sauerkraut fermentation microbial composition analysis ...mywebcreator.org/imagesppt/s.pdf ·...
TRANSCRIPT

1
Sauerkraut Fermentation Microbial
Composition Analysis Colony PCR of 16S
rDNA For DNA sequencing
Lisa Denys, Rachel Hutchins, Kelly McKinne, Lyle Taylor Department of Biology
Eastern Michigan University
Ypsilanti, Michigan, 48197, USA
Class: BIO 415
Term: Winter 2010
Professors: Daniel Clemans, Steven Francoeur

2
Abstract:
In this paper, microbial succession is studied in the wild fermenting of cabbage. Colony PCR of
16S rDNA is performed for DNA sequencing
Introduction:
Sauerkraut has a long shelf-life compared to other foods. Sauerkraut is prepared through an
aerobic chemical process called fermentation. Fermentation provides a process for making
sauerkraut from cabbage. Bacteria play a key role in the fermentation of cabbage. Leuconostoc
and other species produce lactic acids which give sauerkraut its' characteristic flavor. In
sauerkraut fermentation bacterial species found at the beginning of the fermentative process and
are eventually replaced by more acid tolerant species (Peterson 1969). Mineral salts are used in
the process to produce sauerkraut of good sensory and microbiological quality (viander 2003).
The purpose of this work was to observe microbial succession of bacterial species under aerobic
conditions. Microbial succession is governed mainly by pH (linquist 1998). In this experiment,

3
the natural action of lactose fermenting cultures found in raw cabbage, were utilized to acidify
cabbage. The fermentation trials were made in the laboratory and on a small scale. The process is
a wild fermentation in which no starter culture is used.
Hypothesis: We hypothesize that microbial succession will occur in both batches of sauerkraut I
Introduction
Sauerkraut has a long shelf-life compared to other foods. Sauerkraut is prepared through an
aerobic chemical process called fermentation. Fermentation provides a process for making
sauerkraut from cabbage. Bacteria play a key role in the fermentation of cabbage. Leuconostoc
and other species produce lactic acids which give sauerkraut its' characteristic flavor. In
sauerkraut fermentation bacterial species found at the beginning of the fermentative process and
are eventually replaced by more acid tolerant species (peterson 1969). Mineral salts are used in
the process to produce sauerkraut of good sensory and microbiological quality (viander 2003).
The purpose of this work was to observe microbial succession of bacterial species under aerobic
conditions. Microbial succession is governed mainly by pH (linquist 1998). In this experiment,
the natural action of lactose fermenting cultures found in raw cabbage were utilized to acidify
cabbage. The fermentation trials were made in the laboratory and on a small scale. A wild
fermentation which no starter culture is used.
Material and Methods:
Sauerkraut Preparation: Parallel batches of sauerkraut were fermented using red cabbage from
Busch's and green cabbage from Meijer. Potassium chloride (27.5 grams) was mixed with the
chopped cabbage (1kg) to an approximate concentration of 2.5%. Each batch was fermented in a

4
glass vessel.. Cheese cloth was placed over the top of the vessels to allow gases produced during
the fermentative process to escape. A rubber band was placed around the top of the vessel to hold
the cheese cloth in place.
16S rRNA PCR amplification and PCR product purification:
Full length of 16S rDNA gene was amplified with universal bacterial 8F(5’-AGA GTT TGA
TCC TGG CTC AG-3’) primer, and 1492R (5’-GGT TAC CTT GTT ACG ACT T-3’) primer
(IDT DNA Technologies) following the Vincent Young LAB, 16S Clone Libraries protocol
(Vincent Young LAB, University of Michigan) with these modification; New England Bio Labs
Taq 2x master mix was used instead of PuRe Taq Ready-To- GO PCR beads (New England Bio
Labs, Inc., Ipswitch, MA).
Amplified colony PCR preparation for sequencing and analysis:
The amplified colony 16S rDNA genes from the 8 reactions were cleaned up using EXOSAP-
IT® CLEAN-UP (USB Corporation), and then they were set up for sequencing as described in
Vincent Young LAB 16S Clone Library using 10pmol/ μl of 8F primer. Samples were sent to the
University of Michigan sequencing core (www.SeqCore.brcf.med.umich.edu). The sequences
obtained were then converted from .abi format to FASTA format using
Classification and Phylogenetic tree construction:
All 36 sequences were classified using RDP Classifier. The phylogenetic tree (fig.3) of 36
sequences was generated from all of the sauerkraut sequences using RDP Tree Builder
(https://rdp.cme.msu.edu/index.jsp)

