rib fabrication in ostreoidea and plicatuloidea (bivalvia, pteriomorphia)
TRANSCRIPT

Zoomorphology (2003) 122:145–159DOI 10.1007/s00435-003-0080-5
O R I G I N A L A R T I C L E
Antonio G. Checa · Antonio P. Jim�nez-Jim�nez
Rib fabrication in Ostreoidea and Plicatuloidea(Bivalvia, Pteriomorphia) and its evolutionary significance
Received: 24 January 2003 / Accepted: 8 May 2003 / Published online: 24 June 2003� Springer-Verlag 2003
Abstract Ribs of Ostreoidea and Plicatuloidea are de-fined as antimarginal, that is, perpendicular to the marginthroughout growth. Morphogenetically, these ribs areunique, since, unlike radial ribs, they are secreted by ahomogeneous mantle margin. Based also on the recon-structed shell secretion cycle in Bivalvia, we propose thatribs of Ostreoidea and Plicatuloidea are formed by amantle margin which, upon extension from the shellmargin, stretches and folds by taking the preformed ribsas templates. In extending perpendicular to the margin (asin all Bivalvia growing isometrically), such a mantleextends the rib pattern antimarginally. Ribs of this kindare purely mechanical structures, as their arrangementdepends on the mechanical properties of the mantle andon the environmental conditions. This explains the highirregularity of such ribbing patterns. The presence ofantimarginal ribs in both the Ostreoidea and Plicatuloideasheds light on their origin. The first known oyster,Actinostreon cristadifformis, probably derived from anantimarginally ribbed Prospondylidae gen. indet. in theLate Permian or Early Triassic. Antimarginally ribbedTriassic species formerly included in Placunopsis origi-nated both the Dimyidae Atreta in the Late Triassic andEnantiostreon in the Mid Triassic, which was transitionalto Plicatulidae. Therefore, Dimyidae and Plicatulidae areclosely connected and grouped under Plicatuloidea, towhich Ostreoidea is phylogenetically unrelated.
Keywords Bivalves · Oysters · Morphology · Shell ·Evolution
Introduction
Ribs are by far the most common sculptural elements ofthe shell of Bivalvia. They can be classified into threemain morphological categories: radial, commarginal, andoblique (Fig. 1A–C). Radial ribs can be defined ashelicospirals that diverge from the umbo and represent thegrowth trajectories of particular portions of the mantlespecialized for rib secretion, which were identified anddefined by Stone (1998) as corpora spinosa (see below).Commarginal ribs run parallel to the margin and aresecreted by periodic extension of the mantle all along itslength above the shell margin. Commarginal is preferredover the less accurate term concentric (see Cox 1969).Both the radial and commarginal patterns characterizelarge groups of bivalves, for example, radial ribs are theonly ones present in Pectinidae and Arcoidea, whereasCrassatellidae secrete exclusively commarginal ribs. Both
A. G. Checa ()) · A. P. Jim�nez-Jim�nezDepartamento de Estratigraf�a y Paleontolog�a,Facultad de Ciencias,Universidad de Granada,Avenida Fuentenueva s/n, 18071 Granada, Spaine-mail: [email protected].: +34-958-243201Fax: +34-958-248528
Fig. 1A–D The four main morphogenetic varieties of bivalve ribs.A Radial (Pecten, Pectinidae). B Commarginal (Bassina, Veneri-dae). C Oblique (Divaricella, Lucinidae). D Antimarginal (Cu-bitostrea, Ostreidae)

kinds of ribs may coexist in some other groups and,occasionally, on the same shell, thus producing cancellateornamentation. A third, less common type, is the obliqueone. Oblique ribs can be defined as those havingdirections that are intermediate between radial and
commarginal. Several varieties are subsumed under thisgeneral term including the single oblique, divaricate,rasp-like, and straight ornaments (Seilacher 1972; Checa2002), all of which are morphogenetically related, sincethey arise essentially by contact-guidance mechanisms
Fig. 2A–F Examples of antimarginal ribs in Ostreoidea (A, B, D–F) and Plicatuloidea (C). A Ostrea lamellosa Brocchi, 1814, leftvalve, EPUGR.JJ-Ju-10, Pliocene, El Alqui�n, Almer�a (Spain). BHyotissa hyotis (Linnaeus, 1758), right valve, EPUGR.JJ-Ju-12,Pliocene, El Alqui�n, Almer�a (Spain). C Plicatula gibbosaLamarck, 1801, left valve, EPUGR.BV.510, Recent, GasparillaIsland (M�xico). D Oscillopha dichotoma (Bayle, 1849), left valve,
Upper Coniacian, Wadi Sudr, Sinai (Egypt), specimen in Malchus(1990), plate 1, Fig. 7. E Ambigostrea villei (Coquand, 1862), rightvalve, Uppermost Campanian, between Qena and Quseir (Egypt),specimen in Malchus (1990), plate 23, Fig. 3. F Agerostreaungulata (Schlotheim, 1813), left valve, Maastrichtian, AmmoniteHills (Egypt), specimen in Malchus (1990), plate 15, Fig. 8
146

(Checa 2002). Oblique ribs are not exclusive to anysupraspecific taxon with the exception the Divaricellinae(Lucinoidea).
Authors describing ribs of Ostreoidea (mainly Ostrei-dae, since the Gryphaeidae are usually smooth) andPlicatuloidea have invariably considered them to be radial(see, for example, Slack-Smith 1998), but a carefulinspection reveals that ribs in both groups diverge fromthe shell center to remain approximately perpendicular tothe shell margin throughout growth (Figs. 1D, 2). Ribs ofthis kind can be termed antimarginal and were initiallyrecognized and defined in Camptonectes (Pectinidae) byWaller (1986). Antimarginal ribs only follow radialdirections at the ventral margin, whereas toward the sidesthey curve laterally to form broader angles with the radialdirections. Although antimarginal ribs fit into the obliquecondition, they should be kept separate on a morphoge-netic basis.
Antimarginal ribs of Ostreoidea and Plicatuloidea areunique among present-day bivalves. Their pattern oflateral divergence is not easy to understand and, sincetheir singularity has gone unnoticed to earlier authors,there are no morphogenetic studies dealing with them,therefore constituting a morphological enigma. Thepresent study is dedicated to unraveling their morpho-genesis and exploring their systematic and evolutionarysignificance.
Materials and methods
Observations have been made on a large number of Recent andfossil species both from the literature and from collections. Inparticular, we have had access to material housed in the followinginstitutions: Departamento de Estratigraf�a y Paleontolog�a, Uni-versidad de Granada (labeled EPUGR), Museo Nacional deCiencias Naturales, Madrid (MNCN), Geologisch-pal�ontologi-sches Institut, Philipps Universit�t Marburg (GPIUM), and Staat-liches Museum f�r Naturkunde Stuttgart (SMNS).
