sp-303, an antiviral oligomeric proanthocyanidin from the...
TRANSCRIPT

Phytomedicine Vol. 1/1994, pp. 77-106
© 1994 by Gustav FischerVerlag, Stuttgart· Jena . New York Original Papers
SP-303, an Antiviral Oligomeric Proanthocyanidinfrom the Latex of Croton lechleri (Sangre de Drago).
R. UBILLAS1, S.D. JOLAD1, R.C. BRUENING1, M. R. KERNAN1,S.R. KING1, D.F. SESIN1, M. BARRETI1, C. A. STODDART1,1. FLASTER1, J. KU01, F. AYALA1, E. MEZA1, M. CASTANEL1,D. McMEEKIN1, E. ROZHON1, M.S. TEMPESTA1,D. BARNARD2, J. HUFFMAN2, D. SMEE2, R. SIDWELL2,K. SOIKE3,A. BRAZIER4,S. SAFRIN5,R. ORLAND06, P.T.M. KENNy6,N. BEROVA7\ K. NAKANISHI7.
1 Shaman Pharmaceuticals,213 East Grand Avenue, South San Francisco, CA 94080-4812, U.S.A.2 Institute for Antiviral Research, Department of Animal,Dairy and VeterinaryScience, Utah State Univer-
sity,Logan, UT, 84322 U.S.A.3 Tulane Regional Primate center, 18703 Three Rivers Road, Covington, LA, 70433 U.S.A.4 Southern Research Institute, Birmingham, AL, U.S.A.S San Francisco, General Hospital, InfectiousDiseases, Building 80, Ward 84, 998 Potrero Avenue, San
Francisco,CA 94110 U.S.A.6 Suntory Institute for Bioorganic Research (SUNBOR), Shimamoto-cho,Mishima-gun, Osaka 618, JA
PAN7 Department of Chemistry, Columbia University, New York, NY 10027, U.S.A.
t on leave of absence from the BulgarianAcademy of Science, Sofia,Bulgaria.
Summary
SP-303, a large proanthocyanidin oligomer isolated from the latex of the plant species Crotonlechleri (Euphorbiaceae) has demonstrated broad activity against a variety of DNA and RNA viruses. In cell culture, SP-3D3 exhibits potent activity against isolates and laboratory strains of respiratory syncytial virus (RSV), influenza A virus (FLU-A) and parainfluenza virus (PIV). Parallelassays of SP-3D3 and ribavirin showed comparable activity against these viruses. SP-303 also exhibits significant inhibitory activity against herpesvirus (HSV) types 1 and 2, including herpesviruses resistant to acyclovir and foscarnet. Inhibition was also observed against hepatitis A and Bviruses. The antiviral mechanism of SP-303 seems to derive from its direct binding to componentsof the viral envelope, resulting in inhibition of viral attachment and penetration of the plasmamembrane. Antiviral effects of SP-303 were measured by three distinct methods: CPE, MTT andprecursor uptake/incorporation. Cytotoxicity endpoints were markedly greater than the respective antiviral endpoints. SP-303 exhibited activity in RSV-infected cotton rats and African greenmonkeys, PIV-3-infected cotton rats, HSV-2 infected mice and guinea pigs and FLU-A-infectedmice. The most successful routes of SP-303 administration for producing efficacy were: topicalapplication to HSV-2- genital lesions in mice and guinea pigs, aerosol inhalation to FLU-A-infected mice and PIV-3-infected cotton rats, and oral dosage to RSV-infected cotton rats. A variety oftoxicological evaluations demonstrated the safety of SP-303, particularly orally, which was predictable, since condensed tannins are a common dietary component. It is notable that the larger

78 R. Ubillas et al.
proanthocyanidins as a class have high antiviral activity, whereas most of the monomers are inactive. Clinical trials are ongoing to evaluate SP-303 as a therapeutic antiviral agent.
Key words: Croton lechleri (sangre de drago), oligomeric procyanidin SP-303, antiviral activity.
I. Ethnobotanical Background
The sub ject of this study, SP-303, is an oligomeric proanthocyanidin derived from Croton lechleri, one of a variety ofspecies of tre ss within the Croton genus in the family Euph orbiaceae. This family is widely known for many medicinalpurposes (Von Reis and Lipp, 1982; McRae et aI., 1988). Ared, viscous latex is produced by several species within thesection Cyclostigma Griseb . of this genus. These species arewidely distr ibuted throughout Central and South America,ranging from Mexico to Paraguay and southern Brazil.
This red latex is one of the most common traditional medicines in all of Lat in Amer ica. Its diversity of ethnomedicaluses is summarized below. The latex is most commonly referred to as "Sangre de Drago" (" dragons' blood" ) and"Sangre de Grado" Spanish names appl ied to the plant bythe Europeans upon the ir arrival in the New World fivecenturies ago. The origin of the name "Sangre de Drago"comes from a number of species in the botanical genus Dracaena in the family Agavaceae. Stems of several of the 60species in thi s genus produce a red resin (giving rise to thename "dragons' blood" ) that has been used in varnishes.One particular species, Dracaena cinnabari Balf., wa s probabl y known in antiquity. A second species, Dracaena draco (L.), the dragons' blood tree, had been used as a woundhealing agent in Spain, the Canary Island s and other regions of Europe.
Many vernaculars for "Sangre de Drago" exist in man yspecific indigenous langu ages. In the language of the Untsuri shuar, a species of th is genus bearing red latex is referred to as "Uruchnum" (Bennett, 1990). Among the Qu ichua Indians of Amazonian Ecuador, it is "Lan huiqui "(King, pers. obs.). The Cofan Indians call it "Masujuain"(Ceron, 198 7); the Waorani Indians, "Conewe" (King,per s. obs.), In Mexico and Guatemala, one 0 the red latexbearing species, Croton draco Cham & Schltdl., has at least18 different indigenous names including such variations of"dragons' blood" as "Lora Sangre" and "Sangre de Perro" ,as well as a number of non-Spanish derived names such as"Chucum" in the Lacondon language of the region of Chiapas, or "Xixte" in the langu age of the Huasteca in San LuisPotosi, Th is is only a partial listing of the indigenousnames. Clearly, this plant has had widespread and diverseuse for healing among ind igenous groups throughout Central and South Amer ica, including the countries of Mexico,Guatemala, Panama, Colombia, Ecuador, Peru, Bolivia,Paraguay and Brazil.
The most well-known and widespread red latex-bearing
Croton species of the northwest Amazon, and a primar ysource for SP-303 is Croton lechleri Miill. , Arg. Thi s species is distributed in the countries of Colombia, Ecuador, .Peru and Bolivia. A closely related species that may be asynonym, C. draconoides Miill., Arg., is recorded in Peru,and the type specimen was collected in Brazil. There are,however, multiple other species that have yielded SP-303,ranging from Mexico to Paraguay and southern Brazil.Those species include C. draco, Cham & Schltd!., especially wide-spread in Cent ra l America, C. urucurana, Baill. ,well-known in Paraguay and southern Brazil, C. sordidus,Benth ., of the Andean region, and C. gossypifolius Vah!.,common also in the Andean region . Add itional speciesthat yield red latex in a number of countries in Cent raland South Amer ica include C. ferrugineus, c. panamensisS. C. Klotzsch, C. palanostigma Mart. & Baill., and C.salutaris. Th is sub-group of the genus Croton is in need ofrevision because of the many synonym names for the species and the need for taxonomic clarificat ion for th is medicinally important group of New World plant s. The tax onomy and systematics of these species is currently underinspection and revision by Dr. Grady Webster (Webster,1993) and a doctoral candidate at the University of Californ ia, Davis.
Ethn om edical Uses. The source for SP-303, "Sangre deDrago" of Central and South America, is one of the mostwidely found, known and used plant medicinal sourc es inthe entire hemisphere. It has been used for centuries andcontinues to be used by a wide diversity of people, especially in Colombia, Ecuador, Peru, Bolivia, Paragua y, Mexicoand Cenral America. Users include nat ive, urban and mestizo populati ons, as well as urban healers; it is now increasingly being sold in urb an health food sto res. Thi s plantmed icine is extensively sold in Peru and Ecuador (Me za1994, in press). Its most common usages ar e: internally forcoughs, flu, "problems with lungs " , diarrhea, and for"stomach ulcers"; and topically as a wound healing agentfor cut s, open sores, herpes infections, for the gums aftertooth extraction and for oral open sores in and on themouth. In some urban areas, it is appli ed topically after surgery for enhanced wound healing. Th e general dosage forinternal use is 5-10 drops of the red latex in warm or coldwater, milk or in alcohol, 1-3 times a day for 5 days. Th isregimen is often repeated for a period of up to 3 weeks. It isappli ed directly to open wounds to stop blood flow andpromote healing. Diluted with water, it is applied as a vaginal wash for excessive vaginal bleed ing. No side-effectshave been reported after internal use.

In urban regions of many Andean countries and in Central America, the latex is currently being sold under at least20 different labels by a variety of "health food manufacturers". These manufacturers sell small quantities of the liquid in bottles for hepatitis, diabetes, ulcers, cancer prevention, tonsillitis, as an anti-inflammatory, and to enhancefertility and weight loss. It is also recommended for hemorrhoids, acne and as vaginal wash. The predominant usesamong the indigenous populations are limited to topicalwound healing and internally for coughs, colds, diarrheaand stomach problems.
In summary, the Croton species that form a widespreadcomplex known regionally as "Sangre de Drago", plays acritical role in the local and urban traditional medicine ofnumerous countries throughout Latin America. Historically, scientific interest in the wound-healing and antiinfectiveproperties of "Sangre de Drago" is well documented in theliterature (Bettolo and Scarpati, 1979; Persinos, 1972; Persinos-Perdue et al., 1979; Kitazawa et al., 1980; Vaisberg etal., 1979; Vlietinck, 1987), and general scientific interestcontinues to this day (Cai et al., 1991; Lewis et al., 1992;Tempesta, 1993).
Distribution, Density, and Harvesting of the SF-303Source Species. Over the past four years, Shaman Pharmaceuticals has committed itself to the long-term sustainableharvesting and management of SP-303 source species. Wehave conducted 14 distinct studies in four different countries on ecology, distribution, density, growth habits, marketing, agroforestry, management and collection methods.We have worked closely with indigenous peoples and scientists to develop sustainable harvesting methods that willhelp protect and manage this group of plants species. Ourconcerns are both to sustain the source for our lead antiviral products and to assist local populations in continuing tohave direct and easy access to the plant material as a sourceof regional traditional medicine.
In numerous examples, uncontrolled clear-cutting has destroyed the genetic diversity of a number of species throughout the tropics, and particularly in Amazonia. Specificallythe cases of rosewood oil (Aniba rosaeodora) and Oje (Ficus anthelmintica) have shown that intense market demands can cause dramatic ecological consequences bythreatening the gene pool and diversity of economically important species. Since its beginning, Shaman Pharmaceuticals had committed itself to pay very close attention to thedetail of sustainable supply at every stage of the drug development process. Recent work by Cragg et al. (1993), discussing the evolution of taxol and the supply crises that surrounded its source species, has further emphasized the importance of working on the long-term supply aspects of anypotential natural product for drug development. In ouropinion, a protocol incorporating all steps needed to conserve and manage a target species should become an integralpart of any preclinical data package.
Our distribution studies have shown an extremely wide-
SP-303 79
spread occurrence and high natural density of the SP-303 bearing Croton species from Mexico to Paraguay. A density of 3 to 10 species per hectare seems to be common, especially along riparian habitats throughout the northwestAmazon. These specific studies will be published elsewherein collaboration with the primary scientists who conductedthe research. These research data have been provided to local government ministries and conservation organizationsas well as directly to indigenous groups working on themanagement of this species.
Ecological habitat and distribution vary considerably.Most of the C. lechleri grows in the lowland northwesternAmazonian forest region from 100-600 meters in elevation.Related species occur from 700-2500 meters along theeastern and western slopes of the Andean regions of Ecuador, Colombia and Peru. Other Croton species in Mexicoand Central America are more adapted to moist tropicaland in some cases sub-tropical forest habitats in their moisture regime. The species in Paraguay and southern Brazil areadapted to the drier cerrado type of habitats with a muchlower level of annual precipitation.
The species C. lechleri and several related species exhibitclassic pioneer species ecological profiles. They are one ofthe three most successful emergent species appearing incleared fields and on roadsides throughout much of theEcuadorian and Peruvian forest regions between 600 and2500 meters.
The most common harvesting method for C. lechleri hashistorically been the slashing of bark, which initiates theslow release of up to several liters of latex from a standingtree of up to 6 years of age with an average diameter atbreast height (DBH) of 25 centimeters. Traditional harvestpractices utilize small quantities (less than 100 m!) of material for personal and family use. However, the increasingmarket for this material over the past decades has led to amore common harvesting practice of felling of the tree andsubsequent scoring of the bark at intervals of 6-12 inchesaround the circumference. This method yields up to 5-61iters of latex per tree depending upon the age and season ofthe harvest (i.e., latex yield is greater during the rainy season).
Sustainability. Because this tree of the Euphorbiaceae family produces latex continuously, we originally focused onmethods for sustainably "tapping" the latex as it is donefor the harvest of natural rubber. We therefore commissioned a number of studies by experts on the morphologyon the genus of Hevea and Croton. While rubber has beensuccesfully tapped both in the wild and in plantations, thelaticifers of the red latex-bearing Croton species are distinctfrom those of the Hevea species. Rubber trees contain mainlatex "canals" in which the latex restores and flows continuously. In contrast, Croton laticifers contain many tubulesthat must be opened individually to release the latex inside(Rudall, pers. comm.). It is therefore not feasible to "tap"Croton tree species like Hevea rubber trees. However, the

80 R. Ubi lias et at.
rapid gro wth of the trees and high yield of SP-303 (- 1 % )provid es an excellent opportunity for managing these species in agro forestry systems where they only require theminimal management of high yieldi ng mixed-species secondar y forests.
Conservation and Management. M aintaining the integrity of th e tropical rainforest ecosystems requires integratingthe needs of th e people who live in a nd around these en vironments. Long-term conse rva t ion management must beinterwoven with the cultural va lues and needs in reg ion swhere source species occur. Studies have shown that we canmanage and maintain the genet ic diversity of these speciesby cre ating sustainable harvest and management systems incollaboration with forest-dwell ing people.
We recognize this as an ecological and business imperative tomaintain operational and long-term supply and sound ecologicalmanagement for the benefit of local inhabitants. We have enteredinto agreements with indigenous federations such as the ConsejoAguaruna y Huamb isa of the northern Peruvian Amazon to support and develop reforesration and management programs for thistree species. The Consejo Aguaruna y Huambisa provided us withan agreement from their annual congreso which included the signatures of 137 delegates representing some 30 000 individuals ofthese tribal groups in the norrhern Peruvian Amazon (King, 1994 ).This is an ongoing process requiring monitoring and reporrs fromShaman Pharmaceuticals to the various groups with which wework. The specifics on price, purchase and many other details havebeen stipulared by the various indigenous groups with which weare collaborating and are being monitored at the federation levelby organizations such as AIDESEP (the National Peruvian Indigenous Peoples Organization of the Amazonian Region). We havealso initiated similar programs with the Cofan Indians as well asother indigenous groups in the rainforest regions of several Andean countries. We are actively working with these and other individuals to create management plans and conduct reforestationactivities that greatly exceed the amount of harvested material.The governments of the countries in which we are conductingthese activities require routine, long-term management plans toallow us to proceed with the collection and export of raw materials for this drug development program. We have helped create alarge body of information on the conservat ion and management ofthis wide-spread and highly important medicinal plant specieswhere very little baseline information existed previously. To promote these business practices, we are also supporring school programs and encouraging local students to understand the conceptsof reforestat ion and plant uses that are basic to their ethnobotanical and ethnomedical needs. We have been supporring communityseminars in conjunction with the Ministry of Agriculture to informfarmers of the imporrance of this tree and the value of applyingeven minimal management to it. We have also helped organizingseminars to train indigenous federation representat ives to furtherwork with their indigenous colleagues to maintain this imporrantresource for the future.
II. Isolation and Characterization of SP-303
Botanical class characteristics. Proanthocyanidins andproanthocyanidin polymers are found as naturally occurr ing phenolic substances in a wide variety of plants, parti-
cul arl y those with a woody habit of growth. The generalchemical stru ctu re of monom eric and polymeric proanthocyanidins is shown on p. 88 (Harborn e, 1988 ). Thepolym er consists of linear chains of 5,7,3',4'-tet rahydroxy(catechin or epicatechin) o r 5,6,3',4',5'-pentahy droxyflavan-3-ol (gallocatec hin or ga lloep icatechin) units linkedtogether through either C-4 to C-6 and /or C-4 to C-8.
The monomer unit of th e polymer chai n ma y be based oneither of two sterochem istries of th e C rin g, des ignated cisan d termed epicatechin or trans and named catechin, andon eith er of two B ring oxidation patterns, 3',4'-dihydroxyph enyl (designated as a procyanidin unit) or 3',4',5'-trihydroxyphenyl (designated as a p rodelphinidin unit). Therefor e the polymer chains which are based on four differentmonomeric units create a w ide variety of polymeric proanthocya nidins and a large number of possible isomers.
Proanthocyanidins (syn. leucoanthocyanins, or "condensed tannins" ) ar e a class o f co mpounds that yieldcya nidin and or delphinidin [anthoc yan idins] on treatmentwith acid. They encompass all flavan-Ssol monomers,dimers and higher olig omers. Condensed proanthocyanidins are the dimers and higher oligome rs of the variousflavan-Svols, with polymeric proanthocyanidins consistingof fla van-3-ol polym ers. Procyanidins are a subclass of proanthocyanidins, consisting of various flavan- Svols containing only the B ring oxidation pattern of 3 ', 4 '-dihydroxyph en yl. Prodelphinidins ar e also a sublcass of proanthocyanidins, co mposed solel y of B ring oxidation as 3',4',5'trihydroxyph enyI. Leucoanthocyan id ins encompass all flava n-3 ,4-diol monomers as well as flavan-t-ols, and are another subclass of proanthocyanidins. Further examples ofsubclass members have been published by Harborne(1988), who gives the range of oxidation patterns found inth e pr oanthocyanidin class.
Fractionation and isolation. The lat ex of Croton lechleriwas found to have significant in vitro antiviral activity ininfluenza (FLU-A) and respi ra tory syncy tia l virus (RSY)screens and was initially subjected to a bioassay-guidedfract ion at ion over Sephadex to remove inac tive components, and concentrate bio act ivity. This evol ved int o a sea leable isolation scheme using th e seven-step process descri bed below. The product turned out to be the most bioact ive component in the latex, a red-brown proanthocyanidinoligomer, termed SP-303. Flavonoids in general and smallproanthocyanidins ha ve been reported to exhibit antiviralpr op erti es (Beladi et al. , 19 72; Kaul et a l., 1985; Takechi eta l., 1985; Selway, 1986; Vanden Berghe et aI., 1986; Nonaka et aI. , 1989; Hudson, 1990 ; N onaka et aI., 1990). Infact , significa nt clinical effo rts involving a flavan and achalcone were undertak en to trea t rh inovirus infections,although neither were successful (Selway, 1986). Before webegan our work, there had been no published data on th ed irect antiviral effects of proanthocyanidin polymers, alth ough they are currently the subject of considerable inter est as evidenced by patents and patent applications

