scatter hoarding by kangaroo rats (dipodomys … · scatter hoarding by kangaroo rats (dipodomys...
TRANSCRIPT

Scatter hoarding by kangaroo rats(Dipodomys merriami) andpilferage from their caches
Martin DalyLucia F. JacobsMargo I. WilsonPhilip R. BehrendsDepartment of PsychologyMcMaster University,Hamilton, Ontario L8S 4K1,Canada
Address reprint requests to M.Daly. L. F. Jacobs is now at theDepartment of Psychology,University of Utah, Salt LakeCity, UT 84112, USA. P. R.Behrends is now at MichaelBrandman Associates, 7676Hazard Center Drive, San Di-ego, CA 92108, USA.
Received 15 August 1990Revised 27 May 1991Second revision 13 August1991Accepted 14 August 19911045-2249/92/14.00© 1992 International Societyfor Behavioral Ecology
We observed radio-implanted Merriam's kangaroo rats disposing of 10-g bonanzas of rolledoats in 48 trials in the field. The principal determinant of the initial disposition of discoveredfood was apparently its distance from the day burrow: food found within about 10m was mainlylarder hoarded, whereas food encountered farther afield was usually dispersed immediately inshallow caches. Cache sites were newly dug for the purpose and not reused; most caches werenearer the current day burrow than was the food source, but a few were placed far from boththe cacher's day burrow and its habitual nocturnal range. An experiment with artificial cachesindicated that security from discovery increases with spacing and with proximity to perennialshrubs. Nine kangaroo rats cached dyed food, and fecal dye traces revealed extensive pilferagefrom five of them, by both conspecifics and other rodent species. Limited evidence indicatesthat food encountered nearer home and initially larder hoarded was more secure from pilferagethan food initially scattered, and yet kangaroo rats were observed to scatter caches soon afterinitial larder hoarding. A kangaroo rat whose dyed stores escaped pilferage fed from them atintervals for at least 12 days. Even cachers who incurred pilferage made as much, or more, useof their caches as any thief, suggesting that scattering caches may be a defense against cata-strophic losses. [Behav Ecol 3:102-111 (1992)]
Morris (1962) coined the term "scatterhoarding" to describe the behavior of
dispersing stored food in multiple, small pack-ets. Because spatial dispersions of stored foodcan vary continuously, Vander Wall (1990)distinguishes scatter hoarding from larderhoarding on the basis of single versus multipledepositions to the storage site; we refer to thetwo sorts of. stores as "caches" versus "lar-ders." Larder hoarding is more prevalent inmammals, even with foods like seeds that arereadily partible into small caches (Smith andReichman, 1984), but a number of rodentspecies scatter nuts and seeds (Vander Wall,1990).
If the function of hoarding is to sequesterfood from competitors for later use, a con-centrated larder is practical only if it is de-fensible or well hidden. It follows that larderhoarding tends to be associated with territo-rial exclusion and that larders are often sit-uated in occupied dens near the territory'scenter, in accordance with considerations ofboth larder security and central-place forag-ing economics (Orians and Pearson, 1979).
The adaptive logic of scatter-hoarding de-cisions is more complex and less well under-stood (Covich, 1987; Kramer and Nowell,1980; Sherry, 1985; Vander Wall, 1990). Scat-
ter hoarding is apparently more time consum-ing than larder hoarding and certainly placesmore demands on memory (Balda et al., 1987;Jacobs, 1991); moreover, scatter hoardingmakes one's food stores less handy when in-clement weather or other circumstances im-pede excursion. One possible explanation forscatter hoarding is that it reduces the risk ofmajor losses to cache robbers rather than sim-ply minimizing the expected mean loss (Hurlyand Robertson, 1990; Morris, 1962). Deci-sions about the number, size, specific locales,and spatial dispersion of caches influence theexpected costs of scatter hoarding, and thesecosts must be elucidated before the behaviorcan be understood. Theory and research havedealt with the time and travel costs of cachingand cache recovery and with the impact of thespacing of artificial caches on cache survival(reviewed by Vander Wall, 1990). However,almost nothing is known about pilferage fromthe actual caches of scatter hoarders.
Kangaroo rats (Dipodomys; Rodentia: Het-eromyidae) are nocturnal, burrow-dwellingrodents of North American arid zones. Theyforage in open ground and transport the seedsthey find in external fur-lined cheek pouchesto be consumed or stored elsewhere. Hoard-ing practices and use of space vary within the
102 Behavioral Ecology Vol. 3 No. 2
at McM
aster University L
ibrary on July 30, 2016http://beheco.oxfordjournals.org/
Dow
nloaded from

genus. D. spectdbilis builds and occupies de-fended mounds (Jones, 1984; Randall, 1984),around which activity is concentrated and cen-tered (Schroder, 1979) and in which substan-tial stores are amassed (Vorhies and Taylor,1922). InD. merriami, by contrast, home rang-es overlap extensively (Behrends et al., 1986),and nesting burrows (day burrows) are lesselaborate, are typically situated nearer the pe-riphery than the center of the foraging range(present authors' unpublished data), are fre-quently switched or abandoned (Behrends etal., 1986), and seldom contain substantialstores (present authors' unpublished data;Kenagy, 1973; Monson and Kessler, 1940).
As the contrasts above suggest, D. merriamiappears to be mainly a scatter hoarder. Incaptivity, this species buries seeds in cachescontaining about 1 g of seeds and, like otherscatter-hoarding birds and mammals, has anexcellent memory for cache locations (Jacobs,1991). Reynolds (1958) set out small piles ofnon-native seeds and found caches, which heattributed to D. merriami, when seeds sprout-ed after a rain; he reported that the shallowcaches were distributed in "random groups"around feeding stations at a mean distance of14 m (range: 0.6-32 m). The travel necessaryfor establishing and recovering caches is de-monstrably risky activity: in comparisonsamong contemporaneously radio tracked D.merriami of both sexes, individuals who trav-eled the farthest incurred the greatest risk ofbeing preyed upon (Daly et al., 1990). Nev-ertheless, D. merriami pursues a wide-ranging,scatter-hoarding mode of existence that evi-dently demands greater travel than the terri-torial, central-place-foraging alternative prac-ticed by D. spectabilis.