5
Colonies were counted and observed colonies on PCA (Plate Count Agar), MAC (MacConkey
Agar), and HIAS (Heart Infusion Agar) over a thirty five day period using red and green
cabbage. The pH of each batch was measured over the same period. Morphology of the colonies
was observed using light microscopy. The number of lactic acid bacteria was enumerated.
DNA isolation: Total genomic DNA was picked from colonies grown on agar plates. Results
were visualized on .8% gel
16S rDNA PCR amplification, and PCR product purification: full length 16S rRNA gene
was amplified with universal bacterial 8F(5’-AGA GTT TGA TCC TGG CTC AG-3’) primers,
and 1492R(5’-GGT TAC CTT GTT ACG ACT T-3’) primer following Vincent Young LAB,
16S Clone Libraries protocol (Vincent Young LAB, University of Michigan) with these
modification; New England Bio Labs Taq 2x master mix was used instead of PuRe Taq Ready-
To- GO PCR beads (New England Bio Labs, Inc., Ipswitch, MA).
PCR condition was also slightly modified from Vincent Young LAB, 16S Clone Libraries
protocol. Amplification cycle used with 8F, and 1492R were as follow; an initial denaturation
92°C for 2 min, followed by 30 cycles of denaturation 94°C for 30 s, annealing 50°C for 45 s,
extension 72°C for 1.5 min, and final extension step 72 °C for 5 min using Bio-Rad MJ Mini
Personal Thermal Cycler (Bio-Rad Laboratories, Hercules).
Amplified colony PCR preparation for sequencing and analysis: the amplified colony 16S
rRNA genes from the 96 reactions total were cleaned up using EXOSAP-IT® CLEAN-UP (USB
Corporation), and then they were set up for sequencing as described in Vincent Young LAB 16S
Clone Library using 10pmol/ µl of 8F primer. Samples were to the University of Michigan
sequencing core (SeqCore). The sequences obtained were then converted from .abi format to

6
.fasta format. The data were uploaded into and analyzed using the Ribosomal Data Base Project
website (http://rdp.cme.msu.edu/myrdp/overview.spr ).
Results:
Plating:
Acidity decreased rapidly in the beginning of the fermentations (Wiande 2009). Lactic acid
bacteria were "tmtc" on HIAS plates from day 0 in the green cabbage. Lactic acid counts were
4.30E+03 cfu’s/ml on day 0 in the red cabbage. On day 35, in the green cabbage, lactic acid
counts were 8.00E+06 cfu’s/ml. PCA counts were "tmtc" in both red and green cabbage over the
entire range of platings. On the MAC plates which enriched for gram negative bacteria. Gram
negative bacteria showed the greatest number of colonies on day 0 with 8.00E+05 cfu’s/ml), and
day 2 8.00E+06 cfu’s/ml. In the green batch of cabbage, slime forming colonies were observed
on plates from day 35 HIAS medium.
PCR amplification of 16S rRNA gene: Out of 25 colonies (from red and green cabbage)
sequenced, we only obtained 5 sequences due to unsuccessful PCR amplification. Nucleotides
were more than 1000 bp (Table 1) from the NAST analysis (DeSantis 2006). The alignment was
assigned based on minimum sequence length of 900. Looking at the gel, some of the bands are
missing suggesting that the 8F and 1492R primers didn’t amplify the 16S rRNA gene efficiently.
This corresponds with the debris found at the bottom of the gel (Fig. 1, 2). Among the eight 16S
rDNA sequences, five were Leuconostoc. There was one Pantoea agglomerans, one Serratia, and
one Pseudomonas fluorescens was found with > 95% identity match for each. Full fragment
length of 16S rDNA gene (Approximately 1500 bp) was obtained from total genomic DNA
extract when universal bacterial 8F and 1492R primers were used (Figure 1).