(Itsuo et aI., 1989; Cariel and Jean, 1990; Nonaka et aI.,1990; Tempesta, 1993).
After addition of 3 parts isopropanol to the latex (Step 1),the supernatant of the resulting biphasic mixture was further extracted with butanol (Step 2). The residual aqueoussolution was evaporated to dryness, dissolved in methanol,and precipitated with ethyl acetate (Step 3). The filtered supernatant was then purified by a combination of ionexchange chromatography and size exclusion chromatography. Using water as the eluent, alkaloids (primarily taspine) present in the material (Persionos-Perdue et aI., 1979)were removed from the Step 3 intermediate by cationexchange chromatography on CM-Sephadex C-50. Furtherenrichment of the proanthocyanidin polymer-containingfraction was achieved by connecting a Sephadex G-50 column to the outlet of the CM-Sephadex C-50 column. Withwater as the eluent, the proanthocyanidin oligomer adsorbed to the G-50 column while some of the more polarlow molecular weight compounds eluted. The bioactiveproduct was then eluted with 15 % aqueous acetone(Step 4). Fractionation of the Step 4 intermediate on Toyopearl HW-40S and elution with 20 % aqueous acetone removed most of the low molecular weight proanthocyanidins. Further elution with 40 % aqueous acetone gave anintermediate of desired bioactivity (StepS). This intermediate was finally purified on Sephadex LH-20. Elutionwith 90 % aqueous ethanol followed by 20 % aqueousacetone removed remaining monomeric, dimeric, trimericand tetrameric proanthocyanidins, while elution with40-50 % aqueous acetone yielded a fraction comprisingmedium-sized oligomers, designated as SP-303 (Step 6).
The use of three successive gel permeation columns allowed rigorous control of the molecular weight averageand distribution of the final product. The moisture contentand grain size of the final product were controlled by dissolving a given amount of the Step 6 material in twice theamount of water and evaporating the resulting syrup undervacuum for a specific time and temperature, then grindingand sifting the resulting solid material through U.S. Standard Testing Sieves. The overall yield of SP-303 by thismethod was -1 %; this process has been recently improvedto allow larger batches to be prepared increasing the yieldto 12-15 % (wt./v of latex).
An HPLC procedure employing a highly cross-linked polystyrenedivinylbenzene gel permeation column was usedas an in-process control method for evaluating the purity ofmaterial produced in each of the above chromatographicsteps. The same procedure was used to evaluate the purityand molecular weight distribution of the final drug substance. This procedure is described in more detail below.
One method that has been used to determine the numberaveraged molecular weight of a proanthocyanidin is gelpermation chromatography (GPC) of the acetylated polymers (Williams et aI., 1983). The acetylations are normallycarried out by reaction of the polymers with acetic anhy-
SP-303 81
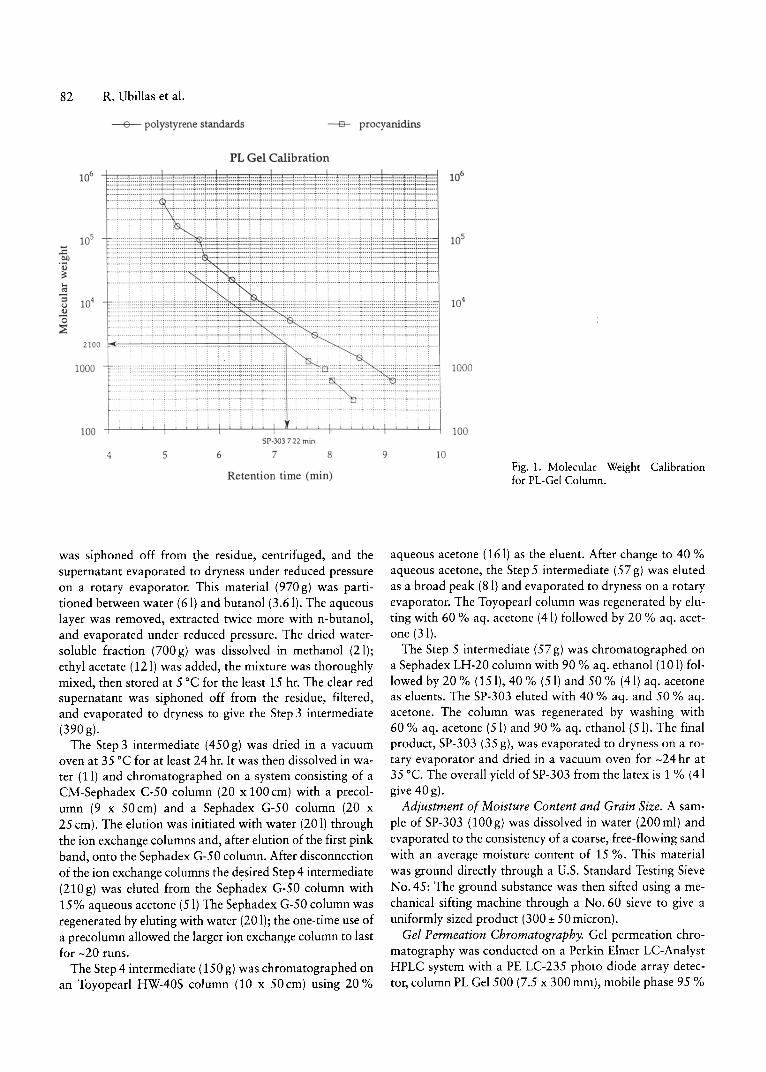
dride in pyridine. Gel permeation chromatograms are obtained using tetrahydrofuran as the mobile phase and polystyrene-divinylbenzene as the stationary phase. Unfortunately, because of the limited solubility of SP-303 in pyridine, this method could not be used for SP-303. Direct injection of SP-303 resulted in adsorption of the materialonto the GPC column. However, by adding water to themobile phase to give 95 % aqueous tetrahydrofuran, it wasfound that all Croton lechleri proanthocyanidins, includingSP-303, eluted from the column with> 99 % recovery. Foraccurate molecular weight determinations by GPC, thestandards must have similar chemical properties to the analyte. The ideal standards are monomers, oligomers andpolymers of the same structure type as the analyte. Polystyrene standards (Polymer Laboratories) and proanthocyanidin oligomers obtained from C. lechleri (Cai et aI., 1991)were used to prepare the calibration curves shown in Figure 1 using a Polymer Laboratories PL Gel 500 column. Toincrease the resolution, this analysis was repeated usingtwo PL Gel Mixed E columns connected in series. The PLGel Mixed E column has a 3 micron particle size, with anefficiency of 50000 plates/m, equivalent to 3 columns ofthe same length containing 10 micron particles.
It is not surprising that the use of two types of polymerstandards resulted in different molecular weight predictions. It was assumed that the calibration curve based uponthe proanthocyanidin standards would give a more accurate estimate of the true molecular weight distribution inSP-303; the data were extrapolated as shown to cover theelution range observed for SP-303. The molecular weightrange and distribution of SP-303 as determined by thismethod is 1500-3300, corresponding to between 5 and 11flavanoid units, with a number-averaged molecular weightfor SP-303 of -2100 daltons and an average chain length of7 flavanoid units. Analysis of the gel permeation chromatogram of SP-303 as shown in Figure 2 was performedusing a photo diode array detector. The UV spectra obtained at 20 % of peak upslope, peak apex and 20 % ofpeak downslope were overlayed and plotted (normalized)from 215-365nm and 250-365nm, as depicted in Figure3. These data clearly show the homogeneity of SP-303.
Chromatography
General. All preparative chromatography was conductedusing FMI pumps and UV/VIS detectors set at 450 nm(linear). Flow rates were 25 -50 rnl/rnin. Moisture contentsof SP-303 were determined using a Karl-Fischer device(Methrohm). Thin-layer chromatography was conductedon HPTLC plates (Merck) developed with EtOAc-HzOformic acid - acetic acid (80 :20: 3: 2) and visualized byspraying with 1 % vanillin in 20 % ethanolic HCI.
Isolation. Isopropanol was added to the cold latex (41) inthe volume ratio of 3 : 1. After mixing, the material wasstored at 5°C for at least 15 hr, the clear red supernatant
82 R. Ubillas et al.
---e- polystyrene standards --i3- pro cyanidins
PL Gel Calibration
100
1000
. ... .,..~ ....';,...~ ....~.....!.....~.....:....~.....
...:.....; .....•....~ ....; ....;.....:.....~.... ~.. ...
SP-303 7.22min
i j i! I i100
lOS:c00
'0~..~
:::l 104u~
'0::E
2100
1000
H:::~:H:F:t::::H:~: :::1::::r~1:~~~f~~~ff:l~~r::1~:::~:::~~~:~i : ::l::~rWH:::t:::l::::~:::~~~:F.~~~f::~
- ; : : - : ic:: i::1JJ~f~j~jl-:; :- : : ~ : ',:;:j,rffj~i; 10'
4 5 6 7 8 9 10
Retenti on time (min)Fig. 1. Molecul ar Weight Calibrationfor PL-Gel Colum n.
was siphoned off from the residue , centri fuged, and thesupernatant evaporated to dryness under reduced pressureon a rotary evaporator. Thi s material (970 g) was partitioned between water (61) and butanol (3.6 1). The aqu eouslayer was removed, extr acted twice more with n-butanol,and evaporated under reduced pressure. The dried watersoluble fraction (700 g) was dissolved in methanol (21);ethyl acetate (121) was add ed, the mixture was thoroughlymixed, then stored at 5°C for the least 15 hr. The clear redsupernatant was siphoned off from the residue, filtered ,and evaporated to dryness to give the Step 3 intermedia te(390g).
The Step 3 intermediate (450 g) was dried in a vacuumoven at 35°C for at least 24 hr. It was then dissolved in water (1 1) and chromatographed on a system consisting of aCM-Sephadex C-50 colum n (20 x 100 cm) with a precolumn (9 x 50 em) and a Sephadex G-50 column (20 x25 ern). The elution was initiated with water (20 I) throughthe ion exch ange columns and , after elution of the first pinkband, onto the Sephadex G-50 column. After disconnectionof the ion exchange columns the desired Step 4 intermediate(210 g) was eluted from the Sephad ex G-50 column with15% aqueous acetone (5 1) The Sephadex G-50 column wasregener at ed by eluting with water (20 I); the one-time use ofa precolumn allowed the larger ion exchange column to lastfor -20 runs .
The Step 4 intermediate (150 g) was chromatographed onan Toyopearl HW-40S column (10 x 50cm) using 20 %
aqueous acetone (16 1) as the eluent. After change to 40 %aque ous aceton e, the Step 5 intermediate (57 g) was elutedas a bro ad peak (8 1) and evaporated to dryness on a rotaryevaporato r. The Toyopearl column was regenerated by eluting with 60 % aq. acetone (41) followed by 20 % aq. acetone (31).
The Step 5 intermediate (57 g) was chromatographed ona Sephadex LH-20 column with 90 % aq . ethanol (10 I) followed by 20 % (15 1), 40 % (5 1) and 50 % (4 1) aq. aceton eas eluents. The SP-303 eluted with 40 % aq. and 50 % aq.aceton e. The column was regenerated by washing with60 % aq. acetone (5 1) and 90 % aq. ethanol (5 1) . Th e finalproduct, SP-303 (35 g), was evaporated to dryness on a rotary evap orator and dr ied in a vacuum oven for - 24 hr at35°C. The overall yield of SP-303 from the latex is 1 % (4 1give 40 g).
Adjustment of Moisture Content and Grain Size. A sample of SP-303 (100 g) was dissolved in water (200 ml) andevaporated to the consistency of a coarse, free-flowing sandwith an average moisture content of 15 %. Thi s materialwas ground directl y through a U.S. Standard Testing SieveNo . 45 : The ground substance was then sifted using a mechanical sifting mach ine through a No.60 sieve to give auniformly sized product (300 ± 50 micron) .
Gel Permeation Chromatography. Gel permeation chromatography was conducted on a Perkin Elmer LC-AnalystHPLC system with a PE LC-235 photo diode array detector, column PL Gel 500 (7.5 x 300mm), mobile phase 95 %

SP-303 83
0.12
II
\\.----
0.11
0 .10
0.09
0.08
0.07
0.06 J
0.05 J
0.04 I
I0.03 -
IIII!
)
1(JIJ11 ·lIJSttml ,..., i.-(. u: hil,.n \1ln'. U l "'"
5.IiIIlt 1C!SIIrI INlm .. lt hilrsts,st..,I
JltOm •••••• "-Ql~ 1I)11C't.l ; , rt rCl 11~:n: tS t!l1Axllhtll hl re 111S:n :tS t!rl
' Lll 7 _ 1 11~1iatt ••• ''' ' ~M ••• I0\n-e1 .. •. . 11 Dlhl\JtIl ••••••• l
lanat , . . .. 1 fcnat 11U• . . . lUll SIXl
II. art.( .... "l~ I: n HitLII: II_ Pwca-c. ~
1 7.111 ~ ~. 11 IQ:I .lKIXl llXUOX
114.l!a lou l WtIl. II IOU I ~ l :t :
o 5
Time (min)
10 15 Fig. 2. Gel. Permeation Chromatogramof SP-303.
ag. THF, flow rate 1 mllmin; diode arra y detector set at 280nm, Analysis of molecular weight range and distribution ofSP-303 was conducted using two PL Mixed E columns(7.5 x 300 mm) connected in series; mobile phase 95 % aq.THF; flow rate 1 milmin; detector 280 nm. Chromatographic fractions were diluted 1 : 9 with THF containing 0.1 %BHT before analysis. SP-303 and dried intermediates weredissolved in 95% THF containing 0.1 % BHT (1 mg/ml) before analysis.
Spectroscopy
Mass Spectrometry: Mass spectra were acquired on thefirst two sectors of a lEOl HX/HX110A tandem four sector mass spectrometer, which was operated at -10 kV accelerating potential. Spectra are averaged profile data of 3scans, which were acquired from 300 to 5000 m/z at a ratethat would scan from 1 to 6000 m/z in one minute as recorded by a lEOl DA5000 data system. Ions were produced by FAB using a lEOl FABgun operated at 6 kV in aconventional FAB ion source. A 3: 1 mixture of dithiothreitol and dithioer ythritol was used as the FAB matrix. Allmeasurements were performed on negatively charged ions.Calibration was performed using CsIlglycerol. Mass spectra were also provided by a Kratos MS50 mass spectrometer equipped with a Xe FAB source. The negative ion FABexperiment was performed with a mass resolution (M/LlM)of 1000, using CsI as calibrant.
NMR: lH and BC NMR spectra were obtained on a
400 MHz Varian instrument in D20 operating at 400 and100 MHz, respectively.
IR: Infrared spectra were obtained on a Perkin-Elmer1605 FT-IR instrument.
CD: Circular dichroism spectra were acquired on alASCO 702 instrument in both H20 and phosphate buffer.
Physical Properties of SP-303
Appearance: dark red-brown powder, light brown fluffysolid after lyophilization.
Solubility: Soluble in H20 and 5 % dextrose up to 50 mg/ml.
Melting point: > 500 "C (decomp.)Elemental analyses: %C 56.1-56.7, %H 4.6-4.9, %N <
0.5, %CI < 0.5, %S < 0.9Specific rotation: [a] D 0.0 0 (H20 , conc.: 0.22, 3.33 &
15.5. g/100 ml).Circular Dichroism: Amax =299 to 305 nm, M:: -1.02 to
1.10; Amax =255.5 to 256. 7nm, .1£: +0.452 to +0.658;(cone. 9.76 x 10-4 to 9.76 X 10-6 M in 0.1 M phosphate buffer @pH 6.7, respectively); Amax = 212.5 nm, Ll£: -2.815;Amax =236 nm, Ll£: +2.774 ; (conc. 385 x 10-6 M in H20 ).
UV-VIS: Amax: 205 , 275 and 460nm (H20 , Abs. 1.07,1.65, 0.25 @conc. 0.0055, 0.2 and 1.0 mg/ml).
FT-IR: 3380, 1690, 1610, 1570, 1450, 1343, 1200,1143, 1100, 1065, 1033, 825 and 725 ern" (KBr press,cone 2 mg SP-0303/200 mg KBr).

