running head: new insights on regulation of stm gene function
TRANSCRIPT

1
Running head: New insights on regulation of STM gene function.
The author responsible for distribution of materials integral to the findings presented in this
article in accordance with the policy described in the instructions for authors
(www.plantphysiol.org) is:
Neelima R. Sinha
Department of Plant Biology, College of Biological Sciences, University of
California, Davis, CA 95616, U. S. A.
Telephone: 530 754 8441
Email: [email protected]
Primary research area:
Genes, Development and Evolution
Plant Physiology Preview. Published on December 18, 2014, as DOI:10.1104/pp.114.248625
Copyright 2014 by the American Society of Plant Biologists
www.plantphysiol.orgon February 14, 2018 - Published by Downloaded from Copyright © 2014 American Society of Plant Biologists. All rights reserved.

2
Transcriptional, post-transcriptional and post-translational
regulation of STM gene expression in Arabidopsis determines
gene function in the shoot apex
José Antonio Aguilar-Martínez1
Naoyuki Uchida1,2
Brad Townsley1
Donnelly Ann West1
Andrea Yanez1
Nafeesa Lynn1
Seisuke Kimura1,3
Neelima Sinha1*
1 Department of Plant Biology, University of California, Davis, CA, USA.
2 WPI-Institute of Transformative Bio-Molecules (WPI-ITbM), Nagoya University, Furo-cho, Chikusa-ku,
Nagoya, 464-8602, Japan.
3 Department of Bioresource and Environmental Sciences, Kyoto Sangyo University, Kyoto 603-8555, Japan
Components of the complex regulation of a homeobox gene key for plant development are dissected at
transcriptional, post-transcriptional and post-translational levels.
www.plantphysiol.orgon February 14, 2018 - Published by Downloaded from Copyright © 2014 American Society of Plant Biologists. All rights reserved.

3
FOOTNOTES
This work was supported by a postdoctoral fellowship from Spanish Ministry of Science
(MICINN) to J.A.A.M. and a grant from the National Science Foundation (Plant Genome
grant no. 1238243) to N.S.
www.plantphysiol.orgon February 14, 2018 - Published by Downloaded from Copyright © 2014 American Society of Plant Biologists. All rights reserved.

4
Abstract
The activity of SHOOT MERISTEMLESS (STM) is required for the functioning of the
shoot apical meristem (SAM). STM is expressed in the SAM but is downregulated at the
site of leaf initiation. STM is also required for the formation of compound leaves.
However, how the activity of STM is regulated at the transcriptional, post-transcriptional
and post-translational levels is poorly understood. We previously found two conserved
noncoding sequences (CNSs) in the promoters of STM-like genes across Angiosperms, the
K-box and the RB-box. Here we characterize the function of the RB-box in Arabidopsis
thaliana. The RB-box, along with the K-box, regulates the expression of STM in leaf
sinuses, which are areas on the leaf blade with meristematic potential. The RB-box also
contributes to restrict STM expression to the SAM. We identified FRS-RELATED
FACTOR1 (FRF1) as a binding factor to the RB-box region. FRF1 is an uncharacterized
member of a subfamily of four truncated proteins related to the FRS (FAR1-Related
Sequences) factors. Internal deletion analysis of the STM promoter identified a region
required to repress the expression of STM in hypocotyls. Expression of STM in leaf
primordia under the control of the JAGGED promoter produced plants with partially
undifferentiated leaves. We further found that the ELK domain has a role in post-
translational regulation of STM by affecting the nuclear localization of STM.
www.plantphysiol.orgon February 14, 2018 - Published by Downloaded from Copyright © 2014 American Society of Plant Biologists. All rights reserved.

5
Introduction
The shoot apical meristem (SAM) contains a group of undifferentiated cells from which
the aerial plant lateral organs derive. Several genes important for the functioning of the
SAM have been characterized (reviewed in Barton, 2010; Aichinger et al., 2012). In
Arabidopsis thaliana, WUSCHEL (WUS) regulates the formation and maintenance of the
SAM (Laux et al., 1996; Mayer et al., 1998) and acts in a negative feedback loop with
CLAVATA (CLV) genes. WUS promotes stem cell activity while CLV genes suppress this
activity (Fletcher et al., 1999; Brand et al., 2001; Schoof et al., 2001). SHOOT
MERISTEMLESS (STM), a class I KNOTTED-LIKE HOMEOBOX (KNOX1) gene, is
required for the maintenance of the SAM to prevent cell differentiation (Endrizzi et al.,
1996; Long et al., 1996). STM and WUS act in different pathways (Lenhard et al., 2002).
The KNOX1 genes KNAT1/BP (KNOTTED1-LIKE IN ARABIDOPSIS THALIANA/
BREVIPEDICELLUS), KNAT2 and KNAT6 show expression in the SAM and are partially
redundant with STM (Byrne et al., 2002; Scofield and Murray, 2006). CUP-SHAPED
COTYLEDON (CUC) genes demarcate the boundaries between emerging leaf primordia
and the SAM and are required for the initial expression of STM during embryo
development (Aida et al., 1997; Aida et al., 1999; Takada et al., 2001).
In addition to these regulatory genes, plant hormones such as cytokinins (CKs) and
gibberellins (GAs) play an important role in the activity of the SAM. CKs are required for
the maintenance of the meristem cells in an undifferentiated state, whereas GAs promote
differentiation. STM promotes cytokinin expression by activating the cytokinin
biosynthesis gene ISOPENTENYL TRANSFERASE7 (IPT7; Jasinski et al., 2005; Yanai et
al., 2005). Simultaneously, STM activates a GA deactivating gene At-GA2ox2 (Jasinski et
al., 2005) and represses the expression of the GA biosynthesis gene At-GA20ox1 (Hay et
al., 2002). In turn, increased CK levels enhance the expression of STM (Rupp et al., 1999;
Kurakawa et al., 2007).
www.plantphysiol.orgon February 14, 2018 - Published by Downloaded from Copyright © 2014 American Society of Plant Biologists. All rights reserved.

6
Leaves are formed on the flanks of the SAM. This process is characterized by the
accumulation of auxins at the site of leaf primordia initiation, the P0 site (Plastochron 0;
Benkova et al., 2003; Reinhardt et al., 2000), and by a down-regulation of STM (Long and
Barton, 1998). ASYMMETRIC LEAVES1/2 (AS1/2) are expressed in leaf primordia but the
activity of STM represses their expression in the SAM (Byrne et al., 2000).
STM also plays a role in the control of leaf morphology. Leaves of plants can be classified
as simple or compound, with simple leaves having a single lamina and compound leaves
having a lamina divided into leaflets. In simple-leaved species such as Arabidopsis, STM is
not expressed in leaf primordia, however STM expression is found in leaflet primordia of
compound-leaved species (Bharathan et al., 2002), with the exception of species within a
derived clade of legumes where the function of STM-like genes in generating leaflet
primordia is taken over by LEAFY-like genes (Bharathan et al., 2002; Champagne et al.,
2007; Hofer et al., 1997; Hofer et al. 2001). Furthermore, ectopic expression of STM-like
genes in compound-leaved species produces leaves with an increased degree of leaf
complexity (Hareven et al., 1996, Janssen et al., 1998). Conversely, a reduction in the
expression of STM-like genes in compound-leaved species reduces leaf complexity (Hay
and Tsiantis, 2006). The function of STM in leaf morphology is in part controlled by its
interaction with the truncated form of the KNOX1 factor PTS/KNATM
(PETROSELINUM/ KNOX ARABIDOPSIS THALIANA MEINOX) through competition
for the formation of dimers with BELL (BEL-like homeodomain) factors (Kimura et al.,
2008; Magnani et al., 2008). KNOX1 levels can also be controlled at the network level by
upstream factors in the PTS network module (Ichihashi et al., 2014).
Despite the importance of STM function in plant development, the regulation of STM
expression and the subsequent transcriptional, post-transcriptional and post-translational
controls that determine effective STM levels are poorly understood. Gene regulation at the
transcriptional level is mediated through the binding of transcription factors to cis-
regulatory sequences. These regulatory sequences are often embedded in conserved
noncoding sequences (CNSs), stretches of noncoding DNA that are evolutionary
preserved. Cis-regulatory sequences can also be located in introns. Importantly, intronic
sequences show direct regulation of KNOX1 genes: several dominant mutants in maize
www.plantphysiol.orgon February 14, 2018 - Published by Downloaded from Copyright © 2014 American Society of Plant Biologists. All rights reserved.

7
knotted1 (kn1) that confer ectopic expression of the gene are caused by events of
transposon insertions in intron 3 in a region particularly rich in CNSs (Greene et al., 1994;
Inada et al., 2003). In grasses, a CNS has been characterized in intron 2 of the class I
KNOX genes liguleless 3 (lg3), lg4a and lg4b (Bauer et al., 2004). In rice, Oryza sativa
homeobox1 (OSH1) positively regulates its own expression through binding to CNSs in
intron 3 (Tsuda et al., 2011). In barley, the hooded mutation increases the expression of
Barley knox3 (Bkn3) due to a duplication of 305 base pairs in intron 4 (Müller et al., 1995).
Yeast one-hybrid experiments showed a number of factors that bind this intronic region
(Santi et al., 2003; Osnato et al., 2010). In rice, exonic sequences of class I KNOX genes
also have regulatory functions (Ito and Kurata, 2008). In general, however, most of the cis-
regulatory sequences are located in the 5’ upstream region of the genes. We have
previously described two CNSs in the STM promoter, the K-box and the RB-box (Uchida
et al., 2007). Transgenic plants harboring a deletion in the K-box did not affect the
expression of STM in the SAM and its down-regulation in P0, but showed an expansion of
the STM expression pattern outside of these domains with consequences on leaf shape
(Uchida et al., 2007). We determined that class I TEOSINTE BRANCHED1-
CYCLOIDEA-PROLIFERATING CELL FACTOR1 (TCP) transcription factors can bind
to the K-box region (Aguilar-Martínez and Sinha, 2013). However, a role for the RB-box
in Arabidopsis has not been defined and other promoter regions important for STM
regulation have yet to be characterized.
The regulation of STM at the protein level has been investigated more extensively. STM
interacts with BELL factors through the MEINOX domain to access the nucleus and
regulate the transcription of target genes (Bellaoui et al., 2001; Smith et al., 2003; Bhatt et
al., 2004; Cole et al., 2006). The homeodomain (HD) is required for DNA binding (Smith
et al., 2002; Viola and Gonzalez, 2009) and intercellular trafficking (Lucas et al., 1995;
Kim et al., 2005; Winter et al., 2007; Bolduc et al., 2008). However the role of the ELK
domain is less understood though it is considered to be required for protein-protein
interaction (Vollbrecht et al., 1991; Kerstetter et al., 1994) and for nuclear localization
signal sequences (NLSs; Meisel and Lam, 1996). Cole et al. (2006) indicated that STM
does not contain a NLS and in the rice KNOX1 gene OSH15, the ELK domain is not
www.plantphysiol.orgon February 14, 2018 - Published by Downloaded from Copyright © 2014 American Society of Plant Biologists. All rights reserved.

8
required for nuclear localization, DNA binding, or homodimer formation, though it was
shown to have a role in suppressing transactivation activity (Nagasaki et al., 2001).
Here we analyze the regulation of STM through the study of promoter reporter constructs
and gene specific misexpression: we i) investigate the role of the RB-box in Arabidopsis,
ii) identify a binding factor to the RB-box region that belongs to a group of
uncharacterized proteins, iii) we delimit other important regions in the STM promoter for
the proper expression of the gene, iv) study the down-regulation of STM in P0 using a
gene-specific misexpression form of STM, and v) also show that the regulation of the
activity of STM in the cell nucleus is mediated by the ELK domain.
Results
The RB-box has a role in the control of STM regulation in Arabidopsis
We have previously shown that the STM gene promoter contains two CNSs, the K-box and
the RB-box (Uchida et al., 2007). The K-box is present in all species analyzed and has a
role in STM regulation (Uchida et al., 2007). A sequence analysis determined that the RB-
box is present in promoter regions of STM genes in all of the species we analyzed in this
study (Figure 1A; Uchida et al., 2007). In most of the species analyzed the RB-box is
located in a region around 500 base pairs (bp) upstream from the translation initiation
codon. However in the Brassicaceae species analyzed, the RB-box is located relatively far
from the translation initiation codon, ranging from -1412 bp in Cardamine hirsuta to -3484
bp in Thellungiella halophila (Fig. 1B).
In Arabidopsis thaliana STM expression is seen in the whole shoot apical meristem except
in the new leaf primordium initiation site (Long et al., 1996; Long and Barton, 2000). The
deletion of the core K-box expands the normal expression domain of STM to the base of
leaf primordia and the midvein of leaf petioles (Uchida et al., 2007). Through reporter
fusion experiments using the uidA gene, we wanted to determine expression patterns
produced by deletions of RB-box elements alone and in combination with deletions of
elements of the K-box (Fig. 2A; Table I). We analyzed 8 days-old seedlings of
representative T3 lines. A promoter fragment of 3379 bp (ProSTM:GUS) recapitulates the
www.plantphysiol.orgon February 14, 2018 - Published by Downloaded from Copyright © 2014 American Society of Plant Biologists. All rights reserved.

