymosis bleeding in the form of melenahematemesis epistaxis [139]
In comparisonwith the nucleocapsid proteins inCCHFVGn and Gc show a higher degree of antigenic variabilityprobably due to their exposition to the host immune systemhypothesis corroborated in the description by Hewson et alin 2004 of four M segment phylogenic groups namely M1M2 M3 and M4 [141 142]
411 mAbs against CCHFV Blackburn et al firstly describedin 1987 murine anti-CCHFV mAbs able to recognize nucle-ocapsid proteins but no neutralizing activity has beendescribed thus far [143]
On the other hand Bertolotti-Ciarlet et al produced apanel of murine mAbs recognizing conformational epitopes
16 BioMed Research International
within theGn andGc glycoproteins expressed by the CCHFVstrain IbAr10200 The effectiveness of the mAbs was studiedby performing PRNT80 and none of the anti-Gn mAbsshowed neutralizing activity although many of the anti-GcmAbs neutralized IbAr10200 strain in vitro with clones 8A15E3 12A9 6C2 and 9H3 showing activity at a gt5120 dilutionMAbs 11E7 and 30F7 exhibited neutralizing activity at 2560dilution A suckling mice protection test was thereforeperformed for these mAbs The previously mentionedanti-Gc Abs were capable to protect mice to an appreciabledegree when applied 24 hours before and in a weakermanner 24 hours after virus challenge Many of the anti-GnmAbs were able to confer significant protection to IbAr10200both 24 hours before and after virus administration Therelevant effectiveness of the anti-Gn mAbs 6B12 10E1113G8 and 10G4 suggests that these mAbs may possess someneutralizing activity due to Ab-based effector mechanisms(eg ADCC) [58]
In further studies the same group characterized thebroadly cross-reactive neutralizing activity of the previouslydescribed murine mAb 11E7 able to bind conformationalepitopes within the C-terminal region of the Gn glycoproteinof all CCHFVM groups [58 141]
42 Rift Valley Fever Virus (RVFV) Similarly to CCHFV RiftValley Fever Virus (RVFV) causes sporadic outbreaks Thelargest epidemic occurred in Egypt in 1977 with an estimated200000 infections 18000 patients manifesting symptomsand 600 fatal prognoses [144] Symptoms include hemor-rhagic episodes fever encephalitis and blindness [145]
The major target of the humoral immune response is theRVFV glycoproteins Gc (G1) and Gn (G2) ranging fromnucleotides 480ndash2090 and 2091ndash3614 respectively [144]
421 mAbs against RVFV Within G2 glycoprotein are des-cribed three neutralizing antigenic sites namely epitope Inucleotides 792ndash893 (amino acid residues 258ndash291) epitopeII nucleotides 1164ndash1196 (amino acid residues 382ndash392) andepitope IV nucleotides 858ndash917 (amino acid residues 280ndash299) [59 145]
Despite the high degree of variability observed inCCHFVglycoproteins RVFV G2 presented an unexpected high con-servation among 22 isolates [59] In the same study onlythe LUNYO and 900060 isolates were resistant to in vitroneutralization by murine mAb 4D4 (recognizing epitope II)All other isolates were neutralized with PRNT80 titers from10240 togt81920On the other hand only SNS isolate showedreduced sensitivity to 4ndash39-CC (epitope IV) All other isolateswere neutralized with PRNT80 titers from 20480 to gt81920Protection studies on mice would be of interest for thedevelopment of a passive immunization prophylaxis [59]
Protection studies from lethal South African RVFVAN1830 strain infection were successfully performed on abattery of anti-G1 and anti-G2 mAbs produced by Besselaarand Blackburn with the strongly neutralizing anti-G1 mAb3E5 and anti-G2mAb 9C4 being the most effective [146]The9C4 antigenic area maps within G2 epitope I and a broadlyneutralizing activity may be investigated
5 Conclusions
Arbovirus infections have acquired increasing interest giventhe augmented globalization and tourism movement all overthe world that could be at the basis of epidemic eventsFurthermore the same vectors can sometimes transmit sev-eral arboviruses concomitantly complicating the diagnosis aswell as the therapy For example confusing mixed epidemicshave occasionally been described such as YFV plus CHIKVDENVplus CHIKV ormore recentlyPlasmodium falciparummalaria plus DENV1 and CHIKV on Madagascarrsquos east coast[1]
In the last decade several efforts have been employed inthe development of effective immunogens and therapeuticsgiving the current absence of specific antiviral drugs as wellas effective vaccines for themost diffused arbovirus infections(ie DENV andWNV)
However at the moment mosquito control is the bestavailable method for preventing arbovirus infections Thusto control the emerging public health problem of arbovirusinfections new antiviral therapeutic strategies that providepotent and broadly cross-protective immunity (especially forflavivirus infections) are an urgent globally medical need
As described in this review the majority of highlyneutralizing mAbs are of murine origin and indeed theirutility in the treatment of these infections is limited In facttheir employment can elicit an immunogenic response (iehuman anti-mouse Abs HAMA) the therapeutic efficacycould be reduced by a relatively faster clearance in humans(compared to human Abs) potentially exacerbated by theHAMA response Finally murine Abs exhibit relatively weakeffector functions (eg ADCC) compared to human AbsIndeed chimerization humanization or better the isolationand characterization of fully human broadly neutralizingmAbs would be the best choice for their possible use in afuture mAb-based therapy against arbovirus infections [147]
Finally possible ADE of infection mechanisms should beevaluated before considering a mAb as a possible candidatetherapeutic in a post-exposure setting as well as in the devel-opment of new Ab-eliciting immunogens
Conflict of Interests
The authors declare no conflict of interests in particular withOmriGam and any other cited Ab-based preparation men-tioned in the paper
References
[1] N Cleton M Koopmans J Reimerink G J Godeke and CReusken ldquoCome fly with me review of clinically importantarboviruses for global travelersrdquo Journal of Clinical Virology vol55 no 3 pp 191ndash203 2012
[2] F X Heinz H Holzmann A Essl and M Kundi ldquoField effec-tiveness of vaccination against tick-borne encephalitisrdquoVaccinevol 25 no 43 pp 7559ndash7567 2007
[3] A D Barrett and D E Teuwen ldquoYellow fever vaccinemdashhowdoes it work and why do rare cases of serious adverse events
BioMed Research International 17
take placerdquo Current Opinion in Immunology vol 21 no 3 pp308ndash313 2009
[4] S B Halstead and S J Thomas ldquoNew vaccines for Japaneseencephalitisrdquo Current Infectious Disease Reports vol 12 no 3pp 174ndash180 2010
[5] R Burioni M Perotti N Mancini and M Clementi ldquoPer-spectives for the utilization of neutralizing human monoclonalantibodies as anti-HCV drugsrdquo Journal of Hepatology vol 49no 2 pp 299ndash300 2008
[6] R A Diotti G A Sautto L Solforosi NManciniM Clementiand R Burioni ldquoNeutralization activity and kinetics of twobroad-range human monoclonal IgG1 derived from recombi-nant Fab fragments and directed against Hepatitis C virus E2glycoproteinrdquo New Microbiologica vol 35 no 4 pp 475ndash4792012
[7] R Burioni F Canducci N Mancini et al ldquoMolecular cloningof the first humanmonoclona antibodies neutralizing with highpotency Swine-origin Influenza A pandemic virus (S-OIV)rdquoNew Microbiologica vol 32 no 4 pp 319ndash324 2009
[8] P K Chandra S Hazari B Poat et al ldquoIntracytoplasmic stableexpression of IgG1 antibody targeting NS3 helicase inhibitsreplication of highly efficient hepatitis C Virus 2a clonerdquoVirology Journal vol 7 article 118 2010
[9] N Clementi D de Marco N Mancini et al ldquoA human mono-clonal antibody with neutralizing activity against highly diver-gent influenza subtypesrdquo PLoS ONE vol 6 no 12 Article IDe28001 2011
[10] D de Marco N Clementi N Mancini et al ldquoA non-VH1-69 heterosubtypic neutralizing human monoclonal antibodyprotects mice against H1N1 and H5N1 virusesrdquo PLoS ONE vol7 no 4 Article ID e34415 2012
[11] L Solforosi N Mancini F Canducci et al ldquoA phage displayvector optimized for the generation of human antibody com-binatorial libraries and the molecular cloning of monoclonalantibody fragmentsrdquoNewMicrobiologica vol 35 no 3 pp 289ndash294 2012
[12] E V Koonin and V V Dolja ldquoEvolution and taxonomy ofpositive-strand RNA viruses implications of comparative anal-ysis of amino acid sequencesrdquo Critical Reviews in Biochemistryand Molecular Biology vol 28 no 5 pp 375ndash430 1993
[13] J S Mackenzie D J Gubler and L R Petersen ldquoEmergingflaviviruses the spread and resurgence of Japanese encephalitisWest Nile and dengue virusesrdquo Nature Medicine vol 10 no 12pp S98ndashS109 2004
[14] T J SmithW E Brandt J L Swanson J MMcCown and E LBuescher ldquoPhysical and biological properties of dengue-2 virusand associated antigensrdquo Journal of Virology vol 5 no 4 pp524ndash532 1970
[15] J M Smit B Moesker I Rodenhuis-Zybert and J WilschutldquoFlavivirus cell entry and membrane fusionrdquo Viruses vol 3 no2 pp 160ndash171 2011
[16] R J Kuhn W Zhang M G Rossmann et al ldquoStructure ofdengue virus implications for flavivirus organization matura-tion and fusionrdquo Cell vol 108 no 5 pp 717ndash725 2002
[17] K Stiasny J H Aberle M Keller B Grubeck-Loebenstein andF X Heinz ldquoAge affects quantity but not quality of antibodyresponses after vaccination with an inactivated flavivirus vac-cine against tick-borne encephalitisrdquo PLoS ONE vol 7 no 3Article ID e34145 2012
[18] LThomasOVerlaetenACabie et al ldquoInfluence of the dengueserotype previous dengue infection and plasma viral load
on clinical presentation and outcome during a dengue-2 anddengue-4 co-epidemicrdquo American Journal of Tropical Medicineand Hygiene vol 78 no 6 pp 990ndash998 2008
[19] Y Zhang J Corver P R Chipman et al ldquoStructures of imma-ture flavivirus particlesrdquo EMBO Journal vol 22 no 11 pp2604ndash2613 2003
[20] A K I Falconar ldquoIdentification of an epitope on the denguevirus membrane (M) protein defined by cross-protective mon-oclonal antibodies design of an improved epitope sequencebased on common determinants present in both envelope (EandM) proteinsrdquo Archives of Virology vol 144 no 12 pp 2313ndash2330 1999
[21] K-J Huang Y-C Yang Y-S Lin et al ldquoThe dual-specific bind-ing of dengue virus and target cells for the antibody-dependentenhancement of dengue virus infectionrdquo Journal of Immunol-ogy vol 176 no 5 pp 2825ndash2832 2006
[22] A K I Falconar ldquoAntibody responses are generated to immun-odominant ELKKLE-typemotifs on the nonstructural-1 glyco-protein during live dengue virus infections inmice and humansimplications for diagnosis pathogenesis and vaccine designrdquoClinical and Vaccine Immunology vol 14 no 5 pp 493ndash5042007
[23] C W Mandl F Guirakhoo H Holzmann F X Heinz and CKunz ldquoAntigenic structure of the flavivirus envelope protein Eat the molecular level using the tick-borne encephalitis virus asa modelrdquo Journal of Virology vol 63 no 2 pp 564ndash571 1989
[24] F A Rey F X Heinz C Mandl C Kunz and S C HarrisonldquoThe envelope glycoprotein from tick-borne encephalitis virusat 2 A resolutionrdquo Nature vol 375 no 6529 pp 291ndash298 1995
[25] S L Allison K Stiasny K Stadler CWMandl and F XHeinzldquoMapping of functional elements in the stem-anchor regionof tick-borne encephalitis virus envelope protein Erdquo Journal ofVirology vol 73 no 7 pp 5605ndash5612 1999
[26] D E Klein J L Choi and S C Harrison ldquoStructure of a denguevirus envelope protein late-stage fusion intermediaterdquo Journal ofVirology vol 87 no 4 pp 2287ndash2293 2013
[27] S L Hanna T C Pierson M D Sanchez A A Ahmed M MMurtadha and R W Doms ldquoN-linked glycosylation of WestNile virus envelope proteins influences particle assembly andinfectivityrdquo Journal of Virology vol 79 no 21 pp 13262ndash132742005
[28] N Mancini G Sautto N Clementi et al ldquoNeutralization inter-fering antibodies a ldquonovelrdquo example of humoral immune dys-function facilitating viral escaperdquoViruses vol 4 no 9 pp 1731ndash1752 2012
[29] Y Modis S Ogata D Clements and S C Harrison ldquoVariablesurface epitopes in the crystal structure of dengue virus type3 envelope glycoproteinrdquo Journal of Virology vol 79 no 2 pp1223ndash1231 2005
[30] N Miller ldquoRecent progress in dengue vaccine research anddevelopmentrdquo Current Opinion in Molecular Therapeutics vol12 no 1 pp 31ndash38 2010
[31] J H Chavez J R Silva A A Amarilla and L T Moraes Figu-eiredo ldquoDomain III peptides from flavivirus envelope proteinare useful antigens for serologic diagnosis and targets forimmunizationrdquo Biologicals vol 38 no 6 pp 613ndash618 2010
[32] B Shrestha J D Brien S Sukupolvi-Petty et al ldquoThe develop-ment of therapeutic antibodies that neutralize homologous andheterologous genotypes of dengue virus type 1rdquo PLoS Pathogensvol 6 no 4 Article ID e1000823 2010
18 BioMed Research International
[33] G D Gromowski N D Barrett and A D T Barrett ldquoChar-acterization of dengue virus complex-specific neutralizing epi-topes on envelope protein domain III of dengue 2 virusrdquo Journalof Virology vol 82 no 17 pp 8828ndash8837 2008
[34] S Sukupolvi-Petty S K Austin W E Purtha et al ldquoType- andsubcomplex-specific neutralizing antibodies against domain IIIof dengue virus type 2 envelope protein recognize adjacentepitopesrdquo Journal of Virology vol 81 no 23 pp 12816ndash128262007
[35] S Sukupolvi-Petty S K Austin M Engle et al ldquoStructure andfunction analysis of therapeutic monoclonal antibodies againstdengue virus type 2rdquo Journal of Virology vol 84 no 18 pp9227ndash9239 2010
[36] Y-Q Deng J-X Dai G-H Ji et al ldquoA broadly flaviviruscross-neutralizingmonoclonal antibody that recognizes a novelepitope within the fusion loop of e proteinrdquo PLoS ONE vol 6no 1 Article ID e16059 2011
[37] J M Costin E Zaitseva K M Kahle et al ldquoMechanistic studyof broadly neutralizing human monoclonal antibodies againstdengue virus that target the fusion looprdquo Journal of Virologyvol 87 no 1 pp 52ndash66 2013
[38] CMMidgley A FlanaganH B Tran et al ldquoStructural analysisof a dengue cross-reactive antibody complexed with envelopedomain III reveals the molecular basis of cross-reactivityrdquoJournal of Immunology vol 188 no 10 pp 4971ndash4979 2012
[39] N J Moreland P Susanto E Lim et al ldquoPhage display appro-aches for the isolation of monoclonal antibodies against denguevirus envelope domain III from human and mouse derivedlibrariesrdquo International Journal ofMolecular Sciences vol 13 no3 pp 2618ndash2635 2012
[40] J J B Cockburn M E Navarro Sanchez N Fretes et alldquoMechanism of dengue virus broad cross-neutralization by amonoclonal antibodyrdquo Structure vol 20 no 2 pp 303ndash3142012
[41] R Rajamanonmani C Nkenfou P Clancy et al ldquoOn a mousemonoclonal antibody that neutralizes all four dengue virusserotypesrdquo Journal of General Virology vol 90 no 4 pp 799ndash809 2009
[42] L H Gould J Sui H Foellmer et al ldquoProtective and herapeuticcapacity of human single-chain Fv-Fc fusion proteins againstWest Nile virusrdquo Journal of Virology vol 79 no 23 pp 14606ndash14613 2005
[43] H Sultana H G Foellmer G Neelakanta et al ldquoFusion LoopPeptide of the West Nile virus envelope protein is essential forpathogenesis and is recognized by a therapeutic cross-reactivehuman monoclonal antibodyrdquo Journal of Immunology vol 183no 1 pp 650ndash660 2009
[44] T Oliphant M Engle G E Nybakken et al ldquoDevelopment ofa humanized monoclonal antibody with therapeutic potentialagainstWest Nile virusrdquoNature Medicine vol 11 no 5 pp 522ndash530 2005
[45] G E Nybakken T Oliphant S Johnson S Burke M S Dia-mond and D H Fremont ldquoStructural basis of West Nile virusneutralization by a therapeutic antibodyrdquo Nature vol 437 no7059 pp 764ndash769 2005
[46] G D Gromowski J T Roehrig M S Diamond J C LeeT J Pitcher and A D T Barrett ldquoMutations of an antibodybinding energy hot spot on domain III of the dengue 2 envelopeglycoprotein exploited for neutralization escaperdquo Virology vol407 no 2 pp 237ndash246 2010
[47] J T Roehrig R A Bolin andR G Kelly ldquoMonoclonal antibodymapping of the envelope glycoprotein of the dengue 2 virusJamaicardquo Virology vol 246 no 2 pp 317ndash328 1998
[48] R de Alwis S A Smith N P Olivarez et al ldquoIdentification ofhumanneutralizing antibodies that bind to complex epitopes ondengue virionsrdquo Proceedings of the National Academy of Sciencesof the United States of America vol 109 no 19 pp 7439ndash74442012
[49] S K Austin K A Dowd B Shrestha et al ldquoStructural basis ofdifferential neutralization of DENV-1 genotypes by an antibodythat recognizes a cryptic epitoperdquo PLoS Pathogens vol 8 no 10Article ID e1002930 2012
[50] M Throsby C Geuijen J Goudsmit et al ldquoIsolation and char-acterization of human monoclonal antibodies from individualsinfected with West Nile Virusrdquo Journal of Virology vol 80 no14 pp 6982ndash6992 2006
[51] A P Goncalvez R H Purcell and C-J Lai ldquoEpitope deter-minants of a chimpanzee Fab antibody that efficiently cross-neutralizes dengue type 1 and type 2 viruses map to inside andin close proximity to fusion loop of the dengue type 2 virusenvelope glycoproteinrdquo Journal of Virology vol 78 no 23 pp12919ndash12928 2004
[52] J T Roehrig J H Mathews and D W Trent ldquoIdentification ofepitopes on the E glycoprotein of Saint Louis encephalitis virususing monoclonal antibodiesrdquo Virology vol 128 no 1 pp 118ndash126 1983
[53] J T Roehrig L A Staudinger A R Hunt J H Mathews andC D Blair ldquoAntibody prophylaxis and therapy for flavivirusencephalitis infectionsrdquo Annals of the New York Academy ofSciences vol 951 pp 286ndash297 2001
[54] A P Goncalvez C-H Chien K Tubthong et al ldquoHumanizedmonoclonal antibodies derived from chimpanzee fabs protectagainst Japanese encephalitis virus in vitro and in vivordquo Journalof Virology vol 82 no 14 pp 7009ndash7021 2008
[55] M Arakawa T Yamashiro G-I Uechi M Tadano and ANishizono ldquoConstruction of humanFab (1205741120581) library and iden-tification of human monoclonal Fab possessing neutralizingpotency against Japanese encephalitis virusrdquo Microbiology andImmunology vol 51 no 6 pp 617ndash625 2007
[56] L Warter C Y Lee R Thiagarajan et al ldquoChikungunya virusenvelope-specific human monoclonal antibodies with broadneutralization potencyrdquo Journal of Immunology vol 186 no 5pp 3258ndash3264 2011
[57] P Pal K A Dowd J D Brien et al ldquoDevelopment of a highlyprotective combination monoclonal antibody therapy againstchikungunya virusrdquo PLoS Pathogens vol 9 no 4 Article IDe1003312 2013
[58] A Bertolotti-Ciarlet J Smith K Strecker et al ldquoCellular local-ization and antigenic characterization of Crimean-Congo hem-orrhagic fever virus glycoproteinsrdquo Journal of Virology vol 79no 10 pp 6152ndash6161 2005
[59] J K Battles and J M Dalrymple ldquoGenetic variation amonggeographic isolates of Rift Valley fever virusrdquo American Journalof Tropical Medicine and Hygiene vol 39 no 6 pp 617ndash6311988
[60] B AThibodeaux N C Garbino NM Liss et al ldquoA humanizedIgG but not IgM antibody is effective in prophylaxis and therapyof yellow fever infection in an AG12917D-204 peripheralchallenge mouse modelrdquo Antiviral Research vol 94 no 1 pp1ndash8 2012
BioMed Research International 19
[61] S Daffis R E Kontermann J Korimbocus H Zeller H-DKlenk and J Ter Meulen ldquoAntibody responses against wild-type yellow fever virus and the 17D vaccine strain charac-terization with human monoclonal antibody fragments andneutralization escape variantsrdquoVirology vol 337 no 2 pp 262ndash272 2005
[62] L N Levanov L E Matveev E P Goncharova et al ldquoChimericantibodies against tick-borne encephalitis virusrdquo Vaccine vol28 no 32 pp 5265ndash5271 2010
[63] A R Hunt S Frederickson C Hinkel K S Bowdish and J TRoehrig ldquoA humanized murine monoclonal antibody protectsmice either before or after challenge with virulent Venezuelanequine encephalomyelitis virusrdquo Journal of General Virologyvol 87 no 9 pp 2467ndash2476 2006
[64] J M da Silva Voorham I A Rodenhuis-Zybert N V A Nunezet al ldquoAntibodies against the envelope glycoprotein promoteinfectivity of immature dengue virus serotype 2rdquo PLoS ONEvol 7 no 3 Article ID e29957 2012
[65] T C Pierson Q Xu S Nelson et al ldquoThe stoichiometry ofantibody-mediated neutralization and enhancement of westnile virus infectionrdquoCell Host andMicrobe vol 1 no 2 pp 135ndash145 2007
[66] G Sautto N Mancini R A Diotti L Solforosi M Clementiand R Burioni ldquoAnti-hepatitis C virus E2 (HCVE2) glyco-proteinmonoclonal antibodies andneutralization interferencerdquoAntiviral Research vol 96 no 1 pp 82ndash89 2012
[67] R Burioni Y Matsuura N Mancini et al ldquoDiverging effectsof human recombinant anti-hepatitis C virus (HCV) antibodyfragments derived from a single patient on the infectivityof a vesicular stomatitis virusHCV pseudotyperdquo Journal ofVirology vol 76 no 22 pp 11775ndash11779 2002
[68] R J Phillpotts J R Stephenson and J S Porterfield ldquoAntibody-dependent enhancement of tick-borne encephalitus virus infec-tivityrdquo Journal of General Virology vol 66 no 8 pp 1831ndash18371985
[69] K A Dowd and T C Pierson ldquoAntibody-mediated neutraliza-tion of flaviviruses a reductionist viewrdquo Virology vol 411 no 2pp 306ndash315 2011
[70] A K I Falconar ldquoMonoclonal antibodies that bind to commonepitopes on the dengue virus type 2 nonstructural-1 andenvelope glycoproteins display weak neutralizing activity anddifferentiated responses to virulent strains implications forpathogenesis and vaccinesrdquo Clinical and Vaccine Immunologyvol 15 no 3 pp 549ndash561 2008
[71] K Meyer A Banerjee S E Frey R B Belshe and R Ray ldquoAweak neutralizing antibody response to hepatitis C virus enve-lope glycoprotein enhances virus infectionrdquo PLoS ONE vol 6no 8 Article ID e23699 2011
[72] R Burioni R AWilliamson P P Sanna F E Bloom and D RBurton ldquoRecombinant human Fab to glycoprotein D neutral-izes infectivity and prevents cell-to-cell transmission of herpessimplex viruses 1 and 2 in vitrordquo Proceedings of the NationalAcademy of Sciences of the United States of America vol 91 no1 pp 355ndash359 1994
[73] M S Rolph A Zaid N E Rulli and S Mahalingam ldquoDown-regulation of interferon-120573 in antibody-dependent enhancementof dengue viral infections of human macrophages is dependenton interleukin-6rdquo Journal of Infectious Diseases vol 204 no 3pp 489ndash491 2011
[74] S S Whitehead J E Blaney A P Durbin and B R MurphyldquoProspects for a dengue virus vaccinerdquoNature ReviewsMicrobi-ology vol 5 no 7 pp 518ndash528 2007
[75] S-M Lok V Kostyuchenko G ENybakken et al ldquoBinding of aneutralizing antibody to dengue virus alters the arrangement ofsurface glycoproteinsrdquoNature Structural andMolecular Biologyvol 15 no 3 pp 312ndash317 2008
[76] A P Goncalvez R Men C Wernly R H Purcell and C-J Lai ldquoChimpanzee Fab fragments and a derived humanizedimmunogiobulin G1 antibody that efficiently cross-neutralizedengue type 1 and type 2 virusesrdquo Journal of Virology vol 78no 23 pp 12910ndash12918 2004
[77] WM P BWahala A A Kraus L B HaymoreM A Accavitti-Loper and A M de Silva ldquoDengue virus neutralization byhuman immune sera role of envelope protein domain III-reactive antibodyrdquo Virology vol 392 no 1 pp 103ndash113 2009
[78] W D Crill H R Hughes M J Delorey and G-J J ChangldquoHumoral immune responses of dengue fever patients usingepitope-specific serotype-2 virus-like particle antigensrdquo PLoSONE vol 4 no 4 Article ID e4991 2009
[79] C Setthapramote T Sasaki O Puiprom et al ldquoHuman mono-clonal antibodies to neutralize all dengue virus serotypes usinglymphocytes from patients at acute phase of the secondaryinfectionrdquo Biochemical and Biophysical Research Communica-tions vol 423 no 4 pp 867ndash872 2012
[80] M Beltramello K L Williams C P Simmons et al ldquoThehuman immune response to dengue virus is dominated byhighly cross-reactive antibodies endowed with neutralizing andenhancing activityrdquo Cell Host andMicrobe vol 8 no 3 pp 271ndash283 2010
[81] C M Midgley M Bajwa-Joseph S Vasanawathana et al ldquoAnin-depth analysis of original antigenic sin in dengue virusinfectionrdquo Journal of Virology vol 85 no 22 pp 410ndash421 2011
[82] B Kaufmann M R Vogt J Goudsmit et al ldquoNeutralizationof West Nile virus by cross-linking of its surface proteins withFab fragments of the human monoclonal antibody CR4354rdquoProceedings of the National Academy of Sciences of the UnitedStates of America vol 107 no 44 pp 18950ndash18955 2010
[83] B Spurrier J M Sampson M Totrov et al ldquoStructural analysisof human and macaque mAbs 2909 and 25B implicationsfor the configuration of the quaternary neutralizing epitope ofHIV-1 gp120rdquo Structure vol 19 no 5 pp 691ndash699 2011
[84] J J SchlesingerMWBrandriss andT PMonath ldquoMonoclonalantibodies distinguish between wild and vaccine strains ofyellow fever virus by neutralization hemagglutination inhibi-tion and immune precipitation of the virus envelope proteinrdquoVirology vol 125 no 1 pp 8ndash17 1983
[85] M W Brandriss J J Schlesinger E E Walsh and M BrisellildquoLethal 17D yellow fever encephalitis in mice I Passive protec-tion by monoclonal antibodies to the envelope proteins of 17Dyellow fever and dengue 2 virusesrdquo Journal of General Virologyvol 67 no 2 pp 229ndash234 1986
[86] H Cho and M S Diamond ldquoImmune responses to West Nilevirus infection in the central nervous systemrdquo Viruses vol 4no 12 pp 3812ndash3830 2012
[87] D W Beasley A D Barrett and R B Tesh ldquoResurgence ofWest Nile neurologic disease in the United States in 2012 whathappened What needs to be donerdquo Antiviral Research vol 99no 1 pp 1ndash5 2013
[88] M Throsby J Ter Meulen C Geuijen J Goudsmit and J DeKruif ldquoMapping and analysis of West Nile virus-specific mon-oclonal antibodies prospects for vaccine developmentrdquo ExpertReview of Vaccines vol 6 no 2 pp 183ndash191 2007
[89] M S Diamond T G Roberts D Edgil B Lu J Ernst and EHarris ldquoModulation of dengue virus infection in human cells
20 BioMed Research International
by alpha beta and gamma interferonsrdquo Journal of Virology vol74 no 11 pp 4957ndash4966 2000
[90] Z Shimoni M J Niven S Pitlick and S Bulvik ldquoTreatmentof West Nile virus encephalitis with intravenous immunoglob-ulinrdquo Emerging Infectious Diseases vol 7 no 4 article 759 2001
[91] AHamdan PGreen EMendelsonM R Kramer S Pitlik andM Weinberger ldquoPossible benefit of intravenous immunoglob-ulin therapy in a lung transplant recipient with West Nile virusencephalitisrdquoTransplant InfectiousDisease vol 4 no 3 pp 160ndash162 2002
[92] M Haley A S Retter D Fowler J Gea-Banacloche and NP OrsquoGrady ldquoThe role for intravenous immunoglobulin in thetreatment of West Nile virus encephalitisrdquo Clinical InfectiousDiseases vol 37 no 6 pp e88ndashe90 2003
[93] K M Chung G E Nybakken B S Thompson et al ldquoAnti-bodies against West Nile virus nonstructural protein NS1prevent lethal infection through Fc 120574 receptor-dependent and-independent mechanismsrdquo Journal of Virology vol 80 no 3pp 1340ndash1351 2006
[94] E A Henchal L S Henchal and J J Schlesinger ldquoSynergisticinteractions of anti-NS1 monoclonal antibodies protect pas-sively immunized mice from lethal challenge with dengue 2virusrdquo Journal of General Virology vol 69 no 8 pp 2101ndash21071988
[95] J T Roehrig ldquoAntigenic structure of flavivirus proteinsrdquoAdvances in Virus Research vol 59 pp 141ndash175 2003
[96] M D Sanchez T C Pierson D McAllister et al ldquoCharacteri-zation of neutralizing antibodies to West Nile virusrdquo Virologyvol 336 no 1 pp 70ndash82 2005
[97] J H Beigel J L Nordstrom S R Pillemer et al ldquoSafety andpharmacokinetics of single intravenous dose of MGAWN1 anovel monoclonal antibody to West Nile virusrdquo AntimicrobialAgents and Chemotherapy vol 54 no 6 pp 2431ndash2436 2010
[98] C A Smeraski V Siddharthan and J D Morrey ldquoTreatmentof spatial memory impairment in hamsters infected with WestNile virus using a humanizedmonoclonal antibodyMGAWN1rdquoAntiviral Research vol 91 no 1 pp 43ndash49 2011
[99] D Lelli AMoreno E Brocchi et al ldquoWestNile virus character-ization and diagnostic applications of monoclonal antibodiesrdquoVirology Journal vol 9 article 81 2012
[100] T S Gritsun V A Lashkevich and E A Gould ldquoTick-borneencephalitisrdquo Antiviral Research vol 57 no 1-2 pp 129ndash1462003
[101] D Ruzek T S Gritsun N L Forrester et al ldquoMutations inthe NS2B and NS3 genes affect mouse neuroinvasiveness of aWestern European field strain of tick-borne encephalitis virusrdquoVirology vol 374 no 2 pp 249ndash255 2008
[102] M Ecker S L Allison T Meixner and F X Heinz ldquoSequenceanalysis and genetic classification of tick-borne encephalitisviruses from Europe and Asiardquo Journal of General Virology vol80 no 1 pp 179ndash185 1999
[103] H Holzmann M S Vorobyova I P Ladyzhenskaya et alldquoMolecular epidemiology of tick-borne encephalitis viruscross-protection between European and Far Eastern subtypesrdquoVaccine vol 10 no 5 pp 345ndash349 1992
[104] C W Mandl ldquoSteps of the tick-borne encephalitis virus repli-cation cycle that affect neuropathogenesisrdquo Virus Research vol111 no 2 pp 161ndash174 2005
[105] A V Atrasheuskaya T M Fredeking and G M IgnatyevldquoChanges in immune parameters and their correction in humancases of tick-borne encephalitisrdquo Clinical and ExperimentalImmunology vol 131 no 1 pp 148ndash154 2003
[106] K L Mansfield N Johnson L P Phipps J R Stephenson AR Fooks and T Solomon ldquoTick-borne encephalitis virusmdashareview of an emerging zoonosisrdquo Journal of General Virologyvol 90 no 8 pp 1781ndash1794 2009
[107] DWVaughn andCHHoke Jr ldquoThe epidemiology of Japaneseencephalitis prospects for preventionrdquo Epidemiologic Reviewsvol 14 pp 197ndash221 1992
[108] T Solomon H Ni D W C Beasley M Ekkelenkamp M JCardosa and A D T Barrett ldquoOrigin and evolution of Japaneseencephalitis virus in Southeast Asiardquo Journal of Virology vol 77no 5 pp 3091ndash3098 2003
[109] M R Wilson ldquoEmerging viral infectionsrdquo Current Opinion inNeurology vol 26 no 3 pp 301ndash306 2013
[110] J N Hanna S A Ritchie D A Phillips et al ldquoAn outbreakof Japanese encephalitis in the Torres Strait Australia 1995rdquoMedical Journal of Australia vol 165 no 5 pp 256ndash260 1996
[111] J N Hanna S A Ritchie D A Phillips et al ldquoJapanese ence-phalitis in north Queensland Australia 1998rdquo Medical Journalof Australia vol 170 no 11 pp 533ndash536 1999
[112] SGeorgeMGourie-Devi and J A Rao ldquoIsolation ofWestNilevirus from the brains of children who had died of encephalitisrdquoBulletin of theWorldHealthOrganization vol 62 no 6 pp 879ndash882 1984
[113] A K Gupta A A Koshy and V J Lad ldquoEnhanced protec-tion of mice against Japanese encephalitis virus infection bycombinations ofmonoclonal antibodies to glycoprotein ErdquoActaVirologica vol 55 no 2 pp 165ndash168 2011
[114] T H Lee B-H Song S-I Yun et al ldquoA cross-protective mAbrecognizes a novel epitope within the flavivirus NS1 proteinrdquoJournal of General Virology vol 93 no 1 pp 20ndash26 2012
[115] S C Weaver ldquoHost range amplification and arboviral diseaseemergencerdquo Archives of Virology Supplementum no 19 pp 33ndash44 2005
[116] M Solignat B Gay S Higgs L Briant and C Devaux ldquoRepli-cation cycle of chikungunya a re-emerging arbovirusrdquoVirologyvol 393 no 2 pp 183ndash197 2009
[117] O Schwartz and M L Albert ldquoBiology and pathogenesis ofchikungunya virusrdquo Nature Reviews Microbiology vol 8 no 7pp 491ndash500 2010
[118] W Akahata Z-Y Yang H Andersen et al ldquoA virus-like particlevaccine for epidemic Chikungunya virus protects nonhumanprimates against infectionrdquo Nature Medicine vol 16 no 3 pp334ndash338 2010
[119] M Kielian and A Helenius ldquopH-Iinduced alterations in thefusogenic spike protein of Semliki Forest virusrdquo Journal of CellBiology vol 101 no 6 pp 2284ndash2291 1985
[120] J White and A Helenius ldquopH-Dependent fusion between theSemliki Forest virus membrane and liposomesrdquo Proceedings ofthe National Academy of Sciences of the United States of Americavol 77 no 6 pp 3273ndash3277 1980
[121] J Dubuisson and C M Rice ldquoSindbis virus attachment isola-tion and characterization of mutants with impaired binding tovertebrate cellsrdquo Journal of Virology vol 67 no 6 pp 3363ndash33741993
[122] C Y Lee Y-W Kam J Fric et al ldquoChikungunya virus neutral-ization antigens and direct cell-to-cell transmission are revealedby human antibody-escapemutantsrdquo PLoS Pathogens vol 7 no12 Article ID e1002390 2011
[123] J E Voss M-C Vaney S Duquerroy et al ldquoGlycoproteinorganization of Chikungunya virus particles revealed by X-raycrystallographyrdquo Nature vol 468 no 7324 pp 709ndash712 2010
BioMed Research International 21
[124] L Li J Jose Y Xiang R J Kuhn and M G Rossmann ldquoStruc-tural changes of envelope proteins during alphavirus fusionrdquoNature vol 468 no 7324 pp 705ndash708 2010
[125] J Fric S Bertin-Maghit C I Wang A Nardin and L WarterldquoUse of human monoclonal antibodies to treat Chikungunyavirus infectionrdquo Journal of Infectious Diseases vol 207 no 2 pp319ndash322 2013
[126] P V Aguilar J G Estrada-Franco R Navarro-Lopez C FerroA D Haddow and S C Weaver ldquoEndemic Venezuelanequine encephalitis in the Americas hidden under the dengueumbrellardquo Future Virology vol 6 no 6 pp 721ndash740 2011
[127] S C Weaver R Winegar I D Manger and N L ForresterldquoAlphaviruses population genetics and determinants of emer-gencerdquo Antiviral Research vol 94 no 3 pp 242ndash257 2012
[128] S Paessler and S C Weaver ldquoVaccines for Venezuelan equineencephalitisrdquo Vaccine vol 27 no 4 pp D80ndashD85 2009
[129] R J Phillpotts ldquoVenezuelan equine encephalitis virus complex-specific monoclonal antibody provides broad protection inmurine models against airborne challenge with viruses fromserogroups I II and IIIrdquoVirus Research vol 120 no 1-2 pp 107ndash112 2006
[130] R J Phillpotts L D Jones and S C Howard ldquoMonoclonalantibody protectsmice against infection anddiseasewhen giveneither before or up to 24 h after airborne challenge with virulentVenezuelan equine encephalitis virusrdquoVaccine vol 20 no 11-12pp 1497ndash1504 2002
[131] J H Mathews J T Roehrig and D W Trent ldquoRole of com-plement and the Fc portion of immunoglobulin G in immunityto Venezuelan equine encephalomyelitis virus infection withglycoprotein-specific monoclonal antibodiesrdquo Journal of Virol-ogy vol 55 no 3 pp 594ndash600 1985
[132] L M OrsquoBrien C D Underwood-Fowler S A Goodchild AL Phelps and R J Phillpotts ldquoDevelopment of a novel mon-oclonal antibody with reactivity to a wide range of Venezuelanequine encephalitis virus strainsrdquoVirology Journal vol 6 article206 2009
[133] L M OrsquoBrien S A Goodchild R J Phillpotts and S D Per-kins ldquoA humanised murine monoclonal antibody protectsmice from Venezuelan equine encephalitis virus Evergladesvirus and Mucambo virus when administered up to 48h afterairborne challengerdquo Virology vol 426 no 2 pp 100ndash105 2012
[134] N Clementi N Mancini L Solforosi M Castelli M Clementiand R Burioni ldquoPhage display-based strategies for cloning andoptimization of monoclonal antibodies directed against humanpathogensrdquo International Journal of Molecular Sciences vol 13no 7 pp 8273ndash8292 2012
[135] A R Hunt R A Bowen S Frederickson T Maruyama JT Roehrig and C D Blair ldquoTreatment of mice with humanmonoclonal antibody 24h after lethal aerosol challenge withvirulent Venezuelan equine encephalitis virus prevents diseasebut not infectionrdquo Virology vol 414 no 2 pp 146ndash152 2011
[136] W-G Hu A L Phelps S Jager et al ldquoA recombinant human-ized monoclonal antibody completely protects mice againstlethal challenge with Venezuelan equine encephalitis virusrdquoVaccine vol 28 no 34 pp 5558ndash5564 2010
[137] T Rulker L Voss P Thullier et al ldquoIsolation and characterisa-tion of a human-like antibody fragment (scFv) that inactivatesVEEV in vitro and in vivordquo PLoS ONE vol 7 no 5 Article IDe37242 2012
[138] C T Walter and J N Barr ldquoRecent advances in the molecularand cellular biology of bunyavirusesrdquo Journal of General Virol-ogy vol 92 no 11 pp 2467ndash2484 2011
[139] C A Whitehouse ldquoCrimean-Congo hemorrhagic feverrdquo Anti-viral Research vol 64 no 3 pp 145ndash160 2004
[140] M Mertens K Schmidt A Ozkul and M H Groschup ldquoTheimpact of Crimean-Congo hemorrhagic fever virus on publichealthrdquo Antiviral Research vol 98 no 2 pp 248ndash260 2013
[141] A A Ahmed J M McFalls C Hoffmann et al ldquoPresence ofbroadly reactive and group-specific neutralizing epitopes onnewly described isolates of Crimean-Congo hemorrhagic fevervirusrdquo Journal of General Virology vol 86 no 12 pp 3327ndash33362005
[142] R Hewson A Gmyl L Gmyl et al ldquoEvidence of segment reas-sortment in Crimean-Congo haemorrhagic fever virusrdquo Journalof General Virology vol 85 no 10 pp 3059ndash3070 2004
[143] N K Blackburn T G Besselaar A J Shepherd and R Swane-poel ldquoPreparation and use of monoclonal antibodies for iden-tifying Crimean-Congo hemorrhagic fever virusrdquo AmericanJournal of Tropical Medicine and Hygiene vol 37 no 2 pp 392ndash397 1987
[144] T Ikegami ldquoMolecular biology and genetic diversity of RiftValley fever virusrdquoAntiviral Research vol 95 no 3 pp 293ndash3102012
[145] KKeegan andM S Collett ldquoUse of bacterial expression cloningto define the amino acid sequences of antigenic determinantson the G2 glycoprotein of Rift Valley fever virusrdquo Journal ofVirology vol 58 no 2 pp 263ndash270 1986
[146] T G Besselaar and N K Blackburn ldquoTopological mapping ofantigenic sites on the Rift Valley fever virus envelope glycopro-teins using monoclonal antibodiesrdquo Archives of Virology vol121 no 1ndash4 pp 111ndash124 1991
[147] L G Presta ldquoEngineering of therapeutic antibodies to mini-mize immunogenicity and optimize functionrdquo Advanced DrugDelivery Reviews vol 58 no 5-6 pp 640ndash656 2006
Submit your manuscripts athttpwwwhindawicom
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anatomy Research International
PeptidesInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporation httpwwwhindawicom
International Journal of
Volume 2014
Zoology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Molecular Biology International
GenomicsInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioinformaticsAdvances in
Marine BiologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Signal TransductionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Evolutionary BiologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Biochemistry Research International
ArchaeaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Genetics Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in
Virolog y
Hindawi Publishing Corporationhttpwwwhindawicom
Nucleic AcidsJournal of
Volume 2014
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Enzyme Research
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Microbiology

BioMed Research International 13
3 Alphaviruses
Major human arboviruses in the Togaviridae family Chikun-gunya (CHIKV) and Venezuelan equine encephalomyelitis(VEEV) viruses belong to the Alphavirus genus which iscomposed of viruses with an icosahedral nucleocapsid sur-rounded by a lipid envelope and glycoprotein spikes Thestructural proteins of alphaviruses arise through co- andpost-translational processing of a polyprotein encoded by asingle positive-stranded RNAproducing the capsid (C) PE26 K and E1 The PE2 glycoprotein is a precursor containingthe E3 glycoprotein fused to the amino terminus of the E2envelope glycoprotein The PE2 glycoprotein is followed by6K a small membrane-associated protein and E1 the secondpolypeptide component of glycoprotein spikes
Trimerized heterodimers of the E1 and E2 viral glyco-proteins form the surface spikes and contain determinantsof viral tropism and virulence The E3 glycoprotein acts as asignal for transport of PE2 across themembranes of the roughendoplasmic reticulum and may promote the formation andintracellular transport of E1-PE2 heterodimers to the cellsurface During transport to the cell surface PE2 undergoesa maturational cleavage event by a furin-like protease toproduce E2 and E3 [116]
The E2 glycoprotein promotes specificity of virus bindingto the host cell surface and is a target of the humoral immuneresponse The E1 glycoprotein mediates fusion of the virionenvelope with the membranes of acidified endosomes allow-ing release of the nucleocapsid into the cytoplasm and theonset of viral replicationAlphavirus E1 shares no appreciablesequence identity to Flavivirus E protein Despite differencesin their amino acid sequences and arrangements on the viralparticle the structures of E and E1 are remarkably similarIndeed the conservation of three domains has been welldocumented in crystal structures of flavivirus E ectodomainsfrom TBEV DENV JEV and WNV as well as Alphavirus E1from Semliki Forest virus (SFV) and CHIKV Abs to the E1glycoprotein do not typically neutralize virus infectivity invitro but can protect against lethal challenge in animals
