rickettsia rickettsii-induced cellular injury of human vascular endothelium in vitro
TRANSCRIPT

INFECTION AND IMMUNITY, June 1984, p. 545-5530019-9567/84/060545-09$02.00/0Copyright © 1984, American Society for Microbiology
Vol. 44, No. 3
Rickettsia rickettsii-Induced Cellular Injury of Human VascularEndothelium In Vitro
DAVID J. SILVERMANDepartment of Microbiology, University of Maryland School of Medicine, Baltimore, Maryland 21201
Received 3 January 1984/Accepted 17 February 1984
The endothelial cell is the putative primary target cell in humans infected with Rickettsia rickettsii, theetiological agent of Rocky Mountain spotted fever. Although the clinical manifestations of infection by thisorganism are well documented, the mechanism of injury to the endothelial cell is not understood. The abilityto culture human endothelial cells in vitro provides a unique system with which to study this host-parasiteinteraction directly. Human vascular endothelial cells derived from the umbilical vein, when infected by R.rickettsii, became severely damaged within a few days postinfection. The primary lesion observed at theultrastructural level appeared to occur at intracellular membranes, specifically, the rough-surfacedendoplasmic reticulum. Widespread dilatation of these membranes eventually led to the creation of largeintracellular cisternae and the apparent circumscription of rickettsiae and cellular organelles by the rough-surfaced endoplasmic reticulum. Small membrane-bound fragments of host cytosol created by dilatingmembranes also were present within the cisternae. Within 5 to 6 days postinfection, cells lost theirosmoregulatory control and lysed. Some possible mechanisms of cell injury directed at the level ofintracellular membranes are discussed.
The infection, proliferation, and cellular destructioncaused by Rickettsia rickettsii, the etiological agent of RockyMountain spotted fever, have been documented in studiesutilizing numerous host cell types (4, 13, 16, 18-20, 25, 27).As a result of these in vitro studies, R. rickettsii was shownto exhibit specific properties which served to distinguish thisorganism, and possibly other members of the spotted fevergroup, from other species of the genus Rickettsia. Intranu-clear infection and replication and the ability of the organismto move bidirectionally through the plasma membrane, re-sulting in the rapid spread of the rickettsiae in cultured cells,are well-established properties of R. rickettsii which havebeen discovered through the use of light microscopy.
Rickettsiae-induced cellular destruction at the electron-microscopic level has also been studied with R. rickettsii andR. prowazekii. The cell damage elicited in chicken embryofibroblasts in response to infection by these two organisms isstrikingly different (18-20). R. prowazekii, because of itsapparent inability to escape from the host cell during initialphases of infection, remains confined to its intracellularenvironment, replicates to large numbers in the host cellcytoplasm, and, as a rule, is released only when the celllyses. R. rickettsii, on the other hand, with its capacity forbidirectional movemnent through the plasma membrane,spreads rapidly to adjacent cells, does not accumulate tolarge numbers within the infected host cell, and inducesearly cytopathic changes which eventually lead to cell lysisand death (18, 25).
Recent studies in my laboratory have focused on thecharacterization of the infection of human vascular endothe-lial cells by R. rickettsii, with the purpose of using thissystem as a possible in vitro model for studying the interac-tion of R. rickettsii with the putative target cell in humaninfections (16a). We have recently shown that the basicgrowth properties of R. rickettsii in human endothelial cellsare similar to those observed in the chicken embryo cellmodel system previously described by Wisseman et al. (25).There are a number of advantages in using human endo-
thelial cells as a model for infection by R. rickettsii: (i)545
endothelial cells are easily obtained from the human umbili-cal vein; (ii) they represent cells which are of the samespecies and ontogenic type as those which are thought to beprincipally parasitized in vivo; (iii) they retain many of theirin vivo properties in an in vitro culture system, e.g., synthe-sis of prostaglandins (22), expression of angiotensin convert-ing enzyme activity (8), synthesis of factor VIII-associatedprotein (9), and retention of the Weibel-Palade body, anendothelial cell-specific cytoplasmic organelle (5); and (iv)they provide the opportunity to examine the interactionbetween the infected target cell and other blood-derived cellssuch as granulocytes, macrophages, and platelets whichlikely modulate the course of infection in humans.
I report here electron-microscopic studies of the cellinjury induced by R. rickettsii in human endothelial cellswith the purpose of extending knowledge of this host-parasite interaction. I propose some possible mechanismsinvolved in cell injury and advocate the potential usefulnessof this in vitro system for studying the pathogenesis ofRocky Mountain spotted fever at the cellular level.
MATERIALS AND METHODS
Rickettsiae. The rickettsial seed used in these experimentswas a plaque-purified isolate of the Sheila Smith strain of R.rickettsii. The organism was propagated in the yolk sacs of 5-day-old embryonated chicken eggs and harvested and puri-fied on Renografin gradients as previously described (7).Quantitation of rickettsiae in the seed stock was carried outby the method of Silverman et al. (17), and infectivity wasdetermined by plaque titration in primary cultures of chickenembryo cells and umbilical vein endothelial cells by amodification of the method of Wike and Burgdorfer (23). Thepurified seed was found to have a rickettsial body count of2.2 x 109/ml and a titer of 1.1 x 109PFU/ml for a rickettsialbody/PFU ratio of 2.
Isolation and culture of human umbilical vein endothelialcells. Endothelial cells were isolated from human umbilicalveins by the methods of Jaffe et al. (9) and Gimbrone (5). The
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/i
ai o
n 17
Dec
embe
r 20
21 b
y 10
6.16
7.54
.236
.
