soil biology and biochemistry - hidrobiologia.ufrj.br et al2018_sbb.pdf · contents lists available...
TRANSCRIPT

Contents lists available at ScienceDirect
Soil Biology and Biochemistry
journal homepage: www.elsevier.com/locate/soilbio
Prokaryotic assemblages within permafrost active layer at Edmonson Point(Northern Victoria Land, Antarctica)
Maria Papalea, Antonella Contea, Anu Mikkonenb, Luigi Michauda,1, Rosabruna La Ferlac,Maurizio Azzaroc, Gabriella Carusoc, Rodolfo Paranhosd, S. Cabral Andersond,Giovanna Maimonec, Alessandro Ciro Rappazzoc, Carmen Rizzoa, Nunziacarla Spanòa,Angelina Lo Giudicea,c,∗, Mauro Guglielmine
a Department of Chemical, Biological, Pharmaceutical and Environmental Sciences, University of Messina, Messina, ItalybDepartment of Biological and Environmental Sciences, University of Jyvaskyla, Jyvaskyla, Finlandc Institute for Coastal Marine Environment, National Council of Research (IAMC-CNR), Messina, Italyd Institute of Biology, Federal University of Rio de Janeiro, Prédio do CCS, Cidade Universitária do Fundao, Rio de Janeiro, Brazile Dipartimento di Scienze Teoriche e Applicate, University of Insubria, Varese, Italy
A R T I C L E I N F O
Keywords:Seasonally thawed active layerAntarcticaProkaryotic communityMetabolic activities
A B S T R A C T
This study was aimed at gaining insights on the prokaryotic community (in terms of both taxonomic compositionand activities) inhabiting the active layer at Edmonson Point, an ice-free area on the eastern slope at the foot ofMount Melbourne (Northern Victoria Land, Antarctica). Samples were collected during the thawing period,when microbial physiological activities are restored to utilize previously frozen organic substrates. Despite thevery small cell sizes (< 0.1 μm3), indicating the occurrence of stressed, dormant and/or starved cells, the pro-karyotic communities appeared to be metabolically active in the decomposition of high molecular weight(> 600 Da) substrates, as indicated also by the obtained rates of enzymatic hydrolytic activities over proteolytic,glycolitic and phosphoric compounds. Taxonomical composition showed that Proteobacteria, Actinobacteria andFirmicutes dominated the prokaryotic community, with most of their members playing crucial roles in organicmatter turnover, as well as nitrogen cycling, or entering a viable but not cultivable state to cope with con-tinuously changing environmental conditions, such as in the case of the active layer. Finally, non-autochthonousbacteria (mainly of marine origin) were detected and they probably contribute to the organic matter turnoverwithin such cold terrestrial habitat.
This research provides the first comprehensive account of the prokaryotic communities inhabiting theAntarctic permafrost and contributes to existing information on the response of their abundance and metabolismin a permafrost area that undergoes to seasonal changes (e.g. in terms of temperature, water availability and icepresence).
1. Introduction
Polar regions are severely vulnerable to the climate change and arelikely to be more susceptible by global warming than other regions(Ganzert et al., 2011). Warmer temperatures could cause not only in-land ice retreat, but also prokaryotic community shifts (which mayenhance microbial invasion, establishment of cosmopolitan genotypesand loss of endemic taxa) which in turn may impact on the ecosystembiogeochemistry and functioning (Frank-Fahle et al., 2014; Kleinteichet al., 2017). Thus, scientific interest towards such complex microbialcommunities is increasing more and more aiming at understanding/
predicting how they react to current and future environmental changes.Most soils in and around the Arctic and Antarctic regions (as well asthose located in high latitudes) are affected by permafrost. This latter ischaracterized by a near-surface seasonally thawed zone called “activelayer” where ground thaws during the summer and freezes again in thefollowing winter (French et al., 2009). In Continental Antarctica, thethickness of the active layer (ALT) undergoes extensive spatial varia-tions (0–160 cm; Vieira et al., 2010; Guglielmin and Cannone, 2012;Guglielmin et al., 2014), and has increased of 1 cm per year (Guglielminand Cannone, 2012), mainly due to the increase of the incoming ra-diation and the reduction of the lasting snow cover. Among the ice-free
https://doi.org/10.1016/j.soilbio.2018.05.004Received 31 July 2017; Received in revised form 2 May 2018; Accepted 6 May 2018
∗ Corresponding author. Istituto per l’Ambiente Marino Costiero, Consiglio Nazionale delle Ricerche (IAMC-CNR), Spianata S. Raineri 86, 98122, Messina, Italy.
1 Posthumous.E-mail address: [email protected] (A. Lo Giudice).
Soil Biology and Biochemistry 123 (2018) 165–179
0038-0717/ © 2018 Published by Elsevier Ltd.
T

areas of the Continental Antarctica, Edmonson Point is quite peculiarbecause it is an “oasis” in which is possible to find the larger patches ofmosses of all the Victoria Land that in some places can form a carpetthicker than 5 cm. Moreover it is one of the few places in ContinentalAntarctica in which ground water flow through the active layer hasbeen documented. In addition it is also one of the places where snowpatches occur largerly and melt earlier because the black color of thebasalts and the volcanic ash outcropping absorbed large amount ofsolar radiation. The active layer is thus characterized by harsh condi-tions in terms of prolonged subzero temperatures (at least 340 days peryear), strong temperature fluctuations, wide water content fluctuationswith very long dry periods and poor nutrient availability, and long-termbackground radiation exposure (e.g. Steven et al., 2006). Nevertheless,most ecological, hydrological, and biogeochemical activities take placewithin such extreme ecological niche (Kane et al., 1991; Hinzman et al.,2003), which is colonized by cold-adapted psychrotolerant rather thantrue psychrophilic microbial communities (Steven et al., 2009; Wagnerand Liebner, 2009; Wilkins et al., 2013). Such microorganisms possesspeculiar physiological flexibility and adopt efficient survival mechan-isms (e.g. the development of resilience/resistance mechanisms, lowmetabolic rates or stages of anabiosis or dormancy) over geologicaltimes to cope with extreme environmental conditions (Steven et al.,2006, 2007; Ponder et al., 2008; Wagner, 2008).
Permafrost thawing due to the ALT thickening has global implica-tions for carbon biogeochemical cycling since it restores microbialphysiological activities (Gilichinsky et al., 2008) and microorganismsare the main drivers of carbon mineralization (Mann et al., 2014). The
mineralization of organic matter and the release of previously frozencarbon as carbon dioxide and methane in thawing permafrost areamong the most important potential feedbacks from terrestrial ecosys-tems to the atmosphere (Dutta et al., 2006; Rodionow et al., 2006;Zimov et al., 2006; Schuur et al., 2008, 2009). However, the currentknowledge of the activity, structure and ecology of the microbialcommunities and their influence on carbon dynamics and ecosystemstability within the active layer remain poorly understood and mainlylimited to investigations of methane-cycling communities of Arctic andAlpine regions (Martineau et al., 2010; Yergeau et al., 2010, 2012;Graef et al., 2011; Goordial et al., 2016).
The active layer is particularly sensitive to ecological changes inresponse to climate seasonal and long-term variations. Accordingly, themicrobial populations inhabiting are supposed to play a key role in theecosystem biogeochemistry and functioning. In particular, during thethawing period previously frozen organic substrates are released, be-coming again available, thus allowing the recovery of microbial phy-siological activities In this context, two main questions are: i) Whichkinds of microbial activities occur in a freshly-thawed permafrost activelayer? ii) Which are the main prokaryotic phylogenetic groups playingsuch activities? To find answers to these questions, during a thawingperiod (Antarctic summer) the prokaryotic communities inhabiting thepermafrost active layer of an underexplored Antarctic area, likeEdmonson Point, were analyzed. This is a wide ice-free area on theeastern slope at the foot of Mount Melbourne (Antarctica). which waschosen as the study site owing to its particular surface and subsurfacecharacteristics. The site - designated Antarctic Specially Protected Area
Fig. 1. Sampling site.
M. Papale et al. Soil Biology and Biochemistry 123 (2018) 165–179
166

(ASPA 165) - beside of the occurrence of Adélie penguins and southpolar skuas colonies, is characterized by diverse terrestrial and fresh-water ecosystems exhibiting biologically diverse habitats alongmoisture and nutrient gradients. Here, a combined analytical approach,which included the estimation of prokaryotic abundances and meta-bolic activities, as well as the community structure and diversity, wasapplied. To integrate information gained by the application of the next-generation sequencing, the cultivable heterotrophic bacterial diversitywas also analyzed.
2. Materials and methods
2.1. Site description and sample collection
Edmonson Point (coordinates: Latitude 74°19′44.2″S- Longitude165°07′59.7″E) in the Northern Victoria Land is a volcanic area close toMt. Melbourne (Fig. 1). The area is characterized by alluvial sedimentsand weathered basaltic outcrops with some ponds and not far penguinrookeries. Here, soils are quite rich in organic content (0.5–1.6% C org)with respect to the surrounding areas and are Aquic Haplorthels over-lying continuous permafrost (Cannone and Guglielmin, 2009; Cannoneet al., 2008). Temperature generally ranges between −30.3and + 18.6 °C, with an amplitude of about 49 °C (Cannone andGuglielmin, 2009). Active layer thickness (ALT) ranges between 26 and81 cm in relation to the different percentage of mosses cover and watercontent (generally higher than 25%; Cannone et al., 2008; Cannone andGuglielmin, 2009) and the different yearly climatic conditions.
The soil samples (4 replicates) were collected directly from thedeeper part (35 cm in depth; just above the permafrost table) of theactive layer at Edmonson Point during the maximum thawing period inearly January 2014. Here, water is unable to drain through the un-derlying permafrost zone, thus remaining trapped and favouring mi-crobial activities. The samples were shipped to the IAMC CNR ofMessina in a thermally insulated box and maintained at −20 °C untillab processing (approx. four months). Sample handling and processingwere carried out aseptically (under sterile conditions) in a dedicatedlaminar flow hood to avoid contamination. The core samples (5 cm indiameter) were subsampled using a sterile disposable chisel, taking onlyinternal fragments by sterile forceps and processed according to stan-dard techniques developed for sediments.
2.2. Phylogenetic composition of the bacterial community
2.2.1. DNA extraction and PCR amplificationDNA was extracted in duplicate from 5 g of each soil sample by
employing the MoBio PowerSoil DNA isolation kit (MoBio Laboratories,Carlsbad, CA, USA) according to the manufacturer's instructions. DNAconcentrations and purity were quantified by using a NanoDrop ND-1000 UV–vis Spectrophotometer (NanoDrop Technologies, USA).
2.2.2. 16S rRNA gene amplicon sequencingThe V1-V2 region of the bacterial 16S rRNA genes was amplified by
PCR. In order to reduce bias in massive sequencing, the two-step PCRprotocol was applied, consisting in a first step of 30 PCR cycles withconventional PCR primers and then using 0.5 μL of first reaction am-plicon for 6 cycles PCR with barcoded primers for Ion Torrent se-quencing. Triplicate PCR reactions of 40 μL were set up at 0 °C under aPCR cabin by using 0.4 μL of Phusion High-Fidelity DNA polymerase(2U μL −1; Thermo Scientific), 8 μL of Phusion buffer (5×), 1 μL of eachdNTP (10mM), 1 μL of SybrGreenI 1:10 000, 1 μL of each primer(10 μM). The universal bacterial primers 27f (5′-AGAGTTTGATCCTGGCTCAG-3′) and 338r (5′-GCT GCCTCCCGTAGGAGT-3′) were used.
