scanning electron microscopy cristispira species … · v41cristispiraspp. in chesapeake bayoysters...
TRANSCRIPT

Vol. 42, No. 2APPLIED AND ENVIRONMENTAL MICROBIOLOGY, Aug. 1981, p. 336-3430099-2240/81/080336-08$02.00/0
Scanning Electron Microscopy of Cristispira Species inChesapeake Bay Oysters
BEN D. TALL AND ROBERT K. NAUMAN*Department ofMicrobiology, University ofMaryland Dental School, Baltimore, Maryland 21201
Received 6 February 1981/Accepted 6 May 1981
Scanning electron microscopy was employed to observe the physical interac-tions between Cristispira spp. and the crystalline style of the Chesapeake Bayoyster (Crassostrea virginica Gmelin 1791). Cristispira organisms were foundassociated with both the inner and outer layers of the posterior two-thirds of thestyle. The spirochetes possessed blunt-tipped ends, a cell diameter range of 0.6 to0.8 ,um, and distended spirochetal envelopes which followed the contour of thecells. Transmission electron microscopy showed that the distension of the enve-lope was probably due to the containment of numerous axial filaments. Inaddition, they were found to possess two distinct spiral shapes which weredependent on whether their location was inside or on the surface of the style.
Cristispira is a genus of large spirochetes col-onizing the crystalline style of a variety of mol-lusks. They have been characterized on the basisof their host species and the fact that theypossess no known pathological involvement withthese hosts. Morphologically, they are distin-guished from other pathogenic and nonpatho-genic spirochetes by the presence of a ridge orcrestlike structure called the "crista." The cristacan be visualized by bright-field (4), dark-field(13), and electron microscopy (15). Transmissionelectron microscopy shows the crista to consistof a closely packed bundle of numerous axialfilaments bound between the outer sheath andthe protoplasmic cylinder (14). Certes (4) wasthe first to document the presence of these spi-rochetes and observed them inhabiting thedigestive tract of several species of oysters of thegenus Ostrea. However, it was Gross (3) who, in1910, proposed the genus Cristispira. Studies ofCristispira spp. have been hampered by theinability to achieve in vitro growth (2, 6, 12, 13).It is probable that the spirochetes found associ-ated with the style are closely tied to the func-tional activity of the style. Therefore, it was feltthat a study of the physical interactions betweenCristispira spp. and the crystalline style of theoyster would provide additional informationabout this unusual ecological relationship. Onetechnique useful in studying these interactionsis scanning electron microscopy (SEM). Thispaper describes SEM observations of the phys-ical interactions between Cristispira spp. andthe crystalline style of the American oyster,Crassostrea virginica (Gmelin 1791).
MATERIALS AND METHODS
Collection of colonized styles. Crystalline styleswere obtained from hand-dredged oysters collectedfrom various oyster beds located in the upper Chesa-peake Bay. The oysters were immediately openedwhen removed from the water, and the presence of astyle was determined by making a lateral incision intothe visceral mass initiating from the mantle cap, con-tinuing along the anterodorsal border (border oppositethe gills), and terminating dorsal to the cloacae. If astyle was found, the anterior end was grasped withforceps and gently removed from the surroundingtissue.The freshly excised style was placed on a clean glass
microscope slide and examined for the presence ofCristispira organisms by bright-field microscopy(x100). Colonized styles were then prepared for SEMas described below.SEM. Freshly excised and colonized styles were
prefixed in 1% glutaraldehyde-filtered bay water (pH6.0; salinity, 13.9 ppt) for 3 h at 4°C. Bay water usedin this study was first adjusted to the desired salinityby the addition of sea salts (Marinemix, HawaiianMarine Imports, Inc., Houston, Tex.) and then filteredthrough a 0.2-,im membrane filter (Millipore Corp.,Bedford, Mass.). The styles were then gently washedwith three changes of filtered bay water (pH 6.0, 13.9ppt) and postfixed in 1% osmium tetroxide-filtered baywater (pH 6.0; 13.9 ppt) for 24 h at 25°C. The styleswere once again washed with three changes of filteredbay water and then dehydrated with a graded series ofethanol (30 to 100%), critical-point dried with C02, andsputter-coated with gold or gold palladium. Specimenswere viewed in a JEOL (JSM T-20) SEM or an AMR1000 SEM. Micrographs were recorded with Polaroidfilm (type 55 P/N, 4 by 5 in. [ca. 10 by 12.5 cm]) orKodak commercial 4127 film.Transmission electron microscopy. Negatively
336
on April 17, 2019 by guest
http://aem.asm
.org/D
ownloaded from

CRISTISPIRA SPP. IN CHESAPEAKE BAY OYSTERS 337
stained specimens of whole cells for electron micros-copy were prepared as follows. A colonized style ex-cised from an oyster by the method described abovewas placed in a 1-dram (ca. 3.9-g) screw-capped vialcontaining the filtered bay water (pH 6.0; 13.9 ppt).The vial was then placed in ice until it was returnedto the laboratory (approximately 3 h, during whichtime the style dissolved). The cell suspension was thencentrifuged at 3,000 x g for 10 min. The supernatantwas discarded, and the pellet was resuspended in 0.1ml of the filtered bay water. A drop of the cell suspen-sion was placed on a 0.25% Formvar carbon-coatedcopper grid (300 mesh). Excess fluid was withdrawnfrom the grid surface with filter paper, and the gridwas immediately stained with 1% (wt/vol) sodiumphosphotungstate (pH 7.3) for 15 to 20 s. Excess stainwas removed from the grid, and the grid was air driedbefore the specimen was examined in a Siemens IAtransmission electron microscope operating at 80 kV.
