review of nutrient efficiency in different breeds of farm · pdf filereview of nutrient...
TRANSCRIPT

Review of nutrient efficiency in different breeds of farm livestock
30 April 2010
Project funded by DEFRA (IF0183)
Bert Tolkamp1*, Eileen Wall2, Rainer Roehe2, Jamie Newbold3, Kostas Zaralis1
¹ SAC Animal Health, Sir Stephen Watson Building, Bush Estate, Penicuik, Midlothian, EH26 0PH, Scotland
2 SAC Sustainable Livestock Systems, Sir Stephen Watson Building, Bush Estate, Penicuik, Midlothian, EH26 0PH, Scotland
3 Welsh Institute of Rural Studies, University of Wales, Aberystwyth SY23 3AL, Wales
*Additional informants and contributors are listed in Appendix 1

2
Table of Contents Executive summary
1. Introduction and background 2. Species specific energy and nutrient efficiencies
1. Meat-production systems a. Broilers b. Turkeys c. Geese d. Ducks e. Pigs f. Sheep g. Beef
2. Egg production 3. Milk production
a. Goats b. Cows
4. Overview and discussion of efficiencies in different systems
3. The feasibility of incorporating lifecycle nutrient efficiency traits into breeding programmes for farmed livestock species
4. General discussion and conclusions
Appendix 1: Informants and contributors to the study
Appendix 2: Tabled modelling assumptions
Appendix 3: Collected references

3
Executive Summary
The purpose of this project was to identify breed or genetic line effects on energy and nutrient efficiency. We have focussed attention on feed energy and protein and have not included any comments on variation in the efficiencies of use of other nutrients. The literature was reviewed and productivity was modelled for ten production systems that produce meat (broilers, turkeys, ducks, geese, pigs, sheep and beef), eggs (laying hens) or milk (dairy goats and cows). Energy and protein efficiencies are defined here as the quantity of energy or protein in the animal product (e.g. a kg of live weight, milk or eggs) as a proportion of the quantity of energy or protein in the feed consumed to produce that product. The term Feed Conversion Ratio (FCR) is the most frequently used estimate of feed efficiency in the industry. It is also quite widely used for breeding purposes and for these reasons is included in this analysis. It quantifies how many kg of feed are consumed for the production of 1 kg animal product. It is, in effect, an inverse measure of efficiency. An improvement in FCR is generally associated with improvements in energy and protein efficiency. Recently, estimates of Residual Feed Intake (RFI) have been applied by parts of the industry as yet another index of efficiency. RFI indicates the difference between the measured feed intake of an animal and its requirements for that feed based on the population average requirements for maintenance and production. Relatively low values for RFI therefore indicate relatively high efficiency.
Our models show, for each of the investigated production systems, the effect of production level alone, i.e. average daily body weight gain or weight of milk or eggs produced by the producing animal per se, on energy and nutrient efficiency. Higher levels of efficiency at higher levels of productivity are the simple consequence of variations in the proportions of consumed energy and protein that are utilised for maintenance requirements of the producing animal. This supports the historic and current livestock breeding approach of reducing the age of animals entering the food chain e.g. growth rate.
Over an animal’s productive lifetime energy and protein efficiency is also affected considerably by variation in the amounts of feed (and therefore energy and protein) required to allow the animal to develop to the point at which it is in its productive state (i.e. yielding meat, eggs or milk). Low prolificacy of the breeding population has a strong negative effect on all efficiencies when breeding animals do not enter the food chain. However, much of the variation in efficiency caused by variation in number of offspring (as a direct result of variation in longevity on efficiency of the system as a whole) disappears if most breeding animals enter the food chain. The efficiency of livestock production systems can thus be improved by maximising the number of animals entering the food chain (e.g. minimising juvenile mortality and controlling disease to reduce morbidity and mortality).
Although information is not available for all species, effects of breed or genetic line on digestive efficiency are generally small if they are observed at all. Even large and small dairy cow breeds (e.g. Holsteins vs. Jerseys) or genetic lines of broilers divergently selected for lean and lipid growth or ‘efficient’ and ‘inefficient’ layer lines are usually observed to extract very similar amounts of digestible energy or ME per kg feed. There is some evidence that local unimproved breeds sometimes extract more digestible energy than international improved breeds from poor quality (i.e. high fibre and/or low protein) feeds but not from high quality feeds. Within the bounds of the research reviewed (i.e. controlled experiments) there

4
is no convincing evidence for useful differences in digestive capacity between improved genotypes fed good quality diets in modern production systems. The existence of genotype x environment interactions for digestive efficiency cannot be ruled out but these are most likely to be relevant in extensive systems that might be found most readily in the developing world. Most research on energy loss via methane emissions from ruminant animals shows that, compared to diet effects, the effects of breed or genetic line effects on methane emissions are small. There is some recent evidence, however, that variation between genotypes in methane release exists (and is sometimes repeatable), which could be related to differences in the structure of the total bacterial population and the methanogenic archeal population between high methane and low methane producing cattle. However, early life nutrition can also significantly affect the microbial population developing in the rumen and the extent to which the basis of such variation in the rumen microbial population is genetic is at present unknown. Whether or not differences in maintenance requirements for energy and protein exist between breeds or genetic lines seems to be affected considerably by the manner in which these requirements are expressed and when these are estimated. Several studies suggest that most variation between breeds or genetic lines in net energy for maintenance disappears when the actual and mature protein (or lean) size of the genotypes are taken into account. There is evidence from various species (including pigs, cattle and laying hens) that variation in metabolisable energy requirements for maintenance does exist and may be related to differences between genotypes in activity and thermal regulation (e.g. energy use for temperature regulation in relation to feather cover in birds). No convincing evidence was found from whole animal studies for useful differences between breeds or genetic lines in the efficiencies of energy and protein utilisation proper (i.e. the efficiency with which animals convert feed energy and protein in the specific functions of generating animal growth, eggs or milk). There is, however, some recent research that suggests differences between genotypes in the functioning of specific mitochondrial respiration chains, especially when genotypes with high or low RFI are compared. This could in principle result in genotype effects on energetic (or protein) efficiency at the level of the animal as a whole. However, more evidence of the existence of such physiological variation is required before exploitation can be considered. Differences between breeds or genetic lines in the lean/lipid composition of gain can have a considerable effect on feed efficiency as measured by FCR. This is because FCR uses weight gain, and not specifically protein or fat gain, as the measure of output productivity. As lean gain has a high content of water, and fat does not, the energy content of a kg of lean gain is very much lower than a kg of fat gain. An increase in the lean/lipid ratio of gain will be associated with increased protein efficiency only if a larger proportion of feed protein is partitioned towards protein retention. However, an increase in lean/lipid ratio in body weight gains will have limited (or even negative) effects on energetic efficiency proper. Variation in productivity of the breeding population (e.g. time between two subsequent parturitions, litter size and survival rate, longevity) has a considerable (indirect) effect on energy and nutrient efficiencies of the system as a whole, especially for systems in which the breeding population consumes a considerable proportion of total feed consumption, such as beef and sheep production.

5
Current selection goals related to nutrient efficiency, such as FCR or RFI remain highly desirable in relation to nutrient efficiency as well as for economic reasons; limitations exist because of the costs of measuring FCR or RFI, especially for ruminants, and more appropriate models for the estimation of RFI may be needed. New technologies now being applied to animal breeding represent a powerful opportunity to prise open the “black box” underlying the response to selection and fully understand the genetic architecture controlling complex polygenic traits such as energy partitioning. The introduction of high density marker (SNP) panels has led to the development of theoretical approaches to selection based on this genomic information. The development of smaller scale SNP arrays that have lower costs should be investigated to identify the genetic control of traits relating to nutrient efficiency. As genetic improvement techniques are refined and become more powerful using modern DNA based methods, it is vital that all participants in food production on-farm are aware of the genetic differences between animals and account for such genetic variation when formulating management strategies such as feeding, housing and health treatments. The use of such tools should enable breeding goals to be achieved more quickly and are thus seen to enhance our ability to further improve the efficiency of production systems (i.e. improve the efficiency of current breeding programmes). The review has highlighted a number of specific research requirements. To further improve energy and nutrient efficiencies, research should be aimed at: (i) Reducing losses, i.e. animals not entering the food chain (e.g. by selection strategies for sow longevity and piglet survival), (ii) Using new genomic tools to further improve efficiency by minimising duration of the period between birth and entering the food chain (i.e. improve growth rates) and increasing output relative to maintenance requirements in dairy and egg production and (iii) Reducing maintenance requirements of animals by selection for optimal behavioural traits, and (iv) development of cheaper tools, for genomic selection (e.g. SNP arrays) as well as for accurate estimation of traits (such as RFI).
This review has described the role of nutrient utilisation in historic and current breeding goals in livestock species, be it by it’s inclusion in breeding programmes directly, as in the case of pigs and poultry, or indirectly via correlated traits such as growth rate and production output, as in the case of many ruminant species. The inclusion of traits related to the lifetime performance of animals (e.g., fertility, health and disease) may also have an additional impact on the overall lifetime nutrient efficiency of the animal itself, as well as the wider system if such traits reduce productive losses. It is important to note that a breeding goal that considers overall lifetime nutrient efficiency may not match directly to system efficiency as considered by the farmer in terms of economic performance and may also not map to environmental benefits or animal welfare goals. Overall we conclude that:
• Gross efficiencies of use of feed energy and protein use are affected by the level of productivity of individuals as a simple consequence of the dilution of maintenance costs in the overall energy or protein demand

6
• There is little evidence that breeds or genetic lines differ measurably in the partial efficiencies of production of specific constituents (lean, fat, milk, eggs) although there is some evidence that suggests that maintenance costs may vary
• The energy and nutrient costs of bringing an individual to its point of productivity have a material effect on efficiencies over lifetimes, but at a system level many of these differences become small if breeding stock also enters the food chain.
• Most selection goals that are currently used for economic reasons are thought to be associated with increased nutrient efficiency and historic and current breeding goals have significantly improved the nutrient efficiency of livestock production.
• New genomic technologies are likely to improve the rates of ‘nutrient efficiency’ return of current industry breeding programmes
• A number of specific research requirements that would benefit the selection of more nutrient efficient animals have been identified

7
Chapter 1: Introduction and background
Approximately three quarters of the UK land area (18.6 m ha) is classed as agricultural land (including woodlands) with an animal population of 10.3 million cattle, 33.9 million sheep, 4.8 million pigs and 167.7 million poultry (Agriculture in the United Kingdom 20071). Livestock systems are an important source of environmental pollution, including phosphorus and greenhouse gas (GHG) emissions such as methane (CH4) and nitrous oxide (N2O). The mitigation of the environmental impact of livestock production systems is increasingly recognised as an important objective and a necessary part of the UK’s overall climate change obligations.
Understanding variation in the way in which different breeds of livestock, or different animals within breeds, utilise the available resources in a system can help to develop tools that can mitigate negative environmental impacts of the supply of animal products.. Different breeds, or different genotypes within a breed, vary in the efficiency with which they convert the resources within a system into animal product and, consequently, in the proportion of used resources that ends up as waste. As a result, the environmental impact associated with the production of a kg of human food of animal origin varies as well. There is a need to understand what variation exists between breeds or between genotypes within breeds in the lifecycle efficiency with which animals utilize nutrients and how efficiency traits can best be incorporated into breeding programmes.
Variation in nutrient efficiency is mainly caused by variation in the proportion of total feed consumed that is actually utilised to synthesise a kg of consumable product such as meat, milk or eggs, i.e. by variation in production efficiency. In addition, nutrient efficiency can be affected by differences in the efficiency of utilisation of the nutrients present in a kg of feed per se, i.e. variation in utilisation efficiency. Breed differences in production and utilisation efficiency may be affected by the environment (e.g. in terms of feed quality, climatic conditions and disease pressure) and for that reason any comparison of breed differences in efficiency can only be considered in relation to genotype by environment (GxE) interactions. In view of the importance of feed energy efficiency in relation to essential parts of lifecycle efficiency (e.g. in terms of the greenhouse gases CO2 and CH4), dietary energy efficiency should be considered in tandem with dietary nutrient efficiency in any review of genetic (breed) differences in lifecycle dietary efficiency. In our view the term ‘breed difference’ should be interpreted to also encompass genetic differences, e.g. between genetic lines within a breed, which seem highly relevant at least for some farm animal species.
Genetic (breed) differences in production efficiency Many traits that are directly and immediately related to production efficiency, such as daily weight gain or milk yield and feed conversion ratio, have long been part of breeding programmes and this has no doubt resulted in genetic differences between genotypes (including breeds) in lifecycle nutrient and energy efficiency. Selection for these direct production traits has generally resulted in an increase in food intake relative to maintenance requirements and, as a result, an increase in the proportion of food energy and nutrients that is utilised directly to synthesise consumable animal product. This in itself has a direct beneficial effect on lifecycle nutrient efficiency. In addition, other, frequently longer-term, traits can be
1 https://statistics.defra.gov.uk/esg/publications/auk/2007/18%20Whole%20publication.pdf

8
part of broader breeding goals that are expected to affect lifecycle nutrient and energy production efficiency. These include:
1. The proportion of total feed requirements that have to be allocated to (mainly female) breeders of meat producing animals. Variation in this proportion for lifecycle energy and nutrient efficiency is especially relevant for species with relatively slow reproduction and low fecundity, such as beef cattle and sheep (Dickerson, 1978). This proportion will, therefore, be affected strongly by fertility traits.
2. Productive lifespan of breeding animals. An increase in productive lifespan of breeders of meat animals will not only directly affect lifecycle nutrient efficiency for breeder animals as mentioned under item 1. but will also have a beneficial effect on lifecycle nutrient efficiency in production systems that directly involve breeding animals such as dairy cows and layers.
3. Nutrient and energy efficiency can be negatively affected by health problems as a result of decreases in food intake (and, therefore, the proportion of feed that is actually utilised to synthesise animal products) and a decrease in feed utilisation efficiency (see below). As a result, there is increasing interest in using genetic (breed) differences in disease resistance traits for broader breeding programmes.
Genetic (breed) differences in nutrient utilisation efficiency The literature shows many examples of genetic (including between breed) variation in efficiency of nutrient and energy utilisation. The following broad categories can be distinguished:
1. Digestive and absorption efficiency. Variation in digestive and absorption efficiency directly affects the amount of energy and nutrients animals can extract from a kg of feed. The term digestive efficiency refers in general to the whole-tract apparent digestibility (of DM, OM, energy, CP, etc.) but in appropriate cases is refined (e.g. in enzymatic an fermentative digestion). There is some evidence for genetic (breed) differences in digestive efficiency for energy as well as amino acids in pigs and poultry (e.g. Carre et al. 2008). It has similarly been suggested that differences in digestive capacity between breeds of ruminant species may be as important as differences between monogastric species (Tolkamp & Brouwer 1993). GxE interactions can be expected to be very important in this respect (see below).
2. Fermentation efficiency. Between-animal variation in fermentation efficiency has been documented for ruminants (e.g. Hegarty et al., 2007). Such variation may have a direct effect on the availability of energy and nutrients to the animal but may also affect the lifecycle environmental consequences of a production system via the resulting variation in the amount of feed energy released as the greenhouse gas CH4.
3. Variation in maintenance requirements. There is evidence for genetic (between breed) variation in maintenance requirements which may be caused by variation in efficiency of N recycling in ruminants (e.g. Reynolds and Kristensen, 2008) or variation in heat loss (e.g. as a result of variation in feather cover or in physical activity in layers; e.g. Luiting et al. 1991). Variation in maintenance requirements in itself contributes directly to variation in production efficiency but should be considered in relation to GxE interactions (see below).
4. Efficiency of partitioning and utilisation of energy and nutrients above maintenance for synthesis. It is generally assumed that the pathways involved in synthesis of proteins and lipids are very much conserved, even across species, and that little

9
variation in efficiency of energy and nutrient utilization for synthesis per se can be expected (Emmans, 1994; Tolkamp and Kyriazakis 2009).
Genomic dissection of feed efficiency into causes such as change in body composition, growth rate, maintenance requirements, etc. can be achieved using genomic markers (Roehe et al. 2003; Mohrmann et al. 2006; Duthie et al. 2008). This dissection will allow to selection directly on the biological traits, which have been shown to be more efficient than the use of composite traits such as feed efficiency (Gunsett, 1984).
The relevance of GxE interactions Breed differences in many of the traits that affect lifecycle nutrient and energy efficiency listed above may only be evident in certain environmental circumstances (in terms of feed quality, climatic conditions and disease pressure). For instance, genetic/breed differences in digestive efficiency may be highly significant for some (class of) feeds but not be observable for others (Carre et al., 2008; Tolkamp & Brouwer, 1993). For that reason, such breed differences require specification of the conditions under which they are observed and under which conditions these differences are absent (or even reversed).
Review of breed differences in lifecycle nutrient efficiency It seems highly unlikely that, on the basis of present knowledge, firm conclusions can be drawn on existing breed differences in lifecycle nutrient efficiency for any animal species and for any production system. An overview of genetic/breed differences in traits that have a large effect on lifecycle nutrient efficiency, and the conditions under which these may be observed would, however, be very useful. Such a review should not only give an, at present, non-existing systematic inventory of our knowledge about genetic/breed differences in traits that are relevant for lifecycle nutrient efficiency (‘state of the art’) but it would also serve to identify more clearly (i) the traits that are most likely to contribute most to genetic/breed differences in lifecycle nutrient efficiency and (ii) identify the type of research that is required to determine with more confidence which genotypes/breeds are most suitable for which environments from a lifecycle nutrient efficiency point of view.
Existing differences between breeds or genotypes in nutrient efficiency are largely related to differences in productivity and cannot be considered in isolation. For that reason, models were included to illustrate such effects of productivity for all species. This serves (i) to illustrate the relevance of productivity for nutrient efficiency and (ii) provide a bench mark for the interpretation of the relevance of observed specific differences in nutrient efficiency between genotypes.
This project will summarise current knowledge on the characteristics of breeds, and genetic variability within breeds, for nutrient and energy efficiency traits and on the incorporation of lifecycle nutrient efficiency targets into animal breeding programmes as per Recommended Action 31 of the UK National Action Plan for FAnGR2. For each of ten production systems the findings from a literature review will be presented first in Chapter 2, in agreement with the first Objective of the current project (Review of current knowledge on lifecycle dietary nutrient efficiency for a range of different breeds). For each production system this will be accompanied by the results of specifically developed models that predict effects of variation
2 “…model/characterise breeds for their lifecycle nutrient efficiency and to investigate the incorporation of lifecycle nutrient efficiency traits into breeding programmes in all livestock species.”

10
in production level proper (i.e. growth of meat producing animals, milk or egg yield per animal) on energy and nutrient efficiency. Where appropriate, the effects of including other relevant aspects of the production system as a whole (especially: effects of including the breeding animals required to produce the production animals themselves) on energy and nutrient efficiency will be included. Although occasionally references are made to other nutrients, the report concentrates on protein as the main nutrient for which sufficient data are available. Three estimators of energetic efficiency are used across all species. Because of its current relevance for the breeding industry, the Feed Conversion Ratio (FCR), which is actually the inverse of efficiency proper (because the FCR quantifies the amount of feed required to produce a kg of animal product) is included. A separate paragraph will be devoted to an alternative to FCR, i.e. the calculation with an animal model of residual feed consumption (RFC). Two real energetic efficiency estimates are included as well: the Gross Energy (GE) and the Metabolisable Energy (ME) efficiency that quantifies the amount of net energy (NE) in the animal or animal product per MJ of GE or ME, respectively, in the required feed for the production process.
Chapter 3 is devoted to the second objective of the current study (i.e. The feasibility of incorporating lifecycle nutrient efficiency traits into breeding programmes for farmed livestock species). Chapter 4 gives a general discussion and conclusions, including gaps in current knowledge and suggested approaches to fill these gaps (Objective 3). Appendix 1 lists informants and contributors to this study, Appendix 3 contains tables with the information used in modelling and Appendix 3 gives the complete list of studied literature.
References
Carre B, Mignon-Grasteau S and Juin H. 2008. Breeding for feed efficiency and adaptation to feed in poultry. World's Poultry Science Journal 64, 377-390.
Dickerson GE 1978. Animal size and efficiency: basic concepts. Animal Production 27, 367-379. Duthie C, Simm G, Doeschl-Wilson A, Kalm E, Knap PW and Roehe R 2008. Quantitative trait loci
for chemical body composition traits in pigs and their positional associations with body tissues, growth and feed intake. Animal Genetic 39, 130-140.
Emmans GC. 1994. Effective energy – a concept of energy-utilization applied across species. British Journal of Nutrition 71, 801-821
Gunsett FC 1984. Linear index selection to improve traits defined as ratios. Journal of Animal Science 59, 1185-1193.
Hegarty RS Goopy JP, Herd RM and B McCorkell 2007. Cattle selection for lower residual feed intake have reduced daily methane production. Journal of Animal Science 85, 1479-1486.
Luiting P, Schrama JW, van der Hel W, Urff EM.1991. Metabolic differences between white leghorns selected for high and low residual food consumption. British Poultry Science, 32: 763 – 782
Mohrmann M, Roehe R, Knap PW, Looft H, Plastow GS and Kalm E 2006. Quantitative trait loci associated with AutoFOM grading characteristics, carcass cuts and chemical body composition during growth of Sus scrofa. Animal Genetics 37, 435-443.
Reynolds CK and Kristensen NB. 2008. Nitrogen recycling through the gut and the nitrogen economy of ruminants: An asynchronous symbiosis. Journal of. Animal Science 86: ,E293-E305
Roehe R, Plastow GS and Knap PW 2003. Quantitative and molecular genetic determination of protein and fat deposition. Journal HOMO of Comparative Human Biology 54:119-131.
Tolkamp BJ and Brouwer BO. 1993. Statistical review of digestion in goats compared with other ruminants. Small Ruminant Research 11,107–123.
Tolkamp BJ and Kyriazakis I. 2009. Toward a new practical energy evaluation system for dairy cows. Animal 3, 307-314.

11
Chapter 2: Species specific energy and nutrient efficiencies
Feed Conversion Ratio (FCR) is by far the most widely used estimator of feed efficiency in the animal industry. The FCR is affected by a large number of factors, some of which will be discussed in detail below. In agreement with common usage in the literature, FCR’s for monogastric species have been expressed in terms of kg feed per kg product but for ruminants (in view of the large variation in water content of the consumed feeds) in terms of kg feed DM per kg animal product.
In the literature, most energetic efficiencies are expressed in terms of NE in the animal (product) divided by the amount of ME consumed. Because the amount of ME derived from feed gross energy (GE) may depend on the digestive efficiency, we have added in all comparisons the estimated GE efficiency, which expresses the amounts of NE as a proportion of the GE consumed. Many different estimates of digestive efficiency are provided in the literature. In the vast majority of cases this refers to apparent digestibilities (i.e. based on the amounts of a diet component that are consumed and excreted in the faeces). Sometimes metabolisability is used as a good estimator of digestive efficiency. Where different aspects of digestive efficiency are considered (e.g. enzymatic digestion vs. lower-tract fermentation), this is clearly indicated in the text. In addition, we reviewed evidence for effects on protein efficiency, i.e. the ratio of the amount of protein in the animal (product) and the protein contained in the consumed feed. The chapter is divided in meat, egg and milk production systems.
It is evident that efficiency is determined to a large extent by productivity and that specific genotype effects can only be evaluated by taking effects of productivity into account. For that reason, effects of productivity of the production animal per se (e.g. broiler, slaughter pig, lamb, dairy cow) on efficiency are illustrated for all species. Where this is relevant, efficiencies for the production system as a whole are also provided by taking into account variation in productivity of the breeder population.
12
2.1 Meat production systems
2.1.1 Broilers
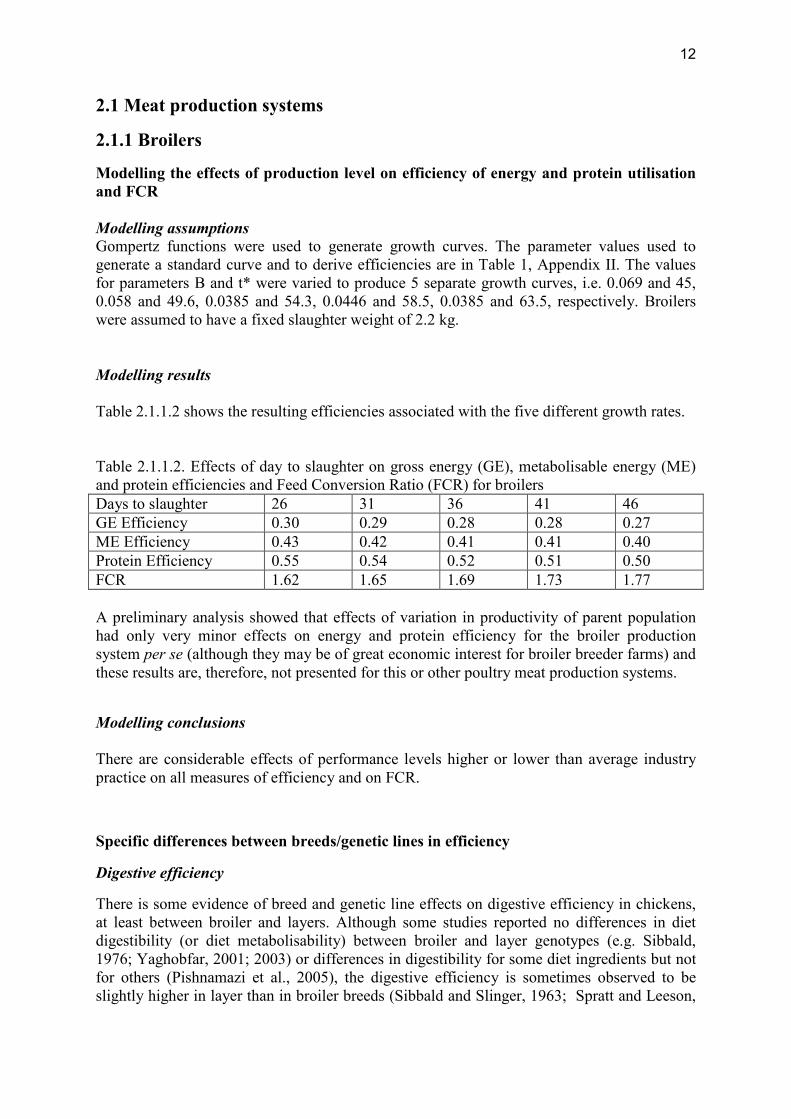
Modelling the effects of production level on efficiency of energy and protein utilisation and FCR Modelling assumptions Gompertz functions were used to generate growth curves. The parameter values used to generate a standard curve and to derive efficiencies are in Table 1, Appendix II. The values for parameters B and t* were varied to produce 5 separate growth curves, i.e. 0.069 and 45, 0.058 and 49.6, 0.0385 and 54.3, 0.0446 and 58.5, 0.0385 and 63.5, respectively. Broilers were assumed to have a fixed slaughter weight of 2.2 kg. Modelling results Table 2.1.1.2 shows the resulting efficiencies associated with the five different growth rates. Table 2.1.1.2. Effects of day to slaughter on gross energy (GE), metabolisable energy (ME) and protein efficiencies and Feed Conversion Ratio (FCR) for broilers Days to slaughter 26 31 36 41 46 GE Efficiency 0.30 0.29 0.28 0.28 0.27 ME Efficiency 0.43 0.42 0.41 0.41 0.40 Protein Efficiency 0.55 0.54 0.52 0.51 0.50 FCR 1.62 1.65 1.69 1.73 1.77 A preliminary analysis showed that effects of variation in productivity of parent population had only very minor effects on energy and protein efficiency for the broiler production system per se (although they may be of great economic interest for broiler breeder farms) and these results are, therefore, not presented for this or other poultry meat production systems.
Modelling conclusions There are considerable effects of performance levels higher or lower than average industry practice on all measures of efficiency and on FCR.
Specific differences between breeds/genetic lines in efficiency
Digestive efficiency
There is some evidence of breed and genetic line effects on digestive efficiency in chickens, at least between broiler and layers. Although some studies reported no differences in diet digestibility (or diet metabolisability) between broiler and layer genotypes (e.g. Sibbald, 1976; Yaghobfar, 2001; 2003) or differences in digestibility for some diet ingredients but not for others (Pishnamazi et al., 2005), the digestive efficiency is sometimes observed to be slightly higher in layer than in broiler breeds (Sibbald and Slinger, 1963; Spratt and Leeson,

13
1987) and Pishnamazi et al. (2005) suggest that layers may derive around 2.5% more ME from their feed than broilers. Part of these differences may be associated with lower voluntary intakes in layer than in broiler genotypes.
Higher digestive efficiency is sometimes (but not always, e.g. Buyse et al., 1998) recorded in comparisons of genetic broiler lines selected for FCR when these are compared with control lines or lines selected for high growth rates (Leenstra and Pit, 1987; Jorgensen et al., 1990; TenDoeschate et al., 1993).
Several reports suggest that there are generally no differences in digestive efficiency, as measured by ME or AME, between genetic lines that have been differently selected for fat content (Leclercq and Saadoun ,1982; Laurin et al.,1985; Geraert et al., 1988; Geraert et al., 1990; Buyse et al., 1998). Boldaji et al. (1981) reported significantly lower TME values for dwarf compared to normal-sized birds.
Heritabilities for digestive efficiency in birds were estimated as reasonable high by Mignon-Grasteaux et al. (2004). Peron et al. (2006), however, recorded differences between broiler lines specifically selected for low or high digestive efficiency and concluded that heritabilities were low; such differences were observed in young birds (but disappeared in older animals), were affected by diet type (Carre et al., 2008) and were probably related to differences between lines in the size of their digestive organs (Rougiere et al., 2009). The digestibilities observed in the high line were stable and according to expectations but in the low line these were highly variable and probably affected by many different factors, including digestive disorders (Peron et al., 2006).
A single study has suggested that only modest improvements could be expected by direct selection on phytate phosphorus bio-availability in chickens as a result of very low heritability estimates but whether or not this is correlated with other (negative) traits is at present unknown (Zhang et al., 2005).
The results of comparisons in digestive efficiency between breeds or lines are quite variable and seem to be affected by differences in animal size, feeding level and feeding regime (Zhang and Aggrey, 2003). Some research has been done in attempts to explain variation between genotype in digestive efficiency, such as variation in size of gastro-intestinal tract, digesta retention time and digestive enzyme activity (Kaminska, 1979; Shires et al., 1987; Buyse et al., 1999) and there is some evidence that selection for fast growth rates can result in higher digestive enzyme activity (Zhang and Aggrey, 2003).
In conclusion, differences between breeds/genetic lines in digestive efficiency of broilers are, if observed at all, generally small. Data show no differences in digestive efficiency between lean and fat lines but there is some evidence that it is slightly increased in lines selected for low FCR and slightly decreased in dwarf genotypes
Maintenance requirements
There is little evidence for variation between breeds/lines in maintenance requirements, at least not between lean and fat lines (Geraert et al., 1990) or between lines selected for fast growth or decreased FCR (Buysse et al., 1998). The suggestion from studies with layers (Luiting 1990; Luiting et al., 1991) that variation between genotypes in maintenance requirements could be due to variation in activity and that, consequently, a higher energetic efficiency in selected broiler lines could be due to a decrease in activity and more docile

14
animals has not been confirmed in a study by Skinner-Noble et al. (2003). They concluded that, in contrast to expectations, broilers from lines selected for good FCRs were actually less lethargic than lines selected for poor FCRs. It is evident that the use of dwarf genotypes is expected to lead to a small decrease in the total maintenance energy and protein requirements of broiler breeding stock.
Efficiency of energy and protein utilisation proper
There is very little information on breed or genetic line effects on efficiency of energy and protein utilisation proper (Lopez and Leeson, 2008). There is no evidence that there are significant differences between lean and fat broiler lines as measured by diet induced thermogenesis (Swennen et al., 2006) and it is frequently assumed that energetic efficiency of protein and lipid retention from a given substrate is very similar across genotypes (or even species; Emmans, 1994; 1997). There is, however, some recent evidence of differences between genetic lines selected for low or high FCR (or residual feed intake, RFI) in some (but not all) mitochondrial functions and, as a result, efficiency of ATP production in broiler muscle, liver and/or duodenum tissue (Bottje and Carstens 2009).
Fractional rates of protein turnover did not differ between a line selected for increased breast yield and a control line (Pym et al., 2004). There were differences between these lines in the proportion of feed protein retained in the body but that was to be expected because the different genotypes received diets with the same protein content (Pym et al., 2004). Differences between high and low abdominal fat lines in fractional protein turnover and deposition were a direct result of differences in intake rather than the result of genetic differences in efficiency of protein utilisation proper (Pym et al., 2004). Variation in oxidative damage to proteins between more and less energy efficient genotypes could affect protein turnover rate and, therefore, protein efficiency (Bottje and Carstens 2009), although no firm evidence for that is at present available. Studies on breed/genetic line effects on activities of specific tissue enzymes that could affect pathways of energy utilisation have started relatively recently (e.g. Collin et al., 2003; 2009) and more information like this can be expected in the near future.
Other factors
The occurrence of disease and variation in liveability/mortality of broilers not only affect the economics of broiler production systems but these factors can also have an important effect on the efficiency of energy and nutrient utilisation at system level (see review by Flock et al., 2005). Considerable progress in this area has been made as typical average mortality percentages have decreased from double digits in the 1930’s to a few percent only today (Havenstein, 2003; Flock et al., 2005) and Emmerson (2003) estimated that improvements in liveability could contribute significantly to efficiency and profitability of the system as a whole. Broiler breeders are working to further reduce over-all broiler mortality, leg disorders and susceptibility to heart/lung insufficiency, which is possible while continuing to select for increased growth rate (Laughly, 2007). The most important broiler diseases have recently been scored with the aim of helping to clarify the host genetic control of resistance (Davies et al. 2009). Exactly how increased disease resistance will affect efficiency of energy and protein utilisation is, however, at present not clear (see general discussion chapter).

15
References
Boldaji F, Roush WB, Nakaue HS, Arscott GH.1981. True metabolizable energy values of corn and different varieties of wheat and barley using normal and dwarf single comb White Leghorn Roosters. Poultry Science 60, 225-227.
Bottje WG and Carstens GE. 2009. Association of mitochondrial function and feed efficiency in poultry and livestock species. Journal of Animal Science 87, E48-E63.
Buyse J, Michels H, Vloeberghs J, Saevels P, Aerts JM, Ducro B, Berckmans D, Decuypere E. 1998. Energy and protein metabolism between 3 and 6 weeks of age of male broiler chickens selected for growth rate or for improved food efficiency. British Poultry Science 39, 264-272.
Buyse J, Michels H, Vloeberghs J, Saevels P, Aerts JM, Ducro B, Berckmans D, Decuypere E. 1998. Energy and protein metabolism between 3 and 6 weeks of age of male broiler chickens selected for growth rate or for improved food efficiency. British Poultry Science 39, 264-272.
Buyse J, Leenstra FR, Zeman M, Rahimi G, Decuypere E. 1999 A comparative study of different selection strategies to breed leaner meat-type poultry. Poultry and Avian Biology Reviews 10, 121-142.
Carre B, Mignon-Grasteau S, Juin, H. 2008. Breeding for feed efficiency and adaptation to feed in poultry. Worlds Poultry Science Journal 64, 377-390.
Collin A, Malheiros RD, Moraes VMB, Van As P, Darras VM, Taouis M, Decuypere E, Buyse J. 2003. Effects of dietary macronutrient content on energy metabolism and uncoupling protein mRNA expression in broiler chickens. British Journal of Nutrition 90, 261-269.
Collin A, Swennen, Q,Skiba-Cassy, S Buyse, J, Chartrin P, Le Bihan-Duval E, Crochet S, Duclos M, Joubert R, Decuypere E, Tesseraud, S. 2009 Regulation of fatty acid oxidation in chicken (Gallus gallus): Interactions between genotype and diet composition. Comparative Biochemistry and Physiology B – Biochemistry and Molecular Biology 153, 171-177.
Davies G, Genini S, Bishop SC, Giuffra E. 2009. An assessment of opportunities to dissect host genetic variation in resistance to infectious diseases in livestock. Animal 3, 415-436.
Emmans GC. 1994. Effective energy – a concept of energy utilization applied across species. British Journal of Nutrition 71, 801-821
Emmans GC. 1997. A method to predict the food intake of domestic animals from birth to maturity as a function of time. Journal of Theoretical Biology 186, 189-199.
Emmerson, D. 2003. Breeding objectives and selection strategies for broiler production in (W.M Muir and S.E. Aggrey, eds.) Poultry Genetics, Breeding and Biotechnology. CABI Publishing, Wallingford, UK.
Geraert, P. A. MacCleod MG, LeClercq, YDB. 1988. Energy metabolism in genetically fat and lean chickens: Diet- and cold-Induced thermogenesis. Journal of Nutrition 118, 1232-1239.
Geraert PA, MacLeod MG, Larbier M, Leclercq B; 1990. Nitrogen metabolism in genetically fat and lean chickens. Poultry Science 69, 1911-1921.
Havenstein GBH, Ferket PR and Qureshi MA. 2003. Growth, livability, and feed conversion of 1957 versus 2001 broilers when fed representative 1957 and 2001 broiler diets. Poultry Science 82, 1500-1508.
Jorgensen H, Sorensen P, Eggum BO. 1990; Protein and energy-metabolism in broiler-chickens selected for either body weight gain or feed efficiency. British Poultry Science 31, 517-524.

