rufino, fabiana de almeida. avaliação de um controle ... rufino.pdf · o sexo feminino,...
TRANSCRIPT

1
RUFINO, Fabiana de Almeida. Avaliação de um controle interno em sistema de multiplex-PCR para a determinação do sexo de embriões bovinos produzidos in vitro. Londrina: UEL, 2005. (Dissertação de Mestrado em Ciência Animal)
RESUMO
A utilização de um controle interno da reação em sistema de multiplex-PCR para a determinação do sexo de embriões bovinos produzidos in vitro foi avaliada em biópsias realizadas em dois grupos de embriões, constituídos por 54 embriões frescos e 44 congelados. Como controle interno foram utilizados os primers BOV 1 e BOV 2 que amplificam um produto com 626 pb do gene ND5 do DNA mitocondrial de bovinos. Os primers BRY.4aF e BRY.4aR (REED et al., 1989) foram utilizados para amplificação de seqüências encontradas no cromossomo Y bovino. A multiplex-PCR apresentou 100% de especificidade na determinação do sexo de 10 embriões machos e 10 embriões fêmeas. Tanto o produto do controle interno quanto o do cromossomo Y foram amplificados até a diluição correspondente a 6,25% do DNA total extraído de um embrião macho. Foi possível a determinação do sexo de 53 (98,1%) embriões frescos e de 40 (90,9%) embriões congelados. Em 34 (63%) embriões frescos e em 27 (61,4%) embriões congelados foram amplificados, simultaneamente, os produtos referentes ao cromossomo Y e ao DNA mitocondrial, caracterizando dessa forma o embrião como macho. O sexo feminino, caracterizado pela amplificação apenas do produto referente ao controle interno, foi detectado em 19 (35,2%) e em 13 (29,5%) embriões frescos e congelados, respectivamente. Em 1 (1,8%) e em 4 (9,1%) biópsias realizadas, respectivamente, em embriões frescos e congelados, nenhum produto foi amplificado provavelmente devido à ausência de células embrionárias. A falta de produto de amplificação em sistemas de PCR na ausência de controle interno caracteriza o embrião como fêmea, gerando resultados errôneos. A multiplex-PCR padronizada nesse trabalho caracterizou-se como sendo de fácil e rápida realização, além de ser sensível, específica e eficiente na identificação de falhas na reação apresentando, com isso, potencial para ser utilizada em escala comercial, na rotina de determinação do sexo de embriões bovinos produzidos in vitro.
Palavras-chave: bovinos, embriões, produção in vitro, sexagem, multiplex-PCR

2
RUFINO, Fabiana de Almeida. Evaluation of an internal control in multiplex-PCR system for sex determination of in vitro produced bovine embryos. Londrina: UEL, 2005. (Msc Dissertation)
ABSTRACT
The use of an internal control in a multiplex-PCR assay for sex determination of in vitro produced bovine embryos was evaluated in biopsies of two groups, constituted by 54 fresh and 44 frozen embryos. It was used as internal control the primers BOV 1 and BOV 2 that amplify a product with 626 bp of bovine mitochondrial DNA ND5 gene. The primers BRY.4a F and BRY.4a R (REED et al., 1989) were used for bovine Y chromosome sequences amplification. The multiplex-PCR presented 100% specificity in sex determination of 10 male and 10 female embryos. Both amplicons of internal control and Y chromosome were amplified until the dilution corresponding to 6.25% of total extracted DNA from a male embryo. Sex determination was possible in 53 (98.1%) fresh embryos and 40 (90.9%) frozen embryos. The products related to Y chromosome and mitochondrial DNA were simultaneously amplified in 34 (63%) fresh embryos and 27 (61.4%) frozen embryos, showing a male embryo. The female sex distinguished by internal control amplification only, was detected in 19 (35.2%) and 13 (29.5%) biopsies, respectively, of fresh and frozen embryos. In 1 (1.8%) and 4 (9.1%) biopsies of fresh and frozen embryos, respectively, neither product was amplified probably due to the absence of embryonic cells. The lack of product amplification in PCR assay in the absence of an internal control can result in a misdiagnosed female embryo, leading to false result. The multiplex-PCR assay developed in this work showed easy and fast application, beside it was sensitive, specific and efficient in reaction failures identification, presenting a great potential to be used in commercial scale in routine sex determination of in vitro produced bovine embryos.
Keywords: bovine, embryos, in vitro production, sexing, multiplex-PCR

3
1. REVISÃO DE LITERATURA

4
1. Produção in vitro de embriões bovinos
Avanços significativos na reprodução animal assistida há algumas décadas
vêm apresentando como principais vantagens, a utilização e a disseminação de
animais geneticamente superiores (VAN WAGTENDONK-DE LEEW et al., 2000).
O desenvolvimento de biotécnicas da reprodução como a produção in vitro (PIV)
de embriões bovinos otimizou o aproveitamento de oócitos, contribuindo para o
incremento na produção de embriões. Com isso, obteve-se aumento no potencial
de exploração do sistema reprodutivo das fêmeas, com grande acréscimo no
número de descendentes (BLONDIN et al., 2002).
A produção in vitro (PIV) de embriões é uma biotécnica para programas de
melhoramento genético animal, devido ao grande potencial e rapidez para a
multiplicação de descendentes de animais geneticamente superiores, quando
comparada a outras biotécnicas (CAMARGO et al., 2003). Embora essa técnica
venha sendo estudada desde 1878 em coelhos e em outros animais de
laboratório, estudos mais específicos na espécie bovina somente ocorreram no
início dos anos 60 (GORDON, 1994). Brackett et al. (1982) relataram o
nascimento do primeiro bezerro obtido pela PIV a partir de oócitos ovulados, e Lu
et al. (1988) descreveram a obtenção de gestação a partir de um embrião
produzido por um processo totalmente in vitro.
Apesar do sucesso da técnica, problemas têm sido descritos, como
distocias, abortamentos e gestações prolongadas. Relatam-se também diversos
problemas com fetos e/ou bezerros provenientes de embriões produzidos in vitro
como maior peso ao nascimento (“síndrome do bezerro grande”), malformações

5
congênitas e elevada taxa de mortalidade neonatal (HASLER et al., 1995; VAN
WAGTENDONK-DE LEEW et al., 1998; VAN WAGTENDONK-DE LEEW et al.,
2000).
Vários estudos têm também demonstrado desvio na proporção macho-
fêmea de embriões produzidos in vitro, com a porcentagem de embriões machos
superior à proporção teoricamente esperada de 50% para cada um dos sexos
(GUTIERREZ-ADÁN et al., 1996; VAN WAGTENDONK-DE LEEW et al., 2000;
GUTIERREZ-ADÁN et al., 2001).
A velocidade de desenvolvimento dos embriões de acordo com o sexo
parece influenciar a desproporção entre esses. Supõe-se que a seleção de
blastocistos em estádios mais avançados do desenvolvimento favorece a escolha
de machos por esses se desenvolverem mais rapidamente em comparação aos
embriões do sexo feminino durante os oito primeiros dias após a fertilização in
vitro (FIV) (AVERY et al., 1991). Esse desenvolvimento precoce poderia ocorrer
devido às condições de cultivo dos embriões. O uso de soro fetal bovino no
cultivo, com alta concentração de glicose, parece acelerar o desenvolvimento dos
machos e retardar o desenvolvimento das fêmeas (GUTIERREZ-ADÁN et al.,
2001). Dentre os fatores relacionados com a diferença na proporção entre os
sexos, pesquisas têm sugerido também o método de preparação dos
espermatozóides na FIV (RHEINGANTZ, 2000). Portanto, a seleção dos embriões
no momento da transferência para as receptoras poderia implicar em maior
número de produtos do sexo masculino (CAMARGO et al., 2003).
A maior incidência de machos nos produtos oriundos da PIV é altamente
indesejável. O impacto de um único touro sobre um plantel é bastante grande e

6
isso exige extremo rigor na seleção de touros para a colheita de sêmen. Como a
produção de gametas é constante, o número de machos com alto potencial
zootécnico necessário em um rebanho é bem menor em relação às fêmeas. Como
a probabilidade de um bezerro tornar-se doador de sêmen é baixa, esses
aspectos justificam a desvalorização dos produtos do sexo masculino. Em relação
às fêmeas, toda bezerra oriunda de um acasalamento bem planejado é,
potencialmente, uma futura doadora. Isto implica em menor risco quanto ao
investimento nos animais desse sexo, justificando assim a maior valorização dos
produtos do sexo feminino.
A falta de receptoras em número e qualidade suficientes, aliada à
ineficiência atual da criopreservação dos embriões produzidos in vitro, também
são fatores relacionados com a desproporção entre os sexos. Praticamente todos
os embriões produzidos in vitro são transferidos a fresco. Assim, na inexistência
de receptoras para todos os embriões, é comum o descarte daqueles menos
adiantados quanto ao desenvolvimento. Segundo a hipótese de que embriões
fêmeas apresentam tendência ao desenvolvimento mais lentamente, poder-se-ia
confundir pior qualidade do embrião com desenvolvimento atrasado e,
conseqüentemente, descarte justamente dos embriões mais valorizados.
2. Identificação do sexo de embriões bovinos pré-implantação
A diferenciação sexual nos bovinos, assim como nos outros mamíferos,
começa na fecundação com a diferenciação do conteúdo cromossômico. O
embrião do sexo feminino possuirá dois cromossomos X enquanto o embrião de

7
um macho possuirá um cromossomo X e um cromossomo Y (HASELTINE e
OHNO, 1981).
A possibilidade de seleção do sexo, previamente à transferência dos
embriões, apresenta grande interesse econômico principalmente devido à
multiplicação de animais do sexo desejado (VAN VLIET et al., 1989). Com o
advento da transferência comercial de embriões nos anos 70, a determinação
seletiva do sexo dos animais antes da transferência tem sido uma meta almejada
pela indústria animal (HERR e REED, 1991; THIBIER e NIBART, 1995). No
entanto, somente no início dos anos 90, a determinação do sexo de embriões
bovinos pré-transferência foi introduzida como uma biotécnica comercialmente
significativa (BREDBACKA, 2001).
Vários métodos têm sido utilizados para a identificação do sexo dos
embriões. De acordo com o procedimento de secção, perfuração e penetração da
zona pelúcida (ZP), esses métodos podem ser classificados como não-invasivos e
invasivos (MC EVOY, 1992).
2.1. Métodos não-invasivos
Os métodos não-invasivos são considerados ideais, pois mantém a
integridade do embrião, reduzindo os prejuízos na transferência e posterior
implantação. Além disso, deve-se considerar também o baixo custo e a facilidade
de interpretação dos resultados (VAN VLIET et al., 1989).

8
2.1.1. Métodos imunológicos
Os métodos imunológicos são utilizados na identificação do sexo de
embriões pela detecção de um antígeno de superfície celular Y-específico
(antígeno H-Y), cuja expressão é controlada por um gene presente no
cromossomo Y dos mamíferos (HASELTINE e OHNO, 1981). O antígeno H-Y é
um antígeno de histocompatibilidade, identificado pela primeira vez em
camundongos do sexo feminino, após rejeição de enxertos de pele provenientes
de machos da mesma linhagem sangüínea (EICHWALD e SILMSER, 1955). No
entanto, embora o termo “antígeno H-Y” tenha sido originalmente aplicado a um
antígeno de rejeição, o uso desse termo freqüentemente inclui antígenos
encontrados somente em células de machos e que induzem a formação de
anticorpos no soro (HASELTINE e OHNO, 1981).
A citotoxicidade e a imunofluorescência indireta (IFI) são dois métodos
utilizados para a detecção do antígeno H-Y nos embriões (VAN VLIET et al.,
1989). Em ensaios citotóxicos, os embriões que expressam esse antígeno
apresentam lise celular ou cessam o desenvolvimento, sendo classificados como
machos (WATCHEL, 1984). White et al. (1982) identificaram o sexo de 1000
embriões murinos de 8-16 células por meio da citotoxicidade. Após a transferência
dos embriões viáveis, 86% (50/58) dos animais nascidos eram fêmeas. No
entanto, o inconveniente nesse tipo de ensaio é a destruição dos embriões
machos.
No método de IFI são utilizados anticorpos mono ou policlonais anti-H-Y
conjugados com o isotiocianato de fluoresceína (FITC). As células portadoras

