skeletal morphology of two controversial poecilosclerid
TRANSCRIPT

HELGOL,~NDER MEERESUNTERSUCHUNGEN Helgol~inder Meeresunters. 50, 369-390 (1996)
Skeletal morphology of two controversial Poecilosclerid genera (Porifera, Demospongiae):
Discorhabde l la and C r a m b e
M. M a l d o n a d o 1'2" & M. J. Uriz 2
1 Harbor Branch Oceanographic Institution, Division of Marine Sciences, Department of Larval Ecology; 5600 U.S. i North, Fort Pierce, FL 34946, USA
2 Centro de Estudios Avanzados de Blanes (C.S.I.C.), Department of Aquat ic Ecology; Camino de Santa BArbara s/n, 17300 Blanes, Girona, Spain
ABSTRACT: The genera Discorhabdella and Crambe are characterized by bearing uncommon spicule types, i.e. pseudoastrose acanthostyles and sphaeroclones, respectively. They have tradition- ally been considered to be unrelated taxa, but the present reexamination made evident that an important amount of skeletal features are shared by both. Some of these morphological features, such as the ornamentation on the point of the ectosomal subtylostyles, are reported for the first time. The study also revealed that a tuberose nature of the tyles of the main choanosomal megascleres could be a common ancestral condition in both genera. The morphology of the multi-toothed anchorate chelae showed a gradual transition across the species, suggesting that the morphological diversity in chelae was generated in these genera through a "palmate-anchorate-arcuate" evolutionary sequence. However, the forward or backward direction of this sequence remained unclear from the available evidence. Important levels of skeletal variability were found to affect many of the skeletal characters, especially in the genus Crambe. In some cases, this variabihty transgressed the limits theoretically defining a species, making evident that the traditional proce- dure just based on comparison of the skeletons becomes unreliable when tackling the taxonomy of these genera. Most of the skeletal variability seemed to correspond to genetic polymorphisms, except in the case of C. acuata. In this taxon, the skeletal variability could be a result of the existence of a cryptic species, originated by a misconceived synonymy be tween C. acuata and C. chelastra. Besides the skeletal variability, the obscure taxonomic meaning of many skeletal features favored the existence of conflicting taxonomic proposals for the suprageneric location of these genera, depending on the author's criteria. This study made evident that any subsequent attempt of phylogenetic inference should be based on an unweighted analysis of the available skeletal information.
I N T R O D U C T I O N
Discorhabdella Dendy , 1924 a n d Crambe V o s m a e r , 1880 are two poec i lo sc l e r id
g e n e r a c h a r a c t e r i z e d by b e a r i n g p s e u d o a s t r o s e a c a n t h o s t y l e s a n d s p h a e r o c l o n e d e s m a s ,
r e spec t ive ly . T h e s e are two sp icu le t y p e s real ly r a r e in l iv ing poec i lo sc l e r ids , bu t
re la t ive ly f r e q u e n t in fossil mater ia l . In gene ra l , Discorhabdella a n d Crambe h a v e
t rad i t ional ly b e e n c o n s i d e r e d to b e u n r e l a t e d , e x c e p t for t he s h a r e d p r e s e n c e of a n c h o -
�9 Addressee for all correspondence: M. Maldonado, Centro de Estudios Avanzados de Blanes (CSIC), Department of Aquatic Biology, Camino de Santa B~rbara s/n, Blanes-17300, Girona, Spain.
�9 Biologische Anstalt Helgoland, Hamburg

370 M. Ma ldonado & M. J. Uriz
rate i sochelae (Hajdu et al., 1994J. However , there is an incomple te puzzle of information sur rounding both genera , which makes a reexamina t ion and re in te rpre ta t ion of the ske le ta l features obligatory. The lack of information to support any s u p r a g e n e n c assign- ment is c lear ly ref lected in the erratic taxonomical history of these taxa, which comprises a list of conflict ing proposals. The genus Crambe has successively been p l a c e d in various poeci loscler id families, such as Desmac idon idae (Vosmaer, 1880; Topsent. 1928), Myxil- hdae (Burton. 1929), Crambidae (L~vi 1963: Boury-Esnault , 1971: M a l d o n a d o & Benito, 1991), Esper iops idae (L~vi, 1973). It has even been ass igned to other orders or sub-orders different from Poecilosclerida, such as Lithistida (Lendenfeld, 1903), Had romer ida (Schmidt, 1879), and Hal ichondr ida (Laubenfels, 1936). The relat ionships of the genus Discorhabdella remain similarly controversial . The first known species of this genus was ass igned to Hymedesmi idae , under the name of Hymeraphya tuberosocapitata Topsent. 1892. The genus Discorhabdella was later e rec ted by Dendy (1924) to ha rbor the species Discorhabdella incrustans. The genus was init ial ly p laced in the H a d r o m e r i d a (family Spirastrel l idae, sub-family Spirastrel l inae) by p resuming an astrose na tu re for its acan~ thostyles. Subsequent ly , the genus was moved into the family Eurypon idae by Topsent (1928), who assumed homology b e t w e e n the pseudoas t rose acanthosty les and the acan- thostyles of Eurypon. Recently, the genus Discorhabdella has been r e loca ted within the family H y m e d e s m i i d a e by Boury-Esnaul t et al. (1992), who descr ibed a third species, Discorhabdella hindei Boury-Esnault , Pansini & Uriz, 1992.
Observat ions on the spicule on togeny of the genera Crambe and Discorhabdella have revea led a po lyaxonid origin of both the sphaeroclones of Crambe and the pseudoas t rose acanthostyles of Discorhabdella (Maldonado, 1993; Uriz & Maldonado , 1995). Never theless , the shape difference b e t w e e n sphaeroclones and pseudoas t rose acanthosty les also indicates that an unknown amount of evolut ionary d i v e r g e n c e has t aken p lace be tween the two genera . This p a p e r invest igates the way in which this evolut ionary process is ref lected by the r ema in ing skele ta l characters. The goa l is to gain a be t te r k n o w l e d g e of the phy logene t i c re la t ionships be tw e e n these controversial poeci loscler ids as well as of the evolut ionary pa thways connect ing dif ferent spicule forms.
MATERIAL AND METHODS
The following holotypes were r eexamined for skele ta l character iza t ion: Discorhab- della tuberosocapitata (Monaco Museum, MN-040323); Discorhabdella mcrustans (Bri- tish Museum of Natura l History, BMNH: 1923.10.1.157); Discorhabdella hindei (Museum Nat ional d 'Histoire Nature l le de Paris, MNHN-LBIM, D-NBE-1991-1); Crambe tuberosa (Centro de Estudios Avanzados de Blanes, CEAB-ALB-44-C); Crambe erecta (Museo Civico de Storia Naturale , MCSN-RN KEN 57C.E. 48311). Other s tudied ma te r i a l was as follows: spec imens of Crambe acuata from Namib ia (Benguela-VI and VII cruises, 1984; see Uriz, 1988); spec imens of Crambe tailliezi from the Ca ta lan coasts of Spa in (Eco- pharm-I cruise, see Uriz et al., 1992) and from the Alboran Is land (Fauna- I cruise, see Maldonado , 1993; Templado et al., 1993); spec imens of Crambe c r a m b e from the Nor theas t coast of Spain and the Balear ic Is lands (CEAB collection), as we l l as numerous sl ides s tored in the collection of the Stat ion Mar ine d 'Endoume (mater ia l col lected by Topsent, Vacele t & Boury-Esnault).
Spicule micrographs were t aken th rough a Hitachi SEM, after c leaning , dehydra t ion