546 SILVERMAN
; ' < < i>n'' WADb aRtSfrV~)'-
W 4 %., e
( <A A,4
4 -~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~4
45~~~~~~~~~~
-4~ ~ ~ ~ ~~ "
' ~~~~~'4, -S% 'RafT :X0
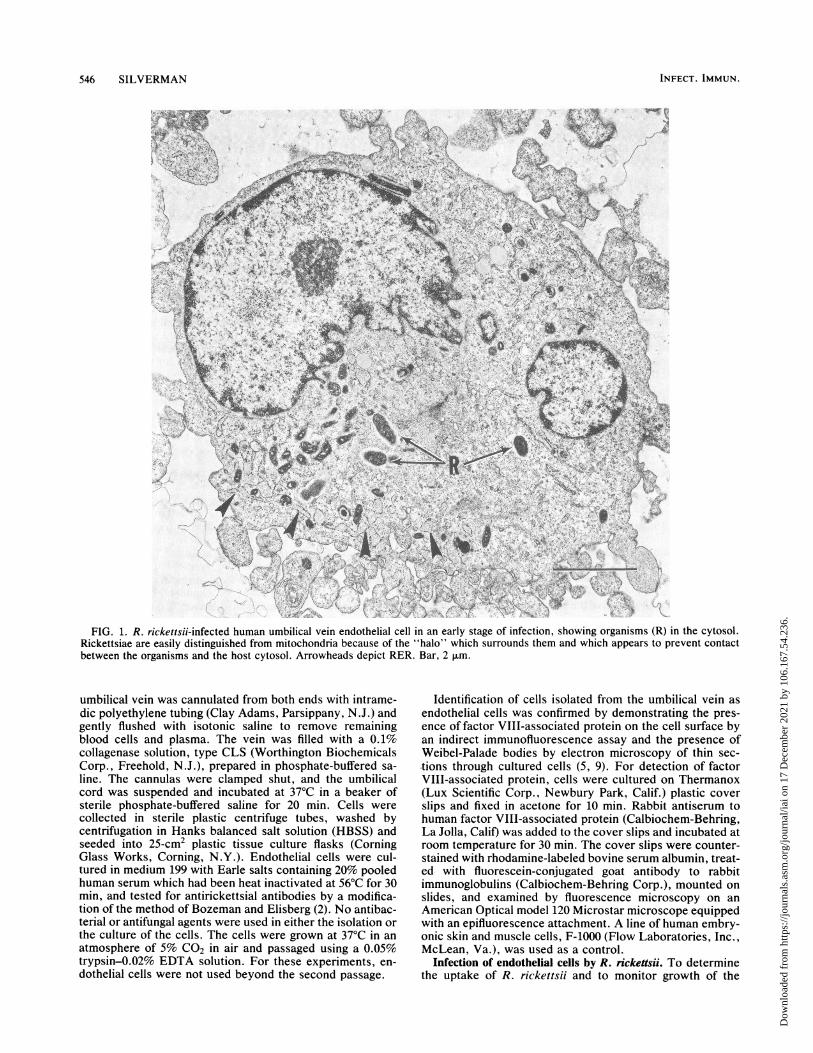
FIG. 1. R. rickettsii-infected human umbilical vein endothelial cell in an early stage of infection, showing organisms (R) in the cytosol.Rickettsiae are easily distinguished from mitochondria because of the "halo" which surrounds them and which appears to prevent contactbetween the organisms and the host cytosol. Arrowheads depict RER. Bar, 2 ,um.
umbilical vein was cannulated from both ends with intrame-dic polyethylene tubing (Clay Adams, Parsippany, N.J.) andgently flushed with isotonic saline to remove remainingblood cells and plasma. The vein was filled with a 0.1%collagenase solution, type CLS (Worthington BiochemicalsCorp., Freehold, N.J.), prepared in phosphate-buffered sa-line. The cannulas were clamped shut, and the umbilicalcord was suspended and incubated at 37°C in a beaker ofsterile phosphate-buffered saline for 20 min. Cells werecollected in sterile plastic centrifuge tubes, washed bycentrifugation in Hanks balanced salt solution (HBSS) andseeded into 25-cm2 plastic tissue culture flasks (CorningGlass Works, Corning, N.Y.). Endothelial cells were cul-tured in medium 199 with Earle salts containing 20% pooledhuman serum which had been heat inactivated at 56°C for 30min, and tested for antirickettsial antibodies by a modifica-tion of the method of Bozeman and Elisberg (2). No antibac-terial or antifungal agents were used in either the isolation orthe culture of the cells. The cells were grown at 37°C in anatmosphere of 5% CO2 in air and passaged using a 0.05%trypsin-0.02% EDTA solution. For these experiments, en-dothelial cells were not used beyond the second passage.
Identification of cells isolated from the umbilical vein asendothelial cells was confirmed by demonstrating the pres-ence of factor VIII-associated protein on the cell surface byan indirect immunofluorescence assay and the presence ofWeibel-Palade bodies by electron microscopy of thin sec-*tions through cultured cells (5, 9). For detection of factorVIII-associated protein, cells were cultured on Thermanox(Lux Scientific Corp., Newbury Park, Calif.) plastic coverslips and fixed in acetone for 10 min. Rabbit antiserum tohuman factor VIII-associated protein (Calbiochem-Behring,La Jolla, Calif) was added to the cover slips and incubated atroom temperature for 30 min. The cover slips were counter-stained with rhodamine-labeled bovine serum albumin, treat-ed with fluorescein-conjugated goat antibody to rabbitimmunoglobulins (Calbiochem-Behring Corp.), mounted onslides, and examined by fluorescence microscopy on anAmerican Optical model 120 Microstar microscope equippedwith an epifluorescence attachment. A line of human embry-onic skin and muscle cells, F-1000 (Flow Laboratories, Inc.,McLean, Va.), was used as a control.
