should we kill the messenger? the role of the surveillance complex in translation termination and...
TRANSCRIPT

Should we kill the messenger?The role of the surveillancecomplex in translationtermination and mRNA turnoverKevin Czaplinski, Maria J. Ruiz-Echevarria,Carlos I. Gonzalez, and Stuart W. Peltz*
Summary
Eukaryotes have evolved conserved mechanisms to rid cells of faulty geneproducts that can interfere with cell function. mRNA surveillance is an example ofa pathway that monitors the translation termination process and promotes degra-dation of transcripts harboring premature translation termination codons. Studieson the mechanism of mRNA surveillance in yeast and humans suggest a commonmechanism where a ‘‘surveillance complex’’ monitors the translation process anddetermines whether translation termination has occurred at the correct positionwithin the mRNA. A model will be presented that suggests that the surveillancecomplex assesses translation termination by monitoring the transition of an RNPas it is converted from a nuclear to a cytoplasmic form during the initial rounds oftranslation. BioEssays 21:685–696, 1999. r 1999 John Wiley & Sons, Inc.
Appropriate expression of genes requires thatthe cellular machinery be highly accurateThe normal flow of gene expression in eukaryotic cellsprogresses from DNA to RNA to protein. Even for genes withthe simplest form of regulation, a large number of events mustoccur with great precision for the accurate production of theprotein product. As a consequence, cells have evolved a‘‘gene expression factory’’ consisting of ‘‘sub-departments’’that are responsible for carrying out the processes of transcrip-tion, splicing, polyadenylation, transport, and translation. Thesteps in this process do not occur in isolated environments,and the ‘‘sub-departments’’ must communicate with eachother to produce the appropriate protein product. Each step inthe process of expressing a gene must be performed accu-
rately and with very low error rates. The fidelity of theseprocesses is a consequence of both the accuracy of thecellular machinery that has evolved to carry out thesefunctions as well as the quality control mechanisms thatsafeguard against faulty products.
Nonsense-mediated mRNA decay is an exampleof a quality control mechanism that ensures highfidelity of gene expressionA clear example of a cellular quality control mechanism is thenonsense-mediated mRNA decay pathway (NMD). This path-way functions to eliminate aberrant mRNAs that containnonsense mutations within the protein coding region thatcould result in a truncated protein product.(1–7) Nonsensemutations can arise from errors during transcription, splicingor DNA replication, or DNA mutagenesis.(4,8) Furthermore,frameshift mutations have been demonstrated to activateNMD by introduction of premature termination codons be-yond the site of the lesion.(9–12) Mutations in DNA that result inpremature translation termination lead to a permanent loss offunction of the given protein. Interestingly, there are close to200 genetic disorders that can result from premature transla-
Department of Molecular Genetics and Microbiology, University ofMedicine and Dentistry of New Jersey, Robert Wood Johnson MedicalSchool, and Cancer Institute of New Jersey, Piscataway, New Jersey.*Correspondence to: Stuart W. Peltz, Department of Molecular Genet-ics and Microbiology, Robert Wood Johnson Medical School,UMDNJ, 675 Hoes Lane, Piscataway, NJ 08854. E-mail: [email protected]
Review articles
BioEssays 21:685–696, r 1999 John Wiley & Sons, Inc. BioEssays 21.8 685

tion termination.(13) Conceivably, understanding how thisprocess affects translation termination and mRNA degrada-tion can lead to rational approaches for the treatment of asubset of these disorders.(14)
The NMD pathway functions in all eukaryotic systemsexamined and appears to have evolved to ensure thattermination of translation occurs at the appropriate codonwithin the transcript.(15–18) Transcripts containing prematurenonsense codons are rapidly degraded, thus preventingsynthesis of incomplete and potentially deleterious pro-teins.(1,4,19) An example of the effect of a nonsense mutationon the decay of the PGK1 transcript in the yeast Saccharomy-ces cerevisiae is shown in Figure 1. The PGK1 mRNA isnormally stable with a half-life of greater than 45 minutes.Insertion of a premature nonsense codon, however, reducesits decay rate to 3 minutes or less. Similar results have beenobserved in a variety of mammalian cell types and in differentorganisms.(1,3,4,19)
The main goal of this review is to describe our currentknowledge about the NMD pathway, focusing primarily on theresults obtained from yeast, but establishing functional com-parisons with higher eukaryotic systems, including mamma-lian cells. Based on these observations, a model to explainhow this pathway recognizes and degrades aberrant mRNAswill be presented. In particular, the model presented will unifythe mechanism of the NMD pathway in yeast and humancells. We suggest that the NMD pathway is a surveillancemechanism that is involved in modulating the fidelity of anumber of cellular processes in translation and mRNA decay.Further, the NMD pathway clearly exemplifies how coopera-tion between seemingly independent processes within a cellis utilized to ensure the appropriate synthesis of a correctproduct.
What differentiates a premature vs. a wild typetermination codon?: the role of the DSEIn order to determine how the NMD pathway degrades anaberrant transcript, it is necessary to understand how the celldiscriminates between a premature vs. a normal terminationcodon (Fig. 2A). Since there are only three terminationcodons (UAA, UAG, and UGA), it is difficult to understandhow they would carry sufficient information to discriminatebetween a wild type and premature termination event. Previ-ous results have demonstrated that a nonsense codon isrecognized as aberrant due to sequences 38 of the nonsensecodon, a sequence defined as a ‘‘downstream element’’(DSE; Fig. 2A constructs 2 and 4).(11,20,21) Transcripts contain-ing a premature termination codon, but lacking a DSE orcontaining a sequence that is not capable of activating NMDare not degraded by the NMD pathway (Fig. 2A constructs 3and 5).(11,20) Thus, the signal to promote NMD is a bipartiteelement consisting of a nonsense codon and a 38 DSE.
The sequences in the DSE important for activating NMDhave been characterized in several transcripts and the detailsof these results have been reviewed previously.(4) Theseexperiments revealed that multiple DSEs can be found withinthe protein coding region of a given gene.(22) For example, atleast three different regions within the PGK1 transcript havebeen shown to function as a DSE (Fig. 2B). These resultssuggest that the sequence of the DSE is flexible and may beinteracting with an RNA binding protein or a set of RNAbinding proteins (see below).
The interplay between the nonsense codon and the DSEhas been analyzed extensively in yeast using the leaderregion of the yeast GCN4 gene as a model system(Fig. 3).(20,23) The GCN4 gene encodes a transcriptionalactivator of amino acid biosynthetic genes and its mRNA
Figure 1. The effect of an early nonsense mutation on the decay of the PGK1 mRNA. Decay rates for the mRNAs encoded by the wildtype and early nonsense-containing PGK1 alleles were determined by Northern blot analysis of RNAs isolated at different times aftertranscription was inhibited by a shift from 24°C to 36°C in a strain harboring a temperature-sensitive RNA polymerase II.
