revision of the coroebine genus coraebosoma...
TRANSCRIPT

Revision of the coroebine genus Coraebosoma Obenberger (Coleoptera: Buprestidae: Agrilinae)
C. L. BELLAMY
Bellamy, C. L.: Revision of the coroebine genus Coraebosoma Obenberger (Coleoptera: Bupres- En scan tidae: Agrilinae). Ent. scand. 21: 197-210. Copenhagen, Denmark June 1990. ISSN 0013-8711.
A
The coroebine genus Coraebosoma Obenberger is revised for the first time with six new species, five described from the Philippine Islands: C. samarense from Samar Is., C. panayense from Pa- nay Is., C. negrosianum from Negros Is., C. sibuyanicum from Romblon and Sibyan Is., C. violaceum from Mindanao Is. and C. indicum, from southern India. The genus is redescribed with the type-species, C. manilense Obenberger, C. carteri Hoscheck and new species diagnosti- cally described. A key to separate the species, illustrations, and a distribution map are presented along with discussion on the generic relationships. The species are cladistically analyzed and sup- ported by an area cladogram.
' \ \ C. L. Bellamy, Dept. of Entomology, NHB 169, Smithsonian Institution, Washington, D. C. 20560, U. S. A.
INTRODUCTION
The Coroebini Bedel is the largest tribal taxon in the Buprestidae, when considering the number of in- cluded genera (approximately 130). As a continua- tion toward my goal of phylogenetic redefinition for the world Coroebini Bedel (e.g. Bellamy 1988a, 1988b for Africa; 1988c for Australia), an under- standing of the coroebine fauna of the Philippines is essential to defining the relationships between the disparate faunas of ~ustral ia , Indomalaysia, southeast Asia and India.
The Philippine Coroebini were last studied in to- tal by Fisher (1921). Several of the genera await revi- sion with the accumulated material available from numerous institutions. This paper attempts to bring together all of the known information on the genus Coraebosoma Obenberger, which has received virtually no attention except the descrip- tion of the two known species.
Methods
Specimens were borrowed from the following col- lections, with the collection abbreviations from, or in the style of, Arnett & Samuelson (1986). MGFT = Museum Georg Frey, Tutzing, W. Germany (via NHMB = Naturhistorisches Museum, Basel);
NMPC = National Museum, Prague; NSMT =
National Science Museum, Tokyo; NMNH = Na- tional Museum of Natural History, Smithsonian Institution, Washington, D. C.; CLBC = my research collection; GHNC = G. H. Nelson collec- tion, Pomona, California; MTCJ = M. Toyama collection, Nishinomiya and KACJ = K. Akiyama, Yokohama, Japan.
The morphometric measurements are taken as follows: overall body proportion (L/W): maximum length, dorsal from midpoint of frontovertex to elytral apex; maximum width, taken at widest point of pronotum; pronotal proportion (PN): length, from midpoint of anterior margin to midpoint of posterior margin; maximum width, taken at widest point.
Label data are listed in a decreasing order of mag- nitude (e.g. country, island, district, etc.) from the exact ordering on the labels. Orthographic correc- tions and coordinates have been determined, where possible, and are listed in [brackets]. Place name or- thography and coordinates were determined using the gazetteer series of the United States Board on Geographic Names, Defense Mapping Agency Topographic Center and the most recent (7th) edi- tion of the Times Atlas of the World, 1988. The cladistic analysis was accomplished using
@ Entomologica scandinavica (Grp. 2)

198 Bellamy, C. L. ENT. SCAND. VOL. 21:2 (1990)
Hennig68 (Farris 1988). Specific routines and cal- culated values are discussed following the descrip- tive portion of the paper.
TAXONOMY
Genus Coraebosoma Obenberger Coraebosoma Obenberger, 1923: 32; Hoscheck 1931: 159;
Obenberger 1934: 815; Bellamy 1985: 425.
Type-species: Coraebosoma manilense Obenberger (by monotypy).
Coroebini; elongate, ovoid, transversely convex above and below; head only slightly visible from above; frontovertex flattened dorsally, broadly shallowly depressed between eyes anterior to anten- nal insertions; eyes large, inner margins slightly divergent dorsally; one narrow arcuate transverse groove dorsal to each antennal cavity, both grooves confluent medially with frontoclypeal depression between antennae; disc of frontoclypeus an invert- ed Y-shape, emarginate distally; gena with narrow, deep groove for basal antennomeres in repose con- fluent with antennal cavity, extends laterally be- tween bottom of eye and subacute marginal tooth on each side; labrum partially visible beyond fron- toclypeal margin; mandibles of moderate size; maxillary palps with 3rd joint widening apically, distal margin obliquely truncate; antennae short, when laid along side not reaching to opposite of an- terior % of pronotum; antennomere 2 shorter, nar- rower than 1; 3 shorter than 2; 4-10 serrate with ser- rate portion of each narrowed, extending obliquely to base; 11 oblong.
Pronotum wider than long; lateral margins crenulate; when viewed from side extending along 45" angle from opposite ventral margin of eye dor- sad to basolateral angle; disc strongly convex an- teriorly, lateral areas explanate; scutellum large, an- teroposteriorly compressed cordiform; elytra with sides subparallel to before apical %, attenuate to apex; margins serrulate, more strongly serrate api- cally; disc convex laterally, flattened medially; pygidium not visible from above; epipleura ventral- ly deflexed, extending almost entire length laterally.
