rflp relationships of a-genome species in the genus oryza
TRANSCRIPT

九州大学学術情報リポジトリKyushu University Institutional Repository
RFLP relationships of A-genome species in thegenus Oryza
Doi, KazuyukiLaboratory of Plant Breeding, Division of Genetics and Plant Breeding, Department of AppliedGenetics and Pest Management, Faculty of Agriculture, Kyushu University
Nakano(Nonomura), MutsukoLaboratory of Plant Breeding, Division of Genetics and Plant Breeding, Department of AppliedGenetics and Pest Management, Faculty of Agriculture, Kyushu University
Yoshimura, AtsushiLaboratory of Plant Breeding, Division of Genetics and Plant Breeding, Department of AppliedGenetics and Pest Management, Faculty of Agriculture, Kyushu University
Iwata, NobuoLaboratory of Plant Breeding, Division of Genetics and Plant Breeding, Department of AppliedGenetics and Pest Management, Faculty of Agriculture, Kyushu University
他
https://doi.org/10.5109/24360
出版情報:九州大学大学院農学研究院紀要. 45 (1), pp.83-98, 2000-11. 九州大学農学部バージョン:権利関係:

よFac. Agr., Kyusゑ疑慧癬.,45(1),83-98(2099)
RFLP relationships of A-genome species in the genus Oryza
Kazuyuki Doi, Mutsuko Nakano Nonomura, Atsushi Yoshimura, Nobuo lwata and D皿ean A. Vauglian*
Laboratory of Plant Breeding, Division of Genetics and Plant Breeding, Department
of Applied GeRetics and Pest Management, Faeulty of Agrieulture,
Kyushu University, Ful(ueka 812-8581, japax
(Received on 」2Lly 31, 2000 and (uccepted on Azegzest 18, 2000)
RFLPs of 192 accessiens ef A-genome rice species were aRalyzed. ORe plant vvas used to
represent each accession assayed. RFLPs were detected for the combinations of Dra 1-digested
total DNA and tweRty-one sir{gle-copy genomic c}ones. A dendrogram was constructed using
塩e{∫PG懸A】ryieth◎d f沁】rrl a geRetic distaxce澱a頴x.αass澁ca雛。轟◎f乞he A-geR◎】me盤ce spec量es
based on RFLP analysis matched well with the conventional class血cation. The African annual
species, O. gtaberrimct and O. barthii, were not clearly differentiated by RFLP loci while the
As懸sρecies, a 3醜vα, O噛)ogen and O.?zivara, f〈)澱ed a c◎即茎ex whe∫e塗簸。轟e of癒e
species can easily be distinguished from each other. An Asian sub-group that is clearly
differentiated from other Asian A-genome germplasm was identified. This consists of some O.
γ麟ρ◎gon accessions that d重d n◎t c沁ster wi撫the cu茎tiva乞ed r呈ce buも罫om篭ed 1◎◎sely k賊
groups. Some accessions of O.蜘(導α蹴were closely a㎏ned to㎞(蓋ca rice cultivars wh丑e the
others, mainly from China, were closely aligned with Japonica cultivars. Most of O.
幽㎜輝漁accessions from Latin America fo㎜ed a group which was more closely rel就ed to
O. glaberri??za aRd O. barShii thak other species. Two accessieRs gf O. ggecmaepatecga,
however, were aligned with lndica cultivars of Asia. lt is possible that the A-genome wild rices
in Latin America are of two types. O. rufipogo?z from the Sepik river of Papua New Guinea was
嶽e◎晦獄aterta1・wh三。鼓had RFLP飴g艶e滋s二二◎R茎y蟄σ爆痴轟。回生.
INTRODUCTION Rice (O?’y2es sativg L.) is oRe ef the most impertaRt fogd crops aRd is the stap}e fogd
for more than haif of the world’s population. There are more than twenty species in the
genus Ory2a. Two of these species, O. sativa and O. glabeMcrua Steud., are cultivated
rices. Wfid OTyga pessess geRes for resistakce to cliseases, insects Emd Stress tolerafice,
but these have not been fully exploited in rice breeding programs.
In erder to use usefu1 gen’es from wild rice speeies, wide-hybridization techniques
have been developed. New rice cellecticRs have beeR added to the already ceRserved
germplasm in recent years (Vaughan, 1994). Molecular markers were identified and used
to select p}aRts carryiRg iRtrogressed useful geRes. lnformatioR on phy}ogeRetic
reiationships ef wild species also help in efficieRt gene transfer. lf useful geRes Were
found in germplasm collection, a donor acces’ 唐奄盾氏@closely related to target breeding lmes
ca油e se王ected based oR the phy至◎genetic蚕f◎㎜aもi◎轟.
Nuclear restriction fragment length polymorphism (RFLP) is now considered a
powerful tool for genetic analysis and plant breeding. RFLP analysis is sensitive to
“ National lnstitute of Agrobiological Resources, Kannondai 2-1-2, Tsukuba, lbaraki 305-8602, Japan
83