9
endogenous STM expression (Uchida et al., 2007; Figs. 2B and 2C). Plants transformed
with the construct ProSTM-ΔLK:GUS lack the large K-box and showed a GUS expression
pattern similar to the plants with just the construct ProSTM:GUS (Figs. 2D and 2E). In
older plants, GUS expression is found in the abaxial side of leaves in ProSTM-ΔLK:GUS
just as in constructs ProSTM-ΔCK:GUS that lack the core K-box (Supplemental Fig. 1A;
Uchida et al., 2007). In ProSTM-ΔCRB:GUS plants there is a deletion in the core RB-box
and the GUS expression pattern is expanded to the abaxial side of the growing leaves and
some expression is also seen in the hypocotyl (Figs. 2F and 2G). A similar pattern, except
for the expression in the hypocotyl, is found in plants with deletions in both the core RB-
box and the core K-box (ProSTM-ΔCRB-CK:GUS; Figs. 2H and 2I). When the large RB-
box is deleted (ProSTM-ΔLRB:GUS), the expression of GUS is detected in patches in the
hypocotyl and the root (Fig. 2J). Analysis of transverse sections indicated that GUS
expression in the pericycle corresponded with the two xylem poles (Fig. 2K). No GUS
expression is detected in the shoot apex. A combined deletion of the large RB-box and the
core K-box (ProSTM-ΔLRB-CK:GUS) produced an expression similar to the pattern of
GUS in hypocotyls seen in ProSTM-ΔLRB:GUS (Figs. 2L and 2N). Furthermore, GUS is
located in the sinuses, or spaces between lobes or teeth, of the growing leaves (Figs. 2L,
2M and 2O). Deleting both the large RB-box and the large K-box (ProSTM-ΔLRB-
LK:GUS; Figs. 2P and 2Q) showed a pattern of GUS expression similar to the construct
with both core RB and core K boxes deleted (Figs. 2H and 2I). GUS expression was not
evident in plants harboring constructs with deletions in the 5’ region of the STM promoter
up to the RB-box, using a native promoter (ProSTM-ΔRB:GUS) nor with the additional
deletion of the core K-box (ProSTM-ΔRB-CK:GUS; Table I; Supplemental Figs. 1B and
1C), indicating that regions further upstream from the RB-box are needed for wild type
expression of STM.
In summary, this promoter analysis shows that the CNS in the RB-box are required for
proper STM expression and that there is interaction between the RB-box and K-box
regulatory functions.
FRF1 binds the RB-box
www.plantphysiol.orgon February 14, 2018 - Published by Downloaded from Copyright © 2014 American Society of Plant Biologists. All rights reserved.

10
In order to gain insight into STM gene regulation, we used yeast one-hybrid screening to
identify factors that interact with the STM promoter region. We previously identified class
I TCP gene products binding to the K-box region (Aguilar-Martínez and Sinha, 2013).
Using a similar approach we wanted to study factors that bind the RB-box. For the yeast
one-hybrid analysis we used a fragment of the STM promoter containing the large RB-box
(-2133 to -1907 bp from the start codon). From the putative binding factors identified
(Supplemental Table I), we focused on At3g59470, which encodes for a putative DNA
binding factor (see below). We used an electrophoretic mobility shift assay (EMSA) and
determined that this factor interacts with the RB-box (Fig. 3A) and with the ΔCRB
fragment (representing a 17 bp deletion of the core, Supplemental Fig. 2). At3g59470 is
related to the FRS (FAR1-Related Sequences) genes, a family of transposase-derived
transcription factors (Hudson et al., 1999; Wang and Deng, 2002; Lin and Wang, 2004).
Well-known members of this family are FHY3 (FAR-RED ELONGATED HYPOCOTYL3;
Whitelam et al., 1993) and FAR1 (FAR-RED IMPAIRED RESPONSE1; Hudson et al.,
1999). We named At3g59470 FRF1 (FRS-RELATED FACTOR1). FRS factors are
characterized by having an N-terminal C2H2-type chelating motif of the WRKY-GCM1
family, a central core transposase domain of MULE transposases and a C-terminal SWIM
zinc-finger domain. The N-terminal domain acts as a DNA binding domain, while the C-
terminal domain is required for transcriptional factor activity (Lin et al., 2007). There are 4
FRF-like genes in the Arabidopsis genome. Interestingly, FRF1, FRF2, FRF3 and FRF4
only share the N-terminal motif with FRS proteins (Fig. 3B and Supplemental Data Set 1)
and form a well supported, though not well resolved, clade. We looked for FRF-like genes
in species other than Arabidopsis (Fig. 3C and Supplemental Fig. 3) and found that these
genes are represented across the dicots (tomato, Medicago truncatula, Vitis vinifera) and
monocots such as rice.
We wanted to study the function of FRF1 by analyzing insertion alleles for this gene. We
characterized one allele frf1-1 that had a tandem insertion located at -700 bp from the
putative start codon. Plants frf1-1 had the same phenotype as wild type plants. We then
made RNAi lines for FRF1 and these transgenic plants showed a normal phenotype.
Additionally, we studied an allele of FRF2, frf2-1, which had a tandem insertion at -140 bp
www.plantphysiol.orgon February 14, 2018 - Published by Downloaded from Copyright © 2014 American Society of Plant Biologists. All rights reserved.

11
from the putative start codon. The frf2-1 allele also showed no phenotypic defects when
compared with the wild type. To the best of our knowledge, no insertion alleles are
available in the coding region for FRF1 nor FRF2. Insertion alleles for FRF3 were not
analyzed in this study. For FRF4 we analyzed an allele named frf4-1 with an insertion at
338 bp from the putative start codon. The phenotype of frf4-1 plants was similar to the
wild type. The small size and low conservation between these gene sequences precludes
simultaneous knock-down by RNAi or amiRNA strategies.
We looked at the expression pattern of FRF genes in the Arabidopsis eFP browser at BAR
(http://bar.utoronto.ca/efp/cgi-bin/efpWeb.cgi; Winter et al., 2007), AtGenExpress
(http://jsp.weigelworld.org/expviz/expviz.jsp; Schmid et al., 2005) and GeneCAT
(http://genecat.mpg.de/899/genecat.html; Mutwill et al., 2008). FRF1 and FRF4 are
broadly expressed in Arabidopsis tissues, particularly in shoot apices (Supplemental Fig.
4). For FRF2 and FRF3 there are no data available in these databases. For these two genes
EST and cDNA matches are described in TAIR (The Arabidopsis Information Resource;
www.arabidopsis.org). According to the Cell eFP Browser (http://
bar.utoronto.ca/cell_efp/cgi-bin/cell_efp.cgi) FRF1 and FRF2 are located in the nucleus,
while FRF3 and FRF4 are present in the nucleus and cytoplasm.
In summary, we determined that FRF1 binds the RB-box and that FRF1 belongs to a small
family of factors phylogenetically conserved and FRS-related family of transcription
factors. Limited analysis of mutant alleles and RNAi lines suggests that these genes may
function redundantly.
Deletion analysis of the promoter of Arabidopsis STM reveals important regions for
its proper regulation
We further analyzed the role of other fragments of the gene promoter in regulating STM
expression. We divided the STM promoter into fragments of about the same size without
including the RB and K boxes, generated internal deletions, fused them to the uidA gene
and analyzed the GUS expression pattern in the transgenic plants from T3 lines (Fig. 4;
Table I). We divided the fragment upstream of the RB-box into three parts, F1, F2 and F3
of 335 bp each one, the fragment between both RB and K-box in 4 parts, F4 to F7 of 388
www.plantphysiol.orgon February 14, 2018 - Published by Downloaded from Copyright © 2014 American Society of Plant Biologists. All rights reserved.

12
bp each one, and we used the fragment F8 between the K-box and the starting codon of
220 bp (Fig. 4A). Compared to the normal expression of STM in the shoot apex (Long et
al., 1996; Long and Barton, 2000; Uchida et al., 2007; Figs. 2B and 2C), the deletion of the
fragment F1 (ProSTM-ΔF1:GUS) showed no changes in the shoot apex localization of
GUS (Figs. 4B and 4C). Deleting the fragment F2 (ProSTM-ΔF2:GUS) lightly expanded
the GUS expression pattern through the hypocotyls (Figs. 4D and 4E). When the F3
fragment is removed (ProSTM-ΔF3:GUS) a strong GUS signal was observed throughout
the hypocotyl, while no expression was found in the shoot apex (Fig. 4F). Analysis of
cross-sections of the hypocotyls showed staining markedly confined to the stele (Fig. 4G).
Light expression in the shoot apex and the hypocotyls was found in constructs that lack the
F4 fragment (ProSTM-ΔF4:GUS, Figs. 4H and 4I) or the F5 fragment (ProSTM-ΔF1:GUS;
Figs. 4J and 4K). An expression similar to the native promoter was found in plants with the
F6 fragment deleted ProSTM-ΔF6:GUS (Figs. 4L and 4M). GUS expression in plants with
the construct ProSTM-ΔF7:GUS (Figs. 4N and 4O) was similar to the constructs ProSTM-
ΔF4:GUS and ProSTM-ΔF5:GUS. The deletion of the F8 fragment (ProSTM-ΔF8:GUS)
produced an expansion of GUS expression pattern mainly to the abaxial side of growing
leaves and petioles (Figs. 4P and 4O).
We also looked for conserved regions other than the K-box and RB-box in the promoters
of STM-like genes. We focused on the Brassicaceae species with sequenced genomes B.
rapa, T. halophila, C. rubella, A. thaliana, A. lyrata and C. hirsuta and in regions from the
RB-box up to -6000 bp from the translation initiation codon (Fig. 5A). Three regions Fa,
Fb and Fc appear greatly conserved. Fa is the most distant (Figs. 5A and 5B), Fb is located
between fragments F1 and F2 (Figs. 5A and 5C), while Fc is located in the fragment F3
close to the RB-box (Figs. 5A and 5D). The functional significance of these conserved
sequences remains to be elucidated.
We also studied the sequence conservation in intronic regions of STM-like genes in the
Brassicaceae species analyzed. Interestingly, sequences of introns I and II are greatly
conserved between species, while sequences of the intron III are poorly preserved
(Supplemental Fig. 5A). In comparison, sequences of introns I, II and IV of BP-like genes
www.plantphysiol.orgon February 14, 2018 - Published by Downloaded from Copyright © 2014 American Society of Plant Biologists. All rights reserved.

13
are not conserved and only one fragment of intron III is well conserved between species
(Supplemetal Fig. 4B).
From this analysis we determined that the sequences in the F3 fragment greatly contribute
to the correct expression of STM, while the role of the other fragments analyzed appears to
be less prominent. Sequence conservation analysis suggests that further upstream
sequences as well as introns I and II may also harbor STM regulatory sequences.
STM expression in initiating leaf primordia
One of the most striking features of STM expression is its down-regulation in the incipient
leaf primordium, the P0. The P0 is characterized as the region containing a group of leaf
founder cells located at the flanks of the SAM. The down-regulation of STM at P0 is
required for the formation of new leaf primordia in a process that appears to be regulated at
the transcriptional level (Long et al., 1996; Long and Barton, 2000). We wanted to
determine the developmental consequences of expression of STM in P0, a site where the
gene is normally down-regulated. Based on mRNA in situ hybridization, the JAGGED
(JAG) gene is expressed in P0 (Dinneny et al., 2004; Ohno et al., 2004). JAG is also
expressed in young leaf primordia. In reproductive development JAG is expressed in all
flower organ primordia (Dinneny et al., 2004; Ohno et al., 2004). We found that 2.2 Kb of
the 5’ region of JAG recapitulates the endogenous expression of the gene (Figs. 6A, 6B
and Supplemental Fig. 6). We generated transgenic plants harboring 2.2 Kb of the JAG
promoter fused to the coding sequences of GFP and STM (ProJAG:GFP-STM) and
analyzed the phenotype (Figs. 6C-H). We expected that expression of STM in P0 would
lead to defects in leaf primordia initiation, as STM promotes the maintenance of the cells in
an undifferentiated stage. Compared to the wild type (Fig. 6C), the transgenic plants
ProJAG:GFP-STM recovered with a strong phenotype were of reduced size and with
epinastic cotyledons (Fig. 6D). Although leaf primordia emerged and show characteristics
of differentiation such as the presence of leaf trichomes, leaf growth and expansion were
arrested (Fig. 6D). These transgenic plants do not produce seeds. Transgenic plants with a
milder phenotype had bigger rosettes and small highly lobed leaves with broad petioles
(Figs. 6E and Supplemental Fig. 7A). Ectopic meristems were not observed on these
www.plantphysiol.orgon February 14, 2018 - Published by Downloaded from Copyright © 2014 American Society of Plant Biologists. All rights reserved.

14
leaves. We used confocal microscopy to characterize GFP distribution in the transgenic
plants generated. In transgenic plants carrying ProSTM:GFP-STM GFP is located in the
SAM (Fig. 6 F and Supplemental Figs. 7B and 7C), similar to ProSTM:STM-VENUS
plants that use VENUS instead of GFP (Heisler et al., 2005). However in plants with STM
under the control of JAG promoter we observed a proliferation of stipule-like structures
that highly express GFP (Fig. 6G and Supplemental Figs. 7D-F). In ProJAG:GFP-STM
plants with less severe phenotype, GFP signal was more distributed throughout the base of
developing leaf primordia (Fig. 6H) and later in development stipule-like structures were
also observed (Supplemental Figs. 7G-I). These results indicate that ectopic expression of
STM in early leaf primordia disrupts normal development of these primordia.
The ELK domain is required for correct nuclear localization and activity of STM
The STM protein is characterized by having four well conserved domains, MEINOX,
subdivided into KNOX1 and KNOX2 subdomains, GSE, ELK and HD (Fig. 7A;
Vollbrecht et al., 1991; Kerstetter et al., 1994; Nagasaki et al., 2001). Despite the
conservation of the ELK domain, its role is poorly understood. To study the function of
this domain, we focused on the serine residue at position 272 (Supplemental Fig. 8). Serine
residues are a common target of protein post-translational modification through
phosphorylation and in addition serine residue at position 272 lies in a putative 14-3-3
protein binding site (http://scansite.mit.edu). We performed a site-directed substitution of
the S-272 to an alanine in ProSTM:GFP-STM (Uchida et al., 2007) to generate transgenic
plants ProSTM:GFP-STM-S272A and analyzed the phenotype. Compared with control
plants ProSTM:GFP-STM that behave as wild type (Uchida et al., 2007), plants carrying
ProSTM:GFP-STM-S272A were characterized by having lobed rosette leaves (Fig. 7B) and
highly lobed cauline leaves (Fig. 7C), while other organs were not affected (Fig. 7D). This
phenotype was different from plants Pro35S:STM-GR that ectopically express STM in a
inducible manner using the glucocorticoid receptor (GR) and are characterized by
producing multiple and very small leaves and ectopic stipules (Gallois et al., 2002; Cole et
al., 2006). Interestingly, plants that express both the BELL factor BLH9 (Pro35S:BLH9)
that interacts with STM to direct the complex to the cell nucleus and STM factor fused to
www.plantphysiol.orgon February 14, 2018 - Published by Downloaded from Copyright © 2014 American Society of Plant Biologists. All rights reserved.