Skele ta l morphology of Discorhabdella and Crambe 371
and coat ing with go ld -pa l l ad ium according to the s tandard method (e.g. Boury-Esnaul t et al., 1992).
SKELETAL CHARACTERIZATION
Genus Crambe Vosmaer
This genus contains five l iving species. In addit ion, a fossil species from New Zea land original ly descr ibed under the name of Vetulina oamaruensi Hinde & Hol- mes,1882 has recent ly been c la imed as be long ing to the genus Crambe (Uriz & Maldo- nado, 1995). The character iza t ion of the Hving species is as follows:
Crambe tuberosa Maldonado & Benito, 1991
Species only known by a unique, small encrust ing spec imen col lected from the coral l igenous facies of Alboran Island. The spicule set consists of tuberose choanosomal acanthosty les (Fig. 1A), ectosomal subtylostyles, sphaeroclone desmas (Fig. 3A, B), and f ive- toothed i soanchorae (Fig. 5A-C), occasional ly with three teeth only. Desmas have a crown of short sp ine- l ike actines on the centrum and be tw e e n four and seven, long, downwards - cu rved act ines n a m e d "arms". A flat expans ion with an i r regular outline occurs at the tip of the arms, usual ly n a m e d "zygomat ic pla te" .
Three ske le ta l features, unnot iced in the original description, have b e e n r evea l ed by the p resen t study: (1) Occurrence of a profuse microspinat ion at the po in ted end of the ec tosomal subtylostyles (Fig. 2A); (2) Partial fusion of the la teral teeth of che lae (Fig. 5B) and occasional reduct ion of the central tooth (Fig. 5A); (3) Spicules with acan those tyles, or iginal ly desc r ibed as tylostyles-II (Maldonado & Benito, 1991), have been conf i rmed to be ontogenet ic s tages of the tuberose tylostyles (see Uriz & Maldonado, 1995).
Crambe tailliezi Vacele t & Boury-Esnault , 1982
Species formerly discovered off the Med i t e r r anean coasts of France and subse- quent ly found in other Western Med i t e r r anean locations (Bibiloni, 1990; Uriz et al., 1992; Maldonado, 1993).
The main monact ines are tylostyles with a sl ightly inf lated tyle that was weak ly tuberose in some specimens from the Cabre ra (Fig. 1B) and Alboran Is land (Fig. 1C). The ectosomal megasc le re s are subtylostyles exhibi t ing a microspiny point, whose o rnamen- tat ion can be sl ightly or in tensely spiny, d e p e n d i n g on the spec imens (Fig. 2B, C). Desmas are a lways presen t in all specimens, a l though somet imes they are rare. Their a p p e a r a n c e ranges from an astroclonal shape to a sphaeroclonal shape (sensu Reid, 1970), d e p e n d i n g on the specimens. The former shows radia t ing arms with weak ly developed, or even absent , zygomat ic p la tes (Fig. 3E, F), w he re a s the la t ter has all arms addres sed to the same direction and large compl ica ted zygomat ic plates (Fig. 4D). The microscleres are th ree- too thed i soanchorae (Fig. 5D), somet imes with r e d u c e d teeth, a long with sl ightly fusiform, microspiny microxeas (Fig. 6D).
Crambe acuata L@vi, 1958
Species k n o w n from the Atlantic and East coasts of Africa, the coasts of Madagasca r , and the Red Sea. We have noted that the whole amount of spec imens a s s igned to this species exhibi ts a grea t skele ta l variation. However , this variat ion can bas ica l ly be

372 M. Ma ldonado & M. J. Uriz
organized into two skele ta l groups (henceforth referred to as C. acuata-I a n d C. acuata-
II). These groups show disjunct b iogeograph ica l distributions, in the South Atlant ic and in the Indian O c e a n - R e d Sea, respect ively.
C. acuata-I is charac ter ized by choanosomal tylostyles with globose tyles (Fig. 1D), ectosomal subtylostyles, scarce as teroid desmas, three- toothed isochelae in one or two size categories , and spiny microxeas (Fig. 6A, B). Desmas are absen t in spec imens descr ibed from M a d a g a s c a r and Aldab ra (L6vi, 1958; Vacelet et al., 1976). Microxeas are also absent in some spec imens (L6vi, 1958, 1961).
C. acuata-II, i.e. South Atlantic spec imens descr ibed by L~vi (1960, 1963) and Uriz (1988), is charac te r ized by the occurrence of abundan t desmas with l a rge zygomat ic plates (Figs 3C, 4C), the occurrence of only one category of isochelae (Fig. 5E), and the presence of s igmoid e lements (Fig. 5G). The ectosomal subtylostyles of these spec imens show a w e a k microspinat ion ident ica l to that i l lustrated in C. taiiliezi in F igure 2C.
Crambe crambe (Schmidt, 1868)
It is one of the most a b u n d a n t sponges in the infralittoral M e d i t e r r a n e a n assem- blages. Its geograph ica l dis t r ibut ion covers the whole Western Med i t e r r anean , even reaching Atlantic zones, such as the Canary Islands and the Saint Vincen t Cape (Portugal). Spec imens display thin or thick encrust ing growth habits. The spicule set usual ly consists of stylesosubtylostyles in two categories of thickness, co r respond ing to choanosomal and ectosomal locat ions (Fig. 1F). These spicules have an i r r egu la r point, usual ly te lescopic (Fig. 2E). The microspinosi ty not iced in other species is he re r e duc e d to a rugose a p p e a r a n c e (Fig. 2F). These spicules may also be t ransformed into a kind of anisos t rongyles or asymmetr ica l styles, especia l ly in oligotrophic env i ronments (Bibiloni, 1990). Other spicule types, such as th ree- too thed anchorate i sochelae and as teroid desmas (Fig. 3G), have only b e e n repor ted in a few specimens (e.g. Topsent , 1925; Boury- Esnault, 1971). These spicules usual ly have aber ran t morphologies , such as unde rdeve l - oped teeth in chelae and r educed arms in desmas. Only those de smas obse rved in Topsent ' s sl ides (specimens from Banyuls) were reasonably well formed, d i sp lay ing a shape similar to that shown by some desmas of C. erecta (Fig. 3D).
C rambe erecta Pulitzer-Finali , 1992
Species known from only one spec imen col lected from North Kenya Banks (East African coasts). Unlike the r ema in ing species of this genus, the ho lo type is an erect, compressed, mass ive specimen. The spicule set consists of choanosomal subtylostyles with a sl ightly m a r k e d tyle (Fig. 1E), ec tosomal subtylostyles, asteroid d e s m a s (Fig. 3D), and anchora te isochelae in two size categories , but without any morpho log ica l differ- ent ia t ion (Fig. 5F). The point of ec tosomal subtylostyles is sl ightly microspiny, d i sp lay ing an ornamenta t ion in te rmedia te b e t w e e n that of C. crambe and those of spec ies Disco- rhabdella (Fig. 2D). The third ca tegory of rud imenta ry i sochelae m e n t i o n e d by Pulitzer- Finali (and sugges t ed as uncer ta in ly proper) was not found in the p r e se n t holotype reexaminat ion . Desmas are astroclons d isp lay ing a morphology t rans i t ional b e t w e e n those of C. acuata and C. crambe. Most of them have long arms whose zygomat i c p la tes are r educed or even absent (Fig. 4D).