84 K ZI)oi et al.
genetic changes at ’the DNA leve} at both codiRg and 一Ron-coding regions. More
importantly, it can be performed easily over many loci. RFLP analysis, therefore, has
beeft widely used te study phy}egeRetic re,latioRships auaeRg populatioRs-aRd species (e.g.
SoRg et al., 1988; Mi}le/r aRd TaRks}ey, 1990).
The A-genome species (O. scttiva eomplex) in the genus OTgea were the most
w董de至y鷺sed㎞rice breedi嚢菖. Th¢y share the same geno搬e asαsα伽αand搬ei◎tic
chromosome pairing of the Fi hybrids is almost complete (Katayama, 199. 7), although
showing various leveis of sterihty (Morishima, 1969). Asian ’wild A一一genome species, O.
rwfipegon semsu gacto was collsidered as wild rice relatives o£ Asian cnitivated rice. lt is
ta,xonomically divided into two species, O. rzLfipogon Griff. semsze stricto and O. nivara
Sharma et Shastry (Sharma and Shastry, 1965), on the basis of morphological and
ecOl◎gic滋charac乞e践s綴cs. in Afr韮ca, a㎜魏a玉cult主vaもed rice,0。99αわθ二曲α, a㎜ua圭w聾d
ぬce,0.わα7号二二A. Chev. and perem醸al w慧d rice,0.乙σ㎎乞蕊α鵬魏α孟αChev.就Roe㎞., are
fozmd. ln Latin America, O. ylzemcte, patzela Steud. is found although they are sometirnes
㎞d嚢ded㎞t◎0.γ藩札)ogo箆. AustraiSaR a㎜uaYt)ie漉a玉A-gertome w宝玉d hc¢is cα漏6{オ0.
men(二二。?zα髭s N9.
Nakano et al. (1992) studied on RFLP variation of the Asian A-genome species., They
ideRtified a group of wild rices which was clear}y distiRct frem O. satives and O.
rztfipogon. The group (cluster 3) did not correspond to any taxonomical classifTication.
Artalyzing RFLPs of accessions from broad range of A-genome rice germplasm collections
heips te understand the geRetic base of the group. lt also prevides detaRed irrformatioR of
phylogenetic relationships of A-geRorae ri¢e species.
Genetics o£ A-genome rices had been extensively studied (for review, ’Oka, 1988).
The present RFLP survey is also a eoRtinuatigR of the previous extensive st“dies (e.g.
Morishima, 1969, for raorphologocal characters; Second, 1985, for isozymes; Da}ly and
Seeond, 1990, for chlQrep}ast genomes).
MATERIALS AND METHODSP叢a蹴t搬ate】蟻a】お
RFLPg. of 67 rice aecessions, consisting of O. sativa, O. rafipqgo7z, Q nivctra, O.
glabeMma, O. barthii, O. longistaminata, O. glzemaepatuta and O. meridionalis,
were aRalyzed. Since Nakano’s Cluster 3 probably beleRgs to O. nivara, seme accessioRs
of O. nivara as well as peremrial O. rLefipogon with quite different’ ecology (ORI-OR 15,
they are pereAnial aRd grew in the area where no eutivated riee was grown) from O.
nivara were added. The data were cerabined with t-hose from the previeu,s}y anaiyzed
materials by Nakano et at. (1992). As a result, 192 accessions of A-genome species were
subjected te data aRa}ysis (Table 1). ’1]he seeds or leaves were previded by the Natiena1
1繋sゆte◎f Agr・bi◎1・gica1 Res◎urces, Ts殺kuba, Japa塗,撫e Natめ轟a11Rstit獲もe◎ごGe纏cs
(NIG), Mishirna, Japan, the lnternanional Rice Germplasm Center (IRGC), lnternatinal
Riee Research lnstitute, Los Bafios, Philippines and Kyushu University, Fukuoka, JapaR.
Species names used were those given by the souree of the seeds. One p}aRt was used to
represent each accession assayed.

£鑑P羅雄認。?zεんゆs(~fA-9θ73〈)me $p8cte$ iR診擁9醗?濡&”99窃 85
Table 1. The 192 accessions of A-geRome species in the genus O?lyza ana}yzed iR the study.
AccessioR
number
V翫ri就yπ織目黛/
accession codeTaxa Origin SOurce i]
Number offragments 2)
re船・。
P4
@ ◎1%345678田◎123娼56789◎12345612345010203840506
聯翻麗瀦朧編譜㎝㎝㎝舗器㎝縄麗麗麗麗麗麗麗麗麗㎜麗㎝㎝㎝㎝㎝㎝
NiPP(痴b段r琶
」臓職三4
1R24
Se礫naddi
Gaw批糠Damge Maruwa}{¢rOs藍]Bo歪&
具縫」●aS厩磁
Ngasein
浦&wDa膿
Mack KhamPulat ’Balaehan
BongorP磁哉も茎3磁鷲(至u
Nang Dum To
N琶oe Cht撒
N撚鷺ToiPUS1.蒸r
加m麓shimsllti
K:am硝i
Ge認d勲鷺Col/鍼kノ纂》a難.S総難ノ1987/1
Khao鷺oD無滋昭aPaR慕kai Kξ}pa至
MashoSltuaba
Ca照bo難ゆ◎Rg
MenalamSipio
Ket鼓難三》鷺.ik
厳a織
DhanRe(孟B融s澱a慧
,Bomsaj
CPSLONel〈1〈en 2
1R26
TKM 6N◎r註.篭8
Shen Shui Lian
Vear KrochakCo. 24
Suok LoryNang Di.}m To
Nang Quot
ααααααα櫛αααα蕊ααα窃αα繕ααααα蕊αα蕊αααααα腐αα撹α窃αα磁αααααα
野冊伽野冊伽伽魏卿野冊伽轡型伽轡型漉轡型伽轡型轡型蜘伽轡型魏轡型甲州轡型轡型轡型轡型伽魏野冊伽勘伽伽
αααααααααααα衡αααααα蕊ααααα蕊αα猛ααααα蕊αααααααα磁αααααα
Ss SS38
SSS3SS3
ρδ8SS8Sρδρδ8ρδρδ8ρδnδSnδ8nδnδρδSρδρδnδnδnδqUnδρδnδ3ρδSρδぐり8nδ
000000000()()000000000000000ハ)00000000000000000 000()00
ぬpauI簸d◎難¢si&
PhMppi,nessO繋縫.lc.撮鍛a
sO犠t量.竃。擁鍛a
Sri Lanka
Yunnan鍼y謎撚ar
N愈pal
Ass四三
B姻臓d¢shMya葺】Mar
Tha宝矯箕d
Laos
Ma蔵aysia
Malay$漁
M:麟ays至a
V蹴namV}.曾も鍛a買峯
Vl醸;三.峯凝轟
1ハdia
I捻d熱
JapaxJapanSo}2tl’i Araerica
P准鰭$嬢熱
1,nos,
Ph鐵興)熱¢s
1数d()鷺㊧s主a
MyanmarPh鎖iPP童ハes
P顯芸)茎)iXles
Ma1撫ysia
Indoxxesia
三田(圭(m¢$重騰
N(きpa}
Nepal,
Ne勲al
BangladeshMa互ay$i&
japaxP}灘やp漁¢s
India
Japan
c}漁a
Carnbodia
Cambo(髄a
C撫臓bo(難a
VietnamVietnewlt
紐欝懸㎜懸㎜膿㎜謙㎜㎜㎜㎜㎜㎜㎜謙㎜懸㎜㎜㎜㎜㎜㎜膿膿㎜㎜㎜臨㎜㎜紐欝欝譲

