sequence and molecular characterization of a dna region
TRANSCRIPT

JOURNAL OF BACTERIOLOGY, June 1993, p. 3394-3400 Vol. 175, No. 110021-9193/93/113394-07$02.00/0Copyright © 1993, American Society for Microbiology
Sequence and Molecular Characterization of a DNA RegionEncoding a Small Heat Shock Protein of
Clostridium acetobutylicumUWE SAUER AND PETER DURRE*
Institut ffir Mikrobiologie, Georg-August-Universitat Gottingen, Gnrsebachstrasse 8,W-3400 Gottingen, Germany
Received 9 November 1992/Accepted 24 March 1993
A DNA region of Clostridium acetobutylicum containing a gene (hspl8) with significant homology to a familyof small eukaryotic heat shock proteins was cloned and sequenced. It is the second reported sequence of alow-molecular-weight heat shock protein from gram-positive bacteria and is induced not only by heat shock butalso at the onset of solventogenesis, as determined by Northern (RNA) blot analysis, thus confirming the resultsof an earlier study performed at the protein level (A. Pich, F. Narberhaus, and H. Bahl, Appl. Microbiol.Biotechnol. 33:697-704, 1990). By primer extension analysis, a transcriptional start site was identified 149 bpupstream of hspl8. This site was preceded by a region that exhibits high homology to the consensus promotersequences of gram-positive bacteria, as well as o70-dependent Escherichia coli. A direct repeat structure wasdetected in the -35 region. The promoter is located 196 bp from the start of a potential regulatory tRNAjjGgene involved in the shift to solventogenesis which is transcribed in the opposite direction. A putativerho-independent transcription termination structure was identified at the 3' end of hspl8.
Clostridium acetobutylicum, a strictly anaerobic, gram-positive, spore-forming bacterium, is able to alter its metab-olism in accordance with growth conditions. During theexponential growth phase in batch cultures, the organismferments sugars to acetate, butyrate, H2, and CO2. With thedecrease of the external pH to below 5 and increase of theproduced butyric acid to certain threshold concentrations,residual sugar and the excreted acids are converted to themain products acetone and butanol shortly before entry intothe stationary phase (for reviews, see references 3, 11, and27). Along with the shift from acid to solvent formation, thecells undergo a series of morphological and physiologicalchanges in motility, shape, and granulose content, culmi-nated by the formation of endospores as the stationary phaseproceeds (14). Furthermore, the shift to solventogenesisseems to be connected to the heat shock response (25, 32).Specific heat shock proteins (HSPs), including DnaK,GroEL, and Hspl8 (originally designated Hspl7 on the basisof protein size determination), are induced during the onsetof solvent formation (25). Since the induction of these HSPswas observed before acetone and butanol were detected inthe medium, their induction might be triggered by the samesignal which is responsible for the shift to solventogenesis.In other microorganisms, both prokaryotic and eukaryotic,synthesis of HSPs seems to be related to developmentalstage or cytodifferentiation as well (12, 13, 15). However, themolecular mechanisms that underlie the metabolic shift tosolventogenesis and its connection to heat shock are stillmostly unknown.
Recently, a transposon-induced mutant of C. acetobutyli-cum that was unable to produce acetone and butanol hasbeen isolated. The effect was shown to be caused byinsertion of a single copy of transposon Tn916, indicatinginactivation of a regulatory locus (5). The DNA region of thismutant has been cloned and partly sequenced. It contains a
* Corresponding author.
gene for a threonine-specific tRNA (30). In this report, wedescribe the further sequencing and molecular characteriza-tion of this DNA region, which was found to encode a smallHSP.
MATERIALS AND METHODS
Bacterial strains, plasmids, and growth conditions. C. ace-tobutylicum DSM 792 (Deutsche Sammlung von Mikroor-ganismen, Braunschweig, Germany), ATCC 824 (kindlysupplied by E. T. Papoutsakis, Northwestern University,Evanston, Ill.), NCIB 8052 (kindly supplied by N. P. Min-ton, Public Health Laboratory Service, Centre for AppliedMicrobiology and Research, Porton Down, United King-dom), and P262 (kindly supplied by D. R. Woods, Universityof Cape Town, Cape Town, South Africa) were used assources of genomic DNA. All C. acetobutylicum strains arephenotypically identical in that they perform butyric acidfermentation and are able to shift their metabolism towardsacetone and butanol production in a later growth phase.Total RNA from heat-shocked cells was isolated from DSM792, and C. acetobutylicum DSM 1731 was used for isolationof total RNA during the shift to solventogenesis. Escherichiacoli JM83 (37) was used as the host and pUC9 (37) was usedas the vector for cloning experiments. In batch culture, C.acetobutylicum was grown under strictly anaerobic condi-tions at 370C in minimal medium as described earlier (5). Forthe mutant, tetracycline was added to a final concentrationof 10 ,ug/ml. Continuous culture experiments were per-formed, as described by Bahl et al. (2), with strain DSM 1731because it can be reliably induced to solvent production.Type strain DSM 792 is more sensitive to lower pH values(8). Solventogenesis was induced by turning off the pHcontrol of the continuous culture. Because of the acidsproduced, the pH fell to 4.3, at which point the pH controlwas turned on again. At this point, solvent formation hadbeen established. E. coli was routinely grown at 370C in LBmedium (28) supplemented with ampicillin (50 ,ug/ml), iso-
3394

SMALL HEAT SHOCK GENE OF C. ACETOBUTYLICUM 3395
propyl-i-D-thiogalactopyranoside (IPTG; 50 jxg/ml), or5-bromo-4-chloro-3-indolyl-3-D-galactopyranoside (X-Gal;40 Rg/ml) if required.