15
GR (Pro35S:STM-GR; Cole et al., 2006) have deeply lobed leaves, resembling
ProSTM:GFP-STM-S272A plants. Furthermore, transgenic plants that overexpress STM
fused to a NLS (Pro35S:NLS-GFP-STM; Cole et al., 2006) also have a similar phenotype
to plants ProSTM:GFP-STM-S272A. We then investigated if the targeted mutation STM-
S272A had an altered nuclear localization of STM which could correlate with the observed
phenotype. We transiently expressed the constructs GFP-STM and GFP-STM-S272A in
Nicotiana benthamiana leaves and observed the localization of STM using confocal laser
scanning microscopy. For the control construct GFP-STM the STM protein was found to
be located in the cytoplasm as previously described (Fig. 7E; Cole et al., 2006). However
the location of STM in STM-S272A was mainly nuclear (Fig. 7F). Taken together these
results suggest that the Serine 272 residue in the ELK domain has a role in the control of
nuclear localization of STM.
Nuclear localization of BELL factors depends on interaction with the nuclear export
receptor AtCRM1/AtEXPO1 (CHROMOSOME REGION MAINTENANCE1/
EXPORTIN1) through the BELL domain, which harbors both a nuclear export signal
(NES) and the STM-binding site (Rutjens et al., 2009). Treatment with leptomycin B
(LMB) specifically inhibits CRM1 activity and BELL factors accumulate in the nucleus
(Rutjens et al., 2009). We found that in N. benthamiana leaves, treatment with LMB also
determined the nuclear accumulation of STM (Supplemental Fig. 9). However, in a yeast
two-hybrid assay, AtCRM1 does not interact with STM (Rutjens et al., 2009) suggesting
that yeast endogenous factors might interfere with this interaction, or other plants factors
might be required for the interaction.
In conclusion, our results highlight the importance of the promoter region RB-box in STM
regulation. The RB-box is required for the expression of STM in the SAM and works with
the K-box in controlling STM expression in leaves. A putative transcription factor FRF1
from a previously uncharacterized subfamily of FRS proteins shows binding to the RB-
box. An initial scanning for important regulatory regions in STM other than the K-box and
the RB-box showed that the region F3 is required to repress the expression of STM in
hypocotyls. The expression of STM in leaf primordia using the JAG promoter does not
completely arrest leaf initiation, suggesting either that expression of STM in the P0 domain
www.plantphysiol.orgon February 14, 2018 - Published by Downloaded from Copyright © 2014 American Society of Plant Biologists. All rights reserved.

16
does not have drastic consequences on leaf initiation or that STM is downregulated post-
transcriptionally in this domain. We also show that the ELK domain is required for a
correct post-translational regulation of STM. Combined, these results show that the
regulation of STM expression is complex with interplay between several promoter binding
TFs like the TCPs, FRFs and other as yet unknown factors in the regions of the promoter
upstream from the RB-box and between the RB- and K-boxes. Further analysis of
subcellular STM protein localization and protein levels in the P0 region of ProJAG:STM
plants is needed to determine the role post-transcriptional regulation plays in determining
STM expression in this domain.
Discussion
Role of the RB-box in STM transcriptional regulation
In Arabidopsis STM expression is normally restricted to the SAM. Our promoter deletion
analyses have shown that in certain constructs lacking the RB-box or both the RB-box and
the K-box, the expression of STM is expanded to the abaxial side of leaves. Interestingly,
the endogenous STM promoter of a species with dissected leaves, such as Cardamine
hirsuta, drives a similar expression pattern in either C. hirsuta or in Arabidopsis, a simple-
leaved species (Hay and Tsiantis, 2006). These authors proposed that the differences in
STM expression between species, and therefore in leaf morphology, can be due to
differences in regulatory regions of the STM promoters, reinforcing a role for the RB-box
and the K-box in this process. Although both species possess RB-box and K-box, subtle
sequence differences, including different spacing between regulatory sequences or
different response of trans-acting factors can account for different promoter activity.
We found that transgenic plants harboring a construct with a deletion of both the large RB-
box and the core K-box showed strong GUS expression in leaf sinuses. KNOX1 genes can
be expressed in leaf sinuses. For instance, in mutants as1-1 and as2-2 the genes KNAT1/BP
and KNAT2 are expressed in the sinuses (Ori et al., 2000). This expression is enhanced in
double mutants as1-1 se (serrate), as2-2 se and as1-1 axr1-3 (auxin resistant1-3) where
deeper sinuses are formed (Ori et al., 2000; Hay et al., 2006). In these double mutants,
www.plantphysiol.orgon February 14, 2018 - Published by Downloaded from Copyright © 2014 American Society of Plant Biologists. All rights reserved.

17
ectopic structures resembling stipules are formed (Ori et al., 2000). STM has been found to
be expressed at low rate and intensity in leaf sinuses, inferred by the analysis of a construct
with the uidA gene fused to the native STM promoter (ProSTM:GUS; Kawamura et al.,
2010). STM is required for leaf serration (Depuydt et al., 2008; Kawamura et al., 2010).
CUC2 is also expressed in leaf sinuses (Nikovics et al., 2006) and mutants with increased
expression of CUC2 show expression of STM as small spots in some sinuses where stipule-
like organs are often observed (Kawamura et al., 2010). Leaf sinuses can be considered as
quiescent domains with the potential for meristem formation (Hu et al., 2011). In fact,
certain Kalanchoë species form plantlets on leaf sinuses (Garces et al., 2007). Our results
stress the importance of both the RB-box and the K-box in determining leaf shape through
regulation of leaf lobes and sinuses.
We have determined that different combinations of deletions of the RB-box and K-box
render different GUS expression patterns. For instance, the expression pattern found when
we combined deletions in the large RB-box and the core K-box is abolished when we used
the large RB-box and the large K-box. This indicates that other sequences close but
different to the core sequences are also important in STM regulation.
Transcription regulation operates in part through DNA bending that closely places
regulatory factors located relatively distant to the core promoter with the RNA polymerase.
It is possible that factors that bind the RB-box and the K-box participate in this bending
mechanism and that specific factors can cause interactions between these sites.
FRF proteins as putative binding factors at the RB-box
Through yeast one-hybrid and EMSA we determined that FRF1 is a putative binding factor
at the RB-box. This factor defines a group of uncharacterized proteins related to the FRS
transposase-derived transcription factors. Interestingly, FRF factors are conserved across
dicots, suggesting a conserved role for them in plant development. Further, FRF factors are
short or truncated versions of FRS factors, with only the N-terminal C2H2 zinc finger
domain sequence conserved. This domain is essential for direct DNA binding (Lin et al.,
2008). One possibility is that FRF factors could regulate the action of FRS factors through
competitive binding for the same DNA binding motifs. HY5 (LONG HYPOCOTYL5)
www.plantphysiol.orgon February 14, 2018 - Published by Downloaded from Copyright © 2014 American Society of Plant Biologists. All rights reserved.

18
interacts with FHY3 and FAR1, interfering with their DNA binding capabilities (Li et al.,
2010). Interestingly, these interactions are mediated by the N-terminal C2H2 of FHY3 and
FAR1. Also FHY3 interacts with CCA1 (CIRCADIAN CLOCK-ASSOCIATED1), LHY
(LATE ELONGATED HYPOCOTYL) and PIF1 (PHYTOCHROME-INTERACTING
FACTOR1) through the N-terminal C2H2 domain (Li et al., 2011; Tang et al., 2012). Thus
another possibility is that FRF factors could interact with factors other than FRS factors.
Further functional studies on FRF factors would help to analyze these possibilities.
The F3 region as controller of the stem and root expression of STM
Our promoter deletion analysis showed that deletion of certain promoter fragments leads to
STM expression located at the pericycle. This is particularly noticeable for the fragment F3
and is also found in part in constructs without the large RB-box. The RB-box and the
fragment F3 are adjacent, indicating that sequences from this region, in particular from F3,
are important to repress the expression of STM in the pericycle. The pericycle is a primary
mersitem tissue and a place for shoot regeneration (Atta et al., 2009). In plants that have
secondary growth, tissue of the pericycle contributes to the development of the vascular
cambium. Furthermore, the pericycle has the capacity to produce lateral roots. Interestingly
the pattern of lateral root distribution is altered in the mutant stm-1 (Casimiro et al., 2001).
Indeed, the STM orthologs in tomato LeT6/TKN2 (Lycopersicum esculentum T6/ Tomato
Knotted2) and in Medicago truncatula Mt-knox-1 are found to be expressed in lateral roots
(Koltai and Bird, 2000; Koltai et al., 2001). The class I KNOX gene KNAT6 acts
redundantly with STM in SAM maintenance and is expressed in roots (Belles-Boix et al.,
2006; Dean et al., 2004). Similarly KNAT1 and the class II KNOX genes KNAT3, KNAT4
and KNAT5 show root specific tissue expression (Truernit et al., 2006). Furthermore
KNAT1 has a specific role in root skewing (Qi and Zheng, 2013).
Arabidopsis STM expression is found not only in the SAM but in stem tissue although not
uniformly (Long et al., 1996). In tomato LeT6 is expressed in the SAM and in vascular
tissue and in poplar the KNOX gene ARBORNOX1 (ARK1) is expressed in the SAM and in
the vascular cambium (Groover et al., 2006). Transcripts from the STM-like gene of the
parasitic plant Cuscuta pentagona are found in the haustoria, a specialized organ derived
www.plantphysiol.orgon February 14, 2018 - Published by Downloaded from Copyright © 2014 American Society of Plant Biologists. All rights reserved.

19
from stems (Alakonya et al., 2012). However stem-tissue expression is more characteristic
of KNAT1-like genes, e.g., in Arabidopsis KNAT1 (Lincoln et al., 1994), tomato TKN1
(Hareven et al., 1996), poplar ARK2 (Du et al., 2009), maize kn1 (Smith et al., 1992;
Jackson et al., 1994) or Prunus persica KNOPE1 (Testone et al., 2012), with these genes
having a more prominent role in regulation of stem tissue differentiation (Townsley et al.,
2013).
We have also located three CNSs in the promoter of STM-like genes upstream of the RB-
box. It will be interesting to analyze if Fa has some function in the regulation of STM
expression. Fb could be the place for enhancer sequences, as constructs with deletions in
this region ProSTM-ΔF1:GUS and ProSTM-ΔF2:GUS have decreased GUS signal in shoot
apices. Fc, included in the F3 fragment, could contain the sequences for the repression of
STM expression in hypocotyls. Functional analyses will determine these possibilities.
Likewise, functional analyses of ProSTM-ΔCRB:STM and ProSTM-ΔF3:STM transgenic
lines will help to understand the roles of RB- and F3-boxes in SAM and leaf development.
Expression of STM in leaf primordia and mechanisms for regulation of STM
expression
Leaves of plants expressing ProJAG:STM with a weak phenotype resemble the leaves of
plants expressing Pro35S:KNAT1 or Pro35S:kn1 (Chuck et al., 1996; Lincoln et al., 1994)
while plants expressing ProJAG:STM with a strong phenotype resemble the phenotype of
plants expressing Pro35S:STM (Gallois et al., 2002; Williams, 1998) or plants that express
STM or Kn1 in a glucocorticoid (GR) inducible system (Pro35S:STM-GR, Pro35S:kn1-
GR) grown from germination on dexamethasone containing media (Gallois et al., 2002;
Hay et al., 2003). AINTEGUMENTA (ANT) is expressed in leaf primordia, along with
expression in other tissues (Elliott et al., 1996; Long and Barton, 2000). Plants expressing
ProANT:STM have a similar phenotype to plants ProJAG:STM (Lenhard et al., 2002). This
would suggest that the expression of STM at the site of presumptive leaf primordia
initiation can still produce rudiments of lateral organs although with suppression of cell
differentiation. Leaf initiation at P0 is characterized by a down-regulation of STM (Long
and Barton, 2000) and by the accumulation of auxin (Benkova et al., 2003; Reinhardt et
www.plantphysiol.orgon February 14, 2018 - Published by Downloaded from Copyright © 2014 American Society of Plant Biologists. All rights reserved.

20
al., 2000), indicating that these processes can be functionally connected. PIN-FORMED1
(PIN1) and PINOID (PID) regulate polar auxin transport and in double mutant embryos
pin1-3 pid-2 the expression of STM is expanded to the cotyledon area, resulting in growth
inhibition of cotyledon primordia, indicating that both PIN1 and PID repress STM
(Furutani et al., 2004).
CUC1 and CUC2 are required for the establishment of organ boundaries in the SAM (Aida
et al., 1997; Aida et al., 1999). In double mutants cuc1 cuc2 the expression of STM is
abolished, indicating that both CUC1 and CUC2 could act as initiators of STM
transcription (Aida et al., 1999). Ectopic expression of CUC1 (Pro35S:CUC1) activates
the expression of STM, producing adventitious shoots at the sinuses of cotyledons. These
shoots are not formed in the stm mutant background (Hibara et al., 2003). However is not
known whether CUC1 and CUC2 directly or indirectly regulate STM. Also single mutants
for PINHEAD (PNH) and the related gene ARGONAUTE1 (AGO1) have defective apical
meristems although expression of STM is detected, however in the double mutant pnh ago1
the expression of STM is absent (Lynn et al., 1999).
Long-term repression of STM in leaves appears to be mediated by the action of Polycomb-
repressive complex2 (PRC2) and PRC1-like. CURLY LEAF (CLF) and SWINGER (SWN)
are members of PRC2. CLF directly binds the promoter of STM and double mutants clf
swn ectopically express STM in leaves (Schubert et al., 2006). Mutations in members of
PRC1-like also missexpress KNOX1 genes (Xu and Shen, 2008). It has been shown that
the leaf adaxial determinants AS1-AS2 interact with members of the PRC2 to mediate
long-term repression of KNOX1 genes KNAT1, KNAT2 and PTS/KNATM but not STM
(Lodha et al., 2013). Histone Deacetylase HDA6 participates in a complex with AS1 to
repress KNAT1, KNAT2 and PTS/KNATM (Luo et al., 2012). Class II TCP factors bind to
the promoters of KNAT1and KNAT2 and interact with AS2 to exert a repressive function
(Li et al., 2012). We determined that class I TCP factors can bind the STM promoter
through the K-box region (Aguilar-Martínez and Sinha, 2013). As regulation of STM
appears to not be mediated by the AS1-AS2 complex, it will be interesting to determine
which TCP interacting factors other than AS1-AS2 act in the regulation of STM.
www.plantphysiol.orgon February 14, 2018 - Published by Downloaded from Copyright © 2014 American Society of Plant Biologists. All rights reserved.

