ribosome display of combinatorial an tibody libraries de...
TRANSCRIPT

Title: Ribosome Display of Combinatorial Antibody Libraries Derived From Mice 1
Immunised With Heat-Killed Xylella fastidiosa and the Selection of MopB-Specific 2
Single-Chain Antibodies. 3
4
Running Title: Anti-Xylella fastidiosa Antibody Libraries 5
Authors: Armaghan Azizi1, Arinder Arora2,3, Anatoliy Markiv1, David J. Lampe4, Thomas 6
Miller2 and Angray S. Kang1,5,* 7
8
1University of Westminster, School of Life Sciences, 115 New Cavendish St, London, 9
W1W 6UW, UK. 10
2University of California, Riverside, Department of Entomology, Riverside, CA 92521, 11
USA. 12
3University of New Mexico, Department of Biology, Albuquerque, NM 87131, USA. 13
4Duquesne University, Department of Biological Sciences, 600 Forbes Ave., Pittsburgh, 14
PA 15282, USA. 15
5Queen Mary University of London, Barts & The London School of Medicine and 16
Dentistry, Institute of Dentistry, London, E1 2AT, UK. 17
18
*Correspondent 19
Dr Angray S Kang 20
Queen Mary University of London, Barts & The London School of Medicine and 21
Dentistry, Institute of Dentistry, London E1 2AT, UK. 22
Tel: +44(0)207 882 7158, Fax: +44(0)207 882 7137, Email: [email protected] 23
Copyright © 2012, American Society for Microbiology. All Rights Reserved.Appl. Environ. Microbiol. doi:10.1128/AEM.07807-11 AEM Accepts, published online ahead of print on 10 February 2012
on July 10, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from

Abstract 24
Pierce's disease is a devastating lethal disease of Vitus vinifera grapevines caused by 25
the bacterium Xylella fastidiosa. There is no cure for Pierce’s disease and control is 26
achieved predominantly by suppressing transmission of the glassy winged sharpshooter 27
insect vector. We present a simple robust approach for the generation of panels of 28
recombinant single chain antibodies against the surface exposed elements of X. 29
fastidiosa that may have potential use in diagnosis and/or disease transmission blocking 30
studies. In vitro combinatorial antibody ribosome display libraries were assembled from 31
immunoglobulin transcripts rescued from the spleens of mice immunized with heat-killed 32
X. fastidiosa. The libraries were used in a single round of selection against an outer-33
membrane protein MopB, resulting in the isolation of a panel of recombinant antibodies. 34
The potential use of selected anti-MopB antibodies was demonstrated by the successful 35
application of the 4XfMopB3 antibody in an ELISA, western blot and 36
immunofluorescence assay. These immortalised in vitro recombinant single chain 37
antibody libraries generated against heat killed X. fastidiosa are a resource for the 38
Pierce’s disease research community that may be readily accessed for the isolation of 39
antibodies against a plethora of X. fastidiosa surface exposed antigenic molecules. 40
41
on July 10, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from

Introduction 42
Xylella fastidiosa includes a group of closely related pathogens each affecting a 43
specific plant target. One subspecies affects grapevines, but little else, another 44
subspecies affects almonds and grapevines and still another affects only oleanders (1). 45
The subspecies Xylella fastidiosa subsp. fastidiosa (Xanthomonadales: 46
Xanthomonadaceae) is the pathogen of the grapevine (14) an economically important 47
crop in California. This bacterium is the causative agent of Pierce’s disease (PD) of 48
grapevines and is vectored by the leaf hopper Homalodisca vitripennis (Germar) 49
(Hemiptera: Cicadellidae) (formerly H. coagulate) also known as the glassy-winged 50
sharp-shooter (GWSS) (39). The precise pathogenic mechanisms resulting in disease 51
are not clearly understood, however the ability of the pathogen to colonize in both the 52
xylem of the grapevine and in the foregut of the GWSS vector is well established (11). 53
The X. fastidiosa genome has been sequenced and comparative analysis has provided 54
valuable information of genes, metabolic pathways and potential virulence factors that 55
may be involved in pathogenicity (43). 56
Investigating the interfaces between the plant-pathogen-insect interactions may 57
reveal sites for molecular interventions that could confer resistance or reduce 58
transmission of the pathogen. It is possible to predict and explore the surface exposed 59
components that may potentially play a role in bacterial virulence and/or be involved in 60
attachment or biofilm formation in either the plant or arthropod. The function of surface 61
displayed targets may also be probed using lectins or polyclonal antibodies (29). 62
However, if monoclonal antibodies tools were available they would allow a more 63
on July 10, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from

detailed study of the assembly, distribution, function and the role in plant-vector 64
interactions. 65
The desire for the efficient rapid generation of monoclonal antibodies to 66
biologically important protein antigens over the past two decades has driven the 67
development of a range of in vitro technologies based around combinatorial 68
immunoglobulin repertoire cloning (26), high throughput screening, phage display (4, 69
27, 37) and ribosome display (22, 23, 36). For ribosome display immunoglobulin mRNA 70
transcripts can be isolated from antibody producing cells converted to cDNA and 71
assembled in vitro to create linear DNA templates encoding libraries of single chain 72
fragment variable antibodies (scFv’s), which can be transcribed in vitro into mRNA that 73
lack a termination codon. Translation of mRNA templates in the absence of a stop 74
codon results in each ribosome stalling at the last codon and retaining the correctly 75
folded antibody polypeptide and the mRNA, creating tri-partite antibody-ribosome-76
mRNA complexes (ARMs) as shown in Figure 2. The ARMs library can then be affinity 77
enriched to select the desired ARM for recovery. The essence of the approach is the 78
linking of the recognition to the replication functions, i.e., linking the phenotype to the 79
genotype (27). Recombinant protein display technologies allows the ready access to 80
genetically encoded ligands or receptors against defined targets, an approach 81
pioneered more than 25 years ago (44). 82
The aim of this study was to create an antibody resource from mice immunized 83
with X. fastidiosa that would permit the isolation of recombinant antibodies against 84
surface accessible molecules on X. fastidiosa and to validate the approach by selecting 85
scFv’s against the outer membrane protein MopB (8). 86
on July 10, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from


Materials and methods 89
Materials 90
Bacterial strains and plasmids 91
The oligonucleotide primers used in this study are listed in Table S1 in the supplemental 92
material. The pSANG10-3F vector (42) was provided by Prof. John McCafferty 93
(University of Cambridge) and used with XL1-Blue E. coli strain (Stratagene) for 94
plasmid construction and BL21(DE3) or Rosetta gamiB (DE3) strain E. coli (Novagen, 95
UK) for recombinant protein expression. Dr. David Lampe provided the pMALc2x_mopB 96
plasmid. Plasmid pAHAHis for scFv bacterial cytoplasmic expression was based on 97
modified pET32a (unpublished). The plasmid pFab1-PhoA-H (47) was provided by 98
Prof. Masataka Takekoshi (Tokai University, Japan). Anti-NANP repeat monoclonal 99
antibody 2A10 (38, 49) VH/VL sequence was provided by Prof. Anthony A. James 100
(University of California Irvine). Xylella fastidiosa Temecula1 strain was prepared at the 101
University of California Riverside in the laboratory of Prof. Thomas A. Miller. 102
Methods 103
MopB 104
The complete amino acid sequence of X. fastidiosa Temecula1 strain (NCBI 105
NP_779898.1) MopB was submitted to web based protein prediction programs, SIG-106
Pred: signal peptide prediction (http://bmbpcu36.leeds.ac.uk/prot_analysis/Signal.html ) 107
(6) and to the PRED TMBB which is a Hidden Markov Model method, capable of 108
predicting transmembrane beta-strands of outer membrane proteins of gram negative 109
on July 10, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from

