spontaneous or natural killer cytotoxicity of k562 ... · james e. nagel, gary d. collins, and...
TRANSCRIPT

[CANCER RESEARCH 41, 2284-2288, June 1981]0008-5472/81 /0041-0000$02.00
Spontaneous or Natural Killer Cytotoxicity of K562 ErythroleukemicCells in Normal Patients
James E. Nagel, Gary D. Collins, and William H. Adler1
Clinical Immunology Section, Gerontology Research Center, National Institute on Aging. NIH, Baltimore, Maryland 21224
ABSTRACT
Peripheral blood mononuclear cells from 200 normal individuals, ages 20 to 95 years, were evaluated for their capabilityto mediate spontaneous or natural killer (NK) cytotoxicity usingthe K562 erythroleukemic cells as targets. The results of a 4-hr 51Crspecific release assay demonstrated that the NK activity
of normal individuals is independent of age, sex, and smokinghabits. Although varying greatly among individuals, the NKactivity of 25 persons restudied after a mean 20-month interval
remained stable. Deficient NK lytic activity is not characteristicof elderly individuals.
INTRODUCTION
The in vitro lysis of tumor cells by lymphocytic effector cellsmay proceed through the activities of at least 3 types ofcytotoxic mononuclear cells: cytotoxic T-lymphocytes; antibody-dependent killer cells; and antibody-independent NK.2
Among this functional classification, NK putatively have theunique ability to recognize, without prior sensitization, nonan-tibody-coated targets and to spontaneously lyse these allo-
genic, syngeneic, and xenogeneic tumor cells. This capabilityto detect and eliminate nonspecifically foreign cells on initialexposure has led to the proposal that NK function as theprimary effectors of immunosurveillance (15, 24, 33). Althoughknown to be nonphagocytic, nonadherent, and heterogeneousfor the presence of Fc receptors for IgG, further morphologicalcharacteristics as well as the origin of human NK have notbeen fully defined. Increasing experimental evidence indicatesthat both the spontaneous (NK) and antibody-dependent cel
lular cytotoxic activities of peripheral blood cells are mediatedby the same subpopulations, which have been characterizedby various investigators to be a T-lymphocyte, promonocyte,or "null" cell or to be of myeloid origin (21, 22, 32, 39).
Many T-cell-mediated immune functions diminish with ad
vancing age; however, the effect of aging on NK activity isunclear. For example, in mice and rats in which the age-related
change is most easily demonstrated, the NK activity of spleencells is absent in the very young, peaks at 5 to 8 weeks of age,and then decreases thereafter. Most studies of human NKactivity have indicated that reactivity does not diminish withage; however, many published investigations have dealt withsmall numbers of individuals or have studied NK activity in ill ortumor-bearing patients of various ages (8, 20, 30, 37, 46, 47).
Despite the lack of conclusive evidence demonstrating an in
1To whom requests for reprints should be addressed.2 The abbreviations used are: NK, natural killer cell(s); BLSA, Baltimore
Longitudinal Study of Aging; RPMI, Roswell Park Memorial Institute; PCS, fetalcalf serum.
Received October 20, 1980; accepted March 2, 1981.
vivo cause and effect relationship between diminished NKactivity and neoplasia, there are sufficient data from the experimental mouse model to support the conclusion that, undercertain circumstances, NK are important in resistance to tumorgrowth (1, 13, 33). The proposed immunosurveillance role ofNK, coupled with recent reports on vital statistics demonstrating that death from malignant neoplasms ranks second tocardiovascular disease as the leading cause of death in individuals over 65 years of age, prompted the present investigation of a large group of well-characterized normal adults on the
effect of age on human NK activity.
MATERIALS AND METHODS
Study Population. The study population was composed of200 adults, ages 20 to 95 years, who are participants in theBLSA. The study group that is reported here included 50females and 150 males. The BLSA is an investigation of normative human aging which began for males in 1958. A companion program for females was initiated in 1978. The self-recruited participants voluntarily return approximately every 18to 24 months to undergo a 3-day inpatient réévaluationof
psychological, biomédical, and physical changes. Essentially,all participants are ambulatory, and most live in their ownresidence with a spouse. Stone and Norris (43) have summarized previously the demographic and social characteristics ofthe male participants. The male subjects included in this reporthave been reevaluated an average of 8 (range, 2 to 19) timesand the female participants have been reevaluated twice. TheBLSA program does not make any overt attempts to alter thelife style or living habits of the participants, other than to advisethe subjects and their personal physicians of any significantmedical abnormalities which may be detected during theirperiodic evaluation.
