ribozyme-targeting elucidates a direct role of pleiotrophin in
TRANSCRIPT

%E JOURNAL OF BIOL~GICAL CHEMISTRS 0 1994 by The American Society for Biochemistry and Molecular Biology, Inc
Vol. 269, No. 33, Issue of August 19, pp. 21358-21363, 1994 Printed in U.S.A.
Ribozyme-targeting Elucidates a Direct Role of Pleiotrophin in Tumor Growth*
(Received for publication, April 27, 1994, and in revised form, May 19, 1994)
Frank Czubayko, Anna Tate RiegelS, and Anton Wellstein8 From the Lombardi Cancer Center and Department of Pharmacology, Georgetown University, Washington, D. C. 20007
The growth and metastasis of solid tumors rely on the activities of polypeptide growth factors. However, nu- merous growth factors are expressed in tumors, and it is difficult to decipher which are essential for tumor pro- gression. We found the secreted growth factor pleiotro- phin (PTN) expressed at high levels in a number of hu- man tumor cell lines as well as in tumor samples. To assess the role of PTN in tumor growth, we inactivated the PTN gene with PTN-targeted hammerhead ribozyme constructs. Cotransfection of PTN and of the ribozymes inhibited PTN-induced colony formation of PTN-respon- sive cells whereas a point mutant, catalytically inactive ribozyme was ineffective. Colony formation induced by transfections with a closely related growth factor (mid- kine) was not affected by the ribozymes. In human mela- noma cells that express high levels of PTN mRNA, stable transfection with PTN-targeted ribozymes quenched production of PTN, inhibited colony formation of the cells, and prevented their tumor growth in mice. This demonstrates that expression of a growth factor can be a rate-limiting step for malignant progression and sug gests that ribozymes could be used therapeutically to target tumor growth factors.
Polypeptide growth factors contribute to the development and maintenance of normal tissues and are essential for the growth and metastasis of solid tumors (1). During tumor pro- gression these factors function as autocrine stimulators of tu- mor cells and/or serve to recruit stromal tissue and blood sup- ply to the expanding tumor mass (2). We purified the heparin- binding growth factor pleiotrophin (PTN)’ from the supernatants of human breast cancer cells (3) and found this gene expressed in a number of human tumor cell lines as well as in human tumor tissues but not in normal endothelial cells, melanocytes, or fibroblasts (4). The present paper addresses the contribution of this factor to tumor growth.
PTN was originally purified and cloned from developing brain, uterus, and placental tissue (5-10). Recent in situ hy- bridization studies in rodents (11, 12) demonstrated that the gene is expressed at the highest levels in the neuroepithelium and ependyma during the second half of intrauterine develop- ment. During the same developmental phase, PTN mRNA was
Healtb/National Cancer Institute (to A. W.) and by a fellowship from the * This work was supported by grants from the National Institutes of
University of Bonn (to F. C.). The costs of publication of this article were
therefore be hereby marked “advertisement” in accordance with 18 defrayed in part by the payment of page charges. This article must
U.S.C. Section 1734 solely to indicate this fact.
NIH. $ Recipient of a research career development award from NIDDW
5 To whom correspondence should be addressed: Lombardi Cancer Center, Georgetown University, 3800 hservoir Rd. N. W., Washington,
The abbreviations used are: PTN, pleiotrophin; MK, midkine; Rz, D. C. 20007 Tel.: 202-687-3672; Fax: 202-687-4821.
ribozyme; CMV, cytomegalovirus.
also detected in the mesenchyme of the lung, reproductive tract, gut, kidney, and bone. With progressing organ differen- tiation, PTN gene expression was increasingly restricted to a few sites and dropped sharply around birth. Finally, in the adult, only few structures such as the hippocampal region and cerebral cortex showed detectable levels of gene expression. The developmental regulation of the PTN protein appears to parallel the expression profile of the mRNA, and the highest protein levels in the growing brain coincide with periods of rapid sprouting of nerve cells (7). This very distinct pattern of gene expression suggests an important role for PTN during brain development and organogenesis. Most interestingly, we found the highest levels of PTN expression in tumors originat- ing from neuroectodermal cell lineages, i.e. in human mela- noma and glioblastoma multiforme tissues’ and in cell lines derived from these malignant lesions (4). This finding supports the notion that developmentally regulated genes may be acti- vated in a preferential fashion during malignant transforma- tion (1).
PTN stimulates neurite outgrowth (6, 10, 13), is mitogenic for fibroblasts (4, 6, 9) and for endothelial cells (4, 14), and could thus serve as an angiogenesis factor for tumors. A poten- tial role of PTN during tumor growth is suggested by studies in which overexpression of PTN in SW-13 cells (4) and in 3T3- fibroblasts (15) supported tumor growth of these non-tumori- genic cells in athymic nude mice. However, a wide variety of growth factors is expressed at any given time in human tumor tissues as well as in cultured cell lines (1, 16), and it is very difficult to decipher which are essential for tumor growth and are ultimately promising targets for specific therapeutic agents. In particular, melanoma cells that express high levels of PTN (4) also express fibroblast growth factors (16) that have biologic activities similar to those of PTN. Much like PTN, fibroblast growth factors stimulate neurites, fibroblasts, and endothelial cells and can also serve as tumor angiogenesis fac- tors (17, 18). To directly assess whether the expression of PTN in tumor cells such as melanoma plays a significant role for their malignant growth, we decided to inactivate the PTN gene product and to study the resulting phenotype of the cells.
