section 12.2 - probes for mitochondria - mobitec innovative … · 2008-07-09 · 473 12.2 probes...
TRANSCRIPT

473
12.2 Probes for MitochondriaMitochondria are found in eukaryotic cells, where they make
up as much as 10% of the cell volume. They are pleomorphicorganelles with structural variations depending on cell type, cell-cycle stage and intracellular metabolic state. The key function ofmitochondria is energy production through oxidative phosphoryla-tion (OxPhos) and lipid oxidation.1 Several other metabolic func-tions are performed by mitochondria, including urea productionand heme, non-heme iron and steroid biogenesis, as well as intra-cellular Ca2+ homeostasis. Mitochondria also play a pivotal rolein apoptosis — a process by which unneeded cells are removedduring development, and defective cells are selectively destroyedwithout surrounding organelle damage in somatic tissues 2–4 (Sec-tion 15.5). For many of these mitochondrial functions, there isonly a partial understanding of the components involved, witheven less information on mechanism and regulation.
Visualizing Mitochondria in Cells and Tissues
The morphology of mitochondria is highly variable. In divid-ing cells the organelle can switch between a fragmented morphol-ogy with many ovoid-shaped mitochondria, as often shown inmost textbooks, and a reticulum in which the organelle is a single,many-branched structure. The cell cycle– and metabolic state–dependent changes in mitochondrial morphology are controlledby a set of proteins that cause fission and fusion of the organellemass. Mutations in these proteins are the cause of several humandiseases, indicating the importance of overall morphology for cellfunctioning (see Mitochondria in Diseases). Organelle morpholo-gy is also controlled by cytoskeletal elements, including actinfilaments and microtubules. In nondividing tissue, overall mito-chondrial morphology is very cell dependent, ranging from spi-raling around the acrosome in spermatozoa, to intercalated bandsof ovoid mitochondria between the actomyosin filaments. Thereis emerging evidence of functionally significant heterogeneity ofmitochondrial forms within individual cells.
The abundance of mitochondria varies with cellular energylevel and is a function of cell type, cell-cycle stage and prolifera-tive state. For example, brown adipose tissue cells,5 hepatocytes 6
and certain renal epithelial cells 7 tend to be rich in active mito-chondria, whereas quiescent immune-system progenitor or pre-cursor cells show little staining with mitochondrion-selectivedyes.8 The number of mitochondria is reduced in Alzheimer’sdisease and their protein and nucleic acids are affected by reactiveoxygen species, including nitric oxide 9 (Chapter 19).
Molecular Probes has a range of mitochondrion-selective dyeswith which to monitor mitochondrial morphology and organellefunctioning. The uptake of most mitochondrion-selective dyes isdependent on the mitochondrial membrane potential; nonyl acri-dine orange and possibly our MitoTracker Green FM, MitoFluorGreen and MitoFluor Red 589 probes are notable exceptions,although their membrane potential–independent uptake andfluorescence has been questioned in some cell types.10,11 Mito-chondrion-selective reagents enable researchers to probe mito-chondrial activity, localization and abundance,12,13 as well as tomonitor the effects of some pharmacological agents, such asanesthetics that alter mitochondrial function.14 Molecular Probes
offers a variety of cell-permeant stains for mitochondria, as wellas subunit-specific monoclonal antibodies directed against pro-teins in the oxidative phosphorylation (OxPhos) system, all ofwhich are discussed below.
MitoTracker Probes: Fixable Mitochondrion-Selective Probes
Although conventional fluorescent stains for mitochondria,such as rhodamine 123 and tetramethylrosamine, are readilysequestered by functioning mitochondria, they are subsequentlywashed out of the cells once the mitochondrion’s membranepotential is lost. This characteristic limits their use in experimentsin which cells must be treated with aldehyde-based fixatives orother agents that affect the energetic state of the mitochondria.To overcome this limitation, Molecular Probes has developedMitoTracker probes 15 — a series of patented mitochondrion-selective stains that are concentrated by active mitochondria andwell retained during cell fixation.16 Because the MitoTrackerOrange, MitoTracker Red and MitoTracker Deep Red probes arealso retained following permeabilization, the sample retains thefluorescent staining pattern characteristic of live cells duringsubsequent processing steps for immunocytochemistry, in situhybridization or electron microscopy. In addition, MitoTrackerreagents eliminate some of the difficulties of working with patho-genic cells because, once the mitochondria are stained, the cellscan be treated with fixatives before the sample is analyzed.
Properties of MitoTracker ProbesMitoTracker probes are cell-permeant mitochondrion-selective
dyes that contain a mildly thiol-reactive chloromethyl moiety.The chloromethyl group appears to be responsible for keepingthe dye associated with the mitochondria after fixation. To labelmitochondria, cells are simply incubated in submicromolar con-centrations of the MitoTracker probe, which passively diffusesacross the plasma membrane and accumulates in active mitochon-dria. Once their mitochondria are labeled, the cells can be treatedwith aldehyde-based fixatives to allow further processing of thesample; with the exception of MitoTracker Green FM, subsequentpermeabilization with cold acetone does not appear to disturb thestaining pattern of the MitoTracker dyes.
Molecular Probes offers seven MitoTracker reagents that differin spectral characteristics, oxidation state and fixability (Table12.1). MitoTracker probes are provided in specially packaged setsof 20 vials, each containing 50 µg for reconstitution as required.
Orange-, Red- and Infrared-Fluorescent MitoTracker DyesWe offer MitoTracker derivatives of the orange-fluorescent
tetramethylrosamine (MitoTracker Orange CMTMRos, M-7510;Figure 12.3) and the red-fluorescent X-rosamine (MitoTrackerRed CMXRos, M-7512; Figure 12.4), as well as our newestderivatives, the MitoTracker Red 580 and MitoTracker Deep Red633 probes (M-22425, M-22426; Figure 12.5, Figure 12.6; seeWidefield Deconvolution Microscopy in Section 12.1). Becausethe MitoTracker Red CMXRos, MitoTracker Red 580 and Mito-
Section 12.2

474 Chapter 12 — Probes for Organelles www.probes.com
Tracker Deep Red 633 probes produce longer-wavelength fluorescence that is well re-solved from the fluorescence of green-fluorescent dyes, they are suitable for multicolorlabeling experiments (Figure 1.41, Figure 8.7, Figure 12.7, Figure 12.8, Figure 12.9).Also available are chemically reduced forms of the tetramethylrosamine (MitoTrackerOrange CM-H2TMRos, M-7511; Figure 12.10) and X-rosamine (MitoTracker RedCM-H2XRos, M-7513; Figure 12.11) MitoTracker probes. Unlike MitoTracker OrangeCMTMRos and MitoTracker Red CMXRos, the reduced versions of these probes do notfluoresce until they enter an actively respiring cell, where they are oxidized to the fluo-rescent mitochondrion-selective probe and then sequestered in the mitochondria (Figure12.12, Figure 15.13). The MitoTracker probes have proven useful for:
• Assaying the role of a kinesin-like protein on germ plasm aggregation in Xenopusoocytes 17
• Detecting early apoptosis (Section 15.5), which is marked by a disruption of mito-chondrial transmembrane potential in all cell types studied 18–20
• Determining the mechanism by which mitochondrial shape is established and main-tained in yeast 21
• Examining the time course of cell swelling in a human collecting-duct cell line usingtotal internal reflection (TIR) microfluorimetry 22
• Localizing a novel kinesin motor protein involved in transport of mitochondria alongmicrotubules 23
• Simultaneously observing fluorescent signals from a green-fluorescent protein (GFP)chimera and from the MitoTracker dye 24–27
• Studying the localization of mitochondria in fibroblasts transformed with cDNA ofwild-type and mutant kinesin heavy chains 28
• Visualizing mitochondria while characterizing the subcellular distribution of calciumchannel subtypes in Aplysia californica bag cell neurons 29 and of the verotoxin Bsubunit in Vero cells 30
MitoTracker Orange CMTMRos and its reduced form CM-H2TMRos have also beenused to investigate the metabolic state of Pneumocystis carinii mitochondria.31 Followingfixation, the oxidized forms of the tetramethylrosamine and X-rosamine MitoTrackerdyes can be detected directly by fluorescence or indirectly with either anti-tetramethyl-rhodamine or anti–Texas Red dye antibodies (A-6397, A-6399; Section 7.4).
MitoTracker Green FM ProbeMitochondria in cells stained with nanomolar concentrations of our patented Mito-
Tracker Green FM dye (M-7514) exhibit bright-green, fluorescein-like fluorescence (Figure12.13, Figure 12.34, Figure 14.62, Figure 16.20). The MitoTracker Green FM probe has theadded advantage that it is essentially nonfluorescent in aqueous solutions and only becomesfluorescent once it accumulates in the lipid environment of mitochondria. Hence, back-ground fluorescence is negligible, enabling researchers to clearly visualize mitochondria inlive cells immediately following addition of the stain, without a wash step.
Figure 12.3 M-7510 MitoTracker OrangeCMTMRos.
Figure 12.4 M-7512 MitoTracker Red CMXRos.
Figure 12.6 Mitochondria of live bovine pulmonaryartery endothelial cells stained with the MitoTrack-er Deep Red 633 dye (M-22426).
Figure 12.5 Bovine pulmonary artery endothelialcells labeled with MitoTracker Red 580 (M-22425)to stain mitochondria. Following fixation and per-meabilization, the Golgi apparatus was labeled withour anti–golgin-97 antibody (A-21270) and detect-ed using the Alexa Fluor 488 goat anti–mouse IgGantibody (A-11001). The cells were counterstainedwith DAPI (D-1306, D-3571, D-21490). The imagewas deconvolved using Huygens software (Scien-tific Volume Imaging, www.svi.nl). 3-D reconstruc-tion was performed using Imaris software (Bit-plane AG).
Table 12.1 Spectral characteristics of the MitoTracker probes.
Cat # MitoTracker Probe Abs * (nm) Em * (nm) Oxidation State
M-7514 MitoTracker Green FM † 490 516 NA
M-7510 MitoTracker Orange CMTMRos 551 576 Oxidized
M-7511 MitoTracker Orange CM-H2TMRos 551 ‡ 576 ‡ Reduced
M-7512 MitoTracker Red CMXRos 578 599 Oxidized
M-7513 MitoTracker Red CM-H2XRos 578 ‡ 599 ‡ Reduced
M-22425 MitoTracker Red 580 581 644 NA
M-22426 MitoTracker Deep Red 633 644 665 NA
* Absorption (Abs) and fluorescence emission (Em) maxima, determined in methanol; values may varysomewhat in cellular environments. † MitoTracker Green FM is nonfluorescent in aqueous environments.‡ These reduced MitoTracker probes are not fluorescent until oxidized. NA = Not applicable.