For the purposes of this study, smokers were defined asthose individuals who have a history of smoking at least 10cigarettes/day. Nonsmokers were considered to be those whowere not smoking at the time of their evaluation, but this groupdoes include former smokers.
Isolation of Mononuclear Cells. Heparinized peripheralblood (20 ml) was diluted 1:4 with RPMI Medium 1640 (GrandIsland Biological Co., Grand Island, N. Y.) and separated bydifferential buoyancy centrifugation on a Ficoll:sodium diatri-
zoate cushion (LSM; Litton Bionetics, Kensington, Md.). Theinterface cells were removed with a Pasteur pipet and washedtwice with RPMI Medium 1640, and the mononuclear cell countwas determined in a hemocytometer.
Cytototoxicity Assay. Cultured cells from the K562 erythroleukemic cell line (provided by Dr. R. B. Herberman, NationalCancer Institute, NIH) grown in RPMI Medium 1640 supplemented with 10% PCS, 10 rriM, 4-(2-hydroxyethyl)-1-piperazi-
2284 CANCER RESEARCH VOL. 41
on April 1, 2020. © 1981 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
NK Activity and Aging
neethanesulfonic acid, 2 mM L-glutamine, and 100 /ig genta-
mycin per ml were used as target cells. The line was routinelysubcultured the day before use in an assay. Prior to an assay,the target cells (2 x 106 in 0.15 ml of Tris buffer, pH 7.4) wereincubated with occasional mixing with 0.2 /iCi of Na251CrO4 in
0.2 ml of sterile isotonic 0.9% NaCI solution (Amersham/SearleCorp., Arlington Heights, III.; specific activity 50 to 100 Ci/mgchromium) for 30 min at 37°. The chromium-labeled K562
target cells were then centrifuged twice through 2 ml of heat-inactivated PCS (to remove unincorporated 51Cr), then resus-
pended in RPMI Medium 1640 containing 10% PCS, andcounted. After adjusting the density to 1 x 10s cells/ml in
RPMI Medium 1640 with 25 mM 4-(2-hydroxyethyl)-1 -pipera-zineethanesulfonic acid and 10% heat-inactivated PCS, 100/il (10" radio-labeled target cells) were dispensed into each
well of a round-bottom microtiter plate (Linbro Chemical, New
Haven, Conn.) containing 100 /il of the serially diluted effectorcells. Eight target:effector ratios from 1:100 to 1:0.78 wereused. Each ratio was run in duplicate. Following the addition ofthe target cells, the microtiter plates were centrifuged for 3 minat 400 x g and then incubated at 37° in a humidified atmo
sphere containing 5% COj:95% air. After 4-hr incubation, 100
/il of supernatant were removed from each well and placed inindividual 12- x 75-mm glass tubes, and the radioactivity was
determined in a gamma counter (Nuclear Chicago, Des Plaines,III). The percentage of specific 51Cr release was calculated as
% of releaseE-S
W- S x 100
where £is cpm in the experimental tube, S is cpm spontaneousrelease, and W is cpm maximal isotope release effected bywater lysis. Dose-response curves were obtained for eachindividual by plotting the percentage of specific 51Cr release
obtained at each of 8 effectortarget ratios and the logic of thenumber of effector cells. The mean spontaneous 51Cr release
by the radiolabeled K562 targets in the absence of effectorcells was 7.1%.