Infection of endothelial cells by R. rickettsii. To determinethe uptake of R. rickettsii and to monitor growth of the
INFECT. IMMUN.
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/i
ai o
n 17
Dec
embe
r 20
21 b
y 10
6.16
7.54
.236
.
R. RICKETTSII AND HUMAN VASCULAR ENDOTHELIUM 547
t .-;~~~0 .,- ' ,
tR...o.-W e..4 ;
~ rRvR'; A,A*a~~~~~~~~~<
At ~~~~~~~~~~~~~~~~~~~~~~~~. !0
1.4~~~~~~4-
vi<.i/ .; .o- ' :'v4A
9 ,{, { ,0
,' X , , %d,s5 < < 4s u . a -, @ y , , f - u
0.
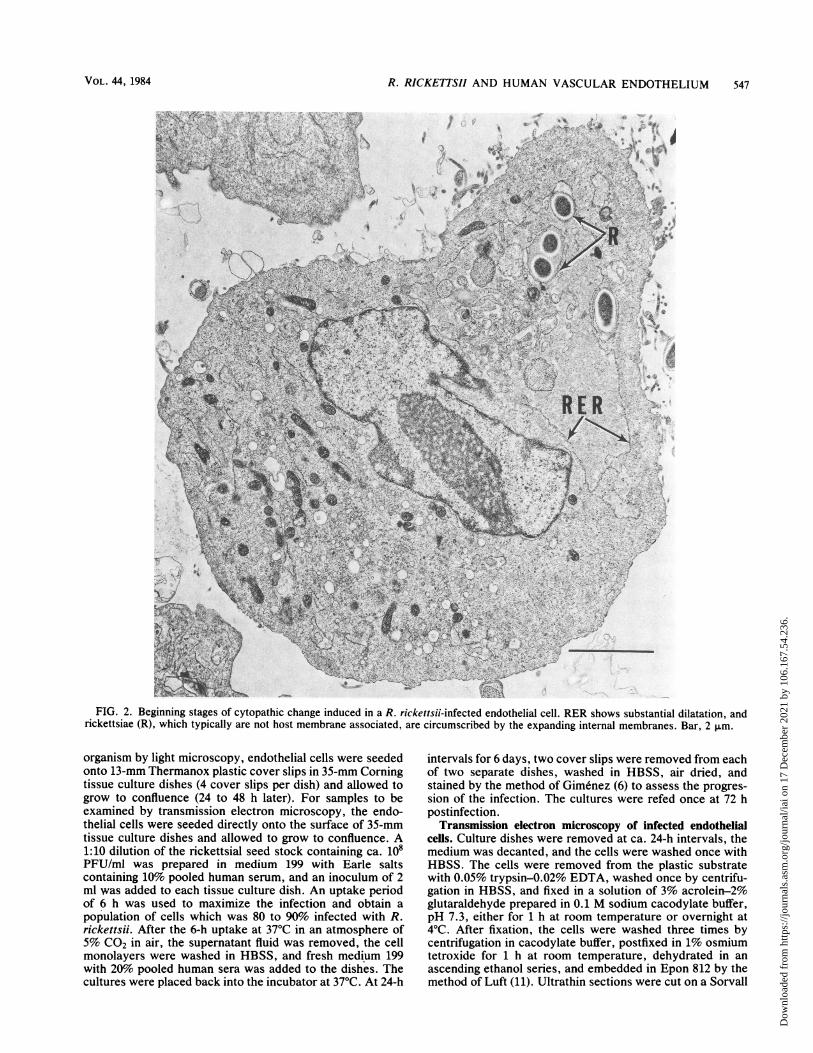
FIG. 2. Beginning stages of cytopathic change induced in a R. rickettsii-infected endothelial cell. RER shows substantial dilatation, andrickettsiae (R), which typically are not host membrane associated, are circumscribed by the expanding internal membranes. Bar, 2 ,.m.
organism by light microscopy, endothelial cells were seededonto 13-mm Thermanox plastic cover slips in 35-mm Comingtissue culture dishes (4 cover slips per dish) and allowed togrow to confluence (24 to 48 h later). For samples to beexamined by transmission electron microscopy, the endo-thelial cells were seeded directly onto the surface of 35-mmtissue culture dishes and allowed to grow to confluence. A1:10 dilution of the rickettsial seed stock containing ca. 108PFU/ml was prepared in medium 199 with Earle saltscontaining 10% pooled human serum, and an inoculum of 2ml was added to each tissue culture dish. An uptake periodof 6 h was used to maximize the infection and obtain apopulation of cells which was 80 to 90% infected with R.rickettsii. After the 6-h uptake at 37°C in an atmosphere of5% CO2 in air, the supernatant fluid was removed, the cellmonolayers were washed in HBSS, and fresh medium 199with 20% pooled human sera was added to the dishes. Thecultures were placed back into the incubator at 37°C. At 24-h
intervals for 6 days, two cover slips were removed from eachof two separate dishes, washed in HBSS, air dried, andstained by the method of Gimenez (6) to assess the progres-sion of the infection. The cultures were refed once at 72 hpostinfection.
Transmission electron microscopy of infected endothelialcells. Culture dishes were removed at ca. 24-h intervals, themedium was decanted, and the cells were washed once withHBSS. The cells were removed from the plastic substratewith 0.05% trypsin-0.02% EDTA, washed once by centrifu-gation in HBSS, and fixed in a solution of 3% acrolein-2%glutaraldehyde prepared in 0.1 M sodium cacodylate buffer,pH 7.3, either for 1 h at room temperature or overnight at4°C. After fixation, the cells were washed three times bycentrifugation in cacodylate buffer, postfixed in 1% osmiumtetroxide for 1 h at room temperature, dehydrated in anascending ethanol series, and embedded in Epon 812 by themethod of Luft (11). Ultrathin sections were cut on a Sorvall
VOL. 44, 1984
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/i
ai o
n 17
Dec
embe
r 20
21 b
y 10
6.16
7.54
.236
.