Review articles
686 BioEssays 21.8

contains four short upstream open reading frames (uORFs)preceding the GCN4 protein coding region (Fig. 3A).(24–27)
Translational initiation/reinitiation events are known to occurin these uORFs, which regulate the expression of the GCN4protein (Fig. 3A, constructs 2,3).(28,29) We have shown thatthe uORFs in the GCN4 leader region do not functionas premature termination codons because they lack a DSE(Fig. 3B).(20) Introduction of a DSE 38 of uORF1 resulted inrapid decay of this mRNA (Fig. 3B).(20) Experiments thatutilized the leader region of the GCN4 transcript(20,23) re-vealed that:
1. DSEs are only functional after at least one translationinitiation/termination cycle has been completed (Fig. 3C,
Figure 3. Using the GCN4 transcript to investigate the rulesthat govern the interplay between translation and NMD. A:Translational control of GCN4. The schematic representationof the wild type GCN4 transcript is shown (construct 1) with thefour uORFs (numbered boxes). When the amino acid pool issufficient (construct 2), ribosomes (double oval) translateuORF1 and reinitiate translation at uORF4. The uORF4represents a very strong barrier to translation reinitiationpreventing synthesis of the Gcn4 protein. Under starvationconditions (construct 3) ribosomal subunits (single oval) scan-ning from uORF1 are not competent to reinitiate at uORF4 dueto low activity of essential components of the translationalapparatus. These subunits become competent to reinitiatetranslation while scanning the interval between uORF4 andGCN4 and reinitiate at GCN4 leading to high levels of Gcn4p.B: Insertion of a DSE 38 of a GCN4 uORF can promote NMD.Schematic representations of the GCN4-PGK1 chimeras andmRNA decay rates are shown. The mRNA decay rates weredetermined as described in Figure 1. The presence of a DSEin construct 2 promotes rapid mRNA decay. C: The activity ofthe DSE requires the process of translation, but not theelongation phase. Insertion of the DSE 38 of a wild type uORFin the GCN4 leader region (construct 1) or 38 of a mutateduORF in which the elongation phase of translation has beenabolished (construct 3) leads to rapid mRNA decay. Mutationof the start site of the uORF prevents translation and rapidmRNA decay (construct 2). The black box at the 38 end of thetranscripts represents a region containing the transcriptiontermination and polyadenylation signals from PGK1.
Figure 2. Identification of cis-acting sequences in the PGK1transcript required for activation of the NMD pathway. A:Schematic representations of the wild type PGK1 allele(construct 1), full-length PGK1 nonsense allele (construct 2),and mini-PGK1 nonsense alleles (constructs 3, 4, and 5) areshown. The mini-PGK1 nonsense allele harbors a 58-proximalnonsense mutation and the sequence between 5.6 and 92.6%of the protein coding region has been deleted and either lacks(construct 3) or contains in either orientation (constructs 4 and5) a DSE (shaded box). The mRNA decay rates of these PGK1alleles were determined as described in Figure 1. B: Aschematic representation of the PGK1 gene containing bothidentified and putative DSEs. DSEs are shown below thePGK1 gene. The second row shows regions with putativeDSEs whose activities have not been tested.
Review articles
BioEssays 21.8 687

construct 2); 2. Although the initiation and termination phasesof translation are required for NMD, the elongation phase isdispensable (Fig. 3C, construct 3); 3. A DSE can activate theNMD pathway when it is located within approximately 150 nts38 of the stop codon (Fig. 3D); this result indicates that a DSEdoes not need to be immediately adjacent to the nonsensecodon; 4. DSEs are not functional if they are part of a proteincoding region that is traversed by ribosomes during thenormal process of translation (Fig. 3E).
‘‘Stabilizer elements’’ have been identified thatcan inactivate the NMD pathwayThe results described above suggest that a translationtermination event in a wild type transcript may not activate theNMD pathway simply because it does not contain a DSE inthe 38-untranslated region. Although this may be the case forsome transcripts, results have shown that there are specificsequences, defined as ‘‘stabilizer elements (STEs),’’ thatinactivate NMD.(11,23,30) Two classes of stabilizer elementshave been identified (Fig. 4). One type of STE is located in theprotein coding region and must be translated for it to inacti-vate the NMD pathway (Fig. 4A).(11,30) A second STE hasrecently been identified in the leader region of the yeastGCN4 transcript (Fig. 4B).(23) This STE is not part of theprotein coding region and inactivates the NMD pathway if it ispositioned downstream of the termination codon and up-
stream of a DSE (Fig. 4B). These results indicate that thereare at least two mechanisms that can inactivate the NMDpathway.
Comparison of the sequences that promote NMDin yeast and mammalian cellsThe sequence requirements for NMD in mammalian cellshave also been investigated. Similar to yeast cells, both anonsense codon and a downstream element are required topromote NMD. Interestingly, although spliced transcriptshave been shown to be substrates for the NMD pathway,(31)
Figure 3. Continued. D: The activity of the DSE depends onits distance from the stop codon. A DSE was inserted atdifferent 38 positions relative to a uORF. The decay rates of themRNAs are shown. Insertion of a DSE proximal to a termina-tion codon (construct 1) leads to rapid decay of the mRNA.Increasing the distance between the stop codon and the DSEinactivates the function of the DSE leading to stabilization ofthe mRNA (construct 2). E: The DSE is not functional when itis translocated by the ribosomes. Schematic representationsof the GCN4- PGK1 fusions utilized in this analysis are shown.In construct 1, the presence of a premature stop codon 58 ofthe DSE activates the NMD and the mRNA is rapidly de-graded. Construct 2 lacks a premature stop codon 58 of theDSE and is not susceptible to NMD.
Figure 4. Two classes of stabilizer elements (STE) thatinactivate the NMD pathway. A: A stabilizer element is locatedin the protein coding region of the PGK1 transcript and mustbe translated to inactivate the NMD pathway. The decay ratesof several PGK1 nonsense-containing mRNAs used in theanalysis are shown. The location of each nonsense mutationin the PGK1 transcript is presented as the percentage of thePGK1 protein coding region that is translated before the earlytermination codon is encountered. mRNA turnover rates forwild type and nonsense-containing PGK1 alleles were deter-mined by Northern blot analysis as described in Figure 1. ThePGK1 alleles shown are as follows: 1. the wild-type PGK1gene; 2. the PGK1 gene with a nonsense mutation at 92.6 %of the coding region; 3. the PGK1 gene with a nonsensemutation at 39 % of the coding region. Failure to translate theSTE (shaded box) leads to rapid mRNA decay (construct 3).B: A stabilizer element is located in the leader region of theGCN4 transcript. Schematic diagrams of the GCN4 andGCN4-PGK1 constructs used for the analysis are shown.Deletion of the STE from the GCN4 transcript (compareconstructs 1 and 2) leads to rapid mRNA degradation. TheSTE is functional when located in hybrid transcripts between atermination codon and a DSE (compare constructs 3 and 4).
Review articles
688 BioEssays 21.8

the splicing of an intron 38 of a nonsense codon can functionas a downstream element.(31–35) Other sequences besidesintrons, however, also function to promote NMD, suggestingthat splicing is not an obligatory event in the NMD pathway inmammalian cells.(32,34) An interesting dilemma is to explainhow, even though the spliced transcript is a substrate forNMD, the presence of an intron 38 of a termination codon candetermine whether the transcript is degraded by this pathway(see below).