RESULTS
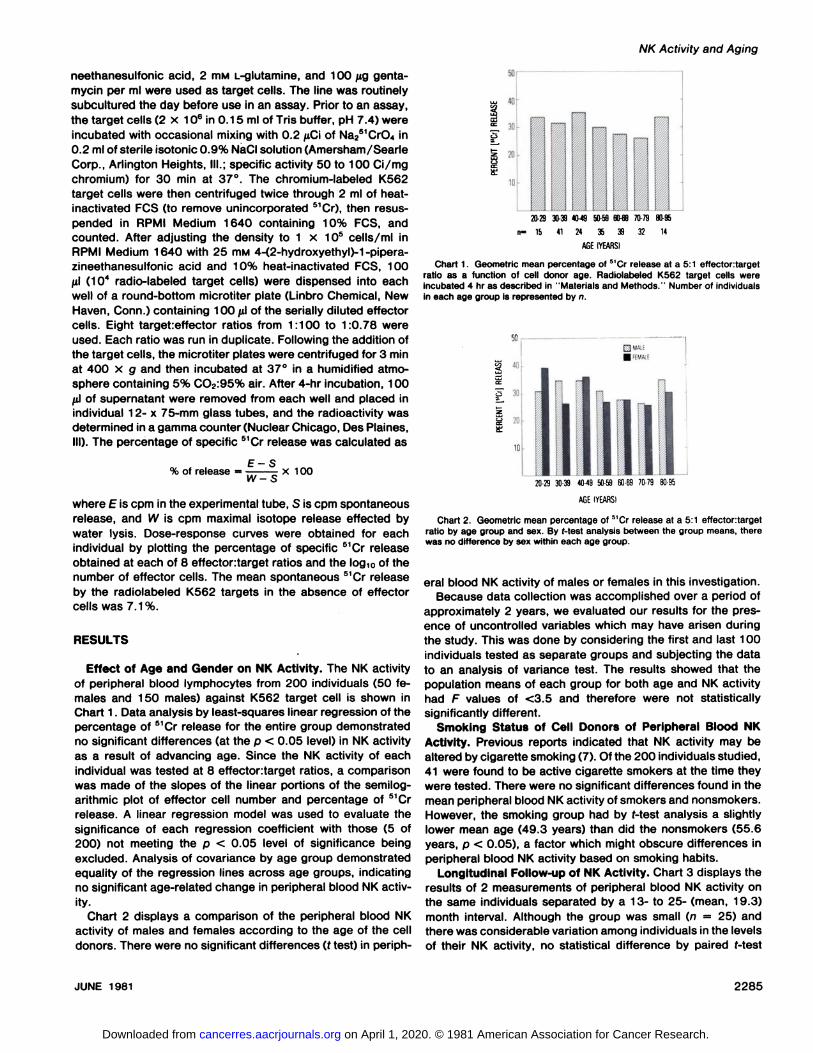
Effect of Age and Gender on NK Activity. The NK activityof peripheral blood lymphocytes from 200 individuals (50 females and 150 males) against K562 target cell is shown inChart 1. Data analysis by least-squares linear regression of thepercentage of 51Cr release for the entire group demonstrated
no significant differences (at the p < 0.05 level) in NK activityas a result of advancing age. Since the NK activity of eachindividual was tested at 8 effectortarget ratios, a comparisonwas made of the slopes of the linear portions of the semilog-arithmic plot of effector cell number and percentage of 51Cr
release. A linear regression model was used to evaluate thesignificance of each regression coefficient with those (5 of200) not meeting the p < 0.05 level of significance beingexcluded. Analysis of covariance by age group demonstratedequality of the regression lines across age groups, indicatingno significant age-related change in peripheral blood NK activ
ity.Chart 2 displays a comparison of the peripheral blood NK
activity of males and females according to the age of the celldonors. There were no significant differences (t test) in periph-
20-2930-394M9 50-59SM9 70-7980-95
n- 15 41 24 35 39 32 14
AGEIYEARSI
Chart 1. Geometric mean percentage of 51Cr release at a 5:1 effectontarget
ratio as a function of cell donor age. Radiolabeled K562 target cells wereincubated 4 hr as described in "Materials and Methods." Number of individuals
in each age group is represented by n.
20-2930-394049 50-5960-697079 80-95
AGEIYEARSI
Chart 2. Geometric mean percentage of 5'Cr release at a 5:1 effectortarget
ratio by age group and sex. By f-test analysis between the group means, therewas no difference by sex within each age group.
eral blood NK activity of males or females in this investigation.Because data collection was accomplished over a period of
approximately 2 years, we evaluated our results for the presence of uncontrolled variables which may have arisen duringthe study. This was done by considering the first and last 100individuals tested as separate groups and subjecting the datato an analysis of variance test. The results showed that thepopulation means of each group for both age and NK activityhad F values of <3.5 and therefore were not statisticallysignificantly different.
Smoking Status of Cell Donors of Peripheral Blood NKActivity. Previous reports indicated that NK activity may bealtered by cigarette smoking (7). Of the 200 individuals studied,41 were found to be active cigarette smokers at the time theywere tested. There were no significant differences found in themean peripheral blood NK activity of smokers and nonsmokers.However, the smoking group had by f-test analysis a slightlylower mean age (49.3 years) than did the nonsmokers (55.6years, p < 0.05), a factor which might obscure differences inperipheral blood NK activity based on smoking habits.
