social cognition and schizophrenia : observing others ... · social cognition and schizophrenia:...
TRANSCRIPT

Social cognition and schizophrenia: Observing others’ actions, rewards and errors
By
Elliot Clayton BROWN
A thesis submitted in partial fulfilment of the requirements for the degree of
Philosophiae Doctoris (PhD) in Neuroscience
from the International Graduate School of Neuroscience
Ruhr University Bochum
June 7thth 2013
This research was conducted at the Research Department of Cognitive Neuropsychiatry,
Psychosomatics and Preventative Medicine, at the LWL University Hospital, within the
Faculty of Medicine at Ruhr University under the supervision of Prof. Dr. Martin Brüne
Printed with the permission of the International Graduate School of Neuroscience, Ruhr University Bochum

Statement
I certify herewith that the dissertation included here was completed and written
independently by me and without outside assistance. References to the work and
theories of others have been cited and acknowledged completely and correctly. The
“Guidelines for Good Scientific Practice” according to § 9, Sec. 3 of the PhD regulations of
the International Graduate School of Neuroscience were adhered to. This work has
never been submitted in this, or a similar form, at this or any other domestic or foreign
institution of higher learning as a dissertation.
The abovementioned statement was made as a solemn declaration. I conscientiously
believe and state it to be true and declare that it is of the same legal significance and
value as if it were made under oath.
Elliot Clayton BROWN
Bochum, 07.06.2013

PhD Commission
Chair:
1st Internal Examiner: Prof. Dr. Martin Brüne
2nd Internal Examiner: Prof. Dr. Boris Suchan
External Examiner:
Non-Specialist:
Date of Final Examination:
PhD Grade Assigned:

Table of Contents
I. List of Figures
II. List of Abbreviations
III. Abstract
1. General background
1.1. Preamble 1
1.2. Social cognitive neuroscience and “mind-reading” 2
1.3. Action observation and mirror neurons 3
1.4. The EEG mu rhythm suppression and action observation 4
1.5. Processing the outcomes of others’ action 7
1.6. Social approach and avoidance motivations 9
1.7. Deficits in social cognition in schizophrenia 12
1.8. Broken mirrors and the observation of others in schizophrenia 14
1.9. General aims of the thesis work 17
2. The relevance of self and other in mirror motor activity
2.1. Introduction 19
2.2. Method
2.2.1. Participants 20
2.2.2. Task design 20
2.2.3. Procedure 22
2.2.4. EEG data acquisition 23
2.2.5. EEG data analysis of the mu rhythm 24
2.3. EEG mu rhythm results 25
2.4. Summary of results 26
3. The influence of reward and punishment on the motor mirror system
3.1. Introduction 28
3.2 Method
3.2.1. Participants 30

3.2.2. Task design 30
3.2.3. Procedure 31
3.2.4. EEG data acquisition 32
3.2.5. Behavioural data analysis 32
3.2.6. EEG mu rhythm suppression analysis 32
3.2.7. Time-course analysis of EEG mu power 33
3.3. Results
3.3.1. Behavioural results 34
3.3.2. EEG mu rhythm suppression results 35
3.3.3. Time-course analysis of EEG mu power results 36
3.4. Summary of results 37
4. Reward-related modulation of the mirror motor system in schizophrenia
4.1. Introduction 39
4.2 Method
4.2.1. Participants 40
4.2.2. Task design 41
4.2.3. Behavioural tests 41
4.2.4. Procedure 42
4.2.5. EEG data acquisition 42
4.2.6. Data analysis
4.2.6.1. Behavioural data analysis 43
4.2.6.2. EEG mu rhythm suppression analysis 43
4.3. Results
4.3.1. Behavioural results 44
4.3.2. EEG mu rhythm results 45
4.4. Summary of results 49
5. Processing the outcomes of others’ action –
sharing others’ rewards, losses and errors
5.1. Introduction 51
5.2 Method
5.2.1. Participants 52

5.2.2. Task design 53
5.2.3. Procedure 55
5.2.4. EEG data acquisition 56
52.5. EEG data analysis 56
5.3. Results 58
5.4. Summary of results 62
6. A possible neurobiological basis of social approach and avoidance
behaviour in schizophrenia
6.1. Introduction 63
6.2 Methods
6.2.1. Participants 66
6.2.2. Tasks and procedure
6.2.2.1. The Approach-Avoidance Task (AAT) 66
6.2.2.2. The AAT effect scores 68
6.2.2.3. Facial emotion recognition and discrimination test 68
6.2.2.4. State and trait anxiety scale 68
6.2.2.5. Plasma oxytocin assessment 69
6.2.3. Data analysis 69
6.3. Results 70
6.4. Summary of results 73
7. General Discussion
7.1. Overall summary of findings 75
7.2. Detailed discussion
7.2.1. Self and other in the mirror motor system 77
7.2.2. Reward and punishment in the motor mirror system 79
7.2.3. Reward-related modulation of the mirror motor system in
schizophrenia 82
7.2.4. Sharing others’ errors, rewards and losses 85
7.2.5. Social approach and avoidance behaviour
and oxytocin in schizophrenia 89
7.3. General conclusions and implications of findings 91

7.4. Limitations 96
7.5. Synthesis 98
7.6. Open questions and future suggestions 107
8. References 109
9. Appendices
9.1. Curriculum Vitae 127
9.2. List of publications 130
9.3. Acknowledgements 133

I. List of Figures
1.1. An illustration of cortical areas found to be homologues of the
mirror neuron system in humans 4
1.2. Plot showing the mu rhythm suppression 6
1.3. A simplified overview of a distinction between 3 categories of
motivation and the associated behaviours 10
2.1. A pictorial description of the experimental design showing
snapshots of the action observation (video) and
action execution parts 23
2.2. Bar chart showing the EEG mu rhythm suppression pooled over
electrodes covering motor cortex, with self and other conditions
shown for action execution and action observation 26
3.1. Subjective ratings of pleasantness, arousal and ease of paying
attention for rewarding, punishing and neutral observed actions 34
3.2. EEG mu rhythm suppression during the observation of rewarding,
punishing and neutral actions pooled over electrodes 36
3.3. Plot showing the change in EEG mu power, averaged from electrodes
over sensorimotor areas over the course of the video,
showing the different reward conditions 37
4.1. Subjective ratings of pleasantness, arousal and ease of paying
attention for rewarding, punishing and neutral observed actions,
showing ratings for the schizophrenia patient group
and healthy control group 45

4.2. Mean mu suppression for schizophrenia patient group and
matched healthy control group during action observation,
pooled across conditions and central electrodes 46
4.3. Mu suppression pooled over electrodes for schizophrenia patients
group and healthy control group during the observation of
rewarding, punishing and neutral actions 47
4.4. Scatter plot for data from only the healthy control group, to
demonstrate the relationship between mean mu rhythm suppression
and empathy scores in the perspective-taking domain 48
4.5. Scatter plot for data from only the schizophrenia patient group,
to demonstrate the relationship between mean mu rhythm
suppression and negative psychotic symptoms, as indicated
by the PANSS 49
5.1. Illustration of experimental design and stimuli for active part,
showing the potential outcomes of either a gain or a loss 54
5.2. Illustration of experimental design for observational / passive
learning part showing the potential outcomes of
either a gain or a loss 55
5.3. Grand-averaged event-related potentials (ERPs) for onset of
positive (wins) and negative (losses) feedback received 59
5.4. Grand-averaged event-related potentials (ERPs) for onset of
response in the passive condition in the 100% condition 60
5.5. P300 amplitudes for active and passive parts showing results of
pairwise comparisons of high and low expectancy condition 61

6.1. a) A bar plot of reaction times from the social approach
avoidance task (AAT).
b) A pictorial illustration of the congruent and incongruent
push-pull responses to angry and happy faces in the
joystick-based AAT. 65
6.2. An illustration of the experimental design for the AAT showing
the order of events in two individual trials 67
6.3. Table showing means and SDs for symptom severity, behavioural
measures and medication between low and high oxytocin groups 71
6.4. AAT effect scores reflecting push minus pull mean RTs for each
condition, comparing RTs for the low and high basal oxytocin groups 72
6.5. Scatter plot showing line of best to demonstrate the relationship
between basal plasma oxytocin levels and the AAT effect score for
angry faces with a straight-gaze 73
7.1. Schematic representation showing the synthesis of topics
explored in this thesis 106

II. List of Abbreviations
AAT Approach-Avoidance Task
ACC Anterior cingulate cortex
ANOVA Analysis of variance
ASD Autism Spectrum Disorder
BCI Brain-Computer-Interface
CLZ Chlorpromazine equivalent
DLPFC Dorsolateral prefrontal cortex
DSM-IV Diagnostic and Statistical Manual of Mental Disorders IV
EDTA Ethylenediaminetetraacetic acid
EEG Electroencephalography
EIA Enzyme immunoassay
EOG Electrooculography
ERN Error-related negativity
ERP Event-related potential
FEDT Face Emotion Discrimination Task
FEIT Face Emotion Identification Task
FFT Fast Fourier Transform
fMRI Functional magnetic resonance imaging
FRN Feedback-related negativity
GLZ General linear model
ICD-10 International Classification of Diseases v.10

IFG Inferior frontal gyrus
IPL Inferior parietal lobule
IPS Inferior parietal sulcus
IRI Interpersonal Reactivity Index
MPFC Medial prefrontal cortex
oERN Observational error-related negativity
OFC Orbitofrontal cortex
oFRN Observational feedback-related negativity
MFC Medial frontal cortex
MNS Mirror Neuron System
PANSS Positive and Negative Syndrome Scale
PCC Precuneus / Posterior cingulate cortex
RT Reaction time
SD Standard deviation
SPF Der Saarbrücker Persönlichkeitsfragebogen
STAI State–Trait Anxiety Inventory
STS Superior temporal sulcus
TMS Transcranial magnetic stimulation
TOM Theory of mind
TPJ Temporoparietal junction
vACC Ventral anterior cingulate cortex

“We may then lay it down for certain that every [mental] representation of a movement awakens in some degree the
actual movement which is its object; and awakens it in a maximum degree whenever it is not kept from so doing by an antagonistic representation present simultaneously to
the mind”
William James - “Principles of Psychology” (1890)

III. Abstract
A fundamental prerequisite for social interaction is the ability to understand the
meaning and intentions of others’ actions. More evidence is emerging to suggest the
presence of shared neural representations of experience in both the first and third-
person stance. Support has come from the discovery of common neural activity during
both action execution and action observation, namely the mirror neuron system (MNS).
The work in this thesis sought to explore shared neural representations of one’s own
and of others’ actions, rewards and errors along a number of different lines, and how
this could be modulated by different contexts. This was also investigated in terms of
schizophrenia, as patients are known to have deficits in social cognition.
In the first three studies, the EEG mu rhythm suppression was used as an index of mirror
neuron-related motor cortex activity. The first two studies demonstrated,
independently, that greater mu suppression was produced when observing actions that
were relevant to the self, as opposed to the other, and to actions that were rewarding as
opposed to those that were neutral. In the third study, it was shown that patients with
schizophrenia also exhibited a reward-related modulation of the mu suppression during
action observation, and furthermore, that the mu suppression was related to psychotic
negative symptoms and empathy. The fourth study also used EEG to investigate reward
and error-related neural activity, and found that the event-related potentials (ERPs)
associated with one’s own rewards and errors (i.e. the feedback-related negativity and
error-related negativity) also resembled the ERPs associated with others’ rewards and
errors. In addition, an ERP associated with others’ feedback (the P300) was substantially
influenced by expectation. Lastly, a study exploring the relationship between the
prosocial neuropeptide oxytocin and social approach and avoidance behaviour in a
schizophrenia population revealed that individual differences in endogenous oxytocin
levels related to the avoidance of negative emotional stimuli.

In conclusion, it is evident that the internal mental and external environmental context
can shape the interpretation of others’ behaviour, and in particular, the perception of
others’ actions, rewards and errors. These studies suggest that some contextual
modulations of the MNS found in previous studies may have been driven by underlying
influences of self-relevance and reward that were intrinsic to the nature of the perceived
stimuli. This also implies that the emergence and development of the MNS, and the
associated cognitive functions related to shared neural representations of one’s own and
of other’s experience, may also be facilitated by the reward-associations made to other
people’s behaviours, which can also be coloured by the context in which other’s actions
are observed. The influence of reward and self-relevance on the MNS may also have
implications on the development and persistence of social cognitive deficits seen in
schizophrenia. To summarize, a synthesis of these findings is put forward that also aims
to integrate other findings and theoretical frameworks related to learning, perception
and action in the frame of social interaction.

1. GENERAL BACKGROUND
1
Chapter 1
General background
1.1. Preamble
Humans are social animals. The evolution of the modern human brain has likely
emerged from the increasing complexity of our social environment over the generations
of our ancestors. Like many other species in the animal kingdom, we spend the majority
of our lives engaged in a social environment, interacting with others, and such that our
actions and behaviours are shaped by socially-driven motivations. We also depend upon
social interaction to maintain our mental wellbeing, as social isolation can lead to
severely detrimental consequences on one’s mental health. Social interaction involves a
multitude of socially-relevant cognitive processes including, to name a few, social
perception, understanding others’ actions, observational learning and social decision-
making. More evidence is emerging that shows “shared neural representations” of
experience when observing others’ behaviours, whereby similar brain activation
patterns are seen for the processing of both one’s own and of others’ experiences, which
may provide the underlying neural basis for understanding, predicting, and empathising
with others’ behaviour. This proposal has largely been fuelled by the discovery of the
mirror neuron system, which is a network of brain regions that are activated both when
performing and observing others’ actions. An important aspect of social interaction is
the context in which social information is processed, as the social context can colour the
interpretation of other people’s behaviour. There is substantial work to indicate that
there is a top-down contextual influence of social information on the modulation of

1. GENERAL BACKGROUND
2
neural activity associated with action observation and observational learning. These
contextual influences on the processing of others’ behaviours have direct and broad
implications on real-world social interaction, but have only begun to be explored in the
field of neuroscience. Dysfunctional social behaviours are often linked with psychiatric
illnesses such as schizophrenia, and impaired social functioning also has strong
associations with deficits in social cognitive skills. Therefore, schizophrenia can act as an
interesting model to explore the neural correlates of social behaviour. By revealing the
neural processes underlying social cognition, we can begin to understand where the
dysfunction is going wrong in schizophrenia, and consequently translate our
understanding of these dysfunctions in the brain to informing therapeutic strategies to
improve social functioning in schizophrenia, and ultimately improve the quality of life of
patients suffering from the illness.
1.2. Social cognitive neuroscience and “mind-reading”
The advancement of modern neuroimaging methods in the last twenty years has opened
up many doors for inquiry in every domain of psychology. The birth of “social
neuroscience” (Cacioppo and Berntson, 1992) uses these newly developed
neuroimaging techniques to reveal the underlying neural mechanisms behind cognitive
processes related to social functioning and behaviour. A seminal paper from Ochsner
and Lieberman (2001) explicitly outlines the emergence of the interdisciplinary field of
“social cognitive neuroscience” that covers a broad scope of investigation. They highlight
the integration of 3 levels of analysis: ‘the social level, which is concerned with the
motivational and social factors that influence behaviour and experience; the cognitive
level, which is concerned with the information-processing mechanisms that give rise to
social-level phenomena; and the neural level, which is concerned with the brain
mechanisms that instantiate cognitive-level processes.’ More generally, the term “social
cognition” often refers to the sum of the cognitive processes required for social
perception and social interaction, and Adolphs (2001) defines social cognition as ‘the
ability to construct representations of the relations between oneself and others, and to
use those representations flexibly to guide social behaviours’.

1. GENERAL BACKGROUND
3
One central concept in the study of social cognition is theory of mind (TOM), or
mentalizing, which is the ability to attribute mental states to others (Premack &
Woodruff, 1978). For someone to be able to understand other people, one must be able
to take the other’s perspective, or put oneself in the “other person’s shoes”. To do this,
one must first understand that other people do not have the same thoughts, opinions,
beliefs or knowledge as oneself. A broad network of brain areas have been consistently
associated with mentalizing / TOM, which includes the medial prefrontal cortex (MPFC),
dorsolateral prefrontal cortex (DLPFC), precuneus / posterior cingulate cortex (PCC),
ventral anterior cingulate cortex (vACC), orbitofrontal cortex (OFC), temporoparietal
junction (TPJ), superior temporal sulcus (STS) and amygdala (Abu-Akel & Shamay-
Tsoory, 2011; Amodio & Frith, 2006; Mitchell, Macrae & Banaji, 2006; Saxe, et al., 2004;
Oschner et al., 2005). This mentalizing / TOM network has been found to be activated
during high-level inference-based mental state reasoning about both self and others. A
further distinction has also been made between cognitive and affective TOM (Shamay-
Tsoory et al., 2007), recruiting different brain areas depending on the nature of the
mental state being shared.
1.3. Action observation and mirror neurons
Another neural system thought to be crucial for social understanding is activated for
both one’s own and others’ motor actions. Since the discovery of an apparently
functionally-specific mirror neuron system (MNS) in the monkey brain that is activated
during both action execution and action observation (Gallese et al., 1996), hypotheses
about the mirror neuron system have made wide speculations about its role in social
cognition. The coupling of action and perception in this mirror neuron system
demonstrates a resonance of motor actions in the observer’s brain, in which shared
neural representations of one’s own and others’ actions are evident. The MNS
hypothesis is compatible with simulation theories of theory of mind (e.g. Davies & Stone,
1995), which in general argues that individuals generate simulations of actions, and
consequently also their own thoughts, intentions, beliefs and emotions, to predict the
mental state of others and therefore ascertain knowledge of other minds. Some authors
propose that this then consequently provides the fundamental elements for the ability to
understand, and empathise with, the social behaviour of others.

1. GENERAL BACKGROUND
4
Numerous fMRI studies have consistently found human homologues of the monkey
mirror neurons (e.g. Gallese et al., 2004; Keysers & Gazzola, 2007; Keysers & Perrett,
2004; Rizzolatti & Craighero 2004), which include premotor cortex, inferior frontal
gyrus (IFG), inferior parietal lobule (IPL), inferior parietal sulcus (IPS), superior parietal
lobule and superior temporal sulcus (STS), as shown in figure 1.1. A more recent review
has highlighted the potential relevance for such fronto-parietal networks in action
understanding (Rizzolatti & Sinigaglia 2010). Interestingly a recent study has also
shown concomitant activation of both the ‘mentalizing’ and the human mirror neuron
network when observing stimuli involving social interaction (Centelles et al., 2011).
Some authors suggest that the mirror neuron system might support the mentalizing /
TOM network (Agnew et al., 2007; Blakemore et al., 2004; Decety & Chaminade,, 2003;
Keysers & Gazzola 2007; Uddin et al., 2007), and therefore may contribute to the neural
basis for higher level social cognitions. However, the functional relevance of this
mirroring network is still under debate.
Figure 1.1: An illustration of cortical areas found to be homologues of the mirror neuron system in
humans. The purple areas are associated with reaching movements, the yellow areas with transitive distal
movements, the orange area with tool use, the green with intransitive movements and the blue with
responding to observation of upper-limb movements (Adapted from Cattaneo & Rizzolatti, 2009)

1. GENERAL BACKGROUND
5
1.4. The EEG mu rhythm suppression and action observation
Evidence for a neurophysiological index of mirror neuron activity in humans has come
from the mu rhythm suppression in EEG recordings in which a reduction (i.e.
suppression) in the power of the mu rhythm occurs during both the execution of an
action, and, though to a lesser degree, during the observation of an action. Rolandic mu
rhythms are EEG oscillations occurring over the motor cortices that are composed of
two main frequency components; one spectral peak in the alpha range (8-13Hz) and the
other in the beta frequency range (around 20Hz) (Hari, 2006). Figure 1.2 shows the
change in the power of the 20Hz frequency band over the motor cortex during the
course of the action. In the plot, action termination is represented at time 0. In this time
/ power plot, one can see a comparable suppression pattern of the power of the mu
rhythm during both action execution (in pink) and observation (in blue), followed by a
sharp rebound. This sharp post-stimulus rebound in the beta frequency range following
termination of the executed (Hari, 2006) and observed action (Muthukumaraswamy et
al., 2004a) has been suggested to reflect an inhibition of motor activity that prevents this
activity from exceeding a threshold for muscle activation, and therefore acting as an
action control mechanism. During rest, neurons in the primary and premotor cortex fire
synchronously, and event-related desynchronization occurs during action execution as
well as during observation of actions, and this suppression of the EEG mu rhythm is
thought to reflect the event-related desynchronization of motor cortex (Pfurtscheller &
Silva, 1999).

1. GENERAL BACKGROUND
6
Figure 1.2: Plot showing the mu rhythm suppression. The plot displays the change in power in the lower
beta band (20Hz) reflecting a part of the EEG mu rhythm that is suppressed during action execution (pink)
and action observation (blue) (modified from Caetano et al., 2007)
It has long been thought that the mu rhythm suppression is specific to motor-related
neural activity (Gastaut & Bert, 1954). Suppression of the mu rhythm has also been
shown for actions that are imagined or heard (e.g. Pineda, 2005). The mu rhythm is a
reliable neurophysiological signature and can be extracted in real-time as it can be seen
with only one trial, without the presence of an overt motor action, and can also be
enhanced through training (Kuhlman, 1978). Due to the presence of the mu rhythm
suppression for imagined actions, and its stability, it is widely used in EEG-based brain-
computer-interface (BCI) work as a control signal for feedback and for external devices
(e.g. Wolpaw et al., 2002). In BCI applications, participants go through a process of
training in which they practice to enhance the strength of their own mu suppression
through imagination of actions. One example of the use of the mu rhythm during motor
imagery as a biofeedback signal is for neuromuscular electrical stimulation to help post-
stroke patients recover from physical disability (Daly et al., 2009). Patients are

1. GENERAL BACKGROUND
7
instructed to imagine moving their dysfunctional limb, and as they do this, the mu
rhythm EEG signal is translated into electrical stimulation of the muscles in the disabled
limb. Take together, the mu rhythm suppression represents a robust index of
sensorimotor cortex activity, which allows exploration of questions related to action
observation, “mirror” motor activity and the coupling of action and perception. The
suggestion that the mu rhythm suppression is a marker of mirror neuron activity is
supported by anatomical and physiological evidence for strong cortico-cortico
connections between ventral premotor cortex and primary sensorimotor cortex (Taylor
et al., 2009), which is where the mu rhythm has been recorded and is thought to be
generated (Hari, 2006). In addition, a study comparing EEG and fMRI results has
confirmed the close relationship between mirror neuron-related activity and the
suppression of the mu rhythm in humans (Perry & Bentin, 2009). Mu rhythm
suppression also appears to be functionally specific to actions that are goal-directed
(Muthukumaraswamy et al., 2004b). A variety of different attentional and contextual
changes in the executed / observed actions have been found to influence the magnitude
of the mu rhythm suppression. Perry and colleagues (2011) demonstrated that the mu
suppression can be enhanced during social interaction. Several studies finding
modulation of the mu rhythm in different social contexts have provided support for the
relevance of the mu suppression in its association with the mirror neuron work, and its
implications in social cognition. These contextual changes in the observed action, and
the relevance to schizophrenia, will form the basis of the first studies documented in this
thesis, and will be discussed in more detail in the later chapters.
1.5. Processing the outcomes of others’ action
It is clear that there is much evidence to demonstrate that the neural areas activated
when performing actions are also coding information about the observed actions of
others. However, it is not only the motor action itself that can be resonated in the
observer’s brain, but also the consequences of the outcome and the implications of the
observed action in terms of error, feedback and reward. Some studies have shown that
the brain activity induced when seeing other people make mistakes is similar to the
activity seen during the processing of one’s own errors, i.e. when an erroneous response
is made, and one is aware of that error. Comparatively, the neural activity produced

1. GENERAL BACKGROUND
8
when one receives feedback following a response choice is also similar to the pattern of
activity seen when observing others receive feedback, as both are modulated by the
valence of the feedback, i.e. if it is positive or negative. Thus these common patterns of
brain activity provide further support for shared neural representations for one’s own
and others’ experiences.
Much evidence for shared representations of self and other’s errors, feedback, rewards
and losses comes from particular patterns of activity seen with EEG. The error-related
negativity or ERN is an event-related potential (ERP) that is elicited shortly (between 50
and 150ms) after a known error response is made (Falkenstein et al., 1990) in a forced
choice response task. A similar ERN has also been found when observing other people
make errors (van Schie et al., 2004; Koban et al., 2010), of which some have referred to
as an observational ERN, or oERN. The FRN is another ERP that is related to the ERN,
and may even be considered a type of ERN, which is elicited when one is given positive
or negative feedback after a choice is made during both performance and observation of
a reinforcement learning task, namely the (observational) feedback related negativity or
(o)FRN (Bellebaum et al., 2010). The FRN is thought to be an index of reward processing
because it is modulated by the valence of the feedback, i.e. positive or negative, gain /
reward or loss (Holroyd & Coles, 2002). Both the ERN and FRN are proposed to
originate from the ACC and are also thought to be indices of the prediction error signal
(Holroyd & Coles, 2002). The prediction error acts like a teaching signal in the brain,
crucial for learning, as it codes the discrepancy between the expected outcome and the
actual outcome, as ascertained by the prediction and the feedback, respectively (Schultz,
2000). Estimations about future outcomes of events, such as the prediction of the
outcome of one’s own and others’ choices, are then updated according to the prediction
error signal. Some studies have shown that the degree of self-relatedness (i.e. how
similar the performer is to the observer) and the interpersonal relationship (i.e. friend
vs. stranger) between the observer and the performer can modulate the magnitude of
the oERN and oFRN when observing others’ errors and feedback (Carp et al., 2009; Ma et
al., 2011). Others have demonstrated that these observational ERPs can also be
modulated by competition and cooperation between the observer and the performer
(Itagaki & Katayama, 2008; Koban et al., 2010; Rigoni et al., 2010). There is a lot of
evidence to support the proposal of shared neural activity in the processing of one’s own
and others’ actions, errors and rewards; however it is still not clear how these shared

1. GENERAL BACKGROUND
9
neural processes may translate to high-level, real-world social behaviour. The oERN and
oFRN are useful candidates for exploring the neural activity associated with social
interaction in different contexts, and could have extensive implications on the capacity
for observational social learning. However, it is still not clear what neural activity these
ERPs actually represent, in which context they can be induced, how necessary these
mechanisms might be to social cognitive processes and to what degree expectancy
influences the neural responses when observing others’ behaviour.
1.6. Social approach and avoidance motivations
So far, the focus of this introduction has been largely put on the “cold” cognitive
processes associated with social interaction. In general, the pathway from underlying
neurobiological and neurophysiological mechanisms has a causal influence on the
processes at the level of cognition, which is ultimately manifested in behaviour.
However, this is obviously not the whole story. Motivation is another important factor
that is central to the translation of neurobiological mechanisms into behaviour, and is
also critical to social interaction. Figure 1.3 shows a simple but illustrative
representation of the different types of basic motivation and how these translate into
behaviour. The figure displays the categorisation of motivation into appetitive
motivation, aversive motivation and quiescence, which parallels the well-known
distinction between fight, flight or freeze responses to threat and stress (Cannon, 1929;
Gray, 1988).

1. GENERAL BACKGROUND
10
Figure 1.3: A simplified overview of a distinction between 3 categories of motivation and the associated
behaviours (modified from McCall & Singer, 2012).
Motivational drives can dictate social behaviours, and in particular, can influence social
approach and avoidance behaviours. Appetitive and aversive motivations underlie the
social motives to have a need for affiliation and a fear of rejection, respectively. These
motives stem from social insecurity, rejection and social isolation (Shipley & Veroff,
1952), and can be deterministic of social functioning and consequently impact on
psychological well-being (Gable, 2006). The willingness to actively engage in social
situations, and the functional success of social interactions, is closely related to
underlying appetitive and aversive motivational factors. Social approach and avoidance
behaviour is directly related with the emotional valence of social stimuli, as healthy
people have a tendency to move away (avoid) from angry facial expressions and
conversely, will move towards (approach) happy emotional stimuli (Marsh et al., 2005;
Roelofs et al., 2009a; Volman et al., 2011). In the other direction, approach and

1. GENERAL BACKGROUND
11
avoidance (AA) movements can also create biases in the evaluation of affective stimuli
(e.g., Cacioppo et al., 1993) and attitudes (Centerbar & Clore, 2006).
Certain pathological conditions, particularly those with impairments in social behaviour,
have been shown to exhibit abnormalities in approach and avoidance responses to
positive and negative social emotional stimuli. For example, socially anxious individuals
have a greater tendency to avoid faces expressing angry emotions (Heuer et al., 2007). A
comparable tendency to avoid angry faces on the AAT has also been found with people
who have Psychogenic Non Epileptic Seizures (Bakvis et al., 2011), who are thought to
have a learned pattern of avoidant behaviour to cope with stressful situations (Ramani
et al., 1980). This learned avoidance is likely to have a detrimental impact on social
functioning, as this may also be accompanied by avoidance of social interaction and
general aversive behaviour in social settings. In contrast to these populations,
psychopaths demonstrate a lack of avoidance to angry faces (Lousie von Borries et al.,
2012). Gaze direction has also been found to be a confounder in AA related behaviour,
with direct gaze inducing a greater tendency for avoidance in socially anxious people
(Roelofs et al., 2010), as negative emotional faces with a direct gaze are generally found
to be more threatening (Adams & Kleck, 2005). These studies demonstrate how the
degree of avoidance for social threat cues indexed by AA behaviour is likely to reflect
tendencies in real-world social behaviour.
Pathological differences in social approach and avoidance of social emotional stimuli are
driven by both cognitive and neurobiological factors (Roelofs, 2005), however it is still
not clear as to how these interact and manifest in terms of coping strategies (Master et
al., 2009). Some recent work has focused on the underlying neurobiological factors that
may be influencing dysfunctional social behaviour and approach and avoidance, with
one popular chemical of interest being oxytocin. Oxytocin is a neuropeptide that has
been found to have a central role in the expression of a multitude of social and affiliative
behaviours across many different species (McCall & Singer, 2012). Most relevantly, some
studies have found that oxytocin can modulate the tendency for social approach and
avoidance, with one study in particular showing that the administration of intranasal
oxytocin in healthy people can decrease aversion behaviour, specifically to angry faces
(Evans et al., 2010). Kemp & Guastella (2010) suggest that oxytocin can increase

1. GENERAL BACKGROUND
12
approach related behaviour and decrease withdrawal, though this was proposed in the
context of the response of an angry person rather than the perception of an angry face.
The study of motivational factors involved in social behaviour, and the underlying
neurobiological influences, provides another route for inquiry that can add further
insights into the understanding of social cognition. This broad view becomes even more
so relevant when applying such research questions to the study of social functioning and
social cognition in schizophrenia. The more “cold” cognitive elements of social cognition,
as associated with the previously mentioned work on mentalizing, action observation
and the observation of others’ errors has been more closely linked to the positive
symptoms of schizophrenia, whereas issues related to motivation are more closely
linked to the negative symptoms.
1.7. Deficits in social cognition in schizophrenia
Schizophrenia is primarily characterized by positive and negative psychotic symptoms.
Positive symptoms refer to extraordinary thoughts and behaviours that are above and
beyond normal experience and include delusions, hallucinations, hyperactivity,
suspiciousness and conceptual disorganization. Negative symptoms include symptoms
that are reductive of normal experience such as a blunted emotional response,
emotional withdrawal, passive social withdrawal, a lack of spontaneity, poor rapport
and a lack of motivation. Other core symptoms associated with schizophrenia include
anxiety, depression, feelings of guilt, motor retardation, disorientation, poor attention,
impulsivity, and disorientation. Deficits in social cognition are well known in
schizophrenia and evidently have a direct impact on functional recovery and rate of
relapse, as well as quality of life during the course of the illness (Green & Leitman,
2008). Impaired social functioning often precedes the onset of the first psychotic
episode and is thought to be a major predictor for the development of psychotic
symptoms (Green et al., 2008). Recent research has revealed that neurocognitive
deficits, such as executive functioning and working memory, do not account for a
majority of the variation in social functioning in schizophrenia (Penn et al., 1997;
McGurk & Meltzer, 2000; Brekke et al., 2005; Nakagami et al., 2008), whereas social
cognition seems to be more strongly associated with social functioning (Lysaker et al.,

1. GENERAL BACKGROUND
13
2004; Brüne et al., 2007). However, there is still little known about the neurological
basis of deficits in social cognition in schizophrenia.
A consensus of five key domains of social cognition that most commonly and
persistently present deficits in schizophrenia have been recently defined (Green et al.,
2008). These domains were listed as: emotion processing, TOM, attributional bias, social
perception and social knowledge. Emotion processing deficits in schizophrenia
encompass multiple impairments including emotion recognition from facial expression
and prosody, emotion discrimination, emotion interpretation and emotional awareness
(for review see Tremeau, 2006). One study has shown a direct relationship between
emotion recognition deficits and the severity of negative and affective symptoms, as well
as poor vocational and global functioning (Hofer et al., 2009). Deficits in mentalizing /
TOM seen in patients with schizophrenia have been directly related to core psychotic
symptoms, and have been used to explain deficits in social functioning (Brüne, 2005). A
recent meta-analysis has shown TOM deficits in both first-episode and remitted patients,
suggesting that these deficits are inherent of the illness (Bora et al., 2009). In the past,
TOM skills were thought to be highly correlated with neurocognitive variables such as
IQ and verbal memory, though recently some have concluded that deficits in theory of
mind may be independent of neurocognitive deficits (Brüne et al., 2009). It is still not
clear as to how or to what extent deficits in TOM contribute to dysfunctional social
behaviour (Brüne, 2005), although an impaired ability for mental state attribution has
been shown to be the single best predictor of social competency when compared to
measures of neurocognitive functioning and psychopathology (Brüne et al., 2007).
Attributional style refers to the tendency to attribute negative or positive situations to
oneself or to others. Pioneering work on abnormal attribution biases in schizophrenia
came from Bentall et al (1994), particularly in reference to the creation and
maintenance of persecutory delusions. They argued that biases in self-representation
are caused by biases in causal attributions, i.e. an over-self-serving attributional bias,
which in turn leads to negative events being attributed to external agents, and
consequently manifesting in paranoid thoughts and persecutory delusions. The two
other domains of social cognition defined by the NIMH workshop (Green et al., 2008),
namely social perception and social knowledge, can be considered more as “umbrella”
terms that largely encompass emotion processing, TOM and attributional style.