Skeletal morphology of Discorhabdella and Crambe 373
Skeletal a r rangement in Crambe
In all five species of the genus Crambe, the choanosomal spicule skeleton is settled on a thin, basal layer of spongin. However, the a r rangement of the spicules shows some differences depend ing on the species and even on the specimens, as follows:
(1) In C. tuberosa and in Atlantic specimens of C. acuata-II, desmas are joined by articulate zygoses to make a basal mult i layered skeleton (Fig. 4A, C), which is just monolayered in some specimens of C. tailliezi (Fig. 4D). In some specimens of C. tailliezi, C. erecta, C. acuata-II, and C. crambe, desmas occur as scattered elements that are not joined one to another by zygoses (Topsent, 1925; L~vi, 1958, 1961; Boury-Esnault, 1971; Vacelet et al., 1976). They can even be absent in most specimens of the last two species.
The general structure of fossil desma-skeletons of Sphaerocladina, as described by Moret (1925: 134), was identical to that hereby illustrated from Crambe tuberosa, even in the minor details of the zygoses. The zygomatic plates of a desma are settled on the centrum and branches of adjacent and subjacent desmas, closely surrounding one or several spine-l ike structures (Fig. 4B, D).
(2) Main monact ines are in a hymedesmioid pattern, traversing the desma layer in C. tuberosa and C. tailliezi. They are in a plumose fashion in C. erecta and C. crambe. Both ar rangements can occur in C. acuata (Fig. 4C), depend ing on the specimens (see L6vi, 1958, 1960, 1963; Vacelet et al., 1976; Uriz, 1988). The amount of spongin in the spiculose bundles becomes important in C. erecta and C. crambe.
(3) Ectosomal subtylostyles are tangent ia l to the sponge surface in C. tuberosa, whereas they are placed obliquely, forming bundles in C. crambe and C. erecta. Both ar rangements occur in C. acuata and C. tailliezi, depend ing on the specimens.
(4) Isochelae and microrhabds, if present, are especially abundan t in the ectosome, al though they can also be scattered throughout the choanosome.
Genus Discorhabdella Dendy
The genus Discorhabdella has three living species. However, a fossil species from New Zealand, originally described in the genus Latrunculia by Hinde & Holmes, 1892, was recently reinterpreted as be longing to the genus Discorhabdella by Boury-Esnault et al. (1992). In general, the scarcity of records does not allow a comprehensive interpreta- tion of either the actual biogeographical or bathymetr ic distribution. The few available data suggest that this genus has epibathyal depth distribution in temperate and subtropi- cal latitudes. The characterization of the hying species is as follows:
Discorhabdella incrustans Dendy, 1924
Indo-Pacific species that is only known from one specimen. The spicule set consists of tuberose tylostyles (Fig. 7D), ectosomal subtylostyles with shghtly microspiny points (Fig.8C), short pseudoastrose acanthostyles without distal point {Fig. 9C), five-toothed isoanchorae usual ly with divided teeth (Fig.10F, G), and microxeas ("oxydiscorhabds" sensu Dendy,1924) with the spines mostly concentrated in two whorls (Fig. 6D, E).
Discorhabdella hindei Boury-t/snault, Pansini & Uriz, 1992
Species known from four specimens collected on the Medi ter ranean side of the Straits of Gibraltar. The spicule set consists of irregularly tuberose tylostyles (Fig. 7A),

374 M. Ma ldonado & M. J. Uriz
ectosomal subtylostyles with microspiny points {Fig. 8B), astrose acanthos ty les with or without a short distal po in t (Fig. 9B), anchora te i sochelae with s even or e ight teeth, somet imes dicho- or t r ichotomical ly d iv ided (Figl 10D, E), and s igmoid spicules.
Discorhabdella tuberosocapitata Topsent, 1892.
Species k n o w n from a s ingle spec imen from the Azores. Its spicule set consists of tuberose tylostyles (Fig~ 7B, C), ec tosomal subtylostyles with microspiny points (Fig. 8A), basa l acanthos ty les with " regular , ( n o n - a s t r o s e ) a p p e a r a n c e (Fig. 9A), and anchora te i sochelae with seven, occasional ly eight, tee th (Fig. 10A-C).
Skele ta l a r r a n g e m e n t in Discorhabdella The ske le ta l a r r angemen t is common to all three l iving species of the genus. It
consists of a thin, basa l layer of spongin on which pseudoas t rose acanthos ty les (or typical acanthosty les in the case of D. tuberososcapitata) are dense ly set t led in ver t ica l orienta- tion, forming a monostrat i f ied layer. I so la ted tuberose tylostyles are spa r se ly sca t tered among the pseudoas t rose acanthostyles . Subtylostyles are g rouped in thin bundles , p redominan t ly p laced in obl ique fashion to the ectosome. Anchora te i sochelae , spiny microrhabds and s igmoids are w i d e s p r e a d in the ectosome, and maybe in the choano- some too, a l though we were unable to corroborate this fact.
REMARKABLE FEATURES
Some morphologica l features deserve special considera t ion because they are ei ther new findings or fea tures shared by both genera , which can help us to u n d e r s t a n d their phylogene t ic relat ionships:
(1) Tuberose tyles are presen t in all species be long ing to Discorhabdella (Fig. 7). In the genus Crambe, this character was ev ident in C. tuberosa and some spicules of some spec imens of C. tailliezi (Fig. 1A-C). In spicules of C. acuata and C. erecta, t he re are only slight swell ings; these, however , can be in te rpre ted as remains denot ing the ancest ra l p resence of a tuberose tyle (Fig. 1D, E). This evolut ionary reduct ion p rocess of the tyle would cu lmina te in its loss in C. c rambe (Fig. 1F).
The shaft of these main megasc le res is apparen t ly smooth, but some scarce and very tiny spines were revea led a round the proximal zone under SEM magnif ica t ion.
(2) Accord ing to the recent confirmation of a po lyaxonid origin for sphae roc lones and pseudoas t rose acanthostyles (Uriz & Maldonado , 1995), it can be said that Discorhabdella and Crambe share the p resence of a basa l ske le ta l layer made of po lyaxon id spicules.
These basa l e lements can be dense ly a r ranged , somet imes even jo ined by zygoses in the case of desmas. In these art iculate zygoses, the perfect fitting of the zygomat i c plates a round the sp ine- l ike structures of ad jacen t desmas sugges ts that g rowth of the spine- l ike s t ructures has to be f inished before the zygomat ic p la tes contact wi th them. From morphologica l observat ions, it has b e e n pos tu la ted that the si l icification process in desmas has two phases (L6vi, 1991; Uriz & Maldonadol 1995). The first phase , where sil icification is gu ided by the axial f i lament, would yield crepis of very cons tan t shape. After this phase , a subsequen t silicification charac te r ized by little control on the silica deposi t ion has been thought to take place, since the final shape of d e s m a s is cer ta inly p leomorphic . However , it must be conceded that to get a perfect f i t t ing b e t w e e n