21
Leaf abaxial polarity determinant YABBY factors FILAMENTOUS FLOWER (FIL) and
YAB3 have a role in the regulation of KNOX1 genes STM, KNAT1 and KNAT2, as double
mutants fil yab3 derepress the expression of these genes in leaves, generating ectopic
meristems (Kumaran et al., 2002). YABBY factors interact with transcriptional
corepressors LEUNIG (LUG) and LEUNIG_HOMOLOG (LUH), as well as with the
LUG/LUH transcriptional adaptor factors SEUSS (SEU) and SEUSS-LIKE (SLKs; Stahle
et al., 2009). In mutants lug -/- luh +/- the expression pattern of STM is expanded (Stahle et
al., 2009), while in mutants seu slk2 the expression levels of STM are greatly reduced (Bao
et al., 2010). In turn, LUG interacts with HDA19 and the Mediator components MED14
and CDK8 (Gonzalez et al., 2007). Moreover, mutant med14 shows an irregular expression
pattern of STM in the SAM (Autran et al., 2002). BASIC PENTACYSTEINE (BPC)
factors bind a promoter region in STM right upstream of the ATG as well as other
promoter regions in several other HOMEOBOX genes (Simonini and Kater, 2014). In rice,
OSH1 is positively autoregulated mainly through CNSs in introns (Tsuda et al., 2011). A
similar mechanism can operate in Arabidopsis STM, although CNSs similar in structure to
the RB-box and K-box are not described in introns of the STM-like genes from the species
analyzed in this study. Thus, a number of factors have a role in STM regulation yet a
comprehensive framework of control of STM gene regulation at transcriptional level
remains to be determined.
Role of the ELK domain in post-translational regulation of STM
The targeted mutation STM-S272A could affect the interaction with STM binding proteins
and hence correct nuclear localization and function. The ELK domain has been suggested
to mediate protein-protein interactions (Mushegian and Koonin, 1996; Sakamoto et al.,
1999). Nagasaki et al. (2001) showed that the ELK domain is not essential for homodimer
formation in rice KNOX factors. The interaction of STM with BELL factors is mediated
by the MEINOX domain (Bellaoui et al., 2001; Smith et al., 2002; Bhatt et al., 2004). It
will be interesting to determine if the activity of STM-S272A and its associated phenotype
is independent on the interaction with BELL factors in vitro and in vivo. Several members
of the OVATE family of factors (AtOFPs) interact with KNOX and BELL factors
www.plantphysiol.orgon February 14, 2018 - Published by Downloaded from Copyright © 2014 American Society of Plant Biologists. All rights reserved.

22
regulating their subcellular localization (Hackbusch et al., 2005). BLH1, BLH7 and
KNAT1 interact with AtOFP1 (Hackbusch et al., 2005), BLH1 and KNAT3 with AtOFP5
(Pagnussat et al., 2007) and KNAT7 with OFP1 and OFP4 (Li et al., 2011). The interaction
of KNAT7 is mediated by the homeodomain (Li et al., 2011). However STM shows no
interaction with AtOFPs (Hackbusch et al., 2005).
Experiments with overexpression of the rice KNOX protein OSH15 carrying a deletion of
the ELK domain produced a novel phenotype in transgenic rice plants, suggesting that this
domain could have a defined function, such as interaction with a specific factor (Nagasaki
et al., 2001). The serine residue used for the targeted mutation in this study lies in a
putative 14-3-3 interacting binding region (http://scansite.mit.edu). Other regions of STM
protein are also putative binding sites for 14-3-3 factors. The role of these interactions is
under study. While the role of STM conserved domains KNOX1, KNOX2, HD and ELK
has been more extensively studied, the proposed function of the GSE domain in regulating
protein degradation (Volbretch et al., 1991; Nagasaki et al., 2001) has not been yet
determined.
In conclusion, while many aspects concerning the control of STM transcriptional regulation
and correct protein localization and stability are still unknown, our findings have revealed
regions and factors important for the correct regulation of STM, a transcription factor with
important roles in plant development.
Materials and methods
Plant materials and growth conditions
Allele frf1-1 (SAIL_292_C04) in Arabidopsis thaliana was obtained from the Syngenta
Arabidopsis Insertion Library (SAIL) (Sessions et al., 2002). Alleles frf2-1
(SALK_076709) and frf4-1 (SALK_048606C) were obtained from the Salk Institute
Genomic Analysis Laboratory (SIGnAL) (Alonso et al., 2003) through the Arabidopsis
Biological Resource Center (ABRC). The mutant alleles were in Columbia-0 (Col-0)
ecotype background. Growth conditions were as described in Aguilar-Martínez and Sinha
(2013).
www.plantphysiol.orgon February 14, 2018 - Published by Downloaded from Copyright © 2014 American Society of Plant Biologists. All rights reserved.

23
Phylogenetic analysis and protein sequence analysis
For the alignment of the RB-box we used the promoter regions of Arabidopsis thaliana
STM (At1g62360.1) and the putative orthologs in A. lyrata gene 338146; Capsella rubella
gene Carubv10022235m.g; Brassica rapa gene Bra027050; Thellungiella halophila gene
Thhalv10023520m; Carica papaya evm.TU.supercontig_152.57; Populus trichocarpa
gene Potri.011G011100; Vitis vinifera gene GSVIVG01004811001; Solanum lycopersicum
gene LeT6 (Solyc02g081120; Janssen et al., 1998); Medicago truncatula gene Mt-knox-1,
Medtr5g085860 (Koltai et al., 2001); Glycine max gene Glyma09g01000; Phaseolus
vulgaris gene Phvul.006G145800 and Ricinus communis gene 29842.t000028. All
sequences were obtained from Phytozome (http://www.phytozome.net/; Goodstein et al.,
2012). For cotton (Gossypium raimondii) we used the geneBank accession number
CO081267, gene Gorai.010G183800; for Antirrhinum majus, geneBank accession number
AY072735.1; for Nicotiana tabacum, gene NTH15 (Tamaoki et al., 2007); for Pisum
sativum, gene Hop1, geneBank accession number AF063307.1 (Giles et al., 1998); for
Cardamine hirsuta, gene C. hirsuta STM (Hay and Tsiantis, 2006); for Asparagus
officinalis, geneBank accession number EF494254 (Uchida et al., 2007).
The sequence alignments were made using Muscle (http://www.ebi.ac.uk/Tools/msa/
muscle/) and Chaos+Dialign (http://dialign.gobics.de/chaos-dialign-submission; Brudno et
al., 2004). The alignment was edited using Mega5 software
(http://www.megasoftware.net/; Tamura et al., 2011) and displayed using Boxshade
version 3.31 in the Mobyle portal (http://mobyle.pasteur.fr; Néron et al., 2009).
The analysis of CNSs in the STM promoter of Brassicaceae species upstream of the RB-
box was made with mVISTA (http://genome.lbl.gov/vista/index.shtml; Frazer et al., 2004).
BP-like genes were Bra000638 in B. rapa, Thhalv10028724m.g in T. halophila,
Carubv10002748m.g in C. rubella and 489727 in A. lyrata. The protein domains of FRF
and FRS factors were analyzed with InterPro (http://www.ebi.ac.uk/interpro/; Hunter et al.,
2011) and drawn using DOG (Domain Graph, version 2.0.1; http:
//dog.biocuckoo.org/index.php; Ren et al., 2009).
www.plantphysiol.orgon February 14, 2018 - Published by Downloaded from Copyright © 2014 American Society of Plant Biologists. All rights reserved.

24
For the analysis of FRF-like proteins, blastp searches were made against genomes of
tomato (ITAG 2.3 release of the sequenced tomato genome,
http://solgenomics.net/organism/Solanum_lycopersicum/ genome), Medicago truncatula
(MtPep-Medicago predicted proteins (Mt3.5) JCVI Medicago truncatula Genome Project
http://www.jcvi.org/cgi-bin/medicago/overview.cgi), Vitis vinifera and rice, both at
Phytozome (http://www.phytozome.net/). The alignments were made with TanslatorX
(Abascal et al., 2010; http://translatorx.co.uk/) using Muscle (http://www.
ebi.ac.uk/Tools/msa/muscle/), see Supplemental Data Set 2 for the alignment. We used the
conserved FAR1 DNA binding domain of the FRF-like proteins to generate a neighbor
joining tree and a parsimony tree with 1000 bootstrap replications. The alignments were
done on the translated amino acid sequence. Only unambiguously alienable portions of the
sequence that were present in all sequences were used for the tree. The region used was
columns 821-873 and 895-933 and all other positions were excluded. Two rice sequences
from the alignment were deleted because even though they were FAR sequences, the
alignment algorithm could not align them unambiguously to infer the relationships to the
other sequences. The phylogenetic trees were generated with PAUP* 4.0 (Swoford D.L.,
2003; paup.csit.fsu.edu) and edited with FigTree v1.4.2 (http://tree.bio.ed.ac.uk/software
/figtree/). Sequences of factors LeT6 (AAC49917.1) in tomato, NTH15 (BAA25546.1) in
tobacco and STM (AAC49148.1) in Arabidopsis were aligned with ClustalW2
(http://www.ebi.ac.uk/Tools/msa/clustalw2/).
Construction of transgenes and plant�transformation
The constructs with selected internal deletions in the STM promoter were generated using
overlapping PCR (Heckman and Pease, 2007). As templates we used constructs with
fusions of the gene uidA with the native STM promoter (ProSTM:GUS, 3379 bp; Uchida et
al., 2007) or the native STM promoter without the K-box (ProSTM-ΔK:GUS; Uchida et al.,
2007). The flanking master primers were STMpFH2 and STMpRBHI. The internal primers
were respectively for ProSTM-ΔLK:GUS, Lkb and Lkc, for ProSTM-ΔCRB:GUS, STMb2
and STMc2, for ProSTM-ΔCRB-CK:GUS, STMb2 and STMc2 on ProSTM-ΔK:GUS as
template, for ProSTM-ΔLRB:GUS, STMb1 and STMb2, for ProSTM-ΔLRB-CK:GUS,
www.plantphysiol.orgon February 14, 2018 - Published by Downloaded from Copyright © 2014 American Society of Plant Biologists. All rights reserved.

25
STMb1 and STMc1 on ProSTM-ΔK:GUS as template, for ProSTM-ΔLRB-LK:GUS,
STMb1 and STMc1 on ProSTM-ΔLK:GUS as template. For ProSTM-ΔF1:GUS, F1b and
F1c, for ProSTM-ΔF2:GUS, F2b and F2c, for ProSTM-ΔF3:GUS, F3b and F3c, for
ProSTM-ΔF4:GUS, F4b and F4c, for ProSTM-ΔF5:GUS, F5b and F5c, for ProSTM-
ΔF6:GUS, F6b and F6c, for ProSTM-ΔF7:GUS, F7b and F7c, and for ProSTM-ΔF8:GUS,
F8b and F8c. For ProSTM-ΔRB:GUS and ProSTM-ΔRB-CK:GUS, we used the pair of
primers STMpRBHIf and STMpRBHI on ProSTM:GUS and ProSTM- ΔK:GUS as
templates, respectively.
For the RNAi construct for FRF1, a fragment of 200 bp of the coding sequence was
amplified using primers FARRNAf (5’-AACGAACACGACAAAATTCAGGAACTGA-
3’) and FARRNAr (5’-CTATACTTGGTGGTTCTGTTGATGATCT-3’) and cloned into
pCR8⁄GW⁄TOPO TA entry vector (Invitrogen). We used pK7GWIWG2(II) (Karimi et al.,
2002) as destination vector.
To generate plants ProJAG:GUS, a 2.2 Kb fragment at 5’of JAGGED in Col-0 ecotype
was isolated using oligos JAGpEcoRIF and JAGpBHIR. The fragment was LR cloned into
pkGWFS7 (Karimi et al., 2002). T3 lines were analyzed. For the generation of plants
ProJAG:GFP-STM, we first isolated a fragment GFP6-STM-T35S from the construct
STMp-STM (Uchida et al., 2007) using oligos GF6FBHI and T35SRBHI. This fragment
was cloned into an entry vector containing the promoter of JAG and LR cloned into the
plasmid pMDC123 with Basta selection in planta (Curtis and Grossniklaus, 2003). Col-0
ecotype plants were transformed with the ProJAG:GFP-STM construct.
Generation of GFP-STM was previously described (Uchida et al., 2007). S272A mutation
was introduced by PCR-based method and the resultant GFP-STM-S272A fragment was
cloned into pCR8/GW/TOPO vector. These constructs served as the entry vectors to
transfer protein coding regions into the pMDC32 vector containing 2 x CaMV 35S
promoter through LR cloning reaction (Invitrogen; Curtis and Grossniklaus, 2003).
ProSTM:GFP-STM expression cassette in pMDC123 was previously reported (Uchida et
al., 2007) and construction of pMDC123/ProSTM:GFP-STM-S272A was carried out using
the same method.
www.plantphysiol.orgon February 14, 2018 - Published by Downloaded from Copyright © 2014 American Society of Plant Biologists. All rights reserved.