The amplification was performed according to the program de-scribed below: denaturation 98 °C for 30 s; annealing 30 cycles at 98 °Cfor 10s, 53 °C for 30 s and 72 °C for 60 s; elongation 72 °C for 10min.Amplified products were visualized by agarose (1.5%, w/v) gel
electrophoresis, using ethidium bromide (EtBr) (1 mgmL−1).The three reactions were pooled and used for the second PCR under
the same conditions. To 0.5 μL of pre-amplified DNA, the followingcomponents of the PCR mixture [0.2 μL of Phusion polymerase (2UμL−1; Thermo Scientific), 4 μL of Phusion buffer, 0.5 μL dNTPs(10mM), 0.5 μL of Sybr, 0.5 μL of each barcoded primer (10 μM)] wereadded to a final volume of 20 μL. The reaction was carried out ac-cording to the program described below: denaturation 98 °C for 30 s;annealing 6 cycles at 98 °C for 10 s, 53 °C for 30 s and 72 °C for 60 s;elongation 72 °C for 10min. The amplified products were visualized bygel electrophoresis as described above, then purified using theAgencourt Ampure XP (Beckman Coulter, Inc.) according to the man-ufacturer's instructions, and quantified using the Qubit fluorometer 2.0(ThermoFisher Scientific). Products were pooled in equimolar con-centrations. Sequencing was performed on an Ion Torrent PersonalGenome Machine™ using the Ion PGM Sequencing 400 Kit and the Ion314™ chip (all Ion Torrent reagents by Thermo Fischer Scientific) fol-lowing manufacturer's protocols.
2.2.3. Post-run analysis of the bacterial communityThe raw data were analyzed using the bioinformatics analysis
software Mothur (version 1.39.5). Barcodes and primers were identifiedwith maximum one base error and trimmed off. Reads were cleaned bylength (reads shorter than 200 bp were discarded) and by quality scoreusing score quality windows (i.e. average 25 and size 10). Remainingsequences were aligned with the Silva reference files (release 123 Fulllength sequences and taxonomy references). Reads were denoised usingthe pre. cluster command in Mothur platform (Schloss et al., 2011) toremove sequences that were likely due to pyrosequencing errors andassemble reads which differed only by 2 bp. Chimeric sequences wereidentified and removed (Edgar et al., 2011). Finally the sequences wereclassified against the same Silva database (Quast et al., 2013) and werecreated the distance matrix (label 0.03) to generate the OTU table forthe subsequently analysis. Shannon diversity index (H’) for each sam-pling site was calculated on the total number of good quality readsusing the Mothur software.
2.3. Estimation of prokaryotic cell abundance and morphology
For the estimation of total prokaryotic abundance and activities,0.5 g of permafrost samples were aseptically treated with the additionof 5 μL of Tween 80 (T80; 10%) and then of tetrasodium pyrophosphate(TSPP; 10mM), following sonication (at 75%, of ultrasounds frequency,3 times for 1min) and finally centrifugation at 800 rpm for 1min(Danovaro et al., 2001; Duhamel and Jacquet, 2006; La Ferla et al.,2017).
2.3.1. Image analysisFor the estimation of prokaryotic cell abundance by Image Analysis
(IA), together with cell size and shape, extracted samples were im-mediately fixed with filter-sterilized formaldehyde (0.22 μm porosity;final conc. 2%) and stored at 4 °C in the dark. The samples were filteredin four replicates through polycarbonate black membranes (porosity0.22 μm; GE Water & Process Technologies) and stained for 10–20minwith 4’,6-diamidino-2-phenylindole (DAPI, Sigma, final concentration10 μgmL−1) according to Porter and Feig (1980). A Zeiss AXIOPLAN 2Imaging (magnification: Plan-Neofluar 100× objective and 10×ocular; HBO 100W lamp; filter sets: G365 exciter filter, FT395 chro-matic beam splitter, LP420 barrier filter) equipped with the digitalcamera AXIOCAMHR (Zeiss) and AXIOVISION 3.1 software for thesubsequent morphometric analysis was used. Details of the technicalfeatures and calibration are reported in La Ferla et al. (2015) and re-ferences therein. Results were expressed in cells g−1 of wet permafrost.
The volume (VOL, expressed in μm3) of individual cell was derivedfrom two linear dimensions (width, W, and length, L) manually ob-tained. Curved objects were drawn by curve spline. VOL of a single cell
M. Papale et al. Soil Biology and Biochemistry 123 (2018) 165–179
167

was calculated according to the geometrical formula adopted by LaFerla et al. (2012) and references therein, i.e. VOL = (π/4)•W2•(L-W/3)(for coccal forms, W=L), assuming that the cells are cylindricalstraight rods with hemispherical or, in the case of coccoid forms,spherical caps. The cell shape was operationally defined as cocci if theirlength and width differed by less than 0.10 μm, coccobacilli if theirlength and width differed by more than 0.10 μm, and rods if their lengthwas at least double their width; V-shaped, C-shaped and S-shaped cellswere defined as vibrios, curved rods and spirillae, respectively.
2.3.2. Flow cytometryThe extracted samples were fixed with sterile paraformaldehyde at
2% (final concentration), incubated for 30min at 4 °C and then frozenat −20 °C. After thawing, four replicates were extracted with sodiumpyrophosphate (Danovaro et al., 2001; Duhamel and Jacquet, 2006),then centrifuged and each supernatant was diluted 10× and thenstained with Sybr Green I (at a final concentration 5×10−4 of thecommercial stock solution; Molecular Probes). The prokaryotic cellswere then analyzed using a FACSCalibur flow cytometer (BD Bios-ciences) equipped with 488 nm argon laser, as described by Gasol andDel Giorgio (2000) and Andrade et al. (2003). Results were expressed incells g−1 of wet permafrost.
2.4. Quantification of viable and respiring cells
2.4.1. Live/dead assayThe viability of prokaryotic cells - in terms of membrane integrity -
was analyzed using the Molecular Probes™ LIVE/DEAD Bac LightBacterial Viability Kits that utilizes SYTO 9 (green-fluorescent nucleicacid stain) and propidium iodide (red-fluorescent nucleic acid stain).Stock solutions were prepared for each dye using 5mL of samples andthe final concentration of each dye was 6 μM SYTO 9 stain and 30 μMpropidium iodide. The two solutions were mixed thoroughly and in-cubated at 4 °C in the dark for 1 h. After filtration of 1mL of mixedsolutions through polycarbonate black membranes, the above referredprocedure for DAPI staining was applied. The microscope cell countswere performed on four replicas using the specific sets for fluoresceine(BP450-490; FT510; LP520) and rhodamine (BP546/12; FT580;LP590). Results were expressed in cells g−1 of wet permafrost.
2.4.2. 5-Cyano-2,3-ditolyl tetrazolium chloride (CTC) assayThe BacLight Redox Sensor CTC Vitality Kit (Molecular probes™)
was adopted to quantify the prokaryotic respiring cells (CTC+).Aliquots of 1 mL of sample was added to 0.1 mL of CTC (5-Cyano-2,3-ditolyl tetrazolium chloride, final concentration 5 mM). The solutionwas incubated at 4 °C in the dark for 5 h (La Ferla et al., 2017) duringwhich respiring cells absorbed and reduced CTC into an water-in-soluble, red-fluorescent formazan product. The incubation was stoppedwith 0.22 μm porosity filter-sterilized formalin (final concentration2%). For each sample, a killed control was prepared by injecting for-malin before CTC addition. Slides were prepared in four replicates ac-cording to the DAPI procedure earlier described using the rhodaminespecific filter set (BP546/12; FT580; LP590). Counts were expressed incells g−1 of wet permafrost.
2.5. Metabolic potentials and ectoenzymatic activities
2.5.1. Physiological profilesBiolog-Ecoplate™ (BE) microplates were used to determine the
functional diversity of EP active layer and the physiological profiles ofmicrobial assemblage by differences in the metabolic potential.Following the standard procedure of Garland and Mills (1991), 96-wellmicrotiter plates, containing 31 carbon sources (namely, complexcarbon sources, carbohydrates, phosphate-carbon compounds, car-boxylic acids, amino acids, amines) and a control in triplicate togetherwith the redox dye tetrazolium violet, were inoculated with 150 μL of
sample treated with phosphate buffered saline (PBS). Each plate wasthen incubated at 4 °C in the dark. The oxidation of the carbon sourcesproduced formazan whose absorbance was quantified at 590 nm byusing a microplate-reader spectrophotometer (MICROTITER ELX-808,Bio Whittaker, Inc.) equipped with an Automatic Microplate Readerand the specific software (WIN KQCL) for data processing. Opticaldensity (OD) was measured after inoculation (at time T0), after 1 day(T1) and thereafter once a week (T7, T14, T21).
The color development in each plate was expressed according toSala et al. (2005), as the averaged substrate color development (ASCD)i.e. ASCD= Σ [(R-C)/31] where R is the averaged absorbance of thethree wells with substrate; C is the averaged absorbance of the controlwells (without substrate). The absorbance percentages of each substratewere determined according to Sala et al. (2006) and the 2% absorbanceof the total absorbance per plate was used as threshold for substrateutilization.
2.5.2. Potential rates of ectoenzymatic activitiesPotential rates of ectoenzymatic activities by the microbial assem-
blage on proteinaceous (leucine-aminopeptidase, LAP) and glucidic (β-glucosidase, β-GLU) organic matter, and on organic phosphates (alka-line-phosphatase, AP) were estimated using specific fluorogenic sub-strates, which are methylcoumarine (MCA) or methylumbelliferone(MUF)-derivates and are analogues to natural compounds, according tothe Hoppe (1993) method. Particularly, L-leucine-4-methylcoumar-inylamide hydrochloride, 4-methylumbelliferyl ß-D-glucopyranosideand 4-methylumbelliferyl phosphate (Sigma) were used for LAP, β-GLUand AP measurements. Permafrost samples were suspended in steriledistilled water in a 1:10 (w/v) ratio and vortexed; the supernatantobtained after sedimentation was used as the enzymatic extract for thesuccessive assays. Increasing amounts of each substrate were added totriplicate 5-mL sub-volumes of each sample and the fluorescence re-leased by substrate hydrolysis was measured with a Turner TD-700model fluorimeter, at 0 time (initial time) and 3 h after incubation at+5 °C. Calibration curves with the standards 7-amino-4-methylcou-marin (MCA) or 4-methylumbelliferone (MUF) were performed for LAPor for β-GLU and AP, respectively. Enzymatic measurements were re-ported in terms of maximum reaction velocity (Vmax) of hydrolysis ofthe substrates, namely as nanomoles of Leucine, Glucoside and PO4
potentially released per gram and per hour, by LAP, β-GLU and AP,respectively, taking into account the initial dilution of each sample.
As a measure of the affinity of each enzyme for its respective sub-strate, the Michaelis-Menten constant (Km) was calculated; the lower isthe Km value, the higher is the affinity of the enzyme for its substrate(Caruso, 2010). Specific enzymatic activity per cell (i.e. cell-specificactivity) was also calculated by dividing activity rates by the cellabundance determined by IA.
To get information on enzymatic stoichiometry, according toSinsabaugh et al. (2008, 2014), the ratios of C-acquiring to P-acquiringenzymes [ln(β-GLU):ln(AP)], C-acquiring to N-acquiring enzymes [ln(β-GLU): ln(LAP)] and N-acquiring to P-acquiring enzymes [ln(LAP): ln(AP)], were also calculated.
2.6. Enumeration, isolation and phylogenetic characterization of cultivableheterotrophic bacteria
2.6.1. Viable countsCulturing was carried out to obtain further and complementary
details on the microbial community inhabiting the permafrost activelayer at Edmonson Point. Two strategies were used to initially cultivatethe permafrost microorganisms: i) direct plating of diluted cell sus-pensions on agar media; and ii) enrichment in liquid media beforeplating (Vishnivetskaya et al., 2000), as specified below. If not other-wise specified, cultures in liquid and solidified media were incubated inthe dark in a thermostat at 4 °C for 8 weeks, and colony forming units(CFU) per gram of wet soil samples were calculated as averages of
M. Papale et al. Soil Biology and Biochemistry 123 (2018) 165–179
168

duplicate plates.For direct plating of soil samples, material (1 g) from the centre of
the core was tenfold diluted in cold filter-sterilized PBS 1×. Each di-lution (100 μL) was spread-plated in duplicate on Tryptic Soy Agar(TSA100; Oxoid) and R2A (Difco) at full strength.