Although the cellular compartment in which NMD occurs inmammalian cells has been controversial,(1,5,35,36) it appearsthat in both yeast and mammalian cells NMD occurs in thecytoplasm when the transcript is associated with ribo-somes.(3,35,37) Thus, an interesting question is to explain howan intron that is removed in the nucleus can affect mRNAdecay in the cytoplasm. A reasonable explanation is that thesplicing event must ‘‘mark’’ the RNA at the location of theintron.(31–35) For these cases, such a mark is most likely aprotein that associates with the mRNA during the splicingprocess. As described below, likely candidates for suchfactors would be RNA binding proteins that can shuttlebetween the nucleus and the cytoplasm.
Identification of the trans-acting factorsinvolved in NMDThree factors involved in NMD have been identified in yeast.Mutations in the UPF1, UPF2, and UPF3 genes were shownto selectively stabilize mRNAs containing early nonsensemutations without affecting the decay rates of most wild typemRNAs.(9,10,38–40) Recent results indicate that the Upf1p,Upf2p, and Upf3p interact and form a complex.(16,38,41) Basedon these results and others, we have suggested the exis-tence of a surveillance complex required for the degradationof nonsense-containing mRNAs that consists of at leastUpf1p, Upf2p, and Upf3p. Deleting combinations of the UPFgenes has the same affect on mRNA levels as deleting asingle UPF gene, consistent with the Upfps functioning as acomplex.(40,41) Therefore, mutations in the surveillance com-plex that inactivate the NMD pathway reduce the fidelity of theprocesses that protect cells from aberrant mRNAs.
The proteins involved in promoting NMD have also beeninvestigated in C. elegans and mammalian cells. In C. ele-gans, seven smg alleles have been identified, which result inan increased abundance of nonsense-containing tran-scripts.(19) A human homologue of the UPF1 gene, calledRENT1 or HUPF1, has been identified, indicating that NMD isan evolutionarily conserved pathway.(42,43) Recent resultssuggest that the human UPF1/rent1 protein also functions inmammalian cells to control the stability of nonsense contain-ing transcripts.(44) In these experiments expression of amutant hupf1 protein confers inhibition of NMD, suggestingthat it can interfere with the normal function of the wild typeHUPF1 in degrading nonsense-containing transcripts.
Mutations in the UPF1 gene demonstrate that it isinvolved in modulating the efficiency oftranslation terminationThe protein product of the yeast UPF1 gene has been the mostextensively investigated factor involved in NMD.(9,10,15–18,45–49)
The Upf1p contains a cysteine- and histidine-rich region nearits amino terminus and all the motifs required to be a memberof the superfamily group I helicases (Fig. 5A).(50) The yeastUpf1p has been purified and demonstrates RNA binding,RNA-dependent ATPase and RNA helicase activities.(15–17,45)
Disruption of the UPF1 gene results in selective stabilizationof nonsense-containing mRNAs. (9,15,16)
In addition to its role in accelerating the decay of nonsense-containing transcripts, recent evidence suggests that theUpf1p enhances translation termination at a nonsensecodon.(10,15–18) Translation termination is the final step ofprotein biosynthesis and consists of a codon-dependenttermination event that results in release of the polypeptidechain from the ribosome.(51,52) In eukaryotes this process isunder the control of two protein factors: eRF1, which cata-lyzes peptide hydrolysis at all three nonsense codons(53) andeRF3, which stimulates the activity of eRF1 in releasing thepolypeptide in a GTP-dependent manner.(54–56)
Nonsense suppression can be used as an in vivo assay forthe measurement of translation termination efficiency.(51,57)
Nonsense suppression results when a near cognate tRNA ora mutant suppressor tRNA successfully competes with thetermination factors at a nonsense mutation so that incorpora-tion of an amino acid into the peptide chain occurs rather thanprematurely terminating translation (Fig. 5B). Increased non-sense suppression in an essential gene harboring a non-sense mutation allows sufficient production of a completedpolypeptide, which can support growth. For example, cellsharboring a nonsense mutation in the LEU2 gene, whoseprotein product is required for the production of the aminoacid leucine, can grow if leucine is present in the media butcannot grow in media lacking leucine (Fig. 5B). If amino acidincorporation occurs at the premature termination codon withhigh enough frequency, the cells harboring the nonsensemutation in the LEU2 gene can grow on media lackingleucine, thus suppressing the nonsense mutation.
The initial result suggesting that the Upf1p is involved intranslation termination was the finding that a upf1D straindemonstrated a nonsense suppression phenotype.(10,15,16,40)
Strains that harbor the leu2–2 nonsense allele and certainmutations in the UPF1 gene can grow on media lackingleucine (Fig. 5A), and demonstrate stabilized mutant LEU2mRNA. More significantly, a set of mutations in the UPF1gene separated its mRNA decay function from its activity inmodulating translation termination at a nonsense codon (Fig.5A, upf1-C and upf1-D).(15,16) A subset of mutations in thehelicase region of the UPF1 gene, that inactivated both its
Review articles
BioEssays 21.8 689

helicase and decay activities (Fig. 5A, upf1-C), still retainedits ability to enhance translation termination at a nonsensecodon and prevented nonsense suppression.(15) Conversely,another subset of mutations in the cysteine- and histidine-richamino terminal region (Fig. 5A, upf1-D), which still retainedhelicase and decay activities, reduced the ability of the Upf1pto enhance translation termination at a nonsense codon,allowing suppression of certain nonsense alleles.(16) Thecorrelation between loss of helicase activity and decayfunction indicates that the helicase activity is essential for therole Upf1 plays in mRNA stability, but is dispensable for itsrole in translation termination.
The genetic results described above suggested that inaddition to its role in NMD, the Upf1p is involved in modulatingthe efficiency of translation termination at a nonsense codon.Consistent with this notion, both the yeast and human Upf1pwere shown to interact with the translation termination factorseRF1 and eRF3.(18) The interaction of Upf1p with the releasefactors affects the biochemical activities of the Upf1p.(18)
Interaction of the release factors with Upf1p inhibited theATPase activity of Upf1p and eRF3 prevented formation of a
Upf1p:RNA complex. These observations are consistent withthe genetic and biochemical data suggesting that the Upf1pATPase/helicase and RNA binding activities are dispensablefor its role in translation termination.(15–17)
The observation that eRF3 prevented formation of aUpf1p:RNA complex suggested that eRF3 and RNA competefor binding to Upf1p.(18) This competition predicts that factorsthat reduce the Upf1p affinity for RNA would consequentlyfavor binding to the release factors. Interestingly, ATP hasbeen shown to bind to Upf1p and reduce its affinity forRNA.(15,17) This suggests that ATP is an important co-factor ofUpf1p that regulates its interaction with the release factors inthe presence of competing RNAs.(18) These observationssuggest that ATP is a co- factor that switches Upf1p betweenits translation termination and NMD activities.(15–17) Consis-tent with this view, a mutant form of the Upf1p that lackedATPase activity, but bound ATP comparably to the wild type,was functional in enhancing translation termination but un-able to promote NMD.(15,17)
Recent data suggest that the human UPF1/rent1 proteinmay also have a role in modulating translation termination.(18)
Figure 5. Mutations in the UPF1 gene separatenonsense-mediated mRNA decay from nonsensesuppression. A: A schematic diagram of the UPF1gene is shown and indicates the cysteine- andhistidine-rich region in yellow and the ATPase andhelicase region, with the seven superfamily groupI helicase motifs boxed in green. The relativelocations of the point mutations in UPF1 whichseparate nonsense suppression from nonsensemediated mRNA decay are indicated and groupedas upf1-C or upf1-D. The table lists the pheno-types associated with upf1D and the point muta-tions indicated above. B: Nonsense Suppressionis an assay for translation termination efficiency.Wild type cells synthesize full-length Leu2 protein(green boxes) and can grow on media lackingleucine. A nonsense mutation in the LEU2 gene(leu2–2) results in a truncated Leu2 protein (browntriangles) and auxotrophy for leucine. Amino acidincorporation instead of termination at the non-sense mutation will result in a full-length Leu2protein, and if incorporation occurs at high levelsthe cells can synthesize enough full-length Leu2protein to support growth on media lacking leu-cine, despite the presence of the premature termi-nation codon. The peptidyl release factors eRF1and eRF3 are required for translation terminationand mutations which reduce the ability of therelease factors to terminate translation will resultin nonsense suppression.