bacteria (http://biophysics.biol.uoa.gr/PRED-TMBB/) (2, 3, 45) to identify the signal 110
peptide and the putative surface exposed domains respectively. The predicted encoded 111
mature (MopB 354 amino acids (aa)) and surface exposed domain (MopB 182 aa) 112
were PCR amplified using primers designed to include NdeI (5′) site encoding an in-113
frame methionine start codon and NotI (3′) site encoding in-frame triple alanine (primers 114
are listed in Table S1. The MopB encoding inserts were PCR amplified using 115
pMALc2x_mopB template. The PCR products were restricted with NdeI and NotI, and 116
inserted into pSANG 10-3F vector to produce proteins with in-frame C-terminal 117
sequence (AAASA(H)6 KLDYKDHDGAYKDHDIAYK(D)4K. The molecular weight and 118
isoelectric points were predicted using ExPASy bioinformatics resource portal 119
(http://web.expasy.org/compute_pi/) (20). The plasmids were used to transform XL1-120
Blue E. coli cells, and confirmed by DNA sequencing. 121
Bacterial expression of recombinant MopB 122
Plasmids encoding recombinant MopB were transformed into BL21 (DE3) E. coli cells 123
(Novagen) and grown in 200 mL of LB medium containing 30 µg/mL kanamycin at 37 °C 124
with shaking at 250 rpm until optical density of 0.4-0.6 at 600 nm. Protein expression 125
was induced by the addition of isopropyl β-D-1-thiogalactopyranoside (IPTG, final 126
concentration 0.3 mM), incubation resumed for 3 h before harvesting by centrifugation 127
5000 rpm, at 4 °C for 20 min (using Sorvall Super T 21 bench top centrifuge, with SL-128
250T rotor). Cell pellets were frozen and stored at -80 °C. 129
Recombinant MopB extraction 130
on July 10, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from

The cell pellet obtained from 200 mL culture was re-suspended in 5 mL denaturing lysis 131
buffer (6 M guanidine hydrochloride, 10 mM Tris–HCl, 500 mM NaCl, 10 mM imidazole, 132
pH 8.0). The cells were lysed by sonication on ice (6×30 sec pulses), in an ultrasonic 133
cell disruptor; and centrifuged at 16,000 rpm for 45 min to pellet the cellular debris. The 134
supernatant containing soluble material was retained for the subsequent purification 135
using Ni2+-NTA agarose affinity resin. One mL of nickel chelating agarose slurry (G-136
Biosciences) was applied to a 10 mL column and equilibrated with 5 mL wash buffer (8 137
M Urea, 10 mM Tris–HCl, 100 mM NaH2PO4, pH 8.0). The supernatant containing the 138
hexa-histidine tagged protein was loaded onto the resin. The flow through was collected 139
and the resin washed with 5 mL of wash buffer (8 M Urea, 10 mM Tris–HCl, 100 mM 140
NaH2PO4, pH 6.3). The bound recombinant MopB fusion proteins were eluted in 5 x 1 141
mL of elution buffer (8 M Urea, 10 mM Tris–HCl, 100 mM NaH2PO4, pH 4.5). Fractions 142
were analyzed by SDS–PAGE (27) using 12 % Tris-Glycine gels and visualized using a 143
fast microwave assisted Coomassie stain technique described elsewhere (34). Protein 144
concentrations were estimated by the Bradford dye binding assay (6). 145
Mouse immunisation 146
X. fastidiosa Temecula strain was grown in PD3 media (13) 100 mL in a 0.5 L flask at 28 147
°C at 180 rpm for 10 days until OD 0.5 at 600 nm. The bacteria were pelleted and re-148
suspending in 100 mL PBS (2x) and the final pellet wet weight determined, then re-149
suspended in PBS at 10 mg/mL w/v and 1 mL aliquot prepared. An aliquot of the X. 150
fastidiosa was incubated at 28 °C (viability, positive control) and the remainder heat 151
treated at 55 °C for 1 h and checked for viability by plating on PD3 agar and incubated 152
for 15 days at 28 °C, the remainder of the bacterial vials were frozen at -80 °C. Once it 153
on July 10, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from

had been confirmed that no viable bacteria were present in the heat-treated ampules, 154
aliquots were sent to ProSci Inc (Poway, CA, USA) for immunisation of Balb/c mice 155
(n=5). Initial immunisation (0.5 mL/mouse) was followed by boosting at weeks 4 and 8 156
and 12. Test bleeds were taken a week after the third boosting for ELISA and dot blot 157
analysis. Following testing, mice were boosted and three days later underwent a 158
splenectomy, each spleen was placed in 10 mL of TRIzol (Invitrogen, CA, USA) for use 159
in RNA isolation. 160
MopB ELISA 161
Recombinant MopB 10 µg/mL or BSA 10 μg/mL in PBS pH 7.4; 0.1 mL/well were used 162
to coat 96-well plate (NUNC) at 4 °C overnight and blocked with 0.2 mL of 2 % BSA in 163
PBS at room temperature for 1 h. Polyclonal sera from mice (n=5) previously immunized 164
with whole heat-killed X. fasitidosa were diluted to 1/20,000 in PBS containing 0.05 % 165
Tween-20 (PBS-T) added in duplicate to the wells (0.1 mL/well) and incubated at 37 ºC 166
for 2 h. The wells were washed with PBS-T and 0.1 mL/well of rabbit anti-mouse 167
antibody (SIGMA-Aldrich) diluted 1:20,000 in 1 % BSA in PBS-T was added and 168
incubated at 37 °C for 2 h. The plates were washed as before and 0.1 mL/well goat-anti-169
rabbit IgG alkaline phosphatase conjugate (SIGMA-Aldrich, diluted 1:40,000 in 1% BSA 170
in PBS-T) was added and incubated at 37 °C for 2 h. The plates were washed as before 171
with an additional final wash with PBS alone and developed with p-nitrophenyl 172
phosphate 1mg/mL in 0.2 M Tris buffer pH 8.0, 0.1 mL/well at 37 °C for 30 min. The 173
absorbance was measured at 405 nm on an ELISA plate reader (VersaMax, Molecular 174
Devices, UK). 175
on July 10, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from

Bacterial dot blot 176
Strips of nitrocellulose ~5 mm wide were spotted with 5 µL of 5 % w/v skimmed milk 177
powder in PBS, 5 µL of bacterial suspension of E. coli and 5 µL X. fastidiosa Temecula1 178
strain, allowed to air dry, blocked with 1 % BSA in PBS and incubated with immune sera 179
(bleed 3) diluted 1/20,000 in PBS-T with 1 % BSA for 1 h. Following washing with PBS-180
T incubated with rabbit anti-mouse alkaline phosphatase antibody (Sigma-Aldrich), 181
diluted 1:20,000 in 1 % BSA in PBS-T for 1 h, washed as before and developed with 182
0.02 %, 5-bromo-4-chloro-3-indolylphosphate (BCIP) and 0.03 % nitroblue tetrazolium 183
(NBT) in 10 mL 100 mM Tris, 100 mM NaCl, 5 mM MgCl2, 0.05 % Tween 20, at pH 9.5. 184
Mouse immunoglobulin library assembly 185
The mouse spleens, each in 10 mL TRIzol (Invitrogen) were shipped on dry ice and 186
stored at -20 °C prior to processing. Upon thawing at room temperature each sample 187
was macerated using an IKA T8 Ultra Turrrax homogeniser and processed following the 188
instructions for total RNA isolation (TRIzol, Invitrogen). The resulting RNA pellets were 189
dissolved in 50 µL RNase-free water and analyzed using a NanoDrop (Thermo-Fisher, 190
UK) to determine relative purity, RNA concentration and stored at -20 °C until required. 191
First strand cDNA was synthesised from ~1 µg of total RNA using ProtoScript® First 192
Strand cDNA Synthesis Kit (M-MuLV) Reverse Transcriptase (New England Biolabs) 193
with Oligo-dT29VN for light chains and MVHLink2 for the heavy chains (all primers are 194
listed in Table 1) following the instructions provided. For antibody library construction 195
the PCR primers were based on published sequences (26, 28) with minor modifications 196
(Table S1). The primers were designed to introduce in-frame NcoI and NotI restriction 197
on July 10, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from