We used enzymatically active RNAs, i.e. ribozymes (191, that were designed to specifically cleave the PTN mRNA and thus should generate cell lines that have significantly reduced PTN levels. The targeting of defined RNAs with engineered ri- bozyme constructs has been used to inactivate viral gene prod- ucts in cells and had been suggested to generate dominant- negative mutants of endogenous cellular genes (20-22) (reviewed in Refs. 23,24). We generated expression vectors for hammerhead ribozymes (20, 21) that were designed to cleave PTN mRNA in the open reading frame 66 and 261 nucleotides downstream of the translation initiation site. The specific cleavage of the target mRNA by these ribozymes was studied in
F. Czubayko and A. Wellstein, unpublished results.
21358

Ribozyme Targeting of Pleiotrophin 21359
vitro after run-off transcription from the respective constructs. The in vivo efficacy of the ribozymes was tested in cotransfec- tion assays with an expression vector for PTN into PTN-sensi- tive cells. Parallel in vivo experiments with the midkine gene (MK) (25) served as a control for nonspecific activity of these ribozymes since PTN and MK belong to the same gene family, and their cDNAs are approximately 50% homologous (reviewed in Ref. 26). Due to the base pairing of ribozymes required for recognition and cleavage of the target site in the substrate RNA, it is important to demonstrate that effects of the ri- bozymes are independent from this antisense activity (27). We generated a point-mutant ribozyme construct that was enzy- matically inactive to assess the extent of the antisense activity inherent t o the mechanism of action of ribozymes.
Our data indicate that the effects of PTN can be suppressed specifically by PTN-targeted ribozymes. Furthermore, expres- sion of these ribozymes in human melanoma cells, that consti- tutively express high levels of F'TN mRNA, inhibited their tu- mor formation in mice. To our knowledge, this is the first report that uses ribozyme targeting of a polypeptide growth factor and demonstrates that a particular growth factor can be a rate- limiting step for melanoma growth in vivo.
MATERIALS AND METHODS Generation of Constructs-The PTN cDNA covering the open reading
frame of the PTN gene was subcloned into the HindIIIIXbaI sites of the pRdCMV plasmid (Invitrogen, San Diego, CA) as described previously (4). To generate the ribozyme constructs, synthetic sense and antisense oligonucleotides containing the catalytic center and the flanking re- gions of each of the ribozymes (see Fig. 1B) were annealed and ligated into the HindIIIINotI sites of the pRc/CMV plasmid. To generate the MK expression vector (pMK), a cDNA fragment covering the open reading frame of the human MK cDNA (gift of Dr. Seddon, see Ref. 28) was inserted into the multiple cloning site of the pRdCMV plasmid (see Fig. 1 and Ref. 4).
Cell Lines, Dansfections, and Growth Assays-SW-13 cells (3, 29) were transfected using a modified calcium phosphate method as de- scribed previously (4). Briefly, SW-13 cells were plated onto 100-mm Petri dishes, grown until they reached 50% confluence, and transfected for 12 h at 35 "C with 2.5% CO, (30) with the amounts of plasmid DNA indicated in the figure legends. The transfection media was replaced with normal growth media, and cells were harvested after 48 h and either used in soft agar colony formation assays (transient transfection experiments) or grown for another 6 weeks in media containing 500 pg/ml of G418 to select stable integrants. Soft agar assays were incu- bated for 10 days and evaluated as described previously (4). Human melanoma cells (WM852, gift from Dr. M. Heerlyn, Wistar Institute, Philadelphia) were transfected with 10 pg of empty vectors or pRz66 or pRz261 (see Fig. 1) using Lipofectaminem (Life Technologies, Inc.). Briefly, WM852 cells at 50% confluence were incubated for 5 h with 10 pg of plasmid DNA mixed with 70 pl of Lipofectaminem reagant in serum-free medium (Optimem, Life Technologies, Inc.) at 37 "C, 5% CO,. The transfection medium was replaced with fresh medium, and 36 h later G418 (1,500 pg/ml) was added to select stably transfected cells for another 8 weeks.
In Vitro Assay for Ribozyme Activity-To generate in vitro tran- scripts, plasmids containing the PTN or the ribozyme cDNA were lin- earized with XbaI. Transcription reactions were carried out with T7 RNA polymerase (31) as recommended by the supplier (Promega, Madi- son, WI). PTN transcripts were labeled by including 50 pCi of [CY-~VICTP in the reaction. The products of the reaction were resus- pended in 50 R ~ M Ms-C1, pH 8.5, with 20 R ~ M MgCl,, heated for 2 min at 85 "C, and incubated for 15 min at 50 "C. Substrate and ribozyme transcripts were then mixed and incubated at 50 "C, and aliquots were removed after 20, 60, and 120 min, boiled briefly, separated on a 6% uredpolyacrylamide gel, and products were detected by autoradiography.