By analogy with the prM protein of dengue Flavivirusfurin cleavage of PE2 may begin after transit of an acidiclate component of the Golgi body where E3 is thought tosuppress the acid pH-triggered activation of glycoprotein E1fusion capability Protective anti-E3 mouse mAbs have alsobeen described [117]
31 Chikungunya Virus (CHIKV) The chikungunya virus(CHIKV) belongs to the Semliki Forest clade and was firstlyisolated in 1953 in Tanzania during an epidemic outbreakfollowing occurred also in Asia and Africa In the last fivedecades several CHIKV outbreaks have been described bothin Africa and Asia separated by gaps lasting from two totwenty years In 2005-2006 about 300000 cases out of785000 inhabitants were reported in La Reunion island witha fatal prognosis for 237 of the patients [117] Neither Europenor the Americas have had outbreaks of CHIKV so far exceptfor imported isolated cases Three genotypes of CHIKV aredescribed Asian EastCentralSouth African (ECSA) andWest African with an amino acid identity spanning from
952 to 998 Recent epidemics in Africa and Indiansubcontinent were caused by strains belonging to the ECSAgenotype Transmitted by Aedes mosquitoes CHIKV ismaintained in the human population by human-mosquito-human transmission [1] The disease is characterized bydengue-like symptoms such as chills high fever headacheand persistent myalgia and a further incapacitating arthralgia(from which the name chikungunya) which affects 40 ofinfected subjects However prognosis is rarely fatal [116]
There is currently no commercial vaccine and antiviraltreatment forCHIKV although some candidate vaccines havebeen tested in humans In this regard in 2000 US Armyperformed a Phase II clinical trial testing a live-attenuatedCHIKV vaccine (TSI-GSD-218) derived from a 1962 strain(15561) of an outbreak in Thailand Out of 58 every patientdeveloped neutralizingAbs and 5 lamentedmild tomoderatejoint pain [117] A phase III trial of this candidate vaccineis ongoing Furthermore a new formulation using virus-likeparticles was able to induce neutralizing Abs in macaquesagainst different CHIKV strains [117 118] Indeed it has beendescribed that infection seems to elicit long-lasting protectiveimmunity and cross-protection among CHIKV and otheralphaviruses
CHIKV entry into host cell is demonstrated to be medi-ated by envelope glycoproteins E1 and E2 which allowvirus fusion to cell membrane in low pH conditions andrecognition of an unknown cellular receptor respectively[116 119ndash121] Even though the genome replication relies onan error-prone RNA-dependent RNA-polymerasis CHIKVstrains S27 and 05115 Reunion showed in E1 and E2 alow amino acid variation of 068 and 33 respectivelyand such a low variability may be needed for an effectivereplication in two phylogenetically distant hosts [116 122]Recently the crystal structure of E1 and E2 heterodimerhas been resolved both at low pH and natural conditionsshedding light on E1 and E2 ectodomainsrsquo structure Inparticular similarly to flaviviruses E protein E1 ectodomainis made of the N-terminal DI the DII containing the fusionloop and the DIII at the C-terminal On the other hand E2ectodomain comprises theN-terminal domainA the domainB supposed to interact with the host cellrsquos unknown receptorand domain C at the C-terminal E1 DIII and E2 domain Care located close to the viral membrane [123 124]
311 mAbs against CHIKV Warter et al described 5F10 and8B10 two human mAbs which strongly neutralized severalCHIKV isolates in vitro without cross-reactivity againstother alphaviruses but to Onyong-nyong virus [56] Mixedpreparation of 5F10 and 8F10 did not show neither synergisticnor addictive effect in vitro and studies upon escape mutantsdemonstrated that 5F10 mAb binds at the tip of the E2domain B while 8B10 recognizes residues close to E1 fusionloop and amino acids within E2 domain A which form atransitional epitope under low pH conditions The authorsspeculated that 5F10 and 8B10 may inhibit CHIKV entry andfusion to the cell membrane respectively It is worth notingthat the previously mentioned escape mutants displayedmutations associated with reduced viral fitness in vitro evenafter 13 neutralizationamplification rounds In the same
14 BioMed Research International
study cell-to-cell transmission was firstly demonstrated as anescapemechanism enhanced by the E2mutation R82G [122]
In further in vivo studies 5F10 and 8B10 significantlydelayed CHIKV-caused death of AGR129 mice both in pro-phylactic and therapeutic tests Interestingly in therapeutictreatment these mAbs showed a synergistic effect whenadministered in combination with the total amount of mAbsinjected being the same of the single-mAbs Possibly thesetreatment did not allow themice to survive CHIKV challengedue to Ig clearing and 6ndash10 days half-life usually described forhuman mAbs [125]
Recently Pal et al cloned thirty-six murine mAbs ableto neutralize the ECSA La Reunion 2006 OPY-1 strain ofCHIKV (CHIKV-LR) the majority of which also neutralizeinfection of other strains corresponding to the Asian andWest African genotypes In particularmAbCHK-152 showedthe highest and broadest neutralizing activity with FRNTvalues indicating an IC50 value of 1 to 3 ngmL dependingon the viral strain used Among the thirty-six mAbs four(CHK-102 CHK-152 and CHK-166 CHK-263) could pre-vent lethality of immunodeficient Ifnarminusminus C57BL6 micewhen administered one day before exposition with CHK-152 and CHK-263 showing the ability to protect mice at thelowest dose (10 120583g) Therapeutic studies were performed onthese mAbs administering a single dose of 100 120583g 24 hoursafter infection The highest activity was described for CHK-166 able to protect 63 of the mice Testing the combinedactivity of the mAbs in therapeutic studies administration ofCHK-166 plus CHK-152 in a dose of 250120583g each turned out tobe themost effective protecting 71 of the IfnarminusminusC57BL6mice 60 hours after infection [57]
32 Venezuelan Equine Encephalomyelitis Virus (VEEV)Venezuelan equine encephalomyelitis virus (VEEV) is main-tained in a natural transmission cycle between mosquitoesand small rodents The first documented outbreaks occurredin the 1930s and several epidemics have been reported so farin Latin-American countries such as Venezuela ColombiaPeru Ecuador Bolivia Honduras Mexico and Panama withhundreds of thousands of human cases being reported anda case-fatality rate up to 1 during the 1969 Ecuador out-break Clinical manifestations of VEE are indistinguishablefrom Dengue [126] Six serogroups (IndashVI) are currentlyrecognized within the VEEV complex VEEV caused humanand equine outbreaks in the Americas for nearly a centuryEquine epizootics have high mortality (38ndash83) leading toa high viremia followed by a lethal encephalitis and often tohuman epidemics involving thousands of cases and hundredsof deaths Indeed VEEV is infectious for humans by theairborne route and has been responsible for a number oflaboratory infections In humans the disease is usually self-limiting with a febrile illness with 1ndash4 of cases progressingto severe encephalitis It is a hazard to laboratory workershas been developed as a biological weapon and is a potentialbioterrorist agent [127] There are no antiviral drugs andvaccines licensed for the treatment of VEEV infection inhumans Indeed although experimental live-attenuated vac-cines have been developed (eg TC-83) with good levels ofprotection in equine and mice in humans they may fail to
give protection in the majority of cases [128] Consideringthe protective role exerted by humoral immune response inanimal models it could be concluded that treatment withspecific IgG may have a beneficial antiviral effect in humanairborne infections with pathogenic strains of VEEV
321 mAbs against VEEV Several works support that mAbsmay protect against airborne VEEV as well as after airborneexposure toVEEVAb reactivity to both surface glycoproteins(E1 and E2) is associated with neutralization of the virusin vitro and passive protection against virus challenge Aseries of anti-E1 and anti-E2 mouse and humanized mAbshave been described recently Among them Phillpotts et alexamined two anti-E2 murine mAbs named 1A4A-1 and1A3A-9 both having potent protective activity against subcu-taneous VEEV challenge in mice Both mAbs had a similarhalf-life (58 and 100 days) in mouse serum after a singleintraperitoneal dose suggesting that mAbs delivered by thisroute at or around the time of VEEV infection persists athigh levels in the blood as well as secretions in respiratorytransudation throughout the clinical course of the disease Inparticular both mAbs administered 24 h prior to airbornechallenges had a substantial protective effect (90ndash100)Treatment of mice 2 or 24 h after airborne infection with asingle intraperitoneal dose of 100 120583g of 1A3A-9 mAb led toapproximately 50 survival There was no beneficial effectwhen mAb treatment was delayed to 72 h Moreover therewas evidence of synergy in vitro in PRNT between 1A3A-9and 1A4A-1 as has been demonstrated for other virusesand mAb pairs However no synergy was found in mouseprotection when mAbs were delivered intraperitoneally as amixture in equal parts to animals challenged 24 h previouslywith VEEV [129]
In a subsequent study Phillpotts et al reported two otherprotective mAbs The 3B2A-9 mAb protected against allthe serogroup I strains while 1A3B-7 mAb protected wellagainst challenge with all of the viruses tested Both 3B2A-9 and 1A3B-7 protected against airborne exposure to theIAB serogroup virus strainTrinidad donkey (TrD)Howeverthere was no evidence of synergistic protection when thesemAbs were combined in equal proportions An intraperi-toneal dose of 10 120583g was sufficient to protect 50 of themice with either mAb [130] According to other data themechanism of protection did not appear to depend uponneutralization [131] Similar results were obtained by OrsquoBrienet al which described a non-neutralizing mAb (IgG2a)named CUF37-2a able to protect 50 of mice from asubcutaneous VEEV challenge when a dose of 915 120583g of mAbwas administered 24 h prior to challenge [132]
In a following study the 1A3B-7 mAb was humanized(and following reported as Hu1A3B-7) maintaining the samefeatures of its murine counterpart In particular evaluation ofin vitro studies indicated that Hu1A3B-7 retained both broadspecificity and neutralizing activity Furthermore in vivoexperiments showed that Hu1A3B-7 successfully protectedmice against lethal subcutaneous and aerosol challenges withVEEV strain TrD Moreover the effectiveness of the human-ization process was determined by assessing proliferationresponses in human T-cells to peptides derived from the
BioMed Research International 15
murine and humanized versions of the VH and VL domainsThis analysis showed that the number of human T-cellepitopes within the humanized Ab had been substantiallyreduced indicating that Hu1A3B-7 may have low immuno-genicity in vivo [133]
Similarly Hunt et al described a humanizedmurinemAbderived from the 3B4C-4 mAb recognizing the E2c epitope(amino acid residues 182ndash209) and following named Hy4-26C showing neutralizing activity similar to the murinemAb Moreover it was protective (70ndash100 of survival) at100 ng dose in mice animal model intraperitoneal inoculatedand at 500120583g for intranasal challenge (80) Therapeuticstudies in mice revealed that Hy4-26C was able to cure miceup to 24 h following infection at 10 120583g on the contrary themouse mAb when given 1 hour after virus challenge [63]
In another work Hunt et al isolated two neutralizinghumanized Fabs F5 and L1A7 employing a blocking strategy[134 135] The anti-E2 specific F5 IgG had a 70 PRNTendpoint of 10 ngmL equivalent to that described for themost effective neutralizing anti-VEEVE2mAbs In particularthe E1-specific hFab L1A7 had a PRNT endpoint of 3120583gmL300-fold lower than F5
Further studies revealed that F5 epitope is located inthe 115ndash199 amino acid region Moreover F5 IgG had potentability to protect mice from infection by either route whenadministered 24 h before exposure however mice treated24 h and 48 h after aerosol exposure developed central ner-vous system infections but exhibited no clinical signs ofdisease [135]
Hu et al reported that passive immunization with thehumanized chimericmousemAbhu1A4A1IgG1-2A inmice at50 120583g 24 h before or after virulent VEEV challenge providedcomplete protection indicating that hu1A4A1IgG1-2A haspotent prophylactic and therapeutic effects against VEEVinfection [136]
Anti-VEEV mAbs isolated from a non-human primategene library has also been reported In particular thehumanized chimeric scFv-Fc ToR67-3B4 recognized viableas well as formalin and 120573-propiolactone inactivated virusparticles It detected specifically the viral E1 envelope proteinof VEEV but did not react with reduced viral glycopro-tein preparations suggesting that recognition depends uponconformational epitopes The recombinant Ab was able todetect multiple VEEV subtypes and displayed only marginalcross-reactivity to otherAlphavirus species except for Easternequine encephalitis virus (EEEV) In addition the scFv-Fcfusion described here might be of therapeutic use since itsuccessfully inactivated VEEV in a murine disease modelIn particular when the recombinant Ab was administered 6hours after challenge 80 to 100 of mice survived lethalVEEV IAB or IE infection Forty to sixty percent of micesurvived when scFv-Fc ToR67-3B4 was applied 6 hours afterchallenge with VEEV subtypes II and former IIIA [137]
4 Bunyaviridae
TheBunyaviridae family contains human arboviruses belong-ing to the Orthobunyavirus Phlebovirus and Nairovirus
genera Bunyaviridae includes enveloped viruses with a frag-mented single-stranded RNA genome of negative polarityTheir tripartite genome consists of a small (S) a medium(M) and a large (L) fragment and the envelope glycoproteinsGn and Gc that are cleaved out of a polyprotein synthesizedby the M fragment These glycoproteins have been provedto mediate the formation of the virus particle to play a rolein the interaction with cell surface receptors to mediate theentry of the virus into cells and to serve as targets for themajority of neutralizing Abs described so far
Orthobunyavirus transmitted through mosquitoes ormidges vectors are divided in 18 serogroups based on cross-titrations in haemagglutination inhibition assays and neutral-ization assays and correlating with main vector preferencesHowever the most clinically important viruses belong onlyto two serogroups the California encephalitis and the Simbuserogroups
Crimean-Congo hemorrhagic fever virus (CCHFV) isconsidered the only one of clinical relevance within theNairovirus genus which uses tick as main vector and is divid-ed in seven serogroups while Toscana virus and Rift Valleyfever virus (RVFV) which are transmitted by sandflies andmosquitoes respectively belong to the Phlebovirus genus[138]
MAbs against theNairovirusCCHFV and the PhlebovirusRVFV have been described
41 Crimean-Congo Hemorrhagic Fever Virus (CCHFV)Crimean-Congo Hemorrhagic Fever Virus (CCHFV) dis-tribution covers the greatest geographic range of any tick-borne virus known as viral isolation andor disease hasbeen reported from more than 30 countries in SoutheasternEurope Africa Asia and Middle East [139 140] Namelyfrom 1953 to 2010 about 6000 human cases were reportedin Southeastern Europe CCHFV causes sporadic outbreakswith mortality rates ranging from 10 to 80 [141 142] Asignificant variation in the time of incubation has beendescribed [139] Prehemorrhagic symptoms include highfever chills headache photophobia and back and abdominalpains Among other symptoms neuropsychiatric changeshave been reported in some patients In severe cases 3ndash6 daysafter the onset of the disease hemorrhagic symptoms occurwith petechiae ecchymosis bleeding in the form of melenahematemesis epistaxis [139]
In comparisonwith the nucleocapsid proteins inCCHFVGn and Gc show a higher degree of antigenic variabilityprobably due to their exposition to the host immune systemhypothesis corroborated in the description by Hewson et alin 2004 of four M segment phylogenic groups namely M1M2 M3 and M4 [141 142]
411 mAbs against CCHFV Blackburn et al firstly describedin 1987 murine anti-CCHFV mAbs able to recognize nucle-ocapsid proteins but no neutralizing activity has beendescribed thus far [143]
On the other hand Bertolotti-Ciarlet et al produced apanel of murine mAbs recognizing conformational epitopes
16 BioMed Research International
within theGn andGc glycoproteins expressed by the CCHFVstrain IbAr10200 The effectiveness of the mAbs was studiedby performing PRNT80 and none of the anti-Gn mAbsshowed neutralizing activity although many of the anti-GcmAbs neutralized IbAr10200 strain in vitro with clones 8A15E3 12A9 6C2 and 9H3 showing activity at a gt5120 dilutionMAbs 11E7 and 30F7 exhibited neutralizing activity at 2560dilution A suckling mice protection test was thereforeperformed for these mAbs The previously mentionedanti-Gc Abs were capable to protect mice to an appreciabledegree when applied 24 hours before and in a weakermanner 24 hours after virus challenge Many of the anti-GnmAbs were able to confer significant protection to IbAr10200both 24 hours before and after virus administration Therelevant effectiveness of the anti-Gn mAbs 6B12 10E1113G8 and 10G4 suggests that these mAbs may possess someneutralizing activity due to Ab-based effector mechanisms(eg ADCC) [58]
In further studies the same group characterized thebroadly cross-reactive neutralizing activity of the previouslydescribed murine mAb 11E7 able to bind conformationalepitopes within the C-terminal region of the Gn glycoproteinof all CCHFVM groups [58 141]
42 Rift Valley Fever Virus (RVFV) Similarly to CCHFV RiftValley Fever Virus (RVFV) causes sporadic outbreaks Thelargest epidemic occurred in Egypt in 1977 with an estimated200000 infections 18000 patients manifesting symptomsand 600 fatal prognoses [144] Symptoms include hemor-rhagic episodes fever encephalitis and blindness [145]
The major target of the humoral immune response is theRVFV glycoproteins Gc (G1) and Gn (G2) ranging fromnucleotides 480ndash2090 and 2091ndash3614 respectively [144]
421 mAbs against RVFV Within G2 glycoprotein are des-cribed three neutralizing antigenic sites namely epitope Inucleotides 792ndash893 (amino acid residues 258ndash291) epitopeII nucleotides 1164ndash1196 (amino acid residues 382ndash392) andepitope IV nucleotides 858ndash917 (amino acid residues 280ndash299) [59 145]
Despite the high degree of variability observed inCCHFVglycoproteins RVFV G2 presented an unexpected high con-servation among 22 isolates [59] In the same study onlythe LUNYO and 900060 isolates were resistant to in vitroneutralization by murine mAb 4D4 (recognizing epitope II)All other isolates were neutralized with PRNT80 titers from10240 togt81920On the other hand only SNS isolate showedreduced sensitivity to 4ndash39-CC (epitope IV) All other isolateswere neutralized with PRNT80 titers from 20480 to gt81920Protection studies on mice would be of interest for thedevelopment of a passive immunization prophylaxis [59]
Protection studies from lethal South African RVFVAN1830 strain infection were successfully performed on abattery of anti-G1 and anti-G2 mAbs produced by Besselaarand Blackburn with the strongly neutralizing anti-G1 mAb3E5 and anti-G2mAb 9C4 being the most effective [146]The9C4 antigenic area maps within G2 epitope I and a broadlyneutralizing activity may be investigated
5 Conclusions
Arbovirus infections have acquired increasing interest giventhe augmented globalization and tourism movement all overthe world that could be at the basis of epidemic eventsFurthermore the same vectors can sometimes transmit sev-eral arboviruses concomitantly complicating the diagnosis aswell as the therapy For example confusing mixed epidemicshave occasionally been described such as YFV plus CHIKVDENVplus CHIKV ormore recentlyPlasmodium falciparummalaria plus DENV1 and CHIKV on Madagascarrsquos east coast[1]
In the last decade several efforts have been employed inthe development of effective immunogens and therapeuticsgiving the current absence of specific antiviral drugs as wellas effective vaccines for themost diffused arbovirus infections(ie DENV andWNV)
However at the moment mosquito control is the bestavailable method for preventing arbovirus infections Thusto control the emerging public health problem of arbovirusinfections new antiviral therapeutic strategies that providepotent and broadly cross-protective immunity (especially forflavivirus infections) are an urgent globally medical need
As described in this review the majority of highlyneutralizing mAbs are of murine origin and indeed theirutility in the treatment of these infections is limited In facttheir employment can elicit an immunogenic response (iehuman anti-mouse Abs HAMA) the therapeutic efficacycould be reduced by a relatively faster clearance in humans(compared to human Abs) potentially exacerbated by theHAMA response Finally murine Abs exhibit relatively weakeffector functions (eg ADCC) compared to human AbsIndeed chimerization humanization or better the isolationand characterization of fully human broadly neutralizingmAbs would be the best choice for their possible use in afuture mAb-based therapy against arbovirus infections [147]
Finally possible ADE of infection mechanisms should beevaluated before considering a mAb as a possible candidatetherapeutic in a post-exposure setting as well as in the devel-opment of new Ab-eliciting immunogens
Conflict of Interests
The authors declare no conflict of interests in particular withOmriGam and any other cited Ab-based preparation men-tioned in the paper
References
[1] N Cleton M Koopmans J Reimerink G J Godeke and CReusken ldquoCome fly with me review of clinically importantarboviruses for global travelersrdquo Journal of Clinical Virology vol55 no 3 pp 191ndash203 2012
[2] F X Heinz H Holzmann A Essl and M Kundi ldquoField effec-tiveness of vaccination against tick-borne encephalitisrdquoVaccinevol 25 no 43 pp 7559ndash7567 2007
[3] A D Barrett and D E Teuwen ldquoYellow fever vaccinemdashhowdoes it work and why do rare cases of serious adverse events
BioMed Research International 17
take placerdquo Current Opinion in Immunology vol 21 no 3 pp308ndash313 2009
[4] S B Halstead and S J Thomas ldquoNew vaccines for Japaneseencephalitisrdquo Current Infectious Disease Reports vol 12 no 3pp 174ndash180 2010
[5] R Burioni M Perotti N Mancini and M Clementi ldquoPer-spectives for the utilization of neutralizing human monoclonalantibodies as anti-HCV drugsrdquo Journal of Hepatology vol 49no 2 pp 299ndash300 2008
[6] R A Diotti G A Sautto L Solforosi NManciniM Clementiand R Burioni ldquoNeutralization activity and kinetics of twobroad-range human monoclonal IgG1 derived from recombi-nant Fab fragments and directed against Hepatitis C virus E2glycoproteinrdquo New Microbiologica vol 35 no 4 pp 475ndash4792012
[7] R Burioni F Canducci N Mancini et al ldquoMolecular cloningof the first humanmonoclona antibodies neutralizing with highpotency Swine-origin Influenza A pandemic virus (S-OIV)rdquoNew Microbiologica vol 32 no 4 pp 319ndash324 2009
[8] P K Chandra S Hazari B Poat et al ldquoIntracytoplasmic stableexpression of IgG1 antibody targeting NS3 helicase inhibitsreplication of highly efficient hepatitis C Virus 2a clonerdquoVirology Journal vol 7 article 118 2010
[9] N Clementi D de Marco N Mancini et al ldquoA human mono-clonal antibody with neutralizing activity against highly diver-gent influenza subtypesrdquo PLoS ONE vol 6 no 12 Article IDe28001 2011
[10] D de Marco N Clementi N Mancini et al ldquoA non-VH1-69 heterosubtypic neutralizing human monoclonal antibodyprotects mice against H1N1 and H5N1 virusesrdquo PLoS ONE vol7 no 4 Article ID e34415 2012
[11] L Solforosi N Mancini F Canducci et al ldquoA phage displayvector optimized for the generation of human antibody com-binatorial libraries and the molecular cloning of monoclonalantibody fragmentsrdquoNewMicrobiologica vol 35 no 3 pp 289ndash294 2012
[12] E V Koonin and V V Dolja ldquoEvolution and taxonomy ofpositive-strand RNA viruses implications of comparative anal-ysis of amino acid sequencesrdquo Critical Reviews in Biochemistryand Molecular Biology vol 28 no 5 pp 375ndash430 1993
[13] J S Mackenzie D J Gubler and L R Petersen ldquoEmergingflaviviruses the spread and resurgence of Japanese encephalitisWest Nile and dengue virusesrdquo Nature Medicine vol 10 no 12pp S98ndashS109 2004
[14] T J SmithW E Brandt J L Swanson J MMcCown and E LBuescher ldquoPhysical and biological properties of dengue-2 virusand associated antigensrdquo Journal of Virology vol 5 no 4 pp524ndash532 1970
[15] J M Smit B Moesker I Rodenhuis-Zybert and J WilschutldquoFlavivirus cell entry and membrane fusionrdquo Viruses vol 3 no2 pp 160ndash171 2011
[16] R J Kuhn W Zhang M G Rossmann et al ldquoStructure ofdengue virus implications for flavivirus organization matura-tion and fusionrdquo Cell vol 108 no 5 pp 717ndash725 2002
[17] K Stiasny J H Aberle M Keller B Grubeck-Loebenstein andF X Heinz ldquoAge affects quantity but not quality of antibodyresponses after vaccination with an inactivated flavivirus vac-cine against tick-borne encephalitisrdquo PLoS ONE vol 7 no 3Article ID e34145 2012
[18] LThomasOVerlaetenACabie et al ldquoInfluence of the dengueserotype previous dengue infection and plasma viral load
on clinical presentation and outcome during a dengue-2 anddengue-4 co-epidemicrdquo American Journal of Tropical Medicineand Hygiene vol 78 no 6 pp 990ndash998 2008
[19] Y Zhang J Corver P R Chipman et al ldquoStructures of imma-ture flavivirus particlesrdquo EMBO Journal vol 22 no 11 pp2604ndash2613 2003
[20] A K I Falconar ldquoIdentification of an epitope on the denguevirus membrane (M) protein defined by cross-protective mon-oclonal antibodies design of an improved epitope sequencebased on common determinants present in both envelope (EandM) proteinsrdquo Archives of Virology vol 144 no 12 pp 2313ndash2330 1999
[21] K-J Huang Y-C Yang Y-S Lin et al ldquoThe dual-specific bind-ing of dengue virus and target cells for the antibody-dependentenhancement of dengue virus infectionrdquo Journal of Immunol-ogy vol 176 no 5 pp 2825ndash2832 2006
[22] A K I Falconar ldquoAntibody responses are generated to immun-odominant ELKKLE-typemotifs on the nonstructural-1 glyco-protein during live dengue virus infections inmice and humansimplications for diagnosis pathogenesis and vaccine designrdquoClinical and Vaccine Immunology vol 14 no 5 pp 493ndash5042007
[23] C W Mandl F Guirakhoo H Holzmann F X Heinz and CKunz ldquoAntigenic structure of the flavivirus envelope protein Eat the molecular level using the tick-borne encephalitis virus asa modelrdquo Journal of Virology vol 63 no 2 pp 564ndash571 1989
[24] F A Rey F X Heinz C Mandl C Kunz and S C HarrisonldquoThe envelope glycoprotein from tick-borne encephalitis virusat 2 A resolutionrdquo Nature vol 375 no 6529 pp 291ndash298 1995
[25] S L Allison K Stiasny K Stadler CWMandl and F XHeinzldquoMapping of functional elements in the stem-anchor regionof tick-borne encephalitis virus envelope protein Erdquo Journal ofVirology vol 73 no 7 pp 5605ndash5612 1999
[26] D E Klein J L Choi and S C Harrison ldquoStructure of a denguevirus envelope protein late-stage fusion intermediaterdquo Journal ofVirology vol 87 no 4 pp 2287ndash2293 2013
[27] S L Hanna T C Pierson M D Sanchez A A Ahmed M MMurtadha and R W Doms ldquoN-linked glycosylation of WestNile virus envelope proteins influences particle assembly andinfectivityrdquo Journal of Virology vol 79 no 21 pp 13262ndash132742005
[28] N Mancini G Sautto N Clementi et al ldquoNeutralization inter-fering antibodies a ldquonovelrdquo example of humoral immune dys-function facilitating viral escaperdquoViruses vol 4 no 9 pp 1731ndash1752 2012
[29] Y Modis S Ogata D Clements and S C Harrison ldquoVariablesurface epitopes in the crystal structure of dengue virus type3 envelope glycoproteinrdquo Journal of Virology vol 79 no 2 pp1223ndash1231 2005
[30] N Miller ldquoRecent progress in dengue vaccine research anddevelopmentrdquo Current Opinion in Molecular Therapeutics vol12 no 1 pp 31ndash38 2010
[31] J H Chavez J R Silva A A Amarilla and L T Moraes Figu-eiredo ldquoDomain III peptides from flavivirus envelope proteinare useful antigens for serologic diagnosis and targets forimmunizationrdquo Biologicals vol 38 no 6 pp 613ndash618 2010
[32] B Shrestha J D Brien S Sukupolvi-Petty et al ldquoThe develop-ment of therapeutic antibodies that neutralize homologous andheterologous genotypes of dengue virus type 1rdquo PLoS Pathogensvol 6 no 4 Article ID e1000823 2010
18 BioMed Research International
[33] G D Gromowski N D Barrett and A D T Barrett ldquoChar-acterization of dengue virus complex-specific neutralizing epi-topes on envelope protein domain III of dengue 2 virusrdquo Journalof Virology vol 82 no 17 pp 8828ndash8837 2008
[34] S Sukupolvi-Petty S K Austin W E Purtha et al ldquoType- andsubcomplex-specific neutralizing antibodies against domain IIIof dengue virus type 2 envelope protein recognize adjacentepitopesrdquo Journal of Virology vol 81 no 23 pp 12816ndash128262007
[35] S Sukupolvi-Petty S K Austin M Engle et al ldquoStructure andfunction analysis of therapeutic monoclonal antibodies againstdengue virus type 2rdquo Journal of Virology vol 84 no 18 pp9227ndash9239 2010
[36] Y-Q Deng J-X Dai G-H Ji et al ldquoA broadly flaviviruscross-neutralizingmonoclonal antibody that recognizes a novelepitope within the fusion loop of e proteinrdquo PLoS ONE vol 6no 1 Article ID e16059 2011
[37] J M Costin E Zaitseva K M Kahle et al ldquoMechanistic studyof broadly neutralizing human monoclonal antibodies againstdengue virus that target the fusion looprdquo Journal of Virologyvol 87 no 1 pp 52ndash66 2013
[38] CMMidgley A FlanaganH B Tran et al ldquoStructural analysisof a dengue cross-reactive antibody complexed with envelopedomain III reveals the molecular basis of cross-reactivityrdquoJournal of Immunology vol 188 no 10 pp 4971ndash4979 2012
[39] N J Moreland P Susanto E Lim et al ldquoPhage display appro-aches for the isolation of monoclonal antibodies against denguevirus envelope domain III from human and mouse derivedlibrariesrdquo International Journal ofMolecular Sciences vol 13 no3 pp 2618ndash2635 2012
[40] J J B Cockburn M E Navarro Sanchez N Fretes et alldquoMechanism of dengue virus broad cross-neutralization by amonoclonal antibodyrdquo Structure vol 20 no 2 pp 303ndash3142012
[41] R Rajamanonmani C Nkenfou P Clancy et al ldquoOn a mousemonoclonal antibody that neutralizes all four dengue virusserotypesrdquo Journal of General Virology vol 90 no 4 pp 799ndash809 2009
[42] L H Gould J Sui H Foellmer et al ldquoProtective and herapeuticcapacity of human single-chain Fv-Fc fusion proteins againstWest Nile virusrdquo Journal of Virology vol 79 no 23 pp 14606ndash14613 2005
[43] H Sultana H G Foellmer G Neelakanta et al ldquoFusion LoopPeptide of the West Nile virus envelope protein is essential forpathogenesis and is recognized by a therapeutic cross-reactivehuman monoclonal antibodyrdquo Journal of Immunology vol 183no 1 pp 650ndash660 2009
[44] T Oliphant M Engle G E Nybakken et al ldquoDevelopment ofa humanized monoclonal antibody with therapeutic potentialagainstWest Nile virusrdquoNature Medicine vol 11 no 5 pp 522ndash530 2005
[45] G E Nybakken T Oliphant S Johnson S Burke M S Dia-mond and D H Fremont ldquoStructural basis of West Nile virusneutralization by a therapeutic antibodyrdquo Nature vol 437 no7059 pp 764ndash769 2005
[46] G D Gromowski J T Roehrig M S Diamond J C LeeT J Pitcher and A D T Barrett ldquoMutations of an antibodybinding energy hot spot on domain III of the dengue 2 envelopeglycoprotein exploited for neutralization escaperdquo Virology vol407 no 2 pp 237ndash246 2010
[47] J T Roehrig R A Bolin andR G Kelly ldquoMonoclonal antibodymapping of the envelope glycoprotein of the dengue 2 virusJamaicardquo Virology vol 246 no 2 pp 317ndash328 1998
[48] R de Alwis S A Smith N P Olivarez et al ldquoIdentification ofhumanneutralizing antibodies that bind to complex epitopes ondengue virionsrdquo Proceedings of the National Academy of Sciencesof the United States of America vol 109 no 19 pp 7439ndash74442012
[49] S K Austin K A Dowd B Shrestha et al ldquoStructural basis ofdifferential neutralization of DENV-1 genotypes by an antibodythat recognizes a cryptic epitoperdquo PLoS Pathogens vol 8 no 10Article ID e1002930 2012
[50] M Throsby C Geuijen J Goudsmit et al ldquoIsolation and char-acterization of human monoclonal antibodies from individualsinfected with West Nile Virusrdquo Journal of Virology vol 80 no14 pp 6982ndash6992 2006
[51] A P Goncalvez R H Purcell and C-J Lai ldquoEpitope deter-minants of a chimpanzee Fab antibody that efficiently cross-neutralizes dengue type 1 and type 2 viruses map to inside andin close proximity to fusion loop of the dengue type 2 virusenvelope glycoproteinrdquo Journal of Virology vol 78 no 23 pp12919ndash12928 2004
[52] J T Roehrig J H Mathews and D W Trent ldquoIdentification ofepitopes on the E glycoprotein of Saint Louis encephalitis virususing monoclonal antibodiesrdquo Virology vol 128 no 1 pp 118ndash126 1983
[53] J T Roehrig L A Staudinger A R Hunt J H Mathews andC D Blair ldquoAntibody prophylaxis and therapy for flavivirusencephalitis infectionsrdquo Annals of the New York Academy ofSciences vol 951 pp 286ndash297 2001
[54] A P Goncalvez C-H Chien K Tubthong et al ldquoHumanizedmonoclonal antibodies derived from chimpanzee fabs protectagainst Japanese encephalitis virus in vitro and in vivordquo Journalof Virology vol 82 no 14 pp 7009ndash7021 2008
[55] M Arakawa T Yamashiro G-I Uechi M Tadano and ANishizono ldquoConstruction of humanFab (1205741120581) library and iden-tification of human monoclonal Fab possessing neutralizingpotency against Japanese encephalitis virusrdquo Microbiology andImmunology vol 51 no 6 pp 617ndash625 2007
[56] L Warter C Y Lee R Thiagarajan et al ldquoChikungunya virusenvelope-specific human monoclonal antibodies with broadneutralization potencyrdquo Journal of Immunology vol 186 no 5pp 3258ndash3264 2011
[57] P Pal K A Dowd J D Brien et al ldquoDevelopment of a highlyprotective combination monoclonal antibody therapy againstchikungunya virusrdquo PLoS Pathogens vol 9 no 4 Article IDe1003312 2013
[58] A Bertolotti-Ciarlet J Smith K Strecker et al ldquoCellular local-ization and antigenic characterization of Crimean-Congo hem-orrhagic fever virus glycoproteinsrdquo Journal of Virology vol 79no 10 pp 6152ndash6161 2005
[59] J K Battles and J M Dalrymple ldquoGenetic variation amonggeographic isolates of Rift Valley fever virusrdquo American Journalof Tropical Medicine and Hygiene vol 39 no 6 pp 617ndash6311988
[60] B AThibodeaux N C Garbino NM Liss et al ldquoA humanizedIgG but not IgM antibody is effective in prophylaxis and therapyof yellow fever infection in an AG12917D-204 peripheralchallenge mouse modelrdquo Antiviral Research vol 94 no 1 pp1ndash8 2012
BioMed Research International 19
[61] S Daffis R E Kontermann J Korimbocus H Zeller H-DKlenk and J Ter Meulen ldquoAntibody responses against wild-type yellow fever virus and the 17D vaccine strain charac-terization with human monoclonal antibody fragments andneutralization escape variantsrdquoVirology vol 337 no 2 pp 262ndash272 2005
[62] L N Levanov L E Matveev E P Goncharova et al ldquoChimericantibodies against tick-borne encephalitis virusrdquo Vaccine vol28 no 32 pp 5265ndash5271 2010
[63] A R Hunt S Frederickson C Hinkel K S Bowdish and J TRoehrig ldquoA humanized murine monoclonal antibody protectsmice either before or after challenge with virulent Venezuelanequine encephalomyelitis virusrdquo Journal of General Virologyvol 87 no 9 pp 2467ndash2476 2006
[64] J M da Silva Voorham I A Rodenhuis-Zybert N V A Nunezet al ldquoAntibodies against the envelope glycoprotein promoteinfectivity of immature dengue virus serotype 2rdquo PLoS ONEvol 7 no 3 Article ID e29957 2012
[65] T C Pierson Q Xu S Nelson et al ldquoThe stoichiometry ofantibody-mediated neutralization and enhancement of westnile virus infectionrdquoCell Host andMicrobe vol 1 no 2 pp 135ndash145 2007
[66] G Sautto N Mancini R A Diotti L Solforosi M Clementiand R Burioni ldquoAnti-hepatitis C virus E2 (HCVE2) glyco-proteinmonoclonal antibodies andneutralization interferencerdquoAntiviral Research vol 96 no 1 pp 82ndash89 2012
[67] R Burioni Y Matsuura N Mancini et al ldquoDiverging effectsof human recombinant anti-hepatitis C virus (HCV) antibodyfragments derived from a single patient on the infectivityof a vesicular stomatitis virusHCV pseudotyperdquo Journal ofVirology vol 76 no 22 pp 11775ndash11779 2002
[68] R J Phillpotts J R Stephenson and J S Porterfield ldquoAntibody-dependent enhancement of tick-borne encephalitus virus infec-tivityrdquo Journal of General Virology vol 66 no 8 pp 1831ndash18371985
[69] K A Dowd and T C Pierson ldquoAntibody-mediated neutraliza-tion of flaviviruses a reductionist viewrdquo Virology vol 411 no 2pp 306ndash315 2011
[70] A K I Falconar ldquoMonoclonal antibodies that bind to commonepitopes on the dengue virus type 2 nonstructural-1 andenvelope glycoproteins display weak neutralizing activity anddifferentiated responses to virulent strains implications forpathogenesis and vaccinesrdquo Clinical and Vaccine Immunologyvol 15 no 3 pp 549ndash561 2008
[71] K Meyer A Banerjee S E Frey R B Belshe and R Ray ldquoAweak neutralizing antibody response to hepatitis C virus enve-lope glycoprotein enhances virus infectionrdquo PLoS ONE vol 6no 8 Article ID e23699 2011
[72] R Burioni R AWilliamson P P Sanna F E Bloom and D RBurton ldquoRecombinant human Fab to glycoprotein D neutral-izes infectivity and prevents cell-to-cell transmission of herpessimplex viruses 1 and 2 in vitrordquo Proceedings of the NationalAcademy of Sciences of the United States of America vol 91 no1 pp 355ndash359 1994
[73] M S Rolph A Zaid N E Rulli and S Mahalingam ldquoDown-regulation of interferon-120573 in antibody-dependent enhancementof dengue viral infections of human macrophages is dependenton interleukin-6rdquo Journal of Infectious Diseases vol 204 no 3pp 489ndash491 2011
[74] S S Whitehead J E Blaney A P Durbin and B R MurphyldquoProspects for a dengue virus vaccinerdquoNature ReviewsMicrobi-ology vol 5 no 7 pp 518ndash528 2007
[75] S-M Lok V Kostyuchenko G ENybakken et al ldquoBinding of aneutralizing antibody to dengue virus alters the arrangement ofsurface glycoproteinsrdquoNature Structural andMolecular Biologyvol 15 no 3 pp 312ndash317 2008
[76] A P Goncalvez R Men C Wernly R H Purcell and C-J Lai ldquoChimpanzee Fab fragments and a derived humanizedimmunogiobulin G1 antibody that efficiently cross-neutralizedengue type 1 and type 2 virusesrdquo Journal of Virology vol 78no 23 pp 12910ndash12918 2004
[77] WM P BWahala A A Kraus L B HaymoreM A Accavitti-Loper and A M de Silva ldquoDengue virus neutralization byhuman immune sera role of envelope protein domain III-reactive antibodyrdquo Virology vol 392 no 1 pp 103ndash113 2009
[78] W D Crill H R Hughes M J Delorey and G-J J ChangldquoHumoral immune responses of dengue fever patients usingepitope-specific serotype-2 virus-like particle antigensrdquo PLoSONE vol 4 no 4 Article ID e4991 2009
[79] C Setthapramote T Sasaki O Puiprom et al ldquoHuman mono-clonal antibodies to neutralize all dengue virus serotypes usinglymphocytes from patients at acute phase of the secondaryinfectionrdquo Biochemical and Biophysical Research Communica-tions vol 423 no 4 pp 867ndash872 2012
[80] M Beltramello K L Williams C P Simmons et al ldquoThehuman immune response to dengue virus is dominated byhighly cross-reactive antibodies endowed with neutralizing andenhancing activityrdquo Cell Host andMicrobe vol 8 no 3 pp 271ndash283 2010
[81] C M Midgley M Bajwa-Joseph S Vasanawathana et al ldquoAnin-depth analysis of original antigenic sin in dengue virusinfectionrdquo Journal of Virology vol 85 no 22 pp 410ndash421 2011
[82] B Kaufmann M R Vogt J Goudsmit et al ldquoNeutralizationof West Nile virus by cross-linking of its surface proteins withFab fragments of the human monoclonal antibody CR4354rdquoProceedings of the National Academy of Sciences of the UnitedStates of America vol 107 no 44 pp 18950ndash18955 2010
[83] B Spurrier J M Sampson M Totrov et al ldquoStructural analysisof human and macaque mAbs 2909 and 25B implicationsfor the configuration of the quaternary neutralizing epitope ofHIV-1 gp120rdquo Structure vol 19 no 5 pp 691ndash699 2011
[84] J J SchlesingerMWBrandriss andT PMonath ldquoMonoclonalantibodies distinguish between wild and vaccine strains ofyellow fever virus by neutralization hemagglutination inhibi-tion and immune precipitation of the virus envelope proteinrdquoVirology vol 125 no 1 pp 8ndash17 1983