Nucleic acid isolation and manipulation. ChromosomalDNA of C. acetobutylicum was isolated by the method ofMarmur (17) as modified by Bertram and Durre (4). Plasmidisolation from E. coli was performed with the Quiagen MidiKit (Diagen GmbH, Dusseldorf, Germany). Total RNA of C.acetobutylicum was isolated by the hot phenol-chloroformprocedure described by Oelmuller et al. (24) with the modi-fications described by Gerischer and Durre (8).DNA was manipulated by standard methods (28). Restric-
tion enzymes and Taq DNA polymerase were obtained fromGIBCO/BRL GmbH (Eggenstein, Germany) and used inaccordance with the instructions of the manufacturer.
Hybridization. Chromosomal DNA of C. acetobutylicumwas digested to completion with the respective restrictionenzymes, and the fragments obtained were separated onagarose gels. Southern blots on nylon membranes (Gene-Screen Plus; Dupont, NEN Research Products, Dreieich,Germany) were prepared in accordance with the manufac-turer's instructions and subjected to prehybridization in0.15% (wtlvol) polyvinylpyrrolidone-0.15% (wtlvol) bovineserum albumin-0.15% (wtlvol) Ficoll-0.9 M NaCl-1% so-dium dodecyl sulfate (SDS)-6 mM EDTA-90 mM Trishydrochloride (pH 7.5)-100 pg of denatured salmon spermDNA per ml (80 RI/cm2 of membrane) for 1 to 3 h at 60'C.Polymerase chain reaction (PCR)-generated DNA probes,made with synthetic oligonucleotides (17-mers) prepared forthe sequencing reactions, were radiolabeled with [a-32P]dATP (Amersham Buchler GmbH, Braunschweig, Ger-many) by using a random-primer DNA labeling kit (GIBCO/BRL). The labeled probes were purified by column purifica-tion on Sephadex G-25, and 0.2 to 0.5 ,uCi/ml was added tothe prehybridization solution. After hybridization at 60°C for10 to 15 h, the membranes were washed twice in 2x SSC (1 xSSC is 0.15 M NaCl plus 0.015 M sodium citrate) and once in2x SSC plus 1% (wt/vol) SDS for 5 min at room temperatureand subjected to autoradiography.RNA for Northern (RNA) blots was separated in denatur-
ing formaldehyde gels and transferred to nylon membranes(GeneScreen Plus). Size determination was done by using anRNA ladder (0.24, 1.4, 2.4, 4.4, 7.5, and 9.5 kb; GIBCO/BRL) as the standard. Hybridization and washing wereperformed as described above for the DNA hybridizationprocedure.DNA sequencing. DNA was sequenced by the dideoxy-
chain termination method (29) with [35S]dATP and a T7sequencing kit from Pharmacia LKB GmbH (Freiburg, Ger-many). Single-stranded templates were prepared from bothstrands of plasmids pK2 and pK42. Synthetic oligonucleo-tides (17-mers) complementary to the ends of already se-quenced templates were prepared by a Gene Assembler Plus(Pharmacia LKB) in accordance with the instructions of themanufacturer. The dideoxy-terminated fragments were sep-arated on 55-cm wedge-shaped thickness gradient gels (0.2 to0.4 mm; 6% [wt/vol] polyacrylamide) with a Macrophorsequencing unit (Pharmacia LKB) as recommended by themanufacturer.Primer extension analysis. Primer extension (PE) analysis
was done as recently described (8), except that SuperScriptreverse transcriptase (GIBCO/BRL) was used. Oligonucleo-tides ON112 (5'-GCAAAGCGAATTGAATC-3') and ON128(5'-ATTAAAACCTCCTAATATT-3'), which are comple-mentary to the 5' end of the primary transcript of hspl8,were radiolabeled at their 5' ends with [-y-32P]ATP by using
ORFB
tRNOORFAl___ tRNA cG ORFA1 ORFC ORFC2
hspl8
1000 200 31
Sau 3AI Hind m 4 Hind E Hind IIISau 3AM
Tn916
000 4000
*In I 1Hind III Hind III Sau 3AI
FIG. 1. Schematic representation of the cloned fragments inplasmids pK2 and pK42. Horizontally oriented arrows and arrow-heads show directions of transcription, positions, and lengths ofORFs. Numbers representing the nucleotide positions and restric-tion endonuclease sites are marked on the bar below. The verticalarrow indicates the insertion point of Tn916 in C. acetobutylicummutant AA2.
T4 polynucleotide kinase (GIBCO/BRL). The lengths ofprimer extension products were calculated by running asequencing reaction with the same primer on the same gel.
In vitro transcription-translation. 35S-labeled proteinswere obtained with a prokaryotic DNA-directed translationkit (Amersham Buchler), separated on a 15% (wt/vol) poly-acrylamide-SDS gel, and visualized by autoradiography.Protein size was determined by using 14C-labeled molecularmass markers (Amersham Buchler).Heat shock pulse-labeling. Pulse-labeling was done essen-
tially as described by Pich et al. (25), except that the cellswere grown to an optical density (600 nm) of 1 and subjectedto 10 min of heat shock. Aliquots were removed only beforeand after the shock. The products were analyzed on thesame gel as the in vitro transcription-translation reactions.Computer programs. The DNA region sequenced and the
deduced amino acid sequences were analyzed with the DNAStrider program (16) and GeneWorks (IntelliGenetics, Inc.,Mountainview, Calif.) on a Macintosh IIsi computer (AppleComputer, Inc., Cupertino, Calif.). Sequence comparisonswere done by using the University of Wisconsin GeneticsComputer Group sequence analysis software package, ver-sion 6.0 (University of Wisconsin Biotechnology Center,Madison) (7).
Nucleotide sequence accession number. The sequence datareported here (see Fig. 2) have been submitted to the EMBLdata base and assigned accession no. X65276.