16
Kaminiska BZ, 1979. Food intake in the young chick. In: K.N, Boorman and Freeman, B.M. (Eds.), Food intake regulation in poultry. Br. Poult. Sci. Ltd. Edinburg, Scotland, PP: 199-206.
Laughlin K 2007. The Evolution of Genetics, Breeding and Production. Temperton Fellowship Report No.15. Harper Adams University College.
Laurin DE, Touchburn SP, Chavez ER and Chan CW. 1985. Methods of measuring energy utilization in broilers: Effect of genetic line and presence of supplemental dietary fat. Poultry Science 64, 969–978.
Leclercq B, and Saadoun A. 1982. Selecting broilers for low or high abdominal fat: comparison of energy metabolism of the lean and fat lines. Poultry Science 61, 1799–1803.
Leenstra FR and Pit R. 1987. Fat deposition in a broiler sires strain. 2. Comparisons among lines selected for less abdominal fat, lower feed conversion ratio and higher body weight after restricted and ad libitum feeding. Poultry Science, 66, 193-202.
Lopez G, Leeson S. 2008. Review: Energy partitioning in broiler chickens. Canadian Journal of Animal Science 88, 205-212
Luiting P. 1990. Genetic variation of energy partitioning in laying hens. Causes of variation in residual feed consumption. Worlds Poultry Science Journal 46, 133-152.
Luiting P, Schrama JW, van der Hel W and Urff EM. 1991. Metabolic differences between White Leghorns selected for high and low residual food consumption. British Poultry Science 32, 763-782.
Mignon-Grasteau S, Muley N, Bastianelli D, Gomez J, Peron A, Sellier N, Millet N, Besnard J, Hallouis JM, Carre B. 2004. Heritability of digestibilities and divergent selection for digestion ability in growing chicks fed a wheat diet. Poultry Science 83, 869-877.
Peron A, Gomez J, Mignon-Grasteau S, Sellier N, Besnard J, Derouet M, Juin H, Carre B. 2006. Effects of wheat quality on digestion differ between the D+ and D- chicken lines selected for divergent digestion capacity. Poultry Science 85, 462-469.
Pishnamazi A, Pourezza J, Edriss MA, Samie AH. 2005. Influence of broiler breeder and laying hen breed on the apparent metabolisable energy of selected feed ingredients. International Journal of Poultry Science 4, 163-166.
Proudman JA, Mellen WJ and Anderson DL. 1970. Utilization of feed in fast and slow-growing lines of chickens. Poultry Science, 49, 961-972.
Pym RAE, Leclercq B, Tomas FM, Tesseraud S. 2004. Protein utilisation and turnover in lines of chickens selected for different aspects of body composition. British Poultry Science 45, 775-786.
Rougiere N, Gomez J, Mignin-Grateau S, Carre B. 2009. Effects of diet particle size on digestive parameters in D+ and D- genetic chicken lines selected for divergent digestion efficiency. Poultry Science 88, 1206-1215.
Shires A, Thompson, JR, Turner, BV, Kennedy PM and Goh, YK. 1987. Rate of passage of corn-canola meal and corn-soybean meal diets through the gastrointestinal tract of broiler and White Leghorn chickens. Poultry Science 66, 289-298.
Sibbald IR. 1976. True metabolisable energy values of several feedingstuffs measured with roosters, laying hens, turkeys and broiler hens. Poultry Science 55, 1459-1463.
Sibbald IR and Slinger SJ. 1963. The effects of breed, sex, an arsenical and nutrient density on the utilization of dietary energy. Poultry Science 42, 1325–1332.
Skinner-Noble DO, Jones RB and Teeter RG. 2003. Components of feed efficiency in broiler breeding stock: Is improved feed conversion associated with increased docility and lethargy in broilers. Poultry Science 82, 532-537.
Spratt RS and Leeson, S. 1987. Determination of metabolizable energy of various diets using Leghorn, Dwarf and regular broiler breeder hens. Poultry Science 66, 314-317.

17
Swennen Q, Janssens GPJ, Collin A, Le Bihan-Duval E, Verbeke K, Decuypere E, Buyse J. 2006. Diet-induced thermogenesis and glucose oxidation in broiler chickens: Influence of genotype and diet composition. Poultry Science 85, 731-742.
Swennen Q, Janssens GPJ, Decuypere E, Buyse J 2004. Effects of substitution between fat and protein on feed intake and its regulatory mechanisms in broiler chickens: Energy and protein metabolism and diet-induced thermogenesis. Poultyry Science 83, 1997-2004.
TenDoeschate RAHM, Scheele CW, Schreurs WAM and van der Klis JD. 1993. Digestibility studies in broiler-chickens – influence of genotype, age, sex and method of determination. British Poultry Science 34, 131-146.
Yaghobfar A. 2001. Effect of genetic line, sex of birds and the type of bioassay on the metabolisable energy value of maize. British Poultry Science 42, 359-353.
Yaghobfar A. 2003. The effects of strain, age and sex of birds on the determined metabolisable energy of maize and fishmeal. British Poultry Science 44, S38-S39.
Zhang W, Aggrey SE, Pesti GM, Bakalli RI, Edwards HM. 2005. Genetic analysis on the direct response to divergent selection for phytate phosphorus bioavailability in a random bred chicken population. Poultry Science 84, 370-375.
Zhang W and Aggrey SE. 2003. Genetic variation in feed utilization efficiency of meat-type chickens. Worlds Poultry Science Journal 59, 328-339.
18
2.1.2 Turkeys
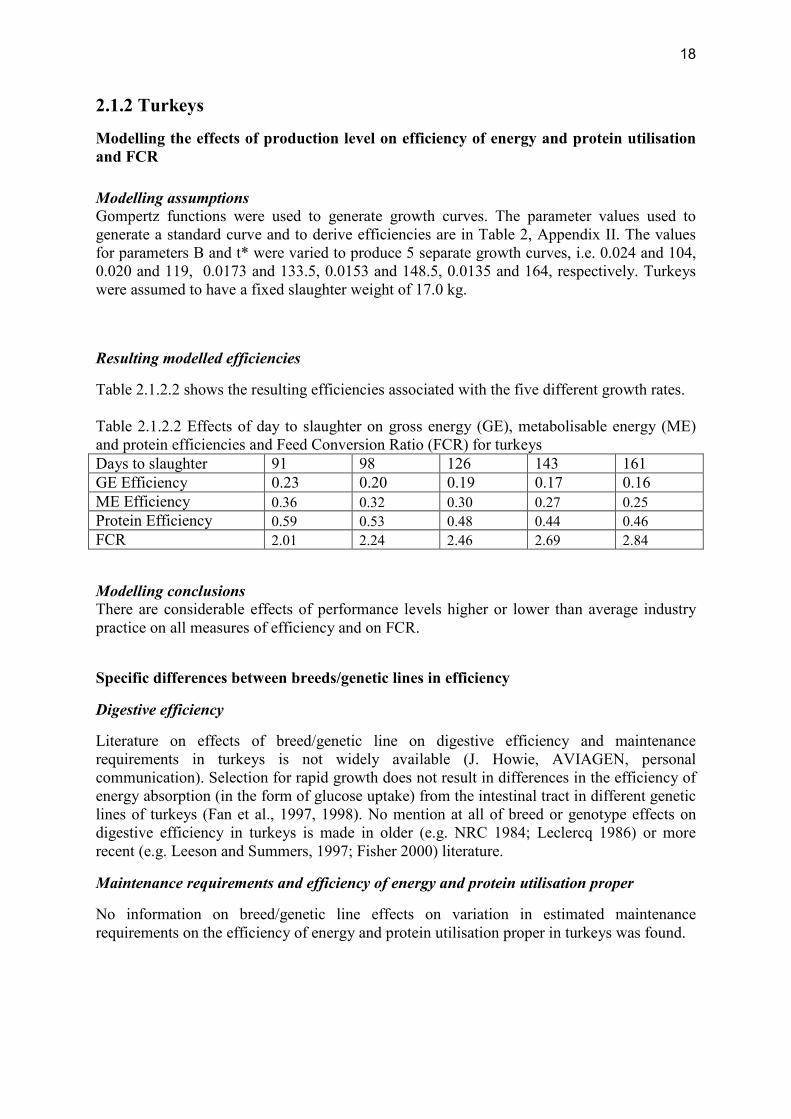
Modelling the effects of production level on efficiency of energy and protein utilisation and FCR Modelling assumptions Gompertz functions were used to generate growth curves. The parameter values used to generate a standard curve and to derive efficiencies are in Table 2, Appendix II. The values for parameters B and t* were varied to produce 5 separate growth curves, i.e. 0.024 and 104, 0.020 and 119, 0.0173 and 133.5, 0.0153 and 148.5, 0.0135 and 164, respectively. Turkeys were assumed to have a fixed slaughter weight of 17.0 kg.
Resulting modelled efficiencies
Table 2.1.2.2 shows the resulting efficiencies associated with the five different growth rates. Table 2.1.2.2 Effects of day to slaughter on gross energy (GE), metabolisable energy (ME) and protein efficiencies and Feed Conversion Ratio (FCR) for turkeys Days to slaughter 91 98 126 143 161 GE Efficiency 0.23 0.20 0.19 0.17 0.16 ME Efficiency 0.36 0.32 0.30 0.27 0.25 Protein Efficiency 0.59 0.53 0.48 0.44 0.46 FCR 2.01 2.24 2.46 2.69 2.84 Modelling conclusions There are considerable effects of performance levels higher or lower than average industry practice on all measures of efficiency and on FCR.
Specific differences between breeds/genetic lines in efficiency
Digestive efficiency
Literature on effects of breed/genetic line on digestive efficiency and maintenance requirements in turkeys is not widely available (J. Howie, AVIAGEN, personal communication). Selection for rapid growth does not result in differences in the efficiency of energy absorption (in the form of glucose uptake) from the intestinal tract in different genetic lines of turkeys (Fan et al., 1997, 1998). No mention at all of breed or genotype effects on digestive efficiency in turkeys is made in older (e.g. NRC 1984; Leclercq 1986) or more recent (e.g. Leeson and Summers, 1997; Fisher 2000) literature.
Maintenance requirements and efficiency of energy and protein utilisation proper
No information on breed/genetic line effects on variation in estimated maintenance requirements on the efficiency of energy and protein utilisation proper in turkeys was found.

19
FCR
Improvements in FCR associated with selection of turkey lines for faster growth is well documented (e.g. Havenstein et al., 2007), but very little information is available on the causes of this improvement, apart from some increase in carcass leanness. Because there is generally a good (inverse) relationship between FCR and efficiency of energy and protein utilisation, such improvements in FCR can be expected to have increased these efficiencies, unless unfavourably correlated with other important factors, such as disease resistance (see below).
Other factors
Differences between genetic lines in disease resistance and physiological response to stress suggest that increasing selection for BW of turkeys is accompanied by changes in the stress response resulting in increased susceptibility to opportunistic bacterial and viral infections, which can be expected to have a negative effect on efficiency as measured by FCR (Nestor et al., 1996; Huff et al., 2005). Similarly, Li et al., (1999) found that genetic selection for increased BW in turkeys may affect disease resistance and, therefore, FCR and efficiency, although such effects are difficult to quantify at present. No significant differences between turkey breeds in mortality up to 18 weeks were observed by Roberson et al. (2003).
References
Fan YK, Croom WJ, Christensen VL, Bird AR, Daniel LR, McBride BW, Eisen EJ. 1998. Apparent energetic efficiency of jejunal glucose uptake in young adult turkeys selected for rapid growth. Canadian Journal of Animal Science 78, 301-306.
Fan YK, Croom J, Christensen VL, Black BL, Bird AR, Daniel LR, McBride BW, Eisen EJ. 1997. Jejunal glucose uptake and oxygen consumption in turkey poults selected for rapid growth. Poultry Science 76, 1738-1745.
Fisher C. 2000. Advances in feed evaluation for poultry. Pp. 243-268 in P.J. Moughan, M.W.A. Verstegen and M.I. Visser-Teyneveld (Eds.) Feed Evaluation: Principles and Practice. Wageningen Pers, Wageningen, The Netherlands.
Havenstein GB , Ferket PR, Grimes JL, Qureshi MA, Nestor KE. 2007. Comparison of the performance of 1966-versus 2003-type turkeys when fed representative 1966 and 2003 turkey diets: Growth rate, livability, and feed conversion. Poultry Science 86, 232-240.
Huff GR, Huff WE, Balog JM, Rath NC, Anthony NB and Nestor KE. 2005. Stress response differences and disease susceptibility reflected by heterophil to lymphocyte ratio in turkeys selected for increased body weight. Poultry Science 84, 709-717.
Li Z, Nestor KE, Saif YM, Bacon WL and Anderson JW. 1999. Effect of selection for increased body weight on mitogenic responses in turkeys. Poultry Science 78, 1532-1535.
Leclerq B. 1986. Energy requirements of avian species. Pp. 125-139 in C. Fisher and K.N. Boorman (Eds.) Nutrient Requirements of Poultry and Nutritional Research.. Butterworth, Sevenoaks, UK.
Leeson S and Summers JD. 1997. Feeding programmes for Turkeys. Pp. 299-323 in Commercial Poultry Nutrition. University Books, Guelph, Canada.

20
Nestor KE, Noble DO, Zhu J and MoritsuY. 1996. Direct and correlated responses to long-term selection for increased body weight and egg production in turkeys. Poultry Science 75, 1180-1191.
NRC 1984. Nutrient Requirements of Poultry. 8th Revised Edition, National Academy Press, Washington, DC, USA
Roberson KD, Rahn AP, Balander RJ, Orth MW, Smith DM, Booren BL, Booren AM, Osburn WN and Fulton RM. 2003. Evaluation of the growth potential, carcass components and meat quality characteristics of three commercial strains of tom turkeys. Journal of Applied Poultry Research 2, 229-236.
21
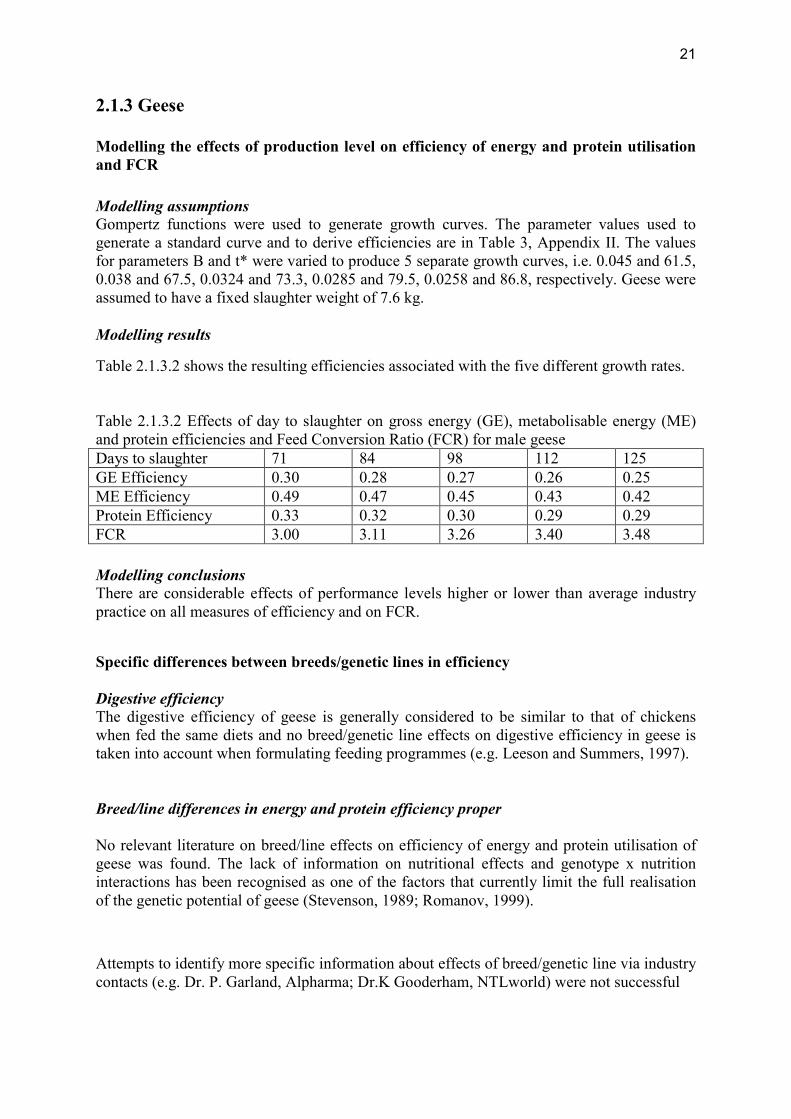
2.1.3 Geese Modelling the effects of production level on efficiency of energy and protein utilisation and FCR Modelling assumptions Gompertz functions were used to generate growth curves. The parameter values used to generate a standard curve and to derive efficiencies are in Table 3, Appendix II. The values for parameters B and t* were varied to produce 5 separate growth curves, i.e. 0.045 and 61.5, 0.038 and 67.5, 0.0324 and 73.3, 0.0285 and 79.5, 0.0258 and 86.8, respectively. Geese were assumed to have a fixed slaughter weight of 7.6 kg. Modelling results
Table 2.1.3.2 shows the resulting efficiencies associated with the five different growth rates. Table 2.1.3.2 Effects of day to slaughter on gross energy (GE), metabolisable energy (ME) and protein efficiencies and Feed Conversion Ratio (FCR) for male geese Days to slaughter 71 84 98 112 125 GE Efficiency 0.30 0.28 0.27 0.26 0.25 ME Efficiency 0.49 0.47 0.45 0.43 0.42 Protein Efficiency 0.33 0.32 0.30 0.29 0.29 FCR 3.00 3.11 3.26 3.40 3.48 Modelling conclusions There are considerable effects of performance levels higher or lower than average industry practice on all measures of efficiency and on FCR.
Specific differences between breeds/genetic lines in efficiency Digestive efficiency The digestive efficiency of geese is generally considered to be similar to that of chickens when fed the same diets and no breed/genetic line effects on digestive efficiency in geese is taken into account when formulating feeding programmes (e.g. Leeson and Summers, 1997). Breed/line differences in energy and protein efficiency proper No relevant literature on breed/line effects on efficiency of energy and protein utilisation of geese was found. The lack of information on nutritional effects and genotype x nutrition interactions has been recognised as one of the factors that currently limit the full realisation of the genetic potential of geese (Stevenson, 1989; Romanov, 1999).
Attempts to identify more specific information about effects of breed/genetic line via industry contacts (e.g. Dr. P. Garland, Alpharma; Dr.K Gooderham, NTLworld) were not successful

22
References
Leeson S and Summers JD 1997. Commercial Poultry Nutrition. University Books, Guelph, Ontaria, Canada.
Romanov, MN. 1999. Goose production efficiency as influenced by genotype, nutrition and production systems. Worlds Poultry Science Journal 55, 281-294.
Stevenson, MH. 1989. Nutrition of domestic geese. Proceedings of the Nutrition Society 48, 103-111.
23
2.1.4 Ducks
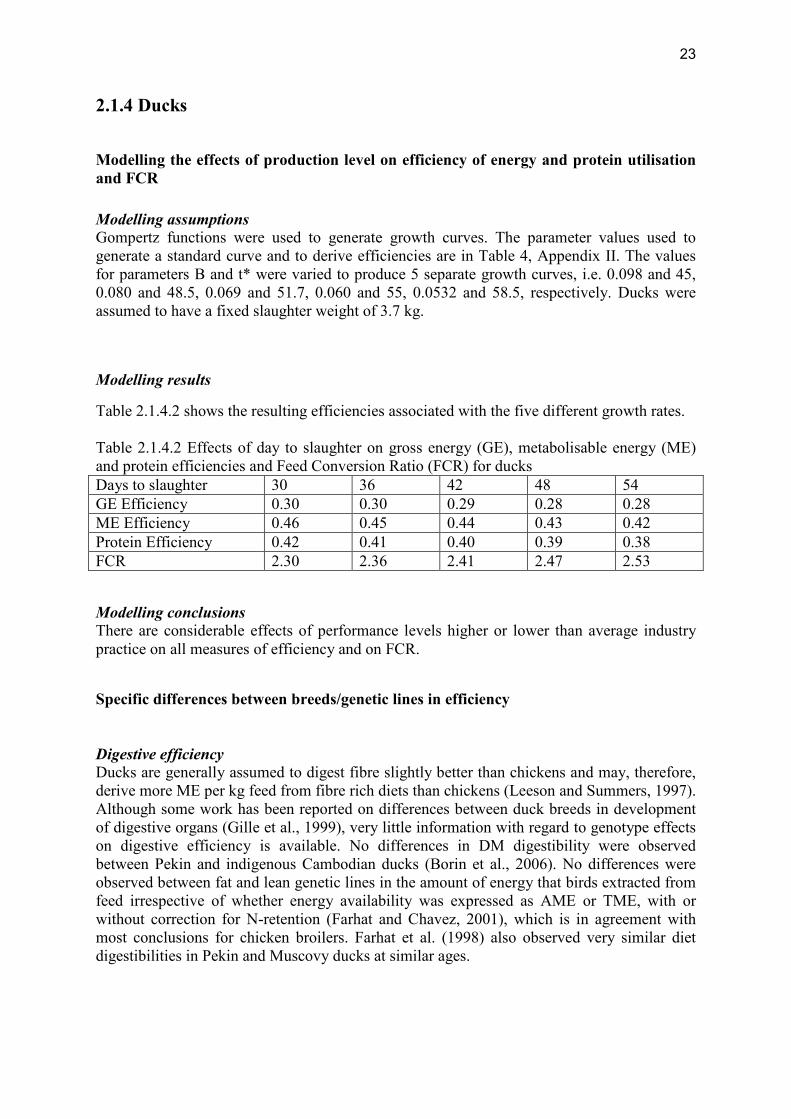
Modelling the effects of production level on efficiency of energy and protein utilisation and FCR Modelling assumptions Gompertz functions were used to generate growth curves. The parameter values used to generate a standard curve and to derive efficiencies are in Table 4, Appendix II. The values for parameters B and t* were varied to produce 5 separate growth curves, i.e. 0.098 and 45, 0.080 and 48.5, 0.069 and 51.7, 0.060 and 55, 0.0532 and 58.5, respectively. Ducks were assumed to have a fixed slaughter weight of 3.7 kg.
Modelling results
Table 2.1.4.2 shows the resulting efficiencies associated with the five different growth rates. Table 2.1.4.2 Effects of day to slaughter on gross energy (GE), metabolisable energy (ME) and protein efficiencies and Feed Conversion Ratio (FCR) for ducks Days to slaughter 30 36 42 48 54 GE Efficiency 0.30 0.30 0.29 0.28 0.28 ME Efficiency 0.46 0.45 0.44 0.43 0.42 Protein Efficiency 0.42 0.41 0.40 0.39 0.38 FCR 2.30 2.36 2.41 2.47 2.53 Modelling conclusions There are considerable effects of performance levels higher or lower than average industry practice on all measures of efficiency and on FCR.
Specific differences between breeds/genetic lines in efficiency Digestive efficiency Ducks are generally assumed to digest fibre slightly better than chickens and may, therefore, derive more ME per kg feed from fibre rich diets than chickens (Leeson and Summers, 1997). Although some work has been reported on differences between duck breeds in development of digestive organs (Gille et al., 1999), very little information with regard to genotype effects on digestive efficiency is available. No differences in DM digestibility were observed between Pekin and indigenous Cambodian ducks (Borin et al., 2006). No differences were observed between fat and lean genetic lines in the amount of energy that birds extracted from feed irrespective of whether energy availability was expressed as AME or TME, with or without correction for N-retention (Farhat and Chavez, 2001), which is in agreement with most conclusions for chicken broilers. Farhat et al. (1998) also observed very similar diet digestibilities in Pekin and Muscovy ducks at similar ages.

24
Maintenance requirements No information was found on possible differences between duck genotypes in maintenance requirements and no specific allowances for specific breeds are made in the literature (e.g. Leeson and Summers, 1997). Efficiency of energy and protein utilisation No information was found on possible differences between duck genotypes on efficiency of ME utilisation. Farhat and Chavez (2001) report a higher efficiency of protein retention in lean compared to fat genetic lines, which is, however, to be expected as both lines were fed the same diet. The lack of information on effects of breed/genetic line on digestive efficiency, maintenance requirements and efficiency of energy and protein utilisation proper is recognised in the duck breeding industry (Dr. David Martin, Cherry Valley, personal communication) FCR Feed Conversion Ratio has been used as a direct selection trait in ducks (Klemm and Pingel, 1992) because FCR can be decreased in ducks by selection, as summarised by Farhat and Chavez (2001). For the Pekin duck h² varied from 0.21 (Clayton and Powell, 1979; Powell, 1984) to 0.52 (Klemm et al., 1994). This ratio increases strongly with the age of animals. For a female mule duck, feed conversion ratio ranges from 2.12 between zero and forty-two days of age, to 4.43 between zero and twelve weeks of age (Guy et al., 1998). Some studies have compared feed efficiency of mule ducks and their parental strains. For growing ducks (based on slaughter age) the male Pekin had the best feed conversion ratio, because, as it is earlier maturing, it is slaughtered younger. Retailleau (1999) estimated the feed conversion ratio at 2.5 and 2.9 for Pekin males and Muscovy males measured between 0 and 49 days of age and between 0 and 84 days of age respectively. When Pekin, Muscovy and mule ducks are measured at the same age, mule ducks have intermediate feed efficiency compared to their parental strains (Guy et al., 1999; Baéza et al., 2005). Although there have been no studies on the genetic variability of mule duck feed efficiency, it seems possible, by selection based on the feed conversion ratio in the common strain (Klemm, 1995) to reduce the feed conversion ratio of mule duck progenies (Larzul et al., 2004). As pointed out by Marie-Etancelin (2008), many of the between-genotype comparisons of FCR in ducks are affected by differences in weight and/or maturity (and consequently body composition) at slaughter. Observed differences in FCR are, therefore, not generally indicative of differences between genotypes in efficiency of energy or protein utilisation proper. References Baéza E, Rideau N, Chartrin P, Davail S, Hoo-Paris R, Mourot J, Guy G, Bernadet MD,
Hermier D. 2005. Canards de Barbarie, Pékin et leurs croisements: aptitude à l'engraissement. Productions Animales 18, 131-141.
Borin K, Lindberg JE, Ogle RB. 2006. Digestibility and digestive organ development in indigenous and improved chickens and ducks fed diets with increasing inclusion levels of cassava leaf meal
Clayton GA and Powell JC. 1979. Growth, food conversion, carcass yields and their heritabilities in ducks (Anas platyrhynchos). British Poultry Science 20, 121-127.
Farhat A, Normand L, Chavez ER, Touchburn SP. 1998. Nutrient digestibility in food waste ingredients for Pekin and Muscovy ducks. Poultry Science 77, 1371-1376.

25
Farhat A. and Chavez ER. 2001. Metabolic studies on lean and fat ducks selected for breast muscle thickness measured by ultrasound scanning. Poultry Science 80, 585-591,
Gille U, Salomon FV, Ronnert J. 1999. Growth of the digestive organs in ducks with considerations on their growth in birds in general. British Poultry Science 40, 194–202
Guy G, Rousselot-Pailley D and Rouvier R. 1998. Description de la croissance de canettes mulardes. Annales de Zootechnie 45, 303-310.
Guy G, Hermier D, Davail S, Bely M, Andre JP and Hoo-Paris R. 1999. Meat production and force-feeding ability of different type of ducks. Proceedings of the 1st World Waterfowl Conference, Taichung, Taiwan, pp 462-468.
Klemm R, Reiter K and Pingel H. 1994. Results and effects of direct selection for feed efficiency in the domestic ducks: 2. Report: Methodical aspects. Archive für Geflügelkunde 58, 176-181.
Klemm, R. 1995. Selection on feed conversion ratio in ducks. Proceedings of the 10th European Symposium on Waterfowl, Halle, Germany, pp. 433-438
Klemm R. and Pingel H. 1992. Results of divergent selection for either low or high feed conversion in duck over six generations. Archives of Animal Breeding 30, 103-110.
Larzul C, Guy G. Bernadet MD. 2004. Feed efficiency, growth and carcass traits in female mule ducks. Archiv fur Geflugelkunde 68, 265-268.
Leeson S. and Summers, J.D.1997. Commercial Poultry Nutrition. University Books, Guelph, Ontaria, Canada.
Marie-Etancelin C, Chapuis H, Brun JM, Larzul C, Mialon-Richard MM and Rouvier R. 2008. Genetics and selection of mule ducks in France: a review. Worlds Poultry Science Journal 64, 187-207.
Powell JC. 1984. Selection for improved feed conversion ratio in the domestic ducks. Proceedings of the17th World Poultry Congress, Helsinki, pp. 108-109.
Retailleau B. 1999. Comparison of the growth and body composition of 3 types of ducks: Pekin, Muscovy and mule. Proceedings of the 1st World Conference on Waterfowl, Taichung, Taiwan, pp. 597-602.
26
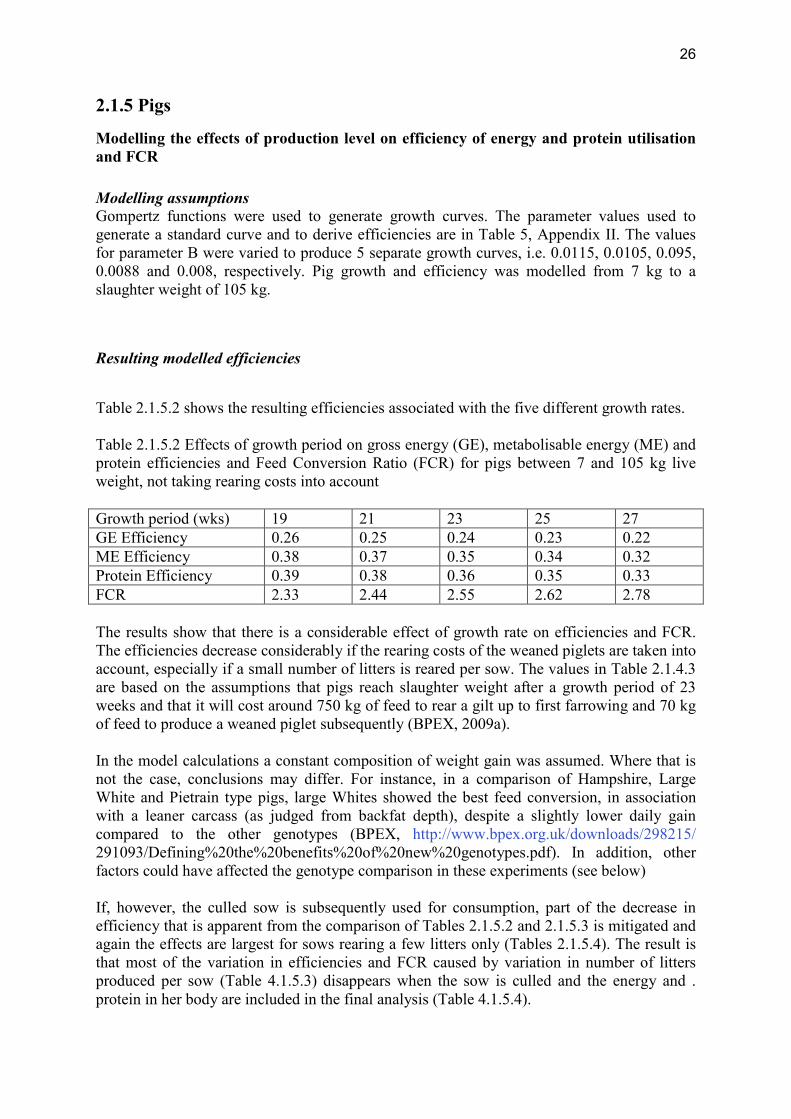
2.1.5 Pigs
Modelling the effects of production level on efficiency of energy and protein utilisation and FCR Modelling assumptions Gompertz functions were used to generate growth curves. The parameter values used to generate a standard curve and to derive efficiencies are in Table 5, Appendix II. The values for parameter B were varied to produce 5 separate growth curves, i.e. 0.0115, 0.0105, 0.095, 0.0088 and 0.008, respectively. Pig growth and efficiency was modelled from 7 kg to a slaughter weight of 105 kg.
Resulting modelled efficiencies
Table 2.1.5.2 shows the resulting efficiencies associated with the five different growth rates. Table 2.1.5.2 Effects of growth period on gross energy (GE), metabolisable energy (ME) and protein efficiencies and Feed Conversion Ratio (FCR) for pigs between 7 and 105 kg live weight, not taking rearing costs into account Growth period (wks) 19 21 23 25 27 GE Efficiency 0.26 0.25 0.24 0.23 0.22 ME Efficiency 0.38 0.37 0.35 0.34 0.32 Protein Efficiency 0.39 0.38 0.36 0.35 0.33 FCR 2.33 2.44 2.55 2.62 2.78 The results show that there is a considerable effect of growth rate on efficiencies and FCR. The efficiencies decrease considerably if the rearing costs of the weaned piglets are taken into account, especially if a small number of litters is reared per sow. The values in Table 2.1.4.3 are based on the assumptions that pigs reach slaughter weight after a growth period of 23 weeks and that it will cost around 750 kg of feed to rear a gilt up to first farrowing and 70 kg of feed to produce a weaned piglet subsequently (BPEX, 2009a). In the model calculations a constant composition of weight gain was assumed. Where that is not the case, conclusions may differ. For instance, in a comparison of Hampshire, Large White and Pietrain type pigs, large Whites showed the best feed conversion, in association with a leaner carcass (as judged from backfat depth), despite a slightly lower daily gain compared to the other genotypes (BPEX, http://www.bpex.org.uk/downloads/298215/ 291093/Defining%20the%20benefits%20of%20new%20genotypes.pdf). In addition, other factors could have affected the genotype comparison in these experiments (see below) If, however, the culled sow is subsequently used for consumption, part of the decrease in efficiency that is apparent from the comparison of Tables 2.1.5.2 and 2.1.5.3 is mitigated and again the effects are largest for sows rearing a few litters only (Tables 2.1.5.4). The result is that most of the variation in efficiencies and FCR caused by variation in number of litters produced per sow (Table 4.1.5.3) disappears when the sow is culled and the energy and . protein in her body are included in the final analysis (Table 4.1.5.4).

27
Table 2.1.5.3 Effects on efficiency of number of litters produced per sow (assuming 10 piglets per litter and 2.2 litters/sow/year), which includes the rearing costs of the sow but not the value of the culled sow.
No of litters/sow Piglets total GE eff. ME eff. Prot. eff FCR 2 20 0.20 0.30 0.27 3.33 3 30 0.21 0.31 0.28 3.25 4 40 0.21 0.32 0.28 3.21 5 50 0.21 0.32 0.29 3.18 6 60 0.21 0.33 0.29 3.17 It is evident that variation around the number of litters reared per sow per year (assumed to be 2.2 in the above calculations) and the average number of pigs reared per litter (assumed to be 10 in the above calculations) will result in additional variation in efficiency estimates. Table 2.1.5.4: Effects on efficiency of number of litters produced per sow (assuming 10 piglets per litter and 2.2 litters/sow/year), which includes the rearing costs and the value of the culled sow.
No of litters/sow Piglets total GE eff. ME eff. Prot. eff FCR 2 20 0.23 0.34 0.30 3.01 3 30 0.23 0.34 0.30 3.03 4 40 0.22 0.34 0.30 3.04 5 50 0.22 0.34 0.30 3.05 6 60 0.22 0.34 0.30 3.05 Specific differences between breeds/genetic lines in efficiency Digestive efficiency
Van Wieren (2000) recorded lower organic matter digestibilities in a domestic pig (Meishan) compared to wild boar fed diets of acorns or forage. In contrast, Hodgkinson et al. (2008) recorded the same energy digestibility for maize and oats, but a lower digestibility in wild boar compared with domestic pigs (a Landrace x Large White cross) on a high forage (alfalfa) diet.
Kemp et al. (1991) recorded significantly higher digestive efficiency for crude fibre in Meishan compared to Landrace pigs fed a standard diet but this was much reduced when pigs were fed a high fibre diet. In contrast (Fevrier et al., 1992), Meishan showed better fibre digestibilities than Large Whites only when they were fed very high fibre diets (in combination with high protein contents), not on a low fibre diet. Yen et al., (2004) concluded that the frequently assumed superior digestive efficiency of Meishan compared to more modern Western genotypes was not confirmed in their comparison with a Duroc x Large White cross in which the Meishan had significantly lower apparent digestibility coefficients for DM, N, NDF and hemicellulose.
No differences in digestive efficiency for energy or nutrients were observed in a comparison of Large White and Creole pigs (Reneaudeau, 2006). Morel et al. (2006) recorded higher

28
energy digestibilities in local Kune Kune pigs than in Landrace x Yorkshire crosses fed a high fibre diet but no differences for more standard (low fibre) diets. Large White pigs tended to digest fibre in pig diets less efficiently than the local Mukota breed and its cross with Large White, especially on high-fibre diets (Kanengoni et al., 2002). Higher whole tract digestibility in Large White than in Iberian pigs was observed by Morales et al. (2002), but this was associated with considerably higher feed intakes by the Iberian pigs. Lower energy and nutrient digestibilities were recorded in Yorkshire pigs compared with the local Mong Cai breed and its cross with Yorkshire (Len et al., 2007). Subsequent research showed that the ileal digestibilities were not different between breeds but total tract digestibilities were (Len et al. 2009a). This suggests that differences in total tract digestibilities were caused by variation in lower tract fermentation, which could be related to a slightly faster development of the gastro-intestinal tract in Mong Cai pigs after they received access to solid feed (Len et al., 2009b). Freire et al. (2003) also concluded that the higher fibre digestibility recorded in Alentejano compared to Duroc*Landrace cross was due to a more extensive cell wall degradation by microflora enzymes in the lower digestive tract.
Windisch et al (2000) observed a reduction in the apparent digestibility of crude protein in Pietrain compared to Landrace pigs, although this had no effect at all on the efficiency of dietary protein utilisation for protein retention as a result of lower urinary N excretion by Pietrains. No differences in ileal digestibility of amino acids was observed between Saddlebacks and Goettingen mini-pigs (Hennig et al., 2004).
In a recent review, Savon (2005) concluded that there is conflicting evidence about differences in digestibility between pig breeds fed high fibre diets and that where differences are observed it is likely mainly due to variation in the size of (and therefore fermentation in) the lower gastro-intestinal tract. These conclusions are in good agreement with earlier reports of digestive efficiency in pigs that also concluded that differences between genotypes are mainly related to fibre digestion and large intestinal function.
Although Fevrier et al. (1992) concluded that there were no differences in the activity of pancreatic enzymes and intestinal disaccharidases between Meishan and Large White pigs, some genotype effects on activities of digestive enzymes (such as maltase and sucrose) have been recorded (Bruininx et al., 2002).
In conclusion, slightly higher total tract digestibilities are frequently (but not universally) recorded in local breeds when compared to modern international pig breeds. In many studies, these differences are most prominent on high fibre diets and disappear when pigs are given access to high quality diets. In some studies this is related to a lower digestive tract that is larger or develops earlier in local compared to modern international breeds, which suggest that observed differences will be related mainly to differences in diet fermentation rather than enzymatic digestion, although information on the latter subject is very scarce. There is no evidence to suggest that in modern pig genotypes variation in digestive efficiency is contributing significantly to variation in efficiency of energy and protein utilisation in the production system as a whole.
Maintenance requirements
Comparisons of maintenance requirements of pigs with different weights and of different genotypes are affected strongly by the manner in which requirements are expressed, as shown by Noblet et al. (1999). Considerable differences in energy maintenance requirements

29
(measured as Fasting Heat Production, FHP) per unit metabolic size (i.e. live weight raised to the power 0.55 or 0.6) have been recorded between Meishan and some other breeds (Large White and Pietrain) (Noblet et al., 1998; van Milgen et al., 1998, 1999). Noblet et al. (1998) conclude that Meishan pigs (a fat genotype) have around 12% lower maintenance requirements, expressed in this way, than the leaner genotypes. Analyses that have taken differences in body composition between breeds into account showed, however, that these between-breed differences disappeared when FHP was regressed on muscle, viscera and fat mass (van Milgen 1998). The negative effect of fat mass in the developed equation was, however, mainly caused by the presence of the Meishan breed in the comparison and van Milgen et al. (1999) speculate that this could have masked real lower energy maintenance requirements for the Meishan breed.
Also Lepron et al. (2007) found lower requirements (as measured via residual ME intake, RMEI) of Meishan compared to Large White and a synthetic breed but expressed some doubt about the accuracy of their estimates. They could, however, link part of the between-genotype variation in RMEI to variation in behaviour and showed that variation in RMEI was caused by variation in estimated maintenance requirements as a result of variation in, for instance, standing, aggressive and locomotory behaviour. Also the work of Henken et al (1991) suggested that differences in maintenance energy requirements between Yorkshire and several lines of Landrace pigs was directly related to the low activity level of the Yorkshires.
As discussed in more detail in the general discussion below, the data obtained with pigs as well as some other species suggest that there is very little variation in NE requirements for maintenance, as measured by resting FHP in a thermoneutral environment when actual (and mature) protein size is taken into account. These results are consistent with the findings of Emmans (1994; 1997). There can, however, be differences between breeds/genetic lines in ME for maintenance requirements that have been linked to real biological differences, the most important of which seems to be the level of animal activity.
Efficiency of utilisation of energy and protein proper
It is generally assumed that the energetic efficiency of lipid and protein deposition is independent of pig weight and genotype for modelling purposes (e.g. Whittemore 2006). Estimation of these efficiencies in different breeds is, however, plagued with a considerable number of methodological issues (e.g. the problem how to properly account for maintenance requirements, as discussed above), which makes the accurate measurements of such efficiencies very difficult. There are, therefore, at present no good arguments for the assumption that variation between breeds/genetic lines of pigs in efficiency of energy utilisation for lipid and protein deposition is (i) present, (ii) can be measured accurately and easily and, therefore (iii) could be a suitable trait for inclusion in pig breeding programmes as part of attempts to improve efficiency of energy and protein utilisation in modern pig production systems.
Comparisons of contrasting genotypes (e.g. Meishan and Large White x Landrace crosses or Landrace versus Pietrain) have concluded that a limiting protein supply is utilised with the same efficiency for protein retention (Kyriazakis et al., 1994; 1995; Susenbeth et al., 1999) and that, therefore, no major genotype effects on this efficiency can be expected for pigs. Lobley et al. (1999) agreed that there is little evidence for genotype effects on efficiency of protein utilisation at the whole body level but also suggest that methodological issues might be a problem here.