[85] M W Brandriss J J Schlesinger E E Walsh and M BrisellildquoLethal 17D yellow fever encephalitis in mice I Passive protec-tion by monoclonal antibodies to the envelope proteins of 17Dyellow fever and dengue 2 virusesrdquo Journal of General Virologyvol 67 no 2 pp 229ndash234 1986
[86] H Cho and M S Diamond ldquoImmune responses to West Nilevirus infection in the central nervous systemrdquo Viruses vol 4no 12 pp 3812ndash3830 2012
[87] D W Beasley A D Barrett and R B Tesh ldquoResurgence ofWest Nile neurologic disease in the United States in 2012 whathappened What needs to be donerdquo Antiviral Research vol 99no 1 pp 1ndash5 2013
[88] M Throsby J Ter Meulen C Geuijen J Goudsmit and J DeKruif ldquoMapping and analysis of West Nile virus-specific mon-oclonal antibodies prospects for vaccine developmentrdquo ExpertReview of Vaccines vol 6 no 2 pp 183ndash191 2007
[89] M S Diamond T G Roberts D Edgil B Lu J Ernst and EHarris ldquoModulation of dengue virus infection in human cells
20 BioMed Research International
by alpha beta and gamma interferonsrdquo Journal of Virology vol74 no 11 pp 4957ndash4966 2000
[90] Z Shimoni M J Niven S Pitlick and S Bulvik ldquoTreatmentof West Nile virus encephalitis with intravenous immunoglob-ulinrdquo Emerging Infectious Diseases vol 7 no 4 article 759 2001
[91] AHamdan PGreen EMendelsonM R Kramer S Pitlik andM Weinberger ldquoPossible benefit of intravenous immunoglob-ulin therapy in a lung transplant recipient with West Nile virusencephalitisrdquoTransplant InfectiousDisease vol 4 no 3 pp 160ndash162 2002
[92] M Haley A S Retter D Fowler J Gea-Banacloche and NP OrsquoGrady ldquoThe role for intravenous immunoglobulin in thetreatment of West Nile virus encephalitisrdquo Clinical InfectiousDiseases vol 37 no 6 pp e88ndashe90 2003
[93] K M Chung G E Nybakken B S Thompson et al ldquoAnti-bodies against West Nile virus nonstructural protein NS1prevent lethal infection through Fc 120574 receptor-dependent and-independent mechanismsrdquo Journal of Virology vol 80 no 3pp 1340ndash1351 2006
[94] E A Henchal L S Henchal and J J Schlesinger ldquoSynergisticinteractions of anti-NS1 monoclonal antibodies protect pas-sively immunized mice from lethal challenge with dengue 2virusrdquo Journal of General Virology vol 69 no 8 pp 2101ndash21071988
[95] J T Roehrig ldquoAntigenic structure of flavivirus proteinsrdquoAdvances in Virus Research vol 59 pp 141ndash175 2003
[96] M D Sanchez T C Pierson D McAllister et al ldquoCharacteri-zation of neutralizing antibodies to West Nile virusrdquo Virologyvol 336 no 1 pp 70ndash82 2005
[97] J H Beigel J L Nordstrom S R Pillemer et al ldquoSafety andpharmacokinetics of single intravenous dose of MGAWN1 anovel monoclonal antibody to West Nile virusrdquo AntimicrobialAgents and Chemotherapy vol 54 no 6 pp 2431ndash2436 2010
[98] C A Smeraski V Siddharthan and J D Morrey ldquoTreatmentof spatial memory impairment in hamsters infected with WestNile virus using a humanizedmonoclonal antibodyMGAWN1rdquoAntiviral Research vol 91 no 1 pp 43ndash49 2011
[99] D Lelli AMoreno E Brocchi et al ldquoWestNile virus character-ization and diagnostic applications of monoclonal antibodiesrdquoVirology Journal vol 9 article 81 2012
[100] T S Gritsun V A Lashkevich and E A Gould ldquoTick-borneencephalitisrdquo Antiviral Research vol 57 no 1-2 pp 129ndash1462003
[101] D Ruzek T S Gritsun N L Forrester et al ldquoMutations inthe NS2B and NS3 genes affect mouse neuroinvasiveness of aWestern European field strain of tick-borne encephalitis virusrdquoVirology vol 374 no 2 pp 249ndash255 2008
[102] M Ecker S L Allison T Meixner and F X Heinz ldquoSequenceanalysis and genetic classification of tick-borne encephalitisviruses from Europe and Asiardquo Journal of General Virology vol80 no 1 pp 179ndash185 1999
[103] H Holzmann M S Vorobyova I P Ladyzhenskaya et alldquoMolecular epidemiology of tick-borne encephalitis viruscross-protection between European and Far Eastern subtypesrdquoVaccine vol 10 no 5 pp 345ndash349 1992
[104] C W Mandl ldquoSteps of the tick-borne encephalitis virus repli-cation cycle that affect neuropathogenesisrdquo Virus Research vol111 no 2 pp 161ndash174 2005
[105] A V Atrasheuskaya T M Fredeking and G M IgnatyevldquoChanges in immune parameters and their correction in humancases of tick-borne encephalitisrdquo Clinical and ExperimentalImmunology vol 131 no 1 pp 148ndash154 2003
[106] K L Mansfield N Johnson L P Phipps J R Stephenson AR Fooks and T Solomon ldquoTick-borne encephalitis virusmdashareview of an emerging zoonosisrdquo Journal of General Virologyvol 90 no 8 pp 1781ndash1794 2009
[107] DWVaughn andCHHoke Jr ldquoThe epidemiology of Japaneseencephalitis prospects for preventionrdquo Epidemiologic Reviewsvol 14 pp 197ndash221 1992
[108] T Solomon H Ni D W C Beasley M Ekkelenkamp M JCardosa and A D T Barrett ldquoOrigin and evolution of Japaneseencephalitis virus in Southeast Asiardquo Journal of Virology vol 77no 5 pp 3091ndash3098 2003
[109] M R Wilson ldquoEmerging viral infectionsrdquo Current Opinion inNeurology vol 26 no 3 pp 301ndash306 2013
[110] J N Hanna S A Ritchie D A Phillips et al ldquoAn outbreakof Japanese encephalitis in the Torres Strait Australia 1995rdquoMedical Journal of Australia vol 165 no 5 pp 256ndash260 1996
[111] J N Hanna S A Ritchie D A Phillips et al ldquoJapanese ence-phalitis in north Queensland Australia 1998rdquo Medical Journalof Australia vol 170 no 11 pp 533ndash536 1999
[112] SGeorgeMGourie-Devi and J A Rao ldquoIsolation ofWestNilevirus from the brains of children who had died of encephalitisrdquoBulletin of theWorldHealthOrganization vol 62 no 6 pp 879ndash882 1984
[113] A K Gupta A A Koshy and V J Lad ldquoEnhanced protec-tion of mice against Japanese encephalitis virus infection bycombinations ofmonoclonal antibodies to glycoprotein ErdquoActaVirologica vol 55 no 2 pp 165ndash168 2011
[114] T H Lee B-H Song S-I Yun et al ldquoA cross-protective mAbrecognizes a novel epitope within the flavivirus NS1 proteinrdquoJournal of General Virology vol 93 no 1 pp 20ndash26 2012
[115] S C Weaver ldquoHost range amplification and arboviral diseaseemergencerdquo Archives of Virology Supplementum no 19 pp 33ndash44 2005
[116] M Solignat B Gay S Higgs L Briant and C Devaux ldquoRepli-cation cycle of chikungunya a re-emerging arbovirusrdquoVirologyvol 393 no 2 pp 183ndash197 2009
[117] O Schwartz and M L Albert ldquoBiology and pathogenesis ofchikungunya virusrdquo Nature Reviews Microbiology vol 8 no 7pp 491ndash500 2010
[118] W Akahata Z-Y Yang H Andersen et al ldquoA virus-like particlevaccine for epidemic Chikungunya virus protects nonhumanprimates against infectionrdquo Nature Medicine vol 16 no 3 pp334ndash338 2010
[119] M Kielian and A Helenius ldquopH-Iinduced alterations in thefusogenic spike protein of Semliki Forest virusrdquo Journal of CellBiology vol 101 no 6 pp 2284ndash2291 1985
[120] J White and A Helenius ldquopH-Dependent fusion between theSemliki Forest virus membrane and liposomesrdquo Proceedings ofthe National Academy of Sciences of the United States of Americavol 77 no 6 pp 3273ndash3277 1980
[121] J Dubuisson and C M Rice ldquoSindbis virus attachment isola-tion and characterization of mutants with impaired binding tovertebrate cellsrdquo Journal of Virology vol 67 no 6 pp 3363ndash33741993
[122] C Y Lee Y-W Kam J Fric et al ldquoChikungunya virus neutral-ization antigens and direct cell-to-cell transmission are revealedby human antibody-escapemutantsrdquo PLoS Pathogens vol 7 no12 Article ID e1002390 2011
[123] J E Voss M-C Vaney S Duquerroy et al ldquoGlycoproteinorganization of Chikungunya virus particles revealed by X-raycrystallographyrdquo Nature vol 468 no 7324 pp 709ndash712 2010
BioMed Research International 21
[124] L Li J Jose Y Xiang R J Kuhn and M G Rossmann ldquoStruc-tural changes of envelope proteins during alphavirus fusionrdquoNature vol 468 no 7324 pp 705ndash708 2010
[125] J Fric S Bertin-Maghit C I Wang A Nardin and L WarterldquoUse of human monoclonal antibodies to treat Chikungunyavirus infectionrdquo Journal of Infectious Diseases vol 207 no 2 pp319ndash322 2013
[126] P V Aguilar J G Estrada-Franco R Navarro-Lopez C FerroA D Haddow and S C Weaver ldquoEndemic Venezuelanequine encephalitis in the Americas hidden under the dengueumbrellardquo Future Virology vol 6 no 6 pp 721ndash740 2011
[127] S C Weaver R Winegar I D Manger and N L ForresterldquoAlphaviruses population genetics and determinants of emer-gencerdquo Antiviral Research vol 94 no 3 pp 242ndash257 2012
[128] S Paessler and S C Weaver ldquoVaccines for Venezuelan equineencephalitisrdquo Vaccine vol 27 no 4 pp D80ndashD85 2009
[129] R J Phillpotts ldquoVenezuelan equine encephalitis virus complex-specific monoclonal antibody provides broad protection inmurine models against airborne challenge with viruses fromserogroups I II and IIIrdquoVirus Research vol 120 no 1-2 pp 107ndash112 2006
[130] R J Phillpotts L D Jones and S C Howard ldquoMonoclonalantibody protectsmice against infection anddiseasewhen giveneither before or up to 24 h after airborne challenge with virulentVenezuelan equine encephalitis virusrdquoVaccine vol 20 no 11-12pp 1497ndash1504 2002
[131] J H Mathews J T Roehrig and D W Trent ldquoRole of com-plement and the Fc portion of immunoglobulin G in immunityto Venezuelan equine encephalomyelitis virus infection withglycoprotein-specific monoclonal antibodiesrdquo Journal of Virol-ogy vol 55 no 3 pp 594ndash600 1985
[132] L M OrsquoBrien C D Underwood-Fowler S A Goodchild AL Phelps and R J Phillpotts ldquoDevelopment of a novel mon-oclonal antibody with reactivity to a wide range of Venezuelanequine encephalitis virus strainsrdquoVirology Journal vol 6 article206 2009
[133] L M OrsquoBrien S A Goodchild R J Phillpotts and S D Per-kins ldquoA humanised murine monoclonal antibody protectsmice from Venezuelan equine encephalitis virus Evergladesvirus and Mucambo virus when administered up to 48h afterairborne challengerdquo Virology vol 426 no 2 pp 100ndash105 2012
[134] N Clementi N Mancini L Solforosi M Castelli M Clementiand R Burioni ldquoPhage display-based strategies for cloning andoptimization of monoclonal antibodies directed against humanpathogensrdquo International Journal of Molecular Sciences vol 13no 7 pp 8273ndash8292 2012
[135] A R Hunt R A Bowen S Frederickson T Maruyama JT Roehrig and C D Blair ldquoTreatment of mice with humanmonoclonal antibody 24h after lethal aerosol challenge withvirulent Venezuelan equine encephalitis virus prevents diseasebut not infectionrdquo Virology vol 414 no 2 pp 146ndash152 2011
[136] W-G Hu A L Phelps S Jager et al ldquoA recombinant human-ized monoclonal antibody completely protects mice againstlethal challenge with Venezuelan equine encephalitis virusrdquoVaccine vol 28 no 34 pp 5558ndash5564 2010
[137] T Rulker L Voss P Thullier et al ldquoIsolation and characterisa-tion of a human-like antibody fragment (scFv) that inactivatesVEEV in vitro and in vivordquo PLoS ONE vol 7 no 5 Article IDe37242 2012
[138] C T Walter and J N Barr ldquoRecent advances in the molecularand cellular biology of bunyavirusesrdquo Journal of General Virol-ogy vol 92 no 11 pp 2467ndash2484 2011
[139] C A Whitehouse ldquoCrimean-Congo hemorrhagic feverrdquo Anti-viral Research vol 64 no 3 pp 145ndash160 2004
[140] M Mertens K Schmidt A Ozkul and M H Groschup ldquoTheimpact of Crimean-Congo hemorrhagic fever virus on publichealthrdquo Antiviral Research vol 98 no 2 pp 248ndash260 2013
[141] A A Ahmed J M McFalls C Hoffmann et al ldquoPresence ofbroadly reactive and group-specific neutralizing epitopes onnewly described isolates of Crimean-Congo hemorrhagic fevervirusrdquo Journal of General Virology vol 86 no 12 pp 3327ndash33362005
[142] R Hewson A Gmyl L Gmyl et al ldquoEvidence of segment reas-sortment in Crimean-Congo haemorrhagic fever virusrdquo Journalof General Virology vol 85 no 10 pp 3059ndash3070 2004
[143] N K Blackburn T G Besselaar A J Shepherd and R Swane-poel ldquoPreparation and use of monoclonal antibodies for iden-tifying Crimean-Congo hemorrhagic fever virusrdquo AmericanJournal of Tropical Medicine and Hygiene vol 37 no 2 pp 392ndash397 1987
[144] T Ikegami ldquoMolecular biology and genetic diversity of RiftValley fever virusrdquoAntiviral Research vol 95 no 3 pp 293ndash3102012
[145] KKeegan andM S Collett ldquoUse of bacterial expression cloningto define the amino acid sequences of antigenic determinantson the G2 glycoprotein of Rift Valley fever virusrdquo Journal ofVirology vol 58 no 2 pp 263ndash270 1986
[146] T G Besselaar and N K Blackburn ldquoTopological mapping ofantigenic sites on the Rift Valley fever virus envelope glycopro-teins using monoclonal antibodiesrdquo Archives of Virology vol121 no 1ndash4 pp 111ndash124 1991
[147] L G Presta ldquoEngineering of therapeutic antibodies to mini-mize immunogenicity and optimize functionrdquo Advanced DrugDelivery Reviews vol 58 no 5-6 pp 640ndash656 2006
Submit your manuscripts athttpwwwhindawicom
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anatomy Research International
PeptidesInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporation httpwwwhindawicom
International Journal of
Volume 2014
Zoology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Molecular Biology International
GenomicsInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioinformaticsAdvances in
Marine BiologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Signal TransductionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Evolutionary BiologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Biochemistry Research International
ArchaeaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Genetics Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in
Virolog y
Hindawi Publishing Corporationhttpwwwhindawicom
Nucleic AcidsJournal of
Volume 2014
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Enzyme Research
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Microbiology

14 BioMed Research International
study cell-to-cell transmission was firstly demonstrated as anescapemechanism enhanced by the E2mutation R82G [122]
In further in vivo studies 5F10 and 8B10 significantlydelayed CHIKV-caused death of AGR129 mice both in pro-phylactic and therapeutic tests Interestingly in therapeutictreatment these mAbs showed a synergistic effect whenadministered in combination with the total amount of mAbsinjected being the same of the single-mAbs Possibly thesetreatment did not allow themice to survive CHIKV challengedue to Ig clearing and 6ndash10 days half-life usually described forhuman mAbs [125]
Recently Pal et al cloned thirty-six murine mAbs ableto neutralize the ECSA La Reunion 2006 OPY-1 strain ofCHIKV (CHIKV-LR) the majority of which also neutralizeinfection of other strains corresponding to the Asian andWest African genotypes In particularmAbCHK-152 showedthe highest and broadest neutralizing activity with FRNTvalues indicating an IC50 value of 1 to 3 ngmL dependingon the viral strain used Among the thirty-six mAbs four(CHK-102 CHK-152 and CHK-166 CHK-263) could pre-vent lethality of immunodeficient Ifnarminusminus C57BL6 micewhen administered one day before exposition with CHK-152 and CHK-263 showing the ability to protect mice at thelowest dose (10 120583g) Therapeutic studies were performed onthese mAbs administering a single dose of 100 120583g 24 hoursafter infection The highest activity was described for CHK-166 able to protect 63 of the mice Testing the combinedactivity of the mAbs in therapeutic studies administration ofCHK-166 plus CHK-152 in a dose of 250120583g each turned out tobe themost effective protecting 71 of the IfnarminusminusC57BL6mice 60 hours after infection [57]
32 Venezuelan Equine Encephalomyelitis Virus (VEEV)Venezuelan equine encephalomyelitis virus (VEEV) is main-tained in a natural transmission cycle between mosquitoesand small rodents The first documented outbreaks occurredin the 1930s and several epidemics have been reported so farin Latin-American countries such as Venezuela ColombiaPeru Ecuador Bolivia Honduras Mexico and Panama withhundreds of thousands of human cases being reported anda case-fatality rate up to 1 during the 1969 Ecuador out-break Clinical manifestations of VEE are indistinguishablefrom Dengue [126] Six serogroups (IndashVI) are currentlyrecognized within the VEEV complex VEEV caused humanand equine outbreaks in the Americas for nearly a centuryEquine epizootics have high mortality (38ndash83) leading toa high viremia followed by a lethal encephalitis and often tohuman epidemics involving thousands of cases and hundredsof deaths Indeed VEEV is infectious for humans by theairborne route and has been responsible for a number oflaboratory infections In humans the disease is usually self-limiting with a febrile illness with 1ndash4 of cases progressingto severe encephalitis It is a hazard to laboratory workershas been developed as a biological weapon and is a potentialbioterrorist agent [127] There are no antiviral drugs andvaccines licensed for the treatment of VEEV infection inhumans Indeed although experimental live-attenuated vac-cines have been developed (eg TC-83) with good levels ofprotection in equine and mice in humans they may fail to
give protection in the majority of cases [128] Consideringthe protective role exerted by humoral immune response inanimal models it could be concluded that treatment withspecific IgG may have a beneficial antiviral effect in humanairborne infections with pathogenic strains of VEEV
321 mAbs against VEEV Several works support that mAbsmay protect against airborne VEEV as well as after airborneexposure toVEEVAb reactivity to both surface glycoproteins(E1 and E2) is associated with neutralization of the virusin vitro and passive protection against virus challenge Aseries of anti-E1 and anti-E2 mouse and humanized mAbshave been described recently Among them Phillpotts et alexamined two anti-E2 murine mAbs named 1A4A-1 and1A3A-9 both having potent protective activity against subcu-taneous VEEV challenge in mice Both mAbs had a similarhalf-life (58 and 100 days) in mouse serum after a singleintraperitoneal dose suggesting that mAbs delivered by thisroute at or around the time of VEEV infection persists athigh levels in the blood as well as secretions in respiratorytransudation throughout the clinical course of the disease Inparticular both mAbs administered 24 h prior to airbornechallenges had a substantial protective effect (90ndash100)Treatment of mice 2 or 24 h after airborne infection with asingle intraperitoneal dose of 100 120583g of 1A3A-9 mAb led toapproximately 50 survival There was no beneficial effectwhen mAb treatment was delayed to 72 h Moreover therewas evidence of synergy in vitro in PRNT between 1A3A-9and 1A4A-1 as has been demonstrated for other virusesand mAb pairs However no synergy was found in mouseprotection when mAbs were delivered intraperitoneally as amixture in equal parts to animals challenged 24 h previouslywith VEEV [129]
In a subsequent study Phillpotts et al reported two otherprotective mAbs The 3B2A-9 mAb protected against allthe serogroup I strains while 1A3B-7 mAb protected wellagainst challenge with all of the viruses tested Both 3B2A-9 and 1A3B-7 protected against airborne exposure to theIAB serogroup virus strainTrinidad donkey (TrD)Howeverthere was no evidence of synergistic protection when thesemAbs were combined in equal proportions An intraperi-toneal dose of 10 120583g was sufficient to protect 50 of themice with either mAb [130] According to other data themechanism of protection did not appear to depend uponneutralization [131] Similar results were obtained by OrsquoBrienet al which described a non-neutralizing mAb (IgG2a)named CUF37-2a able to protect 50 of mice from asubcutaneous VEEV challenge when a dose of 915 120583g of mAbwas administered 24 h prior to challenge [132]
In a following study the 1A3B-7 mAb was humanized(and following reported as Hu1A3B-7) maintaining the samefeatures of its murine counterpart In particular evaluation ofin vitro studies indicated that Hu1A3B-7 retained both broadspecificity and neutralizing activity Furthermore in vivoexperiments showed that Hu1A3B-7 successfully protectedmice against lethal subcutaneous and aerosol challenges withVEEV strain TrD Moreover the effectiveness of the human-ization process was determined by assessing proliferationresponses in human T-cells to peptides derived from the
BioMed Research International 15
murine and humanized versions of the VH and VL domainsThis analysis showed that the number of human T-cellepitopes within the humanized Ab had been substantiallyreduced indicating that Hu1A3B-7 may have low immuno-genicity in vivo [133]
Similarly Hunt et al described a humanizedmurinemAbderived from the 3B4C-4 mAb recognizing the E2c epitope(amino acid residues 182ndash209) and following named Hy4-26C showing neutralizing activity similar to the murinemAb Moreover it was protective (70ndash100 of survival) at100 ng dose in mice animal model intraperitoneal inoculatedand at 500120583g for intranasal challenge (80) Therapeuticstudies in mice revealed that Hy4-26C was able to cure miceup to 24 h following infection at 10 120583g on the contrary themouse mAb when given 1 hour after virus challenge [63]
In another work Hunt et al isolated two neutralizinghumanized Fabs F5 and L1A7 employing a blocking strategy[134 135] The anti-E2 specific F5 IgG had a 70 PRNTendpoint of 10 ngmL equivalent to that described for themost effective neutralizing anti-VEEVE2mAbs In particularthe E1-specific hFab L1A7 had a PRNT endpoint of 3120583gmL300-fold lower than F5
Further studies revealed that F5 epitope is located inthe 115ndash199 amino acid region Moreover F5 IgG had potentability to protect mice from infection by either route whenadministered 24 h before exposure however mice treated24 h and 48 h after aerosol exposure developed central ner-vous system infections but exhibited no clinical signs ofdisease [135]
Hu et al reported that passive immunization with thehumanized chimericmousemAbhu1A4A1IgG1-2A inmice at50 120583g 24 h before or after virulent VEEV challenge providedcomplete protection indicating that hu1A4A1IgG1-2A haspotent prophylactic and therapeutic effects against VEEVinfection [136]
Anti-VEEV mAbs isolated from a non-human primategene library has also been reported In particular thehumanized chimeric scFv-Fc ToR67-3B4 recognized viableas well as formalin and 120573-propiolactone inactivated virusparticles It detected specifically the viral E1 envelope proteinof VEEV but did not react with reduced viral glycopro-tein preparations suggesting that recognition depends uponconformational epitopes The recombinant Ab was able todetect multiple VEEV subtypes and displayed only marginalcross-reactivity to otherAlphavirus species except for Easternequine encephalitis virus (EEEV) In addition the scFv-Fcfusion described here might be of therapeutic use since itsuccessfully inactivated VEEV in a murine disease modelIn particular when the recombinant Ab was administered 6hours after challenge 80 to 100 of mice survived lethalVEEV IAB or IE infection Forty to sixty percent of micesurvived when scFv-Fc ToR67-3B4 was applied 6 hours afterchallenge with VEEV subtypes II and former IIIA [137]
4 Bunyaviridae
TheBunyaviridae family contains human arboviruses belong-ing to the Orthobunyavirus Phlebovirus and Nairovirus
genera Bunyaviridae includes enveloped viruses with a frag-mented single-stranded RNA genome of negative polarityTheir tripartite genome consists of a small (S) a medium(M) and a large (L) fragment and the envelope glycoproteinsGn and Gc that are cleaved out of a polyprotein synthesizedby the M fragment These glycoproteins have been provedto mediate the formation of the virus particle to play a rolein the interaction with cell surface receptors to mediate theentry of the virus into cells and to serve as targets for themajority of neutralizing Abs described so far
Orthobunyavirus transmitted through mosquitoes ormidges vectors are divided in 18 serogroups based on cross-titrations in haemagglutination inhibition assays and neutral-ization assays and correlating with main vector preferencesHowever the most clinically important viruses belong onlyto two serogroups the California encephalitis and the Simbuserogroups
Crimean-Congo hemorrhagic fever virus (CCHFV) isconsidered the only one of clinical relevance within theNairovirus genus which uses tick as main vector and is divid-ed in seven serogroups while Toscana virus and Rift Valleyfever virus (RVFV) which are transmitted by sandflies andmosquitoes respectively belong to the Phlebovirus genus[138]
MAbs against theNairovirusCCHFV and the PhlebovirusRVFV have been described
41 Crimean-Congo Hemorrhagic Fever Virus (CCHFV)Crimean-Congo Hemorrhagic Fever Virus (CCHFV) dis-tribution covers the greatest geographic range of any tick-borne virus known as viral isolation andor disease hasbeen reported from more than 30 countries in SoutheasternEurope Africa Asia and Middle East [139 140] Namelyfrom 1953 to 2010 about 6000 human cases were reportedin Southeastern Europe CCHFV causes sporadic outbreakswith mortality rates ranging from 10 to 80 [141 142] Asignificant variation in the time of incubation has beendescribed [139] Prehemorrhagic symptoms include highfever chills headache photophobia and back and abdominalpains Among other symptoms neuropsychiatric changeshave been reported in some patients In severe cases 3ndash6 daysafter the onset of the disease hemorrhagic symptoms occurwith petechiae ecchymosis bleeding in the form of melenahematemesis epistaxis [139]
In comparisonwith the nucleocapsid proteins inCCHFVGn and Gc show a higher degree of antigenic variabilityprobably due to their exposition to the host immune systemhypothesis corroborated in the description by Hewson et alin 2004 of four M segment phylogenic groups namely M1M2 M3 and M4 [141 142]
411 mAbs against CCHFV Blackburn et al firstly describedin 1987 murine anti-CCHFV mAbs able to recognize nucle-ocapsid proteins but no neutralizing activity has beendescribed thus far [143]
On the other hand Bertolotti-Ciarlet et al produced apanel of murine mAbs recognizing conformational epitopes
16 BioMed Research International
within theGn andGc glycoproteins expressed by the CCHFVstrain IbAr10200 The effectiveness of the mAbs was studiedby performing PRNT80 and none of the anti-Gn mAbsshowed neutralizing activity although many of the anti-GcmAbs neutralized IbAr10200 strain in vitro with clones 8A15E3 12A9 6C2 and 9H3 showing activity at a gt5120 dilutionMAbs 11E7 and 30F7 exhibited neutralizing activity at 2560dilution A suckling mice protection test was thereforeperformed for these mAbs The previously mentionedanti-Gc Abs were capable to protect mice to an appreciabledegree when applied 24 hours before and in a weakermanner 24 hours after virus challenge Many of the anti-GnmAbs were able to confer significant protection to IbAr10200both 24 hours before and after virus administration Therelevant effectiveness of the anti-Gn mAbs 6B12 10E1113G8 and 10G4 suggests that these mAbs may possess someneutralizing activity due to Ab-based effector mechanisms(eg ADCC) [58]
In further studies the same group characterized thebroadly cross-reactive neutralizing activity of the previouslydescribed murine mAb 11E7 able to bind conformationalepitopes within the C-terminal region of the Gn glycoproteinof all CCHFVM groups [58 141]
42 Rift Valley Fever Virus (RVFV) Similarly to CCHFV RiftValley Fever Virus (RVFV) causes sporadic outbreaks Thelargest epidemic occurred in Egypt in 1977 with an estimated200000 infections 18000 patients manifesting symptomsand 600 fatal prognoses [144] Symptoms include hemor-rhagic episodes fever encephalitis and blindness [145]
The major target of the humoral immune response is theRVFV glycoproteins Gc (G1) and Gn (G2) ranging fromnucleotides 480ndash2090 and 2091ndash3614 respectively [144]
421 mAbs against RVFV Within G2 glycoprotein are des-cribed three neutralizing antigenic sites namely epitope Inucleotides 792ndash893 (amino acid residues 258ndash291) epitopeII nucleotides 1164ndash1196 (amino acid residues 382ndash392) andepitope IV nucleotides 858ndash917 (amino acid residues 280ndash299) [59 145]
Despite the high degree of variability observed inCCHFVglycoproteins RVFV G2 presented an unexpected high con-servation among 22 isolates [59] In the same study onlythe LUNYO and 900060 isolates were resistant to in vitroneutralization by murine mAb 4D4 (recognizing epitope II)All other isolates were neutralized with PRNT80 titers from10240 togt81920On the other hand only SNS isolate showedreduced sensitivity to 4ndash39-CC (epitope IV) All other isolateswere neutralized with PRNT80 titers from 20480 to gt81920Protection studies on mice would be of interest for thedevelopment of a passive immunization prophylaxis [59]
Protection studies from lethal South African RVFVAN1830 strain infection were successfully performed on abattery of anti-G1 and anti-G2 mAbs produced by Besselaarand Blackburn with the strongly neutralizing anti-G1 mAb3E5 and anti-G2mAb 9C4 being the most effective [146]The9C4 antigenic area maps within G2 epitope I and a broadlyneutralizing activity may be investigated
5 Conclusions
Arbovirus infections have acquired increasing interest giventhe augmented globalization and tourism movement all overthe world that could be at the basis of epidemic eventsFurthermore the same vectors can sometimes transmit sev-eral arboviruses concomitantly complicating the diagnosis aswell as the therapy For example confusing mixed epidemicshave occasionally been described such as YFV plus CHIKVDENVplus CHIKV ormore recentlyPlasmodium falciparummalaria plus DENV1 and CHIKV on Madagascarrsquos east coast[1]
In the last decade several efforts have been employed inthe development of effective immunogens and therapeuticsgiving the current absence of specific antiviral drugs as wellas effective vaccines for themost diffused arbovirus infections(ie DENV andWNV)
However at the moment mosquito control is the bestavailable method for preventing arbovirus infections Thusto control the emerging public health problem of arbovirusinfections new antiviral therapeutic strategies that providepotent and broadly cross-protective immunity (especially forflavivirus infections) are an urgent globally medical need
As described in this review the majority of highlyneutralizing mAbs are of murine origin and indeed theirutility in the treatment of these infections is limited In facttheir employment can elicit an immunogenic response (iehuman anti-mouse Abs HAMA) the therapeutic efficacycould be reduced by a relatively faster clearance in humans(compared to human Abs) potentially exacerbated by theHAMA response Finally murine Abs exhibit relatively weakeffector functions (eg ADCC) compared to human AbsIndeed chimerization humanization or better the isolationand characterization of fully human broadly neutralizingmAbs would be the best choice for their possible use in afuture mAb-based therapy against arbovirus infections [147]
Finally possible ADE of infection mechanisms should beevaluated before considering a mAb as a possible candidatetherapeutic in a post-exposure setting as well as in the devel-opment of new Ab-eliciting immunogens
Conflict of Interests
The authors declare no conflict of interests in particular withOmriGam and any other cited Ab-based preparation men-tioned in the paper
References
[1] N Cleton M Koopmans J Reimerink G J Godeke and CReusken ldquoCome fly with me review of clinically importantarboviruses for global travelersrdquo Journal of Clinical Virology vol55 no 3 pp 191ndash203 2012
[2] F X Heinz H Holzmann A Essl and M Kundi ldquoField effec-tiveness of vaccination against tick-borne encephalitisrdquoVaccinevol 25 no 43 pp 7559ndash7567 2007
[3] A D Barrett and D E Teuwen ldquoYellow fever vaccinemdashhowdoes it work and why do rare cases of serious adverse events
BioMed Research International 17
take placerdquo Current Opinion in Immunology vol 21 no 3 pp308ndash313 2009
[4] S B Halstead and S J Thomas ldquoNew vaccines for Japaneseencephalitisrdquo Current Infectious Disease Reports vol 12 no 3pp 174ndash180 2010
[5] R Burioni M Perotti N Mancini and M Clementi ldquoPer-spectives for the utilization of neutralizing human monoclonalantibodies as anti-HCV drugsrdquo Journal of Hepatology vol 49no 2 pp 299ndash300 2008
[6] R A Diotti G A Sautto L Solforosi NManciniM Clementiand R Burioni ldquoNeutralization activity and kinetics of twobroad-range human monoclonal IgG1 derived from recombi-nant Fab fragments and directed against Hepatitis C virus E2glycoproteinrdquo New Microbiologica vol 35 no 4 pp 475ndash4792012
[7] R Burioni F Canducci N Mancini et al ldquoMolecular cloningof the first humanmonoclona antibodies neutralizing with highpotency Swine-origin Influenza A pandemic virus (S-OIV)rdquoNew Microbiologica vol 32 no 4 pp 319ndash324 2009
[8] P K Chandra S Hazari B Poat et al ldquoIntracytoplasmic stableexpression of IgG1 antibody targeting NS3 helicase inhibitsreplication of highly efficient hepatitis C Virus 2a clonerdquoVirology Journal vol 7 article 118 2010
[9] N Clementi D de Marco N Mancini et al ldquoA human mono-clonal antibody with neutralizing activity against highly diver-gent influenza subtypesrdquo PLoS ONE vol 6 no 12 Article IDe28001 2011
[10] D de Marco N Clementi N Mancini et al ldquoA non-VH1-69 heterosubtypic neutralizing human monoclonal antibodyprotects mice against H1N1 and H5N1 virusesrdquo PLoS ONE vol7 no 4 Article ID e34415 2012
[11] L Solforosi N Mancini F Canducci et al ldquoA phage displayvector optimized for the generation of human antibody com-binatorial libraries and the molecular cloning of monoclonalantibody fragmentsrdquoNewMicrobiologica vol 35 no 3 pp 289ndash294 2012
[12] E V Koonin and V V Dolja ldquoEvolution and taxonomy ofpositive-strand RNA viruses implications of comparative anal-ysis of amino acid sequencesrdquo Critical Reviews in Biochemistryand Molecular Biology vol 28 no 5 pp 375ndash430 1993
[13] J S Mackenzie D J Gubler and L R Petersen ldquoEmergingflaviviruses the spread and resurgence of Japanese encephalitisWest Nile and dengue virusesrdquo Nature Medicine vol 10 no 12pp S98ndashS109 2004
[14] T J SmithW E Brandt J L Swanson J MMcCown and E LBuescher ldquoPhysical and biological properties of dengue-2 virusand associated antigensrdquo Journal of Virology vol 5 no 4 pp524ndash532 1970
[15] J M Smit B Moesker I Rodenhuis-Zybert and J WilschutldquoFlavivirus cell entry and membrane fusionrdquo Viruses vol 3 no2 pp 160ndash171 2011
[16] R J Kuhn W Zhang M G Rossmann et al ldquoStructure ofdengue virus implications for flavivirus organization matura-tion and fusionrdquo Cell vol 108 no 5 pp 717ndash725 2002
[17] K Stiasny J H Aberle M Keller B Grubeck-Loebenstein andF X Heinz ldquoAge affects quantity but not quality of antibodyresponses after vaccination with an inactivated flavivirus vac-cine against tick-borne encephalitisrdquo PLoS ONE vol 7 no 3Article ID e34145 2012
[18] LThomasOVerlaetenACabie et al ldquoInfluence of the dengueserotype previous dengue infection and plasma viral load
on clinical presentation and outcome during a dengue-2 anddengue-4 co-epidemicrdquo American Journal of Tropical Medicineand Hygiene vol 78 no 6 pp 990ndash998 2008
[19] Y Zhang J Corver P R Chipman et al ldquoStructures of imma-ture flavivirus particlesrdquo EMBO Journal vol 22 no 11 pp2604ndash2613 2003
[20] A K I Falconar ldquoIdentification of an epitope on the denguevirus membrane (M) protein defined by cross-protective mon-oclonal antibodies design of an improved epitope sequencebased on common determinants present in both envelope (EandM) proteinsrdquo Archives of Virology vol 144 no 12 pp 2313ndash2330 1999
[21] K-J Huang Y-C Yang Y-S Lin et al ldquoThe dual-specific bind-ing of dengue virus and target cells for the antibody-dependentenhancement of dengue virus infectionrdquo Journal of Immunol-ogy vol 176 no 5 pp 2825ndash2832 2006
[22] A K I Falconar ldquoAntibody responses are generated to immun-odominant ELKKLE-typemotifs on the nonstructural-1 glyco-protein during live dengue virus infections inmice and humansimplications for diagnosis pathogenesis and vaccine designrdquoClinical and Vaccine Immunology vol 14 no 5 pp 493ndash5042007
[23] C W Mandl F Guirakhoo H Holzmann F X Heinz and CKunz ldquoAntigenic structure of the flavivirus envelope protein Eat the molecular level using the tick-borne encephalitis virus asa modelrdquo Journal of Virology vol 63 no 2 pp 564ndash571 1989
[24] F A Rey F X Heinz C Mandl C Kunz and S C HarrisonldquoThe envelope glycoprotein from tick-borne encephalitis virusat 2 A resolutionrdquo Nature vol 375 no 6529 pp 291ndash298 1995
[25] S L Allison K Stiasny K Stadler CWMandl and F XHeinzldquoMapping of functional elements in the stem-anchor regionof tick-borne encephalitis virus envelope protein Erdquo Journal ofVirology vol 73 no 7 pp 5605ndash5612 1999
[26] D E Klein J L Choi and S C Harrison ldquoStructure of a denguevirus envelope protein late-stage fusion intermediaterdquo Journal ofVirology vol 87 no 4 pp 2287ndash2293 2013
[27] S L Hanna T C Pierson M D Sanchez A A Ahmed M MMurtadha and R W Doms ldquoN-linked glycosylation of WestNile virus envelope proteins influences particle assembly andinfectivityrdquo Journal of Virology vol 79 no 21 pp 13262ndash132742005
[28] N Mancini G Sautto N Clementi et al ldquoNeutralization inter-fering antibodies a ldquonovelrdquo example of humoral immune dys-function facilitating viral escaperdquoViruses vol 4 no 9 pp 1731ndash1752 2012
[29] Y Modis S Ogata D Clements and S C Harrison ldquoVariablesurface epitopes in the crystal structure of dengue virus type3 envelope glycoproteinrdquo Journal of Virology vol 79 no 2 pp1223ndash1231 2005
[30] N Miller ldquoRecent progress in dengue vaccine research anddevelopmentrdquo Current Opinion in Molecular Therapeutics vol12 no 1 pp 31ndash38 2010
[31] J H Chavez J R Silva A A Amarilla and L T Moraes Figu-eiredo ldquoDomain III peptides from flavivirus envelope proteinare useful antigens for serologic diagnosis and targets forimmunizationrdquo Biologicals vol 38 no 6 pp 613ndash618 2010
[32] B Shrestha J D Brien S Sukupolvi-Petty et al ldquoThe develop-ment of therapeutic antibodies that neutralize homologous andheterologous genotypes of dengue virus type 1rdquo PLoS Pathogensvol 6 no 4 Article ID e1000823 2010
18 BioMed Research International
[33] G D Gromowski N D Barrett and A D T Barrett ldquoChar-acterization of dengue virus complex-specific neutralizing epi-topes on envelope protein domain III of dengue 2 virusrdquo Journalof Virology vol 82 no 17 pp 8828ndash8837 2008
[34] S Sukupolvi-Petty S K Austin W E Purtha et al ldquoType- andsubcomplex-specific neutralizing antibodies against domain IIIof dengue virus type 2 envelope protein recognize adjacentepitopesrdquo Journal of Virology vol 81 no 23 pp 12816ndash128262007
[35] S Sukupolvi-Petty S K Austin M Engle et al ldquoStructure andfunction analysis of therapeutic monoclonal antibodies againstdengue virus type 2rdquo Journal of Virology vol 84 no 18 pp9227ndash9239 2010
[36] Y-Q Deng J-X Dai G-H Ji et al ldquoA broadly flaviviruscross-neutralizingmonoclonal antibody that recognizes a novelepitope within the fusion loop of e proteinrdquo PLoS ONE vol 6no 1 Article ID e16059 2011
[37] J M Costin E Zaitseva K M Kahle et al ldquoMechanistic studyof broadly neutralizing human monoclonal antibodies againstdengue virus that target the fusion looprdquo Journal of Virologyvol 87 no 1 pp 52ndash66 2013
[38] CMMidgley A FlanaganH B Tran et al ldquoStructural analysisof a dengue cross-reactive antibody complexed with envelopedomain III reveals the molecular basis of cross-reactivityrdquoJournal of Immunology vol 188 no 10 pp 4971ndash4979 2012
[39] N J Moreland P Susanto E Lim et al ldquoPhage display appro-aches for the isolation of monoclonal antibodies against denguevirus envelope domain III from human and mouse derivedlibrariesrdquo International Journal ofMolecular Sciences vol 13 no3 pp 2618ndash2635 2012
[40] J J B Cockburn M E Navarro Sanchez N Fretes et alldquoMechanism of dengue virus broad cross-neutralization by amonoclonal antibodyrdquo Structure vol 20 no 2 pp 303ndash3142012
[41] R Rajamanonmani C Nkenfou P Clancy et al ldquoOn a mousemonoclonal antibody that neutralizes all four dengue virusserotypesrdquo Journal of General Virology vol 90 no 4 pp 799ndash809 2009
[42] L H Gould J Sui H Foellmer et al ldquoProtective and herapeuticcapacity of human single-chain Fv-Fc fusion proteins againstWest Nile virusrdquo Journal of Virology vol 79 no 23 pp 14606ndash14613 2005
[43] H Sultana H G Foellmer G Neelakanta et al ldquoFusion LoopPeptide of the West Nile virus envelope protein is essential forpathogenesis and is recognized by a therapeutic cross-reactivehuman monoclonal antibodyrdquo Journal of Immunology vol 183no 1 pp 650ndash660 2009