We observed specimens with soft parts of the oysters Ostreaedulis Linnaeus, 1758, provided at Centro de Cultivos MarinosPEMARES, El Rompido (Huelva, Spain), Crassostrea gigas(Thunberg, 1793), sampled in the intertidal of Roche (C�diz,Spain) and Santa Luz�a (Algarve, Portugal), and Saccostreaechinata (Quoy and Gaimard, 1835), from Ehime (Matsuyama,Japan). Specimens of two unidentified species of Plicatula withpreserved soft parts from Bah�a (Brazil) and Perim Island (Yemen)were on loan from the Mus�um National d’Histoire Naturelle deParis (MNHN). Soft parts of bivalves of other groups alreadypresent in the EPUGR collection were also studied: Pectenmaximus (Linnaeus, 1758), Aequipecten opercularis (Linnaeus,1758), Mimachlamys varia (Linnaeus, 1758) (Pectinidae), Anomiaephippium Linnaeus, 1758 (Anomiidae), Acanthocardia aculeata(Linnaeus, 1758), Cerastoderma edule (Linnaeus, 1758) (Cardi-idae), Venus verrucosa Linnaeus, 1758, Venus nux Gmelin, 1791,Dosinia exoleta (Linnaeus, 1758), Chamelea gallina (Linnaeus,1758) (Veneridae), Solecurtus strigilatus (Linnaeus, 1758) (Sole-curtidae), and Digitaria digitaria (Linnaeus, 1758) (Astartidae).Mantles were observed both under reflected light and scanningmicroscopy. Samples were fixed in buffered cacodylate (0.1 M,pH 7.4) 2.5% glutaraldehyde and dehydrated by washing them withincreasing concentrations of ethanol (30%, 50%, 70%, 90%, and100%) at room temperature, each dehydration step taking 20 min.They were then critical-point CO2 dried (Polaron CPD7501) and
sputter-coated with gold (Polaron E5000). Electron micrographswere obtained with a Zeiss DSM 950.
Results
Comparative morphology of radial and antimarginal ribsof Ostreoidea and Plicatuloidea
Besides Camptonectes (Waller 1986), Ostreoidea, andPlicatuloidea, we have encountered antimarginal sculp-tures in other Pectinidae (such as Cyclopecten), Galeom-matidae (Galeomma, Ephippodonta), Mytilidae (Septifer,Crenella, Urumella, Lithophaga), Limidae (Divarilima),Lucinidae (Epicodakia), and in juvenile Neoleptonidae(Neolepton), Nuculidae (Nucula), and Phyllobryidae(Phyllobrya) (see illustrations in Gofas 1991; Hayamiand Kase 1993; Gofas and Salas 1996; Salas and Gofas1998; Checa 2002; N. Malchus personal communication).
An extensive review of literature and museum materialreveals that antimarginal ribs are the only kind found inRecent and fossil Ostreoida. The same applies to Plicat-uloidea, with the exception of radially ribbed speciesassigned to Plicatula, which were relatively commonbetween the Hauterivian and the Eocene (see, forexample, Squires and Saul 1997; El-Hedeny et al.2001). Whether or not they belonged to a single orseveral offshoots of antimarginally ribbed Plicatulidae is amatter of future study. Antimarginal ribs of these twogroups show features that make them distinct from theantimarginal sculpture of other Bivalvia, which isrestricted to the outer shell layers. In oysters, on thecontrary, ribs are plicae (following the terminology ofStenzel 1971) that affect the entire thickness of the shellmargin and replicate mutually on both valves. There arenevertheless two ways in which the shell corrugates. Inmost Ostreoida and in many species of Plicatula (partic-ularly thick-shelled ones) folds of the shell margin fadeout quickly toward the shell interior on the inner shellsurface (Fig. 3A). The position of the mantle marginwithin the shell in the resting position is usually markedby an internal shell deposit forming a marginal flange, theshell interior being flat or almost flat along this line. Onthe contrary, in a few Ostreoida, and in thin-shelledspecies of Plicatula, shell folds affect the whole shellthickness (Fig. 3B). From here on, we will use the termantimarginal to refer to ribs of the kind found inOstreoidea and Plicatuloidea.
Antimarginal ribs can be easily distinguished fromradial ribs (which are the closest morphological, but notmorphogenetic, pattern) by the aforementioned differ-ences in orientation. It is noteworthy, however, thatantimarginal and radial ribs approach each other inappearance when valves become vaulted, because, asvalves approach sphericity, radial ribs tend to meet themargin perpendicularly.
Antimarginal ribs have other particular features. Forexample, ribs of one valve invariably constitute replicasof the intercostal valley of the opposing valve, both valves
147

fitting tightly at the margin. Exceptions include someoysters having a reduced right valve that fits somewhatback from the margin of the left valve (for example,Ostrea and closely related taxa; Stenzel 1971). A briefsurvey of Bivalvia with truly radial ribs reveals instancesin which ribs do not fit at the margin because the profilesof ribs and corresponding intercostal valleys are not fullycomplementary. Additionally, all groups show cases inwhich ribs project beyond the opposing intercostal valleysat the margin, such that they become advanced within theplane of the valve margin. In fact, it can be said that closefitting of valve margins in radially ribbed Bivalvia is theexception rather than the rule.
Antimarginal ribs of oysters may coexist with radialcolor lines, as is frequently observed in O. edulis. Bothstructures have similar directions at the ventral marginand diverge progressively toward the umbo (Fig. 4).
Radial ribs may be of more than one order within thesame shell, for example, Pectinidae commonly developsmall riblets superimposed on high amplitude ribs, whilein Spondylidae and Chamidae small amplitude ribs may
be intercalated between more pronounced ribs (see, forexample, Fig. 5A). Antimarginal ribs all tend to besimilarly strong.
It is interesting to note that ornaments of Bivalviaprojecting extensively beyond the margin (spines, lamel-lae), even if arranged in radial rows, are also antimarginalin projecting clearly perpendicular to the margin (Fig. 5).
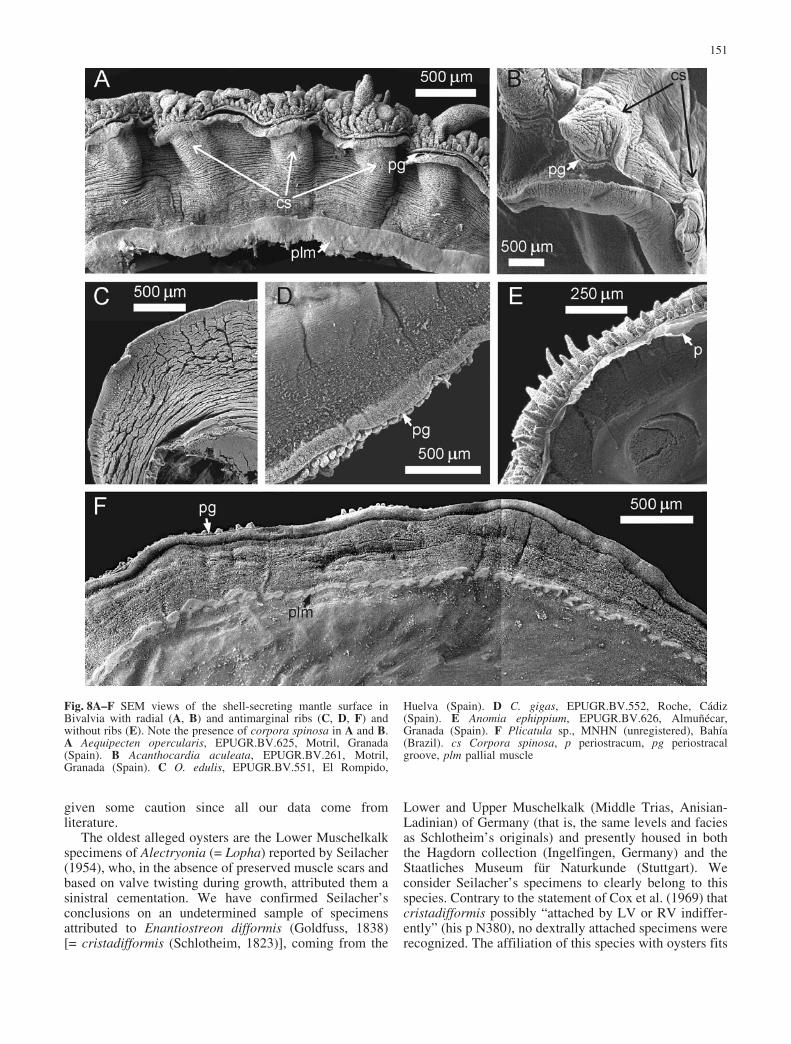
We should emphasize the high irregularity of anti-marginal ribs in profile and distribution, when comparedto radial ribs. Antimarginal ribs may display from kink (asin Lopha) to rounded profiles (for example, Ostrea).Extremely irregular profiles are achieved sometimes atthe distal ends of extensive growth lamellae (Fig. 6A). Aconsequence of antimarginality is that, if rib wavelengthis to be kept, new ribs intercalate profusely where theshell margin noticeably angles (Figs. 1D, 2D, E), thisnever being the case with radial ribs. More strikingly, ribsmay fuse during growth (Fig. 6B), when, due toxenomorphic growth, the margin becomes straight orconcave whereas the ribs, given their antimarginalcondition, converge toward the margin. In rare instances,
Fig. 4A–C In three left valves of Ostrea edulis (Recent, El Rompido, Huelva, Spain) radial color lines shift ventralward with respect toantimarginal ribs during growth. A EPUGR.BV.552. B EPUGR.BV.553. C EPUGR.BV.554
Fig. 3 A In H. hyotis (rightvalve, EPUGR.BV.513, Recent,locality unknown, South Africa)the position of the retractedmantle margin (arrow) ismarked by a deposit of foliatedcalcite underlying the vesicularlayer; this band is smooth andnot affected by corrugations ofthe outer shell. B In Lophacristagalli (Linnaeus, 1758)(right valve, EPUGR.BV.514,Recent, locality unknown,Philippines) the edge of theretracted mantle margin coin-cides with a secondary foliatedlayer (arrow); the inner shellsurface and mantle are affectedby shell plications at this line
148

a rib may end and be relayed progressively by another ribbeginning a little bit to the side (Fig. 6C). In O. edulis, itis frequent for the whole rib arrangement to becomereorganized between growth episodes (Fig. 6D).