9
desse antígeno podem ser visualizadas por meio de microscopia de
imunofluorescência (WATCHEL, 1984). Booman et al. (1989) realizaram a IFI
utilizando anticorpos monoclonais. Embora a identificação dos machos e das
fêmeas tenha sido possível, a avaliação da fluorescência foi subjetiva devido aos
sítios de ligação inespecíficos dos anticorpos com as células dos embriões,
alcançando apenas 77% de acurácia. Uma vantagem encontrada nessa técnica,
em contraposição à citotoxicidade é a preservação dos embriões machos.
Embora os métodos de detecção do antígeno H-Y sejam simples e rápidos
a acurácia é insatisfatória (BREDBACKA, 1998). Esses métodos também não têm
demonstrado repetibilidade e possuem reduzida aplicação para a determinação do
sexo realizada com propósitos comerciais (VEERHUIS et al., 1994).
2.1.2. Métodos bioquímicos
Devido à presença de dois cromossomos X nas células das fêmeas de
mamíferos e para manter o número equivalente de genes entre os sexos no início
do estádio embrionário, um dos cromossomos X da fêmea é inativado em cada
célula (LYON, 1972). Embora não se tenha certeza do momento exato da
inativação do cromossomo X, estudos têm sugerido um breve período entre a
ativação do genoma embrionário e a inativação de um dos cromossomos X
(MONK e HARPER, 1978). Por este motivo, no início do desenvolvimento, a
concentração celular e a atividade de certas enzimas ligadas ao cromossomo X
podem ser duas vezes mais altas nos embriões fêmeas que nos machos. Com
isso, a mensuração da concentração celular e/ou da atividade das enzimas ligadas

10
ao cromossomo X indica o dobro da atividade quando comparada com as enzimas
autossômicas. Essa maior atividade pode ser utilizada para a identificação dos
embriões do sexo feminino. Com base nessa hipótese, Williams (1986) identificou
o sexo de embriões murinos por meio da atividade da enzima glicose-6-fosfato
desidrogenase (G6PD) e obteve eficiência de 64%. Monk e Handyside (1988)
mensuraram a atividade da fosforribosil hipoxantina transferase (FHT), outra
enzima ligada ao cromossomo X e obtiveram 91% de acurácia para fêmeas e
100% para os machos.
O principal fator que inviabiliza a aplicação comercial desses métodos que
se baseiam na determinação da concentração celular e/ou da atividade das
enzimas ligadas ao cromossomo X é a alta taxa de mortalidade embrionária,
sugerindo toxicidade da reação para os embriões (WILLIAMS, 1986).
Adicionalmente, essa técnica exige o conhecimento exato do período de
inativação do cromossomo X bem como a idade exata dos embriões no momento
da análise (RIEGER, 1984), pois as enzimas monitoradas podem resultar da
tradução do RNA mensageiro materno estocado no embrião e não da transcrição
do genoma embrionário, levando a resultados errôneos (MC EVOY, 1992).
2.2. Métodos invasivos
Os métodos mais precisos para a determinação do sexo são invasivos, isto
é, requerem a realização de uma biópsia de células dos embriões. A aplicação dos
métodos invasivos torna possível atingir taxas de até 100% na determinação do
sexo. No entanto, além da dificuldade na manipulação durante o processo de

11
biópsia devido à característica invasiva, esses métodos podem acarretar redução
nos índices de gestações (BREDBACKA, 1998).
2.2.1. Métodos citogenéticos
As análises citogenéticas implicam na visualização da cromatina sexual
(cromossomo X inativado ou corpúsculo de Barr) ou na identificação dos
cromossomos sexuais pela técnica de cariotipagem (BONDIOLI et al., 1989)
2.2.1.1. Identificação do sexo de embriões pela visualização da cromatina
sexual
A análise citogenética para a identificação de embriões do sexo feminino
por meio da visualização da cromatina sexual, realizada em células de embriões
de coelhos com cinco dias de desenvolvimento, foi a primeira técnica utilizada
para a identificação do sexo (EDWARDS e GARDNER, 1967). No entanto, em
outras espécies animais, a técnica não obteve o mesmo sucesso devido à
dificuldade de visualização da cromatina sexual causada pela natureza granular
do citoplasma das células embrionárias (KING, 1984).
2.2.1.2. Identificação do sexo de embriões pela técnica de cariotipagem
Posteriormente, biópsias embrionárias foram submetidas à outra técnica
citogenética para a identificação do par de cromossomos sexuais XX ou XY na

12
fase de metáfase. A identificação do sexo por cariotipagem de células
provenientes de biópsia é realizada por meio da utilização de substâncias
mitostáticas que atuam paralisando o ciclo celular para posterior análise de placas
metafásicas (KING, 1984). Hare et al. (1976) demonstraram a possibilidade da
determinação do sexo em biópsias de embriões bovinos com 14 e 15 dias de
desenvolvimento. No entanto, a necessidade da utilização de blastocistos mais
adiantados, eclodidos e elongados, inviabiliza o uso da técnica em programas
comerciais. Singh e Hare (1980) demonstraram que a média do índice mitótico foi
de apenas 9% quando mórulas de seis dias de desenvolvimento foram
cariotipadas.
Embora a acurácia dessa técnica seja alta, sua eficiência, avaliada pela
proporção de embriões que podem ter o sexo determinado, permanece em níveis
insatisfatórios (BREDBACKA, 1998). Na maioria das vezes, somente poucas
células estão em divisão e pode haver dificuldade na obtenção de células em
metáfases bem expandidas que possibilitem a visualização dos cromossomos
sexuais. Devido também ao grande número de células necessárias para a análise,
a viabilidade dos embriões é seriamente comprometida (SETIABUDI e
GUSTAVSSON, 1991). Como desvantagem ainda, a cariotipagem é um
procedimento demorado e que exige grande habilidade para a sua realização.
Atualmente, a não ser como método de referência para a verificação da eficiência
de outros métodos de identificação do sexo, a cariotipagem não tem sido utilizada
rotineiramente (VAN VLIET et al., 1989).

13
2.2.2. Métodos moleculares
Os estudos do mapeamento do cromossomo Y proporcionaram o
desenvolvimento de sondas de DNA específicas para a determinação do sexo de
embriões pré-transferência (SETIABUDI e GUSTAVSSON, 1991). As sondas Y-
específicas são utilizadas para a marcação de seqüências do cromossomo Y por
meio das técnicas de hibridização in situ, hibridização in situ fluorescente (FISH) e
dot blot.
2.2.2.1. Identificação do sexo de embriões pela técnica de hibridização in situ
A utilização de técnicas de hibridização in situ permite a localização de
seqüências Y-específicas por meio de marcação radioativa. A hibridização positiva
indica a presença do cromossomo Y e permite identificar os embriões machos
(VAN VLIET et al., 1989). Cotinot et al. (1991) desenvolveram e utilizaram uma
sonda Y-específica (BC1.2) para a identificação do sexo em 150 biópsias (10-20
células) de embriões por meio da hibridização in situ, obtendo resultado em 57%
das biópsias. A acurácia da técnica, confirmada por análise citogenética, foi de
95%. Setiabudi e Gustavsson (1991) também utilizaram a sonda BC1.2 com
marcação radioativa e revelação por auto-radiografia, e obtiveram 100% de
acurácia, confirmada por avaliação direta dos cromossomos. Embora a acurácia
dessa técnica seja considerada aceitável, a análise requer longos períodos para
as etapas de hibridização (12 a 16h) e de auto-radiografia (1 a 6 dias)
(SETIABUDI e GUSTAVSSON, 1991). Com isso, é necessário o congelamento ou

14
o abortamento seletivo, com redução da viabilidade comercial da identificação do
sexo de embriões bovinos produzidos in vitro.
2.2.2.2. Identificação do sexo de embriões pela técnica de FISH
Uma variação da técnica de hibridização in situ é a FISH, que utiliza
marcadores de sondas não-radioativas. Em comparação com a hibridização in
situ, o uso de sondas fluorescentes não-radioativas facilita a execução da técnica
e possibilita maior sensibilidade (GAVIO et al., 2001). Embora a aplicação esteja
difundida para o uso em embriões humanos, existe pouca informação sobre a
determinação do sexo em embriões bovinos (SHEA, 1999). Gavio et al. (2001)
analisaram 92 embriões produzidos in vitro por meio de remoção da ZP e posterior
desagregação dos blastômeros. A partir da fixação de um ou dois blastômeros, a
microscopia de fluorescência revelou 79 núcleos em interfase, com 42 (53,2%)
núcleos identificados como machos e 37 (46,8%) núcleos com sinais negativos
caracterizados como fêmeas. Lee et al. (2004) utilizaram a seqüência BtY2
(GenBank accesion nº A18483) em blastômeros individuais e em hemi-embriões
produzidos in vitro correspondentes a cada blastômero e obtiveram taxa de
coincidência de 96% na determinação do sexo.
Embora a técnica de FISH alcance resultados satisfatórios, existe pouca
informação sobre a eficiência da determinação do sexo em células embrionárias
provenientes de biópsias (KOBAYASHI et al., 1998). Devido às inúmeras etapas
requeridas, a aplicação dessa técnica na rotina é praticamente inviável
(KIRSZENBAUM et al., 1990).

15
2.2.2.3. Identificação do sexo de embriões pela técnica de dot blot
Bondioli et al. (1989) realizaram o primeiro ensaio em larga escala para
testar a acurácia da determinação do sexo pela técnica de dot blot. Por meio de
biópsias de 101 embriões bovinos de seis e sete dias de desenvolvimento,
congelados/descongelados e transferidos, 91 (90%) embriões tiveram o sexo
determinado. Esses embriões resultaram em 35 (40%) gestações, com 97% de
acurácia do teste. Os resultados desse estudo experimental indicaram que
elevada acurácia do ensaio e taxa de sobrevivência dos embriões após o
procedimento de biópsia, e mostrou-se aceitável para a aplicação comercial da
técnica. Em procedimento subseqüente, utilizado para a transferência comercial
de embriões bovinos, os autores alcançaram acurácia de 100% na identificação do
sexo com 45 (41%) gestações produzidas. Apesar dos resultados satisfatórios,
alguns fatores como os demorados protocolos de hibridização e a complexidade
do ensaio, assim como o uso de isótopos radioativos, inviabilizam o uso comercial
da técnica, principalmente em embriões produzidos in vitro (HASLER et al., 2002).
2.2.2.4. Identificação do sexo de embriões pela Reação em Cadeia da
Polimerase (PCR)
A identificação do sexo de embriões por meio de métodos moleculares foi
simplificada com o advento da PCR. O primeiro estudo para a determinação do
sexo de embriões por meio da utilização da PCR foi realizado por Handyside et al.
(1989) com embriões humanos. Em bovinos, um passo marcante no

16
desenvolvimento da técnica foi a descrição da determinação do sexo pela PCR a
partir de biópsias de embriões com seis ou sete dias de desenvolvimento
(SCHRÖDER et al., 1990).
A PCR amplifica exponencialmente pequenos fragmentos de DNA de
regiões Y-específicas. A presença ou ausência de amplificação da seqüência Y-
específica indica o sexo do embrião (HERR e REED, 1991). A técnica caracteriza-
se pelo alto grau de eficiência e acurácia (BREDBACKA et al., 1995; HASLER,
2003), além de ser considerada um procedimento rápido e de fácil realização
(SCHRÖDER et al., 1990; MACHÁTY et al., 1993; SHEA, 1999).
Para a determinação do sexo pela PCR podem ser utilizados embriões em
vários estádios de desenvolvimento, desde mórula compacta até blastocisto
expandido, e de qualidade excelente a boa, pois esse quesito é um fator crítico
sobre os resultados (HASLER et al., 2002). Como método invasivo, a principal
desvantagem dessa técnica é a necessidade de remoção de um pequeno número
de células embrionárias (BREDBACKA, 1998).
A biópsia é o componente mais crítico e laborioso do procedimento para a
determinação do sexo de embriões pela PCR e o seu tamanho é um compromisso
entre a eficiência da técnica e a viabilidade do embrião (BREDBACKA, 2001). A
orientação da biópsia não tem relevância em uma mórula compacta. Entretanto,
em blastocistos, a massa celular interna deve permanecer intacta e as amostras
devem ser retiradas do trofoblasto (BREDBACKA, 1998). O tamanho da biópsia
pode variar entre um único blastômero (MACHÁTY et al., 1993; GUTIERREZ-
ADÁN et al., 1996) até aproximadamente 10-30% da massa celular embrionária
(BREDBACKA, 1998). A quantidade de células deve ser compatível com alta

17
eficiência na identificação do sexo e com taxas de gestações aceitáveis
(BREDBACKA, 1998).
No entanto, embriões produzidos in vitro são mais frágeis à
micromanipulação e mais sensíveis à criopreservação sendo esses os principais
obstáculos para o uso difundido da técnica na PIV (BREDBACKA et al., 1994;
HASLER et al., 1995). Devido a esse inconveniente e por ser um procedimento
rápido, a PCR pode ser realizada em laboratórios de PIV de embriões bovinos,
proporcionando resultados em até seis horas, sem a necessidade de
criopreservação (LUZ et al., 2000).
Os protocolos realizados comercialmente para a determinação do sexo de
embriões geralmente envolvem a utilização de uma seqüência Y-específica em co-
amplificação com uma seqüência autossômica bovina (controle interno da reação)
caracterizando, dessa forma, a multiplex-PCR. Eficiência de 90-95% e acurácia de
93-98% foram relatadas por meio da utilização desse sistema (THIBIER e
NIBART, 1995; SHEA,1999).
A inclusão de uma seqüência autossômica é muito importante, pois detecta
tanto o DNA de células de embriões masculino quanto feminino, funcionando
como controle para assegurar a presença de material coletado na biópsia e
também para detectar problemas na amplificação. Com a utilização da seqüência
autossômica são evitados resultados errôneos (“falso-fêmeas”), passíveis de
ocorrer quando se utiliza apenas a amplificação de fragmentos Y-específicos
(PEURA et al., 1991; MACHÁTY et al., 1993).
Por meio da utilização da seqüência autossômica na reação obtém-se a
presença de duas bandas visíveis após eletroforese em gel de agarose e/ou