Prosternum with ventrally produced discal area as in Figs 1,7; mentonierre with two small angulate lobes separated by angular emargination; process narrowed ventrad to procoxae, then widens before apical attenuation; lateroapical notch, depression along hypomeral margin for antennomeres in re- pose; mesosternum with small lateral lobes hidden
beneath anterior lobes of metasternal base which form most of sternal cavity; suture between meta- and metepisternum arcuate anteriorly; metepime- ron partially visible beyond epipleuron metacoxal plate with distal margin nearly straight to acutely projecting apicolateral angle; abdominal sternite 1 longer than 2, subequal to 3 +4+ 5; suture between 1 and 2 feeble, arcuate medially; sutures otherwise transverse; 5 with explanate preapical margin.
Legs with femora slightly swollen, sides sub- parallel, inner margin longitudinally excavated for tibiae in repose; tibiae with two short distal spines; protibiae flattened; metatibiae with projecting tooth near midpoint, setal comb on outer edge; tarsi short; tarsomeres 1-4 with ventral pulvilli; tarsal claws assymetrical with one claw simple, opposite bifid; protarsi with inner claw bifid, meso- and metatarsi with outer claw bifid.
Hind wing as in Figs 11, 12: radial cell quadrate; radial sector short; radiomedial crossvein vein ex- tends from radial cell; short spurious anal vein (Fig. 11, s) feebly indicated between second branch of 3rd and 4th anal veins.
Genitalia: male (Figs 21-25), with apicolateral portion of parameres membranous, explanate; fe- male, of normal buprestid type, without dorsoven- tral differentiation (see Bellamy 1988c: 416 for dis- cussion of coroebine ovipositors).
Remarks. - Coraebosoma may be distinguished from all known coroebines by the very distinctively shaped and ventrally produced prosternum, espe- cially the strongly angularly bilobed mentonierre (e.g. Figs 1,7), a character state which defines this as a monophyletic group. The single bifid tarsal claw of each tarsus is a character state shared, to my knowledge, with only the Indian Mundaria Kerre- mans. Otherwise Coraebosoma shows closer affini- ty to other Philippine coroebines currently placed in the genera Cisseicoraebus Kerremans, Coroebus Laporte & Gory and Obenbergerula Strand. The Philippine species of Coroebus which are most closely related to this complex will eventually be transferred to Cisseicoraebus. Obenbergerula is currently being revised (Bellamy in prep.). Further comments on these relationships are made in the section on phylogeny.
Key to the species of Coraebosoma 1. Anterolateral area of prosternum entire, flat-
tened (Fig. 6); integument melanistic; black se-

ENT. SCAND. VOL. 21:2 (1990) Revision of Coraebosoma 199
Figs 1-11. Key and general morphology of Coraebosoma: (I) C. violaceum; (2, 3, 4, 6 ,9 , 11) C. indicum; (5, 8, 10, 12) C. sibuyanicum; (7) C. panayense: (I) ventral aspect; (2) lateral aspect; (3) head, right front aspect; (5,6) antenna, dorsal aspects; (8) profemur, protibia, dorsal aspect; (9) metafemur, dorsal aspect; (10) protarsus, anterior aspects: IOa, male; lob, female; (11, 12) left hind wing, s = spurious vein. (Scale bars = 1 mm)

200 Bellarny, C. L. ENT. SCAND. VOL. 21:2 (1990)
tae covering all portions of upper surface not covered by golden setal patches (India) . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . indicum sp. n.
- Anterolateral area of prosternum concave, de- pressed between mentonierre and hypomeron for antennae in repose (Fig. 7); integument otherwise colored, if black, with greenish or aeneous reflection, upper surface appearing glabrous except for setal patches and fasciae (Philippines) . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 2
2. Upper surface without concentrated setal pat- ches and fasciae; integument violet blue (Min- danao) . . . . . . . . . . . . . . . . . . . . . . . . violaceum sp. n.
- Upper surface with concentrated setal patches and fasciae; integument black, various shades of green or aeneous reflections (Islands north of N9"). . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 3
3. Upper surface bicolorous, head and pronotum differently colored than elytra; setal patches on dorsal surface small, fewer. . . . . . . . . . . . . . . . . . . . . 4
- Upper surface more or less unicolorous, ground color of head, pronotum and elytra similar al- though reflected elytral colors may be different; setal patches on dorsum more extensive, nu- merous . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 6
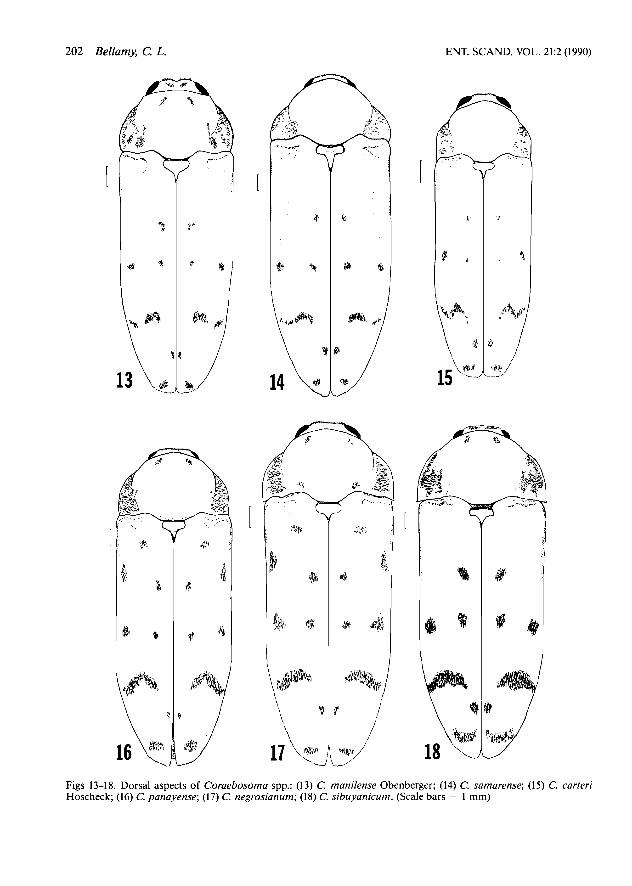
4. Head, pronotum bright cupreous; elytra brigh- ter green aeneous; more robust, L/W < 2.7 (Samar) . . . . . . . . . . . . . . . . . . . . . . . samarense sp. n.