86
Table l. (Continued)
Accession
RwnberVariety name/
accessio難¢O(墨e
じま さノ
B◎ng Se纂
Chaw Ma-gawkLeuang Plah Sew
K1セao Pr隷9uad
Yena滅囎TaungdiBadobjota
jhul Digha
Asha通Bachogad/i
B司ramuri
Chakla-59
MadhukarKa茎ar Har$a縫
Taichng 65As◎m蹟◎yi
Kasalathvv’・ e2s
WIO6W12gW137W168]VSiT558
W593W596W600W625W629W630W1244W1297W1299W1625W1626W1627W1629W163iW1633W1634W1636wr1637
W1654W168gW1724wlsee(s)
W18e2-1
W1802-2
W1806-1
W1807W1811
K. Dot et aL.
Taxa Or㎏無
0.sα誕りα
0.5ati砂a
O. sativa
O.8αti雷α
0.s{蕊由り《茎
O. sativa
O.8α孟勿α
o.8attむa
O. saSivas
O. $ativa
Q sativaO.5撹認磐(満
O. sativa・
o.3α認餐α
0.sαt乞∂α
0。3α赫眉α
0.8α翻㊧α
o.gs・αわ8擁・蹴
O. rufipogon
O.蜘ogo7zO.γ吻)ogo箆
0.顧po9ぐ)鴛
0.吻取>agon
o.蘭)09㈱0.7吻)ogon.
O. rafipo.qon
O.蜘◎gono.謡歌)〈>9《〉%
0.吻ogonO. rLefipogon
o.me劉d乞07bdsgis
O.?駕γdl漁㎜£誌
0.7Zθア「d¢0㎜認8
0.meγ(オ乞〈)nαlts
O.耀箕オ乞。澱、認3
0. merdtonetds
O.耀7rd寂〉灘翻8
0. merdionaLts
O、me7(オ¢o綴認s
O. merdio’uatis
O。?舵γ(オ乞。?鶴ε器
0.晶晶ogomO. rzifipogon
O. rufipogon
O。吻og伽O. rztfipagon
O. rzefipogon
O.r2Lfipo9(ン泥
O. rafipogon
O.rufiメ)◎9《)銘
Vie猟a臓
Thaiユand
Tha簸歌nd
Tha且絃nd
MyamaarMyanniarBangladesh
BakgladeshBaxxg},actesh
Bai}gl,adesh
BangladeshIRclla
Ind壌
Ind重a
TaiwanJapaxx
In(戴a
WesもAf擁《)a
India (East Coast)
1紬畿(£asもCoasも)
India (East Coast)
Tha滋難dCarnbodia
Maiaysia
MalaysiaMalayg., ia
MyanmarMyanmarMyairmarNepal
A鷲sもr蘭a
Austr醗aAusもr漉a
Austra髄a
A幾sもra難a
Austral,ia
A慧sも獄蕪a
Australia
A秘stra難a
Australia
A慧str濾段
China王轟(慧a(Ea$t Cca$t)
China
Cambod重aBar唱諏(:i《≧sh
Ba㎎姶(蓬es鼓
Sri Lanka
Sri La罫.ゼka
Sri L川竹ka
SOurce i)
「
齪賀縦齪蟹劔昭轡型紐賀羅贋羅縦齪轡型㎜㎜㎜㎜㎜㎜㎜㎜齢㎜㎜㎜醗㎜㎜㎜㎜㎜㎜㎜㎜㎜醗㎜㎜㎜㎜㎜㎜㎜㎜㎜
Number offragmeRts 2)

飛砿Pretationshiρs(~ブ∠4-ge?zome spec乞es in thθ gen2ms OγZlxα
Taむ霊eL (C()譲;撫ミ1e(蓬)
Accession
Rwmber
Vanet,y nimxe/
acec?.ssi,ozz codeTaxa Origin Source s)
ヨノ
ミ ユリの ゆ ゑ ユゑ
937◎213706444448284052356
12δ118愈14954793344816666S
鶴顛覆強飯瀧魏欝蕩餅出端粥㏄㏄㏄㏄㏄㏄㏄㏄㏄㏄㏄㏄㏄㏄㏄㏄㏄㏄㏄㏄㏄㏄㏄㏄㏄
WWWWWWWWWWWWWWWWWWWWWWWWW脈旧獣蹴臓臓撫膿職臓獣翫m獣盤mm獄獄眠臓職膿撒獄
0.膿ρ090鴛O. r2tfipogon
O.麟塗)ogon
O. rufipegen
O.蘭)ogon
O.吻ogonO. mflpgg(m
O.蜘ogonO. rufipogon
O.蜘ogαno.膿茎)ogo?3
0. rafipogon
O. rLefipogon
O.γ感官)ogon
O.吻ogo?zO. rufipogon
O. rufipogon
o.噸ゆ◎gon
O.蜘ogenO. rufipogon
O. rufipogon
o.蜘ρgonO.鳩ρogonO. rufipogon
O.顧鈴ogo露。.わα?慧~乞乞乞
0.わαγthi乞
O. barthii
O. bairthii
O. barthii
O. barthii
O. barthii
O.わαγth乞i
O.わαγth乞i
O.わαrth乞i
O. glaberrima
O. gtaberrima
O. glaber7’ima
O.99αわθ綱灘
O. glabentma
O. glabe7yima
O. glabentmo
O.9脇わθ携灘
O. gt2Atmaepatula
O. glzemaepatzLea
O.gSac灘御ご%臨
。.9ど%徽銀伽鴛‘α
O. glnmcvepatz,ela.
O. gluwaepat2kla
o.99ze’脚厩鴛諭
Bariglades}i
Thailand
Tha猛aRdTkailaRd
China
ChinaCl*}a
China
China
China
Chi鍛a
ChinaChina
ChnaCh嶽aIndonesia
Indonesia
IRd◎嚢es玉a
IRdoRe$ia
Indonesia
In(懸a(Wes£C◎翫st)
IRdia (West Ceast)
IRdia rwest Coast)
Indja (West Coast)
擁ya㎜a.r
鍵a蓋
GuineaCha(美
Ta嚢zaRia
TanzaniaNigeria
C難a(圭
ChadCameroonCamerooRC()登go
Senegal
Burkina Faso
CarnerooR
Ivory Coast
Senegal
ChadCa応酬er◎◎益
CubaVenezuela
French Guiana
Braz銭
Braz銭
Braz且
Br3zg
ぼ ヨ エ エ ユ ス エ ズ ま ユ エ ぼ ユ ヨ ユ ユ
エ ズ
ヨ
ヨ
エ
ぼ
ま
エ
エ
黙臨黙器購黙鵬齢㎜購鷲㎜㎜㎜鵬灘㎜黙㎜㎜綴㎜綴㎜㎜綴㎜㎜綴㎜黙㎜㎜隙
87
Number of翻撫帥¢黙総融)