548 SILVERMAN
a
~~~~fr~~ ~0IP
V~~~~~~
iA~~~~~~~~~~~~i
14.1 4&_~~~~~~~~~~~~~~~~~~.w
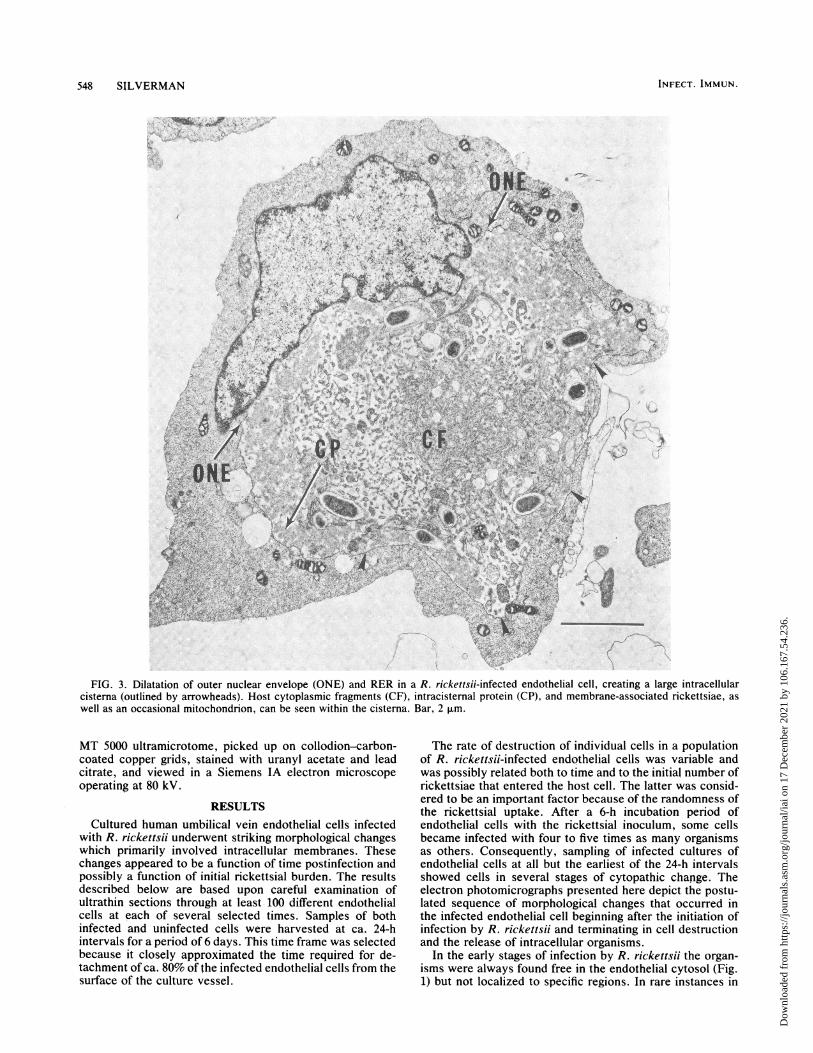
FIG. 3. Dilatation of outer nuclear envelope (ONE) and RER in a R. rickettsii-infected endothelial cell, creating a large intracellularcisterna (outlined by arrowheads). Host cytoplasmic fragments (CF), intracisternal protein (CP), and membrane-associated rickettsiae, aswell as an occasional mitochondrion, can be seen within the cisterna. Bar, 2 *m.
MT 5000 ultramicrotome, picked up on collodion-carbon-coated copper grids, stained with uranyl acetate and leadcitrate, and viewed in a Siemens IA electron microscopeoperating at 80 kV.
RESULTSCultured human umbilical vein endothelial cells infected
with R. rickettsii underwent striking morphological changeswhich primarily involved intracellular membranes. Thesechanges appeared to be a function of time postinfection andpossibly a function of initial rickettsial burden. The resultsdescribed below are based upon careful examination ofultrathin sections through at least 100 different endothelialcells at each of several selected times. Samples of bothinfected and uninfected cells were harvested at ca. 24-hintervals for a period of 6 days. This time frame was selectedbecause it closely approximated the time required for de-tachment of ca. 80% of the infected endothelial cells from thesurface of the culture vessel.
The rate of destruction of individual cells in a populationof R. rickettsii-infected endothelial cells was variable andwas possibly related both to time and to the initial number ofrickettsiae that entered the host cell. The latter was consid-ered to be an important factor because of the randomness ofthe rickettsial uptake. After a 6-h incubation period ofendothelial cells with the rickettsial inoculum, some cellsbecame infected with four to five times as many organismsas others. Consequently, sampling of infected cultures ofendothelial cells at all but the earliest of the 24-h intervalsshowed cells in several stages of cytopathic change. Theelectron photomicrographs presented here depict the postu-lated sequence of morphological changes that occurred inthe infected endothelial cell beginning after the initiation ofinfection by R. rickettsii and terminating in cell destructionand the release of intracellular organisms.
In the early stages of infection by R. rickettsii the organ-isms were always found free in the endothelial cytosol (Fig.1) but not localized to specific regions. In rare instances in
INFECT. IMMUN.
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/i
ai o
n 17
Dec
embe
r 20
21 b
y 10
6.16
7.54
.236
.

R. RICKETTSII AND HUMAN VASCULAR ENDOTHELIUM 549
.v~~~~~.f ~mw-I,<nAt~~~~~~~~w
~~4~ Mk.