Review articles
690 BioEssays 21.8

The human UPF1 protein harbors both the cystine-histidine-rich and helicase regions, which are homologous to the yeastUPF1 gene.(42,43) In addition, hupf1 protein contains aminoand carboxyl terminal domains that are divergent from thosein the yeast counterpart. The human Upf1 protein is notexpressed in yeast whereas a human/yeast chimera thatlacks the amino and carboxyl terminal extensions from thehuman gene can be expressed in yeast.(42) Expression of theyeast/human Upf1 protein in a upf1D strain prevents suppres-sion of nonsense alleles although it is unable to promote rapiddecay of nonsense containing transcripts.(18) FurthermoreHUPF1 was demonstrated to interact with the peptidyl re-lease factors.(18) This result suggests that the human Upf1pmay also be involved in modulating translation termination ata nonsense codon by an interaction with eRF1 and eRF3.
The results described above indicate that the surveillancecomplex is required for efficient translation termination at anonsense codon. Consistent with this view, cells harboringupf2D or upf3D alleles demonstrate a nonsense suppressionphenotype.(39,40) Thus, mutations in the UPF genes reducethe fidelity of the translation termination event at a nonsensecodon, resulting in nonsense suppression. These results alsodemonstrate a cross-talk between the processes of transla-tion termination and mRNA decay. As described in more detailbelow, these results suggest an ordered set of events inwhich the surveillance complex ensures efficient terminationat a nonsense codon and then is subsequently involved indegrading the aberrant nonsense-containing transcript. Cur-rently no evidence definitively demonstrates a role for the Upfproteins in termination at a normal stop codon. This raises theinteresting question for future experiments of whether or notthe termination machinery recognizes premature terminationcodons differently than normal termination codons, and arethe Upf genes involved specifically at premature terminationcodons.
Mutations in the RNA binding protein Hrp1affect NMDThe results from studies on the sequence requirements forthe degradation of nonsense-containing transcripts demon-strated that recognition of the DSE is a critical event in theNMD pathway (Fig. 3). We have recently identified the Hrp1protein as a factor that specifically interacts with a DSE in thePGK1 transcript (C.I.G. and S.W.P, unpublished results). TheHRP1 gene is an essential gene that is homologous topreviously identified mammalian RNA binding proteins(hnRNPA1 and hnRNPD) and contains two RNA recognitionmotifs.(58) The Hrp1 protein has been shown to shuttlebetween the nucleus and the cytoplasm.(59,60) Mutations inHRP1 gene specifically stabilize nonsense-containing mRNAsin vivo (unpublished results). In addition, we have recentlyfound that Hrp1p interacts with the Upf1p (unpublishedresults). Taken together, these results indicate that the RNA
binding protein Hrp1 may be involved in NMD by interactingwith the DSE and signaling to the surveillance complex,through at least Upf1p, that an aberrant termination event hasoccurred.
The decision to ‘‘kill the messenger’’Once the nonsense-containing transcript is recognized asaberrant, it is rapidly degraded. An important rate determiningstep in the decay of an aberrant transcript is the removal ofthe 58 cap by the Dcp1p decapping enzyme.(30,61) Unlikemany wild type transcripts in which poly(A) shortening pre-cedes decapping,(3) a nonsense-containing mRNA is de-capped with a long poly(A) tract.(61) The uncapped mRNA issubsequently degraded by the Xrn1 protein, a 58=38 exoribo-nuclease.
Although both the wild type and NMD pathways utilize theDcp1p and Xrn1p to degrade mRNAs,(30,61,62) differences inhow these pathways recruit the decapping machinery havebeen observed. For example, the mutant upf alleles specifi-cally slow decapping and stabilize nonsense containingmRNAs.(4) Conversely, several mutations have been isolatedthat specifically inhibit decapping of deadenylated mRNAsbut do not affect the decapping of nonsense-containingtranscripts.(63,64) These results suggest that there are differ-ences in how the factors in these pathways recruit andsubsequently decap the mRNA. Interestingly, both of thesepathways may be utilized to regulate decay rates of specifictranscripts. For example, the a1-tubulin mRNA in Chlamydo-monas reinhardtii utilizes both modes of turnover.(65)
Upf1 homologues have been identified in many otherhigher eukaryotes and recent results indicate that the role ofthis factor is likely to be conserved throughout evolution.(18,44)
At present, however, no homologues in other organisms havebeen identified for the UPF2 and UPF3 genes. Thus, onepossibility is that the role of the Upf2 and Upf3 proteins inNMD in yeast may be carried out by different proteins in otherorganisms. The conservation of the Upf1 protein, combinedwith the observation that NMD pathway is conserved through-out evolution, may suggest that the activities required toactivate the NMD pathway may be contained entirely withinUpf1 protein and that the other factors in NMD (i.e., Upf2 andUpf3) may regulate the activity of the Upf1 protein. If this turnsout to be the case, then studies of the Upf1p will helpelucidate the mechanism of translation termination and mRNAturnover in higher eukaryotes.
A unified model of how NMD occurs in yeast andin mammalian cellsThe studies described above help to develop a model for howNMD occurs in yeast and mammalian cells. A unified modelwill need to account for the role of an intron in this process.However, this model must also explain how nonsense-
Review articles
BioEssays 21.8 691

containing transcripts lacking an intron are degraded by theNMD pathway. The results that identify the Hrp1 protein as afactor involved in NMD gives a clue to help resolve thisdilemma. The observation that Hrp1p is an RNA bindingprotein capable of recognizing the DSE, and shuttling be-tween the nucleus and the cytoplasm suggests that it maybind an RNA in the nucleus, be transported to the cytoplasmand, consequently, be capable of affecting NMD in thecytoplasm. In the case of requiring an intron in order topromote NMD, it is possible that the splicing event brings anRNA binding protein to the mRNA that remains associatedwith the transcript as it is transported from the nucleus to thecytoplasm. Thus, we hypothesize that in mammalian cells,the ‘‘mark’’ on the mRNA from splicing(31–35) is an RNA bindingprotein that can also shuttle from the nucleus to the cyto-plasm. Recent results have demonstrated that SR proteinsinvolved in splicing are also shuttling proteins and may bepotential candidates for factors that mark the transcript.(66,67)
The important point is that it is the ability of an appropriateRNA binding protein to interact with the RNA 38 of thetermination event, and not a splicing event per se, that iscritical for activating the NMD pathway.