RESULTSCloning and sequencing of the Tn916 insertion region of C.
acetobutylicum mutant AA2. As reported earlier (30), a PCR-generated DNA fragment from the insertion point of Tn916in C. acetobutylicum DSM 792 mutant AA2 was used as aprobe to screen a partially Sau3AI-digested gene bank of C.acetobutylicum DSM 792 DNA in pUC9 (8). Two identicalclones, pK2 and pK42, containing 1,645- and 2,427-bpSau3AI fragments as inserts (Fig. 1), were detected andsequenced. These two Sau3AI fragments are also originallycontiguous on the chromosome, as proved by Southernhybridization and PCR (data not shown).The 2,427-bp Sau3AI fragment encoded five open reading
frames (ORFs) with no significant homology to any sequencein the EMBL, GenBank, or Swissprot data base. All fiveORFs read in the same direction, and the positions of theirstart and stop codons in the 5'-to-3' direction are as follows:ORFA1, 1567 to 1857; ORFA, 1826 to 2377; ORFB, 2436 to
VOL. 175, 1993

3396 SAUER AND DURRE
1645 GATCG ATTTCTTCTT TAGATTGAAT ATAAACCTGA GCTTGAGATT 1601
TTAACTCTTC AGGTGTTACA GATATTTGAG CCATAATAAC TCCTCCTTAT ATATATCTTT GGTATTGTTT GCTACAGTTA AATACGAAAT TTAATACCAT 1501a TAAT AT A CAGTT
TCGTATTCAT TACCAAACTT TTCCCATTGC AGATTTATTA TATTACTCTC ATCTTCGACT GTCAATGGTT TATTTCAAGG GTTTTCAACA GTTTACATCT 1401
TATTGATAAT TATAACGAAA TAATTAATAT TGTCTTTTTG TGTAAATCCC TAAAAGTTGC TTTACGTATT GTATTATATA TAAATATATT AGGTAGAAAT 1301
TCAGTTACCT TTTACATTTT ATATTGGGGC TTTTCACCAC ATAAAAAAAT GCCAAAGCTC TAAAAGCTTT GGCATTAACT CTCTTtGTGC TCCAGATAGG 1201
ATTTGAACCT ACGACCAACT GATTACGAAT CAGCTGCGCT ACCAACTGCG CTACAGGAGC ATACTTACGA AATATATTTT ACTACATTAA TCCCTTTTAC 1101wTn916
ACAACTTTTT ATGAAAAAAA TTGAAGTTCT AACATTGTAA TTTTTATTTC TGATGAAATG CTACTAAAAC TTTTTATATT TTTTGAAAAA GTAAATTTAT 1001
CCCACAATCG CTTTGAATAC CTCCTAAAAT CTTAATATTT AAAATCCTTT TTGATTTTGA CTTTCTTTGA CTTTCGTACTLACAATACT TQTAAGGTGA 901
AGAGAAGAAA AACTTCAAAC ATTCAGTATT TTCTAAAAAT ATTTTGATTC AATTCGCTTT GCACCTTTAA TGTATACATT GAATAAAAAA TTAAAATAAG 801
CTAATAACTA AAAAATATTT AATATpGGA GitTTAATT ATGTTTGGAA TGGTTCCTTT TAGAAGAAAT AATAATGGTT TAATGAGAAG AGAGGATTTT 701MetPheGlyM etValProPh eArgArgAsn AsnAsnGlyL euMetArgAr gGluAspPhe
TTCGATAAGA TGTTTGATAA CTTTTTCTCA GATGACTTTT TCCCTACTAC TACATTCAAT GGAAATGCAG GCTTCAAAGT AGATATAAAA GAAGATGATGPheAspLysM etPheAspAs nPhePheSer AspAspPheP heProThrTh rThrPheAsn GlyAsnAlaG lyPheLysVa lAspIleLys GluAspAspA
ATAAATATAC AGTTGCAGCA GATTTACCTG GTGTTAAAAA AGATAACATT GAGCTTCAAT ACGAAAACAA CTATTTAACT ATAAATGCAA AAAGAGATGAspLysTyrTh rValAlaAla AspLeuProG lyValLysLy sAspAsnIle GluLeuGlnT yrGluAsnAs nTyrLeuThr IleAsnAlaL ysArgAspAs
601
501
CATTGTTGAG ACTAAAGATG ATAACAATAA TTTTGTAAGA CGTGAAAGAA GTTATGGAGA ACTTAGACGA AGCTTTTATG TAGATAATAT TGATGACTCA 401pIleValGlu ThrLysAspA spAsnAsnAs nPheValArg ArgGluArgS erTyrGlyGl uLeuArgArg SerPheTyrV alAspAsnIl eAspAspSer
AAAATTGATG CATCTTTTCT TGATGGAGTT CTTAGAATAA CTTTACCTAA GAAGGTTAAG GGAAAAGATA ATGGAAGAAG AATTGATATC CACTAAGTAALysIleAspA laSerPheLe uAspGlyVal LeuArgIleT hrLeuProLy sLysValLys GlyLysAspA snGlyArgAr gIleAspIle His
AAAAGTTATA CACTGTTGAT AACTTTCTTC CCCCTACTTA TCCACAGTAG TATGATTTTT ATATTAACAA TGTTAATAAC TATAAAATAT AAAAAACAAC
301
201
TTGTGTTATC CACAAGTTGT TTTTTATTAT TCTTGATGAA TTACATCTAA CCTACCAAAC TTACTGTACT GACCAAGCCA AGCAAGATTA ACAGTTCCTG 101
TAGGACCATT TCTCTGTTTT GCTATTATGC ATTCTGCAAC ATTCTTTTGT TCAGTTTCCT TATTGTAATA TTCATCTCTA TAAAGGAACA TAACAAGATC 1
FIG. 2. Nucleotide sequence of the 1,645-bp Sau3AI fragment of C. acetobutylicum. Only the antisense strand is shown. The deducedamino acid sequence of Hspl8 is shown below the DNA sequence, and the tRNA coding sequence, which is transcribed in the oppositedirection, is marked with an arrow above the DNA sequence. All features above the DNA sequence refer to the antisense strand of thedisplayed sequence. The putative ribosome-binding site is boxed. Arrowheads show regions of dyad symmetry, forming the stems of putativerho-independent transcription termination structures. The transcription start sites identified by primer extension analysis and the -10 and -35regions of the corresponding promoters are marked by bold letters above the sequence for ORFA1 and are thickly underlined for hspl8,respectively. The sequence duplication covering the -35 region of the hspl8 promoter is thinly underlined. The start codon for ORFA1, beingtranscribed from the complementary strand, is shown by three plus signs. The insertion point of Tn916 in mutant AA2 is marked by a verticalarrowhead.