4~~~~~~~~~~~~~~~~~~~~~4
~~~~~~~i~~ ~ ~ ~ V
;'2 ' '3%!'' '4 > 3 aMr e,~~~~- ' s '
;~~ ~ ~ ~ ~~. Sf;0 ,V,Ss . o
I,'s:;r 5 v 8 Xx +g,
1'.
..0
.1 .riL1
I
FIG. 4. Advanced stage of RER dilation in a R. Rickettsii-infected endothelial cell. Note the reduced electron opacity in the cisterna,resulting from the loss of fine granular intracisternal protein. Physically intact mitochondria (MI) appear normal. CF, Cytoplasmic fragments.Bar, 2 ,.m.
which intranuclear organisms were observed, they appearedto be either free in the cell nucleoplasm or within invagina-tions of the nuclear envelope. The rickettsiae, whether in thecytoplasm or the nucleus, always were surrounded by thecharacteristic electron-lucent zone of up to 200 nm whichfacilitated their identification at low magnification andserved to distinguish the organisms from mitochondria andlysosomes, which occasionally appeared similar in size andshape. At 24 h postinfection, rickettsiae were difficult to findby electron microscopy, and no evidence of cell damage wasdetected. However, evidence of rickettsiae-induced cyto-pathic change, although modest in most cells, could bedemonstrated in the form of dilatation of the rough-surfacedendoplasmic reticulum (RER) at 48 h postinfection. Theswollen mitochondrial cristae and increased electron opacityof the mitochondrial matrices, which could be demonstratedthroughout these studies in endothelial cells, probably re-flect hyperactivity of the mitochondria. This observation
was made in both infected and uninfected cells and thus doesnot constitute a rickettsiae-induced cellular change.The dilatation of the RER progressively increased in both
dimension and frequency with respect to time postinfection.Cytoplasmic rickettsiae which were located in regions of thedilating RER became circumscribed by these membranes(Fig. 2), whereas others in areas where RER dilatation wasnot apparent remained free in the host cytosol. Because ofthe continuity that exists between the outer nuclear envelopeand the RER, it was frequently possible to demonstratedistention and dissociation of the outer nuclear envelopefrom its intrinsic association with the nucleus (Fig. 3).Distention of the RER and outer nuclear envelope created alarge intracellular cisterna that often encompassed two-thirds or more of the thin-sectioned surface area of theaffected cells. The following were found within these dilatedcisternae: (i) rickettsiae, and occasionally mitochondriabounded by ribosome-studded membranes of RER origin;
VOL. 44, 1984
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/i
ai o
n 17
Dec
embe
r 20
21 b
y 10
6.16
7.54
.236
.
Xitw, * *4.. 4 )
4.j~~~AW
D>'is; (s X .!w t,
<S S iX,,R 9JA
IF,~~~~~~~~~~~~~~~~~~~~
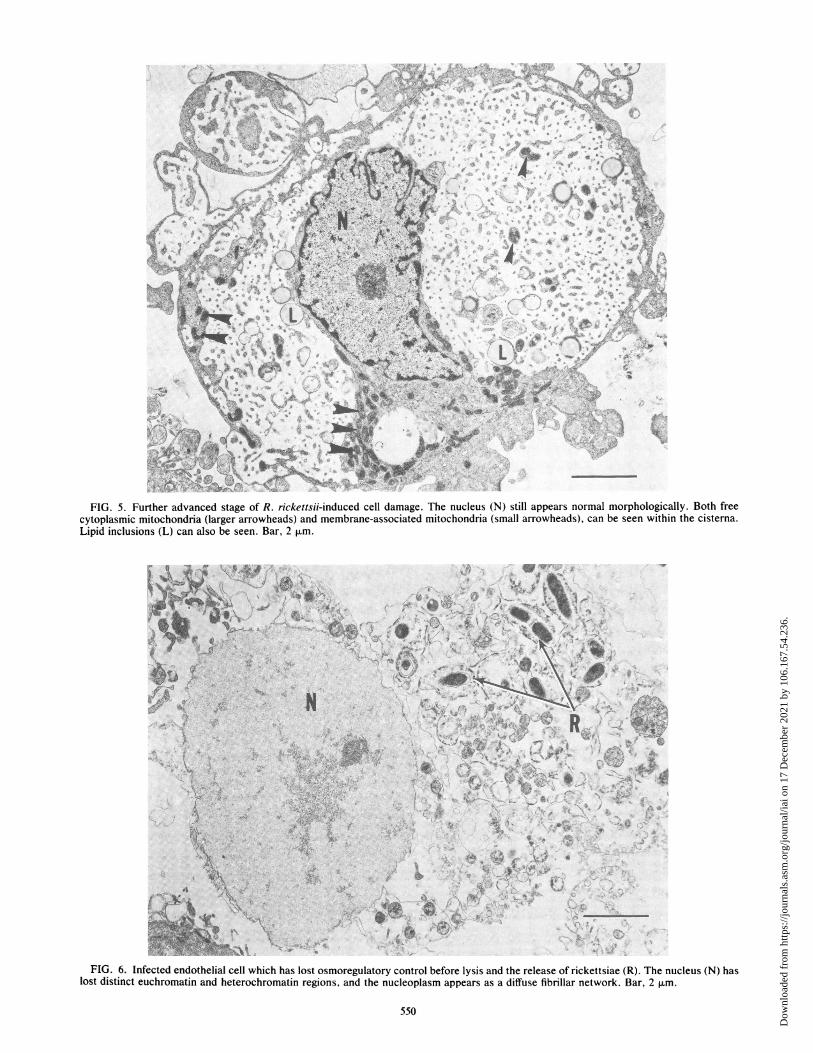
FIG. 5. Further advanced stage of R. rickettsii-induced cell damage. The nucleus (N) still appears normal morphologically. Both freecytoplasmic mitochondria (larger arrowheads) and membrane-associated mitochondria (small arrowheads), can be seen within the cisterna.Lipid inclusions (L) can also be seen. Bar, 2 p.m.