18
acrilamida, correspondentes aos produtos amplificados, referentes às seqüências
do cromossomo Y e autossômica, específicas para a espécie bovina. Dessa forma
os embriões machos são identificados, enquanto a ausência da banda Y-
específica caracteriza um embrião do sexo feminino (BREDBACKA, 1998).
Macháty et al. (1993) identificaram o sexo de 87 embriões produzidos in
vitro, cinco dias pós-fertilização, por meio da multiplex-PCR, utilizando apenas um
blastômero removido no estádio de 16-32 células. Do total de embriões, foi
possível a determinação do sexo em 83 deles, conferindo à técnica 95,4% de
eficiência.
Alta acurácia foi relatada por Bousquet et al. (1999) na determinação do
sexo pela multiplex-PCR, de embriões de graus 1 e 2, produzidos in vitro e
biopsiados. A acurácia da técnica, confirmada por meio de avaliação ultra-
sonográfica do sexo fetal entre 60 e 65 dias de gestação, foi de 97,4% com taxa
de gestação de 53,4% (319/597). A taxa de identificação dos sexos diferiu
estatisticamente, com 56,8% (1047/1844) de machos e 37,5% (691/1844) de
fêmeas.
Costa et al. (2002) identificaram o sexo de embriões produzidos in vitro nos
estádios de mórula e blastocisto. Do total de 50 embriões, 48 tiveram o sexo
determinado pela multiplex-PCR resultando em 96% de eficiência (4% de não-
amplificação).
Camargo et al. (2003) analisaram 109 embriões inteiros, produzidos in vitro,
na fase de blastocisto, por meio da multiplex-PCR. Foi possível determinar o sexo
de 99 embriões demonstrando eficiência de 90,8%. Nesse experimento, que visou
a investigação da diferença entre os sexos, os embriões machos representaram

19
59,6% do total de embriões possíveis de terem o sexo determinado. Luz et al.
(2000) também determinaram o sexo de embriões inteiros produzidos in vitro
utilizando a seqüência macho–específica BC1.2 (COTINOT et al., 1991) e a
seqüência satélite 1.715 (PLUCIENNICZAK et al., 1982) para o controle interno da
reação. Do total de 92 blastocistos, 86 (93,5%) embriões tiveram o sexo
determinado pela multiplex-PCR.
Alguns estudos relataram como desnecessária a utilização do controle
autossômico, pois esses pareceram interferir na amplificação da seqüência Y-
específica (PEURA et al., 1991; BREDBACKA et al., 1994). No entanto, a
eficiência da determinação do sexo de embriões produzidos in vitro encontrada
por Lopes et al. (1999) foi de 90% (34/38) quando nenhuma seqüência
autossômica foi empregada e 94% (48/51) com a utilização das seqüências Y-
específica e autossômica. No mesmo estudo, a acurácia da técnica, relacionada
com o diagnóstico correto do sexo dos animais nascidos foi, respectivamente, de
79% (15/19) e de 100% (30/30) com os dois tipos de reação. Garcia (2001), com o
objetivo de tornar a PCR mais simples, identificou o sexo de 133 embriões
utilizando apenas a seqüência Y-específica. Para a avaliação do desempenho da
técnica foi aceita a margem de erro de 14% para as possíveis fêmeas e de 7%
para os machos. A confirmação por ultra-sonografia, ou mesmo após o
nascimento dos animais, revelou que a taxa correta de embriões machos obtida foi
de 93,7% (74/79) e para os embriões classificados como “possíveis” fêmeas foi
de 75,9% (41/54).

20
3. Considerações finais
A seleção do sexo anteriormente à transferência dos embriões bovinos tem
gerado interesse tanto por parte de pesquisadores quanto de criadores que
buscam aprimorar a qualidade de seus rebanhos.
Nos últimos anos, os grandes avanços observados em biotécnicas da
reprodução como a PIV, possibilitaram o incremento no desenvolvimento de
métodos para a identificação do sexo de embriões. Uma metodologia de rotina,
que tenha por objetivo a determinação do sexo de embriões deve incluir altas
taxas de eficiência e de acurácia, e ser seguida por um processo eficiente na
transferência desses embriões, além da obtenção de altas taxas de gestações.
Visando a aceitação comercial, a técnica de identificação do sexo deve ser
rápida, de fácil realização e de custo aceitável. Nesse contexto, a PCR possui
muitas vantagens sobre as demais técnicas, possibilitando assim sua utilização
para a determinação do sexo de embriões bovinos em escala comercial.

21
4. REFERÊNCIAS
AVERY, B.; MADISON, V.; GREVE, T. Sex and development in bovine in vitro embryo production. Theriogenology, v.35, p.953-963, 1991. BLONDIN, P.; BOUSQUET, D.; TWAGIRAMUNGU, H.; BARNES, F.; SIRARD, M.A. Manipulation of follicular development to produce developmentally competent bovine oocytes. Biology of Reproduction, v.66, p.38-43, 2002. BONDIOLI, K.R.; ELLIS, S.B.; PRYOR, J.H.; WILLIAMS, M.W.; HARPOLD, M.M. The use of male-specific chromosomal DNA fragments to determine the sex of bovine preimplantation embryos. Theriogenology, v.31, n.1, p. 95-103, 1989. BOOMAN, P.; KRUIJT, L.; VERRHUIS, R.; HENGST, A.M.; TIEMAN, M.; RUCH, F.E. Sexing bovine embryos with monoclonal antibodies against the H-Y antigen. Livestock Production Science, v.23, p.1-16, 1989. BOUSQUET, D.; TWAGIRAMUNGU, H.; MORIN, N.; BRISSON, C.; CARBONNEAU, G.; DUROCHER, J. In vitro embryo production in the cow: an effective alternative to the conventional embryo production approach. Theriogenology, v.51, p.59-70, 1999. BRACKETT, R.G.; BOUSQUET, D.; BOICE, M.L.; DONAWICK, W.J.; EVANS, J.F.; DRESSEL, M.A. Normal development following in vitro fertilization in the cow. Biology of Reproduction, v.27, p.147-158, 1982 BREDBACKA, P.; VELMALA, R.; PEIPPO, J.; BREDBACKA, K. Survival of biopsied and sexed bovine demi-embryos. Theriogenology, v.41, p.1023-1031, 1994. BREDBACKA, P; KANKAANPÄÄ, A.; PEIPO, J. PCR-sexing of bovine embryos: a simplified protocol. Theriogenology, v.44, p.167-176, 1995. BREDBACKA, P. Recent development in embryos sexing and its field application. Reproduction Nutrition Development, v.38, p.605-613, 1998. BREDBACKA, P. Progress on methods of gene detection in preimplantation embryos. Theriogenology, v.55, p.23-34, 2001. CAMARGO, L.S.A.; SÁ, W.F.; VIANA, J.H.M.; FERREIRA, A.M.; SERAPIÃO, R.V.; RAMOS, A.A.; MACHADO, M.A.; VALE FILHO, V.R.; ANDRADE, V.J. Identificação do sexo de embriões bovinos fecundados in vitro e cultivados com células do cumulus na presença de soro. Revista Brasileira de Reprodução Animal, v.27, n.3, p.407-409, 2003.

22
COSTA, L.L.; SILVA, J.C.; DINIZ, P.; CIDADÃO, R. Preliminary report on sexing bovine pre-implantation embryos under the conditions of Portugal. Revista Portuguesa de Ciências Veterinárias, v.97, p.95-98, 2002. COTINOT, C.; KIRSZEMBAUM, M.; LEONARD, M.; GIANQUINTO, L.; VAIMAN, M. Isolation of bovine Y-derived sequence: potential use in embryo sexing. Genomics, v.10, p.646-653, 1991. EDWARDS, R.G.; GARDNER, R.L. Sexing of live rabbit blastocysts. Nature, v.214, p.576-577, 1967. EICHWALD, E.J.; SILMSER, C.R. Untitled publication. Transplantation Bulletin, v.2, p.148-149, 1955. GAVIO, D.; FERNANDES, M.B.; VILLA, R.A.; GALERANI, M.A.; MARTELLI, L.R.; LÔBO, R.B. Aplicação do método de FISH para determinação do sexo em embriões zebuínos produzidos in vitro. Revista Brasileira de Reprodução Animal, v.25, n.3, p.404-407, 2001. GARCIA, J.F. Practical considerations of embryo manipulation: preimplantation genetic typing. Theriogenology, v.56, p.1393-1399, 2001. GORDON, I. Laboratory production of cattle embryo. CAB International, ed., University Press, Cambridge, 640p, 1994. GUTIÉRREZ-ADÁN, A.; BEHBOODI, E.; ANDERSON, G.B.; MEDRANO, J.F.; MURRIA, J.D. Relationship between stage of development and sex of bovine IVM-IVF embryos cultured in vitro versus in the sheep oviduct. Theriogenology, v.46, p.515-525, 1996. GUTIÉRREZ-ADÁN, A.; LONERGAN, P.; RIZOS, D.; WARD, F.A.; BOLAND, M.P.; PINTADO, B.; FUENTE, J. Effect of the in vitro culture system on the kinetics of blastocyst development and sex ratio of bovine embryos. Theriogenology, v.55, p.1117-1126, 2001. HANDYSIDE, A.H.; PENKETH, R.M.L.; WINSTON, J.K.; PATTINSON, J.D.A.; DELHANTY, J.D.; TUDDENHAM, E.G.D. Biopsy of human pre-implantation embryos and sexing by DNA amplification. The Lancet, v.333, n.8634, p.347-349, 1989. HARE, W.C.D.; MITCHELL, D.; BETTERIDGE, K.J.; EAGLESOME, M.D.; RANDALL, G.C.B. Sexing 2-week-old bovine embryos by chromosomal analysis prior to surgical transfer: preliminary methods and results. Theriogenology, v.5, p.243-253, 1976. HASELTINE, F.P.; OHNO, S. Mechanisms of gonadal differentiation. Science, v.211, p.12721278, 1981.

23
HASLER, J.F.; HENDERSON, W.B.; HURTGEN, P.J.; JIN, Z.Q.; MC CAULEY, A.D.; MOWER, S.A.; NEELY, B.; SHUEY, L.S.; STOKES, J.E.; TRIMMER, S.A. Production, freezing and transfer of bovine IVF embryos and subsequent calving results. Theriogenology, v.43, p.141-152, 1995. HASLER, J.F.; CARDEY, E.; STOKES, J.E.; BREDBACKA, P. Nonelectrophoretic PCR-sexing of bovine embryos in a commercial environment. Theriogenology, v.58, 1457-1469, 2002. HASLER, J.F. The current status and future of commercial embryo transfer in cattle. Theriogenology, v.79, p.245-264, 2003. HERR, C.M.; REED, K.C. Micromanipulation of bovine embryos for sex determination. Theriogenology, v.35, p.45-54, 1991. KING, W.A. Sexing embryos by cytological methods. Theriogenology, v.21, p.7-17, 1984. KIRSZENBAUM, M.; COTINOT, C.; LEONARD, M.; VAIMAN, M.; FELLOUS, M. Diagnosis of the sex of bovine embryos using molecular biology. Reproduction Nutrition Development, v.1, p.125S-132S, 1990 (supplement). KOBAYASHI, J.; SEKIMOTO, A.; UCHIDA, H.; WADA, T.; SASAKI, K.; SASADA, H.; UMEZU M, SATO E. Rapid detection of male-specific DNA sequence in bovine embryos using fluorescence in situ hibridization. Molecular Reproduction and Development, v.51, p.390-394, 1998. LEE, J.H.; PARK, J.H.; LEE, S.H.; PARK, C.S.; JIN, D. Sexing using single blastomere derived from IVF bovine embryos by fluorescence in situ hibridization (FISH). Theriogenology, v.62, p.1452-1458, 2004. LOPES, R.F.F; SENNA, J.P.M.; CHIES, J.M.; RODRIGUES, J.L. Pit-stop PCR: an approach to increase final product yield of multiplex PCR. Biotechniques, v.26, p.638-639, 1999. LU, K.H.; GORDON, I.; CHEN, H.B. Birth of twins after transfer of cattle embryos produced by in vitro techniques. Veterinary Records, v.122, p.540-593, 1988. LUZ, M.R.; WATANABE, Y.F.; FERRO, J.A.; FERRO, M.I.T.; MAURO, S.M.S.; HOSSEPIAN DE LIMA, V.F.M.; FRANCESCHINI, P.H. Identificação do sexo de embriões bovinos fecundados in vitro pela técnica de PCR multiplex. Brazilian Journal of Veterinary Research and Animal Science, v.37, n.6, p.453-456, 2000. LYON, M.F. X-chromosome inactivation and developmental patterns in mammals. Biological Reviews, v.47, p.1-35, 1972.