84 R. Ubillas et ai.
below, the structure of SP-303 has been assigned as a heterogeneous, oligomeric pro anthocyanidin (condensed tannin) having an average chain length of seven units (rangingto 11), the monomers of which are the four related catechinand gallocatechin stereoisomers. This results in an averagemolecular weight of -2100 a.m.u. A very small amount ofa sub-unit with an anthocyanin character is also present based on UV-VIS spectral dat a, but its precise location withinthe individual chains remains to be determ ined.
The experimental range of cis/trans (catechin/epicatechin) stereochemistries was shown to cover from 1: 2 to
2 : 1, employing l3C-NMR, CD and specific rotation data.The absolute stereochemistries of the monomers determined from CD spectral data are consistent with the 2S stereochemistry of the monomers [(+l-catechin, (+)-gallocatechin, H -epicatechin and (-)-gallepicatechin]. The oligomerunits are formed by linkages of monomers through 4-6 andlor 4-6 and/or 4-8 C-A ring positions, but the exact linkage pattern could not be determ ined spectroscopically.
Spectral Data: The UV dat a of SP-303 are closely relatedin appearance to those of the known proanthocyanidin s.However, the visible dat a are clearly different. The knownpro anthocyanidins are colorless (A 205, 240 and 275 nm)having no absorption in the visible range, while SP-303 is acolored compound with UV absorption in H20 at Amax 205(Abs. 1.07@ 5.5mg/ml ) and 275 (Abs. 1.65 @ 0.2mg/ml )
nm virtually identical to the known proanthocyanidins, butwith additional weak visible absorpt ion at 460 (Abs. 0.25@ 1.0 mg/ml) nm. Structurally, the visible data of SP-303suggest the presence of a flavylium moiety within the proanthocyanidin polymer, accounting for the observed redcolor. This is consistent with the visible spectroscopic datareported (Markham, 1982) for the closely related red-colored anthocyanins, which contain the flavylium/quinonemoiety (A 460-560 nm). Also, the visible absorption at A460 nm is ~ 500 times weaker in absorbence than the ultraviolet absorption at Amax 205 nm; this indicates that thechromophore is present at trace levels « 1 %) . Thus, whilecontributing the characteristic color, the f1avylium moietyrepresents a very minor portion of the overall structure ofSP-303.
The FT-IR data show a strong, broad band for OH stretching at v 3379; C=C stretching band s appropriate forphenyl units at v 1690, 1610 and 1570, phenolic C-OHstretching vibration at v 1200 , C-OH stretching bands at v1143 , 1102 , 1065, 1033, and a band at em" 826 consistent with a 1,2,3,5-tetra substituted benzene ring. The dataare consistent with a polyphenol structure for SP-303.
The l3C-NMR data of SP-303 are indicative of the proanthocyanidin class of polymers. This can be seen when thel3C NMR spectrum of SP-303 is compared to various published 13C NMR spectra of proanth ocyanidins (Harborneand Mabry, 1982; Haslam, 1989). In particular, the l3CNMR chemical shift data of SP-303 are consistent with anoligomer composed of both procyanid in [3',4'-dihydroxy-
366
w ,H
330 350 366 ..
om
om
...........--- - - - - - - -1
~-="""- - - 1
274 .3 290 310
•. u ' • "Cll i t " L" aJ. 1 I
250
274 .3 290 300 310 320 330 340 350
HII
215 230
250 260
II
file (ri l l Display Opt ions Inst".,enl s lash Status fmlVSI
Fig. 3. Diode Array Spectra of SP-303. Spectra taken at PeakApex, 20 % Upslope, and 20 % Downslope.
\H --
H --
.,\
HIe (rlit Display ~tions Inslnooents lesks Stat llS ImlVSl
IH-NMR: 15 in D20 @ 400 MHz: 2.2-3.0, 3.2-5.4,5.5-7.3 (all pekas very broad, ref. to HOD at 15 4.63).
13C-NMR: 15 in D20 @ 100 MHz: 28.7, 36.4, 38.2 , 66.7,72.6, 73.7, 76.6, 78.8, 82.9, 96.5, 97.6, 107.4, 109.2 ,115.5 , 116.8 , 120.0, 121.1, 13104, 133.9, 146.2, 15504,157.6 (all peaks broad, ref. to 1,4-dioxane @ 15 67.6).
Evidence for Structural Assignment. Extensive effortsemploying a wide range of anal ytical techniques have beendirected at determining the structure of SP-303. This workhas succeeded in identifying the types of monomeric unitspresent in the polymer, the stereochemical ratio of compo nent monomers as well as their absolute stereochemistries,and the range and distribution of chain lengths within theoligomer. Based on the inform ation obtained as described

phenyl] (C-1' = 133.9 ppm, C-2' = 115.5ppm, C-3' & C-4'= 146.2 ppm, C-5' = 116.8 ppm, C-6' = 120.0 ppm) andprodelphinidin [3' ,4',5'-trihydroxyphenyl] (C-1'131.4 ppm, C-2' & C-6' = 109.2 ppm, C-3' & C-5' = 146.2ppm, C-4' = 133.9 ppm) B-ring moieties with the individual flavanol C-ring units possessing both the 2,3-trans-3,4trans [similar to catechin/gallocatechin; C-2 = 82.9 ppm, C3 = 72.6 ppm, C-4 = 38.2 ppm] and 2,3-cis3,4-trans [similar to epicatechin/galloepicatechin; C-2 = 76.6 ppm, C-3 =73.7ppm., C-4 = 36.4ppm] stereochemistries when compared to literature NMR assignments of related compounds (Haslam, 1989). The A-ring 13C-NMR data of SP303 (C-4a = -102 ppm (buried), C-5 = 157.6ppm, C-6 =97.6 ppm, C-7 = 157.6 ppm, C-8 = 96.5 ppm, C-8a =155.4 ppm) closely match published phloroglucinol (5,7dihydroxy)-substitution NMR data (Harborne, 1988).However, the 13C-NMR data do not allow for unambiguous assignment of 4-6 and/or 4-8 linkages between theindividual units within the SP-303 polymer due to peakbroadening and peak overlap in the 100-110 ppm regionof the 13C-NMR spectrum (the chemical shifts of C-6 andC-8 when C-substituted lie between 106-108 ppm). Theterminal unit is consistent with the published epicatechin/galloepicatechin (C-2 = 78.8 ppm, C-3 = 66.7 ppm, C4 =28.7ppm) 13C NMR data for related compounds.
The multiplicities [13C NMR: s in DzO @ 100 MHz:28.7, t; 36.4, d; 38.2, d; 66.7, d; 72.6, d; 73.7, d; 76.6, d;78.8, d; 82.9, d; 96.5, d; 97.6, d; 107.4, d; 109.2, d; 115.5,d; 116.8, d; 120,0, d; 121.1, d; 131.4, s; 133.9, s; 146.2, s;155.4, s; 157.6, s (all peaks broad, ref. to l,4-dioxane @ 867.6)], as determined by the Distortionless Enhancementthrough Polarization Transfer (DEPT) spectra, are furtherverification of the 13C-NMR assignments based on chemical shifts as described above. The absence of small peaks inthe range 8 100-115 in the DEPT, when compared to thenormal broad-band decoupled spectrum, is consistent withtheir assignment as quaternary carbons buried under theprotonated carbons in this region of the normal 13C-NMRspectrum.
An Inverse-Gated Decoupled 13C-NMR experiment, performed with SP-303, removes the nuclear Overhauser effects (nOe) normally observed on individual lC peaks, andthus allows for an accurate integration of the 13C-NMRspectrum. According to previous research, this informationallows measurement of the relative ratios of cis and transstereochemistries of the C-ring monomeric units throughdetermination of the integrated area for the C-2, C-3 peaksof catechin (lit. ()83.4, 73.1; obs. ()82.9, 73.7) and epicatechin (lit. 8 76.6, 72.6; obs. () 76.6, 72.6). Due to overlap,only the 8 82.9 C-2 signal of catechin and the 8 76.6 signalfor C-2 of epicatechin are useful in determining the ratio,which is -2:1 in catechin!epicatechin. The method is alsouseful in determining the ratio of the terminal monomericunit to the rest of the units in the oligomer by looking at theratio of the C-2 signal of the terminal unit to all the other
SP-303 85
C-2 signals in the compound. These signals are readily distinguished both by chemical shift and by multiplicity asdetermined by DEPT (doublet at 8 36.4,38.2 vs. a triplet at()28.7). Using the relative areas of these signals, the ratio ofterminal unit/internal units becomes -1:6, which is consistent with an heptamer being the average chain length ofthe polymer. In summary, integration of selected 13C-signalsdetermined SP-303 to be a proanthocyanidin polymer withthe relative stereochemical ratio of approximately 2:1 catechin!epicatechin, as well as an average length of sevenunits.
The lH-NMR spectrum is notable for the extremebroadness of the resonances, which is consistent with theoligomeric nature of SP-303. The three very broad lH-peakscan be assigned on the basis of chemical shifts as follows: ()2.2-3.0, -CHz adjacent to an sp2 carbon, H-4 for the terminal unit; ()3.2-5.4, -CH adjacent to two sp2 carbons forall internal H-4's & -CH-O, H-2 for all units; 8 5.5-7.3,aromatic -CH for all units. The observed line-broadeningprecludes the application of 2-D NMR experiements suchas COSY, NOESY,]-Resolved, etc.; these were tried withoutsuccess under a variety of conditions on SP-303 in DzO,MeOH, DMSO, aetone, pyridine, and combinations thereof, at temperatures ranging from 20 to 80°C. Raising thetemperature, as well as lowering the viscosity of the NMRsolvent did little to sharpen the lH-NMR spectra. This behavior is consistent with many polymeric materials.
A slightly better situation was found in a two-dimensional heteronuclear lH_13C one-bond experiment, Heteronuclear Multiple Quantum Correlation (HMQC), inDzO. The spectra were poorly resolved because of thebroadness of both the parent lH_ and 13C-NMR spectra.However, the data did confirm the proton assignments made above on the basis of lH chemical shifts. For example,the cross-correlation peaks observed confirmed the protonassignments of () 5.5-7.3 as aromatic protons, attached to
carbons ranging from () 96-121. This verified the proton!carbon assignments, and also is completely consistent withthe structural features of the C-ring, as well as the A- and Brings found within the oligomer.
The mass spectrometry of SP-303 proved to be challenging, and many methods and instruments were involved inthe attempt to get significant results. After failing to obtainspectra using CIMS (Chemical Ionization), FDMS (FieldDesorption), SIMS (Secondary Ionization) TOFMS (Timeof Flight) and a variety of FABMS (Fast Atom Bombardment) techniques, and after talking with experts in the areaof proanthocyanidin mass spectrometry (Prof. Barofsky,pers. comm.), we decided to use negative ion FABMS utilizing a matrix consisting of a 5:1 mixture of dithiothreitoland dithioerythritol known as "magic bullet". The spectra(Figs. 4, 5) were obtained under many different conditionsbut were consistently too weak to perform high resolutionand many MS/MS measurements. However, the spectra didallow for confirmation of a gross structure of the proantho-

86 R. Ubillas et al.
913138131371313613135131340a
0ll2 . 7
0
13
0
£I613c .9457 . 9
• 91}" 93~ .4 761.8-llJl. J. ~ j .l J. j j. ...I. 1 .. "'" ... ..I. .. ..6.
" . .. . .. ...~
~ aI ,
2
8
tlUC
'" 6"0C:l
.D
'"tl> 4'c;tl
0:::
18
3
Fig. 4. Negative ion fast atom bombardment mass spectrum (FABMS) of SP-303 in magic bullet mat rix from 300 to 1000 mlz.
2S-,--- - - - - - - ------------------------- -y
213
tlUC
'""0 l Sc:l
.D<'l
tl>'c;tl
0:::
1213 . 1 1594.6 1813 . 7
209 .9
Fig. 5. Negative ion fast atom bombardment mass spectrum (FABMS) of SP-303 in magic bullet matrix from 1000 to 4000 miz,
cyanidin oligomer made up of catechinlepicatechin andgallocatechinlepigallocatechin units. Examinatio n of theFABMS spectrum from 1000 -4000 m/z (Figure 5) revealsclusters of odd-numbered peaks corresponding to M-Hpeaks for a pentamer (5 x approx. 300 mol. wt. of monomer) at m/z 1505 (reI. abund 15), a hexamer (m/z 1809, reI.abund. 14), heptamer (m/z 2097, reI. abund. 19) an octamer (m/z 2401, reI. abund. 12), a nonamer (m/z 2689, reI.
abund. 8), a decamer (m/z 2993, reI. abund. 7) and an undecamer (m/z 3297, reI. abu nd. 5). The even-numberedpeaks seen in Figures 4 and 5 at m/z 1200, 916, 762, 610and 458 are apparent fragments of larger parent(s). Also,each peak corresponding to an individual unit, for examplethe M-H peak at mlz 1809 for a hexamer, is really a clusterof peaks each differing by 16 a.m.u , or oxygen (i.e., M-Hm/z 1777, 1793, 1809, 1825) as shown in Figure 6. This is

SP-303 87
30 179 .17
1809 .1825 jliU 1777.20
182117ue<::l
"0 20c::l
I..0
~ ! Iii<::l
u
1\;;- 15
"<=J i"176 .22 I Ii
iU
Ili f! I'0:: Hl
!~
~5 I I ~1~ I i I I I I . I I -=r==;:---
1750 1760 17713 1780 1790 1800 1810 1820 18313 18 <:0
t'V Z
Fig. 6. Enlargement of the negative ion fast atom bombardment mass spectrum (FABMS) of SP-3D3 in magic bullet matrix form.
consistent with the hexamer being made up of varying ratios of B-ring oxidations (procyanidinlprodelphinidin),with the prodelphinidin occurring as the dominant oxidation pattern (i.e., 1778 a.m.u. corresponds to 3:3 procyanidinlprodelphinidin units, 1794 a.rn.u, corresponds to 2:4procyanidin/prodelphinidin units, 1810 a.m.u. correspondsto 1:5 procyanidinlprodelphinidin and 1826 a.m.u. corresponds to 0:6 procyanidinlprodelphinidin-entirely prodelphinidin B-rings). If the assumption is made that the surfaceactivitity and ionization energies of the individual units inthe matrix are not substantially different, then the relativeabundances of each of the parent M-H ions given abovecan be used to estimate relative percent of each of thevarious pentamer-to-undecamer oligomers in SP-303. Theaverage ratio of procyanidin/prodelphinidin units determined by this method reflects a predominance of prodelphinidin B-ring oxidation in SP-303. The above negativeion FABMS data define a range of oligomers for SP-303 upto the undecamer, each made up of catechinlepicatechinand gallocatechinl epigallocatechin monomers. The rangeand average molecular weight distribution of SP-303, clearly a heterogenous oligomer, are in accord with those determined from the 13C NMR data. Overall, the MS data are inagreement with the 13C-NMR data and the HPLC-GPCmolecular weight determination by substantiating an average oligomer chain length of seven, and an average molecular weight of approximately 2100 a.rn.u.
Elemental analyses were performed on samples of SP303. dried at 35-40°C under high vacuum. To avoid thepossibility of decomposition, no attempt was made to remove all the H20 (e.g., by using P20S under high vacuum
and high temperature conditions) so that typically, 3-7%H20 remained after drying or lyophilization. The analysesare consistent with the following average formula for SP303: (C15H120 6.S)n . 8 H20 , which represents approximately 7% moisture. The calculated %C (56.8%) and %H(4.6%) using the formula are consistent with the observedranges (%C 56.1-56.7; %H 4.6-4.9). It should be notedthat the average formula is in accord with the mass spectralobservation that SP-303 is made up of varying ratios ofprocyanidinlprodelphinidin units. This is reflected in theformula ClsH1206.S, indicating an average distribution ofprocyanidinlprodelphinidin units with C15H120 6 for an allprocyanidin oligomer and C1sH120 7 for an all prodelphinidin oligomer.
The relative stereochemistry of the internal units of SP303 was determined from the 13C-NMR discussion aboveas consisting of -2: 1 catechin (2,3-trans-3,4-trans )/epicatechin (2,3-cis-3,4-trans) stereochemistries, with the terminalunit having the epicatechin (2,3-cis) configuration. Whatremained was to examine the absolute stereochemistries ofthe units themselves. Virtually all of the natural occurringf1avanoids including proanthocyanidins have the same absolute configuration at C-2, apparently following a common biosynthetic pathway conserved in widely differingplants. The two stereochemical forms commonly found area 2,3-trans isomer (2R,3S) as in (+ )-catechin, and a 2,3-cisisomer (2R,3R) as in (-)-epicatechin. Based on biosyntheticconsiderations, SP-303 consists of monomeric units madeup of (+)-catechin, (+ )-gallocatechin, (-)-epicatechin and (-)galloepicatechin. To confirm this, two sets of experimentswere performed. The specific rotation of SP-303 was mea-

IoJVv'
88 R. Ubillas et aI.
HOI
oJVv'
OH
OH
....6(HOH
HO
R = H, OHOH
techin, with (+)-gallocatechin and (-)-galloepicatechin predominating. The sequence of the individual monomers witheach chain could not be determined. Also, the visible spectral data strongly indicate the presence of a f1avylium (anthocyanidin) moiety, although the relat ively small fraction(<1% ) of this component has been an impediment to determining its position within the oligomer. The average chainlength of SP-303 has been determined by 13C-NMR via integration of the terminal unit/internal units as being a heptamer. Negative FABMShas also allowed estimation of the re-
sured in H20 over a range of concentrations, without anyrotation observed. By itself, this suggests that SP-303 isachiral or racemic. However, the 13C NMR shows an average of 2:1 catechinlepicatechin (trans/cis) stereoisomers asdescribed previously. Using the specific rotation, the molefraction of the cis isomer X cis can be calculated by the following formula: X cis = ([a] 578 + 320)/494 from the literature, to give Xcis =(0 + 320)/ =0.65, which translates into a2:1 ratio of cis/trans (which is the same as a ratio of 1:2 catechinlepicatechin). When coupled with the 13C-NMR data, the specific rotation data just described give an experimental range of X cis = 0.33-to-0.66 (±0.05). CD spectrawere obtained to confirm the chirality inherent in SP-303.The circular dichroism spectra confirm the chirality ofSP-303. In addition, the sign of the 280 nm circular dichroic bands is in accord with the C-2 stereochemistry assignedas 2R from literature precedent with other f1avanols, confirming (+)-catechin, (+)-gallocatechin, (-)-epicatechin and(-)-galloepicatechin as the units making up the oligomerSP-303. Additionally, based on precedent, the positive CDat 236/212 nm indicates that the 4b stereochemistry[(-)-epicatechin & galloepicatechin or cis] is dominant overthe 4a stereochemistry [(+)-catechin &-gallocatechin ortrans]. Thus, the CD data support the interpretation of thespecific rotation data given above. In summary, the experimentally determined range of monomer stereochemistry ofSP-303 can be defined as Xcis = 0.33-0.66 of (-)-epicatechin& galloepicatechinl(+)-catechin & gallocatechin.
Structure Conclusions: The basic components of SP-303, aheterogeneous oligomer, have been established as (+)-catechin, (+)-gallocatechin, (-)-epicatechin and (-)-galloepica-
HO
OH
HO
R
h OH
...··UOH
R
h OH
...··UOH
OH
NR
h OH
...··UOH
OH
N =3-9R = H. OH
HO
OH
R =G =CatechinR = OH = Gallocatechin
HO
OH
R =H =EpicatechinR = OH = Epigallocatechin