Northern BZots and Immunoassay for PTN-For the Northern blots 20 pg of total RNA from each cell line were separated in an agarose gel and blotted onto nitrocellulose, hybridized with a F'TN cDNAprobe, and autoradiographed for 48 h (4). For quantitation, the 1.5-kilobase bands were excised and counted in a scintillation counter. For the immunoas- says, conditioned media from the differently transfected cells were con- centrated and partially purified by heparin-affinity chromatography.
100 pl of the 0.9 M NaCl eluates from the columns were assayed for PTN using a rabbit antiserum raised against PTN and a secondary antibody coupled to alkaline phosphatase as a detection system as described previously (4).
RESULTS
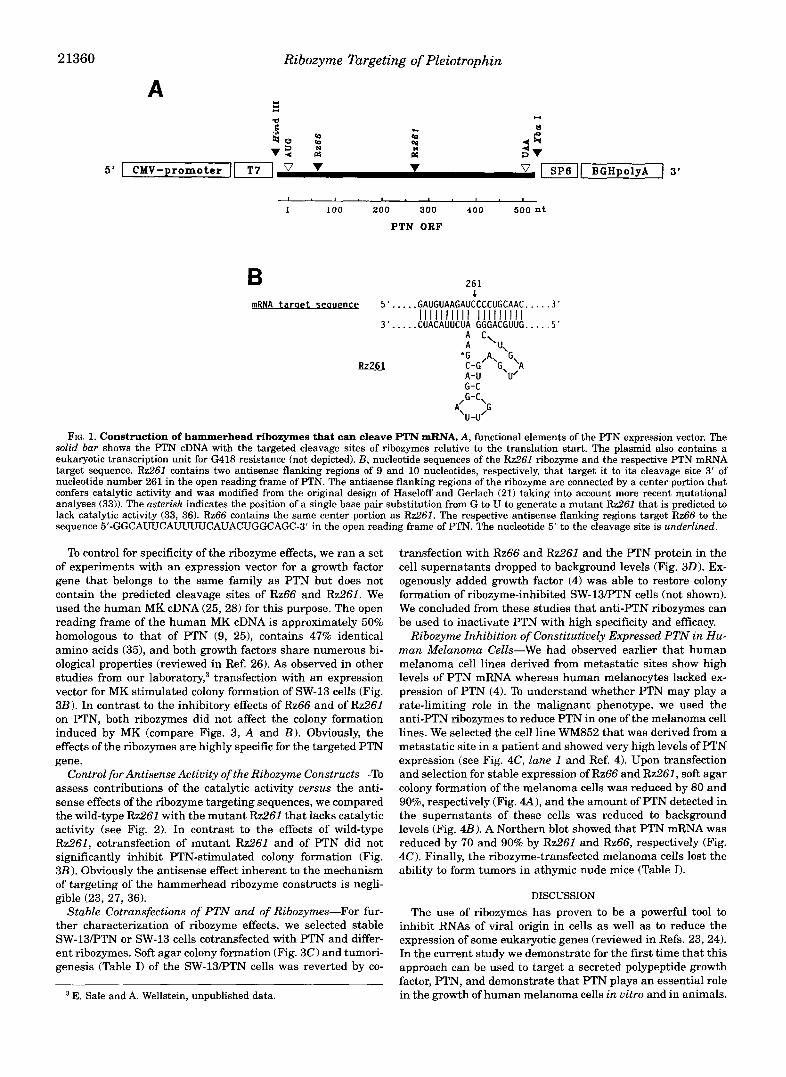
Selection of Ribozyme Target Sequences in PTN-To select regions in the growth factor mRNA for targeting by hammer- head ribozymes (20, 21), we searched the predicted secondary structure of the PTN mRNA (32) for open loops that also con- tained the consensus sequence for ribozyme cleavage. The con- sensus sequence for ribozyme cleavage is a 5'-iVUX-3' sequence that is cleaved 3' of X (N is any nucleotide and X is either A, U, or C; see Ref. 23). From these potential sites, we selected two within the open reading frame of the PTN mRNA that were 66 and 261 nucleotides downstream of the translation initiation site (targeted by Rz66 and Rz261 respectively; see Fig. lA).
Generation of Ribozyme Constructs-The ribozymes that we generated were modeled along the hammerhead structure (Fig. LB) that is derived from the original design by Haseloff and Gerlach (21). We modified and minimized the catalytic center portion to 22 nucleotides based on the mutational analysis that had revealed the essential elements for a catalytically active RNA (33). As illustrated for our Rz261 (Fig. 1B), the molecule is predicted to form three double helices ("hammerhead") which meet at the targeted cleavage site. In addition, two stretches of nucleotides with the sequences 5"CUGAXGA and 5'-GAAA ( X is either A, U, or C) are required for the catalytic activity. The cleavage is directed to a specific site in the targeted RNAby two stretches of 8-12 nucleotides each that flank the catalytic cen- ter on its 3'- and 5'-side. These flanking sequences hybridize with the targeted RNA by Watson-Crick base pairing (reviewed in Ref. 23).