RESULTSStyle morphology. The style possessed an
overall club-shaped appearance, with the foodtassel end (anterior) tapering towards the stylesac end (posterior) (Fig. 1). The wrap-aroundappearance of the style (Fig. 1) is thought to bean artifact produced during specimen prepara-tion, since other styles examined did not dem-onstrate this appearance. Close examination ofthe styles (Fig. 2) revealed a spongy inner layercovered by a much smoother outer layer.Spirochete-style interaction. SEM re-
vealed these organisms to possess blunt-tippedends, a cell diameter range of 0.6 to 0.8 ,um, anddistended spirochetal envelopes which followedthe contour of the cells (see Fig. 5). Negativelystained specimens suggested that the distensionof the envelope was probably due to the contain-ment of the numerous axial filaments foundassociated with the cell (Fig. 3 and 4).
Critsispira organisms were seen in all areas ofthe crystalline style except the area encompass-ing the food tassel end. They were seen associ-ated with both the inner and outer layers of thestyle (Fig. 5, 6, and 7). Closer examination of thespirochetes within these two layers revealedsome spirochetes to be tightly coiled, whereasothers were loosely coiled. Those organisms ob-served just below the outer surface and, possibly,within the style inner matrix were tightly coiled(Fig. 7 and 8), whereas those emerging throughand adhering to the style surface were looselycoiled (Fig. 7, and see Fig. 10). Spirochetes ob-served in the interior of the style occupied pore-like openings and were seen to protrude fromthese openings (Fig. 9). The inner portion ofthese openings or tunnels displayed a smootherappearance than that seen for the style innermatrix, thus suggesting these openings or tun-nels resulted from the movement by the spiro-
chetes within the inner layers of the style. Othermarine microorganisms were observed alongwith Cristispira organisms, primarily at the pos-terior end of the style (Fig. 10).
DISCUSSIONThe observations reported here were of C.
virginica (Gmelin 1791) with its host-associatedspirochetes. These spirochetes, as determinedfrom their characteristic appearance, fine struc-ture, and unusual habitat, are believed to bemembers of the genus Cristispira, and the con-clusions drawn here are likely to be pertinent toother Cristispira spp.-style interactions found inother species of mollusks which harbor theseorganisms. Furthermore, evidence presented inthis study correlates nicely with the work ofRyter and Pilot (14) and with the descriptionpresented in the 8th edition of Bergey's ManualofDeterminative Bacteriology (10). Principally,the reports of early investigators centeredaround taxonomic, staining, and morphologicalproperties (3-6, 12, 14). The conclusions whichevolved from these early studies were that Cris-tispira organisms are spiral shaped and live inassociation with the intestinal tract of bivalvemollusks and that these organisms, because oftheir shape and possession of axial filaments, arespirochetes and should be classified as such.The early literature described a structure
which was referred to as the crista. Earlier in-vestigators believed this structure to be an un-dulating membrane (4). However, the studies ofRyter and Pilot (14) and the observations re-ported here indicate that the crista is a disten-sion of the outer layer of these organisms createdby the numerous axial filaments (Fig. 3 and 4)found between the outer layer and the proto-plasmic cylinder.More recently, Ingham (E. R. Ingham, Abstr.