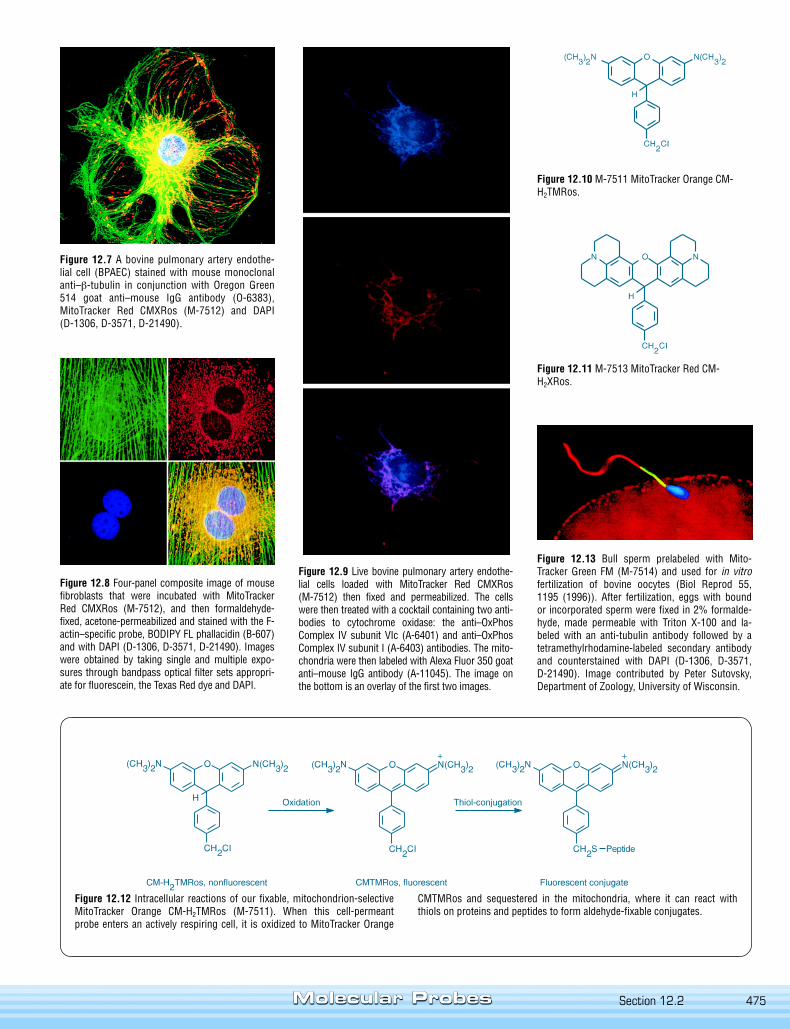
475
Figure 12.7 A bovine pulmonary artery endothe-lial cell (BPAEC) stained with mouse monoclonalanti–β-tubulin in conjunction with Oregon Green514 goat anti–mouse IgG antibody (O-6383),MitoTracker Red CMXRos (M-7512) and DAPI(D-1306, D-3571, D-21490).
Figure 12.8 Four-panel composite image of mousefibroblasts that were incubated with MitoTrackerRed CMXRos (M-7512), and then formaldehyde-fixed, acetone-permeabilized and stained with the F-actin–specific probe, BODIPY FL phallacidin (B-607)and with DAPI (D-1306, D-3571, D-21490). Imageswere obtained by taking single and multiple expo-sures through bandpass optical filter sets appropri-ate for fluorescein, the Texas Red dye and DAPI.
Figure 12.10 M-7511 MitoTracker Orange CM-H2TMRos.
Figure 12.11 M-7513 MitoTracker Red CM-H2XRos.
Figure 12.9 Live bovine pulmonary artery endothe-lial cells loaded with MitoTracker Red CMXRos(M-7512) then fixed and permeabilized. The cellswere then treated with a cocktail containing two anti-bodies to cytochrome oxidase: the anti–OxPhosComplex IV subunit VIc (A-6401) and anti–OxPhosComplex IV subunit I (A-6403) antibodies. The mito-chondria were then labeled with Alexa Fluor 350 goatanti–mouse IgG antibody (A-11045). The image onthe bottom is an overlay of the first two images.
Figure 12.12 Intracellular reactions of our fixable, mitochondrion-selectiveMitoTracker Orange CM-H2TMRos (M-7511). When this cell-permeantprobe enters an actively respiring cell, it is oxidized to MitoTracker Orange
Figure 12.13 Bull sperm prelabeled with Mito-Tracker Green FM (M-7514) and used for in vitrofertilization of bovine oocytes (Biol Reprod 55,1195 (1996)). After fertilization, eggs with boundor incorporated sperm were fixed in 2% formalde-hyde, made permeable with Triton X-100 and la-beled with an anti-tubulin antibody followed by atetramethylrhodamine-labeled secondary antibodyand counterstained with DAPI (D-1306, D-3571,D-21490). Image contributed by Peter Sutovsky,Department of Zoology, University of Wisconsin.
CMTMRos and sequestered in the mitochondria, where it can react withthiols on proteins and peptides to form aldehyde-fixable conjugates.
Section 12.2

476 Chapter 12 — Probes for Organelles www.probes.com
Unlike MitoTracker Orange CMTMRos and MitoTracker Red CMXRos, the Mito-Tracker Green FM probe appears to preferentially accumulate in mitochondria regardlessof mitochondrial membrane potential in certain cell types, making it a possible tool fordetermining mitochondrial mass 32,33 (see Estimating Mitochondrial Mass). Furthermore,the MitoTracker Green FM dye is substantially more photostable than the widely usedrhodamine 123 fluorescent dye and produces a brighter, more mitochondrion-selectivesignal at lower concentrations. Because its emission maximum is blue-shifted approxi-mately 10 nm relative to the emission maximum of rhodamine 123, the MitoTrackerGreen FM dye produces a fluorescent staining pattern that should be better resolved fromthat of red-fluorescent probes in double-labeling experiments. The MitoTracker GreenFM probe has been used to:
• Assay the differentiation state of Trypanosoma brucei bloodstream forms 34
• Demonstrate mitochondrion-selective labeling by avidin, streptavidin and anti-biotinantibodies 35
• Identify mitochondria in immunolocalization experiments in CHO cells 36
• Label sperm in order to determine the fate of sperm mitochondria during fertilizationand subsequent embryo development 37–39 (Figure 12.13, Figure 12.14)
• Monitor mitochondrial distribution and transport in Tau-expressing CHO cells 40
• Study the regulation of calcium signaling by mitochondria in T lymphocytes 41
MitoFluor Probes: Nonfixable Mitochondrion-Selective Probes
MitoFluor Green ProbeAs a companion to the MitoTracker Green FM derivative, we have developed the
MitoFluor Green probe 10 (M-7502), which has a structure similar to MitoTracker GreenFM (Figure 12.15) but lacks its reactive chloromethyl moieties (Figure 12.16) and is notas well retained following fixation. As with MitoTracker Green FM, the MitoFluor Greenprobe can selectively stain mitochondria in both live and fixed cells.10,42 The MitoFluorGreen probe is also substantially more photostable than rhodamine 123 (Figure 12.17);produces a brighter, more mitochondrion-selective signal at lower concentrations; andexhibits a blue-shifted emission maximum relative to that of rhodamine 123 that is betterresolved from that of red-fluorescent probes in double-labeling experiments. Like Mito-Tracker Green FM, the MitoFluor Green probe does not appear to be retained after cellpermeabilization.
MitoFluor Red 589 ProbeThe MitoFluor Red 589 probe (M-22424) appears to accumulate in mitochondria
regardless of the mitochondria’s membrane potential, making it a useful stain for mito-chondria in both live (Figure 12.18) and fixed cells. This probe has absorption and emis-sion peaks at 588 nm and 622 nm, respectively, and can be viewed with filter sets appro-priate for the Texas Red dye. Like our other MitoFluor Red dyes, the MitoFluor Red 589probe provides a clear spectral window below 600 nm for dual labeling with green-fluorescent probes.
MitoFluor Red 594 and MitoFluor Far Red 680 ProbesWe offer two mitochondrial membrane potential–sensing dyes that have long-wave-
length fluorescence emission: the MitoFluor Red 594 (M-22422, Figure 12.19) andMitoFluor Far Red 680 (M-22423, Figure 12.20) probes. The MitoFluor Red 594 dyeis a mitochondrial potential sensor that has been designed for optimal excitation by the594 nm spectral line of the He–Ne laser. This long-wavelength probe can be combinedwith green-fluorescent labels, including other site-selective probes or GFP chimeras.The MitoFluor Far Red 680 dye, also known as rhodamine 800, is a mitochondrialpotential sensor with absorption and fluorescence emission in the near-infrared spectralregion. Accumulation of the MitoFluor Far Red 680 dye by active mitochondria pro-duces a slight red shift in the probe’s absorption and fluorescence emission peaks,accompanied by a marked decrease in fluorescence intensity.43 Although the dye’sresponse is not directly visible to the human eye and must be captured using an infra-red light–sensitive detector such as a CCD camera, this long-wavelength probe is ad-
Figure 12.15 M-7514 MitoTracker Green FM.
Figure 12.16 M-7502 MitoFluor Green.
Figure 12.14 Live bull sperm stained simulta-neously with MitoTracker Green FM (M-7514) andHoechst 33342 (H-1399, H-3570, H-21492), as re-ported in Mol Reprod Dev 47, 79 (1997). Imagecontributed by Peter Sutovsky, Oregon RegionalPrimate Resource Center, Oregon Health SciencesUniversity. Used with permission of Wiley-Liss,Inc., a subsidiary of John Wiley & Sons, Inc.

477
Estimating Mitochondrial MassAccurate measurements of mitchondrial mass require a probe that will accumulate
in mitochondria regardless of the mitochondrial membrane potential, a property dis-played by several of our MitoTracker and MitoFluor dyes — MitoFluor Green (M-7502),MitoFluor Red 589 (M-22424), MitoTracker Red 580 (M-22425) and MitoTracker DeepRed 633 (M-22426). Mitochondrial uptake of nonyl acridine orange (NAO, A-1372) andMitoTracker Green FM (M-7514) has also been reported to be independent of mitochon-drial membrane potential,1,2 although studies have shown that this may not true for all
cell types.3,4 Mitochondrial staining bynonyl acridine orange is attributed tobinding of cardiolipin in the innermitochondrial membrane,5 suggestingthat the observed fluorescence signalmay not correlate with the “mass” ofthe entire mitochondria but instead maymeasure the amount of inner mem-brane present. Indeed, one study hasshown that mitochondrial mass canincrease while the mitochondrial vol-ume remains unchanged.6
Mitochondrial mass varies with celltype and can be useful is separatingpopulations of healthy cells (see Fig-ure). The mitochondrial mass a celldoes change with time, as demonstrat-ed by the loss of mitochondrial mass instudies of aging cells.7 Changes inmitochondrial mass are also observedin apoptotic cells, but are not necessar-ily associated with the loss of mito-chondrial membrane potential.6,8
References
1. Histochemistry 82, 51 (1985); 2. Immunol Lett 61, 157 (1998); 3. Basic Appl Histochem 33, 71(1989); 4. Cytometry 39, 203 (2000); 5. Eur J Biochem 209, 267 (1992); 6. Proc Natl Acad Sci U S A98, 9505 (2001); 7. Mech Ageing Dev 77, 83 (1994); 8. Exp Cell Res 221, 281 (1995).
TECHNICAL NOTE
Figure 12.17 Photostability comparison of mitochondrial staining by MitoFluor Green (upper series,M-7502) and rhodamine 123 (lower series; R-302, R-22420). HeLa cells were stained with 100 nMMitoFluor Green or 500 nM rhodamine 123 in growth medium for 20 minutes at 37°C. Cells were thenrinsed in Hanks’ Balanced Salt Solution (HBSS) with 10% calf serum. Samples were continuously illumi-nated and viewed on a fluorescence microscope using an Omega Optical fluorescein longpass optical fil-ter set and Image-1 software (Universal Imaging Corp.). Images were acquired 0, 10, 30 and 60 secondsafter the start of illumination (as indicated in the top left-hand corner of each panel) and clearly demon-strate the superior photostability of MitoFluor Green.