- Dorsal coloration otherwise; more slender, L/W > 2.9(Luzon) . . . . . . . . . . . . . . . . . . . . . . . . . . 5
5. Head, pronotum bluish purple; elytra dark green aeneous . . . . . . . . . . . . . manilense Obenberger
- Head, pronotum roseocupreous; elytra black with dark red cupreous reflections . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . carteriHoscheck
6. More robust; upper surface feebly green aene- ous; each elytron with no more than 5 setal pat- ches and 1 transverse fascia (Sibuyan, Romblon) . . . . . . . . . . . . . . . . . . . . . . . . . . . . sibuyanicum sp. n.
- More slender; upper surface darker colored; each elytron with 7 setal patches and 1 trans- verse fascia. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 7
7. Color darker, very slight feeble blue iridescence on dorsal surface (Panay) . . . . . . . . panayense sp. n.
- Green or green blue iridescence quite evident on dorsal surface (Negros) . . . . . . . . negrosianum sp. n.
Four species groupings are apparent and are cor- roborated both by morphological and geographical data. The first group is based on the type-species rnanilense and also includes carteri and sarnarense, these three from the two north and east islands of Luzon and Samar, respectively. The second group is composed of the three species from the central is- lands, sibuyanicurn from Sibuyan and Romblon, panayense from Panay and negrosianum from Negros. C. carteri seems to bridge the gap between the first two groups. The third group is solely com- posed of violaceum from Mindanao. The final enti- ty, indicum from southern India, is apparently the least derived member of the genus.
Coraebosoma manilense Obenberger (Figs 13; 21)
Coraebosoma manilenseobenberger, 1923: 33; Hoscheck 1931: 159; Obenberger 1934: 815.
Material examined. - Holotype 0 (NMPC #23673): Philippines, Manilla; 1 Q (NMNH): Mt Makiling [Ma- quiling N14O08' E121°121, Laguna, 21.v.1931, F.C. Had- den.
Holotype male: 16.6 x 5.6 mm, L/W 2.96; head, pronotum dark purple, elytra green with aeneous reflections; underside dark aeneous; frontovertex with two small oblique setal bands between anterior portion; median depression anterior to antenna1 in- sertions, moderately clothed with recumbent setae; pronotum with two oblique setal bands on anterior !A of discal convexity; lateral explanate areas of pronotum moderately covered with short recurved setae; elytra each with setal concentrations forming five small patches and one transverse fascia as in Fig. 13; ventral surface sparsely yet evenly covered with short golden setae, these becoming longer and more dense on mesepimeron, lateral !A metacoxal plate, at apicolateral angles of abdominal sternites 1-4; small area on disc of metasternum and sternite 1 feebly depressed, setae slightly more concentrat- ed, apparently to act as pad for dorsal surface of respective femora; entire surface generally shallow- ly irregularly punctate, each puncture with one short rufobrunneous recurved seta on dorsum; lateral portion of each elytron transversely rugose; lateral margins of elytra subparallel to before apical !A; PN 1 . 8 ~ wide as long; anteromedial portion of pronotal disc strongly convex; elytral apicolateral margins slightly more serrate at separately angulate apices; inner tooth of bifid claw, subequal to outer tooth; genitalia as in Fig. 21.
Female differs from male: 16.1 x 5.6, L/W 2.87; tarsi the same except the inner portion of the bifid claw only reaches to near mid-point of outer claw.
Remarks. - This species comes closest in form to sarnarense described below and can be distin- guished as indicated below and in the key.
Coraebosoma samarense sp. n. (Fig. 14)
Material examined. - Holotype Q (NSMT): Philippines, Samar, Rawis, Basey [N11°17' E125O041, 17.v.1967, F. Schoenig.
Holotype female: 19.0 x 7.1 mm, L/W 2.67; differs from manilense: head, pronotum, cupreous, elytra

ENT. SCAND. VOL. 21:2 (1990) Revision of Coraebosoma 201
brassy green; head with vestiture sparse; lateral areas of pronotum only moderately setose; each elytron with five setal patches and 1 broken fascia as in Fig. 14; frontovertex wider between eyes; prono- tum with lateral marginal costae only subcrenulate.
Remarks. - C. samarense differs from its apparent sister species manilense by being more robust and the differences in the coloration. Further material from Samar will better define this species.
Coraebosoma carteri Hoscheck (Figs 15; 22)
Coraebosoma carteri Hoscheck, 193 1: 159; Obenberger 1934: 815.
Material examined. - Holotype 0 (MGFT): M. Imugan (?) - Luzon.