24
MACHÁTY, Z.; PÁLDY, A.; CSÁKI, T; VARGA, Z.; KISS, I.; BÁRÁNDI, Z.; VAJTA, G. Biopsy and sex determination by PCR of IVF bovine embryos. Journal of Reproduction and Fertility, v.98, p.467-470, 1993. MC EVOY, J.D. Alteration of sex ratio. Animal Breeding Abstracts, v.60, n.2, p.97-111, 1992. MONK, M.; HARPER, M. X-chromosome activity in preimplantation mouse embryos from XX and X0 mothers. Journal of Embryology and Experimental Morphology., v.46, p.53-64, 1978. MONK, M.; HANDYSIDE, A.H. Sexing of preimplantation mouse embryos by measurement of X-linked gene dosage in a single blastomere. Journal of Reproduction and Fertility, v.82, p.365-368, 1988. PEURA , T.; HYTTINEN, J.M.; TURUNEN, M.; JÄNNE, J. A reliable sex determination assay for bovine preimplantation embryos using the polimerase chain reaction. Theriogenology, v.35, p.547-555, 1991. PLUCIENNICZAK, A.; SKOWRONSKI, J.; JAWORSKI, J. Nucleotide sequence of bovine 1.715 satellite DNA and its relation to other bovine satellite sequences. Journal of Molecular Biology, v.158, p.293-304, 1982. RHEINGANTZ, M.G.T. Efeito do método de preparação dos espermatozóides e das condições de cultivo sobre a proporção macho:fêmea dos embriões bovinos produzidos in vitro. Pelotas: UFPel, 2000. Tese (Doutorado) - Programa de Pós-graduação em Biotecnologia, Faculdade de Medicina Veterinária, Universidade Federal de Pelotas, Pelotas, 2000. RIEGER, D. The measurement of metabolic activity as an approach to evaluating viability and diagnosing sex in early embryos. Theriogenology, v.21, p.138-149, 1984. SCHRÖDER, A.; MILLER, J.R.; THOMSENP.D.; ROSCHLAUD, K.; AVERY, B.; POULSEN, P.H.; SCHMIDT, M.; SCHWERIN, M. Sex determination of bovine embryos using the polymerase chain reaction. Animal Biotechnology, v.1, n.1-2, p.121-133, 1990. SETIABUDI, R.; GUSTAVSSON, I. Establishment of embryo sexing techniques in Sweden. Reproduction of Domestic Animals, v.26, p.78-81, 1991. SHEA, B.F. Determining the sex of bovine embryos using the polimerase chain reaction results: a six-year retrospective study. Theriogenology, v.51, p.841-854, 1999. SINGH, E.L.; HARE, W.C.D. The feasibility of sexing bovine morula stage embryos prior to embryo transfer. Theriogenology, v.14, p.421-427, 1980.

25
THIBIER,M.; NIBART, M. The sexing of bovine embryos in the field. Theriogenology, v.43, p.71-80, 1995. VAN VLIET, R.A.; GIBBINS, A.M.V.; WALTON, J.S. Livestock embryo sexing: a review of current methods, with emphasis on Y-specific DNA probes. Theriogenology, v.32, n.3, p.421-438, 1989. VAN WAGTENDONK-DE LEEUW, A.M.; AERTS, B.J.D.; DEN DASS, J.H.G. Abnormal offspring following in vitro production of bovine preimplantation embryos: a field study. Theriogenology, v.49, p.883-894, 1998. VAN WAGTENDONK-DE LEEUW, A.M.; MULLAART, E.; ROOS, A.P.W.; MERTON, J.S.; DEN DASS, J.H.G; KEMP, B.; RUIGH, L. Effects of different reproduction techniques: AI, MOET or IVP, on health and welfare of bovine offspring. Theriogenology, v.53, p.575-579, 2000. VERRHUIS, R.; HENDRIKSEN, P.J.; HENGST, A.M.; KRUIJT, L.; TIEMAN, M.; BOOMAN, P. The production of anti-H-Y monoclonal antibodies: their potential in use in a sex test for bovine embryos. Veterinary Immunology and Immunopathology, v.42, p.317-330, 1994. WATCHEL, S. H-Y antigen in the study of sex determination and control of sex ratio. Theriogenology, v.21, p.18-28, 1984. WHITE, K.L.; LINDNER, G.M.; ANDERSON, C.B.; BONDURANT, R.H. Survival after transfer of “sexed” mouse embryos exposed to H-Y antisera. Theriogenology, v.18, p.655-662, 1982. WILLIAMS, T.J. A technique for sexing mouse embryos by a visual colorimetric assay of the X-linked enzyme, glucose 6-phosphate dehydrogenase. Theriogenology, v.25, p.733-739, 1986.

26
2. OBJETIVOS
2.1 Objetivo geral
• Avaliar o uso de um controle interno em sistema de multiplex-PCR para a
determinação do sexo de embriões bovinos produzidos in vitro.
2.2 Objetivos específicos
• Otimizar um sistema de multiplex-PCR para a amplificação simultânea de
seqüências do cromossomo Y e do gene ND5 do DNA mitocondrial a partir
de embriões bovinos produzidos in vitro;
• Avaliar e corrigir as possíveis competições que o emprego simultâneo, em
sistema de multiplex-PCR, de primers utilizados para a amplificação de
segmentos de DNA, presentes em concentração e locais distintos, podem
ocasionar no processo de amplificação de seqüências do cromossomo Y
bovino;
• Avaliar a sensibilidade e a especificidade da multiplex-PCR;
• Determinar o sexo de dois grupos de embriões bovinos produzidos in vitro,
constituído por embriões frescos e congelados.

27
3. ARTIGO PARA PUBLICAÇÃO
(CITAR REVISTA??)

28
AVALIAÇÃO DE UM CONTROLE INTERNO EM SISTEMA DE MULTIPLEX-PCR
PARA A DETERMINAÇÃO DO SEXO DE EMBRIÕES BOVINOS PRODUZIDOS
IN VITRO
RESUMO
A utilização de um controle interno da reação em sistema de multiplex-PCR para a determinação do sexo de embriões bovinos produzidos in vitro foi avaliada em biópsias realizadas em dois grupos de embriões, constituídos por 54 embriões frescos e 44 congelados. Como controle interno foram utilizados os primers BOV 1 e BOV 2 que amplificam um produto com 626 pb do gene ND5 do DNA mitocondrial de bovinos. Os primers BRY.4aF e BRY.4aR (REED et al., 1989) foram utilizados para amplificação de seqüências encontradas no cromossomo Y bovino. A multiplex-PCR apresentou 100% de especificidade na determinação do sexo de 10 embriões machos e 10 embriões fêmeas. Tanto o produto do controle interno quanto o do cromossomo Y foram amplificados até a diluição correspondente a 6,25% do DNA total extraído de um embrião macho. Foi possível a determinação do sexo de 53 (98,1%) embriões frescos e de 40 (90,9%) embriões congelados. Em 34 (63%) embriões frescos e em 27 (61,4%) embriões congelados foram amplificados, simultaneamente, os produtos referentes ao cromossomo Y e ao DNA mitocondrial, caracterizando dessa forma o embrião como macho. O sexo feminino, caracterizado pela amplificação apenas do produto referente ao controle interno, foi detectado em 19 (35,2%) e em 13 (29,5%) embriões frescos e congelados, respectivamente. Em 1 (1,8%) e em 4 (9,1%) biópsias realizadas, respectivamente, em embriões frescos e congelados, nenhum produto foi amplificado provavelmente devido à ausência de células embrionárias. A falta de produto de amplificação em sistemas de PCR na ausência de controle interno caracteriza o embrião como fêmea, gerando resultados errôneos. A multiplex-PCR padronizada nesse trabalho caracterizou-se como sendo de fácil e rápida realização, além de ser sensível, específica e eficiente na identificação de falhas na reação apresentando, com isso, potencial para ser utilizada em escala comercial, na rotina de determinação do sexo de embriões bovinos produzidos in vitro.
Palavras-chave: bovinos, embriões, produção in vitro, sexagem, multiplex-PCR

29
EVALUATION OF AN INTERNAL CONTROL IN MULTIPLEX-PCR SYSTEM
FOR SEX DETERMINATION OF IN VITRO PRODUCED BOVINE EMBRYOS.
ABSTRACT
The use of an internal control in a multiplex-PCR assay for sex determination of in vitro produced bovine embryos was evaluated in biopsies of two groups, constituted by 54 fresh and 44 frozen embryos. It was used as internal control the primers BOV 1 and BOV 2 that amplify a product with 626 bp of bovine mitochondrial DNA ND5 gene. The primers BRY.4aF and BRY.4aR (REED et al., 1989) were used for bovine Y chromosome sequences amplification. The multiplex-PCR presented 100% specificity in sex determination of 10 male and 10 female embryos. Both amplicons of internal control and Y chromosome were amplified until the dilution corresponding to 6.25% of total extracted DNA from a male embryo. Sex determination was possible in 53 (98.1%) fresh embryos and 40 (90.9%) frozen embryos. The products related to Y chromosome and mitochondrial DNA were simultaneously amplified in 34 (63%) fresh embryos and 27 (61.4%) frozen embryos, showing a male embryo. The female sex distinguished by internal control amplification only, was detected in 19 (35.2%) and 13 (29.5%) biopsies, respectively, of fresh and frozen embryos. In 1 (1.8%) and 4 (9.1%) biopsies of fresh and frozen embryos, respectively, neither product was amplified probably due to the absence of embryonic cells. The lack of product amplification in PCR assay in the absence of an internal control can result in a misdiagnosed female embryo, leading to false result. The multiplex-PCR assay developed in this work showed easy and fast application, beside it was sensitive, specific and efficient in reaction failures identification, presenting a great potential to be used in commercial scale in routine sex determination of in vitro produced bovine embryos.
Keywords: bovine, embryos, in vitro production, sexing, multiplex-PCR

30
3.3. INTRODUÇÃO
O desenvolvimento de novas biotécnicas da reprodução animal tem
possibilitado aumento significativo no melhoramento genético em bovinos. Nos
últimos anos, têm-se verificado importantes mudanças na indústria da tecnologia
de embriões, com a ascensão da produção in vitro (PIV) de embriões bovinos,
(LUZ et al., 2000; CAMARGO et al., 2003), principalmente no Brasil, onde a
técnica mais se destaca. Apesar de ser uma técnica consolidada, vários estudos
relatam desproporção entre os sexos dos animais nascidos por meio da PIV,
sendo a porcentagem de machos superior à proporção teoricamente esperada de
50% para cada um dos sexos (GUTIERREZ-ADÁN et al., 1996; VAN
WAGTENDONK-DE LEEW et al., 2000; GUTIERREZ-ADÁN et al., 2001).
A maior proporção de embriões machos produzidos pela PIV abre
perspectiva para a implantação de sistemas efetivos de determinação do sexo de
embriões antes da transferência, uma vez que, em termos econômicos, a
produção de fêmeas é mais desejável. Os animais machos, quando obtidos por
meio de biotécnicas onerosas, são de baixo valor comercial, à exceção daqueles
de alto valor genético que poderão ser utilizados como reprodutores (COSTA et
al., 2002). Em contraposição, toda fêmea de elite é potencialmente uma futura
doadora. Essa característica implica em menor risco quanto ao investimento nos
animais desse sexo e justifica a maior valorização dos produtos do sexo feminino.
A PIV de embriões bovinos apresenta ainda como inconveniente a
indisponibilidade de protocolos práticos e eficientes de criopreservação, uma vez
que os embriões produzidos in vitro são mais sensíveis à congelação que aqueles

31
produzidos in vivo (VAJTA, 2000). Com isso, os produtos devem ser transferidos a
fresco. A possibilidade da determinação do sexo dos embriões anteriormente à
transferência, representa um avanço tecnológico significativo e com grande
impacto econômico.
Vários métodos têm sido desenvolvidos para identificar o sexo dos
embriões antes da transferência sendo classificados como invasivos ou não-
invasivos (VAN VLIET et al., 1989). Os métodos não-invasivos são considerados
ideais pois mantém a integridade do embrião, não interferindo em sua viabilidade.
Apesar dessa vantagem, esses métodos ainda não são efetivos para a
determinação do sexo de embriões bovinos com propósitos comerciais (VAN
VLIET et al., 1989; HASLER et al., 2002). Os métodos invasivos, que requerem
uma pequena amostra de células removida dos embriões, são os mais precisos.
Porém, a amostra obtida por biópsia deve ser de tamanho reduzido para não
comprometer a viabilidade dos embriões (AGRAWALA et al., 1992; BREDBACKA,
1998).
Entre os métodos desenvolvidos para a determinação do sexo de embriões,
a reação em cadeia da polimerase (PCR) é o que apresenta os melhores
resultados. A técnica destaca-se por ser a única que oferece resultados rápidos,
além de alta eficiência e acurácia na determinação do sexo. A PCR permite a
determinação do sexo logo após a biópsia do embrião, sem a necessidade de
criopreservação, procedimento exigido pela maioria das outras técnicas utilizadas
para determinação do sexo de embriões bovinos (SHRÖDER et al., 1990; PEURA
et al., 1991; BREDBACKA et al., 1995; HASLER et al., 1995; THIBIER e NIBART,
1995; ALVES et al., 2003).