1. GENERAL BACKGROUND
14
Following illness onset, people suffering from schizophrenia often find it hard to
maintain fulfilling social lives. When asked, most psychotic patients would prioritise the
remediation of social functioning over the recovery of psychotic positive or negative
symptoms (Foldemo et al., 2004). However, remission of symptoms with the use of
antipsychotic medication seems to have little impact on the remission of social skills
(Penn et al., 2009). A variety of new social cognition training programs have emerged in
recent years, and such treatment programs have begun to demonstrate how fruitful the
remediation of social cognitive skills can be in schizophrenia (Brown et al., 2012; Tas et
al., 2011). Psychosocial treatments and social cognition training programs are showing
greater medication adherence, better self-management and lower rates of relapse, as
well as providing strategies for encouraging occupational success and stable social
relationships (Kurtz & Richardson, 2011). Remediation of social cognition has also been
show to indirectly improve positive and negative symptoms (Brown et al., 2012). The
creation and design of such programs has been built on the basis of prior research
findings, however this area of research is still in its infancy. These new treatment
programs are the first of their kind, and it is still under debate as to whether the current
view taken of the domains of deficits in social cognition translates directly to treatment
strategies, and most importantly, whether the current framework allows for the transfer
of learned skills to real-world social functioning. In addition, the research on social
cognitive impairments in schizophrenia has yet to find conclusive evidence of the
aetiology of these deficits. One crucial question related to this issue is how low-level
perceptual, sensory and motor abnormalities seen in schizophrenia may contribute to
high-level deficits in more complex social cognitions and behaviour.
1.8. Broken mirrors and the observation of others in schizophrenia
Frith suggested that deficits in TOM in schizophrenia lead to disorders of agency and
action, such as that of “willed action”, “self-monitoring” and the “monitoring of others”
(Frith, 1992). A few studies have shown that patients with schizophrenia have reduced
motor responses in general and in terms of imitation. Motor cortex response seems to be
diminished in schizophrenia during observation and execution of action (Schürmann et
al., 2007; Enticott et al., 2008). One study demonstrated that schizophrenia patients
were less able than healthy controls to imitate facial expressions from still photographs

1. GENERAL BACKGROUND
15
(Schwartz et al., 2006). Moreover, patients with schizophrenia show reduced
contagiousness to observed yawning (Haker and Rössler, 2009). Rapid mimicry-like
reactions to the observation of angry and happy faces are also reduced in schizophrenia
patients compared to controls (Varcin et al., 2010). Park et al (2008) found that patients
with schizophrenia are poor at imitating hand gestures, mouth movements and
emotional expressions. In their study, imitation errors also correlated with reduced
social competence and increased negative symptoms. Conversely, patients with
schizophrenia show abnormalities in response inhibition (Kiehl et al., 2000), which was
suggested to be related to impulsive behaviour and a lack of self-control. Consequently,
patients may have a reduced ability to inhibit motor actions as well as imitative motor
actions. Some psychotic patients exhibit echopraxia, the pathological imitation of
behaviours seen with catatonic syndromes, which has been suggested to occur due to a
pathologically enhanced “mirroring effect” (Pridmore et al., 2008). These contrasting
hypotheses encourage further investigation.
Behavioural and neuroimaging studies have demonstrated deficits in the perception of
biological motion in schizophrenia (Kim et al., 2005; Takahashi et al., 2010), which may
be an explanation for previous findings of a misunderstanding of nonverbal
communication (Toomey et al., 2002). Some authors have made the proposal that a
dysfunction in the mirror neuron system can been used to explain deficits in social
cognition in psychiatric and developmental disorders, namely schizophrenia (Arbib &
Mundhenk, 2005), autism spectrum disorders (Williams et al., 2001) and Williams
Syndrome (Tager-Flusberg 2000). The “broken-mirror” theory of autism argues that
core social and cognitive deficits, including deficits in TOM, are caused by a dysfunction
of the mirror neuron system. However, the results are far from conclusive with some
studies finding abnormal mu rhythm suppression in people with autism during action
imitation tasks (Bernier et al., 2007; Oberman et al., 2005, 2008), while others have not
found differences (Raymaekers et al., 2009; Fan et al., 2010).
Arbib & Mundhenk (2005) extend the “broken-mirror” hypothesis to account for deficits
in self-monitoring in schizophrenia as well as misattributions of agency, and auditory
hallucinations. They make the suggestion that a dysfunction in the mirror neuron
network may have some underlying causal role in the expression of deficits in social
cognitive skills, and behaviours related to the monitoring of one’s own and others’

1. GENERAL BACKGROUND
16
actions and vocalisations seen in schizophrenia. Some neuroimaging studies have
provided further support to this suggestion (Andreasen et al., 2008), as abnormal
activity has been discovered in parietal areas associated with the human mirror neuron
system, in an unmedicated schizophrenia population (Kato et al., 2011). The mu rhythm
suppression, as an index of motor mirror neuron related activity, has been used to
investigate possible abnormalities in schizophrenia. One study from Singh and
colleagues (2011) compared healthy controls subjects and first-episode schizophrenia
patients while they observed videos of a moving hand, a social interactive game setting
and a point-light biological full-body motion display. They had hypothesised that the
context of the social interaction in the observation of others’ actions may be the most
revealing in terms of a potential abnormality in mirror neuron related activity. However,
a significant difference in the power of the mu suppression between controls and
patients was only found in the point-light biological motion condition, with substantially
lower mu suppression in the patient group. The interpretation of this finding was that
this difference may have reflected a deficit in information processing in the patient
group rather than a difference in the observation of goal-directed actions, as the point-
light display provides limited information to determine the biological motion. Though
interestingly, they found that the mu suppression in the patient group was positively
correlated with negative symptom burden and social adjustment.
One other study from McCormick and colleagues (2012) investigated the mu rhythm
suppression in patients with schizophrenia, comparing patients who were actively
psychotic to those who had only residual symptoms, along with a healthy control group.
They used three different action conditions, one in which participants watched a video
of an action, one with the observation of a live action and thirdly, an action execution
condition. In contrast to the study from Singh and colleagues, the McCormick study
found greater mu rhythm suppression in patients that were actively psychotic, as
compared to those with residual symptoms and the healthy controls across all action
conditions. Psychotic symptoms were also positively correlated with the mu
suppression, or more specifically, higher left-sided mu rhythm suppression was greatest
in subjects with more severe psychotic symptoms.
The findings from McCormick and colleagues (2012) are in line with the suggestion
made by Pridmore and colleagues (2008) that psychotic symptoms, such as catatonic

1. GENERAL BACKGROUND
17
ones, in schizophrenia may result from an enhanced or excessive activity in the mirror
system. However, the results from Singh and colleagues (2011) is more complementary
of the proposals associated with “broken mirror” hypothesis, which purports a
diminished mirror neuron system, and therefore would predict lower mu rhythm
suppression in patients with schizophrenia, that would also be related to diminished
social skills.
Despite the numerous suggestions of mirror neuron dysfunction as an explanation for
the symptoms of schizophrenia and the associated deficits in social cognition, the work
done so far to investigate mirror neuron function in patients with schizophrenia is
unclear and contradictory. There also still appears to be insufficient evidence for a direct
link between abnormalities in the mirror neuron system and deficits in social
functioning. It is also not clear what the function of the mirror neuron system
involvement is in social cognition (Heyes, 2010a). Due to the specificity of deficits in
social cognition and self-referential processing seen in psychotic patients, schizophrenia
is an intriguing model to explore abnormalities in neural activity related to the mirror
neuron system, and to help try to identify specific aspects of social cognition that may
directly relate to possible neurophysiological abnormalities.
1.9. General aims of the thesis work
The overarching aim for the work in this thesis was to explore low-level sensory,
perceptual and motor processes associated with the basic elements of social cognition
and social observational learning in healthy people, and in patients with schizophrenia.
The major part of this work sought to investigate how the underlying neurophysiological
mechanisms recruited during the observation of others’ actions, rewards and errors
may have a role in higher level complex social cognitions. More specifically, the EEG mu
rhythm was used as an index of motor mirror activity to study contextual influences on
the oscillatory neural activity during action observation. EEG event-related potentials
were also used to investigate the effects of others’ errors and rewards on one’s own
error- and reward-related neural system. As an additional line of inquiry, the
neurobiological factors related to social approach and avoidance behaviours in
schizophrenia were explored.

1. GENERAL BACKGROUND
18
Schizophrenia was used here as a model disorder for which deficits in social cognition
are well-known. The hope is that these studies will also help to inform therapeutic
strategies for the remediation of social cognitive skills.

2. THE RELEVANCE OF SELF AND OTHER IN MIRROR MOTOR ACTIVITY
19
Chapter 2
The relevance of self and other in mirror motor
activity
2.1. Introduction
The EEG mu rhythm suppression is evoked when an action is performed and when an
action is observed, also referred to as motor resonance (Marshall & Meltzoff, 2011). As a
result of this property of the mu rhythm, it has been closely linked with work on the
human mirror neuron system (Pineda, 2005). A number of studies have demonstrated
that the mu rhythm suppression is sensitive only to biological motion and actions that
are intentional or goal-directed, as compared to non-biological motion and actions
without intention (Iacoboni et al., 2005; Muthukumaraswamy et al., 2004b; Ulloa &
Pineda, 2007). As mentioned earlier, the administration of oxytocin, a “prosocial”
neuropeptide, has been shown to have a modulatory effect on the suppression of the mu
rhythm (Perry et al., 2010b). Social interaction can also enhance the mu suppression
during action observation (Oberman et al., 2007). There is much evidence to suggest
that the mu rhythm suppression has a functionally specific role in action understanding
and in social interaction, however this is still under vigorous debate. To simulate or
understand others’ actions involves some form of perspective-taking, and in motor
resonance, one must be able to recognise that the perspective being taken is not one’s
own. This is also the case for TOM and mentalizing. Although this distinction between
self and other in motor resonance can refer not only to perspective-taking, but may also

2. THE RELEVANCE OF SELF AND OTHER IN MIRROR MOTOR ACTIVITY
20
be relevant in terms of the referential framing of others’ actions to self or others. The
distinction between self and other is intrinsic to the concept of motor resonance,
however, the framing of actions in terms of self and other has not been previously
investigated. By manipulating the self-related context of the action we may gain more
insight into the functional specificity of the mu rhythm suppression, and its relevance in
social interaction. The aim of this study was to see if there was a difference in mu
rhythm suppression for executed and observed actions that were framed as self- and
other-relevant actions. It was hypothesized that self-relevant actions would induce
greater mu rhythm suppression, as compared to actions that were relevant to the other.
2.2 Method
2.2.1. Participants
5 right-hand dominant (according to the Edinburgh Handedness Inventory (Oldfield,
1971) females were recruited in Bochum, Germany. The mean age of participants was
26.6 years (SD = 1.34) and individuals with a history of neurological damage or
psychiatric illness were excluded. Informed consent was acquired from all subjects
before the experimental procedure began.
2.2.2. Task design
The experimental task consisted of three main parts: action execution, action
observation and a baseline task.
The observation part of the experiment used videos that depicted a person seen sitting
at a desk facing the camera, transferring objects with a spoon from one bowl to another.
There were three bowls on the table, one bowl to the right of the person in the video,
and the other two to the left of the person in the video (i.e. the performer), with one of
these two being closer to the camera and the other being closer to the performer (see
Fig. 2.1). Participants were told that the bowl placed closest to the camera was “their”
bowl, and the bowl closest to the performer was the performer’s bowl. In each video, the

2. THE RELEVANCE OF SELF AND OTHER IN MIRROR MOTOR ACTIVITY
21
performer was first seen in a resting position, with hands flat on the table, and then
would pick up the spoon from the table to transfer an object from the bowl to his right to
one of the two bowls to his left (“self”, i.e. the one closest to the camera, or the “other”
bowl, i.e. the one closest to the performer). Importantly, in the “self” condition, the
performer looked towards the camera, whereas in the “other” condition the performer
did not look at the camera. Therefore the performer in the “self" condition also had
direct eye gaze (i.e. simulated eye contact) but the gaze of the performer in the “other”
condition followed the object being transferred. Once the transfer had been made, the
performer would put the spoon back on the table and then return to the original resting
position. Therefore, each video clip consisted of 3 parts: 1) pre-movement (resting
state), 2) movement (action is performed) and 3) post-movement (resting state –
performer has returned to original position and is sitting still). The pre-movement stage
lasted for 1 sec, the movement stage lasted for 7 secs and the post-movement stage last
for 5 secs, however, each trial appeared as one continuous video. The length of the post-
movement time window allows for neurons in the motor cortex to fully resynchronise
(i.e. refractory period), and therefore allowing time for the mu rhythm to return to a
baseline resting state (as in Honaga et al., 2010). A fixation cross was presented for 1 sec
between trials. This part of the experiment began with a block of practice trials in which
7 observation trials (3 “self” and 4 “other” trials) were presented in a pseudorandom
order. The experimental block then followed, which consisted of 30 trials in total
whereby “self” and “other” actions were presented in a pseudorandom order.
For the action execution part of the experiment, three bowls were placed in front of the
participant, just as they were in the action observation part. These three bowls were
arranged in the same spatial configuration as seen in the action observation videos.
During the action execution part, a still image from the action observation video was
displayed on screen to control for visual stimulus. Cues on screen were also given to
participants to instruct them to which bowl they should transfer the object. These cues
came in the form of the words “yours” (i.e. “self” condition) or “theirs” (i.e. “other”
condition) appearing on screen when the action was to be executed. A countdown clock
was also presented on screen while the participants performed the action to try to keep
the duration of participants’ actions the same as those in the videos used in the action
observation part. The inter-movement interval was also kept consistent with the action
observation videos. An auditory cue was used to signal when the action should start and

2. THE RELEVANCE OF SELF AND OTHER IN MIRROR MOTOR ACTIVITY
22
when it should be terminated. 12 practice trials were completed before the
experimental block began. The experimental block consisted of 30 trials in total in which
the order of “self” and “other” trials were presented in a pseudorandom order.
The baseline task used an animated video of a black circle bouncing along a white
background, like a bouncing ball. The circle was made to look like it was bouncing
diagonally across the screen, appearing to start in the background and bouncing
towards the foreground. This was intended to mimic the timing and trajectory of the
observed actions in the action observation part. From video onset, the screen stayed
white for one second (to correspond to the pre-movement part), then the movement of
the circle lasted for 7 secs, with a refractory period of 5 secs after the circle movement
had terminated.
2.2.3. Procedure
The order of the action execution, action observation and baseline parts of the
experiment were counterbalanced across subjects to minimise order effects. Subjects
were sat in front of a computer screen during the whole experiment. In the action
execution part, movements were paced according to auditory cues and a countdown
clock. The participants were required to perform a series of practice trials until the
timing of movement was sufficiently synchronised to cues, particularly with movement
onset and offset. Participants performed a minimum of 7 practice trials, and more if the
experimenter deemed it necessary to have sufficient synchrony with the movement
onset and offset cues. In the action observation part, participants were asked to sit still
with their hands flat on the table, and to only count the number of items that were put
into “their” (i.e. “self” condition) bowl and the “other’s” bowl. At the end of the action
observation part, participants were asked how many items had been placed into “their”
bowl and how many had been placed in the “other’s” bowl. Both performer and
participant used their right arm for all trials.

2. THE RELEVANCE OF SELF AND OTHER IN MIRROR MOTOR ACTIVITY
23
Figure 2.1: A pictorial description of the experimental design showing snapshots of the action
observation (video) and action execution parts, with a timeline representing the duration of each part of
the videos.
2.2.4. EEG data acquisition
EEG was recorded at a sampling rate of 512 Hz from 32 channels with a Brain Products
BrainAmp system, using passive AgCl electrodes held in a BrainCap elasticated EEG cap
(from EasyCap). The 32 electrode positions were distributed over the scalp according to
the international 10-20 EEG system. An additional electrode was placed below the left
eye (EOG) to capture eye blinks. According to the EasyCap 32-channel standard
electrode configuration, the ground electrode was located along the midline, anterior to
electrode Fz. The ground electrode was located along the midline, more posteriorly,
between electrodes Cz and Fz. Following acquisition, the raw data were processed
offline with BrainVision Analyzer 2 (Brain Products GmbH). Firstly, the data was visually
inspected for segments with obvious artifacts that were then removed. The data was re-

2. THE RELEVANCE OF SELF AND OTHER IN MIRROR MOTOR ACTIVITY
24
referenced to all electrodes and submitted to a band-pass filter of 0.1 Hz to 100 Hz, with
a 50 Hz notch filter applied. Ocular correction was performed with the EOG as a
reference.
2.2.5. EEG data analysis of the mu rhythm
The mu suppression was extracted from the central electrodes overlaying sensorimotor
cortex; electrode positions C3, Cz, and C4. From the EEG data collected in the action
execution and action observation parts, the 7 sec action / movement epoch was selected
and segmented into 1000 msec segments, and further analysis was done with these
1000 msec segments. Automatic artifact rejection was used to identify and reject
segments if they exceeded ±100 µV. A Fast Fourier Transform (FFT) with a 10 %
Hamming window was performed separately on each of the 1000 msec baseline and
action observation epochs and an average was then taken for each condition, and
consequently powers in the alpha frequency band (8-13Hz) were exported. The same
procedure was applied to the 7 sec movement epoch from the EEG data collected in the
baseline bouncing ball part and this was used as the baseline for mu extraction (as
Oberman et al., 2005). To calculate the mu suppression, and control for individual
variability in alpha power, a natural log transform (ln) was calculated for the ratio of the
power of the alpha band of the action observation condition over the baseline condition
epochs accordingly (Oberman et al., 2005; Raymaekers et al., 2009).
Repeated measures ANOVA was used with the exported log ratio mu rhythm
suppression values with the action execution / observation conditions, self / other
conditions and electrode position (C3, Cz, C4) as within-subject factors. To test the
hypothesis, pair-wise comparisons were then conducted to compare self and other
conditions for action execution and action observation conditions independently at each
electrode position. The appropriate assumptions for performing an ANOVA were
checked for.

2. THE RELEVANCE OF SELF AND OTHER IN MIRROR MOTOR ACTIVITY
25
2.3. EEG mu rhythm results
Significant main effects for the execution / observation condition was found for the EEG
mu rhythm suppression (F(1,4)=21.27, p=0.01). However, no significant main effects
were found for self & other conditions (F(1,4)=2.90, p=0.16) and electrodes
(F(2,3)=2.76, p=0.17). Though the 2-way interaction effects of self & other and execution
& observation were significant (F(1,4)=10.91, p=0.03). As there was no significant main
effect of electrode found, data from the 3 electrodes were pooled. To explore the 2-way
interaction further, post-hoc comparisons were made between self & other and action
execution & observation. The difference between self and other conditions for action
execution was close to being significant (t(4)=2.66, p=0.06), with actions for the other
condition having greater mu suppression. Although it does not appear to be the case
from superficially looking at the bar chart in figure 2.2, there was a significant difference
revealed between self and other conditions for action observation (t(4)=-3.03, p=0.04)
with the self condition producing greater mu suppression. In addition, the difference
between self and other conditions for the mu suppression during action execution was
not at all far from significance (t(4)=2.65, p=0.057), with actions performed in the other
condition producing greater mu suppression. Figure 2.2 shows the mu suppressions
results comparing self and other conditions for the action execution and action
observation. It is important to note that a more negative mu rhythm means greater mu
suppression, and therefore, more mu suppression reflects greater motor cortex activity.

2. THE RELEVANCE OF SELF AND OTHER IN MIRROR MOTOR ACTIVITY
26
Figure2.2: Bar chart showing the EEG mu rhythm suppression pooled over electrodes covering motor
cortex (C3, Cz, C4), with self and other conditions shown for action execution and action observation.
Error bars reflect standard error of the mean. (*p=<0.05)
2.4. Summary of results
This study aimed to investigate the modulation of the mu rhythm during action
execution and action observation for actions that were either related to the “self” or to
the “other”. As expected action observation evoked substantially less mu rhythm
suppression than during the execution of an action. Though most interestingly, the self
and other conditions did seem to have a substantial effect on the mu suppression in both
action execution and action observation, though this difference only reached significance
in the action observation condition. Although, the difference between self and other in
the action execution task was not far at all from reaching significance, which may have
been due to the small sample size. However, due to the fact that a mu suppression was

2. THE RELEVANCE OF SELF AND OTHER IN MIRROR MOTOR ACTIVITY
27
found in only 5 subjects, as well as an effect of the self and other manipulation, this
implies that the effect is likely to be quite strong and robust, as this was also seen in both
electrodes C3 and C4. In summary, observed actions that were associated with the self
induced greater mu suppression than actions that were associated with the other,
whereas executed actions that were associated with the self induced less mu
suppression than executed actions for the other.

3. THE INFLUENCE OF REWARD AND PUNISHMENT ON THE MOTOR MIRROR SYSTEM
28
Chapter 3
The influence of reward and punishment on the motor
mirror system
3.1. Introduction
In social psychology and behavioural economics, there has been a long tradition of
theorists that have explored the interactions between reward and social behaviour
(Fehr & Camerer, 2007; Gintis & Fehr, 2012). In fact, many psychologists have referred
to “social rewards”, i.e. rewards framed in the social context (Festinger, 1954; Johns &
Quay, 1962), such as receiving social approval (Morris & Coady, 1974). However,
empirical work in neuroscience has only begun to explore the neural correlates of social
rewards (Behrens et al., 2008; Krach et al., 2010). It is known that the neural coding of
reward is crucially involved in action selection and is therefore also intrinsic to goal-
directed behaviour (Schultz, 2000). As the activity in the mirror neuron system has been
shown to be specific only to observed actions that are goal-directed (Rizzolatti et al.,
1996), it follows that reward and punishment are likely to have reciprocal interaction
effects on the neural activity associated with action observation. Consequently, this may
also influence the degree to which action understanding and observational learning take
place. Emerging evidence is suggesting that problems in social functioning seen in
autism may be founded upon an impaired response to social rewards (Dichter &
Adolphs, 2012). A recent study from Sugawara and colleagues (2012) demonstrated that
motor skill learning can be improved if positive, or rewarding feedback is given, even in
offline consolidation of the motor skill. Spontaneous mimicry of facial expressions has

3. THE INFLUENCE OF REWARD AND PUNISHMENT ON THE MOTOR MIRROR SYSTEM
29
also been shown to be enhanced when the emotional face being mimicked has been
associated with a high reward, as compared to those faces associated with a lower
reward (Sims et al., 2012). These studies, though indirectly, point to a likely role for
reward in motor cortex activity during action observation. However, it is still unclear
how reward and punishment may interact with the processing of others’ actions at the
neural level.
In addition to the potential sensitivity of the mirror neuron system to reward-related
contextual differences, there is evidence to suggest that the system may also be
influenced by the perspective in which actions are viewed. Primate work has identified
some mirror neurons in area F5 of the macaque premotor cortex that are tuned
specifically to actions that are presented in an egocentric (1st-person) point of view,
whereas others respond only to actions presented in an allocentric (3rd-person) point of
view (Caggiano et al., 2011). Human behavioural studies have also provided indirect
evidence that the perspective from which an action is viewed can influence motor
resonance through visuomotor priming (Vogt et al., 2003) with actions seen from a 1st-
person perspective leading to greater action identification (Libby, et al., 2009),. This is
thought to be due to the reason that actions seen from an egocentric, as opposed to an
allocentric perspective, may be easier to transpose onto the motor cortex of the
observer (Jeannerod & Anquetil, 2008). However, studies looking at motor cortex
excitability during action observation show mixed results on the effect of perspective
(Alaerts et al., 2009; Maeda et al., 2002).
The primary aim of this study was to compare the mu rhythm suppression during
observation of actions that are rewarding, punishing or neutral for the observer. A
secondary aim was to explore the effect of perspective-taking on the mu suppression.
Finally, a further goal of the study was to better characterize the temporal dynamic
associated with changes in the mu rhythm, in relation to potentially different processing
stages during action observation, given that most previous studies have typically
overlooked the temporal component of the mu suppression. It was hypothesised that
when rewards were associated with observed actions, this would induce greater mu
rhythm suppression as opposed to punishing and neutral actions. It was also predicted
that actions seen from a 1st-person perspective would lead to greater mu suppression, as
compared to actions observed from a 3rd-person perspective.

3. THE INFLUENCE OF REWARD AND PUNISHMENT ON THE MOTOR MIRROR SYSTEM
30
3.2 Method
3.2.1. Participants
17 right-handed (according to the Edinburgh Handedness Inventory (Oldfield, 1971))
females were recruited from Ghent University, Belgium. The mean age of participants
was 20.3 years (SD = 1.99) and potential participants with a history of neurological
damage or psychiatric illness were excluded. Informed consent was acquired from all
subjects before the experiment began.
3.2.2. Task design
Participants sat with their hands flat on a table in front of the screen while they watched
a series of videos. For each participant, 6 different videos were displayed 20 times each,
with actions either from a 1st-person or 3rd-person point-of-view, being rewarding,
punishing or neutral actions (i.e. 2x3 conditions). To control for possible effects of a
spatial bias, 6 different video sets were created in which different configurations of the
“+”, “-” and “0” labels were superimposed each bowl. Participants were randomly
assigned to one of the 6 video sets. 120 videos were presented in total in each testing
session, in a pseudorandom order. Trials were split into 6 blocks, with a single video
used in each trial. A fixation cross was presented for 1sec before each video. A block of 8
practice trials came before the 6 blocks of main trials. Each video lasted for a total of 11
secs, and the movement started 1 sec after the start of the video. The performer in the
video started the movement in a resting position with hands flat on the table, and
returned to this position at movement offset. The movement lasted for 6 secs and after
movement termination and the video continued for a further 4 sec, with the performer
in the resting position.

3. THE INFLUENCE OF REWARD AND PUNISHMENT ON THE MOTOR MIRROR SYSTEM
31
3.2.3. Procedure
Participants watched the video clips, which displayed two people sitting at a table and
transferring coins from one bowl to one of three other bowls in the middle of the table.
In the videos, one person was seen from a 1st-person and the other from a 3rd-person
point-of-view. The three bowls with the “+”, “-” and “0” on them were arranged on the
table along one plane and were all equidistant from the performer. The bowl that the
performer picked the coin up from was closer to each of the performers than the other
three bowls. One of the three bowls was labeled with a “+”, one with a “-” and one with a
“0”. Participants were told that each time a coin was put into the “+” bowl, they would
receive plus one euro (rewarding), when a coin was put into the “-” bowl, they would
have one less euro (punishing), and when a coin was put into the “0” bowl there would
be no change (neutral). Participants were asked to only sit still, watch and count the
number of coins transferred to each bowl and keep track of the amount of money they
would win or lose at the end. Participants were told they had €20 “in the bank” to start
with, which would go up or down according to the number of rewarding and punishing
actions observed.
After each block of trials, participants were asked to report the amount of money “in the
bank” and also rated the previous action for subjective pleasantness, arousal and how
easy it was to pay attention according to a visual analogue scale from 0 to 10 (with the
pleasantness rating from-10 to 10, with -10 being “very unpleasant” and 10 being “very
pleasant”).
According to visual inspection of the vides, a critical 3 sec epoch was selected for the mu
rhythm analysis, in which the reward-related conditions (reward, punishment or
neutral) differ, i.e. when the action begins to diverge to one of the rewarding, punishing
or neutral bowls.