26
Constructs were introduced into Agrobacterium strain GV3101 (pMP90) (Koncz and
Schell, 1986) by electroporation. For plant transformation we used the “floral dip” method
(Clough and Bent, 1998). Primer sequences are indicated in Supplemental Table II.
Characterization of the alleles
Allele frf1-1 was genotyped using the combination NS264_S26Lp and NS264_S26Rp for
the wild type and NS264_S26Lp and LB1SAIL and NS264_S26Rp and LB1SAIL for the
insertions. For genotyping the allele frf2-1 we used the combination NS288_S31Lp and
NS289_S31Rp for the wild type copy and NS288_S31Lp and LBa1 and NS289_S31Rp
and LBa1 for the insertions. The genotype of the allele frf4-1 was made using primers
S6Lp and S6Rp for the wild type and S6Rp and LBa1 for the insertion. The sequences of
the primers were retrieved from the web tool at http://signal.salk.edu/tdnaprimers.2.html
and the positions of the insertions were determined by sequencing. Primer sequences can
be found in Supplemental Table II.
GUS histochemical assays
We used 8 day-old plants of T3 transgenic lines. The staining was for 5 hours and as
described in Aguilar-Martinez and Sinha (2013).
Yeast one-hybrid assay
The Yeast One-Hybrid assay was made according to the MATCHMAKER One‐Hybrid
System (Clontech, Palo Alto, CA) and to Kooiker et al., (2005). 226bp (-2133 to -1907bp)
of the Arabidopsis STM promoter including the RB-box was introduced into the EcoRI and
SacI sites of the pHisi plasmid using oligos RBRIF and STMpRBSacIR to obtain the
pHisi-RB plasmid. The plasmid was sequenced and introduced upstream of the HIS3
reporter gene of the Y187 yeast strain (MAT α) using small scale transformation according
to the Yeast Protocols Handbook (Clontech). In order to assess background due to leaky
HIS3 expression, 3-aminotriazole (3-AT, Sigma A-8056) was used as a competitive
inhibitor of the His3 protein. The reporter strain was titrated on SD/-His plates with
varying amounts of 3-AT (0–60 mM) to determine the optimal concentration of 3-AT for
www.plantphysiol.orgon February 14, 2018 - Published by Downloaded from Copyright © 2014 American Society of Plant Biologists. All rights reserved.

27
background suppression. 15 mM 3-AT was sufficient for background suppression in the
yeast strains. We mated with a yeast strain AH-109 (MAT a) containing a normalized
library of cDNA from Arabidopsis in the plasmid pGADT7 kindly offered by Simona
Masiero (University of Milan). The growing colonies were plated again to confirm the
interactions. The pGADT7 plasmids were isolated and sequenced with the primer V7 (5’-
CTATTCGATGATGAAGATACCCC).
EMSA
The EMSA analysis was performed as in Aguilar-Martínez and Sinha, (2013). Briefly, the
coding sequence of FRF1 was cloned and used to generate a GST-fusion–protein-
expressing construct. The probes were generated with oligos NS171_RBabiot (5’-A
ATCCAGTTTGGTAATATATAAATATG-3’) and NS169_Rbdbiot (5’-GCTAATTTT
TCTTGCTTTTAGGTTAAG-3’). For the fragment of 246 bp covering the RB-box of
STM the plasmid with the construct ProSTM:GUS (Uchida et al., 2007) and ProSTM-
ΔCRB:GUS for the RBΔcore were used as templates. For competition we amplified the
same fragments using unlabeled oligos. We used the LightShift Chemiluminescent EMSA
Kit (Pierce).
Microscopy and chemical treatment
For analysis of ProJAG:GFP-STM and ProSTM:GFP-STM transgenic plants a LSM 710
laser scanning microscope (Carl Zeiss) was used. Chlorophyll autofluoresce collected at
wavelengths 647-721nm and GFP fluorescence collected at wavelengths 495-528nm were
excited using a combination of excitation from 488nm and 633nm lasers. In addition, the
range from 416nm-728nm was collected to increase the visibility of non-fluorescing
features. Light and fluorescence microscopy on vibratome sections of ProJAG:GFP-STM
plants was also employed according to Koenig et al., (2009), using bandpass filters at
525nm (GFP1) and 500nm (GFP2).
Detection of GFP-tagged proteins was performed by agro-infiltration method according to
the previous report (Voinnet et al., 2003). Briefly, Agrobacterium strains harboring
pMDC32/GFP-STM or pMDC32/GFP-STM-S272A were infiltrated into the abaxial air
www.plantphysiol.orgon February 14, 2018 - Published by Downloaded from Copyright © 2014 American Society of Plant Biologists. All rights reserved.

28
spaces of Nicotiana benthamiana plants together with Agrobacterium C58C1 strain
expressing p19, a silencing suppressor encoded by tomato bushy stunt virus. After 2 days,
GFP fluorescence was observed using Leica TCS SP2 confocal microscope (Leica). When
required, leaves were treated with 20 nM LMB (Sigma) 2 hours before observation as
previously reported (Igarashi et al., 2001; Ishida et al., 2004).
Accession numbers
The Arabidopsis Information Resource (TAIR; http://www.arabidopsis.org/) locus
identifiers for the FRF genes are: FRF1 (At3g59470), FRF2 (At3g07500), FRF3
(At4g12850) and FRF4 (At2g43280). FRF-like proteins in tomato are
Solyc06g073200.2.1, Solyc06g073210.2.1, Solyc09g057880.2.1 and Solyc09g057870.2.1,
in Medicago truncatula Medtr2g098920.1, Medtr2g098880.1, in contig_103701_3.1 and in
contig_49211_1.1, in Vitis vinifera GSVIVT01010765001, GSVIVT01010762001,
GSVIVT01001896001, GSVIVT01010761001, GSVIVT01010768001,
GSVIVT01010788001, GSVIVT01032951001, GSVIVT01036426001,
GSVIVT01036422001 and GSVIVT01012789001 and in rice LOC_Os03g15040.1,
LOC_Os10g06860.1, LOC_Os11g19030.1, LOC_Os11g02964.1, LOC_Os07g18260.1,
LOC_Os05g40680.1 and LOC_Os07g39490.1.
Supplemental material
Supplemental Figure 1. Expression pattern of GUS in plants ProSTM-ΔLK:GUS,
ProSTM-ΔRB:GUS and ProSTM-ΔRB-CK:GUS.
Supplemental Figure 2. FRF1 binds the RB-box.
Supplemental Figure 3. Phylogenetic tree of FRF factors.
Supplemental Figure 4. Expression pattern of FRF genes.
www.plantphysiol.orgon February 14, 2018 - Published by Downloaded from Copyright © 2014 American Society of Plant Biologists. All rights reserved.

29
Supplemental Figure 5. Sequence conservation on introns of KNOX1 genes in
Brassicaceae species.
Supplemental Figure 6. GUS expression pattern in plants ProJAG:GUS.
Supplemental Figure 7. Leaf phenotype in plants expressing STM under the JAG
promoter and GFP distribution under the endogenous STM promoter or the JAG promoter.
Supplemental Figure 8. Sequence alignment of STM ortholog factors.
Supplemental Figure 9. Effect of LMB treatment on STM cellular distribution.
Supplemental Table I. Sequenced clones from Yeast One-Hybrid assay.
Supplemental Table II. List of primers used in this study.
Supplemental Data Set 1. Sequence alignment of the FRF and FRS factors.
Supplemental Data Set 2. Sequence alignment of FRF-like factors in eudicot species.
Acknowledgements
We thank Lucia Colombo and Simona Masiero (University of Milan) for the cDNA library
for the yeast one-hybrid assays and members of Sinha lab for discussions and comments
on the manuscript and Preetveer Kaur and Katrina Torres for technical assistance with the
confocal image analysis.
Author contributions
J.A.A.M., N.U. and N.S. designed the research. J.A.A.M., N.U., S.K., B.T., D.A.W., A.Y.
and N.L. performed the research. J.A.A.M., N.U. and N.S. analyzed the data and wrote the
paper.
www.plantphysiol.orgon February 14, 2018 - Published by Downloaded from Copyright © 2014 American Society of Plant Biologists. All rights reserved.

30
Literature cited
Abascal F, Zardoya R, Telford MJ (2010) TranslatorX: multiple alignment of nucleotide
sequences guided by amino acid translations. Nucleic Acids Res. 38:W7-13
Aguilar-Martínez JA, Sinha NR (2013) Analysis of the role of Arabidopsis class I TCP
genes AtTCP7, AtTCP8, AtTCP22 and AtTCP23 in leaf development. Front Plant
Sci 4: 406
Aichinger E, Kornet N, Friedrich T, Laux T (2012) Plant stem cell niches. Annual
Review of Plant Biology 63: 615-636
Aida M, Ishida T, Fukaki H, Fujisawa H, Tasaka M (1997) Genes involved in organ
separation in Arabidopsis, an analysis of the cup-shaped cotyledon mutant. Plant
Cell 9: 841–857
Aida M, Ishida T, Tasaka M (1999) Shoot apical meristem and cotyledon formation
during Arabidopsis embryogenesis, interaction among the CUPSHAPED
COTYLEDON and SHOOTMERISTEMLESS genes. Development 126: 1563–1570
Alakonya A, Kumar R, Koenig D, Kimura S, Townsley B, Runo S, Garces HM, Kang
J, Yanez A, David-Schwartz R, Machuka J, Sinha N (2012) Interspecific RNA
interference of SHOOTMERISTEMLESS-like disrupts Cuscuta pentagona plant
parasitism. Plant Cell 24: 3153-3166
Alonso JM, Stepanova AN, Leisse TJ, Kim C J, Chen H, Shinn P, et al. (2003)
Genome-Wide Insertional Mutagenesis of Arabidopsis thaliana. Science 301: 653-
657
Atta R, Laurens L, Boucheron-Dubuisson E, Guivarc’h A, Carnero E, Giraudat-
Pautot V, Rech P, Chriqui D (2009) Pluripotency of Arabidopsis xylem pericycle
underlies shoot regeneration from root and hypocotyl explants grown in vitro. The
Plant Journal 57: 626–644
www.plantphysiol.orgon February 14, 2018 - Published by Downloaded from Copyright © 2014 American Society of Plant Biologists. All rights reserved.

31
Autran D, Jonak C, Belcram K, Beemster GT, Kronenberger J, Grandjean O, Inzé
D, Traas J (2002) Cell numbers and leaf development in Arabidopsis: a functional
analysis of the STRUWWELPETER gene. EMBO J 21: 6036-49
Bao F, Azhakanandam S, Franks RG (2010) SEUSS and SEUSS-LIKE transcriptional
adaptors regulate floral and embryonic development in Arabidopsis. Plant Physiol
152: 821-836
Barton KM (2010) Twenty years on: the inner workings of the shoot apical meristem, a
developmental dynamo. Developmental Biology 341: 95-113
Bauer P, Lubkowitz M, Tyers R, Nemoto K, Meeley RB, Goff SA, Freeling M (2004)
Regulation and a conserved intron sequence of liguleless3/4 knox class-I homeobox
genes in grasses. Planta 219: 359-68
Bellaoui M, Pidkowich MS, Samach A, Kushalappa K, Kohalmi SE, Modrusan Z,
Crosby WL, Haughn GW (2001) The Arabidopsis BELL1 and KNOX TALE
homeodomain proteins interact through a domain conserved between plants and
animals. Plant Cell 13: 2455–2470
Belles-Boix E, Hamant O, Witiak SM, Morin H, Traas J, Pautot V (2006) KNAT6: An
Arabidopsis Homeobox Gene Involved in Meristem Activity and Organ Separation.
Plant Cell 18: 1900–1907
Benkova E, Michniewicz M, Sauer M, Teichmann T, Seifertova D, Jurgens G, Friml J
(2003) Local efflux dependent auxin gradients as a common module for plant organ
formation. Cell 115: 591–602
Beschtold N, Ellis J, Pelletier G (1993) In planta Agrobacterium-mediated gene transfer
by filtration of adult Arabidopsis thaliana plants. C R Acad Sci (Paris) 316: 1194–
1199
Bharathan G, Goliber TE, Moore C, Kessler S, Pham T, Sinha NR (2002) Homologies
in leaf form inferred from KNOXI gene expression during development. Science
296: 1858–1860
Bhatt AM, Etchells JP, Canales C, Lagodienko A, Dickinson H (2004) VAAMANA–a
BEL1-like homeodomain protein, interacts with KNOX proteins BP and STM and
regulates inflorescence stem growth in Arabidopsis. Gene 328: 103–111
www.plantphysiol.orgon February 14, 2018 - Published by Downloaded from Copyright © 2014 American Society of Plant Biologists. All rights reserved.

32
Bolduc N, Hake S (2009) The maize transcription factor KNOTTED1 directly regulates
the gibberellin catabolism gene ga2ox1. Plant Cell 21: 1647-1658
Brand U, Fletcher JC, Hobe M, Meyerowitz EM, Simon R (2001) Dependence of stem
cell fate in Arabidopsis on a feedback loop regulated by CLV3 activity. Science
289: 617–619
Brudno M, Steinkamp R, Morgenstern B (2004) The CHAOS/DIALIGN WWW server
for multiple alignment of genomic sequences Nucl Acids Res 32: W41-W44
Byrne ME, Barley R, Curtis M, Arroyo JM, Dunham M, Hudson A, Martienssen RA
(2000) Asymmetric leaves1 mediates leaf patterning and stem cell function in
Arabidopsis. Nature 408: 967–971
Byrne ME, Simorowski J, Martienssen RA (2002) ASYMMETRIC LEAVES1 reveals
knox gene redundancy in Arabidopsis. Development 129: 1957–1965
Casimiro I, Marchant A, Bhalerao RP, Beeckmand T, Dhooged S, Swarupb R,
Graham N, Inzé D, Sandberg G, Casero PJ, Bennett M (2001) Auxin transport
promotes Arabidopsis lateral root initiation. Plant Cell 13: 843-852
Champagne CEM, Goliber TE, Wojciechowskib MF, Mei RW, Townsley BT, Wang
K, Paz MM, Geeta R, Sinha NR (2007) Compound leaf development and
evolution in the legumes. Plant Cell 19: 3369-3378
Chuck G, Lincoln C, Hake S (1996) KNAT1 induces lobed leaves with ectopic meristems
when overexpressed in Arabidopsis. Plant Cell 8: 1277-1289
Clough SJ, Bent AF (1998) Floral dip: a simplified method for Agrobacterium-mediated
transformation of Arabidopsis thaliana. Plant J 16: 735-743
Cole M, Nolte C, Werr W (2006) Nuclear import of the transcription factor SHOOT
MERISTEMLESS depends on heterodimerization with BLH proteins expressed in
discrete sub-domains of the shoot apical meristem of Arabidopsis thaliana. Nucleic
Acids Res 34: 1281-1292
Curtis MD, Grossniklaus U (2003) A gateway cloning vector set for high-throughput
functional analysis of genes in planta. Plant Physiol 133: 462–469
Dean G, Casson S, Lindsey K (2004) KNAT6 gene of Arabidopsis is expressed in roots
and is required for correct lateral root formation. Plant Mol Biol 54: 71-84
www.plantphysiol.orgon February 14, 2018 - Published by Downloaded from Copyright © 2014 American Society of Plant Biologists. All rights reserved.

