shape and distribution of astrocytes in the cat retina. presumably being involved in the...
TRANSCRIPT

Reports
Shape and Distribution of Astrocytes in the Cot Retina
Andreas Karschin, Heinz Wassle, and Jutta Schnitzer
Astrocytes in the adult cat retina were stained by immuno-cytochemical localization of glial fibrillary acidic protein(GFAP), a major constituent of the astrocytic intermediatefilaments. Their density in whole mounted retinae showed apeak of about 2000 astrocytes/mm2 at the optic nerve headand dropped to approximately 200 astrocytes/mm2 in the farperiphery. At the central area a prominent local minimum ofastrocyte density was found. The shape of astrocytes changedfrom a stellate form in the outermost retinal periphery to anelongated form in the central part of the retina with the ma-jority of astroglial processes aligned in parallel with the gan-glion cell axons. Results in the cat retina suggest a closecorrelation between astrocytes and optic nerve fibers, the latterpresumably being involved in the establishment of the astro-cytic network. Invest Ophthalmol Vis Sci 27:828-831,1986
Recent studies of astrocytes in the mammalian retinausing classical anatomical methods showed some de-tails of astroglial morphology and also gave evidenceof a characteristic density distribution.1"4 Whereas mostclasses of retinal neurons have their maximum densityin the central area, the number of astrocytes in theprimate retina is roughly proportional to the thicknessof the nerve fiber layer and peaks at the optic nervehead. Although the function of retinal astrocytes re-mains speculative, a definite correlation between as-trocytes and axons in the retina might give further in-sight into their functional role. In contrast to Nissl- orselective Golgi-staining, applied in previous studies, weused immunocytochemical methods to label astrocytes.Application of antibodies directed against GFAP5 re-vealed, in whole-mount preparations, both the shapeand density distribution of astrocytes in the entireretina.
Materials and Methods. Animal care and treatmentin the present study conforms to the ARVO Resolutionon the Use of Animals in Research. Twelve adult catsused for electrophysiological recording experimentswere deeply anesthetized with 4% halothane and per-fused through the heart with 0.9% saline followed bya freshly prepared fixative containing 4% paraformal-dehyde, 0.1 M DL-lysine monohydrochloride, and 0.01
M sodium-m-periodate in 0.1 M sodium phosphatebuffer (PB) at pH 7.4.6 After enucleation, both retinaewere removed from the eyecup and postfixed for an-other 2 hr. Immunohistochemical staining included theperoxidase antiperoxidase (PAP) method. All antibodydilutions were carried out in 0.5% Triton-X-100 in PB.After each step, free floating whole retinae were washedthree times for 15 min in PB. In brief, the procedureis as follows: (1) Pretreatment with 3% hydrogen per-oxide/10% methanol in PB for 30 min at room tem-perature. (2) Incubation for 1 hr at room temperaturein 10% normal goat serum (NGS)/0.1 M DL-lysinemonohydrochloride. (3) Rabbit antiserum directedagainst glial fibrillary acidic protein (GFAP; a gift fromDrs. D. Dahl and A. Bignami) was used at a dilutionof 1:300. All incubation steps were carried out at 4°Cfor 3-4 days. (4) Whole retinae were further treatedwith the immunoglobulin fraction of a goat anti-rabbitserum (Miles Scientific; Naperville, IL) diluted 1:50followed by (5) incubation with a rabbit peroxidaseantiperoxidase complex (Miles Scientific; Naperville,IL) at a dilution of 1:100. Demonstration of the anti-body binding was carried out by (6) preincubation in0.05% 3,3'-diaminobenzidine tetrahydrochloride(DAB; Sigma Chemical; St. Louis, MO) in PB for 5min, followed by (7) incubation in 0.05% DAB/0.01%hydrogen peroxide in PB for an additional 10 min.Retinae were flat-mounted (ganglion cell-side up) onglass slides, dehydrated in ascending alcohols, clearedin xylene and mounted with Permount after counter-staining with the Lillie modification (1968) of WeigertIron Hematoxylin.7 In addition we used the indirectimmunofluorescence technique in which the GFAPantiserum was followed by goat anti-rabbit antibodiesconjugated to fluorescein-isothiocyanate (FITC)(Cappel Laboratories; West Chester, PA). Cell countswere made using a Zeiss (Oberkochen, West Germany)drawing tube with a X 1/0.04 objective for constructingretinal maps and a X40/0.75 (oil-immersion, Zeiss,Planapo) for scanning the whole mounts and countingthe densities of all GFAP-positive astrocytes by meansof a 10 X 10-mm grating in the eyepiece.