Several authors have claimed that kangaroorats' caching behavior makes seeds unavail-able to ants and birds. Caching is unlikely,however, to be completely effective againstother rodents. Reichman and Oberstein (1977)found that captive D. merriami could detectand excavate 0.8-g seed packets buried at adepth of 20 cm, whereas caches of rolled oats,which we have uncovered in the field afterseeing kangaroo rats make them, are usuallyburied only about 2-3 cm deep, and the an-imals allocate up to 1 g to each cache. Boydand Brum (1983) claim that Mojave Desert D.merriami cache naturally collected Larrea tri-dentata seeds at depths of 2.5-5 cm.
In this paper, we report observations andfield experiments with Merriam's kangaroorats aimed at characterizing scatter-hoardingbehavior and detecting pilferage. Where arecaches situated relative to the food source,the home burrow, and one another? Is fooddifferentially disposed of according to whereit is discovered within the home range? Is there
significant pilferage? By whom? And does spa-tial dispersion of caches protect them fromdiscovery?
METHODSStudy site and habitat
This research was conducted at the Universityof California's Boyd Deep Canyon Desert Re-search Station, about 3 km south of Palm Des-ert, California, USA (33°42' N, 116°22' W).The study site is located on the alluvial plainof Deep Canyon at about 250 m elevation andis centered upon a 1-ha grid of 100 trap sta-tions in a 10 x 10 array at 10-m intervals.Each station is the site of a single Shermanlive trap (8 x 9 x 30 cm). Beyond the trappinggrid, markers (labeled stone cairns or stakes)extend in all directions at 10 m intervals, per-mitting researchers to quickly identify any lo-cus in the mapped area (about 11 ha) as a pairof X and Y coordinates to a precision of 1 m.
The substrate is a mixture of pebbly "desertpavement" and sandy washes up to 40 m wide,with rocky patches and a few boulders. Meanannual precipitation at the Boyd Center Lab-oratory (1.4 km from our study site, at 290 melevation) from 1961 to 1989 was 14.6 cm(SD = 10.0; range = 3.4-47.8 cm). The meandaily maximum temperature ranges fromabout 20°C in January to about 40°C in July,and the mean daily minimum is from about11°C to 27°C. Vegetation is denser and ofmore diverse species composition than in mostD. merriami habitats. Essentially similar to oursite is one nearby described in detail by Za-briskie (1979), at which 14% of the soil surfacelay under the canopy of perennial shrubs, ofwhich creosote bush {Larrea tridentata) was thecommonest species.
Radio tracking and general fieldworkpractices
We radio tracked kangaroo rats at the DeepCanyon field site annually from 1980 to 1990,during research periods of 1-7 months, be-ginning in November or December. At thebeginning of each research period, we trappedon 4 to 6 consecutive nights to assess survivalof marked animals, to mark those newly cap-tured, and to select animals for implantationwith radio transmitters emitting individuallyidentifiable signals. We concentrated radiotracking efforts on centrally located, neigh-boring resident animals, tracking up to 25 si-multaneously. For detailed accounts of radioimplantation and tracking procedures, seeBehrends et al. (1986) and Daly et al. (1990).We left the 100 grid traps in place throughouteach research period, but opened them only
Daly et al. • Kangaroo rat scatter hoarding 103
at McM
aster University L
ibrary on July 30, 2016http://beheco.oxfordjournals.org/
Dow
nloaded from

on certain trapping nights (usually once ortwice per week after the initial 4 to 6 trappingnights; see Daly et al., 1990). A trapping nightentailed baiting each of the 100 traps at duskwith about 1 g of rolled oats, checking forcaptures and closing the traps 2—5 h later.We recorded trap site, species, sex, individualidentity, weight, and reproductive conditionof each captured animal, then released it (un-less scheduled for radio removal or implan-tation) at the capture site. Rodents were dis-tinctively marked by toe clipping until 1988,and by passive integrated transponders (PITtags, Destron) after 1988.
Primary radio tracking data consist of dailylocations of each animal's day burrow andhourly locations for 6-14 h on most nights.In addition, we collected more than 1000 hof nocturnal focal-animal follows (durations15 min-6 h), during which we radio locateda target animal and followed on foot, avoidingsudden or rapid movements, while recordingwhereabouts as continuously as possible (typ-ically by speaking softly into a tape recorder),observing the subject under dim headlampillumination, and making behavioral obser-vations opportunistically. Although it must beconceded that individuals vary in observabilityand that it is difficult to determine to whatextent human presence influences the behav-ior of even a well-habituated animal, most kan-garoo rats seem unconcerned about humanobservers after a few hours of focal following.
lowed the provisioned kangaroo rat to seewhere the food was taken. For each round-trip excursion from the food dish, we record-ed whether the animal returned to its day bur-row, as well as the coordinates of successivelocations visited by the animal, whether or notcaching was observed. Caching behavior is dis-tinctive; the animal inserts its snout in a shal-low depression and rocks as it empties its cheekpouches. Kangaroo rats on our study siteplaced about 0.3-1.0 g of oats in each cache,then covered it with soil to a depth of 0.5-3.0 cm. (We have verified that this behaviorrepresents caching by temporarily exposingthe cache's contents after seeing kangaroo ratscache pouched oats upon release from traps,but only a few caches were verified in this wayduring the 48 provisioning trials because ofconcern that our interference might affect thecache's discovery by pilferers or its treatmentby the cacher; see "Pilferage observed di-rectly" in the Results section.) Observation ofcaching was limited, however, by our effortsto avoid startling the animals or approachingtoo closely and by the apparent secretivenessof cachers, who make rapid zig-zag runs, duckthrough shrubbery, and seem reluctant tocache while being watched. Thus, most cachesites could not be determined precisely.
Some further details of procedure changedover years, and these changes can be de-scribed with reference to three "series" oftrials.
General procedure of provisioning trials
To observe hoarding behavior, we conducted48 trials in which radio-implanted kangaroorats already habituated to observers were pro-vided with small food bonanzas in the field.These trials were conducted over the courseof 6 years with 26 subject animals provisioned1-3 times each. In each trial, we radio locatedthe target kangaroo rat, either in its day bur-row before emergence at dusk or during itsnocturnal travels, and placed a petri dish con-taining 10 g of rolled oats on the ground near-by. We then stood back and kept the dishunder watch with headlamp illumination, ver-ifying by radio signal that the target animalwas indeed the discoverer and collector of thefood, and shooing away any interlopers (otherkangaroo rats and pocket mice) until the tar-get animal had emptied the dish. Provisionedanimals usually collected the full 10 g, but onfive occasions we terminated trials when provi-sionees failed to return to the dish for a pro-longed period after taking 5.7-8.3 g.
