sexual maturity and senescence of the male california...
TRANSCRIPT

SEXUAL MATURITY AND SENESCENCE OF THE MALE CALIFORNIA SEA OTTER
(ENHYDRA LUTRIS)
A Thesis
Presented to
the Faculty of the Department of Biological Sciences
San Jose State Univerity
In Partial Fulfillment
of the Requirements for the Degree
~laster of Arts
by
Bonnie Doris Green
r~ay 1978

I dedicate this thesis to my
father, Arthur Green, for his
help along the way and for the
magnificent photography and
printing of the photos herein.

ACKNOWLEDGMENTS
I am deeply grateful for G. Victor Morejohn, my Committee Chairman,
who suggested this interesting and important topic. His support and con
fidence along the way. were greatly appreciated, as was his critical analy
sis in the end. I also owe thanks to Howard Shellhammer and Henry Weston,
my other committee members, for their input and encouragement.
A special thanks is owed to Henry 11urphy, who confirmed my histo
logical interpretations and supplied me with adequate background in order
to make those determinations. Thanks are also due to Geology Department
members Calvin Stevens, who introduced me to the petrographic grinding
technique, and Bil1 Raub, who was ah1ays eager to share his time and
ski1ls with me.
I am indebted to King Wah Moberg and Loren Molliere at the San Jose
State University library and Doris Baron at the Moss Landing library for
their unselfish dedication to their jobs and to me as a student. With
out their help, I would never have been able to accomplish the exhaus
tive literature search.
Peggy Woodin and her associates at San Jose State Biology service
room provided me VJith chemicals, equipment and books Vlithout which the
research could never have been completed.
For assistance in obtaining specimens, I am especially grateful to
Jack Ames of the California Department of Fish and Game. Also helpful
v1ere David Huckaby of Long Beach State University, Dr. Cornell of Sea
World San Diego, and Chuck Woodhouse of the Santa Barbara County museum.

iv
I am indebted to Karl Schneider of the Alaska Department of Fish
and Game for his interpretations, counseling and correspondence. David
Sergeant of the Arctic Biological Station was also invaluable for his
correspondence, suggestions and attempts at processing my sea otter
teeth.
Special thanks are o~1ed to Lynn McMasters for her excellent draft
ing job, Signe Johnsen for aiding me in microtechniques and photomicro
scopy and to Rosie Keegan for typing the thesis.
Finally, I wish to thank the professors, staff and my fellow students
at Moss Landing Marine Laboratories for sharing their experience, their
knowledge and themselves with me and especially Bruce Ross, my fiance',
whose understanding, love and support carried me through this past year.

TABLE OF CONTENTS
Page
Ackno11l edgements. iii
List of Tables. . vii
List of Figures ....•.......•............ viii
List of Plates ....•..................... ix
CHAPTER
l INTRODUCTION. 1
2 MATERIALS AND METHODS •••....••.•.•.•••.•. 7
Skeletal Materials. . . . . • . . . . • . . • . . • . • • • 7 Teeth - Decalcification ..........•.•...... 11 Teeth - Grinding. . . . . . 13 Teeth - Staining. . . . . . . . . . . . . . . . . . • . . . 15 Teeth- Reading the Annuli .......•..•...... 16 Reproductive Organs ..................... 17 Fertility Measurements. . . . . . . . . . . . . . . . . . . 21 Photography. 23
3 RESULTS .........••................ 24
4
Pups. . . . . . • • . . . • . . . . . . . . . . . . . . . . 24 Immatures. . . • . • • . . . . . . • • . . . • • . • . . . . 32 Subadults ..•..•.......•.....••.... : 34 Adults. . . . . . • . . . . . . . . . . . . . • . • . . . . 37 Aged Adults. • • . • . . . • . . . . . 38 Measurements. . . . . . . 40 Distribution. 41
DISCUSS ION. . 44
Reproductive Aspects. 44 Teeth and Tooth Studies ................... 54
5 CONCLUSION. . . . . . . . . . . . . . . . . . . . . . . . . 60
Literature Cited.
Appendices. . . .
61
66

Appendix A.
Appendix B.
Sample Data Sheet. . ..
Frequency Distributions.
Appendix C. Plates of Sea Otter Skeletal Material and Gonadal
Page
67
68
Tissues. . . . . . . . . . . . . . . . . . . . . . 73

LIST OF TABLES
Table Page
A tabulation of the cementum layer counts obtained using different teeth and different methods of observation. 18
2 A summary of data collected from study specimens. . . 29
3 Pup ages based upon tooth eruption sequences proposed by Kenyon (1969) and Lensink (1962). . . . . . . . . . . 30
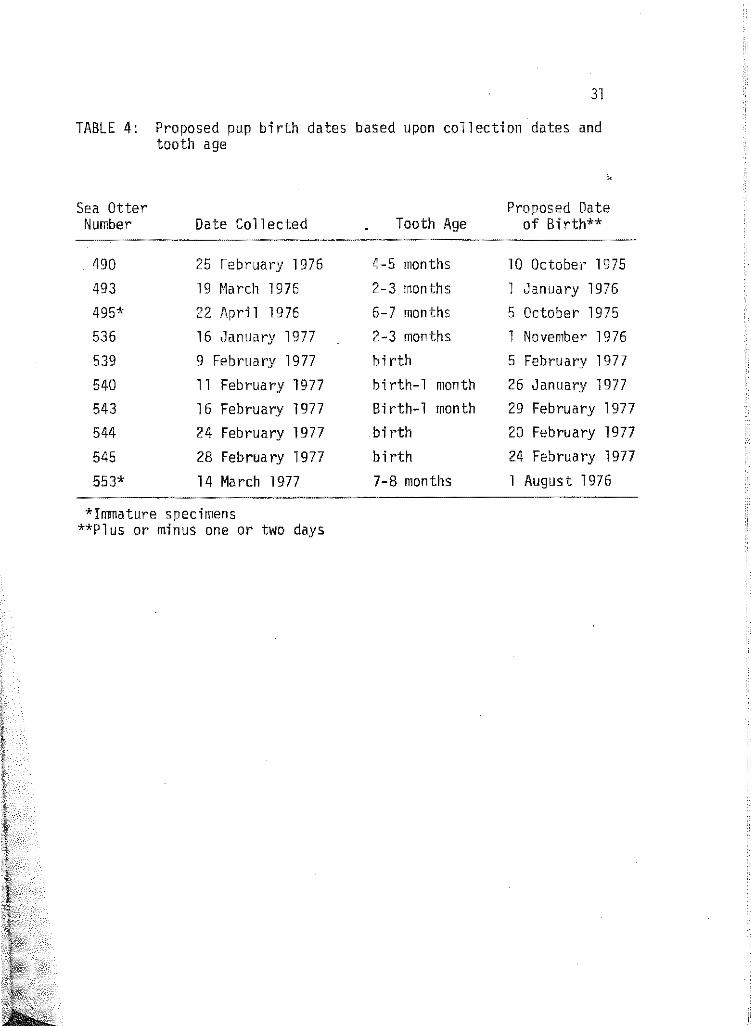
4 Proposed pup birth dates based upon collection dates and teeth age. . . . . . . . . . . . . . . . 31
5 A comparison between tl~o immature specimens. 33
6 Summary of selected reproductive cycles of male Mustelids (after Wright, 1969). . . . . . . . . . . . . . . . . . . . 45
vii

LIST OF FIGURES
Figure Page
1 Distribution of sea ottet" carcasses along the central California coast. (p:pup; i mmature; s:subadult; a=adult; o:aged adult). . . . . . . . . .. . . . . . . . . . . . . . 8
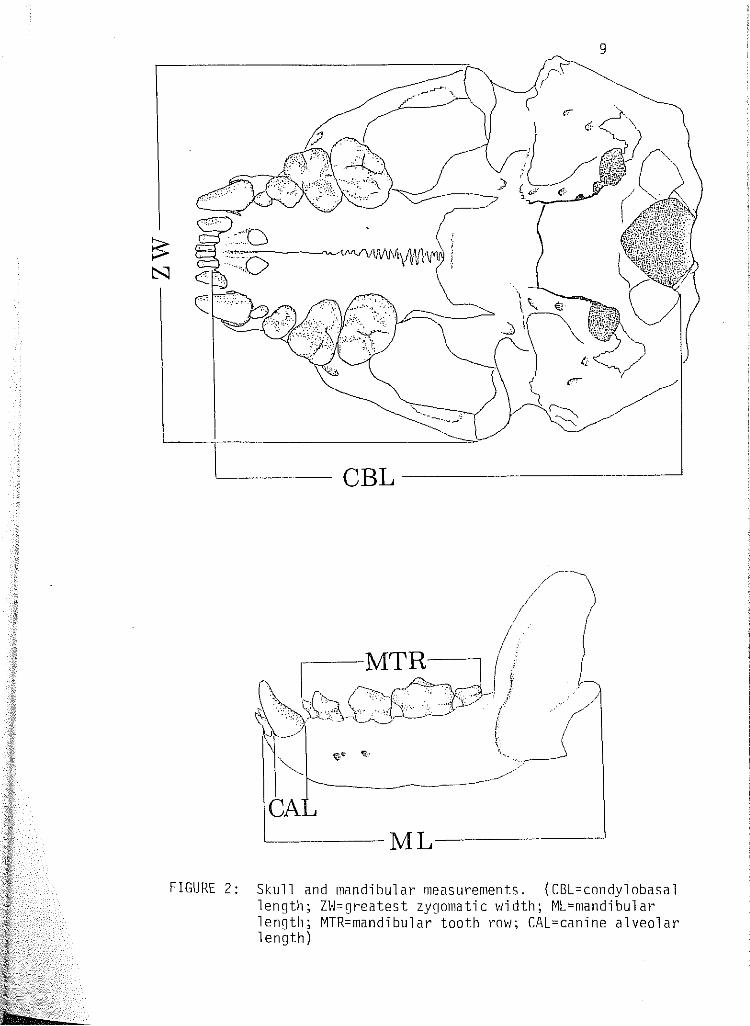
2 Skull and mandibular measurements. (CBL=condylobasal length; ZVJ:greatest zygomatic width; ML:mandibular length; MTR:mandibular tooth row; CAL=canine alveolar length). . . 9
3 Diagram of a medial longi-section of a sea otter upper right canine (a) showing hard tissue morphology, and diagram of a sea otter upper left canine (b). (A:greatest tooth width; B:root length; C:greatest cementum width). . . . . . . . . 10
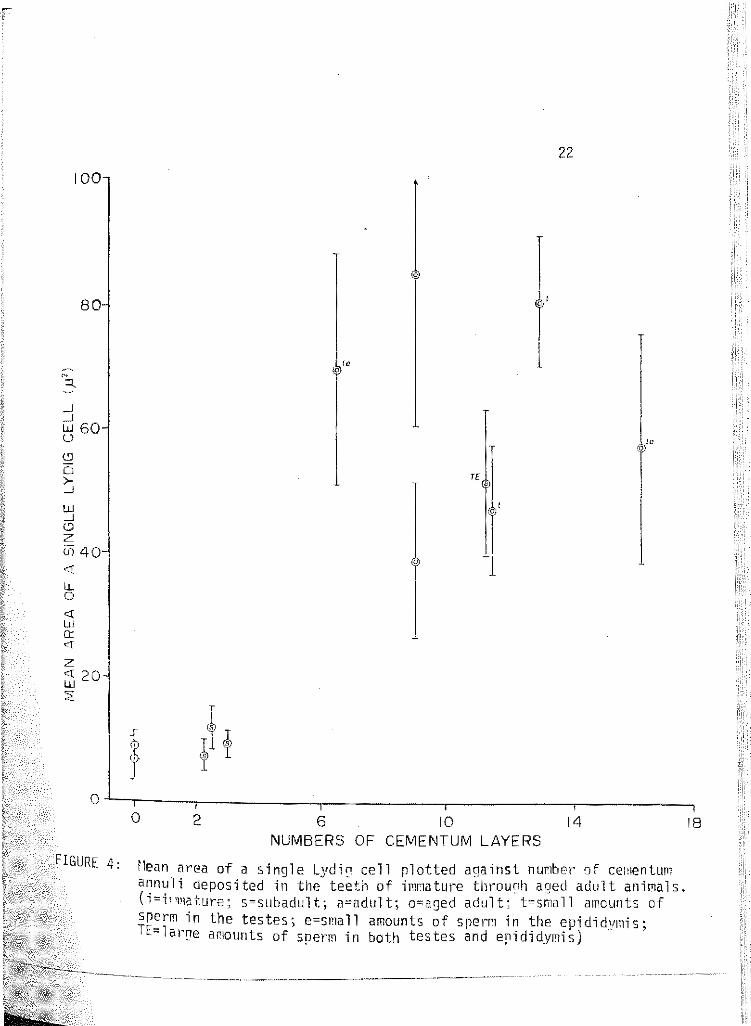
4 f•lean area of a single Lydig cell plotted against number of cementum annuli deposited in the teeth of immature through aged adult animals. (!=immature; s=subadult; a=adult; o= aged adult; t=sma 11 amounts of sperm in the testes; e=sma 11 amounts of sperm in the epididymis; TE=large amounts of sperm in both the testis and epididymis). . . . . . . . . . 22
5 Root canal opening in mm of deciduous and permanent canine teeth plotted against mandibular length ........... 25
6 Diameter of the root canal opening plotted against length of the growing permanent canine tooth. . . . . . .... 26
7 Diameter of root canal opening plotted against age in months. Line drawn between immature sea otters 495 and 553 represents a theoretical rate of root canal closure (3 mm/ 2 mo.). Sea otter. 527, a known-age female, was plotted to test for accuracy. . . . . . . . . . . . . . . . . . 27
8 Number of cementum annuli plotted against mandibular length (nrrn). • . . • • . . • • . • . • • • . • • . • . . • • • • . 42
9 Baculum weight (a) and numbet· of cementum layers (b) plot-ted against baculum length. . . . . . . . . . . . . . . . . 49
vi i i

LIST OF PLATES
Plate
1 Maxillaries and dentary of 539 (above). Maxillaries and dentary of 544 (below). Both animals are ''birth" aged. Top of photograph is anterior. . . . . . ....
2 Maxillaries and dentary of 545 (above) and 540 (below).
3
Sea otter 545 is "birth" aged, 540 is aged birth to 1 month. Top of photograph is anterior ............... .
Maxillaries and dentary of 543, aged ~1axillaries and dentary of 493, aged Top of photograph is anterior ....
birth to 1 month (above). 2 to 3 months (below).
4 Maxillaries and dentary of 536, a 2 to 3 month old (above) 490, a 4 to 5 month old (below). Top of photograph is anter-i or. . . . . . . .
5 Dorsal (above) and ventral (below) views of the skull, dentary and baculum of 495, an immature animal. . . . ..... .
6 Dorsal (above) and ventral (below) views of the skull, dentary and baculum of 553, an immature animal. . . . .....
7 Dorsal (above) and ventral (below) views of the skull, dentary and baculum of 593, a subadult animal ........ .
8 Longi-section of root apex, upper right canine of 593, a sub-adult. Note the 3+ translucent zones ........ .
9 Dorsal (above) and ventral (below) vie~1s of the skull, dentary and baculum of 529, a subadult animal ........ .
10 Longi-section of root apex, upper right canine of 529, a subadult. Note 3- 3-l/2 translucent zones and multiple lining indicated by split arrows .............. .
11 Dorsal (above) and ventral (below) views of the skull, dentary and baculum of 497, a subadult animal ........... .
12 Longi-section of root apex, upper right third incisor (above) of 497, a subadult. Longi-section of root apex, upper right can1ne (below) of the same animal. Note 3-1/4 translucent zones in both photographs ................. .
ix