828
Downloaded From: https://iovs.arvojournals.org/pdfaccess.ashx?url=/data/journals/iovs/933130/ on 08/27/2018
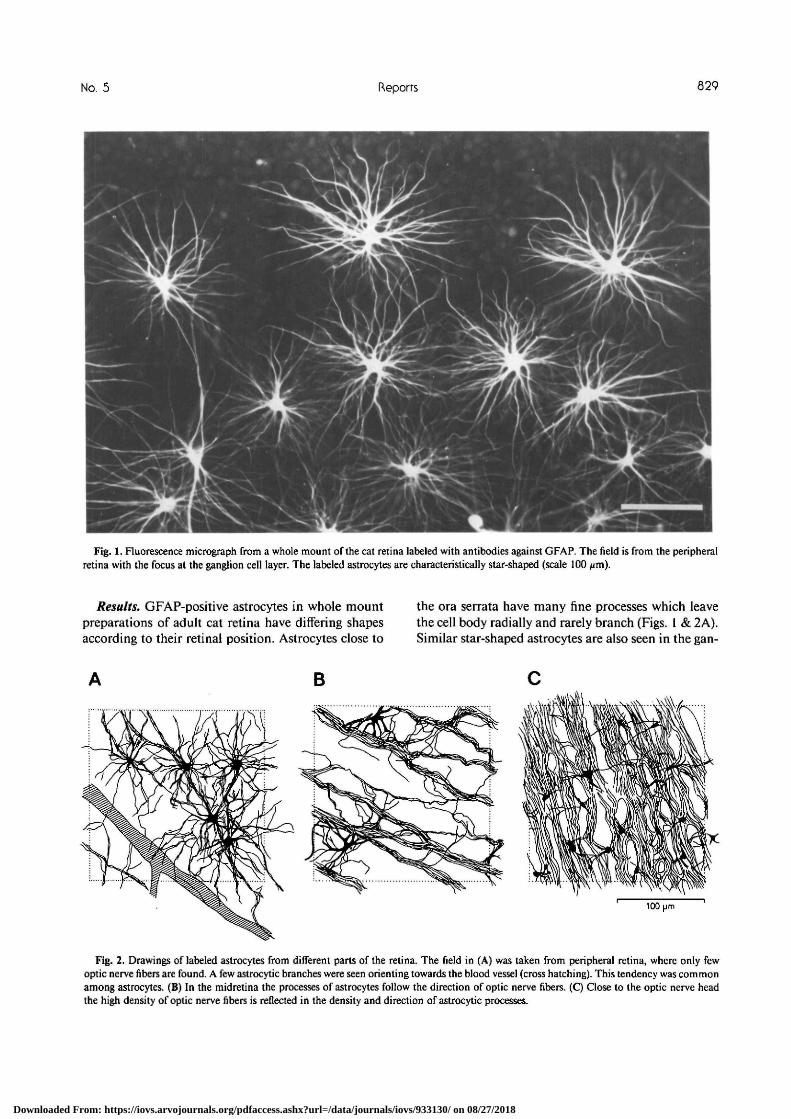
No. 5 Reports 629
Fig. 1. Fluorescence micrograph from a whole mount of the cat retina labeled with antibodies against GFAP. The field is from the peripheralretina with the focus at the ganglion cell layer. The labeled astrocytes are characteristically star-shaped (scale 100 /im).
Results. GFAP-positive astrocytes in whole mountpreparations of adult cat retina have differing shapesaccording to their retinal position. Astrocytes close to
the ora serrata have many fine processes which leavethe cell body radially and rarely branch (Figs. 1 & 2A).Similar star-shaped astrocytes are also seen in the gan-
100 \im
Fig. 2. Drawings of labeled astrocytes from different parts of the retina. The field in (A) was taken from peripheral retina, where only fewoptic nerve fibers are found. A few astrocytic branches were seen orienting towards the blood vessel (cross hatching). This tendency was commonamong astrocytes. (B) In the midretina the processes of astrocytes follow the direction of optic nerve fibers. (C) Close to the optic nerve headthe high density of optic nerve fibers is reflected in the density and direction of astrocytic processes.
Downloaded From: https://iovs.arvojournals.org/pdfaccess.ashx?url=/data/journals/iovs/933130/ on 08/27/2018

830 INVESTIGATIVE OPHTHALMOLOGY 6 VISUAL SCIENCE / Moy 1986 Vol. 27
5mm
Fig. 3. Isodensity lines of astrocyte distribution in a left cat retina.The numbers indicate densities in cells/mm2. Densities steeply in-crease towards the optic nerve head (shown as black dot) up to ap-proximately 2000 cells/mm2. The local minimum of astrocyte countsin the central area is indicated by a star.
glion cell layer of the central retina. Concomitant withincreasing density of optic nerve fibers toward the opticdisc, processes of fiber layer astrocytes tend to orientthemselves parallel to ganglion cell axons (Figs. 2B, C).Thus, apart from some short radiating processes, themajority of processes are aligned in thick bundles andform a characteristic astroglial network. Various tran-sitions between these two extreme features are foundat all locations between the ora serrata and the opticdisc.