Skeletal morphology of DiscorhabdeHa and Crambe 375
zygomatic plates and spine-l ike structures, an accurate control of the silica deposition is needed. This assumption is also reinforced by other invariant features in the morphology of desmas, such as the presence of spines on the arms following a regular pat tern (Fig. 4A) and the occurrence of smooth surfaces at the inferior side of desmas (Fig. 3B).
Desmas show a gradual morphological simplification across the set of species of Crambe. The change in shape, which ranges from sphaeroclone to astroclone morpholo- gies (Fig. 3), is associated to a rarefaction in the number of desmas. This process seems to be culminated in C. crambe, where desmas are absent in most specimens.
As for the astrose acanthostyles, the most remarkable fact is the apparent evolution- ary transfiguration from a pseudoastrose morphology, present in D. hindei, D. incrustans and fossil spicules, to a typical acanthostilic morphology, such as that occurring in D. tuberosocapitata (Fig. 9). Acanthostyles of D. tuberosocapitata have the "proximal spines" more developed than the remaining spines on the shaft (Fig. 9). This is also known in many other poecilosclerid groups. However, it has been demonstrated that the proximal spines, at least in Discorhabdella, are different from the remaining spines on the shaft because they are actually reduced actines containing an axial canal (see Uriz & Maldonado, 1995).
13) Straight ectosomal subtylostyles were present in all species of both taxa. Choano- somal and ectosomal styles have not usually been described separately in C. crambe, but we have observed that they are of two kinds that can be dist inguished by their skeletal a r rangement and by their thickness.
There is a peculiar microspination surrounding the point of these ectosomal subtylo- styles. This ornamentat ion, recently reported for the first time in Crambe tuberosa (Maldonado, 1993), has also been found in all species of Crambe and Discorhabdella. The intensi ty of the ornamenta t ion displays a variation, especially in Crambe, where it ranges from a strong microspinosity to a rugose outl ine (Fig. 2). The spiny character of the point is moderate in all three species of Discorhabdella (Fig. 8). This pattern of ornamentat ion, characterized by spines around the point, clearly differs from that observed in species of Myxilla, where spines are located at the top of the points and sometimes even splitting the point of the spicule.
(4) Both genera share the presence of anchorate isochelae whose n u m b e r of teeth ranges from three to eight across the species. Morphological evidence suggests that the n u m b e r of teeth in chelae is currently subjected to a process of evolution. However, it is not clear whether the process involves an increase or a decrease in n u m b e r of teeth. The three-toothed isoanchorae of C. crambe, C. tailh'ezi, C. acuata and C. erecta show interest ing peculiarities. In Crambe acuata, the three-teeth isoanchorae have one or two additional small teeth (either vestigial or new formation?) located at the base of the lateral or central teeth (Fig. 5E). The isoanchorae of C. erecta sometimes show a small fissure on the alae (Fig. 5F) which could mean either remains of the fusion of a lateral tooth or the beg inn ing of neogenesis of a lateral tooth. Isochelae of some specimens of C. crambe sometimes show all three teeth reduced to unguiferous structures; most f requently isochelae are absent in this species. It is noteworthy that, in the specimen of C. taitliezi from Alboran, some chelae are three-toothed at one end but five-toothed at the other end (Fig. 5D).
In general, in species having chelae with five or more teeth, the central tooth of these chelae was more clearly individualized than the remain ing ones, which were fused in

376 M. Maldonado & M. J. Uriz
combinations of two or three at their bases (Figs 5B, C, 10A, C). Isoanchorae with seven
teeth usually occurred in C. tuberosa, Discorhabdella tuberosocapitata and D. hindei. In
such chelae, the central tooth remained isolated, whereas the lateral tee th were also
diversely fused in groups of two or three (Fig. 10A, D). The central tooth was only found to
be dichotomous in some eight- toothed isoanchorae of Discorhabdella hindei. Types with three, five and seven teeth exhibi ted a symmetrical a r r angemen t of teeth
and were notably more abundant in the species than types with four, six or e ight teeth.
Isoanchorae with four and six teeth s eemed to be occasional irregularit ies der ived from
the three* and f ive-toothed major models. Types with five and seven tee th also seem to
arise from or converge to the three- toothed model, since lateral teeth are always fused
with each other at their bases (Figs 5B, 10C). This three- toothed morphology, in turn, may be related with the sigmoid spicules occurring in some species, at least in the case of
Crambe. In this genus, the central tooth, unlike the lateral ones, is a real prolongat ion of
the central shaft of the chela (Fig. 5B). Thus, any underdeve lopment of tee th would most
likely originate s igmoid elements. However , s igmoid elements are also present in some
species of Discorhabdella even though the central tooth does not appea r to be a real prolongation of the central shaft of the chelae (Fig. 10C). In general, s igmoid spicules of
both genera look somewhat different from the usual, typical poecilosclerid s igmata
because of their blunt ends and the rugose outline of the shaft. As a curiosity, note the presence of a spine on the shaft of a chelae of D. hmdei in
Figure 10E. (5) Microxeas, either with spines a r ranged in two distal whorls or scat tered through-
out the shaft, occur in several species of both genera (Fig. 6). At first sight, oxydisco-
rhabds might be considered as streptastrose spicules (Fig. 6D). However , after a SEM study of the spination in several ontogenet ic stages no axial canal was observed. Thus,
both types of microrhabds are assumed to be monaxonid microscleres, but having
different patterns of ornamentat ion (Uriz & Maldonado, 1995).
DISCUSSION
The main aim of this study was to find new morphological information to solve the
controversial relationships surrounding the genera Crambe and Discorhabdella. How- ever, a l though a detai led skeletal description of both genera has been accomplished, the
taxonomic interpretat ion of these skeletal features must be tackled cautiously. There is a
high skeletal variabili ty affecting most of the spicule types of Crambe, especial ly in C. crambe and C. acuata. We have not iced that there is almost no skeletal difference
be tween, for instance, some specimens of C. c rambe and some other ass igned to C.
acuata: both lacking isoanchorae and desmas. Similar levels of skeletal c loseness occur be tween specimens of the latter species and C. tailliezi. An additional complicat ion is that
the variability in C. tuberosa and C. erecta can not be assessed since there is only one
spec imen known from each of these two species. On the other hand, most of the skeletal variabil i ty found in C. acuata occurs in the
group of Indian specimens (C. acuata-I), Specimens from the West coasts of Africa (C.
acuata-II) have relat ively stable skeletal features. It is interest ing to r e m e m b e r that the
current concept of Crambe acuata is based on a taxonomic fusion be tw een two previous species: Crambe acuata L~vi, 1958 and Crambe chelastra L~vi, 1960. Al though this