In most trials, one observer kept watch atthe food dish and recorded the duration ofcheek-pouch loading episodes and of absencesbetween loadings, while a second observer fol-
Variable procedures of successive seriesof provisioning trials
Series 1We conducted 18 trials with 6 subjects in De-cember 1983. Five kangaroo rats (3 females,2 males) were randomly selected from among13 then being radio tracked, and each wasprovisioned on 3 successive nights. We pre-sented food on the first night at about 2000h (after an initial four or five hourly radiolocations) and on the second and third nightsimmediately after dusk. Provisioning sites weresituated 2, 2, 3, 10, and 17 m from the fiverecipients' initial day burrows. (One animalswitched day burrows daily over the three tri-als.) We recorded locations hourly from duskto midnight for all radio-implanted animalson provisioning nights, as well as on severalsuccessive nights before and after.
A sixth animal with exceptional spatial hab-its was also provisioned for 3 successive nights,after the others. This female's day burrow sitelay outside her nocturnal range, to which sheran directly upon emergence after sunset eachnight. During 91 scheduled hourly radio lo-cations between dusk and midnight and dur-ing focal follows totaling 270 min, she never
104 Behavioral Ecology Vol. 3 No. 2
at McM
aster University L
ibrary on July 30, 2016http://beheco.oxfordjournals.org/
Dow
nloaded from

returned to her day burrow or approachedwithin 24 m of it, although she was found tohave returned there each day. To see what ananimal who so assiduously avoided her dayburrow area during her nocturnal travelswould do upon finding food in the farthestcorner of her habitual range, we provisionedher at a site which she visited nightly 109 mfrom her day burrow on 3 successive nights.
Series 2To compare hoarding behavior when food wasdiscovered near home versus farther away, weran 20 trials with 11 kangaroo rats (6 females,5 males) in December 1985. Seven subjectsencountered food near the day burrow en-trance on one occasion (mean distance 1 m)and at another shrub 8-39 m away (mean 24m) on another occasion; repeat trials for thesame subject were separated by 3-9 nights(mean 5.9). For two other animals, efforts toprovision near the day burrow entrance failed,and they received two trials each at differentsites between 10 and 17m from the day bur-row. Two additional females were each pro-visioned only once, at distances of 1 and 2 mfrom their respective day burrows.
Series 3We conducted the final 10 trials with dyedfood to detect pilferage. Nine subjects wereused. We provisioned the first (female 1000)with red-dyed oats for 2 successive nights inDecember 1986; food was presented 1 h aftersunset at sites 13 and 12m from her day bur-row, and the caching trips and post-trial be-havior of this well-habituated animal wereclosely observed. We then trapped the 1-hagrid according to our standard routine (seeabove), with undyed rolled oats as bait, on thefirst, third, and fifth nights after provisioningand assessed fecal dye presence.
We conducted eight further dyed-food tri-als in the next two Decembers, with each an-imal provisioned only once. Subjects (5 fe-males, 3 males) were animals who had beenreliably captured on prior trapping nights.Distances between food sources and day bur-rows were 0, 3, 4, 9, 15, 18, 19, and 31 m.We separated provisioning trials in time andspace to facilitate correct attribution of traceamounts of dyes to their sources, conductingtwo red-dye trials and two green-dye trials eachyear. In each case, the grid was trapped onthe two nights immediately after provisioningand on several subsequent nights, and fecaldye was assessed in all captured rodents.
Y, Fisher) or green dye (Fast Green FCF, Fish-er) and spread them on paper towels until dry.Feeding experiments with captive animals inthe laboratory demonstrated that (1) feces col-lected 24 h after providing an animal withdyed oats strongly colored water into whichthe feces were dropped; (2) dye was conspic-uous in the feces of an animal provided withjust 1 g of the dyed food; (3) dye was scarcelydetectable in feces passed 48 h after dyed oatswere removed and replaced with ad libitumundyed oats; and (4) kangaroo rats accepteddyed oats as readily as undyed and when pro-vided with both ate them in proportion to theavailable quantities. (Some other dyes werefound to affect palatability in laboratory testsand were therefore not used in field trials.)
On each trapping night during provisioningseries 3 (see below), we collected two to eightfecal boluses from each captured rodent, inaddition to the usual data (species and indi-vidual identity, sex, weight, and conditionmeasures). We placed collected feces in glassvials labeled with the trap number and tookthe vials to the laboratory, where we added afew drops of tap water to each vial.
After 20-30 min, we examined vials for ev-idence of dye, which was subjectively judgedstrong, medium, faint, or absent by judgesblind to rodent identity. Fewer than 10% ofvials exhibited any such evidence, and thosethat did were usually unequivocal; some faintsamples were deemed ambiguous in regard tothe presence of red dye (see Results), but nev-er in regard to green dye. We determined thatthese judgments were reliable in two ways: (1)in several trials, two or three judges indepen-dently categorized the vials as "definite," "faintand ambiguous," or "none" with perfectagreement; and (2) split-sample trials, in whichthe feces of more than 100 captured animalswere split between two vials and judged blind,produced unanimously concordant judg-ments (i.e., no split sample ever produced onedye-present and one dye-absent judgment).Evidence for the validity of these judgmentsis that the samples in which dye was detectedconsistently proved to be those of the provi-sioned animal and its neighbors (see Results).
Thus, although the method may have al-lowed "misses" (failures to detect existing dyetraces), we conclude that it did not produce"false positives" and hence that the resultsprovide minimum estimates of the numbersof animals who consumed some of the oatsfrom a given provisioning trial.