Figure 12.19 Viable bovine pulmonary artery en-dothelial cells incubated simultaneously with thered-fluorescent mitochondrial stain MitoFluor Red594 (M-22422) and the blue-fluorescent nuclearstain Hoechst 33342 (H-1399, H-3570, H-21492).The image is a composite of two micrographs, us-ing appropriate filter sets.
Figure 12.18 Mitochondria of live bovine pulmo-nary artery endothelial cells stained with MitoFluorRed 589 (M-22424).
Figure 12.20 Live bovine pulmonary artery endo-thelial cells were incubated simultaneously withMitoFluor Far Red 680 (M-22423), LysoTrackerGreen DND-26 (L-7526) and Hoechst 33342(H-1399, H-3570, H-21492). MitoFluor Far Red 680accumulated in mitochondria, and its fluorescence(emission maximum ~700 nm) was pseudocoloredred. Green-fluorescent LysoTracker Green dye accu-mulated in the lysosomes, and blue-fluorescent Ho-echst 33342 dye stained the nuclei.
Peripheral blood mononuclear cells (PBMCs)stained with an R-phycoerythrin–labeled anti-CD3antibody (A-21333) and the MitoTracker Greendye (M-7514). For comparison, CD3+ PBMCswithout MitoTracker Green staining are shown onthe same plot (lower right population). This figuredemonstates that the mitochondrial mass ofPBMCs is approximately the same for CD3+ andCD3- individuals.
Section 12.2
478 Chapter 12 — Probes for Organelles www.probes.com
vantageous when working with tissue, blood and other biologi-cal fluids that give rise to high absorbance or autofluorescenceat shorter wavelengths.44
RedoxSensor Red CC-1 Stain
RedoxSensor Red CC-1 (R-14060) stain is a unique probewhose fluorescence localization appears to be based on a cell’scytosolic redox potential. Scientists at Molecular Probes havefound that RedoxSensor Red CC-1 stain passively enters livecells. Once inside, this nonfluorescent probe is either oxidizedin the cytosol to a red-fluorescent product (excitation/emissionmaxima ~540/600 nm), which then accumulates in the mitochon-dria, or is transported to the lysosomes, where it is oxidized.The differential distribution of the oxidized product betweenmitochondria and lysosomes appears to depend on the redoxpotential of the cytosol.45 In proliferating cells, staining of mito-chondria predominates; whereas in contact-inhibited cells thestaining is primarily lysosomal. The best method we have foundto quantitate the distribution of the oxidized product is to use themitochondrion-selective MitoTracker Green FM stain (M-7514)in conjunction with the RedoxSensor Red CC-1 stain.
JC-1 and JC-9: Dual-EmissionPotential-Sensitive Probes
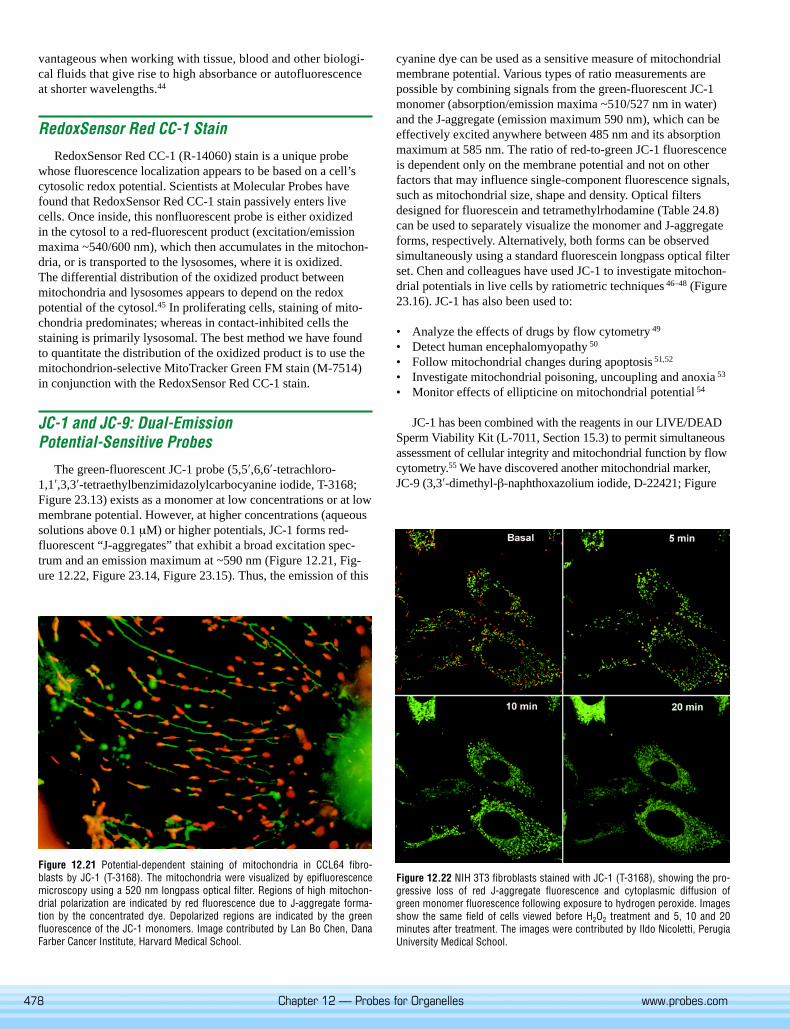
The green-fluorescent JC-1 probe (5,5′,6,6′-tetrachloro-1,1′,3,3′-tetraethylbenzimidazolylcarbocyanine iodide, T-3168;Figure 23.13) exists as a monomer at low concentrations or at lowmembrane potential. However, at higher concentrations (aqueoussolutions above 0.1 µM) or higher potentials, JC-1 forms red-fluorescent “J-aggregates” that exhibit a broad excitation spec-trum and an emission maximum at ~590 nm (Figure 12.21, Fig-ure 12.22, Figure 23.14, Figure 23.15). Thus, the emission of this
Figure 12.22 NIH 3T3 fibroblasts stained with JC-1 (T-3168), showing the pro-gressive loss of red J-aggregate fluorescence and cytoplasmic diffusion ofgreen monomer fluorescence following exposure to hydrogen peroxide. Imagesshow the same field of cells viewed before H2O2 treatment and 5, 10 and 20minutes after treatment. The images were contributed by Ildo Nicoletti, PerugiaUniversity Medical School.
Figure 12.21 Potential-dependent staining of mitochondria in CCL64 fibro-blasts by JC-1 (T-3168). The mitochondria were visualized by epifluorescencemicroscopy using a 520 nm longpass optical filter. Regions of high mitochon-drial polarization are indicated by red fluorescence due to J-aggregate forma-tion by the concentrated dye. Depolarized regions are indicated by the greenfluorescence of the JC-1 monomers. Image contributed by Lan Bo Chen, DanaFarber Cancer Institute, Harvard Medical School.
cyanine dye can be used as a sensitive measure of mitochondrialmembrane potential. Various types of ratio measurements arepossible by combining signals from the green-fluorescent JC-1monomer (absorption/emission maxima ~510/527 nm in water)and the J-aggregate (emission maximum 590 nm), which can beeffectively excited anywhere between 485 nm and its absorptionmaximum at 585 nm. The ratio of red-to-green JC-1 fluorescenceis dependent only on the membrane potential and not on otherfactors that may influence single-component fluorescence signals,such as mitochondrial size, shape and density. Optical filtersdesigned for fluorescein and tetramethylrhodamine (Table 24.8)can be used to separately visualize the monomer and J-aggregateforms, respectively. Alternatively, both forms can be observedsimultaneously using a standard fluorescein longpass optical filterset. Chen and colleagues have used JC-1 to investigate mitochon-drial potentials in live cells by ratiometric techniques 46–48 (Figure23.16). JC-1 has also been used to:
• Analyze the effects of drugs by flow cytometry 49
• Detect human encephalomyopathy 50
• Follow mitochondrial changes during apoptosis 51,52
• Investigate mitochondrial poisoning, uncoupling and anoxia 53
• Monitor effects of ellipticine on mitochondrial potential 54
JC-1 has been combined with the reagents in our LIVE/DEADSperm Viability Kit (L-7011, Section 15.3) to permit simultaneousassessment of cellular integrity and mitochondrial function by flowcytometry.55 We have discovered another mitochondrial marker,JC-9 (3,3′-dimethyl-β-naphthoxazolium iodide, D-22421; Figure

479
23.18), with a very different chemical structure (Figure 23.17) butsimilar potential-dependent spectroscopic properties. However, thegreen fluorescence of JC-9 is essentially invariant with membranepotential while the red fluorescence is significantly increased athyperpolarized membrane potentials.
Mitochondrion-Selective Rhodamines andRosamines
Rhodamine 123Rhodamine 123 (R-302; FluoroPure Grade, R-22420; Figure
12.23) is a cell-permeant, cationic, fluorescent dye that is readilysequestered by active mitochondria without inducing cytotoxiceffects.56 Uptake and equilibration of rhodamine 123 is rapid (afew minutes) compared to dyes such as DASPMI, which maytake 30 minutes or longer.13 Viewed through a fluorescein long-pass optical filter (Table 24.8), fluorescence of the mitochondriaof cells stained by rhodamine 123 appears yellow-green. Viewedthrough a tetramethylrhodamine longpass optical filter, however,these same mitochondria appear red. Unlike the lipophilic rhod-amine and carbocyanine dyes, rhodamine 123 apparently does notstain the endoplasmic reticulum.