X
Plate Page
13 Dorsal (above) and ventral (belm1) views of the skull, dentary and baculum of 592, a subadult animal .....
14 Longi-section of root apex, upper right canine of 592, a subadult. Note 3+ translucent zones ......... .
15 Dorsal (above) and ventral (below) views of the skull, dentary and baculum of 520, a subadult animal .....
16 Longi-section of root apex, upper right canine of 520, a subadult. Note how difficult it is to read the annuli.
17 Dorsal (above) and ventral (below) views of the skull, dentary and baculum of 516, a subadult animal .....
18 Longi-section of root apex, upper right late subadult. Note that 3 annuli can apex, 5 as you move up the root neck ..
canine of 538, a be counted near the
19 Dorsal (above) and ventral (belo~l) vie1·1s of the skull, dentary and baculum of 538, a late subadult ..... .
20 Dorsal (above) and ventral (below) views of the skull, dentary and baculum of 550, an adult animal ..... .
21 Longi-section of root apex, upper right canine of 550, an adult. 11-l/2 annuli are visible .......... .
22 Dorsal (above) and ventral (below) views of the skull, dentary and baculum of 590, an aged adult ...... .
23 Dorsal (above) and ventral (below) views of the skull, dentary and baculum of 521, an aged adult. Note the extreme sagittal and lambdoidal crest development and tooth wear.
24 Dorsal (above) and ventral (below) views of the skull, dentary and baculum of 509, an aged adult. Note necrosis of l01•1er left canine alveolus ............... .
25 Dorsal (above) and ventral (bel011) views of the skull, dentary and baculum of 528, an aged adult. Note dense deposition of cementum up the root neck of extracted upper can1ne (lower photograph) ..............•..

xi
Plate Page
26 Dorsal (above) and ventral (below) vie~1s of the skull fragments, dentary and baculum of 535, an aged adult. Note tooth wear and caries, several of which extend into the root cana 1 s. . . . . . . . . . . . . . . . . . . . . .
27 Longi-section of root apex, upper right canine of 535, an aged adult. 10-1/2 annuli were counted ....... .
28 Dorsal (above) and ventral {belovl) views of the skull, dentary and baculum of 515, an aged adult. Note upper left molar alveolus is filled in with alveolar bone. Canines are worn to gum line. . ............ .
29 Longi-section of root apex, left upper third incisor of 515, an aged adult. 16 translucent zones are echinochrome-stained ....................... .
30 Dorsal {above) and ventral (below) vievts of the skull, dentary and baculum of 577, an aged adult. Note the ex-treme tooth ~1ear. . . . . . . . . . . .
31 Longi-section of root apex, upper right canine of 577, an aged adult. There are 13 to 16 annuli, depending upon where count is made. . . . . ........ .
32 Bacula from pups, immature and subadult sea otters (from bottom to top). . . . . . . . . . . . . . . . . . . . .
33 Bacula from one subadult {538, bottom), one adult {550, top) and seven aged adults. Note rugosity of proximal ends (left) ...............•... · · · ·
34 Cross-sectional view of the testis of pup 543, showing septuli testis (ST) forming a thin partition which divides the testis into lobules. H&E stain, x 125. (sc=Sertoli cell; sg=spermatogonia) ................ .
35 Cross-sectional view of the testis of immature sea otter 553 showing a portion of the tunica albuginea (ta) overlying seminiferous tubules. H&E stain, xl25. (sg=spermatogon i a; ps=primary spermatocyte; ss=secondary sperma tacyte; l=Lydig cell) ................. · · · · · ·
36 C~ose-up view (x500) of basement membrane (BH) of a seminlfet·ous tubule and Lydig cells (L) of sea otter 529, a subadult. K&E stain. (sg=spermatogonia; sc=Sertoli cells; ss= secondary s r t t · · · ) pe ma ocy e 1 n mews 1 s . . . . . . . . . . . . .

xi i
Plate Page
37 Adult testis of sea otter 550 showing basement membrane (bm) separating seminiferous tubule and Lydig cells (L). Note large number of spermatogenic cells in various stages of spermatogenesis and a Sertoli cell (sc) cytoplasmic exten-sion extending toward the lumen (Lu). H&E stain, x550. (sg= spermatogonia; ps=primary spermatocytes; ss=secondary spermatocytes; s=maturing sperm aligned to Sertoli cell) .....
38 Cross-section of l-l/2 epididymal canals \•Jithin epididymis of 550, an adult sea otter, showing a high concentration of sperm (s) in the canals and cilia (c) of the pseudostratified steriocilia cells extending into the canal to move the sperm along. H&E stain, xl25 ............. .
39 Cross-section of aged testis of 494 showing high numbers of Sertoli cells (sc) and occasional spermatogonia (sg). Spermatogenesis has ceased. H&E stain, xl25 ....... .
40 Close-up (x500) of an aged adult, 509, seminiferous tubule in cross-section. Note lack of organization to reproductive cells and sporadic occurrence of spermatids (sd). H&E stain ........................ .
41 Section of testis of aged adult, 521, showing ''Lydig cell complex" (LSC) and collapsed seminiferous tubules (st). H&E stain, x125 ..................... .

ABSTRACT
Unstained, undecalcified 25-60 u sections of the upper right
canine teeth of 27 pup through aged adult sea otters were examined. As
many as 16 cementum annuli were counted, but no teeth were found with
fewer than 2-1/2 annuli. Pups and irnmatures with open root canals
had no cementum. Puberty comrnenses at 4-1/2-5 years and senescence
may occur as early as 7-1/2 years. Bacular growth parallels that of
the other rnustelids and may be used to calculate the onset of puberty
in individual specimens.

CHAPTER l
INTRODUCTION
An essential requirement in the study of vertebrate populations is
a reliable method of age determinat~on. This is particularly true with
animal populations of economic and social import, such as the sea otter
(Enhydra lutris L.). So important was it in early California history
that it very nearly became extinct in the early twentieth century from
over-exploitation by man (Kenyon, 1969).
Now sea otter populations are on the rise. It might even be con
sidered crowded throughout some portions of its range (Lensink, 1962;
Kenyon, 1969; Peterson and Odemar, 1969). This population expansion
with its concomitant decrease in marine invertebrate fauna has resulted
in the sea otter becoming a political issue as well as an economic one.
Commercial and sport fishermen seeking urchins, abalone, Pismo clams,
and crabs are vying with the sea otter for some of its food. But the
sea otter is "relatively" safe, since it has been afforded complete pro-
tection since 1911 (Miller, 1974).
Because of the "hands off" policy, the California Department of
Fish and Game and the federal government maintain, very little is known
of the male sea otter's reproductive biology, particularly areas relating
to life span, age at sexual maturity, how long males remain reproductively
active seasonally or in their lifetimes, and age at which sexual potency
begins to decline (Morejohn, Ames and Lewis, 1975).
The Alaskan and California populations of the sea otter (Enhydra
lutris) are considered geographical clines of the same species and not

2
two subspecies (Roest, 1971). Therefore, any reference I make to ani
mals in my sample will be prefaced with California or southern. Any
sea otters coming from locations outs ide of California will be appro
priately labeled.
Several researchers have attempted to solve the sea otter aging
problem in order to resolve the mysteries surrounding its reproductive
biology. All efforts to date have been centered around tooth and skull
morphology and the various changes which take place ~lith growth.
The first lengthy study on the northern sea otter was conducted by
Barabash-Nikoforov (1947). It included most aspects of the natural his
tory of the animal, but only briefly touched on aging. His method was
one of assigning lengths and weights to the various age groups. One
limitation with this method ~las that some animals were skinned first
and then measured. The second problem lies in the sample sizes he used
for each age group. Only two of the eleven groupings had statistically
adequate sample sizes. Finally, the ~1eights and lengths were based upon
hunters' determinations and not on his own actual measurements.
The use of teeth to determine age in marine mammals began approxi
mately twenty-five years ago with studies by Laws (1952) and Scheffer
(i950a) on pinnipeds.
Lensink's (1962) study on the northern sea otter was far more ex
haustive than Barabash-Nikoforov's (1947). His field observations, how
ever, did not provide the precise information on sex and age composition
essential for a total evaluation of reproductive biology. Polymodal age

3
distributions as exist for most seasonal species (Harding, 1949; Cassie,
1950) were lacking because births may occur throughout the year. Be
cause of this, lensink assigned three broad age groupings: pups, sub
adults and adults.
He defined pups as young, dependent u~on adult females. The sub
adult animals were defined as no longer being in need of parental care,
but were not capable of reproduction. The adults were sexually mature
individuals. Pups and subadults were further divided into groups I to
IX on the basis of skeletal and dental development. lensink stated
that, "since the intervals covered by each age class were based on in
tuitive reasoning from field observations, they were subject to error".
Lensink (1962) also discovered that he could distinguish adult
males from immature males by the weight of their bacula. Among imma
ture animals, however, overlap of bacula weights between age classes
precluded their use as an age criterion. Morejohn, Ames and lewis (1975)
studying the southern sea otter, came up with similar results.
In 1966 Klevezal and Marakov did a preliminary study on the life
span of the sea otters around Copper Island. They reported an age limit
of twelve years based on their study of three carcasses.
Kenyon (1969) was also engaged in a thorough study of the northern
sea otter. His information on aging, however, was very limited. like
Lensink (1962}, he proposed th1·ee age groupings: nev1born, juvenile and
adult.
Again like Lensink (1962), his age classification was based upon

4
tooth eruption. The major flaw is that it provided very little infor
mation relating to absolute age. For instance, his juvenile class
covered almost a two and a half year period.
Schneider was engaged (1973) in a project, one aspect of which was
to develop a technique for aging the Alaskan sea otter. As with Lensink
(1962) and Kenyon (1969) before him, Schneider used teeth, but he re
moved them for study. After removal, teeth were decalcified, sectioned
and stained.
Prior to the removal of teeth, the stage of eruption of each tooth
in all skulls was recorded. When possible, each animal was assigned to
one of Lensink's (1962) nine age classes. Pups that had no annuli were
subjectively classified into one of five "cementum age" categories.
Tooth sections appeared layered when viewed through a microscope
with transmitted light. These layers appear as fine, dark bands separ
ated by broad, light bands. The dark lines, laid down in the winter,
represent periods of low cementum deposition. The first line is laid
down during the pup's first winter. Animals younger than eight months
old during their first winter would not form a dark line for another
year. Therefore, it is possible that an animal is anywhere from eight
to nineteen months old when the first line forms. Here lies a major
source of error. Schneider (1973) feels that if Lensink's (1962) age
classes could be assigned to a relatively accurate age sequence, it
vmuld be possible to use his system to age infants and subadults and
cementum layer counts to age adults.

5
The most recent research done on sea otter aging was performed by
Morejohn, Ames and Lewis in 1975. They developed a relative age guide
based upon skull morphology and tooth eruption using a nearly complete
grov;th series of skulls from fetuses .tht·ough aged adults. These re
searchers took Lensink's (1962) "pup" category and divided it into pup
and immature age classes vlith overlap of the immature age class into
Lensink's "subadult" category. They also added an aged adult class.
Morejohn, et ~.{1975) added new aging criteria to the techniques
already established by previous researchers. These new techniques in
cluded estimating the degree of interdigitation of the sutures between
the palatine bones and the degree of development of the posterior lips
of the glenoid fossae. Furthermore, they found that the pelves could
also be used for aging (and sexing) sea otters. With the above criteria
at hand, they described a progression of osseous changes which take
place from newborn through the various relative age classes to the aged
adult animal.
The one vital element missing in all of the early research endeav
ors was an attempt to correlate relative aging methods with chronologi
cal age. This in turn should be correlated with ontogenetic stages of
the reproductive system so that management issues relative to sea otter
reproduction can be carried out realistically.
Since previous workers have already reported on osseous and mor
phometric changes which take place as the sea otter grows and matures
into adulthood, this paper will address itself to histologic changes

6
taking place in the testes from maturation to senescence and relate
them to chronological age ascertained from cementum annuli. Senes
cense, as used in this paper, refers to infertility accompanying old
age and does not necessarily imply degeneration or incapacitation.
The existence of cementum annuli and their accurate and effective
use as an indicator of chronological age has been verified for such
diverse mammalian orders as Chiroptera (Christian, 1956), Carnivora
(Crowe, 1972; Crowe and Strickland, 1974; Craighead, Craighead and r~c
Cutchen, 1970; Grau, Sanderson and Rogers, 1970; Klevezal and Marakov,
1966; Linhart and Knowlton, 1967; Marks and Erickson, 1966; Mundy and
Fuller, 1964; Sauer, Free and BrO\·me, 1966; Schneider, 1973; Stoneberg
and Jonke1, 1975; and Wright, 1969), Pinnipedia (Big, 1969; Bishop,
1967; Mansfield and Fisher, 1960) and Artiodactyla (Gilbert, 1966; Low
and Cowan, 1963; McEv1an, 1963; Sergeant and Pimlot, 1959).
In addition to the single species studies listed above under or
ders, several researchers have done multi-species and multi-order
studies on aging with cementum annuli. Klevezal and Kleinenberg (1967)
published an extensive work on age determination from annual layers in
the teeth and bones of all the mammalian orders found within the U.S.S.R.
Sergeant (1967) reviewed aging methods on terrestrial and marine mammals
up to 1967. Finally, Stone, Clauson, Slingerlands and Weber (197~) dis
cussed effective sectioning and staining methods used on red and gray
fox, coyote, black bear, fisher, marten, raccoon, bobcat and the white
tailed deer.

CHAPTER 2
~1ATERIALS AND METHODS
All specimens used for this study v1ere acquired through the Cali-
fornia Department of Fish and Game. ·\4ith the exception of one or two
moribund animals, all vlere found dead on beaches along centt·al Califor
nia. (See Figure l for coastal distribution of carcasses.)
Freshly dead otters v1ere vleighed to the nearest kilogram, total
length taken and the head and baculum removed. Gonads were also removed,
and testes sliced approximately every 5 mm to allow for optimum fixative
penetration in Bouin's fluid. Testes weights and lengths were not taken.
Deteriorating corpses were vleighed and measured only 1·1hen feasible;
the head and baculum were removed from such animals.
Skeletal Materials
Skulls and bacula were mascerated in water for at least a month or
until the flesh fell easily away from the bone. They were then scrubbed
in running water and allowed to air day.
The skeletal material was numbered and the follovling measurements,
outlined by DeBiase and Martin (1974), taken with a helios caliper (see
Figure 2).
Baculum weight (Hettler Tl60m analytical and torsion balance) Greatest bacul urn length Condylobasal length (CBL) Mandibular length (ML) Greatest zygomatic width (ZvJ} Can~ne alveolar length, anterior-posterior (CAL) ~1ax1llary tooth rov1 length (post canines and molars) (MTR)
follm·ling canine tooth measurements were also taken (see Figure 3).
7

PT PINOS 0
:PIGEON PT . 122(1
. PT. ANO NUEVO o·
.. SANTA . . , .· CRUZ - - · 0:
·"'~· · ··fe S-A S . _ -1!. S,S,S. <t'll
MONTEREY s~ BAY (,_
)~; ·.· "\t
.;P«J
)PT SUR
'\.- ~PFE:JFFER PT
0
~ . P p' ~WRECK BEACH
" 0,0,0.0 -· • f.S
. . ·s.A,P
PT_ PIEDRAS
8
I N
37'
I 14
MILES
l. 8LANCAS \.'., ·.-.SAN Slf'I.EON
S O,P.
D I
MIL E:S
P,.,,.
122"'
FIGURE 1: Dist1·ibution of sea otter carcasses along the central California coast. (p~pup; i=imature; s=subadult; a=adult; o= aged adult)
9
CBL -----------'
L-------~L------------'
FIGURE 2: Skull and mandibular measurements. (CBL=condylobasal length; ZW=greatest zygomatic width; ML=mandibular length; MTR=mandibular tooth row; CAL=canine alveolar length)

DENTIN
PULP CAVITY
0.
A
ENAMEL-----;//\
t. I \
b,
ROOT---< NECK
\' ! \,·~
!\l I I I i I
1---A----J I B.
' I
TOOTH AXIS
I
BOSS
FIGURE 3: Diagram of a medial lonni-section of a sea otter upper right canine (a) shoi'Iing hard tissue nor!Jhology, and diagt·am of a seC\ otter upper left canine (b). (A=greatest tooth 1·1idth; G=root len!]th; C=greatest cenentum width)