The other glial population of the cat retina, theMiiller cells, have not been found to be convincinglyGFAP-positive. However, in some whole mounts theirpedicles appeared slightly immunoreactive.
A striking feature of the astroglial density distributionis the lack of astrocytes in the area centralis. In pe-ripheral retina astrocytes form a rather regular mosaic(Fig. 1), whereas in central retina astrocytes are arrayedin bundles that bypass the area centralis in successivearc-like formations and follow ganglion cell axon fas-cicles on their way to the optic disc. The quantitativeanalysis of astrocyte density in Figure 3 reveals a max-imum of about 2000 astrocytes/mm2 at the optic nervehead and densities scattering between 150-250 astro-cytes/mm2 close to the ora serrata. Furthermore thereis a local minimum in the central area dropping to
counts lower than 50 astrocytes/mm2. Thus, mor-phologies and total number of astrocytes in the catretina closely follow the topography of retinal axonbundles.
Discussion. The advantage of staining all astrocytesin a whole mount preparation is that it reveals thegradual change from stellate shape in peripheral retinato elongated shape close to the optic nerve head. Fromthis material it is quite obvious that both are extrememorphological variations of one cell type. It is also ap-parent that the optic nerve fibers are the substrate thatcauses this change. The correlation between the num-ber of astrocytes and thickness of the optic nerve fiberlayer, as described in the primate,2'3 is confirmed herefor the cat retina. Such a correlation or even a closeapposition of glial and neuronal processes might in-dicate intercellular interactions as proved to be the casein axonal guidance during early brain development.89
It remains speculative as to whether astroglial processes,shown in vitro to serve as a template for outgrowingneurites and the neuronal pattern,10" only providemechanical support for neuronal elements or are in-volved in physiological functions. Although only Miil-lerian glia have been shown to take up and direct excesspotassium into the vitreous body of the vertebrate ret-ina,12 astrocytes in general are thought to present aneffective means of removing potassium released intothe extracellular space during axonal activity.13 Bearingthese functional aspects in mind, we consider cat retinalastrocytes to be a special glia for ganglion cell axons,as already suggested by Biissow.4
Key words: astrocytes, cat retina, glial fibrillary acidic protein,nerve fiber layer
Acknowledgment. The gift of antibodies against GFA pro-tein from Drs D. Dahl and A. Bignami is gratefully acknowl-edged.
From the Max-Planck-Institut fur Hirnforschung, Deutschor-denstr. 46, D-6000 Frankfurt a.M. 71, West Germany. Submittedfor publication: August 29, 1985. Reprint requests: Prof. Heinz Was-sle, Max-Planck Institut fur Hirnforschung, NeuroanatomischeAbteilung, 6000 Frankfurt a.M. 71 Niederrad, Deutschordenstrasse,46, Post fach 7104 09.
References
1. Ikui H, Uga S, and Kohno T: Electron microscope study onastrocytes in the human retina using ruthenium red. OphthalmicRes 8:100, 1976.
2. Ogden TE: Nerve fibre layer astrocytes of the primate retina:morphology, distribution, and density. Invest Ophthalmol VisSci 17:499, 1978.
3. Ogden TE: Nerve fibre layer of the primate retina: thickness andglial content. Vision Res 23:581, 1983.
4. Biissow H: The astrocytes in the retina and optic nerve head ofmammals: a special glia for the ganglion cell axons. Cell TissueRes 206:367, 1980.
Downloaded From: https://iovs.arvojournals.org/pdfaccess.ashx?url=/data/journals/iovs/933130/ on 08/27/2018

No. 5 Reports 831
5. Uyeda CT, Eng LF, and Bignami A: Immunological study ofthe glial fibrillary acidic protein. Brain Res 37:81, 1972.
6. McLean JW and Nakane PK: Periodate-lysine-paraformalde-hyde-fixative. A new fixative for immunoelectron microscopy.J Histochem Cytochem 22:1077, 1974.
7. Humason GL: Animal Tissue Techniques. 3rd ed. San Francisco,WH Freemann and Co, 1972.
8. Levitt P and Rakic P: Immunoperoxidase localization of glialfibrillary acidic protein in radial cells and astrocytes of the de-veloping rhesus monkey brain. J Comp Neurol 193:815, 1980.