sites to the 5’ end of the VL and to the 3’ end of the VH respectively as shown in Figure 198
1. Members of the light chain families were individually amplified by PCR using Oligo-199
dT29VN cDNA template using combinations of MVKF1-7 with MVKR. Members of the 200
heavy chain families were individually amplified by PCR using MVHLink2 cDNA 201
templates using combinations of MVHF1-10 and MVHR1.1. The amplified light chain 202
products were purified and pooled and an aliquot subject to another round of PCR 203
amplification using MVKFLink and MVKRLink to introduce part of the T7 site and Kozak 204
sequence (32) on the 5’ end and an overlap extension on the 3’ end to facilitate joining 205
to the variable heavy chain libraries. The amplified heavy chain products were 206
processed and modified in a similar manner using primers MVHFLink and MVHR1.1. 207
The modified variable light and heavy chain products were combined and amplified 208
using MVKFLink and MVHR1.1. A synthetic mouse kappa constant (MKC) domain 209
(optimised for E.coli codon usage, synthesised by Epoch Biolabs, Tx USA) was 210
amplified using MKNotCF and MKRev. The MKC was joined to the VL-link-VH 211
combinations by a PCR overlap extension reaction using MVKFLink and MKRev (Figure 212
1). Finally the PCR product encoding all the variable light chains and heavy chain 213
combinations were amplified with primers RDT7 and MKRev to produce the DNA 214
encoding the anti-X. fastidiosa immunoglobulin scFv libraries. The initial PCR 215
amplification reactions were performed at 50 °C annealing temperature with 30 cycles 216
and subsequent library assembly step used 16 cycles with Taq DNA polymerase and 20 217
pmol of each primer pair per reaction. DNA fragments were resolved by gel 218
electrophoresis on 2 % (w/v) agarose gels. DNA isolation from agarose gels was carried 219
on July 10, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from

out following Qiagen DNA Gel-purification kit instructions. In total five combinatorial 220
scFv antibody DNA libraries were constructed. 221
Antibody ribosome display 222
To select for single-chain antibodies a modified eukaryotic ribosome display (23) was 223
used as outlined in Figure 2. The PCR generated DNA library of antibody coding genes 224
derived from mouse #4 were expressed in a coupled rabbit reticulocyte lysate system 225
(Promega; TNT quick coupled transcription/translation system). An in vitro coupled 226
transcription/translation was set up in a 0.5 mL tube as follow: to 40 µL TNT T7 Quick 227
Master Mix, 2 µL of DNA library (0.1-1.0 μg), 1 µL (1 mM) methionine, 1 µL DNA 228
enhancer and 6 µL water were added and incubated at 30 °C for 90 min. Then, 6 µL of 229
DNase I, RNase free (10,000 u/mL, Roche) was added and incubated for 20 min at 30 230
°C (to degrade input DNA). To select specific antibody fragments a 0.5 mL PCR tube 231
was coated with 0.1 mL of 10 µg/mL of recombinant truncated MopB in PBS at room 232
temperature for 1 h, washed with PBS and blocked with 100 μL of molecular biology 233
grade BSA in PBS (10 mg/mL) (New England Biolabs) for 1 h. The 234
translation/transcription mixture, containing the antibody-ribosome-mRNA complexes 235
was added to MopB-coated tube and incubated on ice for 1 h. The tube was washed by 236
filling once with PBS (0.5 mL) and decanting and carefully removing the residual liquid 237
using a sterile pipette tip. The retained RNA was recovered using RNeasy (350 µL) mini 238
protocol following the manufacturer’s instructions and used in One-step™ (Qiagen) RT-239
PCR with MVKFLink and MKRD2 primers to amplify the antibody encoding DNA in a 50 240
µL reaction, using 6 µL RNA template and 5 µL of each primer (20 pM). The mixture 241
was cycled as follows: 50 ˚C for 30 min, 95 ˚C for 15 min, then 35 cycles 94 ˚C for 30 242
on July 10, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from

sec, 52 ˚C for 30 sec, 72 ˚C for 1 min, 72 ˚C for 5 min and finally kept at 8 ˚C. The RT-243
PCR product from a single round of ribosome display was purified by agarose gel 244
electrophoresis and cloned into pCR®IITOPO vector (TOPO TA Cloning® Kits, 245
Invitrogen) according to manufacturer’s instructions and used to transform XL1-Blue 246
competent cells. Plates containing 5-bromo-4-chloro-3-indolyl-beta-D-galactopyranoside 247
(X-gal) were used to select for disruption of the β-galactosidase. Four white colonies 248
were randomly selected and inoculated in 10 mL of LB media with 100 μg/mL of 249
carbenicillin; plasmid DNA was isolated from E. coli using standard procedures using a 250
QIAprep® Spin Miniprep Kit (Qiagen) following the manufacturer’s instructions, and 251
DNA sequenced at the core facility at The Wolfson Institute for Biomedical Research, 252
University College London. 253
ScFv expression 254
A scFv antibody cytoplasmic expression vector with a carboxyl terminal tag peptide 255
sequence derived from influenza hemagglutinin (HA) (Asp Val Pro Asp Tyr Ala Ser: 256
DVPDYAS) followed by a hexa-histidine tag designated pAHAHis was constructed 257
based on pET32a. The recovered single chain antibody encoding sequences were 258
inserted into pAHAHis plasmid as NcoI/NotI fragments for cytoplasmic expression in 259
Rosetta gami B (DE3) E. coli. The pAHAHis encoding the scFv transformed into Rosetta 260
gami B (DE3) E. coli cells (Novagen), inoculated into 200 mL of LB medium containing 261
100 μg/mL of carbenicillin and 34 μg/mL of chloramphenicol, grown at 37 °C with 262
shaking at 250 rpm until optical density reached 0.4-0.6 at 600 nm. Protein expression 263
induced by the addition of isopropyl β-D-1-thiogalactopyranoside (IPTG, final 264
concentration 1 mM) and cells then incubated for 20 h at 20 °C with shaking at 250 rpm. 265
on July 10, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from

Cells were harvested in 4 x 50 mL tubes by centrifugation 5000 rpm, 4 °C for 20 min 266
(using Hettich Universal 320 Centrifuge, 1617 rotor). Cell pellets were frozen and stored 267
at -80 °C before undergoing further processing. The cell pellet was re-suspended in 3 268
mL of native lysis buffer (20 mM Tris-HCl, 500 mM NaCl, 20 mM imidazole, 0.15 Triton 269
X100, pH 8.0 and 250 µL lysozyme at 1 mg/mL). The cells were lysed by sonication on 270
ice as described earlier and centrifuged at 18000 rpm for 45 min. The supernatant 271
containing the soluble fraction was retained for affinity purification. One mL of nickel 272
chelating resin suspension (G-Biosciences, Mo, USA) was packed in a micro spin 273
column and equilibrated with 600 µL native lysis buffer. The supernatants containing the 274
hexa-histidine-tagged recombinant antibodies were loaded onto the equilibrated 1 mL 275
Ni2+-NTA spin column and centrifuged (using Hettich Universal Centrifuge 320R, 1689-276
A rotor) at 1600 rpm for 5 min. The flow through was collected and the Ni2+-NTA column 277
washed two times with 600 µL of wash buffer (20 mM Tris-HCl, 500 mM NaCl, 20 mM 278
imidazole, pH 8.0) at 2900 rpm for 2 min. The bound recombinant antibodies were 279
eluted into 100 µL fractions using 500 µL of elution buffer (20 mM Tris-HCl, 500 mM 280
NaCl, 500 mM imidazole, pH 8.0) at 2900 rpm for 2 min and the purified scFv stored at 281
4 °C. 282
Functional assays 283
Recombinant scFv binding to MopB was examined by western blot, ELISA and 284
Immunofluorescence analysis. The scFv antibody binding to recombinant MopB, was 285
detected via the DVPDYAS peptide tag using a secondary recombinant antibody scFv 286
with a (Gly4Ser)3 linker based on VH and VL sequences of the mouse monoclonal 287
antibody 26/9 anti-DVPDYAS (12) fused to alkaline phosphatase fusion (PhoA) (46) 288
on July 10, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from