DENTIN
-.:-...-__..---;::::..-·
PULP CAVITY
=-- ~
0.
i b. ENAMEL---------;
:-----_DENTINO--ENAMEL:
JUNCTURE
ROOT NECK
,,,t.·
CEMENTUM---..\
A
...... _.....v./\.
I I
c
TOOTH AXIS
B.
BOSS
FIGURE 3: Diagram of a medial lon~i-section of a sea otter upper right canine (a) showing hard tissue morphology, and diagram of a sea otter upper left canine (b). (A=greatest tooth width; G=root length; C=greatest cementum 1vi dth)
0

Greatest tooth width Greatest cementum width Root length Tooth weight Number of cementum layers
ll
All measurements and comments were recorded on a data sheet for
each animal (see Appendix A).
Teeth ---A. Decalcification
The fo 11 o~li ng is a 1 is t of common dec a 1 ci fyi ng agents and the re-
sults I obtained with each.
1. 3% f!Cl
Decalcification with 3% f!Cl was reported by Schneider (1973)
to be quite effective on Alaskan sea otter teeth. My complaint is that
it tended to decalcify too quickly with concomitant shrinkage and dis-
tertian of the tooth. Also, if extreme care was not taken, over-decalci-
fication vmuld occur. The major advantage of f!Cl is that decalcification
is very short.
While using f!Cl, I developed a method for preparing serial
sections of southern sea otter teeth. Teeth were wrapped in cheese cloth,
placed in individually marked containers, run through the Fisher Tissue
rnaton and treated in the same fashion as the soft tissues discussed under
Reproductive Organs (page 19). As each 20 u section came off the mold,
it was lined up next to the previous section, thus allowing for a com
plete series to be contained on one slide. A drop of hot water facili
tated v;ax l'emoval around the specimen and made it easy to see the thin

12
section, roll it out and orient it properly on the slide. This method
was tried on open rooted teeth that are easily decalcified and may be
an excellent method for studying tooth growth, maturity and root canal
closure.
2. Trichloroacetic Acid/Sodium Acetate 50:50
This method (Dr. H. Murphy, personal communication) is excel
lent for decalcifying teeth, because the process is slow and there is
very little shrinkage and distortion. The major problem was that the
decalcifying process was too slow! After three months, the teeth were
still hard. A further problem with this method is the inconvenience.
Reagents had to be measured and changed often. The process was simply
too time-consuming.
3. EDTA Acid, 41.3 gm; NaOH 4.4 grn and Distilled vlater to Make
a Liter
This decalcifier, reported by Scheffer and Peterson (1967)
had disasterous effects upon the teeth. Prior to decalcifying, teeth
v1ere ground in half medially to allow for greater penetration of the
decalcifying agent. They were placed in small vials, the EDTA being
changed frequently, and when able to monitor, I placed them in a l00°F
oven. After three weeks in this solution, the body of the tooth (i.e.,
the dentin) was still completely hard, whi1e the cementum was completely
dissolved.
4. 5 Parts Formic Acid, l Part Formalin and 20 Parts Hater
These reagents were reported to produce excellent results in

13
fourteen days by Crowe (1972) and in five to seven days by Linhart and
Knowlton (1967). The sea otter teeth reacted similarly to those de
calcified in the EDTA acid with only slightly less damage to the cemen-
tum. The above researchers did report using the formic acid solution
fot· fourteen days on bobcat teeth and five to seven days on coyote teeth,
respectively, while I soaked the otter incisor for almost twenty-one
days. Perhaps bobcat and coyote canines are less resistant to decalci-
fication than the upper third incisors of the sea otter.
5. Differential Etching with 10% Formic Acid
The tooth was halved along the longitudinal axis and etched
with 10% formic acid for twelve hours. This method, used by Gambell and
Grzegorzewska (1967), proved to work well for sperm whale teeth etched
for thirty hours; however, sea otter incisors rapidly shrank and distor-
ted after twelve hours to such a point that it became impossible to
read any annuli.
The tooth originally decalcified was the third upper incisor. It
1<1as chosen because, along with lower premolar I, ha9 been reported by
Kenyon (1969) and Schneider (1973) to yield the best results. Premolars
were not used in my study because some or all were missing from various
specimens.
B. Grinding
flhen research began, the upper left third incisor was removed from
several specimens and ground in order to test the feasibility of the grind
ing method. G · d' r1n 1ng was carried out in two fashions.

14
Firstly, teeth were glued to a petrographic slide with Aremco Crys
tal- Bond #509, a thermoplastic adhesive used by micropaleontologists.
When solidly in place, the teeth were ground longitudinally on a fine
lap wheel using silicon carbide abras_ive (600 grit). When the tooth was
virtually ground in half, it was polished with M-302-l/2 abrasive (alumi
num oxide), washed first in running water and then distilled water and
finally blown dry. The slide was then returned to the hot plate, the
adhesive melted and the tooth turned over so that the flat surface of
the tooth was glued to the slide. The grinding process was continued,
but interrupted often in order that the specimen could be examined
through a dissecting scope. As the tooth became too thin for maximum
control on the lap wheel, grinding was continued manually on a glass
plate for greater control.
Usually when the tooth was translucent enough to allow the light to
pass through (that is, when held up to the light) the grinding process
was complete. At this point, the thin section was approximately between
25 and 40 microns in thickness. The tooth surface was polished with
M-302-l/2 abrasive and a coverslip applied with Adam's histoclad.
The other method used was similar to the one just described, but
only continued to the point at which the specimen was simply ground in
half. The annuli on these teeth were prominent due to heavy echino
chrome pigmentation. Unfortunately, only a few of the sea otters ex
amined had echinochrome-stained annuli.
After several decalcifying agents were tried and failed to produce

15
acceptable results, grinding was chosen as the preferred processing
method. \iith incisors ruined by decalcifiers, I turned to processing
the upper right canine as each specimen had one and results would there
fore be consistent.
c. Staining Tooth Tissues
Several stains were used in an attempt to enhance visibility of
the annuli. Hematoxylin was reported by Craighead, !U_. (1970), Gil-
bert (1966), Klevezal and Kleinenberg (1967), Klevezal and Marakov (1966),
Hurphy (personal communication, 1978), Sauer, et _<U_. (1966), Schour and
Hoffman (1939), Stoneberg and Jonkel (1966) and others as providing ef-
fective results in this regard. It proved to be ineffectual, however,
in staining the otter tooth samples.
Next I tried a Giemsma stain (Schneider, 1973; Stone, et _<U_. 1975).
Although it imparted a deep purple tint to the dental tissues, there was
no differential staining of annuli. This could be because the research
ers referenced above were staining decalcified tissues while mine were
undecalcified and Sergeant (1959) reported getting "variable results
vlith hematoxylin on undecalcified tooth thin sections". Orcein, a stain
used on myelin sheaths and dental and paradental tissues also had no ef
fect.
The one stain that proved to be most effective was alizerine RedS,
a stain used by geologists at the United States Geological Survey, Menlo
Park, California, for staining rocks with a high concentration of calcium
carbonate. It is also reported to be an effective stain for calcium con-

16
taining cartilage and bone (Gurr, 1962). Alizarine RedS is taken up
readily and quickly and care must be taken to avoid over-staining.
This dark red stain was applied by dipping slides into the staining
bottle for 5-15 seconds. They were then removed, examined and, if the
teeth were still light, dropped back into the bottle for five to ten
more seconds.
In the beginning, each tooth slide was examined with both trans-
mitted and reflected 11hite light usin~ a dissecting scope at 10 or 40x.
A polarizing scope was also used. Since all methods yielded the same
results, the teeth were examined with transmitted 1 i ght from that time
forward unless the section v1as thick and required reflected light for
proper visibility. Annuli take on a different appearance with reflected
and transmitted light, so once a technique is developed, it must be
strictly adhered to for the sake of standardization.
D. Reading the Annuli
I gained familiarity with cementum layering by examining methods
reviewed by Klevezal and Kleinenberg (1966) and Sergeant (1967). Also,
most papers on the subject either review previous work or provide an ex
tensive bibliography. My major source of proper technique, however, was
Karl Schneider (1973). His v1ork on aging the Alaskan sea otter by read
ing cementum annuli was explicit in regard to materials, methods and
technique. He and I had a long discussion one evening and in general,
he agrees with my interpretations (K. Schneider, personal communication).
Teeth were examined under a binocular microscope using 10 or 40x

17
magnification and transmitted light. One annuli contained both the
narrow, translucent (winter-spring deposition) and the wide, opaque
(summer-fall deposition) bands. Age was determined by counting the num
ber of narrow bands (n) and adding one (n+l) to account for the time of
tooth eruption, root canal closure and time at first cemental deposit.
Annuli counts were not always consistent, as determinations were
often made in several areas of the root. Each root, however, contained
a posterior, lateral bulge termed the boss by Low and Co~1an (1963, see
Figure 2) which usually possessed the most easily readable annuli.
Not only were several annuli counts made on each tooth, but dif
ferent methods were emp 1 oyed to make those counts and lvhen avail ab 1 e,
different teeth were used for comparison. The teeth compared were
usually the upper third incisors ground in earlier stages of my re
search. Thin sections of the canine teeth were viewed through a micro
scope as well as projected through a photographic enlarger (a technique
suggested by D.E. Sergeant, personal communication). After reviewing
the count obtained for each technique, one count, usually the average,
was accepted (see Count Used column, Table 1).
Reproductive Organs; Embedding and Staining
Gonadal tissues were left in Bouin's fluid until it was convenient
to \•IOrk thern up. This time was always less than two months, but usually
vlithin the second to fourth week of acquisition. The time lapse v1as
necessary, not only for fixative infusion, but also because it was not
economical to run the equipment for one specimen at a time.

18
TABLE 1: Tabulation of cementum layer counts obtained using different teeth and different methods of observation. Numerals fol1ol'ling tooth designation3 indicate number annuli counted. (LI =left upper incisor; RI =right upper incisor; U4=1ight microscope; P=polarizing scope; CL=camera lucida tracings; AS=alizaring red S stain) Canine Study Technique
Animal First Grinding Light No. and Tooth Used Microscope ~hotograph Enlarger Count Used
497
499
509
515 516
520
521
528 529
535
538
550
577
590
592
593
607
3 LI : LM 1; P 1-1/2; CL 4-1/2-5
RI 3 :U~ 13; CL 10
LI 3 :U.1 13-1/4 U 3:LM 3; CL 1-3/4
3 LI 3:LM 3; RI :LM 3
3 U :LM 4, very difficult 13:P&AS 11
RI 3 :LM 3-1/2
RI 3:LM:3-3-1/2
2-1/2-3
8
7-1/4
11-1/2
3
3-1/2
putting on 9
8 to 10
3
10-1/2
4-1/2-5
3-1/4 2-1/2-3
15
13 10-12
3 3-1 I 3
3-1/2 3
10-1/2
11
3-1/2 3
10-1/2 10, very · hard to
read
3-1/2- early 5 5-l/2
11-l/3 areas in- 11-1/2, terrupted 13-1/2,
11-3/4
16, lots of 13-16 11+ \'lear
6-l/2
3-1/2
2-1/2
3-1/2
3-1/2
6, hard to count
3
4
3-1/2
3-1/4
8
13
11-1/2
3
3-1/2
9-10
10
3-3-1/2
10-1/2
5 ?
11-1/2
16
6-1/2
3-1/2 3
3-1/2

19
Two and sometimes three 3 mm slices v1ere removed from the middle
of both left and right testes. These were then cut into quarters and
placed in individually labeled containers. Epididymal samples were
taken across the·center of the corpus epididymis. These were halved
or left whole, depending upon the circumference of the corpus and also
placed in individually-labeled containers. The containers were then
placed in running water for 24 hours to remove the picric acid stain
from the tissues and stored in 70% Etoh until placed in the Tissuematon.
Sections of both the testes and epididimydes were then run through
the Fisher Tissuematon on an 18-hour cycle. The cycle was adjusted to
be complete the next day at a convenient time, so as not to allow the
sections to soak in the liquid tissuemat for very long. The schedule
for dehydrating, infiltrating and embedding (Galiger and Kozloff, 1964)
is outlined below.
Dehydrating, Infiltrating, and Embedding Using Dioxane and Paraffin
I. Dehydrating:
A. l dioxane: 1 95% Etoh. . l hour B. 100% dioxane. .2 hours c. 100% dioxane. . 2 hours
II. Infiltrating:
A. Dioxane saturated with paraffin ... 3 hours B. Paraffin. . .3 hours C. Paraffin ............... 3 hours
D. Tissuemat .............. 4 hours i.
'
.. i

20
III. Embedding:
A. Place a layer of tissuemat in the mold
B. Place tissue sample in the mold
c. Add tissuemat to fill D. Cool in cold-water bath
The final stage of the inf11trating process takes place in a re-
movable thermostatically controlled canister that may be placed at the
work bench, plugged in, and maintained at 57" C.
Time is of the essence during the final embedding procedure. Indi
vidual sample pieces were placed in a mold, into which a layer of hot
tissuemat had already been added. The section was horizontally oriented
and the mold was completely filled. Cooling was accelerated in a cold-
water bath and the tissues were ready for sectioning vlithin one to two
hours.
Seven-micron sections, made on a Spencer AO rotary microtome "820",
were placed on slides already coated with a thin layer of albumin.
Several slides were made from each mold so that every sea otter had at
least five slides from each of three testes and epididymal sections or
about fifteen slides total. The slides were placed on a warming tray and
allowed to dry for at least 24 hours.
The following staining schedule, adopted from Humason (1967), worked
well for sea otter gonadal tissues:
Xylene Absolute alcohol 95% alcohol 7o:~ a 1 coho 1
3 minutes 3 minutes 3 minutes 3 minutes
i i
'!

Running ~later Hematoxylin Running 1·1ater Scotts solution Running ~later Eosin (counter stain) 70% alcohol 95% alcohol Absolute alcohol Absolute alcohol Xylene Xylene
4 minutes 4 minutes 4 minutes 3 minutes 4 minutes 45 seconds-] minute 2 or more dips 3 dips · 3 minutes 3 minutes 3 minutes 3 minutes
21
Slides were removed from the xylene bath, two to three drops of
permount were placed on the slide and a 24x50-60 mm coverslip was af
fixed to the surface. The large coverslip was used in order that the
maximum number of thin sections could be covered.
Tissue sections were examined through a Nikon binocular scope,
generally using 40x magnification and occasionally lOOx (oil immersion)
when looking at the fine detail of spermatogenesis.
Fel'ti 1 i ty ~leasurements
Traditional fertility measurements such as fresh testis weight and
dimensions were impossible to take, as all testes when received by me
~1ere already sliced and in fixative. Measurements, therefore, had to
be taken from prepared thin sections.
An attempt was made to tally the total area (in f12) of a Lydig cell
grouping. Area measurements of single Lydig cells were also taken and
plotted (Figure 4).
Hidth and depth measurements were also taken of the seminiferous
tubules and diameters of individual tubules calculated by using Bigg's
(1969) formula: diameter = width ; depth The number of cementum layers
was Plotted against ~lidth, depth and diameter of tubules.
_] _]
100
80
w 60 (.)
w _] (C)
z Ui 40 <! lL 0 q LlJ 0:: q
z q 20 w ~
0
f 0 2
" 0
0
1
l
6 10
I t' 1
NUMBERS OF CEMENTUM LAYERS
22
14
FIGURE 4: rlean area of a single Lydi<:' cell rlotted against nunber Df ce1nentum annuli deposited in the teeth of imnature tllrounh ac1ed adult anir~al s. (i=i~"mature; s=subadult; a=adult; o=aged adult; t=small amounts of srerm in the testes; e=small amounts of srem in the epididymis; TE=lal'[)e ar1ounts of sperm in both testes and epididymis)
18

23
Photography
Photographs were taken of gonadal tissues using a Hild 14Ka 1
photomicrographic camera and FX 135-Panatomic X fine grain black and
white film with an ASA of 32.
Skulls were photographed with a Hasselblad 500 C using a 50 mm wide
angle lens and tri-X film rated ASA 400. Detail was enhanced by using
reflected light. The bacula were photographed with a 4x5 view camera
using Plus X film rated ASA 125. Exposures were 2 seconds at f 22.
Teeth were photographed through a dissecting microscope using 6x
(l25x) or 12x (500x) magnification, depending on the diameter of the
cementum bulb. Panatomic X film (already described) was used for this
purpose.