We inserted the different ribozymes (Fig. 1B) into the mul- tiple cloning site of the pRc/CMV plasmid (see Fig. lA). This expression vector contains the CMV promoter for eukaryotic cells as well as a polyadenylation signal to enable transcription termination and the addition of a poly(A) tail. These elements should increase RNA stability and transport to the cytoplasm if required (23, 34).
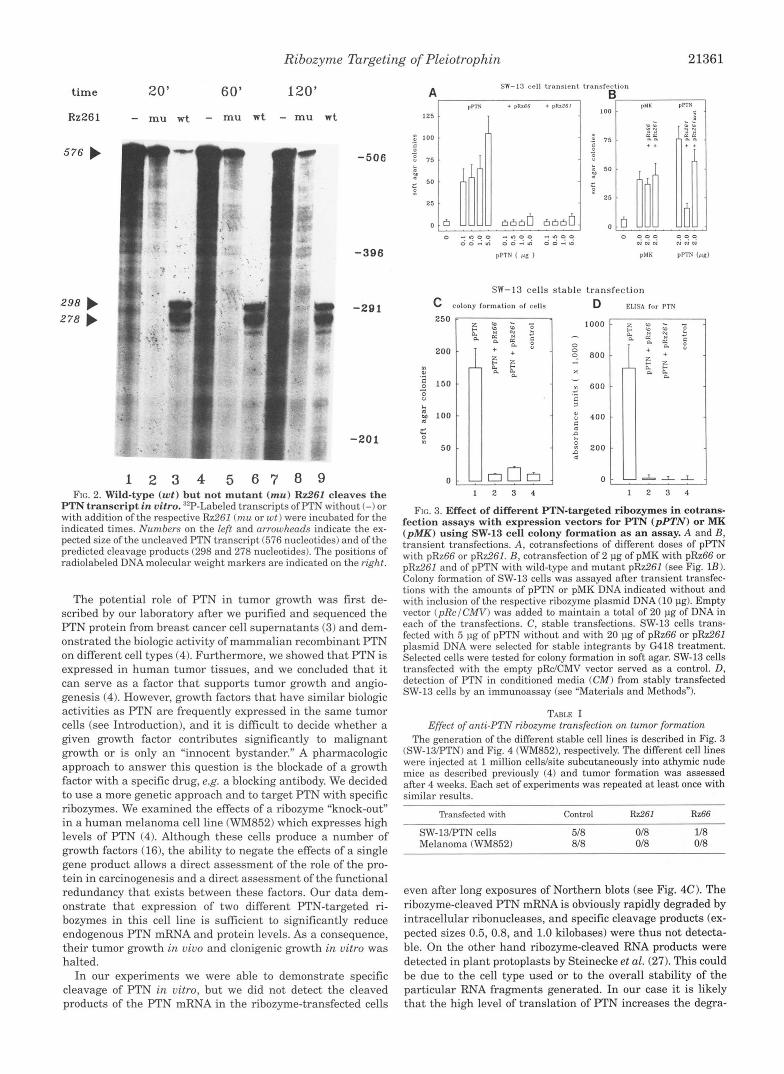
In Vitro Activity of PTN-targeted Ribozymes-In a first set of experiments, we evaluated the catalytic activity of the ri- bozymes with in vitro run-off transcripts from the PTN and ribozyme constructs (31) (Fig. 2). Cleavage of the 576-nucle- otide PTN mRNA fragment (see Fig. lA) by Rz261 was com- plete within 60 min and yielded the predicted fragments of 298 and 278 nucleotides (Fig. 2, lanes 3, 6, and 9). The point mu- tation of G to U in the catalytic domain of Rz261 (Fig. 1B) resulted in a loss of the catalytic activity (Fig. 2, lanes 2 , 5 , and 8) as predicted by the mutational studies of McCall et al. (33). This mutant Rz262 construct was used in in vivo transfection studies to distinguish antisense effects due to the base pairing of the ribozyme targeting sequences from effects on PTN due to the catalytic activity (see below). The in vitro efficacy of Rz66 was similar to that of Rz261 and specifically cleaved the PTN mRNA into the predicted fragments (not shown).
In Vivo Efficacy and Specificity of Ribozymes in Tkansient Cotransfection Assays with PTN-We then studied the efficacy of the ribozyme constructs in vivo using a human cell line (SW-13) that responds to the transient or stable expression of PTN with the formation of colonies in soft agar (4). The data in Fig. 3A illustrate that transient transfections of SW-13 cells with increasing doses of the PTN expression vector resulted in increasing numbers of colonies of these cells in soft agar. This response to PTN was abolished after cotransfection of Rz66 and Rz261 (Fig. 3 A ) , thus demonstrating the efficacy of the cotrans- fected ribozymes in cells in vivo.