Based on the results described above, we present thefollowing model for the mechanism of NMD in yeast andmammalian cells (Fig 6). Concurrent and following synthesisof the transcript the mRNA is packaged into an RNP, whichwe define as a nuclear RNP (Fig. 6A). This RNP is trans-ported from the nucleus to the cytoplasm. During or immedi-ately after transport the protein synthesis machinery interactswith the RNP and ribosomes begin translating the mRNA. Wesuggest that the RNP begins to be remodeled from a nuclearto a cytoplasmic RNP and that the translating ribosome is anactive component of the remodeling process (Fig. 6A). Onerole of the ribosome is to displace the proteins that bound theRNA in the nucleus, such as Hrp1 in yeast, or SR proteinsfrom mammalian cells. Proteins such as Hrp1 or SR proteinsthat disassociate from the RNP are subsequently transportedback to the nucleus.(59,66) We hypothesize that synthesis of anearly full-length polypeptide is important for the successfultransition of a nuclear to cytoplasmic RNP, which will, at leastin part, determine the translatability and stability of themRNA. If, however, the ribosome encounters a prematuretermination codon, then remodeling of the RNP is incomplete,leading to an aberrant RNP structure (Fig. 6B). The aberrantRNP structure is recognized by the degradation machineryand rapid degradation ensues (Fig. 6B).
The NMD pathway demonstrates a link betweentranslation termination and mRNA decayThe studies on how the Upf1 protein functions in translationtermination and mRNA turnover demonstrates a link betweentranslation termination and mRNAdecay. Since the concentra-tion of either release factors or Upfps in cells are less than the
concentration of ribosomes(49,68,69) not every ribosome will beassociated with a surveillance complex. Thus, it is conceiv-able that a paused ribosome at a termination event signalsassembly of this complex (Fig. 6B). Interaction of the releasefactors with the Upf1p and associated factors enhancestranslation termination and also ensures that the RNA-dependent ATPase and/or helicase activities of Upf1p, whichare required to activate NMD, do not function until translationtermination is completed. Once termination is completed, wesuggest that the surveillance complex then searches 38 of thetermination codon, either by scanning or through long rangeinteractions (Fig. 6B). The 58=38 helicase activity of Upf1pmakes it a potential candidate for it being the motor of thescanning process. If the surveillance complex contacts aDSE-associated factor such as Hrp1 the transcript is recog-nized as aberrant due to an incomplete transition fromnuclear RNP to cytoplasmic RNP (Fig. 6B). Thus, the heli-case activity of the Upf1 protein in NMD may also serve as anRNP remodeling factor, analogous to the chromatin (dRNP)remodeling activity of certain transcription factors.(70)
The recognition of whether translation termination hasoccurred prematurely is made by a putative surveillancecomplex interacting with a factor such as the Hrp1 protein(Fig. 6B). Interaction of the surveillance complex with theHrp1 protein, or similar factors, senses that the RNP structureis aberrant and this leads to rapid decapping of the mRNA bythe Dcp1 protein (Fig. 6B). The body of the mRNA issubsequently degraded by the Xrnl 58=38 exoribonuclease.Taken together, the two sequences required for NMD, apremature termination codon, and a DSE, have specific andordered functions in determining whether a transcript isaberrant. The termination event triggers the assembly of thesurveillance complex while the subsequent interaction of thesurveillance complex with the DSE associated with a RNAbinding protein defines an aberrant mRNA that is then rapidlydegraded.
Translation termination may have a broad role inregulating gene expressionThe model described above suggests that the terminationevent is a key point in the assembly of the surveillancecomplex and leads to enhanced translation termination anddegradation of nonsense-containing transcripts. The impor-tant role of the translation termination event may not be onlylimited to degrading aberrant mRNAs. It is conceivable thatthe translation termination event may be an important eventin regulating the stability or translation efficiency of wild typetranscripts. The 38-untranslated regions of many transcriptsencode regulatory elements that modulate the translationefficiency and/or stability of their respective mRNAs.(3,71–74) Itis possible that the termination event is also the signal for theassembly of complexes that subsequently interact with theelements in the 38-UTR that modulate their stability and/or
Review articles
692 BioEssays 21.8
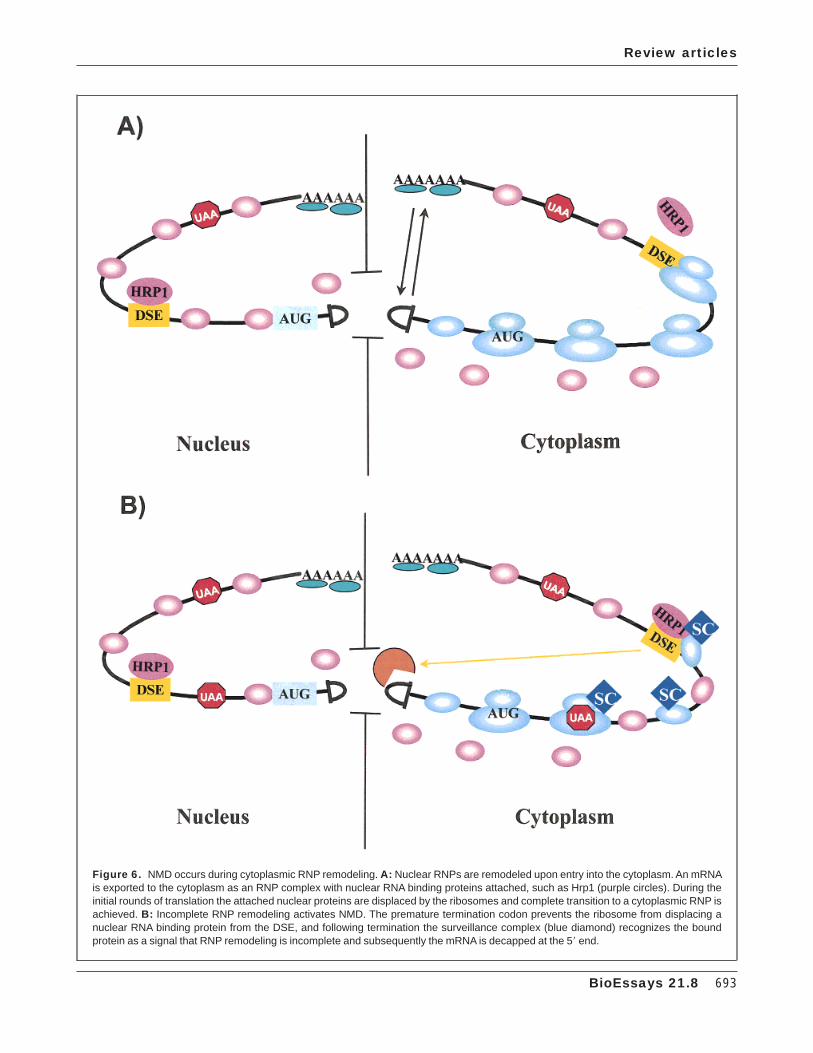
Figure 6. NMD occurs during cytoplasmic RNP remodeling. A: Nuclear RNPs are remodeled upon entry into the cytoplasm. An mRNAis exported to the cytoplasm as an RNP complex with nuclear RNA binding proteins attached, such as Hrp1 (purple circles). During theinitial rounds of translation the attached nuclear proteins are displaced by the ribosomes and complete transition to a cytoplasmic RNP isachieved. B: Incomplete RNP remodeling activates NMD. The premature termination codon prevents the ribosome from displacing anuclear RNA binding protein from the DSE, and following termination the surveillance complex (blue diamond) recognizes the boundprotein as a signal that RNP remodeling is incomplete and subsequently the mRNA is decapped at the 58 end.