88 K Do{ et al.
Table 1. (Contmued)
Access’ion
Ruraber
Variety name/
acceSS圭◎叢Cく)(薫eTaxa 0類募in S◎磁rce且)
Number of癒ag澱e盤総津〉
34R6
R7R8
S1
S2
S3
S4S5
S7T1
T2
T3
T4
T6
T7T8T9U◎
U1
U2
U3
U4U6
U8U9V0
P23456789鐙1112131415
照綴綴照照綴照綴照照綴賑照照蕪蒸賑。R器㎝器㎝。R㎝。R㎝。R㎝㎝㎝
IRGC 105667
1RGC 105670翌RGC 1◎王2()◎
,IRGC 101202
1RGC 191216笠】驚GC lO12爆5
1RGC 104060
1RGC 104075
11RGC 104977
1RGC 1052e4
MGC 105306王】駐(}C1◎6319
1RGC 105428
1RGC 105444
1RGC 105715
1RGC 106e521RGC 1◎61ユ圭
IRGC 106153
1RGC le57eg
IRGC 105726茎】段GC 1()δ889
1RGC 105898
1RGC le59101R’
fC 106036
1RGC 106128
1RGC le6157
1RGC 106i66VN90一脇董~s}
PNG90-183)
PNGgi-73)
P92-3A3)
V92-3B3)
P92-3C3)
MY90-253)
PNGgg-273)
PNG90-343)
PNGge-4ei・>
MV89-863)
PNGge-le3}
即89-653>
PNG90-13r“)
PNGgo-14a,
O. glzkmaepat2Lga Brazil
O.g12smaepatzela Bramio.言。㎎琶8掛川客鶴鋭刃圭ger韮a
O. to’rvgtstamiuata Nigexla
O罵。㎎魏α癬灘翻v◎ry C◎asも
0.5(》㎎お雄?嬬乞?羅編M濾
O. tongistamiw)ata Nigeria
O. to7zgdstamiuata Nigeria
O. to?zgtsta?niuata Kenya
O.lk)ng乞1stcem乞uata Etk,韮○茎)ia
0.7㎜剛乞07zαεお
0.?忌む{蕊磐α
0.?認禦αrα
O. nt’vava
O. ni?nra
O. nivara
o.%証書γα
O. nivara
O。聯(igonO.7隅惣)ogon
O,蜘ogαnO。吻ogonO.噛)ogo%
0.吻ogo箆0.吻ogonO.麟鈴ogo%
0.顧ρogoπ
0.吻agenO.吻ogono.噛)ogαnO. rufmpogon
O.吻ogαnG rafipoge?z
O.糧ノ彰pogo7z
O.噛)◎gon
O、蜘ogonO.・吻09(m
O.噛)og侃O. vafipuoge?z
O.噛)ogonO. rufipuogon
O、噸毎)ogon
Austratia
掻(践a
Sri La醜ka
Sri L澱ka
CambodiaInd玉a
麺(搬
Laos三熟(海
Cambo(亜a
Bangladesh
Ba咽駐deshTha這and
Ma1鼓ysia
In(灘a
taes
Vietn翫m
Vie嶽a撮
㎜㎜魍㎜温麺㎜㎜㎜㎜躍瑠頭嚢㎜脳艦㎜盟認
IIτ三1¥孟董III}三1¥三冨三II頁至1τ11署孟1τ111τ三11
Pap職NewG磁曳ea IRR三Papua New Gninea IRRI.
Ph且ipp血es IRRI
P賊p脚es 茎RRIPh鋤幽es IRRI
Myanmar IRRIPapua New G漁¢a娘R韮
Papua New G漁ea IRR玉
Papua New Guinea IRRI
India IRRIPaP膿New G磁鵬a lRRI二胡{滋a 玉RR!
Papua New Gtmea IRRI
Pap膿New Gu瀧a玉R瓢
互}KY篇Kyushu U通vers童もy;NIAR :Nat圭on撮熱蕊iもuも黛of Agr◎b三◎10g垂cal Resources;MG=Nati◎na}
Institute of Genetics; IRRI=lntem’iational Riee Resear¢h lnstitute.
2) Nimeber of fragneRts equa}s the tota} Rwwber of re$t}ictioR fragraeRt$ present in each accessioR
across 21 clones.
3}0晦1舩ves w琶reμov三d磁.

RFLP・7’e9・αtiひ?wshix)s ofA-gen(nne species ’i.?z the geve”btsLs(:吻雛 89
Table 2. Prebes used in this study.
Probes ChromosomeNumber of unique
£ragraent$ detected
Npb54Npb 113
Npb67Npb132NpblsNpb 129
醤pb49
Npbl14Npb25Npb81Npb12Npb27Npb33.Npbi17
Npb126Npb13NpblO8Npb32Npb37NpbllsNpb88
111圭
1 1
RFLP prebes RFLPs were detected from the combinatioRs of DTa 1-digested total DNA and
twenty-one single-copy genomic clones (Saito et at., 1991) listed in Table 2.
Data a戯田【ysis
Each fragment was treated as a unit character, although fragments detected with one
e}one rRay be alle}ic. The c.haracter was quantified as 2 and O for the preseRce axxd
abseRce of the fragmext, res,peeti’vely.
Genetic distances between accessions X and Y were calculated following the formula,
D雛一伽【2ル鰯κルfx≠砿,)】, where払and嶋were the number of total fragments irし
accessioRs X aRd Y, respeetive}y, aRd M., was the fiiueber ef corarrioR fragmeRts observed,
between accessieRs X and Y. Cl]his foriRula ls based upoR Nei’E, standard geRetic distance
(Nei, 1987, p 220 formula 9.24) wtth the accessions assumed to be homogenous. A
dendrogram was then construeted ustng the UPGMA method (Sokal and Michener, 1958).
Al}d就a analyses as well asも掘。(msも澱。もi◎R of the(ie黙(irogram were do塗e◎nもhe
Micr◎soft Excel apPhc就ion pr(鷹rara.
Measures of genetic vamiation wtthin aRd between groups were done fol}owing that of
Wa鷺9θ孟鰯.(1992). Gen磁e var鼠aもi◎n賊㎞groups was esも㎞襯d as the搬ea鍛ge簸etic
distances of all pair-wise comparisons between differeRt accessions from the same group.
Genetic variation between groups was esttmated as the mean distances from different
groups.