30
Other factors
The occurrence of disease can have a major effect on nutrient efficiency and there are suggestions for important genotype x environment interactions in this regard. Although at present hard data for pigs are not available (S. Bishop, Roslin Institute; personal communication), several strands of evidence suggest that increased disease resistance in pigs may be associated with higher than expected requirements for energy and protein, as has been recorded for sheep (see chapter 2.1.6). For instance, BPEX experiments have suggested that the relatively high feed intake of Hampshire type, compared to Large White or Pietrain types, could not be explained entirely by the relatively high fat level in their carcasses; instead, it could be related to higher costs of their immune system since Hampshire type pigs also showed lower rates of mortality/removal from trial than the other genotypes (BPEX, 2009b; G. Walling, JSR; personal communication). Clapperton et al. (2005, 2006, 2009) showed that higher Acute Phase Proteins, used to diagnose the health status of populations of farm animals, and white blood cell numbers were correlated with pig performance and that the heritability and associations with performance were affected by housing conditions. At present the causality of such relations has not been established (Clapperton et al., 2009) and the possible energy and nutrient costs of increased disease resistance have not been systematically investigated in pigs. This forms, therefore, a major gap in our knowledge of factors that affect nutrient efficiency of different pig genotypes in a variety of (disease) environments (G. Walling, JSR and S. Bishop, Roslin Institute; personal communication).
References
BPEX. 2009. Pig Yearbook 2009. BPEX, Milton Keynes, UK. BPEX. 2009b. Defining the benefits of new genotypes. BPEX, Milton Keynes, UK. Bruininx EMAM, Schellingerhout AB, Lensen EGC, van der Peet-Schwering CMC, Schrama
JW, Everts H, den Hartog LA, Beynen AC. 2002. Associations between individual food intake characteristics and indicators of gut physiology of group-housed weanling pigs differing in genotype. Animal Science 75, 103-113.
Clapperton, M, Bishop SC, Cameron ND and Glass EJ. 2005. Associations of acute phase protein levels with growth performance and with selection for growth performance in Large White pigs. Animal Science 81, 213-220.
Clapperton M, Bishop SC and Glass EJ. 2006. Selection for lean growth and food intake leads to correlated changes in innate immune traits in Large White pigs. Animal Science 82, 867-876.
Clapperton M, Diack AB, Matika O, Glass EJ, Gladney CD, Mellencamp MA, Hoste A and Bishop SC. 2009. Traits associated with innate and adaptive immunity in pigs: heritability and associations with performance under different her health status. Genetics Selection Evolution 41, 54-65.
Emmans GC. 1994. Effective energy – a concept of energy utilization applied across species. British Journal of Nutrition 71, 801-821
Emmans GC. 1997. A method to predict the food intake of domestic animals from birth to maturity as a function of time. Journal of Theoretical Biology 186, 189-199.
Fevrier C, Bourdon D, Aumaitre A. 1992. Effects of level of dietary fiber from wheat bran on digestibility of nutrients, digestive enzymes and performance in the European Large White and Chinese Mei Shan pig. Journal of Animal Physiology and Animal Nutrition 68, 60-72.

31
Flock DK, Laughlin KF and Bentley J. 2005. Minimizing losses in poultry breeding and production: how breeding companies contribute to poultry welfare. Worlds Poultry Science Journal 61, 227-237.
Freire JPB, Dias RIM, Cunha LF and Aumaitre A. 2003. The effect of genotype and dietary fibre level on the caecal bacterial enzyme activity of young piglets: digestive consequences. Animal Feed Science and Technology 106, 119-130.
Henken AM, Vanderhel W, Brandsma HA and Verstegen MWA. 1991. Differences in Energy-Metabolism and Protein Retention of Limit-Fed Growing Pigs of Several Breeds. Journal of Animal Science 69, 14431453.
Hennig U, Metges CC, Berk A., Tuchscherer A. and Kwella M. 2004. Relative ileal amino acid flows and microbial counts in intestinal effluents of Goettingen Minipigs and Saddleback pigs are not different. Journal of Animal Science 82, 1976-1985.
Hodgkinson SM, Schmidt M, Ulloa N. 2008. Comparison of the digestible energy content of maize, oats and alfalfa between the European wild boar (Sus scrofa L.) and Landrace x Large White pig (Sus scrofa domesticus). Animal Feed Science and Technology 144, 167-173.
Kanengoni AT, Dzama K, Chimonyo M, Kusina J, Maswaure SM. 2002. Influence of level of maize cob meal on nutrient digestibility and nitrogen balance in Large White, Mukota and LW x M F-1 crossbred pigs. Animal Science 74, 127-134.
Kemp B, Den Hartog LA, Klok JJ, Zandstra T. 1991. The digestibility of nutrients, energy and nitrogen in the Meishan and Dutch Landrace pig. Journal of Animal Physiology and Animal Nutrition 65, 263-266.
Kyriazakis I, Dotas D, Emmans GC.1994. The effect of breed on the relationship between feed composition and the efficiency of protein-utilization in pigs. British Journal of Nutrition 71, 849-859.
Kyriazakis I, Emmans GC, Anderson D. 1995. Do breeds of pig differ in the efficiency with which they use a limiting protein supply? British Journal of Nutrition 74, 183-195.
Len NT, Lindberg JE and Ogle B. 2007. Digestibility and nitrogen retention of diets containing different levels of fibre in local (Mong Cai), F1 (Mong Cai x Yorkshire) and exotic (Landrace x Yorkshire) growing pigs in Vietnam. Journal of Animal Physiology and Animal Nutrition 91, 297-303.
Len NT, Hong TTT, Ogle B, Lindberg JE. 2009a. Comparison of total tract digestibility, development of visceral organs and digestive tract of Mong Cai and Yorkshire x Landrace piglets fed diets with different fibre sources. Journal of Animal Physiology and Animal Nutrition 93, 181-191.
Len NT, Ngoc TB, Ogle B, Lindberg JE. 2009b. Ileal and total tract digestibility in local (Mong Cai) and exotic (Landrace x Yorkshire) piglets fed low and high-fibre diets, with or without enzyme supplementation. Livestock Science 126, 73-79.
Lepron E, Bergeron R, Robert S, Faucitano L, Bernier JF, Pomar C. 2007. Relationship between residual energy intake and the behaviour of growing pigs from three genetic lines. Livestock Science 111, 104-113.
Morales J, Perez JF, Baucells MD, Mourot J and Gasa J. 2002. Comparative digestibility and lipogenic activity in Landrace and Iberian finishing pigs fed ad libitum corn- and corn-sorghum-acorn-based diets. Livestock Production Science 77, 195-205.
Morel PCH, Lee TS and Moughan PJ. 2006. Effect of feeding level, live weight and genotype on the apparent faecal digestibility of energy and organic matter in the growing pig. Animal Feed Science and Technology 126, 63-74.
Noblet J, Bernier JF, Dubois S, Le Cozler Y, van Milgen J. 1998. Effect of breed and body weight on components of heat production in growing pigs. Pp. 225-228 in

32
(McCracken KJ; Unsworth EF; Wylie ARG, Eds.) Energy Metabolism of Farm Animals. CABI Publishing, Wallingford, UK.
Noblet J, Karege C, Dubois S and van Milgen J. 1999. Metabolic utilization of energy and maintenance requirements in growing pigs: Effects of sex and genotype. Journal of Animal Science 77, 1208-1216.
Qin GX, Verstegen MWA and Bosch MW. 1995. Variation of digestive capacity between genetically different pig populations: a review. Journal of Animal Physiology and Animal Nutrition 73, 233-242.
Renaudeau D, Bocage B, Noblet J. 2006. Influence of energy intake on protein and lipid deposition in Creole and Large White growing pigs in a humid tropical climate. Animal Science 82, 937-945.
Savon L. 2005. Tropical roughages and their effect on the digestive physiology of monogastric species. Cuban Journal of Agricultural Science 39, 463-474.
Susenbeth A, Dickel T, Diekenhorst A, Hohler D. 1999. The effect of energy intake, genotype, and body weight on protein retention in pigs when dietary lysine is the first-limiting factor. Journal of Animal Science 77, 2985-2989.
van Milgen J, Bernier JF, Dubois S, Le Cozler Y, Noblet J. 1998a. Modelling dynamic aspects of heat production in pigs. Pp. 331-334 in (McCracken KJ; Unsworth EF; Wylie ARG, Eds.) Energy Metabolism of Farm Animals. CABI Publishing, Wallingford, UK.
van Milgen J, Bernier JF, Lecozler Y, Dubois S, Noblet J 1998b. Major determinants of fasting heat production and energetic cost of activity in growing pigs of different body weight and breed/castration combination. British Journal of Nutrition 79, 509-517.
Van Wieren SE. 2000. Digestibility and voluntary intake of roughages by wild boar and Meishan pigs. Animal Science 71, 149-156.
Whittemore CT. 2006. Energy value of feedstuffs for pigs. Pp. 317-342 in (I Kyriazakis and CT Whittemore, eds.) Whittemore’s Science and Practice of Pig Production. Blackwell Publishing.
Windisch W, Gotterbarm GG, Roth FX, Schams D, Kirchgessner M. 2000. Effect of genetic provenience of pigs (German Landrace, Pietrain) on parameter of protein metabolism. Züchtungskunde 72, 379-388.
Yen JT, Varel VH, Nienaber JA. 2004. Metabolic and microbial responses in western crossbred and Meishan growing pigs fed a high-fiber diet. Journal of Animal Science 82, 1740-1755
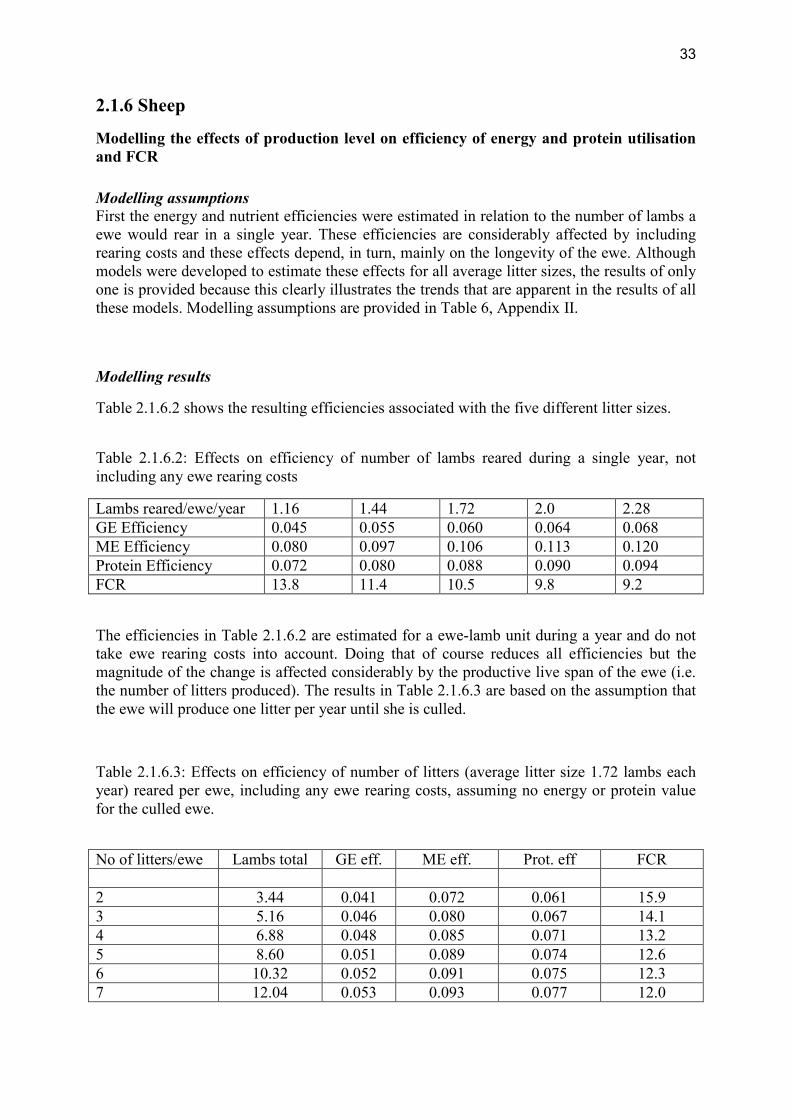
33
2.1.6 Sheep
Modelling the effects of production level on efficiency of energy and protein utilisation and FCR Modelling assumptions First the energy and nutrient efficiencies were estimated in relation to the number of lambs a ewe would rear in a single year. These efficiencies are considerably affected by including rearing costs and these effects depend, in turn, mainly on the longevity of the ewe. Although models were developed to estimate these effects for all average litter sizes, the results of only one is provided because this clearly illustrates the trends that are apparent in the results of all these models. Modelling assumptions are provided in Table 6, Appendix II.
Modelling results
Table 2.1.6.2 shows the resulting efficiencies associated with the five different litter sizes.
Table 2.1.6.2: Effects on efficiency of number of lambs reared during a single year, not including any ewe rearing costs
Lambs reared/ewe/year 1.16 1.44 1.72 2.0 2.28 GE Efficiency 0.045 0.055 0.060 0.064 0.068 ME Efficiency 0.080 0.097 0.106 0.113 0.120 Protein Efficiency 0.072 0.080 0.088 0.090 0.094 FCR 13.8 11.4 10.5 9.8 9.2
The efficiencies in Table 2.1.6.2 are estimated for a ewe-lamb unit during a year and do not take ewe rearing costs into account. Doing that of course reduces all efficiencies but the magnitude of the change is affected considerably by the productive live span of the ewe (i.e. the number of litters produced). The results in Table 2.1.6.3 are based on the assumption that the ewe will produce one litter per year until she is culled.
Table 2.1.6.3: Effects on efficiency of number of litters (average litter size 1.72 lambs each year) reared per ewe, including any ewe rearing costs, assuming no energy or protein value for the culled ewe.
No of litters/ewe Lambs total GE eff. ME eff. Prot. eff FCR 2 3.44 0.041 0.072 0.061 15.9 3 5.16 0.046 0.080 0.067 14.1 4 6.88 0.048 0.085 0.071 13.2 5 8.60 0.051 0.089 0.074 12.6 6 10.32 0.052 0.091 0.075 12.3 7 12.04 0.053 0.093 0.077 12.0

34
The efficiencies as estimated in Table 2.1.6.3 are correct only, however, if the ewe dies or is otherwise destroyed. Since the majority of ewes will be culled and end up in the food chain, the energy and protein value of the ewe has to be taken into account for a correct estimation of energy and protein efficiency of the system as a whole. Table 2.1.6.4 shows that the considerable effects of productive life span on efficiency (as suggested by Table 2.1.6.3) largely disappear when the energy and protein content of the culled ewe is taken into account.
Table 2.1.6.4: Effects on efficiency of number of litters (average litter size 1.72 lambs each year) reared per ewe, including ewe rearing costs and the energy and protein value of the culled ewe.
No of litters/ewe Lambs total GE eff. ME eff. Prot. eff FCR 2 3.44 0.058 0.102 0.084 10.3 3 5.16 0.059 0.103 0.085 10.3 4 6.88 0.059 0.104 0.085 10.4 5 8.60 0.059 0.104 0.085 10.4 6 10.32 0.059 0.104 0.085 10.4 7 12.04 0.060 0.104 0.085 10.4 Specific differences between breeds/genetic lines in efficiency Digestive efficiency
Tests of breed or genetic line effects on digestive efficiency of sheep have shown either no (e.g. Mann et al., 1987; Ranilla et al., 1998; Molina et al., 2001; Kiani et al., 2004; Kamalzadeh and Aouladrabiei, 2009) or statistically significant but small effects (e.g. Givens and Moss, 1994; Lourenco et al., 2000; Lopez et al., 2001; Wildeus et al., 2007), although the latter effects were sometimes confounded by variation in intake level.
Variation between individual sheep in methane release have been recorded as early as 1965 (Blaxter and Clapperton, 1965) and Pinares-Patino et al. (2003b) suggested that such differences were consistent over time in grazing sheep, perhaps determined mainly by variation in rumen organic matter pool (Pinares-Patino et al., 2003a). However, high within-animal variation is also frequently recorded (Pinares-Patino and Clark, 2008) and there are still many questions to be answered about the effects of animal age on methane release per kg feed consumed (Knight et al., 2008) and about employing the most proper (indirect) methodology (Pinares-Patino et al., 2008).
Maintenance requirements
Blaxter (1962) noted considerable variation between individual sheep in maintenance energy requirements as estimated from fasting heat production but these were not affected by the sheep breed. In later work, however, Blaxter et al (1966) concluded that there were breed effects on the maintenance energy requirements of sheep. It seems likely that conclusions from such comparisons are greatly affected by the method of estimation and the age/weight at which breeds are compared (similar to the methodological issues discussed for pigs). For

35
instance, Freetly et al (1995) observed that Texel sheep had lower FHP (an estimate of the NE requirements for maintenance) than Suffolk sheep when compared at the same weight but that Suffolk sheep had lower FHP per kg body weight than Texel sheep when compared at the same age and that all differences between these breeds in FHP per kg body weight disappeared when the two breeds were compared at the same proportion of mature size. Later work of the same group with more breeds reached a very similar conclusion (Freetly et al., 2002). Unless such differences in maturity (usually accompanied by differences in body composition) are taken into account, some scepticism to any recorded breed or genetic line effects on maintenance energy requirements is warranted. The approach taken by Emmans (1994; 1997) in which both mature and actual protein weight are used to estimate maintenance energy and protein requirements is likely to accommodate the variation observed by Freetly et al. (1995; 2002). Such an approach is also consistent with the observation that variation in estimated maintenance energy requirements following different nutritional histories in sheep disappeared when compared on the basis of lean body size (Ball et al., 1998) and with many observations recorded with pigs (see Chapter 2.1.5).
Efficiency of utilisation of energy and protein proper
It is generally assumed that efficiencies of energy and protein utilisation for maintenance and gain can be applied across ruminant species (e.g. ARC, 1980; AFRC, 1993) and in all current major energy or protein evaluation systems worldwide no breed/genotype effects on these efficiencies are included. In fact, there are proposals to use universal material efficiencies of protein and energetic efficiencies of protein and lipid deposition (assuming the same digested substrates) across all species, monogastrics as well as ruminants (Emmans, 1994; 1997).
In ruminants, large effects of feeding level on overall efficiency of ME utilisation in restrictedly fed animals are well documented (e.g. ARC, 1980; AFRC, 1993) but there is good evidence that (almost) all of this variation disappears in ad libitum fed ruminants (Tolkamp and Ketelaars, 1994; Tolkamp and Kyriazakis, 2009; Tolkamp, 2010), which is the dominant feeding regime in almost all ruminant production systems.
There is very limited research on effects of breed/genetic line on efficiency of protein utilisation. Liu et al. (2007) reported no differences in protein synthesis between genetic lines of Merino sheep selected for gastrointestinal nematode resistance and a control line.
Other factors
Disease resistance
It is possible to select sheep genotypes that show increased resistance to certain diseases, such as infection with gastro-intestinal parasites (e.g. Liu et al., 2005). There is conflicting evidence in the literature about how differences between genetic lines in disease resistance affect the efficiency of energy and protein utilisation. In some studies, selection for nematode resistance is associated with negative effects on weight gain or wool growth (e.g. Bisset et al. 2001; Eady et al. 2003), but not in others (e.g. Karlson et al. 1995; see also Liu et al. 2005), which would reduce the expected beneficial effects of nematode resistance on efficiency of energy and protein utilisation. Morris et al (2000) reported that Romney sheep selected for resistance to nematode infection had reduced weight gains and wool growth but more lambs weaned per ewe than sheep selected for susceptibility to nematode infection. In addition, Liu

36
et al (2005) found that infected nematode resistant sheep had significantly higher energy and protein requirements than control sheep when they were young (but not in older sheep).
It is evident that there will be a genotype x environment interaction in this respect because any increased resistance to a specific disease, and its possible beneficial effects on efficiency of energy and protein utilisation, will only be expressed in an environment where the disease is present and confer no benefits when it is absent. It is, therefore, not easy to estimate how differences between genetic lines in nematode resistance in general affect efficiency of energy and protein utilisation, certainly in view of the recorded higher energy and protein requirements for more resistant sheep, especially when there is an energy and protein cost to nematode resistance (Liu et al., 2005).
Recent work suggests that selection against footrot in sheep is possible (Nieuwhof et al., 2008; 2009; Conington et al., 2008) and lower footrot incidence is expected to have a beneficial effect on the efficiency of energy and protein utilisation in this system although at present these effects are difficult to quantify for lack of detailed information.
References
AFRC (Agricultural and Food Research Council). 1993. Energy and Protein Requirements of
Ruminants. An advisory manual prepared by the AFRC Technical Committee on Responses to Nutrients. CAB International, Wallingford, UK.
ARC (Agricultural Research Council). 1980. The Nutrient Requirements of Ruminant Livestock. CAB International, Wallingford, UK.
Ball AJ, Thompson JM, Alston CL, Blakely AR, Hinch GN. 1998. Changes in maintenance energy requirements of mature sheep fed at different levels of feed intake at maintenance, during weight loss and realimentation. Livestock Production Science 53, 191-204.
Bisset SA, Morris CA, McEwan JC, Vlassoff A 2001. Breeding sheep in New Zealand that are less reliant on anthelmintics to maintain health and productivity. New Zealand Veterinary Journal 49, 236–246.
Blaxter KL. 1962. The fasting metabolism, of adult wether sheep. British Journal of Nutrition 16, 615-626.
Blaxter KL, Clapperton JL and Wainman FW. 1966. The extent of differences between six British sheep breeds in their metabolism, feed intake and utilization and resistance to stress. British Journal of Nutrition 20, 283-294.
Conington J, Hosie B, Nieuwhof GJ, Bishop SC 2008. Breeding for resistance to footrot - the use of hoof lesion scoring to quantify footrot in sheep. Veterinary Research Communications 32, 583-589.
Emmans GC. 1994. Effective energy – a concept of energy utilization applied across species. British Journal of Nutrition 71, 801-821
Emmans GC. 1997. A method to predict the food intake of domestic animals from birth to maturity as a function of time. Journal of Theoretical Biology 186, 189-199.
Freetly HC, Nienaber JA, Leymaster KA and Jenkins TG. 1995. Relationships among heat-production, body-weight, and age in Suffolk and Texel ewes. Journal of Animal Science 73, 1030-1037.

37
Eady SJ, Woolaston RR, Barger IA 2003. Comparison of genetic and nongenetic strategies for control of gastrointestinal nematodes of sheep. Livestock Production Science 81, 11 –23.
Freetly HC, Nienaber JA, Brown-Brandl T. 2002. Relationships among heat production, body weight, and age in Finnsheep and Rambouillet ewes. Journal of Animal Science 80, 825-832..
Givens DI and Moss AR. 1994. Effect of breed, age and bodyweight of sheep on the measurement of apparent digestibility of dried grass. Animal Feed Science and Technology 49, 155-162.
Kamalzadeh A and Aouladrabiei MR. 2009. Effects of restricted feeding on intake, digestion, nitrogen balance and metabolizable energy in small and large body sized sheep breeds. Asian-Australasian Journal of Animal Science 22, 667-673.
Karlsson LJE, Greeff JC, Harris JF 1995. Breeding sheep for worm resistance. Journal of. Agricultural Science Western Australia 2, 56–59.
Kiani A, Chwalibog A, Nielsen MO and Tauson AH. 2004. Effect of genetic line on energy and nitrogen metabolism during late gestation in twin-pregnant ewes fed restrictedly. Journal of Animal and Feed Science 13, 417-420.
Knight TW, Molano G, Clark H and Cavanagh A. 2008. Methane emissions from weaned lambs measured at 13, 17, 25 and 35 weeks of age compared with mature ewes consuming a fresh forage diet. Australian Journal of Experimental Agriculture 48, 240-243.
Liu SM, Smith TL, Briegel J, Gao SB, Peng WK. 2007. Fractional protein synthesis rate and polyamine concentrations in tissues of Merino sheep selected for gastrointestinal nematode resistance. Livestock Science 106, 65–75.
Liu S, Smith TL, Briegel J, Murray A, Masters DG, Karlsson LJE, Palmer DG, Greeff JC, Besier RB, Gao SB. 2005. Comparing productive performance of nematode resistant Merino sheep with non-selected control. Livestock Production Science 97, 117– 129.
Liu SM, Smith TL, Karlsson LJE, Palmer DG, Besier RB. 2005. The costs for protein and energy requirements by nematode infection and resistance in Merino sheep. Livestock Production Science 97, 131– 139.
Lopez S, Frutos P, Mantecon AR, Giraldez FJ. 2001. Comparative digestion of herbage by two breeds of sheep: effects of grass maturity stage and level of intake. Animal Science 73, 513-522.
Lourenco ALG, Dias-da-Silva AA, Fonseca AJM, Azevedo JT. 2000. Effects of live weight, maturity and genotype of sheep fed a hay-based diet, on intake, digestion and live weight gain. Livestock Production Science 63, 291-296.
Mann DL, Goode L, Pond KR. 1987. Voluntary intake, digestibility, rate of passage and gastro-intestinal tract fill in tropical and temperate breeds of sheep. Journal of Animal Science 64, 880-886.
Molina E, Ferret A, Caja G, Calsamiglia S, Such X and Gasa J. 2001. Comparison of voluntary food intake, apparent digestibility, digesta kinetics and digestive tract content in Manchega and Lacaune dairy sheep in late pregnancy and early and mid lactation. Animal Science 72, 209-221.
Morris CA, Vlassoff A, Bisset SA, Baker RL, Watson, TG, West CJ, Wheeler, M. 2000. Continued selection of Romney sheep for resistance or susceptibility to nematode infection: estimates of direct and correlated responses. Animal Science 70, 17-27.
Nieuwhof GJ, Conington J, Bishop SC. 2009. A genetic epidemiological model to describe resistance to an endemic bacterial disease in livestock: application to footrot in sheep. Genetics Selection Evolution 41, 1-12.

38
Nieuwhof GJ, Conington J, Bunger L, Haresign W, Bishop SC. 2008. Genetic and phenotypic aspects of foot lesion scores in sheep of different breeds and ages. Animal 2, 1289-1296
Pinares-Patino CS, Ulyatt MJ,, Lassey KR, Barry TN, Holmes CW. 2003a. Rumen function and digestion parameters associated with differences between sheep in methane emissions when fed chaffed lucerne hay. Journal of Agricultural Science 140, 205-214.
Pinares-Patino CS, Ulyatt MJ, Lassey KR, Barry TN, Holmes CW. 2003b. Persistence of differences between sheep in methane emission under generous grazing conditions. Journal of Agricultural Science 140, 227-233.
Pinares-Patino CS, Ulyatt MJ. Lassey KR, Barry TN, Holmes CW. 2003a. Rumen function and digestion parameters associated with differences between sheep in methane emissions when fed chaffed lucerne hay. Journal of Agricultural Science 140, 205-214.
Pinares-Patino CS and Clark CS. 2008. Reliability of the sulphur hexafluoride tracer technique for methane emission measurement from individual animals: an overview Australian Journal of Experimental Agriculture 48, 223-229.
Pinares-Patino CS, Holmes CW, Lassey KR, Ulyatt MJ. 2008. Measurement of methane emission from sheep by the sulphur hexafluoride tracer technique and by the calorimetric chamber: failure and success. Animal 2, 141-148.
Ranilla MJ, Lopez S, Giraldez FJ, Valdes C, Carro MD. 1998. Comparative digestibility and digesta flow kinetics in two breeds of sheep. Animal Science 66, 389-396.
Tolkamp BJ. 2010. Efficiency of energy utilisation and voluntary feed intake in ruminants. Animal, in press.
Tolkamp BJ and Ketelaars JJMH. 1994. Efficiency of energy utilisation in cattle given food ad libitum: predictions according to the ARC system and practical consequences. Animal Production 59, 43-47.
Tolkamp BJ and Kyriazakis I. 2009. Toward a new practical energy evaluation system for dairy cows. Animal 3, 307-314.
Wildeus S, Turner KE and Collins JR. 2007. Growth, intake, diet digestibility, and nitrogen use in three hair sheep breeds fed alfalfa hay. Small Ruminant Research 69, 221-227.
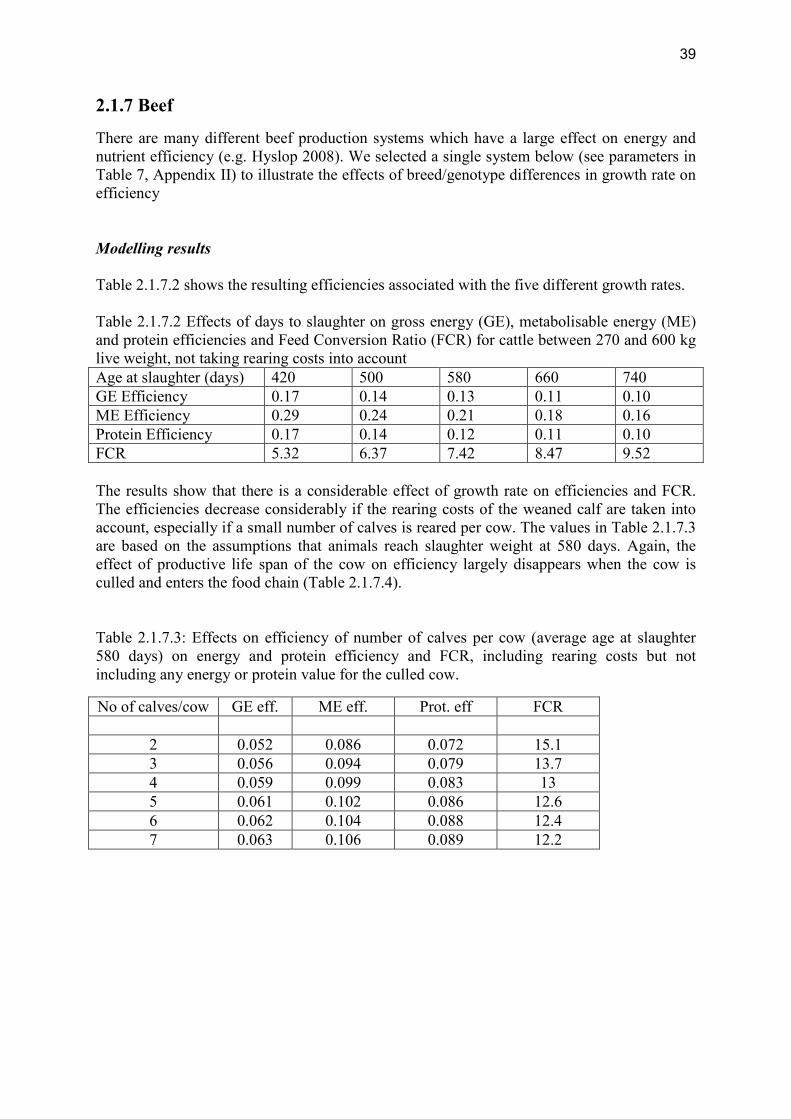
39
2.1.7 Beef
There are many different beef production systems which have a large effect on energy and nutrient efficiency (e.g. Hyslop 2008). We selected a single system below (see parameters in Table 7, Appendix II) to illustrate the effects of breed/genotype differences in growth rate on efficiency Modelling results Table 2.1.7.2 shows the resulting efficiencies associated with the five different growth rates. Table 2.1.7.2 Effects of days to slaughter on gross energy (GE), metabolisable energy (ME) and protein efficiencies and Feed Conversion Ratio (FCR) for cattle between 270 and 600 kg live weight, not taking rearing costs into account Age at slaughter (days) 420 500 580 660 740 GE Efficiency 0.17 0.14 0.13 0.11 0.10 ME Efficiency 0.29 0.24 0.21 0.18 0.16 Protein Efficiency 0.17 0.14 0.12 0.11 0.10 FCR 5.32 6.37 7.42 8.47 9.52 The results show that there is a considerable effect of growth rate on efficiencies and FCR. The efficiencies decrease considerably if the rearing costs of the weaned calf are taken into account, especially if a small number of calves is reared per cow. The values in Table 2.1.7.3 are based on the assumptions that animals reach slaughter weight at 580 days. Again, the effect of productive life span of the cow on efficiency largely disappears when the cow is culled and enters the food chain (Table 2.1.7.4). Table 2.1.7.3: Effects on efficiency of number of calves per cow (average age at slaughter 580 days) on energy and protein efficiency and FCR, including rearing costs but not including any energy or protein value for the culled cow.
No of calves/cow GE eff. ME eff. Prot. eff FCR 2 0.052 0.086 0.072 15.1 3 0.056 0.094 0.079 13.7 4 0.059 0.099 0.083 13 5 0.061 0.102 0.086 12.6 6 0.062 0.104 0.088 12.4 7 0.063 0.106 0.089 12.2

40
Table 2.1.7.4: Effects on efficiency of number of calves per cow (average age at slaughter 580 days) on energy and protein efficiency and FCR, including rearing costs and energy or protein value for the culled cow.
No of calves/cow GE eff. ME eff. Prot. eff FCR 2 0.073 0.121 0.104 10.0 3 0.072 0.120 0.103 10.3 4 0.071 0.119 0.102 10.4 5 0.071 0.119 0.101 10.5 6 0.071 0.119 0.101 10.6 7 0.071 0.118 0.101 10.6
Specific differences between breeds/genetic lines in efficiency Digestive efficiency
In the older literature, significant breed effects on digestive efficiency are frequently observed when low quality feeds were supplied to strongly contrasting beef breeds, especially in comparisons between Bos taurus and Bos indicus breeds or crosses (e.g. Howes et al. 1963; Moore et al., 1975; Warwick and Cobb, 1975). In contrast, Vercoe (1970) found no differences in energy digestibility between a Bos indicus breed and its crosses with British beef cattle breeds, although the Bos indicus did tend to release less methane than the cross-breds. No differences in diet digestibility were observed more recently by Solis et al. (1988) in a comparison of 5 breeds and their crosses that included dairy breeds (Holstein and Jerseys), beef breeds (Angus and Hereford) and a Bos indicus breed (Brahman). There were also no systematic differences in digestive efficiency between Angus and its cross with Brahmans fed a variety of diets in the study of Beaver et al. (1989) or in DM or energy digestibility between non-pregnant and non-lactating Angus and Simmental beef cows (Laurentz et al., 1991). Although some studies suggest that (small) differences in digestive efficiency exist between beef cattle genotypes with low and high Residual Feed Intakes (RFI), Herd and Arthur (2009) urge for caution to attribute much of the variation in RFI to variation in digestion because of the difficulties in reliably measuring small differences in this trait, also in view of the lack of much genetic variation in digestive efficiency in other species.
No differences between dairy and beef breeds were observed in methane production by Boadi and Wittenberg (2002) under either ad libitum or restricted feeding regimes and Ellis et al. (2007) showed that methane production of beef and dairy cows can be predicted with very similar equations based on feed quality characteristics alone. However, Hegarty et al. (2007) observed that cattle selected for lower residual feed consumption also had a reduced methane release, although variation in methane release explained only a very small proportion of the variation in residual feed consumption. Yan et al. (2009) developed predictive equations for methane emissions of beef steers of four different breeds (Friesian, Aberdeen Angus, Simmental and Charolais) on the basis of live weight and diet characteristics alone, i.e. without incorporating breed effects.