(anti-HA scFv-AP) in both the ELISA and western blot. ELISA with either full-length 289
mature, truncated MopB was as described earlier with a modification using 5 % w/v milk 290
in TBS containing 0.1 % NaN3 as a negative control. Purified antibodies, selected scFv 291
antibody for MopB and negative control anti-NANP repeat scFv based on 2A10 mAb 292
diluted 1 in10 using TBS/NaN3 were incubated for 6 h at 8 ˚C. After washing with 293
TBST/NaN3, anti-HA scFv-AP antibody (1:10 diluted in 1 % w/v milk/TBS NaN3) was 294
added and incubated overnight at 8 ˚C, washed as before and developed with p-295
nitrophenyl phosphate as described earlier. 296
For the western blot recombinant MopB proteins were resolved by SDS-PAGE as 297
described earlier and blotted onto polyvinylidene fluoride (PVDF) membrane, blocked in 298
5 % w/v milk in TBS at room temperature for 1 h, then incubated with the scFv 299
4XfMopB3 as the primary antibody (diluted 1:10 in 1 % w/v milk/TBS) for 6 h at 8 ˚C. 300
After washing with TBST incubated with the secondary antibody anti-HA scFv-AP 301
antibody (1:10 diluted in 1 % w/v milk/TBS, 0.1 % NaN3) at room temperature for 1 h on 302
a rocking platform followed by washes as before and a final rinse with TBS (5 min) prior 303
to the addition of substrate BCIP/NBT as described earlier. 304
Heat-killed X. fasitidosa was air dried, on glass slides, blocked with 2 % w/v milk in PBS 305
for 30 min. Purified scFv antibodies (selected scFv antibody for MopB and negative 306
control scFv 2A10 antibody) were diluted 1 in 10 with 1 % w/v milk/PBS. The slides 307
were incubated with either the XfMopB3 (anti-MopB), 2A10 (non-specific) or no scFv for 308
1 h. After washing with PBS anti-poly-histidine-FITC conjugate (AbCam, UK) (diluted 309
1:10,000 in 1 % w/v milk/PBS) was added. The slides were incubated for 1 h at ambient 310
temperature in the dark. Slides were washed with PBS and viewed under oil immersion 311
on July 10, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from

on a fluorescence microscope Axioscop 50 (Zeiss). Images were captured using a CCD 312
camera (PowerShot digital camera, Canon) and AxioVision software (Zeiss). 313
314
on July 10, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from

Results 315
MopB bioinformatic analysis 316
The X. fastidiosa Temecula1 MopB protein sequence downloaded from NCBI data 317
base was analyzed using SIG-PRED to predict the putative signal sequences, Met1-318
Ala36 and Met15-Ala36 were identified (Figure 3A). The β-barrel transmembrane 319
domain identification using PRED TMBB was determined to be between Trp43-Tyr177. 320
A two dimensional representation of the result show the exposed loops with the last 321
loop being the longest (Figure 3B). The recombinant mature and truncated MopB 322
expressed in pSANG 10-3F were predicted to have a pI of 6.36, Mwt of 42.98 kD and pI 323
of 6.79 and Mwt of 24.64 kDa respectively. 324
Expression and purification of the full length mature and truncated MopB protein 325
The gene encoding full length mature Met-Ala36-Asn389 and truncated Met-Ala36-326
Gly252 MopB from X. fastidiosa were PCR amplified and cloned into the pSANG10-3F 327
vector and verified by DNA sequencing. Proteins analyzed by SDS–PAGE, revealed 328
abundant full length and truncated MopB at 3 h at 37 °C post induction with IPTG. The 329
SDS–PAGE indicated proteins with an apparent molecular weight of about 43 kDa for 330
full length MopB and 25 kDa for truncated MopB (Figure 4A,B). The recoveries of 331
recombinant mature and truncated MopB proteins were estimated to be approximately 332
50 mg/L of culture. 333
Sera ELISA and Dot Blot 334
The ELISA with immobilised recombinant truncated MopB protein and 1/20,000 dilution 335
on July 10, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from

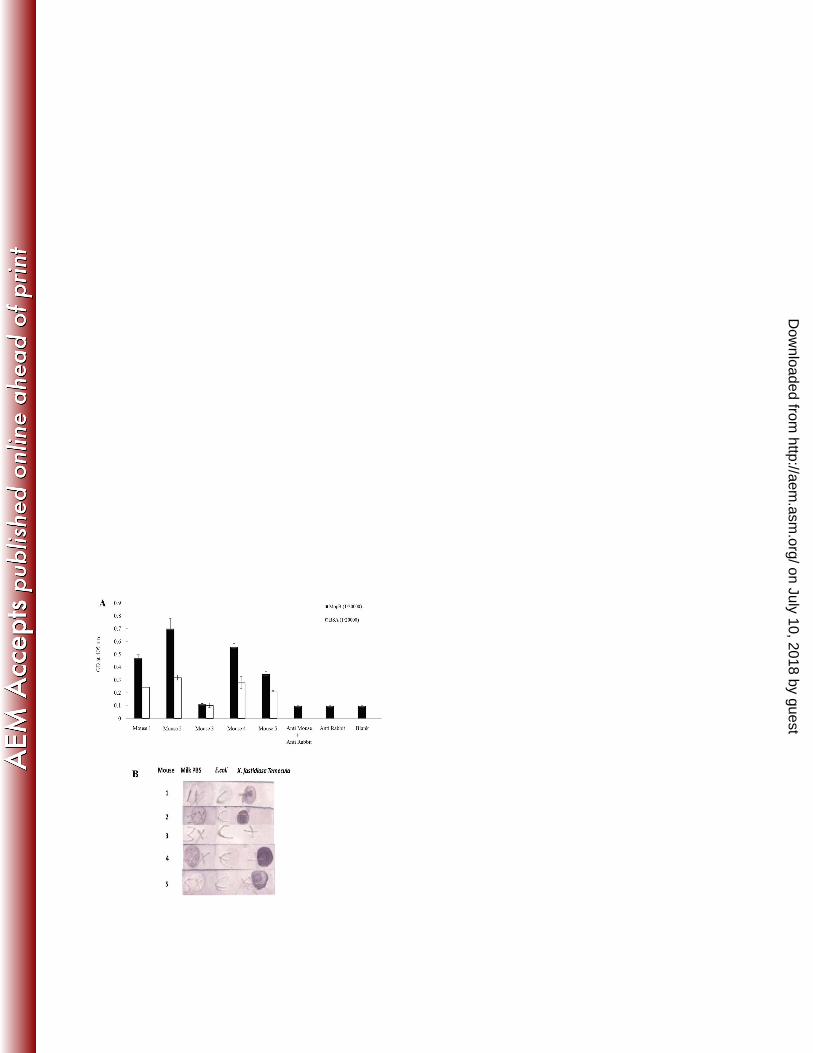
of sera from bleed 3 of the mice immunized with whole heat-killed X. fastidiosa indicated 336
that 4 out of 5 mice produced antibodies that recognise recombinant surface exposed 337
portion of MopB with little or no cross-reactivity with the control protein BSA (Figure 5A). 338
The immune sera from bleed 3 was also evaluated in a dot blot against whole intact E. 339
coli and X. fastidiosa Temecula strain (Figure 5B). Mouse sera #1, 2, 4 and 5 had clear 340
antibody binding to X. fastidiosa at a dilution of 1/20,000 whereas mouse #3 serum did 341
not. The antibody activity against milk proteins was absent or very weak in mouse sera 342
#2 and #4. 343
Mouse Immunoglobulin Library Assembly and Ribosome Display 344
The recovery and purity of total RNA isolated for each of the mice varied with a range of 345
200-50 µg as shown in Table S2 in the supplemental material, providing ample material 346
for the subsequent cDNA reactions. The immunoglobulin libraries were assembled as 347
outlined in the schematic, the PCR amplification results and steps of the library 348
construction using mouse 4# spleen derived cDNA are shown in Figure 1. Mouse Vk6 349
and VH4 and 8 were present at lower levels, all other immunoglobulin families were 350
readily amplified. The final DNA template encoding the library flanked by a T7 site and a 351
synthetic mouse kappa constant chain was used in an in vitro ribosome display with a 352
single selection step with truncated MopB as outlined in Figure 2. The affinity enriched 353
mRNA’s were amplified by RT-PCR and cloned into pCR®IITOPO vector. From 354
approximately one hundred white colonies four were randomly selected to prepare 355
plasmids for sequencing. Three with confirmed intact in-frame VL and VH combinations 356
were sub-cloned into pAINFHis for scFv expression and characterization (4XfMopB1-3). 357
Analysis of the variable region sequences (Table S3 in the supplemental material) 358
on July 10, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from