Ribs of Ostreoidea and Plicatuloidea are not alwaysstrictly antimarginal. Elongated morphotypes (presum-ably characterized by allometric growth) of some speciesof otherwise rounded and antimarginally ribbed Ostreidaeand Plicatulidae (Fig. 7B, C) have ribs that meet themargin at an oblique angle (Fig. 7A, D, E). Allometryleading to elongation is frequent in densely packed,mutually competing oysters and allows the individuals torise rapidly above the substrate in order to maintain aneffective filtering area. Individuals achieve elongation byextruding the mantle preferentially through the ventralmargin, which is maintained in an upright position. Thiscontrasts with the rounded shape of free-growing indi-viduals. That oysters are largely able to control thedirection of mantle extension is inferred also fromtactically growing oysters, which, on falling to thebottom, drastically shifted the direction of mantle exten-sion to continue upward growth (Fig. 7E). An elongatedmorphotype is also typical of other Ostreoidea andPlicatuloidea growing in isolation, and falls within the
limits of intraspecific variability (Fig. 7A–D). Obliqueribs can also be found on the anterior margin ofspecimens that twist noticeably in a posteroventraldirection during growth (Figs. 1D, 2C, D).
Comparative mantle structure
Besides the morphological differences discussed above,there is also a fundamental distinction at a histologicallevel. Observation of the mantle of some species ofBivalvia having radial ribs reveals the presence of bulgingareas, each coinciding with a shell rib. These structurescan be classified as corpora spinosa, defined by Stone(1998) as differentiated mantle tissues surrounding dis-tended portions of the circumpallial artery and whichextend by hemocoelic pressure during the formation ofspines. Stone (1998) identified corpora spinosa in speciesof Spondylidae, Pectinidae, Ostreidae, Chamidae and,with doubt, Limidae. We recognized such structures in thePectinidae P. maximus, A. opercularis (Fig. 8A), and M.varia, and in the Cardiidae A. aculeata (Fig. 8B) and C.edule.
According to our observations, the mantle margins ofthe oysters O. edulis (Fig. 8C) and C. gigas (Fig. 8D), andof two unidentified species of Plicatula (Fig. 8F) arehomogeneous in mesoscopic view and lack any structuresimilar to corpora spinosa. We have observed creasesreminiscent of corpora spinosa in O. edulis and C. gigasdue to mantle shrinking upon fixation. This may explainthe exceptional finding of corpora spinosa in Saccostreacucullata (Born, 1778) by Stone (1998). Corpora spinosaare also lacking in the mantle surfaces of Bivalvia withcommarginal (V. verrucosa, V. nux, D. exoleta) andoblique (C. gallina, D. digitaria, S. strigilatus) ribs and ofother non-ribbed encrusters, such as A. ephippium(Fig. 8E).
Antimarginal ribs in early Ostreoidea, Plicatuloidea,and related taxa.
The use of antimarginal ribs as an additional systematiccriterion may shed light on the origin and early relation-ships between the superfamilies of Ostreina (= Ostreoidea+ Plicatuloidea + Dimyoidea; Waller 1978). The recog-nition of antimarginal ribs in early forms is therefore acrucial issue. An extensive review of museum collectionsand literature has been conducted with this goal. As far asthe Dimyidae are concerned illustrations of the firstmember of the family, the Carnian Atreta richthofeni(Bittner, 1895), only show internal surfaces of the valves.Later fossil species of Atreta and Dimya from the Rhetianto the Cenozoic are antimarginally ribbed [for example,the Rhaetian Atreta intusstriata (Emmrich, 1853), theLate Jurassic Atreta unguis (Loriol ex Merian, 1900), orthe several Cenozoic species figured by Vokes (1979)].The same is seemingly true of the Recent Dimyacorrugata Hedley, 1902. These conclusions must be
Fig. 5A–C Antimarginally projecting spines (A, C) and lamellae(B) in Bivalvia with radial ornaments. A Spondylus wrightianusCrosse, 1872, left valve, EPUGR.BV.511, Recent, near Townsville,Queensland. B Chama pulchella Reeve, 1846, right valve, EPU-GR.BV.512, Recent, Kurrimine Beach, North Queensland. CCardium hians Brocchi, 1814, left valve, EPUGR.JJ-Nb-20,Pliocene, Niebla, Huelva (Spain)
149
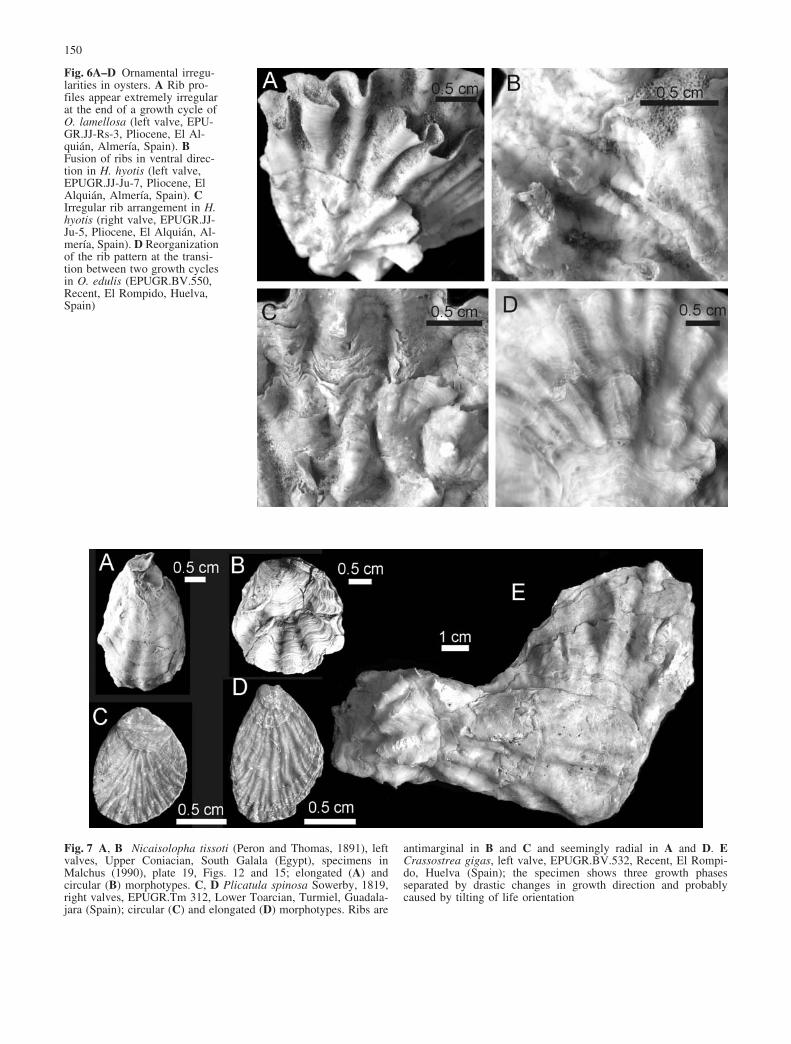
Fig. 7 A, B Nicaisolopha tissoti (Peron and Thomas, 1891), leftvalves, Upper Coniacian, South Galala (Egypt), specimens inMalchus (1990), plate 19, Figs. 12 and 15; elongated (A) andcircular (B) morphotypes. C, D Plicatula spinosa Sowerby, 1819,right valves, EPUGR.Tm 312, Lower Toarcian, Turmiel, Guadala-jara (Spain); circular (C) and elongated (D) morphotypes. Ribs are
antimarginal in B and C and seemingly radial in A and D. ECrassostrea gigas, left valve, EPUGR.BV.532, Recent, El Rompi-do, Huelva (Spain); the specimen shows three growth phasesseparated by drastic changes in growth direction and probablycaused by tilting of life orientation