For broth enrichments, 50 μL of soil suspension were added in du-plicate to 2.5mL of Tryptic Soy Broth (Oxoid) that was used at full(TSB100%), 1/2 (TSB50%) and 1/100 (TSB1%) strength. After incubation,enrichments were then diluted and aliquots (100 μL) plated on the samesolidified (1.5% agar, w/v) TSB used for the enrichment (i.e. TSA100,TSA50 or TSA1) and R2A.
2.6.2. Bacterial isolationColonies were randomly selected from agar plates used for CFU
counts, picked and subcultured almost three times under the sameconditions.
2.6.3. 16S rRNA gene PCR amplification of bacterial isolatesPCR-amplification of 16S rDNA from bacterial isolates was carried
out under the conditions described earlier (Michaud et al., 2004).Briefly, a single colony of each strain was lysed by heating at 95 °C for10min. Amplification of 16S rRNA gene was performed with an ABI9600 thermocycler (PE, Applied Biosystems) using the domain Bacteria-specific primers 27F (5′-AGAGTTTGATCCTGGCTCAG-3′) and 1492R (5′-CTACGGCTACCTTGTTACGA-3′). The reaction mixtures were as-sembled at 0 °C and contained 1–10 ng DNA, 10× buffer, 1.5 mMMgCl2, 150 ng of each forward and reverse primer (MWG, Germany),250 μM dNTP (Polymed, Italy), 0.5 units of PolyTaq polymerase(Polymed, Italy) and sterile distilled water to a final volume of 20 μL.Negative controls for DNA extraction and PCR setup (reaction mixturewithout a DNA template) were also used in every PCR run. The PCRprogram was as follows: denaturation 3min at 95 °C, followed by 30annealing cycles of 1min at 94 °C, 1min at 50 °C, 2min at 72 °C and afinal extension step of 10min at 72 °C.
The products of the amplification reactions were run in agarose gelelectrophoresis (1%, w/v) in TAE buffer (0.04M Tris-acetate, 0.02Macetic acid, 0.001M EDTA), containing 1 μgmL −1 of ethidium bro-mide.
2.6.4. Sequencing and analysis of 16S rRNA geneAutomated sequencing of 16S rRNA gene from isolates was carried
out by cycle sequencing using the dye terminator method. Sequencingwas carried out at the Sequencing Service of the Macrogen Laboratory(The Netherlands). The closest relatives of isolates were determined bycomparison to 16S rRNA gene sequences in the NCBI GenBank and theEMBL databases using BLAST (Altschul et al., 1997), and the
“Seqmatch” and “Classifier” programs of the Ribosomal Database Pro-ject II (http://rdp.cme.msu.edu/). Sequences were further aligned usingthe program Clustal W (Thompson et al., 1994) to the most similarorthologous sequences retrieved from database. Each alignment waschecked manually, corrected and then analyzed using the Neighbour-Joining method (Saitou and Nei, 1987) according to the model of Jukes-Cantor distances. The phylogenetic tree was constructed using theMEGA 5 (Molecular Evolutionary Genetics Analysis) software (Kumaret al., 1993). The robustness of the inferred trees was evaluated by 500bootstrap re-samplings.
The bacterial isolates are part of the Italian Collection of AntarcticBacteria (CIBAN) of the National Antarctic Museum (MNA, www.mna.it) “Felice Ippolito” kept at the University of Messina. They are cur-rently maintained on TSA slopes at 4 °C and routinely streaked on agarplates from tubes every six months to control purity and viability. TheAntarctic strains are also preserved by freezing cell suspensions at –80 °C in TSB to which 20% (v/v) glycerol is added.
2.6.5. Nucleotide sequence accession numbersThe nucleotide sequences have been deposited in the GenBank da-
tabase under the accession nos. KT965154 (isolate D26), KT965159(isolate B23), KT965161 (isolate F25), KT965163 (isolate P3),KT965164 (isolate A5), KT965170 (isolate A2), KT965172 (isolateA29).
3. Results
3.1. Phylogenetic composition of the bacterial community
The Ion Torrent analysis of the V1-V2 region of the bacterial 16SrRNA gene produced a total of 16087 sequences. After the quality checkwithin the RDP pyrosequencing pipeline and the removal of chimeras,2245 high-quality sequences were obtained. The number of high-quality reads per sample ranged from 1109 to 1136. The Shannon di-versity H′ index for all samples ranged between 3.60 and 4.94. Thediversity index and the observed richness (OTUs) (between 667 and710) showed in the replicates a general symmetric pattern.
3.1.1. Bacterial taxaA total of 1219 OTUs belonging to 22 different bacterial phyla were
from the active layer samples (11 of them in percentage< 1%) with thepredominance of the Proteobacteria and Actinobacteria (27.3 and 18.4%of total sequences, respectively), followed by the Acidobacteria,Nitrospirae, Chloroflexi and Bacteroidetes, which ranged between 3.6 and7.8% of total sequences (Fig. 2). The Firmicutes (2.6%), Gemmatimona-detes (2.5%), Parcubacteria (1.9%), Latescibacteria (1.7%), Cyanobacteria
Fig. 2. Phylogenetic affiliation of Antarctic bacteria as determined by the Ion Torrent analysis.
M. Papale et al. Soil Biology and Biochemistry 123 (2018) 165–179
169

(1.5%) constituted a minor component. Differences in relative abun-dances were observed for sequences affiliated to proteobacterial classesas they were mainly referred to the Alpha- (11.6%) and Betaproteo-bacteria (7.2%), whereas the Delta- (4.9%) and Gammaproteobacteria(2.8%) were less represented. A number of sequences (13.3% of totalsequences) were not identified at phylum level.
3.1.2. Bacterial generaOf the total high-quality bacterial sequences, more than 50 genera
were detected from the rest, ranging from 0.1 to 3.1% of total sequences(Table 1). Only six genera occurred at ≥1% of the total bacterial se-quences and were related to the Firmicutes (i.e. genus Lactobacillus;2.0%), Actinobacteria (i.e. genera Propionibacterium and Marmoricola;2.4 and 1.2%, respectively), Alphaproteobacteria (genus Variibacter;1.1%), Chloroflexi (i.e. genus Roseiflexus; 1.0%) and Nitrospirae (i.e.genus Nitrospira; 3.1%).
3.2. Prokaryotic cell abundances and morphometric analysis
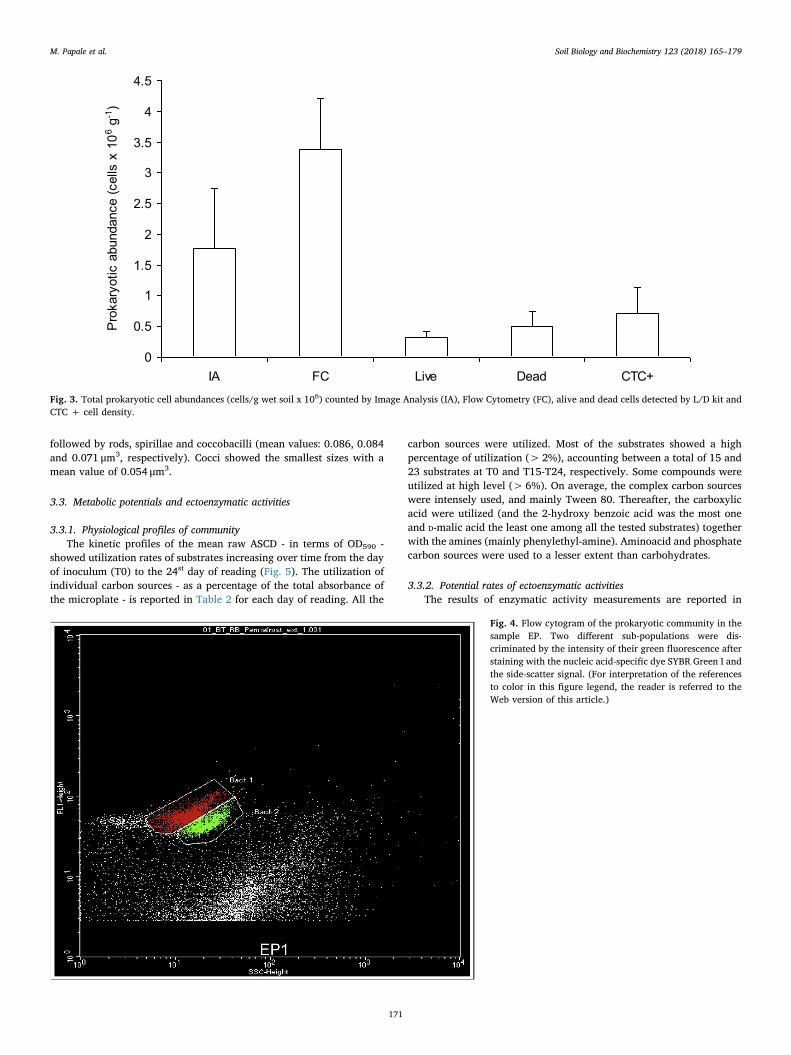
The total prokaryotic cell abundances (counted by Image Analysis,
IA, and Flow Cytometer, FC), the alive and dead cells detected by L/Dkit and the CTC + cell density are reported in Fig. 3. The total pro-karyotic cell abundances were in the order of 106 cells g−1 soil. How-ever, different mean values were detected between IA and FC,amounting at 1.7 ± 0.98×106 and 3.4 ± 0.83×106 cells g−1, re-spectively. The FC analyses clearly pointed out the occurrence of twosub-populations with different apparent DNA contents, as shown inFig. 4.
The total Live and Dead cells abundances were 3.21 ± 1.10× 105
and 5.10 ± 2.42×105 cells g−1, respectively, and alive cells ac-counted for only the 39% of the total count by L/D (ratios L/D=0.6).Respiring cells by CTC amounted to a mean value of7.18 ± 4.21× 105 cells g−1and CTC + accounted for 41% of totalcount.
Cell volume (VOL) widely ranged from 0.017 to 0.883 with a meanvalue of 0.089 ± 0.094 μm3. Cell morphologies mainly included rodsand cocci (37 and 34% of the total, respectively) and in lower per-centages coccobacilli, curved rods, vibrios, and spirillae (11, 9, 7 and2% of the total, respectively). Curved rods and vibrios accounted for thegreatest volumes (mean values: 0.179 and 0.174 μm3, respectively),
Table 1Bacterial genera retrieved within the active layer at Edmonson Point.
M. Papale et al. Soil Biology and Biochemistry 123 (2018) 165–179
170
followed by rods, spirillae and coccobacilli (mean values: 0.086, 0.084and 0.071 μm3, respectively). Cocci showed the smallest sizes with amean value of 0.054 μm3.
3.3. Metabolic potentials and ectoenzymatic activities
3.3.1. Physiological profiles of communityThe kinetic profiles of the mean raw ASCD - in terms of OD590 -
showed utilization rates of substrates increasing over time from the dayof inoculum (T0) to the 24st day of reading (Fig. 5). The utilization ofindividual carbon sources - as a percentage of the total absorbance ofthe microplate - is reported in Table 2 for each day of reading. All the
carbon sources were utilized. Most of the substrates showed a highpercentage of utilization (> 2%), accounting between a total of 15 and23 substrates at T0 and T15-T24, respectively. Some compounds wereutilized at high level (> 6%). On average, the complex carbon sourceswere intensely used, and mainly Tween 80. Thereafter, the carboxylicacid were utilized (and the 2-hydroxy benzoic acid was the most oneand D-malic acid the least one among all the tested substrates) togetherwith the amines (mainly phenylethyl-amine). Aminoacid and phosphatecarbon sources were used to a lesser extent than carbohydrates.