2 1360
A Ribozyme Targeting of Pleiotrophin
5 ’ I CMV-promoter
1 100 200 300 400 500 nt
PTN ORF
B 261 1
mRNA taroet seauence 5 ’ ..... GAUGUAAGAUCCCCUGCAAC . . . . . 3 ’
3 ’ ..... CUACAUUCUA GGGACGUUG ..... 5 ‘ I I I I I I I I I I I I I I I I I I I
* E “
,A\ G C-G G, ‘A A-U ‘U’ G-C G-C,
u -u A< ,G
FIG. 1. Construction of hammerhead ribozymes that can cleave PTN mRNk A, functional elements of the PTN expression vector. The solid bar shows the PTN cDNA with the targeted cleavage sites of ribozymes relative to the translation start. The plasmid also contains a eukaryotic transcription unit for G418 resistance (not depicted). B, nucleotide sequences of the Rz261 ribozyme and the respective PTN mRNA target sequence. Rz261 contains two antisense flanking regions of 9 and 10 nucleotides, respectively, that target it to its cleavage site 3’ of nucleotide number 261 in the open reading frame of PTN. The antisense flanking regions of the ribozyme are connected by a center portion that confers catalytic activity and was modified from the original design of Haseloff and Gerlach (21) taking into account more recent mutational analyses (33)). The asterisk indicates the position of a single base pair substitution from G to U to generate a mutant Rz261 that is predicted to lack catalytic activity (33, 36). Rz66 contains the same center portion as Rz261. The respective antisense flanking regions target Rz66 to the sequence 5’-GGCAWCAUUUU~AUACUGGCAGC-3’ in the open reading frame of PTN. The nucleotide 5’ to the cleavage site is underlined.
To control for specificity of the ribozyme effects, we ran a set of experiments with an expression vector for a growth factor gene that belongs to the same family as PTN but does not contain the predicted cleavage sites of Rz66 and Rz261. We used the human MK cDNA (25,281 for this purpose. The open reading frame of the human MK cDNA is approximately 50% homologous to that of PTN (9, 251, contains 47% identical amino acids (351, and both growth factors share numerous bi- ological properties (reviewed in Ref. 26). As observed in other studies from our lab~ratory,~ transfection with an expression vector for MK stimulated colony formation of SW-13 cells (Fig. 3B). In contrast to the inhibitory effects of Rz66 and of Rz261 on PTN, both ribozymes did not affect the colony formation induced by MK (compare Figs. 3, A and B). Obviously, the effects of the ribozymes are highly specific for the targeted PTN gene.
Control for Antisense Activity of the Ribozyme Constructs-To assess contributions of the catalytic activity versus the anti- sense effects of the ribozyme targeting sequences, we compared the wild-type Rz261 with the mutant Rz261 that lacks catalytic activity (see Fig. 2). In contrast to the effects of wild-type Rz262, cotransfection of mutant Rz261 and of PTN did not significantly inhibit PTN-stimulated colony formation (Fig. 3B). Obviously the antisense effect inherent to the mechanism of targeting of the hammerhead ribozyme constructs is negli- gible (23, 27, 36).
Stable Cotransfections of PTN and of Ribozymes-For fur- ther characterization of ribozyme effects, we selected stable SW-13PTN or SW-13 cells cotransfected with PTN and differ- ent ribozymes. Soft agar colony formation (Fig. 3C) and tumori- genesis (Table I) of the SW-l3/P”N cells was reverted by co-
E. Sale and A. Wellstein, unpublished data.
transfection with Rz66 and Rz261 and the PTN protein in the cell supernatants dropped to background levels (Fig. 30). Ex- ogenously added growth factor (4) was able to restore colony formation of ribozyme-inhibited SW-l3/PTN cells (not shown). We concluded from these studies that anti-PTN ribozymes can be used to inactivate PTN with high specificity and efficacy.
Ribozyme Inhibition of Constitutively Expressed PTN in Hu- man Melanoma Cells-We had observed earlier that human melanoma cell lines derived from metastatic sites show high levels of PTN mRNA whereas human melanocytes lacked ex- pression of PTN (4). To understand whether PTN may play a rate-limiting role in the malignant phenotype, we used the anti-PTN ribozymes to reduce PTN in one of the melanoma cell lines. We selected the cell line WM852 that was derived from a metastatic site in a patient and showed very high levels of PTN expression (see Fig. 4C, lane 1 and Ref. 4). Upon transfection and selection for stable expression of Rz66 and Rz261, soft agar colony formation of the melanoma cells was reduced by 80 and 90%, respectively (Fig. 4A), and the amount of PTN detected in the supernatants of these cells was reduced to background levels (Fig. 4B). A Northern blot showed that PTN mRNA was reduced by 70 and 90% by Rz261 and Rz66, respectively (Fig. 4C). Finally, the ribozyme-transfected melanoma cells lost the ability to form tumors in athymic nude mice (Table I).
DISCUSSION
The use of ribozymes has proven to be a powerful tool to inhibit RNAs of viral origin in cells as well as to reduce the expression of some eukaryotic genes (reviewed in Refs. 23,241. In the current study we demonstrate for the first time that this approach can be used to target a secreted polypeptide growth factor, PTN, and demonstrate that PTN plays an essential role in the growth of human melanoma cells in vitro and in animals.