Fi'{9S ¾-*-- 4j-An..
:
k......
.:
.. ... ... ... .. . . .*:: rv .*:
;4i
.. ..
"
w R -.
.eZ wsRz i , @
^ * s ^=;. i .X .f
... <,
.}.fs
\
y.w '.
w $2,d
; B<;,,,
t- t.e
.s *;.
Axr-Jt1
4t X,, "4t'S,,;
4 5,
* N<ez$;-V
e - .....
FIG. 6. Infected endothelial cell which has lost osmoregulatory control before lysis and the release of rickettsiae (R). The nucleus (N) haslost distinct euchromatin and heterochromatin regions, and the nucleoplasm appears as a diffuse fibrillar network. Bar, 2 p.m.
550
P;X -
ftt
*.fIo
AL r W..: w "-" ''.
O.;,1-'. IV7
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/i
ai o
n 17
Dec
embe
r 20
21 b
y 10
6.16
7.54
.236
.
R. RICKETTSII AND HUMAN VASCULAR ENDOTHELIUM 551
I.rmi 1.
:. Woi.
s
x p
40- .1f. i.
ce,.4
,':.v, ; ,,0
-'9.
.4
4*
4.*
w,1,~ ~ ~ ~ ~ 1
.,e.~~~~~~~~~~~~~~~~~~~~~~~~O*~~~~~i'~~.'-9. ~ q
AML~~~~~~~~-t
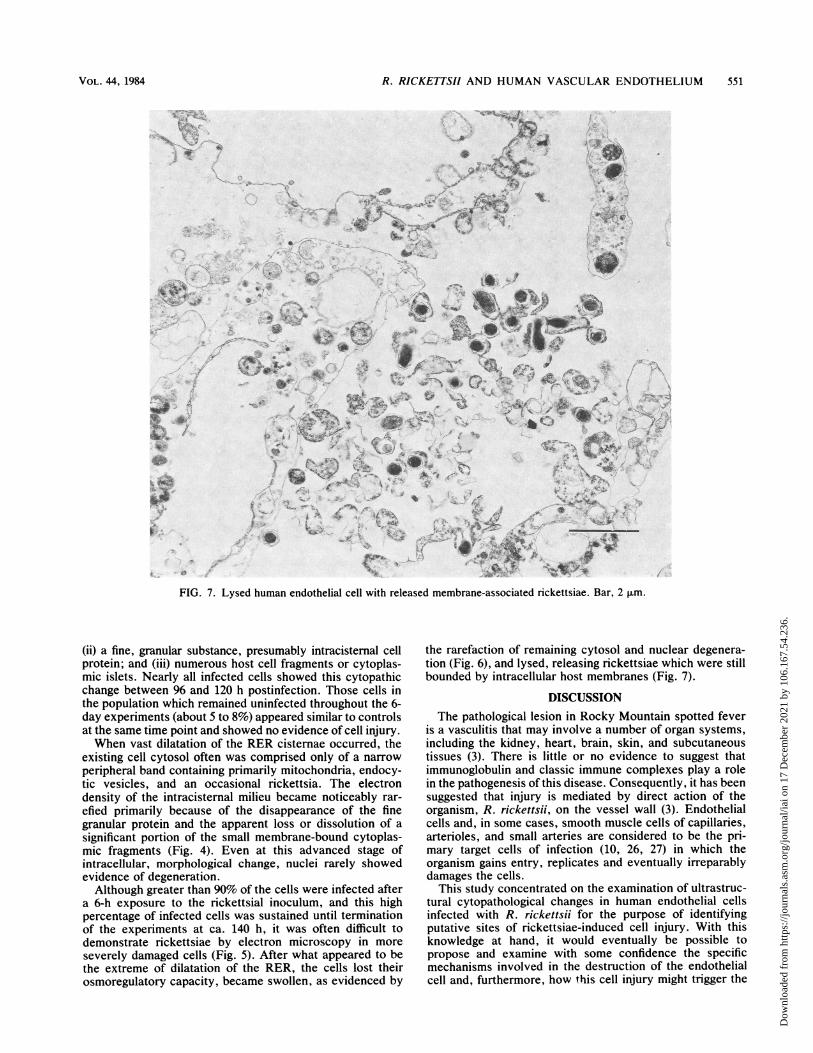
FIG. 7. Lysed human endothelial cell with released membrane-associated rickettsiae. Bar, 2 p.m.
(ii) a fine, granular substance, presumably intracisternal cellprotein; and (iii) numerous host cell fragments or cytoplas-mic islets. Nearly all infected cells showed this cytopathicchange between 96 and 120 h postinfection. Those cells inthe population which remained uninfected throughout the 6-day experiments (about 5 to 8%) appeared similar to controlsat the same time point and showed no evidence of cell injury.When vast dilatation of the RER cisternae occurred, the
existing cell cytosol often was comprised only of a narrowperipheral band containing primarily mitochondria, endocy-tic vesicles, and an occasional rickettsia. The electrondensity of the intracisternal milieu became noticeably rar-
efied primarily because of the disappearance of the finegranular protein and the apparent loss or dissolution of a
significant portion of the small membrane-bound cytoplas-mic fragments (Fig. 4). Even at this advanced stage ofintracellular, morphological change, nuclei rarely showedevidence of degeneration.Although greater than 90% of the cells were infected after
a 6-h exposure to the rickettsial inoculum, and this highpercentage of infected cells was sustained until terminationof the experiments at ca. 140 h, it was often difficult todemonstrate rickettsiae by electron microscopy in more
severely damaged cells (Fig. 5). After what appeared to bethe extreme of dilatation of the RER, the cells lost theirosmoregulatory capacity, became swollen, as evidenced by
the rarefaction of remaining cytosol and nuclear degenera-tion (Fig. 6), and lysed, releasing rickettsiae which were stillbounded by intracellular host membranes (Fig. 7).