Fecal dye assessment and detectionof pilferage
To prepare dyed food, we dipped oat flakesbriefly in a strong solution of red dye (E"c;ri
.osin
Seed survival as a function of spacingof artificial caches
An experiment to determine whether spacingamong caches affects risk of discovery was
Daly et al. • Kangaroo rat scatter hoarding 105
at McM
aster University L
ibrary on July 30, 2016http://beheco.oxfordjournals.org/
Dow
nloaded from

conducted at a site vegetationally and topo-graphically similar to our principal study areabut several hundred meters away. We filledplastic cups 3 cm in diameter and 4 cm deepwith aquarium gravel and buried known num-bers of millet seeds in the cups at a depth of1 cm. We then dug small depressions of thesame dimensions at various sites and placedeach cup snugly within such a depression, itslip flush with the ground. We returned 24 hlater to count the remaining millet seeds.
A total of 360 seed cups were thus situatedin the field, in 120 triplets arranged as equi-lateral triangles. In each triplet, the three cupscontained 4, 8, and 16 millet seeds, respec-tively. We situated the triplets without regardto the proximity of vegetation and recordedthe distance from each cup to the nearest pe-rennial shrub. Twenty triplets were estab-lished in each of six spacings: The sides of thetriangles were 0.25, 0.5, 1, 2, 4, and 8 m inlength. Cups that were not part of the sametriplet were situated at least 50 m apart.
RESULTS
Observations of caching behavior
Provisioned kangaroo rats typically emptiedthe food dish in less than 30 min from thetime of its discovery and occasionally in lessthan 10 min. This required 5-10 "loading"episodes in which the animal stuffed oats intoits cheek pouches at the dish, departed, de-posited the food elsewhere, and returned.
After loading at the food dish, kangaroorats did one of three things: (1) took the fooddirectly to the day burrow (all loads on 19 ofthe 48 trials, some but not all on an additional8); (2) took the food to another nearby burrowwithin 6 m of the food source (all loads on 5trials, some but not all on an additional 6); or(3) immediately scattered the food in caches(all loads on 11 trials, some but not all on anadditional 12).
At least one act of caching was seen in 22trials involving 18 kangaroo rats and 59 cachesites were discovered. Twenty-six of the dis-covered caches (44%) were situated in openterrain away from shrubs (mainly in sandywashes but also under the edges of stones ordead wood); 24 (41%) were placed under orat the edges of shrubs (usually small ones with-out evident burrows); and 9 (15%) were placedunder the drooping canopies of palo verde(Cercidiumjloridum) trees. Up to three caches,separated by as little as 60 cm or as much as40 m, were created from a single load beforereturning to the food dish.
Discovered cache sites were located at a me-dian distance of 8 m from the provisioningsite (range 0.5-49 m), but 9 different kanga-roo rats were observed making at least one
cache more than 20 m from the food source.These distances probably underestimate ac-tual food transport and cache dispersion be-cause we occasionally lost sight of those ani-mals who made longer trips and reestablishedvisual contact only as they returned to theprovisioning site; six different kangaroo ratsmade rapid round trips to points more than40 m from the food dish, but only two of thesix were seen caching on these distant trips.
The lone animal who was provisioned morethan 100 m from her day burrow in series 2produced the most spatially dispersed set ofcaches: although she made most of her cacheswithin 10 m of the food source on all threetrials, she carried some loads much farther,up to a maximum of 65 m, and in severaldirections. The resultant dispersion was suchthat her caches were up to 85 m from oneanother, and a few of her caching trips tookher 20 m or more beyond areas in which shewas ever otherwise radio-located. On no cach-ing trip did she approach within 40 m of herday burrow.
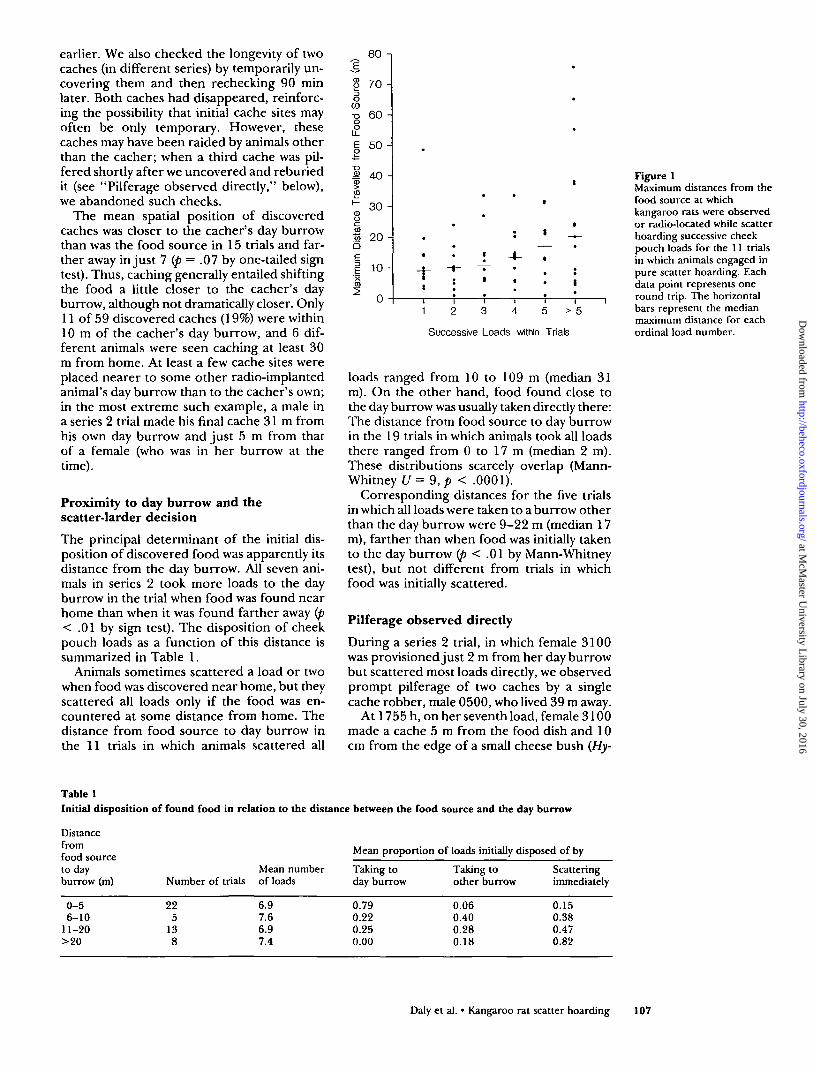
There was a tendency for the distance trav-eled with successive loads to increase (Figure1). An index of this tendency within a trial isthe rank order correlation between the max-imum transport distance for each load and itsordinal number. By this index, the trend wasin the direction of successively greater trans-port in 9 of the 11 trials in which subjectsengaged in pure scatter hoarding and in theopposite direction on 2 (p = .03 by one-tailedsign test); however, in only one trial was thetrend perfect in that every load (N = 5) wastaken farther than the preceding one.