90 K Doi et at.
The relationship of Asian A-genome species was difficult to understand from the
UPGMA deRdrogram. ORe-huxdred forty ene accessicRs frorri Asitt, therefgre, wffre
analyzed by principa], component ana}ysis (PCA). PCA was done using PCA cornrnand of
MacM思(rrhigulegse, 1990)wiもh C¢魏ered PCA◎Pもi◎貧.
RESULTS
Pelymorphism As the 21 probe$ used in thi.s study were single copy clones from O. sutiva, almost al}
the O. saniva acessioxis kad 21 fragmeRts. The tetal riumber of observed fragrRents per
accession ranged from 19 to 34, with an average of 21.6. A total of 111 fragments was
ebserved with a}1 21 probes.
vari就ion of A-genome species
The RFLP-derived dendrograrn is shown in Fig. 1. Groups of O. 77?iewiclionatis, O.
tongistes’nzinata, and O. ggzsmaepatzegg frem Brazil were ideRtified. These RFLV
groupings corresponded well with the conventional classMcation supplied from each
source ef /p}aRt materia}s. However, the African aRriual speeS,es, G ginberwima and O.
barthii, were not clearly differentiated. ‘O. ggesberrimas’ RFLP-group, therefore,
coRtained both O. ylaberrima and O. barthii. The Asian species, Q sativa, O.
rafipogo’re and O. ni’vara, formed a complex wherein none of the species can be clearly
separated from each other. IR Fig. 1, four labels, ‘lnclica’, ‘Ja,penica’, ‘O. ?zivava’ aRd ‘O.
rzLfipogo7z’ wer,e used for the sake of coRveRience. Two RFLP-grgups, ‘IRdica’ and
‘Japenica’ raainiy con,tained lndica and japonie.a cu}tivars, respectively. Each of them
contained f everal wild accessions. The group labeled ‘O. niv(zTa’ was separat,ed from the
other Asia.n greup$ and joiRed O. g. lzemaepautzeta aRd O. ggaberrima. Other Asian
accessions which clustered with neither cultivated rice nor ‘O. nivara’ RFLP-group
formed loosely knit sub-groups.‘0.γゆjpogo?ガR肌P-group is a complex of these
accessions. The three groups labeled ‘lndica’, ‘Japonica’, and ‘O. rztfiipogon’ were the
most c}osel.y related.
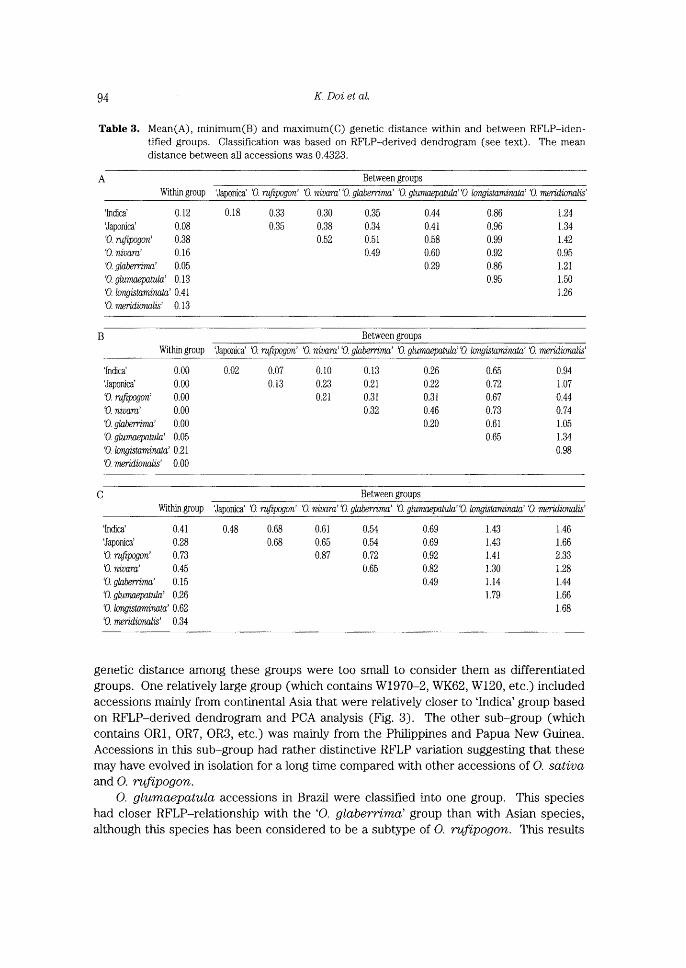
Variation both wit㎞and between the eight groups was eva lu就ed. Mean, m血mum
and max㎞㎜genetic磁stances賦㎞and between the groups are sho㎜血Table 3A,3B and 3C, respectivel,y. O. long’i s’taminata $howed the }argest variation withn group
and O.9♂αわθγワ吃η②α(including O.~)αγ〃麗のthe$∫れanest. ‘0. rufipogり7ガ繕roup s重rnilarly
showed relatively large variation.
Based oR the mean values of the distance be.tween greups (Table 3A), a dendrogram
of all species gro疑ps was co鍛s漁。もed(Fig。2)。至蓑もhまs dendr◎gram,つ.鴛勿α曜
RFLP-group was geneti.cally more ed,osely related to the other Asian groups.
PCA was performed to determine further the relationships of Asian species (Q
8α‘勿α,0。rufig)ogo鴛,0.7乙勿α磐α), b慌exclud㎞蔀ORI2,14 a:朕i l 5(see(灘scussion)(F嬉.
3).至R嶽ePCA,撫e f重rsもaRd sec()簸d axis c◎ぞresp◎難dedも。 IR(iica-Japo擁ca aRdつ.
nivara’ group-other groups differentiation, respectively. The aceessions from ‘Japoniea’
RFLP-greup formed a small group while ‘lndica’ accessions were scattered to a wide area,
refiecting large variation. Accessions of ‘O. 7&twesra’ group forrried a distinct group
comple慧e匪y separated鋤◎m a塁accessi()盤s.

9
鴨
で.
ηO
鯉薯馨0
“.
9
き
^轟 議言語§
91-92
き葦1鑛晶灘ll馨,融嶽1灘書1ぎ聾藤糠量1難葦趨蠣1繍礫1融蕪1紬1§1垂癌葦懸盤llll§§薯騒1雪幽灘垂曇蕪1轟量聲ll灘1馨鑑壽茎1,鑛難lll墾
羅羅羅墾難ll纒纒纒1纒器撫垂鍵櫻羅遡蔓灘鍵鰹餐灘鰹葦灘鍵灘鍵遡窒鰹灘灘翼襲鰯19鑓蝿麟襲襲1灘1縫朧朧1羅難1撚難纒羅懸1鎌
蓬垂塁艶塁垂塁塁塁錘望垂錘垂難垂垂錘垂垂塁垂麗垂垂聾錘難垂垂塁塁塁塁望塁望塁塁望塁望箋塁垂箋難箋箋箋箋箋箋≧1≧鼓鼓塁望塁箋鼓垂望塁垂塁鯉垂鼓難箋箋箋箋鼓鼓並望塁妻玄箋垂箋箋並望i塁呈垂箋箋蓬望塁望蓬望望並箋箋…箋箋箋堂塁1箋箋箋箋箋箋elll箋塁望塁塁箋箋鼓箋雲望塁垂望垂垂聾難垂垂垂垂難垂聾難睡垂難垂垂垂望望塁塁讐塁塁望塁望垂
婁禦難難鍵鍵畢霧ll攣塁難灘睾睾llll峯藝ll蕪ぎ1望朧§麟毒蘇隷1謙麹垂1塁翻藤灘難馨要§馨讃1塁馨馨§塁難lll馨馨§馨E99$§馨§1§灘1馨llll垂馨蓬譲攣華難婁麟朧奮麟選塁難嚢lll難lll塁
O.O
聯.
n
曽.O
8彊置暑ワ毒
ηO
“.
n
,.
n
O.
n
の①O』づOのΦ出一二O屑◎oO一〇刷ρO』堺璃OΦρ【5一一〇り口【一存90一ρ邸7H目餌く同Z
.¢」邸』QQO』で昌Φで①‘9肖O℃①の邸ρ口Oお邸O娼刺ωの面一〇山q』出(Cq .①一つρ5の自円‘O』邸ΦのΦ出ΦO咽餌一当⊆Oお“9』①一⊆一11一図国一…のOお①自①O駒O①一⇒溜θのd[【州邸口O冒邸ブ[110一Z
…あ起g⊃旨①〉唱昌hPづ高の昆〉{11強 (H .oう①自〇一〇〇屑已OqΦbO 円㎝二9膠でΦ一〇Φθ㊤℃①』①≧gり日日』国 .oり口O咽のの①OO邸㎝①円 目病}O∈邸』QOO』づ⊆Φでで①〉刺』①℃一山日』¢
.祠.ヒの個