33
Depuydt S, Doležal K, Lijsebettens MV, Moritz T, Holsters M, Vereecke D (2008)
Modulation of the hormone setting by Rhodococcus fascians results in ectopic
KNOX activation in Arabidopsis. Plant Physiology 146: 1267-1281
Dinneny JR, Yadegari R, Fischer RL, Yanofsky MF, Weigel D (2004) The role of
JAGGED in shaping lateral organs. Development 131: 1101-1110
Du J, Mansfield SD, Groover AT (2009) The Populus homeobox gene ARBORKNOX2
regulates cell differentiation during secondary growth. Plant J 60: 1000-1014
Elliott RC, Betzner AS, Huttner E, Oakes MP, Tucker WQ, Gerentes D, Perez P,
Smyth DR (1996) AINTEGUMENTA, an APETALA2-like gene of Arabidopsis
with pleiotropic roles in ovule development and floral organ growth. Plant Cell 8:
155–168
Endrizzi K, Moussian B, Haecker A, Levin JZ, Laux T (1996) The SHOOT
MERISTEMLESS gene is required for maintenance of undifferentiated cells in
Arabidopsis shoot and floral meristems and acts at a different regulatory level than
the meristem genes WUSCHEL and ZWILLE. Plant Journal 10: 967–979
Fletcher JC, Brand U, Running MP, Simon R, Meyerowitz EM (1999) Signaling of
cell fate decisions by CLAVATA3 in Arabidopsis shoot meristems. Science 283:
1911–1914
Frazer KA, Pachter L, Poliakov A, Rubin EM, Dubchak I (2004) VISTA:
computational tools for comparative genomics. Nucleic Acids Res 32: W273-279
Furutani M, Vernoux T, Traas J, Kato T, Tasaka M, Aida M (2004) PIN-FORMED1
and PINOID regulate boundary formation and cotyledon development in
Arabidopsis embryogenesis. Development 131: 5021-5030
Gallois JL, Woodward C, Reddy GV, Sablowski R (2002) Combined SHOOT
MERISTEMLESS and WUSCHEL trigger ectopic organogenesis in Arabidopsis.
Development 129: 3207-3217
Garces HMP, Champagne CEM, Townsley BT, Park S, Malho R, Pedroso MC,
Harada JJ, Sinha NR (2007) Evolution of asexual reproduction in leaves of the
genus Kalanchoë. PNAS 104: 15578-15583
www.plantphysiol.orgon February 14, 2018 - Published by Downloaded from Copyright © 2014 American Society of Plant Biologists. All rights reserved.

34
Giles JE, Villani PJ, DeMason DA (1998) A class 1 Knox full-length cDNA from pea
shoot tips (Accession No. AF063307) (PGR98-115) Plant Physiol 117: 1125
Gonzalez D, Bowen AJ, Carroll TS, Conlan RS (2007) The transcription corepressor
LEUNIG interacts with the histone deacetylase HDA19 and mediator components
MED14 (SWP) and CDK8 (HEN3) to repress transcription. Mol Cell Biol 27:
5306-5315
Goodstein DM, Shu S, Howson R, Neupane R, Hayes RD, Fazo J, Mitros T, Dirks W,
Hellsten U, Putnam N, Rokhsar DS (2012) Phytozome: a comparative platform
for green plant genomics. Nucleic Acids Res 40: (D1), D1178-D1186
Greene B, Walko R, Hake S (1994) Mutator insertions in an intron of the maize knotted1
gene result in dominant suppressible mutations. Genetics 138: 1275–1285
Groover AT, Mansfield SD, DiFazio SP, Dupper G, Fontana JR, Millar R, Wang Y
(2006) The Populus homeobox gene ARBORKNOX1 reveals overlapping
mechanisms regulating the shoot apical meristem and the vascular cambium. Plant
Mol Biol 61: 917–932
Hackbusch J, Richter K, Müller J, Salamini F, Uhrig JF (2005) A central role of
Arabidopsis thaliana ovate family proteins in networking and subcellular
localization of 3-aa loop extension homeodomain proteins. PNAS 102: 4908–4912
Hareven D, Gutfinger T, Parnis A, Eshed Y, Lifschitz E (1996) The making of a
compound leaf: Genetic manipulation of leaf architecture in tomato. Cell 84: 735–
744
Hay A, Kaur H, Phillips A, Hedden P, Hake S, Tsiantis M (2002) The gibberellin
pathway mediates KNOTTED1-type homeobox function in plants with different
body plans. Current Biology 12: 1557-1565
Hay A, Jackson D, Ori N, Hake S (2003) Analysis of the competence to respond to
KNOTTED1 activity in Arabidopsis leaves using a steroid induction system. Plant
Physiol 131: 1671-1680
Hay A, Tsiantis M (2006) The genetic basis for differences in leaf form between
Arabidopsis thaliana and its wild relative Cardamine hirsuta. Nat Genet 38: 942-
947
www.plantphysiol.orgon February 14, 2018 - Published by Downloaded from Copyright © 2014 American Society of Plant Biologists. All rights reserved.

35
Hay A, Barkoulas M, Tsiantis M (2006) ASYMMETRIC LEAVES1 and auxin activities
converge to repress BREVIPEDICELLUS expression and promote leaf
development in Arabidopsis. Development 133: 3955-3961
Heckman KL, Pease LR (2007) Gene splicing and mutagenesis by PCR-driven overlap
extension. Nature Protocols 2: 924-932
Heisler MG, Ohno C, Das P, Sieber P, Reddy GV, Long JA, Meyerowitz EM (2005)
Patterns of auxin transport and gene expression during primordium development
revealed by live imaging of the Arabidopsis inflorescence meristem. Current
Biology 15: 1899-1911
Hibara K, Takada S, Tasaka M (2003) CUC1 gene activates the expression of SAM-
related genes to induce adventitious shoot formation. Plant J 36: 687-96
Hofer J, Turner L, Hellens R, Ambrose M, Matthews P, Michael A, Ellis N (1997)
UNIFOLIATA regulates leaf and flower morphogenesis in pea. Current Biology 7:
581–587
Hofer J, Gourlay C, Michael A, Ellis THN (2001) Expression of a class 1 knotted1-like
homeobox gene is down-regulated in pea compound leaf primordia. Plant Mol Biol
45: 387–398
Hu W, Feng B, Ma H (2011) Ectopic expression of the Arabidopsis MINI ZINC
FINGER1 and MIF3 genes induces shoot meristems on leaf margins. Plant Mol
Biol 76: 57-68
Hudson M, Ringli C, Boylan MT, Quail PH (1999) The FAR1 locus encodes a novel
nuclear protein specific to phytochrome A signaling. Genes Development 13: 2017-
2027
Hunter S, Jones P, Mitchell A, Apweiler R, Attwood TK, Bateman A, et al. (2012)
InterPro in 2011: new developments in the family and domain prediction database.
Nucleic Acids Res 40: D306-312
Ichihashi Y, Aguilar-Martínez JA, Farhi M, Chitwood DH, Kumar R, Millonc LV,
Peng J, Maloof JN, Sinha NR (2014) Evolutionary developmental transcriptomics
reveals a gene network module regulating interspecific diversity in plant leaf shape
PNAS 111: E2616-E2621
www.plantphysiol.orgon February 14, 2018 - Published by Downloaded from Copyright © 2014 American Society of Plant Biologists. All rights reserved.

36
Igarashi D, Ishida S, Fukazawa J, Takahashi Y (2001) 14-3-3 Proteins regulate
intracellular localization of the bZIP transcriptional activator RSG. Plant Cell 13:
2483-2497
Inada DC, Bashir A, Lee C, Thomas BC, Ko C, Goff, SA, Freeling, M (2003)
Conserved noncoding sequences in the grasses. Genome Res 13: 2030-2041
Ishida S, Fukazawa J, Yuasa T, Takahashi Y (2004) Involvement of 14-3-3 signaling
protein binding in the functional regulation of the transcriptional activator
REPRESSION OF SHOOT GROWTH by gibberellins. Plant Cell 16: 2641-2651
Ito Y, Kurata N (2008) Disruption of KNOX gene suppression in leaf by introducing its
cDNA in rice. Plant Science 174: 357-365
Jackson D, Veit B, Hake S (1994) Expression of maize KNOTTED1 related homeobox
genes in the shoot apical meristem predicts patterns of morphogenesis in the
vegetative shoot. Development 120: 405-413
Janssen BJ, Williams A, Chen JJ, Mathern J, Hake S, Sinha N (1998) Isolation and
characterization of two knotted-like homeobox genes from tomato. Plant Mol Biol
36: 417-425
Jasinski S, Piazza P, Craft J, Hay A, Woolley L, Rieu I, Phillips A, Hedden P, Tsiantis
M (2005) KNOX action in Arabidopsis is mediated by coordinate regulation of
cytokinin and gibberellin activities. Current Biology 15: 1560-1565
Karimi M, Inze D, Depicker A (2002) GATEWAY vectors for Agrobacterium-mediated
plant transformation. Trends Plant Sci 7: 193–195
Kawamura E, Horiguchi G, Tsukaya H (2010) Mechanisms of leaf tooth formation in
Arabidopsis. Plant Journal 62: 429–441
Kerstetter R, Vollbrecht E, Lowe B, Veit B, Yamaguchi J, Hake S (1994) Sequence
analysis and expression patterns divide the maize knotted1-like homeobox genes
into two classes. Plant Cell 6: 1877-1887
Kim JY, Rim Y, Wang J, Jackson D (2005) A novel cell-to-cell trafficking assay
indicates that the KNOX homeodomain is necessary and sufficient for intercellular
protein and mRNA trafficking. Genes Dev 19: 788-793
www.plantphysiol.orgon February 14, 2018 - Published by Downloaded from Copyright © 2014 American Society of Plant Biologists. All rights reserved.

37
Kimura S, Koenig D, Kang J, Yoong F, Sinha N (2008) Natural variation in leaf
morphology results from mutation of a novel KNOX gene. Current Biology 18:
672–677
Koenig D, Bayer E, Kang J, Kuhlemeier C, Sinha N (2009) Auxin patterns Solanum
lycopersicum leaf morphogenesis. Dev 136: 2997-3006
Koltai H, Bird DM (2000) Epistatic repression of PHANTASTICA and class 1 KNOTTED
genes is uncoupled in tomato. Plant Journal 22: 455-459
Koltai H, Dhandaydham M, Opperman C, Thomas J, Bird D (2001) Overlapping plant
signal transduction pathways induced by a parasitic nematode and a rhizobial
endosymbiont. Molecular Plant-Microbe Interactions 14: 1168-1177
Koncz C, Schell J (1986) The promoter of TL -DNA gene 5 controls the tissue-specific
expression of chimeric genes carried by a novel type of Agrobacterium binary
vector. Mol Gen Genet 204: 383-396
Kooiker M, Airoldi CA, Losa A, Manzotti PS, Finzi L, Kater MM, Colombo L (2005)
BASIC PENTACYSTEINE1, a GA binding protein that induces conformational
changes in the regulatory region of the homeotic Arabidopsis gene SEEDSTICK.
Plant Cell 17: 722-729
Kumaran MK, Bowman JL, Sundaresan V (2002) YABBY polarity genes mediate the
repression of KNOX homeobox genes in Arabidopsis. Plant Cell 14: 2761-70
Kurakawa T, Ueda N, Maekawa M, Kobayashi K, Kojima M, Nagato Y, Sakakibara
H, Kyozuka J (2007) Direct control of shoot meristem activity by a cytokinin
activating enzyme. Nature 445: 652–655
Laux T, Mayer KFX, Berger J, Jürgens G (1996) The WUSCHEL gene is required for
shoot and floral meristem integrity in Arabidopsis. Development 122: 87–96
Lenhard M, Jürgens G, Laux T (2002) The WUSCHEL and SHOOTMERISTEMLESS
genes fulfil complementary roles in Arabidopsis shoot meristem regulation.
Development 129: 3195-206
Li J, Li G, Gao S, Martinez C, He G, Zhou Z, Huang X, Lee JH, Zhang H, Shen Y,
Wang H, Deng XW (2010) Arabidopsis transcription factor ELONGATED
www.plantphysiol.orgon February 14, 2018 - Published by Downloaded from Copyright © 2014 American Society of Plant Biologists. All rights reserved.

38
HYPOCOTYL5 plays a role in the feedback regulation of phytochrome A
signaling. Plant Cell 22: 3634-49
Li E, Wang S, Liu Y, Chen JG, Douglas CJ (2011) OVATE FAMILY PROTEIN4
(OFP4) interaction with KNAT7 regulates secondary cell wall formation in
Arabidopsis thaliana. Plant J 67: 328–341
Li G, Siddiqui H, Teng Y, Lin R, Wan XY, Li J, Lau OS, Ouyang X, Dai M, Wan J,
Devlin PF, Deng XW, Wang H (2011) Coordinated transcriptional regulation
underlying the circadian clock in Arabidopsis. Nat Cell Biol 13: 616-22
Li Z, Li B, Shen WH, Huang H, Dong A (2012) TCP transcription factors interact with
AS2 in the repression of class-I KNOX genes in Arabidopsis thaliana. Plant J 71:
99-107
Lin R, Wang H (2004) Arabidopsis FHY3/FAR1 gene family and distinct roles of its
members in light control of Arabidopsis development. Plant Physiology 136: 4010–
4022
Lin R, Ding L, Casola C, Ripoll DR, Feschotte C, Wang H (2007). Transposase-derived
transcription factors regulate light signaling in Arabidopsis. Science 318: 1302-
1305
Lin R, Teng Y, Park HJ, Ding L, Black C, Fang P, Wang H (2008) Discrete and
essential roles of the multiple domains of Arabidopsis FHY3 in mediating
phytochrome A signal transduction. Plant Physiol 148: 981-92
Lincoln C, Long J, Yamaguchi J, Serikawa K, Hake S (1994) A knotted1-like
homeobox gene in Arabidopsis is expressed in the vegetative meristem and
dramatically alters leaf morphology when overexpressed in transgenic plants. Plant
Cell 6: 1859–1876
Lodha M, Marco CF, Timmermans MCP (2013) The ASYMMETRIC LEAVES
complex maintains repression of KNOX homeobox genes via direct recruitment of
Polycomb-repressive complex2. Genes Development 27: 596-601
Long JA, Moan EI, Medford JI, Barton MK (1996) A member of the KNOTTED class
of homeodomain proteins encoded by the STM gene of Arabidopsis. Nature 379:
66–69
www.plantphysiol.orgon February 14, 2018 - Published by Downloaded from Copyright © 2014 American Society of Plant Biologists. All rights reserved.