In four trials in which food was initiallyhoarded to a burrow other than the day bur-row, the animal embarked on scatter hoardingimmediately after sequestering the food. Thefour different kangaroo rats involved were allobserved making caches, at distances rangingfrom 5 to 26 m away, within 6 min of havingdeposited their last loads in the burrow. Fouradditional animals who embarked on widetravels immediately after hoarding to a bur-row appeared to be behaving similarly, al-though they were not seen caching food. Eventhose who initially scatter hoarded were some-times observed making more distal caches alittle later. One female traveled no fartherthan 16 m from the food dish until it wasemptied, then embarked on a series of rapidround trips to several more distant points.Another female, who was tracked to points upto 24 m from the food source while initiallyscattering eight loads, was then seen makinga new cache 33 m away, 47 min after havingcompleted the trial.
In one case, a male was observed re-openingand emptying a cache he had made 52 min
106 Behavioral Ecology Vol. 3 No. 2
at McM
aster University L
ibrary on July 30, 2016http://beheco.oxfordjournals.org/
Dow
nloaded from
earlier. We also checked the longevity of twocaches (in different series) by temporarily un-covering them and then rechecking 90 minlater. Both caches had disappeared, reinforc-ing the possibility that initial cache sites mayoften be only temporary. However, thesecaches may have been raided by animals otherthan the cacher; when a third cache was pil-fered shortly after we uncovered and reburiedit (see "Pilferage observed directly," below),we abandoned such checks.
The mean spatial position of discoveredcaches was closer to the cacher's day burrowthan was the food source in 15 trials and far-ther away in just 7 (p = .07 by one-tailed signtest). Thus, caching generally entailed shiftingthe food a little closer to the cacher's dayburrow, although not dramatically closer. Only11 of 59 discovered caches (19%) were within10 m of the cacher's day burrow, and 6 dif-ferent animals were seen caching at least 30m from home. At least a few cache sites wereplaced nearer to some other radio-implantedanimal's day burrow than to the cacher's own;in the most extreme such example, a male ina series 2 trial made his final cache 31m fromhis own day burrow and just 5 m from thatof a female (who was in her burrow at thetime).
Proximity to day burrow and thescatter-larder decision
The principal determinant of the initial dis-position of discovered food was apparently itsdistance from the day burrow. All seven ani-mals in series 2 took more loads to the dayburrow in the trial when food was found nearhome than when it was found farther away (p< .01 by sign test). The disposition of cheekpouch loads as a function of this distance issummarized in Table 1.
Animals sometimes scattered a load or twowhen food was discovered near home, but theyscattered all loads only if the food was en-countered at some distance from home. Thedistance from food source to day burrow inthe 11 trials in which animals scattered all
1CDO
boCO13
8•I| ULJOJ)
• D
8lle
>
2h-
§Q£
"xCO
ou -
70 -
60 -
50 -
40 -
30 -
20 -
10 -
0 -
•
•
i-i
— i —
•
••
-t- —* 1 •• •
1 l 1
•
*
I
*
•- • -
•
•
:
• i
1 2 3 4 5 > 5
Successive Loads within Trials
loads ranged from 10 to 109 m (median 31m). On the other hand, food found close tothe day burrow was usually taken directly there:The distance from food source to day burrowin the 19 trials in which animals took all loadsthere ranged from 0 to 17 m (median 2 m).These distributions scarcely overlap (Mann-Whitney U = 9, p < .0001).
Corresponding distances for the five trialsin which all loads were taken to a burrow otherthan the day burrow were 9-22 m (median 17m), farther than when food was initially takento the day burrow (p < .01 by Mann-Whitneytest), but not different from trials in whichfood was initially scattered.
Pilferage observed directly
During a series 2 trial, in which female 3100was provisioned just 2 m from her day burrowbut scattered most loads directly, we observedprompt pilferage of two caches by a singlecache robber, male 0500, who lived 39 m away.
At 1755 h, on her seventh load, female 3100made a cache 5 m from the food dish and 10cm from the edge of a small cheese bush (Hy-
Figure 1Maximum distances from thefood source at whichkangaroo rats were observedor radio-located while scatterhoarding successive cheekpouch loads for the 11 trialsin which animals engaged inpure scatter hoarding. Eachdata point represents oneround trip. The horizontalbars represent the medianmaximum distance for eachordinal load number.
Table 1Initial disposition of found food in relation to the distance between the food source and the day burrow
Distancefromfood sourceto dayburrow (m)
Mean proportion of loads initially disposed of by
Number of trialsMean numberof loads
Taking today burrow
Taking toother burrow
Scatteringimmediately
0-56-10
11-20>20
225
138
6.97.66.97.4
0.790.220.250.00
0.060.400.280.18
0.150.380.470.82
Daly et al. • Kangaroo rat scatter hoarding 107
at McM
aster University L
ibrary on July 30, 2016http://beheco.oxfordjournals.org/
Dow
nloaded from

menoclea salsola); when she left, we opened thecache, removed its contents to weigh them(0.70 g), replaced them with about the samequantity of oats, and covered them again. At1759 h, on her eighth load, female 3100 madeanother cache under a log in a sandy wash 7m from the food source and l l m from theprevious cache; we did not disturb this cache.At 1820 h (while female 3100 was more than25 m away), male 0500 opened and emptiedthese two caches, which were 41 and 35 mfrom his own day burrow, in rapid succession,and recached their contents among some smallcheese bushes 27 and 16 m from the siteswhere female 3100 had cached them, 26 mfrom his own day burrow and 22 m from fe-male 3100's burrow.
Pilferage inferred from fecal dye traces
Sixteen to 47 rodents were trapped and as-sessed for fecal dye on trapping nights afterseries 3 trials. At least five of the nine animalsprovisioned with dyed oats were victimized bycache robbers.