shows that different gene segments of the antibody sequences are present in the 359
selected recombinant antibody clones. Antibody 4XfMopB3 scFv detected recombinant 360
mature and truncated MopB in the western blot and ELISA (Figure 6) whereas clone 361
4XfMopB1 detected MopB in the western blot only (data not shown). Furthermore the 362
Xf4MopB3 scFv was used to image X. fastidiosa by immunofluorescence microscopy as 363
shown in Figure 7. GenBank accession numbers for 4XfMopB1 and 4XfMopB3 scFv’s 364
are JQ606804 and JQ606805 respectively. 365
366
on July 10, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from

Discussion 367
Investigations of plant-pathogen and vector-pathogen interfaces in PD would be 368
greatly facilitated with the availability of highly specific monoclonal antibodies. Although 369
it is possible to use conventional hybridoma technology (30) to obtain monoclonal 370
antibodies and then clone and reassemble the corresponding antigen binding site 371
genes, recombinant antibody display technology is a more cost-effective option for the 372
non-specialized laboratory, requiring general molecular biology skills/reagents and 373
techniques to build and access antibody libraries. Moreover, genetically encoded scFv’s 374
are amenable to further engineering to optimize the desired characteristics, such as 375
affinity, specificity (21), formation of multimers (25) or as fluorescent molecules (34, 35). 376
When using a complex immunogen (i.e., whole bacteria) a mixture of antibodies 377
against a mixture of targets (on the cell surfaces) (5), results in combinatorial complexity 378
and invariably requires de-convolution at some point (50). When initially validating a 379
technique, a simpler approach is to anchor one component and search through a 380
complex mixture of potential partners. In this study we assembled combinatorial libraries 381
of recombinant antibodies from mice immunized with whole heat-killed X. fastidiosa. To 382
de-convolute the mixture we chose a surface exposed molecule MopB as bait. 383
Although, the role of MopB on X. fastidiosa is not known, it is an abundant surface 384
protein and may potentially be a target for bacterial agglutination via engineered 385
recombinant antibodies. Earlier reports describing attempts to isolate natural MopB and 386
to make recombinant MopB protein had met with limited success (7, 8). An in silico 387
analysis of the target identified key features that assisted in the design of an appropriate 388
recombinant bait molecule for use in antibody selection. MopB has a characteristic 389
on July 10, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from

signal leader sequence Met15-Ala36 with a classical cleavage motif Ala Ser Ala, 390
followed by a transmembrane β-barrel (with surface exposed loops) Trp43-Tyr177 and a 391
periplasmic domain Arg178-Asn389 (Figure. 3). The protein is extremely toxic to the 392
heterologous host E. coli when inserted into the outer membrane, either using its natural 393
leader sequence or an alternative such as OmpA. A leaderless construct results in 394
proteins not traversing or inserting in the inner membrane but being retained in the 395
bacterial cytoplasm and aggregating due to the hydrophobic nature of the trans-396
membrane β-sheets. Full length mature (43 kDa) and truncated MopB (25 kDa) 397
recombinant proteins were both efficiently expressed without a leader sequence. 398
Extraction using denaturing conditions released the recombinant MopB into the soluble 399
fraction. SDS-PAGE analysis tracking the expression and purification revealed that the 400
recombinant proteins were enriched and the major products eluting from the affinity 401
resin corresponded to the predicted sizes for the mature and truncated MopB (Figure 4), 402
with yield of ~50 mg/L of bacterial culture. 403
Sera from the X. fastidiosa immune mice #1, 2, 4 and 5 diluted 1/20,000 had 404
activity against truncated MopB protein and X. fastidiosa in the ELISA and immuno-dot 405
blot respectively. Mouse #3 serum produced the lowest signal in both assays (Figure 406
5A, B). The very high dilution used in both the evaluations of the sera suggests a very 407
robust response to the whole bacterial antigen preparation. 408
Five antibody libraries were assembled following the steps outlined in Figure 1, 409
and mouse 4# DNA antibody library was used in the subsequent selection procedure. 410
After a single enrichment step using ribosome display, scFv specific for MopB were 411
readily isolated. This rapid enrichment may be a common feature of antibody libraries 412
on July 10, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from

constructed from intentionally immunized or naturally immune sources and is in 413
agreement with our findings when using antibody ribosome display to access human 414
scFv’s against HIV-1 gp120 constructed from an HIV-1 positive donors (48), where only 415
a single round of selection was required. 416
One scFv 4XfMopB3 recognized the mature and truncated recombinant MopB 417
proteins in both the ELISA and western blot (Figure 6) and heat killed X. fastidiosa by 418
IFA (Figure 7) suggesting that the epitope is a non-conformational and possibly one of 419
the three predicted loops exposed on the surface of the bacteria. These results provide 420
evidence that immunization with whole heat-killed X. fastidiosa clearly induces 421
antibodies against surface exposed antigenic molecules such as MopB, indicating that 422
the mild heat treatment used to kill the bacteria, preserved the surface accessible 423
protein immunogenicity. We speculate that the immune sera may also contains 424
antibodies against other putative surface exposed molecules possibly associated with 425
attachment in either the GWSS or the V. vinifera grapevine (31). 426
The in vitro anti-X. fastidiosa scFv libraries generated in this study and the strategy 427
for preparing recombinant putative membrane proteins provides approaches for rapidly 428
discovering additional scFv’s against surface components involved in aggregation (19) 429
and/or motility (15, 24, 33). The anti-MopB or other anti-X. fastidiosa scFv’s molecules 430
may also have use in diagnostic applications for pathogen surveillance and could be 431
assembled with in-built fluorophores as described recently (34, 35). 432
With the availability of genetically encoded molecules that target the surface of X. 433
fastidiosa alternative ways to investigate the biology of X. fastidiosa host vector 434
interactions may be explored to develop interventions. One such approach is 435
on July 10, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from