3.3.2. Potential rates of ectoenzymatic activitiesThe results of enzymatic activity measurements are reported in
0
0.5
1
1.5
2
2.5
3
3.5
4
4.5
IA FC Live Dead CTC+
Pro
kary
otic
abu
ndan
ce (c
ells
x 1
06 g-1
)
Fig. 3. Total prokaryotic cell abundances (cells/g wet soil x 106) counted by Image Analysis (IA), Flow Cytometry (FC), alive and dead cells detected by L/D kit andCTC + cell density.
Fig. 4. Flow cytogram of the prokaryotic community in thesample EP. Two different sub-populations were dis-criminated by the intensity of their green fluorescence afterstaining with the nucleic acid-specific dye SYBR Green I andthe side-scatter signal. (For interpretation of the referencesto color in this figure legend, the reader is referred to theWeb version of this article.)
M. Papale et al. Soil Biology and Biochemistry 123 (2018) 165–179
171

Table 3, where the kinetic parameters (Vmax and Km) of the enzymatichydrolysis, the cell-specific enzymatic activities and the reciprocalmolar ratios of the enzymatic activities are shown.
The mean Vmax values, of leucine aminopeptidase (LAP), β-gluco-sidase (β-GLU) and alkaline phosphatase (AP), showed that enzymatic
activities were in the order AP> β-GLU > LAP, being 1199.2, 71.7and 70.1 μmol g−1 h−1, respectively. The Km values indicated a sig-nificantly higher (p < 0.01) substrate affinity for LAP compared to AP;β-GLU activity showed affinity for polysaccharides at intermediate le-vels (Km=4.39 ± 0.05). The distribution patterns of enzymatic
Fig. 5. Averaged substrate color development (ASCD) with time in the EP sample. The plot of ASCD represented the mean color response for all 96 response wells ineach plate. (For interpretation of the references to color in this figure legend, the reader is referred to the Web version of this article.)
Table 2The utilization of individual carbon sources - as a percentage of the total absorbance of the microplate in EP active layer soil.
T0 T1 T8 T15 T24
complex carbon source c1 Tween 40 4.277 4.173 4.039 3.220 2.085d1 Tween 80 8.336 7.705 9.632 4.889 5.927e1 α-cyclodextrin 1.885 2.809 0.984 2.882 4.393f1 Glycogen 1.269 1.625 5.179 0.863 2.740
carbohydrates g1 D-cellobiose 4.639 3.311 5.075 2.415 3.716h1 α-D-lactose 3.806 2.127 2.434 0.000 0.000a2 β-methyl-D-glucoside 0.942 0.522 0.829 0.000 0.000b2 D-xylose 2.972 4.655 3.936 6.534 5.957c2 i-erythritol 1.957 2.970 2.330 4.352 4.460d2 D-mannitol 4.168 3.471 0.021 3.920 3.507e2 N-acetyl-D-glucosamine 3.516 3.672 1.036 2.485 3.142
phosphate-carbon g2 Glucose-l-phosphste 1.414 4.193 3.055 2.893 3.031h2 D,l-α-glycerol phosphate 0.029 3.652 0.207 2.602 0.000
carboxylic acid b1 Pyruvic acid mèthyl ester 0.217 2.528 0.000 4.294 1.862f2 D-glucosamic acid 3.733 4.193 6.318 6.020 5.712a3 D-galactonic acid γ-lactone 2.863 1.685 3.625 0.012 0.000b3 D-galactonic acid 0.217 2.047 0.000 2.205 4.788c3 2-hydroxy benzoic acid 21.276 10.092 16.572 8.330 5.600d3 4-hydroxy benzoic acid 0.109 3.812 1.036 5.332 5.190e3 γ-hydroxy butyric acid 1.522 2.990 0.031 5.682 3.902f3 Itaconic acid 1.015 0.482 2.382 3.057 6.188g3 α-ketobutyric acid 3.298 6.360 2.900 7.479 5.503h3 D-malic Acid 0.000 0.000 0.000 0.432 0.000
amino acids a4 L-arginine 1.559 1.786 1.657 0.000 0.000b4 L-asparagine 1.631 1.465 0.363 1.995 2.420c4 L-phenylalanine 1.377 1.705 1.243 4.970 2.986d4 L-serine 1.957 2.187 3.263 1.727 3.187e4 L-threonine 2.247 1.685 5.230 4.900 7.067f4 Glycyl-L-glutamic acid 4.277 4.113 7.820 4.329 3.895
amines g4 Phenylethyl-amine 8.844 5.317 7.302 2.182 2.822h4 Putrescine 4.966 2.669 2.071 0.000 0.082
Number of percentages<2: 16 9 13 8 82-4: 7 13 9 11 124-6: 5 6 4 8 9>6: 3 3 5 4 2
M. Papale et al. Soil Biology and Biochemistry 123 (2018) 165–179
172

activities did not reflect those depicted by the values of the living andactively respiring cells. Cell specific enzymatic activities, obtained byscaling the enzymatic rates to the total prokaryotic abundance de-termined by IA, showed similar rates for LAP and β-GLU, while highercell-specific AP were measured, due to high AP activity. The similarVmax values found for LAP and β-GLU were reflected by a C/N ratioclose to 1 as well as in similar C/P and N/P ratios, which were lowerthan 1.
3.4. Enumeration, isolation and phylogenetic characterization of cultivableheterotrophic bacteria
3.4.1. Viable countsFollowing direct plating on both R2A and TSA100 plates and in-
cubation at 4 °C, the active layer sample yielded bacterial concentra-tions of 0.6× 103 and 20.4× 103 CFU g−1 of wet soil, respectively.After enrichment in TSA at different strengths viable counts were fourto six orders of magnitude higher than those obtained by direct plating.Bacterial colonies grew on both R2A (between 7.5× 107 and64.0×109 CFU g−1 of wet soil) and TSA plates (between6.9×107 CFU g−1 of wet soil on TSA1, and 4.1× 109 CFU g−1 of wetsoil on TSA100) (Table 4).
3.4.2. Isolation and phylogenetic characterization of Antarctic bacteriaA total of 68 bacterial strains were isolated from Edmonson Point.
Among them, 14 isolates were obtained by direct plating (ten and fourfrom TSA100 and R2A, respectively). The remaining 54 isolates wereretrieved by the enrichment procedure: 33 from R2A, ten from TSA100,
nine from TSA50, and two from TSA1, respectively.Cultivable bacteria mainly belonged to Firmicutes (39 isolates) and
Actinobacteria (16 isolates), followed by Gamma- (11 isolates), andBetaproteobacteria (two isolates) (Table 5). All sequences with similarity≥97% were considered to represent one phylotype and were groupedinto Operational Taxonomic Units (OTUs). A total of seven phylotypeswere obtained, which five of them that included more isolates (OTUs 1,5, 6, 8 and 11). As shown in Fig. 6, the Actinobacteria formed twodistinct clusters: the first one included Arthrobacter spp. (isolates P3 andA2, and OTU11), while the second cluster included the genus Micro-bacterium (isolate A29). The Firmicutes were all referred to the genusBacillus (39 isolates in the OTU1). The 11 Gammaproteobacteria isolateswere related to the genus Psychrobacter (OTU8), whereas the Betapro-teobacteria included two Polaromonas isolates (OTU6).
4. Discussion
Most studies on microbial diversity in polar environments have fo-cused on soils, permafrost, ice, lakes and marine environments (i.e.Aislabie et al., 2006, 2009; Mosier et al., 2007; Baldi et al., 2010; LoGiudice et al., 2012; Michaud et al., 2012; Pearce et al., 2012;Gugliandolo et al., 2016; Makhalanyane et al., 2016 and referencestherein), while the microbial community structure in permafrost-af-fected soils has only been seldom examined and mainly in the Arctic(i.e. Wagner et al., 2009; Wilhelm et al., 2011; Frank-Fahle et al., 2014;Ganzert et al., 2014; Deng et al., 2015; Hultman et al., 2015). The activelayer is an ecological niche colonized by diverse and functionally cold-adapted microbial assemblages, which fit themselves to the seasonalchanges of temperature, water availability and ice presence. Accordingto Makhalanyane et al. (2016), data on microbial diversity, abundanceand functional processes in polar soils are, at the current stage, largelylacking. Further, the relationships among them remain often ambiguousand deserve to be investigated more in depth.
In this context, this study was aimed at gaining insights on theprokaryotic community occurring within the active layer at EdmonsonPoint, an ice-free area on the eastern slope at the foot of MountMelbourne (Northern Victoria Land, Antarctica). Samples were col-lected during the thawing period, when microbial physiological activ-ities are restored to utilize previously frozen organic substrates.Recently, Hultman et al. (2015), combining several molecular ‘omics’approaches, determined the phylogenetic composition of the microbialcommunities and their functional potential and activity in Alaskan soilsrepresenting different states of thaw. Consistently with other reports(Yergeau et al., 2010; Mackelprang et al., 2011), the seasonally thawedactive layer exhibited both more species and functional diversity thanthe other two soils (i.e. permafrost and thermokarst bog).
Furthermore, we applied a combined analytical approach that in-cluded the estimation of prokaryotic abundances and metabolic activ-ities, and the analysis of the community structure and diversity, asdetermined by culture-dependent and culture-independent methods. Asit was expected, the culturable fraction of the bacterial community didnot reflect that found by pyrosequencing, highlighting once again thewell-known discrepancy between these two approaches (Møller et al.,2013). However, their synergistic application allowed obtaining com-plementary information on the microbial community inhabiting Ant-arctic permafrost-affected soil at Edmonson Point. Even if cultivabletechniques dramatically underestimate microbial numbers and com-position in a sample, the major advantage of these approaches over themodern molecular techniques lies in the fact that they furnish bacterialisolates that can be exploited in further studies on microbial physiologyand biotechnology (Lo Giudice and Fani, 2015).
Our study analyzed samples from just one site and thus our findingscannot be considered representative of Antarctic permafrost-affectedsoils in general. However, overall the composition of the bacterial as-semblages resembled to that found elsewhere, with Proteobacteria,Actinobacteria, Acidobacteria, Bacteroidetes, Planctomycetes,
Table 3Mean values ± standard deviation (sd) of the kinetic parameters (Vmax andKm), cell-specific enzymatic activities and the reciprocal molar ratios of theelements potentially mobilized by the enzymatic activities in the permafrostactive layer at Edmonson Point.
Parameters Ectoenzyme Mean ± sd
Vmax LAP 70.09 ± 0.37(μmol g−1 h−1) AP 1199.25 ± 0.12
β−GLU 71.67 ± 0.05
Km LAP 2.94 ± .0.60(μmol g−1 h−1) AP 112.90 ± 3.52
β−GLU 4.39 ± 0.05
Cell-specific LAP 4.12 ± 0.02 E−05activity AP 7.05 ± 0.03 E−04(μmol h−1 cell−1) β−GLU 4.22 ± 0.02 E−05
C/P 0.60 ± 0.11C/N 1.01 ± 0.03N/P 0.59 ± 0.07
Table 4Viable counts from the active layer at Edmonson Point (EP) (before and afterenrichment).
Samplea Viable counts on (CFUg−1 of wet soil)b
R2A TSA100 TSA50 TSA1
EP 0.6×103 20.4×103
EP (TSB1) 64.0× 109 6.9× 107
EP (TSB50) 7.5×107 7.2× 107
EP (TSB100) 2.8×109 4.1× 108
a EP: natural sample, not enriched; EP (TSB1):sample enriched in TSB at 1%strength;EP (TSB50):sample enriched in TSB at 50% strength;EP(TSB100):sample enriched in TSB at 100% strength.
b TSA100, TSA50 and TSA1:plates of TSA at full, 50% and 1% strength, re-spectively.
M. Papale et al. Soil Biology and Biochemistry 123 (2018) 165–179
173

Table 516S rRNA gene sequence affiliation to their closest phylogenetic neighbors of Antarctic isolates.