Holotype male: 13.3 x 4.5 mm, L/W 2.95; head, pronotum roseocupreous, elytra black with dark red cupreous reflections; underside shining aene- ous; median depression anterior to antenna1 inser- tions, moderately clothed with recumbent setae; pronotal disc glabrous; lateral explanate areas of pronotum moderately covered with short recurved setae; elytra each with setal concentrations forming five small patches and one transverse fascia as in Fig. 13; ventral surface sparsely yet evenly covered with short golden setae, these becoming longer and more dense on mesepimeron, lateral '/z metacoxal plate, at apicolateral angles of abdominal sternites 1-4; a fringe on sternites 3-5; small area on disc of metasternm and sternite 1 feebly depressed, setae slightly more concentrated, apparently to act as pad for dorsal surface of respective femora as in manilense; entire surface generally shallowly ir- regularly punctate, each puncture with one short rufobrunneous recurved seta on dorsum; lateral portion of each elytron transversely rugose; lateral margins of elytra subparallel to before apical !A; PN 1 . 6 ~ wide as long; anteromedial portion of pronotal disc strongly convex; elytral apicolateral margins slightly more serrate at separately angulate apices; inner tooth of bifid claw, subequal to outer tooth; genitalia as in Fig. 22.
Female unknown.
Remarks. - Hoscheck (1931) described this species from possibly Mt Imugan (N16'12' E120°52') a peak in the Zambales Mts north of Manila. One of the two examined specimens of C. manilense is from Mt Maquiling, which is also part of these mountains. This species is related to manilense by
virtue of its dorsal bicoloration and reduced dorsal vestiture but also comes near the sibuyanicum group with the similar ventral coloration and vesti- ture. It can be separated as indicated in the key.
Coraebosoma panayense sp. n. (Figs 7; 16)
Material examined. - Holotype Q (NSMT): Philippines, Panay Is., Culasi Antique, Mt Madja-as, Upaw [Upao? N11°18' E123"05'] (1100 m), 21.iv.1980, N. Kashiwai; 1 9 paratype (KACJ): Panay, 100 m, 14.iv.1984, T. Borromeo.
Holotype female: 18.9 x 6.6 mm, L/W 2.86; integu- ment black with dark green reflection which is most evident on elytra; head with two small oblique setal bands on frontovertex, depression between eyes moderately clothed with recumbent setae; prono- tum with two small obique setal bands on an- teromedial '/z of discal convexity, lateral explanate areas densely setose; each elytron with seven small setal patches and 1 transverse fascia as in Fig. 18; ventral surface sparsely yet evenly covered with short golden setae, these becoming longer and more dense on mesepimeron, lateral '/z metacoxal plate, at apicolateral angles of abdominal sternites 1-4, a fringe on sternite 3; setal pads on metasternum and sternite 1 as in manilense; entire surface generally shallowly irregularly punctate, each puncture with one short rufobrunneous recurved seta on dorsum; lateral portion of each elytron transversely rugose; base of pronotum sparsely subimbricate; PN 1 . 7 ~ wide as long, disc less convex, convexity slightly wider in proportion; tarsi with inner tooth of bifid claw much shorter than outer tooth.
The paratype has the right elytron slightly deformed basally.
Remarks. - With no male specimens known, C. panayense must be differentiated by its difference in overall proportion, integumental coloration and geographic separation from C. negrosianum.
Coraebosoma negrosianum sp. n. (Figs 17; 24)
Material examined. - Holotype 0 (NSMT): Philippines, N. Negros, Mambacal [Mambucal? N10°31' E123O051, iv.1988; 5 paratypes as follows: 1 0 , 1 Q (NSMT): same data as holotype; 3 Q (KACJ): same data except vii.1988.
Holotype male: 15.2 x 5.8 mm, L/W 2.62; integu- ment subnitid black, head pronotum and elytra
202 Bellarny, C. L. ENT. SCAND. VOL. 21:2 (1990)
Figs 13-18. Dorsal aspects of Coraebosoma spp.: (13) C. manilense Obenberger; (14) C. samarense; (15) C. carteri Hoscheck; (16) C. panayense; (17) C. negrosianum; (18) C. sibuyanicum. (Scale bars = 1 mm)

ENT. SCAND. VOL. 21:2 (1990) Revision of Coraebosoma 203
with blue green reflections, strongest on elytra; head with two small oblique setal bands on fron- tovertex, depression between eyes moderately clothed with recumbent setae; pronotum with two small obique setal bands on anteromedial % of dis- cal convexity, lateral explanate areas densely setose; each elytron with seven small setal patches and 1 transverse fascia; ventral surface sparsely yet evenly covered with short golden setae, these becoming longer and more dense on mesepimeron, lateral $5 metacoxal plate, at apicolateral angles of abdomi- nal sternites 1-4, a slightly denser fringe on sternites 3-5; setal pads on metasternum and sternite 1 as in manilense; entire surface generally shallowly ir- regularly punctate, each puncture with one short rufobrunneous recurved seta on dorsum; lateral portion of each elytron transversely rugose; base of pronotum sparsely subimbricate; PN 1 . 7 ~ wide as long, disc less convex, convexity slightly wider in proportion; tarsi with inner tooth of bifid claw subequal to outer claw; genitalia as in Fig. 24.
Variation. - Male, 1 paratype: 16.1 x 6.0, L/W 2.68; female, 4 paratypes: 16.9-19.6 x 6.3-7.5 mm, L/W 2.61-2.68; tarsi with inner tooth of bifid claw much shorter than outer tooth.
Remarks. - As mentioned above, this species is very close to C. panayense and can be differentiated as indicated under that species discussion. Both these species belong to a species group along with C. sibuyanicum, but differ by being slightly more slender, by the generally darker integument and by the increased number of elytral setal patches.