paratransgenesis, which in concept involves the genetic manipulation of vector’s 436
naturally occurring microorganisms such as bacteria, fungi or viruses to compromise 437
competence to transmit a particular agent. 438
Engineering symbiotic bacteria to express and secrete recombinant molecules 439
inside an arthropod was initially demonstrated using the bacteria Rhodococcus rhodnii 440
to express cecropin (16) in the kissing bug Rhodnius prolixus, the vector of Chagas’ 441
disease and subsequently to express a scFv (17). Recently scFv’s that target 442
Trypanosoma cruzi epimastigotes glycans have been developed for use in this 443
paratransgenic application (34). This strategy may also be implemented with symbionts 444
or commensal organism of the GWSS (40). Alternatively, other ‘Trojan horse’ vehicles 445
may be developed. 446
Recently, a scFv against Plasmodium falciparum sporozoite surface protein 447
isolated from a malaria immune individual (10) and shown to inhibit sporozoite invasion 448
of human hepatocytes (9) has been used to reduce P. falciparum sporozoite levels in 449
Anopheles gambiae via an engineered entomopathogenic fungus (18), basically using 450
an infection to fight an infection (41). In the malaria study the recombinant antibody 451
PfNPNA-1 targeted a conserved surface accessible repetitive target on the highly 452
abundant circumsporozoite protein and was engineered to agglutinate the parasite 453
preventing migration to and invasion of the salivary gland. Such an approach is 454
independent of disrupting host cell specific interactions and may be a general strategy 455
against targets that are present at high density on the pathogen surface. Recombinant 456
antibodies against MopB and other abundant surface exposed molecules on X. 457
fastidiosa could also be engineered in a similar way to agglutinate the bacteria and 458
on July 10, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from

evaluated in the GWSS via paratransgenic organisms such as engineered commensals 459
or symbionts or by infection with Metarrhizium spp or Beauvaria bassiana providing 460
novel alternative platforms to investigate in the control of PD. 461
Acknowledgments. 462
This work was supported by a Sub-Award of USDA APHIS Pierces Disease; Symbiotic 463
Control of Pierce’s Disease, #06-8500-0510-GR CA by Thomas A. Miller at UC 464
Riverside to ASK and was in part conducted within the EU FA COST Action FA0701 465
remit, Arthropod Symbiosis: From Fundamental Studies to Pest and Disease 466
Management. We thank Howard Boland for assistance with microscopy. 467
468
on July 10, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from

Legends 469
Figure 1. An illustration of the mouse immunoglobulin DNA library assembly process 470
showing the PCR amplification and assembly steps. Corresponding representative 471
agarose gel resolved PCR products for each of the steps generated using mouse #4 472
spleen total RNA. 473
474
Figure 2. A schematic of the steps involved in in vitro antibody ribosome display. 1: The 475
scFv antibody library DNA template was prepared flanked with a T7 sequence and 476
mouse kappa constant sequence. 2: The antibody library DNA was transcribed to 477
mRNA and translated to form antibody-ribosome-mRNA complex. 3: The mixture 478
incubated with immobilized truncated MopB. 4: The unbound components removed by 479
washing. 5: The retained complexes released. 6: The mRNA reverse transcribed and 480
amplified by PCR. 7: The PCR products cloned into TOPO vectors and recovered 481
plasmids sequenced. Full-length in-frame scFv sequences inserted into pAHAHis vector 482
for recombinant antibody production. 483
484
Figure 3. The amino acid sequence of Xylella fastidiosa temecula1 MopB and the 485
predicted model for the B-barrel outer membrane domain. (A) Underlined black putative 486
leader sequence starting at Met1, overlined black in gray highlight is also a shorter but 487
potential leader sequence starting at Met15 (previously identified by Buening et al.,(8). 488
The arrow between Ala36 and Gln37 denotes signal peptide cleavage site, grey 489
periplasmic regions, blue membrane spanning regions and red surface exposed loops. 490
The yellow highlighted region corresponds to the truncated MopB. Sequence scored a 491
on July 10, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from

value of 2.929, which is lower than the threshold value of 2.965. The difference between 492
the value and the threshold indicates the possibility of the protein being an outer 493
membrane protein. (B) The 2D figure of the predicted MopB transmembrane β-barrel 494
with the protruding surface exposed loops generated using PRED TMBB. The amino 495
acid numbers of the beginning and end of the β-strand traversing the outer membrane 496
are in green. 497
498
Figure 4. SDS-PAGE analysis of (A) Denaturing conditions mature full-length MopB 499
expression and purification. Lane 1, protein molecular weight markers; Lane 2, non 500
induced; Lane 3, induced; Lane 4, final wash; Lanes 5-6, elution 2-3. (B) Denaturing 501
conditions truncated MopB expression and purification. Lane 1: protein marker, Lane 2: 502
uninduced, Lane 3: induced, Lane 4: soluble fraction , Lane 5: flow through, Lanes 6: 503
final wash, lanes 7-10 elution 1 to 4. 504
505
Figure 5. Mouse immune sera evaluation by ELISA and immunodot blot. (A) Mouse 1-5 506
sera from bleed 3 diluted 1/20,000 in the ELISA on truncated MopB (10µg/mL) and the 507
BSA (10µg/mL) , blank, rabbit anti-mouse (1/20,000) + anti-rabbit alkaline phosphatase 508
and anti-rabbit alkaline phosphatase (1/40,000) without anti-mouse. (B) Immunodot blot 509
screen with 1/20,000 dilution of bleed 3 mouse 1-5 sera on 5% w/v milk protein control, 510
or 5µL of E.coli or X.fastidiosa spotted directly onto nitrocellulose strips and probed with 511
rabbit anti-mouse alkaline phosphatase and developed with NBT/BCIP. 512
513
on July 10, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from

Figure 6. Functional Analysis of 4XfMopB3 INF scFv by ELISA and western blot. (A) 514
The ELISA with full-length and truncated MopB and 5% milk protein as a control. 2A10 515
scFv expressed in pAHAHis control antibody (against NANP repeat) was used as an 516
irrelevant negative control scFv. The detection anti-HA- alkaline phosphatase without a 517
first antibody and the substrate alone were used as additional controls. (B) MopB 518
western blot using 4XfMopB3 INF scFv with mature full-length and truncated MopB on 519
nitrocellulose membrane and detected with anti-HA- alkaline phosphatase. 520
521
Figure 7. Immunofluorescence detection of X. fastidiosa with Xf4MopB3 scFv. Panels 522
(A,C & E) viewed in bright field and (B,D & F) and with fluorescence microscopy. (A&B) 523
X. fastidiosa with 1° 4XfMopB3 scFv probed with 2° anti-polyhistidine FITC. (C&D) X. 524
fastidiosa with 1°anti-NPNA scFv probed with 2°anti-polyhistidine FITC and (E&F) X. 525
fastidiosa probed with 1°anti-polyhistidine FITC only. The size bars shown in panels A & 526
B are 5µm. 527
528
529
on July 10, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from