DISCUSSIONThe pathological lesion in Rocky Mountain spotted fever
is a vasculitis that may involve a number of organ systems,including the kidney, heart, brain, skin, and subcutaneoustissues (3). There is little or no evidence to suggest thatimmunoglobulin and classic immune complexes play a rolein the pathogenesis of this disease. Consequently, it has beensuggested that injury is mediated by direct action of theorganism, R. rickettsii, on the vessel wall (3). Endothelialcells and, in some cases, smooth muscle cells of capillaries,arterioles, and small arteries are considered to be the pri-mary target cells of infection (10, 26, 27) in which theorganism gains entry, replicates and eventually irreparablydamages the cells.
This study concentrated on the examination of ultrastruc-tural cytopathological changes in human endothelial cellsinfected with R. rickettsii for the purpose of identifyingputative sites of rickettsiae-induced cell injury. With thisknowledge at hand, it would eventually be possible topropose and examine with some confidence the specificmechanisms involved in the destruction of the endothelialcell and, furthermore, how this cell injury might trigger the
x
".,-: l..
I
.;&I.4V
.iIP-
.P :
t.
1..
VOL. 44, 1984
% .0 )'
A 4
10-0 1 t-A.-lj ',If ....' 01"',
-c -';.11..
iIC_
I
.Ao .'Ito 10
-,". .,
Z .: ;1.4t
.;,
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/i
ai o
n 17
Dec
embe
r 20
21 b
y 10
6.16
7.54
.236
.

552 SILVERMAN
cascade of pathological sequelae usually manifested in hu-mans with severe clinical disease (1, 10, 12, 21).The morphologically detectable injury sustained by cul-
tured human vascular endothelial cells infected in vitro withR. rickettsii is dramatic; it occurs rapidly (within 2 to 3 dayspostinfection) and ultimately leads to cell death. The mostconspicuous cytopathic lesion involved intracellular mem-branes, specifically the RER and outer nuclear envelope,which became highly dilated and created a large intracellularcisterna. Involvement of other cellular organelles could notbe detected. The apparent selectivity of intracellular mem-branes as the primary sites of rickettsial-induced cell injurywas not without precedent. A similar phenomenon in chick-en embryo cells infected with R. rickettsii has been observed(18).The apparent site of the cell injury (intracellular mem-
branes) initiated by R. rickettsii suggests a number ofpotentially destructive effector mechanisms acting directlyon the membrane. Winkler and Miller (24) have proposed arole for phospholipase A in the entry and escape of R.prowazekii in susceptible host cells. This hypothesis wasbased upon the detection of hydrolytic products of mem-brane glycerophospholipids released from cells exposed toand lysed by the organism. This observation demonstratesthat rickettsiae can exert a direct membranolytic action oncells. It must be kept in mind, however, that R. prowazekiibehaves very differently from R. rickettsii in many of itsproperties (18-20, 25), and therefore extrapolation of mecha-nisms of rickettsia-host cell interaction from R. prowazekiito R. rickettsii cannot be made without verification byproper experimentation. Nevertheless, direct action by rick-ettsiae on cellular membranes has some precedent.Another very attractive hypothesis which could explain
the origin of intracellular membrane damage is lipid peroxi-dation. Generation of free radicals as byproducts of cellularmetabolism may lead to the peroxidation of lipids of intracel-lular membranes (14, 15). Peroxidation of lipids damagesmembranes by causing them to become leaky. Increasedmembrane permeability would result in an influx of waterand sodium ions through the damaged membranes, causingdilatation of the intracisternal space. Studies are planned totest the hypothesis that membrane lipid peroxidation is apotential mechanism for the direct injury of endothelial cellsinfected with R. rickettsii. Further studies are also needed toconfirm a possible role for phospholipase in rickettsiae-induced cellular injury.Although cells of nonhuman origin have been used very
effectively to examine specific parameters of rickettsia-hostcell interaction, the human endothelial cell is now alsoavailable for study. The endothelial cell adds the importantdimension of being similar structurally and functionally tothe putative target cell in human infections. The electron-microscopic study reported here, together with the previousreport on the growth kinetics of infection of human endothe-lial cells with R. rickettsii (16a), confirms the availability andcapability of the endothelial cell to serve as an appropriate invitro model system for further study of the pathogenesis ofrickettsial diseases.
ACKNOWLEDGMENTSThis work was supported by Public Health Service grant AI-17416
from the National Institute of Allergy and Infectious Diseases.I thank Elaine Cockroft for her excellent technical assistance,
Barbara A. Hanson and Charles L. Wisseman, Jr., for their helpfuldiscussions and critical review, and Kathleen Montroy and SueWyche for typing the manuscript.
LITERATURE CITED
1. Atkin, M. D., H. S. Strauss, and G. U. Fisher. 1965. A casereport of "Cape Cod" Rocky Mountain spotted fever withmultiple coagulation disturbance. Pediatrics 36:627-632.
2. Bozeman, F. M., and B. L. Elisberg. 1963. Serological diagnosisof scrub typhus by indirect immunofluorescence. Proc. Soc.Exp. Biol. Med. 112:568-573.
3. Bradford, W. D., B. P. Croker, and C. C. Tisher. 1979. Kidneylesions in Rocky Mountain spotted fever-a light-, immunofluo-rescence-, and electron microscopic study. Am. J. Pathol.97:381-392.