lative proportions of oxidation within each of the chainlengths as well as their size range (up to Ll-mer). The sum ofthese data supports an average molecular weight of 21 00.
The relative stereochemistry is shown to range from 2:1to 1:2 ratios of catechin to epicatechin substructures, withthe terminal unit having the epicatechin structure. The absolute stereochemistry determined from CD is consistentwith the majority of naturally occurring flavanoids atC-2. Interestingly, the specific rotation of [a]578 = 0
0
isconsistent with the CD data, confirming the chiral natureof SP-303.
III. Antiviral Studies
Although a variety of plant derived drugs are used successfully in the clinic today, with the exception of podophyllotoxin, no other plant-derived antiviral agent is approved by the FDA for commercial use (Rozhon et aI.,1994). The philosophy of Shaman Pharmaceuticals is toisolate and evaluate pharmacologically active componentsin medicinal plants that have ethnotherapeutic indicationfor certain diseases. Initial efforts in this area resulted in thediscovery of SP-303, a naturally occurring proanthocyanidin oligomer (average molecular weight 2,100 amu). Thelarge body of virological preclinical data obtained onSP-303 is presented below. Some of the data in the reporthave been published previously (Soike et aI., 1992; Gilbertet aI., 1993; Wyde et aI., 1993a & 1993b).
Materials and MethodsViruses and cells. Table 1 lists the sources of RNA and
DNA viruses and cell lines used in this study. Antiviral activities presented here as 50% inhibitory endpoint or 50%effective dose (EDso) are in ug/ml.
Antiviral activity in cell culture. The antiviral activities ofSP-303 and known antiviral control compounds were determined using the viral cytopathic effect (CPE) assay, theplaque neutralization assay, the hemagglutination-inhibition (HAl) assay, and/or the MTT antiviral assay: (i) ViralCPE assay. This assay has previously been described (Wydeet aI., 1993b). (ii) Plaque neutralization assay. The procedure used for the plaque reduction assay was adapted fromthat of Rozhon et al. (1993). Briefly, test compound wasmixed with virus and incubated 1 hr at 22°C before addition to a cellular monolayer (60 mm diameter). Viral adsorption was performed at 3rc for 90 min before the viralinoculum-test compound mixture was aspirated and themonolayers washed with phosphate buffered saline. Anoverlay medium consisting of 0.76% carboxymethyl cellulose, MEM cell culture medium and 2% fetal bovine serum(56°C, 30 min) was added to the cells and incubation wascontinued for 6 days at 3rc. Plaques were visualized by
SP-303 89
staining with crystal violet (0.5% crystal violet, 10% formalin: methanol, 1:1). (iii) HAl assay (for influenza virus Aonly): This assay was performed using a protocol similar tothat described by Dowdle et al. (1979). Briefly, cells thathad been treated with test agent were infected with FLU-Ain 96-well round bottom microtiter plates and the cells were incubated at 37°C. At the end of the day 5 of incubation,an equal volume of 1% washed chicken erythrocytes (BioWhittaker) was added to the culture medium. The platesthen were incubated at 4°C for 1 hr and the individualwells scored for hemadsorption activity. (iv) MTT antiviralassay. This assay was used exclusively for evaluating the activity of antiviral agents against hepatitis A virus (HAV). Asdescribed in the following section, MIT is usually used toassess adverse effects of potential antiviral agents in uninfected cells by measuring cellular viability. However, withHAV-infected cells, MIT was also used as an indirect method to measure the inhibitory activity of antiviral agents;the reagents and protocols used for both assays are identical. Finally, the assay employed to test the activity ofSP-303 against hepatitis B virus (HBV) has been described(Korba and Milman, 1991; Korba and Gerin, 1992).
Selectivity index (SI). An SI (50% cytotoxic concentration +50% antiviral concentration) for each virus testedwith SP-303 or known antiviral control compound in cellculture was calculated using the 50% cytotoxic endpointthat was determined in uninfected cells in an assay run inparallel with the antiviral assay. The cytotoxic measure ofantiviral agents was determined either by visual inspectionor by the MTT assay (Mossman, 1983). In either case, the50% cytotoxic endpoint, 50% inhibitory dose (ID so) is given in ug/rnl, For the MTT assay, a 96-well plate containingthe appropriate cells was incubated with different concentrations of antiviral compound. At the end of the incubation period, the effect of test compound on cellular viabilitywas evaluated by adding 25 ug/ml MTT (5 mg/ml stock inPBS) to each well and incubating for 3 hr at 37°C. The culture medium-MTT mixture was aspirated and the MTTproduct, formazan, was solubilized with addition of 0.1 ml0.04 N HCI in isopropanol. Adsorption at 570 nm was determined for each well using a mikrotiter plate-readingspectrophotometer. A dose-response curve was generatedand the concentration of test compound giving 50% absorption compared to the untreated control represented theICso• In addition, uptake and incorporation of radiolabelled biochemical precursors was used as an independentmeasure of the potential adverse effects of SP-303 in uninfected cells.
Virucidal test: RSV (Long) and SP-303 (0-1000 ug/ml)were mixed and incubated for 1 hr at 37 "C, subjected to serial tenfold dilution, and titered for infectivity in HEp-2cells using the viral CPE assay.

90 R. Ubillas et al.
Table l. Listing of Viruses and Cell Lines Used to Test SP-303 and Their Sources.
Virus Source Cells Source Cell Origin
RespiratorySyncytial Virus HEp -2 ATCC hum. epid. carcinoma
MA-I04 Bio Whittaker Af. green monkey kid.RSV A (Long) ATCCRSV A (Tracy) Baylor U.!RSVA(WydelHouston/88) Baylor U.RSV B (46791) Baylor U.RSV B (47063) Baylor U.RSV B (18537) Baylor U.
Influenza Virus A MD CK ATCC canine kidneyA54 9 ATCC hum. lung carcinoma
FLU A Taiwan(HI Nl) Baylor U.FLU A NWS/33(HI Nl) ATCCFLU A Japan/305157(H2N2) ATCCFLU A Port Chal. 1/73(H3N3) ATCCFLU A Leningrad/86(H3N2) Baylor U.
Influenza Virus B MDCK ATCC canine kidneyFLU B USSR/I00/83 Baylor U.FLU B Yamagata/16/88 Baylor U.Hong Kong 5172 WHO!
Herpes Simplex Virus human foreskinType 1 HFF Clonetics fibroblast
MA-104 Bio Whittaker Af. green monkey kid.MDCK ATCC canine kidney
HSV·l 10186 (TK+) Well Res!HSY·l KOS (TK+) Well ResHSY· l 10166 (TK+) Well ResHSY· l McKrae 555A A.(TK+) NesburneHSV-l ACGr4 (TK) S. SachsHSY·l SC16-R5C1 (TK-) S. SachsHSY·1 BWVL 10168 (TK) Well Res
Herpes Simplex Virus human foreskinType 2 HFF Clonetics fibroblast
MA-104 Bio Whittaker Af. green monkey kid.MDCK ATCC canine kidney
HSY·2 MS ATCCHSY·2 £ 194 (TK+) M. FialaHSY·2 10167 (TK+) Well ResHSY·2 10165 (TK+) Well ResHSY·2 10634 (TK+) Well ResHSY·2 Cl 1252 (TK') S. SachsHSY·2 Cl1182 (TK') S. SachsHSY-2 10161 (TK-) Well Res
Human Cytomegalo-virus (AD-169) ATCC MRC-5 ATCC human lung

Table 1 (continued).
Parainflu enza Virus LLCHEp-2
PIV-l Baylor U.PIY-3 Baylor U.PIY-3 C243 SRP
Measles Virus Baylor U. Yero
Adenovirus-S Baylor U A549
Hepatitis B Virus George-'town U. 2.2.15
Herpes Z oster Utah!(Varicella) Virus State U. HFF
MRC-5
ATCCATCC
ATCC
ATCC
CloneticsATCC
SP-303 91
Rhesus monkey kid.hum. epid. carcinoma
Af. green monk. kid.
hum. lung carcinoma
HBYDNA transfectedhuman livercells
human foreskinfibroblastshuman lung
! Abbreviations used in this table include: WHO, World Health Organization; Well Res, Wellcome Research Laboratories; SRI, Southern Research Institute; Baylor U., Baylor University; Utah State U., Utah State University, Antiviral Research Institute; GeorgetownU., Georgetown University.
2 Sells et aI., 1987.
Mechanism of action studies
Mechanistic studies with SP-303 comprised time-of-addition of test compound relati ve to the viral infectious cycle,effect of test compound on attach ment and penetration ofvirus, and effect of test compound on vira l uncoating: (i)Time- of-addition studies: Test compound was added tocells at various intervals either before or after infection. Depending on the study, test compo und was present throughout the incubation period follow ing its addition, or thecompound was aspirated after a defined period of incubation . Viral replication was assessed by plaquing, except inone study with HSY-1 in which inhibition of viral DNAsynt hesis was the end point (Shipman et aI., 1976; Smee etaI., 1983). (ii) Effect on viral attachm ent: The method employed was a modification of th at of Taylor and Cooper(1989). Briefly, [35S] methi on ine labeled puri fied virus wasincubated with cells for 1 hr in the presence of test compound befor e washin g the cells and determining cell-associated radioactivity by liqu id scint illation spectrophotometry. (iii) Effect on vira l penetrat ion through the plasmamembrane: Virus was adsorbed to cells at 4°C before addin g of test compound and ra ising the incubation temperature to 37° C. After 1 hr incub ation , unadsorbed viru s wa saspir ated and an y rem aining virus was neutralized by theaddition of neutralizing virus antibody. The antiserum wasrem oved and th e resulting virus was quantitat ed by plaqu ing. (iv) Effect on viral unco at ing (HSY-1 only): The meth odology used was that of Feldman et al. (1981) and Harmenberg et al. (1991). Briefly, cells that were pretreatedwith cycloheximide (200 ug/ml ) were infected with [3H]thymidine labeled HSY-1 and incubated for 4 hr at 3rc.Infected cells were subsequently washed, lysed with NP-40,and the cytoplasmic and nuclear components were sepa-
rated by centrifugation at 10,000 x g for 30 min. Radioactivity associated with each fraction was determined byliquid scintillation spectrophotometry. Since herpesviru suncoats at the nuclear membrane, radioacti vity associatedwith nuclei would indicate th at uncoatin g occurred normally.
Determination of the affinity constant for SP-303 bindings to RS V. Methodologies using Scatchard analyses fordetermining affinity con stants for molecules binding to viral part icles have been described (Bennett, 1978; Fox et aI.,1991 ). Briefly, [3H] SP-303 (sp. act. 1llCi/llmol) at concentr ati ons ranging from 0.1-50 ug/rnl and 12.5 ug/ml of unlabeled SP-303 were added to centrifuge tubes that contained purified RSV. The contents of the tubes were mixedand incubated at room temp erature for 40 min. Viralbound SP-303 was separated from unbound SP-303 by subjecting the tubes to two successive rounds of ultracentrifugatio n at 150,000 x g for 1 hr at 4°C. Th e amount of SP303 bound to virus was determ ined by liqu id scintillationcounting. A Scatchard plot and the binding constant, Kd,
were dete rmined using the software program, K· cat™(Biometa llics, Inc. , Princeton , Nj), for ligand binding.
Evaluation of SP-303 in virus-infected animals. Studies inanima ls were performed as recommended in the Guide forthe Care and Use of Laboratory An imals (NIH Publication85-23). Relevant experimental details, including concentrat ions of test agents and regimen s, are provided for all virus/animals models in Table 2. (i) RSY-infected cotton ratsand parainfluenza virus -3: Infect ions of cotton rats andtreatm ents with antiviral test compounds are described byWyde er al. (1993a ). (ii) FLU A-infected mice: Descriptionsof FLU-infected mouse models have been described previously (Grunert et aI., 1965; Schulman, 1968; Walker etal., 1976; Hayden, 1986; Wyde et aI., 1986). (iii) HSV 1- &
92 R. Ubillas et al.
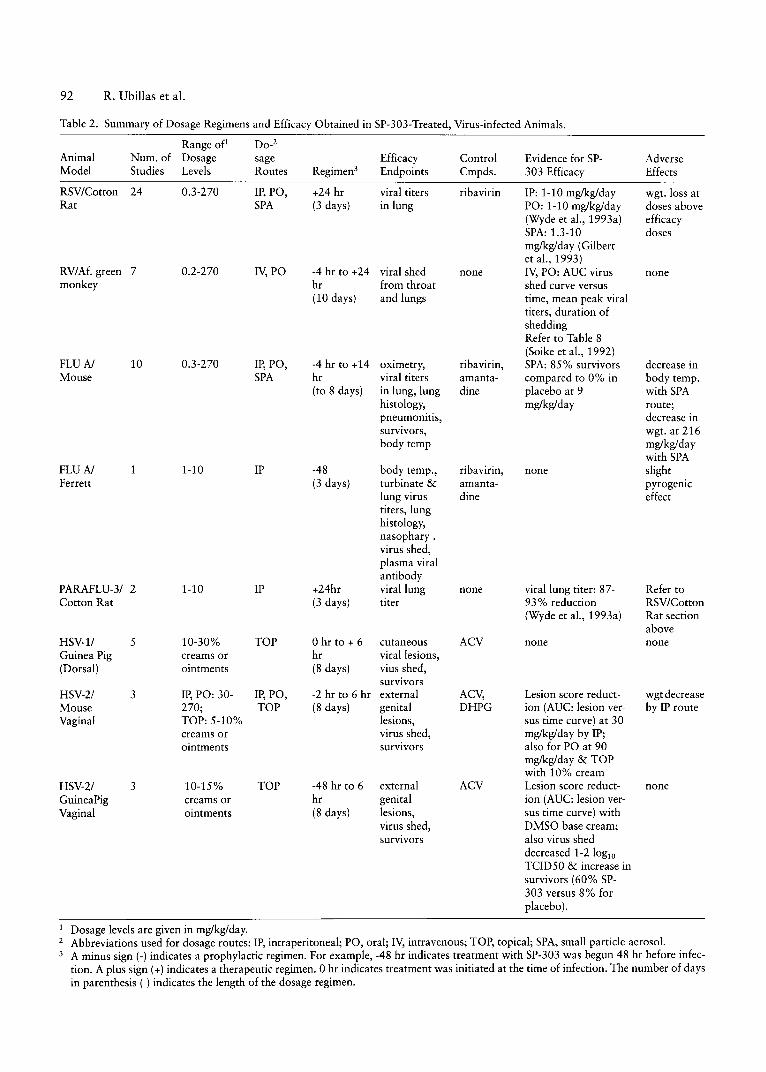
Tabl e 2 . Summary of Dosage Regimens and Efficac y Obtained in SP-30 3-Treated, Virus-infected Animals.
Range of Do-2
Anim al Num. of Dosage sage Efficacy Control Evidence for SP- AdverseM ode l Studies Levels Routes Regimen! Endpoints Cmpds. 30 3 Efficacy Effects
RSv/Cotton 24 0.3-270 IP, PO, +24 hr viral titers ribavirin IP: 1-10 mg/kg/day wgt.loss atRat SPA (3 da ys) in lung PO: 1-10 rug/kg/day do ses above
(Wyde er aI., 1993 a ) efficac ySPA: 1.3 -10 do sesmg/kg/day (Gilbertet aI., 199 3 )
RV/Af. green 7 0.2-2 70 IV, PO -4 hr to +24 viral shed none IV, PO : AV C virus nonemonkey hr from throat shed curve versus
(10 days) and lungs time , mean peak viraltiters, du ration ofsheddingRefer to Tabl e 8(Soike et aI., 1992)
FLUN 10 0.3 -270 IP, PO , -4 hr to +14 oximetry, ribavirin, SPA: 85% sur vivor s decrease inM ouse SPA hr vira l titers amanta- compared to 0% in body temp.
(to 8 da ys) in lung, lun g dine pla cebo at 9 with SPAhistology, mg/kg/day route;pneumonitis, decrease insurvivors, wgt. at 21 6bod y temp mglkg/day
with SPAFLUN 1 1-10 IP -48 bod y temp., ribavirin , none slightFerrett (3 da ys) turbinate & amanta- pyrogenic
lung virus dine effecttiters, lunghistology,nasophar y .virus shed,pla sma viralant ibo dy
PARAFLU-3/ 2 1-10 IP +24hr vira l lung none vira l lung titer: 87- Refer toCotton Rat (3 da ys) titer 93% reduction RSV/Cotton
(Wyde er aI., 1993a) Rat sectionabove
HSV-l / 5 10-30% TOP ohr to + 6 cut ane ous ACV none noneGuinea Pig creams or hr viral lesions ,(Dorsal) ointments (8 days) vius shed,
sur vivorsHSV-2/ 3 IP, PO : 30- IP, PO, -2 hr to 6 hr external ACV, Lesion score reduct- wgt decreaseMouse 270; TOP (8 days ) genita l DHPG ion (AVC : lesion ver- by IP routeVaginal TOP: 5-10 % lesion s, sus time curve) at 30
creams or viru s shed, mg/kg/day by IP;ointments survi vors also for PO at 90
mg/kg/day & TOPwith 10% cream
HSV-2/ 3 10-15% TOP -48 hr to 6 externa l ACV Lesion score reduct- noneGuineaPig creams or hr genita l ion (AVC: lesion ver-
Vaginal ointments (8 da ys) lesions , sus time curve) withviru s shed , DMSO ba se cream;survivors also virus shed
decre ased 1-2 logloTCID 50 & increase insurvivor s (60% SP-303 versus 8% forplacebo) .
I Do sage levels are given in mg/kg/day.2 Abbreviations used for dosage routes: IP, intraperitoneal; PO , or al; IV, intravenous; TOP, topical; SPA, small particle aerosol.3 A minus sign (-) indicates a prophylactic regimen. For example, -48 hr indicates treatment with SP-303 was begun 48 hr before infec-
tion. A plus sign (+) indicates a therapeutic regimen. 0 hr indicates treatment was initiated at the time of infection. The number of daysin parenthesis ( ) indicates the length of the dosage regimen.
SP-303 93
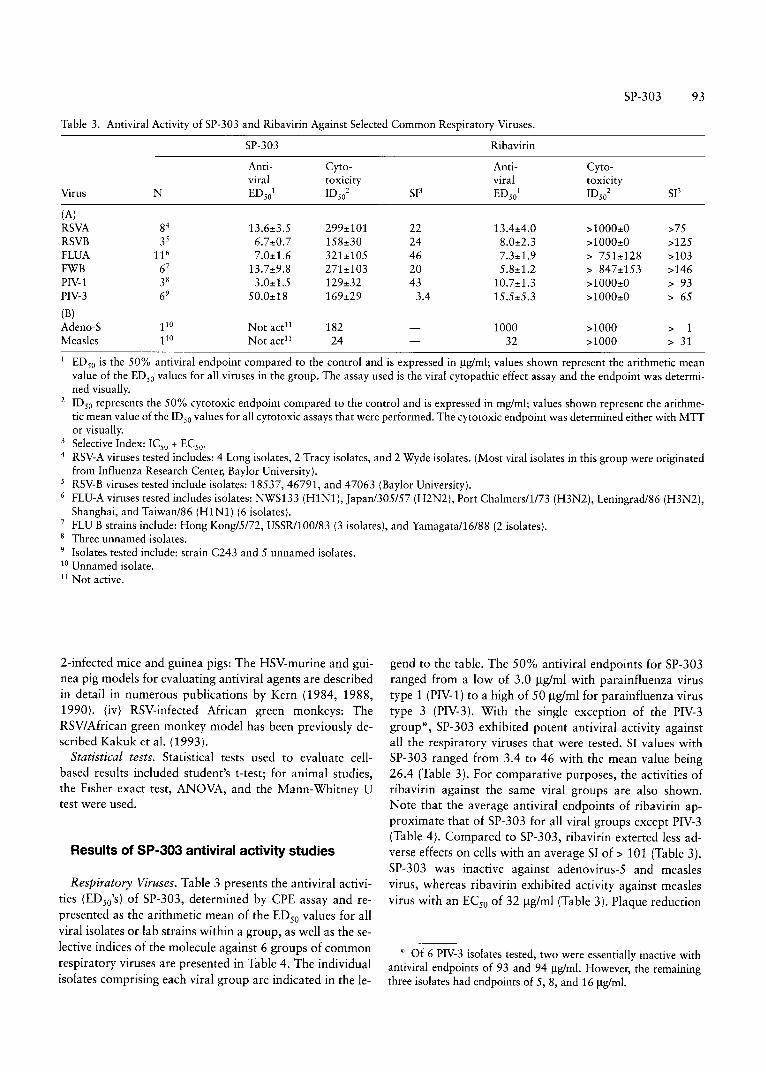
Table 3. Antiviral Activity of SP-303 and Ribavirin Against Selected Common Respiratory Viruses.
SP-303 Ribavirin
Anti- Cyto- Anti- Cyto-viral toxicity viral toxicity
Virus N EDso1 IDso2 SP EDso
1 m.,2 SP
(A)RSVA 84 13.6:!:3.5 299:!:101 22 13.4:!:4.0 >1000:!:0 >75RSVB 35 6.7:!:0.7 158:!:30 24 8.0:!:2.3 >1000:!:0 >125FLUA 116 7.0:!:1.6 32l:!:105 46 7.3:!:1.9 > 751±128 >103FWB 6' 13.7:!:9.8 27l:!:103 20 5.8±1.2 > 847:!:153 >146PIV-1 38 3.0:!:1.5 129:!:32 43 10.7:!:1.3 >1000:!:0 > 93PIV-3 69 50.0:!:18 169:!:29 3.4 15.5:!:5.3 >1000:!:0 > 65
(B)Adcno-S 110 Not act! ' 182 1000 >1000 > 1Measles 110 Not act !' 24 32 >1000 > 31
I EDso is the 50% antiviral endpoint compared to the control and is expressed in ug/ml; values shown represent the arithmetic meanvalue of the EDso values for all viruses in the group. The assay used is the viral cytopathic effect assay and the endpoint was determined visually.
2 IDso represents the 50% cytotoxic endpoint compared to the control and is expressed in mg/ml; values shown represent the arithmetic mean value of the IDso values for all cytotoxic assays that were performed. The cytotoxic endpoint was determined either with MITor visually.
3 Selective Index: ICso + ECso'
4 RSV-A viruses tested includes: 4 Long isolates, 2 Tracy isolates, and 2 Wyde isolates. (Most viral isolates in this group were originatedfrom Influenza Research Center, Baylor University).
5 RSV-B viruses tested include isolates: 18537,46791, and 47063 (Baylor University).6 FLU-A viruses tested includes isolates: NWS133 (HI Nl ), japan/305157 (H2N2), Port Chalmersll /73 (H3N2), Leningradl86 (H3N2),
Shanghai, and Taiwan/86 (H1N1) (6 isolates).7 FLU B strains include: Hong Kon g/SIn, USSR/100/8 3 (3 isolates), and Yamagatall 6/88 (2 isolates).8 Three unnamed isolates.9 Isolates tested include: strain C243 and 5 unnamed isolates.10 Unnamed isolate.11 Not active.
2-infected mice an d guinea pigs: The HSV-murine and guinea pig models for evaluating antiviral agents are describedin detail in numerous publication s by Kern (1984, 1988,1990). (iv) RSV-infected Afr ican green monkeys: TheRSV/African green monkey model ha s been previously described Kakuk et al. (19 93) .
Statistical tests. Statist ical tests used to evaluate cellbased results included student's t-test ; for an imal studies,the Fisher exact test, ANOVA, and th e Mann-Whitney Utest were used .
Results of SP-303 antiviral activity studies
Respiratory Viruses. Table 3 presents the antiviral acti vities (EDso's) of SP-303, determined by CPE assay and represented as the ar ithmetic mean of th e EDso values for allviral isolates or lab strains within a group, as well as the selective indices of the molecule against 6 groups of commonrespiratory viruses are presented in Table 4. The individualisolates comprising each viral group are indicated in the le-
gend to the table. The 50% antiviral endpoints for SP-303ranged from a low of 3.0 ug/rnl with parainfluenza virustype 1 (PIV-1) to a high of 50 ug/rnl for parainfluenza virustype 3 (PIV-3) . With the single exception of the PIV-3gr oup " , SP-303 exhibited potent anti viral activity againsta ll the resp iratory viruses that were tested. SI values withSP-303 ranged from 3.4 to 46 with th e mean value being26.4 (Table 3 ). For comparative purposes, the acti vities ofribavir in against the same viral gro ups are also shown.Note that the average antiviral endpoints of ribavirin ap proximate that of SP-303 for all vira l gro ups except PIV-3(Ta ble 4 ). Compared to SP-303, ribavirin exterted less adverse effect s on cells with an average 51 of> 101 (Table 3 ).SP-303 was inact ive against adenoviru s-5 and measlesviru s, whereas ribavirin exhibited activity against measlesviru s with an ECso of 32 ug/ml (Ta ble 3 ). Plaque reduction
.. Of 6 PIV-3 isolates tested, two were essentially inactive withant iviral endpoints of 93 and 94 ug/ml. However, the remainingthree isolates had endpoints of 5,8, and 1611g/ml.