[44] T Oliphant M Engle G E Nybakken et al ldquoDevelopment ofa humanized monoclonal antibody with therapeutic potentialagainstWest Nile virusrdquoNature Medicine vol 11 no 5 pp 522ndash530 2005
[45] G E Nybakken T Oliphant S Johnson S Burke M S Dia-mond and D H Fremont ldquoStructural basis of West Nile virusneutralization by a therapeutic antibodyrdquo Nature vol 437 no7059 pp 764ndash769 2005
[46] G D Gromowski J T Roehrig M S Diamond J C LeeT J Pitcher and A D T Barrett ldquoMutations of an antibodybinding energy hot spot on domain III of the dengue 2 envelopeglycoprotein exploited for neutralization escaperdquo Virology vol407 no 2 pp 237ndash246 2010
[47] J T Roehrig R A Bolin andR G Kelly ldquoMonoclonal antibodymapping of the envelope glycoprotein of the dengue 2 virusJamaicardquo Virology vol 246 no 2 pp 317ndash328 1998
[48] R de Alwis S A Smith N P Olivarez et al ldquoIdentification ofhumanneutralizing antibodies that bind to complex epitopes ondengue virionsrdquo Proceedings of the National Academy of Sciencesof the United States of America vol 109 no 19 pp 7439ndash74442012
[49] S K Austin K A Dowd B Shrestha et al ldquoStructural basis ofdifferential neutralization of DENV-1 genotypes by an antibodythat recognizes a cryptic epitoperdquo PLoS Pathogens vol 8 no 10Article ID e1002930 2012
[50] M Throsby C Geuijen J Goudsmit et al ldquoIsolation and char-acterization of human monoclonal antibodies from individualsinfected with West Nile Virusrdquo Journal of Virology vol 80 no14 pp 6982ndash6992 2006
[51] A P Goncalvez R H Purcell and C-J Lai ldquoEpitope deter-minants of a chimpanzee Fab antibody that efficiently cross-neutralizes dengue type 1 and type 2 viruses map to inside andin close proximity to fusion loop of the dengue type 2 virusenvelope glycoproteinrdquo Journal of Virology vol 78 no 23 pp12919ndash12928 2004
[52] J T Roehrig J H Mathews and D W Trent ldquoIdentification ofepitopes on the E glycoprotein of Saint Louis encephalitis virususing monoclonal antibodiesrdquo Virology vol 128 no 1 pp 118ndash126 1983
[53] J T Roehrig L A Staudinger A R Hunt J H Mathews andC D Blair ldquoAntibody prophylaxis and therapy for flavivirusencephalitis infectionsrdquo Annals of the New York Academy ofSciences vol 951 pp 286ndash297 2001
[54] A P Goncalvez C-H Chien K Tubthong et al ldquoHumanizedmonoclonal antibodies derived from chimpanzee fabs protectagainst Japanese encephalitis virus in vitro and in vivordquo Journalof Virology vol 82 no 14 pp 7009ndash7021 2008
[55] M Arakawa T Yamashiro G-I Uechi M Tadano and ANishizono ldquoConstruction of humanFab (1205741120581) library and iden-tification of human monoclonal Fab possessing neutralizingpotency against Japanese encephalitis virusrdquo Microbiology andImmunology vol 51 no 6 pp 617ndash625 2007
[56] L Warter C Y Lee R Thiagarajan et al ldquoChikungunya virusenvelope-specific human monoclonal antibodies with broadneutralization potencyrdquo Journal of Immunology vol 186 no 5pp 3258ndash3264 2011
[57] P Pal K A Dowd J D Brien et al ldquoDevelopment of a highlyprotective combination monoclonal antibody therapy againstchikungunya virusrdquo PLoS Pathogens vol 9 no 4 Article IDe1003312 2013
[58] A Bertolotti-Ciarlet J Smith K Strecker et al ldquoCellular local-ization and antigenic characterization of Crimean-Congo hem-orrhagic fever virus glycoproteinsrdquo Journal of Virology vol 79no 10 pp 6152ndash6161 2005
[59] J K Battles and J M Dalrymple ldquoGenetic variation amonggeographic isolates of Rift Valley fever virusrdquo American Journalof Tropical Medicine and Hygiene vol 39 no 6 pp 617ndash6311988
[60] B AThibodeaux N C Garbino NM Liss et al ldquoA humanizedIgG but not IgM antibody is effective in prophylaxis and therapyof yellow fever infection in an AG12917D-204 peripheralchallenge mouse modelrdquo Antiviral Research vol 94 no 1 pp1ndash8 2012
BioMed Research International 19
[61] S Daffis R E Kontermann J Korimbocus H Zeller H-DKlenk and J Ter Meulen ldquoAntibody responses against wild-type yellow fever virus and the 17D vaccine strain charac-terization with human monoclonal antibody fragments andneutralization escape variantsrdquoVirology vol 337 no 2 pp 262ndash272 2005
[62] L N Levanov L E Matveev E P Goncharova et al ldquoChimericantibodies against tick-borne encephalitis virusrdquo Vaccine vol28 no 32 pp 5265ndash5271 2010
[63] A R Hunt S Frederickson C Hinkel K S Bowdish and J TRoehrig ldquoA humanized murine monoclonal antibody protectsmice either before or after challenge with virulent Venezuelanequine encephalomyelitis virusrdquo Journal of General Virologyvol 87 no 9 pp 2467ndash2476 2006
[64] J M da Silva Voorham I A Rodenhuis-Zybert N V A Nunezet al ldquoAntibodies against the envelope glycoprotein promoteinfectivity of immature dengue virus serotype 2rdquo PLoS ONEvol 7 no 3 Article ID e29957 2012
[65] T C Pierson Q Xu S Nelson et al ldquoThe stoichiometry ofantibody-mediated neutralization and enhancement of westnile virus infectionrdquoCell Host andMicrobe vol 1 no 2 pp 135ndash145 2007
[66] G Sautto N Mancini R A Diotti L Solforosi M Clementiand R Burioni ldquoAnti-hepatitis C virus E2 (HCVE2) glyco-proteinmonoclonal antibodies andneutralization interferencerdquoAntiviral Research vol 96 no 1 pp 82ndash89 2012
[67] R Burioni Y Matsuura N Mancini et al ldquoDiverging effectsof human recombinant anti-hepatitis C virus (HCV) antibodyfragments derived from a single patient on the infectivityof a vesicular stomatitis virusHCV pseudotyperdquo Journal ofVirology vol 76 no 22 pp 11775ndash11779 2002
[68] R J Phillpotts J R Stephenson and J S Porterfield ldquoAntibody-dependent enhancement of tick-borne encephalitus virus infec-tivityrdquo Journal of General Virology vol 66 no 8 pp 1831ndash18371985
[69] K A Dowd and T C Pierson ldquoAntibody-mediated neutraliza-tion of flaviviruses a reductionist viewrdquo Virology vol 411 no 2pp 306ndash315 2011
[70] A K I Falconar ldquoMonoclonal antibodies that bind to commonepitopes on the dengue virus type 2 nonstructural-1 andenvelope glycoproteins display weak neutralizing activity anddifferentiated responses to virulent strains implications forpathogenesis and vaccinesrdquo Clinical and Vaccine Immunologyvol 15 no 3 pp 549ndash561 2008
[71] K Meyer A Banerjee S E Frey R B Belshe and R Ray ldquoAweak neutralizing antibody response to hepatitis C virus enve-lope glycoprotein enhances virus infectionrdquo PLoS ONE vol 6no 8 Article ID e23699 2011
[72] R Burioni R AWilliamson P P Sanna F E Bloom and D RBurton ldquoRecombinant human Fab to glycoprotein D neutral-izes infectivity and prevents cell-to-cell transmission of herpessimplex viruses 1 and 2 in vitrordquo Proceedings of the NationalAcademy of Sciences of the United States of America vol 91 no1 pp 355ndash359 1994
[73] M S Rolph A Zaid N E Rulli and S Mahalingam ldquoDown-regulation of interferon-120573 in antibody-dependent enhancementof dengue viral infections of human macrophages is dependenton interleukin-6rdquo Journal of Infectious Diseases vol 204 no 3pp 489ndash491 2011
[74] S S Whitehead J E Blaney A P Durbin and B R MurphyldquoProspects for a dengue virus vaccinerdquoNature ReviewsMicrobi-ology vol 5 no 7 pp 518ndash528 2007
[75] S-M Lok V Kostyuchenko G ENybakken et al ldquoBinding of aneutralizing antibody to dengue virus alters the arrangement ofsurface glycoproteinsrdquoNature Structural andMolecular Biologyvol 15 no 3 pp 312ndash317 2008
[76] A P Goncalvez R Men C Wernly R H Purcell and C-J Lai ldquoChimpanzee Fab fragments and a derived humanizedimmunogiobulin G1 antibody that efficiently cross-neutralizedengue type 1 and type 2 virusesrdquo Journal of Virology vol 78no 23 pp 12910ndash12918 2004
[77] WM P BWahala A A Kraus L B HaymoreM A Accavitti-Loper and A M de Silva ldquoDengue virus neutralization byhuman immune sera role of envelope protein domain III-reactive antibodyrdquo Virology vol 392 no 1 pp 103ndash113 2009
[78] W D Crill H R Hughes M J Delorey and G-J J ChangldquoHumoral immune responses of dengue fever patients usingepitope-specific serotype-2 virus-like particle antigensrdquo PLoSONE vol 4 no 4 Article ID e4991 2009
[79] C Setthapramote T Sasaki O Puiprom et al ldquoHuman mono-clonal antibodies to neutralize all dengue virus serotypes usinglymphocytes from patients at acute phase of the secondaryinfectionrdquo Biochemical and Biophysical Research Communica-tions vol 423 no 4 pp 867ndash872 2012
[80] M Beltramello K L Williams C P Simmons et al ldquoThehuman immune response to dengue virus is dominated byhighly cross-reactive antibodies endowed with neutralizing andenhancing activityrdquo Cell Host andMicrobe vol 8 no 3 pp 271ndash283 2010
[81] C M Midgley M Bajwa-Joseph S Vasanawathana et al ldquoAnin-depth analysis of original antigenic sin in dengue virusinfectionrdquo Journal of Virology vol 85 no 22 pp 410ndash421 2011
[82] B Kaufmann M R Vogt J Goudsmit et al ldquoNeutralizationof West Nile virus by cross-linking of its surface proteins withFab fragments of the human monoclonal antibody CR4354rdquoProceedings of the National Academy of Sciences of the UnitedStates of America vol 107 no 44 pp 18950ndash18955 2010
[83] B Spurrier J M Sampson M Totrov et al ldquoStructural analysisof human and macaque mAbs 2909 and 25B implicationsfor the configuration of the quaternary neutralizing epitope ofHIV-1 gp120rdquo Structure vol 19 no 5 pp 691ndash699 2011
[84] J J SchlesingerMWBrandriss andT PMonath ldquoMonoclonalantibodies distinguish between wild and vaccine strains ofyellow fever virus by neutralization hemagglutination inhibi-tion and immune precipitation of the virus envelope proteinrdquoVirology vol 125 no 1 pp 8ndash17 1983
[85] M W Brandriss J J Schlesinger E E Walsh and M BrisellildquoLethal 17D yellow fever encephalitis in mice I Passive protec-tion by monoclonal antibodies to the envelope proteins of 17Dyellow fever and dengue 2 virusesrdquo Journal of General Virologyvol 67 no 2 pp 229ndash234 1986
[86] H Cho and M S Diamond ldquoImmune responses to West Nilevirus infection in the central nervous systemrdquo Viruses vol 4no 12 pp 3812ndash3830 2012
[87] D W Beasley A D Barrett and R B Tesh ldquoResurgence ofWest Nile neurologic disease in the United States in 2012 whathappened What needs to be donerdquo Antiviral Research vol 99no 1 pp 1ndash5 2013
[88] M Throsby J Ter Meulen C Geuijen J Goudsmit and J DeKruif ldquoMapping and analysis of West Nile virus-specific mon-oclonal antibodies prospects for vaccine developmentrdquo ExpertReview of Vaccines vol 6 no 2 pp 183ndash191 2007
[89] M S Diamond T G Roberts D Edgil B Lu J Ernst and EHarris ldquoModulation of dengue virus infection in human cells
20 BioMed Research International
by alpha beta and gamma interferonsrdquo Journal of Virology vol74 no 11 pp 4957ndash4966 2000
[90] Z Shimoni M J Niven S Pitlick and S Bulvik ldquoTreatmentof West Nile virus encephalitis with intravenous immunoglob-ulinrdquo Emerging Infectious Diseases vol 7 no 4 article 759 2001
[91] AHamdan PGreen EMendelsonM R Kramer S Pitlik andM Weinberger ldquoPossible benefit of intravenous immunoglob-ulin therapy in a lung transplant recipient with West Nile virusencephalitisrdquoTransplant InfectiousDisease vol 4 no 3 pp 160ndash162 2002
[92] M Haley A S Retter D Fowler J Gea-Banacloche and NP OrsquoGrady ldquoThe role for intravenous immunoglobulin in thetreatment of West Nile virus encephalitisrdquo Clinical InfectiousDiseases vol 37 no 6 pp e88ndashe90 2003
[93] K M Chung G E Nybakken B S Thompson et al ldquoAnti-bodies against West Nile virus nonstructural protein NS1prevent lethal infection through Fc 120574 receptor-dependent and-independent mechanismsrdquo Journal of Virology vol 80 no 3pp 1340ndash1351 2006
[94] E A Henchal L S Henchal and J J Schlesinger ldquoSynergisticinteractions of anti-NS1 monoclonal antibodies protect pas-sively immunized mice from lethal challenge with dengue 2virusrdquo Journal of General Virology vol 69 no 8 pp 2101ndash21071988
[95] J T Roehrig ldquoAntigenic structure of flavivirus proteinsrdquoAdvances in Virus Research vol 59 pp 141ndash175 2003
[96] M D Sanchez T C Pierson D McAllister et al ldquoCharacteri-zation of neutralizing antibodies to West Nile virusrdquo Virologyvol 336 no 1 pp 70ndash82 2005
[97] J H Beigel J L Nordstrom S R Pillemer et al ldquoSafety andpharmacokinetics of single intravenous dose of MGAWN1 anovel monoclonal antibody to West Nile virusrdquo AntimicrobialAgents and Chemotherapy vol 54 no 6 pp 2431ndash2436 2010
[98] C A Smeraski V Siddharthan and J D Morrey ldquoTreatmentof spatial memory impairment in hamsters infected with WestNile virus using a humanizedmonoclonal antibodyMGAWN1rdquoAntiviral Research vol 91 no 1 pp 43ndash49 2011
[99] D Lelli AMoreno E Brocchi et al ldquoWestNile virus character-ization and diagnostic applications of monoclonal antibodiesrdquoVirology Journal vol 9 article 81 2012
[100] T S Gritsun V A Lashkevich and E A Gould ldquoTick-borneencephalitisrdquo Antiviral Research vol 57 no 1-2 pp 129ndash1462003
[101] D Ruzek T S Gritsun N L Forrester et al ldquoMutations inthe NS2B and NS3 genes affect mouse neuroinvasiveness of aWestern European field strain of tick-borne encephalitis virusrdquoVirology vol 374 no 2 pp 249ndash255 2008
[102] M Ecker S L Allison T Meixner and F X Heinz ldquoSequenceanalysis and genetic classification of tick-borne encephalitisviruses from Europe and Asiardquo Journal of General Virology vol80 no 1 pp 179ndash185 1999
[103] H Holzmann M S Vorobyova I P Ladyzhenskaya et alldquoMolecular epidemiology of tick-borne encephalitis viruscross-protection between European and Far Eastern subtypesrdquoVaccine vol 10 no 5 pp 345ndash349 1992
[104] C W Mandl ldquoSteps of the tick-borne encephalitis virus repli-cation cycle that affect neuropathogenesisrdquo Virus Research vol111 no 2 pp 161ndash174 2005
[105] A V Atrasheuskaya T M Fredeking and G M IgnatyevldquoChanges in immune parameters and their correction in humancases of tick-borne encephalitisrdquo Clinical and ExperimentalImmunology vol 131 no 1 pp 148ndash154 2003
[106] K L Mansfield N Johnson L P Phipps J R Stephenson AR Fooks and T Solomon ldquoTick-borne encephalitis virusmdashareview of an emerging zoonosisrdquo Journal of General Virologyvol 90 no 8 pp 1781ndash1794 2009
[107] DWVaughn andCHHoke Jr ldquoThe epidemiology of Japaneseencephalitis prospects for preventionrdquo Epidemiologic Reviewsvol 14 pp 197ndash221 1992
[108] T Solomon H Ni D W C Beasley M Ekkelenkamp M JCardosa and A D T Barrett ldquoOrigin and evolution of Japaneseencephalitis virus in Southeast Asiardquo Journal of Virology vol 77no 5 pp 3091ndash3098 2003
[109] M R Wilson ldquoEmerging viral infectionsrdquo Current Opinion inNeurology vol 26 no 3 pp 301ndash306 2013
[110] J N Hanna S A Ritchie D A Phillips et al ldquoAn outbreakof Japanese encephalitis in the Torres Strait Australia 1995rdquoMedical Journal of Australia vol 165 no 5 pp 256ndash260 1996
[111] J N Hanna S A Ritchie D A Phillips et al ldquoJapanese ence-phalitis in north Queensland Australia 1998rdquo Medical Journalof Australia vol 170 no 11 pp 533ndash536 1999
[112] SGeorgeMGourie-Devi and J A Rao ldquoIsolation ofWestNilevirus from the brains of children who had died of encephalitisrdquoBulletin of theWorldHealthOrganization vol 62 no 6 pp 879ndash882 1984
[113] A K Gupta A A Koshy and V J Lad ldquoEnhanced protec-tion of mice against Japanese encephalitis virus infection bycombinations ofmonoclonal antibodies to glycoprotein ErdquoActaVirologica vol 55 no 2 pp 165ndash168 2011
[114] T H Lee B-H Song S-I Yun et al ldquoA cross-protective mAbrecognizes a novel epitope within the flavivirus NS1 proteinrdquoJournal of General Virology vol 93 no 1 pp 20ndash26 2012
[115] S C Weaver ldquoHost range amplification and arboviral diseaseemergencerdquo Archives of Virology Supplementum no 19 pp 33ndash44 2005
[116] M Solignat B Gay S Higgs L Briant and C Devaux ldquoRepli-cation cycle of chikungunya a re-emerging arbovirusrdquoVirologyvol 393 no 2 pp 183ndash197 2009
[117] O Schwartz and M L Albert ldquoBiology and pathogenesis ofchikungunya virusrdquo Nature Reviews Microbiology vol 8 no 7pp 491ndash500 2010
[118] W Akahata Z-Y Yang H Andersen et al ldquoA virus-like particlevaccine for epidemic Chikungunya virus protects nonhumanprimates against infectionrdquo Nature Medicine vol 16 no 3 pp334ndash338 2010
[119] M Kielian and A Helenius ldquopH-Iinduced alterations in thefusogenic spike protein of Semliki Forest virusrdquo Journal of CellBiology vol 101 no 6 pp 2284ndash2291 1985
[120] J White and A Helenius ldquopH-Dependent fusion between theSemliki Forest virus membrane and liposomesrdquo Proceedings ofthe National Academy of Sciences of the United States of Americavol 77 no 6 pp 3273ndash3277 1980
[121] J Dubuisson and C M Rice ldquoSindbis virus attachment isola-tion and characterization of mutants with impaired binding tovertebrate cellsrdquo Journal of Virology vol 67 no 6 pp 3363ndash33741993
[122] C Y Lee Y-W Kam J Fric et al ldquoChikungunya virus neutral-ization antigens and direct cell-to-cell transmission are revealedby human antibody-escapemutantsrdquo PLoS Pathogens vol 7 no12 Article ID e1002390 2011
[123] J E Voss M-C Vaney S Duquerroy et al ldquoGlycoproteinorganization of Chikungunya virus particles revealed by X-raycrystallographyrdquo Nature vol 468 no 7324 pp 709ndash712 2010
BioMed Research International 21
[124] L Li J Jose Y Xiang R J Kuhn and M G Rossmann ldquoStruc-tural changes of envelope proteins during alphavirus fusionrdquoNature vol 468 no 7324 pp 705ndash708 2010
[125] J Fric S Bertin-Maghit C I Wang A Nardin and L WarterldquoUse of human monoclonal antibodies to treat Chikungunyavirus infectionrdquo Journal of Infectious Diseases vol 207 no 2 pp319ndash322 2013
[126] P V Aguilar J G Estrada-Franco R Navarro-Lopez C FerroA D Haddow and S C Weaver ldquoEndemic Venezuelanequine encephalitis in the Americas hidden under the dengueumbrellardquo Future Virology vol 6 no 6 pp 721ndash740 2011
[127] S C Weaver R Winegar I D Manger and N L ForresterldquoAlphaviruses population genetics and determinants of emer-gencerdquo Antiviral Research vol 94 no 3 pp 242ndash257 2012
[128] S Paessler and S C Weaver ldquoVaccines for Venezuelan equineencephalitisrdquo Vaccine vol 27 no 4 pp D80ndashD85 2009
[129] R J Phillpotts ldquoVenezuelan equine encephalitis virus complex-specific monoclonal antibody provides broad protection inmurine models against airborne challenge with viruses fromserogroups I II and IIIrdquoVirus Research vol 120 no 1-2 pp 107ndash112 2006
[130] R J Phillpotts L D Jones and S C Howard ldquoMonoclonalantibody protectsmice against infection anddiseasewhen giveneither before or up to 24 h after airborne challenge with virulentVenezuelan equine encephalitis virusrdquoVaccine vol 20 no 11-12pp 1497ndash1504 2002
[131] J H Mathews J T Roehrig and D W Trent ldquoRole of com-plement and the Fc portion of immunoglobulin G in immunityto Venezuelan equine encephalomyelitis virus infection withglycoprotein-specific monoclonal antibodiesrdquo Journal of Virol-ogy vol 55 no 3 pp 594ndash600 1985
[132] L M OrsquoBrien C D Underwood-Fowler S A Goodchild AL Phelps and R J Phillpotts ldquoDevelopment of a novel mon-oclonal antibody with reactivity to a wide range of Venezuelanequine encephalitis virus strainsrdquoVirology Journal vol 6 article206 2009
[133] L M OrsquoBrien S A Goodchild R J Phillpotts and S D Per-kins ldquoA humanised murine monoclonal antibody protectsmice from Venezuelan equine encephalitis virus Evergladesvirus and Mucambo virus when administered up to 48h afterairborne challengerdquo Virology vol 426 no 2 pp 100ndash105 2012
[134] N Clementi N Mancini L Solforosi M Castelli M Clementiand R Burioni ldquoPhage display-based strategies for cloning andoptimization of monoclonal antibodies directed against humanpathogensrdquo International Journal of Molecular Sciences vol 13no 7 pp 8273ndash8292 2012
[135] A R Hunt R A Bowen S Frederickson T Maruyama JT Roehrig and C D Blair ldquoTreatment of mice with humanmonoclonal antibody 24h after lethal aerosol challenge withvirulent Venezuelan equine encephalitis virus prevents diseasebut not infectionrdquo Virology vol 414 no 2 pp 146ndash152 2011
[136] W-G Hu A L Phelps S Jager et al ldquoA recombinant human-ized monoclonal antibody completely protects mice againstlethal challenge with Venezuelan equine encephalitis virusrdquoVaccine vol 28 no 34 pp 5558ndash5564 2010
[137] T Rulker L Voss P Thullier et al ldquoIsolation and characterisa-tion of a human-like antibody fragment (scFv) that inactivatesVEEV in vitro and in vivordquo PLoS ONE vol 7 no 5 Article IDe37242 2012
[138] C T Walter and J N Barr ldquoRecent advances in the molecularand cellular biology of bunyavirusesrdquo Journal of General Virol-ogy vol 92 no 11 pp 2467ndash2484 2011
[139] C A Whitehouse ldquoCrimean-Congo hemorrhagic feverrdquo Anti-viral Research vol 64 no 3 pp 145ndash160 2004
[140] M Mertens K Schmidt A Ozkul and M H Groschup ldquoTheimpact of Crimean-Congo hemorrhagic fever virus on publichealthrdquo Antiviral Research vol 98 no 2 pp 248ndash260 2013
[141] A A Ahmed J M McFalls C Hoffmann et al ldquoPresence ofbroadly reactive and group-specific neutralizing epitopes onnewly described isolates of Crimean-Congo hemorrhagic fevervirusrdquo Journal of General Virology vol 86 no 12 pp 3327ndash33362005
[142] R Hewson A Gmyl L Gmyl et al ldquoEvidence of segment reas-sortment in Crimean-Congo haemorrhagic fever virusrdquo Journalof General Virology vol 85 no 10 pp 3059ndash3070 2004
[143] N K Blackburn T G Besselaar A J Shepherd and R Swane-poel ldquoPreparation and use of monoclonal antibodies for iden-tifying Crimean-Congo hemorrhagic fever virusrdquo AmericanJournal of Tropical Medicine and Hygiene vol 37 no 2 pp 392ndash397 1987
[144] T Ikegami ldquoMolecular biology and genetic diversity of RiftValley fever virusrdquoAntiviral Research vol 95 no 3 pp 293ndash3102012
[145] KKeegan andM S Collett ldquoUse of bacterial expression cloningto define the amino acid sequences of antigenic determinantson the G2 glycoprotein of Rift Valley fever virusrdquo Journal ofVirology vol 58 no 2 pp 263ndash270 1986
[146] T G Besselaar and N K Blackburn ldquoTopological mapping ofantigenic sites on the Rift Valley fever virus envelope glycopro-teins using monoclonal antibodiesrdquo Archives of Virology vol121 no 1ndash4 pp 111ndash124 1991
[147] L G Presta ldquoEngineering of therapeutic antibodies to mini-mize immunogenicity and optimize functionrdquo Advanced DrugDelivery Reviews vol 58 no 5-6 pp 640ndash656 2006
Submit your manuscripts athttpwwwhindawicom
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anatomy Research International
PeptidesInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporation httpwwwhindawicom
International Journal of
Volume 2014
Zoology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Molecular Biology International
GenomicsInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioinformaticsAdvances in
Marine BiologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Signal TransductionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Evolutionary BiologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Biochemistry Research International
ArchaeaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Genetics Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in
Virolog y
Hindawi Publishing Corporationhttpwwwhindawicom
Nucleic AcidsJournal of
Volume 2014
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Enzyme Research
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Microbiology

BioMed Research International 15
murine and humanized versions of the VH and VL domainsThis analysis showed that the number of human T-cellepitopes within the humanized Ab had been substantiallyreduced indicating that Hu1A3B-7 may have low immuno-genicity in vivo [133]
Similarly Hunt et al described a humanizedmurinemAbderived from the 3B4C-4 mAb recognizing the E2c epitope(amino acid residues 182ndash209) and following named Hy4-26C showing neutralizing activity similar to the murinemAb Moreover it was protective (70ndash100 of survival) at100 ng dose in mice animal model intraperitoneal inoculatedand at 500120583g for intranasal challenge (80) Therapeuticstudies in mice revealed that Hy4-26C was able to cure miceup to 24 h following infection at 10 120583g on the contrary themouse mAb when given 1 hour after virus challenge [63]
In another work Hunt et al isolated two neutralizinghumanized Fabs F5 and L1A7 employing a blocking strategy[134 135] The anti-E2 specific F5 IgG had a 70 PRNTendpoint of 10 ngmL equivalent to that described for themost effective neutralizing anti-VEEVE2mAbs In particularthe E1-specific hFab L1A7 had a PRNT endpoint of 3120583gmL300-fold lower than F5
Further studies revealed that F5 epitope is located inthe 115ndash199 amino acid region Moreover F5 IgG had potentability to protect mice from infection by either route whenadministered 24 h before exposure however mice treated24 h and 48 h after aerosol exposure developed central ner-vous system infections but exhibited no clinical signs ofdisease [135]
Hu et al reported that passive immunization with thehumanized chimericmousemAbhu1A4A1IgG1-2A inmice at50 120583g 24 h before or after virulent VEEV challenge providedcomplete protection indicating that hu1A4A1IgG1-2A haspotent prophylactic and therapeutic effects against VEEVinfection [136]
Anti-VEEV mAbs isolated from a non-human primategene library has also been reported In particular thehumanized chimeric scFv-Fc ToR67-3B4 recognized viableas well as formalin and 120573-propiolactone inactivated virusparticles It detected specifically the viral E1 envelope proteinof VEEV but did not react with reduced viral glycopro-tein preparations suggesting that recognition depends uponconformational epitopes The recombinant Ab was able todetect multiple VEEV subtypes and displayed only marginalcross-reactivity to otherAlphavirus species except for Easternequine encephalitis virus (EEEV) In addition the scFv-Fcfusion described here might be of therapeutic use since itsuccessfully inactivated VEEV in a murine disease modelIn particular when the recombinant Ab was administered 6hours after challenge 80 to 100 of mice survived lethalVEEV IAB or IE infection Forty to sixty percent of micesurvived when scFv-Fc ToR67-3B4 was applied 6 hours afterchallenge with VEEV subtypes II and former IIIA [137]
4 Bunyaviridae
TheBunyaviridae family contains human arboviruses belong-ing to the Orthobunyavirus Phlebovirus and Nairovirus
genera Bunyaviridae includes enveloped viruses with a frag-mented single-stranded RNA genome of negative polarityTheir tripartite genome consists of a small (S) a medium(M) and a large (L) fragment and the envelope glycoproteinsGn and Gc that are cleaved out of a polyprotein synthesizedby the M fragment These glycoproteins have been provedto mediate the formation of the virus particle to play a rolein the interaction with cell surface receptors to mediate theentry of the virus into cells and to serve as targets for themajority of neutralizing Abs described so far
Orthobunyavirus transmitted through mosquitoes ormidges vectors are divided in 18 serogroups based on cross-titrations in haemagglutination inhibition assays and neutral-ization assays and correlating with main vector preferencesHowever the most clinically important viruses belong onlyto two serogroups the California encephalitis and the Simbuserogroups
Crimean-Congo hemorrhagic fever virus (CCHFV) isconsidered the only one of clinical relevance within theNairovirus genus which uses tick as main vector and is divid-ed in seven serogroups while Toscana virus and Rift Valleyfever virus (RVFV) which are transmitted by sandflies andmosquitoes respectively belong to the Phlebovirus genus[138]
MAbs against theNairovirusCCHFV and the PhlebovirusRVFV have been described
41 Crimean-Congo Hemorrhagic Fever Virus (CCHFV)Crimean-Congo Hemorrhagic Fever Virus (CCHFV) dis-tribution covers the greatest geographic range of any tick-borne virus known as viral isolation andor disease hasbeen reported from more than 30 countries in SoutheasternEurope Africa Asia and Middle East [139 140] Namelyfrom 1953 to 2010 about 6000 human cases were reportedin Southeastern Europe CCHFV causes sporadic outbreakswith mortality rates ranging from 10 to 80 [141 142] Asignificant variation in the time of incubation has beendescribed [139] Prehemorrhagic symptoms include highfever chills headache photophobia and back and abdominalpains Among other symptoms neuropsychiatric changeshave been reported in some patients In severe cases 3ndash6 daysafter the onset of the disease hemorrhagic symptoms occurwith petechiae ecchymosis bleeding in the form of melenahematemesis epistaxis [139]
In comparisonwith the nucleocapsid proteins inCCHFVGn and Gc show a higher degree of antigenic variabilityprobably due to their exposition to the host immune systemhypothesis corroborated in the description by Hewson et alin 2004 of four M segment phylogenic groups namely M1M2 M3 and M4 [141 142]
411 mAbs against CCHFV Blackburn et al firstly describedin 1987 murine anti-CCHFV mAbs able to recognize nucle-ocapsid proteins but no neutralizing activity has beendescribed thus far [143]
On the other hand Bertolotti-Ciarlet et al produced apanel of murine mAbs recognizing conformational epitopes
16 BioMed Research International
within theGn andGc glycoproteins expressed by the CCHFVstrain IbAr10200 The effectiveness of the mAbs was studiedby performing PRNT80 and none of the anti-Gn mAbsshowed neutralizing activity although many of the anti-GcmAbs neutralized IbAr10200 strain in vitro with clones 8A15E3 12A9 6C2 and 9H3 showing activity at a gt5120 dilutionMAbs 11E7 and 30F7 exhibited neutralizing activity at 2560dilution A suckling mice protection test was thereforeperformed for these mAbs The previously mentionedanti-Gc Abs were capable to protect mice to an appreciabledegree when applied 24 hours before and in a weakermanner 24 hours after virus challenge Many of the anti-GnmAbs were able to confer significant protection to IbAr10200both 24 hours before and after virus administration Therelevant effectiveness of the anti-Gn mAbs 6B12 10E1113G8 and 10G4 suggests that these mAbs may possess someneutralizing activity due to Ab-based effector mechanisms(eg ADCC) [58]
In further studies the same group characterized thebroadly cross-reactive neutralizing activity of the previouslydescribed murine mAb 11E7 able to bind conformationalepitopes within the C-terminal region of the Gn glycoproteinof all CCHFVM groups [58 141]
42 Rift Valley Fever Virus (RVFV) Similarly to CCHFV RiftValley Fever Virus (RVFV) causes sporadic outbreaks Thelargest epidemic occurred in Egypt in 1977 with an estimated200000 infections 18000 patients manifesting symptomsand 600 fatal prognoses [144] Symptoms include hemor-rhagic episodes fever encephalitis and blindness [145]
The major target of the humoral immune response is theRVFV glycoproteins Gc (G1) and Gn (G2) ranging fromnucleotides 480ndash2090 and 2091ndash3614 respectively [144]
421 mAbs against RVFV Within G2 glycoprotein are des-cribed three neutralizing antigenic sites namely epitope Inucleotides 792ndash893 (amino acid residues 258ndash291) epitopeII nucleotides 1164ndash1196 (amino acid residues 382ndash392) andepitope IV nucleotides 858ndash917 (amino acid residues 280ndash299) [59 145]
Despite the high degree of variability observed inCCHFVglycoproteins RVFV G2 presented an unexpected high con-servation among 22 isolates [59] In the same study onlythe LUNYO and 900060 isolates were resistant to in vitroneutralization by murine mAb 4D4 (recognizing epitope II)All other isolates were neutralized with PRNT80 titers from10240 togt81920On the other hand only SNS isolate showedreduced sensitivity to 4ndash39-CC (epitope IV) All other isolateswere neutralized with PRNT80 titers from 20480 to gt81920Protection studies on mice would be of interest for thedevelopment of a passive immunization prophylaxis [59]
Protection studies from lethal South African RVFVAN1830 strain infection were successfully performed on abattery of anti-G1 and anti-G2 mAbs produced by Besselaarand Blackburn with the strongly neutralizing anti-G1 mAb3E5 and anti-G2mAb 9C4 being the most effective [146]The9C4 antigenic area maps within G2 epitope I and a broadlyneutralizing activity may be investigated
5 Conclusions
Arbovirus infections have acquired increasing interest giventhe augmented globalization and tourism movement all overthe world that could be at the basis of epidemic eventsFurthermore the same vectors can sometimes transmit sev-eral arboviruses concomitantly complicating the diagnosis aswell as the therapy For example confusing mixed epidemicshave occasionally been described such as YFV plus CHIKVDENVplus CHIKV ormore recentlyPlasmodium falciparummalaria plus DENV1 and CHIKV on Madagascarrsquos east coast[1]
In the last decade several efforts have been employed inthe development of effective immunogens and therapeuticsgiving the current absence of specific antiviral drugs as wellas effective vaccines for themost diffused arbovirus infections(ie DENV andWNV)
However at the moment mosquito control is the bestavailable method for preventing arbovirus infections Thusto control the emerging public health problem of arbovirusinfections new antiviral therapeutic strategies that providepotent and broadly cross-protective immunity (especially forflavivirus infections) are an urgent globally medical need
As described in this review the majority of highlyneutralizing mAbs are of murine origin and indeed theirutility in the treatment of these infections is limited In facttheir employment can elicit an immunogenic response (iehuman anti-mouse Abs HAMA) the therapeutic efficacycould be reduced by a relatively faster clearance in humans(compared to human Abs) potentially exacerbated by theHAMA response Finally murine Abs exhibit relatively weakeffector functions (eg ADCC) compared to human AbsIndeed chimerization humanization or better the isolationand characterization of fully human broadly neutralizingmAbs would be the best choice for their possible use in afuture mAb-based therapy against arbovirus infections [147]
Finally possible ADE of infection mechanisms should beevaluated before considering a mAb as a possible candidatetherapeutic in a post-exposure setting as well as in the devel-opment of new Ab-eliciting immunogens
Conflict of Interests
The authors declare no conflict of interests in particular withOmriGam and any other cited Ab-based preparation men-tioned in the paper
References
[1] N Cleton M Koopmans J Reimerink G J Godeke and CReusken ldquoCome fly with me review of clinically importantarboviruses for global travelersrdquo Journal of Clinical Virology vol55 no 3 pp 191ndash203 2012
[2] F X Heinz H Holzmann A Essl and M Kundi ldquoField effec-tiveness of vaccination against tick-borne encephalitisrdquoVaccinevol 25 no 43 pp 7559ndash7567 2007
[3] A D Barrett and D E Teuwen ldquoYellow fever vaccinemdashhowdoes it work and why do rare cases of serious adverse events
BioMed Research International 17
take placerdquo Current Opinion in Immunology vol 21 no 3 pp308ndash313 2009
[4] S B Halstead and S J Thomas ldquoNew vaccines for Japaneseencephalitisrdquo Current Infectious Disease Reports vol 12 no 3pp 174ndash180 2010
[5] R Burioni M Perotti N Mancini and M Clementi ldquoPer-spectives for the utilization of neutralizing human monoclonalantibodies as anti-HCV drugsrdquo Journal of Hepatology vol 49no 2 pp 299ndash300 2008
[6] R A Diotti G A Sautto L Solforosi NManciniM Clementiand R Burioni ldquoNeutralization activity and kinetics of twobroad-range human monoclonal IgG1 derived from recombi-nant Fab fragments and directed against Hepatitis C virus E2glycoproteinrdquo New Microbiologica vol 35 no 4 pp 475ndash4792012
[7] R Burioni F Canducci N Mancini et al ldquoMolecular cloningof the first humanmonoclona antibodies neutralizing with highpotency Swine-origin Influenza A pandemic virus (S-OIV)rdquoNew Microbiologica vol 32 no 4 pp 319ndash324 2009
[8] P K Chandra S Hazari B Poat et al ldquoIntracytoplasmic stableexpression of IgG1 antibody targeting NS3 helicase inhibitsreplication of highly efficient hepatitis C Virus 2a clonerdquoVirology Journal vol 7 article 118 2010
[9] N Clementi D de Marco N Mancini et al ldquoA human mono-clonal antibody with neutralizing activity against highly diver-gent influenza subtypesrdquo PLoS ONE vol 6 no 12 Article IDe28001 2011
[10] D de Marco N Clementi N Mancini et al ldquoA non-VH1-69 heterosubtypic neutralizing human monoclonal antibodyprotects mice against H1N1 and H5N1 virusesrdquo PLoS ONE vol7 no 4 Article ID e34415 2012
[11] L Solforosi N Mancini F Canducci et al ldquoA phage displayvector optimized for the generation of human antibody com-binatorial libraries and the molecular cloning of monoclonalantibody fragmentsrdquoNewMicrobiologica vol 35 no 3 pp 289ndash294 2012
[12] E V Koonin and V V Dolja ldquoEvolution and taxonomy ofpositive-strand RNA viruses implications of comparative anal-ysis of amino acid sequencesrdquo Critical Reviews in Biochemistryand Molecular Biology vol 28 no 5 pp 375ndash430 1993
[13] J S Mackenzie D J Gubler and L R Petersen ldquoEmergingflaviviruses the spread and resurgence of Japanese encephalitisWest Nile and dengue virusesrdquo Nature Medicine vol 10 no 12pp S98ndashS109 2004
[14] T J SmithW E Brandt J L Swanson J MMcCown and E LBuescher ldquoPhysical and biological properties of dengue-2 virusand associated antigensrdquo Journal of Virology vol 5 no 4 pp524ndash532 1970
[15] J M Smit B Moesker I Rodenhuis-Zybert and J WilschutldquoFlavivirus cell entry and membrane fusionrdquo Viruses vol 3 no2 pp 160ndash171 2011
[16] R J Kuhn W Zhang M G Rossmann et al ldquoStructure ofdengue virus implications for flavivirus organization matura-tion and fusionrdquo Cell vol 108 no 5 pp 717ndash725 2002
[17] K Stiasny J H Aberle M Keller B Grubeck-Loebenstein andF X Heinz ldquoAge affects quantity but not quality of antibodyresponses after vaccination with an inactivated flavivirus vac-cine against tick-borne encephalitisrdquo PLoS ONE vol 7 no 3Article ID e34145 2012
[18] LThomasOVerlaetenACabie et al ldquoInfluence of the dengueserotype previous dengue infection and plasma viral load
on clinical presentation and outcome during a dengue-2 anddengue-4 co-epidemicrdquo American Journal of Tropical Medicineand Hygiene vol 78 no 6 pp 990ndash998 2008
[19] Y Zhang J Corver P R Chipman et al ldquoStructures of imma-ture flavivirus particlesrdquo EMBO Journal vol 22 no 11 pp2604ndash2613 2003
[20] A K I Falconar ldquoIdentification of an epitope on the denguevirus membrane (M) protein defined by cross-protective mon-oclonal antibodies design of an improved epitope sequencebased on common determinants present in both envelope (EandM) proteinsrdquo Archives of Virology vol 144 no 12 pp 2313ndash2330 1999
[21] K-J Huang Y-C Yang Y-S Lin et al ldquoThe dual-specific bind-ing of dengue virus and target cells for the antibody-dependentenhancement of dengue virus infectionrdquo Journal of Immunol-ogy vol 176 no 5 pp 2825ndash2832 2006
[22] A K I Falconar ldquoAntibody responses are generated to immun-odominant ELKKLE-typemotifs on the nonstructural-1 glyco-protein during live dengue virus infections inmice and humansimplications for diagnosis pathogenesis and vaccine designrdquoClinical and Vaccine Immunology vol 14 no 5 pp 493ndash5042007
[23] C W Mandl F Guirakhoo H Holzmann F X Heinz and CKunz ldquoAntigenic structure of the flavivirus envelope protein Eat the molecular level using the tick-borne encephalitis virus asa modelrdquo Journal of Virology vol 63 no 2 pp 564ndash571 1989
[24] F A Rey F X Heinz C Mandl C Kunz and S C HarrisonldquoThe envelope glycoprotein from tick-borne encephalitis virusat 2 A resolutionrdquo Nature vol 375 no 6529 pp 291ndash298 1995
[25] S L Allison K Stiasny K Stadler CWMandl and F XHeinzldquoMapping of functional elements in the stem-anchor regionof tick-borne encephalitis virus envelope protein Erdquo Journal ofVirology vol 73 no 7 pp 5605ndash5612 1999
[26] D E Klein J L Choi and S C Harrison ldquoStructure of a denguevirus envelope protein late-stage fusion intermediaterdquo Journal ofVirology vol 87 no 4 pp 2287ndash2293 2013
[27] S L Hanna T C Pierson M D Sanchez A A Ahmed M MMurtadha and R W Doms ldquoN-linked glycosylation of WestNile virus envelope proteins influences particle assembly andinfectivityrdquo Journal of Virology vol 79 no 21 pp 13262ndash132742005
[28] N Mancini G Sautto N Clementi et al ldquoNeutralization inter-fering antibodies a ldquonovelrdquo example of humoral immune dys-function facilitating viral escaperdquoViruses vol 4 no 9 pp 1731ndash1752 2012
[29] Y Modis S Ogata D Clements and S C Harrison ldquoVariablesurface epitopes in the crystal structure of dengue virus type3 envelope glycoproteinrdquo Journal of Virology vol 79 no 2 pp1223ndash1231 2005