Fig. 6A–D Ornamental irregu-larities in oysters. A Rib pro-files appear extremely irregularat the end of a growth cycle ofO. lamellosa (left valve, EPU-GR.JJ-Rs-3, Pliocene, El Al-qui�n, Almer�a, Spain). BFusion of ribs in ventral direc-tion in H. hyotis (left valve,EPUGR.JJ-Ju-7, Pliocene, ElAlqui�n, Almer�a, Spain). CIrregular rib arrangement in H.hyotis (right valve, EPUGR.JJ-Ju-5, Pliocene, El Alqui�n, Al-mer�a, Spain). D Reorganizationof the rib pattern at the transi-tion between two growth cyclesin O. edulis (EPUGR.BV.550,Recent, El Rompido, Huelva,Spain)
150
given some caution since all our data come fromliterature.
The oldest alleged oysters are the Lower Muschelkalkspecimens of Alectryonia (= Lopha) reported by Seilacher(1954), who, in the absence of preserved muscle scars andbased on valve twisting during growth, attributed them asinistral cementation. We have confirmed Seilacher’sconclusions on an undetermined sample of specimensattributed to Enantiostreon difformis (Goldfuss, 1838)[= cristadifformis (Schlotheim, 1823)], coming from the
Lower and Upper Muschelkalk (Middle Trias, Anisian-Ladinian) of Germany (that is, the same levels and faciesas Schlotheim’s originals) and presently housed in boththe Hagdorn collection (Ingelfingen, Germany) and theStaatliches Museum f�r Naturkunde (Stuttgart). Weconsider Seilacher’s specimens to clearly belong to thisspecies. Contrary to the statement of Cox et al. (1969) thatcristadifformis possibly “attached by LV or RV indiffer-ently” (his p N380), no dextrally attached specimens wererecognized. The affiliation of this species with oysters fits
Fig. 8A–F SEM views of the shell-secreting mantle surface inBivalvia with radial (A, B) and antimarginal ribs (C, D, F) andwithout ribs (E). Note the presence of corpora spinosa in A and B.A Aequipecten opercularis, EPUGR.BV.625, Motril, Granada(Spain). B Acanthocardia aculeata, EPUGR.BV.261, Motril,Granada (Spain). C O. edulis, EPUGR.BV.551, El Rompido,
Huelva (Spain). D C. gigas, EPUGR.BV.552, Roche, C�diz(Spain). E Anomia ephippium, EPUGR.BV.626, Almu��car,Granada (Spain). F Plicatula sp., MNHN (unregistered), Bah�a(Brazil). cs Corpora spinosa, p periostracum, pg periostracalgroove, plm pallial muscle
151

in with the presence of an outer foliated layer and,possibly, an internal aragonitic layer (De Renzi andM�rquez-Aliaga 1980). The first reported species of“Ostrea” come from the Carnian (Upper Trias): montis-caprilis Klipstein, 1845, calceoformis Broili, 1904, andmedicostata W�hrmann, 1889. These species were laterclassified as ?Palaeolopha by Malchus (1990), althoughHautmann (2001) considered this genus as a juniorsynonym of Actinostreon. On the basis of its morphologyand hinge structure, cristadifformis should also be trans-ferred to Actinostreon.
Earlier in time, there are two Late Permian cases ofantimarginally ribbed, though dextrally attached, bi-valves. Specimens of the Late Permian Prospondylusacinetus Newell and Boyd, 1970 display typical anti-marginal ribs, particularly those in Newell and Boyd(1970), Figs. 22 (holotype) and 23A, G. Direct examina-tion of the specimen in Fig. 23C (AMNH 28933) confirmsthis statement. From this species, it is only known that ithas an outer calcitic and an inner aragonitic layer (Newelland Boyd 1970).
The second instance is constituted by a set ofantimarginally ribbed forms, which were present sincethe Late Permian. Nakazawa and Newell (1968) describedand illustrated two new Upper Permian species of Japan,which they referred to as Lopha? (later considered as?Enantiostreon by Cox et al. 1969). Their specimensdefinitely display antimarginal ribs, apparently confirm-ing Nakazawa and Newell’s attribution. Nevertheless,these authors determined sinistral attachment by compar-ing their specimens with other oysters, but erroneouslypresumed the tendency to “expand more or less an-teroventrally”. Yet, as easy to observe, Ostreoidea andPlicatuloidea twist posteroventrally during growth(Figs. 1D, 2C, D, F), except when growth is hinderedby irregularities of the substrate. Therefore, the specimensreported by Nakazawa and Newell (1968), particularlythose in their plate 7, Figs. 3 and 8, could have beendextrally attached, but with fully developed antimarginalribs. Xu (1976) recorded similar forms from the UpperPermian of China. He described three species of Enan-tiostreon (two of them new), which he refers to haveattached dextrally. The absence of crura is evident inthose specimens in which the hinge is visible (his plate 39,Figs. 9, 13), which prevents inclusion in Enantiostreon.Until a genus name is available, both Japanese andChinese forms are grouped as Prospondylidae gen. indet.