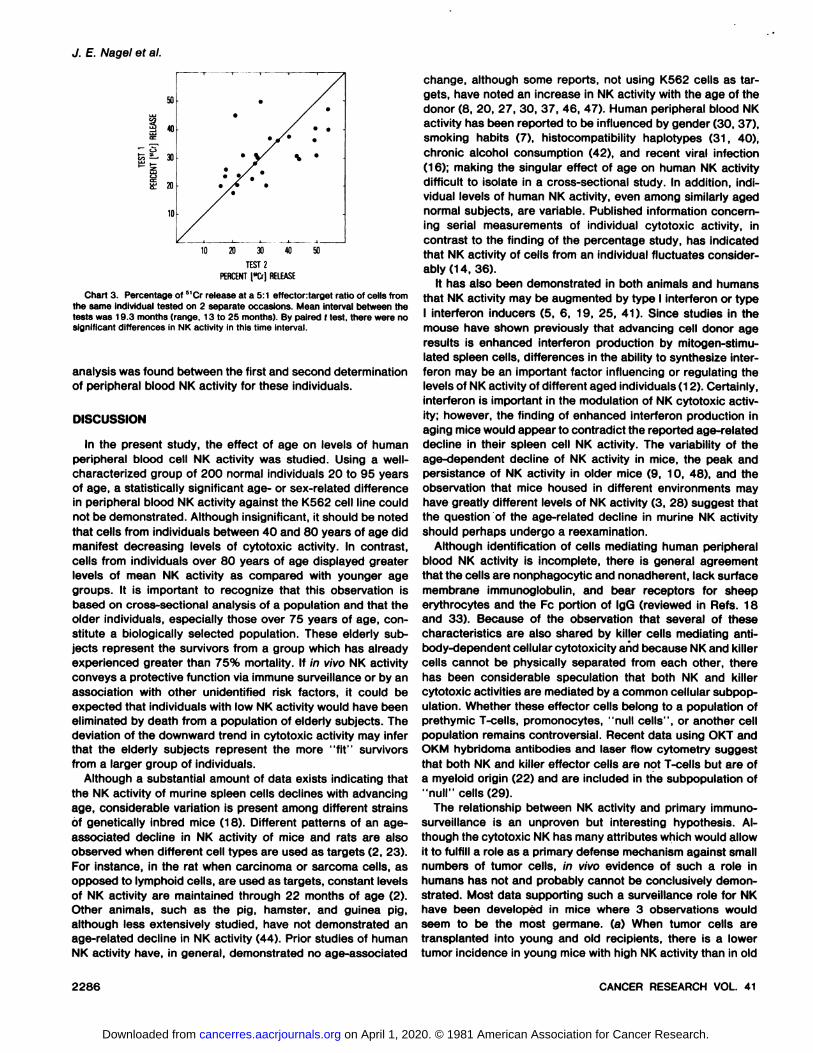
Longitudinal Follow-up of NK Activity. Chart 3 displays theresults of 2 measurements of peripheral blood NK activity onthe same individuals separated by a 13- to 25- (mean, 19.3)month interval. Although the group was small (n = 25) andthere was considerable variation among individuals in the levelsof their NK activity, no statistical difference by paired f-test
JUNE 1981 2285
on April 1, 2020. © 1981 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
J. E. Nagel et al.
M
§ 40K
^ »
È 20
10
10 20 30 « 50
TEST2PfRCENT[»&)RELEASE
Chart 3. Percentage of 51Cr release at a 5:1 effectortarget ratio of cells from
the same individual tested on 2 separate occasions. Mean interval between thetests was 19.3 months (range, 13 to 25 months). By paired ( test, there were nosignificant differences in NK activity in this time interval.
analysis was found between the first and second determinationof peripheral blood NK activity for these individuals.