[30] N Miller ldquoRecent progress in dengue vaccine research anddevelopmentrdquo Current Opinion in Molecular Therapeutics vol12 no 1 pp 31ndash38 2010
[31] J H Chavez J R Silva A A Amarilla and L T Moraes Figu-eiredo ldquoDomain III peptides from flavivirus envelope proteinare useful antigens for serologic diagnosis and targets forimmunizationrdquo Biologicals vol 38 no 6 pp 613ndash618 2010
[32] B Shrestha J D Brien S Sukupolvi-Petty et al ldquoThe develop-ment of therapeutic antibodies that neutralize homologous andheterologous genotypes of dengue virus type 1rdquo PLoS Pathogensvol 6 no 4 Article ID e1000823 2010
18 BioMed Research International
[33] G D Gromowski N D Barrett and A D T Barrett ldquoChar-acterization of dengue virus complex-specific neutralizing epi-topes on envelope protein domain III of dengue 2 virusrdquo Journalof Virology vol 82 no 17 pp 8828ndash8837 2008
[34] S Sukupolvi-Petty S K Austin W E Purtha et al ldquoType- andsubcomplex-specific neutralizing antibodies against domain IIIof dengue virus type 2 envelope protein recognize adjacentepitopesrdquo Journal of Virology vol 81 no 23 pp 12816ndash128262007
[35] S Sukupolvi-Petty S K Austin M Engle et al ldquoStructure andfunction analysis of therapeutic monoclonal antibodies againstdengue virus type 2rdquo Journal of Virology vol 84 no 18 pp9227ndash9239 2010
[36] Y-Q Deng J-X Dai G-H Ji et al ldquoA broadly flaviviruscross-neutralizingmonoclonal antibody that recognizes a novelepitope within the fusion loop of e proteinrdquo PLoS ONE vol 6no 1 Article ID e16059 2011
[37] J M Costin E Zaitseva K M Kahle et al ldquoMechanistic studyof broadly neutralizing human monoclonal antibodies againstdengue virus that target the fusion looprdquo Journal of Virologyvol 87 no 1 pp 52ndash66 2013
[38] CMMidgley A FlanaganH B Tran et al ldquoStructural analysisof a dengue cross-reactive antibody complexed with envelopedomain III reveals the molecular basis of cross-reactivityrdquoJournal of Immunology vol 188 no 10 pp 4971ndash4979 2012
[39] N J Moreland P Susanto E Lim et al ldquoPhage display appro-aches for the isolation of monoclonal antibodies against denguevirus envelope domain III from human and mouse derivedlibrariesrdquo International Journal ofMolecular Sciences vol 13 no3 pp 2618ndash2635 2012
[40] J J B Cockburn M E Navarro Sanchez N Fretes et alldquoMechanism of dengue virus broad cross-neutralization by amonoclonal antibodyrdquo Structure vol 20 no 2 pp 303ndash3142012
[41] R Rajamanonmani C Nkenfou P Clancy et al ldquoOn a mousemonoclonal antibody that neutralizes all four dengue virusserotypesrdquo Journal of General Virology vol 90 no 4 pp 799ndash809 2009
[42] L H Gould J Sui H Foellmer et al ldquoProtective and herapeuticcapacity of human single-chain Fv-Fc fusion proteins againstWest Nile virusrdquo Journal of Virology vol 79 no 23 pp 14606ndash14613 2005
[43] H Sultana H G Foellmer G Neelakanta et al ldquoFusion LoopPeptide of the West Nile virus envelope protein is essential forpathogenesis and is recognized by a therapeutic cross-reactivehuman monoclonal antibodyrdquo Journal of Immunology vol 183no 1 pp 650ndash660 2009
[44] T Oliphant M Engle G E Nybakken et al ldquoDevelopment ofa humanized monoclonal antibody with therapeutic potentialagainstWest Nile virusrdquoNature Medicine vol 11 no 5 pp 522ndash530 2005
[45] G E Nybakken T Oliphant S Johnson S Burke M S Dia-mond and D H Fremont ldquoStructural basis of West Nile virusneutralization by a therapeutic antibodyrdquo Nature vol 437 no7059 pp 764ndash769 2005
[46] G D Gromowski J T Roehrig M S Diamond J C LeeT J Pitcher and A D T Barrett ldquoMutations of an antibodybinding energy hot spot on domain III of the dengue 2 envelopeglycoprotein exploited for neutralization escaperdquo Virology vol407 no 2 pp 237ndash246 2010
[47] J T Roehrig R A Bolin andR G Kelly ldquoMonoclonal antibodymapping of the envelope glycoprotein of the dengue 2 virusJamaicardquo Virology vol 246 no 2 pp 317ndash328 1998
[48] R de Alwis S A Smith N P Olivarez et al ldquoIdentification ofhumanneutralizing antibodies that bind to complex epitopes ondengue virionsrdquo Proceedings of the National Academy of Sciencesof the United States of America vol 109 no 19 pp 7439ndash74442012
[49] S K Austin K A Dowd B Shrestha et al ldquoStructural basis ofdifferential neutralization of DENV-1 genotypes by an antibodythat recognizes a cryptic epitoperdquo PLoS Pathogens vol 8 no 10Article ID e1002930 2012
[50] M Throsby C Geuijen J Goudsmit et al ldquoIsolation and char-acterization of human monoclonal antibodies from individualsinfected with West Nile Virusrdquo Journal of Virology vol 80 no14 pp 6982ndash6992 2006
[51] A P Goncalvez R H Purcell and C-J Lai ldquoEpitope deter-minants of a chimpanzee Fab antibody that efficiently cross-neutralizes dengue type 1 and type 2 viruses map to inside andin close proximity to fusion loop of the dengue type 2 virusenvelope glycoproteinrdquo Journal of Virology vol 78 no 23 pp12919ndash12928 2004
[52] J T Roehrig J H Mathews and D W Trent ldquoIdentification ofepitopes on the E glycoprotein of Saint Louis encephalitis virususing monoclonal antibodiesrdquo Virology vol 128 no 1 pp 118ndash126 1983
[53] J T Roehrig L A Staudinger A R Hunt J H Mathews andC D Blair ldquoAntibody prophylaxis and therapy for flavivirusencephalitis infectionsrdquo Annals of the New York Academy ofSciences vol 951 pp 286ndash297 2001
[54] A P Goncalvez C-H Chien K Tubthong et al ldquoHumanizedmonoclonal antibodies derived from chimpanzee fabs protectagainst Japanese encephalitis virus in vitro and in vivordquo Journalof Virology vol 82 no 14 pp 7009ndash7021 2008
[55] M Arakawa T Yamashiro G-I Uechi M Tadano and ANishizono ldquoConstruction of humanFab (1205741120581) library and iden-tification of human monoclonal Fab possessing neutralizingpotency against Japanese encephalitis virusrdquo Microbiology andImmunology vol 51 no 6 pp 617ndash625 2007
[56] L Warter C Y Lee R Thiagarajan et al ldquoChikungunya virusenvelope-specific human monoclonal antibodies with broadneutralization potencyrdquo Journal of Immunology vol 186 no 5pp 3258ndash3264 2011
[57] P Pal K A Dowd J D Brien et al ldquoDevelopment of a highlyprotective combination monoclonal antibody therapy againstchikungunya virusrdquo PLoS Pathogens vol 9 no 4 Article IDe1003312 2013
[58] A Bertolotti-Ciarlet J Smith K Strecker et al ldquoCellular local-ization and antigenic characterization of Crimean-Congo hem-orrhagic fever virus glycoproteinsrdquo Journal of Virology vol 79no 10 pp 6152ndash6161 2005
[59] J K Battles and J M Dalrymple ldquoGenetic variation amonggeographic isolates of Rift Valley fever virusrdquo American Journalof Tropical Medicine and Hygiene vol 39 no 6 pp 617ndash6311988
[60] B AThibodeaux N C Garbino NM Liss et al ldquoA humanizedIgG but not IgM antibody is effective in prophylaxis and therapyof yellow fever infection in an AG12917D-204 peripheralchallenge mouse modelrdquo Antiviral Research vol 94 no 1 pp1ndash8 2012
BioMed Research International 19
[61] S Daffis R E Kontermann J Korimbocus H Zeller H-DKlenk and J Ter Meulen ldquoAntibody responses against wild-type yellow fever virus and the 17D vaccine strain charac-terization with human monoclonal antibody fragments andneutralization escape variantsrdquoVirology vol 337 no 2 pp 262ndash272 2005
[62] L N Levanov L E Matveev E P Goncharova et al ldquoChimericantibodies against tick-borne encephalitis virusrdquo Vaccine vol28 no 32 pp 5265ndash5271 2010
[63] A R Hunt S Frederickson C Hinkel K S Bowdish and J TRoehrig ldquoA humanized murine monoclonal antibody protectsmice either before or after challenge with virulent Venezuelanequine encephalomyelitis virusrdquo Journal of General Virologyvol 87 no 9 pp 2467ndash2476 2006
[64] J M da Silva Voorham I A Rodenhuis-Zybert N V A Nunezet al ldquoAntibodies against the envelope glycoprotein promoteinfectivity of immature dengue virus serotype 2rdquo PLoS ONEvol 7 no 3 Article ID e29957 2012
[65] T C Pierson Q Xu S Nelson et al ldquoThe stoichiometry ofantibody-mediated neutralization and enhancement of westnile virus infectionrdquoCell Host andMicrobe vol 1 no 2 pp 135ndash145 2007
[66] G Sautto N Mancini R A Diotti L Solforosi M Clementiand R Burioni ldquoAnti-hepatitis C virus E2 (HCVE2) glyco-proteinmonoclonal antibodies andneutralization interferencerdquoAntiviral Research vol 96 no 1 pp 82ndash89 2012
[67] R Burioni Y Matsuura N Mancini et al ldquoDiverging effectsof human recombinant anti-hepatitis C virus (HCV) antibodyfragments derived from a single patient on the infectivityof a vesicular stomatitis virusHCV pseudotyperdquo Journal ofVirology vol 76 no 22 pp 11775ndash11779 2002
[68] R J Phillpotts J R Stephenson and J S Porterfield ldquoAntibody-dependent enhancement of tick-borne encephalitus virus infec-tivityrdquo Journal of General Virology vol 66 no 8 pp 1831ndash18371985
[69] K A Dowd and T C Pierson ldquoAntibody-mediated neutraliza-tion of flaviviruses a reductionist viewrdquo Virology vol 411 no 2pp 306ndash315 2011
[70] A K I Falconar ldquoMonoclonal antibodies that bind to commonepitopes on the dengue virus type 2 nonstructural-1 andenvelope glycoproteins display weak neutralizing activity anddifferentiated responses to virulent strains implications forpathogenesis and vaccinesrdquo Clinical and Vaccine Immunologyvol 15 no 3 pp 549ndash561 2008
[71] K Meyer A Banerjee S E Frey R B Belshe and R Ray ldquoAweak neutralizing antibody response to hepatitis C virus enve-lope glycoprotein enhances virus infectionrdquo PLoS ONE vol 6no 8 Article ID e23699 2011
[72] R Burioni R AWilliamson P P Sanna F E Bloom and D RBurton ldquoRecombinant human Fab to glycoprotein D neutral-izes infectivity and prevents cell-to-cell transmission of herpessimplex viruses 1 and 2 in vitrordquo Proceedings of the NationalAcademy of Sciences of the United States of America vol 91 no1 pp 355ndash359 1994
[73] M S Rolph A Zaid N E Rulli and S Mahalingam ldquoDown-regulation of interferon-120573 in antibody-dependent enhancementof dengue viral infections of human macrophages is dependenton interleukin-6rdquo Journal of Infectious Diseases vol 204 no 3pp 489ndash491 2011
[74] S S Whitehead J E Blaney A P Durbin and B R MurphyldquoProspects for a dengue virus vaccinerdquoNature ReviewsMicrobi-ology vol 5 no 7 pp 518ndash528 2007
[75] S-M Lok V Kostyuchenko G ENybakken et al ldquoBinding of aneutralizing antibody to dengue virus alters the arrangement ofsurface glycoproteinsrdquoNature Structural andMolecular Biologyvol 15 no 3 pp 312ndash317 2008
[76] A P Goncalvez R Men C Wernly R H Purcell and C-J Lai ldquoChimpanzee Fab fragments and a derived humanizedimmunogiobulin G1 antibody that efficiently cross-neutralizedengue type 1 and type 2 virusesrdquo Journal of Virology vol 78no 23 pp 12910ndash12918 2004
[77] WM P BWahala A A Kraus L B HaymoreM A Accavitti-Loper and A M de Silva ldquoDengue virus neutralization byhuman immune sera role of envelope protein domain III-reactive antibodyrdquo Virology vol 392 no 1 pp 103ndash113 2009
[78] W D Crill H R Hughes M J Delorey and G-J J ChangldquoHumoral immune responses of dengue fever patients usingepitope-specific serotype-2 virus-like particle antigensrdquo PLoSONE vol 4 no 4 Article ID e4991 2009
[79] C Setthapramote T Sasaki O Puiprom et al ldquoHuman mono-clonal antibodies to neutralize all dengue virus serotypes usinglymphocytes from patients at acute phase of the secondaryinfectionrdquo Biochemical and Biophysical Research Communica-tions vol 423 no 4 pp 867ndash872 2012
[80] M Beltramello K L Williams C P Simmons et al ldquoThehuman immune response to dengue virus is dominated byhighly cross-reactive antibodies endowed with neutralizing andenhancing activityrdquo Cell Host andMicrobe vol 8 no 3 pp 271ndash283 2010
[81] C M Midgley M Bajwa-Joseph S Vasanawathana et al ldquoAnin-depth analysis of original antigenic sin in dengue virusinfectionrdquo Journal of Virology vol 85 no 22 pp 410ndash421 2011
[82] B Kaufmann M R Vogt J Goudsmit et al ldquoNeutralizationof West Nile virus by cross-linking of its surface proteins withFab fragments of the human monoclonal antibody CR4354rdquoProceedings of the National Academy of Sciences of the UnitedStates of America vol 107 no 44 pp 18950ndash18955 2010
[83] B Spurrier J M Sampson M Totrov et al ldquoStructural analysisof human and macaque mAbs 2909 and 25B implicationsfor the configuration of the quaternary neutralizing epitope ofHIV-1 gp120rdquo Structure vol 19 no 5 pp 691ndash699 2011
[84] J J SchlesingerMWBrandriss andT PMonath ldquoMonoclonalantibodies distinguish between wild and vaccine strains ofyellow fever virus by neutralization hemagglutination inhibi-tion and immune precipitation of the virus envelope proteinrdquoVirology vol 125 no 1 pp 8ndash17 1983
[85] M W Brandriss J J Schlesinger E E Walsh and M BrisellildquoLethal 17D yellow fever encephalitis in mice I Passive protec-tion by monoclonal antibodies to the envelope proteins of 17Dyellow fever and dengue 2 virusesrdquo Journal of General Virologyvol 67 no 2 pp 229ndash234 1986
[86] H Cho and M S Diamond ldquoImmune responses to West Nilevirus infection in the central nervous systemrdquo Viruses vol 4no 12 pp 3812ndash3830 2012
[87] D W Beasley A D Barrett and R B Tesh ldquoResurgence ofWest Nile neurologic disease in the United States in 2012 whathappened What needs to be donerdquo Antiviral Research vol 99no 1 pp 1ndash5 2013
[88] M Throsby J Ter Meulen C Geuijen J Goudsmit and J DeKruif ldquoMapping and analysis of West Nile virus-specific mon-oclonal antibodies prospects for vaccine developmentrdquo ExpertReview of Vaccines vol 6 no 2 pp 183ndash191 2007
[89] M S Diamond T G Roberts D Edgil B Lu J Ernst and EHarris ldquoModulation of dengue virus infection in human cells
20 BioMed Research International
by alpha beta and gamma interferonsrdquo Journal of Virology vol74 no 11 pp 4957ndash4966 2000
[90] Z Shimoni M J Niven S Pitlick and S Bulvik ldquoTreatmentof West Nile virus encephalitis with intravenous immunoglob-ulinrdquo Emerging Infectious Diseases vol 7 no 4 article 759 2001
[91] AHamdan PGreen EMendelsonM R Kramer S Pitlik andM Weinberger ldquoPossible benefit of intravenous immunoglob-ulin therapy in a lung transplant recipient with West Nile virusencephalitisrdquoTransplant InfectiousDisease vol 4 no 3 pp 160ndash162 2002
[92] M Haley A S Retter D Fowler J Gea-Banacloche and NP OrsquoGrady ldquoThe role for intravenous immunoglobulin in thetreatment of West Nile virus encephalitisrdquo Clinical InfectiousDiseases vol 37 no 6 pp e88ndashe90 2003
[93] K M Chung G E Nybakken B S Thompson et al ldquoAnti-bodies against West Nile virus nonstructural protein NS1prevent lethal infection through Fc 120574 receptor-dependent and-independent mechanismsrdquo Journal of Virology vol 80 no 3pp 1340ndash1351 2006
[94] E A Henchal L S Henchal and J J Schlesinger ldquoSynergisticinteractions of anti-NS1 monoclonal antibodies protect pas-sively immunized mice from lethal challenge with dengue 2virusrdquo Journal of General Virology vol 69 no 8 pp 2101ndash21071988
[95] J T Roehrig ldquoAntigenic structure of flavivirus proteinsrdquoAdvances in Virus Research vol 59 pp 141ndash175 2003
[96] M D Sanchez T C Pierson D McAllister et al ldquoCharacteri-zation of neutralizing antibodies to West Nile virusrdquo Virologyvol 336 no 1 pp 70ndash82 2005
[97] J H Beigel J L Nordstrom S R Pillemer et al ldquoSafety andpharmacokinetics of single intravenous dose of MGAWN1 anovel monoclonal antibody to West Nile virusrdquo AntimicrobialAgents and Chemotherapy vol 54 no 6 pp 2431ndash2436 2010
[98] C A Smeraski V Siddharthan and J D Morrey ldquoTreatmentof spatial memory impairment in hamsters infected with WestNile virus using a humanizedmonoclonal antibodyMGAWN1rdquoAntiviral Research vol 91 no 1 pp 43ndash49 2011
[99] D Lelli AMoreno E Brocchi et al ldquoWestNile virus character-ization and diagnostic applications of monoclonal antibodiesrdquoVirology Journal vol 9 article 81 2012
[100] T S Gritsun V A Lashkevich and E A Gould ldquoTick-borneencephalitisrdquo Antiviral Research vol 57 no 1-2 pp 129ndash1462003
[101] D Ruzek T S Gritsun N L Forrester et al ldquoMutations inthe NS2B and NS3 genes affect mouse neuroinvasiveness of aWestern European field strain of tick-borne encephalitis virusrdquoVirology vol 374 no 2 pp 249ndash255 2008
[102] M Ecker S L Allison T Meixner and F X Heinz ldquoSequenceanalysis and genetic classification of tick-borne encephalitisviruses from Europe and Asiardquo Journal of General Virology vol80 no 1 pp 179ndash185 1999
[103] H Holzmann M S Vorobyova I P Ladyzhenskaya et alldquoMolecular epidemiology of tick-borne encephalitis viruscross-protection between European and Far Eastern subtypesrdquoVaccine vol 10 no 5 pp 345ndash349 1992
[104] C W Mandl ldquoSteps of the tick-borne encephalitis virus repli-cation cycle that affect neuropathogenesisrdquo Virus Research vol111 no 2 pp 161ndash174 2005
[105] A V Atrasheuskaya T M Fredeking and G M IgnatyevldquoChanges in immune parameters and their correction in humancases of tick-borne encephalitisrdquo Clinical and ExperimentalImmunology vol 131 no 1 pp 148ndash154 2003
[106] K L Mansfield N Johnson L P Phipps J R Stephenson AR Fooks and T Solomon ldquoTick-borne encephalitis virusmdashareview of an emerging zoonosisrdquo Journal of General Virologyvol 90 no 8 pp 1781ndash1794 2009
[107] DWVaughn andCHHoke Jr ldquoThe epidemiology of Japaneseencephalitis prospects for preventionrdquo Epidemiologic Reviewsvol 14 pp 197ndash221 1992
[108] T Solomon H Ni D W C Beasley M Ekkelenkamp M JCardosa and A D T Barrett ldquoOrigin and evolution of Japaneseencephalitis virus in Southeast Asiardquo Journal of Virology vol 77no 5 pp 3091ndash3098 2003
[109] M R Wilson ldquoEmerging viral infectionsrdquo Current Opinion inNeurology vol 26 no 3 pp 301ndash306 2013
[110] J N Hanna S A Ritchie D A Phillips et al ldquoAn outbreakof Japanese encephalitis in the Torres Strait Australia 1995rdquoMedical Journal of Australia vol 165 no 5 pp 256ndash260 1996
[111] J N Hanna S A Ritchie D A Phillips et al ldquoJapanese ence-phalitis in north Queensland Australia 1998rdquo Medical Journalof Australia vol 170 no 11 pp 533ndash536 1999
[112] SGeorgeMGourie-Devi and J A Rao ldquoIsolation ofWestNilevirus from the brains of children who had died of encephalitisrdquoBulletin of theWorldHealthOrganization vol 62 no 6 pp 879ndash882 1984
[113] A K Gupta A A Koshy and V J Lad ldquoEnhanced protec-tion of mice against Japanese encephalitis virus infection bycombinations ofmonoclonal antibodies to glycoprotein ErdquoActaVirologica vol 55 no 2 pp 165ndash168 2011
[114] T H Lee B-H Song S-I Yun et al ldquoA cross-protective mAbrecognizes a novel epitope within the flavivirus NS1 proteinrdquoJournal of General Virology vol 93 no 1 pp 20ndash26 2012
[115] S C Weaver ldquoHost range amplification and arboviral diseaseemergencerdquo Archives of Virology Supplementum no 19 pp 33ndash44 2005
[116] M Solignat B Gay S Higgs L Briant and C Devaux ldquoRepli-cation cycle of chikungunya a re-emerging arbovirusrdquoVirologyvol 393 no 2 pp 183ndash197 2009
[117] O Schwartz and M L Albert ldquoBiology and pathogenesis ofchikungunya virusrdquo Nature Reviews Microbiology vol 8 no 7pp 491ndash500 2010
[118] W Akahata Z-Y Yang H Andersen et al ldquoA virus-like particlevaccine for epidemic Chikungunya virus protects nonhumanprimates against infectionrdquo Nature Medicine vol 16 no 3 pp334ndash338 2010
[119] M Kielian and A Helenius ldquopH-Iinduced alterations in thefusogenic spike protein of Semliki Forest virusrdquo Journal of CellBiology vol 101 no 6 pp 2284ndash2291 1985
[120] J White and A Helenius ldquopH-Dependent fusion between theSemliki Forest virus membrane and liposomesrdquo Proceedings ofthe National Academy of Sciences of the United States of Americavol 77 no 6 pp 3273ndash3277 1980
[121] J Dubuisson and C M Rice ldquoSindbis virus attachment isola-tion and characterization of mutants with impaired binding tovertebrate cellsrdquo Journal of Virology vol 67 no 6 pp 3363ndash33741993
[122] C Y Lee Y-W Kam J Fric et al ldquoChikungunya virus neutral-ization antigens and direct cell-to-cell transmission are revealedby human antibody-escapemutantsrdquo PLoS Pathogens vol 7 no12 Article ID e1002390 2011
[123] J E Voss M-C Vaney S Duquerroy et al ldquoGlycoproteinorganization of Chikungunya virus particles revealed by X-raycrystallographyrdquo Nature vol 468 no 7324 pp 709ndash712 2010
BioMed Research International 21
[124] L Li J Jose Y Xiang R J Kuhn and M G Rossmann ldquoStruc-tural changes of envelope proteins during alphavirus fusionrdquoNature vol 468 no 7324 pp 705ndash708 2010
[125] J Fric S Bertin-Maghit C I Wang A Nardin and L WarterldquoUse of human monoclonal antibodies to treat Chikungunyavirus infectionrdquo Journal of Infectious Diseases vol 207 no 2 pp319ndash322 2013
[126] P V Aguilar J G Estrada-Franco R Navarro-Lopez C FerroA D Haddow and S C Weaver ldquoEndemic Venezuelanequine encephalitis in the Americas hidden under the dengueumbrellardquo Future Virology vol 6 no 6 pp 721ndash740 2011
[127] S C Weaver R Winegar I D Manger and N L ForresterldquoAlphaviruses population genetics and determinants of emer-gencerdquo Antiviral Research vol 94 no 3 pp 242ndash257 2012
[128] S Paessler and S C Weaver ldquoVaccines for Venezuelan equineencephalitisrdquo Vaccine vol 27 no 4 pp D80ndashD85 2009
[129] R J Phillpotts ldquoVenezuelan equine encephalitis virus complex-specific monoclonal antibody provides broad protection inmurine models against airborne challenge with viruses fromserogroups I II and IIIrdquoVirus Research vol 120 no 1-2 pp 107ndash112 2006
[130] R J Phillpotts L D Jones and S C Howard ldquoMonoclonalantibody protectsmice against infection anddiseasewhen giveneither before or up to 24 h after airborne challenge with virulentVenezuelan equine encephalitis virusrdquoVaccine vol 20 no 11-12pp 1497ndash1504 2002
[131] J H Mathews J T Roehrig and D W Trent ldquoRole of com-plement and the Fc portion of immunoglobulin G in immunityto Venezuelan equine encephalomyelitis virus infection withglycoprotein-specific monoclonal antibodiesrdquo Journal of Virol-ogy vol 55 no 3 pp 594ndash600 1985
[132] L M OrsquoBrien C D Underwood-Fowler S A Goodchild AL Phelps and R J Phillpotts ldquoDevelopment of a novel mon-oclonal antibody with reactivity to a wide range of Venezuelanequine encephalitis virus strainsrdquoVirology Journal vol 6 article206 2009
[133] L M OrsquoBrien S A Goodchild R J Phillpotts and S D Per-kins ldquoA humanised murine monoclonal antibody protectsmice from Venezuelan equine encephalitis virus Evergladesvirus and Mucambo virus when administered up to 48h afterairborne challengerdquo Virology vol 426 no 2 pp 100ndash105 2012
[134] N Clementi N Mancini L Solforosi M Castelli M Clementiand R Burioni ldquoPhage display-based strategies for cloning andoptimization of monoclonal antibodies directed against humanpathogensrdquo International Journal of Molecular Sciences vol 13no 7 pp 8273ndash8292 2012
[135] A R Hunt R A Bowen S Frederickson T Maruyama JT Roehrig and C D Blair ldquoTreatment of mice with humanmonoclonal antibody 24h after lethal aerosol challenge withvirulent Venezuelan equine encephalitis virus prevents diseasebut not infectionrdquo Virology vol 414 no 2 pp 146ndash152 2011
[136] W-G Hu A L Phelps S Jager et al ldquoA recombinant human-ized monoclonal antibody completely protects mice againstlethal challenge with Venezuelan equine encephalitis virusrdquoVaccine vol 28 no 34 pp 5558ndash5564 2010
[137] T Rulker L Voss P Thullier et al ldquoIsolation and characterisa-tion of a human-like antibody fragment (scFv) that inactivatesVEEV in vitro and in vivordquo PLoS ONE vol 7 no 5 Article IDe37242 2012
[138] C T Walter and J N Barr ldquoRecent advances in the molecularand cellular biology of bunyavirusesrdquo Journal of General Virol-ogy vol 92 no 11 pp 2467ndash2484 2011
[139] C A Whitehouse ldquoCrimean-Congo hemorrhagic feverrdquo Anti-viral Research vol 64 no 3 pp 145ndash160 2004
[140] M Mertens K Schmidt A Ozkul and M H Groschup ldquoTheimpact of Crimean-Congo hemorrhagic fever virus on publichealthrdquo Antiviral Research vol 98 no 2 pp 248ndash260 2013
[141] A A Ahmed J M McFalls C Hoffmann et al ldquoPresence ofbroadly reactive and group-specific neutralizing epitopes onnewly described isolates of Crimean-Congo hemorrhagic fevervirusrdquo Journal of General Virology vol 86 no 12 pp 3327ndash33362005
[142] R Hewson A Gmyl L Gmyl et al ldquoEvidence of segment reas-sortment in Crimean-Congo haemorrhagic fever virusrdquo Journalof General Virology vol 85 no 10 pp 3059ndash3070 2004
[143] N K Blackburn T G Besselaar A J Shepherd and R Swane-poel ldquoPreparation and use of monoclonal antibodies for iden-tifying Crimean-Congo hemorrhagic fever virusrdquo AmericanJournal of Tropical Medicine and Hygiene vol 37 no 2 pp 392ndash397 1987
[144] T Ikegami ldquoMolecular biology and genetic diversity of RiftValley fever virusrdquoAntiviral Research vol 95 no 3 pp 293ndash3102012
[145] KKeegan andM S Collett ldquoUse of bacterial expression cloningto define the amino acid sequences of antigenic determinantson the G2 glycoprotein of Rift Valley fever virusrdquo Journal ofVirology vol 58 no 2 pp 263ndash270 1986
[146] T G Besselaar and N K Blackburn ldquoTopological mapping ofantigenic sites on the Rift Valley fever virus envelope glycopro-teins using monoclonal antibodiesrdquo Archives of Virology vol121 no 1ndash4 pp 111ndash124 1991
[147] L G Presta ldquoEngineering of therapeutic antibodies to mini-mize immunogenicity and optimize functionrdquo Advanced DrugDelivery Reviews vol 58 no 5-6 pp 640ndash656 2006
Submit your manuscripts athttpwwwhindawicom
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anatomy Research International
PeptidesInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporation httpwwwhindawicom
International Journal of
Volume 2014
Zoology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Molecular Biology International
GenomicsInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioinformaticsAdvances in
Marine BiologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Signal TransductionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Evolutionary BiologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Biochemistry Research International
ArchaeaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Genetics Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in
Virolog y
Hindawi Publishing Corporationhttpwwwhindawicom
Nucleic AcidsJournal of
Volume 2014
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Enzyme Research
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Microbiology

16 BioMed Research International
within theGn andGc glycoproteins expressed by the CCHFVstrain IbAr10200 The effectiveness of the mAbs was studiedby performing PRNT80 and none of the anti-Gn mAbsshowed neutralizing activity although many of the anti-GcmAbs neutralized IbAr10200 strain in vitro with clones 8A15E3 12A9 6C2 and 9H3 showing activity at a gt5120 dilutionMAbs 11E7 and 30F7 exhibited neutralizing activity at 2560dilution A suckling mice protection test was thereforeperformed for these mAbs The previously mentionedanti-Gc Abs were capable to protect mice to an appreciabledegree when applied 24 hours before and in a weakermanner 24 hours after virus challenge Many of the anti-GnmAbs were able to confer significant protection to IbAr10200both 24 hours before and after virus administration Therelevant effectiveness of the anti-Gn mAbs 6B12 10E1113G8 and 10G4 suggests that these mAbs may possess someneutralizing activity due to Ab-based effector mechanisms(eg ADCC) [58]
In further studies the same group characterized thebroadly cross-reactive neutralizing activity of the previouslydescribed murine mAb 11E7 able to bind conformationalepitopes within the C-terminal region of the Gn glycoproteinof all CCHFVM groups [58 141]
42 Rift Valley Fever Virus (RVFV) Similarly to CCHFV RiftValley Fever Virus (RVFV) causes sporadic outbreaks Thelargest epidemic occurred in Egypt in 1977 with an estimated200000 infections 18000 patients manifesting symptomsand 600 fatal prognoses [144] Symptoms include hemor-rhagic episodes fever encephalitis and blindness [145]
The major target of the humoral immune response is theRVFV glycoproteins Gc (G1) and Gn (G2) ranging fromnucleotides 480ndash2090 and 2091ndash3614 respectively [144]
421 mAbs against RVFV Within G2 glycoprotein are des-cribed three neutralizing antigenic sites namely epitope Inucleotides 792ndash893 (amino acid residues 258ndash291) epitopeII nucleotides 1164ndash1196 (amino acid residues 382ndash392) andepitope IV nucleotides 858ndash917 (amino acid residues 280ndash299) [59 145]
Despite the high degree of variability observed inCCHFVglycoproteins RVFV G2 presented an unexpected high con-servation among 22 isolates [59] In the same study onlythe LUNYO and 900060 isolates were resistant to in vitroneutralization by murine mAb 4D4 (recognizing epitope II)All other isolates were neutralized with PRNT80 titers from10240 togt81920On the other hand only SNS isolate showedreduced sensitivity to 4ndash39-CC (epitope IV) All other isolateswere neutralized with PRNT80 titers from 20480 to gt81920Protection studies on mice would be of interest for thedevelopment of a passive immunization prophylaxis [59]
Protection studies from lethal South African RVFVAN1830 strain infection were successfully performed on abattery of anti-G1 and anti-G2 mAbs produced by Besselaarand Blackburn with the strongly neutralizing anti-G1 mAb3E5 and anti-G2mAb 9C4 being the most effective [146]The9C4 antigenic area maps within G2 epitope I and a broadlyneutralizing activity may be investigated
5 Conclusions
Arbovirus infections have acquired increasing interest giventhe augmented globalization and tourism movement all overthe world that could be at the basis of epidemic eventsFurthermore the same vectors can sometimes transmit sev-eral arboviruses concomitantly complicating the diagnosis aswell as the therapy For example confusing mixed epidemicshave occasionally been described such as YFV plus CHIKVDENVplus CHIKV ormore recentlyPlasmodium falciparummalaria plus DENV1 and CHIKV on Madagascarrsquos east coast[1]
In the last decade several efforts have been employed inthe development of effective immunogens and therapeuticsgiving the current absence of specific antiviral drugs as wellas effective vaccines for themost diffused arbovirus infections(ie DENV andWNV)
However at the moment mosquito control is the bestavailable method for preventing arbovirus infections Thusto control the emerging public health problem of arbovirusinfections new antiviral therapeutic strategies that providepotent and broadly cross-protective immunity (especially forflavivirus infections) are an urgent globally medical need
As described in this review the majority of highlyneutralizing mAbs are of murine origin and indeed theirutility in the treatment of these infections is limited In facttheir employment can elicit an immunogenic response (iehuman anti-mouse Abs HAMA) the therapeutic efficacycould be reduced by a relatively faster clearance in humans(compared to human Abs) potentially exacerbated by theHAMA response Finally murine Abs exhibit relatively weakeffector functions (eg ADCC) compared to human AbsIndeed chimerization humanization or better the isolationand characterization of fully human broadly neutralizingmAbs would be the best choice for their possible use in afuture mAb-based therapy against arbovirus infections [147]
Finally possible ADE of infection mechanisms should beevaluated before considering a mAb as a possible candidatetherapeutic in a post-exposure setting as well as in the devel-opment of new Ab-eliciting immunogens
Conflict of Interests
The authors declare no conflict of interests in particular withOmriGam and any other cited Ab-based preparation men-tioned in the paper
References
[1] N Cleton M Koopmans J Reimerink G J Godeke and CReusken ldquoCome fly with me review of clinically importantarboviruses for global travelersrdquo Journal of Clinical Virology vol55 no 3 pp 191ndash203 2012
[2] F X Heinz H Holzmann A Essl and M Kundi ldquoField effec-tiveness of vaccination against tick-borne encephalitisrdquoVaccinevol 25 no 43 pp 7559ndash7567 2007
[3] A D Barrett and D E Teuwen ldquoYellow fever vaccinemdashhowdoes it work and why do rare cases of serious adverse events
BioMed Research International 17
take placerdquo Current Opinion in Immunology vol 21 no 3 pp308ndash313 2009
[4] S B Halstead and S J Thomas ldquoNew vaccines for Japaneseencephalitisrdquo Current Infectious Disease Reports vol 12 no 3pp 174ndash180 2010
[5] R Burioni M Perotti N Mancini and M Clementi ldquoPer-spectives for the utilization of neutralizing human monoclonalantibodies as anti-HCV drugsrdquo Journal of Hepatology vol 49no 2 pp 299ndash300 2008
[6] R A Diotti G A Sautto L Solforosi NManciniM Clementiand R Burioni ldquoNeutralization activity and kinetics of twobroad-range human monoclonal IgG1 derived from recombi-nant Fab fragments and directed against Hepatitis C virus E2glycoproteinrdquo New Microbiologica vol 35 no 4 pp 475ndash4792012
[7] R Burioni F Canducci N Mancini et al ldquoMolecular cloningof the first humanmonoclona antibodies neutralizing with highpotency Swine-origin Influenza A pandemic virus (S-OIV)rdquoNew Microbiologica vol 32 no 4 pp 319ndash324 2009
[8] P K Chandra S Hazari B Poat et al ldquoIntracytoplasmic stableexpression of IgG1 antibody targeting NS3 helicase inhibitsreplication of highly efficient hepatitis C Virus 2a clonerdquoVirology Journal vol 7 article 118 2010
[9] N Clementi D de Marco N Mancini et al ldquoA human mono-clonal antibody with neutralizing activity against highly diver-gent influenza subtypesrdquo PLoS ONE vol 6 no 12 Article IDe28001 2011
[10] D de Marco N Clementi N Mancini et al ldquoA non-VH1-69 heterosubtypic neutralizing human monoclonal antibodyprotects mice against H1N1 and H5N1 virusesrdquo PLoS ONE vol7 no 4 Article ID e34415 2012
[11] L Solforosi N Mancini F Canducci et al ldquoA phage displayvector optimized for the generation of human antibody com-binatorial libraries and the molecular cloning of monoclonalantibody fragmentsrdquoNewMicrobiologica vol 35 no 3 pp 289ndash294 2012
[12] E V Koonin and V V Dolja ldquoEvolution and taxonomy ofpositive-strand RNA viruses implications of comparative anal-ysis of amino acid sequencesrdquo Critical Reviews in Biochemistryand Molecular Biology vol 28 no 5 pp 375ndash430 1993
[13] J S Mackenzie D J Gubler and L R Petersen ldquoEmergingflaviviruses the spread and resurgence of Japanese encephalitisWest Nile and dengue virusesrdquo Nature Medicine vol 10 no 12pp S98ndashS109 2004
[14] T J SmithW E Brandt J L Swanson J MMcCown and E LBuescher ldquoPhysical and biological properties of dengue-2 virusand associated antigensrdquo Journal of Virology vol 5 no 4 pp524ndash532 1970
[15] J M Smit B Moesker I Rodenhuis-Zybert and J WilschutldquoFlavivirus cell entry and membrane fusionrdquo Viruses vol 3 no2 pp 160ndash171 2011
[16] R J Kuhn W Zhang M G Rossmann et al ldquoStructure ofdengue virus implications for flavivirus organization matura-tion and fusionrdquo Cell vol 108 no 5 pp 717ndash725 2002
[17] K Stiasny J H Aberle M Keller B Grubeck-Loebenstein andF X Heinz ldquoAge affects quantity but not quality of antibodyresponses after vaccination with an inactivated flavivirus vac-cine against tick-borne encephalitisrdquo PLoS ONE vol 7 no 3Article ID e34145 2012
[18] LThomasOVerlaetenACabie et al ldquoInfluence of the dengueserotype previous dengue infection and plasma viral load
on clinical presentation and outcome during a dengue-2 anddengue-4 co-epidemicrdquo American Journal of Tropical Medicineand Hygiene vol 78 no 6 pp 990ndash998 2008
[19] Y Zhang J Corver P R Chipman et al ldquoStructures of imma-ture flavivirus particlesrdquo EMBO Journal vol 22 no 11 pp2604ndash2613 2003
[20] A K I Falconar ldquoIdentification of an epitope on the denguevirus membrane (M) protein defined by cross-protective mon-oclonal antibodies design of an improved epitope sequencebased on common determinants present in both envelope (EandM) proteinsrdquo Archives of Virology vol 144 no 12 pp 2313ndash2330 1999
[21] K-J Huang Y-C Yang Y-S Lin et al ldquoThe dual-specific bind-ing of dengue virus and target cells for the antibody-dependentenhancement of dengue virus infectionrdquo Journal of Immunol-ogy vol 176 no 5 pp 2825ndash2832 2006
[22] A K I Falconar ldquoAntibody responses are generated to immun-odominant ELKKLE-typemotifs on the nonstructural-1 glyco-protein during live dengue virus infections inmice and humansimplications for diagnosis pathogenesis and vaccine designrdquoClinical and Vaccine Immunology vol 14 no 5 pp 493ndash5042007
[23] C W Mandl F Guirakhoo H Holzmann F X Heinz and CKunz ldquoAntigenic structure of the flavivirus envelope protein Eat the molecular level using the tick-borne encephalitis virus asa modelrdquo Journal of Virology vol 63 no 2 pp 564ndash571 1989
[24] F A Rey F X Heinz C Mandl C Kunz and S C HarrisonldquoThe envelope glycoprotein from tick-borne encephalitis virusat 2 A resolutionrdquo Nature vol 375 no 6529 pp 291ndash298 1995
[25] S L Allison K Stiasny K Stadler CWMandl and F XHeinzldquoMapping of functional elements in the stem-anchor regionof tick-borne encephalitis virus envelope protein Erdquo Journal ofVirology vol 73 no 7 pp 5605ndash5612 1999
[26] D E Klein J L Choi and S C Harrison ldquoStructure of a denguevirus envelope protein late-stage fusion intermediaterdquo Journal ofVirology vol 87 no 4 pp 2287ndash2293 2013
[27] S L Hanna T C Pierson M D Sanchez A A Ahmed M MMurtadha and R W Doms ldquoN-linked glycosylation of WestNile virus envelope proteins influences particle assembly andinfectivityrdquo Journal of Virology vol 79 no 21 pp 13262ndash132742005
[28] N Mancini G Sautto N Clementi et al ldquoNeutralization inter-fering antibodies a ldquonovelrdquo example of humoral immune dys-function facilitating viral escaperdquoViruses vol 4 no 9 pp 1731ndash1752 2012
[29] Y Modis S Ogata D Clements and S C Harrison ldquoVariablesurface epitopes in the crystal structure of dengue virus type3 envelope glycoproteinrdquo Journal of Virology vol 79 no 2 pp1223ndash1231 2005
[30] N Miller ldquoRecent progress in dengue vaccine research anddevelopmentrdquo Current Opinion in Molecular Therapeutics vol12 no 1 pp 31ndash38 2010
[31] J H Chavez J R Silva A A Amarilla and L T Moraes Figu-eiredo ldquoDomain III peptides from flavivirus envelope proteinare useful antigens for serologic diagnosis and targets forimmunizationrdquo Biologicals vol 38 no 6 pp 613ndash618 2010
[32] B Shrestha J D Brien S Sukupolvi-Petty et al ldquoThe develop-ment of therapeutic antibodies that neutralize homologous andheterologous genotypes of dengue virus type 1rdquo PLoS Pathogensvol 6 no 4 Article ID e1000823 2010
18 BioMed Research International
[33] G D Gromowski N D Barrett and A D T Barrett ldquoChar-acterization of dengue virus complex-specific neutralizing epi-topes on envelope protein domain III of dengue 2 virusrdquo Journalof Virology vol 82 no 17 pp 8828ndash8837 2008
[34] S Sukupolvi-Petty S K Austin W E Purtha et al ldquoType- andsubcomplex-specific neutralizing antibodies against domain IIIof dengue virus type 2 envelope protein recognize adjacentepitopesrdquo Journal of Virology vol 81 no 23 pp 12816ndash128262007
[35] S Sukupolvi-Petty S K Austin M Engle et al ldquoStructure andfunction analysis of therapeutic monoclonal antibodies againstdengue virus type 2rdquo Journal of Virology vol 84 no 18 pp9227ndash9239 2010
[36] Y-Q Deng J-X Dai G-H Ji et al ldquoA broadly flaviviruscross-neutralizingmonoclonal antibody that recognizes a novelepitope within the fusion loop of e proteinrdquo PLoS ONE vol 6no 1 Article ID e16059 2011
[37] J M Costin E Zaitseva K M Kahle et al ldquoMechanistic studyof broadly neutralizing human monoclonal antibodies againstdengue virus that target the fusion looprdquo Journal of Virologyvol 87 no 1 pp 52ndash66 2013
[38] CMMidgley A FlanaganH B Tran et al ldquoStructural analysisof a dengue cross-reactive antibody complexed with envelopedomain III reveals the molecular basis of cross-reactivityrdquoJournal of Immunology vol 188 no 10 pp 4971ndash4979 2012
[39] N J Moreland P Susanto E Lim et al ldquoPhage display appro-aches for the isolation of monoclonal antibodies against denguevirus envelope domain III from human and mouse derivedlibrariesrdquo International Journal ofMolecular Sciences vol 13 no3 pp 2618ndash2635 2012
[40] J J B Cockburn M E Navarro Sanchez N Fretes et alldquoMechanism of dengue virus broad cross-neutralization by amonoclonal antibodyrdquo Structure vol 20 no 2 pp 303ndash3142012