Next relative by GenBank alignment (ANa, organism) RIb ANa OTUc Isolation medium Treatmentd Hom (%)e Origin of next relative organism N° of isolates
BetaproteobacteriaKC433619, Polaromonas sp. L19.9 B23 KT965159 6 TSA1 TSB1 98 Antarctica 2GammaproteobacteriaJQ684240, Psychrobacter cryohalolentis strain HWG-
A17F25 KT965161 8 TSA50 TSB50 100 Permafrost soil, Tibet Plateau 11
ActinobacteriaFN377733, Arthrobacter sp. SH-61B P3 KT965163 na TSA100 DP 99 Marine sediment, Svalbard 1GU176043, Arthrobacter sp. HY11 A2 KT965170 na R2A DP 97 Antarctic seawater 1JN662538, Arthrobacter sulfonivorans A5 KT965164 11 R2A DP 99 Soils, Tianshan Glacier 11KR007639, Microbacterium sp. SPO40 A29 KT965172 5 R2A TSA1 100 Polar oceanic water 3FirmicutesKF818647, Bacillus simplex strain 265XG8 D26 KT965154 1 TSA100 TSB100 100 Alpine grassland 39
a AN: Accession Number.b RI: representative isolate.c Na: not assigned.d Treatment: DP, direct plating.e Hom: sequence homology.
Fig. 6. Rooted phylogenetic tree calculated by Jukes-Cantor distance estimation algorithm showing affiliation of isolates to closest-related sequences from eithercultivated or cloned bacteria. Percentages of 400 bootstrap resampling that supported the branching orders in each analysis are shown above or near the relevantnodes. The tree was outgrouped with 16S rRNA gene sequence of Methanocaldococcus jannaschii DSM 2661.
M. Papale et al. Soil Biology and Biochemistry 123 (2018) 165–179
174

Chloroflexi and Verrucomicrobia that appear to be frequent when usingculture-independent approaches, while Proteobacteria, Actinobacteria,Bacteroidetes and Firmicutes have been isolated by cultivation-basedmethods (Shivaji et al., 2004; Aislabie et al., 2006; Hansen et al., 2007;Steven et al., 2007; Frank-Fahle et al., 2014; Ganzert et al., 2014; Denget al., 2015; Hultman et al., 2015). The main bacterial compositionobserved in the Edmonson Point active layer reflects the main processes(e.g. biogeochemical cycling, degradation of high-molecular weightcompounds) occurring in this extreme habitat under complex and harshenvironmental conditions. Proteobacteria play crucial roles in globalcarbon, nitrogen, and sulphur cycling, thus resulting really relevant tothe geochemical process of Antarctic soils (Rampelotto et al., 2015) andbeing among the major players of degradation. The successful coloni-zation of the Edmonson Point active layer by Firmicutes and Actino-bacteria (as previously reported by several authors for polar soils; e.g.Shivaji et al., 2004; Aislabie et al., 2006; Steven et al., 2007; Gilichinskyet al., 2007; Liu et al., 2009; Wagner et al., 2009; Jansson and Tas2014) is dependent on their ability to form spores (e.g. Bacillus mem-bers) or enter a metabolically quiescent but viable state, respectively, toadapt to stress in harsh environments (in terms of low temperature, UVradiation, heat, and desiccation), so that they can revive and growunder favourable conditions in a continuous changing habitat, such asthe active layer (Jansson and Tas 2014). Both Firmicutes and Actino-bacteria can metabolize a wide array of substrates as sole carbon source,with significant implications on the carbon cycle in Antarctica soils(Aislabie et al., 2006; Rampelotto et al., 2015). Finally, Bacteroidetes,which include several psychrophilic members commonly found inpermafrost-affected soil ecosystems (Steven et al., 2008; Wagner et al.,2009), were less represented in the Edmonson Point active layer.However, they were generally positively correlated with substrates richin organic carbon (Fierer et al., 2007) and it could be supposed that, astypical decomposers, they could contribute to the turnover of polymericcarbon in the Edmonson Point soil, together with other found groups(Fazi et al., 2005; Aislabie et al., 2009).
Further, the input of organic substrates due to moss vegetation andpenguin rookeries (Ganzert et al., 2011) in the Edmonson Point areacan sustain the nitrogen cycling. This was confirmed by the occurrenceof a high number of sequences related to the genus Nitrospira (amongNitrospirae) and Variibacter (among Alphaproteobacteria) (Lee et al.,2016). Conversely, Acidobacteria are known to have an oligotrophiclifestyle, so high nutrient content might be a limiting factor for mem-bers of this phylum. This result was in agreement with a previous studyby Yergeau et al. (2012), who reported that a higher nutrient avail-ability due to global warming could determine an increase in the Al-phaproteobacteria:Acidobacteria ratio in Antarctic soils. Acidobacteriabelonging to subgroup 4, such as Blastocatella spp. detected in thisstudy, are able to utilize complex substrates. In addition, a number ofphylogenetic lineages detected in the present study (e.g. Actinobacteria,Bacteroidetes, Firmicutes, Chloroflexi, Chlorobi, Cyanobacteria and Pro-teobacteria) include nitrogen-fixing members. As suggested by Ganzertet al. (2011), these autotrophic microorganisms might play a key-role inestablishing a functional microbial food web in cold soils, together withdetected CO2-fixing bacteria from the phyla Chloroflexi, Chlorobi, Cya-nobacteria, Firmicutes and Proteobacteria (in the subphyla Alpha, Beta-,and Gamma-).
The bacterial community was mainly composed by membersstrongly related to nearest phylogenetic neighbors that have generallybeen retrieved in cold habitats (e.g. permafrost soils and glaciers), alsolocated in polar areas, thus suggesting their occurrence at both Poles.This finding is not surprising as similar observations have been oftenreported in polar microbiology studies (e.g. Pearce et al., 2007; LoGiudice et al., 2012; Michaud et al., 2012; Sul et al., 2013; Papale et al.,2017). Our results also highlighted the potential influence of externalinputs, mainly of marine origin. For example, among the Gram-negativeisolates Psychrobacter cryohalolentis is a new species from Siberianpermafrost which was first described by Bakermans et al. (2006).
Psychrobacter species are cold-adapted and halo-tolerant, and they havebeen isolated from a variety of low-temperature (under frozen condi-tions, below 0 °C) marine environments (including Antarctic sea ice,ornithogenic soils and sediments, invertebrates, seawater).
In Edmonson Point other bacteria that are generally associated withthe marine environment were also detected, thus indicating a probableexternal input by marine aerosols and/or birds (mainly penguins andskuas) visiting the site, as previously observed for maritime Antarctica(Papale et al., 2017). This was the case of the two Polaromonas isolateswhich were first isolated from sea ice brine (Irgens et al., 1996). Thisfinding suggests that microorganisms of marine origins could find afavourable environment for their growth in the active layer (Bakermanset al., 2014). In particular, the genus Polaromonas is widely distributedworldwide, probably depending on air-dispersion process, and gen-erally dominates in polar and high-elevation environments, mainly inglacial ice and sediment samples (Darcy et al., 2011). Interestingly, itspresence has been reported in cold substrates, such as the active layer atEdmonson Point (Frey et al., 2010; Darcy et al., 2011; Michaud et al.,2012; Franzetti et al., 2013).
Furthermore, in this study we attempted to relate structure withfunction by analyzing in parallel the phylogenetic analysis by both theBIOLOG assay and the determination of enzymatic activity rates.Although the applied methods did not allow to link a specific phylotypeto a certain activity, the obtained data are fundamental to establish thestatus of microbial communities, which represent the main drivers ofbiogeochemical processes in polar ecosystems. The overall metabolicresponse (as determined by the BIOLOG assay) of the analyzed micro-bial population resulted low. This finding could be attributable to en-vironmental constrains strictly linked to the main characteristics of theactive layer (e.g. extreme climatic conditions in terms of temperaturefluctuations, extreme cold temperature, drying and wetting cycles andfreezing-thawing cycles), since stress is recognized to inhibit or reducemicrobial community functionality (Gryta et al., 2014; Jałowiecki et al.,2016), mostly in Antarctica (Gilichinsky et al., 2008). The bestexploited carbon sources were the polymeric substances, carboxylicacids and amines. These finding are in line with results obtained forpermafrost affected soils in North-Eastern Siberia (Wagner et al., 2009),in Antarctic soils (Kenarova et al., 2013) and in deep Antarctic per-mafrost (La Ferla et al., 2017) where potential metabolic activity to-ward Tween® 80 (polyoxyethylene sorbitan mono-oleate - a nonionicsurfactant and easily degradable antifreeze substance), benzoic acids (acommon metabolite detected in almost all microbial habitats) andphenolic compounds (synthesized by mosses as antioxidants or allelo-pathic agents) have been detected. Differently to Wagner et al. (2009)findings in Siberian permafrost horizons, only the D-xylose was wellused among carbohydrates by the active layer microbial community,while in accordance with La Ferla et al. (2017) and Wagner et al.(2009), amino acids were not intensively metabolized.
The enzymatic activity measurements carried out in the EdmonsonPoint active layer pointed out that the microbial communities played asignificant role in the degradation of high molecular weight substrates.This ability in such a cold terrestrial habitat appeared to be strictlylinked to the production of an efficient battery of cold-active extra-cellular enzymes, which provided psychrophilic microbes with anadequate and prompt uptake of nutrients during the thawing period, asalso observed by previous studies (Vazquez et al., 2004; Margesin andMiteva, 2011; La Ferla et al., 2017). The prevalence of AP over β-GLUand LAP, depicted by the enzymatic distribution patterns, suggestedthat in the Edmonson Point active layer the microbial communitieswere able to degrade preferentially organic phosphates compared topolysaccharides and proteins. Since phosphate cannot be synthesizedby microbes, AP, which hydrolyzes phosphate esters into inorganicphosphate is an important enzyme for phosphorus regeneration; itssynthesis and activity is under bacterial regulation, especially undernutrient deficiency (Dhaked et al., 2005). In the Edmonson Point activelayer, however, high AP activity was measured even without a direct
M. Papale et al. Soil Biology and Biochemistry 123 (2018) 165–179
175

relationship with bacterial abundance, suggesting for this enzyme a notstrictly bacterial origin, such as from algae or fungi. Moreover, in ter-restrial ecosystems of maritime Antarctica, Zhu et al. (2014) reportedthat organic phosphorus compounds from penguin guano or seal ex-creta contributed significantly to phosphorus cycle, increasing AP ac-tivity. Together with AP, extracellular proteases are among the mostwidespread hydrolytic enzymes. As the polymers degradation by ex-tracellular enzymes is a limiting step in organic matter metabolism(Caruso, 2010), proteases such as LAP play a crucial ecological role inorganic matter recycling. In the Edmonson Point, the similar activitylevels of β-GLU and LAP suggested that these two enzymes equallycontributed to the decomposition of organic substrates within the activelayer. Moreover, the Km values indicated that, in spite of the high ac-tivity rates, AP attached its substrates (organic phosphoric esters) witha low affinity, while comparatively higher was the affinity of both LAPand β-GLU for their reciprocal substrates (i.e. proteins and poly-saccharides, respectively). As high substrate affinity is considered as astrategy to support bacterial growth under low nutrient conditions(Rath et al., 1993), such finding suggested that the prokaryotic com-munity in the permafrost active layer was actively metabolizing C- andN-organic compounds to get both these elements for its needs.