R」FTLP retationshipus ofA一 ge?zome spectes tn the genzts Orgxa 93
RFLP c韮縦ss圭f茎cati◎霊
。.o
Ge燕e重至。 dis捻難ce
O.1 O.2 O.3
警1簸dica嚢
饗Japonica聖
’0.r吻080パto. nivara’
’0.glaber伽a’
り.8Z麗〃saep伽1ゴ
’0.tongistamin磁’
’O. meridienalis’
F㎏.2.De慮r◎奮r撚of A-geRolke species ik the geRus()耀羅碗説d◎就鼓e recalぐ幽轍:i geRet呈。
distarlce a膿◎盈g RFLP-gro鋤$s}1◎wn類Tab圭e 3A(}職yわ段rs類d量caもe sもaRd欲rd d¢v繍◎蓑
of genet,ic distance at each branch.
DISCUSSION
AR average of 5.3 (lll/21) mique fragmeRts per prebe is betweeit the 3.4 aRd l l.2
0bserved in previeus studies which ,iRc}“ded 70 acce$sioRs ef O. sativa (WaRg aRd
TaRksley, 1989) and 93 accessioms from the whele genus Ory2a (Wang et ag., 1992),
respectively. This value is reasonable because the germplasm used in the present study
covers A-genome rices.
NakaRe et ag. (1992) ideRtlfied, a uniqge grgup of wild, rice accessioRs (Clxster 3). ‘G
7zivara’ RFLP-group tR the present study corresponded to Nakano’s cluster 3 (Nakano et
al., 1992). lt contains 4 out of 7 aeeessions of O. nivara identified at IRRI whil,e only one
out of 24 ac¢essions of O. rafipogon from IRRI was included. The results of this study,
therefore, suppert the coRcept of the taxoRomic speeies O. iyzivara. RI‘’”LP aRalysis
shewed that thi,s greup is clearly differentiated from other Asian A-geRome germplasra,
although we have not yet been able to find any clear key morphological eharacters to
distinguish all members of ‘O. ntvara’ RFLP-group from O. rafipogon. However, since
ge鼓e fbw◎ccurs be榊ee費A-gerの澱ff O型gαspecies g灘ow沁g sympa頃。認y㎞As重a, c王ear
differentiation ef these taxoRomie species is quite diffuu}t. RFLP markers, as shoiwn
here, were usefu1 in identifying specMe groups within the species complex.
In this study eight accessions of O, rafipogon were m’ the ‘Japonica’ group. Six of
these accessions came from Chna. This irnplies that the geRetic base of Japoniea is iR
Chirta. Accessioms of O. rufipegon frora raaiiy Sgnth and, Southeast Asian cowwtries were
aligned with lndica varieties. This may refiect the fact that in the lowland tropics of South
and Southeast Asia today most varieties are lndica varieties and in many areas, grow
sympatrically witk wild rice. ReceRt $tudies (WaRg et ag., 1992; NakaRe et ag., 1992) have
shewn that some populatieRs of O. r?ofipogon and O. nt?,xxras cofr{plex have a very close
relationship to cultivated rice.
Accessions of ‘Q rztfipogon’ RFLP-group were mainly from Southeast Asia. They
appeared to forin two relatively }arge sub-groups aRd other small groups, although
94 K Doi et at.
Table 3. Mean(A), minimum(B)and maximum(C)genetic distance within and between R肌P-iden一
麟ed gro贈s.αass漁caも沁蓑was b我seδoR RFLP-der呈veごdendrograra(seeもexの. The meaR
distaRce between all aceessioRs was O.4323.
A Between groups
Wlも}療9罫()競P ‘JaP◎搬(殺’‘0.離ρρρo冨 ‘().?zivara’‘σ9㎞7澱’‘O. 99u?㈱暮露’‘(〉晦stSMiuatCl’‘◎.繍鰍奮’
‘indica’
‘Japonica’
‘σ聯90π’
‘α箆魏α澱’
‘o.9㎞徽’
‘0.g伽脚漁’
O.12
0.08
0.38
0.}6
e.es
O.13
‘σ晦5ごα魏琶胤α’0.41
‘o.meri(2rmtis’ e.13
e.18 3民」
◎0り0
◎nU
o.3g
O.38
0,52
O.35
0.34
0,51
g.49
O,44
0.41
0.58
e.6g
O.29
O.86
0,96
0.99
g.92
e860.95
1.24
1.34
1.42
愈.95
i.21
1.50
1.26
B Between groups
Within group ‘Japonica’‘α顧Pρσ(m’て), nivαrcゴ‘O. glαberrt??zα’て).σ1㈲剛掘α’て), Longismm乞uαtα’ ‘O. r dmais’
‘熱磁(譲’
‘japonica’
‘σ傾卿(m’
tO. nivara’
‘Q ggctherrima’
り.轟灘餓’
g.ge
o.eo
o.oo
o.oo
g.oo
e.es
‘σ脚3εα鵬勿αεα’0.21
‘ση認翻窃’0.00
e.g27りO
O11
Aり0
◎、鎗
O.23
021
o。圭3
0.2i
O,31
0.32
g260.22
0.31
0.46
e.2g
倉.65
e.72
0.67
0.73
C.61
0.65
C.94
1.e7
0.44
0.74
1D5王.34
0.98
c Between grogps
Wlthin group ‘Japonica’‘σγ頃Pq9㎝’て), Mvαrα’‘σ9励漉㎜’‘O. giuWWzdα’‘(λ幡じα禰ηαα’‘σ燃枷お’
tindical
‘蜘。題(x’
り,顧卿伽’
to. nivara’
‘O, gim7n(1’
り.轟鰍漁’
O.41
C.28
0.73
0.45
0.15
e.26
‘G白川緬櫨’・9.62
‘O.mendtonatts’ O.34
O.48 8◎0
突U6
0《り
O.61
&65
0.87
O,54
g.54
e,72
0.65
O.69
倉.69
e.92
0.82
0,49
1.43
1.43
1.41
1.30
L14玉.79
1.46
1.66
2.33
1.28
1.44
1.66
1.68
genetic distance among these groups were too small to consider them as differentiated
groups. One relatively large group(which conta血s W1970-2, WK62, W120, etc.)included
accessions mamiy from continental Asia that were relatively closer to ‘lndica’ group based
eR RFLPderived deRdrcgram aRd PCA aka}ysis (Fig. 3). f17he ether sub-group (which
contains ORI, OR7, OR3, etc.) was mainly from the Philippines and Papua New Gtmea.
Accessions in this sub-group had rather distinctive RFLP variation suggesting that these
may have evo}ved in isolatieR for a }oRg time compared with other acces$ioRs of O. $ativa
aitd O. rafipogon.
O. glzemcvejpatzeLa accessions in Brazil were classjfied into one group. This species
had closer RFLP-relationship with the ‘O. ggaberrima’ group than with Asian species,
althgxgh this speeie$ has beex ccRsidered te be a subtype gf O. rufipogo?L. This results