39
Long J, Barton MK (2000) Initiation of axillary and floral meristems in Arabidopsis. Dev
Biol 218: 341-53
Lucas WJ, Bouche-Pillon S, Jackson DP, Nguyen L, Baker L, Ding B, Hake S (1995)
Selective trafficking of KNOTTED1 homeodomain protein and its mRNA through
plasmodesmata. Science 270: 1980-1983
Luo M, Yu CW, Chen FF, Zhao L, Tian G, Liu X, Cui Y, Yang JY, Wu K (2012)
Histone Deacetylase HDA6 Is Functionally Associated with AS1 in Repression of
KNOX Genes in Arabidopsis. PLoS Genet 8: e1003114
Lynn K, Fernandez A, Aida M, Sedbrook J, Tasaka M, Masson P, Barton MK (1999)
The PINHEAD/ZWILLE gene acts pleiotropically in Arabidopsis development and
has overlapping functions with the ARGONAUTE1 gene. Development 126: 469-
481
Magnani E, Hake S (2008) KNOX lost the OX: the Arabidopsis KNATM gene defines a
novel class of KNOX transcriptional regulators missing the homeodomain. Plant
Cell 20: 875–887
Mayer KF, Schoof H, Haecker A, Lenhard M, Jurgens G, Laux T (1998) Role of
WUSCHEL in regulating stem cell fate in the Arabidopsis shoot meristem. Cell 95:
805-815
Meisel L, Lam E. (1996) The conserved ELK homeodomain of KNOTTED-1 contains
two regions that signal nuclear localization. Plant Mol Biol 30: 1–14
Müller KJ, Romano N, Gerstner O, Garcia-Maroto F, Pozzi C, Salamini F, Rohde W
(1995) The barley Hooded mutation caused by a duplication in a homeobox gene
intron. Nature 374: 727-30
Mushegian A, Koonin EV (1996) Sequence analysis of eukaryotic developmental
proteins: Ancient and novel domains. Genetics 144: 817–828
Mutwil M, Obro J, Willats WG, Persson S (2008) GeneCAT: novel Webtools that
combine BLAST and co-expression analyses. Nucleic Acids Res 36: W320–W326
Nagasaki H, Sakamoto T, Sato Y, Matsuoka, M (2001) Functional analysis of the
conserved domains of a rice KNOX homeodomain protein, OSH15. Plant Cell 13:
2085-2098
www.plantphysiol.orgon February 14, 2018 - Published by Downloaded from Copyright © 2014 American Society of Plant Biologists. All rights reserved.

40
Néron B, Ménager H, Maufrais C, Joly N, Maupetit J, Letort S, Sébastien Carrere S,
Tuffery P, Letondal C (2009) Mobyle: a new full web bioinformatics framework.
Bioinformatics 25: 3005-3011
Nikovics K, Blein, T, Peaucelle, A, Ishida T, Morin H, Aida M, Laufs P (2006) The
balance between the MIR164A and CUC2 genes controls leaf margin serration in
Arabidopsis. Plant Cell 18: 2929–2945
Ohno CK, Reddy GV, Heisler MG, Meyerowitz EM (2004) The Arabidopsis JAGGED
gene encodes a zinc finger protein that promotes leaf tissue development.
Development 131: 1111-1122
Ori N, Eshed Y, Chuck G, Bowman JL, Hake S (2000) Mechanisms that control knox
gene expression in the Arabidopsis shoot. Development 127: 5523-5532
Osnato M, Stile MR, Wang Y, Meynard D, Curiale S, Guiderdoni E, Liu Y, Horner
DS, Ouwerkerk PBF, Pozzi C, Müller KJ, Salamini F, Rossini L (2010) Cross
talk between the KNOX and ethylene pathways is mediated by intron-binding
transcription factors in barley. Plant Physiology 154: 1616-1632
Pagnussat GC, Yu HJ, Sundaresan V (2007) Cell-fate switch of synergid to egg cell in
Arabidopsis eostre mutant embryo sacs arises from misexpression of the BEL1-like
homeodomain gene BLH1. Plant Cell 19: 3578–3592
Qi B, Zheng H (2013) Modulation of root-skewing responses by KNAT1 in Arabidopsis
thaliana. Plant Journal 76: 380-392
Reinhardt D, Mandel T, Kuhlemeier C (2000) Auxin regulates the initiation and radial
position of plant lateral organs. Plant Cell 12: 507–518
Ren J, Wen L, Gao X, Jin C, Xue Y, Yao X (2009) DOG 1.0: Illustrator of protein
domain structures. Cell Research 19: 271–273
Rupp HM, Frank M, Werner T, Strnad M, Schmulling T (1999) Increased steady state
mRNA levels of the STM and KNAT1 homeobox genes in cytokinin overproducing
Arabidopsis thaliana indicate a role for cytokinins in the shoot apical meristem.
Plant Journal 18: 557–563
www.plantphysiol.orgon February 14, 2018 - Published by Downloaded from Copyright © 2014 American Society of Plant Biologists. All rights reserved.

41
Rutjens B, Bao D, van Eck-Stouten E, Brand M, Smeekens S, Proveniers M (2009)
Shoot apical meristem function in Arabidopsis requires the combined activities of
three BEL1-like homeodomain proteins. Plant J 58: 641-54
Sakamoto T, Nishimura A, Tamaoki M, Kuba M, Tanaka H, Iwahori S, Matsuoka M
(1999) The conserved KNOX domain mediates specificity of tobacco
KNOTTED1-type homeodomain proteins. Plant Cell 11: 1419–1432
Santi L, Wang Y, Stile MR, Berendzen K, Wanke D, Roig C, Pozzi C, Müller K,
Müller J, Rohde W, Salamini F (2003) The GA octodinucleotide repeat binding
factor BBR participates in the transcriptional regulation of the homeobox gene
Bkn3. Plant J 34: 813-26
Schmid M, Davison TS, Henz SR, Pape UJ, Demar M, Vingron M, Schölkopf B,
Weigel D, Lohmann J (2005) A gene expression map of Arabidopsis
development. Nature Genetics 37: 501-506
Schoof H, Lenhard M, Haecker A, Mayer KF, Jurgens G, Laux T (2001) The stem cell
population of Arabidopsis shoot apical meristems is maintained by a regulatory
loop between the CLAVATA and WUSCHEL genes. Cell 100: 635–644
Schubert D, Primavesi L, Bishopp A, Roberts G, Doonan J, Jenuwein T, Goodrich J
(2006) Silencing by plant Polycomb-group genes requires dispersed trimethylation
of histone H3 at lysine 27. EMBO J 25: 4638–4649
Scofield S, Murray JAH (2006). KNOX gene function in plant stem cell niches. Plant Mol
Biol 60: 929–946
Sessions A, Burke E, Presting G, Aux G, McElver J, Patton D, Dietrich B, Ho P,
Bacwaden J, Ko C, Clarke JD, Cotton D, Bullis D, Snell J, Miguel T,
Hutchison D, Kimmerly B, Mitzel T, Katagiri F, Glazebrook J, Law M, Goff
SA (2002) A High-Throughput Arabidopsis Reverse Genetics System. Plant Cell
14: 2985-2994
Simonini S, Kater MM (2014) Class I BASIC PENTACYSTEINE factors regulate
HOMEOBOX genes involved in meristem size maintenance. J Exp Bot 65: 1455-
1465
www.plantphysiol.orgon February 14, 2018 - Published by Downloaded from Copyright © 2014 American Society of Plant Biologists. All rights reserved.

42
Smith HM, Boschke I, Hake S (2002) Selective interaction of plant homeodomain
proteins mediates high DNA- binding affinity. PNAS 99: 9579-9584
Smith HM, Hake S (2003) The interaction of two homeobox genes,
BREVIPEDICELLUS and PENNYWISE, regulates internode patterning in the
Arabidopsis inflorescence. Plant Cell 15: 1717–1727
Smith LG, Greene B, Veit B, Hake S (1992) A dominant mutation in the maize
homeobox gene, Knotted-1, causes its ectopic expression in leaf cells with altered
fates. Development 116: 21-30
Stahle MI, Kuehlich J, Staron L, von Arnim AG, Golz JF (2009) YABBYs and the
transcriptional corepressors LEUNIG and LEUNIG_HOMOLOG maintain leaf
polarity and meristem activity in Arabidopsis. Plant Cell 21: 3105-3118
Swoford, D.L. (2003) PAUP*. Phylogenetic analysis using parsimony (*and other
methods). Version 4. Sinauer associates, Sunderland, Massachussets.
Takada S, Hibara K, Ishida T, Tasaka M (2001) The CUP-SHAPED COTYLEDON1
gene of Arabidopsis regulates shoot apical meristem formation. Development 128:
1127–1135
Tamaoki M, Sato Y, Matsuoka M (1997) Dorsoventral pattern formation of tobacco leaf
involves spatial expression of a tobacco homeobox gene, NTH15. Genes Genet Syst
72: 1-8
Tamura K, Peterson D, Peterson N, Stecher G, Nei M, Kumar S (2011) MEGA5:
molecular evolutionary genetics analysis using likelihood, distance, and parsimony
methods. Mol Biol Evol 28: 2731-2739
Tang W, Wang W, Chen D, Ji Q, Jing Y, Wang H, Lin R (2012) Transposase-derived
proteins FHY3/FAR1 interact with PHYTOCHROME-INTERACTING FACTOR1
to regulate chlorophyll biosynthesis by modulating HEMB1 during deetiolation in
Arabidopsis. Plant Cell 24: 1984-2000
Testone G, Condello E, Verde I, Nicolodi C, Caboni E, Dettori MT, Vendramin E,
Bruno L, Bitonti MB, Mele G, Giannino D (2012) The peach (Prunus persica L.
Batsch) genome harbours 10 KNOX genes, which are differentially expressed in
www.plantphysiol.orgon February 14, 2018 - Published by Downloaded from Copyright © 2014 American Society of Plant Biologists. All rights reserved.

43
stem development, and the class 1 KNOPE1 regulates elongation and lignification
during primary growth. J Exp Bot 63: 5417-35
Townsley BT, Sinha NR, Kang J (2013) KNOX1 genes regulate lignin deposition and
composition in monocots and dicots. Front Plant Sci 4: 121
Truernit E, Siemering KR, Hodge S, Grbic V, Haseloff J (2006) A map of KNAT gene
expression in the Arabidopsis root. Plant Mol Biol 60: 1-20
Tsuda K, Ito Y, Sato Y, Kurataa N (2011) Positive autoregulation of a KNOX gene is
essential for shoot apical meristem maintenance in rice. Plant Cell 23: 4368–4381
Uchida N, Townsley B, Chung K, Sinha N (2007) Regulation of SHOOT
MERISEMLESS genes via an upstream-conserved noncoding sequence coordinates
leaf development. PNAS 104: 15953-15958
Viola IL, Gonzalez, DH (2009) Binding properties of the complex formed by the
Arabidopsis TALE homeodomain proteins STM and BLH3 to DNA containing
single and double target sites. Biochimie 91: 974–981
Voinnet O, Rivas S, Mestre P, Baulcombe D (2003) An enhanced transient expression
system in plants based on suppression of gene silencing by the p19 protein of
tomato bushy stunt virus. Plant J 33: 949-56
Vollbrecht E, Veit B, Sinha N, Hake S (1991) The developmental gene Knotted-1 is a
member of a maize homeobox gene family. Nature 350: 241-243
Wang H, Deng XW (2002) Arabidopsis FHY3 defines a key phytochrome A signaling
component directly interacting with its homologous partner FAR1. EMBO J 21:
1339–1349
Wang H, Chen J, Wen J, Tadege M, Li G, Liu Y, Mysore KS, Ratet P, Chen R (2008)
Control of compound leaf development by FLORICAULA/LEAFY ortholog
SINGLE LEAFLET1 in Medicago truncatula. Plant Physiol 146: 1759–1772
Whitelam GC, Johnson E, Peng J, Carol P, Anderson ML, Cowl JS, Harberd NP
(1993) Phytochrome A null mutants of Arabidopsis display a wild-type phenotype
in white light. Plant Cell 5: 757–768
Williams RW (1998) Plant homeobox genes: many functions stem from a common motif.
Bioessays 20: 280-282
www.plantphysiol.orgon February 14, 2018 - Published by Downloaded from Copyright © 2014 American Society of Plant Biologists. All rights reserved.