2633; ORFC, 2620 to 3735; ORFC2, 3745 to 3978 (Fig. 1).The first 78 bp of ORFA1 are encoded on the 1,645-bpSau3AI fragment. With PCR-generated radiolabeled probescomplementary to ORFA, ORFB, and ORFC, RNA tran-scripts 1.3 and 1.7 kb long, plus an additional but muchweaker signal of 2.7 kb found only with the probe comple-mentary to ORFC, were detected on Northern blots ofclostridial RNA (data not shown). No differences in signalintensity or length were detected with RNA isolated duringthe shift to solventogenesis, indicating that these ORFs areexpressed but not regulated during the shift; thus, theirfunction remains unclear. A transcription start site wasmapped by PE with clostridial and E. coli clone pK2 RNAsto position 1471, 105 bp upstream of ORFAL. This site waspreceded by a typical gram-positive consensus promoter,5'-TTGACA(17 bp)TATAAT-3' (Fig. 2). Two putative rho-independent transcription termination structures were de-tected. The first is located between positions 1864 and 1888,8 bp downstream of the stop codon of ORFA1, and thesecond is between positions 3995 and 4026, 16 bp down-stream of ORFC2. Reasonable ribosome-binding sites werefound in front of ORFA1, ORFC, and ORFC2.
Figure 2 shows the DNA sequence obtained for the1,645-bp Sau3AI fragment that encodes an ORF and itsputative amino acid sequence. For clarity, the alreadypublished tRNAThG sequence (30), which is transcribed inthe opposite direction, is also shown. A putative ribosome-binding site with reasonable homology to the E. coli proto-type (31) and already sequenced C acetobutylicum genes (8,20, 21, 35, 38) was identified in front of the ORF. Anine-nucleotide palindrome followed by a poly(U) sequence,resembling a prokaryotic rho-independent transcriptional
terminator (26) (AG, -53.5 kJ/mol [33]), is located 100 bpdownstream from the stop codon (Fig. 2).
Identification of the hspl8 gene from C. acetobutylicum. Theamino acid sequence deduced from this ORF was alignedwith sequences available in the EMBL and GenBank databases and was identified and accordingly designated as thehspl8-homologous gene of C. acetobutylicum. Table 1shows the similarity of C. acetobutylicum protein Hspl8 tothe corresponding proteins of other organisms. Since thesmall HSPs exhibit much more diversity in size and aminoacid sequence than the high-molecular-weight HSPs (19),alignment of the amino acid sequences of these proteins wasrestricted to a conserved stretch of 36 amino acids in theC-terminal part (Fig. 3). A hydropathy profile of the aminoacid sequence deduced from hspl8 revealed the majorhydrophilic peak characteristic of the HSP20 family at the Cterminus (data not shown).mRNA analysis of hspl8. Total RNA was isolated from C.
acetobutylicum cells grown at 30°C and subjected to heatshock from 30 to 42°C for 10 min, from cells during the shiftto solventogenesis in continuous culture, and from therespective E. coli clone carrying the 1,645-bp Sau3AI frag-ment from C. acetobutylicum in pUC9. First, this RNA wasused for Northern blot analysis with a radiolabeled 404-bpPCR-generated fragment complementary to hspl8 as theprobe. A single signal, 700 to 800 nt long, was detected (Fig.4). Transcription was transiently induced during the shift tosolventogenesis, with maximum mRNA levels between 4and 9 h after the pH control was switched off. This is exactlythe period of time when the shift to solventogenesis takesplace at the mRNA level, as judged by induction of the adcand ctfgenes (8). It is also in good agreement with the results
J. BACTERIOL.

SMALL HEAT SHOCK GENE OF C. ACETOBUTYLICUM 3397
TABLE 1. Amino acid similarity of Hsp18 from C. acetobutylicum to corresponding proteins of other organisms
Organism Protein % Identity' % Homology' Reference(s)
Glycine max (soybean) Hsp 17.3 35.6 57.3 19Glycine max Hsp 18.5 35.4 58.8 23Pisum sativum (pea) Hsp 18.1 34.0 57.1 23Arabidopsis thaliana Hsp 17.6 33.6 56.2 23Arabidopsis thaliana Hsp 18.1 32.9 54.8 23Triticum aestivum (wheat) Hsp 16.9 30.6 52.1 23Glycine max Hsp 17.6 29.3 53.7 19Mycobactenum leprae Hsp 16.7 28.1 60.2 6, 22Saccharomyces cerevisiae Hsp 23.9 26.7 57.3 23Drosophila melanogaster Hsp 23.0 26.0 53.7 23Mycobacterium tuberculosis Hsp 16.3 25.9 51.8 34Escherichia coli IbpB 25.4 54.6 1Drosophila melanogaster Hsp 19.7 24.1 42.1 23Caenorhabditis elegans Hsp 16.3 21.1 51.9 23Escherichia coli IbpA 20.9 56.0 1Streptomyces albus Hsp 18 14.8 38.3 18Homo sapiens Hsp 22.3 14.2 46.1 23
a The values shown are similarity data derived from alignments of the complete amino acid sequence of Hspl8 from C. acetobutylicum (deduced from thenucleotide sequence) with those of low-molecular-weight HSPs of other organisms.
obtained by others with two-dimensional protein gels, show-ing transient induction of Hspl8 during the shift to solven-togenesis (25). Induction of transcription was also shown inheat-shocked cells (Fig. 4). The same RNA was subjected toprimer extension analysis to determine the exact start pointof the transcript (Fig. 5) by using oligonucleotides ON112and ON128, which are complementary to sequences up-
Consensus ..D ...DSI_
14.