RFI乙P 7㌢脇認(〉?欝~z.塑s《~ブ滋一98鴛《)耀5カθcies客鴛書h8 ge捻zes oγ鋸9α・ 95
ゆEXIS 2
+許
÷
◇●◇
◎ ◇
0 ◇ ◇ axis 1
◇
◇◇◆ 〆◇
●
◆
◇●
◇◇ ■
● ●
XXX
XX
B ◎cX叢 3
十
十
手 ÷÷
X◇ ÷
葵 ◇ ÷虐
X十
X ● ◇÷
◇詑◇
0o
◆ ● ◇
◇×
●
◇●◇● &X董S蓋
O o♂)◇
00O ◇
◇
蓼◇
’◇◇3幅◇
◇
◇◇
O : ’Japonica’ in Fig. 1.
e : ’lndica’ in Fig. 1.
e ×: ’O. rufipogon’in Fig. 1.(e:continental Asia, ×: Philippines, Papua New Guinea)
十;’0.nivartゴ圭1}F曼9.蓋..
Fig. 3. Principal cornponent, analyg. is (1’CA) of 141 accessioris frorn the O. s.’atiwa, O. rzofi.pqqon
and O. nivara. The first axis extract 190/o of total variation, the seeond, ],20/) and the
third, l l OA,. Accessioms were pkotted in the p}ane def}ned by axes i aRd 2 of PCA (A),
a鼓dエa難d3(B).

96 K Doi et at.
(OZ廿日O碧コOgり)〇一出O
(邸
シ邑=娼O
(Oブ【山懐0§O楼)N一属○
(衷℃ε。・髭O
(OZ議日O調仁O麟)寸一桜O
(由
jZ窪盲0』岩O蛋)い一出O
1纏滋・圏 ・諺1野∵ ’
0. rufipogon
O. meridionatis
Fig. 4. Autoradiogram of six C). TzLfipogon accessions frorn lndia
and Papua New Guinea (ORIO・・OR15) probed with Npb81.
0斑2,0R14鱗d OR獅s熱owもwo b麟ds,◎難.e is tylpictt} cf O.
糧伽ogo?z wl.曲the oも糞竃er is typi¢al of O.澱腕d乞。骸傭.
support previous, reports based on ribosomal DNA variation (Sano and Sano, 1989) and
RFLP variation (Wang erml., 1992).
WK24 and WK:28 were◎rigixxaliy c玉as$i貸ed as O. g伽㎜αθpα溜α, but頽伽s s繊dy,
they were classifj,ed with Asian sl)ecies. ltis possible that there are actually two types of
O. gezLmaepatntas in Latin America, aR Brazilian race of Q ggzemasepatzsl.a whi(th is
edfferent’ from the types fouRd in other parts of La’tin Ainer2ca. Second (1985) regarded
the American form of O. jperenMs as a E ubtype of O. 7zefiipogon. However, he did not
test an: Braziliazz a,ccession. As the origin of O. etumaepuest2Lga is Rot, c}ear (Vaughan,
1989), it is ne¢essary to determiRe the full range of variation of wild A-genome
germplasm from Latin America.
Seven O. lo7aytstaminata aceessions (WK37, 38, 41, 42, 44, 45 and 47) had multiple
RFLP fragments with 4 to 8 probes and no sigRal with two probes, which indicates their
high hete. rozygosity. This species group also showed the largest intra-group variat;ion
(,Table 3). ln this study, howeve.r, Ro information oR intra-populatioR variation was
obtained as only eRe plant per accession was tested. Sjnee some useful, genes have
already been found in O. lo7zgist(xminata (Khush et aL., 1990; Maekawa and Tsunoda,
1994), a better ur}derstaRding of the divers,ity in this African perennial species is needed.