41
Maintenance requirements
No significant differences between eight beef-type cattle breeds and crosses in ME requirements for maintenance were observed by Reid et al (1991), although there was a tendency for slightly lower values for crosses with the Brahman (a Bos indicus breed). Laurentz et al. (1991) estimated slightly higher maintenance energy requirements for Simmental compared to Angus non-pregnant and non-lactating beef cows. Herd et al. (1991) observed that low maintenance energy requirements relative to size in Angus steers from lines selected for high, compared to low, yearling weight gains disappeared completely when the differences between genetic lines in mature size were taken into account. Similarly, differences in estimated maintenance energy requirements between Angus cows of these same lines disappeared when the difference in body composition was taken into account and maintenance energy requirements were related to the protein content of the animals (Herd, 1995), which is in direct agreement with the conclusions of Russel and Wright (1983) from a comparison of contrasting beef cow crosses. It also agrees well with similar observations made for pigs and sheep (see Chapters 2.1.5 and 2.1.6). A single equation for the estimation of maintenance requirements for Angus, Simmental and Charolais cross steers was derived by Dawson and Steen (1998). No effects of breed or genetic line are incorporated in the estimation of maintenance energy or protein requirements for beef cattle in ARC (1980) or AFRC (1993) but there is some evidence of relatively high maintenance requirements of double-muscled cattle, even when their high protein content is taken into account (de Campeneere et al., 2001). Montanobermudez et al (1990) observed that the maintenance requirements of Angus cross beef cows increased with the dairy merit of the breed the Angus cows were crossed with. This could, however, again be related to differences in body composition, especially protein content, between genotypes, which was not included as a variable in this study. No differences in FHP or maintenance ME requirements were recorded between different Hereford cross types by Hotovi et al. (1992) but this study, involving twins, did show the presence of a genetic component to maintenance energy metabolism and suggested that selection to change these traits might be effective. Bottje and Carstens (2008) concluded from a review of mitochondrial functions in steers selected for high or low Residual Feed Intake (RFI) that some links in the electron transport chain were more efficient in animals with low RFI than in animals with high RFI. However, the latter animals were able to carry out other mitochondrial functions as well as, or even better, than the former animals and, therefore, the overall genetic effects on energetic efficiency are presently unclear. Nevertheless, this might offer an opportunity for selecting more efficient animals in the future (Bottje and Carstens, 2008).
Efficiency of energy and protein utilisation proper
Current energy and protein evaluation systems (e.g. ARC, 1980; AFRC, 1993; NRC, 2000)) assume that efficiencies of energy and protein utilisation proper are similar in all ruminants (see discussion under sheep) and no differences between breed/genotype are incorporated.
References
AFRC (Agricultural and Food Research Council). 1993. Energy and Protein Requirements of Ruminants. An advisory manual prepared by the AFRC Technical Committee on Responses to Nutrients. CAB International, Wallingford, UK.
ARC (Agricultural Research Council). 1980. The Nutrient Requirements of Ruminant Livestock. CAB International, Wallingford, UK.

42
Beaver EE, Williams JE, Miller SJ, Hancock DL, Hannah SM and O'Connor DL. 1989. Influence of breed and diet on growth, nutrient digestibility, body composition and plasma hormones of Brangus and Angus steers. Journal of Animal Science 67, 2415-2425.
Boadi DA, Wittenberg KM. 2002. Methane production from dairy and beef heifers fed forages differing in nutrient density using the sulphur hexafluoride (SF6) tracer gas technique. Canadian Journal of Animal Science 82, 201-206.
Bottje WG and Carstens GE. 2009. Association of mitochondrial function and feed efficiency in poultry and livestock species. Journal of Animal Science 87, E48-E63.
Dawson LER and Steen R.J. 1998. Estimation of maintenance energy requirements of beef cattle and sheep. Journal of Agricultural Science, Cambridge 131, 477-485.
De Campeneere S, Fiems LO and Boucque ChV. 2001. Energy and protein requirements of double-muscled Belgian Blue bulls. Animal Feed Science and Technology 90, 153-167.
Hegarty RS, Goopy JP, Herd RM, and McCorkell B. 2007. Cattle selected for lower residual feed intake have reduced methane production. Journal of Animal Science 85, 1479-1486.
Herd RM. 1995. Effect of divergent selection for yearling growth rate on the maintenance feed requirements of mature Angus cows. Livestock Production Science 41, 39-49.
Herd RM and Arthur PF. 2009. Physiological basis for residual feed intake. Journal of Animal Science 87, E64-E71.
Herd RM, Speck PA, Wynn PC. 1991. Feed requirements for maintenance and growth of one year old Angus steers selected for either fast or slow yearling growth-rate. Australian Journal of Experimental Agriculture 31, 591-595.
Hotovy SK, Johnson KA, Johnson DE, Carstens .E, Bourdon RM, Seidel GE. 1991. Variation among twin beef-cattle in maintenance energy-requirements. Journal of Animal Science 69, 940-946.
Howes JR, Hentges JF, Davis, GK. 1963. Comparative digestive powers of Hereford and Brahman cattle, Journal of Animal Science 22, 22.
Ellis J.E., Kebreab E, Odongo NE, McBride BW, Okine EK and France J. 2007. Prediction of methane production from dairy and beef cattle. Journal of Dairy Science 90, 3456-3467.
Laurenz JC, Byers FM, Schelling GT, Greene LW. 1991. Effects of season on the maintenance requirements of mature beef-cows. Journal of Animal Science 69, 2168-2176.
Montanobermudez M, Nielsen M.K and Deutscher GH. 1990. Energy-Requirements for Maintenance of Crossbred Beef-Cattle with Different Genetic Potential for Milk. Journal of Animal Science 68, 2279-2288.
Moore RL, Essig HW, Smithson LJ. 1975. Influence of breeds of cattle on ration utilization. Journal of Animal Science 41.
National Research Council 2000. Nutrient Requirements of Beef Cattle. Update 2000 of the 7th revised edition 1996. National Academy Press, Washington DC, USA.
Russel AJF and Wright IA. 1983. Factors affecting maintenance requirements of beef cows. Animal Production 37, 329-334.
Solis JC, Byers FM, Schelling GT, Longs CR Greene LW. 1988. Maintenance requirements and energetic efficiency of cows of different breed types. Journal of Animal Science 66, 764-773.
Vercoe JE. 1970. Fasting metabolism and heat increment of feeding in Brahman x British cross cattle. In: Energy Metabolism of Farm Animals. EAAP Publication 13.

43
Warnick AC and Cobb EH. 1976. Genetic variation in nutrition of cattle for meat production. In: The genetic variance of nutritional requirements of animals. National Academy of Sciences, Maryland, USA.
Yan T, Porter MG and Mayne CS. 2009. Prediction of methane emission from beef cattle using data measured in indirect open-circuit respiration calorimeters. Animal 3, 1455-1462.
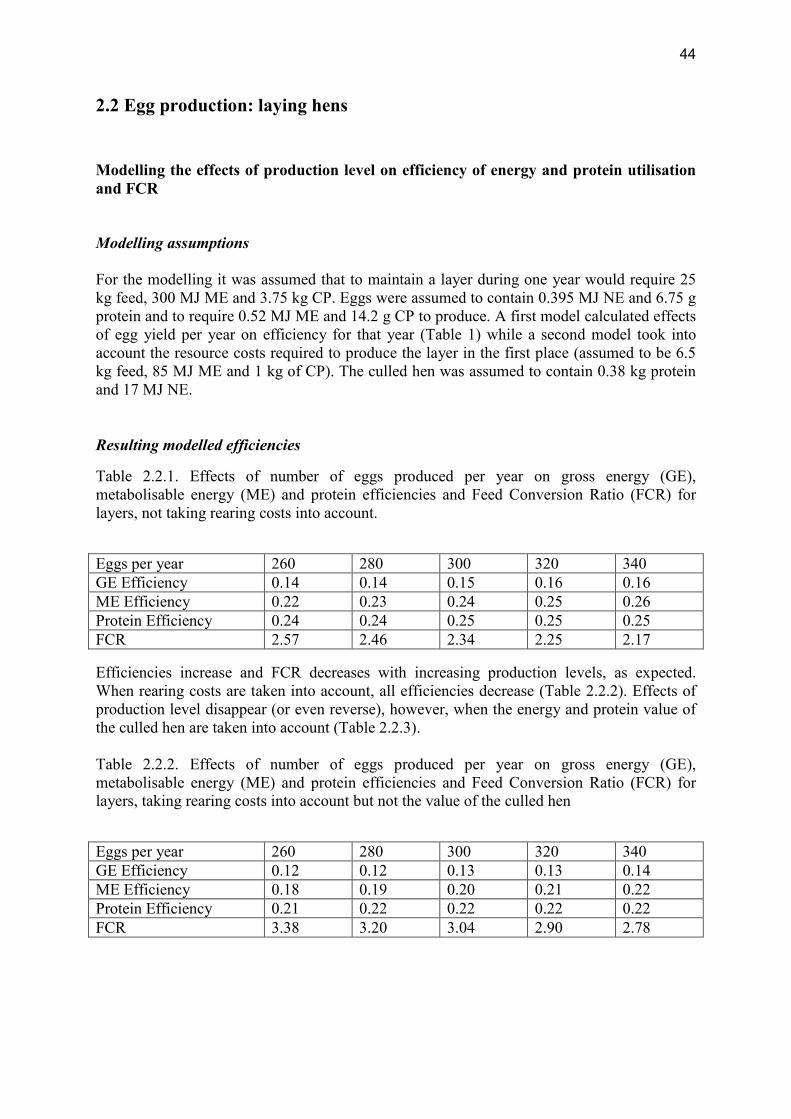
44
2.2 Egg production: laying hens
Modelling the effects of production level on efficiency of energy and protein utilisation and FCR
Modelling assumptions For the modelling it was assumed that to maintain a layer during one year would require 25 kg feed, 300 MJ ME and 3.75 kg CP. Eggs were assumed to contain 0.395 MJ NE and 6.75 g protein and to require 0.52 MJ ME and 14.2 g CP to produce. A first model calculated effects of egg yield per year on efficiency for that year (Table 1) while a second model took into account the resource costs required to produce the layer in the first place (assumed to be 6.5 kg feed, 85 MJ ME and 1 kg of CP). The culled hen was assumed to contain 0.38 kg protein and 17 MJ NE. Resulting modelled efficiencies
Table 2.2.1. Effects of number of eggs produced per year on gross energy (GE), metabolisable energy (ME) and protein efficiencies and Feed Conversion Ratio (FCR) for layers, not taking rearing costs into account.
Eggs per year 260 280 300 320 340 GE Efficiency 0.14 0.14 0.15 0.16 0.16 ME Efficiency 0.22 0.23 0.24 0.25 0.26 Protein Efficiency 0.24 0.24 0.25 0.25 0.25 FCR 2.57 2.46 2.34 2.25 2.17 Efficiencies increase and FCR decreases with increasing production levels, as expected. When rearing costs are taken into account, all efficiencies decrease (Table 2.2.2). Effects of production level disappear (or even reverse), however, when the energy and protein value of the culled hen are taken into account (Table 2.2.3). Table 2.2.2. Effects of number of eggs produced per year on gross energy (GE), metabolisable energy (ME) and protein efficiencies and Feed Conversion Ratio (FCR) for layers, taking rearing costs into account but not the value of the culled hen
Eggs per year 260 280 300 320 340 GE Efficiency 0.12 0.12 0.13 0.13 0.14 ME Efficiency 0.18 0.19 0.20 0.21 0.22 Protein Efficiency 0.21 0.22 0.22 0.22 0.22 FCR 3.38 3.20 3.04 2.90 2.78
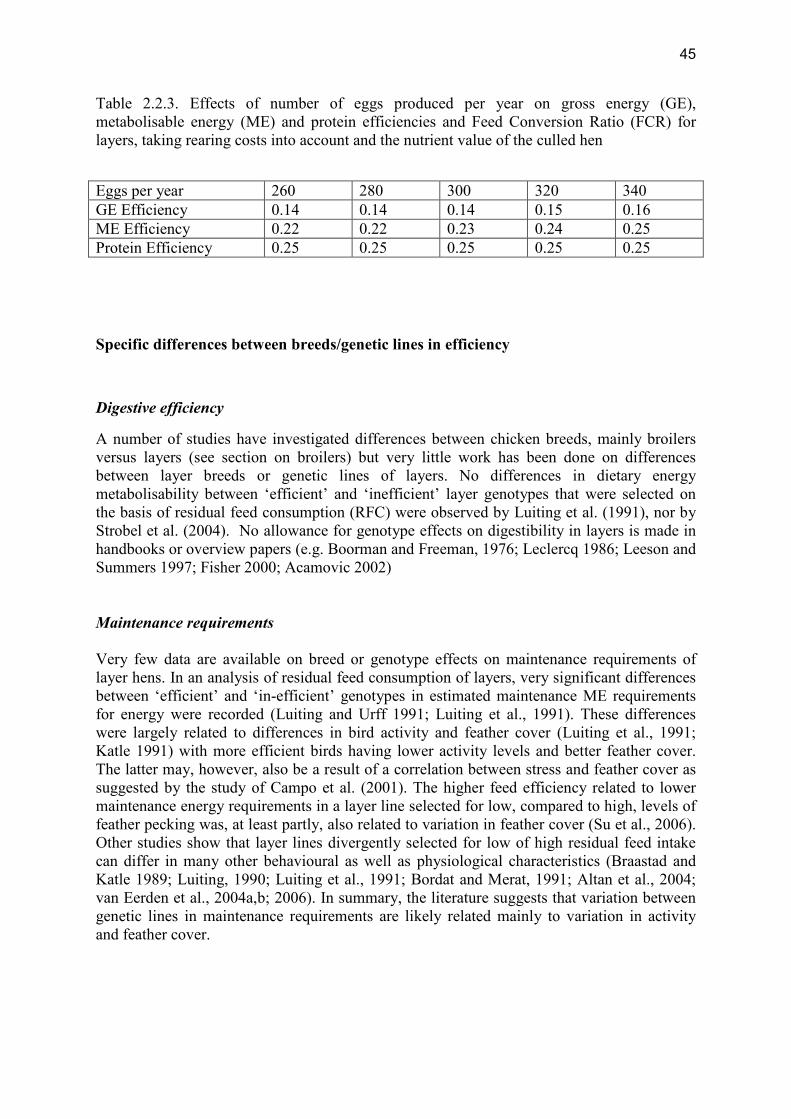
45
Table 2.2.3. Effects of number of eggs produced per year on gross energy (GE), metabolisable energy (ME) and protein efficiencies and Feed Conversion Ratio (FCR) for layers, taking rearing costs into account and the nutrient value of the culled hen
Eggs per year 260 280 300 320 340 GE Efficiency 0.14 0.14 0.14 0.15 0.16 ME Efficiency 0.22 0.22 0.23 0.24 0.25 Protein Efficiency 0.25 0.25 0.25 0.25 0.25
Specific differences between breeds/genetic lines in efficiency
Digestive efficiency
A number of studies have investigated differences between chicken breeds, mainly broilers versus layers (see section on broilers) but very little work has been done on differences between layer breeds or genetic lines of layers. No differences in dietary energy metabolisability between ‘efficient’ and ‘inefficient’ layer genotypes that were selected on the basis of residual feed consumption (RFC) were observed by Luiting et al. (1991), nor by Strobel et al. (2004). No allowance for genotype effects on digestibility in layers is made in handbooks or overview papers (e.g. Boorman and Freeman, 1976; Leclercq 1986; Leeson and Summers 1997; Fisher 2000; Acamovic 2002) Maintenance requirements Very few data are available on breed or genotype effects on maintenance requirements of layer hens. In an analysis of residual feed consumption of layers, very significant differences between ‘efficient’ and ‘in-efficient’ genotypes in estimated maintenance ME requirements for energy were recorded (Luiting and Urff 1991; Luiting et al., 1991). These differences were largely related to differences in bird activity and feather cover (Luiting et al., 1991; Katle 1991) with more efficient birds having lower activity levels and better feather cover. The latter may, however, also be a result of a correlation between stress and feather cover as suggested by the study of Campo et al. (2001). The higher feed efficiency related to lower maintenance energy requirements in a layer line selected for low, compared to high, levels of feather pecking was, at least partly, also related to variation in feather cover (Su et al., 2006). Other studies show that layer lines divergently selected for low of high residual feed intake can differ in many other behavioural as well as physiological characteristics (Braastad and Katle 1989; Luiting, 1990; Luiting et al., 1991; Bordat and Merat, 1991; Altan et al., 2004; van Eerden et al., 2004a,b; 2006). In summary, the literature suggests that variation between genetic lines in maintenance requirements are likely related mainly to variation in activity and feather cover.

46
Efficiency of energy and protein utilisation proper
No information on differences between breeds or genetic lines in efficiency of energy and protein utilisation proper has been found for laying hens. No allowance for genotype effects is made in handbooks or overview papers (e.g. Boorman and Freeman, 1976; Leclercq 1986; Leeson and Summers 1997; Fisher 2000; Acamovic 2002). FCR Considerable differences in FCR between four genotypes in an organic production system were recorded by Sorensen and Kjaer (1991). FCRs ranged from 2.45 (ISA-Brown) to 4.05 (New Hampshire) kg feed/kg eggs. The variation was mainly due to the lower rate of lay and higher age at first egg in the New Hampshire breed (and therefore a direct result of differences in productivity) and occurred despite higher mortality (mainly due to cannibalism) in the ISA-Brown. Other factors The occurrence of disease and liveability/mortality of laying hens not only affect the economics of eggs production systems but these factors can also have an important effect on the efficiency of energy and nutrient utilisation at system level (Flock et al., 2005). References Acamovic T. 2002. Nutrient requirements and standards for poultry. Report of the British
Society of Animal Science Nutritional Standards Working Group, March 2002. Altan O, Aguz I, Akbas Y, Aksit M. 2004. Genetic variability of residual feed consumption
(RFC) and its relationships with some production traits and fear response in Japanese quail hens (Coturnic coturnix japonica). Archiv fur Geflügelkunde 68, 223-229.
Bordas A and Merat P. 1991. Selection divergente pour la consommation alimentaire residuelle de la poule en periode de ponte: reponse au taux proteique de l’aliment. Genetics Selection Evolution 23, 249-256.
Braastad BO and Katle J. 1989. Behavioral differences between laying hen populations selected for high and low efficiency of food utilization. British Poultry Science 30, 533-544.
Campo JL, Gil MG, Torres O, Davila SG. 2001. Association between plumage condition and fear and stress levels in five breeds of chickens. Poultry Science 80, 549-552.
Fisher C. 2000 Advances in feed evaluation for poultry. Pp. 243-268 in P.J. Moughan, M.W.A. Verstegen and M.I. Visser-Teyneveld (Eds.) Feed Evaluation: Principles and Practice. Wageningen Pers, Wageningen, The Netherlands.
Flock DK, Laughlin KF and Bentley J. 2005. Minimizing losses in poultry breeding and production: how breeding companies contribute to poultry welfare. Worlds Poultry Science Journal 61, 227-237.
Katle J. 1991. Selection for efficiency of food utilization in laying hens – causal factors for variation in residual feed consumption. Poultry Science 32, 955-969.
Leclerq B. 1986. Energy requirements of avian species. Pp. 125-139 in C. Fisher and K.N. Boorman (Eds.) Nutrient Requirements of Poultry and Nutritional Research.. Butterworth, Sevenoaks, UK.

47
Leeson S and Summers JD. 1997. Feeding programmes for laying hens. Pp. 143-206 in Commercial Poultry Nutrition. University Books, Guelph, Canada.
Luiting P. 1990. Genetic variation of energy partitioning in laying hens. Causes of variation in residual feed consumption. Worlds Poultry Science Journal 46, 133-152.
Luiting P and Urff EM. 1991. Optimization of A Model to Estimate Residual Feed Consumption in the Laying Hen. Livestock Production Science 27, 321-338.
Luiting P, Schrama JW, van der Hel W and Urff E.M. 1991. Metabolic differences between White Leghorns selected for high and low residual food consumption. British Poultry Science 32, 763-782.
Su G, Kjaer JB and Sorensen P. 2006. Divergent selection on feather pecking behavior in laying hens has caused differences between lines in egg production, egg quality, and feed efficiency. Poultry Science 85, 191-197.
Van Eerden E, Van Den Brand H, Parmentier HK, de Jong MCM, Kemp B. 2004a. Phenotypic selection for residual feed intake and its effect on humoral immune responses in growing layer hens. Poultry Science 83, 1602-1609.
Van Eerden E, Van Den Brand H, Reilingh GD, Parmentier HK, de Jong MCM, Kemp B. 2004b. Residual feed intake and its effect on Salmonella enteritidis infection in growing layer hens. Poultry Science 83, 1904-1910.
Van Eerden E, Van Den Brand H, Heetkamp MJW, Decuypere E and Kemp B. 2006. Energy partitioning and thyroid hormone levels during Salmonella enteritidis infections in pullets with high or low residual feed intake. Poultry Science 85, 1775-1783.
Strobel E, Franke E, Manner K. 2004. Energy exchange in white laying hens with differences in residual food consumption. Archiv fur Geflügelkunde 68, 62-70.
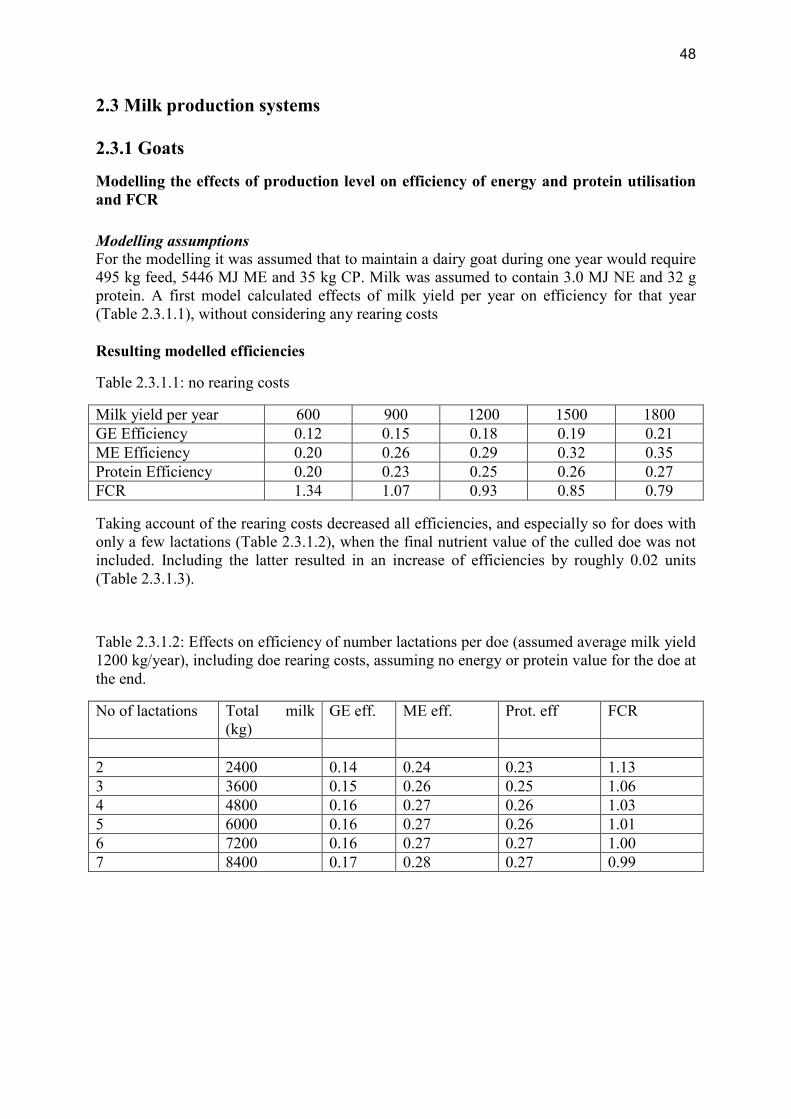
48
2.3 Milk production systems 2.3.1 Goats
Modelling the effects of production level on efficiency of energy and protein utilisation and FCR Modelling assumptions For the modelling it was assumed that to maintain a dairy goat during one year would require 495 kg feed, 5446 MJ ME and 35 kg CP. Milk was assumed to contain 3.0 MJ NE and 32 g protein. A first model calculated effects of milk yield per year on efficiency for that year (Table 2.3.1.1), without considering any rearing costs Resulting modelled efficiencies
Table 2.3.1.1: no rearing costs
Milk yield per year 600 900 1200 1500 1800 GE Efficiency 0.12 0.15 0.18 0.19 0.21 ME Efficiency 0.20 0.26 0.29 0.32 0.35 Protein Efficiency 0.20 0.23 0.25 0.26 0.27 FCR 1.34 1.07 0.93 0.85 0.79 Taking account of the rearing costs decreased all efficiencies, and especially so for does with only a few lactations (Table 2.3.1.2), when the final nutrient value of the culled doe was not included. Including the latter resulted in an increase of efficiencies by roughly 0.02 units (Table 2.3.1.3).
Table 2.3.1.2: Effects on efficiency of number lactations per doe (assumed average milk yield 1200 kg/year), including doe rearing costs, assuming no energy or protein value for the doe at the end.
No of lactations Total milk (kg)
GE eff. ME eff. Prot. eff FCR
2 2400 0.14 0.24 0.23 1.13 3 3600 0.15 0.26 0.25 1.06 4 4800 0.16 0.27 0.26 1.03 5 6000 0.16 0.27 0.26 1.01 6 7200 0.16 0.27 0.27 1.00 7 8400 0.17 0.28 0.27 0.99

49
Table 2.3.1.2: Effects on efficiency of number lactations per doe (assumed average milk yield 1200 kg/year), including doe rearing costs and final energy and protein value for the doe.
No of lactations Total milk (kg)
GE eff. ME eff. Prot. eff
2 2400 0.14 0.27 0.25 3 3600 0.15 0.28 0.26 4 4800 0.16 0.28 0.27 5 6000 0.16 0.28 0.27 6 7200 0.16 0.28 0.28 7 8400 0.17 0.29 0.28
Specific differences between breeds/genetic lines in the efficiency of energy and protein utilisation
Digestive efficiency
Considerable differences in DM digestibility may be recorded between local unimproved goats breeds (such the Black Bedouin) and improved international breeds (such as the Saanen; e.g. Silanikove, 1986), which is likely related to longer rumen retention times of digesta in Bedouin goats (Silanikove et al. 1993). For better quality forage (alfalfa), the lower digestibility observed in Saanen goats was more than offset by the considerably higher feed intake (and resulting higher performance) of Saanen, but that was not the case for low quality (i.e. high fibre, low protein) feed such as straw. No differences, however, were observed more recently between four contrasting goat breeds (Alpine, Angora, Boer and Spanish) for DM, OM, CP, NDF or GE on either a high concentrate or a high forage diet. (Tovar-Luna et al., 2007a,b) and also Urge et al. (2004) found that in these same genotypes OM digestibility was similar, independent of the proportion of concentrate in the diet. In general, digestibilities measured with one ruminant species are frequently applied to other species, sometimes after a correction for feeding level (see discussion under dairy cows). Although goats may have a reputation of being able to digest forages better than other ruminant species (e.g. Knights and Garcia, 1997), this is frequently based on experiments in which variation in digestibility was confounded with variation in diet composition (i.e. goats selected better parts from the diet on offer than cattle). After correction for such confounding effects, goats’ digestive ability was only marginally higher than that of sheep and, if anything, slightly lower than that of cattle (as reviewed by Tolkamp and Brouwer, 1993). At present, there is no convincing evidence for important differences between goat breeds in digestive efficiency, at least not when they are offered medium to good quality diets (Isac et al., 1994; Tovar-Luna et al., 2007a,b). Methane energy as a proportion of total energy intake decreases with increasing feed intake level in goats (Aguilera and Prieto, 1991), which would be an additional beneficial effect on energy efficiency of using highly productive genotypes. Maintenance requirements The literature shows considerable variation in estimated energy maintenance requirements of different goat breeds as estimated from regression of animal performance on variation in ME intake or vice versa. The work of Zemmelink et al. (1991) has shown, however, that most of this variation disappears when such data sets are analysed with the same model. There is

50
some evidence, as for other ruminant species, that specialised dairy goat breeds have higher maintenance energy requirements as measured from fasting heat production than meat breeds (Roy-Smith, 1980; Mohammed, 1982; Ketelaars and Tolkamp 1992; Tolkamp et al., 1994; Sahlu et al., 2004; Tovar-Luna, 2007a). Whether or not such differences can be explained by differences in body composition alone (as observed earlier for pigs and sheep), with dairy types generally being leaner than meat type goats, has, as far as we know, not been investigated. Other factors Considerable differences may exist under adverse environmental conditions between goat breeds in fertility (Bradford et al., 1989) with traditional dairy breeds (such as Saanen, Toggenburg and French Alpine) being more negatively affected than more adapted breeds such as Nubian and Granadina goats (Mellado et al., 2006; Mellado and Meza-Herrera 2002). Also under similar conditions, breed affects on fertility and litter size do exist (Amoah et al., 1996; Goonewardene et al., 1997). Breed does not have an important effect on the shape (in contrast to the overall level) of lactation curves in dairy goats (Gipson and Grossman 1990).
References
Aguilera JF, Prieto C. 1991. Methane Production in Goats Given Diets Based on Lucerne Hay and Barley. Archives of Animal Nutrition 41, 77-84.
Amoah EA, Gelaye S, Guthrie P and Rexroad CE. 1996. Breeding season and aspects of reproduction of female goats. Journal of Animal Science 74, 723-728.
Bradford GE, Burfening PJ, Cartwright TC. 1989. Evaluation of production and reproduction of sheep, goat and alpaca genotypes in the small ruminant collaborative research support program. Journal of Animal Science 67, 3058-3067.
Gipson TA and Grossman M. 1990. Lactation curves in dairy goats - A review. Small Ruminant Research 3, 383-396.
Goonewardene LA, Whitmore W, Jaeger S, Borchert T, Okine E, Ashmawy O and Emond S. 1997. Effect of prebreeding maintenance diet on subsequent reproduction by artificial insemination in Alpine and Saanen goats. Theriogenology 48, 151-159.
Isac M D, Garcia MA Aguilera JF and Alcaide EM. 1994. A comparative-study of nutrient digestibility, kinetics of digestion and passage and rumen fermentation pattern in goats and sheep offered medium quality forages at the maintenance level of feeding. Archives of Animal Nutrition 46, 37-50.
Knights M and Garcia GW. 1997. The status and characteristics of the goat (Capra hircus) and its potential role as a significant milk producer in the tropics: A Review. Small Ruminant Research 26, 203-215.
Mellado M and Meza-Herrera CA. 2002. Influence of season and environment on fertility of goats in a hot-arid environment. Journal of Agricultural Science 138, 97-101.
Mellado M, Valdez R, Garcia JE, Lopez R and Rodriguez A. 2006. Factors affecting the reproductive performance of goats under intensive conditions in a hot arid environment. Small Ruminant Research 63, 110-118.
Mohammed HH. 1982. Energy requirements for maintenance and growth: comparison of goats and sheep. PhD thesis, University of Reading.

51
Roy-Smith F. 1980. The fasting metabolism and relative energy intake of goats compared to sheep. Animal Production 30, 491.
Sahlu T, Goetsch AL, Luo J, Nsahlai IV, Moore JE, Galyean ML, Owens FN, Ferrell CL, Johnson ZB. 2004. Nutrient requirements of goats: developed equations, other considerations and future research to improve them. Small Ruminant Research 53, 191-219.
Silanikove N. 1986. Interrelationships between feed quality, digestibility, feed consumption and energy requirements in desert (Bedouin) and temperate (Saanen) goats. Journal of Dairy Science 69, 2157-2162.
Silanikove N, Tagari H and Shkolnik A. 1993. Comparison of rate of passage, fermentation rate and efficiency of digestion of high-fiber diet in desert bedouin goats compared to Swiss Saanen goats. Small Ruminant Research 12, 45-60.
Tolkamp BJ, Ketelaars JJMH, Hofs, P. 1994. Voluntary intake of digestible organic matter and fasting heat production of West African Dwarf Goats and Swifter sheep. Small Ruminant Research 15, 45-54.
Tovar-Luna I Goetsch AL Puchala R Sahlu T Carstens GE, Freetly, HC, Johnson ZB. 2007a. Effects of diet quality on energy expenditure by 20-month-old Alpine, Angora, Boer, and Spanish wethers. Small Ruminant Research 72, 18-24.
Tovar-Luna I, Goetsch AL, Puchala R, Sahlu T, Carstens GE, Freetly HC and Johnson ZB. 2007b. Efficiency of energy use for maintenance and gain by growing crossbred Boer and Spanish Goats consuming diets differing in forage level. Small Ruminant Research 72, 20-27.
Zemmelink G, Tolkamp BJ, Ogink NWM. Energy requirements for maintenance and gain of West African Dwarf goats. Small Ruminant Research 5, 205-215.