Ribozyme Targeting of Pleiotrophin 21361
time 20’ 60’ 120’
Rz261 - mu w t - mu w t - m u r t
298 278
1 2 3 4 5 6 7 8 9
-506
-396
-29 1
-201
FIG. 2. Wild-type (wt) but not mutant (mu) Rz261 cleaves the PTN transcript in vitro. ”P-Labeled transcripts of PTN without (-) or with addition of the respective Rz261 ( n u or wt) were incubated for the indicated times. Numbers on the left and arrowheads indicate the ex- pected size of the uncleaved FTN transcript (576 nucleotides) and of the predicted cleavage products (298 and 278 nucleotides). The positions of radiolabeled DNAmolecular weight markers are indicated on the right.
The potential role of PTN in tumor growth was first de- scribed by our laboratory after we purified and sequenced the PTN protein from breast cancer cell supernatants (3) and dem- onstrated the biologic activity of mammalian recombinant PTN on different cell types (4). Furthermore, we showed that PTN is expressed in human tumor tissues, and we concluded that it can serve as a factor that supports tumor growth and angio- genesis (4). However, growth factors that have similar biologic activities as PTN are frequently expressed in the same tumor cells (see Introduction), and i t is difficult to decide whether a given growth factor contributes significantly to malignant growth or is only an “innocent bystander.” A pharmacologic approach to answer this question is the blockade of a growth factor with a specific drug, e.g. a blocking antibody. We decided to use a more genetic approach and to target PTN with specific ribozymes. We examined the effects of a ribozyme “knock-out” in a human melanoma cell line (WM852) which expresses high levels of PTN (4). Although these cells produce a number of growth factors (161, the ability to negate the effects of a single gene product allows a direct assessment of the role of the pro- tein in carcinogenesis and a direct assessment of the functional redundancy that exists between these factors. Our data dem- onstrate that expression of two different PTN-targeted ri- bozymes in this cell line is sufficient to significantly reduce endogenous PTN mRNA and protein levels. As a consequence, their tumor growth in vivo and clonigenic growth in vitro was halted.
In our experiments we were able to demonstrate specific cleavage of PTN in vitro, but we did not detect the cleaved products of the FTN mRNA in the ribozyme-transfected cells
L SW- I 3 cell transient t l
SW-13 cells stable transfection C colony formation of cells D EL!% for PTN
250
200
m .- 150 -
0
L s 100
=: y1
50
0 1 2 3 4
000
800
600
400
200
0
1 2 3 4
FIG. 3. Effect of different FTN-targeted ribozymes in cotrans- fection assays with expression vectors for PTN (pPTN) or MK ( p M K ) using SW-13 cell colony formation as an assay. A and B , transient transfections. A, cotransfections of different doses of pPTN with pRz66 or pRz261. B, cotransfection of 2 pg of pMK with pRz66 or pR7261 and of pFTN with wild-type and mutant pRz261 (see Fig. 1B). Colony formation of SW-13 cells was assayed after transient transfec- tions with the amounts of pFTN or pMK DNA indicated without and with inclusion of the respective ribozyme plasmid DNA (10 pg). Empty vector (pRcICMV) was added to maintain a total of 20 pg of DNA in each of the transfections. C , stable transfections. SW-13 cells trans- fected with 5 pg of pFTN without and with 20 pg of pRz66 or pRz261 plasmid DNA were selected for stable integrants by G418 treatment.
transfected with the empty pRc/CMV vector served as a control. D, Selected cells were tested for colony formation in soft agar. SW-13 cells
detection of FTN in conditioned media ( C M ) from stably transfected SW-13 cells by an immunoassay (see “Materials and Methods”).
TABLE I Effect of anti-PTN ribozyme transfection on tumor formation
The generation of the different stable cell lines is described in Fig. 3 (SW-l3/PTN) and Fig. 4 (WM8521, respectively. The different cell lines were injected a t 1 million cellslsite subcutaneously into athymic nude mice as described previously (4) and tumor formation was assessed after 4 weeks. Each set of experiments was repeated at least once with similar results.
Transfected with Control Rz261 Rz66
SW-l3/PTN cells 518 018 118 Melanoma (WM852) 818 018 018
even after long exposures of Northern blots (see Fig. 4 0 . The ribozyme-cleaved PTN mRNA is obviously rapidly degraded by intracellular ribonucleases, and specific cleavage products (ex- pected sizes 0.5, 0.8, and 1.0 kilobases) were thus not detecta- ble. On the other hand ribozyme-cleaved RNA products were detected in plant protoplasts by Steinecke et al. (27). This could be due to the cell type used or to the overall stability of the particular RNA fragments generated. In our case it is likely that the high level of translation of PTN increases the degra-

21362 Ribozyme Targeting of Pleiotrophin
melanoma cell stable transfection A colony formation of cells B ELISA for PTN
2 200
2 3 0
2 2 100 0 II) e
btl I 0
1 2 3 1 2 3
PTN -1440
-950
-780
1 2 3
-28s
-18s
. . 1 FIG. 4. Effect of stable transfections with FTN-targeted ri-
bozymes on human melanoma cells (WM862) that express PTN constitutively. A, soft agar colony formation of the cells after 14 days of incubation. B, detection of FTN in conditioned media from the cells by immunoassay. C, detection of FTN mRNA in the cells by Northern blot. Top, the position of the 1.5-kilobase FTN mRNA and of radiolabeled DNA markers are shown. Bottom, the ethidium bromide stain of the
the 18 S and 28 S ribosomal RNA are shown. agarose gel indicates equivalent loading of the lanes. The positions of
dation rate of the cleaved mRNA fragments (reviewed in Ref. 34).