[41] R Rajamanonmani C Nkenfou P Clancy et al ldquoOn a mousemonoclonal antibody that neutralizes all four dengue virusserotypesrdquo Journal of General Virology vol 90 no 4 pp 799ndash809 2009
[42] L H Gould J Sui H Foellmer et al ldquoProtective and herapeuticcapacity of human single-chain Fv-Fc fusion proteins againstWest Nile virusrdquo Journal of Virology vol 79 no 23 pp 14606ndash14613 2005
[43] H Sultana H G Foellmer G Neelakanta et al ldquoFusion LoopPeptide of the West Nile virus envelope protein is essential forpathogenesis and is recognized by a therapeutic cross-reactivehuman monoclonal antibodyrdquo Journal of Immunology vol 183no 1 pp 650ndash660 2009
[44] T Oliphant M Engle G E Nybakken et al ldquoDevelopment ofa humanized monoclonal antibody with therapeutic potentialagainstWest Nile virusrdquoNature Medicine vol 11 no 5 pp 522ndash530 2005
[45] G E Nybakken T Oliphant S Johnson S Burke M S Dia-mond and D H Fremont ldquoStructural basis of West Nile virusneutralization by a therapeutic antibodyrdquo Nature vol 437 no7059 pp 764ndash769 2005
[46] G D Gromowski J T Roehrig M S Diamond J C LeeT J Pitcher and A D T Barrett ldquoMutations of an antibodybinding energy hot spot on domain III of the dengue 2 envelopeglycoprotein exploited for neutralization escaperdquo Virology vol407 no 2 pp 237ndash246 2010
[47] J T Roehrig R A Bolin andR G Kelly ldquoMonoclonal antibodymapping of the envelope glycoprotein of the dengue 2 virusJamaicardquo Virology vol 246 no 2 pp 317ndash328 1998
[48] R de Alwis S A Smith N P Olivarez et al ldquoIdentification ofhumanneutralizing antibodies that bind to complex epitopes ondengue virionsrdquo Proceedings of the National Academy of Sciencesof the United States of America vol 109 no 19 pp 7439ndash74442012
[49] S K Austin K A Dowd B Shrestha et al ldquoStructural basis ofdifferential neutralization of DENV-1 genotypes by an antibodythat recognizes a cryptic epitoperdquo PLoS Pathogens vol 8 no 10Article ID e1002930 2012
[50] M Throsby C Geuijen J Goudsmit et al ldquoIsolation and char-acterization of human monoclonal antibodies from individualsinfected with West Nile Virusrdquo Journal of Virology vol 80 no14 pp 6982ndash6992 2006
[51] A P Goncalvez R H Purcell and C-J Lai ldquoEpitope deter-minants of a chimpanzee Fab antibody that efficiently cross-neutralizes dengue type 1 and type 2 viruses map to inside andin close proximity to fusion loop of the dengue type 2 virusenvelope glycoproteinrdquo Journal of Virology vol 78 no 23 pp12919ndash12928 2004
[52] J T Roehrig J H Mathews and D W Trent ldquoIdentification ofepitopes on the E glycoprotein of Saint Louis encephalitis virususing monoclonal antibodiesrdquo Virology vol 128 no 1 pp 118ndash126 1983
[53] J T Roehrig L A Staudinger A R Hunt J H Mathews andC D Blair ldquoAntibody prophylaxis and therapy for flavivirusencephalitis infectionsrdquo Annals of the New York Academy ofSciences vol 951 pp 286ndash297 2001
[54] A P Goncalvez C-H Chien K Tubthong et al ldquoHumanizedmonoclonal antibodies derived from chimpanzee fabs protectagainst Japanese encephalitis virus in vitro and in vivordquo Journalof Virology vol 82 no 14 pp 7009ndash7021 2008
[55] M Arakawa T Yamashiro G-I Uechi M Tadano and ANishizono ldquoConstruction of humanFab (1205741120581) library and iden-tification of human monoclonal Fab possessing neutralizingpotency against Japanese encephalitis virusrdquo Microbiology andImmunology vol 51 no 6 pp 617ndash625 2007
[56] L Warter C Y Lee R Thiagarajan et al ldquoChikungunya virusenvelope-specific human monoclonal antibodies with broadneutralization potencyrdquo Journal of Immunology vol 186 no 5pp 3258ndash3264 2011
[57] P Pal K A Dowd J D Brien et al ldquoDevelopment of a highlyprotective combination monoclonal antibody therapy againstchikungunya virusrdquo PLoS Pathogens vol 9 no 4 Article IDe1003312 2013
[58] A Bertolotti-Ciarlet J Smith K Strecker et al ldquoCellular local-ization and antigenic characterization of Crimean-Congo hem-orrhagic fever virus glycoproteinsrdquo Journal of Virology vol 79no 10 pp 6152ndash6161 2005
[59] J K Battles and J M Dalrymple ldquoGenetic variation amonggeographic isolates of Rift Valley fever virusrdquo American Journalof Tropical Medicine and Hygiene vol 39 no 6 pp 617ndash6311988
[60] B AThibodeaux N C Garbino NM Liss et al ldquoA humanizedIgG but not IgM antibody is effective in prophylaxis and therapyof yellow fever infection in an AG12917D-204 peripheralchallenge mouse modelrdquo Antiviral Research vol 94 no 1 pp1ndash8 2012
BioMed Research International 19
[61] S Daffis R E Kontermann J Korimbocus H Zeller H-DKlenk and J Ter Meulen ldquoAntibody responses against wild-type yellow fever virus and the 17D vaccine strain charac-terization with human monoclonal antibody fragments andneutralization escape variantsrdquoVirology vol 337 no 2 pp 262ndash272 2005
[62] L N Levanov L E Matveev E P Goncharova et al ldquoChimericantibodies against tick-borne encephalitis virusrdquo Vaccine vol28 no 32 pp 5265ndash5271 2010
[63] A R Hunt S Frederickson C Hinkel K S Bowdish and J TRoehrig ldquoA humanized murine monoclonal antibody protectsmice either before or after challenge with virulent Venezuelanequine encephalomyelitis virusrdquo Journal of General Virologyvol 87 no 9 pp 2467ndash2476 2006
[64] J M da Silva Voorham I A Rodenhuis-Zybert N V A Nunezet al ldquoAntibodies against the envelope glycoprotein promoteinfectivity of immature dengue virus serotype 2rdquo PLoS ONEvol 7 no 3 Article ID e29957 2012
[65] T C Pierson Q Xu S Nelson et al ldquoThe stoichiometry ofantibody-mediated neutralization and enhancement of westnile virus infectionrdquoCell Host andMicrobe vol 1 no 2 pp 135ndash145 2007
[66] G Sautto N Mancini R A Diotti L Solforosi M Clementiand R Burioni ldquoAnti-hepatitis C virus E2 (HCVE2) glyco-proteinmonoclonal antibodies andneutralization interferencerdquoAntiviral Research vol 96 no 1 pp 82ndash89 2012
[67] R Burioni Y Matsuura N Mancini et al ldquoDiverging effectsof human recombinant anti-hepatitis C virus (HCV) antibodyfragments derived from a single patient on the infectivityof a vesicular stomatitis virusHCV pseudotyperdquo Journal ofVirology vol 76 no 22 pp 11775ndash11779 2002
[68] R J Phillpotts J R Stephenson and J S Porterfield ldquoAntibody-dependent enhancement of tick-borne encephalitus virus infec-tivityrdquo Journal of General Virology vol 66 no 8 pp 1831ndash18371985
[69] K A Dowd and T C Pierson ldquoAntibody-mediated neutraliza-tion of flaviviruses a reductionist viewrdquo Virology vol 411 no 2pp 306ndash315 2011
[70] A K I Falconar ldquoMonoclonal antibodies that bind to commonepitopes on the dengue virus type 2 nonstructural-1 andenvelope glycoproteins display weak neutralizing activity anddifferentiated responses to virulent strains implications forpathogenesis and vaccinesrdquo Clinical and Vaccine Immunologyvol 15 no 3 pp 549ndash561 2008
[71] K Meyer A Banerjee S E Frey R B Belshe and R Ray ldquoAweak neutralizing antibody response to hepatitis C virus enve-lope glycoprotein enhances virus infectionrdquo PLoS ONE vol 6no 8 Article ID e23699 2011
[72] R Burioni R AWilliamson P P Sanna F E Bloom and D RBurton ldquoRecombinant human Fab to glycoprotein D neutral-izes infectivity and prevents cell-to-cell transmission of herpessimplex viruses 1 and 2 in vitrordquo Proceedings of the NationalAcademy of Sciences of the United States of America vol 91 no1 pp 355ndash359 1994
[73] M S Rolph A Zaid N E Rulli and S Mahalingam ldquoDown-regulation of interferon-120573 in antibody-dependent enhancementof dengue viral infections of human macrophages is dependenton interleukin-6rdquo Journal of Infectious Diseases vol 204 no 3pp 489ndash491 2011
[74] S S Whitehead J E Blaney A P Durbin and B R MurphyldquoProspects for a dengue virus vaccinerdquoNature ReviewsMicrobi-ology vol 5 no 7 pp 518ndash528 2007
[75] S-M Lok V Kostyuchenko G ENybakken et al ldquoBinding of aneutralizing antibody to dengue virus alters the arrangement ofsurface glycoproteinsrdquoNature Structural andMolecular Biologyvol 15 no 3 pp 312ndash317 2008
[76] A P Goncalvez R Men C Wernly R H Purcell and C-J Lai ldquoChimpanzee Fab fragments and a derived humanizedimmunogiobulin G1 antibody that efficiently cross-neutralizedengue type 1 and type 2 virusesrdquo Journal of Virology vol 78no 23 pp 12910ndash12918 2004
[77] WM P BWahala A A Kraus L B HaymoreM A Accavitti-Loper and A M de Silva ldquoDengue virus neutralization byhuman immune sera role of envelope protein domain III-reactive antibodyrdquo Virology vol 392 no 1 pp 103ndash113 2009
[78] W D Crill H R Hughes M J Delorey and G-J J ChangldquoHumoral immune responses of dengue fever patients usingepitope-specific serotype-2 virus-like particle antigensrdquo PLoSONE vol 4 no 4 Article ID e4991 2009
[79] C Setthapramote T Sasaki O Puiprom et al ldquoHuman mono-clonal antibodies to neutralize all dengue virus serotypes usinglymphocytes from patients at acute phase of the secondaryinfectionrdquo Biochemical and Biophysical Research Communica-tions vol 423 no 4 pp 867ndash872 2012
[80] M Beltramello K L Williams C P Simmons et al ldquoThehuman immune response to dengue virus is dominated byhighly cross-reactive antibodies endowed with neutralizing andenhancing activityrdquo Cell Host andMicrobe vol 8 no 3 pp 271ndash283 2010
[81] C M Midgley M Bajwa-Joseph S Vasanawathana et al ldquoAnin-depth analysis of original antigenic sin in dengue virusinfectionrdquo Journal of Virology vol 85 no 22 pp 410ndash421 2011
[82] B Kaufmann M R Vogt J Goudsmit et al ldquoNeutralizationof West Nile virus by cross-linking of its surface proteins withFab fragments of the human monoclonal antibody CR4354rdquoProceedings of the National Academy of Sciences of the UnitedStates of America vol 107 no 44 pp 18950ndash18955 2010
[83] B Spurrier J M Sampson M Totrov et al ldquoStructural analysisof human and macaque mAbs 2909 and 25B implicationsfor the configuration of the quaternary neutralizing epitope ofHIV-1 gp120rdquo Structure vol 19 no 5 pp 691ndash699 2011
[84] J J SchlesingerMWBrandriss andT PMonath ldquoMonoclonalantibodies distinguish between wild and vaccine strains ofyellow fever virus by neutralization hemagglutination inhibi-tion and immune precipitation of the virus envelope proteinrdquoVirology vol 125 no 1 pp 8ndash17 1983
[85] M W Brandriss J J Schlesinger E E Walsh and M BrisellildquoLethal 17D yellow fever encephalitis in mice I Passive protec-tion by monoclonal antibodies to the envelope proteins of 17Dyellow fever and dengue 2 virusesrdquo Journal of General Virologyvol 67 no 2 pp 229ndash234 1986
[86] H Cho and M S Diamond ldquoImmune responses to West Nilevirus infection in the central nervous systemrdquo Viruses vol 4no 12 pp 3812ndash3830 2012
[87] D W Beasley A D Barrett and R B Tesh ldquoResurgence ofWest Nile neurologic disease in the United States in 2012 whathappened What needs to be donerdquo Antiviral Research vol 99no 1 pp 1ndash5 2013
[88] M Throsby J Ter Meulen C Geuijen J Goudsmit and J DeKruif ldquoMapping and analysis of West Nile virus-specific mon-oclonal antibodies prospects for vaccine developmentrdquo ExpertReview of Vaccines vol 6 no 2 pp 183ndash191 2007
[89] M S Diamond T G Roberts D Edgil B Lu J Ernst and EHarris ldquoModulation of dengue virus infection in human cells
20 BioMed Research International
by alpha beta and gamma interferonsrdquo Journal of Virology vol74 no 11 pp 4957ndash4966 2000
[90] Z Shimoni M J Niven S Pitlick and S Bulvik ldquoTreatmentof West Nile virus encephalitis with intravenous immunoglob-ulinrdquo Emerging Infectious Diseases vol 7 no 4 article 759 2001
[91] AHamdan PGreen EMendelsonM R Kramer S Pitlik andM Weinberger ldquoPossible benefit of intravenous immunoglob-ulin therapy in a lung transplant recipient with West Nile virusencephalitisrdquoTransplant InfectiousDisease vol 4 no 3 pp 160ndash162 2002
[92] M Haley A S Retter D Fowler J Gea-Banacloche and NP OrsquoGrady ldquoThe role for intravenous immunoglobulin in thetreatment of West Nile virus encephalitisrdquo Clinical InfectiousDiseases vol 37 no 6 pp e88ndashe90 2003
[93] K M Chung G E Nybakken B S Thompson et al ldquoAnti-bodies against West Nile virus nonstructural protein NS1prevent lethal infection through Fc 120574 receptor-dependent and-independent mechanismsrdquo Journal of Virology vol 80 no 3pp 1340ndash1351 2006
[94] E A Henchal L S Henchal and J J Schlesinger ldquoSynergisticinteractions of anti-NS1 monoclonal antibodies protect pas-sively immunized mice from lethal challenge with dengue 2virusrdquo Journal of General Virology vol 69 no 8 pp 2101ndash21071988
[95] J T Roehrig ldquoAntigenic structure of flavivirus proteinsrdquoAdvances in Virus Research vol 59 pp 141ndash175 2003
[96] M D Sanchez T C Pierson D McAllister et al ldquoCharacteri-zation of neutralizing antibodies to West Nile virusrdquo Virologyvol 336 no 1 pp 70ndash82 2005
[97] J H Beigel J L Nordstrom S R Pillemer et al ldquoSafety andpharmacokinetics of single intravenous dose of MGAWN1 anovel monoclonal antibody to West Nile virusrdquo AntimicrobialAgents and Chemotherapy vol 54 no 6 pp 2431ndash2436 2010
[98] C A Smeraski V Siddharthan and J D Morrey ldquoTreatmentof spatial memory impairment in hamsters infected with WestNile virus using a humanizedmonoclonal antibodyMGAWN1rdquoAntiviral Research vol 91 no 1 pp 43ndash49 2011
[99] D Lelli AMoreno E Brocchi et al ldquoWestNile virus character-ization and diagnostic applications of monoclonal antibodiesrdquoVirology Journal vol 9 article 81 2012
[100] T S Gritsun V A Lashkevich and E A Gould ldquoTick-borneencephalitisrdquo Antiviral Research vol 57 no 1-2 pp 129ndash1462003
[101] D Ruzek T S Gritsun N L Forrester et al ldquoMutations inthe NS2B and NS3 genes affect mouse neuroinvasiveness of aWestern European field strain of tick-borne encephalitis virusrdquoVirology vol 374 no 2 pp 249ndash255 2008
[102] M Ecker S L Allison T Meixner and F X Heinz ldquoSequenceanalysis and genetic classification of tick-borne encephalitisviruses from Europe and Asiardquo Journal of General Virology vol80 no 1 pp 179ndash185 1999
[103] H Holzmann M S Vorobyova I P Ladyzhenskaya et alldquoMolecular epidemiology of tick-borne encephalitis viruscross-protection between European and Far Eastern subtypesrdquoVaccine vol 10 no 5 pp 345ndash349 1992
[104] C W Mandl ldquoSteps of the tick-borne encephalitis virus repli-cation cycle that affect neuropathogenesisrdquo Virus Research vol111 no 2 pp 161ndash174 2005
[105] A V Atrasheuskaya T M Fredeking and G M IgnatyevldquoChanges in immune parameters and their correction in humancases of tick-borne encephalitisrdquo Clinical and ExperimentalImmunology vol 131 no 1 pp 148ndash154 2003
[106] K L Mansfield N Johnson L P Phipps J R Stephenson AR Fooks and T Solomon ldquoTick-borne encephalitis virusmdashareview of an emerging zoonosisrdquo Journal of General Virologyvol 90 no 8 pp 1781ndash1794 2009
[107] DWVaughn andCHHoke Jr ldquoThe epidemiology of Japaneseencephalitis prospects for preventionrdquo Epidemiologic Reviewsvol 14 pp 197ndash221 1992
[108] T Solomon H Ni D W C Beasley M Ekkelenkamp M JCardosa and A D T Barrett ldquoOrigin and evolution of Japaneseencephalitis virus in Southeast Asiardquo Journal of Virology vol 77no 5 pp 3091ndash3098 2003
[109] M R Wilson ldquoEmerging viral infectionsrdquo Current Opinion inNeurology vol 26 no 3 pp 301ndash306 2013
[110] J N Hanna S A Ritchie D A Phillips et al ldquoAn outbreakof Japanese encephalitis in the Torres Strait Australia 1995rdquoMedical Journal of Australia vol 165 no 5 pp 256ndash260 1996
[111] J N Hanna S A Ritchie D A Phillips et al ldquoJapanese ence-phalitis in north Queensland Australia 1998rdquo Medical Journalof Australia vol 170 no 11 pp 533ndash536 1999
[112] SGeorgeMGourie-Devi and J A Rao ldquoIsolation ofWestNilevirus from the brains of children who had died of encephalitisrdquoBulletin of theWorldHealthOrganization vol 62 no 6 pp 879ndash882 1984
[113] A K Gupta A A Koshy and V J Lad ldquoEnhanced protec-tion of mice against Japanese encephalitis virus infection bycombinations ofmonoclonal antibodies to glycoprotein ErdquoActaVirologica vol 55 no 2 pp 165ndash168 2011
[114] T H Lee B-H Song S-I Yun et al ldquoA cross-protective mAbrecognizes a novel epitope within the flavivirus NS1 proteinrdquoJournal of General Virology vol 93 no 1 pp 20ndash26 2012
[115] S C Weaver ldquoHost range amplification and arboviral diseaseemergencerdquo Archives of Virology Supplementum no 19 pp 33ndash44 2005
[116] M Solignat B Gay S Higgs L Briant and C Devaux ldquoRepli-cation cycle of chikungunya a re-emerging arbovirusrdquoVirologyvol 393 no 2 pp 183ndash197 2009
[117] O Schwartz and M L Albert ldquoBiology and pathogenesis ofchikungunya virusrdquo Nature Reviews Microbiology vol 8 no 7pp 491ndash500 2010
[118] W Akahata Z-Y Yang H Andersen et al ldquoA virus-like particlevaccine for epidemic Chikungunya virus protects nonhumanprimates against infectionrdquo Nature Medicine vol 16 no 3 pp334ndash338 2010
[119] M Kielian and A Helenius ldquopH-Iinduced alterations in thefusogenic spike protein of Semliki Forest virusrdquo Journal of CellBiology vol 101 no 6 pp 2284ndash2291 1985
[120] J White and A Helenius ldquopH-Dependent fusion between theSemliki Forest virus membrane and liposomesrdquo Proceedings ofthe National Academy of Sciences of the United States of Americavol 77 no 6 pp 3273ndash3277 1980
[121] J Dubuisson and C M Rice ldquoSindbis virus attachment isola-tion and characterization of mutants with impaired binding tovertebrate cellsrdquo Journal of Virology vol 67 no 6 pp 3363ndash33741993
[122] C Y Lee Y-W Kam J Fric et al ldquoChikungunya virus neutral-ization antigens and direct cell-to-cell transmission are revealedby human antibody-escapemutantsrdquo PLoS Pathogens vol 7 no12 Article ID e1002390 2011
[123] J E Voss M-C Vaney S Duquerroy et al ldquoGlycoproteinorganization of Chikungunya virus particles revealed by X-raycrystallographyrdquo Nature vol 468 no 7324 pp 709ndash712 2010
BioMed Research International 21
[124] L Li J Jose Y Xiang R J Kuhn and M G Rossmann ldquoStruc-tural changes of envelope proteins during alphavirus fusionrdquoNature vol 468 no 7324 pp 705ndash708 2010
[125] J Fric S Bertin-Maghit C I Wang A Nardin and L WarterldquoUse of human monoclonal antibodies to treat Chikungunyavirus infectionrdquo Journal of Infectious Diseases vol 207 no 2 pp319ndash322 2013
[126] P V Aguilar J G Estrada-Franco R Navarro-Lopez C FerroA D Haddow and S C Weaver ldquoEndemic Venezuelanequine encephalitis in the Americas hidden under the dengueumbrellardquo Future Virology vol 6 no 6 pp 721ndash740 2011
[127] S C Weaver R Winegar I D Manger and N L ForresterldquoAlphaviruses population genetics and determinants of emer-gencerdquo Antiviral Research vol 94 no 3 pp 242ndash257 2012
[128] S Paessler and S C Weaver ldquoVaccines for Venezuelan equineencephalitisrdquo Vaccine vol 27 no 4 pp D80ndashD85 2009
[129] R J Phillpotts ldquoVenezuelan equine encephalitis virus complex-specific monoclonal antibody provides broad protection inmurine models against airborne challenge with viruses fromserogroups I II and IIIrdquoVirus Research vol 120 no 1-2 pp 107ndash112 2006
[130] R J Phillpotts L D Jones and S C Howard ldquoMonoclonalantibody protectsmice against infection anddiseasewhen giveneither before or up to 24 h after airborne challenge with virulentVenezuelan equine encephalitis virusrdquoVaccine vol 20 no 11-12pp 1497ndash1504 2002
[131] J H Mathews J T Roehrig and D W Trent ldquoRole of com-plement and the Fc portion of immunoglobulin G in immunityto Venezuelan equine encephalomyelitis virus infection withglycoprotein-specific monoclonal antibodiesrdquo Journal of Virol-ogy vol 55 no 3 pp 594ndash600 1985
[132] L M OrsquoBrien C D Underwood-Fowler S A Goodchild AL Phelps and R J Phillpotts ldquoDevelopment of a novel mon-oclonal antibody with reactivity to a wide range of Venezuelanequine encephalitis virus strainsrdquoVirology Journal vol 6 article206 2009
[133] L M OrsquoBrien S A Goodchild R J Phillpotts and S D Per-kins ldquoA humanised murine monoclonal antibody protectsmice from Venezuelan equine encephalitis virus Evergladesvirus and Mucambo virus when administered up to 48h afterairborne challengerdquo Virology vol 426 no 2 pp 100ndash105 2012
[134] N Clementi N Mancini L Solforosi M Castelli M Clementiand R Burioni ldquoPhage display-based strategies for cloning andoptimization of monoclonal antibodies directed against humanpathogensrdquo International Journal of Molecular Sciences vol 13no 7 pp 8273ndash8292 2012
[135] A R Hunt R A Bowen S Frederickson T Maruyama JT Roehrig and C D Blair ldquoTreatment of mice with humanmonoclonal antibody 24h after lethal aerosol challenge withvirulent Venezuelan equine encephalitis virus prevents diseasebut not infectionrdquo Virology vol 414 no 2 pp 146ndash152 2011
[136] W-G Hu A L Phelps S Jager et al ldquoA recombinant human-ized monoclonal antibody completely protects mice againstlethal challenge with Venezuelan equine encephalitis virusrdquoVaccine vol 28 no 34 pp 5558ndash5564 2010
[137] T Rulker L Voss P Thullier et al ldquoIsolation and characterisa-tion of a human-like antibody fragment (scFv) that inactivatesVEEV in vitro and in vivordquo PLoS ONE vol 7 no 5 Article IDe37242 2012
[138] C T Walter and J N Barr ldquoRecent advances in the molecularand cellular biology of bunyavirusesrdquo Journal of General Virol-ogy vol 92 no 11 pp 2467ndash2484 2011
[139] C A Whitehouse ldquoCrimean-Congo hemorrhagic feverrdquo Anti-viral Research vol 64 no 3 pp 145ndash160 2004
[140] M Mertens K Schmidt A Ozkul and M H Groschup ldquoTheimpact of Crimean-Congo hemorrhagic fever virus on publichealthrdquo Antiviral Research vol 98 no 2 pp 248ndash260 2013
[141] A A Ahmed J M McFalls C Hoffmann et al ldquoPresence ofbroadly reactive and group-specific neutralizing epitopes onnewly described isolates of Crimean-Congo hemorrhagic fevervirusrdquo Journal of General Virology vol 86 no 12 pp 3327ndash33362005
[142] R Hewson A Gmyl L Gmyl et al ldquoEvidence of segment reas-sortment in Crimean-Congo haemorrhagic fever virusrdquo Journalof General Virology vol 85 no 10 pp 3059ndash3070 2004
[143] N K Blackburn T G Besselaar A J Shepherd and R Swane-poel ldquoPreparation and use of monoclonal antibodies for iden-tifying Crimean-Congo hemorrhagic fever virusrdquo AmericanJournal of Tropical Medicine and Hygiene vol 37 no 2 pp 392ndash397 1987
[144] T Ikegami ldquoMolecular biology and genetic diversity of RiftValley fever virusrdquoAntiviral Research vol 95 no 3 pp 293ndash3102012
[145] KKeegan andM S Collett ldquoUse of bacterial expression cloningto define the amino acid sequences of antigenic determinantson the G2 glycoprotein of Rift Valley fever virusrdquo Journal ofVirology vol 58 no 2 pp 263ndash270 1986
[146] T G Besselaar and N K Blackburn ldquoTopological mapping ofantigenic sites on the Rift Valley fever virus envelope glycopro-teins using monoclonal antibodiesrdquo Archives of Virology vol121 no 1ndash4 pp 111ndash124 1991
[147] L G Presta ldquoEngineering of therapeutic antibodies to mini-mize immunogenicity and optimize functionrdquo Advanced DrugDelivery Reviews vol 58 no 5-6 pp 640ndash656 2006
Submit your manuscripts athttpwwwhindawicom
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anatomy Research International
PeptidesInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporation httpwwwhindawicom
International Journal of
Volume 2014
Zoology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Molecular Biology International
GenomicsInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioinformaticsAdvances in
Marine BiologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Signal TransductionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Evolutionary BiologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Biochemistry Research International
ArchaeaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Genetics Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in
Virolog y
Hindawi Publishing Corporationhttpwwwhindawicom
Nucleic AcidsJournal of
Volume 2014
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Enzyme Research
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Microbiology

BioMed Research International 17
take placerdquo Current Opinion in Immunology vol 21 no 3 pp308ndash313 2009
[4] S B Halstead and S J Thomas ldquoNew vaccines for Japaneseencephalitisrdquo Current Infectious Disease Reports vol 12 no 3pp 174ndash180 2010
[5] R Burioni M Perotti N Mancini and M Clementi ldquoPer-spectives for the utilization of neutralizing human monoclonalantibodies as anti-HCV drugsrdquo Journal of Hepatology vol 49no 2 pp 299ndash300 2008
[6] R A Diotti G A Sautto L Solforosi NManciniM Clementiand R Burioni ldquoNeutralization activity and kinetics of twobroad-range human monoclonal IgG1 derived from recombi-nant Fab fragments and directed against Hepatitis C virus E2glycoproteinrdquo New Microbiologica vol 35 no 4 pp 475ndash4792012
[7] R Burioni F Canducci N Mancini et al ldquoMolecular cloningof the first humanmonoclona antibodies neutralizing with highpotency Swine-origin Influenza A pandemic virus (S-OIV)rdquoNew Microbiologica vol 32 no 4 pp 319ndash324 2009
[8] P K Chandra S Hazari B Poat et al ldquoIntracytoplasmic stableexpression of IgG1 antibody targeting NS3 helicase inhibitsreplication of highly efficient hepatitis C Virus 2a clonerdquoVirology Journal vol 7 article 118 2010
[9] N Clementi D de Marco N Mancini et al ldquoA human mono-clonal antibody with neutralizing activity against highly diver-gent influenza subtypesrdquo PLoS ONE vol 6 no 12 Article IDe28001 2011
[10] D de Marco N Clementi N Mancini et al ldquoA non-VH1-69 heterosubtypic neutralizing human monoclonal antibodyprotects mice against H1N1 and H5N1 virusesrdquo PLoS ONE vol7 no 4 Article ID e34415 2012
[11] L Solforosi N Mancini F Canducci et al ldquoA phage displayvector optimized for the generation of human antibody com-binatorial libraries and the molecular cloning of monoclonalantibody fragmentsrdquoNewMicrobiologica vol 35 no 3 pp 289ndash294 2012
[12] E V Koonin and V V Dolja ldquoEvolution and taxonomy ofpositive-strand RNA viruses implications of comparative anal-ysis of amino acid sequencesrdquo Critical Reviews in Biochemistryand Molecular Biology vol 28 no 5 pp 375ndash430 1993
[13] J S Mackenzie D J Gubler and L R Petersen ldquoEmergingflaviviruses the spread and resurgence of Japanese encephalitisWest Nile and dengue virusesrdquo Nature Medicine vol 10 no 12pp S98ndashS109 2004
[14] T J SmithW E Brandt J L Swanson J MMcCown and E LBuescher ldquoPhysical and biological properties of dengue-2 virusand associated antigensrdquo Journal of Virology vol 5 no 4 pp524ndash532 1970
[15] J M Smit B Moesker I Rodenhuis-Zybert and J WilschutldquoFlavivirus cell entry and membrane fusionrdquo Viruses vol 3 no2 pp 160ndash171 2011
[16] R J Kuhn W Zhang M G Rossmann et al ldquoStructure ofdengue virus implications for flavivirus organization matura-tion and fusionrdquo Cell vol 108 no 5 pp 717ndash725 2002
[17] K Stiasny J H Aberle M Keller B Grubeck-Loebenstein andF X Heinz ldquoAge affects quantity but not quality of antibodyresponses after vaccination with an inactivated flavivirus vac-cine against tick-borne encephalitisrdquo PLoS ONE vol 7 no 3Article ID e34145 2012
[18] LThomasOVerlaetenACabie et al ldquoInfluence of the dengueserotype previous dengue infection and plasma viral load
on clinical presentation and outcome during a dengue-2 anddengue-4 co-epidemicrdquo American Journal of Tropical Medicineand Hygiene vol 78 no 6 pp 990ndash998 2008
[19] Y Zhang J Corver P R Chipman et al ldquoStructures of imma-ture flavivirus particlesrdquo EMBO Journal vol 22 no 11 pp2604ndash2613 2003
[20] A K I Falconar ldquoIdentification of an epitope on the denguevirus membrane (M) protein defined by cross-protective mon-oclonal antibodies design of an improved epitope sequencebased on common determinants present in both envelope (EandM) proteinsrdquo Archives of Virology vol 144 no 12 pp 2313ndash2330 1999
[21] K-J Huang Y-C Yang Y-S Lin et al ldquoThe dual-specific bind-ing of dengue virus and target cells for the antibody-dependentenhancement of dengue virus infectionrdquo Journal of Immunol-ogy vol 176 no 5 pp 2825ndash2832 2006
[22] A K I Falconar ldquoAntibody responses are generated to immun-odominant ELKKLE-typemotifs on the nonstructural-1 glyco-protein during live dengue virus infections inmice and humansimplications for diagnosis pathogenesis and vaccine designrdquoClinical and Vaccine Immunology vol 14 no 5 pp 493ndash5042007
[23] C W Mandl F Guirakhoo H Holzmann F X Heinz and CKunz ldquoAntigenic structure of the flavivirus envelope protein Eat the molecular level using the tick-borne encephalitis virus asa modelrdquo Journal of Virology vol 63 no 2 pp 564ndash571 1989
[24] F A Rey F X Heinz C Mandl C Kunz and S C HarrisonldquoThe envelope glycoprotein from tick-borne encephalitis virusat 2 A resolutionrdquo Nature vol 375 no 6529 pp 291ndash298 1995
[25] S L Allison K Stiasny K Stadler CWMandl and F XHeinzldquoMapping of functional elements in the stem-anchor regionof tick-borne encephalitis virus envelope protein Erdquo Journal ofVirology vol 73 no 7 pp 5605ndash5612 1999
[26] D E Klein J L Choi and S C Harrison ldquoStructure of a denguevirus envelope protein late-stage fusion intermediaterdquo Journal ofVirology vol 87 no 4 pp 2287ndash2293 2013
[27] S L Hanna T C Pierson M D Sanchez A A Ahmed M MMurtadha and R W Doms ldquoN-linked glycosylation of WestNile virus envelope proteins influences particle assembly andinfectivityrdquo Journal of Virology vol 79 no 21 pp 13262ndash132742005
[28] N Mancini G Sautto N Clementi et al ldquoNeutralization inter-fering antibodies a ldquonovelrdquo example of humoral immune dys-function facilitating viral escaperdquoViruses vol 4 no 9 pp 1731ndash1752 2012
[29] Y Modis S Ogata D Clements and S C Harrison ldquoVariablesurface epitopes in the crystal structure of dengue virus type3 envelope glycoproteinrdquo Journal of Virology vol 79 no 2 pp1223ndash1231 2005
[30] N Miller ldquoRecent progress in dengue vaccine research anddevelopmentrdquo Current Opinion in Molecular Therapeutics vol12 no 1 pp 31ndash38 2010
[31] J H Chavez J R Silva A A Amarilla and L T Moraes Figu-eiredo ldquoDomain III peptides from flavivirus envelope proteinare useful antigens for serologic diagnosis and targets forimmunizationrdquo Biologicals vol 38 no 6 pp 613ndash618 2010
[32] B Shrestha J D Brien S Sukupolvi-Petty et al ldquoThe develop-ment of therapeutic antibodies that neutralize homologous andheterologous genotypes of dengue virus type 1rdquo PLoS Pathogensvol 6 no 4 Article ID e1000823 2010
18 BioMed Research International
[33] G D Gromowski N D Barrett and A D T Barrett ldquoChar-acterization of dengue virus complex-specific neutralizing epi-topes on envelope protein domain III of dengue 2 virusrdquo Journalof Virology vol 82 no 17 pp 8828ndash8837 2008
[34] S Sukupolvi-Petty S K Austin W E Purtha et al ldquoType- andsubcomplex-specific neutralizing antibodies against domain IIIof dengue virus type 2 envelope protein recognize adjacentepitopesrdquo Journal of Virology vol 81 no 23 pp 12816ndash128262007
[35] S Sukupolvi-Petty S K Austin M Engle et al ldquoStructure andfunction analysis of therapeutic monoclonal antibodies againstdengue virus type 2rdquo Journal of Virology vol 84 no 18 pp9227ndash9239 2010
[36] Y-Q Deng J-X Dai G-H Ji et al ldquoA broadly flaviviruscross-neutralizingmonoclonal antibody that recognizes a novelepitope within the fusion loop of e proteinrdquo PLoS ONE vol 6no 1 Article ID e16059 2011
[37] J M Costin E Zaitseva K M Kahle et al ldquoMechanistic studyof broadly neutralizing human monoclonal antibodies againstdengue virus that target the fusion looprdquo Journal of Virologyvol 87 no 1 pp 52ndash66 2013
[38] CMMidgley A FlanaganH B Tran et al ldquoStructural analysisof a dengue cross-reactive antibody complexed with envelopedomain III reveals the molecular basis of cross-reactivityrdquoJournal of Immunology vol 188 no 10 pp 4971ndash4979 2012
[39] N J Moreland P Susanto E Lim et al ldquoPhage display appro-aches for the isolation of monoclonal antibodies against denguevirus envelope domain III from human and mouse derivedlibrariesrdquo International Journal ofMolecular Sciences vol 13 no3 pp 2618ndash2635 2012
[40] J J B Cockburn M E Navarro Sanchez N Fretes et alldquoMechanism of dengue virus broad cross-neutralization by amonoclonal antibodyrdquo Structure vol 20 no 2 pp 303ndash3142012
[41] R Rajamanonmani C Nkenfou P Clancy et al ldquoOn a mousemonoclonal antibody that neutralizes all four dengue virusserotypesrdquo Journal of General Virology vol 90 no 4 pp 799ndash809 2009
[42] L H Gould J Sui H Foellmer et al ldquoProtective and herapeuticcapacity of human single-chain Fv-Fc fusion proteins againstWest Nile virusrdquo Journal of Virology vol 79 no 23 pp 14606ndash14613 2005
[43] H Sultana H G Foellmer G Neelakanta et al ldquoFusion LoopPeptide of the West Nile virus envelope protein is essential forpathogenesis and is recognized by a therapeutic cross-reactivehuman monoclonal antibodyrdquo Journal of Immunology vol 183no 1 pp 650ndash660 2009
[44] T Oliphant M Engle G E Nybakken et al ldquoDevelopment ofa humanized monoclonal antibody with therapeutic potentialagainstWest Nile virusrdquoNature Medicine vol 11 no 5 pp 522ndash530 2005
[45] G E Nybakken T Oliphant S Johnson S Burke M S Dia-mond and D H Fremont ldquoStructural basis of West Nile virusneutralization by a therapeutic antibodyrdquo Nature vol 437 no7059 pp 764ndash769 2005
[46] G D Gromowski J T Roehrig M S Diamond J C LeeT J Pitcher and A D T Barrett ldquoMutations of an antibodybinding energy hot spot on domain III of the dengue 2 envelopeglycoprotein exploited for neutralization escaperdquo Virology vol407 no 2 pp 237ndash246 2010
[47] J T Roehrig R A Bolin andR G Kelly ldquoMonoclonal antibodymapping of the envelope glycoprotein of the dengue 2 virusJamaicardquo Virology vol 246 no 2 pp 317ndash328 1998
[48] R de Alwis S A Smith N P Olivarez et al ldquoIdentification ofhumanneutralizing antibodies that bind to complex epitopes ondengue virionsrdquo Proceedings of the National Academy of Sciencesof the United States of America vol 109 no 19 pp 7439ndash74442012
[49] S K Austin K A Dowd B Shrestha et al ldquoStructural basis ofdifferential neutralization of DENV-1 genotypes by an antibodythat recognizes a cryptic epitoperdquo PLoS Pathogens vol 8 no 10Article ID e1002930 2012
[50] M Throsby C Geuijen J Goudsmit et al ldquoIsolation and char-acterization of human monoclonal antibodies from individualsinfected with West Nile Virusrdquo Journal of Virology vol 80 no14 pp 6982ndash6992 2006
[51] A P Goncalvez R H Purcell and C-J Lai ldquoEpitope deter-minants of a chimpanzee Fab antibody that efficiently cross-neutralizes dengue type 1 and type 2 viruses map to inside andin close proximity to fusion loop of the dengue type 2 virusenvelope glycoproteinrdquo Journal of Virology vol 78 no 23 pp12919ndash12928 2004
[52] J T Roehrig J H Mathews and D W Trent ldquoIdentification ofepitopes on the E glycoprotein of Saint Louis encephalitis virususing monoclonal antibodiesrdquo Virology vol 128 no 1 pp 118ndash126 1983
[53] J T Roehrig L A Staudinger A R Hunt J H Mathews andC D Blair ldquoAntibody prophylaxis and therapy for flavivirusencephalitis infectionsrdquo Annals of the New York Academy ofSciences vol 951 pp 286ndash297 2001
[54] A P Goncalvez C-H Chien K Tubthong et al ldquoHumanizedmonoclonal antibodies derived from chimpanzee fabs protectagainst Japanese encephalitis virus in vitro and in vivordquo Journalof Virology vol 82 no 14 pp 7009ndash7021 2008
[55] M Arakawa T Yamashiro G-I Uechi M Tadano and ANishizono ldquoConstruction of humanFab (1205741120581) library and iden-tification of human monoclonal Fab possessing neutralizingpotency against Japanese encephalitis virusrdquo Microbiology andImmunology vol 51 no 6 pp 617ndash625 2007
[56] L Warter C Y Lee R Thiagarajan et al ldquoChikungunya virusenvelope-specific human monoclonal antibodies with broadneutralization potencyrdquo Journal of Immunology vol 186 no 5pp 3258ndash3264 2011
[57] P Pal K A Dowd J D Brien et al ldquoDevelopment of a highlyprotective combination monoclonal antibody therapy againstchikungunya virusrdquo PLoS Pathogens vol 9 no 4 Article IDe1003312 2013
[58] A Bertolotti-Ciarlet J Smith K Strecker et al ldquoCellular local-ization and antigenic characterization of Crimean-Congo hem-orrhagic fever virus glycoproteinsrdquo Journal of Virology vol 79no 10 pp 6152ndash6161 2005
[59] J K Battles and J M Dalrymple ldquoGenetic variation amonggeographic isolates of Rift Valley fever virusrdquo American Journalof Tropical Medicine and Hygiene vol 39 no 6 pp 617ndash6311988
[60] B AThibodeaux N C Garbino NM Liss et al ldquoA humanizedIgG but not IgM antibody is effective in prophylaxis and therapyof yellow fever infection in an AG12917D-204 peripheralchallenge mouse modelrdquo Antiviral Research vol 94 no 1 pp1ndash8 2012
BioMed Research International 19
[61] S Daffis R E Kontermann J Korimbocus H Zeller H-DKlenk and J Ter Meulen ldquoAntibody responses against wild-type yellow fever virus and the 17D vaccine strain charac-terization with human monoclonal antibody fragments andneutralization escape variantsrdquoVirology vol 337 no 2 pp 262ndash272 2005
[62] L N Levanov L E Matveev E P Goncharova et al ldquoChimericantibodies against tick-borne encephalitis virusrdquo Vaccine vol28 no 32 pp 5265ndash5271 2010
[63] A R Hunt S Frederickson C Hinkel K S Bowdish and J TRoehrig ldquoA humanized murine monoclonal antibody protectsmice either before or after challenge with virulent Venezuelanequine encephalomyelitis virusrdquo Journal of General Virologyvol 87 no 9 pp 2467ndash2476 2006
[64] J M da Silva Voorham I A Rodenhuis-Zybert N V A Nunezet al ldquoAntibodies against the envelope glycoprotein promoteinfectivity of immature dengue virus serotype 2rdquo PLoS ONEvol 7 no 3 Article ID e29957 2012
[65] T C Pierson Q Xu S Nelson et al ldquoThe stoichiometry ofantibody-mediated neutralization and enhancement of westnile virus infectionrdquoCell Host andMicrobe vol 1 no 2 pp 135ndash145 2007
[66] G Sautto N Mancini R A Diotti L Solforosi M Clementiand R Burioni ldquoAnti-hepatitis C virus E2 (HCVE2) glyco-proteinmonoclonal antibodies andneutralization interferencerdquoAntiviral Research vol 96 no 1 pp 82ndash89 2012
[67] R Burioni Y Matsuura N Mancini et al ldquoDiverging effectsof human recombinant anti-hepatitis C virus (HCV) antibodyfragments derived from a single patient on the infectivityof a vesicular stomatitis virusHCV pseudotyperdquo Journal ofVirology vol 76 no 22 pp 11775ndash11779 2002
[68] R J Phillpotts J R Stephenson and J S Porterfield ldquoAntibody-dependent enhancement of tick-borne encephalitus virus infec-tivityrdquo Journal of General Virology vol 66 no 8 pp 1831ndash18371985
[69] K A Dowd and T C Pierson ldquoAntibody-mediated neutraliza-tion of flaviviruses a reductionist viewrdquo Virology vol 411 no 2pp 306ndash315 2011
[70] A K I Falconar ldquoMonoclonal antibodies that bind to commonepitopes on the dengue virus type 2 nonstructural-1 andenvelope glycoproteins display weak neutralizing activity anddifferentiated responses to virulent strains implications forpathogenesis and vaccinesrdquo Clinical and Vaccine Immunologyvol 15 no 3 pp 549ndash561 2008
[71] K Meyer A Banerjee S E Frey R B Belshe and R Ray ldquoAweak neutralizing antibody response to hepatitis C virus enve-lope glycoprotein enhances virus infectionrdquo PLoS ONE vol 6no 8 Article ID e23699 2011
[72] R Burioni R AWilliamson P P Sanna F E Bloom and D RBurton ldquoRecombinant human Fab to glycoprotein D neutral-izes infectivity and prevents cell-to-cell transmission of herpessimplex viruses 1 and 2 in vitrordquo Proceedings of the NationalAcademy of Sciences of the United States of America vol 91 no1 pp 355ndash359 1994
[73] M S Rolph A Zaid N E Rulli and S Mahalingam ldquoDown-regulation of interferon-120573 in antibody-dependent enhancementof dengue viral infections of human macrophages is dependenton interleukin-6rdquo Journal of Infectious Diseases vol 204 no 3pp 489ndash491 2011
[74] S S Whitehead J E Blaney A P Durbin and B R MurphyldquoProspects for a dengue virus vaccinerdquoNature ReviewsMicrobi-ology vol 5 no 7 pp 518ndash528 2007
[75] S-M Lok V Kostyuchenko G ENybakken et al ldquoBinding of aneutralizing antibody to dengue virus alters the arrangement ofsurface glycoproteinsrdquoNature Structural andMolecular Biologyvol 15 no 3 pp 312ndash317 2008
[76] A P Goncalvez R Men C Wernly R H Purcell and C-J Lai ldquoChimpanzee Fab fragments and a derived humanizedimmunogiobulin G1 antibody that efficiently cross-neutralizedengue type 1 and type 2 virusesrdquo Journal of Virology vol 78no 23 pp 12910ndash12918 2004