Skeletal morphology of Discorhabdella and Crambe 377
second species was claimed to be a junior synonymy of C. acuata (L6vi, 1963; Vacelet & Boury-Esnault, 1982), we find from the present study that a reconsideration of such a synonymy would be advisable.
The intraspecific skeletal variability in Crambe, except for the above men t ioned case of C. acuata, seems to be a result of genetic polymorphisms, since the same characters appear and disappear at interspecific levels, as typically done by rampant parallelisms. This variability, traditionally considered to be taxonomically useless, can be a potential source of information to clarify the phylogenetic relationships (e.g. Saether, 1979, 1983; Sluys, 1989). However, from a phenet ic analysis it is not possible to dist inguish be tween parallelism and convergence in a reliable way. Consequently, we can not objectively decide a priori which subset of skeletal features is the best to infer the phylogeny in Crambe.
The taxonomical mean ing of some skeletal features (i.e. the morphological gradation in chelae, the presence-absence of sigmoids and microxeas, and the sphaeroclone- acanthostyle relationship) is also too controversial to infer the relationships at the suprageneric level. For example, note that al though we have illustrated a gradual variation in the morphology of the isochelae, this evidence is not enough to demonstrate whether the evolutionary trend is a reduction or an increase in number of teeth. The polarized sequence "palmate --~ arcuate --, anchorate" has recently been suggested by Hajdu et al. (1994) in a revision of this character at the ordinal level. However, our present findings at the generic level rather support the ordination "paImate-anchora te-arcuate" , whereby the direction remains unclear.
Sigmoid e lements occur in some species of both genera. The low frequencies of these elements in the specimens along with some peculiarities in their shape give rise to doubt the homology be tween these "sigmoids" and the typical sigmata of other poecilosclerid sponges such as, for instance, Myxilla or Pronax. Besides, there is the fact that these sigmoid elements have sometimes been interpreted as reduced states of isoanchorae (e.g. Vacelet & Boury-Esnault, 1982; Van Soest, pets. comm.). This interpretat ion agrees with our observations on isoanchorae of Crambe, but it is not supported by observations on isoanchorae of Discorhabdella. Thus, the interpretat ion of this information also remains controversial.
As for the monaxonid microscleres, they only occur in some species of the two genera, and their presence is known to be affected by a high intraspecific variabili ty (e.g. Vacelet et al., 1976). These spicules cannot be directly used to establish the taxonomy because it is u n k n o w n whether this character reflects the pattern of int rageneric specia- tion; or whether it is rather a rampant parallelism indicat ing that Crambe and Discorhab- della are sister groups, or just a morphological convergence in the shape of the micro- scleres.
The taxonomic level at which Crambe and Discorhabdefla are related remains unclear, even taking into account the homology be tween sphaeroclones and pseudo- astrose acanthostyles. It is obvious that these spicules have primitive morphologies. This agrees with the results of some recent studies suggest ing some species of both genera to be very ancient, probably Tethyan Medi te r ranean relicts (Boury-Esnault et al., 1992; Maldonado & Uriz, 1995). According to the fossil record, sphaeroclones and pseudo- astrose acanthostyles already showed their current shape in the Lower TerNary (Hinde & Holmes, 1892). The amount of morphological divergence be tween sphaeroclones, pseu-

378 M. Maldonado & M. J, Uriz
doastrose acanthostyles, and typical acanthostyles (present in D. tuberosocapitata) can-
not be clearly t ranslated into taxonomy. The question remains whether the amount of
change b e t w e e n sphaeroclones and astrose acanthostyles is more impor tant than the
whole set of shared morphological features in determining the phylogene t ic - taxonomic
relationships be tween both genera. There are several conflicting sugges t ions for the
supragener ic taxonomic location of these taxa, depending on the di f ferent taxonomic
value that different authors concede to every particular skeletaI feature. So, for instance, by paying special at tention to the sphaeroclone skeleton, a tentat ive relat ionship of the
genus Crambe with some fossil genera was sugges ted by Maldonado & Benito (1991). This proposal also leads us to the problem of the relationships be tween Crambe and the
enigmat ic genus Vetulina Schmidt, 1879. Such relationships are extremely difficult to be
corroborated since only desma skeletons are conserved from fossil material. Furthermore,
there are two controversial reports about the probable presence of ec tosomal strongyles in living spec imens of Vetulina stalactites (Laubenfels, 1955; Soest & Stentoft, 1988),
which need to be reexamined. As for the genus Discorhabdella, it has recent ly moved
into the family Hymedesmi idae (Boury-Esnault et al., 1992) by consider ing mostly the
skeletal a r rangement . In contrast, Hajdu et al. (1994) suggest a close re la t ionship among Crambe, Nlonanchora, Discorhabdella, and Myxilla because of the shared occurrence of
anchorate isochelae. From the present study, some morphological fea tures suggest a relationship be tween Crambe and Discorhabdella, but any assessment of their relation-
ships with the genera Monanchora and Myxilla is beyond the scope of this study.
It is obvious that to advance in the inference of the taxonomic re la t ionships it is necessary to falsify some of these compet ing taxonomic hypotheses us ing a method
different from the traditional phenet ic we igh ted inspection. This aim is i n t ended in a
separate paper that will invest igate the congruence level among different subsets of skeletal features as well as the stability of the different taxonomic classifications der ived
from them (authors, current research).
Acknowledgements. We thank Dr. N. Boury-Esnault, J. Vacelet, Dr. C. Carpine, Dr. V. Raineri, and Ms. C. Valentine for loaning specimens and/or skeletal slides. For their valuable assistance with SEM and micrographs, we are indebted to the technical staff of the Servicio de Microscopia of the Universidad de Barcelona, Centro de Investigaciones Pesqueras de Barcelona (CSIC~ and Harbor Branch Oceanographic Institution. We thank Dr. R. W. M van Soest and Dr. E. Hajdu for their interesting supply of unpublished information and comments, and Dr. C. M. Young for support in his laboratory. This study was supported with funds from CICYT-MAR-91-0528, and Fauna Ib6rica Projects, along with a Fulbright-MEC grant to M.M. Figures 1B, 3A, 3F, 4D and 7D were taken from Uriz & Maldonado (1995) with permission of the Biological Journal of the Linnean Society. This is contribution number 1084 from Harbor Branch Oceanographic Institution.
LITERATURE CITED
Bibiloni, M. A., 1990. Fauna de esponjas de las Islas Baleares. Variaci6n cualitativa y cuantitativa de la poblacidn de esponjas en un gradiente batim~trico. Comparaci6n Baleares-Costa Catalana. Diss., Univ. Barcelona, 483 pp.
Boury-Esnault, N., 1971. Spongiaires de la zone rocheuse de Banyuls-sur-Mer. II. Syst~matique. - Vie Milieu 22, 287-350.
Boury-Esnault, N., Pansini, M. & Uriz, M. J., 1992. A new Discorhabdella (Porifera, Demospongiae), a new Tethyan relict of pre-Messinian biota? - J. nat. Hist. 26, 1-27.