Regarding the Plicatulidae, the rib pattern of the firstspecies recorded, Eoplicatula imago (Bittner, 1895) fromthe Carnian of the Southern Alps, is also clearlyantimarginal. This also applies to subsequent species ofthis early genus (the Norian E. parvadehensis Hautmann,2001 and the Rhaetian E. difficilis Healey, 1908) and tothe plicatulid genus Pseudoplacunopsis (initiating withthe Carnian P. affixa Bittner, 1895). Ladinian formsattributed to Enantiostreon flabellum (Schmidt, 1935) areno doubt related to Plicatulidae, not only in theirantimarginal ribs, but also in their hinge structure. Ourpersonal observations have revealed that the right valve
has two thin, elongated crura, which diverge from theumbo and border an elongated ligament pit. The left valvecontains corresponding sockets. There is no ligamentaryarea, and E. flabellum can hardly be distinguished fromthe type species of Enantiostreon (the Late Triassic E.hungaricum Bittner, 1901), the hinge structure of which isunknown.
The group of Triassic species recently excluded fromPlacunopsis by Todd and Palmer (2002) (plana Giebel,1856, matercula Quenstedt, 1852, and ostracina vonSchlotheim, 1822), is closely allied to Enantiostreon inattachment polarity, ribbing pattern, and hinge structure.We will further refer to them as “Placunopsis”.
The hinge of the Dimyidae also consists of crura andsockets, being therefore similar to that of the “Placunop-sis”–Enantiostreon–Plicatulidae group. Early and somesubsequent Dimyidae are moreover characterized by apattern of antimarginal, rather thin ribs, of the kind foundin “P.” plana or ostracina, with which they also share anextensively cemented right valve.
Discussion
Mantle differentiation and rib secretion
The aforementioned bulges of the mantle margin found inBivalvia with radial ribs can unequivocally be interpretedas areas specialized in the secretion of ribs and of spines.These were identified and named corpora spinosa inspiny bivalves by Stone (1998), who attributed to themthe ability of spine formation. We propose that corporaspinosa are responsible for the formation, not only ofspines, but also of ribs of the radial type, therefore beingpermanent structures.
According to our observations, the mantle margin ofBivalvia with antimarginal, commarginal, and obliqueribs is homogeneous and lacks any structure similar tocorpora spinosa. Their absence implies that there are nomantle areas strictly linked to shell ribs, so that both themantle and ribs are free to slide past each other duringgrowth. The development of corpora spinosa appearstherefore to be exclusive of radially ribbed Bivalvia.
Morphogenetic meaning of antimarginal directions
The traces followed by radial ribs are easily understand-able from the morphogenetic standpoint, since theyrepresent the trajectories left on the shell by corporaspinosa during growth. In the case of antimarginal ribs,the absence of corpora spinosa leaves but one possibilityfor their origin: they must be the traces left by foldsformed by a stretched mantle margin. A preliminary clueto the morphogenetic meaning of antimarginal ribs isprovided by the fact that projections from the shellsurfaces of Bivalvia, such as spines and lamellae, arecoincidentally always antimarginal (Fig. 5). This featureadmits a constructional explanation. According to current
152

fabricational theories (see, for example, Morita 1991) themantle can be modeled as a membrane supported byinternal hydrostatic pressure. The stress on such amembrane can be calculated as S=RP/2t, where R is thelocal curvature radius, P the fluid pressure, and t thethickness of the membrane (see, for example, Gordon1978). If such a mantle bulged locally to produce a spineor lamella, perpendicular bulging would be the only oneequalizing stresses on the mantle membrane (Fig. 9),which is a major physical imposition. Such a modelwould be feasible from the biophysical perspective, sincespine-producing structures (corpora spinosa) extend hy-draulically by increasing hemocoelic pressure (Stone1998). Therefore, a major morphogenetic constraintshould be that extension of the corpora spinosa beyondthe shell margin occurs perpendicular to the margin, i.e.,along antimarginal directions.
Independently of spines, a similar antimarginalityprinciple can be applied to the entire mantle membrane.Each time a new shell-growth increment is to be secreted,mantle extension must occur along antimarginal direc-tions. Once the secretion stage ends, the mantle mustretract inside the shell along local perpendicular direc-tions. This implies that the same stepped arrangement ofspines and lamellae (Fig. 5) should be appreciated at thescale of growth increments, although these are too smallfor that pattern to be appreciated.
Secretion of radial and antimarginal ribs
The aforementioned differences between radial andantimarginal ribs argue for profound morphogeneticdifferences. As explained above, radial ribs are secretedby a mantle with differentiated corpora spinosa, eachbeing responsible for the secretion of a rib. We canpropose the following sequence of events for the behaviorof the soft parts during the corresponding secretion cycle(Fig. 10A–D): (1) the mantle extends slightly from theshell margin along directions perpendicular to the shellmargin and the new growth increment is calcified(Fig. 10B); this might represent a low percentage of thetotal growth time, (2) the mantle retracts in directionsopposite to those followed upon extension (Fig. 10C), (3)a presumably prolonged stage of growth of the bivalvesoft parts takes place, during which all mantle sectorsmove in radial directions (Fig. 10C), given the logarith-mic spiral growth of the bivalve shell (see, for example,Raup 1966), and (4) when shell secretion is to resume,sectors responsible for rib secretion are again projectedperpendicular to the margin, but their net movement isradial compared to the previous secretion phase (Fig. 10D;compare to Fig. 10B). Therefore, growth increments ofradial ribs on the sides of the shell (note that radial andantimarginal directions coincide toward the shell’s ventralmargin) should have a stepped appearance, with eachincrement being arranged perpendicularly to the shellmargin, but with successive increments aligned radially.
On the contrary, the mantle of Bivalvia secretingantimarginal ribs is homogeneous, so that the relationshipof ribs and the underlying mantle sectors may easilychange with growth. The secretion cycle could be brokendown into the following sequence of events (Fig. 10E–H):(1) the mantle extends beyond the shell margin inantimarginal directions and secretes a new growthincrement (Fig. 10F), (2) the mantle then retracts inantimarginal directions (Fig. 10G), (3) during growth ofthe soft parts, mantle sectors move in radial directions(Fig. 10G), and (4) when shell secretion is to reinitiate,sectors extend perpendicular to the margin, but the sectorwhich formerly corresponded to a given rib has nowdisplaced slightly toward the ventral margin with respectto the rib (Fig. 10H) (net displacement decreases towardthe ventral margin, where radial and antimarginal direc-tions coincide; Fig. 10G). During (1) and (4) the alreadysecreted ribs exert a tunnel effect on the extruding mantle,so that, upon extrusion, ribs serve as rails for the mantlemargin, which adapts to the folded shell margin andpropagates the rib pattern somewhat with the secretion ofeach growth increment.
Our model is supported by the reported progressivedivergence from the ventral margin to the umbo betweenradial color lines (secreted by particular portions of themantle) and antimarginal ribs in O. edulis (Fig. 4).