Review articles
BioEssays 21.8 693

translation efficiency. Interestingly, one subunit of the proteinphosphatase 2A (PP2A) is the translation termination factoreRF1.(75) It is possible that one role of eRF1 is to bring thePP2A phosphatase into the ribosome at the terminationevent. The PP2A may then be positioned in the appropriatelocation to modulate the activity of factors that regulate thetranslation efficiency or stability of the given transcripts.Interestingly, this scenario is very similar to how we perceivefunctioning of the NMD pathway. The basic premise for bothwild type and NMD is that termination is a rate limiting eventthat pauses the ribosome and signals the assembly ofcomplexes that regulate subsequent events in the life span ofa given transcript. Interestingly, although the role of PP2A intranslation has not been investigated, mutations in the yeastSAL6 gene, that encodes a putative phosphatase, have beenshown to promote suppression of nonsense mutations.(76)
The surveillance complex may have a role inmodulating the fidelity of a number of processesin translationThe results described above implicate the putative surveil-lance complex in having a role in regulating the processes oftranslation termination and mRNA turnover.(9,10,38–40) In addi-tion, recent results have suggested that the surveillancecomplex may also be involved in modulating the fidelity ofcertain aspects of the translation elongation process. Recentresults have shown that mutations in the UPF1 and UPF3genes result in increased programmed -1 ribosomal frame-shifting efficiency.(47,77) Programmed -1 ribosomal frameshift-ing is a unique mode of gene regulation that is usedpredominantly by certain RNA viruses to induce elongatingribosomes to shift reading frame in response to specificmRNA signals.(78–81) In addition, frameshifting efficiency canbe used as an assay to monitor translational fidelity. In-creased frameshifting efficiencies are an indication of re-duced translation fidelity. The fact that mutations in the UPF1and UPF3 genes cause increased frameshifting efficiencysuggests that they are also involved in maintaining fidelity ofthe translation elongation process.
The fact that mutations in factors in the putative surveil-lance complex affect multiple steps in the translation processsuggest it may function as a ‘‘translational checkpoint’’.Analogous to cell cycle control checkpoints, the UPF genesare not essential, but ensure that the processes in which theyare involved occur with high fidelity. In the absence of thiscomplex a number of processes occur with reduced fidelity,putting the viability of the cell at greater risk.
ConclusionsNMD is an interesting pathway because it exemplifies a linkbetween many different cellular processes. This may includeRNP biogenesis in the nucleus, mRNA transport from thenucleus to the cytoplasm, translation elongation and termina-
tion, RNP remodeling and mRNA decay. Thus, NMD isanother example of a process demonstrating a cross-talkbetween multiple pathways. Investigating its mechanismshould give insights into how these multiple processesoverlap to regulate cellular gene expression.
References1. Maquat LE. When cells stop making sense: effects of nonsense codons on
RNA metabolism in vertebrate cells. RNA 1995;1:453–465.2. Caponigro G, Parker R. Mechanisms and control of mRNA turnover in
Saccharomyces cerevisiae. Microbiol Rev 1996;60:233–249.3. Jacobson A, Peltz SW. Interrelationships of the pathways of mRNA decay
and translation in eukaryotic cells. Ann Rev Biochem 1996;65:693–739.4. Ruiz-Echevarria MJ, Czaplinski K, Peltz SW. Making sense of nonsense in
yeast. Trends Biochem Sci 1996;21:433–438.5. Weng Y, Ruiz-Echevarria MJ, Zhang S, Cui Y, Czaplinski K, Dinman JD,
Peltz SW. Characterization of the nonsense-mediated mRNA decaypathway and its effect on modulating translation termination and pro-grammed frameshifting. In mRNA Metabolism and Post-transcriptionalGene Regulation. Modern Cell Biology 1997;17:241–263.
6. Gozalbo D, Hohmann S. Nonsense suppressors partially revert thedecrease of the mRNA levels of a nonsense mutant allele in yeast. CurrGenetics 1990;17:77–79.
7. Losson R, Lacroute F. Interference of nonsense mutations with eukaryoticmessenger RNA stability. Proc Natl Acad Sci USA 1979;76:5134–5137.
8. He F, Peltz SW, Donahue JL, Rosbash M, Jacobson A. Stabilization andribosome association of unspliced pre-mRNAs in a yeast upf1- mutant.Proc Natl Acad Sci USA 1993;90:7034–7038.
9. Leeds P, Peltz SW, Jacobson A, Culbertson MR. The product of the yeastUPF1 gene is required for rapid turnover of mRNAs containing apremature translational termination codon. Genes Dev 1991;5:2303–2314.
10. Leeds P, Wood JM, Lee BS, Culbertson MR. Gene products that promotemRNA turnover in Saccharomyces cerevisiae. Mol Cell Biol 1992;12:2165–2177.
11. Peltz SW, Brown AH, Jacobson A. mRNA destabilization triggered bypremature translational termination depends on three mRNA sequenceelements and at least one trans-acting factor. Genes Dev 1993;7:1737–1754.
12. Baumann B, Potash MJ, Kohler G. Consequences of frameshift mutationsat the immunoglobulin heavy chain locus of the mouse. EMBO J 1985;4:351–359.
13. McKusick VA (with the assistance of Francomano CA, Antonarakis SE,Pearson PL). Mendelian inheritance in man: a catalog of human genesand genetic disorders. Baltimore, MD: Johns Hopkins University Press;1994. (Web site: http://www.ncbi.nlm.nih.gov/Omim/)
14. Howard M, Frizzell RA, Bedwell DM. Aminoglycoside antibiotics restoreCFTR function by overcoming premature stop mutations. Nature Med1996;2:467–469.
15. Weng Y, Czaplinski K, Peltz SW. Genetic and biochemical characteriza-tion of the mutations in the ATPase and helicase regions of Upf1 protein.Mol Cell Biol 1996;16:5477–5490.
16. Weng Y, Czaplinski K, Peltz SW. Identification and characterization ofmutations in the UPF1 gene that affect nonsense suppression and theformation of the Upf protein complex, but not mRNA turnover. Mol Cell Biol1996;16:5491–5506.