For three subjects, the food dish was placedwithin 4 m of the day burrow, and none ofthese three incurred any detected pilferage.Females 2005 and 3020 took all food to theirday burrows within 19 min of discovery,whereas male 0220 quickly deposited five loadsin his day burrow before scattering two loads(caches were located 14 and 13 m away). Male0220's feces exhibited dye 1 and 2 days afterprovisioning, but not at 5,8, or 9 days. Female2005's feces exhibited dye 1 day and 8 daysafter provisioning, but not on days 2 and 5.Female 3020's feces exhibited dye strongly ondays 1, 4, 7, and 13, less intensely on days 2and 10, and not at all on days 8 and 11, im-plying that she fed on dyed stores 12 days afterprovisioning.
For the other six animals, the food dish wasplaced at distances of 9-31 m from the dayburrow, and five of these six animals incurreddetected pilferage. None of these six animalstook any food directly to their day burrows.Four animals (females 1000 and 4030; males2400 and 2003) scattered all loads, taking 20-55 min to complete the task. The other twoanimals (females 1300 and 3040) initially tookall loads down nearby holes other than theday burrow, but then set about scatteringcaches within 5 min of emptying the dish. Onlymale 2400, who exhibited dye on days 1 and2 but not on days 5 and 8, apparently escapedpilferage.
This difference (pilferage from 0 of 3 whoinitially larder hoarded to the day burrow ver-sus 5 of 6 who did not) is marginally significant(p = .05 by one-tailed Fisher's Exact test). Itis unclear, however, whether the relevant fac-
tor was the food dish's placement (near versusfar) or the hoarding behavior (day burrow ver-sus scatter) because the two were perfectlycorrelated.
Ten individual rodents were unambiguouslyidentified as pilferers, representing every noc-turnal rodent species captured on the studysite: three kangaroo rats (D. merriami; one fe-male, two males), three cactus mice (Peromys-cus eremicus; two females, one male), three spi-ney pocket mice (Chaetodipusfallax; all males),and one silky pocket mouse (Chaetodipus for-mosus; male). Six additional possible pilferers(fecal dye traces judged "faint and ambigu-ous") were a female D. merriami, a male P.eremicus, and four male C.fallax. Thus, a singleanimal's caches might be raided by several pil-ferers. On the first trapping night after female1000's two nights of provisioning, dye wasevident in the feces of 5 of the 39 rodentscaptured: female 1000 herself, two male kan-garoo rats, a pocket mouse, and a cactusmouse. Female 4030 may have been robbedby as many as six pilferers after scattering eightloads on her single provisioning, for on thefollowing night dye was evident in the fecesof one pocket mouse and was ambiguous inthree more, and on the second trapping nightdye was also evident in one cactus mouse andambiguous in one kangaroo rat, neither ofwhom had been trapped the previous night.Female 3040 was raided by two pocket mice;male 2003 was raided by two pocket mice andperhaps one cactus mouse; and female 1300was raided by a conspecific female.
Even the most heavily raided animals, how-ever, apparently were able to make as muchor more use of their caches as any pilferer. Inthree cases, dye faded or disappeared fromthe feces of retrapped pilferers while remain-ing evident in the provisioned animal's feces.(However, not every known or suspected pil-ferer was recaptured every night.) In a fourthcase, dye disappeared from pilferers' feces af-ter night 2, but the provisioned animal (female4030) and one pilferer were not retrappedfor purposes of comparison. In the fifth case,dye disappeared from the provisionee and thesole pilferer together. Most importantly, therewas not a single trapping night on which dyewas detected in a pilferer's feces while absentfrom the provisionee's, nor even a case inwhich the provisionee's fecal dye trace was ofa lesser intensity than that of any other animal.
Seed survival as a function of spacing ofartificial caches
After 24 h, 37% of the 3360 millet seeds in360 artificial caches had disappeared. Mostcups were undisturbed (N = 198; 55.0%); asingle seed was missing from another 22
108 Behavioral Ecology Vol. 3 No. 2
at McM
aster University L
ibrary on July 30, 2016http://beheco.oxfordjournals.org/
Dow
nloaded from

(6.1%). If more than one seed was gone (N =140), usually all were gone (N = 121) for allbut one (N= 13). Cups from which more thanone seed had disappeared were thus consid-ered "discovered."
A cup's risk of discovery was unrelated tothe number of seeds it contained: 47 4-seedcups (39%), 46 8-seed cups (38%), and 47 16-seed cups (39%) were discovered. Cups far-ther from perennial shrubs were more likelyto be discovered: 67/196 (34%) of those with-in 1 m of the nearest shrub; 54/134 (40%) if1-2 m away; and 19/30 (63%) if more than 2m away (p < .01 by Moses's ordered categoriestest; Moses, 1986: 416).
Seed removal appeared to be mainly thework of rodents. Gravel had been dug out ofalmost every cup from which seeds were miss-ing; 10 cups were pulled from the ground (butmoved no more than a few centimeters). Wesaw ants in a few emptied cups, but in eachsuch case the gravel had evidently been dugout by a rodent; only one cup appeared un-disturbed and yet was missing a seed.
Increased spacing within triplets reducedlosses. At 2 5-50 cm spacing, 51 % of cups werediscovered; 40% were discovered at 1-2 m,and 26% at 4-8 m {p < .01 by Moses's orderedcategories test). The probability that a tripletwould completely escape discovery was notsignificantly related to spacing (38% of tripletsat 25-50 cm; 45% at 1-2 m; 55% at 4-8 m;p > .10), but the probability that at least oneof the three cups would escape discovery wassignificantly related to spacing (63% of tripletsat 25-50 cm; 73% at 1-2 m; 90% at 4-8 m;p < .01). Given that a particular triplet wasdiscovered at all, increased spacing loweredthe risk that all would be lost (Figure 2).
DISCUSSION
The initial choice between scatter- and larder-hoarding tactics depended primarily on thedistance between the food source and home(Table 1). It is perhaps surprising that theswitch from taking food home to taking it else-where occurred as little as 8-10 m from theday burrow, a distance these animals can tra-verse in less than 2 s (Kenagy, 1973). If larderhoarding is only a stop-gap en route to scatter,however, it may be worthwhile only when itcan be achieved inconspicuously as well asquickly. Vander Wall (1990) notes the "secre-tiveness" of animals engaged in food caching,and kangaroo rats habituated to our presenceseemed to become more evasive when scatterhoarding. In the only acts of cache pilferagethat we observed directly, the robber raidedin less than 1 min two caches that had beenestablished successively in distinct microhab-itats l l m apart; a possible explanation is that
07 -,
O
.25 . 5 1 2 4
Distance between Artificial Caches within Triplet (rru
the pilferer had witnessed the caches beingmade.