4. Burgdorfer, W., R. L. Anacker, R. G. Bird, and D. S. Bertram.1968. Intranuclear growth of Rickettsia rickettsii. J. Bacteriol.96:1415-1418.
5. Gimbrone, M. A. 1976. Culture of vascular endothelium. Prog.Hemostasis Thomb. 3:2745-2756.
6. Gimenez, D. F. 1964. Staining rickettsiae in yolk-sac cultures.Stain Technol. 39:135-140.
7. Hanson, B. A., C. L. Wisseman, Jr., A. Waddell, and D. J.Silverman. 1981. Some characteristics of heavy and light bandsof Rickettsia prowazekii on Renografin gradients. Infect. Im-mun. 34:596-604.
8. Hial, V., M. A. Gimbrone, M. P. Peyton, G. M. Wilcox, and J. J.Pisano. 1979. Angiotensin metabolism by cultured human vascu-lar endothelium and smooth muscle cells. Microvasc. Res.17:314-329.
9. Jaffe, E. A., L. W. Hoyer, and R. L. Nachman. 1973. Synthesisof antihemophilic factor antigen by cultured human endothelialcells. J. Clin. Invest. 52:2757-2764.
10. Lillie, R. D. 1941. The pathology of Rocky Mountain spottedfever. Natl. Inst. Health Bull. 177:1-46.
11. Luft, J. H. 1961. Improvements in epoxy resin embeddingmethods. J. Biophys. Biochem. Cytol. 9:409-414.
12. Mengel, C. E., and C. Trygstad. 1963. Thrombocytopenia inRocky Mountain spotted fever. J. Am. Med. Assoc. 183:886-887.
13. Pinkerton, H., and G. M. Hass. 1932. Spotted fever. I. Intranu-clear rickettsiae in spotted fever studied in tissue culture. J.Exp. Med. 56:151-156.
14. Recknagel, R. O., and E. A. Glende, Jr. 1977. Lipid peroxida-tion: a specific form of cellular injury, p. 591-601. In D. H. K.Lee, H. L. Falk, S. D. Murphy, and S. R. Geiger (ed.),Handbook of physiology. American Physiological Society, Be-thesda, Md.
15. Scarpelli, D. G., and B. F. Trump. 1971. Cell injury, p. 9-65.The Upjohn Co., Kalamazoo, Mich.
16. Schaechter, M., F. M. Bozeman, and J. E. Smadel. 1957. Studyon the growth of rickettsiae. II. Morphologic observations ofliving rickettsiae in tissue culture cells. Virology 3:160-172.
16a.Silverman, D. J., and S. B. Bond. 1984. Infection of humanvascular endothelial cells by Rickettsia rickettsii. J. Infect. Dis.149:201-206.
17. Silverman, D. J., P. Fiset, and C. L. Wisseman, Jr. 1979. Simple,differential staining technique for enumerating rickettsiae inyolk sac, tissue culture extracts, or purified suspensions. J.Clin. Microbiol. 9:437-440.
18. Silverman, D. J., and C. L. Wisseman, Jr. 1979. In vitro studiesof rickettsia-host cell interactions: ultrastructural changes in-duced by Rickettsia rickettsii infection of chicken embryofibroblasts. Infect. Immun. 26:714-727.
19. Silverman, D. J., C. L. Wisseman, Jr., and A. Waddell. 1980. Invitro studies of rickettsia-host cell interactions: ultrastructuralstudy of Rickettsia prowazekii-infected chicken embryo fibro-blasts. Infect. Immun. 29:778-790.
20. Silverman, D. J., C. L. Wisseman, Jr., and A. Waddell. 1981.Envelopment and escape of Rickettsia rickettsii from hostmembranes, p. 241-253. In W. Burgdorfer and R. L. Anacker(ed.), Rickettsiae and rickettsial diseases. Academic Press, Inc.,New York.
21. Trigg, J. W., Jr. 1964. Hypofibrinogenemia in Rocky Mountainspotted fever. N. Engl. J. Med. 270:1042-1044.
22. Weksler, B. B., A. J. Marcus, and E. A. Jaffe. 1977. Synthesis of
INFECT. IMMUN.
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/i
ai o
n 17
Dec
embe
r 20
21 b
y 10
6.16
7.54
.236
.

R. RICKETTSII AND HUMAN VASCULAR ENDOTHELIUM 553
prostaglandin 12 (prostacyclin) by cultured human and bovineendothelial cells. Proc. Natl. Acad. Sci. U.S.A. 74:3922-3926.
23. Wike, D. A., and W. Burgdorfer. 1972. Plaque formation intissue cultures by Rickettsia rickettsii isolated directly fromwhole blood and tick hemolymph. Infect. Immun. 6:736-738.
24. Winkler, H. H., and E. T. Miller. 1982. Phospholipase A and theinteraction of Rickettsia prowazekii and mouse fibroblasts (L-929 cells ). Infect. Immun. 38:109-113.
25. Wisseman, C. L., Jr., E. A. Edlinger, A. D. Waddell, and M. R.Jones. 1976. Infection cycle of Rickettsia rickettsii in chicken
embryo and L-929 cells in culture. Infect. Immun. 14:1052-1064.
26. Wolbach, S. B. 1948. The pathology of the rickettsial diseases ofman, p. 118-125. In F. R. Moulton (ed.), Rickettsial diseases ofman. American Association for the Advancement of Science,Washington, D.C.
27. Wolbach, S. B., and M. J. Schlesinger. 1923. The cultivation ofthe microorganisms of Rocky Mountain spotted fever (Dermo-centroxenus rickettsi) and of typhus (Rickettsia prowazeki) intissue plasma cultures. J. Med. Res. 44:231-256.
VOL. 44, 1984
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/i
ai o
n 17
Dec
embe
r 20
21 b
y 10
6.16
7.54
.236
.