R泓Pretati◎7zship3(~ブノ4「9{?7ZO?ne speciθ5乞n君擁98?’tzes O瑠£矯 97
O. geabe7’rima and O. barthii showed little RFLP variation. O. ylaberwimo has been
reporもed沁have h撮e掻もra-spec撞。 variat韮◎鍛(M:iezan,1986;lsh,ti et a9.,エ988). Res嘘s
む◎m撫epresent study revealed that O.δαγ出品also h畿d誠ttle r1聖1cl鰍r RFLP variat沁n.
Q rztfipogon in Papua New Guinea formed two distmet groups. One group (OR3, I O,
2, 8 aRd 9) originated from the south of the country. The o’ther group (OR12, 14 aRd 15)
w’as cellected along t・he Sepik river iR the fiorth of Papua New Guinea. The aceessieRs
from Northern Papua New Gninea have RFLP fragments whi,ch are present in both O.
rztfipogon aRd O. meTidionalis (Fig. 4). These access,ions (OR12, i 4 and 15) are
m()rpho}◎gica翌y disも無。もhavi㎎玉arge sp汰eleもs a黙(蓬sterile}erwwa a鍛d c()nsidere(墨貸oat㎏
perennial wtld rices (VaughaR, 199e). OR the other hand, t・he aeeessions £rom Scuthem
Papua New G血ea have short genetic distance to other Asian acces$ions. This$uppQrts
earlier reports that two types of O. rufipogon are present on New Guineax} island, the
Asia鍛a捻dもhe Oeea癒a凱ypes(M◎r韮sh㎞a,1969)。
Both aRRuaYbieRnial O. meridionagis and its perermia} relatives have mot been
reported from Papua New Guinea. ln this study O. rufigeogo7?, frem the Sepik river of
Papua New Guinea have RFLP fragments which were found only in O. mewidionatis (Fig.
4). Addltioftal survey ef germp}asm from New Guiriean is},aRd and A’ustra}ia, includmg O.
?「ezLfi夢ogo?z from Australia Rot inc}uded in this就udy, theref()re, is necessaryも。 clar晦the
evolution and diversity of A-genome germplasm in this region.
Presently, new information on the A-genome species has been shown. However, to
f銭11y uR(lerstaRd もhe divers呈もy 重血 this spedes e◎類P紬x, i繋te暮どat呈○識 of data fr◎Σ嚢
hybridization, ecologica}, merphological, isozyme, organelle DNA and maclear DNA studies
ls necessary.
ACKNOWLEDGMENTS We would like to thank Dr. M. Kawase, Shikoku National Agricultural E xperimenta}
Station, Dr. J. lnouye, Kyushu University, Dr. H. Morishnna, National lnstitute of Genetics,
Dr. Y. 1. Sato, Shizuoka URiversity aAd Dr. T. Ogawa, Kyushu National Agrieultura}
ExperimeRtal StatioR, who kndly prcvided us the plant materia.ls.
REFERENCESChen, W. B., 1. Nakaraura, Y. 1. Sato and H. Nakai 1993 Pistribution of delet,ion type i,n epDNA of
cultivated and “ald rice. Jpm. 」. Genet., 68: 597-603
Dally, A. M. and G. SecoRd 1990 Chlorop}ast DNA diversity in wiid and cult.ivated speeies of riee (Genus
O惣鋤sec塾i◎勲Oryza). C圭a(Xstic-mutat,i◎R and ge難eも圭。一(慧sもa瓢}黛一寸ys重s.7短。凱細μ, G醗砿80:
2e9-222
1shii, T., T. Terachi and K. Tsunevssaki 1988 Restrict,ion endonuc}ease analysis of chloropla$t DNA from
A-genome diploid species of rice. Jpm. 」. Genet., 63: 523-536
Ka卿ama, T.1997 Re隻就ioRshipS between chro1薮◎SO獄e猟ミ搬berS axid gei}emic e◎黙S山脚穏eRS in geRUS
O摺9α.ノ麗稀Sc童ence◎fも}}e貧圭ce P圭aRt”. V◎王。3. Ge難eも玉¢s, e(薫. by「r.!瞭蕊ミ.…s(), Y。熟蝿$曲翫ra,響. Kまk登ch重
aRd H. Yamaguchi, Food aRd Agriculture Policy Research Cent,er, Tokyo, pp. 3, 9一一48
Khush, G. S., E. Bacalangco and T. Ogawa 1990 A new gene for resistar’ice to bacterial blight from O.
gongistamiuata. RZce Genet. Newsgett., 7: 121-122
Maekaw3,瓢.隷d T。 Tstm◎da 1994 Possib臨y◎ぎbreed聴。掘ghly. cOO茎舶ler蹴}rice at bo◎撫g sもage by
using O. to?zgtsta??ziuata. Jp?z. 」. Breed., 44(suppl 2):218
Miezan, K. 1986 Genetic structure of Afriean traditional rice cnitivars. fn “Rice (imnetics”, International

98 K. Doi et al.
汽lce R礁searcMnst重tuも¢,簸an瓶P購P脚es, PP.91-1◎7
Miller, J. C. and S. D. Tanksley 1990 RFLff’ analysis of phylogenetic relationships and genetic variation in
縫ミege数縫s乙茎gc《)メ)ersicon. TheoγL〆隻即9. Genet.,80:437一一448
Morishima, H. 1969 Phenetic $imilarity and phylogenetic relationships araong strains of Oryxa
perennis, estimated by methods of numerical taxonomy. Evolution, 2t3: 429一一443
Nakano, M., A. Yoshirnura aiid N. lwata 1992 PhylogeReti¢ study ef cultivated rice and it;s wild relatives
by RFLP. Rice Genet. Newslett., 9: 132-134
Nei, M. 1987 MogeeutaT Evcg2ttiona?一g GenetZc$. Ce}wnbia gniver$ity Press, New Yerk (VSA)
Oka, H.1. i 988 Origin of c2Lltivated rice. Jpn. Sc.i. Soc. Press. (Tokyo);Elsevier (Amsterdam, Oxford,
Nevty York, aRd Tokyo)
Saito, A., M. ¥ano, N. Kishaneto, M. Nakagahra, A. Yoshiraura, K. Saito, S. Kuhara, Y. Ukai, M. Kawase, T.
Nagamine, S. Yoshimura, O. ldeta, R. Oh$awa, Y. Hayano, N. lvvata and’M. Sugiura 1991 Linkage map
of yestrictioR fragmeRt lengtk po}ygierphi$i¥} loc} iR rice. Jp?z. Jl Breeal., 41; 66.r>一67g
Sano, Y and R. Sano 1990 Variation in the intergenic spacer region of ribosomal Z)NA in cultivated and
wild rice specle$. Geno’Rze, 33: 2e9-218,
SecoRd, G. 1985 Evolutionary relationships in the Sativa group o£Oryga ’based onisozynie data. Genet.
Sel. Ewol., 17: 89-l14
Sharma, S. 1]〉, and S. V. S. Shastry 1965 Taxonomic studies iR genus Ocr:yxa L. ax O. rztfipogon Grlf£
senszt s厩。‘o and O.鴛勿αrαSharma et ShasL】ry.?zom.?zov.勉d;乞α%JGenet. P‘α伽Bree(メ.,25:
157一王67
Sokal, R. R. and C. D. Mich,ener 190”8 A stat.istica1 method for eva!uating systematie relati onships. Sci.
Bzeil.,び》z”θγs魔シqブ、Kdl?躍α5,38:1409一】,438
Soiig, K. M., T. C. Osbom amd P. 1”1. Williams 1988 BTaissica taxonomy based on nuclear reswiction
fragnent length polymerphisms (RFLPs). tT7heox Appg. Ge?zet., 75: 784・一794
Th三〇u圭09se,」.199◎蟹ac漁茎a認Graρ慰綬:twO Maci魏◎sh prograras for康e(滋splay a難d a認ysis◎f
multivariate data. Computers and Geosciences, 16: 1235-1240
Vaugtkan, D. A. 1989 The geRus OTyee L. CurreRt status of taxemomy. IRRI Res. Pmper Ser. 138: i-2 i
Vaug. han, D. A. 1990 Wild relative of rice in Papua New Guinea. Report of coilaborative collecting in
Papua ,New Guii}ea betweeR the Depart,’iuent of Priraary lfidustry aRd IRRI. 9 August 一 2 September
l990. Miraeographed.
Vaughan, D. A. 1994 The wild relative of rice: A genetic resources handbook. lnternational Rice
Reseaych IRstitute, Manda (Philippines)
Wal鷹, Z. Y. and S. D. Tanksley 1989 Restriction fragm¢nt le㎎th polymQrphis翻n O㈱αsα伽αL.
Genome 32: K i 3-ll18
Wang, Z. Y., G. Seeond and S. D. Ta}tksley 1992 Polymorphism aiRd phylogeRetic relationships araong
species in the genus Orzlga as determmed by amalysis of nuelear RFLPs. Theoz Ap pl. Genet., 83:
565-581