Cell specific enzymatic activities reflected the same magnitudeorder as that measured for the overall microbial assemblage, suggestingthat at single cell level LAP and β-GLU were potentially working atsimilar activity rates. According to Sinsabaugh et al. (2008, 2014), thestoichiometric ratios between the enzymes, in terms of molar ratios,provide an index of metabolic activities directed towards the acquisi-tion of organic N (such as LAP) and P (such as AP) with respect to C. Inthe examined active layer sample, the N/P ratio, close to 1, indicatedthat the amount of nitrogen and the one of phosphorus potentiallymobilized by microorganisms through their enzymatic activities werecomparable. Shifts in enzyme ratios also can be associated to changes inelement-use efficiency (Sinsabaugh and Follstad Shah, 2012). In theEdmonson Point sample, N/P ratio was higher than C/P and C/N ones,indicating that phosphorus was preferentially mobilized with respect tocarbon.
A survey on prokaryotic abundances and morphometry was alsocarried out as, at current stage, few papers exist on the quantification ofthe prokaryotes in permafrost active layer (e.g. Hansen et al., 2007;Gilichinsky et al., 2007; Ponder et al., 2008; Blanco et al., 2012). Mostof the studies focused on the viable cell counts rather than on the directcell counts. In the present study, the cultivation on the oligotrophicmedium R2A was the best method for the recovery of both the highestcolony number and diversity from Edmonson Point active layer. In linewith previous observations (Vishnivetskaya et al., 2000), full strengthmedia showed a wider variability in colony morphology. The pre-liminary liquid enrichment of samples resulted in a pronounced in-crease in the viable counts and, in turn, it allowed obtaining highernumbers of bacterial isolates. It is possible that enrichment treatmentfacilitated the recovery of cells from a viable but non-culturable state(Steven et al., 2006). The direct plating showed an amount of103 CFU g−1 on R2A and TSA media in accordance with Soina et al.(2004) for Antarctic permafrost samples. Our results on viable abun-dances are in line with those obtained for Siberian and Arctic perma-frost habitats and they are also similar to tundra and ornithogenic soils(e.g. Ramsay and Stannard, 1986; Rivkina et al., 1998).
As determined by DAPI-staining, the order of 106 cells g−1 washigher than that found in the Antarctic Dry Valleys permafrost(Gilichinsky et al., 2007; La Ferla et al., 2017). In the same order(106 cells g−1) but higher than IA, the FC analysis underlined the ex-istence of two different microbial subpopulations, as previously ob-served in Boulder Clay (Northern Victoria Land) permafrost samples (LaFerla et al., 2017).
The prokaryotic community revealed cell volumes relatively small(mean value of 0.089 ± 0.094 μm3). The occurrence of cell sizessmaller than 0.1 μm3 has often been associated with life cycle changes
of stressed, starved, dormant cells in extremely cold and salty en-vironments (Ponder et al., 2008; Kuhn et al., 2014). These assumptionswere confirmed by the predominance of Gram-positives (i.e. Firmicutesand Actinobacteria) among bacterial isolates and by the low metabolicresponse obtained by BIOLOG assay. As concerns the cell morphologies,the numeric prevalence of rods and cocci confirmed the few determi-nations performed by Bölter (2003) in Arctic dry and mesic sites. Coccishowed the smallest sizes, while curved rods and vibrios accounted forthe greatest volumes.
The percentage of the living cells was lower than the dead ones (L/Dratio) as well as the percentage of respiring cells by CTC resulted lowerthan that of total cells. The comparison with the results obtained indeep Antarctic permafrost of Victoria Valley by La Ferla et al. (2017)showed that lived or respiring cells overwhelmed dead cells below the60–74 cm and 265–275 cm depth horizon, respectively. The increase ofviability with increasing depth in permafrost can be explicated by theactive layer proper characteristics. In fact, it is an open system subduedto high climate variability in terms of both thermal and water regimes,and samples were collected during the Antarctic summer, when thehigh U.V. radiation and the drying conditions strongly stressed lifecondition of the microbial community (Smith et al., 2009; Pèrez et al.,2017).
5. Conclusion
The permafrost active layer is severely affected by climate changes,resulting in microbial community (main actor in the biogechemicalprocesses) structural shifts with repercussions on the ecosystem func-tioning. In this study we analyzed the prokaryotic community of anAntarctic permafrost-affected soils (in the Edmonson Point area) duringthe thawing period, when microbial physiological activities are restoredto utilize previously frozen organic substrates. The application of dif-ferent methodologies allowed us obtaining a reasonably consistentsurvey of microbial diversity, activities, and abundances in the in-vestigated area. Even if single time-point analyses were carried out, ourfindings provide, at best, a baseline for future research aimed at as-sessing the effects of environmental change. As suggested byMakhalanyane et al. (2016), taxonomic and functional data could beintegrated into climatic models to better understand the role of climatein dictating changes in the soil microbial community structure andfunction, as well as the contributions of these communities to climate-linked processes.
Interestingly, even if the very small cell size (< 0.1 μm3) suggestedthe occurrence of stressed and/or starved cells, the prokaryotic com-munities appeared to be particularly involved in the decomposition ofhigh molecular weight substrates, thanks to the production of cold-ac-tive extracellular enzymes. The phylogenetic analysis suggested that themajor players of organic matter turnover might be Proteobacteria,Actinobacteria and Firmicutes, with most of their affiliates playing cru-cial roles in biogeochemical cycles, such as the nitrogen one. In addi-tion, such taxonomic groups included species that, by entering a viablebut not cultivable state, are able to survive and cope with extreme andcontinuous changing environmental conditions, such as in the case ofthe active layer. Further, the active layer at Edmonson Point hosts non-autochthonous bacteria which mainly derive from the next marineenvironments (i.e. through bird drop or marine aerosol) and even find afavourable environment for their growth, probably contributing to theorganic matter turnover within such cold terrestrial habitat.
In conclusion, consistently with previous reports, the environmentalconditions of the permafrost-affected soil at Edmonson Point sustain amultifaceted and metabolically active microbial community, capable ofutilizing a wide spectrum of substrates, thus endorsing the great role ofprokaryotes in the emission and removal of carbon and other biogeo-chemical elements acting as a pulsating pump in relation to climatevariability.
M. Papale et al. Soil Biology and Biochemistry 123 (2018) 165–179
176

Conflicts of interest
The authors declare they have no conflict of interests.
Acknowledgements
This work was supported by grants from the National AntarcticResearch Program (PNRA), Italian Ministry of Education and Research(Research Project PNRA 2013/AZ1.05) and from CNR Short TermMobility (STM AMMCNT. CNR prot. N. 0058167–02/09/2015). TheAuthors thank to all the staff at “Mario Zucchelli” Station, for the lo-gistic help and support, which made the expedition possible. They wishto thank Mr. Anderson Aquino of Laboratory of Hydrobiologia of Rio deJaneiro and Mr. Michele Furnari of IAMC of Messina for technical la-boratory helps, and Dr. Giulia Maricchiolo for instrument availability.Thanks are also due to Simona Caputo and Federica Moscheo whocontributed in sample treatment for bacterial isolation.
References
Aislabie, J.M., Chhour, K., Saul, D.J., Miyauchi, S., Ayton, J., Paetzold, R.F., Balks, M.R.,2006. Dominant bacterial groups in soils of marble point and wright valley, VictoriaLand, Antarctica. Soil Biology and Biochemistry 38, 3041–3056.
Aislabie, J., Jordan, S., Ayton, J., Klassen, J.L., Barker, G.M., Turner, S., 2009. Bacterialdiversity associated with ornithogenic soil of the Ross Sea region. Canadian Journalof Microbiology 55, 21–36.
Altschul, S.F., Madden, T.I., Schäffer, A.A., Zhang, J., Zhang, Z., Miller, W., Lipman, D.J.,1997. Gapped blast and psi-blast: a new generation of protein database search pro-grams. Nucleic Acids Research 25, 3389–3402.
Andrade, L., Gonzalez, A.M., Araujo, F.V., Paranhos, R., 2003. Flow cytometry assessmentof bacterioplankton in tropical marine environments. Journal of MicrobiologicalMethods 55, 841–850.
Bakermans, C., Ayala-del-Río, H.L., Ponder, M.A., Vishnivetskaya, T., Gilichinsky, D.,Thomashow, M.F., Tiedje, J.M., 2006. Psychrobacter cryohalolentis sp. nov. andPsychrobacter arcticus sp. nov., isolated from Siberian permafrost. InternationalJournal of Systematic and Evolutionary Microbiology 56, 1285–1291.
Bakermans, C., Skidmore, M.L., Douglas, S., McKay, C.P., 2014. Molecular characteriza-tion of bacteria from permafrost of the Taylor Valley, Antarctica. FEMS MicrobiologyEcology 89, 331–346.
Baldi, F., Marchetto, D., Pini, F., Fani, R., Michaud, L., Lo Giudice, A., Berto, D., Giani, M.,2010. Biochemical and microbial features of shallow marine sediments along theterra nova bay (Ross sea, Antarctica). Continental Shelf Research 30, 1614–1625.
Blanco, Y., Prieto-Ballesteros, O., Gómez, M.J., Moreno-Paz, M., García-Villadangos, M.,Rodríguez-Manfredi, J.A., Cruz-Gil, P., Sánchez-Román, M., Rivas, L.A., Parro, V.,2012. Prokaryotic communities and operating metabolisms in the surface and thepermafrost of Deception Island (Antarctica). Environmental Microbiology 14,2495–2510.
Bölter, M., 2003. Microbiological communities and properties of arctic soils: results of thetundra northwest expedition 1999 (nunavut and northwest territories, Canada).Polarforschung 73, 103–110.
Cannone, N., Guglielmin, M., 2009. Influence of vegetation on the ground thermal regimein continental Antarctica. Geoderma 151, 215–223.
Cannone, N., Wagner, D., Hubberten, H.W., Guglielmin, M., 2008. Biotic and abioticfactors influencing soil properties across a latitudinal gradient in Victoria Land,Antarctica. Geoderma 144, 50–65.
Caruso, G., 2010. Leucineaminopeptidase, beta-glucosidase and alkaline phosphataseactivity rates and their significance in nutrient cycles in some coastal Mediterraneansites. Marine Drugs 8, 916–940.
Danovaro, R., Dell'Anno, A., Trucco, A., Serresi, M., Vanucci, S., 2001. Determination ofvirus abundance in marine sediments. Applied and Environmental Microbiology 67,1384–1387.
Darcy, J.L., Lynch, R.C., King, A.J., Robeson, M.S., Schmidt, S.K., 2011. Global dis-tribution of Polaromonas phylotypes—evidence for a highly successful dispersal ca-pacity. PLoS One 6, e23742.
Deng, J., Gu, Y., Zhang, J., Xue, K., Qin, Y., Yuan, M., Yin, H., He, Z., Wu, L., Schuur, E.A.,Tiedje, J.M., Zhou, J., 2015. Shifts of tundra bacterial and archaeal communitiesalong a permafrost thaw gradient in Alaska. Molecular Ecology 24, 222–234.
Dhaked, R.K., Alam, S.I., Dixit, A., Singh, L., 2005. Purification and characterization ofthermo-labile alkaline phosphatase from an Antarctic psychrotolerant Bacillus sp. P9.Enzyme and Microbial Technology 36, 855–861.
Duhamel, S., Jacquet, S., 2006. Flow cytometric analysis of bacteria- and virus-like par-ticles in lake sediments. Journal of Microbiological Methods 64, 316–332.
Dutta, K., Schuur, E.A.G., Neff, J.C., Zimov, S.A., 2006. Potential carbon release frompermafrost soils of Northeastern Siberia. Global Change Biology 12, 2336–2351.
Edgar, R.C., Haas, B.J., Clemente, J.C., Quince, C., Knight, R., 2011. UCHIME improvessensitivity and speed of chimera detection. Bioinformatics 27, 2194–2200.
Fazi, S., Amalfitano, S., Pernthaler, J., Puddu, A., 2005. Bacterial communities associatedwith benthic organic matter in headwater stream microhabitats. EnvironmentalMicrobiology 7, 1633–1640.