17. Weng Y, Czaplinski K, Peltz SW. ATP is a cofactor of the Upf1 protein thatmodulates it translation termination and RNA binding activities. RNA1998;4:205–214.
18. Czaplinski K, Ruiz-Echevarria MJ, Paushkin SV, Han X, Weng Y, PerlickHA, Dietz HC, Ter-Avanesyan MD, Peltz SW. The surveillance complexinteracts with the translation release factors to enhance termination anddegrade aberrant mRNAs. Genes Dev 1998;12:1665–1677.
19. Pulak R, Anderson P. mRNA surveillance by the Caenorhabditis eleganssmg genes. Genes Dev 1993;7:1885–1897.
20. Ruiz-Echevarria MJ, Peltz SW. Utilizing the GCN4 leader region toinvestigate the role of the sequence determinants in nonsense-mediatedmRNA decay. EMBO J 1996;15:2810–2819.
Review articles
694 BioEssays 21.8

21. Peltz SW, Trotta C, He F, Brown AH, Donahue JL, Welch EW, Jacobson A.Identification of the cis-acting sequences and trans-acting factors in-volved in nonsense-mediated mRNA decay. In: Tuite M, McCarthy J,Brown A, Sherman F, editors. Protein synthesis and targeting in yeast.New York: Springer-Verlag; 1993. p H71:1–10.
22. Zhang S, Ruiz-Echevarria MJ, Quan Y, Peltz SW. Identification andcharacterization of a sequence motif involved in nonsense-mediatedmRNA decay. Mol Cell Biol 1995;15:2231–2244.
23. Ruiz-Echevarria MJ, Gonzalez CI, Peltz SW. Identifying the right stop:determining how the surveillance complex recognizes and degrades anaberrant mRNA. EMBO J 1998;17:575–589.
24. Hinnebusch AG. Evidence for translational regulation of the activator ofgeneral amino acid control in yeast. Proc Natl Acad Sci USA 1984;81:6442–6446.
25. Hinnebusch AG. Mechanism of gene regulation in the general control ofamino acid biosynthesis in Saccharomyces cerevisiae. Microbiol Rev1988;52:248–273.
26. Hinnebusch AG. Translational control of GCN4: and in vivo barometer ofinitiation-factor activity. Trends Biochem Sci 1994;19:409–414.
27. Thireos G, Driscoll-Penn M, Greer H. 58-untranslated sequences arerequired for the translational control of a yeast regulatory gene. Proc NatlAcad Sci USA 1984;81:5096–5100.
28. Abastado JP, Miller PF, Jackson BM, Hinnebusch AG. Suppression ofribosomal reinitiation at upstream open reading frames in amino acidstarved cells form the basis for GCN4 translational control. Mol Cell Biol1991;11:486–496.
29. Dever TE, Feng L, Wek RC, Cigan AM, Donahue TD, Hinnebusch AG.Phosphorylation of initiation factor 2a by protein kinase Gcn2 mediatesspecific translational control of GCN4 in yeast. Cell 1992;68:585–596.
30. Hagan KW, Ruiz-Echevarria MJ, Quan Y, Peltz SW. Characterization of cis-acting sequences and decay intermediates involved in nonsense-mediated mRNA turnover. Mol Cell Biol 1995;15:809–823.
31. Carter MS, Li S, Wilkinson MF. A splicing-dependent regulatory mecha-nism that detects translation signals. EMBO J 1996;15:5965–5975.
32. Cheng J, Belgrader P, Zhou X, Maquat LE. Introns are cis-effectors of thenonsense codon-mediated reduction in nuclear mRNA abundance. MolCell Biol 1994;14:6317–6325.
33. Zhang J, Sun X, Qian Y, LaDuca JP, Maquat LE. At least one intron isrequired for the nonsense-mediated decay of triosephosphate isomerasemRNA: a possible link between nuclear splicing and cytoplasmic transla-tion. Mol Cell Bio 1998;18:5272–5283.
34. Zhang J, Sun X, Qian Y, Maquat LE. Intron function in the nonsense-mediated decay of the b-globin mRNA: indications that pre-mRNAsplicing in the nucleus can influence mRNA translation in the cytoplasm.RNA 1998;4:801–815.
35. Thermann R, Neu-Yilik G, Deters A, Frede U, Wehr K, Hagemeier C,Hentze MW, Kulozik AE. Binary specification of nonsense codons bysplicing and cytoplasmic translation. EMBO J 1998;17:3484–3894.
36. Zhang J, Maquat LE. Evidence that translation reinitiation abrogatesnonsense-mediated mRNA decay in mammalian cells. EMBO J 1997;16:826–833.
37. Zhang S, Welch EW, Hagan KW, Brown AH, Peltz SW, Jacobson A.Polysome associated mRNAs are substrates for the nonsense mediatedmRNA decay pathway in Saccharomyces cerevisiae. RNA 1997;3:234–244.
38. He F, Jacobson A. Identification of a novel component of the nonsense-mediated mRNA decay pathway using an interacting protein screen.Genes Dev 1995;9:437–454.
39. Lee BS, Culbertson MR. Identification of an additional gene required foreukaryotic nonsense mRNA turnover. Proc Natl Acad Sci USA 1995;92:10354–10358.
40. Cui Y, Hagan KW, Zhang S, Peltz SW. Identification and characterization ofgenes that are required for the accelerated degradation of mRNAscontaining a premature translational termination codon. Genes Dev1995;9:423–436.
41. He F, Brown AH, Jacobson A. Upf1p, Nmd2p, and Upf3p are interactingcomponents of the yeast nonsense-mediated mRNA decay pathway. MolCell Biol 1997;17:1580–1594.
42. Perlick HA, Medghalchi SM, Spencer FA, Kendzior RJ Jr, Dietz HC.Mammalian orthologues of a yeast regulator of nonsense-transcriptstability. Proc Natl Acad Sci USA 1996;93:10928–10932.
43. Applequist SE, Selg M, Roman C, Jack HM. Cloning and characterizationof HUPF1, a human homologue of the Saccharomyces cerevisiae non-sense mRNA- reducing UPF1 protein. Nucleic Acids Res 1997;25:814–821.
44. Sun W, Perlick HA, Deitz HC, Maquat LE. The mutated human homologueof yeast Upf1 protein has a dominant negative effect on the decay ofnonsense-containing mRNAs in mammalian cells. Proc Natl Acad Sci USA1998;95:10009–10014.
45. Czaplinski K, Weng Y, Hagan KW, Peltz SW. Purification and characteriza-tion of the Upf1p: a factor involved in translation and mRNA degradation.RNA 1995;1:610–623.
46. Altamura N, Groudinsky O, Dujardin G, Slonimski PP. NAM7 nuclear geneencodes a novel member of a family of helicases with a Z1–6n-ligand motifand is involved in mitochondrial functions in Saccharomyces cerevisiae. JMol Biol 1992;224:575–587.
47. Cui Y, Dinman JD, Peltz SW. mof4–1 is an allele of the UPF1/IFS2 genewhich affects both mRNA turnover and -1 ribosomal frameshifting effi-ciency. EMBO J 1996;15:5726–5736.