Stapanian and Smith (1984) proposed thatin order to space caches adequately to thwartpilferers while minimizing expected losses atthe food source, a scatter hoarder should car-ry successive loads farther. Clarkson et al.(1986) used a model with more continuouscosts of increasing cache density to predictthat successive caches would be establishedfarther from the food source only on average,as animals would continue to situate some nearthe source while maintaining a gradient ofdecreasing density. Marsh tits (Parus palustris)behave in contradiction to both models bycarrying early items significantly farther thanlater ones (Sherry et al., 1982). Kangaroo ratbehavior is more consistent with these models,matching Clarkson et al.'s (1986) analysissomewhat better than Stapanian and Smith's(1984) (Figure 1). However, a few animals de-fied both models by making long trips andestablishing distant caches early in the cachingsequence.
The utility of scatter hoarding is compro-mised to the degree that a pilferer who dis-covers a cache becomes likelier to discoverothers. In particular, pilferers might improvetheir detection rates by adopting area-local-ized search tactics after a discovery (Tinber-gen et al., 1967), so caches should be spacedsufficiently to eliminate this tactic's utility. Ourfinding that increased spacing of artificialcaches protected the caches from discovery issimilar to that of other studies, although notall experiments have found such effects (re-view by Vander Wall, 1990). Reduced risk oflosing all caches within a triplet was conspic-uous at nearest-neighbor distances of 2 m ormore (Figure 2), and provisioned kangaroorats indeed typically distributed caches at least2 or 3 m from one another, although pairs ofcaches were twice created as close as 60 cm.
The finding that artificial caches were more
Figure 2Effect of the spacing ofartificial caches upon theconditional probability that allthree caches in an equilateraltriangular array would bediscovered within 24 h if anyone of them was discovered.
Daly et al. • Kangaroo rat scatter hoarding 109
at McM
aster University L
ibrary on July 30, 2016http://beheco.oxfordjournals.org/
Dow
nloaded from

vulnerable when situated farther from peren-nial shrubs is surprising in view of consider-able evidence that kangaroo rats escape com-petition from pocket mice in such open areas(Price and Brown, 1983). It remains to be seenwhether the same is true of natural caches andalso whether kangaroo rats show adaptivepreferences for relatively safe cache sites. Ob-served cache sites in the present study areunlikely to be representative: Differential vis-ibility probably biased us toward a better de-tection rate for caching in open terrain,whereas our presence may have biased be-havior in the opposite direction, as animalssought cover to cache in secrecy. Also sur-prising was the fact that cache discovery wasunaffected by cache size over the range of 4—16 millet seeds.
An incidental implication of the present re-sults is that the conclusions drawn from cer-tain studies using dyed foods to characterizespecies differences in foraging microhabitatsmay need revision. Price (1977) and Lemenand Rosenzweig (1978) scattered differential-ly marked seeds in different microhabitats andinferred foraging preferences from fecal res-idues; kangaroo rats had eaten mainly seedsfound in open spaces, while pocket mice hadapparently fed more widely. The possibility ofpilferage was not considered, however, and itis possible that pocket mice used a narrowerrange of foraging microhabitats than theseauthors inferred, never foraging far fromshrubs but raiding kangaroo rat stores oftenenough to create the impression of havingdone so.
The fecal dye results and the intensity ofpilferage imply that Merriam's kangaroo ratsare short-term hoarders (Vander Wall, 1990)who cache food for recovery within hours toa few days, rather than for use months later.When stores escaped pilferage, however, dyesdisappeared and reappeared in cachers' fecesup to 13 days after provisioning, and we neverchecked for even longer storage.
This study provides the first evidence ofwhich we are aware that scatter hoarders suf-fer significant pilferage. The short survival ofartificial caches could have turned out to beartifactual: caches created by the animalsthemselves might have proven to be secure.But such was not the case. Five of nine kan-garoo rats provisioned with dyed food lostcache contents to other rodents within 24 h.
The results suggest that scattering is morelikely to incur pilferage than larder hoarding,raising the question why the animals ever scat-ter hoard. A hypothesis for future study is thatscattering entails acceptance of lower expect-ed yield in exchange for reduced risk of majorloss. This would be analogous to the variance-reducing function of "risk-sensitive" foraging
tactics (Real and Caraco, 1986): If the ex-pected fitness value of increments in storedfood exhibits diminishing returns, then strat-egies that reduce the variance in expectedlosses to rivals can be favored over those thatsimply minimize the mean loss. The fact thatscatter hoarders who suffered pilferage nev-ertheless retained a large share of their storesis consistent with this possibility, as is the find-ing that increased spatial dispersion of arti-ficial cache triplets did not affect the risk ofpartial loss but lowered the risk of total loss.
Three animals who larder hoarded dyedfood to the day burrow incurred no detectedpilferage, and the day burrow is indeed anarea of relatively exclusive use as comparedto most of the home range (Behrends et al.,1986). However, other observations suggestthat day burrows are not completely secure.Kangaroo rats on our site typically plug theirburrows by day but leave them open by night,and outside the context of provisioning trials,we have seen both kangaroo rats and pocketmice enter the current day burrows of tem-porarily absent residents; we once saw a pock-et mouse emerge from such an intrusion withbulging cheekpouches. At present, we cannotconclude that larder hoarding by Merriam'skangaroo rats is anything other than a short-term, rapid-sequestering tactic employed be-fore scattering, since at least a few animalsembarked on scattering expeditions shortlyafter larder hoarding. Only further study candetermine whether larders ever persist,whether they really incur less pilferage on av-erage than scattered caches, and, if so, wheth-er scatter hoarding reduces the risk of majorloss.
Other questions about the adaptive logic ofkangaroo rat scatter hoarding raised by ourresults and demanding further research in-clude whether the interspecific flow of pil-fered resources is balanced or parasitic;whether pilferage is a distinct mode of be-havior from foraging for uncached seeds and,if so, whether some animals specialize as"scroungers" (Barnard, 1984); and why kan-garoo rats disperse their caches so widely, in-cluding even placing them beyond areas thatthe cacher routinely visits or passes near.