Fierer, N., Bradford, M.A., Jackson, R.B., 2007. Toward an ecological classification of soil
bacteria. Ecology 88, 1354–1364.Frank-Fahle, B.A., Yergeau, È., Greer, C.W., Lantuit, H., Wagner, D., 2014. Microbial
functional potential and community composition inpermafrost-affected soils of theNW Canadian Arctic. PLoS One 9, e84761.
Franzetti, A., Tatangelo, V., Gandolfi, I., Bertolini, V., Bestetti, G., Diolaiuti, G., D'Agata,C., Mihalcea, C., Smiraglia, C., Ambrosini, R., 2013. Bacterial community structureon two alpine debris-covered glaciers and biogeography of Polaromonas phylotypes.The ISME Journal 7, 1483–1492.
French, H., Demitroff, M., Newell, W.L., 2009. Past permafrost on the mid-Atlantic coastalplain, eastern United States. Permafrost and Periglacial Processes 20, 285–294.
Frey, B., Rieder, S., Brunner, I., Plötze, M., Koetzsch, S., Lapanje, A., Brandl, H., Furrer,G., 2010. Weathering-associated bacteria from the Damma glacier forefield: physio-logical capabilities and impact on granite dissolution. Applied and EnvironmentalMicrobiology 76, 4788–4796.
Ganzert, L., Bajerski, F., Mangelsdorf, K., Lipski, A., Wagner, D., 2011. Arthrobacter li-vingstonensis sp. nov. and Arthrobacter cryotolerans sp. nov., salt-tolerant and psy-chrotolerant species from Antarctic soil. International Journal of Systematic andEvolutionary Microbiology 61, 979–984.
Ganzert, L., Bajerski, F., Wagner, D., 2014. Bacterial community composition and di-versity of five different permafrost-affected soils of Northeast Greenland. FEMSMicrobiology Ecology 89, 426–441.
Garland, J.L., Mills, A.L., 1991. Classification and characterization of heterotrophic mi-crobial communities on the basis of patterns of community-level sole-carbon-sourceutilization. Applied and Environmental Microbiology 57, 2351–2359.
Gasol, J.M., Del Giorgio, P.A., 2000. Using flow cytometry for counting natural planktonicbacteria and understanding the structure of planktonic bacterial communities.Scientia Marina 64, 197–224.
Gilichinsky, D.A., Wilson, G.S., Friedmann, E.I., McKay, C.P., Sletten, R.S., Rivkina, E.M.,Vishnivetskaya, T.A., Erokhina, L.G., Ivanushkina, N.E., Kochkina, G.A.,Shcherbakova, V.A., Soina, V.S., Spirina, E.V., Vorobyova, E.A., Fyodorov-Davydov,D.G., Hallet, B., Ozerskaya, S.M., Sorokovikov, V.A., Laurinavichyus, K.S.,Shatilovich, A.V., Chanton, J.P., Ostroumov, V.E., Tiedje, J.M., 2007. Microbial po-pulations in Antarctic permafrost: biodiversity, state, age, and implication for as-trobiology. Astrobiology 7, 275–311.
Gilichinsky, D., Vishnivetskaya, T., Petrova, M., 2008. Bacteria in permafrost. In:Margesin, R., Schinner, F., Marx, J.C., Gerday, C. (Eds.), Psychrophiles: fromBiodiversity to Biotechnology. Springer-Verlag, Berlin Heidelberg, pp. 83–102.
Goordial, J., Davila, A., Lacelle, D., Pollard, W., Marinova, M.M., Greer, C.W.,DiRuggiero, J., McKay, C.P., Whyte, L.G., 2016. Nearing the cold-arid limits of mi-crobial life in permafrost of an upper dry valley, Antarctica. The ISME Journal 10,1613–1624.
Graef, C., Hestnes, A.G., Svenning, M.M., Frenzel, P., 2011. The active methanotrophiccommunity in a wetland from the High Arctic. Environmental Microbiology Reports3, 466–472.
Gryta, A., Frąc, M., Oszust, K., 2014. The application of the Biolog EcoPlate approach inecotoxicological evaluation of dairy sewage sludge. Applied Biochemistry andBiotechnology 174, 1434–1443.
Gugliandolo, C., Michaud, L., Lo Giudice, A., Lentini, V., Rochera, C., Camacho, A.,Maugeri, T.L., 2016. Prokaryotic community in lacustrine sediments of byersPeninsula (Livingston Island, maritime Antarctica). Microbial Ecology 71, 387–400.
Guglielmin, M., Cannone, N., 2012. A permafrost warming in a cooling Antarctica?Climatic Change 111, 177–195.
Guglielmin, M., Dalle Fratte, M., Cannone, N., 2014. Permafrost warming and vegetationchanges in continental Antarctica. Environmental Research Letters 9, 045001.
Hansen, A.A., Herbert, R.A., Mikkelsen, K., Jensen, L.L., Kristoffersen, T., Tiedje, J.M.,Lomstein, B.A., Finster, K.W., 2007. Viability, diversity and composition of the bac-terial community in a high Arctic permafrost soil from Spitsbergen, NorthernNorway. Environmental Microbiology 9, 2870–2884.
Hinzman, L.D., Kane, D.L., Yoshikawa, K., Carr, A., Bolton, W.R., Fraver, M., 2003.Hydrological variations among watersheds with varying degrees of permafrost. In:Proceedings of the Eighth International Conference on Permafrost, pp. 21–25.
Hoppe, H.G., 1993. Use of fluorogenic model substrates for extracellular enzyme activity(EEA) measurement of bacteria. In: Kemp, P.F., Sherr, B.F., Sherr, E.B., Cole, J.J.(Eds.), Handbook of Methods in Aquatic Microbial Ecology. Lewis Publisher, BocaRaton, FL, pp. 423–432.
Hultman, J., Waldrop, M.P., Mackelprang, R., David, M.M., McFarland, J., Blazewicz, S.J.,Harden, J., Turetsky, M.R., McGuire, A.D., Shah, M.B., 2015. Multi-omics of per-mafrost, active layer and thermokarst bogsoil microbiomes. Nature 521, 208–212.
Irgens, R.L., Gosink, J.J., Staley, J.T., 1996. Polaromonas vacuolata gen. nov., sp. nov., apsychrophilic, marine, gas vacuolate bacterium from Antarctica. InternationalJournal of Systematic Bacteriology 46, 822–826.
Jałowiecki, Ł., Chojniak, J.M., Dorgeloh, E., Hegedusova, B., Ejhed, H., Magnér, J., Płaza,G.A., 2016. Microbial community profiles in wastewaters from onsite wastewatertreatment systems technology. PLoS One 11, e0147725.
Jansson, J., Tas, N., 2014. The microbial ecology of permafrost. Nature ReviewsMicrobiology 12, 414–425.
Kane, D.L., Hinzman, L.D., Zarling, J.P., 1991. Thermal response of the active layer toclimatic warming in a permafrost environment. Cold Regions Science and Technology19, 111–122.
Kenarova, A., Encheva, M., Chipeva, V., Chipev, N., Hristov, P., Moncheva, P., 2013.Physiological diversity of bacterial communities from different soil locations onLivingston Island, South Shetland archipelago, Antarctica. Polar Biology 36,223–233.
Kleinteich, J., Hildebrand, F., Bahram, M., Voigt, A.Y., Wood, S.A., Jungblut, A.D.,Küpper, F.C., Quesada, A., Camacho, A., Pearce, D.A., Convey, P., Vincent, W.F.,Zarfl, C., Bork, P., Dietrich, D.R., 2017. Pole-to-Pole connections: similarities between
M. Papale et al. Soil Biology and Biochemistry 123 (2018) 165–179
177

arctic and antarctic microbiomes and their vulnerability to environmental change.Frontiers in Ecology and Evolution 5, 137.
Kuhn, E., Ichimura, A.S., Peng, V., Fritsen, C.H., Trubl, G., Doran, P.T., Murray, A.E.,2014. Brine assemblages of ultrasmall microbial cells within the ice cover of LakeVida, Antarctica. Applied and Environmental Microbiology 80, 3687–3698.
Kumar, S., Tamura, K., Nei, M., 1993. Mega: Molecular Evolutionary Genetics Analysis,Version 1.02. The Pennsylvania State University 1993, University Park, Pa 16802.
La Ferla, R., Maimone, G., Azzaro, M., Conversano, F., Brunet, C., Cabral, A.S., Paranhos,R., 2012. Vertical distribution of the prokaryotic cell size in the Mediterranean Sea.Helgoland Marine Research 66, 635–650.
La Ferla, R., Maimone, G., Lo Giudice, A., Azzaro, F., Cosenza, A., Azzaro, M., 2015. Cellsize and other phenotypic traits of prokaryotic cells in pelagic areas of the Ross Sea(Antarctica). Hydrobiologia 761, 181–194.
La Ferla, R., Azzaro, M., Michaud, L., Caruso, G., Lo Giudice, A., Paranhos, R., Cabral, A.,Conte, A., Cosenza, A., Maimone, G., Papale, M., Rappazzo, A.C., Guglielmin, M.,2017. Prokaryotic abundance and activity in permafrost of the Northern VictoriaLand and Upper Victoria Valley (Antarctica). Microbial Ecology 74, 402–415.
Lee, J.S., Lee, K.C., Kim, K.K., Lee, B., 2016. Complete genome sequence of the Variibactergotjawalensis GJW-30(T) from soil of lava forest, Gotjawal. Journal of Biotechnology218, 64–65.
Liu, F.H., Wang, S.B., Zhang, J.S., Zhang, J., Yan, X., Zhou, H.K., Zhao, G.P., Zhou, Z.H.,2009. The structure of the bacterial and archaeal community in a biogas digester asrevealed by denaturing gradient gel electrophoresis and 16S rDNA sequencing ana-lysis. Journal of Applied Microbiology 106, 952–966.
Lo Giudice, A., Fani, R., 2015. Cold-adapted bacteria from a coastal area of the Ross Sea(Terra Nova Bay Antarctica): linking microbial ecology to biotechnology.Hydrobiologia 761, 417–441.
Lo Giudice, A., Caruso, C., Mangano, S., Bruni, V., De Domenico, M., Michaud, L., 2012.Marine bacterioplankton diversity and community composition in an Antarcticcoastal environment. Microbial Ecology 63, 210–223.
Mackelprang, R., Waldrop, M.P., DeAngelis, K.M., David, M.M., Chavarria, K.L.,Blazewicz, S.J., Rubin, E.M., Jansson, J.K., 2011. Metagenomic analysis of a per-mafrostmicrobial community reveals a rapid response to thaw. Nature 480, 368–371.
Makhalanyane, T.P., Warwick, M., Goethem, V., Cowan, D.A., 2016. Microbial diversityand functional capacity in polar soils. Current Opinion in Biotechnology 38, 159–166.
Mann, P.J., Sobczak, W., Larue, M.M., Bulygina, E., Davydova, A., Vonk, J.E., Schade, J.,Davydov, S., Zimov, N., Holmes, R.M., Spencer, R.G., 2014. Evidence for key enzy-matic controls on metabolism of Arctic river organic matter. Global Change Biology20, 1089–1100.
Margesin, R., Miteva, V., 2011. Diversity and ecology of psychrophilic microorganisms.Research in Microbiology 162, 346–361.
Martineau, C., Whyte, L.G., Greer, C.W., 2010. Stable isotope probing analysis of thediversity and activity of methanotrophic bacteria in soils from the Canadian HighArctic. Applied and Environmental Microbiology 76, 5773–5784.
Michaud, L., Di Cello, F., Brilli, M., Fani, R., Lo Giudice, A., Bruni, V., 2004. Biodiversityof cultivable antarctic psychrotrophic marine bacteria isolated from terra nova bay(Ross sea). FEMS Microbiology Letters 230, 63–71.
Michaud, L., Caruso, C., Mangano, S., Interdonato, F., Bruni, V., Lo Giudice, A., 2012.Predominance of Flavobacterium, Pseudomonas, and Polaromonas within the prokar-yotic community of freshwater shallow lakes in the northern Victoria Land, EastAntarctica. FEMS Microbiology Ecology 82, 391–404.