DISCUSSION
In the present study, the effect of age on levels of humanperipheral blood cell NK activity was studied. Using a well-characterized group of 200 normal individuals 20 to 95 yearsof age, a statistically significant age- or sex-related difference
in peripheral blood NK activity against the K562 cell line couldnot be demonstrated. Although insignificant, it should be notedthat cells from individuals between 40 and 80 years of age didmanifest decreasing levels of cytotoxic activity. In contrast,cells from individuals over 80 years of age displayed greaterlevels of mean NK activity as compared with younger agegroups. It is important to recognize that this observation isbased on cross-sectional analysis of a population and that theolder individuals, especially those over 75 years of age, constitute a biologically selected population. These elderly subjects represent the survivors from a group which has alreadyexperienced greater than 75% mortality. If in vivo NK activityconveys a protective function via immune surveillance or by anassociation with other unidentified risk factors, it could beexpected that individuals with low NK activity would have beeneliminated by death from a population of elderly subjects. Thedeviation of the downward trend in cytotoxic activity may inferthat the elderly subjects represent the more "fit" survivors
from a larger group of individuals.Although a substantial amount of data exists indicating that
the NK activity of murine spleen cells declines with advancingage, considerable variation is present among different strainsof genetically inbred mice (18). Different patterns of an age-
associated decline in NK activity of mice and rats are alsoobserved when different cell types are used as targets (2, 23).For instance, in the rat when carcinoma or sarcoma cells, asopposed to lymphoid cells, are used as targets, constant levelsof NK activity are maintained through 22 months of age (2).Other animals, such as the pig, hamster, and guinea pig,although less extensively studied, have not demonstrated anage-related decline in NK activity (44). Prior studies of humanNK activity have, in general, demonstrated no age-associated
change, although some reports, not using K562 cells as targets, have noted an increase in NK activity with the age of thedonor (8, 20, 27, 30, 37, 46, 47). Human peripheral blood NKactivity has been reported to be influenced by gender (30, 37),smoking habits (7), histocompatibility haplotypes (31, 40),chronic alcohol consumption (42), and recent viral infection(16); making the singular effect of age on human NK activitydifficult to isolate in a cross-sectional study. In addition, indi
vidual levels of human NK activity, even among similarly agednormal subjects, are variable. Published information concerning serial measurements of individual cytotoxic activity, incontrast to the finding of the percentage study, has indicatedthat NK activity of cells from an individual fluctuates considerably (14, 36).
It has also been demonstrated in both animals and humansthat NK activity may be augmented by type l Interferon or typel Interferon inducers (5, 6, 19, 25, 41). Since studies in themouse have shown previously that advancing cell donor ageresults is enhanced Interferon production by mitogen-stimu-lated spleen cells, differences in the ability to synthesize inter-feron may be an important factor influencing or regulating thelevels of NK activity of different aged individuals (12). Certainly,Interferon is important in the modulation of NK cytotoxic activity; however, the finding of enhanced Interferon production inaging mice would appear to contradict the reported age-relateddecline in their spleen cell NK activity. The variability of theage-dependent decline of NK activity in mice, the peak and
persistance of NK activity in older mice (9, 10, 48), and theobservation that mice housed in different environments mayhave greatly different levels of NK activity (3, 28) suggest thatthe question of the age-related decline in murine NK activity
should perhaps undergo a reexamination.Although identification of cells mediating human peripheral
blood NK activity is incomplete, there is general agreementthat the cells are nonphagocytic and nonadherent, lack surfacemembrane immunoglobulin, and bear receptors for sheeperythrocytes and the Fc portion of IgG (reviewed in Refs. 18and 33). Because of the observation that several of thesecharacteristics are also shared by killer cells mediating antibody-dependent cellular cytotoxicity and because NK and killer
cells cannot be physically separated from each other, therehas been considerable speculation that both NK and killercytotoxic activities are mediated by a common cellular subpop-ulation. Whether these effector cells belong to a population ofprethymic T-cells, promonocytes, "null cells", or another cell
population remains controversial. Recent data using OKT andOKM hybridoma antibodies and laser flow cytometry suggestthat both NK and killer effector cells are not T-cells but are of
a myeloid origin (22) and are included in the subpopulation of"null" cells (29).
The relationship between NK activity and primary immuno-surveillance is an unproven but interesting hypothesis. Although the cytotoxic NK has many attributes which would allowit to fulfill a role as a primary defense mechanism against smallnumbers of tumor cells, in vivo evidence of such a role inhumans has not and probably cannot be conclusively demonstrated. Most data supporting such a surveillance role for NKhave been developed in mice where 3 observations wouldseem to be the most germane, (a) When tumor cells aretransplanted into young and old recipients, there is a lowertumor incidence in young mice with high NK activity than in old
2286 CANCER RESEARCH VOL. 41
on April 1, 2020. © 1981 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from

NK Activity and Aging
mice with diminished NK activity (11). (Jb) There is a higherincidence of spontaneous tumors and leukemias among strainsof mice with genetically low NK activity compared with strainshaving high NK activity (48). (c) NK activity may be increasedby the injection of tumor cells (16). The possible relationship ofthese findings to the increased incidence of neoplasia in elderlyhumans without demonstrably lower levels of NK activity remains speculative. While the present cross-sectional investi
gation has demonstrated that peripheral blood NK activity didnot diminish with age or gender, it is important to consider 2elements of its experimental design which may be relevant tothe results. Since only one cell type, the K562 erythroleukemiacell line, was used as a target, it is important to note that in themouse model, spleen cell NK activity directed against targetsof either solid tumor or lymphoid origin displays different age-
related patterns. Another important consideration is that, sinceour study did not evaluate the NK activity of individuals whowere less than 20 years of age, the possibility exists that peakNK activity may occur in younger individuals, and the levelsseen in our study group represent decreases from earlier peakactivity. This possibility has been suggested by results of otherstudies (8).