48. Atkin AL, Altamura N, Leeds P, Culbertson MR. The majority of yeast UPF1co-localizes with polyribosomes in the cytoplasm. Mol Biol Cell 19956:611–625.
49. Atkin AL, Schenkman LR, Eastham M, Dahlseid JN, Lelivelt MJ, Culbert-son MR. Relationship between yeast polyribosomes and Upf proteinsrequired for nonsense mediated mRNA decay. J Biol Chem 1997;272:22163–22172.
50. Koonin EV. A new group of putative RNA helicases. Trends Biochem Sci1992;17:495–497.
51. Stansfield I, Tuite MF. Polypeptide chain termination in Saccharomycescerevisiae. Curr Genet 1994;25:385–395.
52. Buckingham R, Grentzmann G, Kisselev L. Polypeptide chain releasefactors. Mol Microbiol 1997;24:449–456.
53. Frolova L, Le Goff X, Rasmussen HH, Cheperegin S, Drugeon G, Kress M,Arman I, Haenni AL, Celis LE, Phillippe M, Justesen J, Kisselev L. A highlyconserved eukaryotic protein family possessing properties of a polypep-tide chain release factor. Nature 1994;372:701–703.
54. Frolova L, Le Goff X, Zhouravleva G, Davydova E, Philippe M, Kisselev L.Eukaryotic polypeptide chain release factor eRF3 is an eRF1- andribosome-dependent guanosine triphosphatase. RNA 1996;4:334–341.
55. Stansfield I, Jones KM, Kushnirov VV, Dagakesamanskaya AR, PoznyakovAI, Paushkin SV, Nierras CR, Cox BS, Ter-Avanesyan MD, Tuite MF. Theproducts of the SUP45(eRF1) and SUP35 genes interact to mediatetranslation termination in Saccharomyces cerevisiae. EMBO J 1995;14:4365–4373.
56. Zhouravleva G, Frolova L, LeGoff X, LeGuellec R, Inge-Vechtomov S,Kisselev L, Phillippe M. Termination of translation in eukaryotes isgoverned by two interacting polypeptide chain release factors, eRF1 andeRF3. EMBO J 1995;14:4065–4072.
57. Sherman F. Suppression in the yeast Saccharomyces cerevisiae. In:Strathern J, Jones E, Broach J, editors. The molecular biology of the yeastSaccharomyces cerevisiae: metabolism and gene expression. Cold-Spring Harbor, New York: Cold Spring Harbor Laboratory; 1982. p463–486.
58. Henry M, Borland CZ, Bossie M, Silver PA. Potential RNA binding proteinsin Saccharomyces cerevisiae identified temperature-sensitive mutationsin NPL3. Genetics 1996;142:103–115.
59. Kessler MM, Henry MF, Shen E, Zhao J, Gross S, Silver PA, Moore CL.Hrp1, a sequence specific RNA binding protein that shuttles between thenucleus and the cytoplasm, is required for mRNA 38 end formation inyeast. Genes Dev 1997;11:2545–2556.
60. Shen EC, Henry MF, Weiss VH, Valentini SR, Silver PA, Lee MS. Argininemethylation facilitates the nuclear export of hnRNP proteins. Genes Dev1998;12:679–691.
61. Muhlrad D, Parker R. Premature translational termination triggers mRNAdecapping. Nature 1994;370:578–581.
62. Hsu C, Stevens A. Yeast cells lacking 58=38 exoribonuclease I containmRNA species that are poly(A) deficient and partially lack the 58 capstructure. Mol Cell Biol 1993;13:4826–4835.
63. Hatfield L, Beelman CA, Stevens CA, Parker R. Mutations in trans-actingfactors affecting mRNA decapping in Saccharomyces cerevisiae. MolCell Biol 1996;16:5830–5838.
Review articles
BioEssays 21.8 695

64. Boeck R, Lapeyre B, Brown C, Sachs AB. Capped mRNA degradationintermediates accumulate in the yeast spb8–2 mutant. Mol Cell Biol1998;18:5062–5072.
65. Gera JF, Baker EJ. Deadenylation-dependent and -independent decaypathways for a1-tubulin mRNA in Chlamydomonas reinhardtii. Mol CellBiol 1998;18:1498–1505.
66. Caceres JF, Screaton GR, Krainer AR. A specific subset of SR proteinsshuttles continuously between the nucleus and the cytoplasm. Genes Dev1998;12:55–66.
67. Blencowe BJ, Issner R, Kim J, McCaw P, Sharp PA. New proteins relatedto the Ser-Arg family of splicing factors RNA 1995;1:852–865.
68. Didichenko SA, Ter-Avanesyan MD, Smirnov VN. EF-1a-like ribosome-bound protein of yeast Saccharomyces cerevisiae. Eur J Biochem1991;198:705–711.
69. Stansfield I, Grant CM, Akhmaloka, Tuite MF. Ribosomal association of theyeast SAL4(SUP45) gene product: implications for its role in translationfidelity and termination. Mol Microbiol 1992;6:3469–3478.
70. Cairns BS. Chromatin remodeling machines: similar motors, ulteriormotives. Trends Bioch Sci 1998;23:20–25.
71. Ross J. mRNA stability in mammalian cells. Microbiol Rev 1995;59:423–450.
72. Jacobson A. Poly(A) metabolism and translation: the closed loop model.In: Hershey JWB, Mathews MB, Sonenberg N, editors. Translational
control. Cold Spring Harbor, New York: Cold Spring Harbor LaboratoryPress; 1996. p 451–480.
73. Caponigro G, Parker R. Multiple functions for the poly(A)-binding protein inmRNA decapping and deadenylation in yeast. Genes Dev 1995;9:2421–2432.
74. Wickens M, Anderson P, Jackson RJ. Life and death in the cytoplasm:messages from the 38 end. Curr Opin Genet Dev 1997;7:220–232.
75. Andjelkovic N, Zolnierowicz S, Van Hoof C, Goris J, Hemmings BA. Thecatalytic subunit of protein phosphatase 2A associates with the translationtermination factor eRF1. EMBO J 1996;15:7156–7167.
76. Vincent A, Newnam G, Liebman SW. The yeast translational allosuppres-sor, SAL6: a new member of the PP1-like phosphatase family with a longserine-rich N-terminal extension. Genetics 1994;138:597–608.
77. Ruiz-Echevarria MJ, Yasenchak JM, Han X, Dinman JD, Peltz SW. TheUpf3 protein is a component of the surveillance complex that monitorsboth translation and mRNA turnover and affects viral propagation. ProcNatl Acad Sci USA 1998;95:8721–8726.
78. Dinman JD. Ribosomal frameshifting in yeast viruses. Yeast 1995;11:1115–1127.
79. Farabaugh PJ. Programmed translational frameshifting. Microbiol Rev1996;60:103–134.
80. Gesteland RF, Atkins JF. Recoding: Dynamic reprogramming of transla-tion. Ann Rev Biochem 1996;65:741–768.
81. Brierley I. Ribosomal frameshifting on viral RNAs. J Gen Virol 1995;76:1885–1892.
Review articles
696 BioEssays 21.8