Rhodamine 123 has been used with a variety of cell types suchas presynaptic nerve terminals,57 live bacteria,58,59 plants 60,61 andhuman spermatozoa.62 Using flow cytometry, researchers em-ployed rhodamine 123 to sort respiratory-deficient yeast cells 63,64
and to isolate those lymphocytes that are responsive to mitogenstimulation.65 Rhodamine 123 has also been used to study:
• Apoptosis 52,66
• Axoplasmic transport of mitochondria 67
• Bacterial viability and vitality 58
• Mitochondrial enzymatic activities 68,69
• Mitochondrial transmembrane potential and other membraneactivities 14,60,70–73
• Multidrug resistance 74–81 (Section 15.6)• Mycobacterial drug susceptibility 82,83
• Oocyte maturation 84
Although rhodamine 123 is usually not retained by cells whenthey are washed, a variety of human carcinoma cell lines (but notsarcomas or leukemic cells) retain the dye for unusually longperiods 85 (>24 hours), making rhodamine 123 a potential antican-cer agent for photodynamic therapy.86–91 Rhodamine 123 is
Figure 12.23 R-302 rhodamine 123. Figure 12.24 T-639 tetramethylrosamine chloride. Figure 12.25 R-634 rhodamine 6G chloride.
known to be preferentially taken up and retained by mitochondriaof carcinoma cells 92 and to inhibit their proliferation; 93,94 cardiacmuscle cells also retain rhodamine 123 for days.95
Tetrabromorhodamine 123The brominated analog of rhodamine 123, tetrabromorhod-
amine 123 (T-7539), is also potentially useful for photodynamictherapy. Rhodamine 123 is a relatively weak phototoxin 90 witha quantum yield for singlet oxygen (1O2) generation of <0.01.Continuous illumination of rhodamine 123–stained cells causesmitochondrial fragmentation,96 possibly due to this release ofactivated oxygen. Tetrabromorhodamine 123 is a much moreeffective 1O2 generator,90 with a quantum yield of 0.7. Althoughthe dye is not localized strictly to mitochondria in cells, tetrabro-morhodamine 123 is highly phototoxic to carcinoma cells 90 andits photoproduct is well retained. Tetrabromorhodamine 123 doesnot stain the nucleus in live cells; however, it binds to DNA insolution, where it has been used as a probe for DNA internalflexibility on the microsecond time scale.97
Rosamines and Other Rhodamine Derivatives, Including TMRMand TMRE
Other mitochondrion-selective dyes include tetramethyl-rosamine (T-639, Figure 12.24), whose fluorescence contrastswell with that of fluorescein for multicolor applications, andrhodamine 6G 89,98–100 (R-634, Figure 12.25), which has an ab-sorption maximum between that of rhodamine 123 and tetra-methylrosamine. Tetramethylrosamine and rhodamine 6G haveboth been used to examine the efficiency of P-glycoprotein–mediated exclusion from multidrug-resistant cells 74 (Section15.6). Rhodamine 6G has been employed to study microvascularreperfusion injury 101 and the stimulation and inhibition ofF1-ATPase from the thermophilic bacterium PS3.102
At low concentrations, certain lipophilic rhodamine dyesselectively stain mitochondria in live cells.103 Molecular Probes’researchers have observed that low concentrations of the hexylester of rhodamine B (R 6, R-648) accumulate selectively inmitochondria and appear to be relatively nontoxic. We haveincluded this probe in our Yeast Mitochondrial Stain Sampler Kit(Y-7530, see below for description). At higher concentrations,rhodamine B hexyl ester and rhodamine 6G stain the endoplasmicreticulum of animal cells 103 (Section 12.4).
The accumulation of tetramethylrhodamine methyl and ethylesters (TMRM, T-668; TMRE, T-669) in mitochondria and theendoplasmic reticulum has also been shown to be driven by their
Section 12.2

480 Chapter 12 — Probes for Organelles www.probes.com
membrane potential 104,105 (Section 23.3). Moreover, because oftheir reduced hydrophobic character, these probes exhibit poten-tial-independent binding to cells that is 10 to 20 times lower thanthat seen with rhodamine 6G.106 Tetramethylrhodamine ethylester has been described as one of the best fluorescent dyes fordynamic and in situ quantitative measurements — better thanrhodamine 123 — because it is rapidly and reversibly takenup by live cells.107–109 TMRM and TMRE have been used tomeasure mitochondrial depolarization related to cytosolic Ca2+
transients 110 and to image time-dependent mitochondrial mem-brane potentials.108 A high-throughput assay utilizes TMRE andour low-affinity Ca2+ indicator fluo-5N AM (F-14204, Section20.3) to screen inhibitors of the opening of the mitochondrialtransition pore.111 Researchers have also taken advantage of thered shift exhibited by TMRM, TMRE and rhodamine 123 uponmembrane potential–driven mitochondrial uptake to develop aratiometric method for quantitating membrane potential.70
Reduced Rhodamines and RosaminesInside live cells, the colorless dihydrorhodamines and dihy-
drotetramethylrosamine are oxidized to fluorescent products thatstain mitochondria.112 However, the oxidation may occur in or-ganelles other than the mitochondria. Dihydrorhodamine 123(D-632, D-23806; Figure 12.26) reacts with hydrogen peroxide inthe presence of peroxidases,113 iron or cytochrome c 114 to formrhodamine 123. This reduced rhodamine has been used to moni-tor reactive oxygen intermediates in rat mast cells 115 and to mea-sure hydrogen peroxide in endothelial cells.114 Dihydrorhodamine6G (D-633, Figure 12.27) is another reduced rhodamine that hasbeen shown to be taken up and oxidized by live cells.116–118
Chloromethyl derivatives of reduced rosamines (MitoTrackerOrange CM-H2TMRos, M-7511; MitoTracker Red CM-H2XRos,M-7513), which can be fixed in cells by aldehyde-based fixatives,have been described above. The acetoxymethyl (AM) ester ofdihydrorhod-2, which is prepared by chemical reduction of thecalcium indicator rhod-2 AM (R-1244, R-1245; Section 20.3) has
Figure 12.26 D-632 dihydrorhodamine 123.
Figure 12.27 D-633 dihydrorhodamine 6G.
been extensively used to measure the relatively slow changes inintramitochondrial Ca2+ (Figure 20.28, Figure 20.34).
Other Mitochondrion-Selective Probes
CarbocyaninesMost carbocyanine dyes with short (C1–C6) alkyl chains
(Section 23.3) stain mitochondria of live cells when used at lowconcentrations (~0.5 µM or ~0.1 µg/mL); those with pentyl orhexyl substituents also stain the endoplasmic reticulum whenused at higher concentrations (~5–50 µM or ~1–10 µg/mL).DiOC6(3) (D-273) stains mitochondria in live yeast 21,119–121 andother eukaryotic cells,100,122 as well as sarcoplasmic reticulum inbeating heart cells.123 It has also been used to demonstrate mito-chondria moving along microtubules.23 Photolysis of mitochon-drion- or endoplasmic reticulum–bound DiOC6(3) specificallydestroys the microtubules of cells without affecting actin stressfibers, producing a highly localized inhibition of intracellularorganelle motility.124 Several other potential-sensitive carbocya-nine probes described in Section 23.3 also stain mitochondria inlive cultured cells.100
The carbocyanine DiOC7(3) (D-378), which exhibits spectrasimilar to those of fluorescein, is a versatile dye that has beenreported to be a sensitive probe for mitochondria in plant cells.125
Its other uses include:
• Distinguishing cycling and noncycling fibroblasts 126 andviable and nonviable bacteria 127
• Following the reorganization of the endoplasmic reticulumduring fertilization in the ascidian egg 128
• Identifying functional vasculature in murine tumors 129,130
• Studying multidrug resistance 131 (Section 15.6)• Visualizing the detailed morphology of neurites of Alz-
heimer’s disease neurons 132
Styryl DyesThe styryl dyes DASPMI (4-Di-1-ASP, D-288) and DASPEI
(D-426) can be used to stain mitochondria in live cells.13 Thesedyes have large fluorescence Stokes shifts and are taken up rela-tively slowly as a function of membrane potential. The kineticsof mitochondrial staining with styrylpyridinium dyes has beeninvestigated using the concentration jump method.133 DASPMIand DASPEI have been shown to be useful for:
• Determining the distribution of mitochondria in yeast mutants 63
• Long-term imaging of live mammalian nerve cells and theirconnections 134–136
• Monitoring the metabolic state of Pneumocystis carinii mito-chondria 31
• Screening aberrant mitochondrial distribution and morphologyin yeast 137
Nonyl Acridine OrangeNonyl acridine orange (A-1372) is well retained in the mito-
chondria of live HeLa cells for up to ten days, making it a usefulprobe for following mitochondria during isolation and after cellfusion.138–140 The mitochondrial uptake of this metachromatic dyeis reported not to depend on membrane potential. It is toxic at
481
high concentrations 141 and apparently binds to cardiolipin in allmitochondria, regardless of their energetic state.142–145 This deriv-ative has been used to analyze mitochondria by flow cytometry,146
to characterize multidrug resistance 147 (Section 15.6) and tomeasure changes in mitochondrial mass during apoptosis in ratthymocytes.52
Carboxy SNARF-1 pH IndicatorA special cell-loading technique permits ratiometric measure-
ment of intramitochondrial pH with our SNARF dyes. Cell load-ing with 10 µM 5-(and -6)-carboxy SNARF-1, acetoxymethylester, acetate (C-1271, C-1272; Section 21.2), followed by 4hours of incubation at room temperature leads to highly selectivelocalization of the carboxy SNARF-1 dye in mitochondria (Fig-ure 21.15), where it responds to changes in mitochondrial pH.148
CoroNa Red ChlorideAs shown by colocalization with MitoTracker Green FM, our
newest Na+ indicator, CoroNa Red chloride (C-24430, C-24431;Section 22.1), spontaneously localizes in the mitochondria (Fig-ure 22.11) and may be useful for measuring intramitochondrialNa+ transients.
LucigeninThe well-known chemiluminescent probe lucigenin (L-6868)
accumulates in mitochondria of alveolar macrophages.149 Rela-tively high concentrations of the dye (~100 µM) are required toobtain fluorescent staining; however, low concentrations report-edly yield a chemiluminescent response to stimulated superoxidegeneration within the mitochondria.149 Lucigenin from MolecularProbes has been highly purified to remove a bright blue-fluores-cent contaminant that is found in some commercial samples.
Yeast Mitochondrial Stain Sampler Kit
Fluorescence microscopy has been extensively used to studyyeast.21,121 Molecular Probes offers a Yeast Mitochondrial StainSampler Kit (Y-7530). This kit contains sample quantities of fivedifferent probes that have been found to selectively label yeastmitochondria. Both well-characterized and proprietary mitochon-drion-selective probes are provided:
• Rhodamine 123 64,150–152
• Rhodamine B hexyl ester 103
• MitoTracker Green FM• SYTO 18 yeast mitochondrial stain 153
• DiOC6(3) 21,120,121,154–160
The mitochondrion-selective nucleic acid stain included inthis kit — SYTO 18 yeast mitochondrial stain — exhibits apronounced fluorescence enhancement upon binding to nucleicacids, resulting in very low background fluorescence even in thepresence of dye. SYTO 18 is an effective mitochondrial stain inlive yeast but neither penetrates nor stains the mitochondria ofhigher eukaryotic cells. Each of the components of the YeastMitochondrial Stain Sampler Kit is also available separately,including the SYTO 18 yeast mitochondrial stain (S-7529).