Coraebosoma sibuyanicum sp. n. (Figs 5; 8; 10; 18; 23)
Material examined. - Holotype o (NSMT): Philippines, Sibuyan I., Espana [N12"23' E122'301, vi.1983; 31 para- types as follows: 4 o , 2 9 : same data as holotype; 1 o, 2 Q Punong [N12"24' E122'311, v.1981; 2 o same data except, vi.1981; 4 paratypes simply labelled: Sibuyan Is.; 1 o , 2 Q , v-vi.1982; 1 o,3-8.1982; 1 o, 7 Q Romblon Dist. (or Prov.): iii-vii.1980; 3 9 same data except, v.1988; 1 Q , same data except, v.1985; 1 Q , same data except, iv.1986; 1 0, same data except, iii-v.1981; 2 Q simply labelled: Philippines, R. M. Lumawig; paratypes deposited in CLBC, GHNC, KACJ, MTCJ, NSMT, and NMNH.
One o specimen (NSMT) of this species is labelled with the identical data as two Q paratypes of C. violaceum (i.e. Mindanao), which is doubtful and therefore this one specimen is not designated as a paratype.
Holotype male: 16.9 x 6.6 mm, L/W 2.56; integu- ment subnitid black, head pronotum and elytra
with strong aeneous reflections, strongest on elytra; head with two small oblique setal bands on fron- tovertex, depression between eyes moderately clothed with recumbent setae; pronotum with two small obique setal bands on anteromedial % of dis- cal convexity, lateral explanate areas densely setose; each elytron with seven small setal patches and 1 transverse fascia; ventral surface sparsely yet evenly covered with short golden setae, these becoming longer and more dense on mesepimeron, lateral $5 metacoxal plate, at apicolateral angles of abdomi- nal sternites 1-4; fringe on sternites 3-5 denser than in negrosianum; setal pads on metasternum and sternite 1 as in manilense; entire surface generally shallowly irregularly punctate, each puncture with one short rufobrunneous recurved seta on dorsum; lateral portion of each elytron transversely rugose; base of pronotum sparsely subimbricate; PN 1 . 5 ~ wide as long, disc less convex, convexity slightly wider in pr~portion; tarsi with inner tooth of bifid claw subequal to outer claw; genitalia as in Fig. 23.
Variation. - Male, 11 paratypes: 14.9-17.9 x 6.0-7.0, L/W 2.48-2.55; female, 20 paratypes: 17.9-19.9 x 7.0-7.8 mm, L/W 2.55; tarsi with inner tooth of bifid claw much shorter than outer tooth.
Remarks. - The relatively large amount of material allows some analysis of variation for the only spe- cies known from two, albeit close, islands. The uniformity exhibited by this series in comparison to the smaller number of specimens of the other two species-group members, C. panayense and C. negrosianum helped in my decision to separate those two at species, rather than subspecies, level. These three species can be separated as indicated in the key, most noticeably C. sibuyanicum is the most robust, with the strong aeneous coloration, tne reduction to five elytral setal patches and the differ- ences in the genitalia between this species and C. negrosian um.
Coraebosoma violaceum sp. n. (Figs 1; 19; 25)
Material examined. - Holotype o (NSMT): Philippines, Mindanao Is., Mt Apo [N6"59' E125"16], Agko, alt. 1000 m, 4.x.1978, Shinji Nagai; 2 Q paratypes (NSMT): same data as holotype except, 23-30.xi.1982, native collector; 1 Q paratype from same island, Agusan [Ng030' E124'441, x.1976.
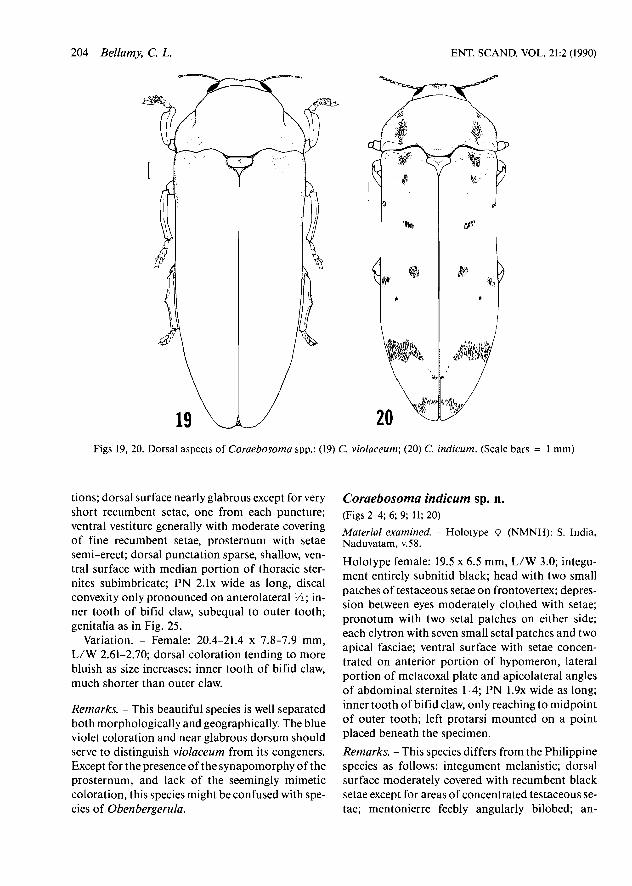
Holotype male: 19.0 x 7.1 mm, L/W 2.67; integu- ment steel blue, dorsum with strong violet reflec-
204 Bellamy, C. L. ENT. SCAND. VOL. 21:2 (1990)
Figs 19, 20. Dorsal aspects of Coraebosoma spp.: (19) C. violaceum; (20) C. indicum. (Scale bars = 1 mm)
tions; dorsal surface nearly glabrous except for very short recumbent setae, one from each puncture; ventral vestiture generally with moderate covering of fine recumbent setae, prosternum with setae semi-erect; dorsal punctation sparse, shallow, ven- tral surface with median portion of thoracic ster- nites subimbricate; PN 2 . 1 ~ wide as long, discal convexity only pronounced on anterolateral V i ; in- ner tooth of bifid claw, subequal to outer tooth; genitalia as in Fig. 25.