CHAPTER 3
RESULTS
Tooth length {crown and developing root) and diameter of root
canal opening were plotted against mandibular length (Figure 5). There
was an inverse relationship betv1een increasing root length and diameter
of root canal opening for deciduous teeth. Permanent canines, on the
other hand, showed a positive correlation between tooth length and root
canal opening until the pups become "immatures" at between six and eight
months of age, or when the tooth was between 27 and 31 mm in length
(Figure 6).
It is impossible to calculate a curve, as there are so few data
points (8 pups, Z immatures). If one assumes, however, that (1) sea
otters 495 and 553 were a minimum of six and a maximum of eight months,
respectively; (2) each was representative of their age group's popula
tion; and {3) their root canals were closing at a constant rate, then
one can draw a 1 i ne between the two data points ( 495 and 533) and obtain
a theoretical rate (3 mm/2 months) of root canal closure (Figure 7).
Since there is no mention of exact age at time of root canal closure or
When the first cementum layer is laid down in any past work dane on sea
otters, I can only postulate that it is some time between nine months
and one year of age.
The canine tooth of sea otter 495, a 6-7-manth old, was decalci
fied in trichloroacetic acid and serially sectioned. There was no depo
sition of cementum.
24

(9 z
8
z 6 w Q.. o ........ ...J E 4 <t E z <( u 1--0 0 a::
2
Ill Ill
Ill
Ill Ill Ill
• I • •
45 47 49 51 53 55 MANDIBULAR LENGTH (mm)
PERMANEf\IT II DECIDUOUS «>
FIGURE 5: RLJt canal openin~ in mm of deciduous and permanent canine teeth . plotted against mandibular length.
57 N U1

16
E E
t.? z z w Q_ ol2 _J <( z <( u f-0 0 8 IX lL 0 IX w f-w :?: <(
4
0
0
/
/ •• /
/
8
FIGURE 6:
--~
~ ~
~~
·~ • - • .. -.. -
16
Diameter of root canal opening
• __ ......... ___ _
24 TOOTH LENGTH (rnm)
plotted against ~ngth at growinn
32
rermanent canine tooth.
40
e N
"'

E E
(:J
z z w (1_ 0 __j
<1 z <1 u 1-0 0 0:: LL 0 0:: w 1-w ~ <1 0
27
9 \ 495 •
• 6 • ,.553
• •
I
~ 527.,
3
0~.---~-------.------~r-------~------, 0 3 5 7 9
AGE IN MONTHS
FIGURE 7: Diameter of root canal openino rlotted against age in months. Line dra11n betv1een imrrature sea otters 495 and 553 represents a theoretical rate of root can~l closure (3 m/2 mo). Sea otter 527, a known age female was plotted to test for accUl'acy.

28
The mandi b 1 es of pups in this samp 1 e ranged between 45 and 55 mm
in length. The diameter of the root canal openings and length of the
growing canine tooth varied between 4.3-6.2 mm and 9.1-13.6 mm, respec
tively (see Table 2, Figures 5 and 6).
Table 3 depicts the tooth eruption sequence in sea otter pups by
incorporating data from Lensink (1962) and Kenyon (1969}. As a check
of its accuracy, a known age female pup ("Rosie", #527) was aged with
the criteria set forward in Table 3 and found to be aged accurately.
After pups were aged from Table 3, their age in months was sub
tracted from their date of collection, and a date or time of approximate
birth was calculated for each (Table 4}. Since all pups were in good
condition when collected (Table 2), they are considered freshly dead
with a minimum of time elapsed between death and recovery. Of ten ani-
mals, three were born in January, three in February, one in August, two
in October and one in November, or 90% were born from October through
February.
The testes of pups were less than 2 em in length. Seminiferous
tubules were lined with Sertoli cells and spermatogonia and there was no
sign of spermatogenesis (Plate 34}. Lydig cells had a mean area of less
than 10 l (Figure 4}. One of the o 1 der pups ( 493, 2 to 3 months) had a
few spermatogonia undergoing mitosis.
Fifty percent of the eight pups collected came with measurable
bacula. These tiny penal bones ranged betv1een .043 and .075 gm and were
from 14.06. to 20.39 rmn in length. Neither the distal nor proximal par-
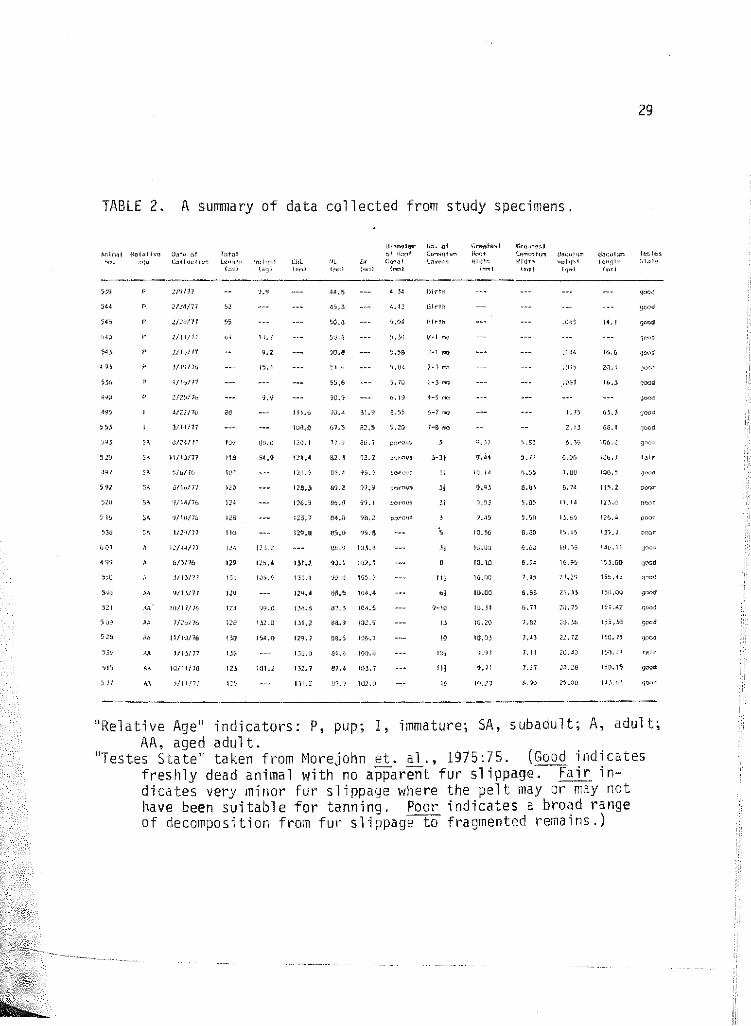
TABLE 2. A summary of data collected from study specimens.
t.",lnql i<ul~tivn J"'" ;,! bt~l '>fl. ·''1" C.-,t!,H;Iicr. t~"11h 'h:I•Jh!
4Vl
,, ,,
:/'l/1}
1/!1!1!
Ulv/11
l/h/P
!/H/17
<J/:·l/P
l\/IJ/77
~/o/lb
jfl,/11
1~/14/11
6/3116
J/13/H
'J/1';/11
Art' 10111/l'i
M !1/hJ/1\1
l!l)/71
lll/11/JG
;0/11/17
;,d lo'jl
li!.l
IOl'
!}4
ll<i
LH
"' "'
II.!
15,1
lU1.2
tUL
·~·
!! l,u
1?1.~
1).1.5
12&.:.
116.5
;:q, 7
1Z9.U
1 '!1. 7
111.2
UI,.,.,.J!::r lb. ;:I Gr,cttf~st
c I >b;lf CwN<I h"' 11""11 bl LM~I L~vnr1 1'1hl'~
(.,·nl ;,..,1 !,.,I
'j,O·l
55.& ). 10
%.·! W.\,il
'l.LI H)2, I
IJ~.5 HM,4
ill.) 104.5
!17.4 HH. 7
llin·
Ulri~
l11t!h
6-1 "'<l
,,
'" ,, -:<-IC
u
"' !Jl
l(,
J .. l5
IJ.%
IO.tlJ
lQ,OU
u.ca
10.20
1.1)
1.11
10.;?1
tru<Oj<!c.l ,;,,.,.,~r""' <JR""'""' Wl<lt'' ~~hj•d
(..,) hp!
,·.H~
iLN
'L51J
6, 71 2.). ])
1.et
l,U
1.11 ;;o,J;)
1.27
!L'}}
ll<lo:ul>l"' vm:;h ,_,
14.1
115.1
111.1
29
!!'"!<'' :d.>h!
"Relative Age" indicators: P, pup; I, immature; SA, subadult; A, adult; AA, aged adult.
"Testes State" taken from ~1orejohn et. al., 1975:75. (Good indicates freshly dead animal with' no apparent fur slippage. Fair indicates very minor fur slippage where the pelt may or may not have been suitable for tanninq. Poor indicates a broad range of decomposition from fur s 1 i ppage"""ta fragmented rerna ins.)

30
TADLE 3: Pup ages based upon tooth eruption sequences propose!i by Kenyon (1969, above) and Lensick (1962, below). Question marks indicate Ken~10n and Lensick v1ere unclear as to hov1 long tooth was erunting before it v1as permanently in place. --.=vesti(leal; ~deciduous tooth eru;Jted at birth; --= deciduous tooth be·ing shed; .::::I=perrwnent tooth erupting; §!!=permanent tooth AGE IN MONTHS fully erupted.
------'·------------ ------------?--- ----- ------·
' I
490 495 533
I
31
TABLE 4: Proposed pup birth dates based upon collection dates and tooth age
Sea Otter Number
490 493 495* 536 539
540 543 544
545
553*
Date Collected
25 February 1976 19 March 1976 22 Apri 1 19 76
16 January 1977 9 February 1977 11 February 1977 16 February 1977 24 February 1977 28 February 1977
14 March 1977
*Immature specimens **Plus or minus one or two days
Tooth Age
4-5 months
2-3 months 6-7 months 2-3 months birth bi rth-1 month Birth-1 month birth
birth 7-8 months
Proposed Date of Birth**
10 October 1975 1 January 1976 5 October 1975 1 November 1976 5 February 1977 26 January 1977 29 February 1977 20 February 1977 24 February 1977
1 August 1976

32
tions of the bacula were developed (Plate 32).
All pup skulls became disarticulated after the masceration process
because sutures between individual bones had not yet fused. However,
individual bones, including the mandibles, were representative of ani
mals in the pup age class already described by Lensink (1962), Kenyon
(1969) and Morejohn, et !D_. (1975).
Immatures
There are only two animals in my sample representing this relative
age class; sea otters 495 and 553. Table 5 demonstrates how skeletal
development may not always follow characters proposed in the literature
(Lensink, 1962; Kenyon, 1969; Morejohn, et !D_. 1975) as definitive for
aging.
In spite of the fact that sea otter 495 was a larger animal with a
seemingly "older" cranial development (at least suturewise), 553 was
classified older because of its more advanced tooth eruption sequence
and bacular development. (553 had a longer, heavier baculum. See
Table 2, Plates 5, 6 and 32.) Neither animal had any cementum deposited
on the upper canines.
The testes of 495 showed no evidence of spermatogenesis. A few
primary spermatogonia were interspersed among the Sertol i cells which
line the seminiferous tubules. Lydig cells, still mere dots, had a mean
area of 9 i ± l. 3.
The testes of 553 represent an advancement over those of 495.

TABLE 5: A comparison between two immature specimens
Character Compared
Permanent teeth open rooted
Wear on lateral anterior surface of lower canines
pm~ :~ still in place
Deciduous post-canines show wear
P~l2, 3
PM2 Presphenoid-pterygoid sutures
Basioccipital-exoccipital sutures
Parietal sutures Area around interparietal and posterior lateral parietals
Post orbital process
Anterior portion of palatine sutures
Maxillary-palatine sutures
Frontal-parietal suture
Maxillary-frontal suture
Parietal-squamosal suture
CBL
ML
zw
Sea Otter 495
All, including PM1 . 1
No
Yes
Some
Not erupted through bone
Not erupted through bone
Closed
Still open
Less interdigitated
Still open Fully developed and pointed
Closed
Sti 11 open
More fully closed
More fully closed Less fully closed
lll. 62 mm
70.36 mm
81.97 mm
Sea Otter 553
1 2 1 11:2 almost closed; PM essentially
closed but still porous
Yes
Raised above erupting permanent premolars and ready to fall out Heavily cavitated
Erupted through bone, not gum
Eruption 1/4-3/4 complete
Sti 11 open
Fused in a few places
Heavily interdigitated
Fusing Poorly calcified and blunt
Fully open but heavily interdigitated
Closing
Less fully closed
Less fully closed
Fully closed
108.00 mm
67.45 mm
85.52 mm
w w

34
Spermatogonia and primary spermatocytes in mitosis predominated (Plate
35). Serto 1 i ce 11 s which 1 i ne an immature tubule 11ere 1 ess evident
than in 495. The mean area of a single Lydig cell was 6.8 i ± 1. 1.
Subadults
There are eight animals in the sample whose skull and dental features
put them into the subadult category as outlined by Morejohn, !ll.· (1975).
Sea otters 538 and 607, which have characters that overlap between the
subadult and adult age classes, will be discussed later.
Each of the first six animals had canines which were fully erupted,
in place, and showed little or no wear. The apex of the root was still
porous and cementum deposition was limited to the bulb surrounding.the
root tip. Each had between 2.5 and 3.5 cementum annuli or were between
3.5 and 4.5 years old.
Although younger subadults could be separated from adults by condy
lobasal and mandibular lengths and zygomatic widths, the older subadults
have measurements which overlap into the adult age group (see Table 2 and
Appendix B). Baculum weights and lengths can be used, however, to achieve
separation (Table 2, Appendix B, Plates 5, 6 and 32).
Gonads were avai1able from three of the six animals under discussion.
Sea otter 593 had spermatogonia, some dividing, lining all of the tubules
examined. Sertoli cells were still present in high numbers. Lydig cells
were singular and there 1·1as no evidence of spermatogenesis.
The tests of 529 still had large numbers of Sertoli cells, but also
contained primary spermatocytes, indicating the beginnings of meiosis

35
(Plate 36). Lydig cells were granular, but still small in size. The
pseudostratified steriocilla cells of the epididymis were mature.
The seminiferous tubules of 497 were filled with Sertoli cells
and only a few Lydig cells were seen dispersed among the tubules. Cells
of the epididymal canals were not yet fully mature.
The total lengths of the six subadults measured varied between 107
and 128 em with a mean of 118 em. Bacula varied between 6.06 and 13.65
gm with a mean \'Ieight of 8.97 gm. The mean bacular length was 113.98 mm,
but lengths ranged from 106.17 to 126.37 mm. The mean area of subadult
Lydig cells varied between 7 and 12 ~2 (Figure 4).
Sea otter 538 presents a real aging problem. Although the physio
logical state of the testes would provide a definitive way to age the
animal, this specimen did not have gonads available. Its total length
is well below the mean of the six subadults and it does have three to
five cementum annuli, depending on v1here the count is made (Plate 18).
However, the skull is fully developed. The lateral sutures between the
parietal and frontal bones are still in evidence, as are the presphenoid
pterygoid sutures. The suture between the parocipittal process and the
squamosal is also evident. The above sutures are all closed on the two
fully adult specimens in the collection (499 and 550). The saggital and
occipital crests are developing. The baculum measurements of 538 are
also intermediate bet1<1een the subadult and the adult and aged adult
specimens (Table 2). So, with some reservation, I am including 538 in
the subadult category.
In general, 607 is comparable to 538. The base of the skull of

36
the former was damaged by a bullet, so some sutures are impossible to
describe. The paroccipital-squamosal sutures are still open, but the
parietal-frontal ones are closed. The ridges on the skull are more
fully developed than 538's, as is the baculum. The zygomatic width is
greater than the other subadults. The gonads provide the evidence for
classifying 607 as an adult. The seminiferous tubules contain cells,
50% of vlhich have advanced only to the secondary spermatocyte stage of
me1osis. Hov1ever, small numbers of spermatids are seen toward the cen
ters of the lumina and Lydig cells, still few in number, are granular.
Small numbers of normal, mature-looking sperm are seen in a few of the
epididymal tubules.
This specimen has an unusual acellular or fibrous growth within the
testis. More peculiar still are the large numbers of granular Lydig
cells which line the growth. This proliferation of interstitial cells
is highly irregular for an animal of this age class, but is seen rather
regularly in the testes of aged males (Plate 41). Another peculiarity
observed was the presence of anomalous "sperm" in the epididymis. When
these darkly staining four and a half by less than one micron objects
were first seen in the epididymis, the possibility of their being bac~
teria v1as explored. However, after careful consideration and discussion
with Drs. Akiyama and Haight of the Microbiology Department of San Jose
State University, it was agreed that bacteriological infection can be
ruled out. The reasons for this are as follows: (l) If there was a
bacterial infection, there should not have been a monoculture of bac-