Several additional points should also be emphasized. Assuggested by findings from our laboratory and others whichindicate that peripheral blood NK activity may be altered byenvironmental factors, diet, and viral infections, a measurementof the ability to generate an NK cytotoxic response to anappropriate stimulus may be more critical than quantitation ofa "basal" level of NK activity in peripheral blood. This would
appear, in particular, to be the case if the modulation of NKactivity by interferon is found to be related to specific in vivopathological conditions. Other reports indicating that the augmenting effects of interferon on NK activity occur by recruitment of precursor cells from the bone marrow (38) and thatcyclic nucleotides may modulate NK cytolysis (34) suggest avariety of other sites at which age-related changes in function
could effect NK activity. Studies that compare human NKactivities against allogeneic and autologous tumors point outthat a critical point in tumor immunosurveillance may be thecapacity of the NK effector cell to recognize the tumor cellantigenic determinants (26). Although distinct target structureshave been reported on lymphoma cell lines (35), their role inNK-mediated cytolysis has been questioned (4). Whether there
are alterations in tumor recognition ability that are secondaryto age-related changes is unknown (45). An additional investi
gation of all of these points will be needed to answer theimportant question of the relevance of NK cytolysis to thefrequent occurrence of neoplasia in the elderly.
REFERENCES
1. Baldwin, R. W. Immune surveillance revisited. Nature (Lond ), 270 5571977.
2. Brooks, C. G., and Flannery, G. R. Quantative studies of natural immunity tosolid tumors in rats. Persistance of natural immunity throughout reproductivelife, and the absence of suppressor cells in infant rats. Immunology, 39.187-194, 1980.
3. Clark, E. A., Russell, P. H., Egghart, M., and Morton, M. A. Characteristicsand genetic control of NK-cell-mediated cytotoxicity activated by naturallyacquired infection in the mouse. Int. J. Cancer, 24: 688-699, 1979.
4. Collins, J. L., Patek, P. Q., and Cohn, M. Tumorigenicity and lysis by naturalkillers. J. Exp. Med., J53: 89-106, 1981.
5. Djeu, J. Y., Heinbaugh, J. A., Holden, H. T., and Herberman, R. B. Augmen
tation of mouse natural killer cell activity by interferon and interferon in-ducers. J. Immunol., 722: 175-181, 1979.
6. Einhorn, S.. Blomgren, H., and Strander, H. Interferon and spontaneouscytotoxicity in man. I. Enhancement of the spontaneous cytotoxicity ofperipheral lympphocytes in human leukocyte inferieron. Int. J. Cancer, 22:405-412, 1978.
7. Person, M., Edwards, A., Lind, A., Milton, G. W., and Mersey, P. Low naturalkiller-cell activity and immunoglobulln levels associated with smoking inhuman subjects. Int. J. Cancer, 23: 603-609, 1979.
8. Forbes, J. T., Greco, F. A., and Oldham, R. K. Natural cell-mediatedcytotoxicity in human tumor patients. In: R. 6. Herberman (ed.), Natural Cell-mediated Immunity against Tumors, pp. 1031-1046. New York: AcademicPress, Inc., 1980.
9. Gomard, E., Ledere, J. C., and Levy, J. P. Spontaneous antilymphomareaction of preleukaemic AKR mice is a non-T-cell killing. Nature (Lond ),250:671-673. 1974.
10. Greenberg, A. H., and Playfair, J. H. L. Spontaneously arising cytotoxicitytotheP-815-Ymastocytomain NZB mice. Clin. Exp. Immunol., 76:99-110,1974.
11. Mailer, O., Hansson, M.. Kiessling, R.. and Wigzell. H. Role of nonconven-tional natural killer cells in resistance against syngeneic tumor cells in vivo.Nature (Lond.), 270: 609-611, 1977.
12. Heine, J.. and Adler, W. H. The quantitative production of interferon bymitogen-stimulated mouse lymphocytes as a function of age and its effecton the lymphocyte proliferative response. J. Immunol., 778: 1366-1369.1977.
13. Henney, C. S., Tracey, D.. Durdik, J. M., and Kumpel, G. Natural killer cells.In vitro and in vivo. Am. J. Pathol., 93: 459-467, 1978.
14. Heppner, G., Stolbach, H. L., Cummings, F., McDonough, E., and Calabresi,P. Problems in the clinical use of the microcytotoxicity assay for measuringcell-mediated immunity to tumor cells. Cancer Res., 35:1931-1937, 1975.