32
Vários sistemas de PCR foram desenvolvidos para a detecção do
cromossomo Y em embriões bovinos. Em vários deles, a ausência de amplificação
representa um embrião do sexo feminino. Porém, a não-amplificação de um
produto esperado na PCR pode também ser conseqüência de falhas no sistema,
particularmente na extração do ácido nucléico das células embrionárias e na
própria PCR. Para o sucesso da técnica todo o sistema de amplificação deve ser
criteriosamente monitorado em todas as etapas analisadas (HERR e REED, 1991;
THIBIER e NIBART, 1995; LOPES et al., 2001).
O uso de controle interno na PCR é a forma mais eficaz para garantir a
eficiência e acurácia da técnica. Para isso vários métodos alternativos têm sido
testados. Porém, a maioria deles apresentam inconvenientes tais como: i) a
necessidade de realização de duas ou até três reações em separado para cada
embrião analisado, ii) competição de amplificação do DNA do controle interno com
o DNA proveniente das células embrionárias; iii) tamanho muito próximo dos
produtos amplificados (Y-específico e controle interno) exigindo que a análise dos
produtos seja realizada em gel de poliacrilamida, que é um processo mais
laborioso e demorado (PEURA et al., 1991; BREDBACKA et al., 1994; LUZ et al.,
2000, LOPES et al., 2001; PARK et al., 2001).
Em virtude de melhores efeitos na determinação do sexo quando se utiliza
um controle interno na reação, o presente trabalho teve como objetivos o
desenvolvimento de um sistema de multiplex-PCR para detecção do cromossomo
Y em embriões bovinos e avaliação da eficácia na determinação do sexo dos
embriões produzidos in vitro.

33
3.4. MATERIAL E MÉTODOS
3.4.1. PIV de embriões bovinos
Os oócitos utilizados para a PIV de embriões foram provenientes de ovários
coletados de matadouro. Os oócitos recuperados foram selecionados e
classificados para a maturação in vitro, que foi realizada durante 24 h em TCM
199 Bicarbonato (InvitrogenTM Life Technologies, EUA) suplementado com soro
fetal bovino (InvitrogenTM Life Technologies, EUA), piruvato de sódio, hormônio
folículo estimulante (FSH – Folltropin-V, Vetrepharm, Canadá) e hormônio
luteinizante (LH – APL, Bélgica). A fertilização in vitro (FIV) foi realizada conforme
protocolo descrito por Parrish et al. (1986), com sêmen previamente preparado em
gradiente de Percoll 45-90% (Sigma Co., EUA). Oócitos e espermatozóides foram
incubados por 20 h e, após esse período, os zigotos foram transferidos para o
meio SOFaa (TERVIT et al., 1972) para o cultivo in vitro (CIV), permanecendo por
7 a 12 dias (D7 a D12). Todas as etapas foram realizadas à temperatura de 38,8º
C e em atmosfera de 5% de CO2 (SENEDA et al., 2001).
3.4.2. Biópsia dos embriões
Para a colheita de células embrionárias foram utilizados dois grupos de
embriões nos estádios de blastocistos (D7) a blastocistos eclodidos (D12). Um
grupo foi constituído por embriões frescos (n=54) e o outro grupo (n=44) por
embriões armazenados individualmente em palhetas e congelados à –196º C em

34
meio de manutenção de embriões (Nutricell, Brasil). Os embriões foram
inspecionados previamente para avaliação da qualidade segundo norma proposta
pela Sociedade Internacional de Transferência de Embriões. Posteriormente à
avaliação, os embriões foram lavados individualmente por três vezes em tampão
salina fosfato (PBS) e dispostos em placas de petri descartáveis para a realização
de biópsias. Os embriões foram biopsiados utilizando a técnica descrita por
Bredbacka (1991), denominada scratched bottom com modificações para biópsias
manuais (BREDBACKA et al., 1995). As biópsias, correspondentes a
aproximadamente 20% do embrião, foram realizadas por meio de agulhas estéreis
e descartáveis, com diâmetro de 30 G. As células biopsiadas foram transferidas
para tubos contendo 5 µL de água ultrapura (MilliQ®) estéril, mantidos em banho
de gelo.
3.4.3. Extração do ácido nucléico de leucócitos de bovinos
Foram selecionados bovinos adultos Bos taurus taurus (n=2) e Bos taurus
indicus (n=2) de ambos os sexos. De cada animal foram obtidos 10 mL de sangue
total com anticoagulante (EDTA 10%) por punção da veia jugular. A extração do
ácido nucléico foi realizada em alíquotas de 100 µL de sangue total por meio do
método da sílica/tiocianato de guanidina, descrito por Boom et al (1990). O ácido
nucléico foi eluído em 40 µL de água ultrapura estéril e conservado a -20º C.
Todos os procedimentos de extração do ácido nucléico foram também realizados
em alíquotas de água ultrapura (MilliQ®) estéril como controle negativo.

35
3.4.4. Extração (ou Lise?) do ácido nucléico de células embrionárias
Para a obtenção do DNA das células embrionárias, em cada tubo contendo
as biópsias foi adicionado 0,8 µg de Proteinase K (InvitrogenTM Life Technologies,
EUA) para a lise das células embrionárias. Os tubos foram incubados em
termociclador (PTC-200, MJ Research Co. Water Town, MA, EUA) a 65º C por 30
min e, posteriormente, a 95º C por 10 min para a inativação da enzima e mantidos
à 4º C até a utilização. (REFERENCIAR?)
3.4.5. Oligonucleotídeos iniciadores (primers)
Para a detecção do cromossomo Y foram utilizados os primers BRY.4aF,
sense, 5'–CTC AGC AAA GCA CAC CAG AC–3' (nt. 1159-1178) e BRY.4aR,
antisense, 5'–GAA CTT TCA AGC AGC TGA GGC–3' (nt. 1439-1459), descritos
por Reed et al. (1989) e que a partir de uma seqüência repetida do cromossomo Y
bovino amplificam um produto com 300 pares de bases (pb).
Como controle interno da PCR foram utilizados os primers BOV1, sense, 5'–
ATA CGC CTT CAT TAC CAG–3' (nt. 12231-12248) e BOV2, antisense, 5'–TTG
AAT GGA GTA GTG CTG–3' (nt. 12856-12839) descritos por Wosiacki et al.
(2005) e que amplificam um produto de 626 pb do gene mitocondrial ND5 de Bos
taurus taurus e Bos taurus indicus (GenBank accesion nº NC_001567).

36
3.4.6. Multiplex-PCR
Anteriormente à padronização da multiplex-PCR foram avaliados vários
protocolos que envolveram desde a extração do ácido nucleico até as condições
(número de ciclos, tempo e temperaturas) da técnica.
A reação foi realizada utilizando 5 µL do DNA extraído de sangue total ou
de células embrionárias e 45 µL de PCR-Mix constituído por 0,4 pmol dos primers
BRY.4aF e BRY.4aR; 0,1 pmol dos primers BOV1 e BOV2; 1x PCR buffer (20 mM
Tris-HCl, pH 8,4; 50 mM KCl); 0,1 mM de cada dNTP, 2,5 mM MgCl2; 2,5 unidades
de Platinum Taq DNA polimerase e água ultrapura estéril para o volume final de
50 µL. Todos os reagentes utilizados foram sintetizados ou elaborados por
InvitrogenTM Life Technologies, EUA. O processo de amplificação foi realizado em
termociclador (PTC-200, MJ Research Co. Water Town, MA, EUA) com as
seguintes condições de tempo e temperatura: i) uma etapa de 5 min/95° C; ii) 30
ciclos de 1 min/95° C, 90 s/63° C e 1 min/72° C; iii) 10 ciclos de 1 min/95° C, 90
s/63° C, 30 s/55° C e 1 min/72° C; e iv) uma etapa de extensão final por 7 min/72°
C. Como controle negativo, em todos os procedimentos, foi incluída uma alíquota
de água ultrapura (MilliQ®) estéril.
3.4.7. Avaliação da sensibilidade da Multiplex-PCR
A determinação da sensibilidade da multiplex-PCR para a detecção das
seqüências do cromossomo Y e do gene mitocondrial bovino ND5 foi realizada por
meio de diluições do DNA extraído. Para o DNA extraído de leucócitos foram
utilizadas diluições seriadas em base 10 (10-1 – 10-7) (Quantificação?) e para o

37
DNA de células embrionárias, a partir de um embrião fresco, anteriormente
caracterizado como macho, foram realizadas oito diluições em base 2,
correspondente às frações de 50% a 0,39% do embrião. (Explicar?)
3.4.8. Avaliação da especificidade da Multiplex-PCR
A especificidade das reações da multiplex-PCR realizadas em biópsias,
correspondentes a aproximadamente 20% de cada embrião fresco, foi avaliada
por meio da realização da técnica no material celular restante (~80%) da biópsia
de embriões identificados como machos (n=10) e fêmeas (n=10). (Explicar?)
3.4.9. Análise dos produtos amplificados
Alíquotas de 7 µL dos produtos amplificados foram analisadas por
eletroforese em gel de agarose a 2% corado com brometo de etídio (0,5 µL/mL)
em solução de TEB pH 8,4 (89 mM TRIS; 89 mM ácido bórico; 2 mM EDTA) e
visualizados sob luz ultravioleta com posterior fotodocumentação digital (KODAK
DC290). Para a avaliação da sensibilidade da multiplex-PCR em células
embrionárias o produto amplificado foi também analisado por eletroforese em gel
de poliacrilamida (EGPA) a 7,5% (espessura do gel?) corado pelo nitrato de
prata (HERRING et al., 1982; DIAS NETO, 1992- não achei essa referência-
Emanuel Dias Neto, 1992).

38
4. RESULTADOS
A análise pela multiplex-PCR do DNA extraído de leucócitos de bovinos
proporcionou a amplificação de uma banda de 626 pb correspondente ao gene
mitocondrial ND5, utilizado como controle interno da reação, tanto em material
proveniente de animais machos quanto de fêmeas. Em DNA extraído de bovinos
machos foi possível também a amplificação da banda de 300 pb correspondente à
uma seqüência do cromossomo Y, que esteve ausente nas amostras provenientes
de animais fêmeas.
Nos experimentos de avaliação da sensibilidade da multiplex-PCR,
realizados em DNA extraído de leucócitos de bovinos machos, foi possível a
visualização em gel de agarose de ambos os produtos (626 e 300 pb) até a
diluição de 10-7 (mensurar?). Em células embrionárias provenientes de embrião
macho, os produtos foram visíveis até as diluições correspondentes a 6,25% e
0,781% do DNA extraído quando analisados, respectivamente, por eletroforese
em gel de agarose (Figura 1) e em gel de poliacrilamida corado pelo nitrato de
prata (Figura 2).
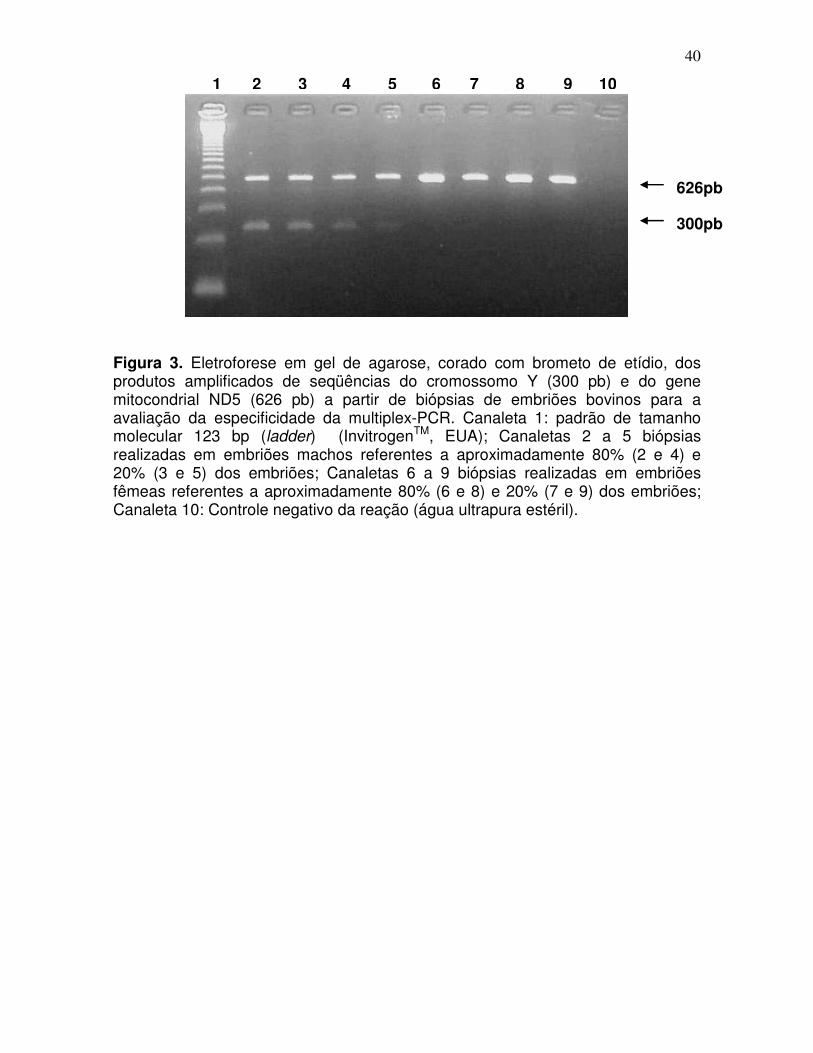
Na avaliação da especificidade da multiplex-PCR houve 100% de
concordância nos resultados utilizando DNA extraído de 20% e de 80% das
células de embriões machos (n=10) e fêmeas (n=10) (Figura 3).

