streptomyces amritsarensis sp. nov., exhibiting broad-spectrum antimicrobial activity
TRANSCRIPT

ORIGINAL PAPER
Streptomyces amritsarensis sp. nov., exhibitingbroad-spectrum antimicrobial activity
Deepika Sharma • Shanmugam Mayilraj •
Rajesh Kumari Manhas
Received: 31 October 2013 / Accepted: 10 March 2014 / Published online: 25 March 2014
� Springer International Publishing Switzerland 2014
Abstract A new actinobacterium strain, designated
2AT, was isolated from a soil sample collected from
Guru Nanak Dev University, Punjab (India) and
characterized using a polyphasic taxonomic approach.
It showed antimicrobial activity against various Gram-
positive and Gram-negative bacteria including drug
resistant bacteria and fungi. The strain had chemotax-
ononomic and morphological properties typical of the
genus Streptomyces. The 16S rRNA gene sequence of
the strain showed 99.9, 99.5 and 99.5 % similarity
with Streptomyces flavotricini DSM 40152T, Strepto-
myces toxytricini DSM 40178T and Streptomyces
globosus DSM 40815T, respectively. This strain
formed a coherent cluster with them and shared
DNA–DNA homology of 37.6 ± 0.6, 34.4 ± 0.5 and
33.1 ± 0.4 % with type strains, S. flavotricini DSM
40152T, S. globosus DSM 40815T and S. toxytricini
DSM 40178T, respectively. Further, the strain was
readily distinguished from the phylogenetic close
relatives in a variety of morphological, physiological
and biochemical properties. Based on the genotypic
and phenotypic characteristics, it is proposed that
strain 2AT represents a novel species in the genus
Streptomyces, for which the name Streptomyces
amritsarensis sp. nov. is proposed, with the type strain
2AT (=MTCC 11845T=JCM 19660T).
Keywords Actinobacterium � Streptomyces
amritsarensis sp. nov � Polyphasic taxonomy
Introduction
The genus Streptomyces, encompassing nearly 600
species with validly published names (Euzeby 2012),
is the largest genus in domain Bacteria. The genus,
first described by Waksman and Henrici (1943),
includes aerobic, Gram-positive, high G?C content
(69–78 mol%) bacteria producing extensively branch-
ing networks of substrate mycelia that give rise to the
vertical branching aerial hyphae bearing chains of
spores. Members of this genus are well known for their
production of antibiotics. However, to combat the
problem of microbial resistance and development of
new antimicrobials to substitute the ineffective ones, it
is imperative to search new, efficient and safe
antibiotics. To discover new therapeutic compounds
we have to screen phylogenetically novel streptomy-
cetes which will avoid the costly rediscovery of
already known compounds from Streptomyces species
Electronic supplementary material The online version ofthis article (doi:10.1007/s10482-014-0151-2) contains supple-mentary material, which is available to authorized users.
D. Sharma � R. K. Manhas (&)
Department of Microbiology, Guru Nanak Dev
University, Amritsar 143005, India
e-mail: [email protected]
S. Mayilraj
Microbial Type Culture Collection and Gene Bank
(MTCC), CSIR-Institute of Microbial Technology
(IMTECH), Chandigarh 160036, India
123
Antonie van Leeuwenhoek (2014) 105:943–949
DOI 10.1007/s10482-014-0151-2