Antisense activity of the ribozymes does not seem to play a significant role in their efficacy and the 70-90% reduction in cellular PTN mRNA due to cleavage by the ribozymes is most likely the predominant mechanism by which the ribozymes are reducing the amount of PTN protein secreted from the cells. Antisense sequences that flank the ribozyme core and target its activity to a specific stretch of nucleotides by base pairing can also block translation of the targeted mRNA and thus act as an antisense molecule. Since our point-mutant construct, which contains the same flanking sequences as wild-type Rz261, was not effective in the cotransfection assays with PTN (see Fig. 3), antisense effects appear to be of minor importance.
A general question is whether the ribozyme effects that we observed could be due to degradation of an RNA species that is related in sequence to PTN and which fortuitously is cleaved by the ribozyme constructs. Several pieces of evidence make this unlikely. First, both of our ribozyme constructs showed similar efficacy in two different systems, i.e. against the cotransfected PTN and against the endogenous gene product. Second, both constructs are targeted against regions in the PTN gene that do not share any homology and both are effective. Third, the lack of an effect of either of these two ribozymes against the closely related MK gene suggests that the effects on PTN are highly sequence-specific. Finally, ribozyme-transfected SW-13 and melanoma cells continued to proliferate and were grown for several weeks to select for stable integrants. Most likely the cells would not have survived nonspecific cleavage of different RNAs by the ribozymes. Taken together our data support the conclusion that only the PTN transcript is directly affected by the ribozymes.
In our current experiments, the level of ribozyme expression was barely detectable on a Northern blot (not shown), but even these low levels of ribozyme RNA in the melanoma cells gen- erated sufficient enzymatic activity to reduce the endogenous PTN mRNA and revert the malignant behavior of the cells. An interesting question is how the levels of ribozyme expression could be increased and how the efficacy of the ribozyme target- ing could be improved further. The expression vector into which our ribozyme constructs were inserted includes a polyadenyla- tion site, and the transcript from this vector should be capped at the 5'-end and thus contains two major elements of RNA stability (34). Furthermore, the CMV promoter driving ri- bozyme expression is highly active in the transfected cells. One additional modification of the constructs to generate higher steady-state expression levels of the ribozymes could be the inclusion of intronic sequences with proper splice junctions (34). As a further potential advantage of this approach, the cellular splicing apparatus may direct the respective ribozyme and the intron-containing endogenous PTN transcript (37) to the same cellular compartments and thus increase the efficacy of the ribozyme. Experiments addressing these questions are currently underway. The significance of the trafficking of ri- bozymes and their substrates was demonstrated in a very re- cent study by Sullenger and Cech (38) in which they showed that viral RNA in cells is only susceptible to cleavage by ri- bozymes when both are colocalized into the same intracellular compartment. With respect to the targeting of eukaryotic gene products, the inclusion of introns in a ribozyme may improve both the expression levels and the correct compartmental localization.
The results from our study have several implications for the role of PTN and the potential usefulness of the ribozyme con- structs in future analyses of the physiological and pathological role of PTN. It will be interesting to determine if the phenotype of other tumor cells which produce PTN can be similarly af- fected by PTN ribozymes and to what extent this growth factor is rate-limiting for their growth in vivo. In this context the efficacy of PTN-targeted ribozymes in glioblastoma multiforme will be very significant, since these tumors are highly malig- nant and may use PTN as their major angiogenesis factor2 Furthermore, it may be construed that in tumors of neuroec- todermal origin PTN contributes significantly to the malignant phenotype and may play a less prominent role in other cancers. As a potential application in oncology it is tempting to specu- late that targeting of a rate-limiting growth factor such as F'TN with specific ribozymes could open up novel avenues in the gene therapy of cancer in particular of such fatal diseases as metastatic melanoma and perhaps glioblastoma multiforme.

Ribozyme Targeting of Pleiotrophin 21363
This approach should be useful if the ribozymes can be deliv- (1991) Biochem. Biophys. Res. Commun. 180,145-151 ered effectively to a significant percentage of the tumor cells. 15. Chauhan,A. K., Li,Y. S., and Deuel, T. F. (1993)Proc. Natl. Acad. Sci. U. S. A.
Finally, the use Of PTN-targeted ribozymes to generate knock- 16. Rode&, u., and Herlyn, M. (1991) Cancer Metastasis Rev. 10,89-101 out transgenic animals should help elucidate the role of F'TN in 17. Burgess, W. H., and T. (1989) Annu. Reu. B k h e m . 589 575-606
embryonal development.