52
2.3.2 Cows
Modelling the effects of production level on efficiency of energy and protein utilisation and FCR Modelling assumptions For the modelling it was assumed that to maintain a cow during one year would require 2347 kg feed, 25812 MJ ME and 170 kg CP. Milk was assumed to contain 3.0 MJ NE and 32 g protein and to require 5.0 MJ ME and 80 g CP to produce. A first model calculated effects of milk yield per year on efficiency for that year only (Table 2.3.2.1). Resulting modelled efficiencies
Table 1: Effects of milk yield per year on energy and protein efficiency and FCR, taking no rearing costs into account
Milk yield per year, kg 4000 6000 8000 10000 12000 GE Efficiency 0.16 0.2 0.24 0.26 0.27 ME Efficiency 0.26 0.32 0.36 0.40 0.42 Protein Efficiency 0.22 0.26 0.29 0.30 0.32 FCR 1.04 0.81 0.69 0.63 0.60 There is a considerable effect of the total number of lactations when efficiencies are calculated to include rearing costs (assumed to be 3595 kg feed, 39540 MJ ME and 450 kg of CP; Table 2.3.2.2). A large part of the effects of lactation number on efficiency disappears, however, when the energy and protein value of the culled cow is included in the efficiency calculations.
Table 2.3.2.2: Effects on efficiency of number lactations per cow (assumed average milk yield 8000 kg/year), including rearing costs, assuming no energy or protein value for the culled cow
No of lactations Total milk (kg)
GE eff. ME eff. Prot. eff FCR
2 16000 0.18 0.28 0.23 0.91 3 24000 0.20 0.30 0.25 0.84 4 32000 0.21 0.32 0.26 0.88 5 40000 0.21 0.32 0.26 0.78 6 48000 0.21 0.33 0.27 0.76 7 56000 0.22 0.34 0.27 0.75
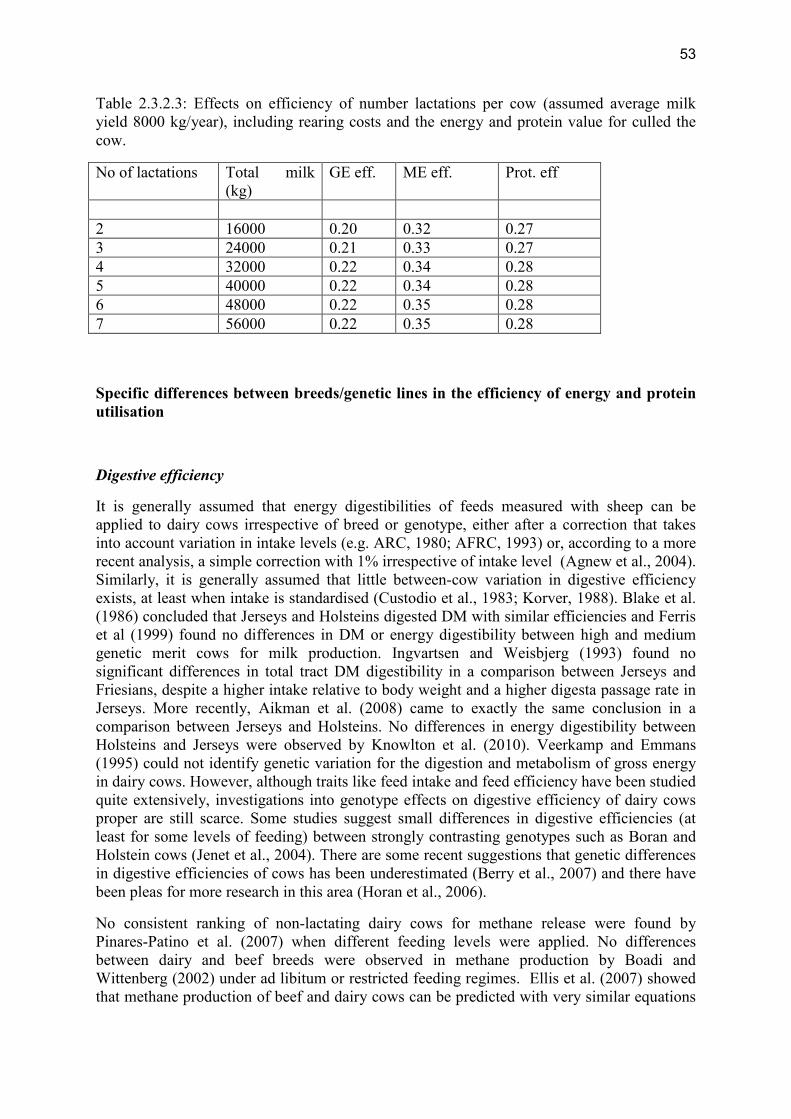
53
Table 2.3.2.3: Effects on efficiency of number lactations per cow (assumed average milk yield 8000 kg/year), including rearing costs and the energy and protein value for culled the cow.
No of lactations Total milk (kg)
GE eff. ME eff. Prot. eff
2 16000 0.20 0.32 0.27 3 24000 0.21 0.33 0.27 4 32000 0.22 0.34 0.28 5 40000 0.22 0.34 0.28 6 48000 0.22 0.35 0.28 7 56000 0.22 0.35 0.28
Specific differences between breeds/genetic lines in the efficiency of energy and protein utilisation
Digestive efficiency
It is generally assumed that energy digestibilities of feeds measured with sheep can be applied to dairy cows irrespective of breed or genotype, either after a correction that takes into account variation in intake levels (e.g. ARC, 1980; AFRC, 1993) or, according to a more recent analysis, a simple correction with 1% irrespective of intake level (Agnew et al., 2004). Similarly, it is generally assumed that little between-cow variation in digestive efficiency exists, at least when intake is standardised (Custodio et al., 1983; Korver, 1988). Blake et al. (1986) concluded that Jerseys and Holsteins digested DM with similar efficiencies and Ferris et al (1999) found no differences in DM or energy digestibility between high and medium genetic merit cows for milk production. Ingvartsen and Weisbjerg (1993) found no significant differences in total tract DM digestibility in a comparison between Jerseys and Friesians, despite a higher intake relative to body weight and a higher digesta passage rate in Jerseys. More recently, Aikman et al. (2008) came to exactly the same conclusion in a comparison between Jerseys and Holsteins. No differences in energy digestibility between Holsteins and Jerseys were observed by Knowlton et al. (2010). Veerkamp and Emmans (1995) could not identify genetic variation for the digestion and metabolism of gross energy in dairy cows. However, although traits like feed intake and feed efficiency have been studied quite extensively, investigations into genotype effects on digestive efficiency of dairy cows proper are still scarce. Some studies suggest small differences in digestive efficiencies (at least for some levels of feeding) between strongly contrasting genotypes such as Boran and Holstein cows (Jenet et al., 2004). There are some recent suggestions that genetic differences in digestive efficiencies of cows has been underestimated (Berry et al., 2007) and there have been pleas for more research in this area (Horan et al., 2006).
No consistent ranking of non-lactating dairy cows for methane release were found by Pinares-Patino et al. (2007) when different feeding levels were applied. No differences between dairy and beef breeds were observed in methane production by Boadi and Wittenberg (2002) under ad libitum or restricted feeding regimes. Ellis et al. (2007) showed that methane production of beef and dairy cows can be predicted with very similar equations

54
based on feed characteristics alone. Munger and Kreuzer (2008) compared methane release per kg of DM consumed by Holstein, Simmental and Jersey cows and found (i) no effects of breed on this variable at all and (ii) no cows with consistently higher or lower than average methane yields. They concluded that the apparent lack of persistence of individual animal differences in methane yields suggests that genetic determination of this trait is of minor importance in dairy cows (Munger and Kreuzer, 2008).
Maintenance requirements
As shown by Birnie et al. (2000), maintenance requirement estimates of dairy cows are, as in other species (see, e.g. the relevant paragraphs on sheep and pigs) affected by body composition with the leanest animals generally recording the highest maintenance energy requirements per unit metabolic size (although rare exceptions to this rule exist; see e.g. Ortigues et al., 1993). As discussed under beef cows, effects of body composition may be involved in the frequently observed increasing estimated maintenance energy requirements with dairy merit of the breed. Also the genotype x nutrient allowance interaction effect on estimated maintenance energy requirements of cows of different breeds observed by Jenkins and Ferell (2007) is likely related to the effect of different maturities on such comparisons (as discussed in the relevant paragraph under sheep). Recent estimates of NE and ME for maintenance requirements of modern genotypes of dairy cows (e.g. Agnew et al., 2004) are considerably higher (by around 40%) than earlier estimates as used in most current international energy evaluation systems but these high estimates are controversial (e.g. Tolkamp and Kyriazakis 2009).
Efficiency of energy and protein utilization proper
At present, energy and protein evaluation systems for dairy cows do not incorporate any effects of breed/genotype on efficiency of energy and protein utilisation proper (e.g. ARC, 1980; AFRC, 1993; Agnew et al., 2004. INRA, 1989; CSIRO, 1990; NRC 2001) and the same efficiencies of energy and protein utilisation are used across ruminant species such as cattle, sheep and goats (e.g. AFRC, 1993). Yan et al. (2006) concluded that Holstein Friesian and Norwegian cows utilised the ME with similar efficiency for milk production on both high- and low-concentrate diets. From calorimetric studies, Ferris et al. (1999) concluded that energy was utilised with similar efficiency for milk production in medium and high-genetic merit cows. Veerkamp and Emmans (1995) could not identify genetic variation in the partial efficiencies with which ME is converted to different products in dairy cows.
References
AFRC (Agricultural and Food Research Council). 1993. Energy and Protein Requirements of Ruminants. An advisory manual prepared by the AFRC Technical Committee on Responses to Nutrients. CAB International, Wallingford, UK.
Agnew RE, Yan T, France J, Kebreab E and Thomas C. 2004. Energy requirement and supply. In Feed into Milk. A new applied feeding system for dairy cows (ed. C Thomas), pp. 11-20. Nottingham University Press, Nottingham, UK.

55
Aikman PC, Reynolds CK, Beever DE. 2008. Diet digestibility, rate of passage, and eating and rumination behavior of Jersey and Holstein cows. Journal of Dairy Science 91, 1103-1114.
ARC (Agricultural Research Council). 1980. The Nutrient Requirements of Ruminant Livestock. CAB International, Wallingford, UK.
Berry DP, Horan B, O'Donovan M, Buckley F, Kennedy E, McEvoy M and Dillon P 2007. Genetics of grass dry matter intake, energy balance, and digestibility in grazing Irish dairy cows. Journal of Dairy Science 90, 4835-4845.
Birnie JW, Agnew RE, Gordon FJ. 2000. The influence of body condition on the fasting energy metabolism of nonpregnant, nonlactating dairy cows. Journal of Dairy Science 83, 1217-1223.
Blake RW, Custodia AA, Howard WH. 1986. Comparative feed-efficiency of Holstein and Jersey cows.. Journal of Dairy Science 69, 1302-1308.
Boadi DA, Wittenberg KM. 2002. Methane production from dairy and beef heifers fed forages differing in nutrient density using the sulphur hexafluoride (SF6) tracer gas technique. Canadian Journal of Animal Science 82, 201-206.
Commonwealth Scientific and Industrial Research Organisation. 1990. Feeding Standards for Australian Livestock: Ruminants. Compiled by the Ruminants Subcommittee of the Standing Committee on Agriculture. CSIRO, East Melbourne, Australia.
Custodio AA, Blake RW, Dahm PF, Cartwright TC, Schelling GT, Coppock CE. 1983. Relationships between measures of feed efficiency and transmitting ability for milk of Holstein cows. Journal of Dairy Science 66, 1937-1946.
Ellis JE, Kebreab E, Odongo NE, McBride BW, Okine EK and France J. 2007. Prediction of methane production from dairy and beef cattle. Journal of Dairy Science 90, 3456-3467.
Ferris CP, Gordon FJ, Patterson DC, Porter, MG and Yan T. 1999. The effect of genetic merit and concentrate proportion in the diet on nutrient utilization by lactating dairy cows. Journal of Agricultural Science 132, 483-490.
Horan B, Faverdin P, Delaby L, Rath M and Dillon P. 2006. The effect of strain of Holstein-Friesian dairy cow and pasture-based system on grass intake and milk production. Animal Science 82, 435-444.
Ingvartsen KL, Weisbjerg MR. 1993. Jersey cows have a higher feed intake capacity and higher rate of passage than Friesian cows. Archives of Animal Breeding 36, 495-498.
Institute National de la Recherche Agronomique. 1989. Ruminant Nutrition, Recommended Allowances & Feed Tables (ed. R Jarrige). John Libbey and Co Ltd, London.
Jenet A, Yimegnuhal A, Fernandez-Rivera S, Tegegne A, Osuji PO, McCrabb G, Kreuzer M 2004. Long-term response to feeding level in lactational performance of Boran (Bos indicus) and Boran X Holstein cows. Animal Science 78, 331-343.
Jenkins TG, Ferrell CL. 2007. Daily dry matter intake to sustain body weight of mature, nonlactating, nonpregnant cows. Journal of Animal Science 85, 1787-1792.
Knowlton KF, Wilkerson VA, Casper DP, Mertens DR. 2010. Manure nutrient excretion by Jersey and Holstein cows. Journal of Dairy Science 93, 407-412.
Korver S. 1988. Genetic aspects of feed-intake and feed-efficiency in dairy cattle – a review. Livestock Production Science 20, 1-13.
Munger A. and Kreuzer M. 2008. Absence of persistent methane emission differences in three breeds of dairy cows. Australian Journal of Experimental Agriculture 48, 77-82.
National Research Council 2001. Nutrient Requirements of Dairy Cattle. 7th revised edition. National Academy Press, Washington DC, USA.
Ortigues I, Petit M and Agabriel J. 1993. Influence of body condition on maintenance energy requirements of Charolais cows. Animal Production 57, 47-53.

56
Pinares-Patino CS, Waghorn GC, Machmueller A, Vlaming B, Molano G, Cavanagh A, Clark H. 2007. Methane emissions and digestive physiology of non-lactating dairy cows fed pasture forage. Canadian Journal of Animal Science 87, 601-613.
Tolkamp BJ and Kyriazakis I. 2009. Toward a new practical energy evaluation system for dairy cows. Animal 3: 307-314.
Van Es AJH 1961. Between-animal variation in the amount of energy required for the maintenance of cows. PhD thesis, Wageningen University.
Veerkamp RF and Emmans GC. 1995. Sources of genetic variation in energetic efficiency in dairy cows. Livestock Production Science 44, 87-97.
Yan T, Mayne CS and Agnew RE. 2006. Effects of dairy cow genotype with two planes of nutrition on energy partitioning between milk and body tissue. Journal of Dairy Science 89, 1031-1042.

57
3 The feasibility of incorporating lifecycle nutrient efficiency traits into breeding programmes for farmed livestock species 3.1 The feasibility of incorporating lifecycle nutrient efficiency traits into breeding programmes for ruminants
Many non genetic farm technologies require ongoing investment of some sort to maintain the commercial benefit (e.g., dietary manipulation to improve fatty acid composition of milk). However, genetic improvement of a livestock population is effectively a permanent change and does not require additional or continuing resources.
In designing a breeding programme, the definition of biological and economic efficiency, and their optimisation, is important. Efficiency can be defined as the ratio of output/input, with output and input given in a variety of units, biological, physical or financial. Using the term “efficiency” requires a clear definition, both of the term chosen and of the system studied. Definitions concerning nutrition and breeding are given and recommended with respect to primary and secondary traits as well as an overall selection index. A few examples are: (1) residual food intake (estimated energy in feed intake minus estimated energy requirements for production and maintenance; (2) feed efficiency; (3) economic efficiency, the net income per unit of product (Ostergaard et al., 1990).
3.1.1. Feed utilisation efficiency and correlated impacts
The pig and poultry industries have incorporated production efficiency in their selection programmes by including traits such as feed conversion (FCR) reducing the time and the amount of feed required to reach market weight. This improvement in efficiency of growing animals has a favourable effect on the emissions from these livestock industries per unit of product. These improvements have mainly been as a result of selective breeding AND improved feeding systems. For example, the time for a broiler to reach 2 kg has reduced from 63 days in 1976 to 36 days in 1999 (McKay et al., 2000). Similar trends can be seen in pigs with Dutch Landrace improving feed efficiency from 3.5 (kg/kg) in 1930 to 2.8 in 1990 (Merks, 2000). These genetic improvements have led to improved productivity parameters for livestock across all world regions, with improvements more marked in industrialised countries. For example, the world kg output/kg biomass of chicken meat has increased from 1.83 in 1980 to 2.47 in 2005 and for pig meat has increased from 0.31 to 0.45 (FAO, 2006). Improving the efficiency with which animals convert feed into saleable product reduces the emissions by producing that livestock commodity with reduced inputs.
Although feed utilisation is routinely considered in the selection programmes for pig and poultry species, due to the nature of many ruminant production systems, with less opportunity for intensive feed recording, the use of such traits in selection has been limited but there have been some examples. Herd et al. (2002) showed that there is a decreased enteric methane production per day in animals selected for reduced residual feed intake. Reduced residual feed intake is akin to selection for high feed efficiency as an animal is eating less but maintaining a similar growth rate (high net feed efficiency) and therefore less feed is required to produce a unit of output. Lines were divergently selected for high and low residual feed intake and showed no significant differences for most production traits. This shows the

58
possibilities for selection of reduced GHG emissions through the selection of animals which use less feed and produce less methane than average to achieve a given level of performance.
Traits related to the efficiency of absorption of dietary nutrients: Direct selection for efficiency of utilisation of the different components of the diet is difficult to achieve as many animal and feed parameters need to be collected. Work on these types of traits has mainly been at an experimental level. Ferris et al. (1999) showed that medium genetic merit (for production) Holstein-Friesian cows have higher nitrogen and methane emissions per unit of N and gross energy intake respectively than high genetic merit cows. This suggests that high genetic merit cows convert the energy and protein components of the feed more effectively than medium genetic merit cows. The estimates of heritability for residual feed intake (RFI) in growing beef animals are moderate (average 0.28; range 0.08-0.43) indicating the existence of genetic variations in the trait (see Herd et al. 2003 for review). Most genetic parameters for RFI were estimated in young cattle in feedlots. Information on RFI in breeding herds is very limited with few studies on adult cattle. Archer et al. (2002) reported an estimate of heritability for mature cows of 0.23.
The feeding efficiency of the animal may also have an impact with low-RFI beef cattle eating less than expected for their live weight and growth rate. Angus cattle divergently selected for RFI currently attain the same growth rates but differ by approximately 15% in their voluntary feed intake (Herd et al. 2003). Not only may selection for low RFI have a favourable effect on energy nutrient efficiency, it may also impact on protein nutrient efficiency. Low RFI animals have been shown to have a reduced manure production (15% to 20% less compared to high RFI cattle) and also on the potential quantity of nitrous oxide liberated from these manures. This potential effect on nitrous oxide can be due to a reduction in the total N intake and a greater efficiency of capturing the dietary N (Hegarty et al. 2007).
A selection experiment for low and high RFI in growing beef cattle has been carried out in Australia since mid 90’s. Results from this experiment not only confirmed that RFI can be genetically improved by selection but only provided interesting evidence on the association of RFI and other traits. The efficiency of beef production systems depend on many traits and the interaction among them, including the feed intake of the breeding herd and slaughter generation, growth traits, as well as other cow traits, such as mature size and reproductive rate (Archer et al. 1999).
Cammack et al. (2005) reported a rather low heritability of 11% for RFI for ram lambs of a composite breed and 25% for their daily feed intake and 26% for their average daily gain. Francois et al. (2002) reported a more substantial heritability for RFI of 30% in ram lambs of a French composite breed with 43% for daily feed intake and 43% for average daily gain.
Increasing the efficiency of production of meat animals reduces emissions per unit output. The studies of Mrode et al. (1990a,b) compared two strategies of selection on lean growth rate and food conversion ratio (FCR = daily food intake/growth rate). The increase in carcass lean and reduction in food conversion ratio with selection for lean growth were similar to the responses with selection for FCR (Mrode et al., 1990b) resulting in an overall improvement in FCR of 7% compared to a control line. However, growth rate was increased with selection for lean growth, but not with selection for improved FCR (Mrode et al., 1990b). Therefore, in terms of genetic improvement for feed efficiency and growth rate of the slaughter generation, selection for lean growth rate is preferable to selection for FCR, such that it is not necessary to measure food intake. However, their conclusions may not hold for maternal lines where

59
selection for lean growth rate is likely to lead to a correlated increase in mature feed maintenance/requirements.
A further example of high vs. low efficiency animals can be seen with Langhill select and control lines of dairy cows. This herd has been selected for maximum production of kg milk fat plus protein (select group) or to remain at the UK national average for production (control group) and has been fed two diets of different energy density (high or low forage). In these data the select group have been shown to have 17% higher yield per lactation, and a 14% higher gross efficiency (Veerkamp et al., 1995). Further studies of the data showed significant differences in how cows lost and regained body energy throughout lactation to support production resulting in the higher gross efficiency, with select group cows on the high forage diet losing the greatest amount of body lipid over three lactations (Coffey et al., 2004). However, it must be noted that the select group of cows achieve this higher efficiency by utilising body energy reserves to support lactation. Although utilising body reserves to support lactation has a favourable effect on methane emissions within the first lactation, it is likely that the longer term reliance on body energy reserves to support lactation will have a longer term unfavourable biological effect and impact on health, fertility and other “fitness” traits in the dairy cow (Friggens and Newbold, 2007; Garnsworthy et al., 2008). Poorer health and fitness in the dairy herd will affect overall systems (and lifetime) nutrient efficiencies as more followers will be required to maintain the herd size as more dairy cows may be culled involuntarily and need to be replaced. However, as shown in section 2, if this nutrient energy from the culled animals is “repaid” to the farming system this difference may be negligible.
Further examination of the metabolic turnover of nutrients in animals may be required to understand the underlying biological differences between high producing animals in their feed utilisation and lower genetic merit animals.
A theoretical study, as part of Defra project IF0149 (Determining strategies for delivering environmentally sustainable production in the UK ruminant industry through genetic improvement) explored the impact of adding feed efficiency traits into the economic indices of beef and sheep breeds in UK production systems. These results showed large net incomes were obtained when genomic selection was applied simultaneously with the inclusion of survival and RFI in the breeding goals in both maternal (62% - 80%) and terminal (33% to 47%) beef breeding programmes. Net farm income benefits in the sheep breeding programmes were also beneficial but smaller in magnitude. Although the net farm income was improved the overall net on farm returns showed that their implementation was not economically viable in all cases until genotyping costs reduce, as they are expected to do.
3.1.2. System nutrient efficiency
Many “fitness” traits have been shown to have a genetic component demonstrating there is scope to improve them via genetic selection. Broader breeding goals that seek to select animals on an optimal combination of production and fitness traits can help to optimise nutrient efficiency in the livestock. However, many of these traits improve overall system nutrient efficiency by assuming that the value of cull animals has no energy value to the system. In theoretical terms, a cull animal from which products are produced (e.g., selling old culled dairy cows for pet food production) means that the energy that went into growing that animal, and maintaining it while it was part of the breeding herd, has an end purpose and

60
therefore is not “wasted” or “lost”. However, in real terms the “economic” value of culled animals means that an involuntary decision to cull an animal (e.g., for poor health and fertility) is seen as a system loss. Therefore looking at things in terms of economic efficiency as opposed to nutrient efficiency may result in a slightly different objective being formed.
Selection for improved fitness traits (lifespan, health, fertility) will help to improve nutrient efficiency by reducing wastage of animals. Improving lifespan in dairy cows and breeding beef cows and ewes will improve the efficiency of the system by reducing the number of followers required to maintain the herd at a given size.
Improving health and fertility will reduce involuntary culling rates and thereby reduce emissions from dairy systems and beef and sheep systems (increased maternal survival) by reducing the numbers of followers required to maintain the herd at a given size. Improving fertility will reduce calving/lambing intervals and inseminations resulting in shorter dry/unproductive periods. This reduces management costs as well as emissions. Garnsworthy (2004) estimated, via modelling, that if cow fertility was restored to 1995 levels from 2003 levels, methane emissions from the dairy industry would reduce by 10-15%, mainly due to reduced follower numbers. Selection for health should reduce treatment costs (and lower antibiotic use), reduce inefficiencies from produce withdrawal during treatment and reducing emissions by maintaining the productivity level of the animal (which is reduced during periods of poor health), all of which contribute to reduced emissions, as well as count to improving animal welfare.
Improving calving/lambing and maternal traits will reduce emissions by improving survival of offspring during the peri-, neo- and post- natal periods. This will reduce wastage in a farming system, thereby improving system wide nutrient efficiency as well as improving calf and dam welfare and survival.
3.1.3. Considering nutrient efficiency in ruminant breeding goals.
Broader breeding goals have become the norm in many livestock species, usually incorporating production and "fitness" (health, fertility, longevity) traits. Breeding goals can be built in a number of ways including the popular method of weighting traits by their relative economic value (REV). These REV's tend to be calculated by estimating the economic benefit to the system of a unit change in the traits being examined. A lot of the example traits given earlier have been incorporated into indices for particular livestock sectors. However, livestock industries have more recently needed to consider societal views of aspects of farming systems, including issues such as welfare, biodiversity, food safety, health properties and environment.
Many traits described earlier, including those routinely included in current selection indices, have an impact on the nutrient efficiency on the animal and the farming system. They also have a cost implication in terms of the (feed) energy required to maintain breeding animals and fuel production. Many of the breeding goals have been developed with feed utilisation components, some more complex than others and these models could be adapted to consider lifetime nutrient efficiency. Farm models could be used to model the nutrient efficiency of animals and from a livestock system and the effect that a change in a trait (e.g., fertility)

61
would have on overall nutrient efficiency. This is similar to the framework used to estimate REVs and weightings ("relative environmental values") derived could be used to express the nutrient efficiency of alternative breeding goals and objectives.
3.1.4. Considering genotype * environment interaction with regard to nutrient efficiency in ruminant breeding goals.
This study has examined the extent to which there is genetic/breed variation in nutrient efficiency in livestock species. However, there is undoubtedly a large nutritional component with much research on the differences between diets in how they are utilised by ruminant animals. However, little work has been done on the potential role of genetics on nutrient efficiency, particularly considering the role of genetics in the whole farming system and it's interaction with including feeding strategy and management policies (e.g., energy balance, housing periods, replacement strategy, economic value of cull animals).
Selection indices have tended to be expressed in terms of a generalised system representative of the “average” farm. The system parameters and relative economics will also differ from production type to production type. For example, recent work has shown that the economics of body tissue mobilisation differs depending on the calving system employed (spring vs. autumn) due to the different costs of feed at grazing opposed to winter feeding (Wall et al., 2008).
Many of the data that have driven the exploration of nutrient efficiency in ruminant systems have focussed on more commercial breeds (growing) of a animals in indoor experimental/testing facilities usually on a higher energy diet. There is gaps in data that explores the role of less numerous/local breeds and considering a range of diets, including grazing on pastures of different quality. This is largely due to a lack of easy recording systems to undertake such studies accurately. This may be an important gap in knowledge in the role that breeds that may be seen a valuable for conversion in playing their part in development of nutrient efficiency systems of production across the range of farming systems that will exit in the future.

62
3.2 The feasibility of incorporating lifecycle nutrient efficiency traits into breeding programmes for pigs
In pig breeding, selection for nutrient efficiency has to be carried out differently for growing-finishing pigs and breeding sows, due to two main different performances (growth versus reproduction), different metabolism (growing versus mature animals), different breeding programmes, etc. In pig production, growing finishing pigs are the result of crossing a sire line (with emphasis on production performances such as growth, leanness, feed efficiency) with a crossbred dam line (with emphasis on both production and reproduction performances such as, growth rate, piglets born alive, survival of piglets, rebreeding interval, longevity of sows, etc.). The nutrient efficiency is determined by several components, which may be genetically improved separately or by combined traits such as feed conversion ratio. In the following the selective ability of important component traits are described in detail, within each component trait firstly for growing finishing pigs and secondly for sows.
3.2.1 Digestive efficiency
Based on the review presented in section 2.1.5 there is no evidence that modern pig genotypes showed genetic differences in efficiency of energy and protein digestibility in growing-finishing pigs. For sows, there may be small difference between local compared to modern pig breeds, because the digestive tract is larger and develops earlier in local breeds. The low differences between breeds may be due to the fact that selection has not been on the difficult to measure trait digestive efficiency or that during evolution the most efficient digestive ability has been developed. Further research on genomics of digestive efficiency is necessary to get more insight into the genetics of digestive efficiency in pigs.
3.2.2 Maintenance requirements
Using Fasting Heat Production (FHP) as indicator for energy maintenance requirements, the literature review in section 2.1.5 suggests that there are differences between breeds. In particular, animal behaviour may be directly associated with energy requirements for maintenance. Several studies have shown that animal behaviour is highly heritable (e.g. Von Felde et al., 1996; Schulze et al., 2003 using feed intake behaviour; Turner et al., 2008,2009; D’Eath et al., 2009 using aggression of pigs) and therefore behavioural traits may be used as indirect selection traits to reduce maintenance requirements and improve animal welfare.
3.2.3 Efficiency of utilisation of energy and protein
Based on the literature review, there are substantial difficulties to measure the efficiency of utilisation of energy and protein for lipid and protein deposition. Therefore, there has to be developed better selection criteria (measured on large number of animals) before a genetic improvement for these traits is possible. Identification of QTL associated with utilisation of energy and protein may be a tool to improve efficiency of utilisation of energy and protein.

63
3.2.4. Body composition
Body composition has a high impact on nutrient efficiency because fat tissue growth requires at least four times more food energy than lean tissue growth due to differences in water content and metabolism (Whittemore, 1993; van Milgen and Noblet, 2003). Selection for reduction in fat tissue and increase in lean tissue has been very successful in pigs, decreasing backfat thickness from 3.2 to 1.9 mm and increase in loin muscle area from 40 to 60 cm2 (Roehe et al., 2003). In particular, the heritability of backfat thickness is high whereas those for lean tissue are moderate (Schulze et al., 2001 and 2002). Furthermore, there have been detected several QTL of body compositions (e.g. Mohrman et al., 2006; Duthie et al., 2008 and 2009). However, the optimal fatness of several dam and sire lines may already achieved by selection so that further reduction in fatness to improve nutrient efficiency may not be an option for most modern pig breeding lines.
3.2.5. Growth rate
Improvement of growth rate will substantially improve nutrient efficiency per kg product because of the reduction in the growing-finishing period and thus reduction in maintenance requirement due to the time the animals will be finished earlier. Table 2.1.5.2 shows that feed conversion ratio improves substantially by increase in growth rate and thus reduction in age at slaughter. Growth rate has a moderate heritability (e.g. Eissen 2000; von Felde et al., 1996, Schulze et al., 2003). In particular increase in lean growth rate will further improve the nutrient efficiency as measured by FCR.
3.2.6. Feed intake and appetite
Feed intake has been reduced in breeding lines selected for lean tissue, because of a negative correlation between feed intake and lean tissue. Reduced feed intake capacity may be the limiting factor for further genetic improvement of growth rate, in particular at early stage of growth (Webb, 1998; van Milgen and Noblet, 2003) and thus reduce the further opportunity to improve nutrient efficiency. However, voluntary feed intake may be driven by energy needed for maintenance requirements and protein and lipid deposition (Emmans, 1997). To optimize feed intake capacity by selection, considering protein as one main driver for feed intake, De Vries and Kanis (1992) developed a bioeconomic growth model using Whittemore and Fawcett’s (1976) linear-plateau model of protein deposition. Consequently, the improvement of feed efficiency by optimization of feed intake capacity with respect to protein deposition curve (considering a minimum lipid deposition) is expected to efficiently improve nutrient efficiency as measured by FCR (Roehe et al., 2002 and 2004).
3.2.7. Feed efficiency
Direct selection for feed efficiency, calculated as a ratio of daily feed intake to daily gain, results in disproportional selection pressure (depending on selection intensity and the genetic

64
parameters) on its two components so that a genetic change is unpredictable (Gunsett 1984; Krieter and Presuhn 1986). Consequently, selection for feed efficiency is inefficient. This inefficiency may have not been recognized in the past, because most of the improvement of feed efficiency was due to reduction in backfat thickness, which now approached its optimum in populations largely selected for lean content. In addition, growth rate has been improved substantially due to breeding, which improved feed efficiency.
3.2.8. Animal reproductive performance
Sow productivity by improving of number of piglets born or piglets born alive are lowly heritable but have been very successful because of the high variation in these traits (Su et al., 2008; Wolf et al., 2009, Kapell et al., 2009) and will improve nutrient efficiency per produced piglet. However, piglet mortality has increased due to selection for number of piglet and therefore selection for piglet survival is expected to improve number of weaned piglets and nutrient efficiency per weaned piglet without high nutritional loss due to loss of piglets (Su et al., 2008; Baxter et al., 2009; Roehe et al., 2009).
3.2.9. Longevity
As shown in section 2.1.5, nutrient efficiency improves with increase in number of parities per sows and thus longevity. Based on a literature review, Serenius and Stadler (2005) concluded that sufficient genetic variation exists for effective selection on longevity, and heritability estimates ranged between 0.02 and 0.25. Traits genetically associated with longevity such as leg conformation can be used to improve the accuracy of estimation of breeding values for longevity and thus its genetic progress.
References
Archer JA, Richardson EC, Herd RM and Arthur PF. 1999. Potential for selection to improve
efficiency of feed use in beef cattle: a review. Australian Journal of Agricultural Research 50, 147-161.
Archer JA., Reverter A, Herd RM, Johnston DJ, and Arthur PF. 2002. Genetic variation in feed intake and efficiency of mature beef cows and relationships with postweaning measurements. Proceedings of the 7th World Congress on Genetics Applied to Livestock Production. Montpellier, France.
Baxter, EM, Jarvis S, Sherwood L, Robson SK, Ormandy E, Farish M, Smurthwaite KM, Roehe R, Lawrence AB, Edwards SA. 2009. Indicators of piglet survival in an outdoor farrowing system. Livestock Science 124, 266-276.
Cammack, KM, Leymaster KA, Jenkins TG, and Nielsen MK. 200). Estimates of genetic parameters for feed intake, feeding behavior, and daily gain in composite ram lambs. Journal of Animal Science 83, 777-785.
Coffey MP, Simm G, Oldham JD, Hill WG and Brotherstone S. 2004. Genotype and diet effects on energy balance in the first three lactations of dairy cows. Journal of Dairy Science 87, 4318-4326.

65
D’Eath RB, Roehe R, Turner SP, Ison SH, Farish M, Jack MC, Lawrence AB. 2009. Genetics of animal temperament: aggressive behaviour at mixing is genetically associated with the response to handling in pigs. Animal 3, 1544-1554.
De Vries, A.G. and Kanis, E. (1992) A growth-model to estimate economic values for food-intake capacity in pigs. Animal Production 55, 241-246.
Duthie C, Simm G, Doeschl-Wilson A, Kalm E, Knap PW, Roehe R. 2008. Quantitative trait loci for chemical body composition traits in pigs and their positional associations with body tissues, growth and feed intake. Animal Genetics 39, 130-140.
Duthie C, Simm G, Pérez-Enciso M, Doeschl-Wilson A, Kalm E, Knap PW, Roehe R. 2009. Genomic scan for quantitative trait loci of chemical and physical body composition and deposition on pig chromosome X considering the pseudoautosomal region. Genetics Selection Evolution 41, 27 (Open Access)
Eissen JJ. 2000. Breeding for feed intake capacity in pigs. Doctoral thesis, University Wageningen, Netherlands.
FAO, 2006. FAO Statistical databases. Rome. [http://faostat.fao.org] Ferris CP, Gordon FJ, Patterson DC, Porter MG and Yan T. 1999. The effect of genetic merit
and concentrate proportion in the diet on the nutrient utlization by lactating dairy cows. Journal of Agricultural Science 132: 483-490.
Francois D, Bibe B, Bouix J, Brunel JC, Weisbecker JL and Ricard E. 2002. Genetic Parameters of Feeding Traits in Meat Sheep. Proceedings of the 7th World Congress on Genetics Applied to Livestock Production. Montpellier, France.
Friggens NC and Newbold JR 2007. Towards a biological basis for predicting nutrient partitioning: the dairy cow as an example. Animal 1, 87-97
Garnsworthy PC 2004. The Environmental impact of fertility in dairy cows: a modeling approach to predict methane and ammonia emissions. Animal Feed Science and Technology 112, 211-223.
Garnsworthy PC, Sinclair KD and Webb R 2008. Integration of physiological mechanisms that influence fertility in dairy cows. Animal 2, 1144-1152
Gunsett FC. 1984. Linear index selection to improve traits defined as ratios. Journal of Animal Science. 59: 1185-1193.
Hegarty, RS, Goopy JP, Herd RM, and McCorkell B. 2007. Cattle selected for lower residual feed intake have reduced daily methane production. Journal of Animal Science 85, 1479-1486.
Herd R, Archer JA, and Arthur PF. 2003. Reducing the cost of beef production through genetic improvement in residual feed intake: opportunities and challenges to application. Journal of Animal Science 81 (E supplement), E9-E17.
Herd RM, Arthur PF, Hegarty RS and Archer JA. 2002. Potential to reduce greenhouse gas emissions from beef production by selection for reduced residual feed intake. 7th World Congress on Genetics Applied to Livestock Production, August 19-23, 2002, Montpellier, France.
Kapell, DNRG, Ashworth CJ, Walling GA, Lawrence AB, Edwards SA, Roehe R. 2009. Estimation of genetic associations between reproduction and production traits based on a sire and dam line with common ancestry. Animal 3, 1354-1362
Lorenzo Bermejo J, Roehe R, Schulze V, Rave G, Looft H, Kalm E. 2003b. Random regression to model genetically the longitudinal data of daily feed intake in growing pigs. Livestock Production Science. 82, 189-199.
McKay JC, Barton NF, Koerhuis ANM and McAdam J. 2000. The challenge of genetic change in the broiler chicken. In "The challenge of genetic change in animal production". BSAS Occasional Publication Number 27.

66
Merks JWM. 2000. One century of genetic change in pigs and the future needs. In "The challenge of genetic change in animal production". BSAS Occasional Publication Number 27.
Mohrmann M, Roehe R, Knap PW, Looft H, Plastow GS, Kalm E. 2006. Quantitative trait loci associated with AutoFOM grading characteristics, carcass cuts and chemical body composition during growth of Sus scrofa. Animal Genetics 37, 435-443.
Moughan PJ. 2003. Journal of Animal Science 81 (E. Suppl. 2), E60-E67. Mrode RA, Smith C and Thompson R 1990a. Selection for rate and efficiency of lean gain in
Hereford cattle. 1. Selection pressure applied and direct response. Animal Production 51, 23-34.
Mrode RA, Smith C and Thompson R 1990b. Selection for rate and efficiency of lean gain in Hereford cattle. 2. Evaluation of correlated responses. Animal Production 51, 35-46.
Ostergaard, V, S. Korver, H. Solbu, B.B. Andersen, J. Oldham and H, Wiktorsson. 1990 Main Report - E.A.A.P. Working Group on: Efficiency in the Dairy Cow. Livestock Production Science, 24: 287-304.
Pomar C, Kyriazakis I, Emmans GC and Knap PW. 2003. Journal of Animal Science 81 (E. Suppl. 2), E178-E186.
Roehe R, Plastow GS, Knap PW. 2003. Quantitative and molecular genetic determination of protein and fat deposition. Journal HOMO of Comparative Human Biology 54, 119-131.
Roehe R, Landgraf S, Mohrmann M, Knap PW, Looft H and Kalm E. 2004. Proc. 55th Annual Meeting EAAP, Bled, Slovenia, paper PG6.7, 1-10.
Roehe R, Lorenzo Bermejo J, Landgraf S, Knap PW and Kalm E. 2002. Proc. 53th Annual Meeting EAAP, Cairo, Egypt, paper G6.3, 1-8.
Roehe R, Shrestha NP, Mekkawy,W, Baxter EM, Knap PW, Smurthwaite KM, Jarvis S, Lawrence AB, Edwards SA. 2009. Genetic analyses of piglet survival and individual birth weight on first generation data of a selection experiment for piglet survival under outdoor conditions. Livestock Science 121, 173-181
Schulze V, Roehe R, Looft H and Kalm E. 1998. Genetic improvement of exterior and leg soundness in pigs. Züchtungskunde 70, 43-60.
Schulze V, Roehe R, Looft H, Kalm E. 2001. Genetic analysis of the course of individual growth and feed intake of group-penned performance tested boars. Archives of Animal Breeding 44, 139-156.
Schulze V, Roehe R, Lorenzo Bermejo J, Looft H, Kalm E. 2002. Genetic associations between observed feed intake measurements during growth, feed intake curve parameters and growing-finishing performances of central tested boars. Livestock Production Science 73, 199-211.
Schulze V, Roehe R, Lorenzo Bermejo J, Looft H, Kalm E. 2003. The influence of feeding behaviour on feed intake curve parameters and performance traits of station-tested boars. Livestock Production Science 82, 105-116.
Su G, Sorensen D, and Lund MS. 2008. Variance and covariance components for liability of piglet survival during different periods. Animal 2:184-189.
Turner SP, Roehe R, Mekkawy W, Farnworth MJ, Knap PW, Lawrence AB. 2008. Bayesian analysis of genetic associations of skin lesions and behavioural traits to identify genetic components of individual aggressiveness in pigs. Behaviour Genetics 38, 67-75.
Turner SP, Roehe R, D’Eath RB, Ison SH, Farish M, Jack MC, Lundeheim N, Rydhmer L, Lawrence AB. 2009. Genetic validation of post-mixing skin injuries in pigs as an indicator of aggressiveness and the relationship with injuries under more stable social conditions. Journal of Animal Science 87, 3076-3082.