S k e l e t a l m o r p h o l o g y of D i s c o r h a b d e l l a a n d C r a m b e 379
Burton, M., 1929. Descr ip t ions of Sou th Afr ican s p o n g e s . Part II: The "Lithist idae", w i th a critical su rvey of the d e s m a - f o r m i n g sponges . - Rep; Fish. mar . biol. Surv. S. Air. 7, 1-12.
Dendy , A., 1924. Porifera. Part l: Non-Anta rc t i c s p o n g e s . - Br. antarct . Ter ra Nova Exped . 1910-1913 (Zool.) 6 (3), 269-392.
Hajdu , E., Soest, R. W. M. van & Hooper , J. N. A., 1994. Proposal for a phy logene t i c subo rd ina l c lass i f icat ion of poeci loscler id sponges . In: S p o n g e s in t ime a n d space . Ed. by R. W. M. v a n Soest, Th. M. G. v a n K e m p e n & J. C. B r a e k m a n . Ba lkema , Rot terdam, 123-139.
Hinde , G. J. & Holmes , W. M., 1892. O n the s p o n g e r e m a i n s in the Lower Ter t ia ry Stra ta nea r O a m a r u , N e w Zea land . - J. Linn. Soc. (Zool.) 24, 177-262.
Laubenfe l s , M. W. de, 1936. A d i scuss ion of the s p o n g e f a u n a of the Dry Tor tugas in par t i cu la r and t he W e s t Ind ies in genera l , wi th ma te r i a l for a revis ion of the famil ies and orders of the Porifera. - Pap. T o r t u g a s Lab. 30 (467), 1-225.
Laubenfe l s , M. W. de, 1955. Porifera. In: T rea t i se on inve r t eb ra t e pa leontology. Part E. Ed. by R. C. Moore. Geol. Soc. Am., N e w York, 21-212.
Lendenfe ld , R. von, 1903. Tet raxonia . - Das T ie r re ich 19, 1-168. L6vi, C., 1958. R6sul ta ts sc ien t i f iques des c a m p a g n e s de la "Calypso" . C a m p a g n e 1951-1952 en
M e t Rouge. Spong ia i re s de M er Rouge. - A n n l s Inst. oceanogr . , Monaco 34, 3-46. L6vi, C., 1960. Les spong i a i r e s h d e s m e s ast~ro~des. - Bull. Inst. oceanogr . Monaco 57 (1179), t -8 . Levi, C., 1961. Resu l ta t s sc ien t i f iques des c a m p a g n e s de la "Calypso" . XIV. C a m p a g n e 1954 dans
l ' O c e a n Indien. 2. Les spong ia i r e s de l'Ile Aldabra . - Ann l s Inst. oc~anogr. , M o n a c o 39, 3-32. Levi, C.,1963. Spong ia i re s d 'Afr ica du Sud (1) Poeci loscler ides . - T r a n s . R. Soc. S. Africa 37 (1), 1-71. Levi, C., 1973. S y s t e m a t i q u e de la c lasse des D e m o s p o n g i a r i a (D~mosponges) . In: Tra i te de zoologie.
T. 3, 1: Spongia i res : Ana tomie , phys io logie , sys t~ma t ique , ~cologie. Ed. by P. P. GrassY. Masson , Paris, 577-631.
L6vi, C., 1991. Lithist id s p o n g e s from the Norfolk Rise. Recen t a n d Mesozo ic gene ra . In: Fossil and Recen t s p o n g e s . Ed. by J. Rei tner & H. Keupp . Spr inger , N e w York, 72-82.
Ma ldonado , M. & Benito, J., 1991. C r a m b e tuberosa n. sp. (Demospong i ae , Poecilosclerida): a n e w M e d i t e r r a n e a n poeci loscler id wi th li thistid affinities. - Cah. Biol. mar . 32, 323-332.
Ma ldonado , M., 1993. D e m o s p o n j a s l i torales de Albor~n. Faun i s t i ca y b iogeograffa . Diss., Univ. Barcelona, 499 pp.
Ma ldonado , M. & Uriz, M. J., 1995. Biotic aff ini t ies in a t rans i t ional zone b e t w e e n the At lant ic and the M e d i t e r r a n e a n : an a p p r o a c h b a s e d on s p o n g e s . - J. B i o g e o g r a p h y 22, 89-110.
Motet , L., I925. Cont r ibu t ion a l '~ tude des s p o n g i a i r e s s i l icieus du Cr~tac~ s u p e r i e u r fran~ais. - M~m. Soc. g~ol. Fr. 5, 1-303.
Puli tzer-Finali , G., 1992. A collection of m a r i n e s p o n g e s f rom East Africa. - Anna l i Mus . civ. Stor. nat. G i acomo Doria 89, 247-350.
Reid, R. E. H., 1970. Te t r axons a n d d e m o s p o n g e p h y l o g e n y . - Syrup. zool. Soc. L o n d o n 25, 63-89. Sae ther , O. A., 1979. Unde r l y i ng s y n a p o m o r p h y a n d a n a g e n e t i c analys is . - Zool. Scr. 8, 305-312. Sae ther , O. A., 1983. T he cana l i zed evo lu t ionary po ten t ia l i ncons i s t enc i e s in p h y l o g e n e t i c r eason-
ing. - Syst. Zool. 32, 343-359. Sluys, R., 1989. R a m p a n t paral le l ism: an app ra i s a l of t he u s e of n o n u n i v e r s a l de r ived cha rac t e r s ta tes
in p h y l o g e n e t i c reconst ruct ion. - Syst. Zool. 38, 350-370. Schmidt , O., 1868. Die S p o n g i e n der Kiiste yon Algier . Mit N a c h t r ~ g e n zu d e n S p o n g i e n des
Adr i a t i s chen Meeres . E n g e l m a n n , Leipzig, 44 pp. Schmidt , O., 1879. Report on the Sponges . I. S p o n g i e n des M e e r b u s e n von Mexico. Fischer , Jena ,
32 pp. Soest , R. W. M. v a n & Stentoft , N., I988. B a r b a d o s d e e p - w a t e r s p o n g e s . - Stud. F a u n a Curaqao
Caribb. Isl. 122, 1-175. Templado , J., Guer ra , A., Bedoya, J., Moreno , D., Rembn, J. M., M a l d o n a d o , M. & Ramos , M. A.,
1993. F a u n a circalitoral del sur de la p e n i n s u l a ib~rica. Resu l t ados de la c ampa f i a oceanogr~f ica " F a u n a I". C.S.I.C., Madr id , 133 pp.
Topsen t , E., 1892. Cont r ibu t ion a l '4 tude de s spong i a i r e s de l 'A t l an t ique N o r d . - R~sult. Camp. scient . Pr ince Alber t I 2, 1-165.
Topsen t , E., 1925. E tude de spong ia i r e s du golfe de Naples . - Archs Zool: exp. gen . 63, 623-725. Topsen t , E., 1928. Spong ia i re s de l 'A t l an t ique et de la M~di te r ran~e , p r o v e n a n t de s croissi~res du
Prince Alber t I de Monaco. - Result. C a m p . scient . Pr ince Alber t I Y4, 1-376.