94 R. Ubill as et a l.
Table 4. Antiviral Activity of SP-303 Against Respirator y Syncytial Virus! Using the Plaque Reduction Assay.
Antiviral .' Cyto toxicity" Selective"PFU Inoculum! EDso IDso Index
48136236366406730
3.33.03.53.04.84.6
83.283.24 1.683.241. 683.2
25281228
918
Ave. SI '"S.E.20", 7.6
1 Different amounts of vira l inocula (RSV,Long Strain; ATCC VR-26) were mixed with dilutions of SP-303 and incubated at room temperature for 1 hr before being added to HEp-2 cellular monolayers. After a 90 min adsorpt ion period, the SP-303/virus mixtur e wasaspirated from the cells, the monolayer washed, an overlay of carboxymethyl cellulose and MEM cell culture medium with 2% FBSwas added, and the monolayers were incubated for 6 days at 37°. Plaques were enumerated by staining with crystal violet.
2 Size of the viral inoculum in plaque forming units (PFU).3 Antiviral activity (EDsoin ug/rnl) is expressed at the 50% inhibitory concentration compared to the non-treated, infected contro ls.4 Cytotoxi city (IDso in lug/ml), detemined by MTT, is expressed as the 50% inhibitory concentration compared to the non-treated con
trols.s Selectivity index =ID so + EDso
TableS. Effect of SP-303 on Uptake and Incorporat ion of [3H] Labeled Precursors.
Average50% Inhibitory Endpoints]Uptake Incorporation
Leu101
Thy122
Urid104
Leu64
Thy65
Urid73
] 50% inhibitory endpoints are expressed in ug/rnl, The period of incubation of SP-303 with HEp-2 cells was 18 hours. Abbreviations:Leu, leucine; Thy, thymidine; Urid, uridine.
analyses using SP-303 and RSV confirmed the antivi ralactivity of SP-303 observed in the cytopathic effect assays(Table 4 ). Six individual assays employing different quantit ies of vira l inocul a resulted in antiviral endpo ints ran gingfrom 3.3-4.8 Ilg/ml and SI va lues, determined by MIT,ranging fro m 9-28 w ith a mean value of 20 .
In addition to visua l and MIT evalu at ion s to assess th epotential adve rse effects of SP-3 03, a third study was doneto measure the effect the compound has on cellula r uptak eand incorporation of ami no acid (leucine ) and nucleic aci d(uridine and thymidine) precursors (Table 6). When different concentrations of SP-3 03 were incubated with uninfected cells in the presence of ra diolabeled precusors, the concentrations th at inhibited 50% uptake and incorpora tionof these precursors were well above the 50% antivira l end points. Thus, the fact that th e antiviral endpoints of SP-30 3(Tables 4 & 5 ) were consistently less than those determinedin cytotoxity assays lends co mpelling evide nce to supportth e assertion th at th e antivi ra l activity of SP-303 ind ependent of th e cytotoxic effect o f the molecul e.
Herp esuiruses. The inhibitory activities of SP-303, deter-
mined by CPE assay, against either th ymidine kinase plus(TK+) or thymidine kinase minus (TK) herpes simplex virustypes 1 and 2 , are shown in Table 6. In addition, in Table 6,the antiviral activites of SP-303 against acyclovir resistant(ACVS)/fosca rnet resistant (FOS') herpes viruses, tha t wedetermined using th e plaque reduct ion assay, are shown.SP-303 was inactive against one strain of herpe s zoster (varicella ) virus which was test ed .
Usin g th e antivira l CPE assay (Ta ble 7), th e EDso va luesfor SP-303 agains t H SV-l , TK + of TK- st ra ins, range froma low of 3 ug/rnl to no activity at the high est concentrationtested. The average ECso for viruses in th e HSV-1 group is35.6 ug/ml, compared to 20.5 ug/ml for the HSV-2group, sugges ting that SP-30 3 has slightly greater potencyaga inst th e HSV-2 serotype. However, th e range of ECsovalues o bserve d for both serotypic groups was simi lar. Inhibition of H SV-1 strain 10168 and H SV-2 strains CI 11 82and CI 1252 suggests the po ssib ility th at SP-303, unl ikeacyclov ir, is act ive against TK- H SV strai ns. In reg ard to
po ssible adve rse effects of SP-303 on cel ls, the molecul eexerts a grea ter cytotoxic effect than acyclovir. W ith the ex-
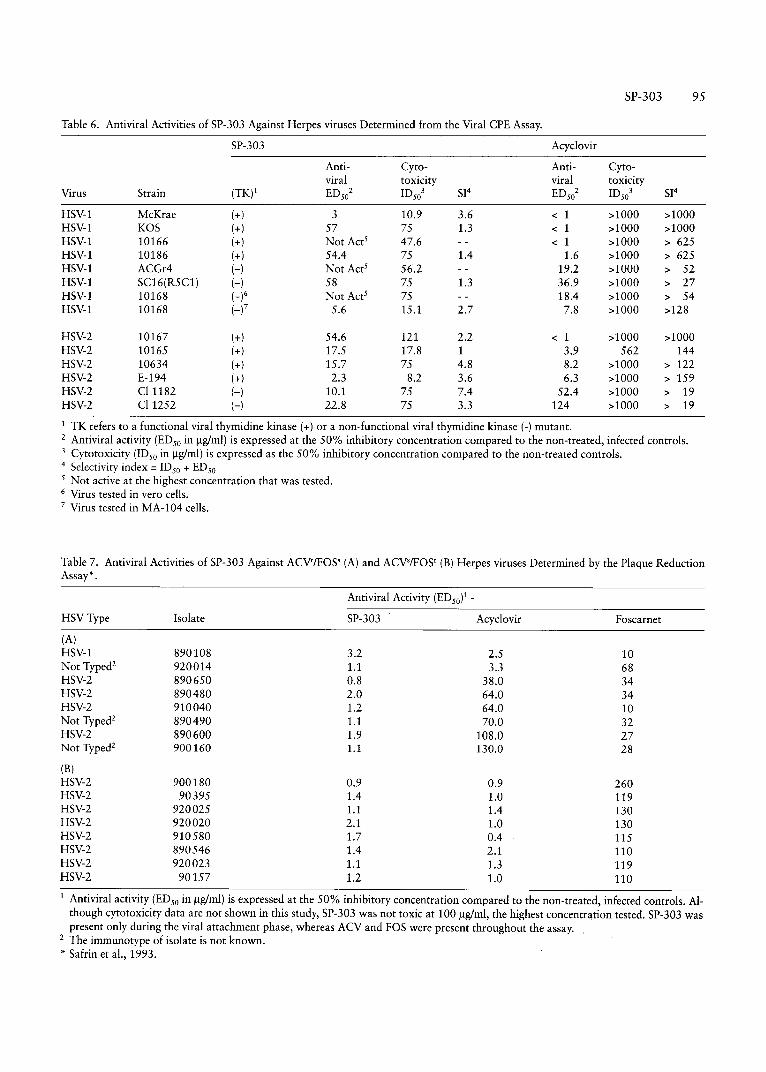
SP-303 95
Table 6. Antiviral Activities of SP-303 Against Herpes viruses Determined from the Viral CPE Assay.
SP-303 Acyclovir
Anti- Cyro- Anti- Cyto-viral toxicity viral toxicity
Virus Strain (TK)l EDs02 ID
s03 SI4 EDs02 IDs03 SI4
HSY-l McKrae (+ ) 3 10.9 3.6 < 1 >1000 >1000HSY-l KOS (+) 57 75 1.3 < 1 >1000 >1000HSY-l 10166 (+) Not Acr! 47.6 < 1 >1000 > 625HSY-l 10186 (+) 54.4 75 1.4 1.6 >1000 > 625HSY-l ACGr4 H Not Acr' 56.2 19.2 >1000 > 52HSY-1 SC16(R5C1) H 58 75 1.3 36.9 >1000 > 27HSY-1 10168 (_)6 Not Act? 75 18.4 >1000 > 54HSY-1 10168 (-f 5.6 15.1 2.7 7.8 >1000 >128
HSY-2 10167 (+) 54.6 121 2.2 < 1 >1000 >1000HSY-2 10165 (+) 17.5 17.8 1 3.9 562 144HSY-2 10634 (+) 15.7 75 4.8 8.2 >1000 > 122HSY-2 E-194 (+) 2.3 8.2 3.6 6.3 >1000 > 159HSY-2 C[1182 H 10.1 75 7.4 52.4 >1000 > 19HSY-2 C[ 1252 H 22.8 75 3.3 124 >1000 > 19
I TK refers to a functional viral thymidine kinase (+) or a non-functional viral thymidine kinase (-) mutant.2 Antiviral activity (EDso in ug/rnl) is expressed at the 50% inhibitory concentration compared to the non-treated, infected controls.3 Cytotoxicity (IDso in ug/rnl) is expressed as the 50% inhibitory concentration compared to the non-treated controls.4 Selectivity index =ID so + EDso5 Not active at the highest concentration that was tested.6 Virus tested in vera cells.7 Virus tested in MA-I04 cells.
Table 7. Antiviral Activities of SP-303 Against ACY'/FOS' (A) and ACY'/FOS' (B) Herpes viruses Determined by the Plaque ReductionAssay' .
Antiviral Activity (EDso)1 -
HSYType Isolate SP-303 Acyclovir Foscarnet
(A)HSY-1 8901 08 3.2 2.5 10Not Typed? 920014 1.1 3.3 68HSY-2 890650 0.8 38.0 34HSY-2 890480 2.0 64.0 34HSY-2 910040 1.2 64.0 10Not Typed? 890490 1.1 70.0 32HSY-2 890600 1.9 108.0 27Not Typed- 900160 1.1 130.0 28
(B)HSY-2 900180 0.9 0.9 260HSY-2 90395 1.4 1.0 119HSY-2 920025 1.1 1.4 130HSY-2 920020 2.1 1.0 130HSY-2 910580 1.7 0.4 115HSY-2 890546 1.4 2.1 110HSY-2 920023 1.1 1.3 119HSY-2 90157 1.2 1.0 110
I Antiviral activity (EDso in ug/ml) is expressed at the 50% inhibitory concentration compared to the non-treated, infected controls. Although cytotoxici ty data are not shown in this study, SP-303 was not toxic at 100 ug/ml, the highest concentration tested. SP-303 waspresent only during the viral attachment phase, whereas ACY and FOS were present throughout the assay.
2 The immunotype of isolate is not known.• Safrin et al., 1993.

96 R. Ubillas et al.
ception of a single assay, IDso endpoints with acyclovirwere not obtained at 1000 ug/rnl (Table 6).
When 5P-303 was tested against ACV'/F05' andACV'/F05r (Table 7) H5V type 1 and 2 viruses using theplaque reduction assay, considerabl y greater potencie s wereobtained than those obtained from the CPE assay (Table6). As seen in Table 7, 5P-303 is highly active against theACV' and ACVs viruses listed in this Table are 1.6 and 1.5ug/ml, respectively. These values compare with 60 and 1.1ug/rnl EDso's for ACV' and ACVs viruses, respectively, andwith EDso's of 15.8 and 36.6 ug/rnl for F05S and F05' viruses, respectively. Thus, as shown with the plaque reduction assay, 5P-303 exhibits highly potent activity against 8ACV' and 8 F05' strains of herp es viruses. Although theseACV' viruses have not been characterized for TK- or TK+genotype, the results of this study stro ngly imply the abilityof 5P-303 to inhibit TK-,Acvr H5V. The major implicationof this study is that 5P-303 exerts its antiviral effect by amechanism different than that of acyclovir and foscarnet,which are both known to inhibit replication of H5V DNA(Hovi, 1988; Mertz, 1990 ).
When 5P-303 was tested against hepatit is A virus (HAV)using MIT to define anti viral and cytotoxic endpoints, anEDsoof 5.0±0.0 ug/ml (n = 2) was obta ined with a ICsoof135 ug/rnl (51 = 27) . Against hepat itis B virus (HBV), SP303 at 50 ug/ml resulted in approximately a 50 % reductionof intra cellular HBV genom ic and replicative intermediateDNAs. No toxicitiy was detected at 50 ug/ml using neutralred stain ing.
Antiviral mechanism of SP-303
Virucidal test: Assessment of R5V infectivity when the virus was incubated for 1 hr with increasing concentrationsof 5P-303 indicated that 5P-303 is virucidal at concentrations of ~ 125 ug/ml: at this concentration, a decrease inviral titer of 1.8 10glO TCID so is observed. Since the virucidal concentrat ion is well above the inhibitory concentrations observed under conditions employed for anti viralassays, the virucidal effect of 5P-303 does not playa role inthe latter assay.
The following observat ions have been made regard ingthe time during the viral infectious cycle that SP-303 exert sits antiviral effect: (i) Time-of-addition stud ies performed inrelation to the viral cycle indicate that the optimal time to
produce an antiviral effect with SP-303 is early in the viralreplicat ive cycle: For example , the most potent anti-R5V effects are observed when 5P-303 (6 ug/rnl) is added to cellsconcomitantly with R5V and as late as 2 hours post-infection : The R5V titer is decreased between 1-2 loglo TCIDsowhen SP-303 is added between -2 hr to +2 hr post-infection. In contrast, if 5P-303 is added to cells at 4 hours postinfection, virtually no anti-R5V effect is observed. Similarfindings were observed when viral DNA synthesis was the
endp oint in H5V-1-infected cells: When SP-303 (75 ug/ml ]was added to cells before infection und allowed to remainwith the cells, viral DNA synthesis was completel y inhibited when measured at 18 hr. If added immediately after a1 hr viral adsorption period and allowed to remain on thecells, 5P-303 blocked HSV DNA synthesis by 30%, but ifadded later than 1 hr, there was no effect on viral DNA synthesis measured at 18 hr:o(i i) 5P-303 retain s its anti viral effect when cells are pretreated with the molecule for onehour and washed before infection. Cells pretreated with5P-303 for 1 and then subjected to as many as 10 washe sprior to infection inhibit R5V plaque formation by 60 %.Strong adherence to the plasm a membrane of cells may account for 5P-303 retaining some of its antiviral activity under these experimental conditions. These findings contrastwith those obtained for ribavirin : If ribavirin is added to
cells during the viral adsorption period and the monolayeris washed to remove comp ound , no antiviral effect is seen.Based on the fact that ribavirin is a nucleoside analoguewhich exert s its inhibitory effect during replication of theviral nucleic acid, this result is not unexpe cted and supportsa different antiviral mechanism of 5P-303 compared to thatof ribavirin. The result s of the above studies indicate that5P-303 exerts its anti viral effect by inhibiting an early viralmechanism such as adsorption and/or penetration throughthe plasma membrane.
In a mechanistic study using [3S5]-labeled R5V that wascoincubated with 5P-303 at 10.5 ug/ml, a 50% inhibiti onof viral attachment to cells at 60 min compared to untreated controls was demonstrated. A second mechani stic study, indicat ing an alternative antiviral mechanism, showedthat 5P-303 (10.5 ug/ml) inhibits nearly 100% of R5V penetr ation through the plasma membrane when the compound is added during the adsorption phase. Thus, R5V ismore sensitive to the effect of 5P-303 in inhibiting penetration than attachment. Confirmation of binding of 5P-303to virus particles was determined using purified R5V and[3H]-labeled 5P-303 , and Scatchard ana lysis gave a Kd
value for 5P-303 binding to R5V of 23±10 x 10-8 M (datanot shown). Although these studies clearly show that 5P303 binds to R5V, and that this interaction likely accountsfor inhibition of viral attachment and penetration, thesestudies do not preclude the possibility that 5P-303 adherence to plasma membranes also plays a role in blocking viral infectivity.
Finally, 5P-303 fails to induce production of interferonwhen incubated with L-92 9 cells (data not shown).
Efficacy of SP-303 in virus-infected animals
Table 2 provides details on the concentrations of 5P-303that were tested, routes of admin istration, experimental parameters for measuring efficacy, and other relevant information on the in vivo evaluati ons of 5P-303.