3. THE INFLUENCE OF REWARD AND PUNISHMENT ON THE MOTOR MIRROR SYSTEM
32
3.2.4. EEG data acquisition
EEG was recorded from 64 channels with a BioSemi ActiveTwo system at a sampling
rate of 2,048 Hz but was later down sampled offline to 512 Hz. The 64 electrode
positions were distributed over the scalp according to the international 10-20 EEG
system. An additional electrode was placed above the right eye in line with the pupil
(vertical EOG), plus one placed at the outer canthus (horizontal EOG). According to the
BioSemi criteria, the predetermined electrode locations CMS and DRL served as the
reference and ground electrodes, respectively. Following acquisition, the raw data were
processed offline with BrainVision Analyzer 2 (Brain Products GmbH). Firstly, the data
was visually inspected and channels that were particularly noisy were identified,
removed and later topographically interpolated. The data was then re-referenced to all
electrodes and submitted to a band-pass filter of 0.1 Hz to 30 Hz, with a 50 Hz notch
filter applied. Ocular correction was performed with the vertical EOG.
3.2.5. Behavioural data analysis
Each score on the visual analogue scale for the subjective rating of arousal and attention
was calculated by measuring the distance from the start of the line to the point at which
participants had marked a cross. For the subjective rating of pleasantness, a mid-point
on the scale was measured and taken as the zero point, with responses falling to the left
of the zero point representing negative scores, and those to the right being positive. For
each question, the mean score was taken for all rewarding, punishing and neutral
actions. Paired t-tests were later performed between scores on all conditions for each
question. A correlation analysis was also performed to investigate whether scores of
pleasantness, arousal and attention related independently to the mu suppression.
3.2.6. EEG mu rhythm suppression analysis
The mu suppression was extracted from the central electrodes overlaying sensorimotor
cortex; electrode positions C3, C1, Cz, C2, and C4. Baseline and action observation
epochs were first determined. For the baseline for the mu extraction, the 1000 msec

3. THE INFLUENCE OF REWARD AND PUNISHMENT ON THE MOTOR MIRROR SYSTEM
33
epoch preceding movement onset (after video onset) was used as the baseline for mu
extraction (as Schuch et al., 2010). The 3 sec action observation critical epoch was
segmented into 1000 msec segments, and further analysis was done with these 1000
msec segments. EEG artifacts were identified and rejected if they exceeded ±100 µV. A
Fast Fourier Transform (FFT) with a 10 % Hamming window was performed separately
on each of the 1000 msec baseline and action observation epochs and an average was
then taken for each condition, and consequently powers in the alpha frequency band (8-
13Hz) were exported. To calculate the mu suppression, and control for individual
variability in alpha power, a natural log transform (ln) was calculated for the ratio of the
power of the alpha band of the action observation condition over the baseline condition
epochs accordingly (Oberman et al., 2005; Raymaekers et al., 2009).
Repeated measures ANOVA was used with the exported log ratio mu rhythm
suppression values with the reward-related conditions (rewarding, punishing, neutral)
and electrode position (C1, C2, C3, C4, Cz) as within-subject factors. Pair-wise
comparisons were conducted for significant main effects (figure 3.2). The appropriate
assumptions for performing an ANOVA were checked for. Whole-head topographical
plots were acquired with the mapping function of BrainVision Analyzer 2 by selecting
the 8-13Hz frequency band for all electrodes following an FFT of the same 3 sec segment
used for the mu rhythm extraction. These were then averaged across subjects for the
reward conditions (i.e. rewarding, punishing, and neutral).
3.2.7. Time-course analysis of EEG mu power
To calculate the change in mu power (8-13 Hz) during the course of the video, averages
were taken for each consecutive 500 msec segment from the start to the end of the
video. This was done for each reward condition (rewarding, punishing, and neutral) (see
figure 3.3). A post-hoc analysis was done on the 3 sec epoch used for the mu rhythm
analysis, which was split into three 1 sec epochs. Paired t-tests were performed to
compare differences between each condition in each of the three 1 sec epochs.

3. THE INFLUENCE OF REWARD AND PUNISHMENT ON THE MOTOR MIRROR SYSTEM
34
3.3. Results
3.3.1. Behavioural results
Results show that the subjective ratings of pleasantness were congruent to the reward
conditions (figure 3.1), with rewarding actions being judged as the most pleasant, and
punishing the least. Paired comparisons revealed significant differences between
pleasantness ratings of rewarding and punishing (t(16)=4.59, p<0.001), rewarding and
neutral (t(16)=5.18, p<0.001), and punishing and neutral actions (t(16)=-2.38, p=0.03).
It was also evident from subjective ratings of arousal that rewarding actions were more
arousing than neutral actions (t(16)=3.23, p=0.005). Importantly, the ratings
demonstrate that differences between reward conditions were not accounted for by
differences in attention, showing no significant differences between conditions. It is also
worth noting that the correlation analyses revealed no significant correlations between
behavioural ratings of pleasantness, arousal nor attention with the mu suppression.
Figure 3.1: Subjective ratings of pleasantness, arousal and ease of paying attention for rewarding,
punishing and neutral observed actions (*=p<0.05, n.s.=p>0.05) (from Brown et al., 2013). Ratings for
pleasantness were scored by using the midpoint of the scale as the zero-point. Error bars represent the
standard error.

3. THE INFLUENCE OF REWARD AND PUNISHMENT ON THE MOTOR MIRROR SYSTEM
35
3.3.2. EEG mu rhythm suppression results
Non-significant effects were found for perspective conditions (F (1,16)=1.50, p=0.24),
nor for the interaction between reward-related conditions and perspective (F
(2,15)=0.21, p=0.82. Therefore, perspective conditions were not included in any of the
further analysis as egocentric and allocentric perspective conditions were pooled
together. Significant main effects for the EEG mu rhythm suppression were found among
the three reward-related conditions (F(2,15)=3.74, p=0.05) and six electrodes
(F(4,13)=4.22, p=0.02). Pairwise comparisons between reward-related conditions
showed significant differences between rewarding and punishing (t(16)=-2.15, p=0.05)
and rewarding and neutral (t(16)=-2.36, p=0.03), however there was no significant
difference between punishing and neutral actions (t(16)=-1.42, p=0.17). Figure 1b
shows the (log ratio relative to baseline) mu rhythm suppression for each reward-
related condition (rewarding, punishing, and neutral) pooled over the electrodes
covering sensorimotor cortex, and perspective conditions (egocentric and allocentric).
The largest mu suppression was found for rewarding and the smallest for neutral
actions (Fig. 3.2). Topographical maps of the mu power (Fig. 3.2) including all 64
channels demonstrated substantial suppression predominantly over medial frontal and
sensorimotor areas, with little overlap between the two.

3. THE INFLUENCE OF REWARD AND PUNISHMENT ON THE MOTOR MIRROR SYSTEM
36
Figure 3.2: EEG mu rhythm suppression during the observation of rewarding, punishing and neutral
actions pooled over electrodes C3, C1, Cz, C2 and C4 (*=p<0.05, n.s.=p>0.05) (from Brown et al., 2013).
Error bars represent the standard error.
3.3.3. Time course analysis of EEG mu power results
Remarkably, a closer look at the time course of the mu power effect (figure 3.3) revealed
a significant suppression at video onset, followed by a second significant suppression
around 3.5 to 4 sec after video onset. In addition to this, the time-plot revealed an
asymmetry between reward conditions following this second suppression. Post-hoc
comparisons confirmed this asymmetry whereby, in the first second of the critical 3 sec
epoch, rewarding actions were significantly different from neutral (t(16)=2.65, p=0.02)
and punishing actions (t(16)=2.61, p=0.02),whereas no significant difference was found
between punishing and neutral actions (t(16)=0.66, p=0.52).The difference between
punishing and neutral actions only reached significance during the third second of this
epoch (t(16)=2.12, p=0.05), and therefore demonstrating a later mu suppression for
punishing than rewarding actions.

3. THE INFLUENCE OF REWARD AND PUNISHMENT ON THE MOTOR MIRROR SYSTEM
37
Figure 3.3: Plot showing the change in EEG mu power, averaged from electrodes over sensorimotor areas
(C3, C1, Cz, C2, C4) over the course of the video, showing the different reward conditions. The critical 3 sec
time window selected for the mu rhythm suppression analysis (used in Figure 3.2) is also highlighted here
between 4 and 7 secs. Stills taken from the video stimuli are presented along the horizontal axis. The
dotted vertical lines mark the 1 sec epochs used to compare the latency of the mu suppression between
conditions (from Brown et al., 2013). Error bars represent the standard error.
3.4. Summary of results
Here, the effect of reward and punishment on the mu rhythm suppression during action-
observation was examined. It was predicted that mu-rhythm suppression would be
greater for rewarding actions, relative to the punishing and neutral conditions, and that
the observed effect would be larger in the 1st, as opposed to 3rd person point-of-view.
These results partly support the hypothesis, with the main finding revealing reward-
related modulation of motor cortex activity, as shown by the systematic changes in the
mu rhythm suppression during action observation. When one observed others’ actions,
it appeared that there was greater motor resonance if the consequence of the action was
associated with a reward for the observer, whereas actions associated with punishment
induced less motor resonance. Importantly, observed actions that did not lead to a
reward or punishment, i.e. were embedded in a neutral context, induced comparatively
the least motor cortex activity. We also hypothesised that a difference in perspective-
taking (1st-person vs. 3rd-person) would have an influence on the expression of the mu

3. THE INFLUENCE OF REWARD AND PUNISHMENT ON THE MOTOR MIRROR SYSTEM
38
suppression; however our results did not confirm this prediction, in contrast to some
previous studies (Libby et al., 2009).
When looking at the distribution of the mu suppression over the whole head with the
topographic mapping, it appears that the effects of the experimental manipulation were
primarily driven by suppression over sensorimotor areas, rather than posterior occipital
areas or frontal alpha suppression. Hence, this rules out the possibility that the effects
reported in this study were confounded by systematic changes in attention-based
posterior alpha. The behavioural results also confirmed that the effect of the reward
manipulation was not driven by attentional differences.
Interestingly, our analysis of the temporal dynamic of the mu rhythm showed a second
suppression component that appeared to be the result of the reward condition effect.
Finally, it was also shown that the mu rhythm suppression occurred later for punishing
actions than rewarding ones. Despite punishing actions inducing greater mirror motor
activity than neutral actions, it appeared that punishing actions were associated with a
somewhat delayed and prolonged mirror motor response.

4. REWARD-RELATED MODULATION OF THE MIRROR SYSTEM IN SCHIZOPHRENIA
39
Chapter 4
Reward-related modulation of the mirror motor
system in schizophrenia
4.1. Introduction
Schizophrenia is disease that is marked by positive and negative symptoms that are
accompanied by deficits in social cognition, which often lead to severely impaired social
functioning. Social cognitive deficits in schizophrenia are also associated with problems
in distinguishing between self and other. This has been associated with the deficits in
TOM and the capacity to understand others’ intentions and emotions. Selective
abnormalities in reward processing are also well-known in schizophrenia, which are
thought to have a central role in the expression of psychotic negative symptoms (Gold et
al., 2008). Considering the concurrent deficits in social cognition and reward processing,
this makes schizophrenia an interesting model to explore the potential effects of reward
and punishment on motor resonance, especially as some authors have already proposed
that a potential underlying deficit in the mirror neuron system may contribute to
explaining deficits in higher level social cognitions such as TOM, emotion processing and
attribution bias (Arbib & Mundhenk, 2005; Enticott et al., 2008).
Intuitively, it seems plausible to argue that patients with schizophrenia and poor social
functioning would also have diminished mirror neuron related activity, as shown by
Singh and colleagues (2011) with the mu rhythm. However, the study from McCormick

4. REWARD-RELATED MODULATION OF THE MIRROR SYSTEM IN SCHIZOPHRENIA
40
and others (2012) found excessive mirror neuron related activity in an actively
psychotic group of patients with schizophrenia. These conflicting results encourage
further exploration of mirror neuron activity, as indexed by the mu rhythm suppression,
in patients with schizophrenia. If reward does play a role in the mu rhythm suppression,
as shown in the previous chapter, then a deficit in reward processing may also have a
detrimental impact on motor resonance during action observation, which has not been
accounted for in previous action observation studies.
The aim of this study was to explore the possible interaction between reward and the
mu rhythm suppression during action observation in a population with schizophrenia,
and to compare this to a healthy control group. It was predicted that there would be a
general difference in the magnitude of the mu rhythm suppression between people with
schizophrenia and healthy control participants. It was also hypothesised that the
modulating effects between the rewarding and punishing conditions on the mu
suppression would also be different in the patient group, as compared to healthy
controls. As a further hypothesis, it was predicted that the mu suppression would have
some association with behavioural measures of self-reported empathy, affective mood
state and psychotic symptoms.
4.2 Method
4.2.1. Participants
13 right-handed participants (7 male) diagnosed with schizophrenia were recruited
from the LWL University Hospital Bochum, with a mean age of 37.9 years (SD=10.3).
Diagnosis was confirmed with the ICD-10 and all patients were considered as clinically
stable and the suitability for participation in the research study was determined by the
psychiatrist responsible for the patient. Inclusion criteria for the patient group only
permitted the selection of participants who had received a F20 (ICD-10) diagnosis of a
schizophrenia disorder that was not drug-induced, had no history of neurological injury
and had no obvious motor deficits. Psychotic symptomatology of all participants was
assessed with the Positive and Negative Syndrome Scale (PANSS (Kay et al., 1987)). The
PANSS is a 30-item semi-structured interview designed to assess five symptom

4. REWARD-RELATED MODULATION OF THE MIRROR SYSTEM IN SCHIZOPHRENIA
41
categories associated with schizophrenia: positive symptoms (i.e., hallucinations and
delusions), negative symptoms (i.e., avolition and anhedonia), cognitive symptoms (i.e.,
thought disorder), hostility, and depression. A qualified and PANSS-trained psychiatrist
assigned a score from 1 to 7 for each item, with higher scores indicating more severe
psychopathology. For the whole group, the PANSS positive mean score was 19.4
(SD=7.9), PANSS negative mean was 25.8 (SD=7.7) and PANSS general was 47.6
(SD=14.5). All patients were taking atypical antipsychotic medication.
13 age matched (6 male) right-handed healthy control participants were also recruited
as a comparison group, with a mean age 34.3 years (SD=11.1). Healthy participants were
only included if they had no history of psychiatric illness, no neurological injury and no
deficits in motor functioning. The study was approved by the local ethics committee and
all participants gave written consent to participate in the study. The capacity to give
consent for each patient was confirmed by a consultant psychiatrist, following
psychiatric assessment.
4.2.2. Task design
The same experimental paradigm used in the previous chapter was also used in this
study, and notably, none of the healthy participants from the study described in the
previous chapter were used in the healthy control group for this study. EEG was
recorded from all participants during the observation of videos, as described in the
methods section in the last chapter (chapter 3).
4.2.3. Behavioural tests
The German version of the Interpersonal Reactivity Index (IRI; Davis 1980) was also
given to all participants: Der Saarbrücker Persönlichkeitsfragebogen (SPF; Paulus,
2009). This self-administered questionnaire is designed to be a test of different
dimensions of empathy. The questionnaire consists of 16 empathy-related statements
for which participants give a score from 1 to 5, with 1 meaning that the statement does
not apply to them at all, and 5 being that this applies very well. Each item score

4. REWARD-RELATED MODULATION OF THE MIRROR SYSTEM IN SCHIZOPHRENIA
42
contributes to one of four factor scores, with these factors being: Fantasy, Perspective-
taking, Empathetic concern and Personal distress. A final score for each of the four
factors is determined as these four factors are considered independent, so a total score
is not considered valid.
4.2.4. Procedure
Firstly, the behavioural test was administered to participants, which was then followed
by the EEG experiment. All instructions for the administration of the experiment used
during the EEG recording were also the same as the procedure describes in the previous
chapter, except they were given in German.
4.2.5. EEG data acquisition
EEG was recorded at a sampling rate of 512 Hz from 32 channels with a Brain Products
BrainAmp system, using passive AgCl electrodes held in a BrainCap elasticated EEG cap
(from EasyCap). The 32 electrode positions were distributed over the scalp according to
the international 10-20 EEG system. An additional electrode was placed below the left
eye (EOG) to capture eye blinks. Following acquisition, the raw data were processed
offline with BrainVision Analyzer 2 (Brain Products GmbH). Firstly, the data was visually
inspected for segments with obvious artifacts that were then removed. The data was re-
referenced to all electrodes and submitted to a band-pass filter of 0.1 Hz to 100 Hz, with
a 50 Hz notch filter applied. Ocular correction was performed using independent
component analysis (Jung et al., 2000).

4. REWARD-RELATED MODULATION OF THE MIRROR SYSTEM IN SCHIZOPHRENIA
43
4.2.6. Data analysis
4.2.6.1. Behavioural data analysis
The scores on the visual analogue scale for the subjective rating of pleasantness, arousal
and attention were calculated using the same method as described in chapter 3. Paired t-
tests were later performed between scores on all conditions for each question.
Correlation analyses were also performed to investigate whether subjective ratings of
pleasantness, arousal and attention related independently to the mu suppression.
Separate ANOVAs were used to test for group differences on the subjective ratings of
pleasantness, arousal and attention for observed actions, and was also used to test for
group differences in empathy scores on the SPF.
4.2.6.2. EEG mu rhythm suppression analysis
The mu suppression was extracted from the central electrodes overlaying sensorimotor
cortex; electrode positions C3, Cz and C4. The same procedure described in chapter 3 for
mu extraction was followed here. Therefore, the mu suppression represented a log
transformed ratio of the alpha power during the critical 3 sec action observation epoch
and the 1 sec epoch preceding movement onset.
After the appropriate assumptions for performing an ANOVA were checked for, an
ANOVA was used to check for group differences on the mean mu suppression values.
Following this, a repeated measures ANOVA was used with the exported log ratio mu
rhythm suppression values with the perspective (1st and 3rd-person perspective / self
and other perspective), reward-related conditions (rewarding, punishing, neutral) and
electrode positions (C3, Cz and C4) as within-subject factors, and group (patient,
control) as the between-subjects factor. Pair-wise comparisons were conducted for
significant main effects, within each group. Whole-head topographical plots were
acquired with the mapping function of BrainVision Analyzer 2 by selecting the 8-13Hz
frequency band for all electrodes following an FFT of the same 3 sec segment used for

4. REWARD-RELATED MODULATION OF THE MIRROR SYSTEM IN SCHIZOPHRENIA
44
the mu rhythm extraction. These were then averaged across subjects, within each group,
for all conditions (i.e. rewarding, punishing, and neutral).
Following this initial analysis of the mu rhythm, a Pearson correlation was performed
with behavioural scores and the EEG data to look for possible relationships between the
behavioural measures and the mu rhythm suppression. This was done with the whole
sample and separately for each group. Furthermore, a correlation analysis was also done
with the PANSS scores and the mu rhythm in the patient group.
4.3. Results
4.3.1. Behavioural results
No significant group differences were found for empathy ratings on the SPF between
patients and matched controls. In regards to the subjective ratings of the observed
actions, the comparison of the patient group and healthy control group did not reveal
any significant differences, as shown in figure 4.1. Pairwise comparisons of rewarding,
punishing and neutral conditions on subjective ratings for the whole sample (including
both patients and controls) showed only significant differences on ratings for
pleasantness between rewarding and punishing actions (t(25)=3.09, p<0.01), and
between punishing and neutral actions (t(25)=-2.27, p=0.035).

4. REWARD-RELATED MODULATION OF THE MIRROR SYSTEM IN SCHIZOPHRENIA
45
Figure 4.1. Subjective ratings of pleasantness, arousal and ease of paying attention for rewarding,
punishing and neutral observed actions, showing ratings for the schizophrenia patient group (blue) and
healthy control group (green). Ratings for pleasantness were scored by using the midpoint of the scale as
the zero-point. Error bars represent the standard error.
4.3.2. EEG mu rhythm results
Most notably, there did appear to be a trend for patients to have lower mu suppression
than healthy controls, although this did not reach significance (F(1,25)=1.392, p=0.25).
The mean mu suppression values for both groups, pooled over all conditions, are
displayed in figure 4.2, along with topographical plots of the mu suppression. The
topographical maps show the most substantial suppression predominantly over left
frontal and bilateral motor areas.
Just as in the previous data presented in chapter 3, no significant effects were found for
perspective conditions (F (1,25)=0.77, p=0.39), nor for the interaction between reward-
related conditions and perspective (F (2,24)=0.78, p=0.46). Therefore, perspective
conditions were not included in any of the further analysis as 1st and 3rd-person
perspective conditions were pooled together.

4. REWARD-RELATED MODULATION OF THE MIRROR SYSTEM IN SCHIZOPHRENIA
46
Figure 4.2: Mean mu suppression for schizophrenia patient group and matched healthy control group
during action observation, pooled across conditions and central electrodes (C3, Cz, C4). Topographical
maps of mu suppression for action observation are also displayed here above the bar chart. A darker
colour in these maps represents lower mu power. Error bars represent the standard error.
A significant main effect of reward was found for the mu rhythm suppression
(F(2,24)=3.86, p<0.01), although the 2-way interaction between reward and group did
not reach significance (F(2,24)=1.10, p=0.35). There were also no significant main
effects found between electrodes. For the patient group, pairwise comparisons between
reward-related conditions revealed significant differences between rewarding and
neutral actions (t(12)=-2.28, p=0.04) but not between rewarding and punishing actions
(t(12)=-0.29, p=0.77), or between punishing and neutral actions (t(12)=-1.59, p=0.14).
For the healthy control group, only a significant difference was found between punishing
and neutral actions (t(12)=-2.78, p=0.02), and not between rewarding and punishing
(t(12)=1.63, p=0.13) or rewarding and neutral actions (t(12)=-1.10, p=0.29). Figure 4.3
shows the mu rhythm suppression in each group for each reward-related condition
(rewarding, punishing, and neutral) pooled over electrodes C3, Cz and C4, and over
perspective conditions (1st and 3rd).

4. REWARD-RELATED MODULATION OF THE MIRROR SYSTEM IN SCHIZOPHRENIA
47
Figure 4.3: Mu suppression over electrodes C3, Cz and C4 for schizophrenia patient group and healthy
control group during the observation of rewarding, punishing and neutral actions. Error bars represent
the standard error.
No significant correlations were found between the mu power and subjective ratings of
pleasantness, arousal and attention for the observed actions. When looking at the
sample as a whole, including both patients and healthy controls, a significant negative
correlation was found between the overall mu power and the perspective–taking factor
on the empathy scale (SPF) (r=-0.41, p=0.04), whereby greater perspective-taking was
associated with greater mu suppression. To explore this relationship further, correlation
analyses were done for each group separately, revealing that the correlation between
the mu power and empathy scores only reached significance in the healthy control
group and not in the patient group. The healthy control group demonstrated a strong
negative correlation between the mu power and the perspective-taking factor on the SPF
(r=-0.80, p<0.01), shown in figure 4.4, which explains the correlation found with the
whole sample.

4. REWARD-RELATED MODULATION OF THE MIRROR SYSTEM IN SCHIZOPHRENIA
48
Figure 4.4: Scatter plot for data from only the healthy control group, to demonstrate the relationship
between mean mu rhythm suppression and empathy scores in the perspective-taking domain, along with
line of best fit.
In the patient group, a strong negative correlation was found between the overall mu
power and negative psychotic symptoms (r=-0.67, p=0.02), as reflected from the PANSS
scores (figure 4.5), which indicated that the patients with greater negative symptoms
exhibited greater mu suppression. This relationship between negative symptoms and
mu suppression did not appear to be specific to any one of the reward-related
conditions.

4. REWARD-RELATED MODULATION OF THE MIRROR SYSTEM IN SCHIZOPHRENIA
49
Figure 4.5: Scatter plot for data from only the schizophrenia patient group, to demonstrate the
relationship between mean mu rhythm suppression and negative psychotic symptoms, as indicated by the
PANSS (Positive and Negative Syndrome Scale), along with line of best fit.
4.4. Summary of results
This study aimed to compare the differences in reward-related EEG mu rhythm
suppression between a patient group with schizophrenia and a matched healthy control
group. The hypothesis that the patient group would exhibit a different degree of mu
rhythm suppression as compared to a healthy control group was not shown to be
statistically significant, although a trend for lower mu suppression in the patient group
was emerging. Though unexpectedly, the effect of reward on the mu suppression in the
healthy group did not follow the finding from the study described in the previous
chapter, as the healthy controls in this study showed the greatest mu suppression for
punishing actions, whereas the patient group seemed to follow a more similar pattern as
those reported in the previous chapter, whereby rewarding actions induced the greatest
mu suppression.

4. REWARD-RELATED MODULATION OF THE MIRROR SYSTEM IN SCHIZOPHRENIA
50
The associations between the mu suppression and the behavioural data revealed a
strong association with empathy scores, though only in the healthy control group.
Healthy controls with greater perspective-taking demonstrated greater overall mu
rhythm suppression. In terms of the relationship between mu suppression and psychotic
symptoms, more severe negative symptoms were found to be related to greater overall
mu suppression in the patient group.

5. PROCESSING THE OUTCOMES OF OTHERS’ ACTIONS
51
Chapter 5
Processing the outcomes of others’ action - sharing
others’ rewards, losses and errors
5.1. Introduction
An early negative going ERP has been found to be elicited when a person makes an
erroneous response (Falkenstein et al., 1990). This is referred to as the error-related
negativity (or ERN). A few studies have also demonstrated that a similar ERN is present
when others’ errors are observed (van Schie et al., 2004), suggesting a form of shared
representation of others’ errors. Similarly, a negative going ERP is also found shortly
after feedback is given following a choice response, i.e. the point in which the subject
discovers whether the outcome of their choice is a loss or gain, namely the feedback
related negativity (FRN). The FRN has been suggested to reflect activity in the reward
system as it is modulated by the valence of the outcome (Hajcak et al., 2006). Holroyd
and colleagues (2003) demonstrated that the FRN can be induced by both monetary
feedback (gain or loss) and performance feedback (i.e. good or bad). There are different
interpretations of what neural activity is actually being reflected in the FRN. Yeung and
colleagues (2005) have suggested that the FRN could actually be reflecting the reward
signal, without the requirement of any overt action. Nieuwenhuis and colleagues (2004)
demonstrated that the FRN is an index of the difference between the expected and the
actual outcome, and therefore may be an index of the prediction error, in which the
largest FRN was generated with unexpected negative feedback. In terms of a social
context, an ERP comparable to the FRN can be elicited when feedback is given from

5. PROCESSING THE OUTCOMES OF OTHERS’ ACTIONS
52
observing another person receiving positive or negative feedback in a reinforcement
learning task. This has been called the observational FRN (oFRN). The ERN and FRN that
are induced when observing other’s performance (i.e. oERN and oFRN) is likely related
to the evaluation of others’ outcomes and could reflect some shared or empathetic
response to someone else’s wins, losses and errors. There is evidently an intimate
relationship between the ERN and FRN and their associated cognitive functions, but this
relationship still requires further clarification. The FRN’s sensitivity to feedback
expectancy has also never been explored in the social context, in terms of the
observation of another person’s performance feedback.
The following study investigated the FRN induced when receiving one’s own feedback
and when observing the feedback outcome of others’ performance, while manipulating
reward expectancy. The study sought to induce an oFRN when observing the outcome of
others’ responses, while also seeking to induce an oERN when observing others’ errors,
in the frame of an associative learning task. Firstly, in accordance with the previous
literature, it was hypothesised that the valence of the feedback during the observation of
others’ performance would modulate the size of the associated oFRN. Secondly, it was
predicted that the observation of others’ errors in an associative learning task, where
the expectation was high, would induce an oERN. As a third hypothesis, it was predicted
that the feedback expectancy would influence the magnitude of the ERPs associated with
the feedback, i.e. the FRN and oFRN. Another intention of this study was to validate a
new paradigm for possible use in a schizophrenia population.
5.2 Method
5.2.1. Participants
8 right-handed participants (4 male) were recruited in Bochum, Germany. The mean age
of participants was 25.2 years (SD = 2.7) and individuals with a history of neurological
damage or psychiatric illness were excluded. Informed consent was acquired from all
subjects before the experimental procedure began.

5. PROCESSING THE OUTCOMES OF OTHERS’ ACTIONS
53
5.2.2. Task design
A reinforcement learning game, based on a task from Frank and colleagues (2004), was
given to participants during the recording of EEG. In this task, two stimulus cues were
presented on screen and participants were required to choose one of the two stimulus
cues. The stimulus cues that were used were unfamiliar symbols (Chinese characters).
Feedback of monetary gain or loss was presented to participants shortly after a choice
was made. There were two parts to the task, one in which the association between the
response and the outcome was essentially random (i.e. 50% chance of receiving reward
and 50% receiving loss), inducing low expectation of the outcome and therefore did not
intend to involve any learning. This permitted all participants to receive the same
frequency of reward (financial gain). In the other part of the task, the association
between response and outcome was at 100% (i.e. the choice of one cue always led to a
reward and the choice of the other always led to a loss), and therefore inducing high
expectation and fast learning of the associations. The participants first played the game
(active part), and then observed a simulation of the game (passive part) in which they
passively observed the game being played by another person. Therefore, the game
consisted of 4 main parts: active high expectancy (100% association), active low
expectancy (50% association), passive high expectancy (100% association) and passive
low expectancy (50% association). The response choice pattern (i.e. the frequency of
choosing each cue in stimulus pairs) put into the simulation of the game for the 50%
part was used as a model for the choice behaviour of the observed performer, with the
previous participant’s choice behaviour being fed into the observed choice behaviour of
performer. The response choice pattern for the 100% part was a predetermined
pseudorandom choice pattern, with approximately 50% gains and 50% losses. An
illustration of the experimental design and stimuli used for the active part is shown in
figure 5.1, and for the passive part in figure 5.2.

5. PROCESSING THE OUTCOMES OF OTHERS’ ACTIONS
54
Figure 5.1. Illustration of experimental design and stimuli for active part, showing the potential outcomes
of either a gain or a loss.
Participants began each part of the game with 8 practice trials, which was followed by
200 experimental trials for each part of the game (i.e. 200 x 4). Each trial began with a
fixation cross (presented for a variable length of time: between 500 and 1000ms),
followed by stimulus presentation (up to 1500ms), then a red circle was displayed
around the choice made by the participant (300 ms), then a blank black screen (500ms)
and finally either positive (gain) or negative (loss) feedback (500ms).
The 50% part of the task presented one of three pairs of stimuli (e.g. A&B, C&D or E&F).
These three stimulus pairs were presented around 67 times each in this part in a
random order. In the 100% part, only one pair (e.g. G&H) of stimulus cues was used to
make learning of the associations faster and more stable. Different stimuli were used for
the active and passive parts of the game. The aim for the 50% part was to have an equal
number of gains and losses, and to control for individual difference in learning to allow
for a more reliable investigation of the FRN and observational FRN (oFRN). The aim of
the 100% part was to evoke an ERN and observational ERN (oERN) by increasing
certainty and therefore inducing known errors.