Fig. 9 The stress at a given point of the surface of a membraneincreases with hydrostatic pressure and the curvature radius anddecreases with membrane thickness (t). When a perpendicularfinger-like bulging is formed, the radii of curvature on both sides(R1, R2) and therefore the corresponding stresses are identical. Thisis not the case when the finger bulges obliquely, since curvatureradii cease to be of the same magnitude
153

Folded-mantle margin model
According to our model, antimarginal ribs are formed bya folded mantle margin, which extends perpendicular tothe shell margin. To form wrinkles, the mantle marginmust grow allometrically, so that its length must exceedthe length of its projection into the plane of thecommissure. The excess mantle length adjusts by pro-ducing folds. The process can be compared to creasing of
a curved ribbon, which produces folds with axes perpen-dicular to the long axis of the ribbon (Fig. 11). Mantlefolding can apparently occur in Ostreoidea and Plicatu-loidea in two ways. Mantle folding is temporary inspecies in which corrugations of the internal shell surfacefade out to the position of the retracted mantle margin(Fig. 3A), being restricted to the moments when the shellmargin is reached to secrete a new growth increment. Asthe mantle advances toward the shell margin, it stretches
Fig. 10A–H Secretion cycles in bivalves with radial (A–D) andantimarginal (E–H) ribs, illustrated in the anteroventral sector ofthe right valve (inner surface) of a hypothetical Cardiidae andOstreidae, respectively. A, E Resting position of the mantle (gray).B, F Rib-secreting mantle points extend along local antimarginaldirections. C, G Once the new shell increment is secreted, themantle retracts along antimarginal directions; during subsequentmantle growth, selected points move along radial directions, which
form decreasing angles with antimarginal directions toward theventral margin (small insets in G). D Upon repeated mantleextension, rib-secreting areas (corpora spinosa) extend alongantimarginal directions, in such a way that the final arrangementif ribs is radial (short arrows). H In oysters, the mantle extendsalong antimarginal directions, but the mantle margin areas formerlycoinciding with ribs (in F) are now located more dorsally withrespect to actual ribs
154

out parallel to the margin and folds. When ribs affect theentire shell thickness the mantle is permanently folded(Fig. 3B).
For ribs to be antimarginal, a sine qua non condition isthat the mantle must also be extruded perpendicular to theshell margin throughout. In allometrically growing Os-treoidea and Plicatuloidea, elongation proceeds by addinga dorsoventral component to the local directions of mantleextension, which therefore cease to be antimarginal(Fig. 7A, D, E). Posteroventral twisting of the shellduring growth may lead to oblique ribs on the anteriormargin (Figs. 1D, 2C, D) for similar reasons. What isimportant is that ribs of allometrically growing formscontinue to match the directions of mantle extension, andtherefore, they do not constitute contradictions to ourmodel. It bears emphasizing that obliquity does not occurin elongated oysters growing isometrically, in which themantle extends antimarginally throughout growth(Fig. 2F).
According to our model, antimarginal ribs are purelymechanical structures. Therefore, their morphologicalproperties must obey the same laws that determine themorphology of folds in nature. In particular, when a layer(of viscosity m1, embedded in a less viscous medium ofviscosity m2) folds by lateral contraction, the wavelengthof the folds is 2pt(m1/6m2)1/3, where t is the thickness ofthe layer (Biot et al. 1961). Accordingly, oyster ribwavelength must be proportional to mantle thickness.This fits in with the fact that the mantle of oysters (andbivalves in general) is a viscoelastic material the marginof which thickens from the umbo to the ventral margin, incoincidence with rib wavelength (Fig. 2). A possible testof the model (not made here for lack of material) wouldbe to check whether Ostreoidea with denser and smallerribs have thinner or less viscous mantles.
The interpretation of antimarginal ribs as generated bya folded mantle also explains the aforementioned irreg-ularity of ribs. This is particularly true with regards to ribprofiles (Figs. 2A, 6A) and intercalation (Figs. 1D,2A, B, D, E), which may be due to the need for new
ribs to appear according to the greater mantle length andto the law of preset wavelength (see above).
Implications for the origin of Ostreoidea and Plicatuloidea
Waller (1978) placed Dimyoidea, Plicatuloidea, andOstreoidea in his suborder Ostreina (= Ostreoida; Carter1990). Alternatively, Yonge (1978, 1980) believed thatthe Dimyidae showed no relationships to the Ostreidae orPectinidae, but that they were likely cemented ancestorsof the Plicatulidae. He placed the Dimyidae within thePlicatuloidea. Hautmann (2001) rejected Waller’s hy-pothesis on the basis that: (1) Ostreidae attach to thesubstrate by their left valve, while Plicatulidae andDimyidae are dextrally attached, (2) the inner shell layerof early oysters was probably nacreous, but crossedlamellar in Dimyidae and Plicatulidae, and (3) theabsence of secondary hinge teeth and secondary hingeligament in Ostreidae. Dimyidae also differ from Plicat-ulidae in their dimyarian condition. Consequently, Haut-mann stated the possibility that the three families ofOstreina evolved independently.
Antimarginal ribs are the only kind of rib ornament inOstreoidea and, possibly, Dimyoidea, and the primitivecondition in the Plicatuloidea. Therefore, they may beused as an additional systematic criterion, which mayshed light on the above hypotheses of relationship.
As stated above, Actinostreon cristadifformis consis-tently attached by the left valve, therefore being the firsttrue oyster. Proposed ancestors of oysters are to be foundwithin fossil Pseudomonotidae (Termier and Termier1949; Newell 1960; Newell and Boyd 1970), Arcoida(Scarlato and Starobogatov 1984; Starobogatov 1992), orPterioida, possibly the Pterineidae (Carter 1990; McRo-berts and Carter 1994). This last hypothesis is based onthe observation that early oysters had an inner nacreouslayer and not crossed lamellar, as in the Pseudomonoti-dae, but it and the hypothesis implying a fossil Arcoida asancestor are not further supported by fossil data. This isnot the case with the Pseudomonotidae-ancestry hypoth-esis, particularly taking into account the existence of aseries of bivalves similar to oysters at the time ofappearance of this group. From among the dextrallyattached oyster-like, Permian to Mid Triassic Bivalvia,two Prospondylidae had already developed antimarginalribs in the Late Permian: P. acinetus, and the specimensreported by Nakazawa and Newell (1968) and Xu (1976)and here attributed to Prospondylidae gen. indet. (seeabove). This unnamed genus differs from P. acinetus inlacking a byssal notch in the juvenile. On the other hand,both forms can reasonably be related on the basis of theirage and ornamental style. In conclusion, data on ribsallow us to propose the evolutionary sequence P.acinetus–Prospondylidae gen.indet.–A. cristadifformis(Fig. 12). The latter species (and oysters, in general)emerged by reversion from dextral to sinistral attachment,without noticeable changes being required in the structureof the ligamental area.
Fig. 11 Folded ribbon model. During extension, the shell-secretingmantle stretches progressively and the excess length accommodatesby producing folds with axes perpendicular to the long axis of theribbon
155

Affiliation of Enantiostreon with early Plicatulidae(Eoplicatula, Pseudoplacunopsis) seems unquestionableon the basis of the ribbing pattern and hinge structure. Theformer genus can also be connected with the coarselyribbed “P.” matercula (Anisian). Enantiostreon appearedin the Ladinian, being thus intermediate in age between“P.” matercula and early Plicatulidae (initiating in theCarnian). The proposed evolutionary sequence is: “P.”matercula–Enantiostreon–Plicatulidae (Fig. 13), alongwhich few changes of the hinge occurred (deepening ofthe resilifer and coarsening of crura). Hautmann (2001)
considered the ancestor of the first Plicatulidae (Eopli-catula) to be a genus similar to the Prospondylidae Persia(“Persia-like genus”). He interpreted the faint ridges andgrooves of the broad ligamental area as tracks of weakcrura and sockets, which is not necessarily the case.Additionally, Persia has radial and not antimarginal ribs(see Repin 1996). Derivation of Plicatulidae from aPersia-like genus would involve: (1) change in ornamen-tation style, (2) development of crura and sockets, and (3)loss of ears. Persia first appears in the Upper Norian(Repin 1996; Hautmann 2001), thus postdating Eoplicat-ula.