380 M. M a l d o n a d o & M. J. Ur i z
Uriz, M. J., 1988. D e e p - w a t e r sponges from the cont inenta l shelf and slope of Namib ia (South-West Africa): Classes Hexact ine l l ida and Demospongia . - Monogr . Zool. mar. 3, 9-157.
Uriz, M. J. & Maldonado , M., 1995. A recons idera t ion of the relat ionship b e t w e e n po lyaxonid and monaxon id spicules in Deraospong iae (Porifera): n e w data from the genera Crambe and Discor- habdella. (Porifera)- Biol. J. Linn. Soc. 55, 1-15~
Uriz, M. J., Rosell, D. & Martin, D., 1992. The sponge popula t ion of the C ab re r a Archipe lago (Balearic Islands): characterist ics, distr ibution and a b u n d a n c e of the most r ep r e sen t a t i ve species.
- Mar. Ecot. 13, 101-117. Vacelet , J, & Boury-Esnault , N., 1982. Une nouvel le ~ponge du genre Crambe (Demospongiae ,
Poecilosclerida) de Medi te r ran6e , C. tailliezi n. sp. - Tray. scient. Parc. na tn . Port-Cros 8, 107-113.
Vacelet , J., Vasseur , P. & Levi, C., 1976. Spongia i res de ta pen te ex terne des r~cifs corall iens de Tul~ar (Sud-Ouest de Madagascar) . - Mem. Mus. natn. Hist. nat. Paris [Ser. A: Zool.) 99, 1-116.
Vosmaer , G. C. J., 1880. The sponges of the Leyden Museum. I. The family of the Desmac id inae . - Not. Leyden Mus. II, 18, 99-154.

O
/.
"++~
X �9
"+
O
/
~8
o4
t5KV ii~ .
...
i~gu
m
:;(<
�9
+'.'
, ..
..
''2
'+.
(/3
o ~r o o o n,
r~
(3"
tb
Fig
. 1.
Ma
in c
ho
an
oso
ma
l m
on
ac
tin
es
of C
ram
be.
A:
C.
tub
ero
sa.
No
te t
he
co
mp
ara
tiv
e s
ize
be
twe
en
ch
oa
no
som
al
an
d e
cto
som
al
mo
na
cti
ne
s.
B:
C.
tail
liez
i, s
pe
cim
en
fro
m t
he
Ca
bre
ra I
slan
d (
fro
m U
riz
& M
ald
on
ad
o,
19
95
by
co
urt
esy
of
Bio
l. J
. L
inn
. S
oc.)
. C
: C
. ta
illi
ezi,
sp
ec
ime
n f
rom
th
e A
lbo
ran
Isl
and
. D
: C
. a
cua
ta.
E:
C.
erec
t& F
: C
. cr
am
be
LO
33