5. PROCESSING THE OUTCOMES OF OTHERS’ ACTIONS
55
Figure 5.2. Illustration of experimental design for observational / passive learning part, showing the
potential outcomes of either a gain or a loss..
5.2.3. Procedure
Participants were sat in front of the computer screen and were instructed to make the
appropriate responses by pressing either the left or right CTRL button on a keyboard
placed on the desk in front of them. They were told to respond as quickly as possible, but
also to try to win as much money as they could. If they did not respond fast enough, then
a warning screen was displayed (1500ms after onset of the stimulus cues) to ask
participants to respond faster. In the active part of the experiment, participants were
told that they would be playing a game in which they could win or lose money. They
were not told about the likelihood of their choices leading to gains or losses. The order
of the high and low expectancy conditions were counterbalanced across subjects. For
the passive part of the experiment, participants were instructed that they would observe
the responses from someone else playing the game, and that this person was a sat in
another room and the computers were connected via a network. Importantly, they were

5. PROCESSING THE OUTCOMES OF OTHERS’ ACTIONS
56
also told that the outcomes they observed from the other person’s choices would not
lead to any financial gains or losses for themselves. Trials in which responses were not
fast enough, and in which the warning screen was presented, were excluded from the
analysis.
5.2.4. EEG data acquisition
EEG was recorded at a sampling rate of 512 Hz from 32 channels with a Brain Products
BrainAmp system, using passive AgCl electrodes held in a BrainCap elasticated EEG cap
(from EasyCap). The 32 electrode positions were distributed over the scalp according to
the international 10-20 EEG system. An additional electrode was placed below the left
eye (EOG) to capture eye blinks. According to the EasyCap 32-channel standard
electrode configuration, the ground electrode was located along the midline, anterior to
electrode Fz. The ground electrode was located along the midline, more posteriorly,
between electrodes Cz and Fz. Following acquisition, the raw data were processed
offline with BrainVision Analyzer 2 (Brain Products GmbH). Firstly, the data was visually
inspected for segments with obvious artifacts that were then removed. The data was re-
referenced to all electrodes and submitted to a band-pass filter of 0.1 Hz to 30 Hz. Ocular
correction was performed with the EOG as a reference.
5.2.5. EEG data analysis
For the analysis of the feedback-related negativity (FRN & oFRN), data was segmented at
100 ms before the onset of the presentation of the feedback, to 600ms after the feedback
onset. For the analysis of the error-related negativity (ERN & oERN), data was
segmented at 100 ms before the onset of the response, to 600ms after the onset of the
response. These segments were baseline corrected using the 100ms epoch preceding the
event (feedback or response). The segmentation for the feedback and responses was
done independently for the four main conditions (active high expectancy (100%
association), active low expectancy (50% association), passive high expectancy (100%
association) and passive low expectancy (50% association)). Peak detection was used on
the feedback-locked segments to identify the FRN/oFRN, which defined the criteria as

5. PROCESSING THE OUTCOMES OF OTHERS’ ACTIONS
57
the negative maximal peak that was reached between the time window of 190-300ms
(as in Frank et al., 2005). The mean latency of the FRN was 284.8 ms (SD=14.5). For the
ERN in the active part, peak detection was used on the response-locked segments with
the criteria as the negative maximal peak that was reached between 40-160 ms (as
Morris et al., 2006). The ERN in the passive part (i.e. the oERN) appeared to be later than
the ERN produced in the active part, consistent with previous literature (van Schie et al.,
2004), and thus the criteria for peak detection used for the oERN in passive trials was
defined more liberally (i.e. 50ms to 200ms). The amplitude and latency of the FRN (only
Fz) and ERN (Fz, Cz and Pz) peaks was exported for statistical analysis. Following visual
inspection of the feedback-locked waveforms, it appeared that a P300 was also present,
and therefore, peak detection was used to identify the P300 as the most positive peak
reached between 250 and 450 ms. The amplitude and latency of the P300 was also
exported for statistical analysis.
A repeated-measures ANOVA (2x2x2) was used with the amplitudes of the FRN, with the
factors: active & passive, high & low expectancy, and wins & losses. The FRN exhibited a
fronto-central topographical distribution, with maximums being reached at electrodes
Fz and Cz, consistent with the previous literature (Baker & Holroyd, 2009; Holroyd &
Krigolson, 2007), although only Fz was used for the analysis of the FRN. For the ERN, a
repeated-measures ANOVA (2x2x3) was performed with the factors: active & passive,
wins & losses, and electrode (Fz, Cz and Pz), as the topographical distribution of the ERN
was less clear. Only the high expectancy trials (100% association) trials were used for
the analysis of the ERN, as the ERN is only generated when a known-error is made, and
therefore it would not be appropriate to use the low-expectancy trials as the
associations between stimulus cues and outcomes were random. Following this, the
exported P300 peak amplitudes were submitted to a repeated-measures ANOVA
(2x2x2x3), using the factors: active & passive, high & low expectancy, wins & losses, and
electrode (Fz, Cz and Pz). Post-hoc tests (with LSD) were also later performed on the
ERN and P300 amplitudes.

5. PROCESSING THE OUTCOMES OF OTHERS’ ACTIONS
58
5.3. Results
Figure 5.3 shows the feedback-locked waveforms for active and passive parts of the task
with a low expectancy (only 50% association trials) in electrodes Fz, Cz, and Pz. The
waveforms for both wins and losses are shown in figure 5.3. The FRN and P300 ERPs are
labeled in the top-left plot in the figure. No significant main effects were found for any of
the conditions for the analysis of the FRN amplitude, and therefore there was also no
significant main effect of active versus passive conditions on amplitude of the FRN.
The ANOVA conducted on the ERN amplitudes for trials with a high expectancy (only
100% association trials) revealed significant main effects of wins & losses (F(1,7)=9.16,
p=0.02), and a significant 2-way interaction between active/passive trials and
wins/losses (F(1,7)=12.48, p≤0.01). The 3-way interaction between active/passive
parts, wins/losses and electrodes was also found to be significant (F(2,6)=6.20, p=0.04).
Post-hoc comparisons of the ERN for the high expectancy trials were performed to
compare wins and losses for each electrode independently. Only significant differences
between wins and losses were found in electrodes Fz (t(7)=3.02, p=0.02) and Cz
(t(7)=3.59, p<0.01) for the active part of the task, but no significant differences were
found between wins and losses in any of the electrodes in the passive part. However, a
trend did seem to be appearing over electrode Cz for the observational ERN, whereby
responses that led to a win produced a larger ERN amplitude (i.e. more negative), as
compared to observed responses that led to a loss. Event-locked waveforms and ERN
amplitudes for the passive task are presented in figure 5.4.

5. PROCESSING THE OUTCOMES OF OTHERS’ ACTIONS
59
Figure 5.3. Feedback-locked grand-average event-related potentials (ERPs) for onset of positive (wins)
and negative (losses) feedback. This compares ERPs for the active and passive tasks in electrodes Fz, Cz
and Pz for the low expectancy (50% association) condition. The oFRN is indicated with the waveforms
shown as dotted lines, labeled in the top left plot.

5. PROCESSING THE OUTCOMES OF OTHERS’ ACTIONS
60
Figure 5.4. Event-locked grand-average event-related potentials (ERPs) for onset of response in the
passive condition in the 100% condition (high expectancy) in electrodes Fz, Cz and Pz. The oERN is
labeled on the waveform of electrode Cz. For responses that lead to positive feedback (wins), the
waveforms are in green and responses that lead to negative feedback (loss) are in red. Error bars
represent the standard error.
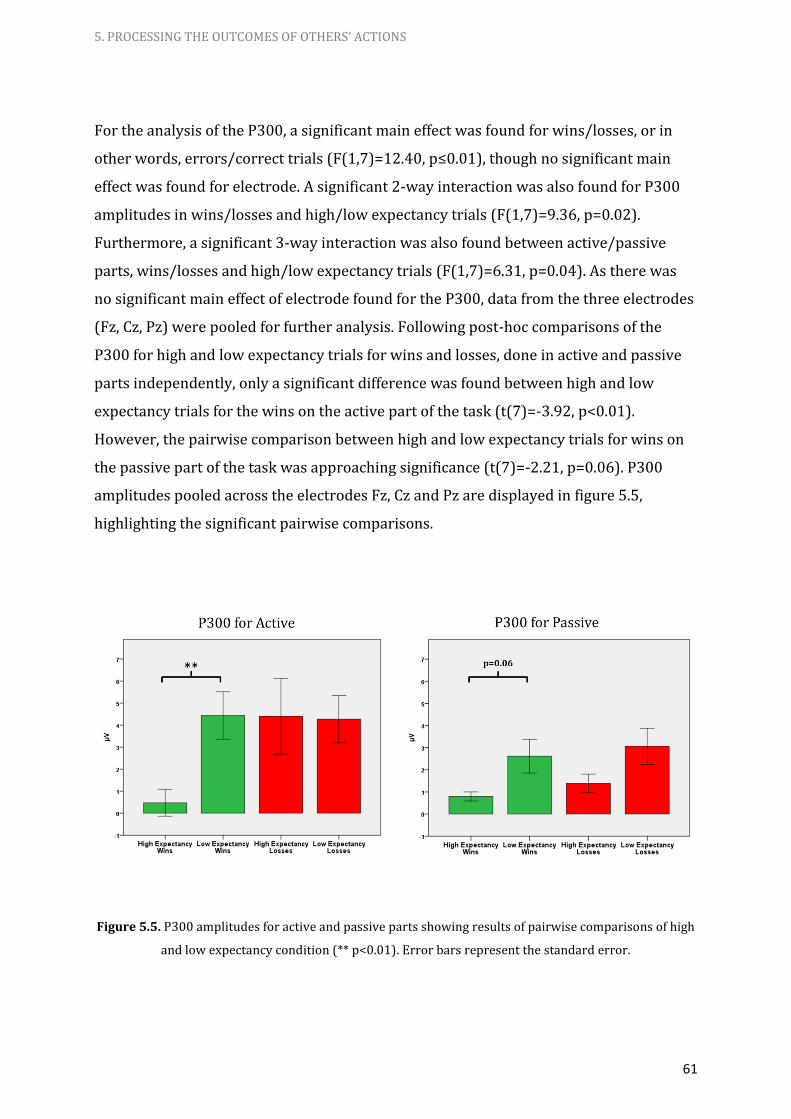
5. PROCESSING THE OUTCOMES OF OTHERS’ ACTIONS
61
For the analysis of the P300, a significant main effect was found for wins/losses, or in
other words, errors/correct trials (F(1,7)=12.40, p≤0.01), though no significant main
effect was found for electrode. A significant 2-way interaction was also found for P300
amplitudes in wins/losses and high/low expectancy trials (F(1,7)=9.36, p=0.02).
Furthermore, a significant 3-way interaction was also found between active/passive
parts, wins/losses and high/low expectancy trials (F(1,7)=6.31, p=0.04). As there was
no significant main effect of electrode found for the P300, data from the three electrodes
(Fz, Cz, Pz) were pooled for further analysis. Following post-hoc comparisons of the
P300 for high and low expectancy trials for wins and losses, done in active and passive
parts independently, only a significant difference was found between high and low
expectancy trials for the wins on the active part of the task (t(7)=-3.92, p<0.01).
However, the pairwise comparison between high and low expectancy trials for wins on
the passive part of the task was approaching significance (t(7)=-2.21, p=0.06). P300
amplitudes pooled across the electrodes Fz, Cz and Pz are displayed in figure 5.5,
highlighting the significant pairwise comparisons.
Figure 5.5. P300 amplitudes for active and passive parts showing results of pairwise comparisons of high
and low expectancy condition (** p<0.01). Error bars represent the standard error.

5. PROCESSING THE OUTCOMES OF OTHERS’ ACTIONS
62
5.4. Summary of results
The aim of this study was to explore the effects of feedback valence on the size of the
oFRN when observing others’ feedback, while also investigating the oERN produced as a
result of observing others’ errors, and the effect of high and low feedback expectancy on
the oFRN and oERN. In relation with the hypotheses, the main findings of the study
reveal that the FRN produced in response to the feedback for both one’s own and for
others’ performance was not significantly affected by the valence nor the expectancy of
the feedback. From inspection of the oERN during the observation of other’s responses,
it appears that a trend for valence-related modulation was appearing in which a greater
oERN was produced when observing the others’ responses that led to a positive
feedback. The most substantial finding comes from the effect of expectancy on a later
ERP associated with the feedback, namely the P300, which was greater for unexpected
than expected positive feedback (wins) in both active and passive parts of the
experiment. It also appears that the P300 was influenced by the interaction between
feedback expectancy and the valence of the feedback in the active condition.

6. SOCIAL APPROACH AND AVOIDANCE IN SCHIZOPHRENIA
63
Chapter 6
A possible neurobiological basis of social approach
and avoidance behaviour in schizophrenia
6.1. Introduction
Schizophrenia is a psychiatric illness diagnosed by the persistence of negative and
positive psychotic symptoms, but is also marked by abnormalities in social behaviour
and impaired social functioning. So far, the underlying social cognitive deficits
associated with social functioning have been discussed, though alongside these cognitive
deficits, abnormalities in the motivational drives underlying social interaction are also
prominent in schizophrenia. It has been hypothesised that many signs and symptoms
associated with schizophrenia can be understood in the context of dysregulated
approach and avoidance (i.e. fight-flight) behaviour (Brüne, 2008), an interpretation
that has received at least partial support through studies using ethological observation
of patients’ nonverbal behaviour (Annen et al., 2012; Geerts & Brüne, 2009). Much
recent work has also focused on the underlying neurobiological factors related to
dysfunctional social behaviour, with oxytocin being one popular neuropeptide for study.
A number of pathological conditions associated with dysfunctions in social behaviour,
have been found to also present differences in social approach and avoidance behaviour.
To the best of my knowledge, there have not been any previous studies exploring social
approach and avoidance with the AAT in a population with schizophrenia, although
there is a large body of work which has shown that people with schizophrenia have

6. SOCIAL APPROACH AND AVOIDANCE IN SCHIZOPHRENIA
64
abnormalities in emotional face processing (Tremeau, 2006). In addition, there have
been a few studies investigating more general social approach and avoidance behaviour
in schizophrenia, relative to positive and negative social stimuli. People with
schizophrenia seem to have a general tendency for avoiding negative social situations
(Liddle, 1987). Evans et al. (2011) presented participants with happy and angry faces in
an associative learning task and found that people with schizophrenia, when compared
to healthy controls, had a greater aversion to angry faces, even when choosing an angry
face led to positive feedback. Furthermore, some recent studies have explored
endogenous levels of oxytocin in schizophrenia, demonstrating negative correlations
between plasma oxytocin and negative symptoms such as social and emotional
withdrawal (Rubin et al, 2010; Sasayama et al., 2012). Much work is now being invested
into exploring the therapeutic value of oxytocin administration in improving social
functioning in schizophrenia (Feifel, 2012). However, most research investigating the
effects of oxytocin on social functioning have focused on social cognitive domains but
have not addressed the effect on social approach and avoidance behaviours, which are
also likely to have substantial if not more direct impact on social functioning. In addition,
little work has investigated individual differences in endogenous basal oxytocin levels
and the potential relationship with social approach and avoidance behaviour in clinical
populations, which would likely have substantial implications on patients’ response to
therapeutic interventions that seek to improve social functioning.
A joystick-based social approach-avoidance task (AAT) has previously been used to
explore motor responses to social approach and avoidance behaviour (Rinck & Becker,
2007), whereby subjects are required to either pull the joystick towards themselves, or
push it away when presented positive or negative emotional faces. By comparing
reaction times for arm flexion or extension movements to different emotional faces,
healthy people tend to have faster congruent responses (i.e. pull (/approach) happy and
push (/avoid) angry) than incongruent responses (i.e. pull-angry and push-happy). This
low-level difference seen in motor responses is thought to reflect associations between
social approach-avoidance behaviour and the processing of emotional valence. Figure
6.1 shows a pictorial representation of the congruent and incongruent conditions in the
AAT. Results from a study by Roelofs and colleagues (2010) are shown on the left-hand
side of the figure, demonstrating that reaction times are lower for the congruent
condition, and higher for the incongruent condition.
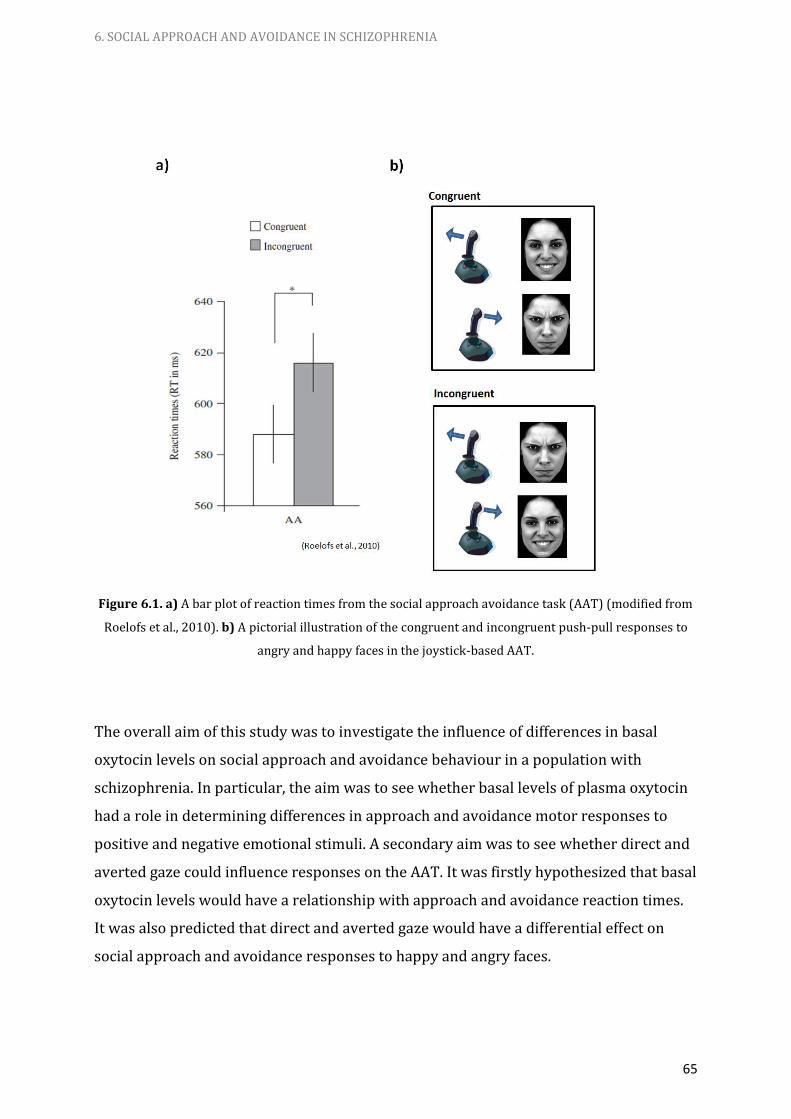
6. SOCIAL APPROACH AND AVOIDANCE IN SCHIZOPHRENIA
65
Figure 6.1. a) A bar plot of reaction times from the social approach avoidance task (AAT) (modified from
Roelofs et al., 2010). b) A pictorial illustration of the congruent and incongruent push-pull responses to
angry and happy faces in the joystick-based AAT.
The overall aim of this study was to investigate the influence of differences in basal
oxytocin levels on social approach and avoidance behaviour in a population with
schizophrenia. In particular, the aim was to see whether basal levels of plasma oxytocin
had a role in determining differences in approach and avoidance motor responses to
positive and negative emotional stimuli. A secondary aim was to see whether direct and
averted gaze could influence responses on the AAT. It was firstly hypothesized that basal
oxytocin levels would have a relationship with approach and avoidance reaction times.
It was also predicted that direct and averted gaze would have a differential effect on
social approach and avoidance responses to happy and angry faces.

6. SOCIAL APPROACH AND AVOIDANCE IN SCHIZOPHRENIA
66
6.2 Methods
6.2.1. Participants
28 right-handed participants (15 male) diagnosed with schizophrenia were recruited
from the Psychosis Unit of the Psychiatry Department at Celal Bayar Hospital University,
Turkey. Diagnosis was confirmed with the DSM-IV SCID and all participants were
remitted patients and considered as clinically stable. The mean age of the sample was
33.62 ±8.61 years and all participants were right-handed. Psychotic symptomatology of
all participants was assessed with the Positive and Negative Syndrome Scale (PANSS;
Kay et al., 1987). The PANSS is a 30-item semi-structured interview designed to assess
five symptom categories associated with schizophrenia: positive symptoms (i.e.,
hallucinations and delusions), negative symptoms (i.e., avolition and anhedonia),
cognitive symptoms (i.e., thought disorder), hostility, and depression. A qualified and
PANSS-trained psychiatrist assigned a score from 1 to 7 for each item, with higher
scores indicating more severe psychopathology.
6.2.2. Tasks and procedure
Self-report questionnaires and blood collection were completed before performance of
the AAT. All tests were performed between 11:00 a.m. and 3:00 p.m., because of the peak
pulsatile release of oxytocin (Amico et al 1983). Patients were asked to refrain from
eating or doing physical exercise 60 minutes before the start of the experiment.
6.2.2.1. The Approach- Avoidance Task (AAT)
The stimuli used in the AAT task were based on a previous set of photographs of faces
used in other AAT tasks (Roelofs et al., 2010). All photographs had been cropped to the
hairline and all were in black and white. Four different male and four female actors were
used with half of the faces expressing anger and the other half expressing happiness. The
eye gaze of the faces had been previously modified (as in Roelofs et al., 2010) resulting
in half of the faces having a direct gaze and the other half having an averted gaze. This
produced a total of 32 different stimuli (8 actors x 2 emotions x 2 gaze directions). The

6. SOCIAL APPROACH AND AVOIDANCE IN SCHIZOPHRENIA
67
task was structured in four blocks: two experimental blocks, and two practice blocks
preceding each experimental block. There were 24 trials in each practice block, making a
total of 48 practice trials. Each stimulus was presented 3 times in a pseudo-random
order in the experimental block making a total of 192 experimental trials, with 96 in
each block.
Figure 6.2. An illustration of the experimental design for the AAT showing the order of events in two
individual trials. (Modified from Roelofs et al., 2009)
Participants were seated in front of the computer screen and a Logitech Attack 3000
joystick was fixed to the table between the participant and the screen. The task was self-
paced so participants triggered the onset of stimulus presentation by pressing the fire
button on the joystick with the index finger of the right-hand while the joystick was in
the resting (central) position. A blank (black) screen was presented between each trial,
and at stimulus onset, pictures appeared in the centre of the screen. In one
practice/main block, participants were asked to push the joystick away from themselves
when they saw an angry face and pull the joystick towards themselves when they saw a
happy face (“congruent” condition). In the other practice/main block, participants were
asked to do the opposite and pull the joystick towards themselves whenever an angry
face was presented and to push the joystick away from themselves when a happy face
was presented (“incongruent” condition). The order of the congruent and incongruent
blocks was counterbalanced across participants. When the joystick was pushed during
stimulus presentation, the face would shrink, getting smaller and smaller till it
disappeared, and when the joystick was pulled, the face would zoom in until it
disappeared after the maximum size was reached. During the practice blocks, faces
shrank and zoomed appropriately, but did not disappear if the response was incorrect,
according to the instructions given at the start of the task, however, stimuli disappeared

6. SOCIAL APPROACH AND AVOIDANCE IN SCHIZOPHRENIA
68
after every response on all trials in the experimental blocks. Participants were asked to
respond as quickly as possible, but also to be as accurate as possible. Initiation response
times (RTs) (i.e. the first deviation from the central resting position of the joystick) were
recorded for all responses and taken as the RTs for all analyses.
6.2.2.2 The AAT effect scores
The AAT effect scores were calculated for mean RTs for each condition by subtracting
pull RTs from push RTs (i.e. push-pull) (as done by Roelofs et al., 2010). This resulted in
4 mean AAT effect scores for each participant for each of the different faces: happy
direct-gaze, happy averted-gaze, angry direct-gaze and angry averted-gaze. A more
negative AAT effect score reflects greater approach and a more negative score reflects
greater avoidance.
6.2.2.3. Facial emotion recognition and discrimination test
To control for deficits in emotion recognition, the Face Emotion Identification Task and
the Face Emotion Discrimination Task (FEIT & FEDT; Kerr and Neale, 1993) were given
to participants. The FEIT consists of 19 photographs of emotional faces presented on
screen, and participants are required to identify which emotion is being expressed in
each photograph. The FEDT presents 30 pairs of photographs of emotional faces and
requires the participant to decide if the two faces in each pair display the same or
different emotions.
6.2.2.4. State and trait anxiety scale
The State–Trait Anxiety Inventory (STAI; Spielberger et al., 1983) was used as a self-
reported measure of anxiety. For the 20 State items, patients were asked to answer with
the response that best describes “how they feel now” according to a 4-point scale: 1 (not
at all), 2 (somewhat), 3 (moderately so), and 4 (very much so). For the 20 Trait items,
patients were asked to answer with the response that best describes “how they
generally feel” according to a 4-point scale: 1 (almost never), 2 (sometimes), 3 (often),
and 4 (almost always).

6. SOCIAL APPROACH AND AVOIDANCE IN SCHIZOPHRENIA
69
6.2.2.5. Plasma oxytocin assessment
Basal oxytocin levels were measured by collecting blood samples from each participant
before the beginning of the experiment. Each sample was drawn into EDTA tubes that
contained the polypeptide aprontinin (EDTA-Aprotinin Tubes, Greiner Bio-One GmbH,
Germany). Shortly after collection, the samples were centrifuged at 4°C at 4000g for 20
min after which plasma was separated into two tubes. Plasma was stored in a freezer at -
80°C until the assessment day and assayed in duplicates. For the analyses, in
consideration of the debate on the plasma extraction procedure, we preferred to use a
novel commercially available extraction-free Elisa kit (Bachem S-1355 Oxytocin - EIA
Kit, Extraction-free CE-marked) in which the samples are incubated for one night. For
human serum or plasma samples, typical sensitivity (Av. IC50) was 0.15 ng/ml, with a
range of 0-10 ng/ml. The limit of detection was 1.2 pg/well and intra- and inter-assay
variability were 9% and 15% as reported by the manufacturers. . It is important to note
here that plasma oxytocin levels collected at a single time point are considered to reflect
a reliable and stable measure of baseline levels which may be used as an index of central
oxytocinergic activity (Amico et al., 1983; Bartz & Hollander, 2006; Pierrehumbert et al.,
2010).
6.2.3. Data analysis
Due to impaired neurocognition commonly seen in a patient group with schizophrenia,
many participants exhibited a slow processing speed during performance in the AAT.
Our RT data was skewed towards higher values and also demonstrated a high variance
between subjects when compared to previous studies. Therefore we used a Repeated
Generalized Linear Model (GLZ) analysis, an extension of repeated measures ANOVA,
which permits analysis on longitudinal data without the assumption of normality. We
selected a gamma with link function for RTs as suggested for the analysis of data skewed
towards higher values (Madsen & Thyregod, 2011). The use of this type of model
accounts for the flexibility on the range of allowable values for predictable functions,
which direct modelling of the response variable cannot address, and also permits
analysis of each trial instead of averaging the data by considering participants with
higher RTs as outliers (Madsen & Thyregod, 2011). In this analysis step we used the raw
RTs for each trial as the dependent variable and emotion (i.e. happy and angry),

6. SOCIAL APPROACH AND AVOIDANCE IN SCHIZOPHRENIA
70
response (i.e. push and pull) and gaze (i.e. direct and averted) as categorical factors and
basal oxytocin levels as a continuous factor. In accordance with the longitudinal nature
of this analysis, 4508 trials for 28 subjects were taken in this step.
To further elucidate the interactions seen between AAT performance and oxytocin
levels, we performed a post-hoc analysis by dividing the subjects into high and low basal
oxytocin level groups using a median split. A one-way ANOVA was used to compare low
and high oxytocin groups in terms of symptomatology with the PANSS positive, PANSS
negative and PANSS general scores. Groups were also compared on behavioural
measures of face recognition and discrimination, state and trait anxiety with respective
STAI scores, and lastly medication was compared between groups using the
chlorpromazine equivalent scores. An ANOVA was also used to compare groups on
demographic data of age, sex and chronicity (i.e. duration of illness). Following this,
group differences in the AAT effect scores were assessed using Mann-Whitney-U tests,
as suggested for data with high variance (Tabachnick & Fidell, 2006). Lastly, inter-
correlations were conducted with Spearman correlational analyses to check for the
consistency of the AAT effect scores across conditions. The same procedure was also
applied to explore the association between oxytocin and AAT effect scores.
6.3. Results
The total group mean for oxytocin levels was 271.1 pg/ml (SD=161.4). The low oxytocin
group exhibited a group mean of 161.1pg/ml (SD=36.6) and the high oxytocin group had
a mean of 381.1pg/ml (SD=163.4). An independent t-test shows that there is a
significant group difference in oxytocin levels between the groups (p≤0.001). Table 1
shows means and SDs for symptom severity, behavioural data and medication for the
low and high oxytocin groups, and the results of an ANOVA comparing groups on these
measures. This shows that there are no significant group differences in symptom
severity, face recognition, state and trait anxiety or medication. Importantly, there were
also no significant differences in age, sex or chronicity (duration of illness) between the
low and high oxytocin groups.

6. SOCIAL APPROACH AND AVOIDANCE IN SCHIZOPHRENIA
71
Low oxytocin group
High oxytocin group
p
Mean SD Mean SD
PANSS Positive 10.79 2.94 10.43 4.13 0.794 PANSS Negative 16.57 4.86 16.21 6.33 0.868 PANSS General 26.07 6.79 28.29 8.89 0.465 FEIT 11.64 2.79 11.92 3.23 0.811 FEDT 23.79 4.41 24.85 4.62 0.547 STAI Trait 48.64 5.97 49.23 7.34 0.821 STAI Sate 46.08 6.78 43.31 4.71 0.238 CPZ 357.14 194.00 514.29 369.21 0.171
Table 6.3: Table showing means and SDs for symptom severity, behavioural measures and medication
between low and high oxytocin groups. P-values show the result of an ANOVA comparing the two groups.
(PANSS= Positive and Negative Syndrome Scale, CLZ=Chlorpromazine equivalent, FEIT= Face Emotion Identification
Task, FEDT= Face Emotion Discrimination Task, STAI= State–Trait Anxiety Inventory)
In the Repeated Generalized Linear Model (GLZ) using the AAT raw RTs, we found
significant main effects of gaze direction (Wald-X2=4.90, p=0.02), response (Wald-
X2=15.13, p=≤0.001), basal oxytocin levels (Wald-X2=7.51, p=0.006). The model also
revealed significant 2-way interaction effects between emotion and response (Wald-
X2=18.33, p=≤0.001) and response and oxytocin level (Wald-X2=5.96, p=0.015), and a 3-
way interaction between gaze direction, emotion and oxytocin level (Wald-X2=22.94,
p=≤0.001). Most critically, the 4-way interaction between gaze direction, emotion,
response and oxytocin level was significant (Wald-X2=8.19, p=0.004).
To further investigate the 4-way interaction between gaze, emotion, response and
oxytocin, we compared the low and high basal oxytocin group on AAT effect scores in
each condition (displayed in Figure 1). Accordingly, there were significant differences
between groups for angry faces with averted gaze (z=-2.07, p=0.039), and for angry
faces with direct gaze (z=-2.11, p=0.035). However, there were no significant differences
between oxytocin groups in AAT effect scores for happy faces. Importantly, we found
that the group differences in AAT effect scores were independent of emotional face
processing skills as performance on the face discrimination (z=-1.22, p=0.22) and face
recognition (z=-0.47, p=0.64) tasks were not significantly different between groups.
Lastly, there were also no significant differences between low and high oxytocin groups
in terms of gender, symptomatology, medication and state/trait anxiety.