9. Silver J, Lorenz SE, Wahlsten D, and Coughlin J: Axonal guid-ance during development of the great cerebral commissures: de-
scriptive and experimental studies, in vivo, on the role of pre-formed glial pathways. J Comp Neurol 210:10, 1982.
10. Noble M, Fok-Seang J, and Cohen J: Glia are a unique substratefor the in-vitro growth of central nervous system neurons. JNeurosci 4:1892, 1984.
11. Fallon JR: Preferential outgrowth of central nervous systemneurites on astrocytes and Schwann cells as compared with non-glial cells in vitro. J Cell Biol 100:198, 1985.
12. Newman EA: Regulation of potassium levels by glial cells in theretina. Trends in Neuroscience 4:156, 1985.
13. Somjen GG: Electrophysiology of neuroglia. Annu Rev Physiol37:163, 1975.
A Monodonol Antibody Thot Binds to Cones
Vance Lemmon
A monoclonal antibody that binds to cones has been produced.This antibody, 50-1 Bl 1, binds to the outer segments of conesin rhesus monkeys. Immunohistochemical experiments in-dicate that 50-1BH binds to a subset of photoreceptors,probably cones, in all vertebrate species tested thus far, in-cluding man. In vitro experiments on chicken retina indicatethat the antigen is intracellular and associated with the plasmamembrane, while electronmicroscopic-immunohistochemicalstudies demonstrate that the antigen is contained in the la-mellae of the outer segments of rhesus cones. InvestOphthalmol Vis Sci 27:831-836, 1986
The ability to use immunohistochemical techniquesto differentiate between different classes of cells hasproven to be an invaluable approach in studying bothnormal cell function and pathological conditions. Overthe past several years a number of monoclonal anti-bodies have been developed that bind to specific classesof cells in the retina of various species.1"3 Until now,there have been no antibodies produced that bind spe-cifically to vertebrate cones. This, combined with thelack of conventional histochemical techniques to dis-criminate between rods and cones, has greatly hinderedstudies of retinal degeneration in both human patho-logical material and in animal models of photoreceptordegeneration. We have recently produced a mono-clonal antibody that binds to cones in monkeys andalso binds to cones in formalin fixed human retina.Therefore, we believe that this antibody will greatlyfacilitate studies of the cell biology and pathology ofcones.
Materials and Methods. Mice were obtained fromJackson Labs (Bar Harbor, ME). White leghorn chickeneggs were obtained from S. Sacks and Son; Evans City,PA. All procedures used in this study conform to theARVO Resolution on the Use of Animals in Research.
To prepare material for immunization of mice, reti-nas were dissected from E19 chick embryos, andplasma membranes were prepared using discontinuoussucrose density gradients with steps of 0.32 M, 0.8 M,and 1.2 M sucrose.4 Plasma membranes were collected,solubilized in 0.5% NP-40 in 20 mM TRIS, saline, pH7.2 and then centrifuged at 100,000Xg.av. for 30'. Thesupernatant was then incubated with Bio-beads (BioradLaboratories; Richmond, CA) to remove the detergent.The supernatant was used to immunize Balb/C miceusing Freund's complete adjuvant. One month laterthe mice were boosted with antigen in Freund's in-complete adjuvant and then rested for at least another4 wk. Prior to fusion a mouse was injected with antigenvia the tail vein on 3 successive days. All immuniza-tions contained approximately 100 fxg of protein.
Hybridomas were prepared following standard pro-cedures using NS-1 cells.5 Supernatants were firstscreened using a dot blot immunoassay with the samematerial that was used to immunize the mice.6 Wellscontaining supernatants that were positive on the dotblot were subsequently tested using immunohistolog-ical procedures with cryostat sections of paraformal-dehyde fixed E19 chick retina. After incubating thesections with hybridoma supernatant the sections werewashed with phosphate buffered saline (PBS) and thenincubated with a 1:500 dilution of fluorescein labeledgoat anti-mouse-IgG (Cappel Labs). Wells containingsupernatants that bound to restricted classes of retinalcells were cloned by limiting dilution.
In order to study the specificity of antibody 50-1B11double label experiments were conducted using tech-niques developed by de Monasterio et al.7 Procion ye-low (obtained from Dr. de Monasterio) was injectedinto the vitreous of rhesus monkeys that were beingused in neuroanatomical experiments for other pur-
Downloaded From: https://iovs.arvojournals.org/pdfaccess.ashx?url=/data/journals/iovs/933130/ on 08/27/2018