Annu. Meet. Am. Soc. Microbiol. 1977, N10, p.230) reported using SEM and observed Cris-tispira organisms adhering to the outer surfaceof the oyster style.The findings presented here, however, showed
not only surface-adhering spirochetes but alsospirochetes within the inner matrix of the style(Fig. 5, 7, and 10). A possible reason for thedifferences observed may simply be the timerequired for the spirochetes to colonize the style.The longer the feeding oyster is left undisturbed,the greater likelihood of the style being formed,and therefore, a greater degree of colonization ofthe style may occur. However, the nature of thisrelationship is not well understood.Judd (9) observed Cristispira organisms
within the crystalline style ofAmphidemsa aus-trale. He used a carbohydrate-specific periodic
VOL. 42, 1981
on April 17, 2019 by guest
http://aem.asm
.org/D
ownloaded from

338 TALL AND NAUMAN
FIG. 1. Scanning electron micrograph of the crystalline style of C. virginica which was cut into two piecesto facilitate mounting onto the specimen stub. Bar = 1.0 mm.
FIG. 2. Scanning electron micrograph ofa crater-like area in a portion of the style illustrating its layerednature. Arrow points to spirochete observed in crater. Bar = 10 tun.
acid-thiosemicarbazide-silver proteinate stain-ing technique (PATSCSP). Using this stain, heconcluded that the style was composed of car-bohydrates and that the reactions he observedwith the PATSCSP stain were indicative of gly-coproteins. Judd believed that Cristispira orga-nisms enzymatically removed the carbohydrate
moiety from these glycoproteins. His interpre-tation was based on clear areas surroundingspirochetes in thin sections of styles. Althoughhis interpretation may be correct, the zones ofclearing may also be due to the inability of thespirochete envelope to take up the PATSCSPstain. Still another possibility is suggested by
APPL. ENVIRON. MICROBIOL.
on April 17, 2019 by guest
http://aem.asm
.org/D
ownloaded from

V1CRISTISPIRA SPP. IN CHESAPEAKE BAY OYSTERS 339
FIG. 3. Electron micrograph of a negativelystained specimen. Note the numerous axial filaments.Bar = 1.0 ,m.
FIG. 4. Electron micrograph of a higher magnifi-cation ofone end of the spirochete seen in Fig. 3. Bar= 1.0 PLm.
close examination of Fig. 9. The clear areasreported by Judd could very well be the tunnelswhich resulted from the spirochetes' movementthrough the style.The two spiral shapes of Cristispira organisms
observed in different areas of the style lendsupport to the studies of Greenberg and Canale-Parola (7). They studied the effect of viscosityon the motility of several spirochetes. Theirstudies revealed a correlation between cell coil-ing and the ability of spirochetes to swim inviscous environments. The two different spiralshapes observed in the present study are prob-ably examples of spirochetes moving in two dif-ferent viscous environments. Cristispira orga-
nisms on the surface and emerging from theinner layers come in contact with a less viscousenvironment; thus, a tightly coiled morphologyis not required for motility. Conversely, thoseorganisms found within the style require atightly coiled morphology to translocate in themore viscous environment. It is possible that thedifferent cell types observed could represent twodifferent populations of cells. However, thescope of this study does not permit us to deter-mine this. Another possibility exists that the twocoiling morphologies observed could be artifac-tual, resulting from SEM preparation proce-dures. This seems unlikely because the orga-nisms were immediately fixed in 1% glutaralde-
VOL. 42, 1981
'41 m4
4, i.