The Dimyidae emerged in the Carnian, most probablyfrom finely ribbed species of “Placunopsis” (plana–ostracina group), which initiated in the Anisian and lasteduntil the Late Triassic. In our hypothesis the dimyariancondition would be the key innovation (Fig. 13).
The common ancestors of the Enantiostreon–Plicatul-idae clade and the Dimyidae are therefore to be foundwithin the group of Triassic “Placunopsis” (Fig. 13),which deserves future revision. Our hypothesis fits in withthe extreme anatomical similarity between Plicatulidaeand Dimyidae observed by Yonge (1975) and supportstheir inclusion into the Plicatuloidea, advocated by thesame author. Enantiostreon is here removed either fromthe Terquemiidae (= Prospondylidae), into which it isgenerally placed from Cox et al. (1969), or from theOstreidae, to which it was tentatively adscribed by Carter(1990). It must be transferred to the Plicatuloidea andplaced close to the Plicatulidae.
The ancestors of Ostreina, the Pseudomonotoidea, wereclosely related to the Aviculopectinoidea (Newell andBoyd 1970), which appeared in the Middle Silurian (datafrom Skelton and Benton 1993). Recent molecular phylo-genies (Adamkiewicz et al. 1997; Campbell 2000; Steinerand Hammer 2000; Giribet and Distel 2002; Giribet andWheeler 2002) group oysters with Pterioida rather than
Fig. 13 Proposed cladogramfor the early phylogeny of thePlicatuloidea and associatedchanges in the ligament andhinge structure. Sketches re-produce upper (left) valves
Fig. 12 Evolutionary sequence leading to the Ostreoidea from theProspondylidae and associated ligamentary changes. Sketchesreproduce upper valves. Arrows indicate posterior direction
156

with Pectinoida (in which they are traditionally includedsince Cox et al. 1969), which is probably due to the earlydivergence of the lineage leading to Ostreina. Thosestudies including the Plicatulidae (Campbell 2000; Steinerand Hammer 2000; Giribet and Distel 2002) place themtogether with the Anomioidea in a single clade, which isusually a sister group of the Pectinoidea. Our hypothesisfits in with molecular data in that: (1) we imply separateorigins for the Ostreoidea and Plicatuloidea and (2)Triassic “Placunopsis” might well be related to Jurassic(true) Placunopsis, which clearly belong to the Anomiidae(Todd and Palmer 2002). In consequence, the suborderOstreina, as conceived by Waller (1978), can no longer bemaintained.
Conclusions
Antimarginal ribs of Ostreoidea and Plicatuloidea aresecreted by a mantle margin, which, upon extension fromthe shell to secrete a new shell growth increment, isstretched and wrinkled by taking the already formed ribsas templates. The rib pattern is propagated antimarginallybecause the mantle extends perpendicular to the margin.Such ribs are therefore mechanical products. Theirarrangement (number, size, and position) is not geneti-cally fixed, but rather depends upon the mechanicalproperties of the mantle (thickness and viscosity) and onenvironmental conditions (substrate, density of individu-als). This explains the high variability and irregularity ofoyster ribbing patterns.
Contrary to other groups developing radial ribs, both inOstreoidea and in Plicatuloidea the shell-secreting mantlesurface is homogeneous and therefore relatively free ofhistological constraints (with the mentioned exception ofsome radially ribbed fossil Plicatulidae). This evidentlycontributes to phenotypic plasticity and, in connectionwith their ability as encrusters, may have helped bothgroups (and, particularly, the Ostreoidea) to adapt tohighly irregular substrates (Fig. 14). In connection with
this, the development of the undifferentiated oystermantle may have been a precondition for their past andpresent ecological success and consequent diversification,particularly in comparison to other encrusters with radialribs and differentiated mantles (for example, Spondylidae,Chamidae).
Other bivalves and mollusks have also occasionallyexploited the mantle-folding strategy. Rib formation bymantle folding processes has been referred to in theInoceramidae (Checa and Crampton 2002) and inammonites (Checa 1994). Active compression of themantle in radial directions and consequent commarginal(rather than antimarginal) folding has been invoked inboth cases. Therefore, the case of antimarginal ribs ofOstreoidea and Plicatuloidea is unique and has theadvantage of being partly testable by appealing to theanatomy of the soft parts. Such ribs constitute a fourthmechanism within the rib morphogenetic space availableto bivalves (Fig. 15).
Acknowledgements Thanks are given to Serge Gofas (Universityof M�laga, Spain), Hans Hagdorn (Hagdorn Museum, Ingelfingen,Germany), Gerd Dietl (Staatliches Museum f�r Naturkunde,Stuttgart), Istvan F�zy (Hungarian Natural History Museum,Budapest), Bushra Hussaini (American Museum of Natural History,New York), and Reinhard Schmidt-Effing (Philipps Universit�tMarburg). All of them contributed in an essential way to the presentstudy either by providing material of Recent and fossil specimensor allowing access to private or institutional collections. Liz Harper(Cambridge University) and Nikolaus Malchus (UniversitatAutnoma de Barcelona) critically reviewed the manuscript. Thelatter one kindly allowed reproduction of specimens in his 1990monograph. Michael Hautmann (Universit�t W�rzburg) providedconstructive criticism on the early phylogeny of oyster-likeBivalvia (although he may not agree with all our statements). Otto
Fig. 15 Morphogenetic classification of bivalve ribs, with indica-tion of the main fabricational strategies. Only in the case of radialribs do corpora spinosa differentiate at the mantle margin
Fig. 14A, B Specimen of C. gigas (EPUGR.BV.533, Recent, SantaLuz�a, Algarve), which grew within the inside corner of a brickhollow (with square section); the left valve replicated exactly thatshape. This is a sample of the ability of oysters to adapt to irregularsubstrates
157

Kraus (Universit�t Hamburg) and two anonymous reviewers madesignificant improvements. This work was funded by ResearchProjects PB97 0790 and BOS2001–3220 of the DGYCIT (Minis-terio de Ciencia y Tecnolog�a) and by the Research GroupRNM0178 of the Junta de Andaluc�a.