6. SOCIAL APPROACH AND AVOIDANCE IN SCHIZOPHRENIA
72
Figure 6.4: AAT effect scores reflecting push minus pull mean RTs for each condition, comparing RTs for
the low and high basal oxytocin groups. A more positive score reflects greater approach and a more
negative score reflects more avoidance. Error bars represent the standard error.
We found significant inter-correlations between happy straight and happy averted AAT
effect scores (rs (28)=0.51; p=0.006). Although, it appears that the correlation between
AAT effect scores for angry straight and angry averted was only approaching
significance (rs (28)=0.35; p=0.07). Furthermore we also found significant negative
correlations between AAT scores for angry-straight vs. happy-straight (rs (28)=-0.479,
p=0.01) and angry-straight vs. happy-averted (rs (28)=-0.413, p=0.03). Such relationship
was also seen in the same trend for angry averted although it was not reaching
significance for happy-straight (rs (28)=-0.256, p=0.18) or happy-averted (rs (28)=-
0.187, p=0.34). A correlation analysis confirmed the association between oxytocin and
the AAT response, as levels of oxytocin were negatively correlated with AAT effect
scores for angry faces with direct gaze (rs(28)= -0.396, p=0.04), as shown in Figure 2.
However, the correlation between basal oxytocin and AAT effect scores for angry faces
with an averted gaze did not reach significance (rs (28)=-0.188, p=0.35) although the
direction was still negative. There were also no significant correlations between AAT

6. SOCIAL APPROACH AND AVOIDANCE IN SCHIZOPHRENIA
73
effect scores and any of the factors of PANSS scores. Additionally, there were no
significant correlations between basal oxytocin levels and the subdomains of the PANSS
scale.
Figure 6.5: Scatter plot showing line of best to demonstrate the relationship between basal plasma
oxytocin levels and the AAT effect score for angry faces with a straight-gaze
6.4. Summary of results
The aim of this study was to investigate the role of basal oxytocin levels in approach and
avoidance responses to emotional pictures with schizophrenia patients. In particular,
the main hypothesis was that differences in endogenous oxytocin would have
differential effects on responses to angry and happy faces, which was evidently
confirmed. It was found that the difference in basal oxytocin levels was only associated
with responses to angry faces but not with happy faces. Further to this, the correlational
analyses showed that this effect was more strongly related to angry faces with a straight

6. SOCIAL APPROACH AND AVOIDANCE IN SCHIZOPHRENIA
74
gaze than to angry faces with an averted gaze. Importantly, these differences were found
to be independent from face discrimination and identification capacities of the patients.
In summary, these results demonstrate that only the patients with a higher level of
oxytocin demonstrate a greater aversion to negative emotions, as compared to positive
emotions.

7. GENERAL DISCUSSION
75
Chapter 7
General Discussion
7.1. Overall summary of findings
This thesis explored how different contextual manipulations can have an influence on
the processing of one’s own and of others’ actions. By looking at the EEG mu rhythm as
an index of motor resonance in the mirror system, it was demonstrated that by framing
observed actions in the context of self or other, or as being rewarding or punishing, this
modulated motor cortex activity to different degrees. Self-related observed actions, i.e.
actions that were associated with the self, led to greater motor cortex activity than
observed actions associated with the other. Though conversely, motor cortex activity
was lower for self-related performed actions. Observed actions that were rewarding for
participants generally led to the greatest degree of motor resonance when compared to
actions that did not lead to a reward. Despite previously suggested deficits in reward-
processing in schizophrenia, patients appeared to show a reward-related modulating
effect on motor resonance. In accordance with the proposal that a dysfunction of the
mirror neuron system occurs in schizophrenia, there did seem to be a trend for patients
with schizophrenia to have lower overall mu suppression when observing others’
actions as compared to the healthy control group, although this was not substantial
enough to reach statistical significance. Furthermore, a relationship between the degree
of motor resonance and empathy, specifically the perspective-taking dimension of
empathy, was found in healthy control participants, but not in schizophrenia patients.
Although a notable relationship between the mu rhythm suppression and negative

7. GENERAL DISCUSSION
76
psychotic symptoms was found in patients with schizophrenia, whereby more severe
negative symptoms were associated with greater mu suppression. In contrast to some
previous studies, differences in the perspective in which the actions were viewed did not
affect the degree of motor resonance in any of the experiments. In an associative
learning task, the reward-related event-related potentials (ERPs) in EEG when
observing others’ gains, losses and errors appeared to be influenced most substantially
by the degree of expectation of the possible outcomes of one’s own and of the others’
actions. The effect of feedback expectancy on reward-related ERPs also interacted with
the reward-related valence of the outcomes. Having a high expectancy for positive
feedback for one’s own and others’ actions produced a weaker ERP (the P300) as
compared to having low expectations for a positive feedback. However, unexpectedly,
the observational feedback-related negativity (FRN) was not modulated by the valence
(gain or loss) of the feedback given. Though one other interesting finding came from a
trend that was beginning to emerge when observing others’ errors, demonstrating a
smaller ERN for observed responses that led to negative feedback (i.e. errors), which is
the opposite of the usual pattern seen for ERN amplitude in self-generated errors. By
investigating social approach and avoidance behaviours in schizophrenia, it was found
that endogenous oxytocin levels had some relationship with patients’ responses to
positive and negative social emotional stimuli. More specifically, oxytocin was
associated with social approach-avoidance responses to angry faces. It seems that
patients with schizophrenia and higher levels of endogenous oxytocin exhibited greater
aversion to negative emotions, in comparison to patients with lower levels of oxytocin.
The interpretation of these findings and the implications of contextual modulations of
one’s own and of others’ actions, and the possible underlying neurobiological
mechanisms, will now be discussed in more detail.

7. GENERAL DISCUSSION
77
7.2. Detailed discussion
7.2.1. Self and other in the mirror motor system
The modulation of the mu rhythm suppression by framing an executed and observed
action in terms of self and other is shown here from the first study. Though
interestingly, the pattern of modulation of the mu rhythm suppression in terms of self
and other were the opposite for executed and observed actions. When looking at the
experimental design of the study, it is clear that the action kinematics of the “self” and
“other” actions in both the action execution and action observation parts were quite
different, as the bowls in which the objects were transferred to were positioned at
different distances from the participant and the performer. This may have had an
influence specifically on the action execution part, particularly as the “other” bowl in the
action execution part was further away from the participant and therefore required a
further reach than the “self” condition. This further reach required for the “other” bowl
may have been an explanation for the greater mu suppression seen in the “other”
condition during action execution, as greater mu suppression reflects greater motor
cortex excitability. Most notably, the modulation of the mu rhythm in reaction to self and
other during the action observation part of the experiment supported the originally
proposed hypothesis, in that observed actions that were relevant to the “self” induced
greater mu suppression. This finding is quite intuitive as it makes sense to engage more
with others’ actions that are relevant to oneself. One explanation for the greater mu
rhythm suppression seen in the “self” condition during action observation may have also
been due to the difference in eye-gaze between conditions, as the performer looked
towards the camera during the “self“ condition but not in the “other” condition. This
direct eye gaze may have led to greater attentional capture, as many studies in the past
have used eye-gaze to direct participants’ focus of attention (e.g. Friesen & Kingstone,
1998). Therefore, this greater attentional capture in the “self” condition may have
resulted in the participant just simply paying more attention to these actions, and
consequently would have been less engaged in actions for the “other”. This finding
highlights the importance of eye-gaze in social interactions and more specifically, the
influence of eye-gaze on motor resonance and MNS activity. Direct eye-gaze may
produce greater motor resonance, and thus, observed actions may be simulated to a

7. GENERAL DISCUSSION
78
greater degree when the person being observed is looking directly at the observer. Eye-
gaze is a highly salient stimulus that plays a central role in social interaction (Kendon,
1967), and the understanding of communicative intention, which provides further
support to this suggestion (John & Mervis, 2010).
Other environmental contextual influences could modulate the degree to which one
determines the relevance of others’ actions to oneself. For example, the self-relevance of
observed actions may be determined by the relationship between the observer and the
person performing the action. One may have more of a bias to think that a friend’s
actions are referring to oneself, when compared to observing the actions of a stranger. In
line with the findings from this study, it appears that the degree of similarity between
observer and performer can influence the activation of the MNS. This is demonstrated by
studies showing differences in the activation of the mirror neuron network when
observing actions of members of an in-group versus observing actions from an out-
group member (Sobhani et al., 2012).
The link between the mu rhythm and the mirror neuron hypothesis originally stemmed
from the finding that suppression of the mu rhythm occurs when one performs an
action, observes an action, imagines an action, hears action-related sounds and sees
objects that have some motor affordance (Pineda, 2005). The functional relevance of the
mu rhythm suppression on interpersonal processes has been further confirmed by
studies revealing different social contextual modulations (Perry et al., 2010a). The
findings from the study presented in this thesis provide further insight into the potential
uses of the mu rhythm suppression as an index of a motor-related process that may have
some specialized function for social interaction. These findings may also have
implications in psychopathology, as the tendency to associate others’ actions to oneself
or to the other could also be determined by a predisposed cognitive bias, and even a
pathological bias caused by a psychiatric disorder, such as schizophrenia. Schizophrenia
can often present with delusions of reference, in which patients believe that unrelated
and irrelevant events and behaviours refer directly to them and have some special
personal significance, and therefore will also consider irrelevant actions as being
relevant to oneself. People with schizophrenia also tend to have attributional biases,
which are associated with paranoid ideations (Bentall et al., 1994), in which there is a
pathological bias for attributing the cause of negative events to other people, often

7. GENERAL DISCUSSION
79
resulting in delusions of persecution. The contextual framing of observed actions,
particularly in terms of the relevance to “self” and “other”, may be dysfunctional in
schizophrenia and may therefore help in explaining such dissociative psychotic
symptoms that are grounded in the understanding of others’ actions, via MNS activity.
The limitations that were revealed from this study helped to inform the design of the
forthcoming studies looking at the mu rhythm suppression, as described here in this
thesis. In particular, the potential confounding variable of eye gaze was removed from
later study designs by not showing the head of the performer in the stimuli presented to
participants. In addition, the potential confounding effect of the different kinematics of
actions in different conditions was also later controlled for by having the actions for
different conditions using essentially the same kinematics, in the same plane of
movement, reach distance and spatial configuration.
7.2.2. Reward and punishment in the motor mirror system
The outcome of the study looking at the influence of reward and punishment on the mu
rhythm has implications on a wide range of themes in social cognition and also raises a
number of methodological and theoretical considerations for future research on the mu
suppression. As already pointed out, the influence of reward and punishment on
observational learning has been primarily investigated at the behavioural level; and thus
this study is the first providing neuroscientific evidence for this link. The results suggest
that action understanding, and thus observational learning, may be driven by the
associations made between rewards, punishments and the observed actions, due to
differences in motor resonance and MNS activity.
In light of these results, we suggest that some previous studies investigating the
contextual differences in mu rhythm suppression and mirror neuron-related activity
could have been influenced by uncontrolled effects of the varying reward value
associated with the observed actions across different experimental conditions. In the
social domain, studies comparing social and non-social stimuli (Pineda & Hecht, 2009),
with the aim of highlighting the social relevance of the MNS, could be accounted for by
the intrinsic reward associated with social stimuli and social interaction, as opposed to

7. GENERAL DISCUSSION
80
stimuli absent of any social meaning or value, which may be inherently less rewarding.
In other words, a social interaction in itself may be rewarding for the observer, as
already proposed by some authors (Krach et al., 2010). The reward value attributed to
observed actions can also depend upon the relationship between confederates in a social
interaction, such as the differences created by in-group and out-group membership
(Gutsell & Inzlicht, 2012), and competitive vs. cooperative scenarios (Koban et al., 2010).
Different social contexts will induce different reward values associated with others’
actions, which could be driven by a wide variety of personal and interpersonal
motivational and environmental factors. The degree of mu suppression produced when
observing actions is also enhanced when the observed action inflicts pain on the
performer (Perry et al., 2010a), which could be comparable to the effect of punishment
in the study described in this thesis. The reward value that is associated with the
observed action, or the consequence of the observed action, could modulate the degree
to which one empathises with others and shares the others’ intentions or concerns.
Regarding the lack of an effect of perspective in this study, the findings in the literature
are rather inconsistent (Alaerts et al., 2009 Maeda et al., 2002). Some studies also found
some lateralisation in the expression of the mu rhythm, with greater mu suppression in
one hemisphere than the other (Stancak & Pfurtscheller, 1996). Although this
lateralisation in terms of the motor cortex activity is likely to be affected by how
participants simulated or transformed the action onto their own motor cortex. There is
mixed evidence to show that some people have a greater tendency to mirror the other
persons’ actions onto their own motor cortex, in which case, one would see greater
activity on the contralateral motor cortex (i.e. contralateral to the observed action). On
the other hand, other people may have more of a tendency to do a mental rotation of the
observed action when simulating or transforming the observed action onto their own
motor cortex, in which case, one would see greater motor activity in the ipsilateral
motor cortex. If there were individual differences in the degree to which participants
tended to perceive the mirrored action or the mentally rotated action, then the effect of
perspective would have been lost after averaging.
The frontal suppression seen in the topographical representation of the alpha power has
also been shown by some studies looking at the mu rhythm (Cochin et al., 1999; Perry et
al., 2010b). However, as the work on the mu rhythm is focused on the activity over the

7. GENERAL DISCUSSION
81
sensorimotor cortex, this analysis is often done only to check for the potential overlap of
frontal alpha suppression with suppression over the motor areas, and therefore to
confirm that the effect seen is not driven by this frontal alpha. A possible explanation for
seeing this clear frontal alpha suppression might be related to studies suggesting
that human mirror neuron areas are thought to reside in the premotor area and inferior
frontal gyrus (Cattaneo & Rizzolatti, 2009). Since frontal electrodes are placed over
these areas, it might be the case that the frontal suppression reflects activity in other
parts of the human MNS. Alternatively, a more widespread suppression might appear,
depending upon the stimuli and the task, and therefore the widespread suppression
might reflect a wider network that also includes the MNS as well as other mechanisms /
systems. Although this is speculative and would require clarification from other studies,
possibly multimodal neuroimaging studies combining EEG and fMRI.
Motor acts and social interactions are dynamic processes. According to the results from
this study, contextual online changes in activity in the observer’s motor cortex are likely
to reflect associated contextual changes during the temporal dynamics of the observed
action. Schuch and colleagues (2010) were one of the first to look at the dynamic
temporal changes in the mu suppression over the course of the whole action. From their
results, it appears that there is only one significant suppression of the mu rhythm,
occurring at the time that the action-related object is presented on screen. This initial
suppression may be an index of an anticipatory or preparatory motor response to the
forthcoming action, as likely also reflected in the results presented here, due to the
expectation or anticipation of the forthcoming action in each trial. The second
suppression seen in the results of the study in this thesis provides evidence for an
independent suppression component, produced by the context or outcome of the action.
The delayed mu rhythm suppression found for punishing actions is a new and puzzling
finding. This delayed response could be a reflection of some kind of aversion to the
negative consequences of others’ actions, in which there may be active inhibition of
motor cortex consequently causing a slower return to baseline. Hence, our findings
show that mirrored motor activity during action observation does not correspond just to
a single or unique motor resonance response, but also shows differential effects in the
neurophysiological time-course and expression that may depend upon situational
changes in affective or motivational factors. This reveals a more fine-grained temporal
dynamic for the mu suppression than previously thought. These findings highlight the

7. GENERAL DISCUSSION
82
importance of looking at the online dynamic changes in neurophysiology over time, as a
more ecologically valid approach to study social interaction, and to gain further insight
into how our brains respond to the dynamic contextual changes in our environment.
7.2.3. Reward-related modulation of the mirror motor system in schizophrenia
According to the study presented here comparing mu rhythm suppression in patients
with schizophrenia and a matched healthy control group, a trend emerged in which
patients appeared to exhibit less mu rhythm suppression, even though this result did not
reach significance. This apparent trend is in line with the proposed hypothesis, although
the lack of significance suggests that this difference may not be as substantial as
originally predicted. In comparison to the two other known studies done looking at mu
suppression in patients with schizophrenia, these results seem to be more in line with
the findings from Singh and colleagues (2012), who also found reduced mu suppression
in patients. However, as already mentioned, the study from Singh and others may have
been confounded by the nature of the stimuli used. Group differences in their study were
only found on the condition that presented point-light biological motion displays, and
not with videos of live hand actions or full-body actions, and thus the diminished mu
suppression may have been a result of an information processing deficit in
schizophrenia, rather than a difference in the processing of others’ actions.
The action ratings for both patients and healthy controls confirm the subjective
experiences of rewarding, punishing and neutral actions as the valence of the ratings
were relatively consistent with the intended valence of the reward manipulation. No
significant differences on the action ratings were found between patients and healthy
controls, and both groups rated rewarding actions as more pleasant than punishing
actions. In terms of the mu suppression and reward manipulation, it is evident in the
findings that patients also showed a reward-related modulation of motor cortex activity
during action observation. This effect seems to be comparable to the reward-related
modulation seen in the earlier study in a healthy population, but however, does differ
from the pattern of reward-related modulation seen in the matched healthy control
group recruited as a comparison group in this study. The finding that the patient group
had the greatest mu suppression for rewarding actions suggests that schizophrenia may

7. GENERAL DISCUSSION
83
be accompanied by a deficit in reward-processing that is not generalized, but may
instead be a more selective deficit, as already suggested by studies looking at
reinforcement learning in schizophrenia (Gold et al., 2008). This is also supported by the
finding that there was no substantial difference in mu suppression between rewarding
and punishing actions in patients, whereas the finding from the earlier study did
demonstrate a difference between rewarding and punishing actions. Therefore, there is
likely to be at least some sensitivity to reward in schizophrenia, but this sensitivity may
not be as specific to the valence of reward, i.e. in terms of the difference between reward
and punishment, as compared to healthy controls. It may also be the case that the
salience of the reward and punishment in schizophrenia may be lower than in healthy
people, and therefore resulting in the apparently similar response to both rewarding
and punishing actions.
The puzzling finding that the matched healthy control group in this study demonstrated
the greatest mu suppression for punishing actions is not consistent with the findings
from the earlier study with a different healthy population. One explanation for this
conflict in the results for the two healthy groups in the earlier and later study may be the
differences in the demographics of these two samples. The average age of the healthy
sample in the earlier study, included in chapter 3, was much lower than that of the
matched healthy control group used as a comparison to the schizophrenia group. This
difference in age may have had a confounding effect on the individual’s subjective value
associated with the financial gains and losses acquired in the experiment. The younger
healthy sample in the earlier study was comprised mostly of students, who were most
likely to have had less money than the older subjects in the later study, and
consequently this could have resulted in a dampened effect of the reward-related
modulation of the mu suppression in the older group. In addition, according to the
action ratings, it does appear that the older sample rated neutral actions more positively
than the younger sample, which may have been a further indication of the dampening of
the effects, or lower subjective value, for financial gains and losses in the experiment.
The association found between the mu rhythm results and empathy provides an
interesting reflection on the relevance of the mu suppression in social cognition. The
relatively strong correlation between perspective-taking and mu suppression in the
healthy group confirms findings from previous studies that also found a relationship

7. GENERAL DISCUSSION
84
between empathy and the mu rhythm (Woodruff et al., 2011). This therefore adds
further support for a link between MNS activity and empathetic ability. The lack of
association between the mu rhythm and empathy scores in the patient group implies
that this relationship may not be present in the schizophrenia group. However, patients
with schizophrenia are known to have problems with insight and self-awareness,
particularly with self-assessment of empathetic ability (Harvey et al., 2013), and
therefore, this may have led to inaccurate self-report scores. If this was the case, then a
statistical relationship between the mu rhythm and self-reported empathy scores would
not be expected.
Possibly the most interesting finding of this study came from the association found
between the degree of mu suppression and the severity of psychotic negative symptoms
in the patient group. Patients with more severe negative symptoms seemed to exhibit
greater mu suppression. The association between negative symptoms and mu
suppression in patients is in line with suggestion made by Pridmore and colleagues
(2008), in that the presence of catatonic symptoms in schizophrenia leads to greater
motor cortex excitability, and therefore a pathologically enhanced mirroring effect. This
would usually be exhibited by symptoms such as echopraxia, being an over-activation of
imitative behaviour, but may not present in patients these days as modern antipsychotic
medication is now more effective in diminishing the overt signs of these catatonic
symptoms, and therefore making them less obvious. However, catatonic symptoms are
generally captured in the negative symptoms subscale of the PANSS, therefore implying
that this hypothesis may hold some weight. Though, unfortunately, it was not able to
have a comprehensive assessment of catatonic symptoms in this study to test such a
hypothesis. The association found in the study presented here could be comparable to
the findings from McCormick and colleagues (2012), who showed that schizophrenia
patients who were actively psychotic exhibited greater mu suppression than patients
who only had residual symptoms, and healthy controls. However, they found that the
association was not specific to negative symptoms, but was more associated with
positive symptoms, and more specifically, with the global hallucinations subscale. The
study from Singh and colleagues (2011) also found a correlation between mu
suppression and negative symptoms, although this was in the opposite direction to the
correlation found in the study presented here, as more severe negative symptoms were
associated with lower mu suppression. This conflict is a curious one, though could be

7. GENERAL DISCUSSION
85
accounted for by the difference in patient populations used, as the study from Singh and
colleagues used only patients that were in the early stages of the illness, i.e. first-episode
patients, whereas the patients used in the study presented here were mostly people who
had suffered from schizophrenia for several years.
According to the previous studies looking at mu suppression in schizophrenia patients,
and the study presented in this thesis, there does seem to be some apparent relationship
between the severity of symptoms in schizophrenia and the magnitude of mu
suppression. This adds further weight to the hypothesis that impairments in the MNS
may be contributing to the pathology of schizophrenia, and most likely also impacting on
the skills required for social functioning. The findings in this thesis also point to a role of
reward in the MNS and in motor resonance, while also highlighting the relevance of
contextual modulations of the mu rhythm on social cognitive skills including empathy
and perspective-taking, and how this relates to the psychopathology of schizophrenia.
7.2.4. Sharing others’ errors, rewards and losses
The main hypothesis of the study looking at observational learning with the
observational feedback-related negativity (oFRN) and observational error-related
negativity (oERN) was not supported by the findings. No effect of feedback valence on
the amplitude of the FRN was found for both the active and passive (i.e. observational)
parts. There is a substantial amount of evidence from previous work that shows that the
FRN generally distinguishes between gains and losses, as well as positive and negative
feedback without any personal gain or loss (Gehring and Willoughby, 2002; Holroyd and
Coles, 2002). Usually, the FRN is more pronounced for negative feedback than for
positive feedback, which includes the responses to financial loss and gain, respectively.
There may have been a number of factors in this study that led to a lack of valence
related modulation of the FRN. A recent study from Bismark and colleagues (2013)
revealed that a FRN was only produced when there was sufficient time for an
expectation for the valence of the feedback to be developed. This is also in line with the
proposal made by Holroyd & Coles (2002) that the FRN reflects the size of a dopamine-
driven teaching signal that may be representative of the (negative) prediction error, and

7. GENERAL DISCUSSION
86
therefore, without expectation or a prediction, the prediction error would not vary for
wins and losses. In the case of the trials that were used for the FRN analysis in this study,
a random association between the stimulus cue and feedback (~50%) was used to
control for individual differences in learning rate, and consequently also to control for
the potential variance in the number of wins and losses across participants. Therefore,
there should not have been any learning involved in this 50% association condition, and
thus, any possible patterns of expectation for the valence of feedback should have been
cancelled out when averaging across trials. This may provide one explanation for why a
valence-related modulation of the FRN may not have occurred in this data. Though
notably, when looking at the waveform of the ERPs produced in relation to the feedback,
it does appear that a clear FRN-like wave (i.e. similar timing and amplitude as previous
reports of the FRN) was produced for both active and passive parts of the task. This
confirms the presence of an oFRN when observing the outcomes of others’ actions. Yu
and Zhou (2006) were the first to find similar FRN amplitudes in both an active and
passive associative learning task, which is also apparent from the findings of the FRN
produced in the study presented here. This has also been shown by more recent studies
from Itagaki & Katayama (2008) and Fukushima & Hiraki (2009).
The ERN and FRN are both thought to originate from the ACC in the medial frontal
cortex (MFC), and the dorsal ACC has also been extensively linked with response conflict
(Kim et al., 2011). Ullsperger and colleagues (2007) demonstrated that the posterior
MFC is sensitive to both internally and externally generated errors. A seminal study
from van Schie and colleagues (2004) was the first to demonstrate an oERN component
produced when observing others’ errors. They showed that the oERN was modulated by
the correctness of the response, in that observing incorrect responses, or errors, led to a
larger oERN (i.e. more negative amplitude), as compared to observing correct responses.
The magnitude of the oERN has also been shown to be modulated by the interpersonal
context of the observed responses. In particular, a few studies have demonstrated that
observed responses in competitive and cooperative, or neutral scenarios lead to
opposite patterns in terms of the size of the ERN response when observing others’
correct and erroneous responses (Koban et al., 2010; van Meel et al., 2010). In a
competitive scenario, a smaller oERN is produced when observing others’ error, as
compared to the oERN when observing a correct response, whereas in a cooperative or
neutral situation, the oERN is larger for others’ errors. In other words, in a cooperative

7. GENERAL DISCUSSION
87
or neutral scenario, the oERN response pattern for observed responses is more similar
to the pattern seen when one is making correct or erroneous responses themselves.
According to the results presented in this thesis, there was a trend appearing for this
oERN to be larger for trials in which positive feedback was expected, i.e. wins for the
other. Importantly, the participants were told that the outcomes of the observed
responses would not lead to any financial gain or loss for themselves. So it seems to be
the case that observed responses that were expected to lead to a win for the other
induced a modulation of the oERN that follows the same pattern as the oERN previously
seen in competitive scenarios. Seeing a larger oERN when observing responses that led
to a win, as compared to the oERN seen for observed responses that led to a loss, may
have reflected some form of “Schadenfreude” in the observer, in which they were
pleased to see the other person losing money. So in this case, observing the other
person’s winning responses was more like experiencing an error. This finding could also
relate to Festinger’s (1954) theory of social comparison, which states that humans are
driven to evaluate themselves by assessing their abilities in comparison to others.
Therefore, the apparent Schadenfreude expressed in the pattern of the oERN response
may have been naturally induced because both the participant and the person being
observed were in with a chance of winning money, and thus the participant assessed the
other person’s financial gains and losses only in relation to their own gains and losses.
This finding may have therefore been different if the context of the observed responses
were outside of the context of one’s own wins and losses, i.e. if participants only
performed the observational part and not the active part.
The effect of feedback expectancy in the results presented here seems to provide the
most substantial effect on the ERPs. Only after visual inspection of the feedback-locked
waveforms was the analysis of the P300 considered, as the investigation of this ERP was
not included in the original hypothesis for this study. In general, larger amplitudes are
produced for the P300 when outcomes or events are highly unexpected (Polich &
Donchin 1988; Picton, 1992), which is also evident from the findings presented here.
The P300 effects also seem to be independent of the FRN, as no significant effect of
expectancy or feedback valence was found for the FRN, as also demonstrated by Kobza
and colleagues (2011). Yeung and Sanfey (2004) argue that the FRN and P300 reflect
different aspects of outcome evaluation, suggesting that the FRN is more sensitive to
feedback valence whereas the P300 encodes the magnitude of reward, but only when

7. GENERAL DISCUSSION
88
the magnitude of reward is defined in terms of a higher level motivational / affective
evaluation. This is a controversial finding that is still under debate, but this may relate to
the findings presented here, as financial gain or loss were the indicators of positive and
negative feedback, and thus would not necessarily be considered as high-level
motivational or affective rewards. Therefore this may also help to explain the apparent
lack of a consistent valence-related pattern of modulation of the P300 when looking
more closely at the pairwise comparisons across the active and passive parts
independently.
In sum, the findings from the oERN reveal that the evaluation of the outcomes of others’
actions may be determined by the context in which they are presented, and in this case,
may be influenced by how the outcomes of others’ actions relate to the outcomes of
one’s own actions. This study is the first to reveal valence-related modulation of the
P300 during the observation of others’ feedback. These results also provide further
insight into the modulation of the P300 by feedback expectancy and valence and how
this effect may be independent from the modulation of the FRN and ERN in both active
and observational learning. It may be the case that the P300 has a more substantial role
in evaluating feedback expectancy than in the valuation of reward or feedback valence.
These results add support to the suggestion that the modulation of the P300 is likely to
reflect a separate mechanism from that involved in the evaluation of outcome as
represented by the FRN. The relationship between the active and observational
components of the FRN, ERN and P300 provide an interesting platform to explore the
processing of reward valence, magnitude and expectation in psychiatric populations that
exhibit differences in reinforcement learning. Schizophrenia would be a particularly
interesting candidate for investigation due to the known dysfunction in the dopamine
system, which could influence social observational learning, and also consequently help
in contributing to psychotic negative symptoms.

7. GENERAL DISCUSSION
89
7.2.5. Social approach and avoidance and oxytocin in schizophrenia
The findings from the study investigating social approach-avoidance behaviour and its
associations with endogenous oxytocin levels confirm previous findings that patients
with schizophrenia demonstrate a greater aversion to negative emotions (Evans et al.,
2011). Though crucially, this effect was only apparent in the subgroup of patients that
had a higher level of basal oxytocin. Therefore it seems to be the case that oxytocin is
playing a fundamental role in the neurophysiological response to negative emotions, but
not positive ones. Averbeck and colleagues (2011) found that oxytocin administration in
patients with schizophrenia has the potential for improving emotion recognition,
although the specificity of the effects of oxytocin on responses to stimuli of different
emotional valences is not completely clear (Bartz et al., 2010). Many studies have found
emotion processing deficits in schizophrenia (e.g. Gur et al., 2002; Schneider et al.,
1995). However, as there were no group differences on emotion recognition and
discrimination in high and low oxytocin groups, the difference in approach and
avoidance responses do not seem to be related to a deficit in emotion processing in
general, but instead reflect a specific response that is determined by the valence of the
emotion presented. This confirms the social implications of oxytocin as a neuropeptide
that may be driving valence specific differences in social approach and avoidance
behaviour that is specific to whether the emotion is positive or negative. In particular,
this study shows that basal levels of oxytocin may be specifically affecting avoidance
behaviour towards threatening social cues in this schizophrenia population.
The main finding that the influence of basal oxytocin was only associated with angry
faces and not happy is an intriguing one. Here two possible explanations are proposed.
Firstly, it may be the case that responses to happy faces in the AAT may actually reflect
quiescence rather than approach, as shown by previous studies (Heinrichs & Domes,
2008). Secondly, there is also evidence that dopaminergic activity, in the ventral
striatum, may be driving positive affective motivational approach states (Burgdorf &
Panksepp, 2006). Considering the known abnormalities in the dopaminergic system in
schizophrenia, this may be disrupting approach motivations, and therefore subsequently
disturbing the congruent approach response seen here in the AAT, which is specific to
approach of positive emotions. In addition to this, the group differences that are seen
between the low and high basal oxytocin groups may be influenced by the possible

7. GENERAL DISCUSSION
90
interactions between the dopamine system and endogenous oxytocin (Baskerville &
Douglas, 2010).
The correlation found between the AAT response for angry faces with a direct gaze, and
not angry faces with an averted gaze supports previous suggestions that direct gaze has
a more threatening, and potentially avoidance inducing effect when presented with
negative emotional stimuli and social threat cues (Adams & Kleck, 2005). The fact that a
significant difference was seen between AAT response and gaze direction contradicts
previous suggestions of active avoidance of attending to the eye regions of faces in
schizophrenia (Morris et al., 2009). However, according to the data, it is suggested that
these previous findings may have be driven by within-group variation in basal oxytocin,
particularly as it has been shown that oxytocin administration can increase gaze
towards the eye region (Guastella et al., 2008). Therefore it may be the case that
individuals with a higher level of basal oxytocin may be paying more attention to the
eyes of faces, and consequently would be more sensitive to social cues.
In light of these findings, it is informative to first look at the effect of
neuroendocrinological factors underlying social behaviour at a low-level of processing,
by investigating the neurophysiological effects of oxytocin on social affective and
motivational systems, and then to look at how these effects may be translated
downstream, through neural activity, and inevitably into social behaviour in
schizophrenia. This adds another perspective on the investigation of social cognitive
deficits in schizophrenia, by looking at the pathways from biology to behaviour, which
complements lines of inquiry that investigate behaviour and then look for underlying
biological driving factors.