15. Herberman, R. B., Djeu, J. Y., Kay. H. D., Ortaldo, J. R., Riccardi, C.,Bonnard, G. D.. Holden, H. T., Fagnani, R., Santoni, A., and Puccetti, P.Natural killer cells: characteristics and regulation of activity. Immunol. Rev.,44:43-70. 1979.
16. Herberman, R. B., and Holden, H. T. Natural cell-mediated immunity. Adv.Cancer Res.. 27: 305-377, 1978.
17. Herberman, R. B., Nunn, M. E., Holden. H. T., Staal, S., and Djeu, J. Y.Augmentation of natural cytotoxic reactivity of mouse lymphoid cells againstsyngeneic and allogeneic targets. Int. J. Cancer, 79: 555-564, 1977.
18. Herberman, R. B., Nunn, M. E.. and Lavrin, D. H. Natural cytotoxic reactivityof mouse lymphoid cells against syngeneic and allogenic tumors. I. Distribution of reactivity and specificity. Int. J. Cancer, 76: 216-229, 1975.
19. Herberman, R. B., Ortaldo, J., and Bonnard, G. Augmentation by interferonof human natural and antibody-dependent cell-mediated cytotoxicity. Nature(Lond.), 277: 221-223, 1979.
20. Mersey, P., Edwards, A., and McCarthy, W. H. Tumor-related changes innatural killer cell activity in melanoma patients. Influence of stage of disease,tumor thickness, and age of patients. Int. J. Cancer, 25: 187-194, 1980.
21. Kaplan, J., and Callewaert, D. M. Expression of human T-lymphocyte antigens by natural killer cells. J. Nati. Cancer Inst., 60: 961 -964, 1978.
22. Kay. H. D., and Horowitz, D. A. Evidence by reactivity with hybridomaantibodies for a probable myeloid origin of peripheral blood cells active innatural cytotoxicity and antibody-dependent cell-mediated cytotoxicity. J.Clin. Invest., 66. 847-851, 1980.
23. Kumar, V., Luevano, E., and Bennett, M. Hybrid resistance to EL-4 lymphomacells. I. Characterization of natural killer cells that lyse EL-4 cells and theirdestruction from marrow-dependent natural killer cells. J. Exp. Med., 750:531-547, 1979.
24. Lotzova, E., and McCredie, K. B. Natural killer cells in mice and man andtheir possible biological significance. Cancer Immunol. Immunother.. 4:215-221, 1978.
25. Moore, M., and Potter, M. R. Enhancement of human natural cell-mediatedcytotoxicity by interferon. Br. J. Cancer, 47: 378-387. 1980.
26. Mukherji, B., Flowers, A., Rothman, L., and Nathanson, L. Spontaneous invitro cytotoxicity against autochthonous human melanoma cells. J. Immunol.,724:412-417, 1980.
27. Oldham, R. K., Djeu, J. Y., Cannon, G. G., Siwarski, D., and Herberman, R.B. Cellular microcytotoxicity in human tumor systems: analysis of results. J.Nati. Cancer Inst., 55: 1305-1318, 1975.
28. Örn,A., Gidlund, M., Ojo, E., Grönvik, K-O., Andersson, J., and Wigzell. H.Factors controlling the augmentation of natural killer cells. In: R. B. Herberman (ed.), Natural Cell-mediated Immunity against Tumors, pp. 581-592.
New York: Academic Press, Inc., 1980.29. Ozer, H., Strelkauskas, A. J., Gallery, R. T., and Schlossman, S.'F. The
functional dissection of human peripheral blood null cells wtih respect toantibody-dependent cellular cytotoxicity and natural killing. Eur. J. Immunol.,9: 112-118, 1979.
30. Penschow, J., and Mackay, I. R. NK and K cell activity of human blood:differences according to sex, age, and disease. Ann. Rheum. Dis., 39: 82-
86, 1980.31. Petranyi. G., Ivanyi, P., and Hollan, S. R. Relations of HL-A and Rh systems
to immune activity. Vox. Sang., 26: 470-482, 1974.
JUNE 1981 2287
on April 1, 2020. © 1981 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from

J. E. Nagel et al.
32. Pichler, W. J., Gendelman, F. W., and Nelson, D. L. Fc receptors on human 40.T lymphocytes. II. Cytotoxic capabilities of human Ty, T/i, B, and L cells.Cell. Immunol., 42. 410-417, 1979.