37
teria. (2) The dark, staining objects are limited to areas also con
taining ''normal'' sperm and sloughed cells, and are not dispersed through
out the tubules as would be the case in infection. (3) There is no in
dication of an inflamatory response. Because this animal does have some
normal looking sperm in the epididymis and few, but nonetheless present
spermatids in the seminiferous tubules, it is classified as an adult!
Adults
There are two animals classified as adults in the sample. Sea otter
499 was 53 pounds when captured by California Department of Fish and Game
in 1976 and given to Sea World San Diego. He died after three and a half
years in captivity. The cranial development is similar to that of the
other adult specimen, but lacks its robust appearance and saggital and
lambdoidal crest development.
Only the canines and upper incisors show any wear. Cementum was de
posited sparsely up the neck of the root; a characteristic of subadult
teeth. The upper right canine was ground and found to have eight annuli.
This would mean that the animal was approximately nine years old upon
death and four and a half when captured.
The testes were not available for inspection by me, but were weighed
upon necropsy and found to be 14 gm each (Dr. Cornell, personal communica
tion). The baculum was longer than all but two adult and aged specimens,
but its weight falls well within the range of the subadult bacula (Table 2).
Sea otter 550, the only other adult in the sample, was one of the
largest males collected. Its teeth showed only minor wear and the upper

38
right canine contained 11+ annuli {see Plate 21). The three skull
measurements taken were all well over the subadult limits and even
out-measured several of the aged specimens {Table 2).
The testes of 550 were in prime condition, showing active spermato
genesis throughout (Plate· 37) and the· epididymis 11as loaded with sperm
(Plate 38). Twenty-five Lydig cell area counts were made, from which a
mean area of 51.33 / ~1as calculated. This mean area fell some11here in
the middle of the five aged adult measurements (see Figure 4). In a
further attempt to achieve separation of the age classes, length and
width measurements of the adult and aged adults' seminiferous tubules
were taken and applied to Bigg's formula. The mean diameter of the
adult seminiferous tubules was 259.8 ~2 ; the mean width was 171.8 u2.
The mean length, width and diameter measurements for each adult and
older animal were plotted against the number of cementum annuli and no
significant differences between these two age groups were observed.
The baculum of 550 was longer and heavier than any of the other males
in the sample.
Aged Adults
Animals ~1ere considered aged if their teeth sho11ed extreme wear or
there was loss of teeth and/or necrosis of the alveoli. Periodontal
disease was commonly evident (see Plates 24, 26, 28, 30). (Kenyon, 1969;
51, has excellent photographs of severe bone and tooth damage.) Their
skulls contain well-developed sagittal and lambdoidal crests and all
sutures are closed.

39
The n~an candylobasal length far this relative age class was 131.29
mm, but lengths ranged from 129.42 to 134:78 mm (Table 2). The mean
mandibular length was 87.88 mm with lav1 and high extremes at 84.81 and
89.85, respectively. Zygomatic width.s ranged from 100.82 to 106.14 mm
with a mean of 103.59 (Table 2).
Roots of the canine teeth had cementum extending up the neck of the
tooth. The cementum bulbs were so large and the teeth so completely
anchored within the alveolus that most had to be cut or sawed out. Two
specimens (515 and 577), having received trauma to the root apex, had a
small or incomplete cementum bulb, 11hich made it difficult to read the
annuli properly. Aged adults had a mean number of 11 annuli, but counts
ranged from 6.5 to 16.
Aged adults have a mean total length of 128.14 em. The shortest
animal was 123 em; the longest 138 em.
In general, aged specimens had a degenerating testis. Seminiferous
tubules seemed to be collapsing or degenerating. In several, spermatogene
sis had slowed dm1n or even stopped and in three specimens (515, 521, 528)
the seminiferous tubules had taken on an immature appearance with Sertoli
cells lining the tubules and the lumina being filled with primary and
sometimes secondary spermatocytes (see Plate 39). In all cases, Lydig
cells were large and granular. The mean area of a single Lydig cell far
this relative age class is 64 u2 Three aged adults (509, 521, 528) had
an unusual proliferation of Lydig cell tissue (Plate 41). All aged ani
mals have heavy bacula (mean weight of 21.55 gm) with rugose proximal

40
ends (Plate 33).
Measurements
Various measurements were taken in an effort to record growth and
separate the five age classes. Frequency distributions were plotted
for the three skull measurements and found to be ineffective for sepa
rating animals at the upper and lower limits of their relative age classes
(see Appendix B). Lensink (1962) had similar results.
Bacular measurements, on the other hand, were useful in attaining
a separation, at least above and below the adult level (Appendix B). It
appears, from the data collected, that the baculum becomes ''adult'' when
sperm production commences. That is to say, subadult and younger speci
mens which have not yet produced mature sperm have a shorter and less
robust baculum than adult and aged specimens (Plates 32, 33). The only
exception to this is 499, an animal which spent three and a half years
in captivity and is suspected of having hormonal problems.
The small sample size, coupled with the fact that two of the three
adults are anomalous, makes it difficult to unequivocally put a lower
limit on the weight or length of adult bacula, but it seems clear at
least that animals with bacula weighing less than 16 gm and having lengths
less than 140 mm are still subadults.
It is equally difficult to separate adult and aged animals on length
or weight of bacula alone. Clearly, skull and dental morphology and
gonadal histology must also be used.
Several gonadal measurements were used in an attempt to achieve age

41
class separation. The first tried 1·1as mean area of a sing'le Lydig
cell. Although this method enables one to easily separate immature
from mature animals (Figure 4), it is inadequate for use with adult
and aged animals. Also ineffective was the taking of seminiferous
tubule measurements using a formula proposed by Bigg (1969). A much
more effective measurement 1·10ul d have been whole testis length and
weight and should be the measurement used in future studies.
Because the mandibles were the only bones consistently available
from each one of the skeletal specimens in the collection, they 1vere
used in an effort to separate the various age groups. When plotted
against root can a 1 opening, permanent and deciduous tooth grov1th pat
terns could be discerned (Figure 5). However, when plotted against num
ber of cementum rings (Figure 8), the graph created had an equal proba
bility of being either a linear or an exponential curve. It was, there
fore, decided not to use mandibular length measurements as a criterion
for growth.
Distribution
Sea otter carcasses were collected along the central California
coast from San Simeon in the south to Point Arto Nuevo (northwest of
Santa Cruz) in the north (Figure 1). And although the sample size is
small, a slight distributional pattern of age classes is observed.
Five of the six pups collected were found in Stillwater Cove and to the
south. Seven of the eight subadults collected were found within f.lon
terey Bay with all but two being found in the north bay area. Sea otter
607, the young "adult" or old subadul t, was found there also. The sub-

• 16
:J 12 ::J • • z
•
:::E ::J 1- • • z w :::E 8 • w • u lL 0 • 0:: w ([) • 2: ::J z 4
• • • •
•• • •
0 .. ... . • • • +> N
45 50 55 60 65 70 75 80 85 90 MANDIBULAR LENGTH IN (mm)
FIGURE 8: Number of cementum annuli plotted against mandibular lenath (m).

43
adult collected alive the Monterey breakwater (499) was the specimen
kept by Sea World San Diego for three and a half years and for this
study is considered an adult. Five of the seven old animals were col
lected in the south bay area with the other two at either end of the
sample range. Four of the five old animals in the south bay area were
found in and around the wharf and breakwater.

Reproductive Aspects
CHAPTER 4
DISCUSSION
The sea otter differs in many respects from the other members of
the family Mustelidae. Its aquatic adaptations have been described at
length by Taylor (1914) and others. The teeth, adapted for crushing
the exoskeletons of hard-shelled invertebrates and scooping them out of
their tests (Fisher, 1941; Hildebrand, 1954) are also unique among the
family, as well as within the order: Carnivora. The reproductive as
pects of its life cycle presented in this paper further contribute to
its uniqueness among the Mustelidae.
Wright (1969) prepared a list of selected male mustelid reproduc
tive cycles to which I have added seven other authorities and four more
species (Table 6). It is clear, after examination of this table, that
most mus te 1 i d ma 1 es mature before the end of their second year. Enhydra,
on the other hand, does not mature until its fifth year.
Sexual maturation can be viewed as two processes: (1) the full
development of the gonads ~lith concomitant active spermatogenesis and
hormone elaboration; and (2) enlargement of the baculum.
It is difficult to state just when the subadu1t sea otter becomes
sexually mature. Fisher (1939) reported that the "partly grown" males
are the most sexually active. It might be that subadults coming into
active spermatogenesis for the first time, such as sea otter 607 with
its granular Lydig cells, are elaborating enough hormone to stimulate
mating behavior without perhaps the sperm count to effect fertilization.
44

46
Enders (1952) reported that female mink (Mustela vison) bred to yearling
males have a higher percentage of non-pregnancies than when bred to
older males.
Very little histological research has been done on male mustelids.
The bulk of the research in America has been conducted by Wright (1947,
1950, Mustela frenata; 1955, Gulo .9_l!_}_Q_; 1969, Taxidea taxus). He stated
that the testes of yearling long-tailed weasels (~. frenata) showed pri
mary and secondary spermatocytes in synthesis, spermatids and a few
sperm heads forming, but they still possessed an immature-type baculum.
Similarly, male wolverines (~ . .9_l!_}_Q_) just becoming active had a few sperm
in the testes, none in the epididymides and immature baculum. These
characters are identical to subadult testes and bacula I have examined
in sea otters.
The researchers listed in Table 6 feel that in Mustelids, the onset
of sexual maturity is extremely rapid. Friley (1949), referring to re
search conducted by Deanseley (1935) on the stoat (Mustela erminea),
reported that change from the immature to the mature-type baculum takes
place within a period of one month. A similar rapid development in the
baculum of the American badger (Taxidea taxus) was reported by Wright
(1969). He suggests this rapid development (of the baculum) is asso
ciated with high levels of androgen being secreted for the first time in
maturing males. Wright (1947) says that juvenile and young male long
tailed weasels exhibit a "small-type baculum", whereas adults in active
spermatogenesis or captive animals known to have been in active sperma
togenesis in the past, exhibit a large, mature-type baculum. In a later

47
experiment on the same animal, Hright (1950) castrated immature weasels
prior to the onset of puberty. Half of the group, implanted with pel
lets of testosterone propionate, developed adult-type bacula. Unimplanted
males did not develop adult-type bacula, even after several months.
All American mustelids, with the exception of the sea otter, are
seasonal breeders (Asdell, 1964; all authorities, Table 6). That is to
say, they exhibit dimensional and hormonal changes in the testes with the
onset and completion of the mating season. The same is true of male
European muste 1 ids, with the exception of the European badger ( t~e 1 es
meles). Audy (1976) reported that Meles showed active spermatogenesis
throughout the year, though it is sexually active only in January and
February. Testosterone leve 1 s 11ere found to vary throughout the sexua 1
cycle, but peaked once during the mating season and then again during
the sexually quiescent period. He also reported (1975) that the testes
of the European badger did not undergo a well-defined period of involu
tion as did the testes of three other mustelids he had studied. In this
respect, the European badger is very similar to the sea otter.
Hy results seem to indicate also that the;·e is a clear demarcation
between the testes and bacula of the adult and the subadult male sea
otter. The testes of a subadult (and younger) animalscontain non-granular
Lydig cells with a mean area less than 12 ~2 . The adult-type Lydig cell,
in contrast, is large and granular and has a mean area of 62 i. In addi
tion to being granular and larger in area, the adult-type Lydig cells are
also more numerous. Asdell (1964) reported that the interstitial cells

48
of the European weasel (t4ustela nivalls L.) increase to tv/ice their
former diameter during the breeding season and those of the ferret
(Mustela putorius furo L.) are also larger during the breeding season.
Immature-type testes, in addition to having an occasional small,
non-granular cell of Lydig, have arrested spermatogenesis. Pups pos
sess seminiferous tubules lined v1ith Sertoli cells and only have a very
few spermatogonia (Plate 34). Immature animals show development only
to the secondary spermatocyte stage, but primary spermatocytes are
equal in number (Plate 35). The subadult animals still have Sertoli
cells lining the tubules and a few spermatogonia, but meiosis is evi
dent within the lumina. Lydig cells have also increased in number and
taken on a granular texture (Plate 36). At this stage of development,
the bacula are still small (Plate 32) and growth has been concerned
with increasing the length of the os penis. These findings agree with
developmental stages of the other mustelids just discussed.
"fi'IO animals under study are in transition between subadult and
adult. 538 is closer to the younger group, 607 closer to the older.
Both animals possess bacula which were increasing in weight at a faster
rate than length (Figure 9). 607 also had a bulkier, more rugos proxi
mal end. These growth characters are similar to those described by
Friley (1949) for the river otter (Lutra c. canadenesis) in transition
bet1-1een subadult and adult.
The growth curve generated when weight of baculum is plotted against
length (Figure 9) is very similar to the pattern of growth of the os
penis in black bear (Ursus americanus) reported by Rausch (1961;95).

16·
49
-n ~
G') c ;u fT1 (fJ
<.0 0::12 w [(1!!1 >-<(
"' ...1 113 "' n :2: l:l <:: ::J
~ 1- I! <:: z 03 w ;:;: 2 f1) w 8 !ill ~. 0
ll.. 0 (fJ
CT' 0:: 0 w rt rt co ISl 0 2 E3 ::J
z 4
"' ::> ISC L'll*
0- Ill ISl
::> ~ llJ c: 3 CT' f1) -;
0 lm!IEl Uli!J ..., 0
n (])
3 CJ
"' rt <:: E3 28
!!I "' '< f1)
,El .., "' 24
;::; rt
P' PUP El 0 -"' 1 'IMMATURE I:J ~
20 s • SUB ADULT [§] '• ADULT
0 E o • 1\GEO ADULT [li rt a> r+ 01 m z 16 "-
"' 1- ISl
"' I ISl "" (:> ~.
::l w 12 "' 3 rt Ill t:r 2 01 :::J n ..J Ill t:: :::J 8 ~ l) !51 c <(
" m ~ m ::l 4 /:) M-:::r mm .
0 !'EimiEl
0 4 8 12 16 BACULUM LENGTH IN em.
*507, in transition between subadult and adult age categories

50
Sea otters 538 and 607 are plotted at points in the curve where weight
is increasing at a faster rate than length. Rausch feels that this
change in rate probably coincides with the onset of puberty. The testes
of 607 confirm this.
Elder (1951;44), in discussing the baculum as an age criterion in
mink (Mustela_ vison), reported that, "the mink baculum does not reach
its full development the first year following sexual maturity, but seems
to increase considerably in succeeding years". He goes on to say that
there is little overlap in bacula weight between juvenile and adult mink.
14right (1969) reports identical growth patterns for bacula of the Ameri
can badger. My graphs of sea otter bacula (Figures 9-A and 9-B) cor
roborate their findings.
There was only one fully adult specimen with gonads in this study.
In cross-section, its testes contain seminiferous tubules in all stages
of active spermatogenesis. The lumina are filled with free sperm
(Plate 37). His baculum is longer and heavier than those of any other
adult or aged male. This seems to indicate that baculum gr01~th continues
throughout life, as there are aged males in the sample with lighter and
shorter bacula. There are no testes weights on this specimen, but 499,
the other adult, had testes weighing 14 gm each. Schneider (personal
communication, unpublished data) told me that the adult testes of Alas
kan sea otters ile studied v1ere a 11 13 gm or greater.
Wright (1969;441) reports that, "the baculum of the adult male bad
ger is large in proportion to the total body size, being surpassed in