Avidin Conjugates for Staining Mitochondria
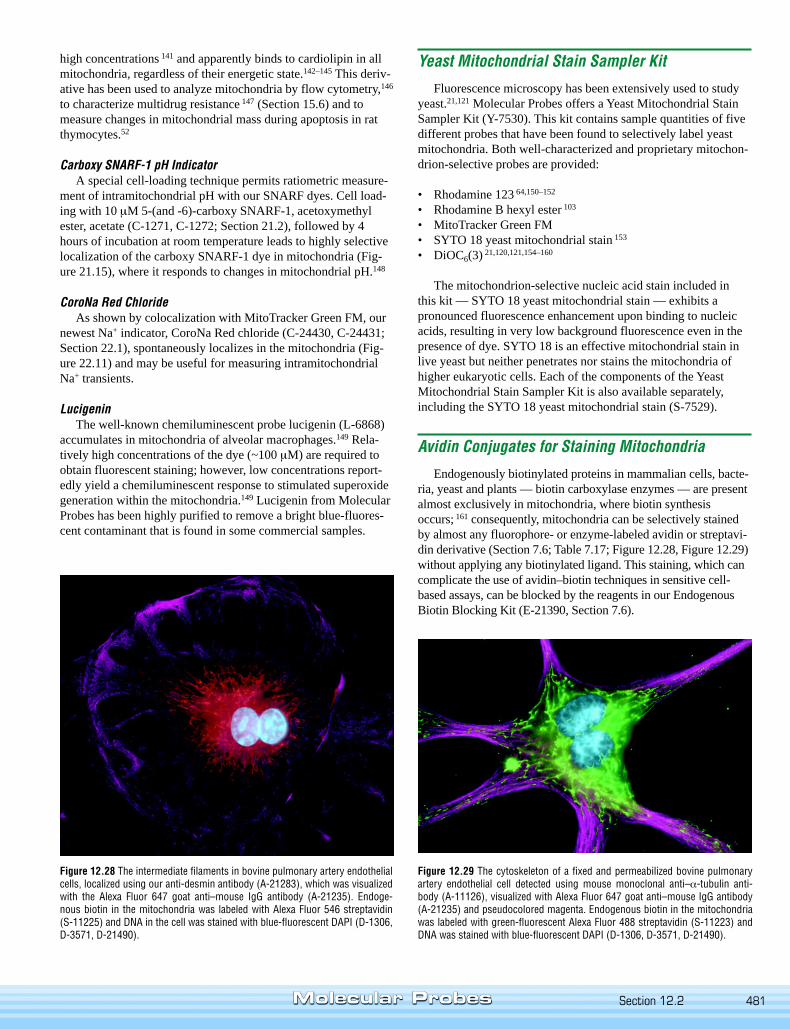
Endogenously biotinylated proteins in mammalian cells, bacte-ria, yeast and plants — biotin carboxylase enzymes — are presentalmost exclusively in mitochondria, where biotin synthesisoccurs; 161 consequently, mitochondria can be selectively stainedby almost any fluorophore- or enzyme-labeled avidin or streptavi-din derivative (Section 7.6; Table 7.17; Figure 12.28, Figure 12.29)without applying any biotinylated ligand. This staining, which cancomplicate the use of avidin–biotin techniques in sensitive cell-based assays, can be blocked by the reagents in our EndogenousBiotin Blocking Kit (E-21390, Section 7.6).
Figure 12.29 The cytoskeleton of a fixed and permeabilized bovine pulmonaryartery endothelial cell detected using mouse monoclonal anti–α-tubulin anti-body (A-11126), visualized with Alexa Fluor 647 goat anti–mouse IgG antibody(A-21235) and pseudocolored magenta. Endogenous biotin in the mitochondriawas labeled with green-fluorescent Alexa Fluor 488 streptavidin (S-11223) andDNA was stained with blue-fluorescent DAPI (D-1306, D-3571, D-21490).
Figure 12.28 The intermediate filaments in bovine pulmonary artery endothelialcells, localized using our anti-desmin antibody (A-21283), which was visualizedwith the Alexa Fluor 647 goat anti–mouse IgG antibody (A-21235). Endoge-nous biotin in the mitochondria was labeled with Alexa Fluor 546 streptavidin(S-11225) and DNA in the cell was stained with blue-fluorescent DAPI (D-1306,D-3571, D-21490).
Section 12.2
482 Chapter 12 — Probes for Organelles www.probes.com
Antibodies to Mitochondrial Proteins
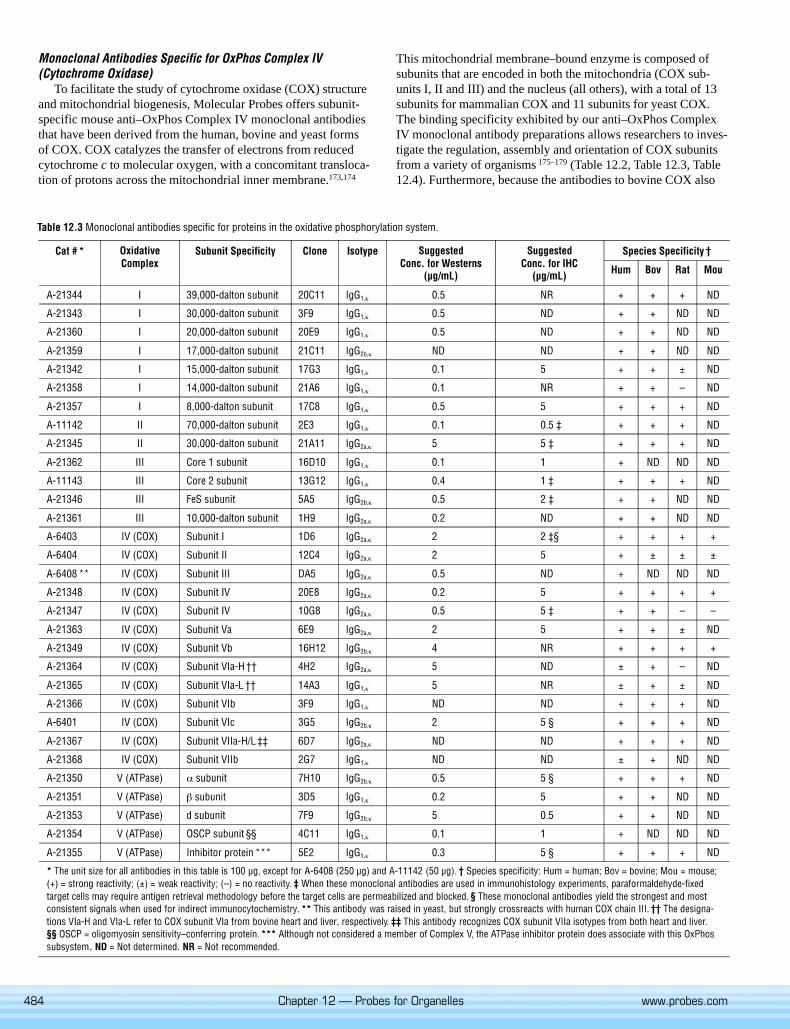
Monoclonal Antibodies Specific for Proteins in the Oxidative Phosphorylation SystemOxidative phosphorylation (OxPhos) activity occurs in the mitochondria and, in mam-
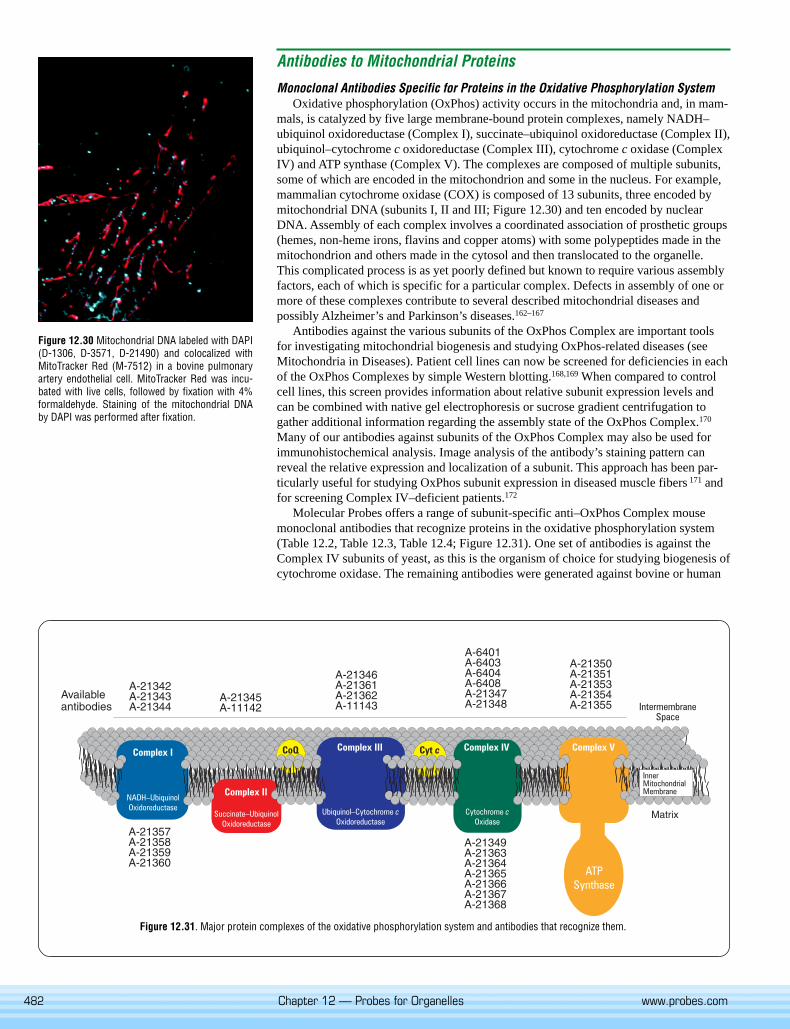
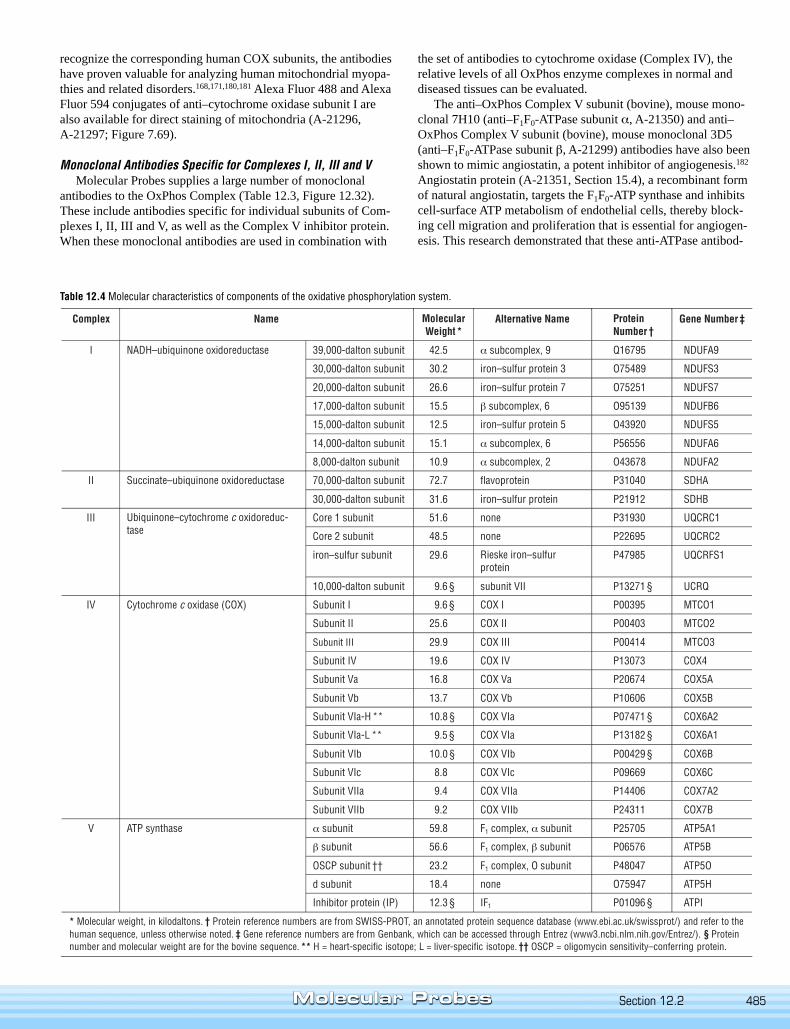
mals, is catalyzed by five large membrane-bound protein complexes, namely NADH–ubiquinol oxidoreductase (Complex I), succinate–ubiquinol oxidoreductase (Complex II),ubiquinol–cytochrome c oxidoreductase (Complex III), cytochrome c oxidase (ComplexIV) and ATP synthase (Complex V). The complexes are composed of multiple subunits,some of which are encoded in the mitochondrion and some in the nucleus. For example,mammalian cytochrome oxidase (COX) is composed of 13 subunits, three encoded bymitochondrial DNA (subunits I, II and III; Figure 12.30) and ten encoded by nuclearDNA. Assembly of each complex involves a coordinated association of prosthetic groups(hemes, non-heme irons, flavins and copper atoms) with some polypeptides made in themitochondrion and others made in the cytosol and then translocated to the organelle.This complicated process is as yet poorly defined but known to require various assemblyfactors, each of which is specific for a particular complex. Defects in assembly of one ormore of these complexes contribute to several described mitochondrial diseases andpossibly Alzheimer’s and Parkinson’s diseases.162–167
Antibodies against the various subunits of the OxPhos Complex are important toolsfor investigating mitochondrial biogenesis and studying OxPhos-related diseases (seeMitochondria in Diseases). Patient cell lines can now be screened for deficiencies in eachof the OxPhos Complexes by simple Western blotting.168,169 When compared to controlcell lines, this screen provides information about relative subunit expression levels andcan be combined with native gel electrophoresis or sucrose gradient centrifugation togather additional information regarding the assembly state of the OxPhos Complex.170
Many of our antibodies against subunits of the OxPhos Complex may also be used forimmunohistochemical analysis. Image analysis of the antibody’s staining pattern canreveal the relative expression and localization of a subunit. This approach has been par-ticularly useful for studying OxPhos subunit expression in diseased muscle fibers 171 andfor screening Complex IV–deficient patients.172
Molecular Probes offers a range of subunit-specific anti–OxPhos Complex mousemonoclonal antibodies that recognize proteins in the oxidative phosphorylation system(Table 12.2, Table 12.3, Table 12.4; Figure 12.31). One set of antibodies is against theComplex IV subunits of yeast, as this is the organism of choice for studying biogenesis ofcytochrome oxidase. The remaining antibodies were generated against bovine or human
Figure 12.31. Major protein complexes of the oxidative phosphorylation system and antibodies that recognize them.