39
Figura 1. Eletroforese em gel de agarose, corado com brometo de etídio, dos produtos amplificados de seqüências do cromossomo Y (300 pb) e do gene mitocondrial ND5 (626 pb) a partir de diluições seriadas de células embrionárias para a avaliação da sensibilidade da multiplex-PCR. Canaleta 1: padrão de tamanho molecular 123 bp (ladder) (InvitrogenTM, EUA); Canaletas 2 a 8: Diluições seriadas em base 2 referentes a 50% (2); 25% (3); 12,5% (4); 6,25% (5); 3,125% (6); 1,562% (7) e 0,781% (8) das células de um embrião bovino macho; Canaleta 9: Controle negativo da reação (água ultrapura estéril).
Figura 2. Eletroforese em gel de poliacrilamida, corado com nitrato de prata, dos produtos amplificados de seqüências do cromossomo Y (300 pb) e do gene mitocondrial ND5 (626 pb) a partir de diluições seriadas de células embrionárias para a avaliação da sensibilidade da multiplex-PCR. Canaleta 1: padrão de tamanho molecular 123 bp (ladder) (InvitrogenTM, EUA); Canaletas 2 a 10: Diluições seriadas em base 2 referentes a 50% (2); 50% (3); 25% (4); 12,5% (5); 6,25% (6); 3,125% (7); 1,562% (8); 0,781% (9); e 0,390 (10) das células de um embrião bovino macho.
626pb
300pb
1 2 3 4 5 6 7 8 9 10
1 2 3 5 6 7 8 9 4
626pb
300pb
40
Figura 3. Eletroforese em gel de agarose, corado com brometo de etídio, dos produtos amplificados de seqüências do cromossomo Y (300 pb) e do gene mitocondrial ND5 (626 pb) a partir de biópsias de embriões bovinos para a avaliação da especificidade da multiplex-PCR. Canaleta 1: padrão de tamanho molecular 123 bp (ladder) (InvitrogenTM, EUA); Canaletas 2 a 5 biópsias realizadas em embriões machos referentes a aproximadamente 80% (2 e 4) e 20% (3 e 5) dos embriões; Canaletas 6 a 9 biópsias realizadas em embriões fêmeas referentes a aproximadamente 80% (6 e 8) e 20% (7 e 9) dos embriões; Canaleta 10: Controle negativo da reação (água ultrapura estéril).
1 3 4 5 6 7 8 9 102
300pb
626pb

41
A multiplex-PCR realizada em células embrionárias obtidas por biópsia
apresentou taxas de 90,9% e 98,1% de eficiência, respectivamente, na
determinação do sexo de embriões congelados (n=44) e frescos (n=54) (Tabela
1).
Tabela 1. Identificação do sexo de embriões bovinos produzidos in vitro,
congelados e frescos, por meio da técnica de multiplex-PCR realizada em
biópsias embrionárias.
EMBRIÕES AVALIADOS (%) SEXO
CONGELADOS FRESCOS
Macho 27 (61,4) 34 (63,0)
Fêmea 13 (29,5) 19 (35,2)
Não Determinado 4 (9,1) 1 (1,8)
TOTAL 44 (100,0) 54 (100,0)
Nas amostras de DNA de embriões machos evidenciaram-se dois
fragmentos, referentes à seqüência do cromossomo Y (300 pb) e ao gene
mitocondrial ND5 (626 pb), correspondente ao controle interno. Por outro lado, nas
amostras com DNA de fêmeas evidenciou-se apenas um fragmento referente à
seqüência do controle interno. Dos 27 embriões congelados e classificados como
machos, 12 embriões apresentaram bandas autossômicas visíveis e fortes, porém
bandas Y-específicas fracas e inconsistentes (n=15) deixaram dúvidas sobre o
diagnóstico do sexo. Nos 34 embriões frescos identificados como machos, as
duas bandas apresentaram-se com a mesma intensidade. O padrão de

42
amplificação do DNA de embriões obtido com a multiplex-PCR pode ser
visualizado nas figuras 4 e 5.

43
Figura 4. Eletroforese em gel de agarose, corado com brometo de etídio, dos produtos amplificados por multiplex-PCR de seqüências do cromossomo Y (300 pb) e do gene mitocondrial ND5 (626 pb) a partir de biópsias de embriões bovinos frescos. Canaleta 1: padrão de tamanho molecular 123 bp (ladder) (InvitrogenTM, EUA); Canaletas 2, 3, 5, 8 e 9: embriões machos; Canaletas 4 6, 7 e 10: embriões fêmeas; Canaleta 11: Controle negativo da reação (água ultrapura estéril).
Figura 5. Eletroforese em gel de agarose, corado com brometo de etídio, dos produtos amplificados por multiplex-PCR de seqüências do cromossomo Y (300 pb) e do gene mitocondrial ND5 (626 pb) a partir de biópsias de embriões bovinos congelados. Canaleta 1: padrão de tamanho molecular 123 bp (ladder) (InvitrogenTM, EUA); Canaletas 2, 4 e 5: embriões machos; Canaletas 3 e 6: embriões fêmeas; Canaleta 7: Controle negativo da reação (água ultrapura estéril).
1 4 5 6 7 8 9 10 11 2 3
300pb
626pb
626pb
300pb
1 2 3 4 5 6 7

44
5. DISCUSSÃO
A padronização de sistemas de multiplex-PCR exige repetições múltiplas
das reações, com variações tanto da concentração dos reagentes quanto das
condições (números de ciclos, tempo e temperatura). A repetibilidade dos
resultados somente é alcançada por meio da utilização de ácido nucléico em
qualidade e concentrações fixas. Com isso, a padronização do sistema
diretamente em DNA proveniente de embriões bovinos seria de difícil realização
uma vez que a concentração do ácido nucléico extraído de um embrião não seria
suficiente para a realização de todas as análises. Para contornar esses
inconvenientes optou-se, inicialmente, por padronizar a multiplex-PCR descrita
nesse trabalho utilizando ácido nucléico extraído diretamente de sangue total de
bovinos machos e fêmeas adultos. Após a determinação da maior diluição do DNA
que, com a realização da técnica por meio de um protocolo padrão, possibilitasse
a amplificação dos produtos da seqüência do cromossomo Y e do gene
mitocondrial ND5, bem como, a visualização por meio da eletroforese em gel de
agarose, foram avaliados diversos protocolos alternativos.
A utilização de primers utilizados para o controle interno das reações,
caracterizando a multiplex-PCR, foi de grande importância considerando que os
resultados das amplificações são mais confiáveis quando tal procedimento é
adotado, excluindo a possibilidade de resultados errôneos originados
principalmente por falhas na amplificação de seqüências do cromossomo Y ou
ausência de produtos amplificados (THIBIER e NIBART, 1995; LOPES et al.,
2001). Apesar de alguns estudos relatarem como desnecessário o uso do controle

45
interno, outros estudos relataram maiores taxas de eficiência e acurácia na
determinação do sexo de embriões produzidos in vitro quando as reações são
realizadas na presença de um controle interno (NIBART et al., 1997; LOPES et al.,
1999; LOPES et al., 2001).
A opção pelo controle interno utilizado no presente trabalho ocorreu
principalmente pelas diferenças de temperatura (10° C) de hibridização dos
primers BRY.4a e BOV e do tamanho (300 e 626 pb) dos produtos amplificados,
pois esse aspecto facilita a diferenciação rápida dos produtos em gel de agarose.
Em testes preliminares, foi constatada a não amplificação de produtos da
seqüência do cromossomo Y na temperatura de anelamento dos primers para o
gene mitocondrial ND5 e vice-versa. Com isso, puderam ser realizados
experimentos avaliando o número de ciclos necessários para a amplificação de
cada produto em uma mesma reação, sem a ocorrência de interferências
passíveis de serem observadas quando as temperaturas de anelamento dos
primers são muito próximas.
O maior número de cópias do gene mitocondrial ND5 por célula, em relação
à seqüência do cromossomo Y, pode favorecer a amplificação do controle interno
em detrimento à do cromossomo Y. Devido a esse inconveniente, foi necessária
também a padronização do número de ciclos ideais para a amplificação de cada
produto. Todas as reações foram realizadas com 40 ciclos térmicos. Porém, o
número de ciclos teve que ser padronizado e consistiu na amplificação apenas da
seqüência do cromossomo Y nos primeiros 30 ciclos. Somente nos últimos 10
ciclos foi realizada a amplificação simultânea das seqüências do cromossomo Y e
do gene mitocondrial ND5. Devido à diferença na temperatura de hibridização dos

46
primers (BRY.4a e BOV) não houve a necessidade de interrupção da reação para
a inclusão dos primers do controle interno, conforme descrito por outros autores
(LOPES et al., 2001; PARK et al., 2001).
Também com o objetivo de favorecer a amplificação da seqüência do
cromossomo Y, o tempo de anelamento dos primers do controle interno foi
reduzido para apenas 30 s, pois com 1 min houve amplificação em excesso do
produto da seqüência do gene mitocondrial ND5 em detrimento à seqüência do
cromossomo Y. Pelo mesmo motivo a concentração dos primers BOV 1 e BOV 2
também teve que ser reduzida de 0,4 pmol para 0,1 pmol.
Somente após a definição das melhores condições com relação à
concentração dos reagentes (primers, dNTP, tampão da PCR, MgCl2) e,
principalmente, do número de ciclos ideais para a amplificação dos produtos foram
realizados os experimentos relativos à avaliação da sensibilidade, especificidade,
bem como de determinação do sexo dos embriões bovinos congelados e frescos.
A especificidade foi determinada pela realização da multiplex-PCR em
biópsias correspondentes a 10-20% de um embrião fresco e como contraprova
foram analisados os 80-90% restantes do embrião. Em todos os experimentos
houve 100% de concordância nos resultados, demonstrando que a falta de
amplificação da seqüência do cromossomo Y nos embriões fêmeas analisados
(n=10) não foi devido a falhas no sistema.
Diluições seriadas realizadas a partir do DNA extraído de um embrião
fresco caracterizado como macho foram realizadas com o intuito de simular a
biópsia embrionária e testar a sensibilidade do sistema. A fácil visualização dos
dois produtos amplificados a partir de diluições correspondentes a 6,25% do DNA

47
extraído de um embrião em gel de agarose e a 0,781% em EGPA demonstra que
a multiplex-PCR desenvolvida pode ser realizada em poucas células embrionárias
correspondentes a menos de 10% das células de um embrião. A maior
sensibilidade da técnica, quando os produtos foram analisados em EGPA seguida
de coloração pela prata, deve-se à maior sensibilidade desse sistema de
revelação quando comparado ao brometo de etídio. Entretanto, a EGPA seguida
da coloração pela prata é uma técnica mais laboriosa e demorada, que inviabiliza
sua implantação como técnica de rotina na determinação pré-implantação do sexo
de embriões bovinos.
A taxa de eficiência da multiplex-PCR na determinação do sexo dos
embriões foi de 90,9% para embriões congelados e 98,1% para embriões frescos.
A taxa de eficiência de 90,9% encontrada demonstrou que, apesar da
degeneração dos embriões congelados, obteve-se uma porcentagem semelhante
a outros trabalhos (~90%) que utilizaram embriões de boa qualidade (SHEA,
1999; CAMARGO et al., 2003). A taxa de eficiência de 98,1% em embriões frescos
e de boa qualidade foi superior à obtida por outros autores que relataram taxas de
94% a 95,4% (MACHÁTY et al., 1993; LOPES et al., 2001).
As freqüências de identificação do cromossomo Y em embriões congelados
(61,4%) e frescos (63,0%) foi equivalente a outros trabalhos que também
relataram desproporção entre os sexos de embriões produzidos in vitro, com taxas
de 56,8% a 59,6% de produtos machos (BOUSQUET et al., 1999; CAMARGO et
al., 2003).
Embora muitas vezes têm-se constatado desproporção entre os dois sexos
de embriões produzidos in vitro, a causa dessa diferença ainda não está