I, I*4
A,
on April 17, 2019 by guest
http://aem.asm
.org/D
ownloaded from
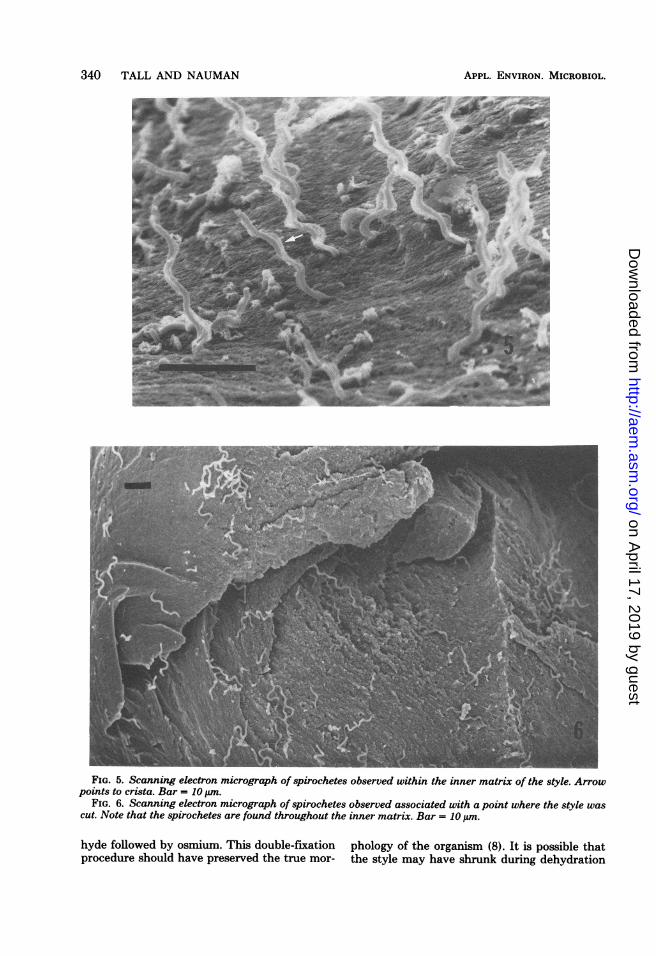
340 TALL AND NAUMAN
FIG. 5. Scanning electron micrograph of spirochetes observed within the inner matrix of the style. Arrowpoints to crista. Bar = 10 ,um.
FIG. 6. Scanning electron micrograph of spirochetes observed associated with a point where the style wascut. Note that the spirochetes are found throughout the inner matrix. Bar = 10 ,tm.
hyde followed by osmium. This double-fixationprocedure should have preserved the true mor-
phology of the organism (8). It is possible thatthe style may have shrunk during dehydration
APPL. ENVIRON. MICROBIOL.
on April 17, 2019 by guest
http://aem.asm
.org/D
ownloaded from
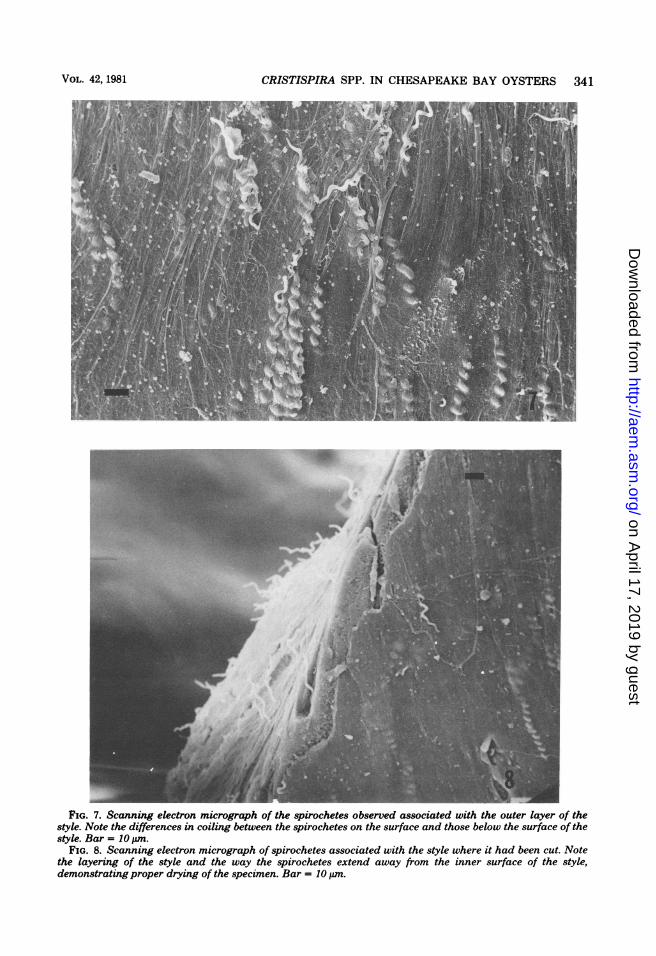
V41CRISTISPIRA SPP. IN CHESAPEAKE BAY OYSTERS 341
FIG. 7. Scanning electron micrograph of the spirochetes observed associated with the outer layer of thestyle. Note the differences in coiling between the spirochetes on the surface and those below the surface of thestyle. Bar = 10 ,um.