[77] WM P BWahala A A Kraus L B HaymoreM A Accavitti-Loper and A M de Silva ldquoDengue virus neutralization byhuman immune sera role of envelope protein domain III-reactive antibodyrdquo Virology vol 392 no 1 pp 103ndash113 2009
[78] W D Crill H R Hughes M J Delorey and G-J J ChangldquoHumoral immune responses of dengue fever patients usingepitope-specific serotype-2 virus-like particle antigensrdquo PLoSONE vol 4 no 4 Article ID e4991 2009
[79] C Setthapramote T Sasaki O Puiprom et al ldquoHuman mono-clonal antibodies to neutralize all dengue virus serotypes usinglymphocytes from patients at acute phase of the secondaryinfectionrdquo Biochemical and Biophysical Research Communica-tions vol 423 no 4 pp 867ndash872 2012
[80] M Beltramello K L Williams C P Simmons et al ldquoThehuman immune response to dengue virus is dominated byhighly cross-reactive antibodies endowed with neutralizing andenhancing activityrdquo Cell Host andMicrobe vol 8 no 3 pp 271ndash283 2010
[81] C M Midgley M Bajwa-Joseph S Vasanawathana et al ldquoAnin-depth analysis of original antigenic sin in dengue virusinfectionrdquo Journal of Virology vol 85 no 22 pp 410ndash421 2011
[82] B Kaufmann M R Vogt J Goudsmit et al ldquoNeutralizationof West Nile virus by cross-linking of its surface proteins withFab fragments of the human monoclonal antibody CR4354rdquoProceedings of the National Academy of Sciences of the UnitedStates of America vol 107 no 44 pp 18950ndash18955 2010
[83] B Spurrier J M Sampson M Totrov et al ldquoStructural analysisof human and macaque mAbs 2909 and 25B implicationsfor the configuration of the quaternary neutralizing epitope ofHIV-1 gp120rdquo Structure vol 19 no 5 pp 691ndash699 2011
[84] J J SchlesingerMWBrandriss andT PMonath ldquoMonoclonalantibodies distinguish between wild and vaccine strains ofyellow fever virus by neutralization hemagglutination inhibi-tion and immune precipitation of the virus envelope proteinrdquoVirology vol 125 no 1 pp 8ndash17 1983
[85] M W Brandriss J J Schlesinger E E Walsh and M BrisellildquoLethal 17D yellow fever encephalitis in mice I Passive protec-tion by monoclonal antibodies to the envelope proteins of 17Dyellow fever and dengue 2 virusesrdquo Journal of General Virologyvol 67 no 2 pp 229ndash234 1986
[86] H Cho and M S Diamond ldquoImmune responses to West Nilevirus infection in the central nervous systemrdquo Viruses vol 4no 12 pp 3812ndash3830 2012
[87] D W Beasley A D Barrett and R B Tesh ldquoResurgence ofWest Nile neurologic disease in the United States in 2012 whathappened What needs to be donerdquo Antiviral Research vol 99no 1 pp 1ndash5 2013
[88] M Throsby J Ter Meulen C Geuijen J Goudsmit and J DeKruif ldquoMapping and analysis of West Nile virus-specific mon-oclonal antibodies prospects for vaccine developmentrdquo ExpertReview of Vaccines vol 6 no 2 pp 183ndash191 2007
[89] M S Diamond T G Roberts D Edgil B Lu J Ernst and EHarris ldquoModulation of dengue virus infection in human cells
20 BioMed Research International
by alpha beta and gamma interferonsrdquo Journal of Virology vol74 no 11 pp 4957ndash4966 2000
[90] Z Shimoni M J Niven S Pitlick and S Bulvik ldquoTreatmentof West Nile virus encephalitis with intravenous immunoglob-ulinrdquo Emerging Infectious Diseases vol 7 no 4 article 759 2001
[91] AHamdan PGreen EMendelsonM R Kramer S Pitlik andM Weinberger ldquoPossible benefit of intravenous immunoglob-ulin therapy in a lung transplant recipient with West Nile virusencephalitisrdquoTransplant InfectiousDisease vol 4 no 3 pp 160ndash162 2002
[92] M Haley A S Retter D Fowler J Gea-Banacloche and NP OrsquoGrady ldquoThe role for intravenous immunoglobulin in thetreatment of West Nile virus encephalitisrdquo Clinical InfectiousDiseases vol 37 no 6 pp e88ndashe90 2003
[93] K M Chung G E Nybakken B S Thompson et al ldquoAnti-bodies against West Nile virus nonstructural protein NS1prevent lethal infection through Fc 120574 receptor-dependent and-independent mechanismsrdquo Journal of Virology vol 80 no 3pp 1340ndash1351 2006
[94] E A Henchal L S Henchal and J J Schlesinger ldquoSynergisticinteractions of anti-NS1 monoclonal antibodies protect pas-sively immunized mice from lethal challenge with dengue 2virusrdquo Journal of General Virology vol 69 no 8 pp 2101ndash21071988
[95] J T Roehrig ldquoAntigenic structure of flavivirus proteinsrdquoAdvances in Virus Research vol 59 pp 141ndash175 2003
[96] M D Sanchez T C Pierson D McAllister et al ldquoCharacteri-zation of neutralizing antibodies to West Nile virusrdquo Virologyvol 336 no 1 pp 70ndash82 2005
[97] J H Beigel J L Nordstrom S R Pillemer et al ldquoSafety andpharmacokinetics of single intravenous dose of MGAWN1 anovel monoclonal antibody to West Nile virusrdquo AntimicrobialAgents and Chemotherapy vol 54 no 6 pp 2431ndash2436 2010
[98] C A Smeraski V Siddharthan and J D Morrey ldquoTreatmentof spatial memory impairment in hamsters infected with WestNile virus using a humanizedmonoclonal antibodyMGAWN1rdquoAntiviral Research vol 91 no 1 pp 43ndash49 2011
[99] D Lelli AMoreno E Brocchi et al ldquoWestNile virus character-ization and diagnostic applications of monoclonal antibodiesrdquoVirology Journal vol 9 article 81 2012
[100] T S Gritsun V A Lashkevich and E A Gould ldquoTick-borneencephalitisrdquo Antiviral Research vol 57 no 1-2 pp 129ndash1462003
[101] D Ruzek T S Gritsun N L Forrester et al ldquoMutations inthe NS2B and NS3 genes affect mouse neuroinvasiveness of aWestern European field strain of tick-borne encephalitis virusrdquoVirology vol 374 no 2 pp 249ndash255 2008
[102] M Ecker S L Allison T Meixner and F X Heinz ldquoSequenceanalysis and genetic classification of tick-borne encephalitisviruses from Europe and Asiardquo Journal of General Virology vol80 no 1 pp 179ndash185 1999
[103] H Holzmann M S Vorobyova I P Ladyzhenskaya et alldquoMolecular epidemiology of tick-borne encephalitis viruscross-protection between European and Far Eastern subtypesrdquoVaccine vol 10 no 5 pp 345ndash349 1992
[104] C W Mandl ldquoSteps of the tick-borne encephalitis virus repli-cation cycle that affect neuropathogenesisrdquo Virus Research vol111 no 2 pp 161ndash174 2005
[105] A V Atrasheuskaya T M Fredeking and G M IgnatyevldquoChanges in immune parameters and their correction in humancases of tick-borne encephalitisrdquo Clinical and ExperimentalImmunology vol 131 no 1 pp 148ndash154 2003
[106] K L Mansfield N Johnson L P Phipps J R Stephenson AR Fooks and T Solomon ldquoTick-borne encephalitis virusmdashareview of an emerging zoonosisrdquo Journal of General Virologyvol 90 no 8 pp 1781ndash1794 2009
[107] DWVaughn andCHHoke Jr ldquoThe epidemiology of Japaneseencephalitis prospects for preventionrdquo Epidemiologic Reviewsvol 14 pp 197ndash221 1992
[108] T Solomon H Ni D W C Beasley M Ekkelenkamp M JCardosa and A D T Barrett ldquoOrigin and evolution of Japaneseencephalitis virus in Southeast Asiardquo Journal of Virology vol 77no 5 pp 3091ndash3098 2003
[109] M R Wilson ldquoEmerging viral infectionsrdquo Current Opinion inNeurology vol 26 no 3 pp 301ndash306 2013
[110] J N Hanna S A Ritchie D A Phillips et al ldquoAn outbreakof Japanese encephalitis in the Torres Strait Australia 1995rdquoMedical Journal of Australia vol 165 no 5 pp 256ndash260 1996
[111] J N Hanna S A Ritchie D A Phillips et al ldquoJapanese ence-phalitis in north Queensland Australia 1998rdquo Medical Journalof Australia vol 170 no 11 pp 533ndash536 1999
[112] SGeorgeMGourie-Devi and J A Rao ldquoIsolation ofWestNilevirus from the brains of children who had died of encephalitisrdquoBulletin of theWorldHealthOrganization vol 62 no 6 pp 879ndash882 1984
[113] A K Gupta A A Koshy and V J Lad ldquoEnhanced protec-tion of mice against Japanese encephalitis virus infection bycombinations ofmonoclonal antibodies to glycoprotein ErdquoActaVirologica vol 55 no 2 pp 165ndash168 2011
[114] T H Lee B-H Song S-I Yun et al ldquoA cross-protective mAbrecognizes a novel epitope within the flavivirus NS1 proteinrdquoJournal of General Virology vol 93 no 1 pp 20ndash26 2012
[115] S C Weaver ldquoHost range amplification and arboviral diseaseemergencerdquo Archives of Virology Supplementum no 19 pp 33ndash44 2005
[116] M Solignat B Gay S Higgs L Briant and C Devaux ldquoRepli-cation cycle of chikungunya a re-emerging arbovirusrdquoVirologyvol 393 no 2 pp 183ndash197 2009
[117] O Schwartz and M L Albert ldquoBiology and pathogenesis ofchikungunya virusrdquo Nature Reviews Microbiology vol 8 no 7pp 491ndash500 2010
[118] W Akahata Z-Y Yang H Andersen et al ldquoA virus-like particlevaccine for epidemic Chikungunya virus protects nonhumanprimates against infectionrdquo Nature Medicine vol 16 no 3 pp334ndash338 2010
[119] M Kielian and A Helenius ldquopH-Iinduced alterations in thefusogenic spike protein of Semliki Forest virusrdquo Journal of CellBiology vol 101 no 6 pp 2284ndash2291 1985
[120] J White and A Helenius ldquopH-Dependent fusion between theSemliki Forest virus membrane and liposomesrdquo Proceedings ofthe National Academy of Sciences of the United States of Americavol 77 no 6 pp 3273ndash3277 1980
[121] J Dubuisson and C M Rice ldquoSindbis virus attachment isola-tion and characterization of mutants with impaired binding tovertebrate cellsrdquo Journal of Virology vol 67 no 6 pp 3363ndash33741993
[122] C Y Lee Y-W Kam J Fric et al ldquoChikungunya virus neutral-ization antigens and direct cell-to-cell transmission are revealedby human antibody-escapemutantsrdquo PLoS Pathogens vol 7 no12 Article ID e1002390 2011
[123] J E Voss M-C Vaney S Duquerroy et al ldquoGlycoproteinorganization of Chikungunya virus particles revealed by X-raycrystallographyrdquo Nature vol 468 no 7324 pp 709ndash712 2010
BioMed Research International 21
[124] L Li J Jose Y Xiang R J Kuhn and M G Rossmann ldquoStruc-tural changes of envelope proteins during alphavirus fusionrdquoNature vol 468 no 7324 pp 705ndash708 2010
[125] J Fric S Bertin-Maghit C I Wang A Nardin and L WarterldquoUse of human monoclonal antibodies to treat Chikungunyavirus infectionrdquo Journal of Infectious Diseases vol 207 no 2 pp319ndash322 2013
[126] P V Aguilar J G Estrada-Franco R Navarro-Lopez C FerroA D Haddow and S C Weaver ldquoEndemic Venezuelanequine encephalitis in the Americas hidden under the dengueumbrellardquo Future Virology vol 6 no 6 pp 721ndash740 2011
[127] S C Weaver R Winegar I D Manger and N L ForresterldquoAlphaviruses population genetics and determinants of emer-gencerdquo Antiviral Research vol 94 no 3 pp 242ndash257 2012
[128] S Paessler and S C Weaver ldquoVaccines for Venezuelan equineencephalitisrdquo Vaccine vol 27 no 4 pp D80ndashD85 2009
[129] R J Phillpotts ldquoVenezuelan equine encephalitis virus complex-specific monoclonal antibody provides broad protection inmurine models against airborne challenge with viruses fromserogroups I II and IIIrdquoVirus Research vol 120 no 1-2 pp 107ndash112 2006
[130] R J Phillpotts L D Jones and S C Howard ldquoMonoclonalantibody protectsmice against infection anddiseasewhen giveneither before or up to 24 h after airborne challenge with virulentVenezuelan equine encephalitis virusrdquoVaccine vol 20 no 11-12pp 1497ndash1504 2002
[131] J H Mathews J T Roehrig and D W Trent ldquoRole of com-plement and the Fc portion of immunoglobulin G in immunityto Venezuelan equine encephalomyelitis virus infection withglycoprotein-specific monoclonal antibodiesrdquo Journal of Virol-ogy vol 55 no 3 pp 594ndash600 1985
[132] L M OrsquoBrien C D Underwood-Fowler S A Goodchild AL Phelps and R J Phillpotts ldquoDevelopment of a novel mon-oclonal antibody with reactivity to a wide range of Venezuelanequine encephalitis virus strainsrdquoVirology Journal vol 6 article206 2009
[133] L M OrsquoBrien S A Goodchild R J Phillpotts and S D Per-kins ldquoA humanised murine monoclonal antibody protectsmice from Venezuelan equine encephalitis virus Evergladesvirus and Mucambo virus when administered up to 48h afterairborne challengerdquo Virology vol 426 no 2 pp 100ndash105 2012
[134] N Clementi N Mancini L Solforosi M Castelli M Clementiand R Burioni ldquoPhage display-based strategies for cloning andoptimization of monoclonal antibodies directed against humanpathogensrdquo International Journal of Molecular Sciences vol 13no 7 pp 8273ndash8292 2012
[135] A R Hunt R A Bowen S Frederickson T Maruyama JT Roehrig and C D Blair ldquoTreatment of mice with humanmonoclonal antibody 24h after lethal aerosol challenge withvirulent Venezuelan equine encephalitis virus prevents diseasebut not infectionrdquo Virology vol 414 no 2 pp 146ndash152 2011
[136] W-G Hu A L Phelps S Jager et al ldquoA recombinant human-ized monoclonal antibody completely protects mice againstlethal challenge with Venezuelan equine encephalitis virusrdquoVaccine vol 28 no 34 pp 5558ndash5564 2010
[137] T Rulker L Voss P Thullier et al ldquoIsolation and characterisa-tion of a human-like antibody fragment (scFv) that inactivatesVEEV in vitro and in vivordquo PLoS ONE vol 7 no 5 Article IDe37242 2012
[138] C T Walter and J N Barr ldquoRecent advances in the molecularand cellular biology of bunyavirusesrdquo Journal of General Virol-ogy vol 92 no 11 pp 2467ndash2484 2011
[139] C A Whitehouse ldquoCrimean-Congo hemorrhagic feverrdquo Anti-viral Research vol 64 no 3 pp 145ndash160 2004
[140] M Mertens K Schmidt A Ozkul and M H Groschup ldquoTheimpact of Crimean-Congo hemorrhagic fever virus on publichealthrdquo Antiviral Research vol 98 no 2 pp 248ndash260 2013
[141] A A Ahmed J M McFalls C Hoffmann et al ldquoPresence ofbroadly reactive and group-specific neutralizing epitopes onnewly described isolates of Crimean-Congo hemorrhagic fevervirusrdquo Journal of General Virology vol 86 no 12 pp 3327ndash33362005
[142] R Hewson A Gmyl L Gmyl et al ldquoEvidence of segment reas-sortment in Crimean-Congo haemorrhagic fever virusrdquo Journalof General Virology vol 85 no 10 pp 3059ndash3070 2004
[143] N K Blackburn T G Besselaar A J Shepherd and R Swane-poel ldquoPreparation and use of monoclonal antibodies for iden-tifying Crimean-Congo hemorrhagic fever virusrdquo AmericanJournal of Tropical Medicine and Hygiene vol 37 no 2 pp 392ndash397 1987
[144] T Ikegami ldquoMolecular biology and genetic diversity of RiftValley fever virusrdquoAntiviral Research vol 95 no 3 pp 293ndash3102012
[145] KKeegan andM S Collett ldquoUse of bacterial expression cloningto define the amino acid sequences of antigenic determinantson the G2 glycoprotein of Rift Valley fever virusrdquo Journal ofVirology vol 58 no 2 pp 263ndash270 1986
[146] T G Besselaar and N K Blackburn ldquoTopological mapping ofantigenic sites on the Rift Valley fever virus envelope glycopro-teins using monoclonal antibodiesrdquo Archives of Virology vol121 no 1ndash4 pp 111ndash124 1991
[147] L G Presta ldquoEngineering of therapeutic antibodies to mini-mize immunogenicity and optimize functionrdquo Advanced DrugDelivery Reviews vol 58 no 5-6 pp 640ndash656 2006
Submit your manuscripts athttpwwwhindawicom
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anatomy Research International
PeptidesInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporation httpwwwhindawicom
International Journal of
Volume 2014
Zoology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Molecular Biology International
GenomicsInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioinformaticsAdvances in
Marine BiologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Signal TransductionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Evolutionary BiologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Biochemistry Research International
ArchaeaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Genetics Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in
Virolog y
Hindawi Publishing Corporationhttpwwwhindawicom
Nucleic AcidsJournal of
Volume 2014
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Enzyme Research
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Microbiology

18 BioMed Research International
[33] G D Gromowski N D Barrett and A D T Barrett ldquoChar-acterization of dengue virus complex-specific neutralizing epi-topes on envelope protein domain III of dengue 2 virusrdquo Journalof Virology vol 82 no 17 pp 8828ndash8837 2008
[34] S Sukupolvi-Petty S K Austin W E Purtha et al ldquoType- andsubcomplex-specific neutralizing antibodies against domain IIIof dengue virus type 2 envelope protein recognize adjacentepitopesrdquo Journal of Virology vol 81 no 23 pp 12816ndash128262007
[35] S Sukupolvi-Petty S K Austin M Engle et al ldquoStructure andfunction analysis of therapeutic monoclonal antibodies againstdengue virus type 2rdquo Journal of Virology vol 84 no 18 pp9227ndash9239 2010
[36] Y-Q Deng J-X Dai G-H Ji et al ldquoA broadly flaviviruscross-neutralizingmonoclonal antibody that recognizes a novelepitope within the fusion loop of e proteinrdquo PLoS ONE vol 6no 1 Article ID e16059 2011
[37] J M Costin E Zaitseva K M Kahle et al ldquoMechanistic studyof broadly neutralizing human monoclonal antibodies againstdengue virus that target the fusion looprdquo Journal of Virologyvol 87 no 1 pp 52ndash66 2013
[38] CMMidgley A FlanaganH B Tran et al ldquoStructural analysisof a dengue cross-reactive antibody complexed with envelopedomain III reveals the molecular basis of cross-reactivityrdquoJournal of Immunology vol 188 no 10 pp 4971ndash4979 2012
[39] N J Moreland P Susanto E Lim et al ldquoPhage display appro-aches for the isolation of monoclonal antibodies against denguevirus envelope domain III from human and mouse derivedlibrariesrdquo International Journal ofMolecular Sciences vol 13 no3 pp 2618ndash2635 2012
[40] J J B Cockburn M E Navarro Sanchez N Fretes et alldquoMechanism of dengue virus broad cross-neutralization by amonoclonal antibodyrdquo Structure vol 20 no 2 pp 303ndash3142012
[41] R Rajamanonmani C Nkenfou P Clancy et al ldquoOn a mousemonoclonal antibody that neutralizes all four dengue virusserotypesrdquo Journal of General Virology vol 90 no 4 pp 799ndash809 2009
[42] L H Gould J Sui H Foellmer et al ldquoProtective and herapeuticcapacity of human single-chain Fv-Fc fusion proteins againstWest Nile virusrdquo Journal of Virology vol 79 no 23 pp 14606ndash14613 2005
[43] H Sultana H G Foellmer G Neelakanta et al ldquoFusion LoopPeptide of the West Nile virus envelope protein is essential forpathogenesis and is recognized by a therapeutic cross-reactivehuman monoclonal antibodyrdquo Journal of Immunology vol 183no 1 pp 650ndash660 2009
[44] T Oliphant M Engle G E Nybakken et al ldquoDevelopment ofa humanized monoclonal antibody with therapeutic potentialagainstWest Nile virusrdquoNature Medicine vol 11 no 5 pp 522ndash530 2005
[45] G E Nybakken T Oliphant S Johnson S Burke M S Dia-mond and D H Fremont ldquoStructural basis of West Nile virusneutralization by a therapeutic antibodyrdquo Nature vol 437 no7059 pp 764ndash769 2005
[46] G D Gromowski J T Roehrig M S Diamond J C LeeT J Pitcher and A D T Barrett ldquoMutations of an antibodybinding energy hot spot on domain III of the dengue 2 envelopeglycoprotein exploited for neutralization escaperdquo Virology vol407 no 2 pp 237ndash246 2010
[47] J T Roehrig R A Bolin andR G Kelly ldquoMonoclonal antibodymapping of the envelope glycoprotein of the dengue 2 virusJamaicardquo Virology vol 246 no 2 pp 317ndash328 1998
[48] R de Alwis S A Smith N P Olivarez et al ldquoIdentification ofhumanneutralizing antibodies that bind to complex epitopes ondengue virionsrdquo Proceedings of the National Academy of Sciencesof the United States of America vol 109 no 19 pp 7439ndash74442012
[49] S K Austin K A Dowd B Shrestha et al ldquoStructural basis ofdifferential neutralization of DENV-1 genotypes by an antibodythat recognizes a cryptic epitoperdquo PLoS Pathogens vol 8 no 10Article ID e1002930 2012
[50] M Throsby C Geuijen J Goudsmit et al ldquoIsolation and char-acterization of human monoclonal antibodies from individualsinfected with West Nile Virusrdquo Journal of Virology vol 80 no14 pp 6982ndash6992 2006
[51] A P Goncalvez R H Purcell and C-J Lai ldquoEpitope deter-minants of a chimpanzee Fab antibody that efficiently cross-neutralizes dengue type 1 and type 2 viruses map to inside andin close proximity to fusion loop of the dengue type 2 virusenvelope glycoproteinrdquo Journal of Virology vol 78 no 23 pp12919ndash12928 2004
[52] J T Roehrig J H Mathews and D W Trent ldquoIdentification ofepitopes on the E glycoprotein of Saint Louis encephalitis virususing monoclonal antibodiesrdquo Virology vol 128 no 1 pp 118ndash126 1983
[53] J T Roehrig L A Staudinger A R Hunt J H Mathews andC D Blair ldquoAntibody prophylaxis and therapy for flavivirusencephalitis infectionsrdquo Annals of the New York Academy ofSciences vol 951 pp 286ndash297 2001
[54] A P Goncalvez C-H Chien K Tubthong et al ldquoHumanizedmonoclonal antibodies derived from chimpanzee fabs protectagainst Japanese encephalitis virus in vitro and in vivordquo Journalof Virology vol 82 no 14 pp 7009ndash7021 2008
[55] M Arakawa T Yamashiro G-I Uechi M Tadano and ANishizono ldquoConstruction of humanFab (1205741120581) library and iden-tification of human monoclonal Fab possessing neutralizingpotency against Japanese encephalitis virusrdquo Microbiology andImmunology vol 51 no 6 pp 617ndash625 2007
[56] L Warter C Y Lee R Thiagarajan et al ldquoChikungunya virusenvelope-specific human monoclonal antibodies with broadneutralization potencyrdquo Journal of Immunology vol 186 no 5pp 3258ndash3264 2011
[57] P Pal K A Dowd J D Brien et al ldquoDevelopment of a highlyprotective combination monoclonal antibody therapy againstchikungunya virusrdquo PLoS Pathogens vol 9 no 4 Article IDe1003312 2013
[58] A Bertolotti-Ciarlet J Smith K Strecker et al ldquoCellular local-ization and antigenic characterization of Crimean-Congo hem-orrhagic fever virus glycoproteinsrdquo Journal of Virology vol 79no 10 pp 6152ndash6161 2005
[59] J K Battles and J M Dalrymple ldquoGenetic variation amonggeographic isolates of Rift Valley fever virusrdquo American Journalof Tropical Medicine and Hygiene vol 39 no 6 pp 617ndash6311988
[60] B AThibodeaux N C Garbino NM Liss et al ldquoA humanizedIgG but not IgM antibody is effective in prophylaxis and therapyof yellow fever infection in an AG12917D-204 peripheralchallenge mouse modelrdquo Antiviral Research vol 94 no 1 pp1ndash8 2012
BioMed Research International 19
[61] S Daffis R E Kontermann J Korimbocus H Zeller H-DKlenk and J Ter Meulen ldquoAntibody responses against wild-type yellow fever virus and the 17D vaccine strain charac-terization with human monoclonal antibody fragments andneutralization escape variantsrdquoVirology vol 337 no 2 pp 262ndash272 2005
[62] L N Levanov L E Matveev E P Goncharova et al ldquoChimericantibodies against tick-borne encephalitis virusrdquo Vaccine vol28 no 32 pp 5265ndash5271 2010
[63] A R Hunt S Frederickson C Hinkel K S Bowdish and J TRoehrig ldquoA humanized murine monoclonal antibody protectsmice either before or after challenge with virulent Venezuelanequine encephalomyelitis virusrdquo Journal of General Virologyvol 87 no 9 pp 2467ndash2476 2006
[64] J M da Silva Voorham I A Rodenhuis-Zybert N V A Nunezet al ldquoAntibodies against the envelope glycoprotein promoteinfectivity of immature dengue virus serotype 2rdquo PLoS ONEvol 7 no 3 Article ID e29957 2012
[65] T C Pierson Q Xu S Nelson et al ldquoThe stoichiometry ofantibody-mediated neutralization and enhancement of westnile virus infectionrdquoCell Host andMicrobe vol 1 no 2 pp 135ndash145 2007
[66] G Sautto N Mancini R A Diotti L Solforosi M Clementiand R Burioni ldquoAnti-hepatitis C virus E2 (HCVE2) glyco-proteinmonoclonal antibodies andneutralization interferencerdquoAntiviral Research vol 96 no 1 pp 82ndash89 2012
[67] R Burioni Y Matsuura N Mancini et al ldquoDiverging effectsof human recombinant anti-hepatitis C virus (HCV) antibodyfragments derived from a single patient on the infectivityof a vesicular stomatitis virusHCV pseudotyperdquo Journal ofVirology vol 76 no 22 pp 11775ndash11779 2002
[68] R J Phillpotts J R Stephenson and J S Porterfield ldquoAntibody-dependent enhancement of tick-borne encephalitus virus infec-tivityrdquo Journal of General Virology vol 66 no 8 pp 1831ndash18371985
[69] K A Dowd and T C Pierson ldquoAntibody-mediated neutraliza-tion of flaviviruses a reductionist viewrdquo Virology vol 411 no 2pp 306ndash315 2011
[70] A K I Falconar ldquoMonoclonal antibodies that bind to commonepitopes on the dengue virus type 2 nonstructural-1 andenvelope glycoproteins display weak neutralizing activity anddifferentiated responses to virulent strains implications forpathogenesis and vaccinesrdquo Clinical and Vaccine Immunologyvol 15 no 3 pp 549ndash561 2008
[71] K Meyer A Banerjee S E Frey R B Belshe and R Ray ldquoAweak neutralizing antibody response to hepatitis C virus enve-lope glycoprotein enhances virus infectionrdquo PLoS ONE vol 6no 8 Article ID e23699 2011
[72] R Burioni R AWilliamson P P Sanna F E Bloom and D RBurton ldquoRecombinant human Fab to glycoprotein D neutral-izes infectivity and prevents cell-to-cell transmission of herpessimplex viruses 1 and 2 in vitrordquo Proceedings of the NationalAcademy of Sciences of the United States of America vol 91 no1 pp 355ndash359 1994
[73] M S Rolph A Zaid N E Rulli and S Mahalingam ldquoDown-regulation of interferon-120573 in antibody-dependent enhancementof dengue viral infections of human macrophages is dependenton interleukin-6rdquo Journal of Infectious Diseases vol 204 no 3pp 489ndash491 2011
[74] S S Whitehead J E Blaney A P Durbin and B R MurphyldquoProspects for a dengue virus vaccinerdquoNature ReviewsMicrobi-ology vol 5 no 7 pp 518ndash528 2007
[75] S-M Lok V Kostyuchenko G ENybakken et al ldquoBinding of aneutralizing antibody to dengue virus alters the arrangement ofsurface glycoproteinsrdquoNature Structural andMolecular Biologyvol 15 no 3 pp 312ndash317 2008
[76] A P Goncalvez R Men C Wernly R H Purcell and C-J Lai ldquoChimpanzee Fab fragments and a derived humanizedimmunogiobulin G1 antibody that efficiently cross-neutralizedengue type 1 and type 2 virusesrdquo Journal of Virology vol 78no 23 pp 12910ndash12918 2004
[77] WM P BWahala A A Kraus L B HaymoreM A Accavitti-Loper and A M de Silva ldquoDengue virus neutralization byhuman immune sera role of envelope protein domain III-reactive antibodyrdquo Virology vol 392 no 1 pp 103ndash113 2009
[78] W D Crill H R Hughes M J Delorey and G-J J ChangldquoHumoral immune responses of dengue fever patients usingepitope-specific serotype-2 virus-like particle antigensrdquo PLoSONE vol 4 no 4 Article ID e4991 2009
[79] C Setthapramote T Sasaki O Puiprom et al ldquoHuman mono-clonal antibodies to neutralize all dengue virus serotypes usinglymphocytes from patients at acute phase of the secondaryinfectionrdquo Biochemical and Biophysical Research Communica-tions vol 423 no 4 pp 867ndash872 2012
[80] M Beltramello K L Williams C P Simmons et al ldquoThehuman immune response to dengue virus is dominated byhighly cross-reactive antibodies endowed with neutralizing andenhancing activityrdquo Cell Host andMicrobe vol 8 no 3 pp 271ndash283 2010
[81] C M Midgley M Bajwa-Joseph S Vasanawathana et al ldquoAnin-depth analysis of original antigenic sin in dengue virusinfectionrdquo Journal of Virology vol 85 no 22 pp 410ndash421 2011
[82] B Kaufmann M R Vogt J Goudsmit et al ldquoNeutralizationof West Nile virus by cross-linking of its surface proteins withFab fragments of the human monoclonal antibody CR4354rdquoProceedings of the National Academy of Sciences of the UnitedStates of America vol 107 no 44 pp 18950ndash18955 2010
[83] B Spurrier J M Sampson M Totrov et al ldquoStructural analysisof human and macaque mAbs 2909 and 25B implicationsfor the configuration of the quaternary neutralizing epitope ofHIV-1 gp120rdquo Structure vol 19 no 5 pp 691ndash699 2011
[84] J J SchlesingerMWBrandriss andT PMonath ldquoMonoclonalantibodies distinguish between wild and vaccine strains ofyellow fever virus by neutralization hemagglutination inhibi-tion and immune precipitation of the virus envelope proteinrdquoVirology vol 125 no 1 pp 8ndash17 1983
[85] M W Brandriss J J Schlesinger E E Walsh and M BrisellildquoLethal 17D yellow fever encephalitis in mice I Passive protec-tion by monoclonal antibodies to the envelope proteins of 17Dyellow fever and dengue 2 virusesrdquo Journal of General Virologyvol 67 no 2 pp 229ndash234 1986
[86] H Cho and M S Diamond ldquoImmune responses to West Nilevirus infection in the central nervous systemrdquo Viruses vol 4no 12 pp 3812ndash3830 2012
[87] D W Beasley A D Barrett and R B Tesh ldquoResurgence ofWest Nile neurologic disease in the United States in 2012 whathappened What needs to be donerdquo Antiviral Research vol 99no 1 pp 1ndash5 2013
[88] M Throsby J Ter Meulen C Geuijen J Goudsmit and J DeKruif ldquoMapping and analysis of West Nile virus-specific mon-oclonal antibodies prospects for vaccine developmentrdquo ExpertReview of Vaccines vol 6 no 2 pp 183ndash191 2007
[89] M S Diamond T G Roberts D Edgil B Lu J Ernst and EHarris ldquoModulation of dengue virus infection in human cells
20 BioMed Research International
by alpha beta and gamma interferonsrdquo Journal of Virology vol74 no 11 pp 4957ndash4966 2000
[90] Z Shimoni M J Niven S Pitlick and S Bulvik ldquoTreatmentof West Nile virus encephalitis with intravenous immunoglob-ulinrdquo Emerging Infectious Diseases vol 7 no 4 article 759 2001
[91] AHamdan PGreen EMendelsonM R Kramer S Pitlik andM Weinberger ldquoPossible benefit of intravenous immunoglob-ulin therapy in a lung transplant recipient with West Nile virusencephalitisrdquoTransplant InfectiousDisease vol 4 no 3 pp 160ndash162 2002
[92] M Haley A S Retter D Fowler J Gea-Banacloche and NP OrsquoGrady ldquoThe role for intravenous immunoglobulin in thetreatment of West Nile virus encephalitisrdquo Clinical InfectiousDiseases vol 37 no 6 pp e88ndashe90 2003
[93] K M Chung G E Nybakken B S Thompson et al ldquoAnti-bodies against West Nile virus nonstructural protein NS1prevent lethal infection through Fc 120574 receptor-dependent and-independent mechanismsrdquo Journal of Virology vol 80 no 3pp 1340ndash1351 2006
[94] E A Henchal L S Henchal and J J Schlesinger ldquoSynergisticinteractions of anti-NS1 monoclonal antibodies protect pas-sively immunized mice from lethal challenge with dengue 2virusrdquo Journal of General Virology vol 69 no 8 pp 2101ndash21071988
[95] J T Roehrig ldquoAntigenic structure of flavivirus proteinsrdquoAdvances in Virus Research vol 59 pp 141ndash175 2003
[96] M D Sanchez T C Pierson D McAllister et al ldquoCharacteri-zation of neutralizing antibodies to West Nile virusrdquo Virologyvol 336 no 1 pp 70ndash82 2005
[97] J H Beigel J L Nordstrom S R Pillemer et al ldquoSafety andpharmacokinetics of single intravenous dose of MGAWN1 anovel monoclonal antibody to West Nile virusrdquo AntimicrobialAgents and Chemotherapy vol 54 no 6 pp 2431ndash2436 2010
[98] C A Smeraski V Siddharthan and J D Morrey ldquoTreatmentof spatial memory impairment in hamsters infected with WestNile virus using a humanizedmonoclonal antibodyMGAWN1rdquoAntiviral Research vol 91 no 1 pp 43ndash49 2011
[99] D Lelli AMoreno E Brocchi et al ldquoWestNile virus character-ization and diagnostic applications of monoclonal antibodiesrdquoVirology Journal vol 9 article 81 2012
[100] T S Gritsun V A Lashkevich and E A Gould ldquoTick-borneencephalitisrdquo Antiviral Research vol 57 no 1-2 pp 129ndash1462003
[101] D Ruzek T S Gritsun N L Forrester et al ldquoMutations inthe NS2B and NS3 genes affect mouse neuroinvasiveness of aWestern European field strain of tick-borne encephalitis virusrdquoVirology vol 374 no 2 pp 249ndash255 2008
[102] M Ecker S L Allison T Meixner and F X Heinz ldquoSequenceanalysis and genetic classification of tick-borne encephalitisviruses from Europe and Asiardquo Journal of General Virology vol80 no 1 pp 179ndash185 1999
[103] H Holzmann M S Vorobyova I P Ladyzhenskaya et alldquoMolecular epidemiology of tick-borne encephalitis viruscross-protection between European and Far Eastern subtypesrdquoVaccine vol 10 no 5 pp 345ndash349 1992
[104] C W Mandl ldquoSteps of the tick-borne encephalitis virus repli-cation cycle that affect neuropathogenesisrdquo Virus Research vol111 no 2 pp 161ndash174 2005
[105] A V Atrasheuskaya T M Fredeking and G M IgnatyevldquoChanges in immune parameters and their correction in humancases of tick-borne encephalitisrdquo Clinical and ExperimentalImmunology vol 131 no 1 pp 148ndash154 2003
[106] K L Mansfield N Johnson L P Phipps J R Stephenson AR Fooks and T Solomon ldquoTick-borne encephalitis virusmdashareview of an emerging zoonosisrdquo Journal of General Virologyvol 90 no 8 pp 1781ndash1794 2009
[107] DWVaughn andCHHoke Jr ldquoThe epidemiology of Japaneseencephalitis prospects for preventionrdquo Epidemiologic Reviewsvol 14 pp 197ndash221 1992
[108] T Solomon H Ni D W C Beasley M Ekkelenkamp M JCardosa and A D T Barrett ldquoOrigin and evolution of Japaneseencephalitis virus in Southeast Asiardquo Journal of Virology vol 77no 5 pp 3091ndash3098 2003
[109] M R Wilson ldquoEmerging viral infectionsrdquo Current Opinion inNeurology vol 26 no 3 pp 301ndash306 2013
[110] J N Hanna S A Ritchie D A Phillips et al ldquoAn outbreakof Japanese encephalitis in the Torres Strait Australia 1995rdquoMedical Journal of Australia vol 165 no 5 pp 256ndash260 1996
[111] J N Hanna S A Ritchie D A Phillips et al ldquoJapanese ence-phalitis in north Queensland Australia 1998rdquo Medical Journalof Australia vol 170 no 11 pp 533ndash536 1999
[112] SGeorgeMGourie-Devi and J A Rao ldquoIsolation ofWestNilevirus from the brains of children who had died of encephalitisrdquoBulletin of theWorldHealthOrganization vol 62 no 6 pp 879ndash882 1984
[113] A K Gupta A A Koshy and V J Lad ldquoEnhanced protec-tion of mice against Japanese encephalitis virus infection bycombinations ofmonoclonal antibodies to glycoprotein ErdquoActaVirologica vol 55 no 2 pp 165ndash168 2011
[114] T H Lee B-H Song S-I Yun et al ldquoA cross-protective mAbrecognizes a novel epitope within the flavivirus NS1 proteinrdquoJournal of General Virology vol 93 no 1 pp 20ndash26 2012
[115] S C Weaver ldquoHost range amplification and arboviral diseaseemergencerdquo Archives of Virology Supplementum no 19 pp 33ndash44 2005
[116] M Solignat B Gay S Higgs L Briant and C Devaux ldquoRepli-cation cycle of chikungunya a re-emerging arbovirusrdquoVirologyvol 393 no 2 pp 183ndash197 2009
[117] O Schwartz and M L Albert ldquoBiology and pathogenesis ofchikungunya virusrdquo Nature Reviews Microbiology vol 8 no 7pp 491ndash500 2010
[118] W Akahata Z-Y Yang H Andersen et al ldquoA virus-like particlevaccine for epidemic Chikungunya virus protects nonhumanprimates against infectionrdquo Nature Medicine vol 16 no 3 pp334ndash338 2010
[119] M Kielian and A Helenius ldquopH-Iinduced alterations in thefusogenic spike protein of Semliki Forest virusrdquo Journal of CellBiology vol 101 no 6 pp 2284ndash2291 1985
[120] J White and A Helenius ldquopH-Dependent fusion between theSemliki Forest virus membrane and liposomesrdquo Proceedings ofthe National Academy of Sciences of the United States of Americavol 77 no 6 pp 3273ndash3277 1980
[121] J Dubuisson and C M Rice ldquoSindbis virus attachment isola-tion and characterization of mutants with impaired binding tovertebrate cellsrdquo Journal of Virology vol 67 no 6 pp 3363ndash33741993
[122] C Y Lee Y-W Kam J Fric et al ldquoChikungunya virus neutral-ization antigens and direct cell-to-cell transmission are revealedby human antibody-escapemutantsrdquo PLoS Pathogens vol 7 no12 Article ID e1002390 2011
[123] J E Voss M-C Vaney S Duquerroy et al ldquoGlycoproteinorganization of Chikungunya virus particles revealed by X-raycrystallographyrdquo Nature vol 468 no 7324 pp 709ndash712 2010
BioMed Research International 21
[124] L Li J Jose Y Xiang R J Kuhn and M G Rossmann ldquoStruc-tural changes of envelope proteins during alphavirus fusionrdquoNature vol 468 no 7324 pp 705ndash708 2010
[125] J Fric S Bertin-Maghit C I Wang A Nardin and L WarterldquoUse of human monoclonal antibodies to treat Chikungunyavirus infectionrdquo Journal of Infectious Diseases vol 207 no 2 pp319ndash322 2013
[126] P V Aguilar J G Estrada-Franco R Navarro-Lopez C FerroA D Haddow and S C Weaver ldquoEndemic Venezuelanequine encephalitis in the Americas hidden under the dengueumbrellardquo Future Virology vol 6 no 6 pp 721ndash740 2011
[127] S C Weaver R Winegar I D Manger and N L ForresterldquoAlphaviruses population genetics and determinants of emer-gencerdquo Antiviral Research vol 94 no 3 pp 242ndash257 2012
[128] S Paessler and S C Weaver ldquoVaccines for Venezuelan equineencephalitisrdquo Vaccine vol 27 no 4 pp D80ndashD85 2009
[129] R J Phillpotts ldquoVenezuelan equine encephalitis virus complex-specific monoclonal antibody provides broad protection inmurine models against airborne challenge with viruses fromserogroups I II and IIIrdquoVirus Research vol 120 no 1-2 pp 107ndash112 2006
[130] R J Phillpotts L D Jones and S C Howard ldquoMonoclonalantibody protectsmice against infection anddiseasewhen giveneither before or up to 24 h after airborne challenge with virulentVenezuelan equine encephalitis virusrdquoVaccine vol 20 no 11-12pp 1497ndash1504 2002
[131] J H Mathews J T Roehrig and D W Trent ldquoRole of com-plement and the Fc portion of immunoglobulin G in immunityto Venezuelan equine encephalomyelitis virus infection withglycoprotein-specific monoclonal antibodiesrdquo Journal of Virol-ogy vol 55 no 3 pp 594ndash600 1985
[132] L M OrsquoBrien C D Underwood-Fowler S A Goodchild AL Phelps and R J Phillpotts ldquoDevelopment of a novel mon-oclonal antibody with reactivity to a wide range of Venezuelanequine encephalitis virus strainsrdquoVirology Journal vol 6 article206 2009
[133] L M OrsquoBrien S A Goodchild R J Phillpotts and S D Per-kins ldquoA humanised murine monoclonal antibody protectsmice from Venezuelan equine encephalitis virus Evergladesvirus and Mucambo virus when administered up to 48h afterairborne challengerdquo Virology vol 426 no 2 pp 100ndash105 2012
[134] N Clementi N Mancini L Solforosi M Castelli M Clementiand R Burioni ldquoPhage display-based strategies for cloning andoptimization of monoclonal antibodies directed against humanpathogensrdquo International Journal of Molecular Sciences vol 13no 7 pp 8273ndash8292 2012
[135] A R Hunt R A Bowen S Frederickson T Maruyama JT Roehrig and C D Blair ldquoTreatment of mice with humanmonoclonal antibody 24h after lethal aerosol challenge withvirulent Venezuelan equine encephalitis virus prevents diseasebut not infectionrdquo Virology vol 414 no 2 pp 146ndash152 2011
[136] W-G Hu A L Phelps S Jager et al ldquoA recombinant human-ized monoclonal antibody completely protects mice againstlethal challenge with Venezuelan equine encephalitis virusrdquoVaccine vol 28 no 34 pp 5558ndash5564 2010
[137] T Rulker L Voss P Thullier et al ldquoIsolation and characterisa-tion of a human-like antibody fragment (scFv) that inactivatesVEEV in vitro and in vivordquo PLoS ONE vol 7 no 5 Article IDe37242 2012
[138] C T Walter and J N Barr ldquoRecent advances in the molecularand cellular biology of bunyavirusesrdquo Journal of General Virol-ogy vol 92 no 11 pp 2467ndash2484 2011
[139] C A Whitehouse ldquoCrimean-Congo hemorrhagic feverrdquo Anti-viral Research vol 64 no 3 pp 145ndash160 2004
[140] M Mertens K Schmidt A Ozkul and M H Groschup ldquoTheimpact of Crimean-Congo hemorrhagic fever virus on publichealthrdquo Antiviral Research vol 98 no 2 pp 248ndash260 2013
[141] A A Ahmed J M McFalls C Hoffmann et al ldquoPresence ofbroadly reactive and group-specific neutralizing epitopes onnewly described isolates of Crimean-Congo hemorrhagic fevervirusrdquo Journal of General Virology vol 86 no 12 pp 3327ndash33362005
[142] R Hewson A Gmyl L Gmyl et al ldquoEvidence of segment reas-sortment in Crimean-Congo haemorrhagic fever virusrdquo Journalof General Virology vol 85 no 10 pp 3059ndash3070 2004
[143] N K Blackburn T G Besselaar A J Shepherd and R Swane-poel ldquoPreparation and use of monoclonal antibodies for iden-tifying Crimean-Congo hemorrhagic fever virusrdquo AmericanJournal of Tropical Medicine and Hygiene vol 37 no 2 pp 392ndash397 1987
[144] T Ikegami ldquoMolecular biology and genetic diversity of RiftValley fever virusrdquoAntiviral Research vol 95 no 3 pp 293ndash3102012
[145] KKeegan andM S Collett ldquoUse of bacterial expression cloningto define the amino acid sequences of antigenic determinantson the G2 glycoprotein of Rift Valley fever virusrdquo Journal ofVirology vol 58 no 2 pp 263ndash270 1986
[146] T G Besselaar and N K Blackburn ldquoTopological mapping ofantigenic sites on the Rift Valley fever virus envelope glycopro-teins using monoclonal antibodiesrdquo Archives of Virology vol121 no 1ndash4 pp 111ndash124 1991
[147] L G Presta ldquoEngineering of therapeutic antibodies to mini-mize immunogenicity and optimize functionrdquo Advanced DrugDelivery Reviews vol 58 no 5-6 pp 640ndash656 2006
Submit your manuscripts athttpwwwhindawicom