Figure 12.30 Mitochondrial DNA labeled with DAPI(D-1306, D-3571, D-21490) and colocalized withMitoTracker Red (M-7512) in a bovine pulmonaryartery endothelial cell. MitoTracker Red was incu-bated with live cells, followed by fixation with 4%formaldehyde. Staining of the mitochondrial DNAby DAPI was performed after fixation.

483
Table 12.2 Monoclonal antibodies to yeast oxidative phosphorylation complex IV (COX).
Figure 12.32 Western blot analysis of bovine heartmitochondrial lysate probed with antibodies againstcomponents of OxPhos Complex III and OxPhosComplex V. Antibodies against OxPhos Complex IIIcore 2 subunit (A-11143) and OxPhos Complex Valpha subunit (A-21350) were labeled with theZenon One Alexa Fluor 488 Mouse IgG1 Labeling Kit(Z-25002, green fluorescence) and the Zenon OneAlexa Fluor 647 Mouse IgG1 Labeling Kit (Z-25008,far-red fluorescence, pseudocolored magenta),respectively, before probing the blot. The lanes inpanel A are 1) MW marker, 2) 8 µg, 3) 4 µg,4) 2 µg, 5) 1 µg, 6) 0.5 µg, 7) 0.25 µg, 8) 0.125 µg,9) 60 ng and 10) 30 ng of the bovine heart mito-chondrial extract. Panel (B) shows lanes 6 through10 imaged with a longer exposure time. The mo-lecular weight standards are Precision PrestainedProtein Standards (Bio-Rad), which display far-redfluorescence similar to that of Alexa Fluor 647.
A
B
Mitochondria in DiseasesGiven the multiple functions and numerous proteins present
in the mitochondria, it is not surprising that genetically inheriteddefects of mitochondrial function are a major cause of morbidityand mortality in humans. In particular, there are several humandiseases that have known defects in the proteins responsible foroxidative phosphorylation (OxPhos) in cells. Typically, such defectsproduce lactic acidemia, exercise intolerance or neurological dis-orders. Diseases of OxPhos are notoriously difficult to diagnose,and it is even more difficult to correlate their phenotype–genotyperelationships. A subset of OxPhos defects is maternally inherited.These defects result from mutations in mitochondrial DNA (mt-DNA), a small, 16-kilobase genome present in hundreds to thou-sands of copies per cell. mtDNA, which encodes 13 polypeptidesof the OxPhos machinery, differs from the nuclear genome in itsabsence of histones, poor repair mechanisms and very limited
TECHNICAL NOTE
recombination frequencies. As a result, mtDNA in somatic cellsbuilds up mutations over time due to errors in replication that arenot repaired and physical insult from a variety of toxins. Suchaccumulated mutations are implicated in a number of neurodegen-erative diseases — notably Parkinson’s and Alzheimer’s diseases— where the mutation load triggers premature apoptotic or necro-tic cell death. For example, a strong link has been establishedbetween exposure to the pesticide rotenone, a well-defined andspecific inhibitor of OxPhos, and Parkinson’s disease. mtDNAmutations function by reducing energy production within the celland are thought to contribute to cancer and to aging. Likewise,mutations in the nuclear-encoded subunits of OxPhos have beenfound to regulate the life span in flies and worms. Many of theproducts listed in this section are useful tools for studying degen-erative conditions.
material and were selected because they react with high specificity for the human formof the various proteins. All of our antibodies work well in Western blots and a majoritycan be used for immunohistochemistry, as listed in Table 12.3. These antibodies mayalso be employed to test other subcellular preparations for mitochondrial contamination.Stringent selection criteria were applied during the development of these monoclonalantibodies, including:
• Ability of the antibodies to detect native protein in solid-phase binding assays suchas particle-concentration fluorescence immunoassays (PCFIAs) and enzyme-linkedimmunosorbent assays (ELISAs)
• Specificity for the appropriate denatured subunit in Western blots of whole-cell ex-tracts and isolated mitochondria
• Where appropriate, specific mitochondrial subcellular localization of immunohis-tochemical reactivity in fixed cultured human cells
Detailed information regarding the IgG isotype and recommended working concen-tration is provided with each product. For detection of these monoclonal antibodies,Molecular Probes offers anti–mouse IgG secondary antibodies labeled with biotin,enzymes, NANOGOLD and Alexa Fluor FluoroNanogold 1.4 nm gold clusters, Capti-vate ferrofluid or a wide range of fluorophores (Section 7.3, Table 7.3). The IgG1 anti-bodies in this group (Table 12.3) can be complexed with the reagents in our Zenon OneMouse IgG1 Labeling Kits (Section 7.2, Table 7.1) for labeling mitochondrial proteins inWestern blots (Figure 12.32) and cells (Figure 15.64).
Section 12.2
484 Chapter 12 — Probes for Organelles www.probes.com
Monoclonal Antibodies Specific for OxPhos Complex IV(Cytochrome Oxidase)
To facilitate the study of cytochrome oxidase (COX) structureand mitochondrial biogenesis, Molecular Probes offers subunit-specific mouse anti–OxPhos Complex IV monoclonal antibodiesthat have been derived from the human, bovine and yeast formsof COX. COX catalyzes the transfer of electrons from reducedcytochrome c to molecular oxygen, with a concomitant transloca-tion of protons across the mitochondrial inner membrane.173,174
This mitochondrial membrane–bound enzyme is composed ofsubunits that are encoded in both the mitochondria (COX sub-units I, II and III) and the nucleus (all others), with a total of 13subunits for mammalian COX and 11 subunits for yeast COX.The binding specificity exhibited by our anti–OxPhos ComplexIV monoclonal antibody preparations allows researchers to inves-tigate the regulation, assembly and orientation of COX subunitsfrom a variety of organisms 175–179 (Table 12.2, Table 12.3, Table12.4). Furthermore, because the antibodies to bovine COX also
Table 12.3 Monoclonal antibodies specific for proteins in the oxidative phosphorylation system.
485
Table 12.4 Molecular characteristics of components of the oxidative phosphorylation system.
recognize the corresponding human COX subunits, the antibodieshave proven valuable for analyzing human mitochondrial myopa-thies and related disorders.168,171,180,181 Alexa Fluor 488 and AlexaFluor 594 conjugates of anti–cytochrome oxidase subunit I arealso available for direct staining of mitochondria (A-21296,A-21297; Figure 7.69).
Monoclonal Antibodies Specific for Complexes I, II, III and VMolecular Probes supplies a large number of monoclonal
antibodies to the OxPhos Complex (Table 12.3, Figure 12.32).These include antibodies specific for individual subunits of Com-plexes I, II, III and V, as well as the Complex V inhibitor protein.When these monoclonal antibodies are used in combination with
the set of antibodies to cytochrome oxidase (Complex IV), therelative levels of all OxPhos enzyme complexes in normal anddiseased tissues can be evaluated.
The anti–OxPhos Complex V subunit (bovine), mouse mono-clonal 7H10 (anti–F1F0-ATPase subunit α, A-21350) and anti–OxPhos Complex V subunit (bovine), mouse monoclonal 3D5(anti–F1F0-ATPase subunit β, A-21299) antibodies have also beenshown to mimic angiostatin, a potent inhibitor of angiogenesis.182
Angiostatin protein (A-21351, Section 15.4), a recombinant formof natural angiostatin, targets the F1F0-ATP synthase and inhibitscell-surface ATP metabolism of endothelial cells, thereby block-ing cell migration and proliferation that is essential for angiogen-esis. This research demonstrated that these anti-ATPase antibod-
Section 12.2
486 Chapter 12 — Probes for Organelles www.probes.com
ies had similar inhibitory effects, implying that they also compro-mised ATP metabolism and may function as angiostatin analogs.