Variation. - Female: 20.4-21.4 x 7.8-7.9 mm, L/W 2.61-2.70; dorsal coloration tending to more bluish as size increases; inner tooth of bifid claw, much shorter than outer claw.
Remarks. - This beautiful species is well separated both morphologically and geographically. The blue violet coloration and near glabrous dorsum should serve to distinguish violaceum from its congeners. Except for the presence of the synapomorphy of the prosternum, and lack of the seemingly mimetic coloration, this species might be confused with spe- cies of Obenbergerula.
Coraebosoma indicum sp. n. (Figs 2-4; 6; 9; 11; 20)
Material examined. - Holotype Q (NMNH): S. India, Naduvatam, v.58.
Holotype female: 19.5 x 6.5 mm, L/W 3.0; integu- ment entirely subnitid black; head with two small patches of testaceous setae on frontovertex; depres- sion between eyes moderately clothed with setae; pronotum with two setal patches on either side; each elytron with seven small setal patches and two apical fasciae; ventral surface with setae concen- trated on anterior portion of hypomeron, lateral portion of metacoxal plate and apicolateral angles of abdominal sternites 1-4; PN 1 . 9 ~ wide as long; inner tooth of bifid claw, only reaching to midpoint of outer tooth; left protarsi mounted on a point placed beneath the specimen.
Remarks. - This species differs from the Philippine species as follows: integument melanistic; dorsal surface moderately covered with recumbent black setae except for areas of concentrated testaceous se- tae; mentonierre feebly angularly bilobed; an-

ENT. SCAND. VOL. 21:2 (1990) Revision of Coraebosoma 205
Figs 21-25. Coraebosoma spp., male genitalia, dorsal aspects: (21) C. manilense Obenberger; (22) C. carteri Hoscheck; (23) C. sibuyanicum; (24) C. negrosianum; (25) C. violaceum. (Scale bar = 1 mm)
terolateral portion of prosternum entire, flattened; PHYLOGENY mid-length projection on outer edge of metafe-
- mora only feebly indicated. C. indicum is most likely closest to the ancestral
stock of Coraebosoma with its less derived charac- ter states in comparison to the Indian Mundaria and the putative Philippine outgroups and con- geners. The zoogeographical quandary with the relatively large distance between the Philippines and the Indian subcontinent (Fig. 23, inset) might be better explained with subgeneric distinction, ex- cept for the lack of any convincing apomorhic fea- tures and is thus not supported by the species and area cladograms which follow.
I have not been able to pinpoint the exact type-lo- cality place name. There is a Naduvattam (N25'24' E8S004') listed in the gazetteer but this can no way be considered as 'southern' India. The only other close possibility is Nadavantam, in northwestern part of the Nellore district in Andhra Pradesh. Fur- ther material will help resolve the distribution of this species, but with the rapid demise of natural habitat in southern India, this is unlikely.
With the extensive buprestid surveys of Baudon for Laos (e.g. 1966) and Descarpentries & Villiers (e.g. 1967) for 'Indochina' not reporting any ap-
With the still rather chaotic state of coroebine tax- onomy awaiting the redefinition of the large and widely distributed nominate genus Coroebus, which is crucial to codifying the world genera, I have used broad tribal parameters in general and compared character states of three Philippine genera, Obenbergerula, Cisseicoraebus and Coroe- bus with which to establish character state polarity within the transformation series. These three genera and the Indian Mundaria belong to the Coroebus generic group and share the plesio- morphic ovipositor type (Bellamy 1988c: 416).
While Mundaria has a similar form, wing vena- tion and the synapomorphy of the single bifid tarsal claw, I have omitted it due to the lack of any male specimen. Obenbergerula has been left out of the outgroup comparison as it awaits revision and monophyletic definition.
I have used Cisseicoraebus grandis Kerremans and Coroebus pullatus Saunders as outgroups in comparison to the defined species of Coraeboso- ma. The characters used are listed in Table 1 and when viewed against the following discussion of polarity yielded the character state matrix (Table 2).
parently related taxa, the search for intermediates should focus in southern Malaysia and Indonesia, Character state polarity most likely Sumatra and Borneo. 1. Body proportions. - In the Neotropical, African and

206 Bellamy, C L. ENT. SCAND. VOL. 21:2 (1990)
Table 1. Adult characters employed (P = plesiomorphic; A, A2, A3 = apomorphic).
Consistency Retention index index
1. Body proportions: L/W r 2.6 (P); slender, L/W > 2.7 (A). 2. Integument color: iridescent (P); melanistic (A).
3. Dorsal coloration: unicolorous (P); bicolorous (A).
4. Dorsal vestiture: generally setose (P); setal patches (A); sparsely setose or glabrous (A2).
5. Lateral pronotal setal patches: present (P); absent (A).
6. Number of setal patches: irregularly fasciate (P); 7 + 1 fascia (A);
5 + 1 fascia (A2); 4 + 1 fascia.