33. Pross, H. F.. and Baines, M. G. Spontaneous human lymphocyte-mediated 41.
cytotoxicity against tumor target cells. VI. A brief review. Cancer Immunol.Immunother., 3: 75-85, 1977.
34. Roder, J. C., and Klein, M. Target-effector interaction in the natural killer 42.cell system. IV. Modulation by cyclic nucleotides. J. Immunol., 723. 2785-
2790, 1979.35. Roder, J. C., Rosen, A., Fenyo, E. M., and Troy, F. A. Target effector 43.
interaction in the natural killer system: isolation of target structures. Proc.Nati. Acad. Sei. U. S. A.. 76: 1405-1409, 1979. 44.
36. Rosenberg. E. B., McCoy, J. L.. Green, S. S.. Donnelly, F. C.. Siwarski, D.F., Levine, P. H., and Herberman, R. B. Destruction of human lymphoidtissue culture cell lines by human peripheral lymphocytes in 5'Cr-release 45.
cellular cytotoxicity assays. J. Nati. Cancer Inst., 52. 345-352, 1974.37. Saksela, E., Timonen, G., Ranki, A., and Hayry, P. Morphological and
functional characterization of isolated effector cells responsible for human 46.natural killer activity to fetal fibroblasts and to cultured cell line targets.Immunol. Rev., 44: 71-123, 1979.
38. Saksela. E., Timonen, T., and Cantell, K. Human natural killer cell activity is 47.augmented by Interferon via recruitment of "pre-NK" cells. Scand. J.
Immunol., 10.257-266. 1979.39. Santoli. D., Trinchieri, G.. Moretta, L., Zmijewski, C. M., and Koprowski, H. 49.
Spontaneous cell-mediated cytotoxicity in humans. Distribution and characterization of the effector cell. Clin. Exp. Immunol.. 33. 309-318, 1978.
Santoli. D.. Trinchieri, G., Zmijewski, C. M., and Koprowski, H. HLA-relatedcontrol of spontaneous and antibody-dependent cell-mediated cytotoxicactivity in humans. J. Immunol., 117: 765-770, 1976.Sato, T.. Fuse, A., and Kuwata. T. Enhancement by interferon of naturalcytotoxic activities of lymphocytes from human cord blood and peripheralblood of aged persons. Cell. Immunol., 45: 458-463, 1979.Saxena, Q. B., Mezey, E., and Adler, W. H. Regulation of natural killeractivity in vivo. II. Effect of alcohol consumption on human peripheral bloodnatural killer activity. Int. J. Cancer, 26. 413-417, 1980.Stone. J. L., and Norris. A. H. Activities and attitudes of participants in theBaltimore Longitudinal Study. J. Gerontol.. 27. 575-580. 1966.Stutman, O. Ontogeny and other age-related effects of natural cytotoxic(NC) cells in mice. In: R. B. Herberman (ed.). Natural Cell-mediated Immunityagainst Tumors, pp. 231-240. New York: Academic Press, Inc., 1980.
Tai. A. S.. and Warner, N. L. Characterization of murine natural killer (NK)cells: analysis of recognition patterns by effector cells. Proc. Am. Assoc.Cancer Res., 27. 229, 1980Takasugi, M., Mickey, M. R., and Terasaki, P. I. Reactivity of lymphocytesfrom normal persons on cultured tumor cells. Cancer Res.. 33. 2898-2902,1973.Takasugi. M., Ramseyer, A., and Takasugi, J. Decline of natural nonselectivecell-mediated cytotoxicity in patients with tumor progression. Cancer Res.,37. 413-418. 1977.
Zarling, J. M., Nowinski. R. C., and Bach, F. H. Lysis of leukemia cells byspleen cells of normal mice. Proc. Nati. Acad. Sei. U. S. A., 72 2780-2784,1975.
2288 CANCER RESEARCH VOL. 41
on April 1, 2020. © 1981 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from

1981;41:2284-2288. Cancer Res James E. Nagel, Gary D. Collins and William H. Adler Erythroleukemic Cells in Normal PatientsSpontaneous or Natural Killer Cytotoxicity of K562
Updated version
http://cancerres.aacrjournals.org/content/41/6/2284
Access the most recent version of this article at:
E-mail alerts related to this article or journal.Sign up to receive free email-alerts
Subscriptions
Reprints and
To order reprints of this article or to subscribe to the journal, contact the AACR Publications
Permissions
Rightslink site. Click on "Request Permissions" which will take you to the Copyright Clearance Center's (CCC)
.http://cancerres.aacrjournals.org/content/41/6/2284To request permission to re-use all or part of this article, use this link
on April 1, 2020. © 1981 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from