51
American mustelids only in the river otter (Friley, 1949)''. I take
exception to this, since my measurements indicate that the weight of
the adult sea otter baculum increases some 600 times that of the pups'.
Scheffer (1950b;391) stated that, ''the baculum of the mature fur seal
may be over 1 ,500 times as heavy as that of the newborn pup" and that
he knows of "no other mammal having a bone which grows so enormously
during the lifetime of the individual". I reviewed his calculations
and found that he was off one order of magnitude. In actuality, accord
ing to his numbers, the fur seal baculum increases only 150 times its
weight from pup to adulthood and is, therefore, far less impressive
than the baculum of the sea otter.
I encountered great difficulty in attempting to quantify differences
between the adult and the aged testes. There is a possibility that this
is related to the extremely small sample size (1 adult testis, 5 aged
testes) that I was dealing with, but I prefer to think otherwise.
In spite of the small sample size, all material was very fresh and
changes resulting from decomposition could not have taken place. Sea
otter 550, when found, was nearly dead from shark wounds. Upon death,
the testes were immediately removed and placed in fixative, so little
error could have resulted from my observations of the testes. Likewise,
the aged adult specimens were collected around Monterey wharf, where
freshly dead and dying animals could be immediately collected and pro
cessed.
Sea otter 499 presented a curious problem. Although his cranial

52
development, bacular length and testis weight were adult, his bacular
11eight was clearly subadult. It seems apparent that he had some hor
monal difficulties, since testosterone is linked to increasing bacular
weight at onset of puberty. In addition, Dr. Cornell (personal communi
cation) of Sea Wm'ld San Diego mentioned that this animal showed very
little interest in sex.
It is clear that separation of the adult and aged adult groups
could not be achieved mathematically (see Results, page ). Histologi
cal examination provided the key to separation. As the aged male testis
became less functional in terms of amount of spermatogenesis observed
and spermatid and sperm production, the seminiferous tubules became
longer and more narrow. Since Bigg's (1969) formula for seminiferous
tubule diameter included a summation of length and width, the diameter
remained constant even as the tubules were collapsing (Plate 41).
Senescence in the sea otter testes is indicated by (1) a lack of
organization to the spermatogenic cycle; (2) reversion to the immature
condition; and (3) proliferation of what I term a "Lydig cell complex"
(Plate 41).
This lack of organization is clearly seen in Plate 40, where the
reproductive cells are scattered within the seminiferous tubule rather
than being ordered with younger spermatogenic stages close to the tubule
~tall and later stages grading toward the lumen center as in the adult
testis (Plate 37).
Reversion to the immature condition in seasonal mustelids is well

53
documented in previous studies shown in Table 6. There is no reason to
doubt that this also occurs in senescent males, as Plate 39 indicates.
\•right (1969) is the only one that referred to Lydig cell build-up
as shown in Plate 41. He described extensive areas of adrenal cortex
like interstitial tissue occurring along the tunica albuginea and in the
region of the rete testis in all male American badgers studied. The
Lydig cell complexes of aged sea otter testes were also concentrated in
these areas.
Senescence is not a subject widely reported in the literature.
Benkovsky (1975), in categorizing developmental periods for the (river)
otter on Sakhalin Island, observed a decline in reproductive potential
after six or seven years. He .went on to say that the teeth of o 1 der
otters displayed hypercementose, a fact 1 observed with the sea otter as
well. He also reported, at least as far as musculature was concerned,
that "sable, American mink, Ussurian raccoon-dogs, fox and other, older
males became feminized". Benkovsky's paper also referred (page 26) to
similar findings by Bromly (1964, 1965) for Ussurian wild boars and
bears and by Gentner (1967) for other predators.
Crowe (1972) uses "senectitude" as an age class category for bob
cat with between 6 and 9 cementum annuli. Ranch mink were found by
Elder (1951) to experience a decline in breeding vigor between 3-l/2 and
5-l/2 years of age. litter sizes of the Siberian sable (Martes zibel
lina 1.) were also shown by Asdell (1964) to decrease after ten years of
age.

54
Karl Schneider (personal communication) wrote to me that he had
recently processed two sea otters found dead on the beach. Both were
adults, but one was quite old. No sperm was found in the epididymal
fluid of either animal and the testes of both (10.8 gm and 12.6 gm)
were below the 13 gm weight he measured (unpublished data) for adult
sea otters in active spermatogenesis.
Teeth and Tooth Studies
It should be emphasized that all known research to date on infant
and immature sea otters relating to growth and development has centered
around relative age studies using cranial development and tooth eruption
sequences.
As noted in the Results section, 90% of the pups in my sample were
born between October and February. If this is a representative sample
of the true population (and not a biased subsample of weak animals not
strong or healthy enough to survive their first winter), then I can con
clude that a· pup is somewhere between nine and twelve months old when
the root canal closes. To test this assumption, I plotted the root canal
diameter of sea otter 527, the known-age female pup, onto Figure 7 to
compare her degree of closure with that of the male pups estimated to
be in her age category.
Although it would seem that the root canal of 527 is closing at a
faster rate than the males, she is still close enough to the rate of
closure line to agree with my estimate. If anything, her rate of closure
would bring the estimate back one month or closer to that of Schneider's

55
(1973). He has determined that sea otter pups in Alaska must be at
least eight months old and possess a certain amount of light-colored
(opaque) cementum before the first dark (translucent) layer is deposited
which he estimates to occur between eigQt and nineteen months of age.
Klevezal and Marakov (1966) simply say that cementum begins to form
only on post-embryonic sea otter teeth, but that is of little help in
solving this problem.
The American badger (Taxidea taxus) does not lay down its first
annulus until 14 to 19 months of age (Wright, 1969; Crowe and Strickland,
1975). The bobcat (Lynx rufus) and the coyote Can two other
small terrestrial carnivores, lay down their first annulus at between 20
and 23 months of age (Crowe, 1972; Linhart and Knowlton, 1967, respec
tively) as does the black bear (Ursus americanus, Sauer, {L. 1966).
The coyote, like the sea otter, has its permanent canines erupting
around four to five months of age. Linhart and Knowlton (1967) have ob
served that its root canal closes around eight to nine months (similar
to the sea otter) and the first opaque layer forms around the twentieth
or twenty-third month. Perhaps the coyote's similarity to the sea otter
in tooth eruption, weight at maturity and life span is also reflected in
the cementum deposition patterns.
It seems obvious that the question of just when the first annulus
is forming will not be answered until data are gathered on tagged, known
age specimens (Schneider, 1973). It is pretty much an accepted procedure,
however, to add one to the number of annuli counted (Crowe, 1972; Linhart
and Knowlton, 1967; Marks and Erickson, 1966; McEwan, 1963; Mundy and

Fuller, 1964; Rausch, 1961; Sauer,
1959; Sergeant, 1967), as I have.
56
al. 1966; Sergeant and Pimlott,
Teeth from animals in the subadult relative age category are very
easy to identify. As alt·eady mentioned, the occlusal surfaces show
little wear; the root apex is still porous and cemental deposition is
limited to the apical region of the root. As the animal becomes older,
cementum deposition extends up the root neck toward the gum line. A
similar pattern of mineralization is seen in the premolar teeth of
dogs (Owens, 1975) and the canines of coyotes (Linhart and Knowlton,
1967).
All subadults, except sea otter 538, have between 3 and 3-l/2 cemen
tum annuli or are between 4 and 4-1/2 years old. The borderline sub
adult-adult, 538, has between 3 and 5 annuli, depending upon how and
where the count is made (see Plate 18). This discrepancy could be ex
plained by the fact that (1) the animal was slow to mature, or; (2)
the lines counted are similar to the "double lines" in Alaskan sea otter
teeth (Schneider, 1973) and the "false separating zones" or "multiple
lining" frequently occurring in bear teeth (Craighead, et .'!.l· 1970;
Sauer, et E]_. 1966). The most likely explanation, however, could simply
be that prior to physical and/or sexual maturation, the annuli are wide
apart, often discontinuous and usually difficult to read (Craighead, ei:.
al. 1970; Crowe, 1972; Crowe and Strickland, 1974; Laws, 1953; Linhart
and Knowlton, 1967; ~1undy and Fuller, 1964; Schneider, 1973; Sauer,~
a 1. 1966, and others).
Except for sea otter 607, a borderline adult with 3-1/2 annuli,

57
adult sea otters in this study have 6-l/2 or more annuli. As the otter
increases in age, the root width and greatest cementum width also in
crease, but not in a linear fashion (see Table 2) as proposed by Ser
geant and Pimiott (1959) for the moose (Alces americanus). These char
acters, therefore, are not acceptable here as a relative or actual age
guide.
Wear also increases with age, but degree of wear does not neces
sarily correlate directly with it. For instance, 590 is classified as
an aged adult. His cementum width is exactly midway between the widths
of all other adult and aged animals (Table 2). Furthermore, he shm·;s a
moderate amount of wear on all of his teeth (Plate ), but there are
aged specimens ~lith less wear. Since the amount of tooth wear and perio
dontal disease in the sea otter (Fisher, 1941; Kenyon, 1969; Klevezal
and Marakov, 1966) and other species (Hooper and Ostenson, 1949; McE1·1an,
1962; Ransom, 1966; Rausch, 1961; Thomas and Bandy, 1975) is related to
diet and not necessarily age, it cannot be used as an aging criterion.
One result relating to teeth and tooth studies needs discussing. I
refer to the low annuli count obtained from sea otter 590, an aged adult.
I have no reasonable explanation for this low count, except perhaps to
suggest that either he matured earlier than most otters or that at some
point during cementum bulb development, deposition may have been inter
rupted. It is well knovm that trauma to the periodontal membrane will
result in disruption of cementum deposition (Gilbert, 1966; Lov1 and Cowan,
1963; both refer to Ham and Leeson, 1961).

58
Most researchers using cementum annuli choose to decalcify and
stain thin sections, rather than to grind and view undecalcified teeth
as I have done. All decalcifying agents tried by me, hov1ever, failed
·to produce adequate results. I can only assume that either the Cali-.
fornia sea otter teeth are inherently different from those of other
mammals or that my technique is suspect. It is gratifying to report
that other researchers (Fisher and Mackenzie, 1954; Frost, 1958; Gaskin
and Blair, 1977) have also obtained usable results with undecalcified,
unstained tooth sections.
There is no general agreement as to why or how the dentin and
cementum annuli form. Schour (1939;91) writes, "Contour lines in enamel
and dentin may reflect the nutritional and metabolic variations that
occur during the growth and calcification of dental tissues''. Laws
(1950) suggests that in the elephant seal (Mirounga leonina), these
annuli result from changes in calcium metabolism and parathyroid activ
ity due to the different amounts of vitamin 0 available ~1hen the seal
is out of the water and fasting during breeding and moulting cycles.
Low and Cowan (1963) propose that the formation of cementun annuli in
deer (Odocoileus hemionus) is related to variations in calcium uptake.
Mansfield and Fisher (1960) suggest that annuli are related to minimal
feed during breeding and moult in the harbor seal (Phoca vitulin~).
Sergeant and Pimiott (1959) relate them in the moose to feeding cycles
or endocrine rhythms for caribou (Rangifer tarandus) McEwan ( 1963)
referred to seasonal differences in the amount of light and nutritional
quantities. Finally, Stoneberg and Jonkel (1966) found that in each

animal studied, the researcher attributes ''layer formation on some
seasonal metabolic disturbance peculiar to that animal''. They go on
59
to suggest that, ''the final answer to the problem of what causes dif
ferential cemental development rests with further investigation'', and I
agree.
More in-depth studies relating to the chemistry and development of
cementum formation may be found in Chovelton, Carmichael and Pearson
(1975), Owens (1975) and Selvig (1964, 1965).

CHAPTER 5
CONCLUSION
The existence of five relative age classes as reported by t~orejohn,
et _i!l. (1975) for the sea otter is confirmed by using cementum annuh
counts and testicular and bacular development.
My studies indicated that the root canal of permanent canine teeth
closes between eight and twelve months of age at a rate of approximately
3 mm in two months and that the first translucent zone is deposited dur
ing the "irmwtures"' second winter. "hen using the number of cementum
annuli as an indicator of chronological age, therefore, one must be
added to account for the time elapsed during tooth eruption, root canal
closure, and build-up of enough opaque cementum to precede translucent
cementa] deposition.
Sexual maturation occurs some time during the subadults' fifth year,
but sexual behavior may precede this. I have determined that sexual po
tency can continue past twelve years of age.
Senescence, indicated by cessation of spermatogenesis and regression
of the sex cells to the "immature" condition, was observed in animals
with as few as 6-1/2 cementum annuli. Counts as high as 13 to 16 annuli
v1ere made. The testes of several senescent males contained areas of ex
treme Lydig cell proliferation.
The sea otter baculum develops similarly to other mustelids, but is
enormously larger in proportion. Prior to the onset of active spermato
genesis, there is an immature type baculum. As sexual maturation {puber
ty) approaches, baculum weight increases at a faster rate than 1 ength.
It appears that the baculum continues to grow throughout life.
60

LITERATURE CITED
Asdell, S. A. 1964. Mustelidae, pp 449-471. In: Patterns of manunalian reproduction. 2nd ed. Cornell Uni v. PresS":"""
Audey, M. C. and M. Bonnin-Laffargue. 1975. The endocrine activity of the European badger's testis (Meles meles L.). (In French, English summary) Arch. Biol. 86(2):223-232.
Audy, M. C. 1976. Seasonal sex cycle of male European mustelids. (In French, English summary) Gen. Camp. Endocrinol. 30(1):117-127.
Barabash-Nikiforov, 1947. The sea otter. Transl. from Russian. Published for the National Science Foundation, Hashington, D.C. by the Israel Program for Scientific Translations, Jerusalem, 1962. 227 pp.
Ben'Kovskii, L. M. halin Island. Zool. 2:20-27.
1975. Age related development of the otter from Sak(In Russian, Russian and English summary) Vestn.
Bigg, M. A. 1969. The harbor seal in British Columbia. Fish. Res. Board Can. Bull. 172, 33 pp,
Bishop, R. H. 1967. Reproduction, age determination and behavior of the harbor seal, Phoca vitulina L., in the Gulf of Alaska. Master's Thesis, Univ. of Alaska, 121 pp.
Cassie, R. M. 1954. Some uses of size frequency distributions. 5(3):510-522.
probability paper in the analysis of Aust. J. Mar. Fresh-water Res.
Chovelton, A., D. J. Carmichael and C. H. Pearson. 1975. The composition of the organic matrix of bovine cementum. Arch. Oral Biol. 20(8): 537-542.
Christian, J. J. 1956. the big brown bat, 66-95.
The natural history of a summer aggregation of Eptericus fuscus. Amer. Midland Natur. 55(1):
Craighead, J. J., F. C. Craighead and H. E. McCutchen. 1970. Age determination of grizzly bears from fourth premolar tooth sections. J. Hildlife Manage. 34:353-363.
Crowe, D. M. layers.
1972. The presence of annuli in bobcat tooth cementum J. Hildlife Manage. 36:1330-1332.
Crowe, D. M. and M. D. Strickland. 1974. Dental annulation in the American badger. J. Mammalogy 56(1):269-272.
61

62
DeBlase, A. F. and R. E. Martin. 1974. to families of the world. William 323 pp.
A manual of mammalogy with keys C. Brown Co., Dubuque, Iowa,
Elder, W. H. 1951. The baculum as an age criterion in mink. J. Mammalogy 32(1):43-50.
Enders, R. K. 1952. Reproduction in the mink (Mustela vison). Proc. Amer. Phil. Soc. 96:691-755.
Fisher, E. M. 1939. Habits of the southern sea otter. J. Mammalogy 20(1) :21-37.
Fisher, E. M. 1941. Notes on the teeth of the sea otter. J. Mammalogy 22(4):428-433.
Fisher, H. D. and B. A. Mackenzie. tions for age determination.
1954. Rapid preparation of tooth secJ. Wildlife Manage. 18(4):535-537.
Friley, C. E., Jr. 1949. Age determination, by use of the baculum, in the river otter Lutra c. canadensis Schreber. J. Mamma logy 30(2): 102-110.
Frost, H. M. 1958. Preparation of thin, undecalcified bone sections by rapid manual methods. Stain Technol. 33(6) :273-277.
Galigher, A. and E. M. Kozloff. 1964. Essentials of practical microtechnique. Lea and Febiger, Philadelphia, 484 pp.
Gambell, R. and C. Grzegorzewska. 1967. The rate of lamina formation in sperm whale teeth. Norsk Hvalfangst-Tidende 56(6):118-121.
Gaskin, D. E. and B. A. Blair. 1977. Age determination of harbour porpoise, Phocoena phocoena (L.), in the western North Atlantic. Can. J. Zool. 55(1) :18-30.
Gilbert, F. F. 1966. Aging white-tailed deer from annuli in the cementum of the first incisor. J. Wildlife Manage. 30(1):200-202.
Grau, A. G., G. C. Sanderson and J. P. Rogers. 1970. of raccoons. J. Wildlife Manage. 34(2):364-372.
Age determination
Gurr, E. 1962. Staining animal tissues practical and theot·etical. Leonard Hill Ltd.·, London, 631 pp.
Harding, J. P. 1949. The use of probability paper for the graphical analysis of polymodal frequency distributions. J. Mar. Biol. Ass. U.K. 28:141-158.