7
15
47
89
5
15
47
89
6
15
47
89
7
Fig. 1
15
47
89
8
1547
899
Figure 2
Table 2 summarizes the species that were found during plating. Leuconostoc mesenteroides are
gram-positive microorganisms with coccoid morphology (Facklam 1995) Weissella cibaria is
associated mainly from Malaysian foods and clinical samples from humans (Björkroth 2002).
Pantoea agglomerans is a plant pathogen causing human disease(Cruz et al 2007). Pseudomonas
fluorescens suppresses black root rot of tobacco, a disease caused by the fungus Thielaviopsis
basicola. The strain CHAO excretes several metabolites with antifungal properties. One such
metabolite is hydrogen cyanide (Voisard et al 1989). Pseudomonas fluorescens is inherently
Figure 1. A successful 1% gel of
standard PCR amplification of 16S
rRNA gene from red and green
cabbage.
Figure 2. Gel Bottom

8
resistant to a variety of antimicrobial agents (Sillankorva et al 2008) Rahnella enterobacterial
species causes bacteremia and septicemia, as well as endocarditis, urinary tract infections, and
wound infections, mostly in immunocompromised patients (Caroff et al). R. aquatilis is resistant
to amoxicillin, ticarcillin, cefuroxime, and cephalothin(Bellais et al 2001) and carry the carry the
blaRAHN gene. Serratia is a gram-negative, facultative anaerobic, rod-shaped bacteria of the
Enterobacteriaceae family
Table 1
* samples borrowed from JEB, no sequence data available
Discussion
The potassium chloride used in the fermentation process serves two major purposes. First, it
causes an osmotic imbalance which results in the release of water and nutrients from the cabbage
leaves. Second, the mineral salt inhibits the growth of many spoilage organisms (Linquist 1998).
Vegetables left exposed to air start to grow molds(Katz 2003). On day 7 we observed what
appeared to be mold growth in the red cabbage fermentation. We added approximately 200 ml
of saline solution to the red cabbage on day 7 and day 14 to increase the distance between the top
of the sauerkraut and the level of mold growth. Mold growth may have been due to pockets of
low or high potassium chloride concentrations in the red cabbage. It is also possible that the
mold growth was due to the red batch being exposed to air (Linquist 1998). In the red batch, an
Erlenmeyer flask was used instead of a beaker to weigh the sauerkraut down in the vessel. Since
the surface are of the bottom of the Erlenmeyer flask was smaller than that of the beaker, the red
cabbage might have been exposed to too much air.

9
Conclusion
Evidence of microbial succession was found. Slime forming pseudomonas appeared on day 0
PCA plates, in both batches, this evidence was further supported by the results of sequence
analysis. The count of slime forming colonies increased from day 0 through day 21 in both
batches. Pantoea agglomerans was identified in day 0 sequence analysis of MAC colonies.
Acidity ranged from 7 on day 0, to 4.5 on day 35 in both the red and green cabbage. Growth on
MAC plates was initially high and decreased to 0 on days 21 and 35.

10
Table 2
Field3 Field5
"1547895" WHITE_PCA_-7_DIL_DAY_2_PH_5_GR_NA_GL_11_rc Leuconostocacea
"1547896" RED_MAC_-3_DIL_DAY_7_PH_4.5_GR_NA_GL_13_gc Leuconostocaceae
"1547897" RED_MAC_-2_DIL_DAY_14_PH_4.5_GR_NA_GL_15_gc Leuconos
"1547898" SLIMY_HIAS_-4_DIL_DAY_21_PH_4.5_GR_NA_GL_22_gc Weissella cibaria Leuconostoc
"1547899" SLIMY_HIAS_-5_DIL_DAY_35_PH_4.5_GR_P_GL_25_rc Leuconostocace
"1547900" DAY2_MAC_RANDAZZO Serratia *
"1547901" DAY2_MAC_RANDAZZO Pantoea agglomerans *
"1547903" DAY0_PCA_MEIJER Pseudomonas fluorescens *
"1547904" JEB Pseudomonas
"1547905" JEB Acinetobacter
"1547909" GA Pseudomonas
"1547910" GA Massilia
"1547911" GA Pseudomonas
"1547912" GA Pantoea agglomerans
"1547914" DEA Pseudomonas
"1547915" DEA Leuconostoc
"1547916" DEA Leuconostoc
"1547917" DEA Leuconostoc
"1547918" DEA Leuconostoc
"1547919" DEA Leuconostoc
"1547920" DEA Leuconostoc
"1547921" DEA Leuconostoc
"1547922" DEA Leuconostoc
"1547923" DEA Micrococcaceae genus Arthro
"1547924" DEA Pseudomonas
"1547925" JEB Brevundimonas
"1547926" JEBS Acinetobacter
"1547927" JEB Massilia
"1547928" JEB Chryseobacterium
"1547929" JEB Pseudomonas
"1547930" JEB Pseudomonas
"1547931" JEB Pseudomonas
"1547932" JEB Pseudomonas
"1547934" JEB Erwinia
"1547935" JEB Erwinia
"1547936" JEB Hymenobacter gelipurpurascens genus Hymeno