382 M. M a l d o n a d o & M. J. Uriz
Fig. 2. Points of ectosomal monactine of Crambe. A: C. tuberosa. B: C. tailliezi, specimen from the Alboran Island. C: C. tailliezi, specimen from the Cabrera Island. D: C. erecta. E, F: C. crambe,
Mediterranean specimens from the Northeastern coast of Spain
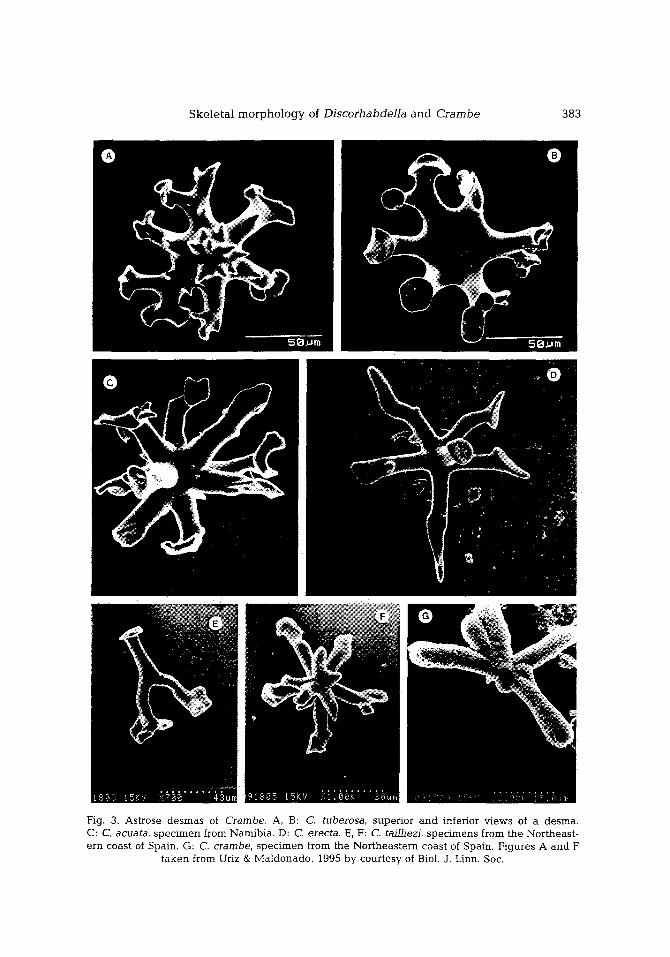
Ske le t a l m o r p h o l o g y of Discorhabdel la a n d Cr a m b e 383
Fig. 3. Astrose desmas of Crambe. A, B: C. tuberosa, superior and inferior views of a desma. C: C. acuata, specimen from Namibia. D: C. erecta. E, F: C. tailliezi, specimens from the Northeast- ern coast of Spain. G: C. crambe, specimen from the Northeastern coast of Spain. Figures A and F
taken from Uriz & Maldonado, 1995 by courtesy of Biol. J. Linn. Soc.
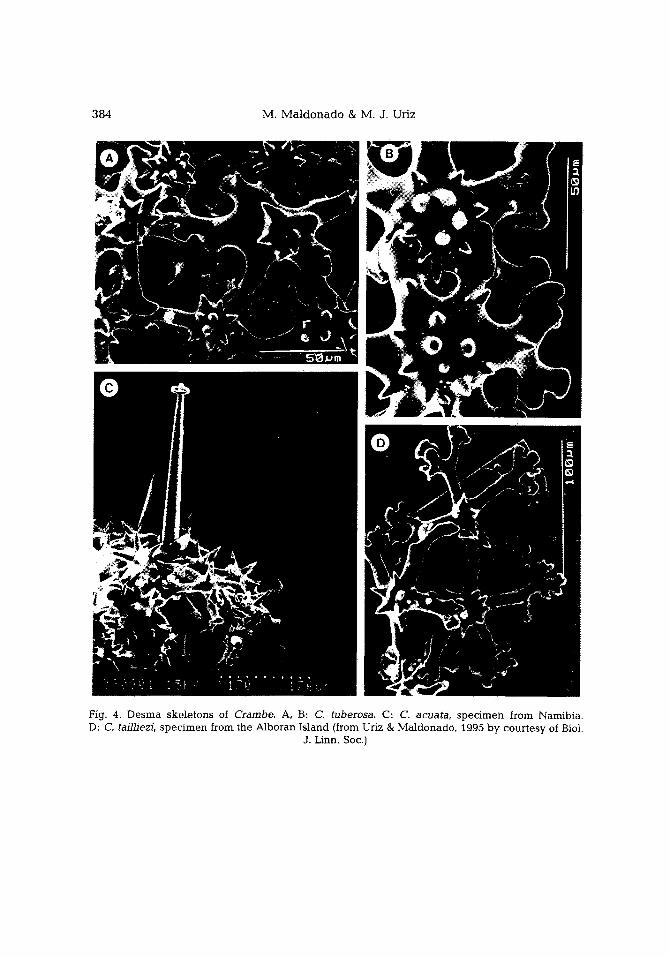
384 M. M a l d o n a d o & M. J. Uriz
"
J
t ~ . j '% ,, f " , .
Fig. 4. Desma skeletons of Crambe. A, B: C. tuberosa. C: C. acuata, specimen from Namibia. D: C. tailhezi, specimen from the Alboran island (from Uriz & Maldonado, 1995 by courtesy of Biol.
J. Linn. Soc.)
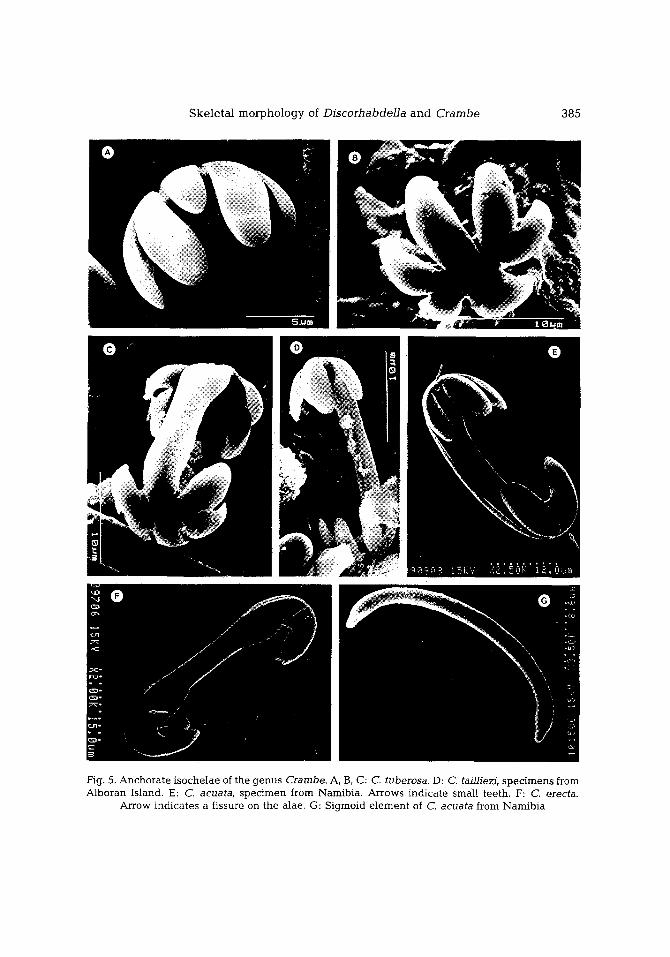
S k e l e t a l m o r p h o l o g y of D i s c o r h a b d e l l a a n d C r a m b e 385
- '4
o-. / ,,2 ~
~ " f / /
Fig. 5. A n c h o r a t e i soche l ae of the g e n u s Crambe. A, B, C: C. tuberosa. D: C. tailfiezi, s p e c i m e n s from Albo ran Island. E: C. acuata, s p e c i m e n from Narnibia. Ar rows ind ica te smal l tee th . F: C. erecta.
Arrow ind ica te s a f issure on the alae. G: S igmoid e l e m e n t of C. acuata f rom N a m i b i a

386 M. M a l d o n a d o & M, J. Uriz
Fig. 6. Diactinal microscleres. A, B: Crambe acuata from Namibia. C: C. tailliezi from Alboran Island. D, E: Discorhabdefla incrustans (oxydiscorhabd)

S k e l e t a l m o r p h o l o g y of Discorhabdel la a n d Crambe 387
Fig. 7. Main choanosomal monactines of Discorhabdelle. A: D. hindei. B, C: D. tuberosocapitata. D: D. incrustans (from Uriz & Maldonado, 1995 by courtesy of Biol. J. Linn. Soc.)

o 0 bl
Fig
. 8.
Poi
nts
of e
cto
som
al m
egas
cler
es o
f D
isco
rhab
dell
a.
A:
D.
tube
roso
capi
tata
. B
: D
. h
ind
ei.
C:
D.
incr
ust
an
s

Fig
. 9.
Pse
ud
oa
stro
se a
ca
nth
ost
yle
s o
f D
isco
rha
bd
ella
, A
: D
. tu
ber
oso
cap
ita
ta,
B:
D.
hin
dei
. C
: D
. in
cro
sta
ns
o ,,~
0 g.
LO
:D

390 M. M a l d o n a d o & M. J. Uriz
Fig. 10. Anchorate isochelae of Discorhabdella. A, B, C: D. tuberosocapitata. D, E: D. hindei. F, G: D. incrustans