7. Fronovertex: broad (P); narrow (A).
8. Pronotal disc: broad, flat (P); strongly convex, compressed laterally (A).
9. Mentonierre: entire (P); feebly bilobed (A); angularly bilobed (A2).
10. Lateral prosternum: entire, flat (P); excavate (A).
11. Setal cushions for femora: absent (P); present (A).
12. Mid-length metafemoral swelling: feeble (P); pronounced (A).
13. Tarsal claw symmetry: symmetrical (P); asymmetrical (A).
14. Tarsal claws: sexually equal (P); sexually dimorphic (A). 15. 2DA2vein: present (P); broken (A); absent (A2).
16. Spurious anal vein: present (P); absent (A).
17. Apicolateral membranous lobe of parameres: small (P); expanded (A).
Table 2. Character state matrix for taxa and characters considered (plesiomorphic = 1; progressively apomorphic = 2, 3, 4; missing data = -).
Taxon Character number 1 2 3 4 5 6 7 8 9 1 0 1 2 3 4 5 6 7
1. Cisseicoraebus grandis 2. Coroebus pullatus 3. Coraebosoma indicum 4. Coraebosoma violaceum 5. Coraebosoma negrosianum 6. Coraebosoma panayense 7. Coraebosoma sibyanicum 8. Coraebosoma samarense 9. Coraebosoma manilense
10. Coraebosoma carteri
Australian coroebine faunas, a trend is apparent with the more derived taxa developing a slender and more cylindri- cal body.
2. Integument color. - In both the tribe as a whole and the family in general, iridescent or metallic coloration is the derived state.
3. Dorsal coloration. - Being more complex, bicoloration of the upper surface is probably mimetic and thus apomophic.
4. Dorsal vestiture. - Throughout the tribe and family, a sparse covering of setae is the logical progression from a glabrous integument. The setal covering can become more dense, the setae can become squamose, or the vesti- ture can form distinctive, probably mimetic, patterns. The complete loss of integument vestiture in an otherwise setose complex is undoubtedly apomorphic as well.
5. Lateral pronotal setal patch. - The trend from a dense patch of setae on either side of the pronotal disc to reduced or absent vestiture in coroebines is apomorphic.
ENT. SCAND. VOL. 21:2 (1990) Revision of Coraebosoma 207
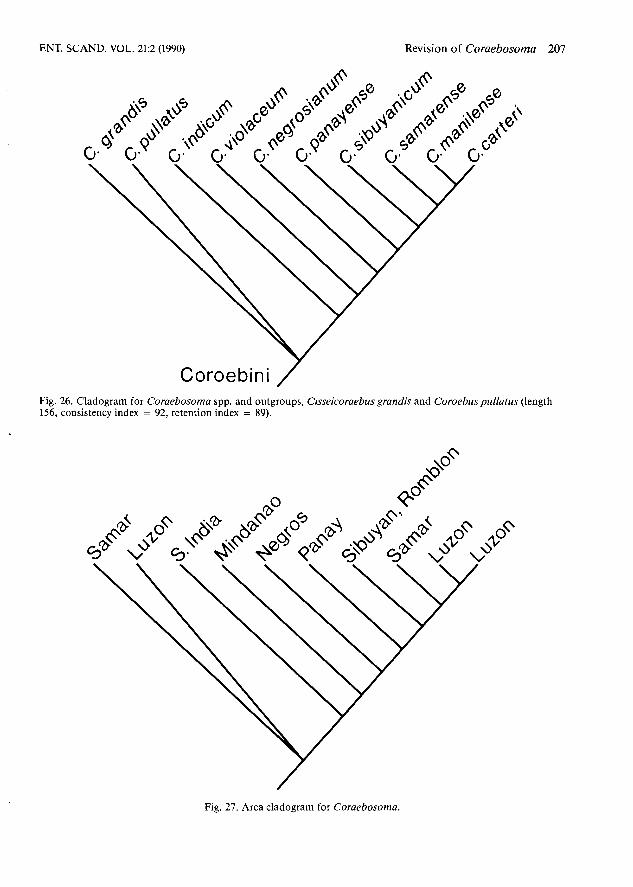
Fig. 26. Cladogram for Coraebosoma spp. and outgroups, Cisseicoraebus grandis and Coroebuspullatus (length = 156, consistency index = 92, retention index = 89).
Fig. 27. Area cladogram for Coraebosoma.
-(dew u!eu~ jo [I/, 'xo~dde jo ale3s) sau~dd~lryd ayl 01 e~pu~ u~o~j eaJe %u!~oys dew lasu! fspuels~ au!ddyyd ayl jo d~yy -82 .%!A

ENT. SCAND. VOL. 21:2 (1990) Revision of Coraebosoma 209
6. Number of elytral setal patches. - The gradual reduc- tion from a general vestiture through the transformation series of an increasingly smaller number of patches or fas- ciae is progressively apomorphic.
7. Frontovertex. - The narrowing of the frontovertex be- tween the eyes correlates with the transformation of body proportions. Thus, a narrow frontovertex is viewed as apo- morphic.
8. Pronotal disc. - The more complex and thus derived state of a strongly convex disc and explanate lateral prono- tum is defined as apomorphic.
9. Mentonierre. - The chronological progression from a simple entire anterior margin of the prosternum to the an- gularly bilobed state of Coraebosoma shows this to be a highly derived state.