67
Van Milgen J and Noblet J. 2003. Journal of Animal Science 81 (E. Suppl. 2), E86-E93. Veerkamp RF, Simm G and Oldham JD. 1995 Genotype by environment interaction -
experience from Langhill. In Breeding and Feeding the High Genetic Merit Dairy Cow. Occasional Publication 19, Lawrence TLJ, Gordon FJ and Carson S (eds) British Society of Animal Science, Edinburgh, United Kingdom.
Von Felde A, Roehe R, Looft H, Kalm E. 1996. Genetic association between feed intake and feed intake behaviour at different stages of growth of group-housed boars. Livestock Production Science 47, 11-22.
Wall E, Coffey MP and Amer PR 2008. A theoretical framework for deriving direct economic values for body tissue mobilization traits. Journal of Diary Science 91, 343-353.
Wolf J, Žáková E, and Groeneveld E. 2008. Within-litter variation of birth weight in hyperprolific Czech Large White sows and its relation to litter size traits, stillborn piglets and losses until weaning. Livestock Science 115, 195-205.
Whittemore CT, Green DM and Knap PW. 2001. Technical Review of the energy and protein requirements of pigs: protein. Animal Science 73, 363-373.

68
4. General discussion and conclusions
4.1 Overview and discussion of breed/genetic line effects on efficiencies in different production systems
4.1.1 Modelled effects of productivity on energy and protein efficiencies
It is evident from the review that there are considerable breed/genotype effects on energy and nutrient efficiency as a direct result of differences in productivity. That includes, first, productivity in terms of growth rate and level of milk and egg production in the animals providing the end product for human consumption. These effects are largely a result of utilising consumed energy and protein to a larger extent for energy and protein retention (in growth, milk or egg yield) and to a lesser extent for maintenance. For many production systems, also breed/genotype effects on productivity of the breeding population (i.e. in terms of fertility, mortality, longevity and disease resistance) can have considerable effects on energy and nutrient efficiency of production systems as a whole. This is especially the case when (a large proportion of) breeding animals do not themselves enter the food chain. When most breeding animals are culled and slaughtered, however, that is much less the case.
It should be emphasised here that that is so in terms of energy and nutrient efficiency, which does not (necessarily) correspond with economic efficiency. This is a result of the fact that a MJ of energy and a kg of protein in breeding animals (e.g. a sow) generally has a much lower value than the same in animals directly produced for consumption (e.g. pigs grown for slaughter at a relatively young age).
For most production systems, breeding programmes already include one or more productivity traits in their selection indices. These have had, therefore, a beneficial effect on energy and nutrient efficiencies of such production systems (and are expected to have similar effects in the future as well).
Many selection programmes also include traits in relation to product composition, most importantly the lean : lipid ratio in the product. An increase in the lean : lipid ratio will generally have a very beneficial effect on FCR because of the relatively low energy costs of depositing 1 g of lean compared to 1 g of lipid. A lower FCR is also of great economic importance. However, the effects of such traits on energy and protein efficiency are not immediately clear. It is evident that selection for higher lean : lipid ratio will be beneficial for the efficiency of protein utilisation if it is associated with partitioning a larger proportion of consumed feed protein to retention, for reasons discussed above. The effects on energetic efficiency per se, however, could well be more variable, or even negative. The amount of metabolisable energy required to retain a gram of protein does not differ much from the amount of ME required to retain a gram of lipid (most estimates vary around 53 kJ/g; see e.g. Whittemore, 2006). Because the net energy NE content of protein is lower than that of lipid (around 24 compared with around 40 kJ/g), leaner genotypes are expected to be less energetically efficient that fatter genotypes when they retain the same amount of NE relative to maintenance requirements.
A comparison of energy and protein efficiencies of the different production systems (Table 2.4.1) shows that broiler production systems are the most efficient and lamb and beef production systems the least. Although not the subject of the current review, it should be emphasised that this is so for the definitions of energy and protein efficiency used in the present report. The situation would be very different if efficiency was defined as the amount

69
of energy or protein in animal product divided by the amount of feed energy and protein that is suitable for human consumption. In some production systems, such as those with broilers, this would have a limited effect on efficiency estimates as shown in this report because a major proportion of feed energy and protein consists of grains and protein meals that are directly suitable for human consumption. However, in other systems, especially ruminant production systems based largely on forage, such an efficiency would be considerable higher than those listed in Table 2.4.1 (values could even exceed 1.0; Van Es, 1975).
Table 2.4.1. Summary of average gross energy (GE), metabolisable energy (ME) and crude protein (CP) efficiencies and feed conversion ratios (FCR) of ten production systems.
Species Production GE eff. ME eff CP eff FCR Broiler 36 d to slaughter, no rearing costs 0.28 0.41 0.52 1.69 Turkey 126 d to slaughter, no rearing costs 0.19 0.30 0.48 2.46 Geese 98 d to slaughter, no rearing costs 0.27 0.45 0.30 3.26 Ducks 42 d to slaughter, no rearing costs 0.29 0.44 0.40 2.41 Pigs 22 wks to slaughter, 4 litters of10 piglets 0.21 0.31 0.31 2.99 Sheep 1.72 lambs/litter, 4 litters 0.06 0.10 0.09 10.4 Beef 4 calves/cow 0.06 0.12 0.10 10.4 Layers 300 eggs/hen, including rearing costs 0.14 0.23 0.25 Goats 1200 kg milk/lactation, 4 lactations 0.16 0.28 0.27 Cows 8000 kg milk/lactation, 4 lactations 0.22 0.34 0.28
4.1.2 Digestive efficiencies
For monogastrics, there is some evidence for breed/genetic line effects on digestive efficiency, although there is a lack of data for some species. Comparisons between layer, broiler and dwarf genotype type chickens sometimes show small differences in the amounts of ME extracted per kg feed but such comparisons are frequently confounded with effects of animal size and feeding level. Even genetic lines of broilers divergently selected for lean and lipid growth or between ‘efficient’ and ‘inefficient’ layer lines are usually observed to extract very similar amounts of ME per kg feed. Recent research has suggested, however, that selection for digestive efficiency per se can be effective perhaps, although it may be associated with other changes, e.g. in body composition. Also in pigs, statistically significant differences in energy digestibility are sometimes observed. Such differences are usually (but not universally) recorded especially when local and relatively unimproved pig breeds are compared with international improved pig breeds fed on poor quality (especially: high-fibre), but not on high-quality, diets. There is scant evidence for important differences between such breeds in enzymatic digestion and any differences are usually related to a larger/earlier developing lower tract in local breeds resulting in slightly higher total-tract digestibilities as a result of lower-tract fermentation. The possible effects of differences in digestive efficiency on total efficiency at the animal

70
level are very small compared to the efficiency gains that can be made by improving productivity. No convincing evidence was found in favour of important differences between improved pig genotypes fed high quality diets in modern production systems. In ruminants it is generally assumed that feed digestibilities measured with one species (usually sheep) can be used (sometimes after a correction, e.g. related to variation in intake level) to other species (such as dairy cows or goats). There is, again, some evidence for breed effects on digestive efficiency in ruminant species, especially when local unimproved breeds are compared with international improved breeds and when animals are fed low quality (but not high quality) diets. Most comparisons of currently common breeds, however, have not led to the view that there are important breed/genotype effects on digestive efficiency in ruminants.
There has been an increase in interest in recent years in a specific aspect of digestion in ruminants as far as that is related to the emission of methane by gastrointestinal micro-organisms. In the previous chapter, only limited attention has been given to the relevant literature in this respect and that is the reason we have devoted the next paragraph to that subject entirely.
4.1.3 Breed/genotype effects on methane release
It has been established by several research groups that within-animal variability in methane production is high. Blaxter and Clapperton (1965) reported highly significant differences in methane emissions between individual sheep fed the same diet when compared to the day to day variation in any individual sheep. However, Munger and Kreuzer 2008 did not find any consistent differences in methane emissions per unit of intake between three breeds of dairy cows. Similarly Blaxter and Clapperton (1965) failed to detect significant differences in methane emissions from six different breeds of sheep, when intake was adjusted based on metabolic body size. Nevertheless, individual animals with extremely low rates of methane energy loss have been identified on both feedlot and pasture diets (Johnson et al. 1991; Joblin 1999). Indeed in grazing sheep consuming the same amount of dry matter Lassey et al. (1997) noted that daily methane emissions varied from 16 to 22.5 g between the 4 lowest and highest emitters respectively. Several studies (Robertson and Waghorn 2002; Pinares et al. 2003; Goopy et al. 2006) have presented evidence that such differences in methane yield persist over time, although other studies could not confirm this (Pinares et al. 2005; Munger and Kreuzer, 2008). Vlaming et al (2008) suggested that diet might affect the repeatability of measurements and that in many studies the precision of the measurement techniques being used might prevent differences being detected. As early as 1966, Hungate noted that between-animal variation in digestion, fermentation and, in particular, rumen turnover rate offered an opportunity to select ruminant livestock for increased productivity (Hungate 1966). A strong association between methane production (% of gross energy intake) and rumen retention time (RRT) was observed in sheep by Pinares et al. (2003), with RRT explaining 57% of the between-sheep variation in methane emission. Pinares et al. (2007) subsequently also confirmed that cows with a low retention time of particles in the rumen produce less methane per unit of intake. In humans, there has been extensive study of methane formation due to concern about hind-gut fermentation and the incidence of colitis (King et al. 1998). These studies routinely partition the human population into methane producers and non-methane producers and only some 28-50% of people in

71
western societies produce methane (Pitt et al. 1980; McKay et al. 1985; Doré et al. 1995). Flatz et al. (1985) determined the heritability of methane excretion using 228 adult twins (h2 = 0.42) and demonstrated that the inheritance of methane production was controlled by a multi-factorial system, not regular Mendelian inheritance. While there is a strong dietary effect on methane production, Pitt et al. (1980) showed methanogenesis could not be induced in non-methane producing people by a lactulose dose which generally promotes methanogenesis. A major determinant of whether people produce methane or not are the mean retention time (MRT) of digesta in the gastrointenstinal tract, which has a large effect on the intestinal populations of methanogens and their competitors. El Oufir et al. (1996) observed that 58% of the variance in faecal methanogen numbers was explained by MRT. While factors other than MRT also influence the pattern of fermentation, the human example provides a clear indication that GIT fermentation may differ greatly between individuals and that fermentation characteristics are heritable. Guan et al (2008) and Zhou et al (2009) have shown that both the structure of total bacterial population and the methanogenic archeal population, respectively, were significant different between high methane and low methane producing cattle. However the extent to which the basis of such a change in the rumen microbial population is genetic is unknown and Yanez et al (2010) have shown that early life nutrition can also significantly affect the microbial population developing in the rumen.
4.1.4 Maintenance requirements
Comparisons of the net energy for maintenance requirements, generally estimated from resting fasting heat production, of different breeds/genetic lines are fraught with a number of methodological issues (see the discussions in the relevant sections, especially under pigs and sheep). These relate specifically to the most appropriate manner in which maintenance requirements of different breeds are compared (at the same age, the same weight, the same maturity? How to account for differences in body composition?). Most of the reviewed data are consistent with the view that observed differences in NE estimates for maintenance tend to diminish or disappear altogether when these take account of the actual protein (or lean) size as well as the mature protein (or lean) size. This view is consistent with the approach taken by Emmans (1994; 1997) that actual and mature protein sizes can account for (most of) the variation between breeds/genotypes and even between species in maintenance NE requirements.
However, even if there seems to be very little effect of breed/genetic line on estimated NE for maintenance as estimated from resting animals in a thermoneutral environment, this does not necessarily mean that also the metabolisable energy for maintenance is unaffected by genotype under farm conditions. There is evidence, especially from chickens and pigs, that there is some real variation in ME requirements for maintenance that is related to differences between genotypes in a number of behavioural or physiological characteristics. Although the present evidence is not entirely conclusive, variation in energy requirements for maintenance have been associated with differences between genotypes in, especially, level of activity and the opportunities for body temperature regulation (as affected by feather cover in poultry). Such differences have been observed in studies that analysed variation between genotypes in residual feed intake, although that is not universally the case (compare e.g. the relevant sections under broilers and layers).

72
4.1.5 Efficiency of energy and protein utilisation ‘proper’.
For poultry and pig production systems there is at present no convincing evidence of important breed/genetic line effects on the efficiency of energy utilisation for protein and lipid retention proper when differences in body composition have been taken into account. Reports of differences in efficiency of protein utilisation between lean and fat genetic lines are frequently confounded because different genotypes were fed the same diets (which will have resulted in ‘protein-overfeeding’ of the fat genotypes) and cannot be considered as evidence for breed effects on the efficiency of protein utilisation proper. Well-conducted studies with contrasting pig breeds have reached the conclusion that as long as protein overfeeding does not occur, there are no important differences between very different genotypes in the efficiency of feed protein utilisation for protein retention proper. Several studies suggest that even across species, there is very little variation in the material efficiency of protein utilisation proper and the energetic efficiency proper of protein and lipid retention (Emmans, 1994; 1997). Current energy and protein evaluation systems all assume that, at least within species, efficiencies for protein and energy utilisation proper can be applied across breed/genetic lines.
There are a very limited number of studies that suggest there might be genetic differences, e.g. at the level of mitochondrial oxidative metabolism, that could affect efficiency of ATP production. Although at present there is no convincing evidence that this results in important differences in energetic efficiency at the level of the animal as a whole, it is possible that more information in this area will become available in the near future.
4.1.6 The use of FCR and RFI
As indicated above, the use of FCR in selection programmes has had a large beneficial effect on the economics of animal production systems and has usually been associated with improvements in energy and nutrient efficiencies as well. FCR’s are, however, affected by a large number of factors and the use of ratio traits for genetic selection may cause problems associated with prediction of change in the component traits in future generations (Hoque and Suzuki, 2009). Residual feed intake (RFI) is a measure of feed efficiency that is independent of animal size and level of production, such as and growth or milk yield. For that reason, there is considerable interest in using this efficiency measure as a useful new trait in selection programmes for a variety of production systems. Herd and Arthur (2009) identified five major physiological processes that are likely to contribute to variation in RFI. These are very similar to the processes that have been discussed in the present report, namely level of feed intake, digestion of feed, metabolism (anabolism and catabolism, including differences in relation to variation in body composition), physical activity, and thermoregulation. At present, the physiological mechanisms identified as affecting RFI are based on relatively few studies, some with small sample sizes, and the genomic basis of variation in these physiological processes remains to be determined (Herd and Arthur, 2009). For that reason, it is not easy to predict exactly how improvements in RFI, will affect energy and nutrient efficiency. It is evident, however, that improvements in RFI resulting from changes in digestion, physical activity and thermo-regulation (as well as several other variables) can be expected to also have a beneficial effect on energy and nutrient efficiency. It is likely that many hundred genes are associated with differences in RFI and further research is needed to link physiological with molecular genetics information before it can become a basis for

73
commercial tests to identify genetically superior animals (Herd and Arthur, 2009) and determine the precise effects on energy and nutrient efficiency. There is some theoretical evidence that selection for RFI will have the additional benefit of reducing greenhouse emissions by livestock but Arthur and Herd (2005) observe that it is still a challenge is to generate the necessary empirical data to support this assertion. 4.2 Incorporating efficiency traits in breeding programmes At present, traits are included in breeding programmes for economic reasons alone. It is evident from the review that improvement in most traits that are currently part of selection programmes for economic reasons alone will also have a beneficial effect on nutrient efficiency. That is valid not only for traits directly related to the production process, such as daily gains or the output of eggs or milk but for all other traits that cause variation in ‘feed wastage’ as a result of low fertility and fecundity, mortality, disease, etc. The issue whether or not traits could be incorporated into breeding programmes with the specific aim of increasing nutrient efficiency then boils down to three questions: 1. Are there traits that are currently used in breeding programmes that have a negative effect on nutrient efficiency? 2. How do traits that are not currently part of breeding programmes but that could/should be part for economic reasons alone affect nutrient efficiency? 3. Are there traits that are not currently part of breeding programmes for economic reasons but that are relevant for nutrient efficiency reasons alone? These will be briefly discussed below. 4.2.1 Are there traits that are currently used in breeding programmes that have a negative effect on nutrient efficiency? Although it is frequently assumed that a decrease in FCR, commonly used in current breeding programmes (especially for monogastrics), will have a beneficial effect on nutrient efficiency, that may not always be the case. As discussed, a shift in the ratio of lean to lipid in animal growth, although highly beneficial from an economic point of view, may not always have a beneficial effect on nutrient efficiency because of the lower partial energetic efficiency for protein retention compared to that for lipid retention. That hardly affects FCR, as a result of the beneficial effect on FCR of all the water that is retained with protein growth. These discrepancies can be avoided by using a more appropriate model for the estimation of RFI (in this case the separation of daily gain in lipid and lean gains). The effects on energetic efficiency are expected to be small, however, and improvements in terms of FCR as a result of increases in the ratio of lean to lipid deposition will remain highly desirable from an economic as well as human nutrition point of view. For other routine biological traits considered in genetic improvement programmes, both production and efficiency traits that are relevant from an economic point of view would appear to have a positive effect on nutrient efficiency. 4.2.2 How do traits that are not currently part of breeding programmes but that could/should be part for economic reasons alone affect nutrient efficiency? At present, traits related to feed efficiency are not commonly part of breeding programmes for ruminants, mainly as a result of the costs associated with intake measurements. There is, however, some evidence that the use of FCR (or perhaps rather RFI) as an additional trait in breeding programmes could have substantial economic benefits. Although at present the

74
exact underlying biological mechanisms that result in variation in RFI are not always well established, improvements in RFI can be expected to result in improved nutrient efficiency as well. This would be an additional argument for including such traits. The main problem, as stated above, is the costs associated with measuring intake and improved cheap methodologies to do that would undoubtedly stimulate inclusion of FCR/RFI into breeding programmes for ruminants. It should be noted that some newer traits that could be included in breeding programmes may have an influence on nutrient efficiency, which have not yet defined. Examples include traits that relate to the composition of the product (e.g., fat:lean content, milk protein content, fatty acid composition). Selection on such traits is likely to influence the manner in which animal partition nutrients towards different biological functions/products. This may have an influence on the efficiency of nutrient utilisation overall. Other traits which may impact on the efficiency with which animals utilise nutrients include immune response traits. The mechanism by which these traits may influence nutrient utilisation is that selection for improved immune function/disease resistance is likely to change the manner by which animals partition energy to their immune system and away from growth. Understanding the relationships on the biological pathways that partition energy in the animal is an important step in understanding the lifetime energy utilisation of an animal. 4.2.3 Are there traits that are not currently part of breeding programmes for economic reasons but that are relevant for nutrient efficiency reasons alone? The literature review has highlighted that, with a few possible exceptions, the variation between genotypes in the partial processes such as digestion and efficiency of energy and protein utilisation proper as currently estimated is small. Some variation in ME for maintenance requirements between genotypes does seem to exist, mainly as a result of variation in activity level and (in birds) variation in feather cover that affects the animal’s temperature regulation energy costs. This can probably be exploited but any variation in ME requirements for maintenance will be automatically covered by breeding programmes that include FCR or RFI as traits. Enormous progress has been made in the selection of animals for specific traits using traditional quantitative genetics approaches. Nevertheless considerable variation in phenotypes remains unexplained, and therefore represents potential additional gain for animal production. In addition, the paradigm shift in new technologies now being applied to animal breeding represents a powerful opportunity to prise open the “black box” underlying the response to selection and fully understand the genetic architecture controlling the traits of interest. These arguments particularly apply to complex traits such as energy partitioning. Unravelling the complexity behind polygenic traits is known as systems biology. A move away from traditional approaches of animal breeding toward systems approaches represents a multitude of exciting opportunities for animal breeding going forward as well as providing alternatives for overcoming some of the limitations of traditional approaches such as the expressed phenotype being an imperfect predictor of the individuals true genetic merit, or the phenotype being only expressed in one gender or late in the lifetime of an animal. The use of genomic information to understand the genetic regulation of complex and/or novel traits provides great opportunity.

75
Genomics is the study of the structure, function and intragenomic interactions within the genome. The potential of genomics in animal breeding relies on the ability to identify the causative mutation giving rise to a difference in phenotype, or to identify a linked genetic marker, and to use the polymorphism in a breeding program with the appropriate breeding goal. Possibly one of the greatest benefits of genomics in animal breeding is that genomic DNA is available from an animal at any age as well as being available from both genders. Knowledge of the “ideal genotype” and the genotype of every animal can facilitate increased genetic gain through a combination of altering accuracy of predicted EBVs and the generation interval. Early work on the utilisation of genomic information in breeding programmes focussed on the development of genetic tests (e.g., CVM, BLAD in dairy bulls, Scrapie in sheep) and more recently on the development of marker panels (up to 10s of markers) for specific traits that may be difficult to record (e.g., meat tenderness). However, with the limited numbers of markers of interest and the complexity and cost in identifying regions of the genome that impacted on quantitative traits (quantitative trait loci, QTL) the further development of should tests has been surpassed with the introduction of high density marker (SNP) panels. This had led to the development of theoretical approaches to selection based on this information, coined genomic selection. Genomic selection, as we have come to know it was first described by Meuwissen et al. (2001) and has been described as “the most promising application of molecular genetics in livestock populations since work began almost 20 years ago” (Sellner et al., 2007). It is based on the simultaneous selection for many thousand of genetic markers that densely cover the entire genome made possible by the development of arrays with many thousands of SNPs for fast genotyping throughput. Several alternative breeding programs are possible to exploit genomic selection and to date no published study has thoroughly investigated the alternatives. The cost of screening many thousands of animals is currently prohibitive and development of a smaller scale SNP array, which is of lower cost should be investigated. This type of methodology may be particularly important in identifying the genetic control of traits relating to nutrient efficiency, where there would be a limit to the numbers of animals with detailed phenotypes. Transcriptomics is the study of the expression levels of messenger RNA (mRNA) in a given tissue at a certain time under different conditions. Messenger RNA is transcribed from DNA and carries the coding information for synthesis of proteins which in turn affect cellular function and ultimately the phenotype. However, we now know that this pathway is fraught with complexity including differences in coding and regulatory DNA sequences, alternate splicing of RNA and mRNA, the regulation by small RNA of translation through transcript degradation, and the post-translational modification of the protein on the way to generating the phenotype. Once a candidate gene (or genome region) is found to have a significant effect on a given phenotype then expression studies on the gene in the population can be undertaken to better understand the role that gene plays in the biological process affecting that trait. Understanding the biological process underpinning the expression of a phenotype can give greater insight into other potential candidate genes and molecular mechanisms regulating the trait of interest. This type of methodology could prove extremely useful for complex traits, likely to be influenced by a number of biological pathways, working differently at different life stages (e.g., growing animals vs. mature animal nutrient partitioning rules). This may be also of particular relevance in understanding how trait expression is influenced in different environments.

76
Proteomics is the large-scale study of the structure and function of proteins and how they interact with the genome while metabolomics is the study of the relationship between the genome and metabolic processes. Up to recently researchers generally concentrated on only a few proteins or metabolites but new platforms have been developed facilitating more rapid quantification of protein and metabolite levels without any bias associated with the protein or metabolite to be studied. The advantage of investigative research on proteins and metabolites over that on the genome is that proteins and metabolites in themselves are inherently biological end-products. Information contained in the nucleic acid sequence of the DNA determines, via transcription to mRNA, the amino acid sequence of a protein which in turn determines it structure and function. All biological process and observed phenotypes are affected by this process and while the basic nucleic acid sequence does not change, factors such as environmental conditions can affect the transcription of DNA to mRNA and eventually to protein. Further steps to utilise these techniques/technologies and apply them to relevant traits in livestock, might help us understand the complexity of traits and how they are expressed in different animals, in different environments and at different stages of life. Also, it may help us understand the interaction between traits at a genomic, genetic and environmental level. As genetic improvement techniques are refined and become more powerful using modern DNA based methods, it is vital that all participants in food production on-farm are aware of the genetic differences between animals and account for such genetic variation when formulating management strategies such as feeding, housing and health treatments. Better still would be a joint approach to breeding animals with their subsequent management environment taken into account. A major challenge to the future of animal breeding, unrelated to the science itself, is the shift in funding, and by default scientists and students, from quantitative genetics to molecular genetics and related disciplines. Detailed economic analyses must be undertaken on the long-term cost-benefit of resources being drained from traditional animal breeding into generation of critical mass in systems biology. This is not to say that resources should not be expended in systems biology, but a more balanced approach should be taken. Without quality statistical know-how on the generation of the most appropriate and accurate phenotypes as well as the most efficient way to exploit additional information into genetic evaluations and the commercial population via an efficient genetic improvement programme, any investment in systems biology to increase genetic gain will be squandered.

77
4.3 Further research requirements/Gaps in knowledge Further research on genomics of digestive efficiency is necessary to get more insight into the genetics of digestive efficiency in pigs. In particular, animal behaviour may be directly associated with energy requirements for maintenance. Several studies have shown that animal behaviour is highly heritable (e.g. Von Felde et al., 1996; Schulze et al., 2003 using feed intake behaviour; Turner et al., 2008,2009; D’Eath et al., 2009 using aggression of pigs) and therefore behavioural traits may be used as indirect selection traits to reduce maintenance requirements and improve animal welfare. Identification of QTL associated with utilisation of energy and protein may be a tool to improve efficiency of utilisation of energy and protein. In particular, the heritability of backfat thickness is high whereas those for lean tissue are moderate (Schulze et al., 2001 and 2002). Furthermore, there have been detected several QTL of body compositions (e.g. Mohrman et al., 2006; Duthie et al., 2008 and 2009). However, the optimal fatness of several dam and sire lines may already achieved by selection so that further reduction in fatness to improve nutrient efficiency may not be an option for most modern pig breeding lines. Consequently, the improvement of feed efficiency by optimization of feed intake capacity with respect to protein deposition curve (considering a minimum lipid deposition) is expected to efficiently improve nutrient efficiency (Roehe et al., 2002 and 2004). Direct selection for feed efficiency, calculated as a ratio of daily feed intake to daily gain, results in disproportional selection pressure (depending on selection intensity and the genetic parameters) on its two components so that a genetic change is unpredictable (Gunsett 1984; Krieter and Presuhn 1986). Consequently, selection for feed efficiency is inefficient. This inefficiency may have not been recognized in the past, because most of the improvement of feed efficiency was due to reduction in backfat thickness, which now approached its optimum in populations largely selected for lean content. In addition, growth rate has been improved substantially due to breeding, which improved feed efficiency. Selection for piglet survival is expected to improve number of weaned piglets and nutrient efficiency per weaned piglet without high nutritional loss due to loss of piglets (Su et al., 2008; Baxter et al., 2009; Roehe et al., 2009). As shown in section 2.1.5, nutrient efficiency improves with increase in number of parities per sows and thus longevity. Based on a literature review, Serenius and Stadler (2005) concluded that sufficient genetic variation exists for effective selection on longevity, and heritability estimates ranged between 0.02 and 0.25. Traits genetically associated with longevity such as leg conformation can be used to improve the accuracy of estimation of breeding values for longevity and thus its genetic progress Herd et al. (2002) showed that there is a decreased enteric methane production per day in animals selected for reduced residual feed intake. Reduced residual feed intake is akin to selection for high feed efficiency as an animal is eating less but maintaining a similar growth rate (high net feed efficiency) and therefore less feed is required to produce a unit of output. Lines were divergently selected for high and low residual feed intake and showed no

78
significant differences for most production traits. This suggests that there are perhaps possibilities for selection of reduced GHG emissions through the selection of animals which use less feed and produce less methane than average to achieve a given level of performance. The estimates of heritability for residual feed intake (RFI) in growing beef animals are moderate (average 0.28; range 0.08-0.43) indicating the existence of genetic variations in the trait (see Herd et al. 2003 for review). Most genetic parameters for RFI were estimated in young cattle in feedlots. Information on RFI in breeding herds is very limited with few studies on adult cattle. Archer et al. (2002) reported an estimate of heritability for mature cows of 0.23. The feeding efficiency of the animal may also have an impact with low-RFI beef cattle eating less than expected for their live weight and growth rate. Angus cattle divergently selected for RFI currently attain the same growth rates but differ by approximately 15% in their voluntary feed intake (Herd et al. 2003). Not only may selection for low RFI have a favourable effect on energy nutrient efficiency, it may also impact on protein nutrient efficiency. Low RFI animals have been shown to have a reduced manure production (15% to 20% less compared to high RFI cattle) and also on the potential quantity of nitrous oxide liberated from these manures. This potential effect on nitrous oxide can be due to a reduction in the total N intake and a greater efficiency of capturing the dietary N (Hegarty et al. 2007). Cammack et al. (2005) reported a rather low heritability of 11% for RFI for ram lambs of a composite breed and 25% for their daily feed intake and 26% for their average daily gain. Francois et al. (2002) reported a more substantial heritability for RFI of 30% in ram lambs of a French composite breed with 43% for daily feed intake and 43% for average daily gain. Therefore, in terms of genetic improvement for feed efficiency and growth rate of the slaughter generation, selection for lean growth rate is preferable to selection for FCR, such that it is not necessary to measure food intake. However, their conclusions may not hold for maternal lines where selection for lean growth rate is likely to lead to a correlated increase in mature feed maintenance/requirements. Although utilising body reserves to support lactation has a favourable effect on methane emissions within the first lactation, it is likely that the longer term reliance on body energy reserves to support lactation will have a longer term unfavourable biological effect and impact on health, fertility and other “fitness” traits in the dairy cow (Friggens and Newbold, 2007; Garnsworthy et al., 2008). Poorer health and fitness in the dairy herd will affect overall systems (and lifetime) nutrient efficiencies as more followers will be required to maintain the herd size as more dairy cows may be culled involuntarily and need to be replaced. In developing breeding programmes that consider lifetime nutrient efficiency it is important that the wider aspects of efficiencies in the whole system are considered as well as the correlated effects on traits both in the breeding goal as well as traits not considered. It is important that breeding goals do not focus on one specific point of efficiency as the wider implications of a favourable genetic change in a specific trait may be cancelled out by wider system impacts. In developing breeding goals it is important to consider a number of factors (i) ability to routinely collect the trait of interest (or strongly correlated proxies) on enough animals in a cost effectively manner; (ii) the scope to improve the trait genetically (i.e., as defined by the

79
genetic standard deviation of the trait); (iii) ability to routinely estimate breeding values and (iv) the relative importance of the trait to the overall breeding goal of the “average” farmer of individual farmers in the case of customised herd/flock indices. This review has highlighted that there is little evidence for major breed and genetic differences in lifetime nutrient efficiencies. However, it should be noted that more recent research may suggest that there evidence of a minor impact of breed/genetics on lifetime nutrient efficiency. Research in this area may be limited in the ability to dissect, in detail the biological partitioning of individual animals, both at a given point in time, but also over their entire lifetime. Key to filling knowledge gaps in this area is the routine availability of accurate and comparable data on traits that underlie nutrient efficiency in livestock species. This requires the development of key and (somewhat) common protocols on the definition of traits, methodology for recording them, and recording them and all relevant times during animals lifetime, across breeds/lines and across systems. Newer genetic/biological technologies (e.g., genomics, sequencing) and ability to record a wider range of traits that relate lifetime nutrient efficiency (e.g., methane production, rumen microbial populations) may help to further understand the mechanisms by which animals utilise nutrients available to them and therefore provide stronger evidence of differences between breeds/animals in nutrient utilisation. 4.4 References Arthur PF and Herd RM. 2005. Efficiency of feed utilisation by livestock – Implications and
benefits of genetic improvement. Canadian Journal of Animal Science 85, 281-290. Blaxter KL and. Clapperton JL 1965 Prediction of the amount of methane produced by
ruminants. British Journal of Nutrition 19, 511-522 Dore J, Pacchart P, Bernalier, A., Goderal, L, Morvan B, Rambaud JC. 1995. Enumeration of
H2 utilising metahanogenic archaea acetogenic and sulphur reducing bacteria from human feces. FEMS Microbiology Ecology 17, 279-284.
El Oufir L, Flourie B, Bruley S., Barry JL, Cloarec D, Bornet F, Galmichie JP. 1996. Relations between transit time fermentation products and hydrogen consuming flora in healthy human. Gut 38, 870-877.
Emmans GC. 1994. Effective energy – a concept of energy utilization applied across species. British Journal of Nutrition 71, 801-821
Emmans GC. 1997. A method to predict the food intake of domestic animals from birth to maturity as a function of time. Journal of Theoretical Biology 186, 189-199.
Flatz G, Czeizelm A, Merneki J, Kuhnau J, Jahn D. 1985. Pulmonary hydrogen and methane excretion following ingestion of an unabsorable carbohydrate: a study of twins. Journal of Paediatric Gastroenterology and Nutrition 4, 936-941.
Goopy JP, Hegarty RS and Dobos RC. 2006. The persistence over time of divergent methane production of lot fed cattle. In ‘Greenhouse gases and animal agriculture: an update’. (Eds CR Soliva,CR, Takahashi,J, Kreuzer), M pp. 111–114. (Elsevier: Amsterdam)
Guan LL, Nkrumah JD, Basarab JA and Moore SS. 2008. Linkage of microbial ecology to phenotype: correlation of rumen microbial ecology to cattle’s feed efficiency. FEMS Microbiology Letters 288, 85–91.
Herd RM and Arthur PF. 2009. Physiological basis for residual feed intake. Journal of Animal Science 87, E64-E71.
Hoque MA and Suzuki K. 2009. Genetics of residual feed intake in cattle and pigs: a review. Asian-Australasian Journal of Animal Sciences 22, 747-755.
Hungate RE .1966. The rumen and it s microbes. Academic Press NY.

80
Joblin KN. 1999. Ruminal acetogens and their potential to lower ruminant methane emissions. Australian Journal of Agricultural Research 50, 1307-1313.
Johnson DE, Hill TM, Carmean BR, Brannie ME, Lodman DW, Ward G. 1991. New perspectives on ruminant methane emissions. In “Energy metabolism of farm animals” (Eds Wenk C and Bossinger M) pp 376-379.
King TS, Elia M, Hunter JO. 1998. Abnormal colonic fermentation in irritable bowel syndrome. Lancet 352, 1187-1189.
Lassey KR, Ulyatt MJ, Martin RJ, Walker CF, Shelton ID. 1997. Methane emissions measured directly from grazing livestock in New Zealand. Atmosphere and Environment 31, 2905–2914.
McKay LF, Eastwood,MA, Brydon WG. 1985. Methane excretion in man- a study of breath, flatus and faeces. Gut 26, 206-214.
Meuwissen THE, Hayes BJ and Goddard ME. 2001 Prediction of total genetic value using genome-wide dense marker maps. Genetics 157, 1819–1829.
Munger A and Kreuzer M. 2008 Absence of persistent methane emission differences in three breeds of dairy cows: Australian Journal of Experimental Agriculture 48, 77-82.
Pinares-Patino CS, Ulyatt MJ, Lassey KR, Barry TN and Holmes CW. 2003. Persistence of differences between sheep in methane emission under generous grazing conditions. Journal of Agricultural Science, Cambridge 140, 227–233.
Pinares-Patino C, Vlaming B, Cavanagh A, Molano G and Clark H. 2005. Persistence of dairy cows in animal-to-animal variation in methane emission. In ‘Working papers of the 2nd international conference on greenhouse gases and animal agriculture’. (Eds Soliva,CR, J Takahashi, J, Kreuzer, M) pp. 401–404. (Institute of Animal Science: ETH Zurich,Switzerland)
Pinares-Patiño CS, Waghorn GC, Machmuller A, Vlaming B, Molano G, Cavanagh A and Clark H. 2007. Methane emissions and digestive physiology of non-lactating dairy cows fed pasture forage Canadian Journal of Animal Science 87, 601-613.
Pitt P, De Bruijn, KM, Beeching MR, Golberg E, Blendis LM. 1980. Studies on breath methane; the effect of ethnic origins and lactulose. Gut 21, 951-959.
Robertson LR and Waghorn GC. 2002. Dairy industry perspectives on methane emissions and production from cattle fed pasture or total mixed rations in New Zealand. Proceedings of the New Zealand Society of Animal Production 62, 213–218.
Sellner EM, Kim JW, McClure MC, Taylor KH, Schnabel RD and Taylor JF. 2007. Application of genomic information in livestock. Journal of Animal Science 85, 3148-3158.
Yanez DR, Macias B, Pinloche E, Newbold CJ. 2010. The persistence of bacterial and methanogenic archaeal communities residing in the rumen of young lambs FEMS Microbiol Ecol On line ahead of publication DOI:10.1111/j.1574-6941.2010.00852.x
Van Es AJH 1975. Heterotrophe production in animals. Pp. 105-127 in (GJ Vervelde, ed.) Productivity in Biological Systems (in Dutch). Wageningen: Centrum voor Landbouw publikaties and Landbouwdocumentatie.
Vlaming JB, Lopez-Villalobos N, Brookes IM, Hoskin SO, Clark H. 2008. Within- and between-animal variance in methane emissions in non-lactating dairy cows Australian Journal of Experimental Agriculture 48, 124–127
Whittemore CT. 2006. Energy value of feedstuffs for pigs. Pp. 317-342 in (I. Kyriazakis and .T. Whittemore, eds.) Whittemore’s Science and Practice of Pig Prduction. Blackwell Publishing.
Zhou M, Herandez-Sanabria E and Guan LL. 2009. Assessment of the microbial ecology of ruminant methanogens in cattle with different feed efficiencies. Applied and Environmental Microbiology, 75, 6524-6533.