RSV-infected cotton rats: In the cotton rat model, SP-303was administered at several different dosage levels by threeroutes of administration: oral, intraperitoneal, and smallparticle aerosol (Wyde et al., 1993a; Gilbert et al., 1993).
(i) Oral evaluations: In a series of 12 efficacy studies inRSV-infected cotton rats (some are published in Wyde et al.1993b), SP-303 was administered in three dosages per study ranging from 0.3 to 270 mg/kg and given either once,twice, or three times daily. The efficacy of SP-303 wasmeasured from RSV recovered from lungs by transpleurallavage. In some of these studies, a ribavirin-treated groupwas included to compare its activity with that of SP-303.Comparing groups of animals receiving different dosagelevels to placebo-treated animals within individual studies,statistically significant reductions in viral titers are seenwith 1 mg/kg doses in 3 studies; with 3.0 mg/kg in 4 studies; and with 10 mg/kg in 3 other studies (data not shown).With the exception of a single study, no dose over 10 mg/kgresulted in statistically significant reductions in viral titersunder the conditions employed. The data from each studywere analyzed by two statistical methods: (i) ANOVA wasused across all dosage groups (in a single study) to determine if the overall reduction at all dosages was significantcompared to the control; and (ii) by linear regression to detect a dose-response related reduction- in RSV lung titers.Using the criteria of both requiring a statistically significanteffect produced by SP-303 for at least one dosage groupand evidence for a positive dosage response, 6 studies provide reasonable evidence that SP-303 is active in RSV-infected cotton rats under the experimental conditions, whileonly 4 studies were clear failures. In two other studies,there was a statistically significant decrease in RSV titers inone dosage group, but evidence for a dose-response waspoor. In summary, 67% (8/12) of the studies exhibitedevidence for efficacy at any dosage level. When correlatingefficacy with the number of dosage groups per study thatresulted in significant reductions in viral titer,S studies hadonly a single dosage which was statistically significant,while 2 studies had 2 dosages that were significant and 1study was significant at all 3 dosages. Analysis of dataresulting from these studies did not enable conclusions tobe drawn about a dose-response effect or the advantages(or disadvantages) of once versus multiple daily dosing.The findings of this series of studies are consistent with thesmall sample sizes (n = 4/group).
(ii) IP dosage route: When SP-303 was evaluated in theRSV/cotton rat model using the IP route of administration,the overall results of nine individual experiments confirmedthe ability of this compound to reduce RSV pulmonary titers. (Some of these studies are published in Wyde et al.,1993a). When the data were pooled, a dose-response relationship to 10 mg/kg of SP-303 was evident. At dosages of30 mg/kg, however, slight toxicity of the compound wasobserved, with weight losses occurring in infected and control animals. In 3 experiments, SP-303 was compared to ri-
SP-303 97
bavirin given at 90 mg/kg, with the latter drug showing asomewhat superior ability to reduce viral titers.
(iii) Inhalation route ofdosage: Administration of SP-303by small particle aerosol to RSV-infected cotton rats resulted in significant reductions in pulmonary titers of RSVwhen given at doses 1.3-10 mg/kg/day (Gilbert et al.,1993). However, at doses of 18.7 mg/kg/day, the compoundwas somewhat toxic, resulting in weight loss in comparisonto control animals. The reductions in RSV titers obtainedwith all groups treated with SP-303 by the inhalation routewere less than those seen in the ribavirin-treated positivecontrols.
RSV-infected African green monkeys: Seven studies wereperformed in which SP-303 was tested for efficacy in RSVinfected African green monkeys, the results of which aresummarized in Table 8. Three of these studies involved intravenous (IV) dosage of SP-303 while the remaining fourstudies employed oral gavage (PO) as a means of delivery.Some of these data were presented at the InternationalAntiviral Meeting in 1992 and subsequently published asan abstract (Soike et al., 1992). In all studies, monkeyswere infected with RSV intratracheally and intranasally.The ability of SP-303 to produce efficacy in RSV-infectedmonkeys was assessed using the following parameters:(i) mean area under the curve (AUC) of RSV titer versustime after inoculation; (ii) mean RSV titer at all time points;(iii) mean peak RSV titer at each dosage level (or for eachexperimental group); (iv) mean time to first RSV shedding;and (v) mean duration of RSV shedding. Except for symptomatic effects which were not quantitatively evaluated,AUC is the most important assessment as it measures theeffect of SP-303 on reduction of RSV titers throughout theexperiment.
(i) IV dosage route: With doses of 0.2, 0.5, or 1.0mg/kg/day given in divided doses starting 4 h prior to infection and continued for 7 days, statistically significant,dose-dependent reductions in throat swab titers of RSVwere obtained in SP-303 treated-groups compared to placebo-treated animals (Table 8). Indeed, animals receiving thehigh dose of 1.0 mg/kg/day had a reduction 3 loglo TCIDsoin peak virus titer, which represents a substantial reductionin oropharyngeal shedding of RSV. Also, these animalsseemed to experience fewer symptoms of infection. At the1 mg/kg dosage, the mean time to initial RSV shedding wassignificantly delayed and the duration of RSV shedding wassignificantly reduced compared to control animals (datanot shown). As a follow up to this study, the same high doseof SP-303 (1 mg/kg/day) was compared in groups treatedeither 4 hr (-4 hr) prior to infection or at 24 (+24) hr and 48(+48) hr post-infection (Table 9). In the -4 hr group, statistically significant reductions in comparison to control animals were again seen in throat swab titers of virus, reduction in mean duration of oropharyngeal shedding of virus(not shown), and a delay in mean time to onset of viralshedding (not shown). For the +24 hr group, there was a
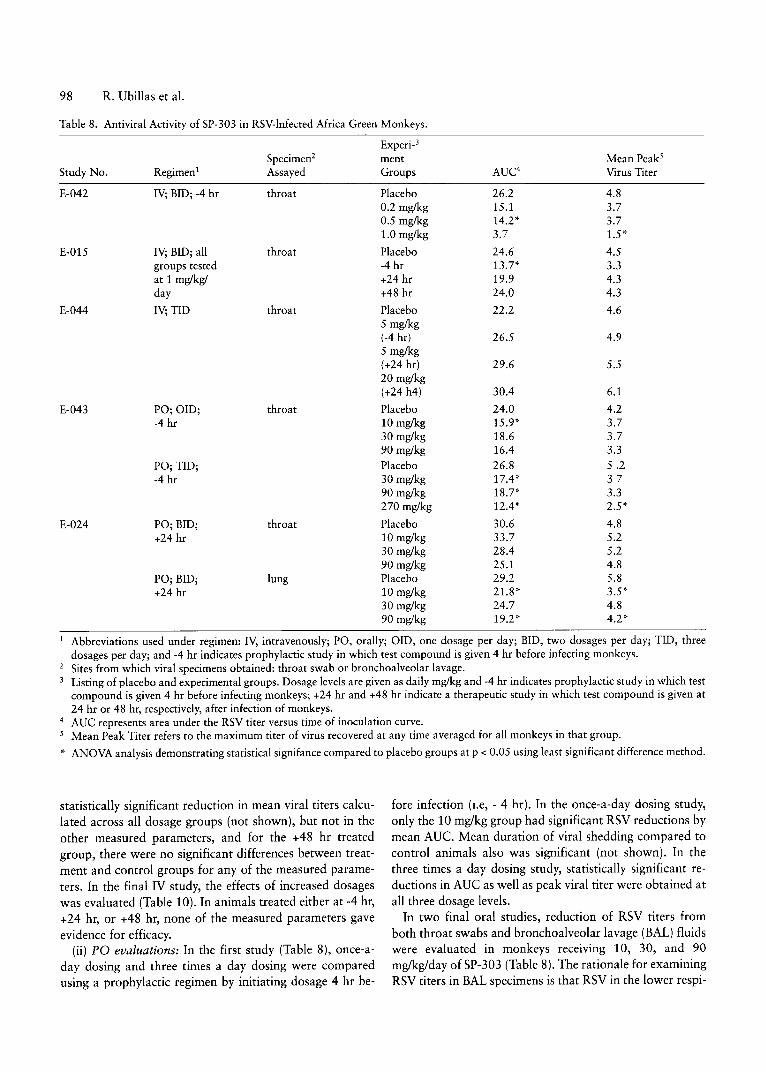
98 R. Ubillas et a l.
Table 8. Antiviral Activity of SP-303 in RSV-lnfected Africa Green Monkeys.
Experi-:'Specimen- ment Mean Peaks
Study No. Regimen! Assayed Groups AUe4 Virus Titer
£-042 IV; BID; -4 hr throat Placebo 26.2 4.80.2 mg/kg 15.1 3.70.5 mg/kg 14.2* 3.71.0 mg/kg 3.7 1.5*
£-015 IV; BID; all throat Placebo 24.6 4.5groups tested -4 hr 13.7* 3.3at 1 mglkg/ +24 hr 19.9 4.3day +48 hr 24.0 4.3
£-044 IV;TID throat Placebo 22.2 4.65 mg/kg(-4 hr) 26.5 4.95 mg/kg(+24 hr) 29.6 5.520 mg/kg(+24 h4) 30.4 6.1
£-043 PO; OlD; throat Placebo 24.0 4.2-4 hr 10 mglkg 15.9* 3.7
30 mg/kg 18.6 3.790 mg/kg 16.4 3.3
PO; TID; Placebo 26.8 5 .2-4 hr 30 rng/kg 17.4* 3 7
90 mg/kg 18.7* 3.3270 mg/kg 12.4* 2.5*
£-024 PO; BID; throat Placebo 30.6 4.8+24 hr 10 mg/kg 33.7 5.2
30 mg/kg 28.4 5.290 mg/kg 25.1 4.8
PO; BID; lung Placebo 29.2 5.8+24 hr 10 mg/kg 21.8* 3.5*
30 mg/kg 24.7 4.890 mg/kg 19.2* 4.2*
! Abbreviations used under regimen: IV, intravenously; PO, orally; OlD, one dosage per day; BID, two dosages per day; TID, threedosages per day; and -4 hr indicates prophylactic study in which test compound is given 4 hr before infecting monkeys.
2 Sites from which viral specimens obtained: throat swab or bronchoa lveolar lavage.J Listing of placebo and experimenta l groups. Dosage levels are given as daily mg/kg and -4 hr indicates prophylactic study in which test
compound is given 4 hr before infecting monkeys; +24 hr and +48 hr indicate a therapeutic study in which test compound is given at24 hr or 48 hr, respectively, after infection of monkeys.
4 AUe represents area under the RSV titer versus time of inoculation curve.S Mean Peak Titer refers to the maximum titer of virus recovered at any time averaged for all monkeys in that group .
* ANOVA analysis demonstrating statistical signifancecompared to placebo groups at p < 0.05 using least significant difference method .
statistically significant reduction in mean viral titers calcula ted across all dosage groups (not shown), but not in theother measured parameters, and for the +48 hr treatedgroup, there were no significant differences between treatment and control g roups for any of the measured parameters. In the final IV study, the effects of increased dosageswas evaluated (Table 10). In animals treated either at -4 hr,+24 hr, or +48 hr, none of the measured parameters gave
evidence for efficacy.(ii) PO evaluations: In the first study (Table 8), once-a
day dosing and three t imes a day dosing were comparedusing a prophylactic regimen by initiating dosage 4 hr be-
fore infection (i.e, - 4 hr). In the once-a-day dosing study,on ly the 10 mg/kg group had significant RSV reductions bymean AUe. Mean duration of viral shedding compared to
control animals also was significant (not shown) . In thethree times a day dosing study, statistically significant re ductions in AUe as well as peak viral titer were obtained atall three dosage levels.
In two final oral studies, reduction of RSV titers fromboth throa t swabs and bronchoalveolar lavage (BAL) fluidswere evaluated in monkeys receiving 10, 30, and 90mg/kg/day of SP-303 (Table 8). The rationale for examiningRSV titers in BAL specimens is tha t RSV in the lower respi-

ratory tract of humans is of greater significance pathologically than virus confined to the nasopharyngeal cavity. Although some efficacy was observed in throat swab specimens at the 30 and 90 mg/kg dosages in regard to meantime to first RSV shedding, respectively (not shown), no effect was observed with AUC and mean peak RSV titers atthe 10-90 mg/kg range. In contrast, for BAL specimens, the10 and 90 mg/kg groups were statistically significant compared to placebo-treated animals in respect to two test parameters: AUC and mean peak RSV titer. In fact, mean RSVtiters in BAL fluids were reduced by approximately 2 10glOTCIDso units. These results are particularly significant because administration of SP-303 was begun 24 hours afterinfection. Thus, the compound demonstrated efficacy in anRSV infection in the lung which represents the most significant clinical scenario of RSV infection in humans.
For either IV or PO studies with African green monkeys,no generalized toxic effect of SP-303 was observed. Occasionally, in the IV studies a small amount of irritation wasobserved at the intravenous site of injection. In the few instances when symptomatology was scored, observers feltSP-303-dosed animals experienced a decrease in rhinorheaand sneezing and coughing.
FLU-A-infected mice and PlV-3-infected cotton rats (Table 2). As resported by Wyde et al. (1993a) and Gilbert etal. (1993), SP-303 was efficacious in PIV-3- and FLU-A- infected animals, respectively (Table 4). Among IP, PO, andinhalation routes of dosage used with SP-303 in FLU-A-infected mice, only the inhalation route resulted in statistically significant efficacy. Evidence for efficacy in FLU-infectedanimals treated with SP-303 by small particle aerosol (SPA)was demonstrated by oximetry (i.e., % saturation of arterial 02), pulmonary FLU titers, survivors, pneumonitis, andhistology of lung tissue. In two studies published using PIV3-infected cotton rats (Wyde et aI., 1993a), SP-303 administered IP at 3 and 10 ug/rnl, reduced virus in the lung by
SP-303 99
90% compared to control animals. Efficacy with other experimental parameters was not reported for this model.
HSV-I- or HSV-2- infected mice and guinea pigs (Table 2).Parameters were used to assess efficacy in the HSV/mouseand guinea pig models: AUC from viral lesion score versustime curve, virus shed from lesions, and the number of surviving animals at the end of the experiment. In the murine/and guinea pig/ vaginal models efficacy due to SP-303 wasdemonstrable by IP, PO, and TOP routes of administration.In mice, the AUC determined from vaginal lesion scoreswas statistically significant for the SP-303-treated groupscompared to placebo groups by all three routes (IP: 6.7 at30 mg/kg/day vs. 14.0 for the placebo; PO: 6.2 at 270mg/kg/day vs. 10.5 for the placebo; TOP: 5.4 with 10%cream vs. 9.3 for the placebo), but a dose-response effectwas not apparent. Fig. 7 compares the effect of topical SP303 and acyclovir treatment on murine genital lesions. Asseen, acyclovir is approximately twice as active as SP-303based on AUC: 6.7 for SP-303 and 1.4 for acyclovir.Neither titers of HSV-2 shed nor percentage of survivors forthe SP-303-treated animals were significantly different thanplacebo groups. Again, acyclovir or DHPG (ganciclovir),used as positive controls, were efficacious by most measures.
Topical application of SP-303 to viral lesions in the HSV2/guinea pig vaginal model resulted in statistically significant efficacy in regard to virus shed in 2 of 3 studies. (In thestudy in which SP-303 failed, acyclovir also was a completefailure.) In one of these studies, the decrease in virus shedcorrelated with a concomitant reduction in AUC as well asan increase in the percentage of survivors compared to thecontrol with one of 3 topical vehicles that was tested (Table2). The results of this study suggest that the formulation ofthe topical vehicle may be significant since vehicle 1, unlikethe other two vehicles, contained 78% dimethyl sulfoxide(w:w) (Fig. 8).
--0-- Vehicle 14
~0o
(J) 3
c.2U)III 2
...J
CcaIII:E
0
2 4 6 8 10 12 14
........<>........ Vehicle 2
····0···· Vehicle 3
----6---- ACV (5%)
- - -EB- - - Placebo
Regimen: +6 hr, 15% SP-303in different ointments, BID, 8days.
'Statistically significant cornpared to placebo @ p < 0.05.
1 6 PFU shed & % survivors alsosignificant'.
Days Post-I nfection
Fig. 7. Effect of Topical Treatment of SP-303 on HSV-2 Vaginal Infection of Mice.