Møller, A.K., Søborg, D.A., Al-Soud, W.A., Sørensen, S.J., Kroer, N., 2013. Bacterialcommunity structure in High-Arctic snow and freshwater as revealed by pyr-osequencing of 16S rRNA genes and cultivation. Polar Research 32 (1).
Mosier, A.C., Murray, A.E., Fritsen, C.H., 2007. Microbiota within the perennial ice coverof Lake Vida, Antarctica. FEMS Microbiology Ecology 59, 274–288.
Papale, M., Rizzo, C., Villescusa, J.A., Rochera, C., Camacho, A., Michaud, L., Lo Giudice,A., 2017. Prokaryotic assemblages in the maritime antarctic lake Limnopolar (byersPeninsula, south Shetland Islands). Extremophiles 21, 947–961.
Pearce, D.A., Cockell, C.S., Lindström, E.S., Tranvik, L.J., 2007. First evidence for a bi-polar distribution of dominant freshwater lake bacterioplankton. Antarctic Science19, 245–252.
Pearce, D.A., Newsham, K.K., Thorne, M.A.S., Calvo-Bado, L., Krsek, M., Laskaris, P.,et al., 2012. Metagenomic analysis of a southern maritime Antarctic soil. Frontiers inMicrobiology 3 (403).
Pérez, V., Hengst, M., Kurte, L., Dorador, C., Jeffrey, W.H., Wattiez, R., Molina, V.,Matallana-Surget, S., 2017. Bacterial survival under extreme UV radiation: a com-parative proteomics study of Rhodobacter sp., isolated from high altitude wetlands inChile. Frontiers in Microbiology 8, 1173.
Ponder, M.A., Thomashow, M.F., Tiedje, J.M., 2008. Metabolic activity of Siberian per-mafrost isolates, Psychrobacter arcticus and Exiguobacterium sibiricum, at low wateractivities. Extremophiles 12, 481–490.
Porter, K.G., Feig, Y.S., 1980. The use of DAPI for identifying and counting aquatic mi-croflora. Limnology & Oceanography 25, 943–948.
Quast, C., Pruesse, E., Yilmaz, P., Gerken, J., Schweer, T., Yarza, P., Peplies, J., Glöckner,F.O., 2013. The SILVA ribosomal RNA gene database project: improved data pro-cessing and web-based tools. Nucleic Acids Research 41, D590–D596.
Rampelotto, P.H., Barboza, A.D., Pereira, A.B., Triplett, E.W., Schaefer, C.E., de OliveiraCamargo, F.A., Roesch, L.F., 2015. Distribution and interaction patterns of bacterialcommunities in an ornithogenic soil of Seymour Island, Antarctica. Microbial Ecology69, 684–694.
Ramsay, A.J., Stannard, R.E., 1986. Numbers and viability of bacteria in ornithogenicsoils of Antarctica. Polar Biology 5, 195–198.
Rath, J., Schiller, C., Herndl, G.J., 1993. Ectoenzymatic activity and bacterial dynamicsalong a trophic gradient in the Caribbean Sea. Marine Ecology Progress Series 102,89–106.
Rivkina, E., Gilichinsky, D., Wagener, S., Tiedje, J., McGrath, J., 1998. Biogeochemicalactivity of anaerobic microorganisms from buried permafrost sediments.Geomicrobiology 15, 187–193.
Rodionow, A., Flessa, H., Kazansky, O., Guggenberger, G., 2006. Organic matter com-position and potential trace gas production of permafrost soils in the forest tundra innorthern Siberia. Geoderma 135, 49–62.
Saitou, N., Nei, M., 1987. The neighbor-joining method: a new method for reconstructingphylogenetic trees. Molecular Biology and Evolution 4, 406–425.
Sala, M.M., Arin, L., Balagué, V., Felipe, J., Guadayol, O., Vaque, D., 2005. Functionaldiversity of bacterioplankton assemblages in western Antarctic seawaters during latespring. Marine Ecology Progress Series 292, 13–21.
Sala, M.M., Estrada, M., Gasol, J.M., 2006. Seasonal changes in the functional diversity ofbacterioplankton in contrasting coastal environments of the NW Mediterranean.Aquatic Microbial Ecology 44, 1–9.
Schloss, P.D., Gevers, D., Westcott, S.L., 2011. Reducing the effects of PCR amplificationand sequencing artifacts on 16S rRNA-based studies. PLoS One 6, e27310.
Schuur, E.A.G., Bockheim, J., Canadell, J.G., Euskirchen, E., Field, C.B., Goryachkin, S.V.,Hagemann, S., Kuhry, P., Lafleur, P.M., Lee, H., Mazhitova, G., Nelson, F.E., Rinke,A., Romanovsky, V.E., Shiklomanov, N., Tarnocai, C., Venevsky, S., Vogel, J.G.,Zimov, S.A., 2008. Vulnerability of permafrost carbon to climate change: implicationsfor the global carbon cycle. BioScience 58, 701–714.
Schuur, E.A.G., Vogel, J.G., Crummer, K.G., Lee, H., Sickman, J.O., Osterkamp, T.E.,2009. The effect of permafrost thaw on old carbon release and net carbon exchangefrom tundra. Nature 459, 556–559.
Shivaji, S., Reddy, G.S., Aduri, R.P., Kutty, R., Ravenschlag, K., 2004. Bacterial diversityof a soil sample from Schirmacher Oasis, Antarctica. Cellular & Molecular Biology 50,525–536.
Sinsabaugh, R.L., Follstad Shah, J.J., 2012. Ecoenzymatic stoichiometry and ecologicaltheory. Annual Review of Ecology Evolution and Systematics 43, 313–343.
Sinsabaugh, R.L., Lauber, C.L., Weintraub, M.N., Ahmed, B., Allison, S.D., Crenshaw, C.,Contosta, A.R., Cusack, D., Frey, S., Gallo, M.E., Gartner, T.B., Hobbie, S.E., Holland,K., Keeler, B.L., Powers, J.S., Stursova, M., Takacs-Vesbach, C., Waldrop, M.P.,Wallenstein, M.D., Zak, D.R., Zeglin, L.H., 2008. Stoichiometry of soil enzyme ac-tivity at global scale. Ecology Letters 11, 1252–1264.
Sinsabaugh, R.L., Belnap, J., Findlay, S.G., Follstad Shah, J.J., Hill, B.H., Kuehn, K.A.,Kuske, C.R., Litvak, M.E., Martinez, N.G., Moorhead, D.L., Warnock, D.D., 2014.Extracellular enzyme kinetics scale with resource availability. Biogeochemistry 121,287–304.
Smith, D.J., Schuerger, A.C., Davidson, M.M., Pacala, S.W., Bakermans, C., Onstott, T.C.,2009. Survivability of Psychrobacter cryohalolentis K5 under simulated Martian sur-face conditions. Astrobiology 9, 221–228.
Soina, V.S., Mulyukin, A.L., Demkina, E.V., Vorobyova, E.A., El-Registan, G.I., 2004. Thestructure of resting bacterial populations in soil and subsoil permafrost. Astrobiology4, 345–358.
Steven, B., Léveillé, R., Pollard, W.H., Whyte, L.G., 2006. Microbial ecology and biodi-versity in permafrost. Extremophiles 10, 259–267.
Steven, B., Briggs, G., McKay, C.P., Pollard, W.H., Greer, C.W., Whyte, L.G., 2007.Characterization of the microbial diversity in a permafrost sample from the Canadianhigh Arctic using culture-dependent and culture-independent methods. FEMSMicrobiology Ecology 59, 513–523.
Steven, B., Pollard, W.H., Greer, C.W., Whyte, L.G., 2008. Microbial diversity and activitythrough a permafrost/ground ice core profile from the Canadian high Arctic.Environmental Microbiology 10, 3388–3403.
Steven, B., Niederberger, T.D., Whyte, L.G., 2009. Bacterial and archaeal diversity inpermafrost. In: Margesin, R. (Ed.), Permafrost Soils. Springer-Verlag, BerlinHeidelberg, pp. 59–72.
Sul, W.J., Oliver, T.A., Ducklow, H.W., Amaral-Zettler, L.A., Mitchell, L.S., 2013. Marinebacteria exhibit a bipolar distribution. Proceedings of the National Academy ofSciences 110, 2342–2347.
Thompson, J.D., Higgins, D.G., Gibson, T.J., 1994. CLUSTAL W: improving the sensitivityof progressive multiple sequence alignment through sequence weighting, position-specific gap penalties and weight matrix choice. Nucleic Acids Research 22,4673–4680.
Vazquez, S.C., Coria, S.H., Mac Cormack, W.P., 2004. Extracellular proteases from eightpsychrotolerant Antarctic strains. Microbiological Research 159, 157–166.
Vieira, G., Bockheim, J., Guglielmin, M., Balks, M., Abramov, A., Boelhouwers, J.,Cannone, N., Ganzert, L., Gilichinsky, D.A., Goryachkin, S., Lopez-Martinez, J., IanMeiklejohn, I., Raffi, R., Ramos, M., Schaefer, C., Serrano, E., Simas, F., Sletten, R.,Wagner, D., 2010. Thermal state of permafrost and active-layer monitoring in theAntarctic: advances during the International Polar Year 2007–2009. Permafrost andPeriglacial Processes 21, 182–197.
Vishnivetskaya, T., Kathariou, S., McGrath, J., Gilichinsky, D., Tiedje, J.M., 2000. Low-temperature recovery strategies for the isolation of bacteria from ancient permafrostsediments. Extremophiles 4, 165–173.
Wagner, D., 2008. Microbial communities and processes in Arctic permafrost environ-ments. In: Dion, P., Nautiyal, C.S. (Eds.), Microbiology of Extreme Soils. SpringerBerlin Heidelberg, pp. 133–154.
Wagner, D., Liebner, S., 2009. Global warming and carbon dynamics in permafrost soils:methane production and oxidation. In: Margesin, R. (Ed.), Permafrost Soils. Springer,Berlin Heidelberg, pp. 219–236.
Wagner, D., Kobabe, S., Liebner, S., 2009. Bacterial community structure and carbonturnover in permafrost-affected soils of the Lena Delta, northeastern Siberia.Canadian Journal of Microbiology 55, 73–83.
Wilhelm, R.C., Niederberger, T.D., Greer, C., Whyte, Lyle, G., 2011. Microbial diversity ofactive layer and permafrost in an acidic wetland from the Canadian High Arctic.Canadian Journal of Microbiology 57, 303–315.
M. Papale et al. Soil Biology and Biochemistry 123 (2018) 165–179
178

Wilkins, D., Yau, S., Williams, T.J., Allen, M.A., Brown, M.V., DeMaere, M.Z., Lauro, F.M.,Cavicchioli, R., 2013. Key microbial drivers in Antarctic aquatic environments. FEMSMicrobiology Reviews 37, 303–335.
Yergeau, E., Hogues, H., Whyte, L.G., Greer, C.W., 2010. The functional potential of highArctic permafrost revealed by metagenomic sequencing, qPCR and microarray ana-lyses. The ISME Journal 4, 1206–1214.
Yergeau, E., Bokhorst, S., Kang, S., Zhou, J., Greer, C.W., Aerts, R., Kowalchuk, G.A.,
2012. Shifts in soil microorganisms in response to warming are consistent across arange of Antarctic environments. The ISME Journal 6, 692–702.
Zhu, R., Wang, Q., Ding, W., Wang, C., Hou, L., Ma, D., 2014. Penguins significantlyincreased phosphine formation and phosphorus contribution in maritime Antarcticsoils. Scientific Reports 4, 7055.
Zimov, S.A., Schuur, E.A., Chapin, F.S., 2006. Permafrost and the global carbon budget.Science 312, 1612–1613.
M. Papale et al. Soil Biology and Biochemistry 123 (2018) 165–179
179