7. GENERAL DISCUSSION
91
7.3. General conclusions and implications of findings
In conclusion to the studies investigating the mu suppression in this thesis, it is evident
that the modulation of the mu rhythm has much potential in exploring the social
contextual factors affecting motor resonance and motor MNS activity. The socially-
relevant modulations demonstrated in these studies also add further support to the
relevance of the mu rhythm suppression in social cognition. There is much evidence
showing that activity in the motor-related areas of the MNS can be affected by the
context and “meaning” of the observed action, and here it is proposed that some of these
previous findings could be accounted for by underlying differences in the contextual
framing of the action, particularly in terms of the self-relevance and subjective value
associated with the seen actions. For example, studies demonstrating modulation of mu
suppression during the observation of painful versus non-painful actions (Cheng et al.,
2008; Perry et al., 2010a) provide one clue to the influence of reward on the MNS,
particularly as pain and reward-processing are closely linked (Leknes & Tracey, 2008).
In terms of differences in social context and self-relevance, actions performed in the
context of a social interaction produce greater mu rhythm suppression than actions
performed outside of an interaction (Perry et al., 2011). The mu rhythm can also be
modulated by the social relevance of the observed action (Kilner et al., 2006; Oberman
et al., 2007). Furthermore, the interpersonal liking between individuals can modulate
MNS areas, as differences in premotor cortex activation were found when observing in-
group versus out-group members’ actions (Sobhani et al., 2012). Some have suggested
that being in a social interaction, in itself, can be rewarding (Krach et al., 2011). If this is
the case, then it would be plausible to argue that differences in motor activity seen in
studies comparing social and non-social settings or stimuli may be confounded by
differences in reward-processing.
The modulation of reward and punishment on motor resonance adds further support to
the notion that the MNS can contribute to understanding others’ goals and intentions.
Furthermore, this also provides neuroscientific support to early behavioural work on
social observational learning from Bandura (1977) whereby the association of
punishment with observed actions led to a discouragement of imitative behaviour.
Therefore, this provides evidence for the dynamic interplay between action observation
and motivational drives, suggesting an online modulation of action understanding

7. GENERAL DISCUSSION
92
depending on the specific reward-related factors involved and perceived in the social
setting at a given moment in time.
It has been suggested that selective dysfunctions of the mirror system may play a key
role in the creation and maintenance of pathological deficits in social cognition,
including those seen in schizophrenia (Arbib & Mundhenk, 2005; Buccino et al., 2008;
Enticott et al., 2008; McCormick et al., 2012; Singh et al., 2011). Schizophrenia has been
referred to as a “loss in ego boundaries” whereby the distinction between self and other
is blurred, as confirmed by substantial experimental evidence, more specifically, in
terms of distinguishing between one’s own and others’ actions, voices and intentions
(e.g. Blakemore et al., 2000; Fisher et al., 2008). This blurring between self and other is a
central aspect of the skills required for social cognition. For effective functioning of the
motor system, there must be a distinction between actions that are self-generated and
those that are generated by external forces. The mechanism that underlies this
distinction between one’s own and others’ actions produces a sense of agency and is
crucial for effective motor control (Frith et al., 2000). To successfully engage with others
in a social interaction, one also needs to determine which actions are relevant to oneself
and those that are relevant to others. There may also be varying tendencies to associate
others’ actions to ourselves and to others, as the degree to which others’ actions are
relevant to oneself may be shaped by the social context in which the actions are seen. As
well as the deficits in social cognition and social functioning known in schizophrenia, it
has also been found that patients have selective abnormalities in reward-processing,
which are likely to be associated with negative symptoms such as a lack of affective
reactivity, loss of motivation and social withdrawal (Gold et al., 2008; Waltz et al., 2013).
According to the findings presented in this thesis, and in light of the known collective
deficits and cognitive impairments seen in schizophrenia, it is proposed here that some
of the previous findings on the abnormalities in the mu rhythm and MNS activity may
have resulted from a more elementary pathological deficit in distinguishing between self
and other and reward-processing during the processing of others’ actions. Therefore, it
may be the case that in schizophrenia, the disruption of this interaction between the
processing of others’ action, and the associations with self and other, and reward, could
perpetuate problems in social functioning. This may also have implications on deficits in
other domains of social cognition seen in schizophrenia. The disturbance of the

7. GENERAL DISCUSSION
93
underlying neural system that associates self-relevance and rewards to social stimuli
could lead to patients having abnormal experiences of reward and punishment when
processing social stimuli, as a cause of an underlying general deficit in self and other and
reward-processing. Alternatively, or in addition to, it may be that cognitive deficits in
processing social stimuli, or a reduced preference for social stimuli, may impoverish the
ability to experience social stimuli as being “intrinsically” rewarding. If the perception of
social stimuli in patients with schizophrenia is somehow degraded, and the self-
relevance of observed actions is disturbed, then these social stimuli may also not be
rewarding for them, as they would be in healthy people. To put this more simply, it could
be that people with schizophrenia do not find social stimuli rewarding, and therefore are
less likely to engage with such stimuli, and would also be less inclined to seek social
interactions, as it is less rewarding for them. As reward has a substantial influence on
vicarious motor activity and motor learning, then it is likely that abnormalities in the
processing of reward could also have a detrimental impact on the development of the
MNS, as well as those cognitive skills associated with MNS function, consequently
leading to the overt impairments in social cognition and social functioning seen in
schizophrenia. This proposal is further supported by emerging evidence from work in
Autism Spectrum Disorders (ASDs) suggesting that problems in social functioning seen
in ASDs may be founded upon an impaired response to social rewards (Chevallier et al.,
2012; Dichter & Adolphs, 2012; Kohls et al., 2012; Lin et al., 2012a,b). A particularly
interesting recent fMRI study (Gromann et al., 2013) has also demonstrated that
psychotic patients exhibit reduced sensitivity to social reward, as seen by the activity in
the caudate nucleus, which further correlated with paranoia scores. The authors
concluded that this reduced sensitivity may help in explaining a basic loss of trust in
schizophrenia, as mediated by brain areas associated with reward-processing and
mentalizing.
The studies included in this thesis are essentially investigating social observational
learning, linking the contextual influences on the online neural processing of other’s
actions with the neural responses associated with the evaluation of the outcomes of
others’ actions. The context created by the reward value associated with others’ actions
can modulate the degree to which we resonate or simulate others’ actions, and
additionally, the social context in which the observed action is seen can modulate the
reward value associated with the action. As suggested by the findings here looking at the

7. GENERAL DISCUSSION
94
ERPs evoked during an observational learning task, the social context and expectation
can influence the valence of the assessment of others’ actions. The evaluation of other’s
errors can be determined by how the other person’s performance or ability relates to
one’s own. It may be the case that if one sees another person’s performance as being
superior, then the valuation of the outcome of the others’ actions and behaviour is likely
to be more negative than if one considers the others’ performance to be inferior. In other
words, if one individual in a social interaction thinks that the other person is winning
more than them, then they are likely to consider their own wins to be less valuable. This
pattern may also apply in the opposite direction, and could also apply to one’s own and
others’ errors or mistakes, in that one’s own mistakes would not be seen as bad if other
people are also making the same mistakes. The proposal that the observation of others’
performance has an influence on the evaluation of one’s own performance is quite an
intuitive one, which seems to be confirmed by the previously mentioned behavioural
and neuroimaging literature, as well as by the results presented in this thesis. The level
of expectation of others’ outcomes is also intrinsically instrumental in this proposal, as
the interaction between the expectation, valence and magnitude of one’s own and
others’ outcomes or feedback can determine the degree to which the outcome is actually
rewarding for the observer. The data presented in this thesis reveals some of the
differences seen in the evaluations of others’ performance in relation to one’s own, and
the influence of expectation, expressed at a low-level of early neural processing. These
low-level contextual modulations of neural processing may also propagate to higher
levels of processing related to evaluation and expectation of others’ behaviour, based on
predetermined beliefs and stereotypes associated with social contexts (Amodio et al.,
2007; 2008).
Rewards not only reinforce behaviours, but also provide an incentive to drive behaviour
and motivate towards pursuing goals (Robbins & Everitt, 1996). The seeking of rewards
through goal-directed behaviour requires the initiation of appetitive approach
locomotor responses, and conversely, is inhibited by aversive or avoidant behaviours
(Balleine & Dickinson, 1998; Mogenson et al., 1980). In the realm of social interaction,
these principles still apply, but are influenced by additional socially relevant
environmental and biological factors (Brown & Brüne, 2012). In terms of the
environmental factors driving behaviour, it is already known that the emotional valence
and saliency of stimuli can substantially influence social approach and avoidance

7. GENERAL DISCUSSION
95
behaviour (Kemp & Guestella, 2010; Roelofs et al., 2009). Neurobiological factors
involving hormones such as cortisol and oxytocin also have a central role in determining
social approach and avoidance tendencies (Roelofs et al., 2005). Deficits in social
motivational drives have substantial relevance in functional recovery in schizophrenia
(Barch & Dowd, 2010). Lower levels of basal plasma oxytocin have previously been
associated with greater symptom severity and poorer social cognition in schizophrenia
(Goldman et al., 2008; Rubin et al., 2010). However, the study presented in this thesis
demonstrates that the effects of the differences in endogenous oxytocin levels on the
motivational and affective systems of social approach and avoidance may be
heterogeneous, with some individuals being affected more than others. Individual
differences in endogenous levels of the neuropeptides related to attachment and social
behaviour are determined by inherited genetic traits (Walum et al., 2012) and past
environmental experiences during development, such as childhood trauma (Heim et al.,
2009). The findings from the study in this thesis imply that these individual differences
in endogenous levels of oxytocin may also play a role in the differences in social
functioning seen across a schizophrenia population. Furthermore, recent interest in the
interactions between the dopaminergic system and oxytocin further underscore this
interaction between rewards, motivation and social behaviour, and the potential
relevance to the psychopathology of schizophrenia (Baskerville & Douglas, 2010;
Rosenfeld et al., 2011; Strathearn, 2011).
Psychosocial interventions that involve imitative and observational learning may benefit
from considering whether an underlying deficit in the processing of reward could
influence the ability to learn by observation. In these cases, differences in reward
processing could also influence the capacity for motor simulation, and therefore also
affect the capacity for social learning and the development of social skills during
childhood, which may also persist into adulthood. The reward or punishment associated
with others’ actions may influence the capacity for understanding others’ actions, their
goals and intentions, and therefore could also directly affect the potential for social
observational learning or its selective breakdown in certain pathological conditions such
as schizophrenia.

7. GENERAL DISCUSSION
96
7.4. Limitations
There were several limitations of the studies included in this thesis, of which some have
already been briefly mentioned, but here will now be discussed in more detail.
The study investigating the effect of the self and other framing of actions on the mu
rhythm suppression was largely flawed due to the low sample size, as this study was
intended as a pilot study that was later found to have some important confounding
variables in the experimental design. Firstly, the effect of direct and averted eye-gaze
added an inherent difference in the experimental conditions that could not be separated
out from the findings of the effect of the intended self and other manipulation across
conditions. A second major confounder of this experiment was the difference in the
spatial location of the bowls used in action execution conditions that most likely would
have had an effect on general motor cortex activity. One of the bowls in which the
objects were required to be transferred to was further than the other bowl, and this was
not counterbalanced across self and other conditions. These confounding variables were
taken into consideration in the experimental design of the later studies looking at the
mu rhythm suppression during action observation.
The first study investigating the effects of reward and punishment on the mu
suppression in the healthy group had some limitations associated with the sample
population and the controlling of confounders potentially arising from individual
differences in cognition and behaviour. To be specific, the sample used in the first study
looking at reward and punishment and the mu suppression used a largely homogenous
population of females within a narrow age range, who were all students. This therefore
may have not been representative of a healthy population as a whole. In addition,
individual differences in working memory were not controlled for, which may have
affected the action ratings given after action observation, as participants with better
working memory may have given more accurate subjective ratings of the previously
seen actions, as compared to participants with poorer working memory.
The study looking at the effects of reward and punishment on the mu rhythm
suppression in schizophrenia patients demonstrated that the matched healthy control
group exhibited a different pattern of reward-related modulation of the mu rhythm. This

7. GENERAL DISCUSSION
97
may have related to the limitation of the earlier study, as mentioned, because the
matched control group in the patient study was quite different in terms of age, and
additionally, was a mixed sex group, including both males and females. This therefore
made the two healthy control groups, i.e. the young female healthy group in the first
study and the older mixed-sex healthy group used in the patient study, less comparable.
Another limitation of this study may have come from the differences in neurocognition
between the patient group and the matched healthy control group. Differences in
neurocognition were not controlled for and may have had some influence on the
understanding of instructions, empathy questionnaires and action ratings. Furthermore,
the sample size of the groups used in the patient study may not have been sufficient to
show the full extent of the effects of the experimental manipulation of reward-related
modulation of the mu suppression. This also made the data from the first reward-related
modulation mu suppression study less comparable to the second, as sample sizes were
not matching.
In the study exploring the observational FRN and observational ERN, one limitation in
the experimental design was the order and sequence of trials presented for each
condition. The trials for high expectancy were presented in a separate block to the trials
for low expectancy. This may have caused some subsequent global contextual effects in
terms of the responses to the positive and negative feedback and the response to others’
errors, whereby the randomness of the valence of the feedback could have led to an
impoverishment of ERP responses to the feedback. In fact, this randomness may have
also induced some frustration in participants, particularly in the block for low
expectancy trials, as some participants actually reported that they tried to look for a
pattern or rule in the associations between stimulus cues and the outcomes in order to
maximise their gains. Consequently, participants may have taken a submissive or
helpless attitude towards the outcomes of their choices, and in this case, this would have
likely affected the magnitude and valence-related modulation of the ERP responses to
feedback, especially in the latter part of the low expectancy blocks of trials. Additionally,
the sample size of this study may not have been large enough to reveal the true effects of
the experimental manipulations, and therefore it is hard to draw conclusive inferences
from this data.

7. GENERAL DISCUSSION
98
For the study investigating social approach-avoidance behaviour in schizophrenia, the
main limitation was the lack of a healthy control group for comparing normal AAT
responses of healthy participants, and therefore the possibility cannot be ruled out that
the effect found between oxytocin and the AAT may not have been specific to
schizophrenia. However, given the generally slow response times and positive skewness
in AAT scores, and differences in emotion perception of this schizophrenia population,
the use of univariate comparisons would have exerted a statistical bias for this clinical
population when compared to healthy participants with a normal (faster) processing
speed. Nevertheless it would still be informative for future studies to look for causal
inferences using multivariate analyses between the oxytocin levels and AAT scores in
healthy population to allow for the generalizability of the findings.
7.5. Synthesis
The overarching aim of the work included in this thesis was to investigate some of the
processes involved in social interactions and observational learning, from biology to
cognition to behaviour, and also, how these processes can go wrong in the pathology of
schizophrenia. In studying the brain-basis for social interaction it is becoming more and
more evident that numerous interacting and parallel neural systems are recruited
during social encounters. In light of the findings presented in this thesis, and by
combining some integrative frameworks recently proposed for explaining some aspects
of social cognition, a synthesis of the empirical and theoretical work on the topics
discussed here will now be put forward. However, before this synthesis is outlined, a
number of concepts will be briefly introduced to permit further cohesion between
topics.
The general principles that will now be introduced could be considered to fall under the
umbrella of a currently popular idea of the “predictive brain”. The concept of the
“predictive brain” can be generally stated in that brains are constantly generating
mental representations to predict future states (Bar, 2007; Friston, 2005). It is thought
that these predictive internal representations of forthcoming events are constantly
being compared with the actual perceived outcome of internal mental and external
environmental events. To allow for learning to take place, one must be able to process

7. GENERAL DISCUSSION
99
one’s own errors to learn from one’s mistakes, and consequently update internal
representations of the predicted future. Reward prediction errors are generated in
dopaminergic neurons during learning and are thought to encode the magnitude of the
discrepancy, as a product of the comparison, between expected reward and the
experienced reward, i.e. actual outcome (Schultz and Dickinson, 2000). These reward
prediction errors thus drive decision-making. In addition to basic learning processes,
this predictive coding framework is also evident in perception and motor control
whereby internal models of a predicted outcome of a visual percept or motor command
are generated, and consequently act as top-down modulators of bottom-up sensory
input (Rao & Ballard, 1999; Wolpert & Miall, 1996), and also producing sensory or
action prediction errors as a result of the matching process between predicted and
actual outcome (i.e. bottom-up sensory input). This matching process is thought to
create a sense of agency and ownership for one’s own actions, perceptions and
intentions, and therefore provides the central underlying neural mechanism for
distinguishing between self and other (Frith et al., 2000). Thus a breakdown in this
matching system, i.e. when a mismatch occurs, can have pathological consequences, such
as the disturbances in distinguishing between self and other seen in schizophrenia
(Feinberg, 1978). These general principles of the “predictive brain” have proven to be a
fruitful foundation for investigating the implementation of cognitive processes in the
underlying neural substrates. This also provides a framework on which mathematical
principles can be applied, and therefore opening up the potential for testing hypotheses
about the neural and cognitive mechanisms of learning, action and perception with
biologically-plausible computational models. Despite being grounded in relatively old
ideas (Helmholtz, 1860; von Holst and Mittelstaedt, 1950; Sperry, 1950) much
experimental work has only recently emerged to provide support to this framework, and
has already started to be applied to the realm of social neuroscience (Brown & Brüne,
2012).
Much controversy still surrounds the mirror neuron hypothesis, as many authors have
questioned the functional specificity of the human MNS (Heyes, 2010a; Hickok, 2009). In
response to these criticisms, some alternative models of the mirror neuron system have
been proposed. One that is relevant here, is a predictive coding account of the MNS
(Kilner et al., 2007a,b), which uses a Bayesian statistical framework for its
implementation. This proposal argues that an internal model of an action (or predicted

7. GENERAL DISCUSSION
100
model of the expected consequence of the action) is generated during the observation of
others’ actions, which in turn transfers an action prediction through backwards
connections, from frontal areas implicated in the mirror system, to action
representations in the temporal and parietal mirror neuron-related areas. This
predicted or generative model is then matched with the sensory input of the observed
action (i.e. the visual input of the seen action), which then results in an action prediction
error. This action prediction error therefore represents the discrepancy between the
predictive model of the observed action and the actual sensory (visual) feedback
received following the observation of the action. As with other predictive systems, the
brain seeks to minimize the prediction error (Friston, 2005). Another alternative
account of the mirror system relevant here is based on associative learning (Heyes,
2001; 2010b), and argues that learned sensorimotor experiences, through self-
observation and the observation of others, actually promotes the formation and
emergence of the human MNS. This is therefore acquired and further refined throughout
development. The learned associations of action contingencies (i.e. the contingency
between actions and their sensory consequences / outcomes) are thought to provide the
basis for action understanding. These models of the MNS, and the concepts surrounding
the predictive brain, and its application in social interactions, provide the basis for the
following synthesis.
Figure 7.1 shows a schematic diagram representing a proposed integration of some of
the topics and associated mechanisms discussed in this thesis. However, this schema is
not intended to illustrate the neural pathways between specific or localised brain
regions, but instead is more of a conceptual cognitive map to display the links between
different cognitive processes. Indeed, these cognitive processes may also be served by
related networks of brain regions, though the pathways shown here are not meant to be
faithful representations of anatomical pathways. For the sake of simplicity, the role of
oxytocin has not been included in this synthesis, as this would require consideration of
other interacting hormones that were not investigated in the work in this thesis.
The diagram illustrates the interplay between two agents in a social interaction, and the
internal cognitive mechanisms underlying this interaction. The pathway labelled (a)
represents the sensory input received from the external environment during an
interaction, which may be through the observation of others’ actions, but could also

7. GENERAL DISCUSSION
101
represent input from other sensory modalities. The coupling of action and perception is
shown here by the pathway labelled (b), which is also portraying the process of motor
resonance and MNS related activity, whereby the perception of others’ actions activates
the observer’s motor cortex. As is shown by the star overlaying the action-perception, or
sensorimotor pathway (b), reward-related processing modulates the activation of motor
cortex when observing others, of which the reward value is tagged to the sensory input
via pathway (i). This tagging (or association) of the reward value to the sensory input is
reminiscent of reward-based perceptual learning (Goldstone, 1998), and also illustrates
the proposal that the reward associated with the perceived stimuli can drive the degree
to which one pays attention or engages with that stimuli. In the social interactive
context, this also encapsulates the idea that rewards can drive social learning through
modulation of sensory and motor cortex activation. The rewards associated with
perceived stimuli in a social interaction, including others actions, is generated as a
product of the predicted model. This predicted model of the expected reward outcome is
subsequently compared or matched with the actual perceived outcome, as shown by
pathways (f) and (c), respectively. This matching process of the expected reward and the
actual reward is represented here by the comparator, which produces the reward
prediction error that subsequently updates the predicted model (pathway (e)). This
updating therefore shapes future reward predictions and the reward associations made
to perceived stimuli.
The reinforcement learning loop represented by pathways (d), (e) and (f), which
receives sensory inputs via pathway (c), has reward intrinsic to its functioning. The
mechanism and neurophysiology underlying this reward-based learning loop is
relatively well-established (Schultz, 2000), though outside of the social interactive
context. This learning mechanism of updating the predicted model of future states of the
external and internal mental environment also extends to the principles of social
observational learning, and learning from others’ behaviour. The work in this thesis
presents data that confirms the similarity between the underlying neural mechanism
responsible for learning from one’s own actions, and from learning through the
observation of others’ actions and behaviours. The similarities seen between the ERN
and FRN, and the oERN and oFRN are clear examples of this, as individuals can either
have empathetic or Schadenfreude-like experiences of others’ errors.

7. GENERAL DISCUSSION
102
As a comparable mechanism, the sensory input received during the social interaction is
also matched to the predicted model of that input, be it for example, from the predicted
outcome of an observed action or a heard speech gesture, producing a sensory or
cognitive prediction error. This feedback mechanism could be comparable to the social
version of the MOSAIC model proposed by Wolpert and colleagues (2003), which
parallels the sensorimotor loop between the predictive model and incoming sensory
information (as shown by pathways (c), (d), (e) and (f)), with the social interactive loop
being between self-generated and observed communicative actions. Communicative
actions are thought to be generated from the actions observed by a confederate, which
consequently causes changes in the observer’s mental state, which in turn initiates
communicative actions from the other person, which are perceived by the observer. This
interpersonal loop of social interaction therefore allows one to make predictions and
learn about the likely behaviour of another person in response to one’s own
communicative behaviour. The internal predictive models of other people are thought to
be decoded and learned through the mappings between our own actions and our own
mental states as a priori information, thereby using one’s own motor system to compute
the internal mental states of others, which is consequently suggested to form a basis for
theory of mind.
In the case of a social interaction, the prediction error produced as a result of the
matching of the expectations of others’ actions, and the actual perceived outcomes of the
other’s actions could be considered more specifically as a “social prediction error” that
serves to update the predicted model of other people’s behaviour. The difference
between a social and non-social prediction error would be that the social prediction
error is mediated by additional factors, including social knowledge and expectations of
others’ behaviour. For example the coding of the social reward prediction error, when
observing the outcomes of others’ actions, may produce a prediction error similar to
that produced when one observes the outcome of one’s own actions, but rather the
reward value would be relative to the consequence of the observed outcome on the
observer, i.e., how rewarding other’s behaviours are to the observer. This social
prediction error could also act as a teaching signal for updating higher-level
expectations of others in a social context, such as those related to predetermined beliefs
and stereotypes about individuals or social categories. In the case of these more high-
level predictions about the state of the world, social prediction errors can be generated

7. GENERAL DISCUSSION
103
as a result of an expectancy violation in a social situation, such as when a person breaks
a promise (Baumgartner et al., 2009) or when a social norm is violated (Harris & Fiske,
2010). Contextual information for social expectancies could come from environmental
cues, and particularly the context of the social situation, or could be generated from
internal contexts such as an individual’s affective state, or from a cognitive bias, such as
an attributional bias, as seen in schizophrenia (Bentall et al., 1994).
The generative internal model of the predicted sensory outcome of an event is also used
in fine motor control, and has been referred to by Friston and colleagues (2011) with the
term “precision”, which is shown in the diagram by pathway (g). The cyclic updating of
the predicted model of an executed action, via comparison with the sensory feedback
(pathway (c), (d), (e) and (f)), is utilised to make fine motor adjustments during the
performance of an action, i.e. contributing to pathway (g). In relation to this, a similar
mechanism involving the predicted reward outcome of one’s own actions could also be
represented conceptually by this pathway and feedback loop. A comparable mechanism
may also be at play in a social interaction, in which this cyclic updating of the expected
outcomes of others actions drives the execution of one’s own actions and behaviour. The
dynamic interplay between the perceived behaviour of others, and the expected
outcome of one’s own and others’ actions, are likely to drive social behaviours, including
social approach and avoidance, as shown here by pathway (j). The motivational aspects
behind social approach and avoidance behaviour are determined by the rewards
associated with one’s own and others actions, and thus the initiation of one’s own motor
actions and behaviour is driven by the interactions between these internal and external
motivational factors in the social scenario.
The figure also highlights some areas of this cognitive map that could potentially be
going wrong in schizophrenia. These are labelled with a red outline. It is known that
people with schizophrenia have some sensory and motor processing dysfunctions
(Javitt, 2009; Schurmann et al., 2007), and thus this is labelled in the diagram. According
to the pathways laid out in this schema, these sensory and motor deficits may also play a
role in the problems in the transmission of reward-related processing in other areas that
are encoding the reward, and thus having more downstream effects that would only be
expressed in motivational signs in behaviour. It has been proposed that differences in
reward-processing in schizophrenia cannot be merely explained by a generalised

7. GENERAL DISCUSSION
104
impairment in the experience of reward, but may be more likely to be a deficit in the
representation of the value of different choices, which can consequently lead to the
impairments seen in decision-making (Gold et al., 2008). Therefore, this deficit in the
representation of value, and the subsequent influence on choice responses, may be
representative of a disturbance in the pathways (b) to (j) illustrated in figure 7.1. Thus, a
breakdown in these pathways could potentially influence social approach and
avoidance, and the initiation of social behaviours. As a result of a breakdown in this
mechanism, motivation to seek social interaction may also be diminished. Eventually,
this is likely to have a detrimental effect on the capacity for successful social functioning
in schizophrenia.
Furthermore, there is a substantial body of work suggesting a disturbance in the
matching process between the predicted outcome or sensory consequence of an event
and the actual sensory input in schizophrenia (Feinberg, 1978; Ford et al., 2001)
Although this work refers instead to the corollary discharge and efference copy, which is
essentially a copy of a motor command, and is represented in the figure by pathway (f).
A dysfunction in the processing of the corollary discharge or efference copy has been
used to explain disturbances in the sense of agency and the generation of auditory
hallucinations in schizophrenia (Ford & Mathalon, 2005). This disturbance in the
processing of the corollary discharge is comparable to a mismatch occurring between
pathways (c) and (f), and is therefore represented in figure 7.1 as a dysfunction in the
comparator. This would therefore also produce inaccurate prediction errors, which may
impact on the reward-related and sensory predictive models. If this was the case in
schizophrenia, then according to this schema illustrated here, a disturbance in the
comparator may have resonating effects on other downstream and connected processes.
In essence, the schematic representation illustrated by figure 7.1 seeks to integrate a
series of cognitive loops associated with the processing of one’s own actions, perception
and learning, and how these processes become extended into the context of social
interaction through the processing of others’ behaviour. The processing of both non-
social and social cognitive mechanisms is represented by these multiple interacting
loops in personal and interpersonal neural systems, which interface at sensory and
motor inputs and outputs. The aim of this schematic representation is also to highlight
the dynamic and parallel nature of the processes that occur during a social interaction

7. GENERAL DISCUSSION
105
between and within two interacting brains. Within the scope of this schema, some areas
are highlighted in which potential breakdowns may be occurring in schizophrenia. In
light of the dynamic and parallel nature of the interactions between personal and
interpersonal cognitive processes, it is therefore easy to see how breakdowns in some
low-level sensory or perceptual sub-processes may propagate downstream, eventually
leading to more overt dysfunctions in high-level social cognition and social functioning.

7. GENERAL DISCUSSION
106

7. GENERAL DISCUSSION
107
7.6. Open questions and future suggestions
It is still under debate as to what degree the MNS is involved in action understanding,
and even whether this activity actually reflects action understanding at all (Hickok,
2009). Evidently, there is still more work to be done to clarify the functional specificity
of the MNS in humans, and to see whether different areas related to the human mirror
neuron network may have more specific subsets of functions related to social cognition.
Further studies exploring the mu rhythm suppression during action observation should
also seek to dissociate the dynamic temporal changes in neural activity when making
inferences about social interaction, which more accurately reflect the dynamic changes
in the environment that occur during everyday social interaction. In light of these new
results, differential “simulated” motor effects may stem from underlying fundamental
situational and contextual differences in the processing of self-relevance, reward or
punishment. More specifically, future studies therefore may need to consider the
potential confounding effects of the associated self-relevance and reward on the
observed action in the experimental condition of interest, and the self/reward-related
associations of actions created by different contexts, whether it is social or not. To gain
further insight into the findings presented here on the mu rhythm, it would be
interesting for a future study to investigate the interaction between self-relevance and
reward in action observation in both a healthy and schizophrenia group. A further
suggestion would be to see if the modulation of motor resonance as a result of the
interaction between self-relevance and reward may be also associated with behavioural
measures of empathy and Schadenfreude.
It is still not clear how deficits in reward-processing may be associated with deficits in
social cognitive processes in psychopathologies such as schizophrenia. Further studies
are required to discern how potential disturbances at the low level of sensory and motor
processing can translate to the poor performance seen in more complex social cognitive
tasks in psychiatric illness. Future work looking at the ERP responses, such as the oFRN
and oERN, in healthy and schizophrenia populations during the observation of others’
behaviour is likely to provide a very useful and informative platform to explore the
underlying neurological causes for higher level social cognitive deficits.