Hsp18 Co R Y _ DS K F sF 14IbpA Ec AER4WQ A _ IHVR GANLV- DLER 14IbpB Ec MN EVS GATFV-- DLR 21Hspl8 Sa AAVAAL D KQV-GELIAE AMDKVG EESNHspl6.7 Ml P ALV-1LDIE Rh Y 31Hspl6.3 Mt A S -- EDD D 16Hsp18.1 Ps -RMD 15Hsp17.6 At FIX ACMEEISME-U 15Hsp18.1 At --(ME 17Hsp16.9 Ta 5VE 6LHspl7.3 Gm LD 15Hspl7.6 Gm -i Ev 15Hspl8.5 Gm --(VE M 15Hsp23.9 Sc IT YPGVDAD N Y 24Hspl6. 3 Ce RSFKMI LPEDVDLTS VKSN (4E 22Hsp197 Dm w "- K VS S LS 36Hsp23 lDn I WQVSQLS 45Hsp22.2 Hs YISRC KY TLPPGVDPTQ VSSSLS EA 30
FIG. 3. Comparison of the conserved C-terminal parts of severallow-molecular-weight HSPs. A consensus amino acid sequence,defined by identical positions in at least 50% of the respective HSPs,is shown at the top. Identical amino acids and conservative replace-ments have been placed in the same boxes. Conservative replace-ments are defined as being within the following groups: I, L, M, andV; D, E, N, and Q; H, K, and R; F, Y, and W; A and G; and S andT. The following six bacterial HSPs are shown at the top: Ca, C.acetobutylicum (this work); Ec, E. coli (1); Sa, Streptomyces albus(18); Ml, Mycobacterium leprae (6, 22); Mt, M. tuberculosis (34).These are followed by seven plant HSPs (Ps, Pisum sativum; At,Arabidopsis thaliana; Ta, Triticum aestivum; Gm, Glycine max[19]), one yeast HSP (Sc, Saccharomyces cerevisiae), one nematodeHSP (Ce, Caenorhabditis elegans), and three HSPs of higherorganisms (Dm, Drosophila melanogaster; Hs, Homo sapiens). Ifnot otherwise indicated, all sequences are from Nover and Scharf(23). The numbers on the right indicate distances from the Cterminus. For S. albus, the distance has not been determined.
stream of hspl8 (ON112, nucleotides 838 to 845; ON128,nucleotides 762 to 780; compare Fig. 2). A single start pointof transcription was mapped 149 bp upstream of the ATGstart codon and 176 bp downstream of the Tn916 insertion inAA2 (compare Fig. 2). The deduced promoter sequence wasidentical to the consensus promoter of gram-positive bacte-ria (10). The same result was obtained with E. coli clonespK2 and pK42 (data not shown). Since the putative clostrid-ial promoter is homologous to the o70-dependent E. coficonsensus promoter, it is not surprising that it is also used inthe respective E. coli clones. Even though the PE signal forheat-shocked C. acetobutylicum mutant AA2 is muchweaker than for the wild type, in Northern blot analysis thesame RNA revealed a signal as strong as that of the wild type
300C HS lh 2h Sh 6h 8h 9h 15h 27h
FIG. 4. Northern hybridization with the radioactively labeledPCR-generated fragment complementary to hspl8. The first twolanes each contained 3 ~Lg of RNA isolated from C. acetobulylicumDSM 792 before (30"C) and after heat~shock (HS). The followinglanes each contained 10 ptg of RNA isolated from C. acetobutylicumDSM 1731 at different time points during the shift to solventogenesisin continuous culture. Fragment size (nucleotides [nt]), as judged bycomparison with size markers, is indicated to the right.
VOL. 175, 1993

3398 SAUER AND DURRE
A B 1 X A 4 ; 6 7
GA T C 12
-35
-10
FIG. 5. Mapping of the 5' end of hspl8 mRNA by primerextension analysis. "Pradiolabeled oligonucleotides 0N112 (A)and 0N128 (B) were hybridized to 10 pg each of total mRNA
isolated from C. acetobutylicum. Lanes: 1, DSM 1731 3 h after the
pH control in continuous culture was shut off; 2, DSM 792 after heat
shock from 30 to 42'C; 3, DSM 792 after heat shock from 30 to 42C;
4, DSM 792 before heat shock from 30 to 420C; 5, DSM 1731 after
heat shock; 6, mutant AA2 after heat shock. The primer extension
products were analyzed on a sequencing gel. G, A, T, and C indicate
the products of sequencing reactions with the same oligonucleotidesas the primer. The sequence shown is the complementary strand.
The -10 and -35 regions are underlined and marked with bold
letters. The arrow points to the base representing the 5' end of the
mRNA, i.e., the transcription start point (5).
(data not shown), indicating that the weaker PE signal of
AA2 is due to the PE procedure rather than weaker induc-
tion. A sequence duplication of 10 nucleotides (TTTGACa'1TIC) was found to cover the -35 region of the putative
promoter, from positions 926 to 945 (Fig. 2).In vitro transcription-translation experiments. A protein
with the expected size of Hspl8 (17.7 kDa) was produced in
vitro in a coupled transcription-translation assay from plas-mid pK2 but not from the pUC9 control (Fig. 6). At least one
other protein, of approximately 10 to 12 kDa, which corre-
sponds to the predicted protein of ORFAl (11 kDa), was
produced. Pulse-labeled proteins of C. acetobulylicum DSM792 and P262 before and after heat shock were run on the
same gel, showing that a protein of the same size is inducedby heat shock, thus demonstrating at the protein level that
the cloned gene is the small HSP Hspl8 of C acetobutyli-cum. Interestingly, the second small HSP (Hspl4) of C
acetobutylicum P262 seems to be either slightly larger than
the one in strain DSM 792 or missing entirely (Fig. 6).
transciptiontransationystm S-labeled proteins were sepa-
rated on a 15% (wtlvol) polyacrylamide-SDS gel and subsequentlyautoradiographed. Lanes: 1, in vitro transcription-translation withcontrol plasmid pUC9; 2, in vitro transcription-translation with pK2,which encodes hspl8; 3, 'C-labeled marker proteins; 4 and 5,pulse-labeled proteins of strain P262 after and before heat shock,respectively; 6 and 7, pulse-labeled proteins of strain DSM 792 afterand before heat shock, respectively. The molecular masses ofmarker proteins are given to the right. The arrowheads point to the14- and 18-kDa HSPs of C. acetobutylicum.