Antibodies Against Other Mitochondrial Proteins
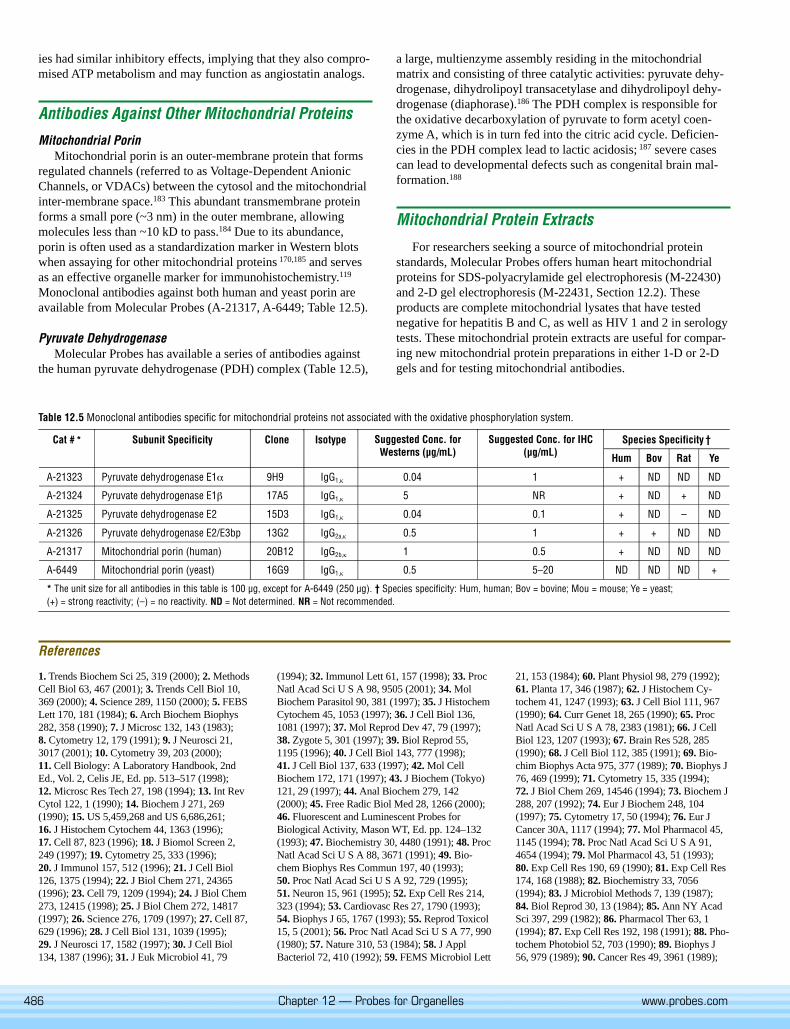
Mitochondrial PorinMitochondrial porin is an outer-membrane protein that forms
regulated channels (referred to as Voltage-Dependent AnionicChannels, or VDACs) between the cytosol and the mitochondrialinter-membrane space.183 This abundant transmembrane proteinforms a small pore (~3 nm) in the outer membrane, allowingmolecules less than ~10 kD to pass.184 Due to its abundance,porin is often used as a standardization marker in Western blotswhen assaying for other mitochondrial proteins 170,185 and servesas an effective organelle marker for immunohistochemistry.119
Monoclonal antibodies against both human and yeast porin areavailable from Molecular Probes (A-21317, A-6449; Table 12.5).
Pyruvate DehydrogenaseMolecular Probes has available a series of antibodies against
the human pyruvate dehydrogenase (PDH) complex (Table 12.5),
a large, multienzyme assembly residing in the mitochondrialmatrix and consisting of three catalytic activities: pyruvate dehy-drogenase, dihydrolipoyl transacetylase and dihydrolipoyl dehy-drogenase (diaphorase).186 The PDH complex is responsible forthe oxidative decarboxylation of pyruvate to form acetyl coen-zyme A, which is in turn fed into the citric acid cycle. Deficien-cies in the PDH complex lead to lactic acidosis; 187 severe casescan lead to developmental defects such as congenital brain mal-formation.188
Mitochondrial Protein Extracts
For researchers seeking a source of mitochondrial proteinstandards, Molecular Probes offers human heart mitochondrialproteins for SDS-polyacrylamide gel electrophoresis (M-22430)and 2-D gel electrophoresis (M-22431, Section 12.2). Theseproducts are complete mitochondrial lysates that have testednegative for hepatitis B and C, as well as HIV 1 and 2 in serologytests. These mitochondrial protein extracts are useful for compar-ing new mitochondrial protein preparations in either 1-D or 2-Dgels and for testing mitochondrial antibodies.
Table 12.5 Monoclonal antibodies specific for mitochondrial proteins not associated with the oxidative phosphorylation system.
References
1. Trends Biochem Sci 25, 319 (2000); 2. MethodsCell Biol 63, 467 (2001); 3. Trends Cell Biol 10,369 (2000); 4. Science 289, 1150 (2000); 5. FEBSLett 170, 181 (1984); 6. Arch Biochem Biophys282, 358 (1990); 7. J Microsc 132, 143 (1983);8. Cytometry 12, 179 (1991); 9. J Neurosci 21,3017 (2001); 10. Cytometry 39, 203 (2000);11. Cell Biology: A Laboratory Handbook, 2ndEd., Vol. 2, Celis JE, Ed. pp. 513–517 (1998);12. Microsc Res Tech 27, 198 (1994); 13. Int RevCytol 122, 1 (1990); 14. Biochem J 271, 269(1990); 15. US 5,459,268 and US 6,686,261;16. J Histochem Cytochem 44, 1363 (1996);17. Cell 87, 823 (1996); 18. J Biomol Screen 2,249 (1997); 19. Cytometry 25, 333 (1996);20. J Immunol 157, 512 (1996); 21. J Cell Biol126, 1375 (1994); 22. J Biol Chem 271, 24365(1996); 23. Cell 79, 1209 (1994); 24. J Biol Chem273, 12415 (1998); 25. J Biol Chem 272, 14817(1997); 26. Science 276, 1709 (1997); 27. Cell 87,629 (1996); 28. J Cell Biol 131, 1039 (1995);29. J Neurosci 17, 1582 (1997); 30. J Cell Biol134, 1387 (1996); 31. J Euk Microbiol 41, 79
(1994); 32. Immunol Lett 61, 157 (1998); 33. ProcNatl Acad Sci U S A 98, 9505 (2001); 34. MolBiochem Parasitol 90, 381 (1997); 35. J HistochemCytochem 45, 1053 (1997); 36. J Cell Biol 136,1081 (1997); 37. Mol Reprod Dev 47, 79 (1997);38. Zygote 5, 301 (1997); 39. Biol Reprod 55,1195 (1996); 40. J Cell Biol 143, 777 (1998);41. J Cell Biol 137, 633 (1997); 42. Mol CellBiochem 172, 171 (1997); 43. J Biochem (Tokyo)121, 29 (1997); 44. Anal Biochem 279, 142(2000); 45. Free Radic Biol Med 28, 1266 (2000);46. Fluorescent and Luminescent Probes forBiological Activity, Mason WT, Ed. pp. 124–132(1993); 47. Biochemistry 30, 4480 (1991); 48. ProcNatl Acad Sci U S A 88, 3671 (1991); 49. Bio-chem Biophys Res Commun 197, 40 (1993);50. Proc Natl Acad Sci U S A 92, 729 (1995);51. Neuron 15, 961 (1995); 52. Exp Cell Res 214,323 (1994); 53. Cardiovasc Res 27, 1790 (1993);54. Biophys J 65, 1767 (1993); 55. Reprod Toxicol15, 5 (2001); 56. Proc Natl Acad Sci U S A 77, 990(1980); 57. Nature 310, 53 (1984); 58. J ApplBacteriol 72, 410 (1992); 59. FEMS Microbiol Lett
21, 153 (1984); 60. Plant Physiol 98, 279 (1992);61. Planta 17, 346 (1987); 62. J Histochem Cy-tochem 41, 1247 (1993); 63. J Cell Biol 111, 967(1990); 64. Curr Genet 18, 265 (1990); 65. ProcNatl Acad Sci U S A 78, 2383 (1981); 66. J CellBiol 123, 1207 (1993); 67. Brain Res 528, 285(1990); 68. J Cell Biol 112, 385 (1991); 69. Bio-chim Biophys Acta 975, 377 (1989); 70. Biophys J76, 469 (1999); 71. Cytometry 15, 335 (1994);72. J Biol Chem 269, 14546 (1994); 73. Biochem J288, 207 (1992); 74. Eur J Biochem 248, 104(1997); 75. Cytometry 17, 50 (1994); 76. Eur JCancer 30A, 1117 (1994); 77. Mol Pharmacol 45,1145 (1994); 78. Proc Natl Acad Sci U S A 91,4654 (1994); 79. Mol Pharmacol 43, 51 (1993);80. Exp Cell Res 190, 69 (1990); 81. Exp Cell Res174, 168 (1988); 82. Biochemistry 33, 7056(1994); 83. J Microbiol Methods 7, 139 (1987);84. Biol Reprod 30, 13 (1984); 85. Ann NY AcadSci 397, 299 (1982); 86. Pharmacol Ther 63, 1(1994); 87. Exp Cell Res 192, 198 (1991); 88. Pho-tochem Photobiol 52, 703 (1990); 89. Biophys J56, 979 (1989); 90. Cancer Res 49, 3961 (1989);

487
References — continued
91. Photochem Photobiol 48, 613 (1988); 92. Can-cer Res 45, 6093 (1985); 93. Science 218, 1117(1982); 94. Biochem Biophys Res Commun 118,717 (1984); 95. Proc Natl Acad Sci U S A 79, 5292(1982); 96. Cancer Res 50, 4167 (1990); 97. ProcNatl Acad Sci U S A 79, 5896 (1982); 98. His-tochemistry 94, 303 (1990); 99. Exp Pathol 31, 47(1987); 100. J Cell Biol 88, 526 (1981); 101. Trans-plantation 58, 403 (1994); 102. J Bioenerg Bio-membr 25, 679 (1993); 103. J Cell Sci 101, 315(1992); 104. Biophys J 56, 1053 (1989); 105. Bio-phys J 53, 785 (1988); 106. J Fluorescence 3, 265(1993); 107. Cell Biology: A Laboratory Hand-book, Vol. 2, Celis JE, Ed. pp. 399–403 (1994);108. Biophys J 65, 2396 (1993); 109. OpticalMicroscopy for Biology, Herman B, Jacobson K,Eds. pp. 131–142 (1990); 110. Proc Natl Acad SciU S A 91, 12579 (1994); 111. Anal Biochem 295,220 (2001); 112. Methods Cell Biol 29, 103(1989); 113. Eur J Biochem 217, 973 (1993);114. Arch Biochem Biophys 302, 348 (1993);115. APMIS 102, 474 (1994); 116. Free Radicals:A Practical Approach, Punchard NA, Ed. pp. 83–99 (1996); 117. Proc Natl Acad Sci U S A 93, 1167(1996); 118. Methods Enzymol 172, 102 (1989);119. Mol Biol Cell 9, 917 (1998); 120. Cell MotilCytoskeleton 25, 111 (1993); 121. Methods Cell
Biol 31, 357 (1989); 122. Methods Cell Biol 29,125 (1989); 123. Exp Cell Res 125, 514 (1980);124. Cancer Res 55, 2063 (1995); 125. Plant Phys-iol 84, 1385 (1987); 126. Nature 290, 593 (1981);127. J Appl Bacteriol 78, 309 (1995); 128. J CellBiol 120, 1337 (1993); 129. Br J Cancer 62, 903(1990); 130. Br J Cancer 59, 706 (1989); 131. Bio-chemistry 34, 3858 (1995); 132. J Cell Biol 107,2703 (1988); 133. Histochemistry 99, 75 (1993);134. J Neurocytol 19, 67 (1990); 135. J Neurosci 7,1207 (1987); 136. Trends Neurosci 10, 398 (1987);137. J Cell Biol 126, 1361 (1994); 138. Histochem-istry 82, 51 (1985); 139. Histochemistry 80, 385(1984); 140. Histochemistry 79, 443 (1983);141. FEBS Lett 260, 236 (1990); 142. J Dent Res74, 1295 (1995); 143. Eur J Biochem 228, 113(1995); 144. Eur J Biochem 194, 389 (1990);145. Biochem Biophys Res Commun 164, 185(1989); 146. Basic Appl Histochem 33, 71 (1989);147. Cancer Res 51, 4665 (1991); 148. Biotech-niques 30, 804 (2001); 149. Free Radic Biol Med17, 117 (1994); 150. J Biol Chem 274, 543 (1999);151. Mol Biol Cell 9, 523 (1998); 152. Yeast 14,147 (1998); 153. Biochim Biophys Acta 1366, 177(1998); 154. J Cell Biol 143, 359 (1998); 155. JCell Biol 143, 333 (1998); 156. J Cell Biol 141,
1371 (1998); 157. Cytometry 23, 28 (1996); 158. JCell Biol 130, 345 (1995); 159. Mol Biol Cell 6,1381 (1995); 160. Biochem Int 2, 503 (1981);161. Histochemistry 100, 415 (1993); 162. Bio-chim Biophys Acta 1366, 199 (1998); 163. Bio-chim Biophys Acta 1366, 211 (1998); 164. CurrOpin Cardiol 13, 190 (1998); 165. Ann Neurol 44,S99 (1998); 166. J Neural Transm 105, 855 (1998);167. Semin Liver Dis 18, 237 (1998); 168. Bio-chim Biophys Acta 1362, 145 (1997); 169. J BiolChem 276, 8892 (2001); 170. J Biol Chem 276,16296 (2001); 171. Biochim Biophys Acta 1315,199 (1996); 172. Brain 123, 591 (2000); 173. Sci-ence 283, 1488 (1999); 174. Annu Rev Biochem59, 569 (1990); 175. Biochemistry 30, 3674(1991); 176. Biochim Biophys Acta 1225, 95(1993); 177. Methods Enzymol 260, 117 (1995);178. J Biol Chem 268, 18754 (1993); 179. J BiolChem 266, 7688 (1991); 180. Pediatr Res 28, 529(1990); 181. Hum Mol Genet 6, 935 (1997);182. Proc Natl Acad Sci U S A 98, 6656 (2001);183. Biochim Biophys Acta 894, 109 (1987);184. J Biol Chem 273, 24406 (1998); 185. BiochimBiophys Acta 1455, 35 (1999); 186. J Biol Chem272, 5757 (1997); 187. Biochem J 239, 89 (1986);188. Neurology 53, 612 (1999).