7. GENERAL DISCUSSION
108
There has recently been much attention paid to the potential therapeutic effects of
oxytocin administration in treating pathological deficits in social cognition (Feifel,
2012). However, it appears that much of the research on oxytocin with schizophrenia
populations has largely focused on improvements in high-level social cognitive skills and
social functioning capacity as the primary outcome measure, with many showing mixed
results (Feifel, 2012). However, it may be more feasible and informative to first look at
the effect of neuroendocrinological factors underlying social behaviour at a low-level of
processing, by investigating the neurophysiological effects of oxytocin on social affective
and motivational systems, and how these effects may be translated downstream,
through neural activity, and inevitably into social behaviour. The findings presented
here encourage replication with future studies using oxytocin administration in clinical
groups, while also considering the endogenous individual differences as confounders
when assessing the effect of exogenous neuroendocrinological manipulation on social
behaviours and motivations.
The abnormalities seen in schizophrenia in the underlying neurophysiological and
neurobiological mechanisms associated with the observation of others’ actions, the
outcomes of others’ actions and the relationship with social motivations leading to
approach and avoidance behaviours may help to serve as signs of risk factors in the
development of the illness, and particular symptom clusters. Using these potential
biomarkers of psychotic characteristics may also aid in predicting treatment efficacy.
This could therefore contribute to the development of more specialised treatment
programs for individuals and subgroups of individuals with schizophrenia that aim to
remediate specific sets of social cognitive skills. The use of brain-based assessment
paradigms also provides an extra tool for the clinician to determine the most effective
treatments for patients and to give more accurate prognoses.

8. REFERENCES
109
Chapter 8
References
Abu-Akel, A., Shamay-Tsoory, S., 2011. Neuroanatomical and neurochemical bases of theory of mind. Neuropsychologia 49, 2971-2984. Adams, R.B., Jr., Kleck, R.E., 2005. Effects of direct and averted gaze on the perception of facially communicated emotion. Emotion 5, 3-11. Adolphs, R., 2001. The neurobiology of social cognition. Current opinion in neurobiology 11, 231-239. Agnew, Z.K., Bhakoo, K.K., Puri, B.K., 2007. The human mirror system: a motor resonance theory of mind-reading. Brain research reviews 54, 286-293. Alaerts, K., Heremans, E., Swinnen, S.P., Wenderoth, N., 2009. How are observed actions mapped to the observer's motor system? Influence of posture and perspective. Neuropsychologia 47, 415-422. Amico, J.A., Tenicela, R., Johnston, J., Robinson, A.G., 1983. A time-dependent peak of oxytocin exists in cerebrospinal fluid but not in plasma of humans. The Journal of clinical endocrinology and metabolism 57, 947-951. Amodio, D.M., Devine, P.G., Harmon-Jones, E., 2008. Individual differences in the regulation of intergroup bias: the role of conflict monitoring and neural signals for control. Journal of personality and social psychology 94, 60-74. Amodio, D.M., Frith, C.D., 2006. Meeting of minds: the medial frontal cortex and social cognition. Nature reviews. Neuroscience 7, 268-277. Amodio, D.M., Jost, J.T., Master, S.L., Yee, C.M., 2007. Neurocognitive correlates of liberalism and conservatism. Nature neuroscience 10, 1246-1247.

8. REFERENCES
110
Andreasen, N.C., Calarge, C.A., O'Leary, D.S., 2008. Theory of mind and schizophrenia: a positron emission tomography study of medication-free patients. Schizophrenia bulletin 34, 708-719. Annen, S., Roser, P., Brune, M., 2012. Nonverbal behavior during clinical interviews: similarities and dissimilarities among schizophrenia, mania, and depression. The Journal of nervous and mental disease 200, 26-32. Arbib, M.A., Mundhenk, T.N., 2005. Schizophrenia and the mirror system: an essay. Neuropsychologia 43, 268-280. Averbeck, B.B., Bobin, T., Evans, S., Shergill, S.S., 2011. Emotion recognition and oxytocin in patients with schizophrenia. Psychological medicine, 1-8. Baker, T.E., Holroyd, C.B., 2009. Which way do I go? Neural activation in response to feedback and spatial processing in a virtual T-maze. Cerebral cortex 19, 1708-1722. Bakvis, P., Spinhoven, P., Zitman, F.G., Roelofs, K., 2011. Automatic avoidance tendencies in patients with psychogenic non-epileptic seizures. Seizure : the journal of the British Epilepsy Association 20, 628-634. Balleine, B.W., Dickinson, A., 1998. Goal-directed instrumental action: contingency and incentive learning and their cortical substrates. Neuropharmacology 37, 407-419. Bandura, A., 1977. Social learning theory. Prentice-Hall, Englewood Cliffs. Bar, M., 2007. The proactive brain: using analogies and associations to generate predictions. Trends in cognitive sciences 11, 280-289. Barch, D.M., Dowd, E.C., 2010. Goal representations and motivational drive in schizophrenia: the role of prefrontal-striatal interactions. Schizophrenia bulletin 36, 919-934. Bartz, J.A., Hollander, E., 2006. The neuroscience of affiliation: forging links between basic and clinical research on neuropeptides and social behavior. Hormones and behavior 50, 518-528. Bartz, J.A., Zaki, J., Bolger, N., Hollander, E., Ludwig, N.N., Kolevzon, A., Ochsner, K.N., 2010. Oxytocin selectively improves empathic accuracy. Psychological science 21, 1426-1428. Baskerville, T.A., Douglas, A.J., 2010. Dopamine and oxytocin interactions underlying behaviors: potential contributions to behavioral disorders. CNS neuroscience & therapeutics 16, e92-123. Baumgartner, T., Fischbacher, U., Feierabend, A., Lutz, K., Fehr, E., 2009. The neural circuitry of a broken promise. Neuron 64, 756-770. Behrens, T.E., Hunt, L.T., Woolrich, M.W., Rushworth, M.F., 2008. Associative learning of social value. Nature 456, 245-249.

8. REFERENCES
111
Bellebaum, C., Kobza, S., Thiele, S., Daum, I., 2010. It was not MY fault: event-related brain potentials in active and observational learning from feedback. Cerebral cortex 20, 2874-2883. Bentall, R.P., Kinderman, P., Kaney, S., 1994. The self, attributional processes and abnormal beliefs: towards a model of persecutory delusions. Behaviour research and therapy 32, 331-341. Bismark, A.W., Hajcak, G., Whitworth, N.M., Allen, J.J., 2013. The role of outcome expectations in the generation of the feedback-related negativity. Psychophysiology 50, 125-133. Blakemore, S.J., Smith, J., Steel, R., Johnstone, C.E., Frith, C.D., 2000. The perception of self-produced sensory stimuli in patients with auditory hallucinations and passivity experiences: evidence for a breakdown in self-monitoring. Psychological medicine 30, 1131-1139. Blakemore, S.J., Winston, J., Frith, U., 2004. Social cognitive neuroscience: where are we heading? Trends in cognitive sciences 8, 216-222. Bora, E., Yucel, M., Pantelis, C., 2009. Theory of mind impairment in schizophrenia: meta-analysis. Schizophrenia research 109, 1-9. Brekke, J., Kay, D.D., Lee, K.S., Green, M.F., 2005. Biosocial pathways to functional outcome in schizophrenia. Schizophrenia research 80, 213-225. Brown, E.C., Brune, M., 2012. The role of prediction in social neuroscience. Frontiers in human neuroscience 6, 147. Brown, E.C., Tas, C., Brune, M., 2012. Potential therapeutic avenues to tackle social cognition problems in schizophrenia. Expert review of neurotherapeutics 12, 71-81. Brown, E.C., Wiersema, J.R., Pourtois, G., Brune, M., 2013. Modulation of motor cortex activity when observing rewarding and punishing actions. Neuropsychologia 51, 52-58. Brüne, M., 2005. "Theory of mind" in schizophrenia: a review of the literature. Schizophrenia bulletin 31, 21-42. Brüne, M., Abdel-Hamid, M., Lehmkamper, C., Sonntag, C., 2007. Mental state attribution, neurocognitive functioning, and psychopathology: what predicts poor social competence in schizophrenia best? Schizophrenia research 92, 151-159. Brüne, M., Abdel-Hamid, M., Sonntag, C., Lehmkamper, C., Langdon, R., 2009. Linking social cognition with social interaction: Non-verbal expressivity, social competence and "mentalising" in patients with schizophrenia spectrum disorders. Behavioral and brain functions : BBF 5, 6.

8. REFERENCES
112
Brüne, M., Sonntag, C., Abdel-Hamid, M., Lehmkamper, C., Juckel, G., Troisi, A., 2008. Nonverbal behavior during standardized interviews in patients with schizophrenia spectrum disorders. The Journal of nervous and mental disease 196, 282-288. Buccino, G., Amore, M., 2008. Mirror neurons and the understanding of behavioural symptoms in psychiatric disorders. Current opinion in psychiatry 21, 281-285. Burgdorf, J., Panksepp, J., 2006. The neurobiology of positive emotions. Neuroscience and biobehavioral reviews 30, 173-187. Cacioppo, J.T., Berntson, G.G., 1992. Social psychological contributions to the decade of the brain. Doctrine of multilevel analysis. The American psychologist 47, 1019-1028. Caetano, G., Jousmaki, V., Hari, R., 2007. Actor's and observer's primary motor cortices stabilize similarly after seen or heard motor actions. Proceedings of the National Academy of Sciences of the United States of America 104, 9058-9062. Cacioppo, J.T., Priester, J.R., Berntson, G.G., 1993. Rudimentary determinants of attitudes. II: Arm flexion and extension have differential effects on attitudes. Journal of personality and social psychology 65, 5-17. Caggiano, V., Fogassi, L., Rizzolatti, G., Pomper, J.K., Thier, P., Giese, M.A., Casile, A., 2011. View-based encoding of actions in mirror neurons of area f5 in macaque premotor cortex. Current biology : CB 21, 144-148. Cannon, W.B., 1929. Bodily changes in pain, hunger, fear and rage : an account of recent researches into the function of emotional excitement. D. Appleton and Company, New York ; London. Carp, J., Halenar, M.J., Quandt, L.C., Sklar, A., Compton, R.J., 2009. Perceived similarity and neural mirroring: evidence from vicarious error processing. Social neuroscience 4, 85-96. Cattaneo, L., Rizzolatti, G., 2009. The mirror neuron system. Archives of neurology 66, 557-560. Centelles, L., Assaiante, C., Nazarian, B., Anton, J.L., Schmitz, C., 2011. Recruitment of both the mirror and the mentalizing networks when observing social interactions depicted by point-lights: a neuroimaging study. PloS one 6, e15749. Centerbar, D.B., Clore, G.L., 2006. Do approach-avoidance actions create attitudes? Psychological science 17, 22-29. Cheng, Y., Yang, C.Y., Lin, C.P., Lee, P.L., Decety, J., 2008. The perception of pain in others suppresses somatosensory oscillations: a magnetoencephalography study. NeuroImage 40, 1833-1840. Chevallier, C., Kohls, G., Troiani, V., Brodkin, E.S., Schultz, R.T., 2012. The social motivation theory of autism. Trends in cognitive sciences 16, 231-239.

8. REFERENCES
113
Cochin, S., Barthelemy, C., Roux, S., Martineau, J., 1999. Observation and execution of movement: similarities demonstrated by quantified electroencephalography. The European journal of neuroscience 11, 1839-1842. Daly, J.J., Cheng, R., Rogers, J., Litinas, K., Hrovat, K., Dohring, M., 2009. Feasibility of a new application of noninvasive Brain Computer Interface (BCI): a case study of training for recovery of volitional motor control after stroke. Journal of neurologic physical therapy : JNPT 33, 203-211. Davis, M. 1980. A multidimensional approach to individual differences in empathy. JSAS catalogue of Selected Documents in Psychology, 10, 85. Decety, J., Chaminade, T., 2003. When the self represents the other: a new cognitive neuroscience view on psychological identification. Consciousness and cognition 12, 577-596. Dichter, G., Adolphs, R., 2012. Reward processing in autism: a thematic series. Journal of neurodevelopmental disorders 4, 20. Enticott, P.G., Hoy, K.E., Herring, S.E., Johnston, P.J., Daskalakis, Z.J., Fitzgerald, P.B., 2008. Reduced motor facilitation during action observation in schizophrenia: a mirror neuron deficit? Schizophrenia research 102, 116-121. Evans, S., Shergill, S.S., Averbeck, B.B., 2010. Oxytocin decreases aversion to angry faces in an associative learning task. Neuropsychopharmacology : official publication of the American College of Neuropsychopharmacology 35, 2502-2509. Evans, S., Shergill, S.S., Chouhan, V., Bristow, E., Collier, T., Averbeck, B.B., 2011. Patients with schizophrenia show increased aversion to angry faces in an associative learning task. Psychological medicine 41, 1471-1479. Falkenstein, M., Hohnsbein, J., Hoormann, J., & Blanke, L., 1990. Effects of errors in choice reaction tasks on the ERP under focused and divided attention. Psychophysiological brain research, 1, 192-195. Fehr, E., Camerer, C.F., 2007. Social neuroeconomics: the neural circuitry of social preferences. Trends in cognitive sciences 11, 419-427. Feifel, D., 2012. Oxytocin as a potential therapeutic target for schizophrenia and other neuropsychiatric conditions. Neuropsychopharmacology : official publication of the American College of Neuropsychopharmacology 37, 304-305. Feinberg, I., 1978. Efference copy and corollary discharge: implications for thinking and its disorders. Schizophrenia bulletin 4, 636-640. Festinger, L., Hutte, H.A., 1954. An experimental investigation of the effect of unstable interpersonal relations in a group. Journal of abnormal psychology 49, 513-522. Fisher, M., McCoy, K., Poole, J.H., Vinogradov, S., 2008. Self and other in schizophrenia: a cognitive neuroscience perspective. The American journal of psychiatry 165, 1465-1472.

8. REFERENCES
114
Foldemo, A., Ek, A.C., Bogren, L., 2004. Needs in outpatients with schizophrenia, assessed by the patients themselves and their parents and staff. Social psychiatry and psychiatric epidemiology 39, 381-385. Ford, J.M., Mathalon, D.H., Heinks, T., Kalba, S., Faustman, W.O., Roth, W.T., 2001. Neurophysiological evidence of corollary discharge dysfunction in schizophrenia. The American journal of psychiatry 158, 2069-2071. Ford, J.M., Mathalon, D.H., 2005. Corollary discharge dysfunction in schizophrenia: can it explain auditory hallucinations? International journal of psychophysiology : official journal of the International Organization of Psychophysiology 58, 179-189. Frank, M.J., Seeberger, L.C., O'Reilly R, C., 2004. By carrot or by stick: cognitive reinforcement learning in parkinsonism. Science 306, 1940-1943. Frank, M.J., Woroch, B.S., Curran, T., 2005. Error-related negativity predicts reinforcement learning and conflict biases. Neuron 47, 495-501. Friesen, C.K., Kingstone, A., 1998. The eyes have it! Reflexive orienting is triggered by nonpredictive gaze. Psychonomic bulletin and review 5, 490–495. Friston, K., 2005. A theory of cortical responses. Philosophical transactions of the Royal Society of London. Series B, Biological sciences 360, 815-836. Friston, K., Mattout, J., Kilner, J., 2011. Action understanding and active inference. Biological cybernetics 104, 137-160. Frith, C.D., 1992. The cognitive neuropsychology of schizophrenia. Erlbaum, Hove. Frith, C.D., Blakemore, S., Wolpert, D.M., 2000. Explaining the symptoms of schizophrenia: abnormalities in the awareness of action. Brain research. Brain research reviews 31, 357-363. Fukushima, H., Hiraki, K., 2009. Whose loss is it? Human electrophysiological correlates of non-self reward processing. Social neuroscience 4, 261-275. Gable, S.L., 2006. Approach and avoidance social motives and goals. Journal of personality 74, 175-222. Gallese, V., Fadiga, L., Fogassi, L., Rizzolatti, G., 1996. Action recognition in the premotor cortex. Brain : a journal of neurology 119 ( Pt 2), 593-609. Gallese, V., Keysers, C., Rizzolatti, G., 2004. A unifying view of the basis of social cognition. Trends in cognitive sciences 8, 396-403. Gastaut, H.J., Bert, J., 1954. EEG changes during cinematographic presentation; moving picture activation of the EEG. Electroencephalography and clinical neurophysiology 6, 433-444.

8. REFERENCES
115
Geerts, E., Brune, M., 2009. Ethological approaches to psychiatric disorders: focus on depression and schizophrenia. The Australian and New Zealand journal of psychiatry 43, 1007-1015. Gehring, W.J., Willoughby, A.R., 2002. The medial frontal cortex and the rapid processing of monetary gains and losses. Science 295, 2279-2282. Gintis, H., Fehr, E., 2012. The social structure of cooperation and punishment. The Behavioral and brain sciences 35, 28-29. Gold, J.M., Waltz, J.A., Prentice, K.J., Morris, S.E., Heerey, E.A., 2008. Reward processing in schizophrenia: a deficit in the representation of value. Schizophrenia bulletin 34, 835-847. Goldman, M., Marlow-O'Connor, M., Torres, I., Carter, C.S., 2008. Diminished plasma oxytocin in schizophrenic patients with neuroendocrine dysfunction and emotional deficits. Schizophrenia research 98, 247-255. Gray, J.A., 1987. The psychology of fear and stress. Cambridge University Press, Cambridge. Green, M.F., Leitman, D.I., 2008. Social cognition in schizophrenia. Schizophrenia bulletin 34, 670-672. Green, M.F., Penn, D.L., Bentall, R., Carpenter, W.T., Gaebel, W., Gur, R.C., Kring, A.M., Park, S., Silverstein, S.M., Heinssen, R., 2008. Social cognition in schizophrenia: an NIMH workshop on definitions, assessment, and research opportunities. Schizophrenia bulletin 34, 1211-1220. Gromann, P.M., Heslenfeld, D.J., Fett, A.K., Joyce, D.W., Shergill, S.S., Krabbendam, L., 2013. Trust versus paranoia: abnormal response to social reward in psychotic illness. Brain : a journal of neurology. Guastella, A.J., Mitchell, P.B., Dadds, M.R., 2008. Oxytocin increases gaze to the eye region of human faces. Biological psychiatry 63, 3-5. Gur, R.E., McGrath, C., Chan, R.M., Schroeder, L., Turner, T., Turetsky, B.I., Kohler, C., Alsop, D., Maldjian, J., Ragland, J.D., Gur, R.C., 2002. An fMRI study of facial emotion processing in patients with schizophrenia. The American journal of psychiatry 159, 1992-1999. Gutsell, J.N., Inzlicht, M., 2012. Intergroup differences in the sharing of emotive states: neural evidence of an empathy gap. Social cognitive and affective neuroscience 7, 596-603. Hajcak, G., Moser, J.S., Holroyd, C.B., Simons, R.F., 2006. The feedback-related negativity reflects the binary evaluation of good versus bad outcomes. Biological psychology 71, 148-154.

8. REFERENCES
116
Haker, H., Rossler, W., 2009. Empathy in schizophrenia: impaired resonance. European archives of psychiatry and clinical neuroscience 259, 352-361. Hari, R., 2006. Action-perception connection and the cortical mu rhythm. Progress in brain research 159, 253-260. Harris, L.T., Fiske, S.T., 2010. Neural regions that underlie reinforcement learning are also active for social expectancy violations. Social neuroscience 5, 76-91. Harvey, P.O., Zaki, J., Lee, J., Ochsner, K., Green, M.F., 2013. Neural substrates of empathic accuracy in people with schizophrenia. Schizophrenia bulletin 39, 617-628. Heim, C., Young, L.J., Newport, D.J., Mletzko, T., Miller, A.H., Nemeroff, C.B., 2009. Lower CSF oxytocin concentrations in women with a history of childhood abuse. Molecular psychiatry 14, 954-958. Heinrichs, M., Domes, G., 2008. Neuropeptides and social behaviour: effects of oxytocin and vasopressin in humans. Progress in brain research 170, 337-350. Helmholtz, H.v., 1867. Handbuch der physiologischen Optik. L. Voss, Leipzig. Heuer, K., Rinck, M., Becker, E.S., 2007. Avoidance of emotional facial expressions in social anxiety: The Approach-Avoidance Task. Behaviour research and therapy 45, 2990-3001. Heyes, C., 2001. Causes and consequences of imitation. Trends in cognitive sciences 5, 253-261. Heyes, C., 2010a. Mesmerising mirror neurons. NeuroImage 51, 789-791. Heyes, C., 2010b. Where do mirror neurons come from? Neuroscience and biobehavioral reviews 34, 575-583. Hickok, G., 2009. Eight problems for the mirror neuron theory of action understanding in monkeys and humans. Journal of cognitive neuroscience 21, 1229-1243. Hofer, A., Benecke, C., Edlinger, M., Huber, R., Kemmler, G., Rettenbacher, M.A., Schleich, G., Wolfgang Fleischhacker, W., 2009. Facial emotion recognition and its relationship to symptomatic, subjective, and functional outcomes in outpatients with chronic schizophrenia. European psychiatry : the journal of the Association of European Psychiatrists 24, 27-32. Holroyd, C.B., Coles, M.G., 2002. The neural basis of human error processing: reinforcement learning, dopamine, and the error-related negativity. Psychological review 109, 679-709. Holroyd, C.B., Krigolson, O.E., 2007. Reward prediction error signals associated with a modified time estimation task. Psychophysiology 44, 913-917.

8. REFERENCES
117
Holroyd, C.B., Nieuwenhuis, S., Yeung, N., Cohen, J.D., 2003. Errors in reward prediction are reflected in the event-related brain potential. Neuroreport 14, 2481-2484. Honaga, E., Ishii, R., Kurimoto, R., Canuet, L., Ikezawa, K., Takahashi, H., Nakahachi, T., Iwase, M., Mizuta, I., Yoshimine, T., Takeda, M., 2010. Post-movement beta rebound abnormality as indicator of mirror neuron system dysfunction in autistic spectrum disorder: an MEG study. Neuroscience letters 478, 141-145. Iacoboni, M., Molnar-Szakacs, I., Gallese, V., Buccino, G., Mazziotta, J.C., Rizzolatti, G., 2005. Grasping the intentions of others with one's own mirror neuron system. PLoS biology 3, e79. Itagaki, S., Katayama, J., 2008. Self-relevant criteria determine the evaluation of outcomes induced by others. Neuroreport 19, 383-387. Javitt, D.C., 2009. Sensory processing in schizophrenia: neither simple nor intact. Schizophrenia bulletin 35, 1059-1064. Jeannerod, M., Anquetil, T., 2008. Putting oneself in the perspective of the other: a framework for self-other differentiation. Social neuroscience 3, 356-367. John, A.E., Mervis, C.B., 2010. Comprehension of the communicative intent behind pointing and gazing gestures by young children with Williams syndrome or Down syndrome. Journal of speech, language, and hearing research : JSLHR 53, 950-960. Johns, J.H., Quay, H.C., 1962. The effect of social reward on verbal conditioning in psychopathic and neurotic military offenders. Journal of consulting psychology 26, 217-220. Jung, T.P., Makeig, S., Humphries, C., Lee, T.W., McKeown, M.J., Iragui, V., Sejnowski, T.J., 2000. Removing electroencephalographic artifacts by blind source separation. Psychophysiology 37, 163-178. Kato, Y., Muramatsu, T., Kato, M., Shibukawa, Y., Shintani, M., Mimura, M., 2011. Magnetoencephalography study of right parietal lobe dysfunction of the evoked mirror neuron system in antipsychotic-free schizophrenia. PloS one 6, e28087. Kay, S.R., Fiszbein, A., Opler, L.A., 1987. The positive and negative syndrome scale (PANSS) for schizophrenia. Schizophrenia bulletin 13, 261-276. Kemp, A.H., Guastella, A.J., 2010. Oxytocin: prosocial behavior, social salience, or approach-related behavior? Biological psychiatry 67, e33-34; author reply e35. Kendon, A., 1967. Some functions of gaze-direction in social interaction. Acta psychologica 26, 22-63. Kerr, S.L., Neale, J.M., 1993. Emotion perception in schizophrenia: specific deficit or further evidence of generalized poor performance? Journal of abnormal psychology 102, 312-318.

8. REFERENCES
118
Keysers, C., Gazzola, V., 2007. Integrating simulation and theory of mind: from self to social cognition. Trends in cognitive sciences 11, 194-196. Keysers, C., Perrett, D.I., 2004. Demystifying social cognition: a Hebbian perspective. Trends in cognitive sciences 8, 501-507. Kiehl, K.A., Smith, A.M., Hare, R.D., Liddle, P.F., 2000. An event-related potential investigation of response inhibition in schizophrenia and psychopathy. Biological psychiatry 48, 210-221. Kilner, J.M., Friston, K.J., Frith, C.D., 2007a. Predictive coding: an account of the mirror neuron system. Cognitive processing 8, 159-166. Kilner, J.M., Friston, K.J., Frith, C.D., 2007b. The mirror-neuron system: a Bayesian perspective. Neuroreport 18, 619-623. Kilner, J.M., Marchant, J.L., Frith, C.D., 2006. Modulation of the mirror system by social relevance. Social cognitive and affective neuroscience 1, 143-148. Kim, J., Doop, M.L., Blake, R., Park, S., 2005. Impaired visual recognition of biological motion in schizophrenia. Schizophrenia research 77, 299-307. Kim, C., Kroger, J.K., Kim, J., 2011. A functional dissociation of conflict processing within anterior cingulate cortex. Human brain mapping 32, 304-312. Koban, L., Pourtois, G., Vocat, R., Vuilleumier, P., 2010. When your errors make me lose or win: event-related potentials to observed errors of cooperators and competitors. Social neuroscience 5, 360-374. Kobza, S., Thoma, P., Daum, I., Bellebaum, C., 2011. The feedback-related negativity is modulated by feedback probability in observational learning. Behavioural brain research 225, 396-404. Kohls, G., Chevallier, C., Troiani, V., Schultz, R.T., 2012. Social 'wanting' dysfunction in autism: neurobiological underpinnings and treatment implications. Journal of neurodevelopmental disorders 4, 10. Krach, S., Paulus, F.M., Bodden, M., Kircher, T., 2010. The rewarding nature of social interactions. Frontiers in behavioral neuroscience 4, 22. Kuhlman, W.N., 1978. Functional topography of the human mu rhythm. Electroencephalography and clinical neurophysiology 44, 83-93. Kurtz, M.M., Richardson, C.L., 2012. Social cognitive training for schizophrenia: a meta-analytic investigation of controlled research. Schizophrenia bulletin 38, 1092-1104. Leknes, S., Tracey, I., 2008. A common neurobiology for pain and pleasure. Nature reviews. Neuroscience 9, 314-320.

8. REFERENCES
119
Libby, L.K., Shaeffer, E.M., Eibach, R.P., 2009. Seeing meaning in action: a bidirectional link between visual perspective and action identification level. Journal of experimental psychology. General 138, 503-516. Liddle, P.F., 1987. Schizophrenic syndromes, cognitive performance and neurological dysfunction. Psychological medicine 17, 49-57. Lin, A., Rangel, A., Adolphs, R., 2012a. Impaired learning of social compared to monetary rewards in autism. Frontiers in neuroscience 6, 143. Lin, A., Tsai, K., Rangel, A., Adolphs, R., 2012b. Reduced social preferences in autism: evidence from charitable donations. Journal of neurodevelopmental disorders 4, 8. Louise von Borries, A.K., Volman, I., de Bruijn, E.R., Bulten, B.H., Verkes, R.J., Roelofs, K., 2012. Psychopaths lack the automatic avoidance of social threat: relation to instrumental aggression. Psychiatry research 200, 761-766. Lysaker, P.H., Davis, L.W., Hunter, N.L., 2004. Neurocognitive, social and clinical correlates of two domains of hopelessness in schizophrenia. Schizophrenia research 70, 277-285. Ma, Q., Shen, Q., Xu, Q., Li, D., Shu, L., Weber, B., 2011. Empathic responses to others' gains and losses: an electrophysiological investigation. NeuroImage 54, 2472-2480. Madsen, H., Thyregod, P., 2011. Introduction to General and Generalized Linear Models. Boca Raton, FL, USA: Chapman and Hall. Maeda, F., Kleiner-Fisman, G., Pascual-Leone, A., 2002. Motor facilitation while observing hand actions: specificity of the effect and role of observer's orientation. Journal of neurophysiology 87, 1329-1335. Marsh, A.A., Ambady, N., Kleck, R.E., 2005. The effects of fear and anger facial expressions on approach- and avoidance-related behaviors. Emotion 5, 119-124. Marshall, P.J., Meltzoff, A.N., 2011. Neural mirroring systems: exploring the EEG mu rhythm in human infancy. Developmental cognitive neuroscience 1, 110-123. Master, S.L., Amodio, D.M., Stanton, A.L., Yee, C.M., Hilmert, C.J., Taylor, S.E., 2009. Neurobiological correlates of coping through emotional approach. Brain, behavior, and immunity 23, 27-35. McCall, C., Singer, T., 2012. The animal and human neuroendocrinology of social cognition, motivation and behavior. Nature neuroscience 15, 681-688. McCormick, L.M., Brumm, M.C., Beadle, J.N., Paradiso, S., Yamada, T., Andreasen, N., 2012. Mirror neuron function, psychosis, and empathy in schizophrenia. Psychiatry research 201, 233-239. McGurk, S.R., Meltzer, H.Y., 2000. The role of cognition in vocational functioning in schizophrenia. Schizophrenia research 45, 175-184.

8. REFERENCES
120
Mitchell, J.P., Macrae, C.N., Banaji, M.R., 2006. Dissociable medial prefrontal contributions to judgments of similar and dissimilar others. Neuron 50, 655-663. Mogenson, G.J., Jones, D.L., Yim, C.Y., 1980. From motivation to action: functional interface between the limbic system and the motor system. Progress in neurobiology 14, 69-97. Morris, P., Coady, H., 1974. The effect of need for approval and incentives upon children's probability learning responses. Child development 45, 862-865. Morris, R.W., Weickert, C.S., Loughland, C.M., 2009. Emotional face processing in schizophrenia. Current opinion in psychiatry 22, 140-146. Muthukumaraswamy, S.D., Johnson, B.W., 2004a. Changes in rolandic mu rhythm during observation of a precision grip. Psychophysiology 41, 152-156. Muthukumaraswamy, S.D., Johnson, B.W., McNair, N.A., 2004b. Mu rhythm modulation during observation of an object-directed grasp. Brain research. Cognitive brain research 19, 195-201. Nakagami, E., Xie, B., Hoe, M., Brekke, J.S., 2008. Intrinsic motivation, neurocognition and psychosocial functioning in schizophrenia: testing mediator and moderator effects. Schizophrenia research 105, 95-104. Nieuwenhuis, S., Yeung, N., Holroyd, C.B., Schurger, A., Cohen, J.D., 2004. Sensitivity of electrophysiological activity from medial frontal cortex to utilitarian and performance feedback. Cerebral cortex 14, 741-747. Oberman, L.M., Hubbard, E.M., McCleery, J.P., Altschuler, E.L., Ramachandran, V.S., Pineda, J.A., 2005. EEG evidence for mirror neuron dysfunction in autism spectrum disorders. Brain research. Cognitive brain research 24, 190-198. Oberman, L.M., Pineda, J.A., Ramachandran, V.S., 2007. The human mirror neuron system: a link between action observation and social skills. Social cognitive and affective neuroscience 2, 62-66. Ochsner, K.N., Beer, J.S., Robertson, E.R., Cooper, J.C., Gabrieli, J.D., Kihsltrom, J.F., D'Esposito, M., 2005. The neural correlates of direct and reflected self-knowledge. NeuroImage 28, 797-814. Ochsner, K.N., Lieberman, M.D., 2001. The emergence of social cognitive neuroscience. The American psychologist 56, 717-734. Oldfield, R.C., 1971. The assessment and analysis of handedness: the Edinburgh inventory. Neuropsychologia 9, 97-113. Park, S., Matthews, N., Gibson, C., 2008. Imitation, simulation, and schizophrenia. Schizophrenia bulletin 34, 698-707.