11
Figure 3
References
Pederson CS, Albury MN (1969) The Sauerkraut Fermentation.
NY Agric Exp Station Tech Bull 824
Viander B, M_ki M, Palva A (2003) Impact of low salt concentration,
salt quality on natural large-scale sauerkraut fermentation.
Food Microbiol 20
Lindquist, J.editor. 1998. Laboratory Manual for the Food Microbiology Laboratory
University of Wisconsin – Madison
Wiande, B., Ryhanen, E.L. Laboratory and large-scale fermentation of white cabbage
into sauerkraut and sauerkraut juice by using starters in combination
with mineral salt with a low NaCl content. 2009. Springer-Verlag.
Hammes WP (1990) Bacterial starter cultures in food production.
Food Biotechnol 4(1)
Björkroth, K. J., U. Schillinger, R. Geisen, N. Weiss, B. Hoste, W. H. Holzapfel, H. J. Korkeala,
and P. Vandamme. 2002. Taxonomic study of Weissella confusa and description of Weissella
cibaria, a novel species detected in food and clinical samples. Int. J. Syst. Evol. Microbiol. 52

12
Facklam R, Elliott JA. Identifi cation, classification, and clinical relevance
of catalase-negative, gram-positive cocci, excluding streptococci
and enterococci. Clin Microbiol Rev. 1995
Cruz AT, Cazacu AC, Allen CH. Pantoea agglomerans, a plant pathogen causing human disease.
J Clin Microbiol. 2007;45:1989–1992
Voisard, C, Keel, C., Haas, D., and Defago, G. (1989). Cyanide pmduction
by Pseudomonas fluorescens helps suppress black root rot of
tobacco under gnotobiotic conditions. EMBO J. 8, 351-358.
Bellais S, Poirel L, Fortineau N, Decousser JW, Nordmann P. Biochemical-genetic
characterization of the chromosomally encoded extended-spectrum class A β-lactamase from
Rahnella aquatilis. Antimicrob Agents Chemother. 2001;45:2965–2968. doi:
10.1128/AAC.45.10.2965-2968.2001.
Rozhon, W., Petutschnig, E., Khan, M., Summers, D., and Poppenberger, B. (2010). Frequency
and diversity of small cryptic plasmids in the genus rahnella. BMC Microbiology, 10:56+.
Sillankorva S, Neubauer P, Azeredo J. Pseudomonas fluorescens Biofilms Subjected to Phage