81
APPENDIX I The authors gratefully acknowledge the information, discussion, contributions and suggestions from the following persons: Dr. S. Avendaňo (Aviagen Ltd) Dr. S. Bishop (University of Edinburgh) Dr. A Carston (AFBINI) Dr. J. Conington (SAC) Dr. P Garland (ALPHARMA) Dr. K. Gooderham (NTLWorld) Mr. E. Hegarty (Norfolk Geese) Ms. J. Howie (Aviagen Ltd) Dr. J. Houdijk (SAC) Dr. J. Hyslop (SAC) Dr. D. Martin (Cherry Valley Ltd) Mr. J. McAdam (Aviagen Ltd) Dr. C. Morgan (SAC) Dr. J. Rooke (SAC) Prof J. Oldham (SAC) Dr. J. Vipond (SAC) Dr. G. Walling (JSR Genetics Ltd)
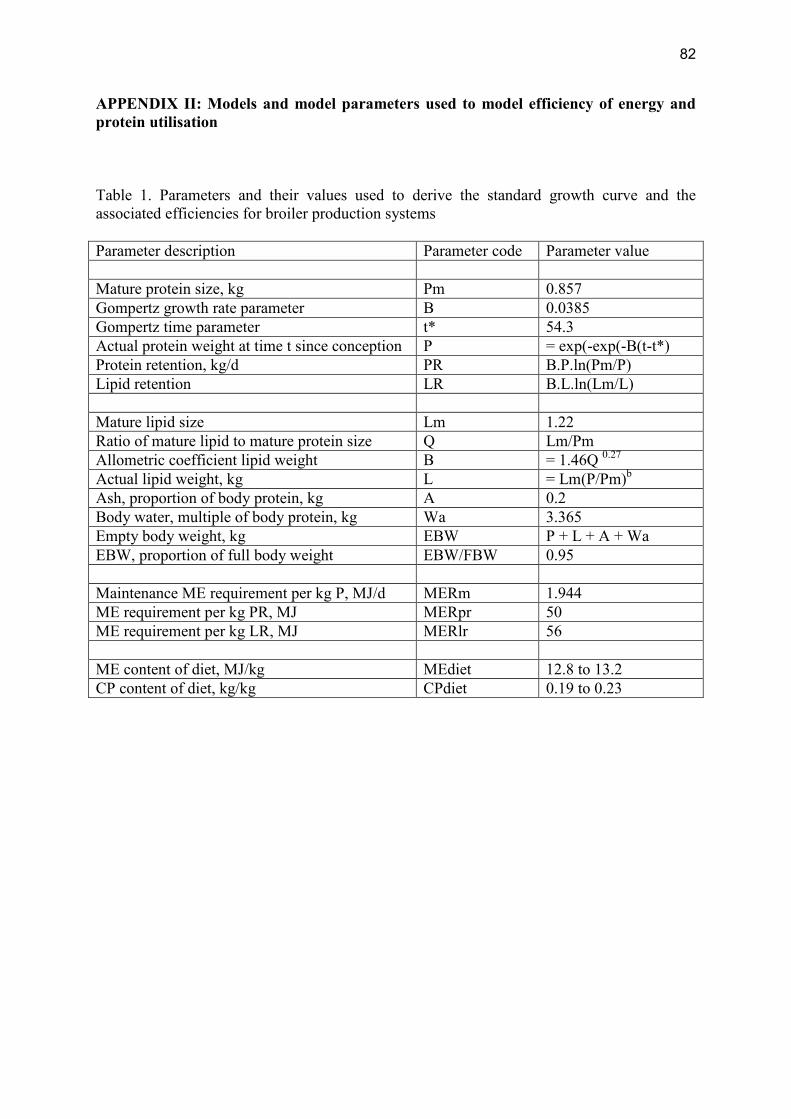
82
APPENDIX II: Models and model parameters used to model efficiency of energy and protein utilisation Table 1. Parameters and their values used to derive the standard growth curve and the associated efficiencies for broiler production systems Parameter description Parameter code Parameter value Mature protein size, kg Pm 0.857 Gompertz growth rate parameter B 0.0385 Gompertz time parameter t* 54.3 Actual protein weight at time t since conception P = exp(-exp(-B(t-t*) Protein retention, kg/d PR B.P.ln(Pm/P) Lipid retention LR B.L.ln(Lm/L) Mature lipid size Lm 1.22 Ratio of mature lipid to mature protein size Q Lm/Pm Allometric coefficient lipid weight B = 1.46Q 0.27 Actual lipid weight, kg L = Lm(P/Pm)b Ash, proportion of body protein, kg A 0.2 Body water, multiple of body protein, kg Wa 3.365 Empty body weight, kg EBW P + L + A + Wa EBW, proportion of full body weight EBW/FBW 0.95 Maintenance ME requirement per kg P, MJ/d MERm 1.944 ME requirement per kg PR, MJ MERpr 50 ME requirement per kg LR, MJ MERlr 56 ME content of diet, MJ/kg MEdiet 12.8 to 13.2 CP content of diet, kg/kg CPdiet 0.19 to 0.23
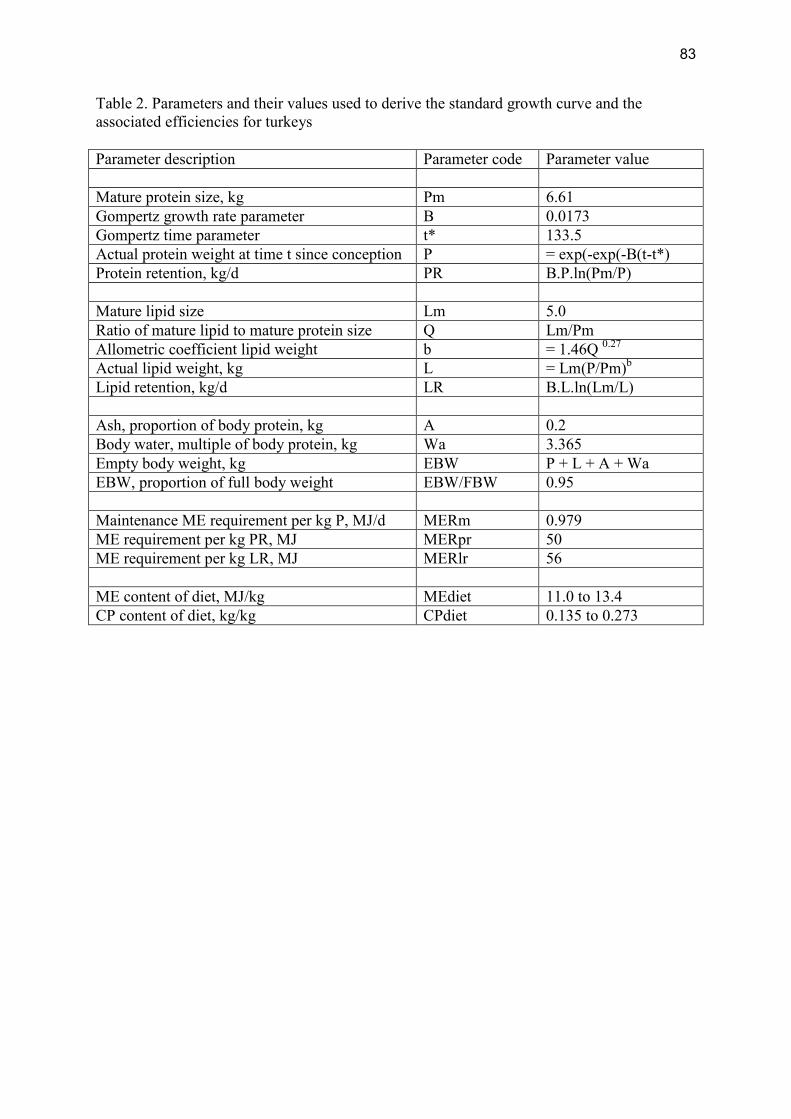
83
Table 2. Parameters and their values used to derive the standard growth curve and the associated efficiencies for turkeys Parameter description Parameter code Parameter value Mature protein size, kg Pm 6.61 Gompertz growth rate parameter B 0.0173 Gompertz time parameter t* 133.5 Actual protein weight at time t since conception P = exp(-exp(-B(t-t*) Protein retention, kg/d PR B.P.ln(Pm/P) Mature lipid size Lm 5.0 Ratio of mature lipid to mature protein size Q Lm/Pm Allometric coefficient lipid weight b = 1.46Q 0.27 Actual lipid weight, kg L = Lm(P/Pm)b Lipid retention, kg/d LR B.L.ln(Lm/L) Ash, proportion of body protein, kg A 0.2 Body water, multiple of body protein, kg Wa 3.365 Empty body weight, kg EBW P + L + A + Wa EBW, proportion of full body weight EBW/FBW 0.95 Maintenance ME requirement per kg P, MJ/d MERm 0.979 ME requirement per kg PR, MJ MERpr 50 ME requirement per kg LR, MJ MERlr 56 ME content of diet, MJ/kg MEdiet 11.0 to 13.4 CP content of diet, kg/kg CPdiet 0.135 to 0.273

84
Table 3. Parameters and their values used to derive the standard growth curve and the associated efficiencies for male geese Parameter description Parameter code Parameter value Mature protein size, kg Pm 1.18 Gompertz growth rate parameter B 0.0324 Gompertz time parameter t* 73.3 Actual protein weight at time t since conception P = exp(-exp(-B(t-t*) Protein retention, kg/d PR B.P.ln(Pm/P) Mature lipid size Lm 3.5 Ratio of mature lipid to mature protein size Q Lm/Pm Allometric coefficient lipid weight B = 1.46Q 0.27 Actual lipid weight, kg L = Lm(P/Pm)b Lipid retention, kg/d LR B.L.ln(Lm/L) Ash, proportion of body protein, kg A 0.2 Body water, multiple of body protein, kg Wa 3.365 Empty body weight, kg EBW P + L + A + Wa EBW, proportion of full body weight EBW/FBW 0.95 Maintenance ME requirement per kg P, MJ/d MERm 1.67 ME requirement per kg PR, MJ MERpr 50 ME requirement per kg LR, MJ MERlr 56 ME content of diet, MJ/kg MEdiet 11.5 CP content of diet, kg/kg CPdiet 0.16 to 0.20
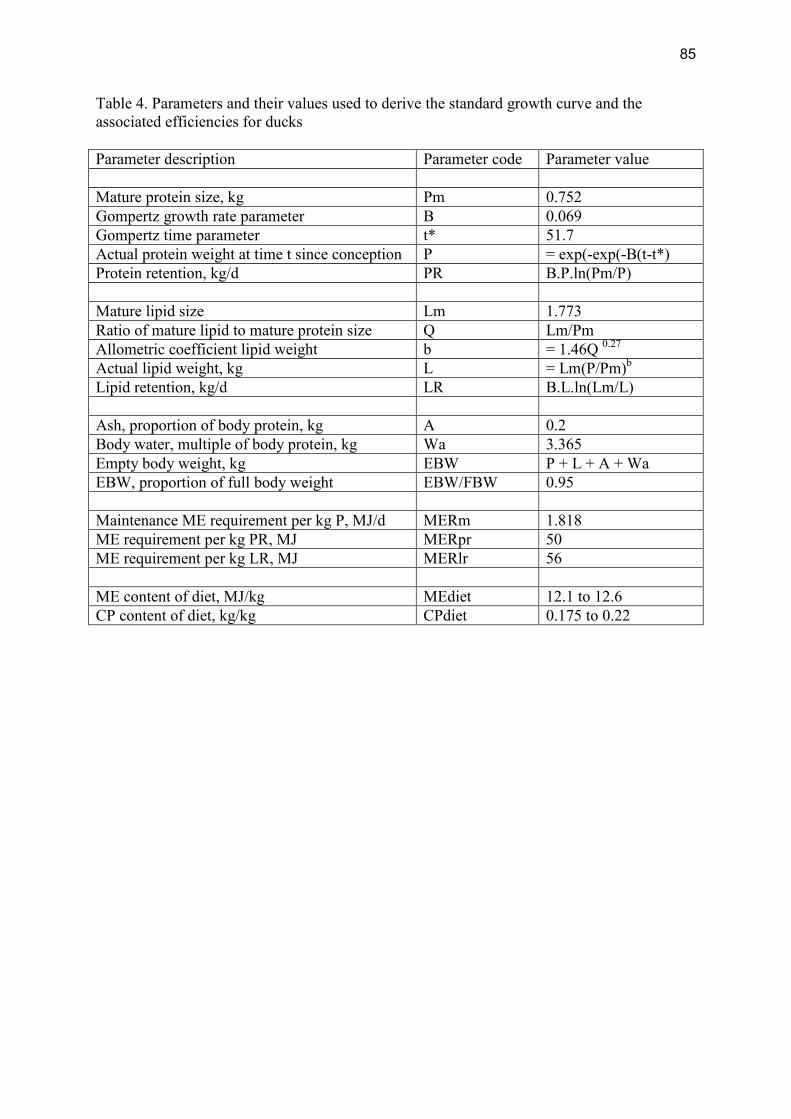
85
Table 4. Parameters and their values used to derive the standard growth curve and the associated efficiencies for ducks Parameter description Parameter code Parameter value Mature protein size, kg Pm 0.752 Gompertz growth rate parameter B 0.069 Gompertz time parameter t* 51.7 Actual protein weight at time t since conception P = exp(-exp(-B(t-t*) Protein retention, kg/d PR B.P.ln(Pm/P) Mature lipid size Lm 1.773 Ratio of mature lipid to mature protein size Q Lm/Pm Allometric coefficient lipid weight b = 1.46Q 0.27 Actual lipid weight, kg L = Lm(P/Pm)b Lipid retention, kg/d LR B.L.ln(Lm/L) Ash, proportion of body protein, kg A 0.2 Body water, multiple of body protein, kg Wa 3.365 Empty body weight, kg EBW P + L + A + Wa EBW, proportion of full body weight EBW/FBW 0.95 Maintenance ME requirement per kg P, MJ/d MERm 1.818 ME requirement per kg PR, MJ MERpr 50 ME requirement per kg LR, MJ MERlr 56 ME content of diet, MJ/kg MEdiet 12.1 to 12.6 CP content of diet, kg/kg CPdiet 0.175 to 0.22
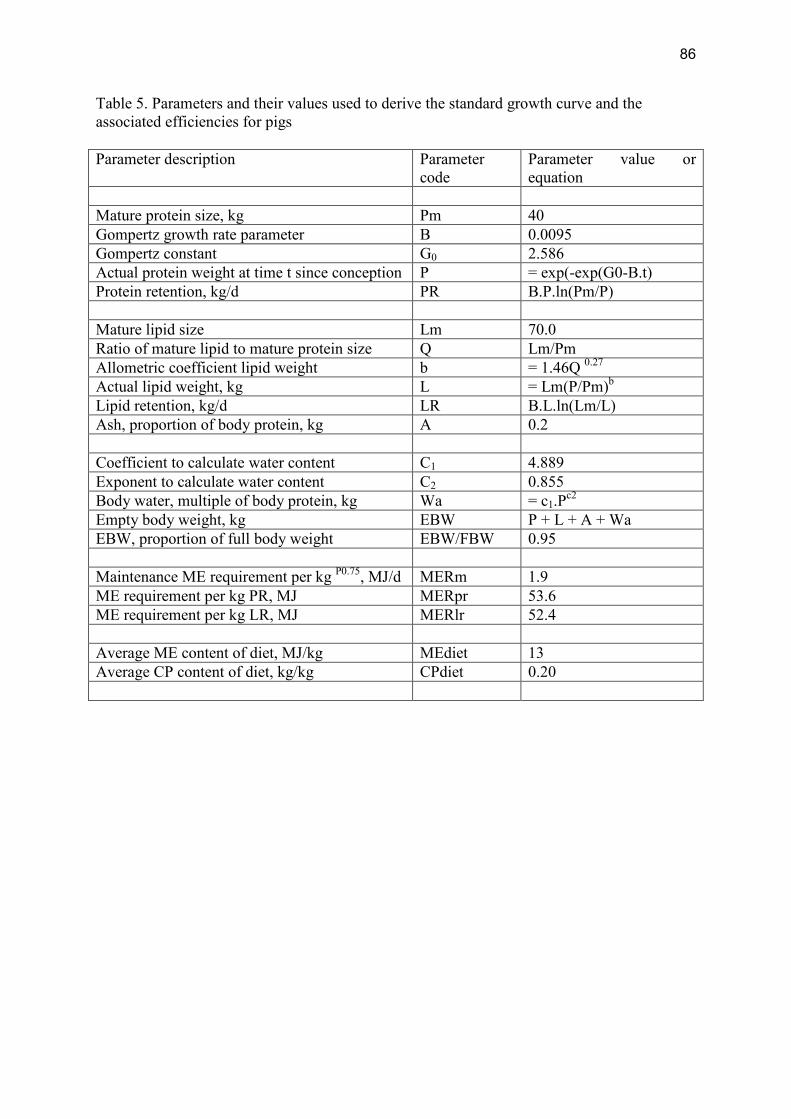
86
Table 5. Parameters and their values used to derive the standard growth curve and the associated efficiencies for pigs Parameter description Parameter
code Parameter value or equation
Mature protein size, kg Pm 40 Gompertz growth rate parameter B 0.0095 Gompertz constant G0 2.586 Actual protein weight at time t since conception P = exp(-exp(G0-B.t) Protein retention, kg/d PR B.P.ln(Pm/P) Mature lipid size Lm 70.0 Ratio of mature lipid to mature protein size Q Lm/Pm Allometric coefficient lipid weight b = 1.46Q 0.27 Actual lipid weight, kg L = Lm(P/Pm)b Lipid retention, kg/d LR B.L.ln(Lm/L) Ash, proportion of body protein, kg A 0.2 Coefficient to calculate water content C1 4.889 Exponent to calculate water content C2 0.855 Body water, multiple of body protein, kg Wa = c1.Pc2 Empty body weight, kg EBW P + L + A + Wa EBW, proportion of full body weight EBW/FBW 0.95 Maintenance ME requirement per kg P0.75, MJ/d MERm 1.9 ME requirement per kg PR, MJ MERpr 53.6 ME requirement per kg LR, MJ MERlr 52.4 Average ME content of diet, MJ/kg MEdiet 13 Average CP content of diet, kg/kg CPdiet 0.20

87
Table 6. Parameters and their values used to derive the efficiencies and FCR for lamb production systems Characteristic Values used in model Ewe live weight (kg) 75 NE maintenance, ewe, MJ/W 3/4 0.275 CP maintenance ewe kg/W 3/4 0.005 NE content milk (MJ) 4.4 Protein content milk (g/kg) 55 Feed ME content 10.2 to 11.1 Overall efficiency ME utilization 0.6 Lamb birth weight (kg) 4 Lamb weaning age (weeks) 16 Lamb finishing age (weeks) 28 Lamb finishing weight (kg) 42.5 NE in finished lamb (MJ) 848 CP in finished lamb (kg) 6 ME costs rearing ewe (MJ) 7875 CP costs rearing ewe (kg) 93 CP in culled ewe (kg) 8 NE in culled ewe (MJ) 750
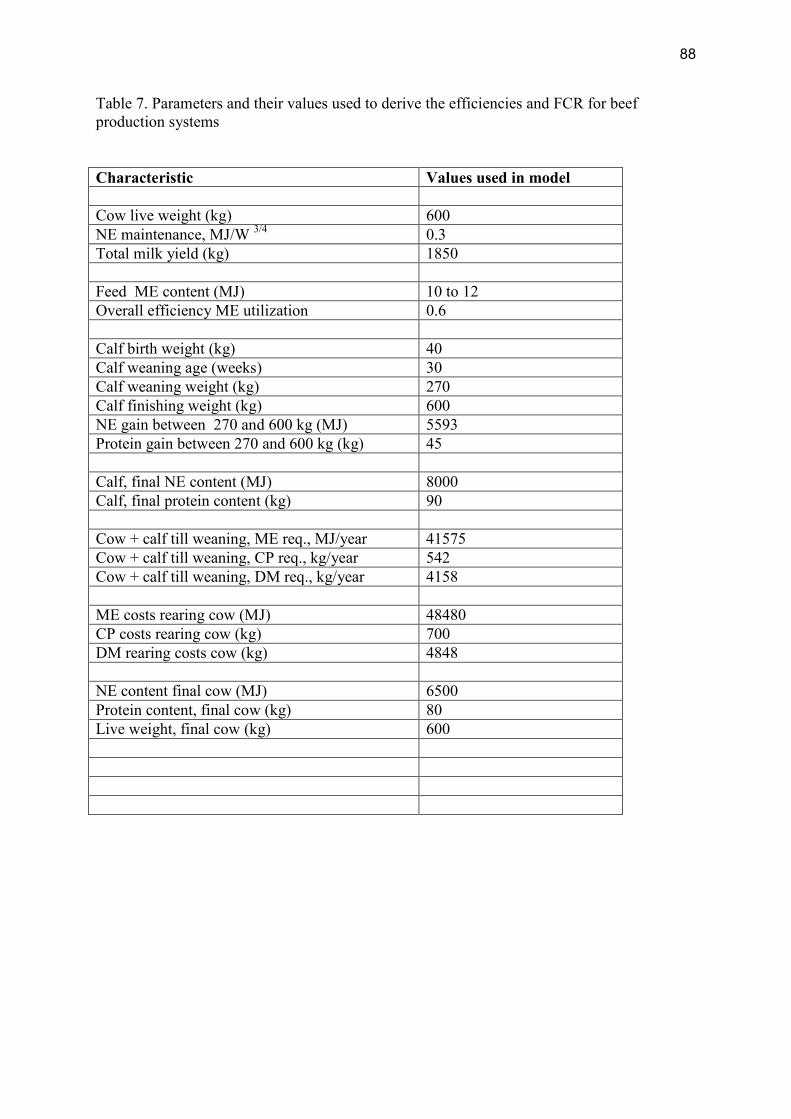
88
Table 7. Parameters and their values used to derive the efficiencies and FCR for beef production systems Characteristic Values used in model Cow live weight (kg) 600 NE maintenance, MJ/W 3/4 0.3 Total milk yield (kg) 1850 Feed ME content (MJ) 10 to 12 Overall efficiency ME utilization 0.6 Calf birth weight (kg) 40 Calf weaning age (weeks) 30 Calf weaning weight (kg) 270 Calf finishing weight (kg) 600 NE gain between 270 and 600 kg (MJ) 5593 Protein gain between 270 and 600 kg (kg) 45 Calf, final NE content (MJ) 8000 Calf, final protein content (kg) 90 Cow + calf till weaning, ME req., MJ/year 41575 Cow + calf till weaning, CP req., kg/year 542 Cow + calf till weaning, DM req., kg/year 4158 ME costs rearing cow (MJ) 48480 CP costs rearing cow (kg) 700 DM rearing costs cow (kg) 4848 NE content final cow (MJ) 6500 Protein content, final cow (kg) 80 Live weight, final cow (kg) 600

89
APPENDIX III: Collected references Acamovic T. 2002. Nutrient requirements and standards for poultry. Report of the British
Society of Animal Science Nutritional Standards Working Group, March 2002. AFRC (Agricultural and Food Research Council). 1993. Energy and Protein Requirements of
Ruminants. An advisory manual prepared by the AFRC Technical Committee on Responses to Nutrients. CAB International, Wallingford, UK.
Agnew RE, Yan T, France J, Kebreab E and Thomas C. 2004. Energy requirement and supply. In Feed into Milk. A new applied feeding system for dairy cows (ed. C Thomas), pp. 11-20. Nottingham University Press, Nottingham, UK.
Aguilera JF and Prieto C. 1991. Methane production in goats given diets based on lucerne hay and barley. Archives of Animal Nutrition 41, 77-84.
Aikman PC, Reynolds CK, Beever DE. 2008. Diet digestibility, rate of passage, and eating and rumination behavior of Jersey and Holstein cows. Journal of Dairy Science 91, 1103-1114.
Altan O, Aguz I, Akbas Y and Aksit M. 2004. Genetic variability of residual feed consumption (RFC) and its relationships with some production traits and fear response in Japanese quail hens (Coturnic coturnix japonica). Archiv fur Geflügelkunde 68, 223-229.
Amoah EA, Gelaye S, Guthrie P, Rexroad CE. 1996. Breeding season and aspects of reproduction of female goats. Journal of Animal Science 74, 723-728.
ARC (Agricultural Research Council). 1980. The Nutrient Requirements of Ruminant Livestock. CAB International, Wallingford, UK.
Baéza E, Rideau N, Chartrin P, Davail S, Hoo-Paris R, Mourot J, Guy G, Bernadet MD and Hermier D. 2005. Canards de Barbarie, Pékin et leurs croisements: aptitude à l'engraissement. Productions Animales 18, 131-141.
Ball AJ, Thompson JM, Alston CL, Blakely AR, Hinch GN. 1998. Changes in maintenance energy requirements of mature sheep fed at different levels of feed intake at maintenance, during weight loss and realimentation. Livestock Production Science 53, 191-204.
Baxter EM, Jarvis S, Sherwood L, Robson SK, Ormandy E, Farish M, Smurthwaite KM, Roehe R, Lawrence AB, Edwards SA. 2009. Indicators of piglet survival in an outdoor farrowing system. Livestock Science 124, 266-276
Beaver EE, Williams JE, Miller SJ, Hancock DL, Hannah SM and O'Connor DL. 1989. Influence of breed and diet on growth, nutrient digestibility, body composition and plasma hormones of Brangus and Angus steers. Journal of Animal Science. 67, 2415-2425
Berry DP, Horan B, O'Donovan M, Buckley F, Kennedy E, McEvoy M, Dillon P. 2007. Genetics of grass dry matter intake, energy balance, and digestibility in grazing Irish dairy cows. Journal of Dairy Science 90, 4835-4845.
Birnie, JW, Agnew RE, Gordon FJ. 2000. The influence of body condition on the fasting energy metabolism of nonpregnant, nonlactating dairy cows. Journal of Dairy Science 83, 1217-1223.

90
Blake RW, Custodia AA and Howard WH. 1986. Comparative feed-efficiency of Holstein and Jersey cows. Journal of Dairy Science 69, 1302-1308.
Blaxter KL and. Clapperton JL. 1965. Prediction of the amount of methane produced by ruminants. British Journal of Nutrition, 19, 511-522
Boadi DA and Wittenberg KM. 2002. Methane production from dairy and beef heifers fed forages differing in nutrient density using the sulphur hexafluoride (SF6) tracer gas technique. Canadian Journal of Animal Science 82, 201-206.
Bisset SA, Morris CA, McEwan JC and Vlassoff A. 2001. Breeding sheep in New Zealand that are less reliant on anthelmintics to maintain health and productivity. New Zealand Veterinary Journal 49, 236–246.
Blaxter KL. 1962. The fasting metabolism, of adult wether sheep. British Journal of Nutrition 16, 615-626.
Blaxter KL, Clapperton JL and Wainman FW. 1966. The extent of differences between six British sheep breeds in their metabolism, feed intake and utilization and resistance to stress. British Journal of Nutrition 20, 283-294.
Bottje WG and Carstens GE. 2009. Association of mitochondrial function and feed efficiency in poultry and livestock species. Journal of Animal Science 87, E48-E63.
BPEX. 2009. Pig Yearbook 2009. BPEX, Milton Keynes, UK. BPEX. 2009b. Defining the benefits of new genotypes. BPEX, Milton Keynes, UK. Bordas A and Merat P. 1991. Selection divergente pour la consommation alimentaire
residuelle de la poule en periode de ponte: reponse au taux proteique de l’aliment. Genetics Selection Evolution 23, 249-256.
Borin K, Lindberg JE and Ogle RB. 2006. Digestibility and digestive organ development in indigenous and improved chickens and ducks fed diets with increasing inclusion levels of cassava leaf meal. Journal of Animal Physiology and Animal Nutrition 90, 230-237.
Braastad BO and Katle J. 1989. Behavioral differences between laying hen populations selected for high and low efficiency of food utilization. British Poultry Science 30, 533-544.
Bradford GE, Burfening PJ and Cartwright TC. 1989. Evaluation of Production and Reproduction of Sheep, Goat and Alpaca Genotypes in the Small Ruminant Collaborative Research Support Program. Journal of Animal Science 67, 3058-3067.
Bruininx EMAM, Schellingerhout AB, Lensen EGC, van der Peet-Schwering CMC, Schrama JW, Everts, den Hartog LA and Beynen AC. 2002. Associations between individual food intake characteristics and indicators of gut physiology of group-housed weanling pigs differing in genotype. Animal Science 75, 103-113.
Buyse J, Michels H, Vloeberghs J, Saevels P, Aerts JM, Ducro B, Berckmans D and Decuypere E. 1998. Energy and protein metabolism between 3 and 6 weeks of age of male broiler chickens selected for growth rate or for improved food efficiency. British Poultry Science 39, 264-272.
Buyse J, Leenstra FR, Zeman M, Rahimi G and Decuypere E. 1999. A comparative study of different selection strategies to breed leaner meat-type poultry. Poultry and Avian Biology Reviews 10, 121-142.

91
Campo JL, Gil MG, Torres O and Davila SG. 2001. Association between plumage condition and fear and stress levels in five breeds of chickens. Poultry Science 80, 549-552.
Carre B, Mignon-Grasteau S, Juin, H. 2008. Breeding for feed efficiency and adaptation to feed in poultry. Worlds Poultry Science Journal 64, 377-390.
Clayton GA and Powell JC. 1979. Growth, food conversion, carcass yields and their heritabilities in ducks (Anas platyrhynchos). British Poultry Science 20, 121-127.
Collin A, Malheiros RD, Moraes VMB, Van As P, Darras VM, Taouis M, Decuypere E and Buyse J. 2003. Effects of dietary macronutrient content on energy metabolism and uncoupling protein mRNA expression in broiler chickens. British Journal of Nutrition 90, 261-269.
Collin A, Swennen, Q,Skiba-Cassy, S Buyse, J, Chartrin P, Le Bihan-Duval E, Crochet S, Duclos M, Joubert R, Davies G, Genini S, Bishop SC and Giuffra E. 2009. An assessment of opportunities to dissect host genetic variation in resistance to infectious diseases in livestock. Animal 3, 415-436.
Conington J, Bishop SC, Grundy B, Waterhouse A, Simm G. 2001. Multi-trait selection indexes for sustainable UK hill sheep production. Animal Science 73, 413-423.
Conington J, Hosie B, Nieuwhof GJ, Bishop SC. 2008. Breeding for resistance to footrot - the use of hoof lesion scoring to quantify footrot in sheep. Veterinary Research Communications 32, 583-589.
CSIRO (Commonwealth Scientific and Industrial Research Organisation). 1990. Feeding Standards for Australian Livestock: Ruminants. Compiled by the Ruminants Subcommittee of the Standing Committee on Agriculture. CSIRO, East Melbourne, Australia.
Custodio AA, Blake RW, Dahm PF, Cartwright TC, Schelling GT and Coppock CE. 1983. Relationships between measures of feed efficiency and transmitting ability for milk of Holstein cows. Journal of Dairy Science 66, 1937-1946.
Dawson LER and Steen RWJ. 1998. Estimation of maintenance energy requirements of beef cattle and sheep. Journal of Agricultural Science, Cambridge 131, 477-485.
D’Eath, RB, Roehe R, Turner SP, Ison SH, Farish M, Jack MC and Lawrence AB. 2009. Genetics of animal temperament: aggressive behaviour at mixing is genetically associated with the response to handling in pigs. Animal 3, 1544-1554.
De Campeneere S, Fiems LO and Boucque ChV. 2001. Energy and protein requirements of double-muscled Belgian Blue bulls. Animal Feed Science and Technology 90, 153-167.
Decuypere E and Tesseraud S. 2009 Regulation of fatty acid oxidation in chicken (Gallus gallus): Interactions between genotype and diet composition. Comparative Biochemistry and Physiology B – Biochemistry and Molecular Biology 153, 171-177.
De Vries A.G. and Kanis E. (1992) A growth-model to estimate economic values for food-intake capacity in pigs. Animal Production 55, 241-246.
Dickerson GE. 1978. Animal size and efficiency: basic concepts. Animal Production 27: 367-379.
Dore J, Pacchart P, Bernalier A., Goderal L, Morvan B, and Rambaud JC. 1995. Enumeration of H2 utilising methanogenic archaea acetogenic and sulphur reducing bacteria from human feces. FEMS Microbiology Ecology, 17, 279-284.

92
Duthie C, Simm G, Doeschl-Wilson A, Kalm E, Knap PW and Roehe R. 2008. Quantitative trait loci for chemical body composition traits in pigs and their positional associations with body tissues, growth and feed intake. Animal Genetic 39: 130-140.
Duthie C, Simm G, Pérez-Enciso M, Doeschl-Wilson A, Kalm E, Knap PW and Roehe R. 2009. Genomic scan for quantitative trait loci of chemical and physical body composition and deposition on pig chromosome X considering the pseudoautosomal region. Genet. Sel. Evol. 41, 27 (Open Access)
Eady SJ, Woolaston RR and Barger IA. 2003. Comparison of genetic and nongenetic strategies for control of gastrointestinal nematodes of sheep. Livestock Production Science 81, 11 –23.
Eissen JJ. 2000. Breeding for feed intake capacity in pigs. Doctoral thesis, University Wageningen, Netherlands.
Ellis JE, Kebreab E, Odongo NE, McBride BW, Okine EK and France J. 2007. Prediction of methane production from dairy and beef cattle. Journal of Dairy Science 90, 3456-3467.
El Oufir, L, Flourie B, Bruley S, Barry JL, Cloarec, D Bornet F and Galmichie JP. 1996. Relations between transit time fermentation products and hydrogen consuming flora in healthy human Gut 38, 870-877.
Emmans GC. 1994. Effective energy – a concept of energy utilization applied across species. British Journal of Nutrition 71, 801-821
Emmans GC. 1997. A method to predict the food intake of domestic animals from birth to maturity as a function of time. Journal of Theoretical Biology 186, 189-199.
Emmerson D. 2003. Breeding objectives and selection strategies for broiler production in (W.M Muir and S.E. Aggrey, eds.) Poultry Genetics, Breeding and Biotechnology. CABI Publishing, Wallingford, UK.
Fan YK, Croom WJ, Christensen VL, Bird AR, Daniel LR, McBride BW, Eisen EJ. 1998. Apparent energetic efficiency of jejunal glucose uptake in young adult turkeys selected for rapid growth. Canadian Journal of Animal Science 78, 301-306.
Fan YK, Croom J, Christensen VL, Black BL, Bird AR, Daniel LR, McBride BW, Eisen EJ. 1997. Jejunal glucose uptake and oxygen consumption in turkey poults selected for rapid growth. Poultry Science 76, 1738-1745.
Farhat A, Normand L, Chavez ER, Touchburn SP. 1998. Nutrient digestibility in food waste ingredients for Pekin and Muscovy ducks. Poultry Science 77, 1371-1376.
Farhat A. and Chavez ER. 2001. Metabolic studies on lean and fat ducks selected for breast muscle thickness measured by ultrasound scanning. Poultry Science 80, 585-591.
Ferris CP, Gordon FJ, Patterson DC, Porter, MG and Yan T. 1999. The effect of genetic merit and concentrate proportion in the diet on nutrient utilization by lactating dairy cows. Journal of Agricultural Science 132, 483-490.
Fevrier C, Bourdon D and Aumaitre A. 1992. Effects of level of dietary fiber from wheat bran on digestibility of nutrients, digestive enzymes and performance in the European Large White and Chinese Mei Shan pig. Journal of Animal Physiology and Animal Nutrition 68, 60-72.

93
Fisher C. 2000. Advances in feed evaluation for poultry. Pp. 243-268 in P.J. Moughan, M.W.A. Verstegen and M.I. Visser-Teyneveld (Eds.) Feed Evaluation: Principles and Practice. Wageningen Pers, Wageningen, The Netherlands.
Flatz G, Czeizelm A, Merneki J, Kuhnau J, Jahn D. 1985. Pulmonary hydrogen and methane excretion following ingestion of an unabsorbable carbohydrate: a study of twins . Journal of Paediatric Gastroenterology and Nutrition 4, 936-941.
Hungate RE. 1966. The rumen and it s microbes. Academic Press NY. Flock DK, Laughlin K.F and Bentley J. 2005. Minimizing losses in poultry breeding and
production: how breeding companies contribute to poultry welfare. Worlds Poultry Science Journal 61, 227-237.
Freetly HC, Nienaber JA, Leymaster KA and Jenkins TG. 1995. Relationships among heat-production, body-weight, and age in Suffolk and Texel ewes. Journal of Animal Science 73, 1030-1037.
Freetly HC, Nienaber JA and Brown-Brandl T. 2002. Relationships among heat production, body weight, and age in Finnsheep and Rambouillet ewes. Journal of Animal Science 80, 825-832.
Freire JPB, Dias RIM, Cunha LF and Aumaitre A 2003. The effect of genotype and dietary fibre level on the caecal bacterial enzyme activity of young piglets: digestive consequences. Animal Feed Science and Technology 106. 119-130.
Geraert PA, MacCleod MG and LeClercq, YDB. 1988. Energy metabolism in genetically fat and lean chickens: Diet- and cold-Induced thermogenesis. Journal of Nutrition 118, 1232-1239.
Geraert PA, MacLeod MG, Larbier M and Leclercq B; 1990. Nitrogen metabolism in genetically fat and lean chickens. Poultry Science 69, 1911-1921.
Gille U, Salomon FV and Ronnert J. 1999. Growth of the digestive organs in ducks with considerations on their growth in birds in general. British Poultry Science 40, 194–202
Gipson TA and Grossman M. 1990. Lactation Curves in Dairy Goats - A Review. Small Ruminant Research 3, 383-396.
Givens DI and Moss AR. 1994. Effect of Breed, Age and Bodyweight of Sheep on the Measurement of Apparent Digestibility of Dried Grass. Animal Feed Science and Technology 49, 155-162.
Goonewardene LA, Whitmore W, Jaeger S, Borchert T, Okine E, Ashmawy O and, Emond S. 1997. Effect of prebreeding maintenance diet on subsequent reproduction by artificial insemination in Alpine and Saanen goats. Theriogenology 48, 151-159.
Goopy JP, Hegarty RS, Dobos RC. 2006. The persistence over time of divergent methane production of lot fed cattle. In ‘Greenhouse gases and animal agriculture: an update’. (Eds CR Soliva,CR, Takahashi,J, Kreuzer), M pp. 111–114. (Elsevier: Amsterdam)
Guan, LL, Nkrumah, JD, Basarab JA and Moore SS. 2008. Linkage of microbial ecology to phenotype: correlation of rumen microbial ecology to cattle’s feed efficiency. FEMS Microbiol. Lett. 288:85–91
Gunsett FC. 1984. Linear index selection to improve traits defined as ratios. Journal of Animal Science 59:1185-1193.