References: 530
1. Almeida, R. P., and A. H. Purcell. 2003. Biological traits of Xylella fastidiosa strains from grapes 531 and almonds. Appl Environ Microbiol 69:7447-7452. 532
2. Bagos, P. G., T. D. Liakopoulos, I. C. Spyropoulos, and S. J. Hamodrakas. 2004. A Hidden Markov 533 Model method, capable of predicting and discriminating beta-barrel outer membrane proteins. 534 BMC Bioinformatics 5:29. 535
3. Bagos, P. G., T. D. Liakopoulos, I. C. Spyropoulos, and S. J. Hamodrakas. 2004. PRED-TMBB: a 536 web server for predicting the topology of beta-barrel outer membrane proteins. Nucleic Acids 537 Res 32:W400-404. 538
4. Barbas, C. F., 3rd, A. S. Kang, R. A. Lerner, and S. J. Benkovic. 1991. Assembly of combinatorial 539 antibody libraries on phage surfaces: the gene III site. Proc Natl Acad Sci U S A 88:7978-7982. 540
5. Bowley, D. R., T. M. Jones, D. R. Burton, and R. A. Lerner. 2009. Libraries against libraries for 541 combinatorial selection of replicating antigen-antibody pairs. Proc Natl Acad Sci U S A 106:1380-542 1385. 543
6. Bradford, J. R. 2001. In silico Methods for Prediction of Signal Peptides and their Cleavage Sites, 544 and Linear Epitopes. University of Leeds, Leeds. 545
7. Bruening, G., E. Civerolo, A. M. Dandekar, and G. Gupta. 2003. ROLES OF XYLELLA FASTIDIOSA 546 PROTEINS IN VIRULENCE. Pierce’s Disease Research Symposium. 547
8. Bruening, G., E. Civerolo, B. Kirkpatrick, and D. Gilchrist. 2002. VIRULENCE ANALYSIS OF THE 548 PIERCE’S DISEASE AGENT XYLELLA FASTIDIOSA. Pierce’s Disease Research Symposium. 549
9. Chappel, J. A., M. R. Hollingdale, and A. S. Kang. 2004. IgG(4) Pf NPNA-1 a human anti-550 Plasmodium falciparum sporozoite monoclonal antibody cloned from a protected individual 551 inhibits parasite invasion of hepatocytes. Hum Antibodies 13:91-96. 552
10. Chappel, J. A., W. O. Rogers, S. L. Hoffman, and A. S. Kang. 2004. Molecular dissection of the 553 human antibody response to the structural repeat epitope of Plasmodium falciparum sporozoite 554 from a protected donor. Malar J 3:28. 555
11. Chatterjee, S., R. P. P. Almeida, and S. Lindow. 2008. Living in two worlds: The plant and insect 556 lifestyles of Xylella fastidiosa. Annu Rev Phytopathol 46:243-271. 557
12. Churchill, M. E., E. A. Stura, C. Pinilla, J. R. Appel, R. A. Houghten, D. H. Kono, R. S. Balderas, G. 558 G. Fieser, U. Schulze-Gahmen, and I. A. Wilson. 1994. Crystal structure of a peptide complex of 559 anti-influenza peptide antibody Fab 26/9. Comparison of two different antibodies bound to the 560 same peptide antigen. J Mol Biol 241:534-556. 561
13. Davis, M. J., A. H. Purcell, and S. V. Thomson. 1980. Isolation Media for the Pierces Disease 562 Bacterium. Phytopathology 70:425-429. 563
14. Davis, M. J., A. H. Purcell, and S. V. Thomson. 1978. Pierce's disease of grapevines: isolation of 564 the causal bacterium. Science 199:75-77. 565
15. De La Fuente, L., T. J. Burr, and H. C. Hoch. 2008. Autoaggregation of Xylella fastidiosa cells is 566 influenced by type I and type IV pili. Appl Environ Microbiol 74:5579-5582. 567
16. Durvasula, R. V., A. Gumbs, A. Panackal, O. Kruglov, S. Aksoy, R. B. Merrifield, F. F. Richards, 568 and C. B. Beard. 1997. Prevention of insect-borne disease: an approach using transgenic 569 symbiotic bacteria. Proc Natl Acad Sci U S A 94:3274-3278. 570
17. Durvasula, R. V., A. Gumbs, A. Panackal, O. Kruglov, J. Taneja, A. S. Kang, C. Cordon-Rosales, F. 571 F. Richards, R. G. Whitham, and C. B. Beard. 1999. Expression of a functional antibody fragment 572 in the gut of Rhodnius prolixus via transgenic bacterial symbiont Rhodococcus rhodnii. Med Vet 573 Entomol 13:115-119. 574
on July 10, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from

18. Fang, W., J. Vega-Rodriguez, A. K. Ghosh, M. Jacobs-Lorena, A. Kang, and R. J. St Leger. 2011. 575 Development of transgenic fungi that kill human malaria parasites in mosquitoes. Science 576 331:1074-1077. 577
19. Feil, H., W. S. Feil, and S. E. Lindow. 2007. Contribution of Fimbrial and Afimbrial Adhesins of 578 Xylella fastidiosa to Attachment to Surfaces and Virulence to Grape. Phytopathology 97:318-579 324. 580
20. Gasteiger, E., A. Gattiker, C. Hoogland, I. Ivanyi, R. D. Appel, and A. Bairoch. 2003. ExPASy: the 581 proteomics server for in-depth protein knowledge and analysis. Nucleic Acids Research 31:3784-582 3788. 583
21. Gram, H., L. A. Marconi, C. F. Barbas, 3rd, T. A. Collet, R. A. Lerner, and A. S. Kang. 1992. In 584 vitro selection and affinity maturation of antibodies from a naive combinatorial immunoglobulin 585 library. Proc Natl Acad Sci U S A 89:3576-3580. 586
22. Hanes, J., and A. Pluckthun. 1997. In vitro selection and evolution of functional proteins by 587 using ribosome display. Proc Natl Acad Sci U S A 94:4937-4942. 588
23. He, M., and M. J. Taussig. 1997. Antibody-ribosome-mRNA (ARM) complexes as efficient 589 selection particles for in vitro display and evolution of antibody combining sites. Nucleic Acids 590 Res 25:5132-5134. 591
24. Hoch, H. C., T. J. Burr, D. A. Cooksey, M. Black, A. Walker, X. Shi, and D. Athinuwat. 2010. 592 Presented at the 2010 Pierce's Disease Research Symposium. California Department of Food and 593 Agriculture, Sacramento, CA., San Diego, CA. 594
25. Holliger, P., T. Prospero, and G. Winter. 1993. "Diabodies": small bivalent and bispecific 595 antibody fragments. Proc Natl Acad Sci U S A 90:6444-6448. 596
26. Huse, W. D., L. Sastry, S. A. Iverson, A. S. Kang, M. Alting-Mees, D. R. Burton, S. J. Benkovic, 597 and R. A. Lerner. 1989. Generation of a large combinatorial library of the immunoglobulin 598 repertoire in phage lambda. Science 246:1275-1281. 599
27. Kang, A. S., C. F. Barbas, K. D. Janda, S. J. Benkovic, and R. A. Lerner. 1991. Linkage of 600 recognition and replication functions by assembling combinatorial antibody Fab libraries along 601 phage surfaces. Proc Natl Acad Sci U S A 88:4363-4366. 602
28. Kettleborough, C. A., J. Saldanha, K. H. Ansell, and M. M. Bendig. 1993. Optimization of primers 603 for cloning libraries of mouse immunoglobulin genes using the polymerase chain reaction. Eur J 604 Immunol 23:206-211. 605
29. Killiny, N., A. Rashed, and R. P. Almeida. 2012. Disrupting the transmission of a vector-borne 606 plant pathogen. Appl Environ Microbiol 78:638-643. 607
30. Kohler, G., and C. Milstein. 1975. Continuous cultures of fused cells secreting antibody of 608 predefined specificity. Nature 256:495-497. 609
31. Koide, T., R. Z. Vencio, and S. L. Gomes. 2006. Global gene expression analysis of the heat shock 610 response in the phytopathogen Xylella fastidiosa. J Bacteriol 188:5821-5830. 611
32. Kozak, M. 1987. An analysis of 5'-noncoding sequences from 699 vertebrate messenger RNAs. 612 Nucleic Acids Res 15:8125-8148. 613
33. Li, Y., G. Hao, C. D. Galvani, Y. Meng, L. De La Fuente, H. C. Hoch, and T. J. Burr. 2007. Type I 614 and type IV pili of Xylella fastidiosa affect twitching motility, biofilm formation and cell-cell 615 aggregation. Microbiology 153:719-726. 616
34. Markiv, A., B. Anani, R. V. Durvasula, and A. S. Kang. 2011. Module based antibody 617 engineering: a novel synthetic REDantibody. J Immunol Methods 364:40-49. 618
35. Markiv, A., R. Beatson, J. Burchell, R. V. Durvasula, and A. S. Kang. 2011. Expression of 619 recombinant multi-coloured fluorescent antibodies in gor -/ trxB- E. coli cytoplasm. BMC 620 Biotechnol 11:117. 621
on July 10, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from