phiIBB-PF7A. BMC Biotechnol. 2008;8:79
Caroff, N., C. Chamoux, F. Le Gallou, E. Espaze, F. Gavini, D. Gautreau, H. Richet, and A.
Reynaud. 1998. Two epidemiologically related cases of Rahnella aquatilis bacteremia. Eur. J.
Clin. Microbiol. Infect. Dis. 17:349-352
Roberts, G.,and Paustian, T. 2009. Through the Microscope Third Edition
Textbook Consortia
Katz, S.E. 2003. Wild Fermentation: The Flavor, Nutrition, and Craft of Live-Culture Foods
Chelsea Green Publishing Company.
DeSantis, T. Z., P. Hugenholtz, K. Keller, E. L. Brodie, N. Larsen, Y. M. Piceno, R. Phan, and
G. L. Andersen. 2006. NAST: a multiple sequence alignment server for comparative analysis of
16S rRNA genes. Nucleic Acids Res 34:W394-9.
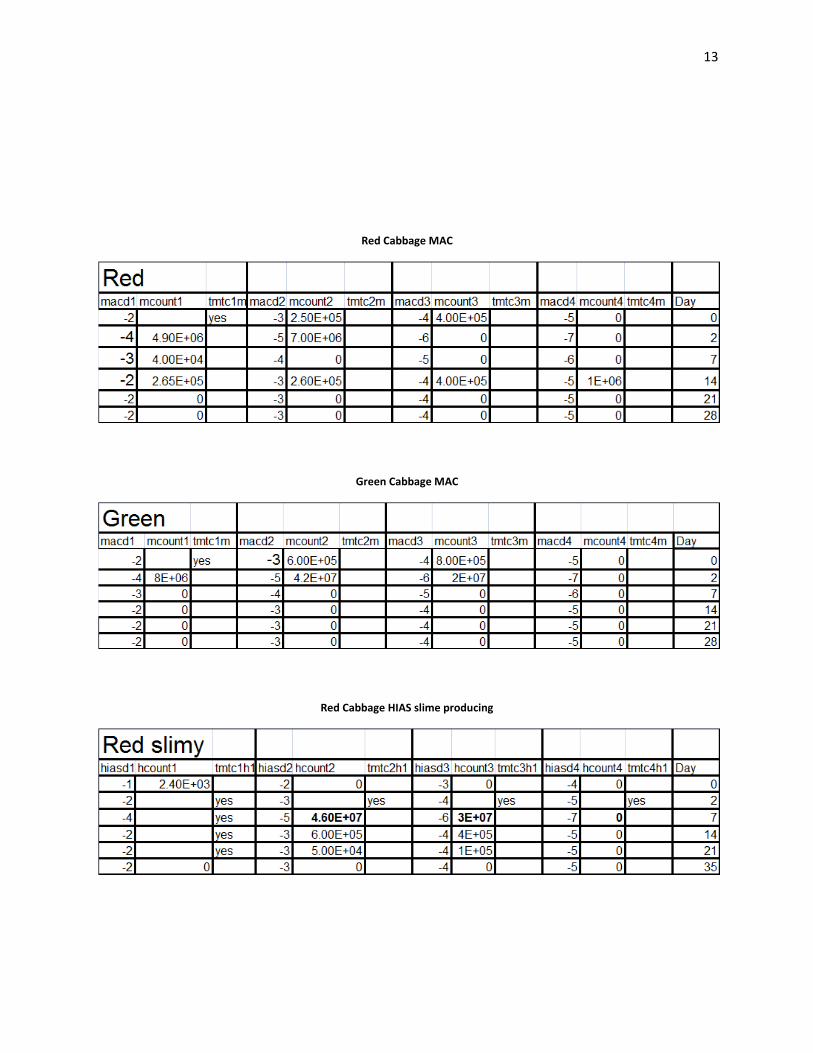
13
Red Cabbage MAC
Green Cabbage MAC
Red Cabbage HIAS slime producing
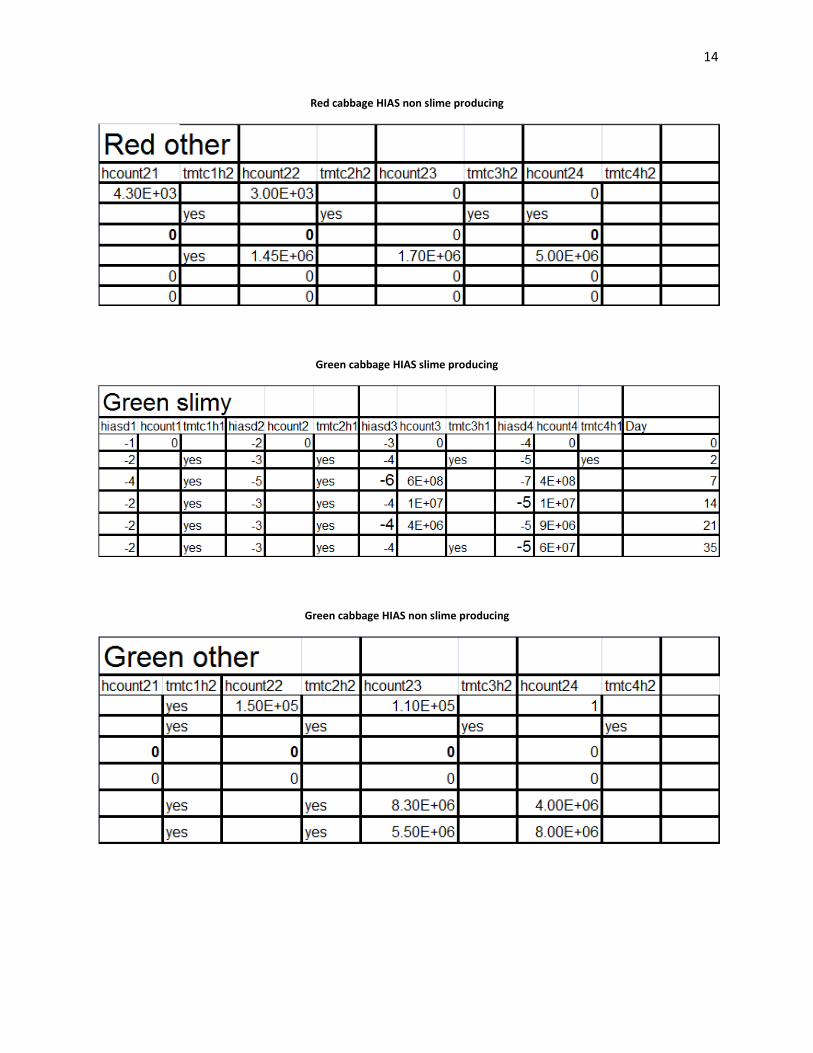
14
Red cabbage HIAS non slime producing
Green cabbage HIAS slime producing
Green cabbage HIAS non slime producing
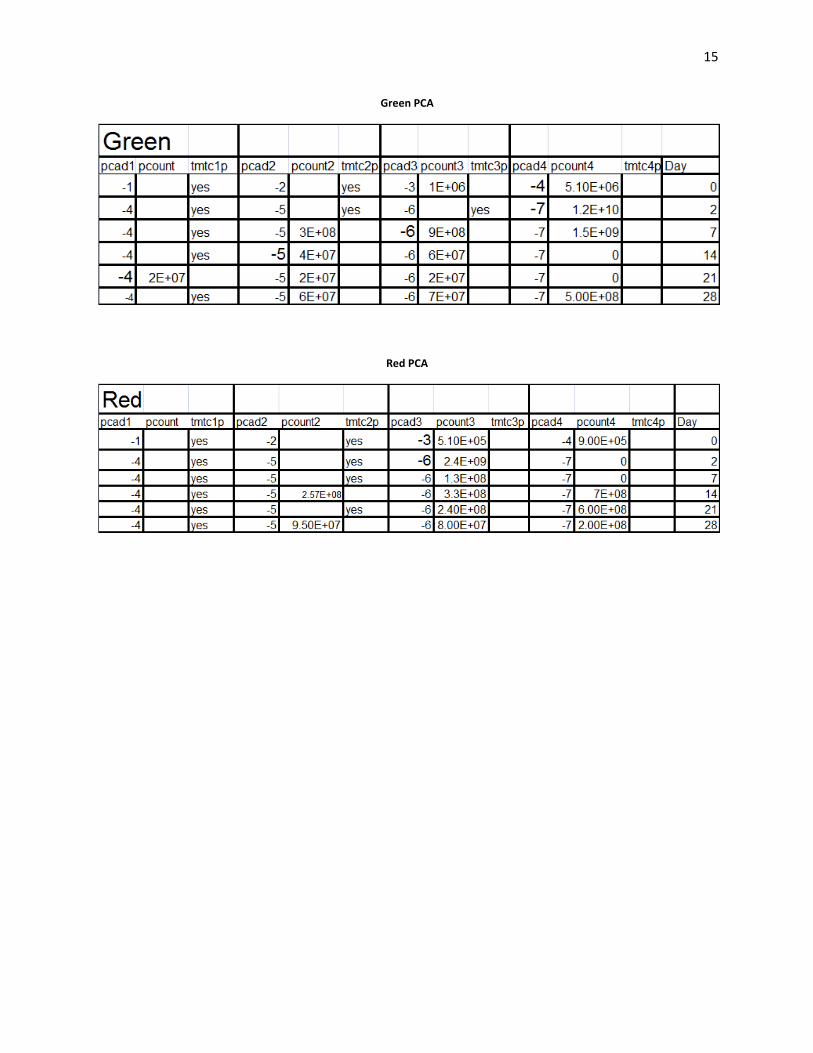
15
Green PCA
Red PCA