References
Adamkiewicz SL, Harasewych MG, Blake J, Saudek D, Bult CJ(1997) A molecular phylogeny of the bivalve mollusks. MolBiol Evol 14:619–628
Biot MA, Od� H, Roever, WL (1961) Experimental verification ofthe theory of folding of stratified viscoelastic media. Bull GeolSoc Am 72:1621–1630
Campbell DC (2000) Molecular evidence on the evolution of theBivalvia. In: Harper EM, Taylor JD, Crame JA (eds) Theevolutionary biology of the Bivalvia. Geological SocietySpecial Publication 177, London, pp 31–46
Carter JG (1990) Evolutionary significance of shell microstructurein the Palaeotaxodonta, Pteriomorphia and Isofilibranchia(Bivalvia: Mollusca). In: Carter JG (ed) Skeletal biomineral-isation: patterns, processes and evolutionary trends, vol 1. VanNostrand Reinhold, New York, pp 135–296
Checa A (1994) A model for the morphogenesis of ribs inammonites inferred from associated microsculptures. Palaeon-tology 37:863–888
Checa A (2002) Fabricational morphology of oblique ribs inbivalves. J Morphol 254:195–209
Checa A, Crampton JS (2002) Mechanics of sculpture formation inMagadiceramus? rangatira rangatira (Inoceramidae, Bivalvia)from the Upper Cretaceous of New Zealand. Lethaia 35:279–290
Cox LR (1969) General features of Bivalvia. In: Moore RC (ed)Treatise on invertebrate paleontology, part N, Mollusca 6,Bivalvia, vol 1. Geological Society of America and Universityof Kansas Press, Boulder, Colorado and Lawrence, Kansas,pp N2–N224
Cox LR, Newell ND, Branson CC, Casey R, Chavan A, CooganAH, Dechaseaux C, Fleming CA, Haas F, Hertlein LG, KeenAM, LaRocque A, McAlester AL, Perkins BF, Puri HS, SmithLA, Soot-Ryen T, Stenzel HB, Turner RD, Weir J (1969)Systematic descriptions. In: Moore RC (ed) Treatise oninvertebrate paleontology, part N, Mollusca 6, Bivalvia, vol 1.Geological Society of America and University of Kansas Press,Boulder, Colorado and Lawrence, Kansas, pp N225–N489
De Renzi M, M�rquez-Aliaga A (1980) Primary and diageneticfeatures in the microstructure of some Triassic bivalves. RevInst Inv Geol 34:101–116
El-Hedeny MM, Abdel Aal AA, Maree M, Seeling J (2001)Plicatulid bivalves from the Coniacian-Santonian MatullaFormation, Wadi Sudr, western Sinai, Egypt. Cretaceous Res22:295–308
Giribet G, Distel D (2002) Bivalve phylogeny and molecular data.In: Lydeard C, Lindberg D (eds) Molecular systematics andphylogeography of mollusks. Smithsonian Institution Press,Washington (in press)
Giribet G, Wheeler W (2002) On bivalve phylogeny: a high-levelanalysis of the Bivalvia (Mollusca) based on combinedmorphology and DNA sequence data. Invertebr Biol121:271–324
Gofas S (1991) The family Galeommatidae (Bivalvia: Leptonacea)in the Eastern Atlantic. Veliger 34:344–353
Gofas S, Salas C (1996) Small Nuculidae (Bivalvia) with functionalprimary hinge in the adults. J Conchol 35:427–435
Gordon JE (1978) Structures, or why things don’t fall down.Penguin Books, Reading, pp 1–395
Hautmann M (2001) Taxonomy and phylogeny of cementingTriassic bivalves (families Prospondylidae, Plicatulidae,Dimyidae and Ostreidae). Palaeontology 44:339–373
Hayami I, Kase T (1993) Submarine cave Bivalvia from theRyukyu islands: systematics and evolutionary significance.Tokyo Univ Mus Bull 35:1–133
Malchus N (1990) Revision del Kreide-Austern (Bivalvia: Pteri-omorphia) gyptens (Biostratigraphie, Systematik). Berl Geo-wiss Abh, Reihe A 125:1–231
McRoberts CA, Carter JG (1994) Nacre in an early gryphaeidbivalve (Mollusca). J Paleontol 68:1405–1408
Morita R (1991) Mechanical constraints on aperture form ingastropods. J Morphol 207:93–102
Nakazawa K, Newell ND (1968) Permian bivalves of Japan. MemFac Sci Kyoto Univ Ser Geol Miner 35:1–108
Newell ND (1960) The origin of the oysters. Proc 21st Int GeolCongr 2:81–86
Newell ND, Boyd DW (1970) Oyster-like Permian Bivalvia. BullAm Mus Nat Hist 143:217–282
Raup DM (1966) Geometric analysis of shell coiling: coiling inammonoids. J Paleontol 41:43–65
Repin YS (1996) New Late Triassic bivalves from Iran and ataxonomy of the superfamily Spondylacea. Paleontol J 30:363–369
Salas C, Gofas S (1998) Description of four new species ofNeolepton Monterosato, 1875 (Mollusca: Bivalvia: Neolep-tonidae), with comments on the genus and on its affinity withthe Veneracea. Ophelia 48:35–70
Scarlato OA, Starobogatov YI (1984) The systematics of thesuborder Mytileina (Bivalvia). Malacol Rev 17:115–116
Seilacher A (1954) �kologie der triassischen Muschel Lima lineata(Schloth.) und ihrer Ep�ken. Neues Jahrb Geol PalaeontolMonatsh 1954:163–183
Seilacher A (1972) Divaricate patterns in pelecypod shells. Lethaia5:325–343
Skelton PW, Benton MJ (1993) Mollusca: Rostroconchia,Scaphopoda and Bivalvia. In: Benton MJ (ed) The fossil record2. Chapman and Hall, London, pp 237–263
Slack-Smith SM (1998) Order Ostreoida. In: Beesley PL, Ross GJB,Wells A (eds) Mollusca: the Southern synthesis, fauna ofAustralia, vol 5, part A. CSIRO Publishing, Melbourne, pp 268–282
Squires RL, Saul LR (1997) Review of the bivalve genus Plicatulafrom Cretaceous and lower Cenozoic strata of California andBaja California. J Paleontol 71:287–298
Starobogatov YI (1992) Morphological basis for the phylogeny andclassification of Bivalvia. Ruthenica 2:1–26
Steiner G, Hammer S (2000) Molecular phylogeny of the Bivalviainferred from the 18S rDNA sequences with particular refer-ence to the Pteriomorphia. In: Harper EM, Taylor JD, CrameJA (eds) The evolutionary biology of the Bivalvia. GeologicalSociety Special Publication 177, London, pp 11–29
Stenzel HB (1971) Oysters. In: Moore RC (ed) Treatise oninvertebrate paleontology, part N, Mollusca 6, Bivalvia, vol 3.Geological Society of America and University of Kansas Press,Boulder, Colorado and Lawrence, Kansas, pp N953–N1224
Stone HMI (1998) The functional morphology and evolution ofpronounced shell ornament in epifaunal bivalves. UnpublishedD. Phil. Thesis, Cambridge University, Cambridge, pp 1–271
Termier H, Termier G (1949) Roles des Aviculopectinidae dans lamorphog�n�se des dysodontes Mesozoiques. Bull Mus NatlHist Nat Paris 21:292–299
Todd JA, Palmer TJ (2002) The Jurassic bivalve genus Placunop-sis: new evidence on anatomy and affinities. Palaeontology45:487–510
Vokes HE (1979) Observations on the genus Dimya (Mollusca,Bivalvia) in the Cenozoic faunas of the Western Atlanticregion. Tulane Stud Geol Paleontol 15:33–53
Waller TR (1978) Morphology, morphoclines and a new classifi-cation of the Pteriomorphia (Mollusca: Bivalvia). Philos TransR Soc Lond Ser B 284:345–365
Waller TR (1986) A new genus and species of scallop (Bivalvia:Pectinidae) from off Somalia, and the definition of a new tribeDecatopectinini. Nautilus 100:39–46
158

Xu J (1976) Terquemiidae Cox, 1964 (in Chinese). In: Compilinggroup of the lamellibranch fossils of China (ed) The lamelli-branch fossils of China. Science Press, Beijing, pp 230–231
Yonge CM (1975) The status of the Plicatulidae and the Dimyidaein relation to the superfamily Pectinacea (Mollusca: Bivalvia). JZool 176:545–553
Yonge CM (1978) On the Dimyidae (Mollusca, Bivalvia) withspecial reference to Dimya corrugata Hedley and Basiliomyagoreaui Bayer. J Moll Stud 44:357–375
Yonge CM (1980) On the Dimyidae and Plicatulidae: proposedsuperfamily Plicatulacea. J Malacol Soc Aust 4:241–242
159