1DD R. Ubillas et al.
However, SP-3D3 was not efficacious in dorsally infectedhairless or shaved guinea pigs when applied topically as a3D% cream or ointment. In contrast, ACV was nearly always efficacious in this model by a least one of the aboveparameters. In summary, SP-303 is capable of producing efficacy in the HSV-2/mouse and guinea pig vaginal models,but a dose-dependent effect correlating to any antiviral parameter was not observed.
Discussion of Virology Test Results
The antiviral activity of SP-3D3 against a variety of DNAand RNA viruses in cell culture and in animal models of infection is presented below. In cell culture, SP-303 is inhibitory against a broad spectrum of respiratory viruses thatcomprise three genera in two viral families: the Paramyxoviridae (PIV-I & RSV) and the Orthomyxoviridae (FLU-A& FLU B). Moreover, SP-3D3 inhibits viruses representingthree non-respiratory viral families: Herpesviridae (HSV-Iand HSV-2), Picornaviridae (hepatitis A virus), and Hepadnaviridae (hepatitis B virus). The antiviral potency ofSP-3D3 in the antiviral CPE assay is similar to that of ribavirin against RSV, whereas against HSV-I and HSV-2 in theCPE assay, SP-3D3 is not as broadly active against acyclovir" herpesviruses as acyclovir". Alternatively, as measured by plaque reduction assay, SP-303 has the advantageover acyclovir- against acyclovir- herpesviruses, as SP-3D3is highly inhibitory to these viruses. Likewise, SP3D3 exhibits significant inhibitory effects against FOS' herpes viruses. In animal models of viral infection, SP-3D3 exhibitsactivity in RSV-infected cotton rats and African green monkeys, PIV-3-infected cotton rats, HSV-2-infected mice andguinea pigs, and FLU-A-infected mice.
With the exception of nucleoside analogues, which mayhave broad antiviral activities, and interferon, which inhibits a variety of viruses (Hovi, 1988; Hirsch & Kaplan,199D), the broad "interfamily" antiviral activtiy of SP3D3,which is not a nucleoside and does not appear to induce interferon, is unusual, if not unique. The results of studiespresented here indicate that SP-3D3 interacts with the viralparticle outside the cell to exert its antiviral effect. In fact, ifSP-3D3 is added to infected cells after RSV or HSV has penetrated the plasma membrane, no viral inhibition is detected. Furthermore, a study using radiolabeled SP-3D3 demonstrated that little or no SP-303 crosses the plasmamembrane (unpublished results). Thus, the antiviral effectof SP-3D3 is manifest through extracellular interaction withvirus. In the case of viruses having lipid envelopes (i.e, RSV,FLU, PIV, & HSV), SP-3D3 must interact with either the lipid moieties comprising the envelope or the virion glycoproteins that protrude from the envelope or both. It is wellknown that tannins strongly bind to basic polymers andproteins (Goldstein and Swain 1965), and this probablyplays a significant role in the antiviral effect of SP-3D3. Re-
gardless of the viral structural component to which SP-3D3binds, binding studies with RSV and [3H]-labeled SP-3D3suggest that SP-3D3 binds in a specific manner to virus: AKd of 2.3 x 10-8 M was determined from the Scatchard analysis, which is similar to affinities of known antiviral ligands that bind to specific receptors on the virion particle.For example, the WIN molecules interact with the nonenveloped capsids of human rhinoviruses with Kd values inthe 10-8 M range (Fox et al., 1991).
In regard to distinguishing lipid constituents from viralglycoproteins as targets for SP-3D3 binding, it can be argued that, since the viral envelope is derived from the plasma membrane, SP-3D3 should be equally active against different viruses replicating in the same cell. Clearly this is notthe case (Tables 3, 6, 7). In light of this argument, it is interesting that SP-3D3 causes a reduction in the intracellularDNAs of hepatitis B virus (HBV) since the antiviral assayfor HBV employs chronically infected hepatocytes. SinceHBV acquires its envelope intracellularly, rather than bybudding through the plasma membrane (Robinson, 199D),it would seem that SP-303 inhibits HBV indirectly by interacting with the plasma membrane of infected hepatocytes.The one nonenveloped virus against which SP-3D3 is inhibitory is hepatitis A virus (HAV). Although antiviral mechanism of action studies were not performed with HAVaswere done with RSV, and to a lesser extent with HSV, thisresult suggests that SP-3D3 interacts with some polypeptidecomponent on the surface of the HAV capsid to result in inhibition. In any event, without further study, it is not possible to define the precise nature of SP-303 interaction withthe viral envelope or-capsid.
The finding that SP-3D3 inhibits RSV-infection of HEp-2cells when the cells are pretreated with SP-3D3 and thenwashed extensively before infection suggests an inhibitorymechanism of SP-3D3 different than that of binding directly to the viral particle. Under these conditions, it is possiblethat SP-3D3 adheres to the plasma membrane to render thecellular receptors for viral attachment inaccessible to virus,and thereby blocks virus attachment. Although the resultsof the binding experiment with purified RSV and SP-3D3clearly demonstrate that SP-3D3 interacts with the viralparticle, none of the cell-based assays that resulted in inhibition of virus discriminate between SP-3D3 binding to thecell or to the virus. In fact, it cannot be discounted thatboth mechanims have a role. It should be emphasized thatthe effect of SP-303 to inhibit RSV binding to the plasmamembrane or penetration through the plasma membrane ifattachment has occurred is consistent with a large body ofdata obtained on picornaviruses treated with the capsidbinding WIN molecules (Fox, 1986; Smith, 1986; Pevear,1989). Finally, precedence exists for plant-derived antiviralagents that interact with viral particles to inhibit early viralevents such as attachment, penetration, and/or uncoating.The best-studied of these agents are flavanoid molecules

SP-303 101
3.-------------------,
'D--iJf-i:J----o--o-{}-Q--G *
----0---· Placebo
........<>.. ...... ACV (5%)
'Statistically significant cornpared to placebo @ p < 0.05.
SP-303 (10%)
Regimen : +6 Hr, SP·30310% ointment BID, B days.(Ointment: Squibb cr. base)
--0--
*.......(). .
1:) •• •0-' -0- --0"0"0··-0
,.0'"
~
...0
2
2.5
0.5
1.5
Q)...oo
(J)
l:.S!IJlQ)...J
l:l'GQ)
:E
4 6 8 10 12 14 16
Days Post-Infection
Fig. 8. Effect of Topical Treatment of Sp·303 on HSV-2 Vaginal Infection of Guinea Pigs.
that interact with the capsids of picornaviruses (Ishitsuka etaI., 1982; Superti et al., 1989; Conti et aI., 1992).
Except for topical application of SP-303 to HSV-Ilesionson the backs of guinea pigs, SP-3D3 exhibited efficacy in allother animal models of viral infection that were tested (Tables 9-11). The routes of SP-3D3 administration that weremost successful in producing efficacy were: topical application to HSV-2 genital lesions in mice and guinea pigs, aerosol inhalation to FLUA-infected mice and PIV-3-infectedcotton rats , and oral dosage to RSV-cotton rats . It shouldbe noted that SP-303 was not always active by all the experimental parameters used to measure efficacy in a singlestudy, and did not always demonstrate clear dose-dependent effects. Despite this, SP-3D3 was clearly efficacious inmost of the virus/animal models used for this report.
Given that SP-3D3 possesses antiviral activity in cell culture that, with few exceptions, approximates that of positive control antiviral compounds, then it is worth considering why the level of efficacy achieved with SP-303 in vivowas not always equivalent to that of the control compounds, or in one model why SP-303 was completely inactive. The best system to examine this question is that of theHSV/animal models that tested acyclovir or DHPG (ganciclovir) in parallel with SP-3D3. (Although ribavirin wasoccasionally orally dosed in parallel with SP-3D3 in cottonrats, the optimal way to dose ribavirin to achieve efficacy isby inhalation.) When topically applied to the backs ofHSV-I infected guinea pigs, no evidence was obtained withSP-3D3 to support efficacy, whereas, ACV almost alwaysafforded efficacy in this model. Moreover, when SP-3D3was applied topically to HSV-2 vaginal lesions in mice, thecompound was uniformly less active than DHPG. Thereappear to be two possible major explanations that explainthese observed differences: (i) SP-3D3 is not reaching thesite of viral replication, and/or (ii) SP-303 is reaching thesite of replication but its extracellular antiviral mechanismis disadvantageous. In regard to (i), evidence obtained with
three different topical formulations of SP-303 implies thatefficacy may be enhanced if the compound can be deliveredto the site of viral replication in sufficiently inhibitoryquantities, (Fig. 3). On the other hand, in regard to (ii),even if more SP-303 is delivered to the site of HSV replication, the extracellular antiviral mechanism of SP-303 maypreclude it from being as efficacious as acyclovir. If the majority of the virus in the lesion is highly cell-associated (i.e.,little extracellular virus), then acyclovir clearly has the advantage with its intracellular antiviral mechanism. In fact,acyclovir is highly efficacious in either the HSV-l or HSV-2model topical models.
When the SP-303 is given to HSV-2 vaginally infected miceby the IP and oral routes, the level of efficacy achieved,based on AUC (lesion score versus time curve), is 4.7- and2.8- fold less than that achieved with DHPG. The explanation for these differences in activity may relate to pharmacokinetic differences between SP-3D3 and DHPG. To affordefficacy in this system, an antiviral molecule must get fromthe peritoneal cavity or the intestinal tract into the circulation and eventually to the site of viral replication. This presumption precludes SP-303 acting as a general immune stimulant or as interferon. Both possibilites have been testedand excluded (unpublished data).
Toxicology
The toxicity of SP-3D3 has been investigated followingparenteral and oral administration in acute and multipledose studies in rats , mice, dogs and monkeys. The acute,single dose toxicity studies conducted for SP-3D3 are summarized in Table 10.
As noted in Table 9, no mortality was observed in any ofthe species following oral administration of SP-3D3 insingle doses up to 30D mg/kg. There were no meaningful

102 R. Ubillas et al.
Table 9. Acute, Single Dose Toxicity Studies of SP-303· .
Species
Mouse
Rat
Rat
Dog
Study LD.
MDR#402
MDR #401
MDR#406
ITR #126
Route Dose Range Number of Animals Results(mg/kg)
LV. 6.25-50.0 10/sex/dose LDso>50.0 mg/kg(x 4 doses SP-303)
LP. 10.0-100.0 10/sex/dose LDso>100.0 mg/kg(x 4 doses SP-303)
LV. 3.0-50.0 6/sex/dose LDso>50.0(x 4 doses SP-303) mglkg**
LP. 10.0-100.0 6/sex/dose LDso>lOO.O mg/kg(x 4 doses SP-303)
P.O. 30.0-300.0 6/sex/dose LDso>300.0 mg/kg(x 3 doses SP-303)
LV. 10.0-18.9 2 LDso>18.9 mg/kg
• In the mouse and rat studies, a single administration of SP-303 was followed by a 14-day observation period. In the dog study, dogswere observed for five days after drug administration.
** There were 5 of 12 (42%) animals that died in this study following intravenous administration. LDsois probably between SO and 100mglkg.
Table 10. Repeated-Dose Toxicit Studies: Oral Administration.
Species Study LD. Route Daily Dose Duratio Number of AnimalsRange n(mg/kg)
Mo use Shaman" LP. 2110/20 30 5 female/dose (x3 doses)P.O. (IP) days 5 females/dose (x3 doses)
10130/90(Po)
Rat Shaman" P.O. 0/100/300/ 5days 4/sex/dose (x 3 doses)a900
Rat MBR #04- P.O. 0/10/200/5 14 5/sex/dose (x 3 doses)07 00 days
Rat VIC #086 P.O. 0/50/200/5 30 12/sex/dose (x 3 doses)b00 days
Dog ITR #218 P.O. 100 5 days 2 males test1 male control
Monkey Pilot P.O. 100/300/90 5 days 2/dose (x3 doses)0
Monkey ITR #226 P.O. 0/10/50110 14 3/sex/dose (x 3 do5es)cITR #25 012001500 days
Monkey ITR #240 P.O. 0/301100/2 30 6/sex/dose (control, high00 days dose)
6/sex/dose (low, mid dose)
• Non-GLP studya 2/sex in controlsb Plus 5/sex in control and high dose groups for recoveryc 6/sex in controls
No ObservableEffect Level(NOEL)
N/A
N/A
500mg/kg/day
>200 and <500mg/kg/day
N/A
N/A
>100 and <200mg/kg/day
>30 and <100mglkg/day
changes seen in food consumption, body weights, clinical
signs or effects observed at necropsy.Following intravenous and intraperitoneal administra
tion in mice, the lowest doses producing letha lity were 25mg/kg and 10 mg/kg respectively. In rats, mortality was observed only in the high dose group (50 mg/kg) following intravenous administ ration, and no lethality was observed
following intraperitoneal administration at doses up to 100
mg/kg.In multiple dose studies, the toxicity of orally adminis
tered SP-303 has been examined in mice, rats, dogs andmonkeys for treatment periods varying . from 5 to 30 days(Table 10). Five-day pilot studies followed by 14 and30-day GLP trials have been co nducted in bo th rats and

SP-303 103
Table 11. Repeated-DoseToxicity Studies: Intravenous Administration.
Species
Rat
Dog
Dog
Design
Study MDR #4030,5,20,35 mg/kg i.v. x 5 Days5/sex/dose
ITR #1240,5,20,35 mg/kg i.v x 5 Days1/sex/dose at 0, 5, 20 mg/kg2/sex/dose at 35a mglkg
2 dogs received 35 mg/kg boluswhich was changed to 30 mglkg infusion (over 15 min) due to rapidloss of consciousness after bolus
ITR #1610,2.5,5, 10 mg/kg i.v.( 15 min infusion) x 5 Days3/sex/dose at 0, 2.5, 5 mg/kg5/sex/dose at 10 mg/kg
One dog of each sex in high groupplaced into a 2- or 4-week recoveryperiod.
Findings
1. Mortality: none2. Clin signs: changes at the inj sites and adjacent tissues3. Chern: Mid/High-increased WBC, Segs, platelets; deer RBCs, Hb, Hct (not considered
biologically significant)4. Path: incr weight in liver (all), kidney (low/mid) (not considered biologically signifi
cant)5. Histo: limited to the inj sites - hemorrhage, necrosis, inflammation, or thrombosis (not
considered biologically significant)6. NOEL: 5.0 mg/kg/day
1. Mortality: none2. Clin signs: following 35 mglkg bolus included ataxia, loss of consciousness, increased
salivation, bradycardia coughing and retching (recovered 30 min post dose); Sx notobserved in same animals following 30 mglkg infusion; deer weight & food consumption (high)
3. Chern: MidlHigh-increased alk phos, ALT, LDH and sorbitol dehydrogenase; thrombocytopenia (max deer 81% - 89% to 24,000-51,000/1l1); prolonged aPTT & PTwithlow fibrinogen;
4. Path: (in MidlHigh) dark discoloration of mucosa in proximal sm intestine, enlargedmesenteric lymph nodes, pigment in the reticuloendothelial system, sinusoidal histiocytosis, pigmented mononuclear cells in centrilobular veins, dilated lymphatics, cytoplasmic vacuoles in splenic RE cells; hemorrhages in serosa of the gall bladder, endocardium, colon, urethra, urinary bladder and skeletal muscle; chronic lymphadenitis,enlarged vacuolated histiocytes in sinuses of mesenteric lymph nodes and thrombosisof the injection/infusion site
5. Histo: Rx-related changes observed in all animals; changes in low group were few andslight in severity
6. NOEL: 5.0 mg/kg/day
1. Mortality: none2. Clin signs: no significant findings3. Chern: Thrombocytopenia (high) - onset 15 minutes post- dose day 2 until study ter
mination, max decr of 52%-87% (nadir 33,000-180,000/1l1); values returned to pretreatment levels within 1 to 5 days of recovery
4. Histopath: dark discoloration of the proximal small intestine (all) with dose-relatedincrease in incidence; hepatic sinusoidal histiocytosis, cytoplasmic vacuoles in splenicsheathed capillaries, enlarged and vacuolated histiocytes in medulla of mesentericlymph nodes, focal thrombosis in lungs (high); pigmentation in hepatic sinusoidal cellsand in histiocytes of prox sm intest villi (mid/high); histiocytic foci in prox sm intestvilli, thrombosis at inj site (all); histologic changes in low-dose animals were slight ormild: in the recovery phase, changes reported in the reticuloendothelial system generally either showed no improvement or, in some cases, were more evident
5. NOEL: 2.5 mg/kg/day
monkeys. In the 14-day trials in rats, groups of 5 male and5 female Sprague-Dawley rats were administered SP-3Q3by oral gavage at doses of 0,10,200, or 500 mg/kg/day for14 days. No treatment-related clinical signs were observedand no animals died during this study. There was no effectof SP-303 on any of the hematological, clotting or clinicalchemistry parameters monitored in this study. There wereno significant findings at gross necropsy, and no histopathologic changes were observed in the high-dose group.
Following this study, groups of 12 male and 12 femaleSprague-Dawley rats were administered SP-303 by gavageat doses of 0, 50, 200, or 500 mg/kg/day for 30 days. Fiveadditional rats of both sexes served as recovery animals and
were assigned to the high dose and control groups. At theend of the 3D-day dosing period, the recovery animals wereobserved for an additional 30 days before sacrifice. Onehigh-dose male was found dead on day 21 but the cause ofdeath was not determined.
Several high dose animals demonstrated rough haircoatduring the latter weeks of treatment but this was not apparent during the 3D-day recovery period. Sporadic reductions in food intake possibly accompanied by marginaldecreases in body weight gains were seen at the high doselevel. This was not seen following cessation of treatment,except for an apparent spurious decrease in body weightgain for high dose females. Slight decreases in urinary spe-