This research was supported by grants to M. Daly fromthe Natural Sciences and Engineering Research Councilof Canada. We are grateful for fellowship support fromthe North Atlantic Treaty Organization and the NationalScience Foundation to L. F.Jacobs, from thej. S. Gug-genheim Foundation to M. Daly, and from the Center forAdvanced Study in the Behavioral Sciences to M. Dalyand M. Wilson. Thanks to Mary Price for tutoring us indyed food techniques; to Don Kramer and two anony-mous referees for useful criticism of an earlier version ofthis article; and to Bill Mayhew, Al Muth, and Jan Za-briskie for countless kindnesses surrounding our use ofthe Boyd Deep Canyon Desert Research Center.
110 Behavioral Ecology Vol. 3 No. 2
at McM
aster University L
ibrary on July 30, 2016http://beheco.oxfordjournals.org/
Dow
nloaded from

REFERENCES
Balda RP, Bunch KG, Kamil AC, Sherry DF, TombackDF, 1987. Cache site memory in birds. In: Foragingbehavior (Kamil AC, Krebs JR, Pulliam HR, eds). NewYork: Plenum; 645-666.
Barnard CJ (ed), 1984. Producers and scroungers: strat-egies of exploitation and parasitism. London: CroomHelm.
Behrends P, Daly M, Wilson MI, 1986. Range use patternsand spatial relationships of Merriam's kangaroo rats(Dipodomys merriami). Behaviour 96:187-209.
Boyd RS, Brum CD, 1983. Predispersal reproductive at-trition in a Mojave Desert population of Larrea triden-tata (Zygophyllaceae). Am Midi Nat 110:14-24.
Clarkson K, Eden SF, Sutherland WJ, Houston AI, 1986.Density dependence and magpie food hoarding. J AnimEcol 55:111-121.
Covich AP, 1987. Optimal use of space by neighboringcentral place foragers: when and where to store surplusresources. In: Advances in behavioral economics, vol 1(Green L, KagelJH, eds). Norwood, New Jersey: Ablex;249-294.
Daly M, Wilson M, Behrends PR, Jacobs LF, 1990. Char-acteristics of kangaroo rats, Dipodomys merriami, asso-ciated with differential predation. Anim Behav 40:380-389.
Hurly TA, Robertson RJ, 1990. Variation in the foodhoarding behaviour of red squirrels. Behav Ecol So-ciobiol 26:91-97.
Jacobs LF, 1991. Memory for cache locations in Mer-riam's kangaroo rats. Anim Behav, in press.
Jones WT, 1984. Survivorship in philopatric and dis-persing kangaroo rats (Dipodomys spectabilis). Ecology67:202-207.
Kenagy GJ, 1973. Daily and seasonal patterns of activityin a heteromyid rodent community. Ecology 54:1201-1219.
Kramer DL, Nowell W, 1980. Central place foraging inthe eastern chipmunk, Tamias striatus. Anim Behav 28:772-778.
LemenCA, RosenzweigML, 1978. Microhabitat selectionin two species of heteromyid rodents. Oecologia 33:127-136.
Monson G, Kessler W, 1940. Life history notes on thebanner-tailed kangaroo rat, Merriam's kangaroo rat,and the white-throated wood rat in Arizona and NewMexico. J Wildl Manage 4:37-43.
Morris D, 1962. The behaviour of the green acouchi(Myoprocta pratti) with special reference to scatterhoarding. Proc Zool Soc Lond 139:701-732.
Moses LE, 1986. Think and explain with statistics. Read-ing, Massachusetts: Addison-Wiley.
Orians GH, Pearson NE, 1979. On the theory of centralplace foraging. In: Analysis of ecological systems (HornDJ, Stairs GR, Mitchell RD, eds). Columbus: Ohio StateUniversity Press; 155-177.
Price MV, 1977. Validity of live trapping as a measure offoraging activity of heteromyid rodents. J Mammal 58:107-110.
Price MV, Brown JH, 1983. Patterns of morphology andresource use in North American desert communities.Great Basin Nat Mem 7:117-134.
Randall JA, 1984. Territorial defense and advertisementby footdrumming in bannertail kangaroo rats (Dipodo-mys spectabilis) at high and low population densities.Behav Ecol Sociobiol 16:11-20.
Real L, Caraco T, 1986. Risk and foraging in stochasticenvironments. Annu Rev Ecol Syst 17:371-390.
Reichman OJ, Oberstein D, 1977. Selection of seed dis-tribution types by Dipodomys merriami and Perognathusamplus. Ecology 58:636-643.
Reynolds HG, 1958. The ecology of the Merriam kan-garoo rat (Dipodomys merriami Mearns) on the grazinglands of southern Arizona. Ecol Monogr 28:111-127.
Schroder GD, 1979. Foraging behavior and home rangeutilization of the bannertail kangaroo rat (Dipodomysspectabilis). Ecology 60:657-665.
Sherry DF, 1985. Food storage by birds and mammals.Adv Study Behav 15:1-68.
Sherry DF, Avery MA, Stevens A, 1982. The spacing ofstored food by marsh tits. Z Tierpsychol 58:153-162.
Smith CC, Reichman OJ, 1984. The evolution of foodcaching by birds and mammals. Annu Rev Ecol Syst 15:329-351.
Stapanian MA, Smith CC, 1984. Density-dependent sur-vival of scatterhoarded nuts: an experimental ap-proach. Ecology 65:1387-1396.
Tinbergen N, Impekoven M, Franck D, 1967. An exper-iment on spacing-out as a defence against predation.Behaviour 28:307-321.
Vander Wall SB, 1990. Food hoarding in animals. Chi-cago: University of Chicago Press.
Vorhies CT, Taylor WP, 1922. Life history of the kan-garoo rat, Dipodomys spectabilis spectabilis Merriam. USDep Agric Bull 1091:1-40.
Zabriskie JG, 1979. Plants of Deep Canyon. Riverside,California: University of California Press.
Daly et al. • Kangaroo rat scatter hoarding 111
at McM
aster University L
ibrary on July 30, 2016http://beheco.oxfordjournals.org/
Dow
nloaded from