63
Hildebrand, M. 1954. Incisor tooth wear in the sea otter. J. Mammalogy 35(4):595.
Hooper, E. T. and B. T. Ostenson. 1949. Age groups in Michigan otter. Occas. Pap. Mus. Zool. Univ. Mich., 518, 22 pp.
Humason, G. L. 1967. Animal tissue techniques. 2nd ed. H. W. Freeman and Co., San Francisco, 569 pp.
Kenyon, K. W. 1969. The sea otter in the Eastern Pacific Ocean. U.S. Fish. Wildlife Serv. N. Amer. Fauna 68, 352 pp.
Klevezal, G. A. and S. E. Kleinenberg. 1967. Age determination of mammals from annual layers in teeth and bones. Transl. from Russian. Published for the U.S. Department of Interior and the National Science Foundation by the Israel Program for Scientific Translations, Jerusalem, 1969. 128 pp.
Klevezal, G. A. and S. V. Marakov. 1966. Age of the sea otter (in Russian) Sb. Nauch-Tekhn. Inform. VNIIZhP 16. 3 pp.
Laws, R. M. 1952. A new method of age determination for mammals. Nature (London) 169:972-973.
Laws, R. M. 1953. A new method of age determination in mammals with special reference to the elephant seal (Mirounga· leonina Linn.). Falkland Isl. Dependencies Surv. Sci. Rep. 2:1-ll.
Lensink, C. J. 1962. The history and status of sea otters in Alaska. PhD Thesis, Purdue Univ., 188 pp.
Linhart, S. B. and F. F. Knowlton. 1967. Determining age of coyotes by tooth cem~ntum layers. J. Wildlife Manage. 31(2):362-365.
Low, W. A. and I. MeT. Cowan. 1963. Age determination of deer by annular structure of dental cementum. J. Wildlife Manage. 27(3):466-471.
Mansfield, A. W. and H. D. Fisher. 1960. Age determination in the harbor seal, Phoca vitulina L. Nature (London) 186:92-93.
Marks, S. A. and A. W. Erickson. 1966. Age determination in black bear. J. Wildlife Manage. 30(2):389-410.
McEwan, E. H. 1963. Seasonal annuli in the cementum of the teeth of barren ground caribou. Can. J. Zool. 41(1):111-113.
Miller, D. J. 1974. The sea otter Enhydra lutris, its onomic status, and some ecological relationships. Game Mar. Resour. Leaf. 7. 13 pp.
life history, taxCalif. Dept. Fish

~1orejohn, G. V., J. A. Ames and D. B. Lewis. 1975. of sea otters, Enhydra lutris L., in California. Game ~1ar. Resour. Tech. Rep. 30. 82 pp.
64
Post-mortem studies Calif. Dept. Fish·
Mundy, K. R. D. and l~. A. Fuller. 1964. Age determination in the grizzly bear. J. Wildlife Manage. 28(4):863-866.
Owens, P. D. A. 1975. Patterns of mineralization in the roots of premolar teeth in dogs. Arch. Oral Bio1. 20:709-712.
Peterson, R. S. and N. W. Odemar. 1969. Population growth of the sea otter in California: Results of aerial censuses and behavioral studies. Pages 69-72 In: Proceedings of the Sixth Annual Conference on Biological Sonar an~Diving Mammals. Stanford Research Institute, Menlo Park, California.
Ransom, A. B. 1966. Determining age of white-tailed deer from layers in cementum of molars. J. Wildlife t1anage. 30(1):197-199.
Rausch, R. 1961. Notes on the black bear, Ursus americanus Pallas, in Alaska, with particular reference to dentition and growth. Z. Saeugetierk 26:77-107.
Roest, A. I. 1971. A systematic study of the sea otter (Enhydra lutris). Pages 133-135 In: Proceedings of the Eighth Annual Conference on Biological Sonar and Diving Mammals. Biological Sonar Laboratory, Marine Mammal Study Center, Fremont, California.
Sauer, P. R., S. Free and S. Browne. 1966. Age determination in black bears from canine tooth sections. N.Y. Fish Game J. 13(2):125-139.
Scheffer, V. B. 1950a. Growth layers on the teeth of pinnipedia as indication 6f age. Science 112(2907):309-311.
Scheffer, V. B. 1950b. Grov1th of the testes and baculum in the fur seal, Callorhinus ursinus. J. t4ammalogy 31 (4) :384-394.
Scheffer, V. B. and R. S. Peterson. 1967. Growth layers in the teeth of suckling fur seals. Growth 31:35-38.
Schneider, K. B. 1973. Age determination of sea otter. Alaska Dept. Fish Game, Federal Aid in Wildlife Restoration, Final Report, Projects W-17-4 and W-17-5, Job B. lOR, 23 pp.
Schour, r. and t1. t4. Hoffman. 1939. Studies in tooth development. I. The 16 microns calcification rhythm in the enamel and dentin from fish to man. J. Dent. Res. 18:91-102.
Selvig, K. A. 1964. An ultrastructural study of cementum formation. Acta Odontol. Scand. 22:105-120.

65
Selvig, K. A. 1965. The fine structure of human cementum. Acta Odontol. Scand. 23:423-441.
Sergeant, D. E. 1967. Age determination of land mamw~ls from annuli. Z. Saeugetiet·k 32(5) :297-300.
Sergeant, D. E. and D. H. Pimlott. 1959. Age determination in moose from sectioned incisor teeth. J. Wildlif~ Manage. 23(3):315-321.
Stone, W. B., A. S. Clauson, D. E. 51 ingerlands and B. L. Weber. 1975. Use of Romanowsky stains topt·epare tooth sections for aging mammals. N.Y. Fish Game J. 22(2):156-158.
Stoneberg, R. P. and C. J. Jonkel. 1966. Age determination of black bear by cementum layers. J. Wildlife Manage. 30(2):411-414.
Taylor, \~. P. 1914. The problem of aquatic adaptation in the carnivora, as illustrated in the osteology and evolution of the sea otter. Univ. Calif. Publ. Geol. 7:465-495.
Thomas, D. C. and P. J. Bandy. 1975. Accuracy of dental-11ear age estimates of black-tailed deer. J. Nildlife Manage. 39(4):674-678.
Wright, P. L. 1947. The sexual cycle of the male long-tailed weasel. J. Mammalogy 28(4):343-352.
Wright, P. L. 1950. Development of the baculum of the long-tailed weasel. Proc. Soc. Exp. Biol. Med. 75:820-822.
Wright, P. L. 1969. Reproductive eye l e of the rna le American badger (Taxidea taxus). J. Reprod. Fert., Suppl. 6:435-445.
Wright, P. L. and R. Rausch. 1955. Reproduction in the wolverine, Gulo ~· J. 14amma 1 ogy 36 ( 3): 346-355.

APPENDICES
66

so
APPENDIX A
SAMPLE DATA SHEET
67
Date collected ______ By ________ _
Where found ---------------------------
Age Weight ___ _ CBL __ _
Sex Total length _____ _ zw r1L ___ _
Condition --------------------------
Remarks ---------------------------
Prior to processing, record:
Permanent ~- J .. L2_[ ___ 3
PM r 'I I Upper
M I
2
pm_ ·-----+·· 1--· I i I I
l±t -...... .. ::::::::: :~ l r : ---- I - ~-~
! : Lower
NE E/1 E/2 E 0 p w
• 1 1 j I c
not erupted through bone erupted through bone, not gum eruption 1/4 to 3/4 complete fully erupted deciduous tooth shed permanent tooth in place wear indicated
Skull measurements:
~-- ... J .. PM M! ,
·t--·1· pm
Canine alveolar length (anterior-posterior) __________ _
Maxillary tooth row length (pc and molars) ___________ _
Greatest width between canines-----------------
Mandibular angle ________ Tooth taken and weight ____ _
Sexual data:
Baculum weight _______ Baculum length _________ _
Degree of apparent sexual development and activity and comments

APPENDIX B
FREQUENCY DISTRIBUTION OF BACULAR LENGTHS
length (mm) Pups lrr:matures Subadul ts Adults Aged Adults
Below 20 4 21-25 26-30
31-35 36-40 41-45 46-50
51 56-60 61-65 1 66-70 1 71-75
76-80 81-85 86-90
91-95 96-100 ~
101-105 106-110 3
111-115 1
116-120 121-125 1 126-130 131-135 136-140 141-145 1
146-150 1 4
151-155 1 2
155-160 1
68

69
APPENDIX B (Continued) FREQUENCY DISTRIBUTION OF BACULAR WEIGHTS
Weight (gm) Pups Immatures Subadults Adults Aged Adults
<1 4
1-2 2
3-4 5-6 2
7-8 2
9-10 ll-12
13-14 15-16 1 1
17-18 1 19-20 4
21-22 2
23-24 25-26
27-28

70
APPENDIX B (Continued)
FREQUENCY DISTRIBUTION OF CONOYLOBASAL LENGTHS
Width (mm) Immature Subadult Adult Aged Adult
108 1
109 110
111 1
112
113 114 115
116
117 118
11 g
120 1
121 2
122 123
124 125 126 1 127 128 2
129 1 2
130 1
131 1 2
132 1
133 1
134 1

71
APPENDIX B (Continued)
FREQUENCY DISTRIBUTION OF ZYGOMATIC WIDTHS
Width (mm) Immature Subadult Adult Aged Adult
81
82
83
84
85
85 1
87
88
89
90
91
92 1
93
94
95 2
96
97 1
98 l 99 1 100 1
101
102 1 2
103 104 2
105 1
106 1

72
APPENDIX B (Continued)
FREQUENCY DISTRIBUTION OF MANDIBULAR LENGTHS
Width (mm) Pups Imma tures Subadu1ts Adults Aged Adults
40-42
43-45 2
46-48 49-51 5
52-54 55-57 58-60 61-63
64-66 67-69 1
70-72 73-75 76-78 1
79-81
82-84 3 1
85-87 3 2
88-90 2 4

APPENDIX C
PLATES OF SEA OTTER SKELETAL MATERIAL
AND GONADAL TISSUES
73

PLATE 1: r~axillaries and dentary of 539 (above). ~1axillaries and dentary of 544 (below). Both animals are "birth" aged. Top of photograph is anterior.


PLATE 2: Maxillaries and dentary of 545 {above) and 540 (below). Sea otter 545 is ''birth'' aged; 540 is aged birth to 1 month. Top of photograph is anterior.


PLATE 3: Maxillaries and dentary of 543, aged birth to 1 month (above). Maxillaries and dentary of 493, aged 2 to 3 months (below). Top of photograph is anterior.


PLATE 4: Maxillaries and dentary of 536, a 2 to 3 month old (above); 490 a 4 to 5 month old (below). Top of photograph is anterior.
! ) '

I ! ' '

PLATE 5: Dorsal (above) and ventral (below) views of the skull, dentary and baculum of 495, an immature animal.
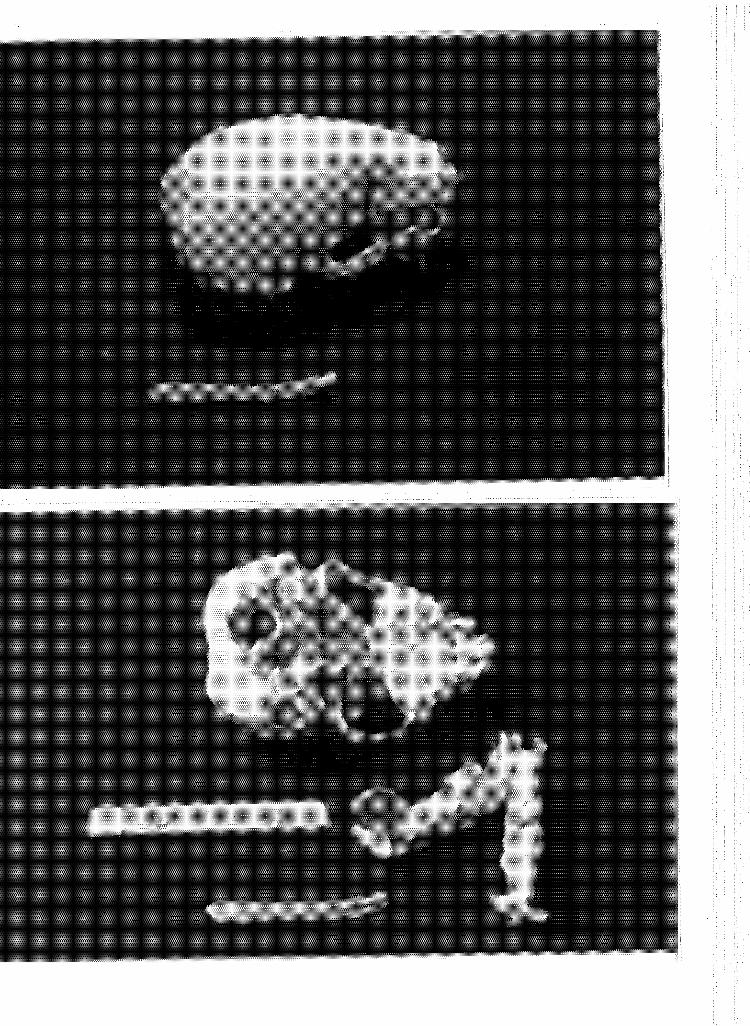

PLATE 6: Dorsal (above) and ventral (below) views of the skull, dentary and baculum of 553, an immature animal.


PLATE 7: Dorsal (above) and ventral (below) views of the skull, dentary and baculum of 593, a subadult animal.


PLATE 8: Longi-section of root apex, upper right canine of 593, a subadult. Note the 3+ translucent zones.


PLATE 9: Dorsal (above) and ventral (below) views of the skull, dentary and baculum of 529, a subadult animal.


PLATE 10: Longi-section of root apex, upper right canine of 529, a subadult. Note 3-3-l/2 translucent zones and multiple lining indicated by split arrows.


PLATE 11: Dorsa1 (above) and ventral (be1ow) views of the sku11, dentary and bacu1um of 497, a subadu1t animal.


PLATE 12: Longi-section of root apex, upper right third incisor (above) of 497, a subadult. Longi-section of root apex, upper right canine {below) of the same animal. Note 3-1/4 translucent zones in both photographs.


PLATE 13: Dorsal (above) and ventral (below) views of the skull, dentary and baculum of 592, a subadult animal.


PLATE 14: Longi-section of root apex, upper right canine of 592, a subadult. Note 3+ translucent zones.


PLATE 15: Dorsal (above) and ventral (below) views of the skull, dentary and baculum of 520, a subadult animal.


PLATE 16: Longi-section of root apex, upper right canine of 520, a subadult. Note how difficult it is to read the annuli.


PLATE 17: Dorsal (above) and ventral (below) views of the skull, dentary and baculum of 516, a subadult animal.


PLATE 18: Longi-section of a 1 ate subadult. near the apex, 5
root apex, upper right· canine of 538, Note that 3 annuli can be counted
as you move up the root neck.


PLATE 19: Dorsal {above) and ventral {belmv) views of the skull, dentary and baculum of 538, a late subadult.