104 R. Ubillas et al.
cific gravity were observed for high dose females and possibly high dose males at week 5. As this effect was slight andwas not seen at other times or accompanied by any otherindications of renal toxic ity, its biological significance is unkno wn. No other urinalysis measurements, or hematologyor clinical chemistry parameters were altered by SP-303treatment .
A 14-day dose ranging study was conducted with SP-303administered orally to Rhesus monk eys. The study was carried out in 2 separate segments; the results of both segmentshave been combined for this summary. In the first segment ,3 monkeys of each sex were assigned to the control group(0 mg/kg), low-dose group (10 mg/kg/day), mid-dose level(200 rug/kg/day), and highdo se group (500 mg/kg/day). Inthe second segment, 3 monkey s of each sex were assignedto the control (0 mg/kg), low-dose gro up (50 mg/kg/day )and high-dose group (100 mg/kg/day). Except for the doselevels administered, the an imals in both segments weretreated essentially ident ically. There were no changes in hematology, blood chemistry, clotti ng test, or IgG or IgE levels that were considered to be biologically meaningful.One female dosed with 500 mg/kg/day SP-303 had an increased eosinophil count on study day 15. Histopathological examination revealed eosinophilic infiltration in the hematopoetic organs. The toxicological significance of thisfind ing is unknown. Pathological lesions possibly associated with administration of SP303 were restr icted to animalsreceiving 200 or 500 mg/kg/day. They consisted of pigmented histiocytes in the small and large intestines, a superficial gastropathy, thymic lymphoid involution. In add ition ,diffuse pigmentation in hepatic sinusoidal cells was observed in the 500 mg/kg group. A marked decrease in meanthymic weights with thymic involution, was evident in animals of both sexes in the 500 mg/kg group.
In the next study, groups of 6 male and 6 female Rhesusmonkeys were administered SP-303 by stomach tube at doses of 0, 30,100 , or 200 mg/kg/day for 30 days. At the endof the dosing period, 4 males and 4 females from each groupwere sacrificed and submitted to complete necro psies, while2 males and two 2 from the contro l and high-dose groupswere maintained for an additiona l 30-days recovery period .One low-dose male died from stra ngulation with its neckchain on day 29. There were no other death s during the study period. Body weights, water consumption, clinicalpath ology, ophthalmology and electrocardiography evaluat ions did not show any evidence of treatment-related findings. The only clinical findings of note were decreased appetite in some animals in all groups and loose or liquidstoo ls, predominantly in the high-dose group. Histologically, an increased numb er of pigmented sinusoidal cells in theliver (high-dose group), greenish pigmented histiocytes in alllevels of the small intestin e, an increased pigmentation inthe histiocytes of the colon and cecum were noted (mid- andhigh-dose groups) were noted. In the recovery animals, allfindings had notably subsided or disappeared completely.
In addition to the oral studies, repeat dosing via the intravenou s route has been conducted in rats and dogs, and issummarized in Table 11. As noted in the table, intravenousadministration of SP-303 was much better tolerated in ratsthan in dogs. Intravenous administra tion of SP-303 for 5days was associated with significant hematologi c toxicity(primarily consisting of thrombocytop enia) in dogs. In addition, dogs exhibited a numb er of histopath ologic changesincluding pigmentation and vacuoliza tion of histiocytesthrough out the reticuloendothelial system. Administrationof a 35 mg/kg bolus injection to dogs precipitated an episode of ataxia, bradycardia and loss of consciousness fromwhich the animals recovered after about 30 minutes.
References
Beladi, I., Pusztan, R., Muesi, I., Bakay, M. and Gabor, M.: Activity of some flavonoids against viruses. Ann. N. Y. Academy ofScience 284; 358-364,1977.
Bennett, B.:Useful Plants of Amazonian Ecuador. Institute of Economic Botany. The New York Botanical Garden, New York,New York, 1990.
Bennett, ]. P.: Methods in binding studies, pp. 57-90. H.I. Yamamura, S.] . Erina , and M.]. Kuhar (ed.), Neurotransmitter receptor binding. Raven Press, New York, 1978.
Betrolo, R. M., and Scarpati, M.L.: Alkaloids of Croton draconoides. Phytochemistry 18: 520; 1979.
Cariel, L. and Jean, D.: Proanthocyanidol-based composition andits pharmacological application. WO 90/133041, 1990.
Cai, Y., Evans, E]., Roberts, M. E, Phillipson, J.D., Zenk , M.H.,and Gleba, Y. Y. : Polyphenolic compounds from Croton lechleri.Phytochemistry 30:2033-2040,1991.
Ceron C« Ethnobotanica de los Cofanes en Dureno Provincia delNapo. Universidad Central del Ecuador. Thesis Doctorate ofBiology, 1987.
Conti, c.,Tomao, P. Genovese, D., Desideri, N., Stein, M.L., andOrsi, N. : Mechanism of action of the antirhinovirus flavanoid4' , 6-dicyanoflavan. Antimicrob. Agents Chemother. 36: 95-99,1992.
Cragg, G., S. Schepartz, M. Suffness and M. Grever: The Taxolsupply crisis. New NCI policies for handling the large scale production of novel natural product anti-cancer and ant i-HIVagents.]. Nat. Prod. 56: 1657-1 668, 1993.
Dowdle, W.A., Kendal, A.P., and Noble, G. R.: In DiagnosticProcedures for Viral, Rickettsial, and Chlamydial Infections, 5thedition, (E.H. Lennette and N.]. Schmidt, eds), pp . 585-609,American Public Health Association, Washington, D.C. , 1979.
Feldman, L., Blankenship, M.L., and Ben-Porat, T.: Isolation andcharacterization of a temperatur e-sensitive uncoating mutant ofpseudorabies virus. ] . Gen. Virol. 54: 333-242, 1981.
Fox, M.P., McKinlay, M.A., Diana, G. P., and Dutko. EJ .: Bindingaffinities of structurally related human rhinovirus capsid-binding compounds are related to their activities against humanrhinovirus type 14. Antimicrob. Agents Chemo ther. 35:1040-1 047, 1991.
Fox, M.P., Otto, M.J., and KcKinlay, M.A.: Prevention of rhinovirus and poliovirus uncoating by WIN 51711, a new antiviraldrug. Antimicrob. Agents Chemother. 30 : 110-116, 1986.
Gilbert, B.E., Wyde, P.R., Wilson, P.Z., and Meyerson, L.: SP-303srnallparticle aerosol treatm ent of influenza A virus infection in

mice and respiratory syncytial virus infection in cotton rats. Antiviral Res. 21: 37-45, 1993.
Goldstein,]. L. and Swain, T.: The Inhibition of Enzymes by Tannins, Phytochemistry 4: 185-192, 1965.
Grunert, R. R., McGahen, ]. W., and Davis, W.L.: The in vivo antiviral activity of 1-adamantanamine (amantadine). 1. Prophylactic and therapeutic activity against influenza viruses. Virology 26: 262-269,1965.
Harborne,]. B.: The Flavonoids-Advances in Research since 1980,Chapman and Hall, New York, 1988.
Harborne, ].B. and Mabry, T.].: The Flavonoids-Advances in Research, Chapman and Hall, New York, Chapt. 2, 1982.
Harmenberg, ]., Akesson-]ohansson, A., Grasland, A., Malmfors,Bergman, ]., Wahren, B., Akerfeldt, S., Lundblad, L., and Cox,S.: The mechanism of action of anti-herpes virus compound2,3-dimethyl-6(2-dimethylaminoethyl)-6H-indolo-(2,3-b)quinoxaline Antiviral Res. 15: 193-204, 1991.
Haslam, E.: Plant Polyphenols-Vegetable Tannins Revisited. In:Chemistry and Pharmacology of Natural Products, CambridgeUniversity Press, New York, 1989.
Haslam, E., Self, R., Eagles, j., Galletti, G. e., Mueller-Harvey, I.,Hartley, R.D., Lea, A.G.H., Magnolato, D., Richli, U., andGujer, R.: Fast Atom Bombardment Mass Spectrometry of Polyphenols (syn. Vegetable Tannins), pp 449-468. John Wiley &Sons, Ltd, 1986.
Hayden, E G.: Animal models of influenza virus infection for evaluation of antiviral agents. In: Experimental Models in Antimicrobial Chemotherapy, vol. 3 (0. Zak and M.A. Sande, eds.),pp. 353-371, Academic Press, New York, 1986.
Hirsch, M. S., and Kaplan, ]. c.. Antiviral agents. In: Virology, second edition (B.N. Fields and D.M. Knipe, eds.), pp. 441-468.Raven Press, Ltd., New York, 1990.
Hovi, T.: Successful selective inhibitors of viruses. In: AntiviralAgents: The Development and Assessment of Antiviral Chemotherapy, vol. 1 (H.]. Field, ed.), pp. 1-21, CRC Press, Boca Raton, Florida, 1988.
Hudson,]., B.: Antiviral Compounds From Plants. CRC Press, Inc.2000 Corporate Blvd., N.W., Boca Raton, Florida, 1990.
Ishitsuka, H., Ninomiya, Y.T., Ohsawa, e., Fujiu, M., and Suhara,Y.: Direct and specific inactivation of rhinovirus by chalconeRo 09-0410. Antimicrob. Agents Chemother. 22: 617-621,1982.
Itsuo, N., Genichiro, N., Michihiro, E and Showa, U.: Tannincompounds. U.S. Patent #4,806,658, 1989.
Kakuk, T.]., Soike, K., Brideau, R.]., Zaya, R.M., Cole, S.L.,Zhang, ].- Y., Roberts, E.D., Wells, P.A., and Wathen, M. W.: Ahuman respiratory syncytial virus primate model of enhancedpulmonary pathology induced with a formalin-inactivated RSVvaccine but not a recombinant FG vaccine. ]. Infect. Dis. 167:553-561,1993.
Kaul, T. N., Middleton, Jr. E., and. Ogra, P. L.: Antiviral Effect ofFlavonoids on Human Viruses. ]. Med. Virology 15: 71-79,1985.
Kern, E. R.: Treatment of genital herpes simplex virus infections inguinea pigs. In: Herpesvirus (E Rapp, ed.), pp. 617-636, AlanR. Liss, Inc., New York, 1984.
Kern, E.R.: Animal models as assay systems for the developmentof antivirals. In: Antiviral Drug Development. A Multidisciplinary Approach (E. De Clercq and R.T. Walker, eds.), pp.149-172, Plenum Press, New York, 1988.
Kern, E.R.: Preclinical evaluation of antiviral agents: in vitro andanimal model testing. In: Antiviral Agents and Viral Diseases ofMan, Third Edition (G.]. Galasso, R.]. Whitley, and T.e. Merigan, eds.), pp. 87-123, Raven Press; New York, 1990.
SP-303 105
King, S.: Establishing reciprocity: Biodiversity, conservation andnew models for cooperation between forest-dwelling peoplesand the pharmaceutical industry. In: Sourcebook on IntellectualProperty, Rights and the Possibilities for Indigenous People (T.Greaves, ed.). Proc. AAAS Meeting San Francisco, December1992.
Kitazawa, E., Sato, A., Takahashi, S., Kuwano, H., and Ogiso, A.:Novel diterpenelactones with anti-peptic ulcer activity fromcroton sublyratus. Chem. Pharm. Bulletin 28: 227-234,1980.
Korba, B.E., and Milman, G.: A cell culture assay for compoundswhich inhibit hepatitis B virus replication. Antiviral Res. 15:217-228,1991.
Korba, B.E., and Gerin,]. L.: Use of a standardized cell culture assay to assess activities of nucleoside analogues against hepatitisB virus replication. Antiviral Res. 19: 55-70, 1992.
Lewis, W. H., Stonard, R. ]., Porras-Reyes, B. and Mustoe, T. A.:U.S. Patent #5,156,847,1992.
Markham, K.R.: Techniques of Flavonoid Identification. Academic Press, p. 24-29. Oval Road, London NW1, 1982.
McRae, W.D., Hudson, ].B., and Towers, G.H.N.: Studies on thePharmacologoical Activity of Amazonian Euphorbiaceae. ].Ethnopharm., 22: 143-172, 1988.
Mertz. G.].: Antiviral agents and viral diseases of man. 3rd Ed. Ed.by Galasso, G.]., Whitley, R.]., Merigan, T.e., Raven Press,Ltd., NewYork; Chapter 8-Herpes Simplex Virus. (1990).
Meza, E.: Estudio preliminar del potencial produccion de sangrede grado (Croton spp.) Selva Alta, Peru; Estudio preliminar demercado de sangre de grado, Lima, Peru., in press, 1994.
Mossmann, T.: (1983). Rapid colorimetric assay for cellular growth and survival: application to proliferation and cytotoxicityassays.]. Immunol. Meth. 65: 55-63.
Nonaka, G.-I., Lee, K.-H., Cheng, Y.-e. and Kilkuskie, R. E.: Inhibition of human retroviruses. Patent #WO 90/04968 (1990).
Persinos, G. ].: Anti-Inflammation Compositions Containing Taspine or Acid Salts Thereof and Method of Use, Patent#3,694,557,1972.
Persinos-Perdue E, Blomster, R.M., Blake, D.A., and Farnsworth,N. R.: South American Plants II: Taspine isolation and anti-inflammatory activity.]. Pharm. Sci. 68: 124-126, 1979.
Pevear, D. e., Fancher, M.]., Felock, P.]., Rossmann, M. G., Miller,M.S., Diana, G., Treasurywalla, A.M., McKinlay, M.A., andDutko, E].: Conformation change in the floor of the humanrhinovirus canyon blocks adsorption to HeLa cell receptors.]. Virol. 63: 2002-2007, 1989.
Robinson, W.S.: Hepadnaviruses and their replication. In: Virology, Second Edition (B.N. Fields and D.M. Knipe, eds.), pp.2137-2169. Raven Press, Ltd., New York, 1990.
Rozhon, E., Albin, R., and Schwartz, ].: Strategies for discoveringantiviral agents from natural products. In: The Discovery of Natural Products with Therapeutic Potential (V.P. Gullo, ed.), pp.223-245. Butterworth-Heinemann, Stoneham, Massachusetts,1994.
Rozhon, E.]., Cox, S., Buontempo, P.]., O'Connell, ]., et al.: SCH38057: a picornavirus capsid-binding molecule with antiviralactivity after the initial stage of uncoating. Antiviral Research21: 15-35, 1993.
Safrin, S., Phan, L. and Elbeik, T.: Evaluation of the in vitro activity of SP-303 against clinical isolates of acyclovir-resistant andfoscarnet-resistant herpes simplex virus. Abstract #136, Antiviral Res. 20 (suppl. 1): 117, 1993.
Schulman, ]. L.: Effect of I-amantanamine hydrochloride (amantadine HCl) on transmission of influenza virus infection in mice.Proc. Soc. Exp. BioI. Med. 128: 1173-1178, 1968.
Sells, M.A., Chen, M.-L., and Acs, G.: Production of hepatitis B

106 R. Ubillas et al.
virus part icles in HepG2 cells transfected with cloned hepatitis Bvirus DNA. Proc. Natl. Acad. Sci. USA 84: 1005-1009, 1987.
Selway, J. W.T.: Antiviral Activity of F1avones and Flavans, PlantFlavonoids in Biology and Medicine: Biochemical, Pharma cological, and Structure-Activity Relations, Alan R. Liss, Inc. NewYork pp. 521-536, 1986.
Shipman, c.,Jr., Smith, S.H., Carlson, R.H., and Drach,]. c.: Antiviral activity of arabinosyladen ine and ara binosylhypoxanthine in herpes simplex virus-infected KB cells: selective inhibition of viral deoxyribonucleic acid synthesis in synchronizedsuspension cultures. Antimicrob. Agents Chemother. 9:120-127,1976.
Smee, D.E, Martin, J. c., Verheyden, J. P.H., and Matthews, T.R.:Antiherpesvirus activity of the acyclic nucleoside 9-(1,3-dihydroxy-2propoxymethyl)guanine. Antimicrob. Agents Cbemother. 23: 676-682, 1983.
Smith, T.]., Kremer, M.J., Luo, M., Vriend, G., Arnold, E., Kamer,G., Rossmann, M. G., McKinlay, M. A., Diana, G.D., and Otto,M.J .: The site of attachment in human rhinovirus 14 for anti viral agents that inhibit uncoating. Science 233 : 1286-1293,1986.
Soikc, K.E, Zhang, ].- Y., and Meyerson, L. R.: Reduction of respiratory syncytial virus (RSV) shedding in African green monkeys treated with SP-303. Antiviral Res. Suppl. 1: 91, 1992.
Sprua nce, S. L. and McKeough, M. B.: 15% SP-303 in 95% DMSO and 5% ACV in PEG ointment in the treatment of dorsalcutaneous herpes simplex virus type 1 infection in guinea pigs.Unpubli shed data .
Superti, E, Seganti, L., Orsi, N., Divizia, M., Gabrieli, R., Pana,A., and Stein, M.L.: Effect of isoflavans and isoflavenes on theinfection of Frpl3 cells by hepatiti s A virus. Antiviral Res. 11:247-254,1989.
Takech i, M., Tanaka, Y., Takehara, M., Nonak a, G.!. , Nishioka,I.: Structure and antih erpetic activity among the tannins. Phytochemistry 24: 2245-2250, 1985.
Taylor, H.P., and Cooper, N.R.: Hum an cytomegalovirus bindingto fibroblasts is receptor mediated. J. Virol. 63: 3991-3998,1989.
Tempesta, M.: Proanthocyanidin Polymers Having AntiviralActivity and Methods of Obtaining Same. U.S. Patent #5, 211,944, 1993.
Vaisberg, A.]., Milia M., Del Carmen Planas M., Cordova J.L. ,Rosas de Agusti E., Ferreyra R., Del Carmen Mustiga M. , Carlin L. and Hammond G.B.: Taspine is the cicatrizant principle insangre de grado extracted from Croton lechleri. Planta Med. 55:140-143,1 989.
Vanden Berghe, D.A., Vlietinck, A.J., and Van Hoof, L.: PlantProducts as Potent ial Antiviral Agents. Bull. Inst. Pasteur.101- 147, 1986.
Vlietinck, A.]. : Biologically active substances from traditionaldrugs. In: K. Hostettmann and P.J . Lea (Eds.), Biologically Active Natural Products. Clarendon Press, Oxford, Chapt. 3,1987.
Von Reis S. and Lipp EJ. , Jr.: New plant sources for drugs andfoods from the New York Botanical Garden . Harvard University Press, Cambridge, Massachusetts, p. 139, p. 155, 1982.
Walker, J. S., Stephen, E.L., Spertzel, R.O.: Small particle aerosolsof antiviral compounds in treatment of type A influenza pneumonia in mice.]. Infect. Dis. 133 (suppl.), A140-144, 1976.
Webster, G.: A provisional synopsis of the sections of the genusCroton (Euphorbiaceae). Taxo n 42: 794-823, 1993.
Williams, V.M., Porter, L.]. and Hemingway, R. W.: Molecularweight profiles of proanth ocyan idin polymers. Phytochemistry22: 569 , 1983.
Wyde, P.R., Ambrose, M.W., Meyerson, L.R., and Gilbert, B. E.:The antiviral activity of SP-303, a natural polyphenolic polymer,against respiratory syncytial and parainfluenza type 3 viruses incotton rats. Antiviral Res. 28: 145-1 54, 1993a.
Wyde, P.R., Meyerson, L.R., and Gilbert . B.E.: In vitro evaluationof the antiviral activity of SP-303, an Euphorbiaceae shrub extract, against a panel of respiratory viruses. Drug. Develop. Res.28: 467-472, 1993b.
Wyde, P.R., Wilson, S.Z., Gilbert, B.E ., and Smith, R.H.A.: Protection of mice- from lethal influenza virus infection with highdose - short duration ribavirin aeroso l. Antimicrob. AgentsChemother. 30 : 942-944, 1986.
Address:
M.S. Tempesta, Shaman Pharm aceuticals, 213 East GrandAvenue, South San Francisco, CA 94080-4812, U.S.A.