Southern blot analysis ofhspl8 with DNAs from different C.acetobutylicum strains. Since there seem to be big differencesbetween the C acetobutylicum strains in use by differentresearch groups (36, 38), we wanted to test whether hspl8 isthe same in all strains. Therefore, we used restrictionendonuclease-digested DNAs from several C. acetobutyli-cum strains for Southern hybridization with the above-mentioned 404-bp PCR-generated fragment complementarytohspl8 as a specific probe at a hybridization temperature of600C (Fig. 7). The resulting bands for ATCC 824 and DSM792 correspond to the fragment size expected on the basis ofsequence analysis. With P262, much weaker signals wereobtained in both digests; however, these proved to beidentical in size to the respective bands of ATCC 824 andDSM 792. Strain NCIB 8052 gave only a weak signal of adifferent size in the Sau3AI digest and none at all in theHindlII-digested DNA, which might be due to a very smallfragment that had already migrated through the gel. For bothP262 and NCIB 8052, the weaker signals indicate somesequence variation with respect to the hspl8 gene clonedfrom C. acetobulylicum DSM 792.
DISCUSSIONThis report presents the DNA sequence and the first
mRNA analysis of a prokaryotic low-molecular-weight HSP.At the mRNA level, it was shown that hspl8 is induced byheat shock, as well as during the onset of solventogenesis,thus confirming earlier reports on transient induction by theshift to solventogenesis (25). A peak of new mRNA synthe-sis was observed between 4 and 9 h after the pH control was
J. BACTERIOL.
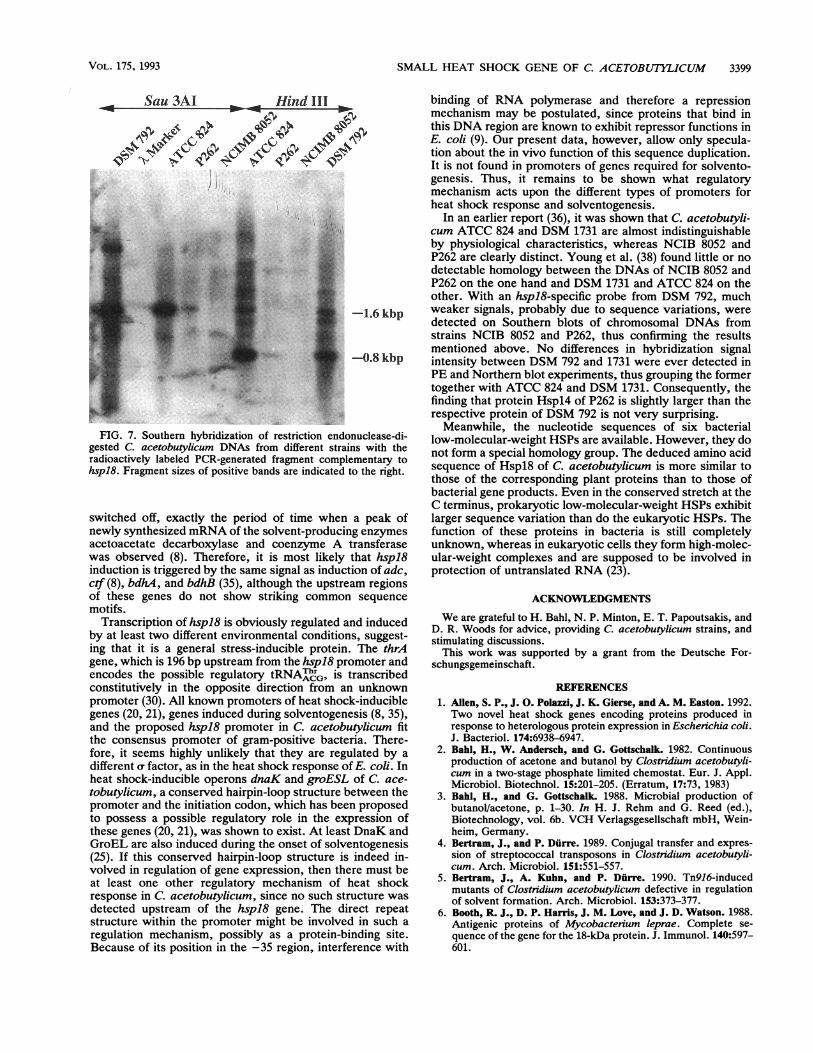
SMALL HEAT SHOCK GENE OF C. ACETOBUTYLICUM 3399
Sau 3AI HindII
0 -)409 s40 40 4 .8.t ,
E~~~~~~ F"PWE:FIG. 7. Southern hybridization of restriction ends
gested C. acetobutylicum DNAs from different strairadioactively labeled PCR-generated fragment complhspl8. Fragment sizes of positive bands are indicated
switched off, exactly the period of time whennewly synthesized mRNA of the solvent-produciacetoacetate decarboxylase and coenzyme Awas observed (8). Therefore, it is most likelyinduction is triggered by the same signal as inducctf (8), bdhA, and bdhB (35), although the upstreof these genes do not show striking commormotifs.