44
Winter D, Vinegar B, Nahal H, Ammar R, Wilson GV, Provart NJ (2007) An
‘Electronic Fluorescent Pictograph’ browser for exploring and analyzing large-
scale biological data sets. PLoS ONE 2: e718.
Winter N, Kollwig G, Zhang S, Kragler F (2007) MPB2C, a microtubule-associated
protein, regulates non-cell-autonomy of the homeodomain protein KNOTTED1.
Plant Cell 19: 3001-3018
Xu L, Shen WH (2008) Polycomb silencing of KNOX genes confines shoot stem cell
niches in Arabidopsis. Current Biology 18: 1966-1971
Yanai O, Shani E, Dolezal K, Tarkowski P, Sablowski R, Sandberg G, Samach A, Ori
N (2005) Arabidopsis KNOXI proteins activate cytokinin biosynthesis. Current
Biology 15: 1566-1571
Table I. Name of the constructs of the STM promoter and fragments deleted.
www.plantphysiol.orgon February 14, 2018 - Published by Downloaded from Copyright © 2014 American Society of Plant Biologists. All rights reserved.
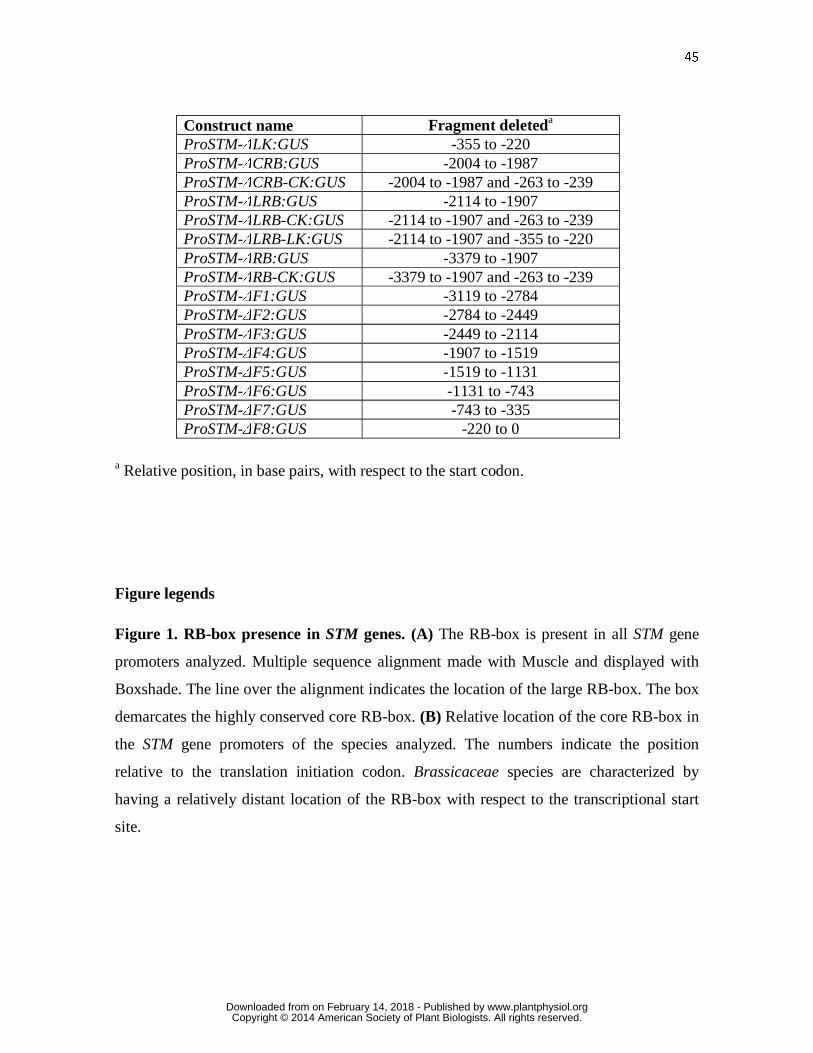
45
Construct name Fragment deleteda
ProSTM-ΔLK:GUS -355 to -220 ProSTM-ΔCRB:GUS -2004 to -1987 ProSTM-ΔCRB-CK:GUS -2004 to -1987 and -263 to -239 ProSTM-ΔLRB:GUS -2114 to -1907 ProSTM-ΔLRB-CK:GUS -2114 to -1907 and -263 to -239 ProSTM-ΔLRB-LK:GUS -2114 to -1907 and -355 to -220 ProSTM-ΔRB:GUS -3379 to -1907 ProSTM-ΔRB-CK:GUS -3379 to -1907 and -263 to -239 ProSTM-ΔF1:GUS -3119 to -2784 ProSTM-ΔF2:GUS -2784 to -2449 ProSTM-ΔF3:GUS -2449 to -2114 ProSTM-ΔF4:GUS -1907 to -1519 ProSTM-ΔF5:GUS -1519 to -1131 ProSTM-ΔF6:GUS -1131 to -743 ProSTM-ΔF7:GUS -743 to -335 ProSTM-ΔF8:GUS -220 to 0
a Relative position, in base pairs, with respect to the start codon.
Figure legends
Figure 1. RB-box presence in STM genes. (A) The RB-box is present in all STM gene
promoters analyzed. Multiple sequence alignment made with Muscle and displayed with
Boxshade. The line over the alignment indicates the location of the large RB-box. The box
demarcates the highly conserved core RB-box. (B) Relative location of the core RB-box in
the STM gene promoters of the species analyzed. The numbers indicate the position
relative to the translation initiation codon. Brassicaceae species are characterized by
having a relatively distant location of the RB-box with respect to the transcriptional start
site.
www.plantphysiol.orgon February 14, 2018 - Published by Downloaded from Copyright © 2014 American Society of Plant Biologists. All rights reserved.

46
Figure 2. Expression of GUS in the Arabidopsis STM promoter constructs harboring
combinations of RB-box and K-box deletions. (A) Schemes of the STM promoter
fragments used. Numbers indicate the position relative to the translation initiation codon
ATG. Solid lines indicates the fragment of the promoter used, dashed boxes indicate the
promoter region deleted for every construct. Vertical dashed lines connect the boxes
between constructs for clarification. (B-Q) Whole-mount and transverse sections of 8 days-
old T3 seedlings harboring the constructs depicted in (A). (B, C) Staining of plants with
the construct ProSTM:GUS with the native fragment of 3379bp. Seedling (B) and
transverse section (C) through the shoot apex. (D, E) Staining of plants with the construct
ProSTM-ΔLK:GUS harboring the STM promoter without the long K-box. (F, G) seedling
(F) and cross-section (G) of plants with the construct ProSTM-ΔCRB:GUS having the STM
promoter without the core RB-box. (H, I) GUS expression pattern in plants ProSTM-
ΔCRB-CK:GUS lacking both the core RB-box and the core K-box. (J, K) Staining of
plants with the construct ProSTM-ΔLRB:GUS without the large RB-box. (J) Seedling and
(K) cross-section through the hypocotyl. (L-O) GUS expression pattern in plants ProSTM-
ΔLRB-CK:GUS lacking both the large RB-box and the core K-box. Seedling (L),
transverse section through the shoot apex (M), the hypocotyl (N) and young leaf with GUS
accumulation in sinuses (O). (P, Q) Staining of plants with the construct ProSTM-ΔLRB-
LK:GUS harboring deletions of the large RB-box and the large K-box. Seedling (P) and
transverse section (Q) through the shoot apex. Dashed lines outline leaf primordia. Bars
are B, D, F, H, J, L and P = 0.5 mm, C, E, G, I, K and N = 50 µm, M and Q = 100 µm.
Figure 3. Characterization of FRF1 and related factors. (A) FRF1 is capable to bind
the RB-box through EMSA analysis. Indicated is also the positive control (C+) from the
manufacture’s kit. Arrows mark the shifted bands and arrowheads the free probe of 246 bp
for the RB-box and 60 bp for C+. (B) FRF factors are related to FRS factors. Protein-
domain structure of FRF and FRS factors. The FAR1 DNA binding domain is indicated by
a pale grey box, the MULE transposase domain by dark grey box and the Zinc finger of the
SWIM-type by a black box. Numbers indicate the size of each factor. (C) Phylogenetic
analysis of FRF factors in Arabidopsis, tomato, medicago, grape and rice. Neighbor
www.plantphysiol.orgon February 14, 2018 - Published by Downloaded from Copyright © 2014 American Society of Plant Biologists. All rights reserved.

47
joining tree with 1000 bootstrap replications using the conserved FAR1 DNA binding
domain of the FRF factors. Branch lengths are indicated. The tree was generated using
PAUP* 4.0 (Swoford D.L., 2003; paup.csit.fsu.edu).
Figure 4. GUS expression pattern in constructs with internal deletions in the
Arabidopsis STM promoter. (A) Schemes of the STM promoter with the internal deletion
fragments used. Numbers indicate the position relative to the translation initiation codon
ATG. The position of the RB-box and the K-box are also indicated. Solid lines indicate the
fragment of the promoter used, dashed boxes indicate the internal deletion on each
construct. Vertical dashed lines connect the boxes between constructs for clarification. (B-
R) Whole-mount and transverse sections of 8 days-old T3 seedlings harboring the
constructs indicated in (A). (B, C) Staining of plants with the construct ProSTM-
ΔF1:GUS. Seedling (B) and transverse section through the shoot apex (C). (D, E) Staining
of plants with the construct ProSTM-ΔF2:GUS. Seedling (D) and cross-section through the
shoot apex (E). (F, G) Seedling (F) and cross-section through the hypocotyl (G) of stained
plants with the construct ProSTM-ΔF3:GUS. (H, I) GUS expression pattern in plants
ProSTM-ΔF4:GUS. Whole-mount stained seedling (H) and cross-section through the shoot
apical region (I). (J, K) Staining of plants with the construct ProSTM-ΔF5:GUS. Seedling
(J) and cross-section through the shoot apex (K). (L, M) GUS expression pattern in plants
ProSTM-ΔF6:GUS. Seedling (L) and transverse section through the shoot apex (M). (N,
O) Staining of plants with the construct ProSTM-ΔF7:GUS. Seedling (N) and cross-section
through the shoot apical region (O). (P-R) Staining of plants with the construct ProSTM-
ΔF8:GUS. Seedling (P) and cross-section through the shoot apex (Q) and through the
hypocotyl (R). Leaf primordia are outlined with dashed lines. Bars are B, D, F, H, J, L, N
and P = 0.5 mm, C, E and O = 25 µm, G, K, M and R = 50 µm, I and Q = 100 µm.
Figure 5. CNSs in STM-like gene promoters of Brassicaceae species upstream of the
RB-box. (A) mVISTA alignment of B. rapa STM promoter compared with the
corresponding genes of T. halophila, C. rubella, A. thaliana, A. lyrata and C. hirsuta.
Three regions, Fa, Fb and Fc are identified. (B) Sequence alignment in the Fa region. (C)
www.plantphysiol.orgon February 14, 2018 - Published by Downloaded from Copyright © 2014 American Society of Plant Biologists. All rights reserved.

48
Alignment of the sequences for the Fb region. (D) Sequence alignment in the Fc region.
Sequence aligments were made with Muscle and displayed with Boxshade.
Figure 6. Expression of STM under the control of JAGGED promoter.
(A, B) Consecutive transverse sections through the shoot apex of transgenic Arabidopsis
plants ProJAG:GUS expressing the gene uidA under the control of the JAGGED promoter.
(A) GUS is expressed in Po leaf primordia (arrowhead). (B) GUS is found in Po
(arrowhead) and in the developing leaves (asterisks). (C-E) Expression of STM directed by
the JAG promoter. (C) Rosette of a wild type plant. (D) Representative rosette of
ProJAG:STM plants with a strong phenotype. Plants are very small with partially
undifferentiated leaves. (E) Rosette of plants ProJAG:STM with a mild phenotype. The
plants are small and have deeply lobed leaves. (F-H) Confocal anlysis of plants expressing
GFP-STM under STM or JAG promoters. (F) Confocal merged image of plants
ProSTM:GFP-STM. GFP signal is found at the base of the shoot apex (arrowhead). (G)
Confocal merged image of the shoot apex of a plant ProJAG:GFP-STM with strong
phenotype showing intense GFP expression in stipule-like structures. (H) Confocal merged
image of a plant ProJAG:GFP-STM with highly lobed leaves and GFP expression in the
base of developing leaves. Bars are A and B = 50 µm, C = 0.5 cm, D and E = 0.25 cm, F =
25 µm, G and H = 100 µm.
Figure 7. Altering the ELK domain affects leaf development and cellular localization
of STM. (A) Diagram depicting the conserved domains of STM KNOX1, KNOX2, GSE,
ELK and HD. Numbers indicate relative position from the first amino-acid. (B-D) Effect of
the S272A mutation in the ELK domain in leaf development. Compared with plants
ProSTM:GFP-STM that have a phenotype similar to wild type (Uchida et al., 2007), plants
ProSTM:GFP-STM-S272A produce smaller and lobed rosette leaves (B) and highly lobed
cauline leaves (C) while overall plant architecture remains unaffected (D). Representative
plants and isolated leaves are five-week-old. (E-F) The mutation S272A alters the cellular
distribution of STM. In agro-infiltration experiments in N. benthamiana leaves, STM
(GFP-STM) is distributed throughout the cytoplasm (E), however in the S272A mutation
(GFP-STM-S272A) STM is located in the cell nucleus (F). Arrow indicates a cell nucleus.
www.plantphysiol.orgon February 14, 2018 - Published by Downloaded from Copyright © 2014 American Society of Plant Biologists. All rights reserved.

49
STM distribution visualized with confocal microscopy. Bars are B = 1 cm, C = 5 mm, D =
2 cm, E and F = 20 µm.
www.plantphysiol.orgon February 14, 2018 - Published by Downloaded from Copyright © 2014 American Society of Plant Biologists. All rights reserved.

www.plantphysiol.orgon February 14, 2018 - Published by Downloaded from Copyright © 2014 American Society of Plant Biologists. All rights reserved.

www.plantphysiol.orgon February 14, 2018 - Published by Downloaded from Copyright © 2014 American Society of Plant Biologists. All rights reserved.

www.plantphysiol.orgon February 14, 2018 - Published by Downloaded from Copyright © 2014 American Society of Plant Biologists. All rights reserved.

www.plantphysiol.orgon February 14, 2018 - Published by Downloaded from Copyright © 2014 American Society of Plant Biologists. All rights reserved.
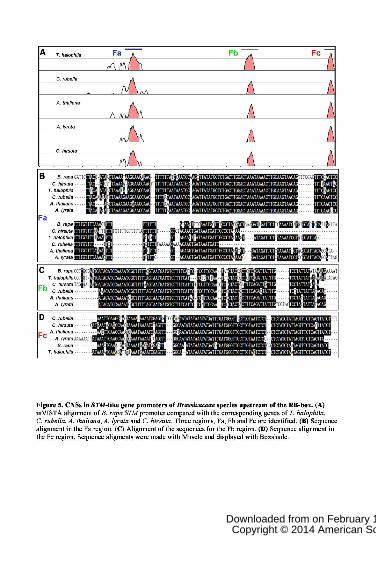
www.plantphysiol.orgon February 14, 2018 - Published by Downloaded from Copyright © 2014 American Society of Plant Biologists. All rights reserved.

www.plantphysiol.orgon February 14, 2018 - Published by Downloaded from Copyright © 2014 American Society of Plant Biologists. All rights reserved.

www.plantphysiol.orgon February 14, 2018 - Published by Downloaded from Copyright © 2014 American Society of Plant Biologists. All rights reserved.