Data Table — 12.2 Probes for Mitochondria
Cat # MW Storage Soluble Abs EC Em Solvent NotesA-1372 472.51 L DMSO, EtOH 495 84,000 519 MeOHD-273 572.53 D,L DMSO 484 154,000 501 MeOHD-288 366.24 L DMF 475 45,000 605 MeOH 1D-378 600.58 D,L DMSO 482 148,000 504 MeOHD-426 380.27 L DMF 461 39,000 589 MeOH 1D-632 346.38 F,D,L,AA DMF, DMSO 289 7,100 none MeOH 2, 3D-633 444.57 F,D,L,AA DMF, DMSO 296 11,000 none MeOH 2, 3D-22421 532.38 D,L DMSO, DMF 522 143,000 535 CHCl3 4D-23806 346.38 F,D,L,AA DMSO 289 7,100 none MeOH 3, 5L-6868 510.50 L H2O 455 7,400 505 H2O 6, 7M-7502 602.99 L DMSO 489 112,000 517 MeOHM-7510 427.37 F,D,L DMSO 551 102,000 576 MeOHM-7511 392.93 F,D,L,AA DMSO 235 57,000 none MeOH 2, 3M-7512 531.52 F,D,L DMSO 578 116,000 599 MeOHM-7513 497.08 F,D,L,AA DMSO 245 45,000 none MeOH 2, 3M-7514 671.88 F,D,L DMSO 490 119,000 516 MeOHM-22422 647.10 F,D,L DMSO 604 97,000 637 MeOHM-22423 495.96 F,D,L DMSO 682 95,000 702 EtOHM-22424 ~550 F,D,L DMSO 588 105,000 622 MeOH 8M-22425 724.00 F,D,L DMSO 588 81,000 645 MeOHM-22426 543.58 F,D,L DMSO 640 194,000 662 MeOHR-302 380.83 F,D,L MeOH, DMF 507 101,000 529 MeOHR-634 479.02 F,D,L EtOH 528 105,000 551 MeOHR-648 627.18 F,D,L DMF, DMSO 556 123,000 578 MeOHR-14060 434.41 F,D,L,AA DMSO 239 52,000 none MeOH 2, 9R-22420 380.83 F,D,L MeOH, DMF 507 101,000 529 MeOH 10S-7529 ~450 F,D,L DMSO 483 64,000 none pH 7 8, 11, 12T-639 378.90 L DMF, DMSO 550 87,000 574 MeOHT-668 500.93 F,D,L DMSO, MeOH 549 115,000 573 MeOHT-669 514.96 F,D,L DMSO, EtOH 549 109,000 574 MeOHT-3168 652.23 D,L DMSO, DMF 514 195,000 529 MeOH 13T-7539 740.87 F,D,L MeOH, DMF 524 91,000 550 MeOH
For definitions of the contents of this data table, see “How to Use This Book” on page viii.
Notes1. Abs and Em of styryl dyes are at shorter wavelengths in membrane environments than in reference solvents such as methanol. The difference is typically 20 nm for absorption and
80 nm for emission, but varies considerably from one dye to another.2. This compound is susceptible to oxidation, especially in solution. Store solutions under argon or nitrogen. Oxidation appears to be catalyzed by illumination.
Section 12.2

488 Chapter 12 — Probes for Organelles www.probes.com
Data Table — 12.2 Probes for Mitochondria — continued
3. These compounds are essentially colorless and nonfluorescent until oxidized. Oxidation products (in parentheses) are as follows: D-632 and D-23806 (R-302); D-633 (R-634);M-7511 (M-7510); M-7513 (M-7512).
4. JC-9 exhibits long-wavelength J-aggregate emission at ~635 nm in aqueous solutions and polarized mitochondria.5. This product is supplied as a ready-made solution in DMSO with sodium borohydride added to inhibit oxidation.6. L-6868 has much stronger absorption at shorter wavelengths (Abs = 368 nm (EC = 36,000 cm-1M-1)).7. This compound emits chemiluminescence (Em = 470 nm) upon oxidation in basic aqueous solutions.8. MW: The preceding ~ symbol indicates an approximate value, not including counterions.9. R-14060 is colorless and nonfluorescent until oxidized. The spectral characteristics of the oxidation product (2,3,4,5,6-pentafluorotetramethylrosamine) are similar to those of T-639.
10. This product is specified to equal or exceed 98% analytical purity by HPLC.11. This product is supplied as a ready-made solution in the solvent indicated under Soluble.12. S-7529 is fluorescent when bound to DNA (Abs = 490 nm, Em = 507 nm).13. JC-1 forms J-aggregates with Abs/Em = 585/590 nm at concentrations above 0.1 µM in aqueous solutions (pH 8.0) (Biochemistry 30, 4480 (1991)).
Product List — 12.2 Probes for Mitochondria
The full citations and, in most cases, links to PubMed for all references in thisHandbook are available at our Web site (www.probes.com/search).
Monoclonal antibody products are listed in Tables 12.2, 12.3 and 12.5.
Cat # Product Name Unit SizeA-1372 acridine orange 10-nonyl bromide (nonyl acridine orange) .................................................................................................................................. 100 mgA-21296 anti-OxPhos Complex IV subunit I, mouse IgG2a, monoclonal 1D6, Alexa Fluor® 488 conjugate
(anti-cytochrome oxidase subunit I, Alexa Fluor® 488 conjugate) *1 mg/mL* *human reactivity* ..................................................................... 100 µLA-21297 anti-OxPhos Complex IV subunit I, mouse IgG2a, monoclonal 1D6, Alexa Fluor® 594 conjugate
(anti-cytochrome oxidase subunit I, Alexa Fluor® 594 conjugate) *1 mg/mL* *human reactivity* ..................................................................... 100 µLD-378 3,3′-diheptyloxacarbocyanine iodide (DiOC7(3)) ................................................................................................................................................... 100 mgD-273 3,3′-dihexyloxacarbocyanine iodide (DiOC6(3)) .................................................................................................................................................... 100 mgD-632 dihydrorhodamine 123 ......................................................................................................................................................................................... 10 mgD-23806 dihydrorhodamine 123 *5 mM stabilized solution in DMSO* .............................................................................................................................. 1 mLD-633 dihydrorhodamine 6G ........................................................................................................................................................................................... 25 mgD-426 2-(4-(dimethylamino)styryl)-N-ethylpyridinium iodide (DASPEI) ......................................................................................................................... 1 gD-288 4-(4-(dimethylamino)styryl)-N-methylpyridinium iodide (4-Di-1-ASP) ................................................................................................................ 1 gD-22421 3,3′-dimethyl-α-naphthoxacarbocyanine iodide (JC-9; DiNOC1(3)) ...................................................................................................................... 5 mgE-21390 Endogenous Biotin-Blocking Kit *100 assays* ..................................................................................................................................................... 1 kitL-6868 lucigenin (bis-N-methylacridinium nitrate) *high purity* ..................................................................................................................................... 10 mgM-22431 mitochondrial proteins (human heart) for 2-D gel electrophoresis *5 mg/mL* ................................................................................................... 100 µLM-22430 mitochondrial proteins (human heart) for SDS-polyacrylamide gel electrophoresis *2 mg/mL* ......................................................................... 100 µLM-22423 MitoFluor™ Far Red 680 ....................................................................................................................................................................................... 10 mgM-7502 MitoFluor™ Green ................................................................................................................................................................................................. 1 mgM-22424 MitoFluor™ Red 589 ............................................................................................................................................................................................. 1 mgM-22422 MitoFluor™ Red 594 ............................................................................................................................................................................................. 1 mgM-22426 MitoTracker® Deep Red 633 *special packaging* ................................................................................................................................................ 20 x 50 µgM-7514 MitoTracker® Green FM *special packaging* ....................................................................................................................................................... 20 x 50 µgM-7511 MitoTracker® Orange CM-H2TMRos *special packaging* .................................................................................................................................... 20 x 50 µgM-7510 MitoTracker® Orange CMTMRos *special packaging* ......................................................................................................................................... 20 x 50 µgM-22425 MitoTracker® Red 580 *special packaging* ......................................................................................................................................................... 20 x 50 µgM-7513 MitoTracker® Red CM-H2XRos *special packaging* ............................................................................................................................................ 20 x 50 µgM-7512 MitoTracker® Red CMXRos *special packaging* ................................................................................................................................................. 20 x 50 µgR-14060 RedoxSensor™ Red CC-1 *special packaging* .................................................................................................................................................... 10 x 50 µgR-302 rhodamine 123 ..................................................................................................................................................................................................... 25 mgR-22420 rhodamine 123 *FluoroPure™ grade* .................................................................................................................................................................. 25 mgR-634 rhodamine 6G chloride ......................................................................................................................................................................................... 1 gR-648 rhodamine B, hexyl ester, perchlorate (R 6) ......................................................................................................................................................... 10 mgS-7529 SYTO® 18 yeast mitochondrial stain *5 mM solution in DMSO*.......................................................................................................................... 250 µLT-7539 2′,4′,5′,7′-tetrabromorhodamine 123 bromide ..................................................................................................................................................... 5 mgT-3168 5,5′,6,6′-tetrachloro-1,1′,3,3′-tetraethylbenzimidazolylcarbocyanine iodide (JC-1; CBIC2(3)) .............................................................................. 5 mgT-669 tetramethylrhodamine, ethyl ester, perchlorate (TMRE) ....................................................................................................................................... 25 mgT-668 tetramethylrhodamine, methyl ester, perchlorate (TMRM) ................................................................................................................................... 25 mgT-639 tetramethylrosamine chloride ............................................................................................................................................................................... 25 mgY-7530 Yeast Mitochondrial Stain Sampler Kit ................................................................................................................................................................. 1 kit