Transcription of hspl8 is obviously regulated a
by at least two different environmental conditioning that it is a general stress-inducible proteingene, which is 196 bp upstream from the hspl8 pr
encodes the possible regulatory tRNAThr is
constitutively in the opposite direction from a]promoter (30). All known promoters of heat shocgenes (20, 21), genes induced during solventogenand the proposed hspl8 promoter in C. acetobithe consensus promoter of gram-positive bactefore, it seems highly unlikely that they are regidifferent cr factor, as in the heat shock response oheat shock-inducible operons dnaK and groESLtobutylicum, a conserved hairpin-loop structure tpromoter and the initiation codon, which has beeto possess a possible regulatory role in the exthese genes (20, 21), was shown to exist. At leastGroEL are also induced during the onset of solv(25). If this conserved hairpin-loop structure isvolved in regulation of gene expression, then th4at least one other regulatory mechanism ofresponse in C. acetobutylicum, since no such stidetected upstream of the hspl8 gene. The distructure within the promoter might be involveregulation mechanism, possibly as a protein-bBecause of its position in the -35 region, interf
binding of RNA polymerase and therefore a repressionmechanism may be postulated, since proteins that bind inthis DNA region are known to exhibit repressor functions inE. coli (9). Our present data, however, allow only specula-tion about the in vivo function of this sequence duplication.It is not found in promoters of genes required for solvento-genesis. Thus, it remains to be shown what regulatorymechanism acts upon the different types of promoters forheat shock response and solventogenesis.
In an earlier report (36), it was shown that C. acetobutyli-cum ATCC 824 and DSM 1731 are almost indistinguishableby physiological characteristics, whereas NCIB 8052 andP262 are clearly distinct. Young et al. (38) found little or nodetectable homology between the DNAs of NCIB 8052 andP262 on the one hand and DSM 1731 and ATCC 824 on theother. With an hspl8-specific probe from DSM 792, much
-1.6 kbp weaker signals, probably due to sequence variations, weredetected on Southern blots of chromosomal DNAs fromstrains NCIB 8052 and P262, thus confirming the resultsmentioned above. No differences in hybridization signal
-0.8 kbp intensity between DSM 792 and 1731 were ever detected inPE and Northern blot experiments, thus grouping the formertogether with ATCC 824 and DSM 1731. Consequently, thefinding that protein Hspl4 of P262 is slightly larger than therespective protein of DSM 792 is not very surprising.Meanwhile, the nucleotide sequences of six bacterial
)nuclease-di- low-molecular-weight HSPs are available. However, they domnswith the not form a special homology group. The deduced amino acidto the right, sequence of Hspl8 of C. acetobutylicum is more similar to
those of the corresponding plant proteins than to those ofbacterial gene products. Even in the conserved stretch at theC terminus, prokaryotic low-molecular-weight HSPs exhibit
a peak of larger sequence variation than do the eukaryotic HSPs. Theng enzymes function of these proteins in bacteria is still completelytransferase unknown, whereas in eukaryotic cells they form high-molec-that hspl8 ular-weight complexes and are supposed to be involved intion of adc, protection of untranslated RNA (23).-am regionsn sequence ACKNOWLEDGMENTS
ind uced We are grateful to H. Bahl, N. P. Minton, E. T. Papoutsakis, andindsugedt- D. R. Woods for advice, providing C. acetobutylicum strains, and, suggest-rA stimulating discussions.The thrA This work was supported by a grant from the Deutsche For-
omoter and schungsgemeinschaft.transcribedn unknown REFERENCESk-inducible 1. Allen, S. P., J. 0. Polazzi, J. K. Gierse, and A. M. Easton. 1992.iesis (8, 35), Two novel heat shock genes encoding proteins produced inutylicum fit response to heterologous protein expression in Escherichia coli.ria. There- J. Bacteriol. 174:6938-6947.ulated by a 2. Bahl, H., W. Andersch, and G. GottschalL 1982. ContinuousfE. coli. In production of acetone and butanol by Clostridium acetobutyli-of C. ace- cum in a two-stage phosphate limited chemostat. Eur. J. Appl.,ete.n the- Microbiol. Biotechnol. 15:201-205. (Erratum, 17:73, 1983)
)etween the 3. Bahl, H., and G. GottschalL 1988. Microbial production ofXn proposed butanol/acetone, p. 1-30. In H. J. Rehm and G. Reed (ed.),pression of Biotechnology, vol. 6b. VCH Verlagsgesellschaft mbH, Wein-t DnaK and heim, Germany.entogenesis 4. Bertram, J., and P. Durre. 1989. Conjugal transfer and expres-indeed in- sion of streptococcal transposons in Clostridium acetobutyli-
ere must be cum. Arch. Microbiol. 151:551-557.heat shock 5. Bertram, J., A. Kuhn, and P. Dfirre. 1990. Tn916-inducedrucrea wasomutants of Clostridium acetobutylicum defective in regulationructure was of solvent formation. Arch. Microbiol. 153:373-377.irect repeat 6. Booth, R. J., D. P. Harris, J. M. Love, and J. D. Watson. 1988.d in such a Antigenic proteins of Mycobacterium leprae. Complete se-finding site. quence of the gene for the 18-kDa protein. J. Immunol. 140:597-erence with 601.
VOL. 175, 1993

3400 SAUER AND DURRE
7. Devereux, J., P. Haeberli, and 0. Smithies. 1984. A comprehen-sive set of sequence analysis programs for the VAX. NucleicAcids Res. 12:387-395.
8. Gerischer, U., and P. Durre. 1992. mRNA analysis of the adcgene region of Clostridium acetobutylicum. J. Bacteriol. 174:426-433.
9. Gralla, J. D. 1991. Transcriptional control-lessons from an E.coli promoter data base. Cell 66:415-418.
10. Graves, M. C., and J. C. Rabinowitz. 1986. In vivo and in vitrotranscription of the Clostridium pasteunanum ferredoxin gene.J. Biol. Chem. 261:11409-11415.
11. Jones, D. T., and D. R. Woods. 1986. Acetone-butanol fermen-tation revisited. Microbiol. Rev. 50:484-524.
12. Killeen, K. P., and D. R. Nelson. 1988. Acceleration of starva-tion and glycerol-induced myxospore formation by prior heatshock in Myxococcus xanthus. J. Bacteriol. 170:5200-5207.