48
estabelecida. Os estudos abordam vários fatores relacionados, entre os quais o
desenvolvimento mais rápido dos embriões machos em detrimento das fêmeas
nos oito primeiros dias após a FIV. Sugere-se também algum efeito no
desenvolvimento dos embriões do sexo masculino durante o CIV, podendo assim
proporcionar a escolha de embriões para implantação em estádios mais
avançados no desenvolvimento e o descarte dos embriões menos adiantados
(AVERY et al., 1991; HASLER, 1998).
Não foi possível determinar o sexo de 4 (9,1%) embriões congelados e de
apenas 1 (1,8%) embrião fresco, uma vez que no material proveniente desses
embriões também não foi possível a visualização do fragmento correspondente ao
gene mitocondrial ND5. A não-amplificação da seqüência utilizada como controle
interno provavelmente foi devido à falha ocorrida durante o procedimento de
biópsia como a não inclusão de células embrionárias.
A viabilidade dos embriões foi fundamental para o sucesso da multiplex-
PCR. Embriões produzidos in vitro, em contraposição àqueles produzidos in vivo,
são mais sensíveis ao processo de congelamento (-196° C). A interferência da
viabilidade do embrião no processo de amplificação ficou evidenciada uma vez
que no grupo de embriões congelados, 12 embriões considerados machos
apresentaram bandas da seqüência do cromossomo Y de fraca intensidade em
gel de agarose, muito provavelmente por estarem degenerados, com conseqüente
degradação do DNA celular. Em contrapartida, os produtos de amplificação da
seqüência do cromossomo Y e do gene mitocondrial ND5, quando provenientes
de células embrionárias de embriões machos que não foram submetidos ao

49
congelamento, foram amplificados em igualdade de condições, uma vez que a
intensidade das bandas visualizadas em gel de agarose foi muito semelhante.
O sistema de multiplex-PCR padronizado no presente trabalho
proporcionou a rápida determinação do sexo de embriões bovinos produzidos in
vitro por meio da realização de uma única reação por embrião. A diferenciação
dos produtos amplificados referentes ao controle interno e ao cromossomo Y foi
de fácil visualização em eletroforese em gel de agarose, particularmente devido a
sua grande diferença de tamanho (pb). A padronização do sistema proporcionou
condições ideais de amplificação específica para cada produto, evitando
interferências passíveis de ocorrer quando há maior disponibilidade de cópias de
DNA do controle interno. A multiplex-PCR apresentou taxas de sensibilidade e
especificidade na determinação do sexo de embriões provenientes de Bos taurus
taurus e Bos taurus indicus que possibilitam a sua implantação para uso comercial
em laboratórios de PIV de embriões bovinos.

50
6. REFERÊNCIAS
ALVES, B.C.A.; HOSSEPIAN DE LIMA, V.F.M.; TEIXEIRA, C.M.; MOREIRA FILHO, C.A. Use of primers derived from a new sequence of the bovine Y Chromosome for sexing Bos taurus and Bos indicus embryos. Theriogenology, v.59, p.1415-1419, 2003. AGRAWALA, P.L.; WAGNER, V.A.; GELDERMANN, H. Sex determination and milk protein genotyping of preimplant stage bovine embryos using multiplex PCR. Theriogenology, v.38, p.969-78, 1992. AVERY, B.; MADISON, V.; GREVE, T. Sex and development in bovine in vitro embryo production. Theriogenology, v.35, p.953-963, 1991. BOOM, R.; SOL, C.J.A.; SALIMANS, M.M.M; JANSEN, C.L.; WERTHEIM-van DILLEN, P.M.E.; NOORDAA, J. van der. Rapid and Simple Method for Purification of Nucleic acids. Journal of Clinical Microbiology, v. 28, n. 3, p. 495-503, 1990. BOUSQUET, D.; TWAGIRAMUNGU, H.; MORIN, N.; BRISSON, C.; CARBONNEAU, G.; DUROCHER, J. In vitro embryo production in the cow: an effective alternative to the conventional embryo production approach. Theriogenology, v.51, p.59-70, 1999. BREDBACKA, P. Biopsy of morulae and blastocysts. Reproduction of Domestic Animal, v.26, p.82-84, 1991. BREDBACKA, P.; VELMALA, R.; PEIPPO, J.; BREDBACKA, K. Survival of biopsied and sexed bovine demi-embryos. Theriogenology, v.41, p.1023-1031, 1994. BREDBACKA, P.; KANKAANPÄÄ, A.; PEIPO, J. PCR-sexing of bovine embryos: a simplified protocol. Theriogenology, v.44, p.167-176, 1995. BREDBACKA, P. Recent development in embryos sexing and its field application. Reproduction Nutrition Develoment, v.38, p.605-613, 1998. CAMARGO, L.S.A.; SÁ, W.F.; VIANA, J.H.M.; FERREIRA, A.M.; SERAPIÃO, R.V.; RAMOS, A.A.; MACHADO, M.A.; VALE FILHO, V.R.; ANDRADE, V.J. Identificação do sexo de embriões bovinos fecundados in vitro e cultivados com células do cumulus na presença de soro. Revista Brasileira de Reprodução Animal, v.27, n.3, p.407-409, 2003. COSTA, L.L.; SILVA, J.C.; DINIZ, P.; CIDADÃO, R. Preliminary report on sexing bovine pre-implantation embryos under the conditions of Portugal. Revista Portuguesa de Ciências Veterinárias, v.97, p.95-98, 2002.

51
GUTIÉRREZ-ADÁN, A.; BEHBOODI, E.; ANDERSON, G.B.; MEDRANO, J.F.; MURRIA, J.D. Relationship between stage of development and sex of bovine IVM-IVF embryos cultured in vitro versus in the sheep oviduct. Theriogenology, v.46, p.515-525, 1996. GUTIÉRREZ-ADÁN, A.; LONERGAN, P.; RIZOS, D.; WARD, F.A.; BOLAND, M.P.; PINTADO, B.; FUENTE, J. Effect of the in vitro culture system on the kinetics of blastocyst development and sex ratio of bovine embryos. Theriogenology, v.55, p.1117-1126, 2001. HASLER, J.F.; HENDERSON, W.B.; HURTGEN, P.J.; JIN, Z.Q.; MC CAULEY, A.D.; MOWER, S.A.; NEELY, B.; SHUEY, L.S.; STOKES, J.E.; TRIMMER, S.A. Production, freezing and transfer of bovine IVF embryos and subsequent calving results. Theriogenology, v.43, p.141-152, 1995. HASLER, J.F. The current status of oocyte recovery in vitro embryo production in domestic animals with emphasis on bovine. Journal of Reproduction and Fertility, v.76, n.3, p.52-74, 1998. HASLER, J.F.; CARDEY, E.; STOKES, J.E.; BREDBACKA, P. Nonelectrophoretic PCR-sexing of bovine embryos in a commercial environment. Theriogenology, v.58, 1457-1469, 2002. HERR, C.M.; REED, K.C. Micromanipulation of bovine embryos for sex determination. Theriogenology, v.35, p.45-54, 1991. HERRING, A.J.; INGLIS, N.F.; OJEH, C.K.; SNODGRASS, D.R.; MENZIES, J.D. Rapid diagnosis of rotavirus infection by direct detection of viral nucleic acid in silver-strained polyacrilamide gels. Journal of Clinical Microbiology, v. 16, p. 473-47, 1982. LAEMMLI, V.K. Cleavage of strutural proteins during the assembly of the head of bacteriophage T4. Nature, v. 227, p. 680-685, 1970. LOPES, R.F.F; SENNA, J.P.M.; CHIES, J.M.; RODRIGUES, J.L. Pit-stop PCR: an approach to increase final product yield of multiplex PCR. Biotechniques, v.26, p.638-639, 1999. LOPES, R.F.F.; FORRELL, F.; OLIVEIRA, A.T.D.; RODRIGUES, J.L. Splitting and biopsy for bovine embryo sexing under field conditions. Theriogenology, v.56, p.1383-1392, 2001. LUZ, M.R.; WATANABE, Y.F.; FERRO, J.A.; FERRO, M.I.T.; MAURO, S.M.S.; HOSSEPIAN DE LIMA, V.F.M.; FRANCESCHINI, P.H. Identificação do sexo de embriões bovinos fecundados in vitro pela técnica de PCR multiplex. Brazilian Journal of Veterinary Research and Animal Science, v.37, n.6, p.453-456, 2000.

52
MACHÁTY, Z.; PÁLDY, A.; CSÁKI, T; VARGA, Z.; KISS, I.; BÁRÁNDI, Z.; VAJTA, G. Biopsy and sex determination by PCR of IVF bovine embryos. Journal of Reproduction and Fertility, v.98, p.467-470, 1993. NIBART, M., MARQUANT LE GUIENNE, B.; HUMBLOT, P.; GUERIN, B. The application of new reproductive technologies in France. Arquivos da Faculdade de Veterinária UFRGS, v.25 (suplemento 1), p.21-35, 1997. PARK, J.H.; LEE, J.H.; CHOI, K.M.; JOUNG, S.Y.; KIM, J.Y.; CHUNG, G.M.; JIN, D.I.; IM, K.S. Rapid sexing of preimplantation bovine embryo using consecutive and multiplex polimerase chain reaction (PCR) with biopsied single blastomere. Theriogenology, v.55, p.1843-1853, 2001.
PARRISH, J.J.; SUSKO-PARRISH, J.L.; LEIBFRIEDGE-RUTHEDGE, M.L.; CRITSER, E.S.; EYESTONE, W.H.; FIRST, N.L. Bovine in vitro fertlization with frozen thawed semen. Theriogenology, v.25, p.591-600, 1986. PEURA , T.; HYTTINEN, J.M.; TURUNEN, M.; JÄNNE, J. A reliable sex determination assay for bovine preimplantation embryos using the polimerase chain reaction. Theriogenology, v.35, p.547-555, 1991. REED, K.C.; MATTHALI, K.I.; MANN. D.A.; BEATON, S. Determination of genetic sex in ruminants using Y-chromossome-specific polynucleotides. Patent Cooperation Treaty Nº WO 89/07154 (1989). SCHRÖDER, A.; MILLER, J.R.; THOMSENP.D.; ROSCHLAUD, K.; AVERY, B.; POULSEN, P.H.; SCHMIDT, M.; SCHWERIN, M. Sex determination of bovine embryos using the polymerase chain reaction. Animal Biotechnology, v.1, n.1-2, p.121-133, 1990. SENEDA, M.M.; ESPER, C.R.; GARCIA, J.M.; VANTINI, R.; OLIVEIRA, J.A. Relationship between follicle size and ultrasound-guided transvaginal oocyte recovery. Animal Reproduction Science, v. 67, p. 37-43, 2001. TERVIT, H.R.; WHITTNGHAM, D.G.; ROWSON, L.E.A. Sucessful culture in vitro of sheep and cattle ova. Journal of Reproduction and Fertility, v.30, p.493-497, 1972. THIBIER, M.; NIBART, M. The sexing of bovine embryos in the field. Theriogenology, v.43, p.71-80, 1995. VAJTA, G. Vitrification of the oocytes and embryos of domestic animals. Animal Reproduction Science, v.60-61, p.357-364, 2000. VAN VLIET, R.A.; GIBBINS, A.M.V.; WALTON, J.S. Livestock embryo sexing: a review of current methods, with emphasis on Y-specific DNA probes. Theriogenology, v.32, n.3, p.421-438, 1989.

53
VAN WAGTENDONK-DE LEEUW, A.M.; MULLAART, E.; ROOS, A.P.W.; MERTON, J.S.; DEN DASS, J.H.G; KEMP, B.; RUIGH, L. Effects of different reproduction techniques: AI, MOET or IVP, on health and welfare of bovine offspring. Theriogenology, v.53, p.575-579, 2000. WOSIACKI, S.R.; BARREIRO, M.A.; ALFIERI, A.F.; ALFIERI, A.A. Semi-nested PCR for detection and typing of bovine papillomavirus type 2 in urinary bladder and whole blood from cattle with enzootic haematuria. Journal of Virological Methods, v.126, p.215-219, 2005.

54
4. CONCLUSÕES
• O emprego simultâneo, em sistema de multiplex-PCR, dos primers
BRY.4a(F) e BRY.4a(R) e BOV 1 e BOV 2 que amplificam,
respectivamente, produtos com 300 e 626 pb a partir de seqüências
encontradas no cromossomo Y bovino e no gene mitocondrial ND5
possibilitou a definição do sexo de embriões bovinos;
• Na determinação do sexo de embrião bovino a utilização de um controle
interno em uma mesma reação (multiplex-PCR) pode ocasionar competição
responsável pela redução ou mesmo inibição da amplificação de
seqüências Y-específicas;
• Alterações realizadas na concentração de reagentes e nas condições
(tempo, temperatura e número de ciclos) da reação possibilitaram a
amplificação simultânea, por multiplex-PCR, de seqüências do cromossomo
Y e do gene mitocondrial ND5 a partir do DNA extraído de células obtidas
por biópsias de embriões bovinos produzidos in vitro;
• As taxas de determinação do sexo de embriões bovinos (eficiência) obtidas
pela multiplex-PCR foram superiores às observadas na literatura;
• A multiplex-PCR apresentou taxas de sensibilidade e especificidade que
possibilitam sua implantação como técnica de rotina para determinação do
sexo de embriões bovinos produzidos in vitro.