90,67%682
18. Folkman, J., and Shing, Y. (1992) J Biol. Chem. 267,10931-10934 19. Kruger, K, Grabowski, P. J., Zaug, A. J., Sands, J., Gottachling, D. E., and
Acknowledgments-We thank Drs. M. Avigan, S. G. Irving, M. E. 20. Uhlenbeck, 0. c. (1987) ~~t~~ 328,59&600 Cech, T. R. (1982) Cell 31, 147-157
Lippman, R. C. Schlegel (Georgetown University), and M. Herlyn 21. Haseloff, J., and Gerlach, W. L. (1988) Nature 334,585-591 (Wistar Institute, Philadelphia) for discussions and helpful suggestions. 22. Cameron, F. H., and Jennings, P. A. (1989) Proc. Natl. Acad. Sei. (I. S. A. 8 8 ,
9139-9143 REFERENCES
2. Liotta, L. A,, Steeg, P. S., and Stetler-Stevenson, W. G. (1991) Cell 64,327-336 1. Cross, M., and Dexter, T. M (1991) Cell 64, 271-280
3. Wellstein, A,, Fang, W. J., Khatri, A., Lu, Y., Swain, S. S., Dickson, R. B., Sasse,
4. Fang, W. J., Hartmann, N., Chow, D., Riegel, A. T., and Wellstein, A. (1992) J J., Riegel, A. T., and Lippman, M. E. (1992) J. B i d . Chem. 267,2582-2587
5. Milner, P. G., Li, Y. S., Hoffman, R. M., Kodner, C. M., Siegel, N. R., and Deuel, Biol. Chem. 267, 25889-25897
T. F. (1989) Biochem. Biophys. Res. Commun. 165,10961103 6. Li,Y. S., Milner, P. G., Chauhan, A. K., Watson, M. A,, Hoffman, R. M., Kodner,
7. Rauvala, H. (1989) EMBO J . 8,2933-2941 C. M., Milbrandt, J., and Deuel, T. F. (1990) Science 260, 1690-1694
9. Kovesdi, I., Fairhurst, J. L., Kretschmer, P. J., and Bohlen, P. (1990) Biochem. 8. Merenmies, J., and Rauvala, H. (1990) JBiol. Chem. 266, 16721-16724
10. Kuo, M. D., Oda, Y., Huang, J. S., and Huang, S. S. (1990) J . B id . Chem. 266,
11. Bloch, B., Normand, E., Kovesdi, I., and Bohlen, P. (1992) Brain Res. Deu.
12. Vanderwinden, J. M., Mailleux, P., Schihann, S. N., and Vanderhaeghen, J.
13. Raulo, E., Julkunen, I., Merenmies, J., Pihlaskari, R., and Rauvala, H. (1992)
14. Courty, J., Dauchel, M. C., Caruelle, D., Perderiset, M., and Barritault, D.
Biophys. Res. Commun. 172,850-854
18749-18752
Brain Res. 70,267-278
J. (1992) Anat. Embryol. 186, 387406
J. Bid . Chem. 267, 11408-11416
23. Symons, R. H. (1992)Annu. Reu. Biochem. 61,641-671 24. Pyle, A. M. (1993) Science 261, 709-714 25. lbmomura, M., Kadomatsu, K., Nakamoto, M., Muramatsu, H., Kondoh, H.,
Imagawa, K., and Muramatau, T. (1990) Biochem. Biophys. Res. Comrnun. 171,603-609
~~
26. Bohlen, P., and Kovesdi, I. (1991) Prog. Growth Factor. Res. 3, 143-157 27. Steinecke, P., Herget, T., and Schreier, P. H. (1992) EMBO J. 11, 1525-1530 28. Kretschmer, P. J., Fairhurst, J. L., Decker, M. M., Chan, C. P., Gluzman, Y.,
BBhlen, P., and Kovesdi, I. (1991) Growth Factors 5, 99-114 29. Wellstein, A., Lupu, R., Zugmaier, G., Flamm, S. L., Cheville, A. L., Delli Bovi,
P., Besilico, C., Lippman, M. E., and Kern, F. G. (1990) Cell Growth D i f m 1,63-71
30. Chen, C., and Okayama, H. (1987) Mol Cell. Biol 7,2745-2752 31. Milligan, J. F., and Uhlenbeck, 0. C. (1989) Methods Enzymol. 180, 5 1 4 2 32. Zuker, M., and Stiegler, P. (1981) Nucleic Acids. Res 9, 133-148 33. McCall, M. J., Hendry, P., and Jennings, P. A. (1992) Proc. Natl. Acad. Sci.
34. Sachs, A. B. (1993) Cell 74,413421 35. Tsutsui, J., Uehara, K., Kadomatsu, K., Matsubara, S., and Muramatsu, T.
37. Lai, S. P., Czubayko, F., Riegel, A. T., and Wellstein, A. (1992) Biochem. Bio- 36. L'Huillier, P. J., Davis, s. R., and Bellamy, A. R. (1992)EMBO J . 11,44114418
38. Sullenger, B. A,, and Cech, T. R. (1993) Science 262,1566-1569
U. S. A. 89,5710-5714
(1991) Biochem. Biophys. Res. Commun. 176,792-797
phys. Res. Commun. 187,1113-1122