94
Guy G, Rousselot-Pailley D and Rouvier, R. 1998. Description de la croissance de canettes mulardes. Annales de Zootechnie 45, 303-310.
Guy G, Hermier D, Davail S, Bely M, Andre J.P. and Hoo-Paris R. 1999. Meat production and force-feeding ability of different type of ducks. Proceedings of the 1st World Waterfowl Conference, Taichung, Taiwan, pp 462-468.
Havenstein GBH, Ferket PR and Qureshi MA. 2003. Growth, livability, and feed conversion of 1957 versus 2001 broilers when fed representative 1957 and 2001 broiler diets. Poultry Science 82: 1500-1508.
Havenstein GB , Ferket PR, Grimes JL, Qureshi MA and Nestor KE. 2007. Comparison of the performance of 1966-versus 2003-type turkeys when fed representative 1966 and 2003 turkey diets: Growth rate, livability, and feed conversion. Poultry Science 86, 232-240.
Hegarty RS, Goopy JP, Herd RM and McCorkell. 2007. Cattle selected for lower residual feed intake have reduced methane production. Journal of Animal Science 85, 1479-1486.
Henken AM, Vanderhel W, Brandsma HA and Verstegen MWA. 1991. Differences in Energy-Metabolism and Protein Retention of Limit-Fed Growing Pigs of Several Breeds. Journal of Animal Science 69, 14431453.
Hennig U, Metges CC, Berk A, Tuchscherer A and Kwella M. 2004. Relative ileal amino acid flows and microbial counts in intestinal effluents of Goettingen Minipigs and Saddleback pigs are not different. Journal of Animal Science 82, 1976-1985.
Herd RM. 1995. Effect of divergent selection for yearling growth rate on the maintenance feed requirements of mature Angus cows. Livestock Production Science 41, 39-49.
Herd RM, Speck PA and Wynn PC. 1991. Feed requirements for maintenance and growth of one year old Angus steers selected for either fast or slow yearling growth-rate. Australian Journal of Experimental Agriculture 31, 591-595.
Hodgkinson SM, Schmidt M and Ulloa N. 2008. Comparison of the digestible energy content of maize, oats and alfalfa between the European wild boar (Sus scrofa L.) and Landrace x Large White pig (Sus scrofa domesticus). Animal Feed Science and Technology 144, 167-173.
Horan B, Faverdin P, Delaby L, Rath M and Dillon P. 2006. The effect of strain of Holstein-Friesian dairy cow and pasture-based system on grass intake and milk production. Animal Science 82, 435-444.
Hotovy S.K, Johnson KA, Johnson DE, Carstens GE, Bourdon RM and Seidel GE. 1991. Variation Among Twin Beef-Cattle in Maintenance Energy-Requirements. Journal of Animal Science 69, 940-946.
Howes JR, Hentges JF and Davis GK. 1963. Comparative digestive powers of Hereford and Brahman cattle, Journal of Animal Science 22, 22.
Huff GR, Huff WE, Balog JM, Rath NC, Anthony NB and Nestor KE. 2005. Stress response differences and disease susceptibility reflected by heterophil to lymphocyte ratio in turkeys selected for increased body weight. Poultry Science 84, 709-717.
Ingvartsen KL and Weisbjerg MR. 1993. Jersey cows have a higher feed intake capacity and higher rate of passage than Friesian cows. Archives of Animal Breeding 36, 495-498.
INRA (Institute National de la Recherche Agronomique). 1989. Ruminant Nutrition,

95
Recommended Allowances & Feed Tables (ed R Jarrige). John Libbey and Co Ltd, London.
Isac MD, Garcia MA, Aguilera JF and Alcaide EM. 1994. A Comparative-Study of Nutrient Digestibility, Kinetics of Digestion and Passage and Rumen Fermentation Pattern in Goats and Sheep Offered Medium Quality Forages at the Maintenance Level of Feeding. Archives of Animal Nutrition 46, 37-50.
Jenet A, Yimegnuhal A, Fernandez-Rivera S, Tegegne A, Osuji PO, McCrabb G and Kreuzer M. 2004. Long-term response to feeding level in lactational performance of Boran (Bos indicus) and Boran X Holstein cows. Animal Science 78, 331-343.
Jenkins TG and Ferrell CL. 2007. Daily dry matter intake to sustain body weight of mature, nonlactating, nonpregnant cows. Journal of Animal Science 85, 1787-1792.
Joblin KN. 1999. Ruminal acetogens and their potential to lower ruminant methane emissions. Australian Journal of Agricultural Research 50, 1307-1313.
Johnson DE, Hill TM, Carmean BR, Brannie ME, Lodman DW and Ward G. 1991 Mew perspectives on ruminant methane emissions. In “Energy metabolism of farm animals (Eds Wenk C and Bossinger M) pp 376-379.
Jorgensen H, Sorensen P and Eggum BO. 1990; Protein and energy-metabolism in broiler-chickens selected for either body weight gain or feed efficiency. British Poultry Science 31, 517-524.
Kamalzadeh A and Aouladrabiei MR. 2009. Effects of restricted feeding on intake, digestion, nitrogen balance and metabolisable energy in small and large body sized sheep breeds. Asian-Australasian Journal of Animal Science 22, 667-673.
Kaminiska BZ. 1979. Food intake in the young chick. In: K.N, Boorman and Freeman, B.M. (Eds.), Food intake regulation in poultry. British Poultry Science, Ltd. Edinburg, Scotland, PP: 199-206.
Kanengoni AT, Dzama K, Chimonyo M, Kusina J, Maswaure SM. 2002. Influence of level of maize cob meal on nutrient digestibility and nitrogen balance in Large White, Mukota and LW x M F-1 crossbred pigs. Animal Science 74, 127-134.
Kapell DNRG., Ashworth CJ, Walling GA. Lawrence AB, Edwards SA and Roehe R. 2009. Estimation of genetic associations between reproduction and production traits based on a sire and dam line with common ancestry. Animal 3, 1354-1362
Karlsson LJE, Greeff JC and Harris JF 1995. Breeding sheep for worm resistance. Journal of. Agricultural Science Western Australia 2, 56–59.
Katle J. 1991. Selection for efficiency of food utilization in laying hens – causal factors for variation in residual feed consumption. Poultry Science 32, 955-969.
Kemp B, Den Hartog LA, Klok JJ and Zandstra T. 1991. The digestibility of nutrients, energy and nitrogen in the Meishan and Dutch Landrace pig. Journal of Animal Physiology and Animal Nutrition 65, 263-266.
Kiani A, Chwalibog A, Nielsen MO and Tauson AH. 2004. Effect of genetic line on energy and nitrogen metabolism during late gestation in twin-pregnant ewes fed restrictedly. Journal of Animal and Feed Science 13, 417-420.
King, TS, Elia, M and Hunter, JO.1998. Abnormal colonic fermentation in irritable bowel syndrome. Lancet 352, 1187-1189.

96
Klemm R, Reiter K and Pingel H. 1994. Results and effects of direct selection for feed efficiency in the domestic ducks: 2. Report: Methodical aspects. Archiv für Geflügelkunde 58, 176-181.
Klemm R. 1995. Selection on feed conversion ratio in ducks. Proceedings of the 10th European Symposium on Waterfowl, Halle, Germany, pp. 433-438
Klemm R. and Pingel H. 1992. Results of divergent selection for either low or high feed conversion in duck over six generations. Archives of Animal Breeding 30, 103-110.
Knights M, Garcia GW. 1997. The status and characteristics of the goat (Capra hircus) and its potential role as a significant milk producer in the tropics: A Review. Small Ruminant Research 26, 203-215.
Knight TW, Molano G, Clark H and Cavanagh A. 2008. Methane emissions from weaned lambs measured at 13, 17, 25 and 35 weeks of age compared with mature ewes consuming a fresh forage diet. Australian Journal of Experimental Agriculture 48, 240-243.
Knowlton KF, Wilkerson VA, Casper DP and Mertens DR. 2010. Manure nutrient excretion by Jersey and Holstein cows. Journal of Dairy Science 93, 407-412.
Korver S. 1988. Genetic aspects of feed-intake and feed-efficiency in dairy cattle – a review. Livestock Production Science 20, 1-13.
Kyriazakis I, Dotas D and Emmans GC.1994. The effect of breed on the relationship between feed composition and the efficiency of protein-utilization in pigs. British Journal of Nutrition 71, 849-859.
Kyriazakis I, Emmans GC and Anderson D. 1995. Do breeds of pig differ in the efficiency with which they use a limiting protein supply? British Journal of Nutrition 74, 183-195.
Larzul C, Guy G and Bernadet MD. 2004. Feed efficiency, growth and carcass traits in female mule ducks. Archiv fur Geflugelkunde 68, 265-268.
Lassey KR, Ulyatt MJ, Martin RJ, Walker CF, Shelton ID. 1997. Methane emissions measured directly from grazing livestock in New Zealand. Atmosphere and Environment 31, 2905–2914.
Laughlin K. 2007. The Evolution of Genetics, Breeding and Production. Temperton Fellowship Report No.15. Harper Adams University College.
Laurenz JC, Byers FM, Schelling GT and Greene LW. 1991. Effects of Season on the Maintenance Requirements of Mature Beef-Cows. Journal of Animal Science 69, 2168-2176.
Laurin DE, Touchburn SP, Chavez ER and Chan CW. 1985. Methods of measuring energy utilization in broilers: Effect of genetic line and presence of supplemental dietary fat. Poultry Science 64:969–978.
Leclercq B and Saadoun A. 1982. Selecting broilers for low or high abdominal fat: comparison of energy metabolism of the lean and fat lines. Poultry Science 61:1799–1803.
Leclerq B. 1986. Energy requirements of avian species. Pp. 125-139 in C. Fisher and K.N. Boorman (Eds.) Nutrient Requirements of Poultry and Nutritional Research. Butterworth, Sevenoaks, UK.

97
Leenstra FR and Pit R. 1987. Fat deposition in a broiler sire strain. 2. Comparisons among lines selected for less abdominal fat, lower feed conversion ratio and higher body weight after restricted and ad libitum feeding. Poultry Science 66, 193-202.
Leeson S and Summers JD 1997. Commercial Poultry Nutrition. University Books, Guelph, Ontaria, Canada.
Leeson S and Summers JD. 1997. Feeding programmes for laying hens. Pp. 143-206 in Commercial Poultry Nutrition . University Books, Guelph, Canada.
Len NT, Lindberg JE and Ogle B. 2007. Digestibility and nitrogen retention of diets containing different levels of fibre in local (Mong Cai), F1 (Mong Cai x Yorkshire) and exotic (Landrace x Yorkshire) growing pigs in Vietnam. Journal of Animal Physiology and Animal Nutrition 91, 297-303.
Len NT, Hong TTT, Ogle B and Lindberg JE. 2009a. Comparison of total tract digestibility, development of visceral organs and digestive tract of Mong Cai and Yorkshire x Landrace piglets fed diets with different fibre sources. Journal of Animal Physiology and Animal Nutrition 93, 181-191.
Len NT, Ngoc TB, Ogle B and Lindberg JE. 2009b. Ileal and total tract digestibility in local (Mong Cai) and exotic (Landrace x Yorkshire) piglets fed low and high-fibre diets, with or without enzyme supplementation. Livestock Science 126, 73-79.
Lepron E, Bergeron, R, Robert S, Faucitano L, Bernier J. F and Pomar C. 2007. Relationship between residual energy intake and the behaviour of growing pigs from three genetic lines. Livestock Science 111, 104-113.
Li Z, Nestor KE, Saif YM, Bacon WL and Anderson JW. 1999. Effect of selection for increased body weight on mitogenic responses in turkeys. Poultry Science 78, 1532-1535.
Leeson S and Summers JD. 1997. Feeding programmes for Turkeys. Pp. 299-323 in Commercial Poultry Nutrition. University Books, Guelph, Canada.
Liu SM, Smith TL, Briegel J, Gao SB and Peng WK 2007. Fractional protein synthesis rate and polyamine concentrations in tissues of Merino sheep selected for gastrointestinal nematode resistance. Livestock Science 106, 65–75.
Liu S, Smith TL, Briegel J, Murray A, Masters DG, Karlsson LJE, Palmer DG, Greeff JC, Besier RB and Gao SB 2005. Comparing productive performance of nematode resistant Merino sheep with non-selected control. Livestock Production Science 97, 117– 129.
Liu SM, Smith TL, Karlsson LJE, Palmer DG and Besier RB 2005. The costs for protein and energy requirements by nematode infection and resistance in Merino sheep. Livestock Production Science 97, 131– 139.
Lopez S, Frutos P, Mantecon AR and Giraldez FJ. 2001. Comparative digestion of herbage by two breeds of sheep: effects of grass maturity stage and level of intake. Animal Science 73, 513-522.
Lopez G, Leeson S. 2008. Review: Energy partitioning in broiler chickens. Canadian Journal of Animal Science 88, 205-212
Lorenzo Bermejo J, Roehe R, Schulze V, Rave G, Looft H and Kalm E. 2003b. Random regression to model genetically the longitudinal data of daily feed intake in growing pigs. Livestock Production Science 82, 189-199.

98
Lourenco ALG, Dias-da-Silva AA, Fonseca AJM and Azevedo JT. 2000. Effects of live weight, maturity and genotype of sheep fed a hay-based diet, on intake, digestion and live weight gain. Livestock Production Science 63, 291-296.
Luiting P. 1990. Genetic variation of energy partitioning in laying hens. Causes of variation in residual feed consumption. Worlds Poultry Science Journal 46, 133-152.
Luiting P and Urff EM. 1991. Optimization of a model to estimate residual feed consumption in the laying hen. Livestock Production Science 27, 321-338.
Luiting P, Schrama JW, van der Hel W and Urff EM. 1991. Metabolic differences between White Leghorns selected for high and low residual food consumption. British Poultry Science 32, 763-782.
Mann DL, Goode L and Pond KR. 1987. Voluntary intake, digestibility, rate of passage and gastro-intestinal tract fill in tropical and temperate breeds of sheep. Journal of Animal Science 64, 880-886.
Marie-Etancelin C, Chapuis H, Brun JM, Larzul C, Mialon-Richard MM and Rouvier R. 2008. Genetics and selection of mule ducks in France: a review. Worlds Poultry Science Journal 64, 187-207.
McKay LF, Eastwood MA and Brydon WG. 1985. Methane excretion in man- a study of breath, flatus and faeces. Gut 26, 206-214
Mellado M and Meza-Herrera CA. 2002. Influence of season and environment on fertility of goats in a hot-arid environment. Journal of Agricultural Science 138, 97-101.
Mellado M, Valdez R, Garcia JE, Lopez R and Rodriguez A. 2006. Factors affecting the reproductive performance of goats under intensive conditions in a hot arid environment. Small Ruminant Research 63, 110-118.
Meuwissen THE, Hayes BJ and Goddard ME. 2001 Prediction of total genetic value using genome-wide dense marker maps. Genetics 157, 1819–1829.
Mignon-Grasteau S, Muley N, Bastianelli D, Gomez J, Peron A, Sellier N, Millet N, Besnard J, Hallouis JM and Carre B. 2004. Heritability of digestibilities and divergent selection for digestion ability in growing chicks fed a wheat diet. Poultry Science 83, 869-877.
Mohammed HH. 1982. Energy requirements for maintenance and growth: comparison of goats and sheep. PhD thesis, University of Reading.
Mohrmann M, Roehe R, Knap PW, Looft H, Plastow GS and Kalm E 2006. Quantitative trait loci associated with AutoFOM grading characteristics, carcass cuts and chemical body composition during growth of Sus scrofa. Animal Genetics 37:435-443.
Molina E, Ferret A, Caja G, Calsamiglia S, Such X and Gasa J. 2001. Comparison of voluntary food intake, apparent digestibility, digesta kinetics and digestive tract content in Manchega and Lacaune dairy sheep in late pregnancy and early and mid lactation. Animal Science 72, 209-221.
Montanobermudez M, Nielsen MK and Deutscher GH. 1990. Energy-Requirements for maintenance of crossbred beef-cattle with different genetic potential for milk. Journal of Animal Science 68, 2279-2288.
Moore RL, Essig HW and Smithson LJ. 1975. Influence of breeds of cattle on ration utilization. Journal of Animal Science 41, 203-207.

99
Morales J, Perez JF, Baucells M.D, Mourot J and Gasa J. 2002. Comparative digestibility and lipogenic activity in Landrace and Iberian finishing pigs fed ad libitum corn- and corn-sorghum-acorn-based diets. Livestock Production Science 77, 195-205.
Morel PCH, Lee TS and Moughan PJ. 2006. Effect of feeding level, live weight and genotype on the apparent faecal digestibility of energy and organic matter in the growing pig. Animal Feed Science and Technology 126, 63-74.
Morris CA, Vlassoff A, Bisset SA, Baker RL, Watson TG, West CJ and Wheeler M 2000. Continued selection of Romney sheep for resistance or susceptibility to nematode infection: estimates of direct and correlated responses. Animal Science 70, 17-27.
Moughan P.J. 2003. Journal of Animal Science 81 (E. Suppl. 2), E60-E67. Munger A. and Kreuzer M. 2008. Absence of persistent methane emission differences in
three breeds of dairy cows. Australian Journal of Experimental Agriculture 48, 77-82. NRC (National Research Council). 1984. Nutrient Requirements of Poultry. 8th Revised
Edition, National Academy Press, Washington, DC, USA NRC (National Research Council). 2000. Nutrient Requirements of Beef Cattle. Update 2000
of the 7th revised edition 1996. National Academy Press, Washington DC, USA. NRC (National Research Council). 2001. Nutrient Requirements of Dairy Cattle. 2001. 7th
revised edition. National Academy Press, Washington DC, USA. Nieuwhof GJ, Conington J and Bishop SC. 2009. A genetic epidemiological model to
describe resistance to an endemic bacterial disease in livestock: application to footrot in sheep. Genetics Selection Evolution 41, 1-12.
Nieuwhof GJ, Conington J, Bunger L, Haresign W and Bishop SC 2008. Genetic and phenotypic aspects of foot lesion scores in sheep of different breeds and ages. Animal 2, 1289-1296
Noblet J. Bernier JF, Dubois S, Le Cozler Y and van Milgen J. 1998. Effect of breed and body weight on components of heat production in growing pigs. Pp. 225-228 in (McCracken KJ; Unsworth EF; Wylie ARG, Eds.) Energy Metabolism of Farm Animals. CABI Publishing, Wallingford, UK.
Noblet J, Karege C, Dubois S and van Milgen J. 1999. Metabolic utilization of energy and maintenance requirements in growing pigs: Effects of sex and genotype. Journal of Animal Science 77, 1208-1216.
Nestor KE, Noble DO, Zhu J and Moritsu Y. 1996. Direct and correlated responses to long-term selection for increased body weight and egg production in turkeys. Poultry Science 75, 1180-1191.
Oddy VH. 1999. Protein metabolism and nutrition in farm animals: an overview. P. 7-23 in (Lobley GE, White A and MacRae JC, eds) Protein Metabolism and Nutrition. EAAP Publication 96.
Ortigues I, Petit M and Agabriel J. 1993. Influence of body condition on maintenance energy requirements of Charolais cows. Animal Production 57, 47-53.
Peron A, Gomez J, Mignon-Grasteau S, Sellier N, Besnard J, Derouet M., Juin H and Carre B. 2006. Effects of wheat quality on digestion differ between the D+ and D- chicken lines selected for divergent digestion capacity. Poultry Science 85, 462-469.
Pinares-Patino CS, Ulyatt MJ, Lassey KR, Barry TN and Holmes CW. 2003a. Rumen function and digestion parameters associated with differences between sheep in

100
methane emissions when fed chaffed lucerne hay. Journal of Agricultural Science 140, 205-214.
Pinares-Patino CS and Clark D. 2008. Reliability of the sulphur hexafluoride tracer technique for methane emission measurement from individual animals: an overview Australian Journal of Experimental Agriculture 48, 223-229.
Pinares-Patino CS, Holmes CW, Lassey KR and Ulyatt MJ. 2008. Measurement of methane emission from sheep by the sulphur hexafluoride tracer technique and by the calorimetric chamber: failure and success. Animal 2, 141-148.
Pinares-Patino CS, Waghorn GC, Machmueller A, Vlaming B, Molano G, Cavanagh A and Clark H. 2007. Methane emissions and digestive physiology of non-lactating dairy cows fed pasture forage. Canadian Journal of Animal Science 87, 601-613.
Pinares-Patino CS, Ulyatt MJ, Lassey KR, Barry TN and Holmes CW. 2003b. Persistence of differences between sheep in methane emission under generous grazing conditions. Journal of Agricultural Science, Cambridge 140, 227–233.
Pinares-Patino C, Vlaming B, Cavanagh A, Molano G and Clark H. 2005. Persistence of dairy cows in animal-to-animal variation in methane emission. In ‘Working papers of the 2nd international conference on greenhouse gases and animal agriculture’. (Eds Soliva CR, Takahashi J, Kreuzer, M) pp. 401–404. (Institute of Animal Science: ETH Zurich, Switzerland)
Pishnamazi A, Pourezza J, Edriss MA and Samie AH. 2005. Influence of broiler breeder and laying hen breed on the apparent metabolisable energy of selected feed ingredients. International Journal of Poultry Science 4, 163-166.
Pitt P, De Bruijn KM, Beeching MR, Golberg E and Blendis LM. 1980. Studies on breath methane; the effect of ethnic origins and lactulose. Gut 21, 951-959.
Pomar C, Kyriazakis I, Emmans, GC and Knap PW. 2003. Journal of Animal Science 81 (E. Suppl. 2), E178-E186.
Roehe R, Plastow GS and Knap PW. 2003. Quantitative and molecular genetic determination of protein and fat deposition. Journal HOMO of Comparative Human Biology 54, 119-131.
Powell JC. 1984. Selection for improved feed conversion ratio in the domestic ducks. Proceedings of the17th World Poultry Congress, Helsinki, pp. 108-109.
Proudman JA, Mellen WJ and Anderson DL. 1970. Utilization of feed in fast and slow-growing lines of chickens. Poultry Science, 49, 961-972.
Pym RAE, Leclercq B, Tomas FM and Tesseraud S. 2004 Protein utilisation and turnover in lines of chickens selected for different aspects of body composition. British Poultry Science 45, 775-786.
Qin GX, Verstegen MWA and Bosch MW. 1995. Variation of digestive capacity between genetically different pig populations: a review. Journal of Animal Physiology and Animal Nutrition 73, 233-242.
Ranilla MJ, Lopez S, Giraldez FJ, Valdes C and Carro MD. 1998. Comparative digestibility and digesta flow kinetics in two breeds of sheep. Animal Science 66, 389-396.
Renaudeau D, Bocage B and Noblet J. 2006. Influence of energy intake on protein and lipid deposition in Creole and Large White growing pigs in a humid tropical climate. Animal Science 82, 937-945.

101
Retailleau B. 1999. Comparison of the growth and body composition of 3 types of ducks: Pekin, Muscovy and mule. Proceedings of the 1st World Conference on Waterfowl, Taichung, Taiwan, pp. 597-602.
Reynolds CK and Kristensen NB. 2008. Nitrogen recycling through the gut and the nitrogen economy of ruminants: An asynchronous symbiosis. Journal of Animal Science 86, E293-E305
Roberson KD, Rahn AP, Balander RJ, Orth MW, Smith DM, Booren BL, Booren AM, Osburn WN and Fulton RM. 2003. Evaluation of the growth potential, carcass components and meat quality characteristics of three commercial strains of tom turkeys. Journal of Applied Poultry Research 2, 229-236.
Robertson LR and Waghorn GC. 2002. Dairy industry perspectives on methane emissions and production from cattle fed pasture or total mixed rations in New Zealand. Proceedings of the New Zealand Society of Animal Production 62, 213–218.
Roehe R, Plastow GS and Knap PW 2003. Quantitative and molecular genetic determination of protein and fat deposition. Journal HOMO of Comparative Human Biology 54, 119-131.
Roehe R, Landgraf S, Mohrmann M, Knap PW, Looft H and Kalm E. 2004 Proc. 55th Annual Meeting EAAP, Bled, Slovenia, paper PG6.7, 1-10.
Roehe R, Lorenzo Bermejo J, Landgraf S, Knap PW and Kalm E. 2002. Proc. 53th Annual Meeting EAAP, Cairo, Egypt, paper G6.3, 1-8.
Roehe R, Shrestha NP, Mekkawy W, Baxter EM, Knap PW, Smurthwaite KM, Jarvis S, Lawrence AB and Edwards SA. 2009. Genetic analyses of piglet survival and individual birth weight on first generation data of a selection experiment for piglet survival under outdoor conditions. Livestock Science 121, 173-181
Romanov MN. 1999. Goose production efficiency as influenced by genotype, nutrition and production systems. Worlds Poultry Science Journal 55, 281-294.
Rougiere N, Gomez J, Mignin-Grateau S and Carre B. 2009. Effects of diet particle size on digestive parameters in D+ and D- genetic chicken lines selected for divergent digestion efficiency. Poultry Science 88, 1206-1215.
Roy-Smith F. 1980. The fasting metabolism and relative energy intake of goats compared to sheep. Animal Production 30, 491.
Russel AJF and Wright IA. 1983. Factors affecting maintenance requirements of beef cows. Animal Production 37, 329-334.
Sahlu T, Goetsch AL, Luo J, Nsahlai IV, Moore JE, Galyean ML, Owens FN, Ferrell CL and Johnson ZB. 2004. Nutrient requirements of goats: developed equations, other considerations and future research to improve them. Small Ruminant Research 53, 191-219.
Savon L. 2005. Tropical roughages and their effect on the digestive physiology of monogastric species. Cuban Journal of Agricultural Science 39, 463-474.
Schulze V, Roehe R, Looft H and Kalm E. 1998. Genetic improvement of exterior and leg soundness in pigs. Züchtungskunde 70, 43-60.
Schulze V, Roehe R, Looft H and Kalm E. 2001. Genetic analysis of the course of individual growth and feed intake of group-penned performance tested boars. Archives of Animal Breeding 44, 139-156.

102
Schulze V, Roehe R, Lorenzo Bermejo J, Looft H and Kalm E. 2002. Genetic associations between observed feed intake measurements during growth, feed intake curve parameters and growing-finishing performances of central tested boars. Livestock Production Science 73, 199-211.
Schulze V, Roehe R, Lorenzo Bermejo J, Looft H, Kalm E. 2003. The influence of feeding behaviour on feed intake curve parameters and performance traits of station-tested boars. Livestock Production Science 82, 105-116.
Sellner EM, Kim JW, McClure MC, Taylor KH, Schnabel RD and Taylor JF. 2007. Application of genomic information in livestock. Journal of Animal Science 85, 3148-3158.
Shires A, Thompson JR, Turner BV, Kennedy PM and Goh YK. 1987. Rate of passage of corn-canola meal and corn-soybean meal diets through the gastrointestinal tract of broiler and White Leghorn chickens. Poultry Science 66, 289-298.
Sibbald IR. 1976. True metabolisable energy values of several feeding stuffs measured with roosters, laying hens, turkeys and broiler hens. Poultry Science 55, 1459-1463.
Sibbald IR, and SJ Slinger. 1963. The effects of breed, sex, an arsenical and nutrient density on the utilization of dietary energy. Poultry Science 42, 1325–1332.
Silanikove N. 1986. Interrelationships between feed quality, digestibility, feed consumption and energy requirements in desert (Bedouin) and temperate (Saanen) goats. Journal of Dairy Science 69, 2157-2162.
Silanikove N, Tagari H and Shkolnik A. 1993. Comparison of rate of passage, fermentation rate and efficiency of digestion of high-fiber diet in desert bedouin goats compared to Swiss Saanen goats. Small Ruminant Research 12, 45-60.
Skinner-Noble DO, Jones RB and Teeter RG. 2003. Components of feed efficiency in broiler breeding stock: Is improved feed conversion associated with increased docility and lethargy in broilers. Poultry Science 82, 532-537.
Solis JC, Byers FM, Schelling GT, Longs CR and Greene LW. 1988. Maintenance requirements and energetic efficiency of cows of different breed types. Journal of Animal Science 66, 764-773
Spratt RS and Leeson S. 1987. Determination of metabolizabkle energy of various diets using Leghorn, Dwarf and regular broiler breeder hens. Poultry Science 66, 314-317.
Stevenson MH. 1989. Nutrition of domestic geese. Proceedings of the Nutrition Society 48, 103-111.
Su G, Sorensen D and Lund MS. 2008. Variance and covariance components for liability of piglet survival during different periods. Animal 2, 184-189.
Susenbeth A, Dickel T, Diekenhorst A and Hohler D. 1999. The effect of energy intake, genotype, and body weight on protein retention in pigs when dietary lysine is the first-limiting factor. Journal of Animal Science 77, 2985-2989.
Swennen Q, Janssens GPJ, Collin A, Le Bihan-Duval E, Verbeke K, Decuypere E and Buyse J. 2006. Diet-induced thermo-genesis and glucose oxidation in broiler chickens: Influence of genotype and diet composition. Poultry Science 85, 731-742.
Swennen Q, Janssens GPJ, Decuypere E, Buyse J. 2004. Effects of substitution between fat and protein on feed intake and its regulatory mechanisms in broiler chickens: Energy

103
and protein metabolism and diet-induced thermogenesis. Poultry Science 83, 1997-2004.
Strobel E, Franke E and Manner K. 2004. Energy exchange in white laying hens with differences in residual food consumption. Archiv fur Geflügelkunde 68, 62-70.
Su G, Kjaer JB and Sorensen P. 2006. Divergent selection on feather pecking behavior in laying hens has caused differences between lines in egg production, egg quality, and feed efficiency. Poultry Science 85, 191-197.
TenDoeschate RAHM, Scheele CW, Schreurs WAM and van der Klis JD 1993. Digestibility studies in broiler-chickens – influence of genotype, age, sex and method of determination. British Poultry Science 34, 131-146.
Tolkamp BJ. 2010. Efficiency of energy utilisation and voluntary feed intake in ruminants. Animal 4, 1084-1092.
Tolkamp BJ and Brouwer BO. 1993. Statistical review of digestion in goats compared with other ruminants. Small Ruminant Research 11:107–123.
Tolkamp BJ and Ketelaars JJMH. 1994. Efficiency of energy utilisation in cattle given food ad libitum: predictions according to the ARC system and practical consequences. Animal Production 59, 43-47.
Tolkamp BJ, Ketelaars JJMH and Hofs, P. 1994. Voluntary intake of digestible organic matter and fasting heat production of West African Dwarf Goats and Swifter sheep. Small Ruminant Research 15, 45-54.
Tolkamp BJ and Kyriazakis I. 2009. Toward a new practical energy evaluation system for dairy cows. Animal 3, 307-314.
Tovar-Luna I, Goetsch AL, Puchala R, Sahju T, Carstens GE, Freetly HC and Johnson ZB. 2007a. Effects of diet quality on energy expenditure by 20-month-old Alpine, Angora, Boer, and Spanish wethers. Small Ruminant Research 72, 18-24.
Tovar-Luna I, Goetsch AL, Puchala R, Sahlu T, Carstens GE, Freetly HC and Johnson ZB. 2007b. Efficiency of energy use for maintenance and gain by growing crossbred Boer and Spanish Goats consuming diets differing in forage level. Small Ruminant Research 72, 20-27.
Turner SP, Roehe R, Mekkawy W, Farnworth M.J, Knap PW and Lawrence AB. 2008. Bayesian analysis of genetic associations of skin lesions and behavioural traits to identify genetic components of individual aggressiveness in pigs. Behaviour Genetics 38, 67-75.
Turner SP, Roehe R, D’Eath RB, Ison SH, Farish M, Jack MC, Lundeheim N, Rydhmer L and Lawrence AB. 2009. Genetic validation of post-mixing skin injuries in pigs as an indicator of aggressiveness and the relationship with injuries under more stable social conditions. Journal of Animal Science 87, 3076-3082.
Van Eerden E, Van Den Brand H, Parmentier HK, de Jong MCM and Kemp B. 2004a. Phenotypic selection for residual feed intake and its effect on humoral immune responses in growing layer hens. Poultry Science 83, 1602-1609.
Van Eerden E, Van Den Brand H, Heetkamp MJW, Decuypere E and Kemp B. 2006. Energy partitioning and thyroid hormone levels during Salmonella enteritidis infections in pullets with high or low residual feed intake. Poultry Science 85, 1775-1783.

104
Van Eerden E, Van Den Brand H, Reilingh GD, Parmentier HK, de Jong MCM and Kemp B. 2004b. Residual feed intake and its effect on Salmonella enteritidis infection in growing layer hens. Poultry Science 83, 1904-1910.
Van Es AJH 1961. Between-animal variation in the amount of energy required for the maintenance of cows. PhD thesis, Wageningen University.
Van Es AJH 1975. Heterotrophe production in animals. Pp. 105-127 in (GJ Vervelde, ed.) Productivity in Biological Systems (in Dutch). Wageningen: Centrum voor Landbouw Publikaties and Landbouwdocumentatie.
Van Milgen J and Noblet J. 2003. Journal of Animal Science 81 (E. Suppl. 2), E86-E93. van Milgen J, Bernier JF, Dubois S, Le Cozler Y, Noblet J. 1998a. Modelling dynamic
aspects of heat production in pigs. Pp. 331-334 in (McCracken KJ; Unsworth EF; Wylie ARG, Eds.) Energy Metabolism of Farm Animals. CABI Publishing, Wallingford, UK.
van Milgen J, Bernier JF, Lecozler Y, Dubois S, Noblet J. 1998b. Major determinants of fasting heat production and energetic cost of activity in growing pigs of different body weight and breed/castration combination. British Journal of Nutrition 79, 509-517.
Van Wieren SE. 2000. Digestibility and voluntary intake of roughages by wild boar and Meishan pigs. Animal Science 71, 149-156.
Veerkamp RF and Emmans GC. 1995. Sources of genetic variation in energetic efficiency in dairy cows. Livestock Production Science 44, 87-97.
Vercoe JE. 1970. Fasting metabolism and heat increment of feeding in Brahman x British cross cattle. In: Energy Metabolism of Farm Animals. EAAP Publication 13.
Vlaming JB, Lopez-Villalobos N, Brookes IM, Hoskin SO and Clark H. 2008. Within- and between-animal variance in methane emissions in non-lactating dairy cows Australian Journal of Experimental Agriculture 48, 124–127
Von Felde A., Roehe R, Looft H and Kalm E. 1996. Genetic association between feed intake and feed intake behaviour at different stages of growth of group-housed boars. Livestock Production Science 47, 11-22.
Warnic AC and Cobb EH. 1976. Genetic variation in nutrition of cattle fro meat production. In: The genetic variance on nutritional requirements of animals. National Academy of Sciences, Maryland, USA.
Whittemore CT, Green DM and Knap PW. 2001. Technical Review of the energy and protein requirements of pigs: protein. Animal Science 73, 363-373.
Whittemore CT. 2006. Energy value of feedstuffs for pigs. Pp. 317-342 in (I. Kyriazakis and .T. Whittemore, eds.) Whittemore’s Science and Practice of Pig Production. Blackwell Publishing.
Wildeus S, Turner KE and Collins JR. 2007. Growth, intake, diet digestibility, and nitrogen use in three hair sheep breeds fed alfalfa hay. Small Ruminant Research 69, 221-227.
Windisch W, Gotterbarm GG, Roth FX, Schams D and Kirchgessner M. 2000. Effect of genetic provenience of pigs (German Landrace, Pietrain) on parameter of protein metabolism. Züchtungskunde 72, 379-388.
Wolf J, Žáková E, and Groeneveld E. 2008. Within-litter variation of birth weight in hyperprolific Czech Large White sows and its relation to litter size traits, stillborn piglets and losses until weaning. Livestock Science 115,195-205.

105
Yaghobfar A. 2001; Effect of genetic line, sex of birds and the type of bioassay on the metabolisable energy value of maize. British Poultry Science 42, 359-353.
Yaghobfar A. 2003. The effects of strain, age and sex of birds on the determined metabolisable energy of maize and fishmeal. British Poultry Science 44, S38-S39.
Yan T, Mayne CS and Agnew RE. 2006. Effects of dairy cow genotype with two planes of nutrition on energy partitioning between milk and body tissue. Journal of Dairy Science 89, 1031-1042.
Yan T, Porter MG and Mayne CS 2009. Prediction of methane emission from beef cattle using data measured in indirect open-circuit respiration calorimeters. Animal 3, 1455-1462.
Yanez-Ruiz DR, Macias B, Pinloche E and Newbold CJ. 2011. The persistence of bacterial and methanogenic archaeal communities residing in the rumen of young lambs FEMS Microbiology Ecology 72, 272-278.
Yen JT, Varel VH, Nienaber JA. 2004. Metabolic and microbial responses in western crossbred and Meishan growing pigs fed a high-fiber diet Journal of Animal Science 82, 1740-1755
Zemmelink G, Tolkamp BJ, Ogink NWM. Energy requirements for maintenance and gain of West African Dwarf goats. Small Ruminant Research 5, 205-215.
Zhang W, Aggrey SE, Pesti GM, Bakalli RI and Edwards HM. 2005. Genetic analysis on the direct response to divergent selection for phytate phosphorus bioavailability in a random bred chicken population. Poultry Science 84, 370-375.
Zhang W and Aggrey SE. 2003. Genetic variation in feed utilization efficiency of meat-type chickens. Worlds Poultry Science Journal 59, 328-339.
Zhou M, Herandez-Sanabria E and Guan LL. 2009. Assessment of the microbial ecology of ruminant methanogens in cattle with different feed efficiencies. Applied and Environmental Microbiology 75, 6524-6533.