10. Lateral prosternum. - The simple state of the flat and entire prosternal process is succeeded in complexity by the excavate apomorphic situation.
11. Setal cushions for femora. - The reduction of the general ventral vestiture to dense areas beneath the area of articulation for the meso- and metafemora is certainly a derived situation with the coroebines.
12. Mid-length metafemoral swelling. - The configura- tion, degree and placement of the setal comb on the hind femora proves to be at least generically distinct through- out the coroebines. Therefore, the more pronounced this femoral appendix the more apomorphic it is.
13. Tarsal claw symmetry. - The situation of only a single bifid claw on each tarsus is apomorphic in comparison to the commonality of the reciprocal state. This is the ex- treme of a transformation series from simple to appendic- ulate to bifid, with the appendices being symmetrical to asymmetrical.
14. Tarsal claw sexual dimorphism. - Sexual dimorphism is generally held to be apomorphic.
15. Second branch of second anal vein. - A transforma- tion series starts goes from the vein being present and at- tached basally to being free to being absent.
16. Spurious anal vein. - The presence of this anal vein be- tween the 3rd and 4th branches is more complex and thus plesiomorphic in terms of buprestid wings. The absence of this spurious vein is the more derived condition.
17. Parameres, lateral lobes. - The presence of mem- branous lateral lobes on the parameres of coroebines is apomorphic in comparison to the tribe in general. The further expansion of these lobes is more apomorphic.
Cladogram calculation and construction
Hennig86 (Farris 1988) is a very compact yet power- ful program and the reader is referred to the review of Fitzhugh (1989) for commentary on the various routines and cladogram-calculating commands.
Data from Thble 2 was input and the implicit enumeration algorithm (ie) was summoned and ini- tially produced ten equally parsimonious trees of length = 31; consistency index (CI) = 70 and reten- tion index (ri) = 60. Two runs of the successive ap- proximations a posteriori weighting routine (xs w ) yielded one tree (Fig. 26) of length = 156; CI = 92; (ri) = 89 with the individual character values for both indices listed in Table 1.
By following the methodology detailed in Hum- phries & Parenti (1985), the area cladogram (Fig. 27) was produced to corroborate both the actual distribution (Fig. 28) and the single tree from Hennig86 (Fig. 26).
Acknowledgments
I would like to thank the following curators and colleagues who searched their collections, some without success, for specimens of Coraebosoma for this study: S. Bily, NMPC, Prague, for the Obenberger type; M. Brancucci, NHMB, for the Hoscheck type from MGFT; R. P. Dechambre, MNHM, Paris; M. D. Kerley, BMNH, Lon- don; G. A. Samuelson, BPBM, Honolulu; Gayle Nelson, GHNC; and especially so to my colleagues from Japan for providing me with the bulk of material described herein: Y. Kurosawa and A. Shinohara, NSMT, Tokyo; M. Toya- ma (MTCJ), Nishinomiya and K. Akiyama (KACJ), Yokohama.
References
Arnett, R. H., Jr. & Samuelson, G. A. 1986. The insect and spider collections of the world. 220 pp. Leiden.
Baudon, A. 1966. Catalogue commente des Buprestidae recoltes au Laos. Mem. Soc. r. ent. Belg. 30: 1-80.
Bellamy, C. L. 1985. A catalogue of the higher taxa of the family Buprestidae (Coleoptera). Navors. nas. Mus. Bloemfontein 4: 405-472.
- 1988a. New genera, species and synonymy of African Coroebini (Coleoptera: Buprestidae: Agrilinae). Coleopts Bull. 42: 173-192.
- 1988b. Studies in the African Agrilinae, Coroebini VI (Coleoptera: Buprestidae). J. ent. Soc. sth. Afr. 51: 101-114.
- 1988c. The classification and phylogeny of the Aus- tralian Coroebini Bedel, with a revision of the genera Paracephala, Meliboeithon and Dinocephalia (Coleoptera: Buprestidae: Agrilinae). Invertebr. Tax- on. 2: 413-453.
Descarpentries, A. & Villiers, A. 1967. Catalogue raisonne des Buprestidae d'Indochine. XIV. Coraebini (4e partie). Annls Soc. ent. Fr. 3: 471-492.
Farris, J. S. 1988. Hennig86reference. Documentation for version 1.5.
Fisher, W. S. 1921. New Coleoptera from the Philippine Is- lands, family Buprestidae, tribe Agrilini. Philipp. L Sci. 18: 349-447.
Fitzhugh, K. 1989. Cladistics in the fast lane. J l N. Y; ent. SOC. 97: 234-241.

'4JemuaU '8loqPua~s OOLS-XU '9t Ca~LqPunl 'SX009 0170dV :woq palaplo aq 01 sdnoi8 ielns!lied 30 s~u!idai 01 uo!~d!nsqns pue Ivuinol
.u!l 'p"3XO 'dd 86 + !!x iiydel -1aa 'PE6-Z8L '103 ~ld0al03 'p a~p!lsa.~dna 'PC61 - -%Oa%O!q 3!ls!p~13 '5861 '8 '7 '!lualed '1 '3 '~a!"ddmn~ 'PP-EI :I azmd .snm .~pu 'ppo yua .q~ .sap!lsa~dna '991-EEI :LI pa8 'snm .tooz .~l!m '111 ('103) uapy
ap sama% xneamou ap spas aun '~~61 '1 '~a%~aquaqo -saldna lap s!uluuay 1nz a%~llyaa '1~61 '8 'V '~3aq3~0~