13. Kurtz, S., J. Rossi, L. Petko, and S. Lindquist. 1986. An ancientdevelopmental induction: heat-shock proteins induced in sporu-lation and oogenesis. Science 231:1154-1157.
14. Long, S., D. T. Jones, and D. R. Woods. 1984. Initiation ofsolvent production, clostridial stage and endospore formation inClostridium acetobutylicum P262. Appl. Microbiol. Biotechnol.20:256-261.
15. Maniak, M., and W. Nellen. 1988. A developmentally regulatedmembrane protein gene in Dictyostelium discoideum is alsoinduced by heat shock and cold shock. Mol. Cell. Biol. 8:153-159.
16. Marck, C. 1988. 'DNA Strider': a 'C' program for the fastanalysis ofDNA and protein sequences on the Apple Macintoshfamily of computers. Nucleic Acids Res. 16:1829-1836.
17. Marmur, J. 1960. A procedure for the isolation of deoxyribo-nucleic acid from microorganisms. J. Mol. Biol. 3:208-218.
18. Mazodier, P., G. Guglielmi, J. Davies, and C. J. Thompson.1991. Characterization of the groEL-like genes in Streptomycesalbus. J. Bacteriol. 173:7382-7386.
19. Nagao, R. T., E. Czarnecka, W. B. Gurley, F. Schoffl, and J. L.Key. 1985. Genes for low-molecular-weight heat shock proteinsof soybeans: sequence analysis of a multigene family. Mol. Cell.Biol. 5:3417-3428.
20. Narberhaus, F., and H. Bahl. 1992. Cloning, sequencing, andmolecular analysis of the groESL operon of Clostridium aceto-butylicum. J. Bacteriol. 174:3282-3289.
21. Narberhaus, F., K. Giebeler, and H. Bahl. 1992. Molecularcharacterization of the dnaK gene region of Clostridium aceto-butylicum, including grpE, dnaJ, and a new heat shock gene. J.Bacteriol. 174:3290-3299.
22. Nerland, A. H., A. S. Mustafa, D. Sweetser, T. Godal, and R. A.Young. 1988. A protein antigen of Mycobacterium leprae isrelated to a family of small heat shock proteins. J. Bacteriol.170:5919-5921.
23. Nover, L., and K.-D. Scharf. 1991. Heat shock proteins, p.41-128. In L. Nover (ed.), Heat shock response. CRC Press,
Inc., Boca Raton, Fla.24. Oelmuller, U., N. Krfiger, A. Steinbuchel, and C. Friedrich.
1990. Isolation of prokaryotic RNA and detection of specificmRNA with biotinylated probes. J. Microbiol. Methods 11:73-81.
25. Pich, A., F. Narberhaus, and H. Bahl. 1990. Induction of heatshock proteins during solvent formation in Clostridium aceto-butylicum. Appl. Microbiol. Biotechnol. 33:697-704.
26. Platt, T. 1986. Transcription termination and the regulation ofgene expression. Annu. Rev. Biochem. 55:339-372.
27. Rogers, P. 1986. Genetics and biochemistry of Clostridiumrelevant to development of fermentation processes. Adv. Appl.Microbiol. 31:1-60.
28. Sambrook, J., E. F. Fritsch, and T. Maniatis. 1989. Molecularcloning: a laboratory manual, 2nd ed. Cold Spring HarborLaboratory, Cold Spring Harbor, N.Y.
29. Sanger, F., S. Nicklen, and A. R. Coulson. 1977. DNA sequenc-ing with chain-terminating inhibitors. Proc. Natl. Acad. Sci.USA 74:5463-5467.
30. Sauer, U., and P. Durre. 1992. Possible function of tRNAhcr inregulation of solvent formation in Clostidium acetobutylicum.FEMS Microbiol. Lett. 100:147-154.
31. Shine, J., and L. Dalgarno. 1974. The 3'-terminal sequence ofEscherichia coli 16S ribosomal RNA: complementarity to non-sense triplets and ribosome binding sites. Proc. Natl. Acad. Sci.USA 71:1342-1346.
32. Terracciano, J. S., E. Rapaport, and E. R. Kashket. 1988.Stress- and growth phase-associated proteins of Clostridiumacetobutylicum. Appl. Environ. Microbiol. 54:1989-1995.
33. Tinoco, J. R., P. N. Borer, B. Dengler, M. D. Levine, 0. C.Uhlenbeck, D. M. Crothers, and J. Gralla. 1973. Improvedestimation of secondary structure in ribonucleic acid. Nature(London) New Biol. 346:40-41.
34. Verbon, A., R. A. Hartskeerl, A. Schuitema, A. H. J. Kolk, D. B.Young, and R. Lathigra. 1992. The 14,000-molecular-weightantigen of Mycobactercum tuberculosis is related to the alpha-crystallin family of low-molecular-weight heat shock proteins. J.Bacteriol. 174:1352-1359.
35. Walter, K. A., G. B. Bennett, and E. T. Papoutsakis. 1992.Molecular characterization of two Clostridium acetobutylicumATCC 824 butanol dehydrogenase isozyme genes. J. Bacteriol.174:7149-7158.
36. Woolley, R. C., and J. G. Morris. 1990. Stability of solventproduction by Clostridium acetobutylicum in continuous cul-ture: strain differences. J. Appl. Bacteriol. 69:718-728.
37. Yanisch-Perron, C., J. Vieira, and J. Messing. 1985. ImprovedM13 phage cloning vectors and host strains: nucleotide se-quences of the M13mpl8 and pUC19 vectors. Gene 33:103-119.
38. Young, M., N. P. Minton, and W. L. Staudenbauer. 1989.Recent advances in the genetics of the clostridia. FEMS Micro-biol. Rev. 63:301-326.
J. BACTERIOL.