PLATE 20: Dorsal (above) and ventral (below) views of the skull, dentary and bacul urn of 550, an adult animal.


PLATE 21: Longi-section of root apex, upper right canine of 550, an adult. ll-l/2 annuli are visible.


PLATE 22: Dorsal (above) and ventral (below) views of the skull, dentary and baculum of 590, an aged adult.


PLATE 23: Dorsal (above) and ventral (below) views of the skull, dentary and baculum of 521, an aged adult. Note the extreme sagittal and lambdoidal crest development and tooth wear.
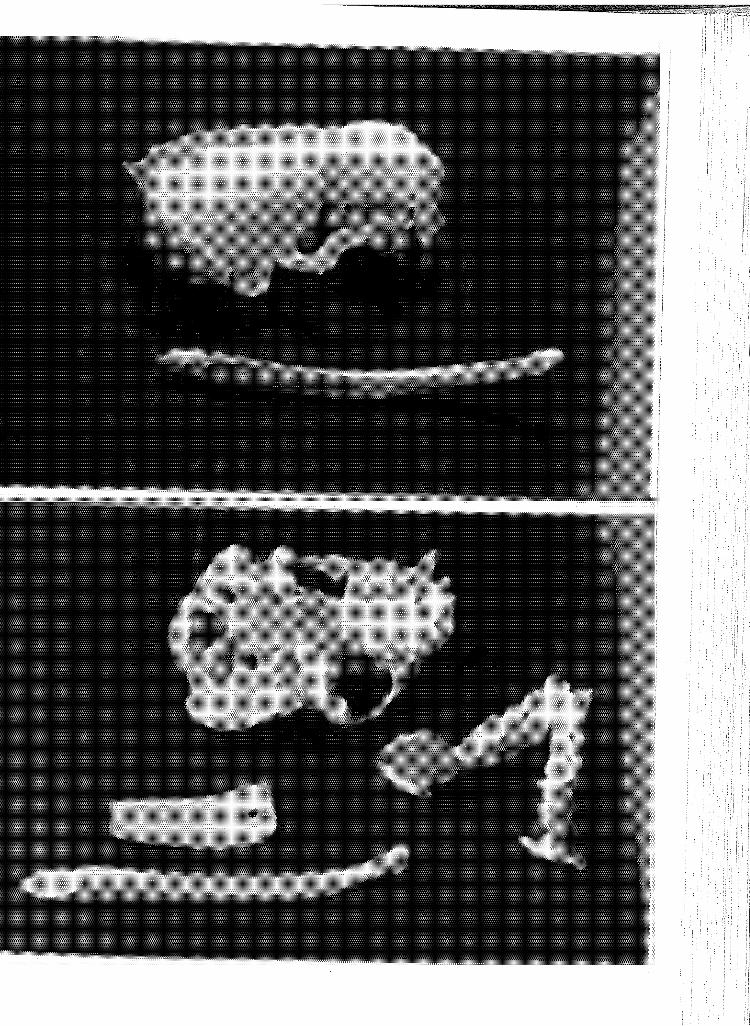

PLATE 24: Dorsal (above) and ventral (below) views of the skull, dentary and baculum of 509, an aged adult. Note necrosis of the lower left canine alveolus.
"~~~. '
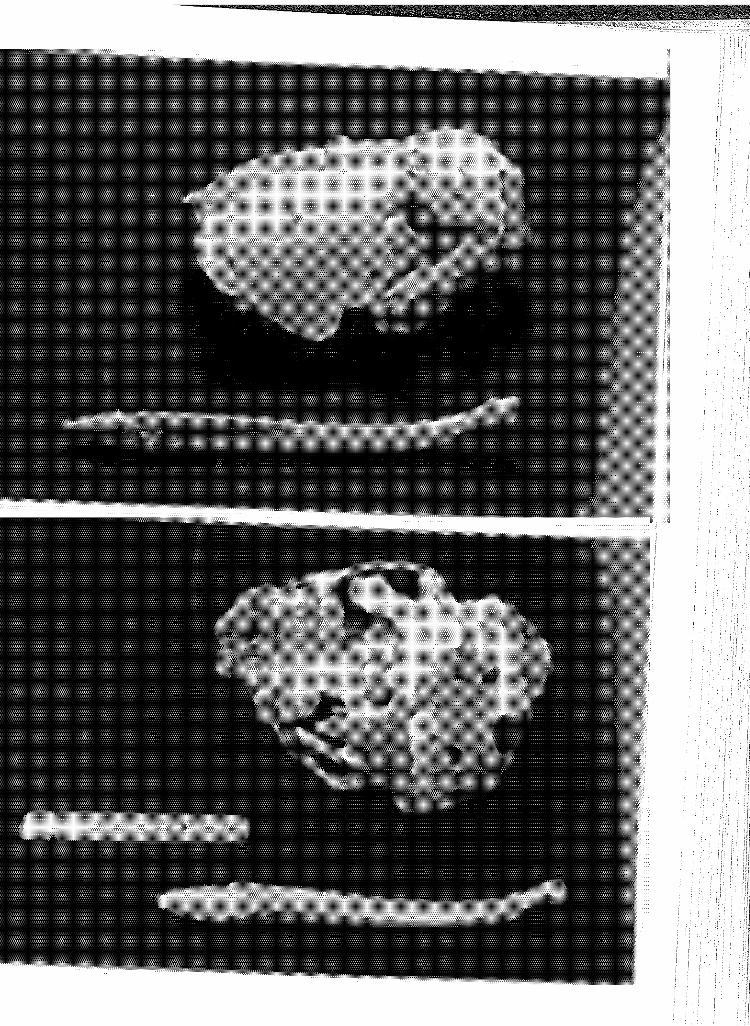
I I
I '!i 'i!

PLATE 25: Dorsal (above) and ventral (below) views of the skull, dentary and baculum of 528, an aged adult. Note dense deposition of cementum up the root neck of the extracted upper canine (lower photograph).
! i '
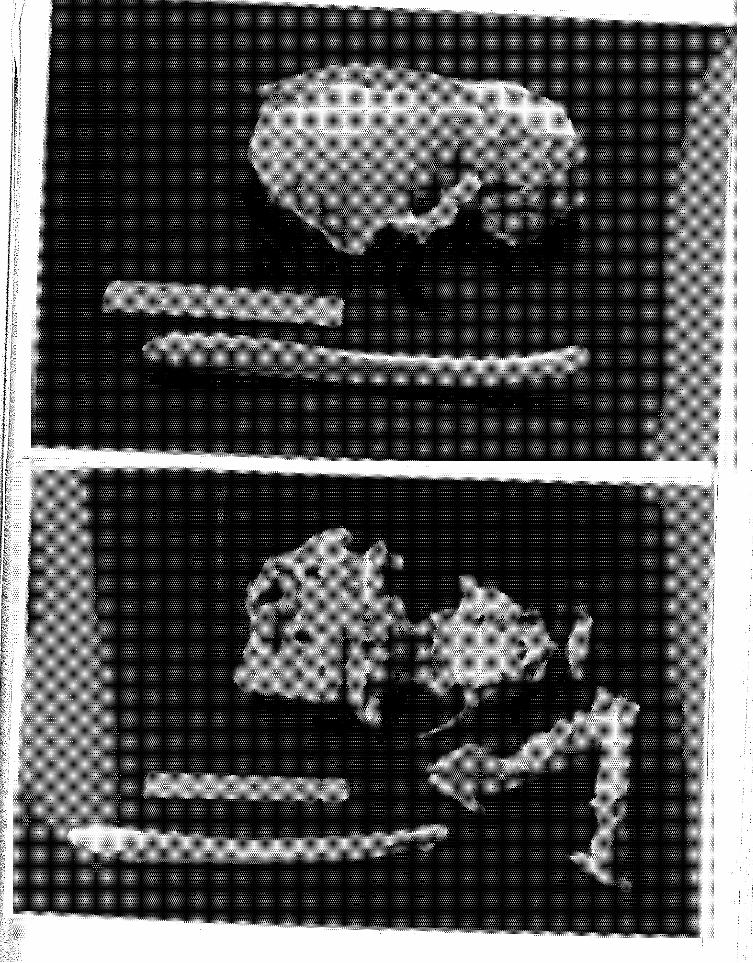

PLATE 26: Dorsal (above) and ventral (below) views of the skul1 fragments, dentary and baculum of 535, an aged adult. Note tooth 1~ear and caries, severa 1 of 1~hi ch extend into the root canals.
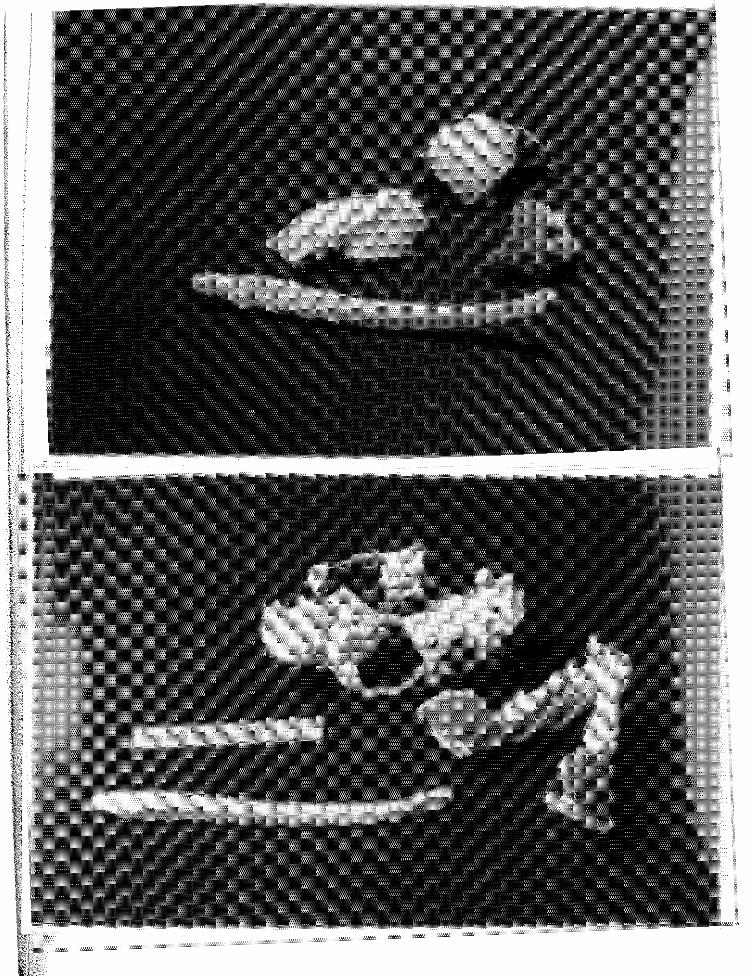

PLATE 27: Longi-section of root apex, upper right canine of 535, an aged adult. 10-l/2 annuli were counted.


PLATE 28: Dorsal (above) and ventral (belmv) viev1s of the skull, dentary and baculum of 515, an aged adult. Note upper left molar alveolus is filled in with.alveolar bone. Canines are v1orn to the gum line.


PLATE 29: Longi-section of root apex, left upper third incisor of 515, an aged adult. 16 translucent zones are echinochrome-stained.


PLATE 30: Dorsal (above) and ventral (below) vie\vs of the skull, dentary and baculum of 577, an aged adult. Note the extreme tooth wear.


PLATE 31: Longi-section of root apex, upper right canine of 577, an aged adult. There are 13 to 16 annuli, depending upon ~there the count is made.


PLATE 32: Bacula from pups, immature and subadult sea otters (from bottom to top).


PLATE 33: Bacula from one subadult (538, bottom), one adult (550, top) and seven aged adults. Note rugosity of proximal ends (left).

I if r: I ;r

PLATE 34: Cross-sectional view of the testis of pup 543 showing septuli testis (ST) forming a thin partition which divides the testis into lobules. H&E stain x 125. (sc=Sertoli cell; sg=spermatogonia).


PLATE 35: Cross-sectional view of the testis of immature sea otter 553 showing a portion of the tunica albuginea (ta) overlying the seminiferous tubules. H&E stain xl25. (sg= spermatogonia; ps=primary spermatocyte; ss=secondary spermatocyte; L=Lydig cell).


PLATE 36: Close-up view (x500) of basef'lent mel'lb.rane (bm) of a seminiferous tubule and Lydig cells (L) of sea otter 529, a subadult, H&E stain. (sg=spermatogonia; sc=Sertoli cells; ss= sec.ondary spermatocyte in meiosis).

1 I
I
PLATE 37: Adult testis of sea otter 550 showing basement membrane (bm) separating seminiferous tubule and Lydig cells (L). Note large number of spermatogenic cells in various stages of spermatogenesis and a Sertoli cell (sc) cytoplasmic extension extending toward lumen (Lu). H&E stain x500. (sg;spermatogonia; ps;primary spermatocytes; ss; secondary spermatocytes; s;maturing sperm aligned to Sertoli cell).
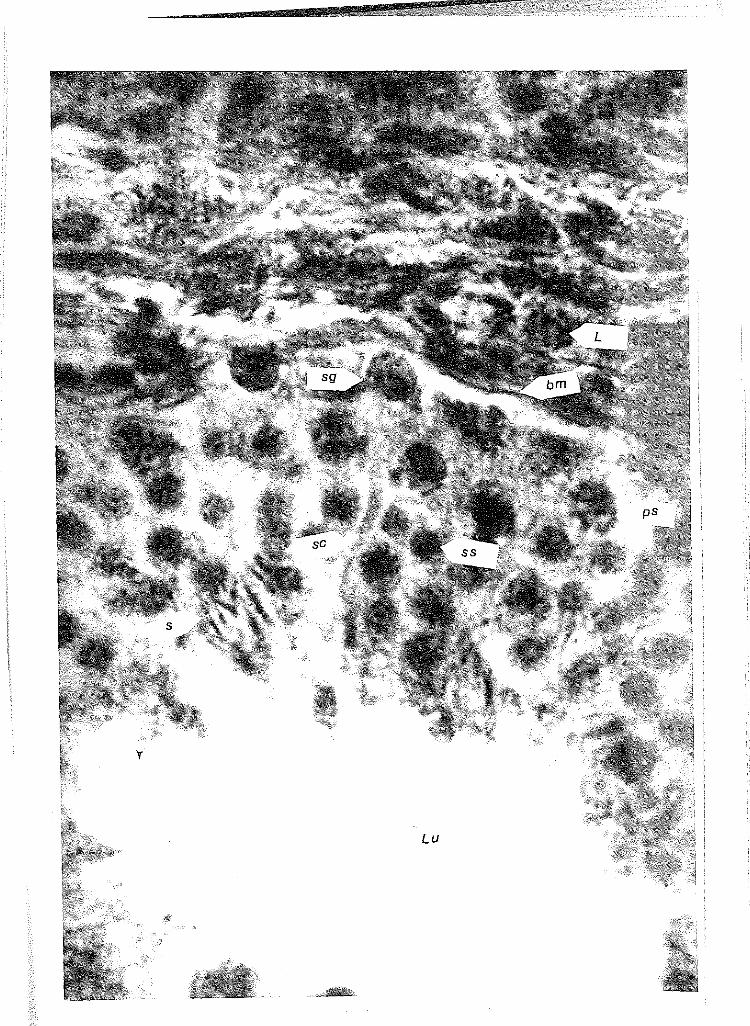

PLATE 38: Cross-section of one and a half epididymal canals within epididymis of 550, an adult sea otter, showing a high concentration of sperm (s) in canals and cilia (c) of pseudostra~ified steriocilia cells extending into canal to move sperm along. H&E stain xl25.

l : ~
I ~ I
' h 1: ' , II !I
~ !
1

PLATE 39: Cross-section of aged testis of 494 showing high numbers of Sertoli cells (sc) and occasional spermatogonia (sg). Spermatogenesis has ceased. H&E stain xl25.

PLATE 40: Close-up (xSOO) of an aged adult, 509, seminiferous tubule in cross-section. Note lack of organization to reproductive cells and sporadic occurrence of spermatids (sd). H&E stain.


PLATE 41: Section of the testis of aged adult, 521, showing ''Lydig cell complex'' (LSC) and collapsed seminiferous tubules (st). H&E stain xl25.