55
5. APÊNDICES
5.1. LISTA DE REAGENTES
5.2. LISTA DE SOLUÇÕES E TAMPÕES
5.3. PROTOCOLOS DE TÉCNICAS

56
5.1. LISTA DE REAGENTES
1. Acetona, P.A. (CH3COCH3) PM 58,08 (Dinâmica)
2. Ácido acético glacial, P.A. (CH3COOH) PM 60,05 (Nuclear)
3. Ácido bórico (H3BO3) PM 61,83 (Sicalab)
4. Ácido clorídrico (HCl) PM 36,46 (Reagen)
5. Ácido etilenodiaminotetraacético SAL DI-SÓDICO – EDTA, P.A.
(C10H14N2O8Na2.2H2O) PM 372,24 (Reagen)
6. Ácido sulfúrico, P.A. (H2SO4) PM 98,08 (Synth)
7. Acrilamida (C3H5NO) PM 71,08 (Vetec)
8. Agar Noble (Difco)
9. Agarose (Invitrogen TM Life Technologies)
10. Álcool etílico absoluto (C2H2OH) PM 46,07 (Nuclear)
11. Álcool etílico hidratado 92,8o INPM – 95,0o GL (Da Ilha e D’aldeia)
12. Álcool isoamílico
13. Azul de Bromofenol (Sigma)
14. Brometo de etídeo (C21H20N3Br) PM 394,3 (Sigma)
15. Cloreto de Magnésio (MgCl2) 50 mM (Invitrogen TM Life Technologies)
16. Cloreto de potássio, P.A. (KCl) PM 74,56 (Reagen)
17. Cloreto de sódio, P.A. (NaCl) P.M. 58,45 (Reagen)
18. Clorofórmio, P.A. (CHCl3) PM 119,38 (Dinâmica)
19. Dimetil sulfóxido (Sigma)
dNTP (100 mM, 4 x 250 µL; 25 µmol cada - 100 mM dATP, 100 mM dCTP,
100 mM dGTP, 100 mM dTTP) (InvitrogenTM Life Technologies)
20. Dodecilsulfato de sódio – Lauril sulfato de sódio – SDS (C12H25NaO4S) PM
288,38 (BDH e Synth)
21. Fenol saturado
22. Folmoldeído (CH2O) PM 30,03 (Biotec)
23. Glicerol
24. Glicina, P.A. (Nuclear)

57
25. Hidróxido de sódio P.A. (NaOH) PM 40,00 (Dinâmica)
26. Hidroximetil amino metano – TRIS 99% PM 121,14 (Inlab)
27. Isotiocianato de guanidina PM 118,16 (Invitrogen TM Life Technologies)
28. Metanol, P.A. (CH3OH) PM 32,04 (Allkimia)
29. 2-Mercapto-etanol (C2H6O5) PM 78,13 (Fluka)
30. N,N’-Metileno-bis-acrilamida (Sigma)
31. Nitrato de prata, P.A. (AgNO3) (Symylar)
32. Persulfato de amônio P.A. ((NH4)2S2O8) PM 288,2 (Sigma)
33. Platinum Taq DNA Polymerase (Invitrogen TM Life Technologies)
34. Potássio fosfato monobásico, P.A. (KH2PO4) PM 136,09 (Labsynth)
35. Proteinase K (Fungal) (Invitrogen TM Life Technologies)
36. Sódio fosfato di-básico anidro, P.A. (Na2HPO4) PM 141,96 (Synth)
37. TEMED PM 116,21 (Invitrogen TM Life Technologies)
38. Triton X-100

58
5.2. LISTA DE SOLUÇÕES E TAMPÕES
1. Tampão fosfato salina PBS 1x [ ]
2. Sílica
3. L6
4. L2
5. Álcool etílico 70o
6. Tampão de amostra para agarose
7. Tampão de corrida para agarose TEB (TRIS – Ácido bórico – EDTA) 1x[ ]
8. Ágar
9. Persulfato de Amônio P.A. 0,2%
10. Acrilamida
11. Lower Tris
12. Upper Tris
13. Tampão de amostra para acrilamida
14. Tampão de corrida para acrilamida
15. Solução Fixadora
16. Solução de Prata
17. Solução Stop
18. Solução de Coloração
19. Solução de Conservação

59
5.3. PREPARO DE SOLUÇÕES E TAMPÕES
Álcool Etílico 70%
- Álcool etílico hidratado 92,8o INPM – 95,0o GL (C2H2OH)
- 100 mL de H2O MiliQ® q.s.p.
Proteinase K
- 100 mg de Proteinase K (> 20 unidades/mg)
- 5 mL de H2O MiliQ® autoclavada q.s.p.
Fonte: segundo o fabricante
Sílica
- 60 g de sílica (Dióxido de sílica - SiO2)
- 500 mL de H2O MilliQ® autoclavada q.s.p.
- agitar lentamente e manter em repouso durante 24 horas
- desprezar 430 mL do sobrenadante por sucção
- ressuspender 500 mL da sílica em H2O MiliQ® autoclavada q.s.p.
- permanecer em repouso por 5 horas
- desprezar 440 mL do sobrenadante
- adicionar 600 µL de ácido clorídrico (HCl) - ajustar o pH (2,0)
- aliqüotar e autoclavar
Tampão L6
- 120 g de isotiocianato de guanidina
- 100 mL de Tris-HCl 0,1M (pH 6,4)
- 22 mL de EDTA (ácido etilenodiaminotetraacético) 0,2M (pH 8,0)
- 2,6 g de Triton X-100

60
Fonte: BOOM et al. (1990)
Tampão L2
- 120 g de isotiocianato de guanidina
- 100 mL de Tris-HCl 0,1M (pH 6,4) Fonte: BOOM et al. (1990)
Tampão Fostato Salina - PBS 1x [ ]
- Cloreto de sodio, P.A. (NaCl) 8,0 g (137 mM)
- Cloreto de potássio, P.A. (KCl) 0,2 g (3 mM)
- Sódio fosfato dibásico anidro, P.A. (Na2HPO4) 1,2 g (8 mM)
- Potássio fosfato monobásico, P.A. (KH2PO4) 0,2 g (15 mM)
- H2O bd q.s.p. 1.000 mL
Tampão de corrida para eletroforese em gel de agarose - TEB (Tris – ácido
bórico – EDTA) 10x[ ]
- 10,778 g de Tris (Hidroximetil amino metano) - 89 mM
- 5,503 g de ácido bórico (H3BO3) - 89 mM
- 0,747 g de EDTA - 2 mM
- H2O bd q.s.p. 1.000 mL
- pH 8,4
Tampão de amostra para Agarose
- Azul de bromofenol 0,25%
- Sacarose, P.A. – sucrose (C12H22O11) 45%

61
Persulfato de Amônio P.A. 0,2%
- Persulfato de Amônio P.A. ((NH4)2S2O8) 0,04 g
- H2O bd 2 mL
Acrilamida (50:1,3)
- N,N’-Metileno-bis-acrilamida 1,3 g
- Acrilamida 50 g
- H2O bd q.s.p. 100 mL
Lower Tris
- Hidroximetil amino metano – TRIS 36,34 g
- H2O bd q.s.p. 200 mL
- Ácido clorídrico (HCl) para ajustar o pH (8,8)
Upper Tris
- Hidroximetil amino metano – TRIS 12,12 g
- H2O bd q.s.p. 200 mL
- Ácido clorídrico (HCl) para ajustar o pH (6,8)
Tampão de amostra para Acrilamida
- Azul de bromofenol (0,05%) 0,4 mL
- SDS 10% 2,0 mL
- 2-Mercapto-ethanol (C2H6O5) 0,2 mL
- Hidroximetil amino metano – TRIS-HCl 0,5M 2,0 mL (pH 6,8)
- Glicerol 2,0 mL
- H2O bd 4,7 mL

62
Tampão de corrida para Acrilamida
- Hidroximetil amino metano – TRIS 3 g
- Glicina, P.A. 14,4 g
- H2O bd q.s.p. 1.000 mL
Solução Fixadora
- Álcool etílico absoluto (C2H2OH) 30 mL
- Ácido acético glacial, P.A. (CH3COOH) 1,5 mL
- H2O bd q.s.p. 300 mL
Solução de Prata
- Nitrato de prata, P.A. (AgNO3) 0,55 g
- H2O bd q.s.p. 300 mL
Solução Stop
- Ácido acético glacial, P.A. (CH3COOH) 15 mL
- H2O bd q.s.p. 300 mL
Solução de Coloração
- Hidróxido de sódio, P.A. (NaOH) 9 g
- Folmoldeído (CH2O) 2,5 mL
- H2O bd q.s.p. 300 mL
- Sodium Borohydride 0,06 g
Solução de Conservação

63
- Álcool etílico absoluto (C2H2OH) 30 mL
- H2O bd q.s.p. 300 mL

64
5.4. PROTOCOLOS DE TÉCNICAS
Extração do DNA de leucócitos pela técnica de sílica/tiocianato de guanidina
- Adicionar 100 µL de sangue total
- Adicionar 20 µL de sílica hidratada
- Adicionar 1000 µL de tampão L6
- Homogeneizar em vortex
- Agitar durante 30 minutos em temperatura ambiente
- Centrifugar a 10.000 x g durante 30 segundos
- Desprezar o sobrenadante em solução contendo NaOH 10 M
- Adicionar 500 µL de tampão L2
- Homogeneizar em vórtex
- Centrifugar a 10.000 x g durante 30 segundos
- Desprezar o sobrenadante em solução contendo NaOH 10 M
- Adicionar 500 µL de tampão L2
- Homogeneizar em vórtex
- Centrifugar a 10.000 x g durante 30 segundos
- Desprezar o sobrenadante em solução contendo NaOH 10 M
- Adicionar 1000 µL de etanol 70% gelado
- Homogeneizar em vórtex
- Centrifugar a 10.000 x g durante 30 segundos
- Desprezar o sobrenadante
- Adicionar 1000 µL de etanol 70% gelado
- Homogeneizar em vórtex
- Centrifugar a 10.000 x g durante 30 segundos
- Desprezar o sobrenadante
- Adicionar 1000 µL de acetona PA gelada
- Homogeneizar em vórtex
- Desprezar o sobrenadante
- Secar o pellet em banho-maria a 56o C durante 15 minutos

65
- Adicionar 50 µL de água ultrapura autoclavada
- Homogeneizar em vórtex
- Deixar em banho-maria à 56o C durante 15 minutos
- Homogeneizar em vórtex
- Centrifugar a 10.000 x g durante 2 minutos
- Recolher o sobrenadante (40 µL)
- Estocar a –20o C
Fonte: BOOM et al., 1990
Extração do DNA de embriões bovinos produzidos in vitro
- 5,0 µL de água ultrapura estéril
- biópsias dos embriões
- 2,0 µL de proteinase K
- Programa PROTK no termociclador: 65º C/30min e 95º C/10min
Multiplex-PCR
- 5,0 µL de buffer 10x (concentração final de 1x)
- 2,5 µL de Cloreto de magnésio 50 mM (concentração final 2,5 mM)
- 2,0 µL da solução de uso de dNTPs (concentração final 0,1 mM de cada)
- 0,5 µL de Platinum Taq DNA polimerase (concentração final 2,5 unidades)
- 2,0 µL do primer BRY.4aF
- 2,0 µL do primer BRY.4aR
- 2,0 µL dos primers BOV1/BOV2
- 29 µL de água ultrapura MilliQ® autoclavada
Programa SEXAGEM no termociclador
- 1 ciclo (95o C durante 5min)

66
- 30 ciclos (95o C durante 1min; 63o C durante 1min e 30s; 72o C durante 1 min)
- 10 ciclos (95o C durante 1min; 63o C durante 1min e 30s; 55º C durante 30 s.; 72o
C durante 1 min)
- 1 ciclo (72o C durante 7 minutos)
Gel de Agarose a 2%
- Agarose 1,0 g
- TEB 1x[ ] 50 mL
- Brometo de etídeo (C21H20N3Br) 15 µL
- aplicar 8 µL do produto com 2 µL do tampão de amostra para agarose e
submeter à eletroforese no tampão de corrida
- visualizar em luz ultravioleta
Gel de Poliacrilamida
- montar a forma com solução de acrilamida
Lower Gel
- H2O bd 11,44 mL
- Solução de Lower Tris 5,0 mL
- Solução de Acrilamida 3,0 mL
- Solução de Persulfato de amônio - 2% 0,56 mL
- TEMED 100 µL
Upper Gel
- H2O bd 6,2 mL
- Solução de Upper Tris 2,5 mL
- Solução de Acrilamida 0,7 mL

67
- Solução de Persulfato de amônio 2% 0,6 mL
- TEMED 50 µL
- aplicar 10 µL do produto com 5 µL de H2O ultrapura miliQ® autoclavada e 10 µL
do tampão de corrida para acrilamida e submeter a eletroforese em tampão de
corrida para acrilamida
- agitar o gel na solução fixadora por 30 min, na solução de prata por 30 min, corar
na solução de coloração até obter a coloração desejada, aplicar a solução stop por
10 a 15 min e manter na solução de conservação.