36. Mattheakis, L. C., R. R. Bhatt, and W. J. Dower. 1994. An in vitro polysome display system for 622 identifying ligands from very large peptide libraries. Proc Natl Acad Sci U S A 91:9022-9026. 623
37. McCafferty, J., A. D. Griffiths, G. Winter, and D. J. Chiswell. 1990. Phage antibodies: 624 filamentous phage displaying antibody variable domains. Nature 348:552-554. 625
38. Nardin, E. H., V. Nussenzweig, R. S. Nussenzweig, W. E. Collins, K. T. Harinasuta, P. Tapchaisri, 626 and Y. Chomcharn. 1982. Circumsporozoite proteins of human malaria parasites Plasmodium 627 falciparum and Plasmodium vivax. J Exp Med 156:20-30. 628
39. Purcell, A. H., A. H. Finlay, and D. L. McLean. 1979. Pierce's Disease Bacterium: Mechanism of 629 Transmission by Leafhopper Vectors. Science 206:839-841. 630
40. Ramirez, J. L., T. M. Perring, and T. A. Miller. 2008. Fate of a genetically modified bacterium in 631 foregut of glassy-winged sharpshooter (Hemiptera: Cicadellidae). J Econ Entomol 101:1519-632 1525. 633
41. Rasgon, J. L. 2011. Using infections to fight infections: paratransgenic fungi can block malaria 634 transmission in mosquitoes. Future Microbiol 6:851-853. 635
42. Schofield, D. J., A. R. Pope, V. Clementel, J. Buckell, S. Chapple, K. F. Clarke, J. S. Conquer, A. 636 M. Crofts, S. R. Crowther, M. R. Dyson, G. Flack, G. J. Griffin, Y. Hooks, W. J. Howat, A. Kolb-637 Kokocinski, S. Kunze, C. D. Martin, G. L. Maslen, J. N. Mitchell, M. O'Sullivan, R. L. Perera, W. 638 Roake, S. P. Shadbolt, K. J. Vincent, A. Warford, W. E. Wilson, J. Xie, J. L. Young, and J. 639 McCafferty. 2007. Application of phage display to high throughput antibody generation and 640 characterization. Genome Biol 8:R254. 641
43. Simpson, A. J., F. C. Reinach, P. Arruda, F. A. Abreu, M. Acencio, R. Alvarenga, L. M. Alves, J. E. 642 Araya, G. S. Baia, C. S. Baptista, M. H. Barros, E. D. Bonaccorsi, S. Bordin, J. M. Bove, M. R. 643 Briones, M. R. Bueno, A. A. Camargo, L. E. Camargo, D. M. Carraro, H. Carrer, N. B. Colauto, C. 644 Colombo, F. F. Costa, M. C. Costa, C. M. Costa-Neto, L. L. Coutinho, M. Cristofani, E. Dias-Neto, 645 C. Docena, H. El-Dorry, A. P. Facincani, A. J. Ferreira, V. C. Ferreira, J. A. Ferro, J. S. Fraga, S. C. 646 Franca, M. C. Franco, M. Frohme, L. R. Furlan, M. Garnier, G. H. Goldman, M. H. Goldman, S. L. 647 Gomes, A. Gruber, P. L. Ho, J. D. Hoheisel, M. L. Junqueira, E. L. Kemper, J. P. Kitajima, J. E. 648 Krieger, E. E. Kuramae, F. Laigret, M. R. Lambais, L. C. Leite, E. G. Lemos, M. V. Lemos, S. A. 649 Lopes, C. R. Lopes, J. A. Machado, M. A. Machado, A. M. Madeira, H. M. Madeira, C. L. Marino, 650 M. V. Marques, E. A. Martins, E. M. Martins, A. Y. Matsukuma, C. F. Menck, E. C. Miracca, C. Y. 651 Miyaki, C. B. Monteriro-Vitorello, D. H. Moon, M. A. Nagai, A. L. Nascimento, L. E. Netto, A. 652 Nhani, Jr., F. G. Nobrega, L. R. Nunes, M. A. Oliveira, M. C. de Oliveira, R. C. de Oliveira, D. A. 653 Palmieri, A. Paris, B. R. Peixoto, G. A. Pereira, H. A. Pereira, Jr., J. B. Pesquero, R. B. Quaggio, P. 654 G. Roberto, V. Rodrigues, M. R. A. J. de, V. E. de Rosa, Jr., R. G. de Sa, R. V. Santelli, H. E. 655 Sawasaki, A. C. da Silva, A. M. da Silva, F. R. da Silva, W. A. da Silva, Jr., J. F. da Silveira, et al. 656 2000. The genome sequence of the plant pathogen Xylella fastidiosa. The Xylella fastidiosa 657 Consortium of the Organization for Nucleotide Sequencing and Analysis. Nature 406:151-159. 658
44. Smith, G. P. 1985. Filamentous fusion phage: novel expression vectors that display cloned 659 antigens on the virion surface. Science 228:1315-1317. 660
45. Spyropoulos, I. C., T. D. Liakopoulos, P. G. Bagos, and S. J. Hamodrakas. 2004. TMRPres2D: high 661 quality visual representation of transmembrane protein models. Bioinformatics 20:3258-3260. 662
46. Suzuki, C., H. Ueda, E. Suzuki, and T. Nagamune. 1997. Construction, bacterial expression, and 663 characterization of hapten-specific single-chain Fv and alkaline phosphatase fusion protein. J 664 Biochem 122:322-329. 665
47. Tachibana, H., M. Takekoshi, X. J. Cheng, Y. Nakata, T. Takeuchi, and S. Ihara. 2004. Bacterial 666 expression of a human monoclonal antibody-alkaline phosphatase conjugate specific for 667 Entamoeba histolytica. Clin Diagn Lab Immunol 11:216-218. 668
on July 10, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from

48. Tang, J., L. Wang, A. Markiv, S. A. Jeffs, H. Dreja, A. McKnight, M. He, and A. S. Kang. 2012. 669 Accessing of Recombinant Human Monoclonal Antibodies from Patient Libraries by Eukaryotic 670 Ribosome Display. Human Antibodies in press. 671
49. Zavala, F., J. P. Tam, M. R. Hollingdale, A. H. Cochrane, I. Quakyi, R. S. Nussenzweig, and V. 672 Nussenzweig. 1985. Rationale for development of a synthetic vaccine against Plasmodium 673 falciparum malaria. Science 228:1436-1440. 674
50. Zhang, H., A. Torkamani, T. M. Jones, D. I. Ruiz, J. Pons, and R. A. Lerner. 2011. Phenotype-675 information-phenotype cycle for deconvolution of combinatorial antibody libraries selected 676 against complex systems. Proc Natl Acad Sci U S A 108:13456-13461. 677
678
679
680
on July 10, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from