8. REFERENCES
121
Paulus, C., 2009. DER SAARBRÜCKER PERSÖNLICHKEITSFRAGEBOGEN SPF(IRI) ZUR MESSUNG VON EMPATHIE: Psychometrische Evaluation der deutschen Version des Interpersonal Reactivity Index. Universitäts- und Landesbibliothek, Saarbrücken. Penn, D.L., Corrigan, P.W., Bentall, R.P., Racenstein, J.M., Newman, L., 1997. Social cognition in schizophrenia. Psychological bulletin 121, 114-132. Penn, D.L., Keefe, R.S., Davis, S.M., Meyer, P.S., Perkins, D.O., Losardo, D., Lieberman, J.A., 2009. The effects of antipsychotic medications on emotion perception in patients with chronic schizophrenia in the CATIE trial. Schizophrenia research 115, 17-23. Perry, A., Bentin, S., 2009. Mirror activity in the human brain while observing hand movements: a comparison between EEG desynchronization in the mu-range and previous fMRI results. Brain research 1282, 126-132. Perry, A., Bentin, S., Bartal, I.B., Lamm, C., Decety, J., 2010a. "Feeling" the pain of those who are different from us: Modulation of EEG in the mu/alpha range. Cognitive, affective & behavioral neuroscience 10, 493-504. Perry, A., Bentin, S., Shalev, I., Israel, S., Uzefovsky, F., Bar-On, D., Ebstein, R.P., 2010b. Intranasal oxytocin modulates EEG mu/alpha and beta rhythms during perception of biological motion. Psychoneuroendocrinology 35, 1446-1453. Perry, A., Stein, L., Bentin, S., 2011. Motor and attentional mechanisms involved in social interaction--evidence from mu and alpha EEG suppression. NeuroImage 58, 895-904. Pfurtscheller, G., Lopes da Silva, F.H., 1999. Event-related EEG/MEG synchronization and desynchronization: basic principles. Clinical neurophysiology : official journal of the International Federation of Clinical Neurophysiology 110, 1842-1857. Picton, T.W., 1992. The P300 wave of the human event-related potential. Journal of clinical neurophysiology : official publication of the American Electroencephalographic Society 9, 456-479. Pierrehumbert, B., Torrisi, R., Laufer, D., Halfon, O., Ansermet, F., Beck Popovic, M., 2010. Oxytocin response to an experimental psychosocial challenge in adults exposed to traumatic experiences during childhood or adolescence. Neuroscience 166, 168-177. Pineda, J.A., 2005. The functional significance of mu rhythms: translating "seeing" and "hearing" into "doing". Brain research. Brain research reviews 50, 57-68. Pineda, J.A., Hecht, E., 2009. Mirroring and mu rhythm involvement in social cognition: are there dissociable subcomponents of theory of mind? Biological psychology 80, 306-314. Polich, J., Donchin, E., 1988. P300 and the word frequency effect. Electroencephalography and clinical neurophysiology 70, 33-45. Premack, D., Woodruff, G., 1978. Chimpanzee problem-solving: a test for comprehension. Science 202, 532-535.

8. REFERENCES
122
Pridmore, S., Brune, M., Ahmadi, J., Dale, J., 2008. Echopraxia in schizophrenia: possible mechanisms. The Australian and New Zealand journal of psychiatry 42, 565-571. Ramani, S.V., Quesney, L.F., Olson, D., Gumnit, R.J., 1980. Diagnosis of hysterical seizures in epileptic patients. The American journal of psychiatry 137, 705-709. Rao, R.P., Ballard, D.H., 1999. Predictive coding in the visual cortex: a functional interpretation of some extra-classical receptive-field effects. Nature neuroscience 2, 79-87. Raymaekers, R., Wiersema, J.R., Roeyers, H., 2009. EEG study of the mirror neuron system in children with high functioning autism. Brain research 1304, 113-121. Rigoni, D., Polezzi, D., Rumiati, R., Guarino, R., Sartori, G., 2010. When people matter more than money: an ERPs study. Brain research bulletin 81, 445-452. Rinck, M., Becker, E.S., 2007. Approach and avoidance in fear of spiders. Journal of behavior therapy and experimental psychiatry 38, 105-120. Rizzolatti, G., Craighero, L., 2004. The mirror-neuron system. Annual review of neuroscience 27, 169-192. Rizzolatti, G., Fadiga, L., Gallese, V., Fogassi, L., 1996. Premotor cortex and the recognition of motor actions. Brain research. Cognitive brain research 3, 131-141. Rizzolatti, G., Sinigaglia, C., 2010. The functional role of the parieto-frontal mirror circuit: interpretations and misinterpretations. Nature reviews. Neuroscience 11, 264-274. Robbins, T.W., Everitt, B.J., 1996. Neurobehavioural mechanisms of reward and motivation. Current opinion in neurobiology 6, 228-236. Roelofs, K., Elzinga, B.M., Rotteveel, M., 2005. The effects of stress-induced cortisol responses on approach-avoidance behavior. Psychoneuroendocrinology 30, 665-677. Roelofs, K., Minelli, A., Mars, R.B., van Peer, J., Toni, I., 2009. On the neural control of social emotional behavior. Social cognitive and affective neuroscience 4, 50-58. Roelofs, K., Putman, P., Schouten, S., Lange, W.G., Volman, I., Rinck, M., 2010. Gaze direction differentially affects avoidance tendencies to happy and angry faces in socially anxious individuals. Behaviour research and therapy 48, 290-294. Rosenfeld, A.J., Lieberman, J.A., Jarskog, L.F., 2011. Oxytocin, dopamine, and the amygdala: a neurofunctional model of social cognitive deficits in schizophrenia. Schizophrenia bulletin 37, 1077-1087. Rubin, L.H., Carter, C.S., Drogos, L., Pournajafi-Nazarloo, H., Sweeney, J.A., Maki, P.M., 2010. Peripheral oxytocin is associated with reduced symptom severity in schizophrenia. Schizophrenia research 124, 13-21.

8. REFERENCES
123
Sasayama, D., Hattori, K., Teraishi, T., Hori, H., Ota, M., Yoshida, S., Arima, K., Higuchi, T., Amano, N., Kunugi, H., 2012. Negative correlation between cerebrospinal fluid oxytocin levels and negative symptoms of male patients with schizophrenia. Schizophrenia research 139, 201-206. Saxe, R., Xiao, D.K., Kovacs, G., Perrett, D.I., Kanwisher, N., 2004. A region of right posterior superior temporal sulcus responds to observed intentional actions. Neuropsychologia 42, 1435-1446.
Schneider, F., Gur, R.C., Gur, R.E., Shtasel, D.L., 1995. Emotional processing in schizophrenia: neurobehavioral probes in relation to psychopathology. Schizophrenia research 17, 67-75.
Schuch, S., Bayliss, A.P., Klein, C., Tipper, S.P., 2010. Attention modulates motor system
activation during action observation: evidence for inhibitory rebound. Experimental
brain research. 205, 235-249.
Schultz, W., 2000. Multiple reward signals in the brain. Nature reviews. Neuroscience 1, 199-207. Schultz, W., Dickinson, A., 2000. Neuronal coding of prediction errors. Annual review of neuroscience 23, 473-500. Schurmann, M., Jarvelainen, J., Avikainen, S., Cannon, T.D., Lonnqvist, J., Huttunen, M., Hari, R., 2007. Manifest disease and motor cortex reactivity in twins discordant for schizophrenia. The British journal of psychiatry : the journal of mental science 191, 178-179. Schwartz, B.L., Mastropaolo, J., Rosse, R.B., Mathis, G., Deutsch, S.I., 2006. Imitation of facial expressions in schizophrenia. Psychiatry research 145, 87-94. Shamay-Tsoory, S.G., Shur, S., Barcai-Goodman, L., Medlovich, S., Harari, H., Levkovitz, Y., 2007. Dissociation of cognitive from affective components of theory of mind in schizophrenia. Psychiatry research 149, 11-23. Shipley, T.E., Jr., Veroff, J., 1952. A projective measure of need for affiliation. Journal of experimental psychology 43, 349-356. Sims, T.B., Van Reekum, C.M., Johnstone, T., Chakrabarti, B., 2012. How reward modulates mimicry: EMG evidence of greater facial mimicry of more rewarding happy faces. Psychophysiology 49, 998-1004. Singh, F., Pineda, J., Cadenhead, K.S., 2011. Association of impaired EEG mu wave suppression, negative symptoms and social functioning in biological motion processing in first episode of psychosis. Schizophrenia research 130, 182-186. Sobhani, M., Fox, G.R., Kaplan, J., Aziz-Zadeh, L., 2012. Interpersonal liking modulates motor-related neural regions. PloS one 7, e46809.

8. REFERENCES
124
Sperry, R.W., 1950. Neural basis of the spontaneous optokinetic response produced by visual inversion. Journal of comparative and physiological psychology 43, 482-489. Spielberger, C.D., Gorssuch, R.L., Lushene, P.R., Vagg, P.R., & Jacobs, G.A., 1983. Manual for the State-Trait Anxiety Inventory. Consulting Psychologists Press, Inc. Stancak, A., Jr., Pfurtscheller, G., 1996. The effects of handedness and type of movement on the contralateral preponderance of mu-rhythm desynchronisation. Electroencephalography and clinical neurophysiology 99, 174-182. Stone, T., Davies, M., 1995. Mental simulation : evaluations and applications. Blackwell, Oxford. Strathearn, L., 2011. Maternal neglect: oxytocin, dopamine and the neurobiology of attachment. Journal of neuroendocrinology 23, 1054-1065. Sugawara, S.K., Tanaka, S., Okazaki, S., Watanabe, K., Sadato, N., 2012. Social rewards enhance offline improvements in motor skill. PloS one 7, e48174. Tabachnick, B.G., Fidell, L.S., 2006. Using Multivariate Statistics, 5th ed.Pearson Education. Takahashi, H., Kato, M., Sassa, T., Shibuya, T., Koeda, M., Yahata, N., Matsuura, M., Asai, K., Suhara, T., Okubo, Y., 2010. Functional deficits in the extrastriate body area during observation of sports-related actions in schizophrenia. Schizophrenia bulletin 36, 642-647. Taylor, P.C., Walsh, V., Eimer, M., 2008. Combining TMS and EEG to study cognitive function and cortico-cortico interactions. Behavioural brain research 191, 141-147. Toomey, R., Schuldberg, D., Corrigan, P., Green, M.F., 2002. Nonverbal social perception and symptomatology in schizophrenia. Schizophrenia research 53, 83-91. Tremeau, F., 2006. A review of emotion deficits in schizophrenia. Dialogues in clinical neuroscience 8, 59-70. Uddin, L.Q., Iacoboni, M., Lange, C., Keenan, J.P., 2007. The self and social cognition: the role of cortical midline structures and mirror neurons. Trends in cognitive sciences 11, 153-157. Ulloa, E.R., Pineda, J.A., 2007. Recognition of point-light biological motion: mu rhythms and mirror neuron activity. Behavioural brain research 183, 188-194. Ullsperger, M., Nittono, H., von Cramon, D.Y., 2007. When goals are missed: dealing with self-generated and externally induced failure. NeuroImage 35, 1356-1364. Van Meel, C.S., Van Heijningen, C.A., 2010. The effect of interpersonal competition on monitoring internal and external error feedback. Psychophysiology 47, 213-222.

8. REFERENCES
125
van Schie, H. T., Mars, R. B., Coles, M. G., & Bekkering, H., 2004. Modulation of activity in medial frontal and motor cortices during error observation. Nature neuroscience, 7(5), 549-554. Varcin, K.J., Bailey, P.E., Henry, J.D., 2010. Empathic deficits in schizophrenia: the potential role of rapid facial mimicry. Journal of the International Neuropsychological Society 16, 621-629. Vogt, S., Taylor, P., Hopkins, B., 2003. Visuomotor priming by pictures of hand postures: perspective matters. Neuropsychologia 41, 941-951.
Volman, I., Roelofs, K., Koch, S., Verhagen, L., Toni, I., 2011. Anterior prefrontal cortex
inhibition impairs control over social emotional actions. Current biology 21, 1766-1770.
von Holst, E., and Mittelstaedt, H., 1950. Das Reafferenzprinzip (Wechselwirkungen
zwischen Zentralnervensystem und Peripherie). Naturwissenschaften 37, 464-476.
Waltz, J.A., Kasanova, Z., Ross, T.J., Salmeron, B.J., McMahon, R.P., Gold, J.M., Stein, E.A., 2013. The roles of reward, default, and executive control networks in set-shifting impairments in schizophrenia. PloS one 8, e57257. Walum, H., Lichtenstein, P., Neiderhiser, J.M., Reiss, D., Ganiban, J.M., Spotts, E.L., Pedersen, N.L., Anckarsater, H., Larsson, H., Westberg, L., 2012. Variation in the oxytocin receptor gene is associated with pair-bonding and social behavior. Biological psychiatry 71, 419-426. Wolpaw, J.R., Birbaumer, N., McFarland, D.J., Pfurtscheller, G., Vaughan, T.M., 2002. Brain-computer interfaces for communication and control. Clinical neurophysiology : official journal of the International Federation of Clinical Neurophysiology 113, 767-791. Wolpert, D.M., Doya, K., Kawato, M., 2003. A unifying computational framework for motor control and social interaction. Philosophical transactions of the Royal Society of London. Series B, Biological sciences 358, 593-602. Wolpert, D.M., Miall, R.C., 1996. Forward Models for Physiological Motor Control. Neural networks : the official journal of the International Neural Network Society 9, 1265-1279. Woodruff, C.C., Martin, T., Bilyk, N., 2011. Differences in self- and other-induced Mu suppression are correlated with empathic abilities. Brain research 1405, 69-76. Yeung, N., Holroyd, C.B., Cohen, J.D., 2005. ERP correlates of feedback and reward processing in the presence and absence of response choice. Cerebral cortex 15, 535-544. Yeung, N., Sanfey, A.G., 2004. Independent coding of reward magnitude and valence in the human brain. The Journal of neuroscience : the official journal of the Society for Neuroscience 24, 6258-6264. Yu, R., Zhou, X., 2006. Brain responses to outcomes of one's own and other's performance in a gambling task. Neuroreport 17, 1747-1751.

9. APPENDICES
126
Chapter 9
Appendices

9. APPENDICES
127
9.1 Curriculum Vitae
Elliot Clayton BROWN
Research Department of Cognitive Neuropsychiatry, LWL University Hospital
Alexandrinenstrasse 1-3, Bochum 44791, Germany
EDUCATION
2013 PhD in Neuroscience - International Graduate School of Neuroscience (IGSN), Ruhr University
Bochum, Germany (ongoing).
2008 MSc (Dist) in Clinical & Cognitive Neuroscience - Goldsmiths, University of London, UK.
2007 Advanced Certificate in Psychology - Birkbeck, University of London, UK.
2004 BSc (Hons) in Biological Sciences - University of Reading, UK.
2000 Higher National Certificate in Computing - Middlesex University, London, UK.
PROFESSIONAL APPOINTMENTS
2010-2013 Graduate Student - IGSN, Ruhr University Bochum, Germany. Supervised by Prof Dr Martin
Brüne (PhD title: “Social cognition and schizophrenia: Observing others’ actions, rewards and
errors”)
2012 Visiting Research Fellow - Dept of Psychiatry, Celal Bayar University Hospital, Manisa, Turkey.
Supervised by Prof Dr Aysen Esen-Danaci (2 months)
2011 Visiting Research Fellow - Dept of Experimental Clinical and Health Psychology, Ghent
University, Belgium. Supervised by Prof Roeljan J Wiersema & Prof Gilles Pourtois (2 months)
2009-2010 Research Assistant - HHSRI, School of Psychology, University of Hertfordshire, Herts, UK.
Supervised by Dr D John Done & Prof Anthony J Marcel.
2008-2009 Honorary Research Associate - Institute of Psychiatry, Kings College London, UK. Supervised
by Prof Anthony S David.
2008-2009 Research Assistant - Blackheath Brain Injury Rehabilitation Unit, Huntercombe Group,
London, UK (Visual awareness case-study). Supervised by Prof Gianna Cocchini
2008-2009 Assistant Psychologist - Neuropsychology Department, Huntercombe Group, Blackheath
Brain Injury Rehabilitation Unit, London, UK.
2007-2008 Healthcare Assistant - Maudsley Hospital, South London and Maudsley (SLaM) NHS Trust,
London, UK.

9. APPENDICES
128
TEACHING EXPERIENCE & SUPERVISION
2012 Student supervision of visiting medical student on project: “The EEG mu rhythm in patients with
schizophrenia” (RUB).
2011/2 Student supervision of visiting doctoral candidate on project: “EEG medial frontal negativities in
economic decision-making in schizophrenia and Borderline Personality Disorder” (RUB).
2011 Student supervision of MSc project: “The EEG P300 in patients with schizophrenia” (RUB).
2010 Student supervision of MSc project: “Sensitivity to social rejection in individuals with persecutory
delusions and a clinically anxious group” (UHerts).
2010 Student supervision of MSc projects: “Estimates and plausibility in an older population” (UHerts).
2010 Student supervision of BSc 3rd
year student group project: “Empathy in reaction time? - Inter- and
intra-individual sequential repetition effects in reaction time” (UHerts).
2010 Teaching of “Topics in psychopathology” module in BSc Psychology course (UHerts).
2010 Teaching Statistics for Social Sciences in Undergraduate Psychology Bridging Course (UHerts).
INVITED TALKS
“Living with schizophrenia.” (2013) Christ’s College Finchley, London, UK.
“Reward-related modulation of motor cortex during action observation in schizophrenia.” (2013) Maryland
Psychiatric Research Centre, University of Maryland Baltimore, USA.
“Reward-related modulation of motor cortex during action observation.” (2013) Action and Neurocognition
Group, Donders Institute for Brain, Cognition and Behaviour, Radboud University, Nijmegen, The Netherlands.
“Social cognition in schizophrenia.” (2012) Klinik für Kinder- und Jugendpsychiatrie, Psychotherapie und
Psychosomatik im LWL-Psychiatrieverbund Westfalen, Hamm, Germany.
“The neurobiology of social approach and avoidance behaviour in schizophrenia.” (2012) University Clinic and
Polyclinic for Psychiatry and Psychotherapy, Münster, Germany.
“Social approach and avoidance behaviour in schizophrenia.” (2012) The Psychosis Studies and Intervention
Team, Department of Psychiatry, The University of Hong Kong, Hong Kong.
“Social cognition in movement and the relevance to schizophrenia.” (2012) Yasar University, Izmir, Turkey.
AWARDS & GRANTS
2012 Scientific and Technological Research Council of Turkey (TUBITAK) - Research Fellowship for Foreign
Citizens.
2011 International Brain Research Organisation (IBRO) - CEERC-WERC InEurope Travel Fellowship.
2011 Ruhr University Bochum, Research School Graduate Funding.
2010 International Graduate School of Neuroscience (IGSN), Ruhr University Bochum - 3-year PhD
Scholarship.

9. APPENDICES
129
PROFESSIONAL MEMBERSHIPS
Schizophrenia International Research Society (SIRS) since 2011.
WORKSHOPS & SCHOOLS ATTENDED
2012 Toolkit of Cognitive Neuroscience 2012: Advanced analysis and source modelling of EEG and MEG data
- Donders Institute for Brain, Cognition and Behaviour, Nijmegen, Netherlands.
2011 ESCON Workshop on Social Neuroscience - Brussels / Ghent, Belgium.
2011 Autumn School 'Perception, Action and Control: Methods, Concepts and Challenges - Donders
Institute for Brain, Cognition and Behaviour - Nijmegen, Netherlands.
2010 Centre for Integrative Neuroscience and Neurodynamics Summer School - Reading, UK.
2010 Combined EEG & fMRI, and EEG & TMS, Brain Products workshop - Reading, UK.
2008 “Transcranial Magnetic Stimulation (TMS) in Plasticity and Rehabilitation”, MAGSTIM TMS Summer
School - Institute of Cognitive Neuroscience, University College London, UK.
2008 Neuroscience in Education Workshop, Institute of Cognitive Neuroscience - University College London,
UK.
AD-HOC REVIEWER FOR
Brain Imaging and Behaviour, Frontiers in Human Neuroscience, Journal of Neurology & Neurophysiology,
Psychological Reports, Schizophrenia Research.
LANGUAGES
English (Native) German (Basic) Cantonese (Intermediate)

9. APPENDICES
130
9.2 List of Publications
Peer-reviewed journals
1. Gonzalez-Liencres, C., Tas, C., Brown, E.C., Erdin, S., Onur, E., Cubukcuoglu, Z., Esen-Danaci, A., Brüne, M. (2013) Oxidative stress in schizophrenia: The effects on social cognition and neurocognition (in preparation).
2. Brown, E.C., Tas, C., Yilmaz, H., Esen-Danaci, A., Roelofs, K., Brüne, M. (2013). A role for resting-state frontal EEG activity as a trait marker for social approach and avoidance behaviour in schizophrenia (in preparation).
3. Tas, C., Brown, E.C., Onur, E., Aydin, O., Brüne, M. (2013). The effects of endogenous oxytocin levels on the perception of emotions in bipolar disorder (submitted).
4. Brown, E.C., Tas, C., Esen-Danaci, A., Roelofs, K., Brüne, M. (2013). Social approach and avoidance behaviour for negative emotions is modulated by endogenous oxytocin in schizophrenia (submitted).
5. Tas, C., Brown, E.C., Imak, S., Aydin, O., Esen-Danaci, A., Brüne, M. (2013). Cortisol response to psychosocial stress predicted by plasma oxytocin levels facilitates social functioning in schizophrenia (submitted).
6. Brown, E.C., Tas, C., Esen-Danaci, A., Brüne, M. (2013). The relationship between the subdomains of social cognition, social functioning and symptomatology in a clinically stable group of schizophrenia patients (under review).
7. Brune, M., Tas, C., Brown, E.C., Armgart, C., Dimaggio, G., Lysaker, P. (2013). Metakognitive und sozial-kognitive Defizite bei Schizophrenien. Funktionelle Bedeutung und Behandlungsstrategien. Zeitschrift für Psychiatrie, Psychologie und Psychotherapie (in press).
8. Brown, E.C., Brüne, M. (2013). Reward in the mirror neuron system, social context and the implications on psychopathology. Behavioral and Brain Sciences (in press).
9. Brown, E.C., Wiersema, J.R., Pourtois, G., Brüne, M. (2013). Modulation of motor cortex activity when observing rewarding and punishing actions. Neuropsychologia 51 (1), 52-58.
10. Tas, C., Brown, E.C., Cubukcuoglu, Z., Aydemir, O., Esen-Danaci, A., Brüne, M. (2013). Towards an integrative approach to understanding quality of life in schizophrenia: The role of neurocognition, social cognition and psychopathology. Comprehensive Psychiatry 54 (3), 262-268.
11. Brown, E.C., Brüne, M. (2012). Evolution of social predictive brains? Frontiers in Psychology 3, (414).
12. Tas, C., Brown, E.C., Esen-Danaci, A., Lysaker, P.H., Brüne, M. (2012). Intrinsic motivation and metacognition as predictors of learning potential in patients with remitted schizophrenia. Journal of Psychiatric Research 46 (8), 1086-1092.
13. Brown, E.C., Brüne, M. (2012). The role of predictive coding in social neuroscience. Frontiers in Human Neuroscience 6 (147).
14. Brown, E.C., Tas, C., Brüne, M. (2011). Potential therapeutic avenues to tackle social cognition problems in schizophrenia. Expert Review of Neurotherapeutics 12 (1), 71-81.

9. APPENDICES
131
Book chapters
15. Brown, E.C., Tas, C., Gonzalez-Liencres, C., Brüne, M. (2013) Neurological underpinnings of social cognition and metacognition in schizophrenia spectrum disorders. In P. Lysaker, G. Dimaggio and M. Brüne, Social cognition and metacognition in schizophrenia: Psychopathology and treatment approaches. San Diego, CA, USA: Academic Press / Elsevier (contract approved)
16. Tas, C., Brown, E.C., Gonzalez-Liencres, C., Brüne, M. (2013) Experimental usage of oxytocin to combat deficits in social cognition in schizophrenia. In P. Lysaker, G. Dimaggio and M. Brüne, Social cognition and metacognition in schizophrenia: Psychopathology and treatment approaches. San Diego, CA, USA: Academic Press / Elsevier (contract approved)
Published conference abstracts
17. Brown, E.C., Wiersema, J.R., Pourtois, G., Brüne, M. (2012). Catharsis in the motor cortex: Reward and punishment when observing others’ actions. Belgian Brain Congress, Liege, Belgium (Published in Frontiers in Neuroscience. (doi: 10.3389/conf.fnhum.2012.210.00052))
18. Brown E.C., Tas C., Danaci A.E., Brüne M. (2012). How separable are the domains of social cognitive deficits in schizophrenia? 3rd Schizophrenia International Research Conference, Florence, Italy (Published in Schizophrenia Research 136 (S1), 180 (doi: 10.1016/S0920-9964(12)70545-5))
19. Tas C., Brown E.C., Danaci A.E., Lysaker P., Brüne M. (2012). Motivation and metacognition as predictors of occupational functioning in remitted schizophrenia patients. 3rd Schizophrenia International Research Conference, Florence, Italy. (Published in Schizophrenia Research 136 (S1), 180 (doi: 10.1016/S0920-
9964(12)70544-3))
20. Brown, E.C., Brüne, M. (2011). Theory of mind in at-risk stages of schizophrenia. 3rd European Conference on Schizophrenia Research (ECSR): Facts and Visions, Berlin, Germany (Published in European Archives of Psychiatry and Clinical Neurosciences 261 (S1), S26 (TALK))
21. Brown, E.C., Brüne, M. (2011). Social cognition and violence in schizophrenia: Is there a link? 3rd European Conference on Schizophrenia Research (ECSR): Facts and Visions, Berlin, Germany (Published in European Archives of Psychiatry and Clinical Neurosciences 261 (S1), S39 (TALK))

9. APPENDICES
132
Other conference abstracts
22. Brown, E.C., Pourtois, G., Wiersema, J.R., Brüne, M. (2013). The influence of reward and punishment in action observation. UCL Conference on “Implications of Research on the Neuroscience of Affect, Attachment, and Social Cognition”, University College London, UK. (POSTER)
23. Brown, E.C., Brüne, M. (2013). Neural evidence for the potential use of rewards in observational learning in schizophrenia? Neurex Symposium on “Cognitive disorders and remediation in schizophrenia and other mental disorders”, Strasbourg, Switzerland. (TALK)
24. Brown E.C., Tas C., Danaci A.E., Brüne M. (2012). Social approach and avoidance in schizophrenia: The relationship with paranoia, social cognition and oxytocin. The German Society for Biological Psychiatry Congress (DGBP), Heidelberg, Germany. (POSTER)
25. Tas C., Brown E., Esen-Danacı A., Brüne M. (2012). The role of plasma oxytocin and cortisol levels and metacognition on social learning and social stress response in schizophrenia patients. The German Society for Biological Psychiatry Congress (DGBP), Heidelberg, Germany. (POSTER)
26. Brown, E.C., Pourtois, G., Wiersema, J.R., Tas, C., Brüne, M. (2012). Reward-related changes in motor cortex excitability during action observation of others. 8th FENS Forum of Neuroscience, Barcelona, Spain. (POSTER)
27. Brown, E.C., Brüne, M. (2011). Mirror neuron activity in schizophrenia. Deutsche
Gesellschaft für Psychiatrie, Psychotherapie, und Nervenheilkunde Kongress
(DGPPN), Berlin, Germany. (TALK)
28. Brown, E.C., Brüne, M. (2011). How ’social’ is the mirror neuron system: Insights from the EEG mu rhythm. Donders Discussions, Nijmegen, Netherlands. (POSTER)
29. Brown, E.C., Cocchini, G. (2010). Perception without awareness: A case study. International Conference on Parietal Lobe Function, European Science Foundation (ESF) Congress, Amsterdam, Netherlands. (POSTER)
30. Brown, E.C., Cocchini, G. (2009). A study of error awareness and insight in schizophrenia using the Sustained Attention to Response Task (SART). British Psychological Society (BPS) Annual Conference, Brighton, UK. (POSTER)
Other Publications
31. Tas, C., Brown, E.C., Enzi, B., Brüne, M. (2013). Social cognitive deficits in schizophrenia: Are they acquired or developmental? IGSN Report (in press).
(“in preparation” refers to manuscripts that are completed and are awaiting submission)

9. APPENDICES
133
9.3 Acknowledgements
I would like to dedicate this thesis to my beloved Godfather, Con Conway, who sadly
passed away in September 2012. May he rest in peace.
Firstly I would like to express my utmost gratitude to my Doktorvater, Martin Brüne, for
his positive support and guidance throughout my PhD. I wholeheartedly appreciate the
freedom and encouragement he provided me, which kept me motivated and focused.
Secondly, I would like to give a huge thanks to Cumhur Tas, as without his working
partnership, I would not have been half as productive in the last 3 years, or enjoyed my
work half as much. I feel that I have gained a life-long research collaborator and a new
brother.
I give my thanks to my mother, as without her unconditional and unwavering support
over the last few years (well, last 32 years to be precise), I would not have been able to
have the opportunity to follow my passion and my dream.
I would also like to give my thanks to Manu Schütze for her helpful feedback on my
thesis, but most of all, for all her loving support and care, which gave me strength to
carry on.
My thanks also go to Burak Erdeniz, for all the times we shared talking about the brain,
and everything related to it. His inspiration has been invaluable.
Many thanks also to Cristina Gonzalez for bringing lots of laughter and fresh new ideas
to our office.
I also want to extend my gratitude to my supervisors and collaborators that I have
worked with over the last 3 years, as without them, I would not have been able to write
this thesis. To Christian Bellebaum, Roeljan Wiersema, Gilles Pourtois, Aysen Esen-
Danaci and Karin Roelofs, thank you so much for all of your help and support. Thanks
also to my friends at the Psychology Department of Ghent University, and the Psychiatry
Department at Celal Bayar University Hospital, and particularly Duygu Kuzu for her
incomparable motivation and help with collecting data in Turkey.
I would also like to express my thanks to all my other friends and family along the way
for giving more meaning to my life.
Last, but not least, I would like to extend my appreciation to all of the people that kindly
participated in the studies presented here, as without them I would have no data.