FIG. 8. Scanning electron micrograph of spirochetes associated with the style where it had been cut. Notethe layering of the style and the way the spirochetes extend away from the inner surface of the style,demonstrating proper drying of the specimen. Bar = 10 tum.
VOL. 42, 1981
on April 17, 2019 by guest
http://aem.asm
.org/D
ownloaded from

342 TALL AND NAUMAN
u +_:.FIG. 9. Scanning electron micrograph illustrating the pore-like openings occupied by the spirochetes
(arrows). Bar = 5 tmn.FIG. 10. Scanning electron micrograph of the posterior end of the style. Note the loosely coiled shape of
spirochetes adhering to the surface in addition to other marine microorganisms. Bar = 10 um.
and critical-point drying; however, the orga-nisms should have maintained their prefixationmorphology during this process.The findings presented here generally corrob-
orate those of other investigators. Most impor-
tantly, they confirm that Cristispira organismsadhere to the outside of the style besides beingassociated with the inner layers. Therefore, theirmovement within the style suggests they obtainenergy from either the style itself or some part
APPL. ENVIRON. MICROBIOL.
on April 17, 2019 by guest
http://aem.asm
.org/D
ownloaded from

VL421CRISTISPIRA SPP. IN CHESAPEAKE BAY OYSTERS 343
of the style. However, the reason this organelleharbors Cristispira organisms remains un-known.
ACKNOWLEDGMENTSWe thank M. J. Garreis and her co-workers, G. Johnson
and W. Beatty, from the State of Maryland Department ofGeneral Sanitation, for their excellent technical assistance inobtaining oyster specimens. We also like to express our grati-tude to G. Centola and R. Pendergass for their technicalassistance concerning SEM.
LITERATURE CITED1. Bailey, K., and B. D. Worboys. 1960. The lamelibranch
crystalline style. Biochem. J. 76:487-491.2. Berkeley, C. 1959. Some observations on Cristispira in
the crystalline style of Saxidomus gigonteus (De-shayes) and in that ofsome other lamellibrachiata. Can.J. Zool. 37:53-58.
3. Breznak, J. A. 1973. Biology of non-pathogenic host-associated spirochetes. Crit. Rev. Microbiol. 2:457-489.
4. Certes, A. 1882. Les parasites et les commensaux del'huite. Bull. Soc. Zool. Fr. 7:347-353.
5. Dobell, C. 1911. On Cristispira veneris nov. species andthe affinities and classification of spirochetes. Q. J.Microbiol. Sci. 56:507-541.
6. Fantham, H. B. 1908. Spirochaeta (Trypanosoma) bal-
bianii (Certes) and Spirochaeta anodontae (Keysse-litz); their movements, structure, and affinities. Q. J.Microbiol. Sci. 52:1-69.
7. Greenberg, E. P., and E. Canale-Parola. 1977. Rela-tionship between cell coiling and motility of spirochetesin viscous environments. J. Bacteriol. 131:960-969.
8. Hayat, M. A. 1970. Principles and techniques of electronmicroscopy. Biological applications, p. 65-107. Van Nos-trand Reinhold Co., New York.
9. Judd, W. 1979. The secretions and fine structure of thebivalve crystalline style sacs. Ophelia 18:205-233.
10. Kuhn, D. 1974. Order I Spirochaetales nomen novum, p.171. In R. E. Buchanan and N. E. Gibbons (ed.), Ber-gey's manual of determinative bacteriology, 8th ed. TheWilliams & Wilkins Co., Baltimore.
11. Nelson, T. C. 1918. On the origin, nature and function ofthe crystalline style of lamellibrachiata. J. Morphol. 31:53-111.
12. Noguchi, H. 1921. Cristispira in North American shell-fish; a note on a spirillum found in oysters. J. Exp. Med.34:295-315.
13. Perrin, W. A. 1906. Researches upon the life history ofTrypanosoma balbianii (certes). Arch. Protistenkd. 7:131-156.
14. Ryter, A., and J. Pilot. 1965. Structure des spirochetes.H. Etude der genre Cristispira au microscope optiqueet au microscope electronique. Ann. Inst. Pasteur Paris109:552-562.
VOL. 42, 1981
on April 17, 2019 by guest
http://aem.asm
.org/D
ownloaded from