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anatomy Research International
PeptidesInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporation httpwwwhindawicom
International Journal of
Volume 2014
Zoology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Molecular Biology International
GenomicsInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioinformaticsAdvances in
Marine BiologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Signal TransductionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Evolutionary BiologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Biochemistry Research International
ArchaeaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Genetics Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in
Virolog y
Hindawi Publishing Corporationhttpwwwhindawicom
Nucleic AcidsJournal of
Volume 2014
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Enzyme Research
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Microbiology

BioMed Research International 19
[61] S Daffis R E Kontermann J Korimbocus H Zeller H-DKlenk and J Ter Meulen ldquoAntibody responses against wild-type yellow fever virus and the 17D vaccine strain charac-terization with human monoclonal antibody fragments andneutralization escape variantsrdquoVirology vol 337 no 2 pp 262ndash272 2005
[62] L N Levanov L E Matveev E P Goncharova et al ldquoChimericantibodies against tick-borne encephalitis virusrdquo Vaccine vol28 no 32 pp 5265ndash5271 2010
[63] A R Hunt S Frederickson C Hinkel K S Bowdish and J TRoehrig ldquoA humanized murine monoclonal antibody protectsmice either before or after challenge with virulent Venezuelanequine encephalomyelitis virusrdquo Journal of General Virologyvol 87 no 9 pp 2467ndash2476 2006
[64] J M da Silva Voorham I A Rodenhuis-Zybert N V A Nunezet al ldquoAntibodies against the envelope glycoprotein promoteinfectivity of immature dengue virus serotype 2rdquo PLoS ONEvol 7 no 3 Article ID e29957 2012
[65] T C Pierson Q Xu S Nelson et al ldquoThe stoichiometry ofantibody-mediated neutralization and enhancement of westnile virus infectionrdquoCell Host andMicrobe vol 1 no 2 pp 135ndash145 2007
[66] G Sautto N Mancini R A Diotti L Solforosi M Clementiand R Burioni ldquoAnti-hepatitis C virus E2 (HCVE2) glyco-proteinmonoclonal antibodies andneutralization interferencerdquoAntiviral Research vol 96 no 1 pp 82ndash89 2012
[67] R Burioni Y Matsuura N Mancini et al ldquoDiverging effectsof human recombinant anti-hepatitis C virus (HCV) antibodyfragments derived from a single patient on the infectivityof a vesicular stomatitis virusHCV pseudotyperdquo Journal ofVirology vol 76 no 22 pp 11775ndash11779 2002
[68] R J Phillpotts J R Stephenson and J S Porterfield ldquoAntibody-dependent enhancement of tick-borne encephalitus virus infec-tivityrdquo Journal of General Virology vol 66 no 8 pp 1831ndash18371985
[69] K A Dowd and T C Pierson ldquoAntibody-mediated neutraliza-tion of flaviviruses a reductionist viewrdquo Virology vol 411 no 2pp 306ndash315 2011
[70] A K I Falconar ldquoMonoclonal antibodies that bind to commonepitopes on the dengue virus type 2 nonstructural-1 andenvelope glycoproteins display weak neutralizing activity anddifferentiated responses to virulent strains implications forpathogenesis and vaccinesrdquo Clinical and Vaccine Immunologyvol 15 no 3 pp 549ndash561 2008
[71] K Meyer A Banerjee S E Frey R B Belshe and R Ray ldquoAweak neutralizing antibody response to hepatitis C virus enve-lope glycoprotein enhances virus infectionrdquo PLoS ONE vol 6no 8 Article ID e23699 2011
[72] R Burioni R AWilliamson P P Sanna F E Bloom and D RBurton ldquoRecombinant human Fab to glycoprotein D neutral-izes infectivity and prevents cell-to-cell transmission of herpessimplex viruses 1 and 2 in vitrordquo Proceedings of the NationalAcademy of Sciences of the United States of America vol 91 no1 pp 355ndash359 1994
[73] M S Rolph A Zaid N E Rulli and S Mahalingam ldquoDown-regulation of interferon-120573 in antibody-dependent enhancementof dengue viral infections of human macrophages is dependenton interleukin-6rdquo Journal of Infectious Diseases vol 204 no 3pp 489ndash491 2011
[74] S S Whitehead J E Blaney A P Durbin and B R MurphyldquoProspects for a dengue virus vaccinerdquoNature ReviewsMicrobi-ology vol 5 no 7 pp 518ndash528 2007
[75] S-M Lok V Kostyuchenko G ENybakken et al ldquoBinding of aneutralizing antibody to dengue virus alters the arrangement ofsurface glycoproteinsrdquoNature Structural andMolecular Biologyvol 15 no 3 pp 312ndash317 2008
[76] A P Goncalvez R Men C Wernly R H Purcell and C-J Lai ldquoChimpanzee Fab fragments and a derived humanizedimmunogiobulin G1 antibody that efficiently cross-neutralizedengue type 1 and type 2 virusesrdquo Journal of Virology vol 78no 23 pp 12910ndash12918 2004
[77] WM P BWahala A A Kraus L B HaymoreM A Accavitti-Loper and A M de Silva ldquoDengue virus neutralization byhuman immune sera role of envelope protein domain III-reactive antibodyrdquo Virology vol 392 no 1 pp 103ndash113 2009
[78] W D Crill H R Hughes M J Delorey and G-J J ChangldquoHumoral immune responses of dengue fever patients usingepitope-specific serotype-2 virus-like particle antigensrdquo PLoSONE vol 4 no 4 Article ID e4991 2009
[79] C Setthapramote T Sasaki O Puiprom et al ldquoHuman mono-clonal antibodies to neutralize all dengue virus serotypes usinglymphocytes from patients at acute phase of the secondaryinfectionrdquo Biochemical and Biophysical Research Communica-tions vol 423 no 4 pp 867ndash872 2012
[80] M Beltramello K L Williams C P Simmons et al ldquoThehuman immune response to dengue virus is dominated byhighly cross-reactive antibodies endowed with neutralizing andenhancing activityrdquo Cell Host andMicrobe vol 8 no 3 pp 271ndash283 2010
[81] C M Midgley M Bajwa-Joseph S Vasanawathana et al ldquoAnin-depth analysis of original antigenic sin in dengue virusinfectionrdquo Journal of Virology vol 85 no 22 pp 410ndash421 2011
[82] B Kaufmann M R Vogt J Goudsmit et al ldquoNeutralizationof West Nile virus by cross-linking of its surface proteins withFab fragments of the human monoclonal antibody CR4354rdquoProceedings of the National Academy of Sciences of the UnitedStates of America vol 107 no 44 pp 18950ndash18955 2010
[83] B Spurrier J M Sampson M Totrov et al ldquoStructural analysisof human and macaque mAbs 2909 and 25B implicationsfor the configuration of the quaternary neutralizing epitope ofHIV-1 gp120rdquo Structure vol 19 no 5 pp 691ndash699 2011
[84] J J SchlesingerMWBrandriss andT PMonath ldquoMonoclonalantibodies distinguish between wild and vaccine strains ofyellow fever virus by neutralization hemagglutination inhibi-tion and immune precipitation of the virus envelope proteinrdquoVirology vol 125 no 1 pp 8ndash17 1983
[85] M W Brandriss J J Schlesinger E E Walsh and M BrisellildquoLethal 17D yellow fever encephalitis in mice I Passive protec-tion by monoclonal antibodies to the envelope proteins of 17Dyellow fever and dengue 2 virusesrdquo Journal of General Virologyvol 67 no 2 pp 229ndash234 1986
[86] H Cho and M S Diamond ldquoImmune responses to West Nilevirus infection in the central nervous systemrdquo Viruses vol 4no 12 pp 3812ndash3830 2012
[87] D W Beasley A D Barrett and R B Tesh ldquoResurgence ofWest Nile neurologic disease in the United States in 2012 whathappened What needs to be donerdquo Antiviral Research vol 99no 1 pp 1ndash5 2013
[88] M Throsby J Ter Meulen C Geuijen J Goudsmit and J DeKruif ldquoMapping and analysis of West Nile virus-specific mon-oclonal antibodies prospects for vaccine developmentrdquo ExpertReview of Vaccines vol 6 no 2 pp 183ndash191 2007
[89] M S Diamond T G Roberts D Edgil B Lu J Ernst and EHarris ldquoModulation of dengue virus infection in human cells
20 BioMed Research International
by alpha beta and gamma interferonsrdquo Journal of Virology vol74 no 11 pp 4957ndash4966 2000
[90] Z Shimoni M J Niven S Pitlick and S Bulvik ldquoTreatmentof West Nile virus encephalitis with intravenous immunoglob-ulinrdquo Emerging Infectious Diseases vol 7 no 4 article 759 2001
[91] AHamdan PGreen EMendelsonM R Kramer S Pitlik andM Weinberger ldquoPossible benefit of intravenous immunoglob-ulin therapy in a lung transplant recipient with West Nile virusencephalitisrdquoTransplant InfectiousDisease vol 4 no 3 pp 160ndash162 2002
[92] M Haley A S Retter D Fowler J Gea-Banacloche and NP OrsquoGrady ldquoThe role for intravenous immunoglobulin in thetreatment of West Nile virus encephalitisrdquo Clinical InfectiousDiseases vol 37 no 6 pp e88ndashe90 2003
[93] K M Chung G E Nybakken B S Thompson et al ldquoAnti-bodies against West Nile virus nonstructural protein NS1prevent lethal infection through Fc 120574 receptor-dependent and-independent mechanismsrdquo Journal of Virology vol 80 no 3pp 1340ndash1351 2006
[94] E A Henchal L S Henchal and J J Schlesinger ldquoSynergisticinteractions of anti-NS1 monoclonal antibodies protect pas-sively immunized mice from lethal challenge with dengue 2virusrdquo Journal of General Virology vol 69 no 8 pp 2101ndash21071988
[95] J T Roehrig ldquoAntigenic structure of flavivirus proteinsrdquoAdvances in Virus Research vol 59 pp 141ndash175 2003
[96] M D Sanchez T C Pierson D McAllister et al ldquoCharacteri-zation of neutralizing antibodies to West Nile virusrdquo Virologyvol 336 no 1 pp 70ndash82 2005
[97] J H Beigel J L Nordstrom S R Pillemer et al ldquoSafety andpharmacokinetics of single intravenous dose of MGAWN1 anovel monoclonal antibody to West Nile virusrdquo AntimicrobialAgents and Chemotherapy vol 54 no 6 pp 2431ndash2436 2010
[98] C A Smeraski V Siddharthan and J D Morrey ldquoTreatmentof spatial memory impairment in hamsters infected with WestNile virus using a humanizedmonoclonal antibodyMGAWN1rdquoAntiviral Research vol 91 no 1 pp 43ndash49 2011
[99] D Lelli AMoreno E Brocchi et al ldquoWestNile virus character-ization and diagnostic applications of monoclonal antibodiesrdquoVirology Journal vol 9 article 81 2012
[100] T S Gritsun V A Lashkevich and E A Gould ldquoTick-borneencephalitisrdquo Antiviral Research vol 57 no 1-2 pp 129ndash1462003
[101] D Ruzek T S Gritsun N L Forrester et al ldquoMutations inthe NS2B and NS3 genes affect mouse neuroinvasiveness of aWestern European field strain of tick-borne encephalitis virusrdquoVirology vol 374 no 2 pp 249ndash255 2008
[102] M Ecker S L Allison T Meixner and F X Heinz ldquoSequenceanalysis and genetic classification of tick-borne encephalitisviruses from Europe and Asiardquo Journal of General Virology vol80 no 1 pp 179ndash185 1999
[103] H Holzmann M S Vorobyova I P Ladyzhenskaya et alldquoMolecular epidemiology of tick-borne encephalitis viruscross-protection between European and Far Eastern subtypesrdquoVaccine vol 10 no 5 pp 345ndash349 1992
[104] C W Mandl ldquoSteps of the tick-borne encephalitis virus repli-cation cycle that affect neuropathogenesisrdquo Virus Research vol111 no 2 pp 161ndash174 2005
[105] A V Atrasheuskaya T M Fredeking and G M IgnatyevldquoChanges in immune parameters and their correction in humancases of tick-borne encephalitisrdquo Clinical and ExperimentalImmunology vol 131 no 1 pp 148ndash154 2003
[106] K L Mansfield N Johnson L P Phipps J R Stephenson AR Fooks and T Solomon ldquoTick-borne encephalitis virusmdashareview of an emerging zoonosisrdquo Journal of General Virologyvol 90 no 8 pp 1781ndash1794 2009
[107] DWVaughn andCHHoke Jr ldquoThe epidemiology of Japaneseencephalitis prospects for preventionrdquo Epidemiologic Reviewsvol 14 pp 197ndash221 1992
[108] T Solomon H Ni D W C Beasley M Ekkelenkamp M JCardosa and A D T Barrett ldquoOrigin and evolution of Japaneseencephalitis virus in Southeast Asiardquo Journal of Virology vol 77no 5 pp 3091ndash3098 2003
[109] M R Wilson ldquoEmerging viral infectionsrdquo Current Opinion inNeurology vol 26 no 3 pp 301ndash306 2013
[110] J N Hanna S A Ritchie D A Phillips et al ldquoAn outbreakof Japanese encephalitis in the Torres Strait Australia 1995rdquoMedical Journal of Australia vol 165 no 5 pp 256ndash260 1996
[111] J N Hanna S A Ritchie D A Phillips et al ldquoJapanese ence-phalitis in north Queensland Australia 1998rdquo Medical Journalof Australia vol 170 no 11 pp 533ndash536 1999
[112] SGeorgeMGourie-Devi and J A Rao ldquoIsolation ofWestNilevirus from the brains of children who had died of encephalitisrdquoBulletin of theWorldHealthOrganization vol 62 no 6 pp 879ndash882 1984
[113] A K Gupta A A Koshy and V J Lad ldquoEnhanced protec-tion of mice against Japanese encephalitis virus infection bycombinations ofmonoclonal antibodies to glycoprotein ErdquoActaVirologica vol 55 no 2 pp 165ndash168 2011
[114] T H Lee B-H Song S-I Yun et al ldquoA cross-protective mAbrecognizes a novel epitope within the flavivirus NS1 proteinrdquoJournal of General Virology vol 93 no 1 pp 20ndash26 2012
[115] S C Weaver ldquoHost range amplification and arboviral diseaseemergencerdquo Archives of Virology Supplementum no 19 pp 33ndash44 2005
[116] M Solignat B Gay S Higgs L Briant and C Devaux ldquoRepli-cation cycle of chikungunya a re-emerging arbovirusrdquoVirologyvol 393 no 2 pp 183ndash197 2009
[117] O Schwartz and M L Albert ldquoBiology and pathogenesis ofchikungunya virusrdquo Nature Reviews Microbiology vol 8 no 7pp 491ndash500 2010
[118] W Akahata Z-Y Yang H Andersen et al ldquoA virus-like particlevaccine for epidemic Chikungunya virus protects nonhumanprimates against infectionrdquo Nature Medicine vol 16 no 3 pp334ndash338 2010
[119] M Kielian and A Helenius ldquopH-Iinduced alterations in thefusogenic spike protein of Semliki Forest virusrdquo Journal of CellBiology vol 101 no 6 pp 2284ndash2291 1985
[120] J White and A Helenius ldquopH-Dependent fusion between theSemliki Forest virus membrane and liposomesrdquo Proceedings ofthe National Academy of Sciences of the United States of Americavol 77 no 6 pp 3273ndash3277 1980
[121] J Dubuisson and C M Rice ldquoSindbis virus attachment isola-tion and characterization of mutants with impaired binding tovertebrate cellsrdquo Journal of Virology vol 67 no 6 pp 3363ndash33741993
[122] C Y Lee Y-W Kam J Fric et al ldquoChikungunya virus neutral-ization antigens and direct cell-to-cell transmission are revealedby human antibody-escapemutantsrdquo PLoS Pathogens vol 7 no12 Article ID e1002390 2011
[123] J E Voss M-C Vaney S Duquerroy et al ldquoGlycoproteinorganization of Chikungunya virus particles revealed by X-raycrystallographyrdquo Nature vol 468 no 7324 pp 709ndash712 2010
BioMed Research International 21
[124] L Li J Jose Y Xiang R J Kuhn and M G Rossmann ldquoStruc-tural changes of envelope proteins during alphavirus fusionrdquoNature vol 468 no 7324 pp 705ndash708 2010
[125] J Fric S Bertin-Maghit C I Wang A Nardin and L WarterldquoUse of human monoclonal antibodies to treat Chikungunyavirus infectionrdquo Journal of Infectious Diseases vol 207 no 2 pp319ndash322 2013
[126] P V Aguilar J G Estrada-Franco R Navarro-Lopez C FerroA D Haddow and S C Weaver ldquoEndemic Venezuelanequine encephalitis in the Americas hidden under the dengueumbrellardquo Future Virology vol 6 no 6 pp 721ndash740 2011
[127] S C Weaver R Winegar I D Manger and N L ForresterldquoAlphaviruses population genetics and determinants of emer-gencerdquo Antiviral Research vol 94 no 3 pp 242ndash257 2012
[128] S Paessler and S C Weaver ldquoVaccines for Venezuelan equineencephalitisrdquo Vaccine vol 27 no 4 pp D80ndashD85 2009
[129] R J Phillpotts ldquoVenezuelan equine encephalitis virus complex-specific monoclonal antibody provides broad protection inmurine models against airborne challenge with viruses fromserogroups I II and IIIrdquoVirus Research vol 120 no 1-2 pp 107ndash112 2006
[130] R J Phillpotts L D Jones and S C Howard ldquoMonoclonalantibody protectsmice against infection anddiseasewhen giveneither before or up to 24 h after airborne challenge with virulentVenezuelan equine encephalitis virusrdquoVaccine vol 20 no 11-12pp 1497ndash1504 2002
[131] J H Mathews J T Roehrig and D W Trent ldquoRole of com-plement and the Fc portion of immunoglobulin G in immunityto Venezuelan equine encephalomyelitis virus infection withglycoprotein-specific monoclonal antibodiesrdquo Journal of Virol-ogy vol 55 no 3 pp 594ndash600 1985
[132] L M OrsquoBrien C D Underwood-Fowler S A Goodchild AL Phelps and R J Phillpotts ldquoDevelopment of a novel mon-oclonal antibody with reactivity to a wide range of Venezuelanequine encephalitis virus strainsrdquoVirology Journal vol 6 article206 2009
[133] L M OrsquoBrien S A Goodchild R J Phillpotts and S D Per-kins ldquoA humanised murine monoclonal antibody protectsmice from Venezuelan equine encephalitis virus Evergladesvirus and Mucambo virus when administered up to 48h afterairborne challengerdquo Virology vol 426 no 2 pp 100ndash105 2012
[134] N Clementi N Mancini L Solforosi M Castelli M Clementiand R Burioni ldquoPhage display-based strategies for cloning andoptimization of monoclonal antibodies directed against humanpathogensrdquo International Journal of Molecular Sciences vol 13no 7 pp 8273ndash8292 2012
[135] A R Hunt R A Bowen S Frederickson T Maruyama JT Roehrig and C D Blair ldquoTreatment of mice with humanmonoclonal antibody 24h after lethal aerosol challenge withvirulent Venezuelan equine encephalitis virus prevents diseasebut not infectionrdquo Virology vol 414 no 2 pp 146ndash152 2011
[136] W-G Hu A L Phelps S Jager et al ldquoA recombinant human-ized monoclonal antibody completely protects mice againstlethal challenge with Venezuelan equine encephalitis virusrdquoVaccine vol 28 no 34 pp 5558ndash5564 2010
[137] T Rulker L Voss P Thullier et al ldquoIsolation and characterisa-tion of a human-like antibody fragment (scFv) that inactivatesVEEV in vitro and in vivordquo PLoS ONE vol 7 no 5 Article IDe37242 2012
[138] C T Walter and J N Barr ldquoRecent advances in the molecularand cellular biology of bunyavirusesrdquo Journal of General Virol-ogy vol 92 no 11 pp 2467ndash2484 2011
[139] C A Whitehouse ldquoCrimean-Congo hemorrhagic feverrdquo Anti-viral Research vol 64 no 3 pp 145ndash160 2004
[140] M Mertens K Schmidt A Ozkul and M H Groschup ldquoTheimpact of Crimean-Congo hemorrhagic fever virus on publichealthrdquo Antiviral Research vol 98 no 2 pp 248ndash260 2013
[141] A A Ahmed J M McFalls C Hoffmann et al ldquoPresence ofbroadly reactive and group-specific neutralizing epitopes onnewly described isolates of Crimean-Congo hemorrhagic fevervirusrdquo Journal of General Virology vol 86 no 12 pp 3327ndash33362005
[142] R Hewson A Gmyl L Gmyl et al ldquoEvidence of segment reas-sortment in Crimean-Congo haemorrhagic fever virusrdquo Journalof General Virology vol 85 no 10 pp 3059ndash3070 2004
[143] N K Blackburn T G Besselaar A J Shepherd and R Swane-poel ldquoPreparation and use of monoclonal antibodies for iden-tifying Crimean-Congo hemorrhagic fever virusrdquo AmericanJournal of Tropical Medicine and Hygiene vol 37 no 2 pp 392ndash397 1987
[144] T Ikegami ldquoMolecular biology and genetic diversity of RiftValley fever virusrdquoAntiviral Research vol 95 no 3 pp 293ndash3102012
[145] KKeegan andM S Collett ldquoUse of bacterial expression cloningto define the amino acid sequences of antigenic determinantson the G2 glycoprotein of Rift Valley fever virusrdquo Journal ofVirology vol 58 no 2 pp 263ndash270 1986
[146] T G Besselaar and N K Blackburn ldquoTopological mapping ofantigenic sites on the Rift Valley fever virus envelope glycopro-teins using monoclonal antibodiesrdquo Archives of Virology vol121 no 1ndash4 pp 111ndash124 1991
[147] L G Presta ldquoEngineering of therapeutic antibodies to mini-mize immunogenicity and optimize functionrdquo Advanced DrugDelivery Reviews vol 58 no 5-6 pp 640ndash656 2006
Submit your manuscripts athttpwwwhindawicom
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anatomy Research International
PeptidesInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporation httpwwwhindawicom
International Journal of
Volume 2014
Zoology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Molecular Biology International
GenomicsInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioinformaticsAdvances in
Marine BiologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Signal TransductionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Evolutionary BiologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Biochemistry Research International
ArchaeaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Genetics Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in
Virolog y
Hindawi Publishing Corporationhttpwwwhindawicom
Nucleic AcidsJournal of
Volume 2014
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Enzyme Research
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Microbiology

20 BioMed Research International
by alpha beta and gamma interferonsrdquo Journal of Virology vol74 no 11 pp 4957ndash4966 2000
[90] Z Shimoni M J Niven S Pitlick and S Bulvik ldquoTreatmentof West Nile virus encephalitis with intravenous immunoglob-ulinrdquo Emerging Infectious Diseases vol 7 no 4 article 759 2001
[91] AHamdan PGreen EMendelsonM R Kramer S Pitlik andM Weinberger ldquoPossible benefit of intravenous immunoglob-ulin therapy in a lung transplant recipient with West Nile virusencephalitisrdquoTransplant InfectiousDisease vol 4 no 3 pp 160ndash162 2002
[92] M Haley A S Retter D Fowler J Gea-Banacloche and NP OrsquoGrady ldquoThe role for intravenous immunoglobulin in thetreatment of West Nile virus encephalitisrdquo Clinical InfectiousDiseases vol 37 no 6 pp e88ndashe90 2003
[93] K M Chung G E Nybakken B S Thompson et al ldquoAnti-bodies against West Nile virus nonstructural protein NS1prevent lethal infection through Fc 120574 receptor-dependent and-independent mechanismsrdquo Journal of Virology vol 80 no 3pp 1340ndash1351 2006
[94] E A Henchal L S Henchal and J J Schlesinger ldquoSynergisticinteractions of anti-NS1 monoclonal antibodies protect pas-sively immunized mice from lethal challenge with dengue 2virusrdquo Journal of General Virology vol 69 no 8 pp 2101ndash21071988
[95] J T Roehrig ldquoAntigenic structure of flavivirus proteinsrdquoAdvances in Virus Research vol 59 pp 141ndash175 2003
[96] M D Sanchez T C Pierson D McAllister et al ldquoCharacteri-zation of neutralizing antibodies to West Nile virusrdquo Virologyvol 336 no 1 pp 70ndash82 2005
[97] J H Beigel J L Nordstrom S R Pillemer et al ldquoSafety andpharmacokinetics of single intravenous dose of MGAWN1 anovel monoclonal antibody to West Nile virusrdquo AntimicrobialAgents and Chemotherapy vol 54 no 6 pp 2431ndash2436 2010
[98] C A Smeraski V Siddharthan and J D Morrey ldquoTreatmentof spatial memory impairment in hamsters infected with WestNile virus using a humanizedmonoclonal antibodyMGAWN1rdquoAntiviral Research vol 91 no 1 pp 43ndash49 2011
[99] D Lelli AMoreno E Brocchi et al ldquoWestNile virus character-ization and diagnostic applications of monoclonal antibodiesrdquoVirology Journal vol 9 article 81 2012
[100] T S Gritsun V A Lashkevich and E A Gould ldquoTick-borneencephalitisrdquo Antiviral Research vol 57 no 1-2 pp 129ndash1462003
[101] D Ruzek T S Gritsun N L Forrester et al ldquoMutations inthe NS2B and NS3 genes affect mouse neuroinvasiveness of aWestern European field strain of tick-borne encephalitis virusrdquoVirology vol 374 no 2 pp 249ndash255 2008
[102] M Ecker S L Allison T Meixner and F X Heinz ldquoSequenceanalysis and genetic classification of tick-borne encephalitisviruses from Europe and Asiardquo Journal of General Virology vol80 no 1 pp 179ndash185 1999
[103] H Holzmann M S Vorobyova I P Ladyzhenskaya et alldquoMolecular epidemiology of tick-borne encephalitis viruscross-protection between European and Far Eastern subtypesrdquoVaccine vol 10 no 5 pp 345ndash349 1992
[104] C W Mandl ldquoSteps of the tick-borne encephalitis virus repli-cation cycle that affect neuropathogenesisrdquo Virus Research vol111 no 2 pp 161ndash174 2005
[105] A V Atrasheuskaya T M Fredeking and G M IgnatyevldquoChanges in immune parameters and their correction in humancases of tick-borne encephalitisrdquo Clinical and ExperimentalImmunology vol 131 no 1 pp 148ndash154 2003
[106] K L Mansfield N Johnson L P Phipps J R Stephenson AR Fooks and T Solomon ldquoTick-borne encephalitis virusmdashareview of an emerging zoonosisrdquo Journal of General Virologyvol 90 no 8 pp 1781ndash1794 2009
[107] DWVaughn andCHHoke Jr ldquoThe epidemiology of Japaneseencephalitis prospects for preventionrdquo Epidemiologic Reviewsvol 14 pp 197ndash221 1992
[108] T Solomon H Ni D W C Beasley M Ekkelenkamp M JCardosa and A D T Barrett ldquoOrigin and evolution of Japaneseencephalitis virus in Southeast Asiardquo Journal of Virology vol 77no 5 pp 3091ndash3098 2003
[109] M R Wilson ldquoEmerging viral infectionsrdquo Current Opinion inNeurology vol 26 no 3 pp 301ndash306 2013
[110] J N Hanna S A Ritchie D A Phillips et al ldquoAn outbreakof Japanese encephalitis in the Torres Strait Australia 1995rdquoMedical Journal of Australia vol 165 no 5 pp 256ndash260 1996
[111] J N Hanna S A Ritchie D A Phillips et al ldquoJapanese ence-phalitis in north Queensland Australia 1998rdquo Medical Journalof Australia vol 170 no 11 pp 533ndash536 1999
[112] SGeorgeMGourie-Devi and J A Rao ldquoIsolation ofWestNilevirus from the brains of children who had died of encephalitisrdquoBulletin of theWorldHealthOrganization vol 62 no 6 pp 879ndash882 1984
[113] A K Gupta A A Koshy and V J Lad ldquoEnhanced protec-tion of mice against Japanese encephalitis virus infection bycombinations ofmonoclonal antibodies to glycoprotein ErdquoActaVirologica vol 55 no 2 pp 165ndash168 2011
[114] T H Lee B-H Song S-I Yun et al ldquoA cross-protective mAbrecognizes a novel epitope within the flavivirus NS1 proteinrdquoJournal of General Virology vol 93 no 1 pp 20ndash26 2012
[115] S C Weaver ldquoHost range amplification and arboviral diseaseemergencerdquo Archives of Virology Supplementum no 19 pp 33ndash44 2005
[116] M Solignat B Gay S Higgs L Briant and C Devaux ldquoRepli-cation cycle of chikungunya a re-emerging arbovirusrdquoVirologyvol 393 no 2 pp 183ndash197 2009
[117] O Schwartz and M L Albert ldquoBiology and pathogenesis ofchikungunya virusrdquo Nature Reviews Microbiology vol 8 no 7pp 491ndash500 2010
[118] W Akahata Z-Y Yang H Andersen et al ldquoA virus-like particlevaccine for epidemic Chikungunya virus protects nonhumanprimates against infectionrdquo Nature Medicine vol 16 no 3 pp334ndash338 2010
[119] M Kielian and A Helenius ldquopH-Iinduced alterations in thefusogenic spike protein of Semliki Forest virusrdquo Journal of CellBiology vol 101 no 6 pp 2284ndash2291 1985
[120] J White and A Helenius ldquopH-Dependent fusion between theSemliki Forest virus membrane and liposomesrdquo Proceedings ofthe National Academy of Sciences of the United States of Americavol 77 no 6 pp 3273ndash3277 1980
[121] J Dubuisson and C M Rice ldquoSindbis virus attachment isola-tion and characterization of mutants with impaired binding tovertebrate cellsrdquo Journal of Virology vol 67 no 6 pp 3363ndash33741993
[122] C Y Lee Y-W Kam J Fric et al ldquoChikungunya virus neutral-ization antigens and direct cell-to-cell transmission are revealedby human antibody-escapemutantsrdquo PLoS Pathogens vol 7 no12 Article ID e1002390 2011
[123] J E Voss M-C Vaney S Duquerroy et al ldquoGlycoproteinorganization of Chikungunya virus particles revealed by X-raycrystallographyrdquo Nature vol 468 no 7324 pp 709ndash712 2010
BioMed Research International 21
[124] L Li J Jose Y Xiang R J Kuhn and M G Rossmann ldquoStruc-tural changes of envelope proteins during alphavirus fusionrdquoNature vol 468 no 7324 pp 705ndash708 2010
[125] J Fric S Bertin-Maghit C I Wang A Nardin and L WarterldquoUse of human monoclonal antibodies to treat Chikungunyavirus infectionrdquo Journal of Infectious Diseases vol 207 no 2 pp319ndash322 2013
[126] P V Aguilar J G Estrada-Franco R Navarro-Lopez C FerroA D Haddow and S C Weaver ldquoEndemic Venezuelanequine encephalitis in the Americas hidden under the dengueumbrellardquo Future Virology vol 6 no 6 pp 721ndash740 2011
[127] S C Weaver R Winegar I D Manger and N L ForresterldquoAlphaviruses population genetics and determinants of emer-gencerdquo Antiviral Research vol 94 no 3 pp 242ndash257 2012
[128] S Paessler and S C Weaver ldquoVaccines for Venezuelan equineencephalitisrdquo Vaccine vol 27 no 4 pp D80ndashD85 2009
[129] R J Phillpotts ldquoVenezuelan equine encephalitis virus complex-specific monoclonal antibody provides broad protection inmurine models against airborne challenge with viruses fromserogroups I II and IIIrdquoVirus Research vol 120 no 1-2 pp 107ndash112 2006
[130] R J Phillpotts L D Jones and S C Howard ldquoMonoclonalantibody protectsmice against infection anddiseasewhen giveneither before or up to 24 h after airborne challenge with virulentVenezuelan equine encephalitis virusrdquoVaccine vol 20 no 11-12pp 1497ndash1504 2002
[131] J H Mathews J T Roehrig and D W Trent ldquoRole of com-plement and the Fc portion of immunoglobulin G in immunityto Venezuelan equine encephalomyelitis virus infection withglycoprotein-specific monoclonal antibodiesrdquo Journal of Virol-ogy vol 55 no 3 pp 594ndash600 1985
[132] L M OrsquoBrien C D Underwood-Fowler S A Goodchild AL Phelps and R J Phillpotts ldquoDevelopment of a novel mon-oclonal antibody with reactivity to a wide range of Venezuelanequine encephalitis virus strainsrdquoVirology Journal vol 6 article206 2009
[133] L M OrsquoBrien S A Goodchild R J Phillpotts and S D Per-kins ldquoA humanised murine monoclonal antibody protectsmice from Venezuelan equine encephalitis virus Evergladesvirus and Mucambo virus when administered up to 48h afterairborne challengerdquo Virology vol 426 no 2 pp 100ndash105 2012
[134] N Clementi N Mancini L Solforosi M Castelli M Clementiand R Burioni ldquoPhage display-based strategies for cloning andoptimization of monoclonal antibodies directed against humanpathogensrdquo International Journal of Molecular Sciences vol 13no 7 pp 8273ndash8292 2012
[135] A R Hunt R A Bowen S Frederickson T Maruyama JT Roehrig and C D Blair ldquoTreatment of mice with humanmonoclonal antibody 24h after lethal aerosol challenge withvirulent Venezuelan equine encephalitis virus prevents diseasebut not infectionrdquo Virology vol 414 no 2 pp 146ndash152 2011
[136] W-G Hu A L Phelps S Jager et al ldquoA recombinant human-ized monoclonal antibody completely protects mice againstlethal challenge with Venezuelan equine encephalitis virusrdquoVaccine vol 28 no 34 pp 5558ndash5564 2010
[137] T Rulker L Voss P Thullier et al ldquoIsolation and characterisa-tion of a human-like antibody fragment (scFv) that inactivatesVEEV in vitro and in vivordquo PLoS ONE vol 7 no 5 Article IDe37242 2012
[138] C T Walter and J N Barr ldquoRecent advances in the molecularand cellular biology of bunyavirusesrdquo Journal of General Virol-ogy vol 92 no 11 pp 2467ndash2484 2011
[139] C A Whitehouse ldquoCrimean-Congo hemorrhagic feverrdquo Anti-viral Research vol 64 no 3 pp 145ndash160 2004
[140] M Mertens K Schmidt A Ozkul and M H Groschup ldquoTheimpact of Crimean-Congo hemorrhagic fever virus on publichealthrdquo Antiviral Research vol 98 no 2 pp 248ndash260 2013
[141] A A Ahmed J M McFalls C Hoffmann et al ldquoPresence ofbroadly reactive and group-specific neutralizing epitopes onnewly described isolates of Crimean-Congo hemorrhagic fevervirusrdquo Journal of General Virology vol 86 no 12 pp 3327ndash33362005
[142] R Hewson A Gmyl L Gmyl et al ldquoEvidence of segment reas-sortment in Crimean-Congo haemorrhagic fever virusrdquo Journalof General Virology vol 85 no 10 pp 3059ndash3070 2004
[143] N K Blackburn T G Besselaar A J Shepherd and R Swane-poel ldquoPreparation and use of monoclonal antibodies for iden-tifying Crimean-Congo hemorrhagic fever virusrdquo AmericanJournal of Tropical Medicine and Hygiene vol 37 no 2 pp 392ndash397 1987
[144] T Ikegami ldquoMolecular biology and genetic diversity of RiftValley fever virusrdquoAntiviral Research vol 95 no 3 pp 293ndash3102012
[145] KKeegan andM S Collett ldquoUse of bacterial expression cloningto define the amino acid sequences of antigenic determinantson the G2 glycoprotein of Rift Valley fever virusrdquo Journal ofVirology vol 58 no 2 pp 263ndash270 1986
[146] T G Besselaar and N K Blackburn ldquoTopological mapping ofantigenic sites on the Rift Valley fever virus envelope glycopro-teins using monoclonal antibodiesrdquo Archives of Virology vol121 no 1ndash4 pp 111ndash124 1991
[147] L G Presta ldquoEngineering of therapeutic antibodies to mini-mize immunogenicity and optimize functionrdquo Advanced DrugDelivery Reviews vol 58 no 5-6 pp 640ndash656 2006
Submit your manuscripts athttpwwwhindawicom
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anatomy Research International
PeptidesInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporation httpwwwhindawicom
International Journal of
Volume 2014
Zoology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Molecular Biology International
GenomicsInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioinformaticsAdvances in
Marine BiologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Signal TransductionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Evolutionary BiologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Biochemistry Research International
ArchaeaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Genetics Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in
Virolog y
Hindawi Publishing Corporationhttpwwwhindawicom
Nucleic AcidsJournal of
Volume 2014
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Enzyme Research
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Microbiology

BioMed Research International 21
[124] L Li J Jose Y Xiang R J Kuhn and M G Rossmann ldquoStruc-tural changes of envelope proteins during alphavirus fusionrdquoNature vol 468 no 7324 pp 705ndash708 2010
[125] J Fric S Bertin-Maghit C I Wang A Nardin and L WarterldquoUse of human monoclonal antibodies to treat Chikungunyavirus infectionrdquo Journal of Infectious Diseases vol 207 no 2 pp319ndash322 2013
[126] P V Aguilar J G Estrada-Franco R Navarro-Lopez C FerroA D Haddow and S C Weaver ldquoEndemic Venezuelanequine encephalitis in the Americas hidden under the dengueumbrellardquo Future Virology vol 6 no 6 pp 721ndash740 2011
[127] S C Weaver R Winegar I D Manger and N L ForresterldquoAlphaviruses population genetics and determinants of emer-gencerdquo Antiviral Research vol 94 no 3 pp 242ndash257 2012
[128] S Paessler and S C Weaver ldquoVaccines for Venezuelan equineencephalitisrdquo Vaccine vol 27 no 4 pp D80ndashD85 2009
[129] R J Phillpotts ldquoVenezuelan equine encephalitis virus complex-specific monoclonal antibody provides broad protection inmurine models against airborne challenge with viruses fromserogroups I II and IIIrdquoVirus Research vol 120 no 1-2 pp 107ndash112 2006
[130] R J Phillpotts L D Jones and S C Howard ldquoMonoclonalantibody protectsmice against infection anddiseasewhen giveneither before or up to 24 h after airborne challenge with virulentVenezuelan equine encephalitis virusrdquoVaccine vol 20 no 11-12pp 1497ndash1504 2002
[131] J H Mathews J T Roehrig and D W Trent ldquoRole of com-plement and the Fc portion of immunoglobulin G in immunityto Venezuelan equine encephalomyelitis virus infection withglycoprotein-specific monoclonal antibodiesrdquo Journal of Virol-ogy vol 55 no 3 pp 594ndash600 1985
[132] L M OrsquoBrien C D Underwood-Fowler S A Goodchild AL Phelps and R J Phillpotts ldquoDevelopment of a novel mon-oclonal antibody with reactivity to a wide range of Venezuelanequine encephalitis virus strainsrdquoVirology Journal vol 6 article206 2009
[133] L M OrsquoBrien S A Goodchild R J Phillpotts and S D Per-kins ldquoA humanised murine monoclonal antibody protectsmice from Venezuelan equine encephalitis virus Evergladesvirus and Mucambo virus when administered up to 48h afterairborne challengerdquo Virology vol 426 no 2 pp 100ndash105 2012
[134] N Clementi N Mancini L Solforosi M Castelli M Clementiand R Burioni ldquoPhage display-based strategies for cloning andoptimization of monoclonal antibodies directed against humanpathogensrdquo International Journal of Molecular Sciences vol 13no 7 pp 8273ndash8292 2012
[135] A R Hunt R A Bowen S Frederickson T Maruyama JT Roehrig and C D Blair ldquoTreatment of mice with humanmonoclonal antibody 24h after lethal aerosol challenge withvirulent Venezuelan equine encephalitis virus prevents diseasebut not infectionrdquo Virology vol 414 no 2 pp 146ndash152 2011
[136] W-G Hu A L Phelps S Jager et al ldquoA recombinant human-ized monoclonal antibody completely protects mice againstlethal challenge with Venezuelan equine encephalitis virusrdquoVaccine vol 28 no 34 pp 5558ndash5564 2010
[137] T Rulker L Voss P Thullier et al ldquoIsolation and characterisa-tion of a human-like antibody fragment (scFv) that inactivatesVEEV in vitro and in vivordquo PLoS ONE vol 7 no 5 Article IDe37242 2012
[138] C T Walter and J N Barr ldquoRecent advances in the molecularand cellular biology of bunyavirusesrdquo Journal of General Virol-ogy vol 92 no 11 pp 2467ndash2484 2011
[139] C A Whitehouse ldquoCrimean-Congo hemorrhagic feverrdquo Anti-viral Research vol 64 no 3 pp 145ndash160 2004
[140] M Mertens K Schmidt A Ozkul and M H Groschup ldquoTheimpact of Crimean-Congo hemorrhagic fever virus on publichealthrdquo Antiviral Research vol 98 no 2 pp 248ndash260 2013
[141] A A Ahmed J M McFalls C Hoffmann et al ldquoPresence ofbroadly reactive and group-specific neutralizing epitopes onnewly described isolates of Crimean-Congo hemorrhagic fevervirusrdquo Journal of General Virology vol 86 no 12 pp 3327ndash33362005
[142] R Hewson A Gmyl L Gmyl et al ldquoEvidence of segment reas-sortment in Crimean-Congo haemorrhagic fever virusrdquo Journalof General Virology vol 85 no 10 pp 3059ndash3070 2004
[143] N K Blackburn T G Besselaar A J Shepherd and R Swane-poel ldquoPreparation and use of monoclonal antibodies for iden-tifying Crimean-Congo hemorrhagic fever virusrdquo AmericanJournal of Tropical Medicine and Hygiene vol 37 no 2 pp 392ndash397 1987
[144] T Ikegami ldquoMolecular biology and genetic diversity of RiftValley fever virusrdquoAntiviral Research vol 95 no 3 pp 293ndash3102012
[145] KKeegan andM S Collett ldquoUse of bacterial expression cloningto define the amino acid sequences of antigenic determinantson the G2 glycoprotein of Rift Valley fever virusrdquo Journal ofVirology vol 58 no 2 pp 263ndash270 1986
[146] T G Besselaar and N K Blackburn ldquoTopological mapping ofantigenic sites on the Rift Valley fever virus envelope glycopro-teins using monoclonal antibodiesrdquo Archives of Virology vol121 no 1ndash4 pp 111ndash124 1991
[147] L G Presta ldquoEngineering of therapeutic antibodies to mini-mize immunogenicity and optimize functionrdquo Advanced DrugDelivery Reviews vol 58 no 5-6 pp 640ndash656 2006
Submit your manuscripts athttpwwwhindawicom
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anatomy Research International
PeptidesInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporation httpwwwhindawicom
International Journal of
Volume 2014
Zoology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Molecular Biology International
GenomicsInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioinformaticsAdvances in
Marine BiologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Signal TransductionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Evolutionary BiologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Biochemistry Research International
ArchaeaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Genetics Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in
Virolog y
Hindawi Publishing Corporationhttpwwwhindawicom
Nucleic AcidsJournal of
Volume 2014
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Enzyme Research
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Microbiology

Submit your manuscripts athttpwwwhindawicom
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anatomy Research International
PeptidesInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporation httpwwwhindawicom
International Journal of
Volume 2014
Zoology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Molecular Biology International
GenomicsInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioinformaticsAdvances in
Marine BiologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Signal TransductionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Evolutionary BiologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Biochemistry Research International
ArchaeaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Genetics Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in
Virolog y
Hindawi Publishing Corporationhttpwwwhindawicom
Nucleic AcidsJournal of
Volume 2014
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Enzyme Research
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Microbiology