(Antony-Babu and Goodfellow 2008). In a screening
program for novel antimicrobial compounds from
actinobacteria, isolate 2AT which demonstrated potent
antimicrobial activity against various organisms
including drug resistant bacteria [methicillin resistant
Staphylococcus aureus (MRSA), vancomycin-resis-
tant Enterococcus (VRE) and multi drug resistant
E. coli] was isolated from a soil sample collected from
Guru Nanak Dev University, Amritsar, Punjab (India).
The present study was undertaken to establish the
taxonomic status of this strain. The polyphasic taxo-
nomic study showed that isolate 2AT belonged to a
new Streptomyces species, for which the name Strep-
tomyces amritsarensis sp. nov. is proposed.
Materials and methods
Microorganisms, maintenance and culture
conditions
Strain 2AT was isolated from a soil sample collected
from Guru Nanak Dev University, Punjab, India
(31.63�N, 74.87�E) using standard dilution plating
method. Soil sample was serially diluted up to 10-6
and 0.1 ml aliquot from each dilution was spread onto
starch casein nitrate agar supplemented with cyclo-
heximide (50 lg/ml) and nystatin (25 lg/ml, HiMe-
dia, India). The plates were incubated at 28 �C for
28 days (Sharma et al. 2011).
The isolate and the type strains of S. flavotricini
DSM 40152T, S. globosus DSM 40815T and S.
toxytricini DSM 40178T were maintained on modified
Bennett’s agar (Jones 1949) slopes at 4 �C and as
mycelial fragments and spores in 20 % v/v glycerol at
-80 �C.
Morphological, physiological and biochemical
characterization
Cultural characteristics were determined as per meth-
ods prescribed in International Streptomyces Project
(Shirling and Gottlieb 1966). The pigmentation of
aerial and substrate mycelia, soluble pigment and
sporulation which are highly characteristic and useful
in the classification of the genus Streptomyces were
observed by cultivating the strain on starch casein
nitrate agar, Bennett’s agar and different ISP media at
28 �C for 1 week. Morphological properties of the
strain were observed by light microscope (Olympus)
and spore surface was determined by scanning elec-
tron microscopy following desiccation of the cells and
gold coating, using a Carl Zeiss model EVOLS 10.
Physiological tests such as growth at different
temperatures (12–42 �C), pH (5.0–10.0) and NaCl
(1–7 %, w/v) concentrations were performed by
growing the strain on starch casein nitrate agar at
28 �C. Assimilation of sugars as carbon sources (1 %)
was studied according to Shirling and Gottlieb (1966).
D-Glucose, sucrose, xylose, inositol, D-mannitol, D-
fructose, L-rhaminose, D-raffinose and cellulose (1 %)
(HiMedia, India) were added to basal medium after
filter sterilization. Catalase production, citrate utiliza-
tion (Simmons) and urea hydrolysis were determined
according to Cowan and Steel (1965). Hydrolysis of
casein, gelatin and starch, indole production, MR-VP
and oxidase tests were performed as recommended by
Smibert and Krieg (1994). Nitrate reduction was
investigated according to Lanyi (1987).
Chemotaxonomic characterization
Freeze-dried cells for chemotaxonomic analysis
(except for fatty acid study) were prepared by
harvesting the mycelium in the late exponential phase
following growth in trypticase soy broth (TSB;
HiMedia, India) at 28 �C for 3 days. Analysis of
isomer of diaminopimelic acid in the cell wall and
sugars in the whole-cell hydrolysate was done accord-
ing to method given by Lechevalier and Lechevalier
(1980). Menaquinones were extracted and purified as
per method given by Saha et al. (2005). The purified
menaquinones were separated by reversed phase
HPLC (SCL-10AVP, Shimadzu) using the solvent
system of acetonitrile and isopropanol in a ratio of
65:35 with a flow rate of 1 ml/min and monitored at a
wavelength of 269 nm. For the determination of
cellular fatty acids, strain was grown on TSB medium
at 30 �C for 2 days; cellular fatty acids were extracted,
methylated and analysed using Gas Chromatography
according to the instructions of the Sherlock Microbial
Identification System (MIDI, USA Version 4.0; Ac-
tino library) as described previously (Sasser 1990;
Pandey et al. 2002). Extraction of polar lipids was
done based on the modified protocol of Bligh and Dyer
(1959). Two-dimensional TLC was run for identifica-
tion of polar lipids according to procedures described
by Komagata and Suzuki (1987). Lipid spots were
944 Antonie van Leeuwenhoek (2014) 105:943–949
123

detected using the following spray reagents: molyb-
datophosphoric acid (5 % w/v) in absolute ethanol,
molybdenum blue spray reagent (1.3 %, Sigma),
ninhydrin (0.2 % w/v) in acetone and anisaldehyde
reagent (Sigma) for detection of total lipids, phospho-
lipids, aminolipids and glycolipids, respectively.
Phylogenetic analysis and genomic relatedness
For 16S rRNA gene sequencing the genomic DNA
extraction and amplification was performed according
to the method described by Mayilraj et al. (2006).
Identification of phylogenetic neighbours and the
calculation of pairwise 16S rRNA gene sequence
similarities were achieved using the EzTaxon server
(http://eztaxon-e.ezbiocloud.net/; Kim et al. 2012).
Phylogenetic trees were constructed according to the
neighbour-joining and maximum-parsimony using
bootstrap values based on 1,000 replications with the
MEGA6 software (Tamura et al. 2013). The G?C
content of genomic DNA was determined spectro-
photometrically (Lambda 35, Perkin Elmer, Waltham,
MA, USA) using thermal denaturation method
(Mandel and Marmur 1968). DNA–DNA hybridiza-
tion was performed with freshly isolated genomic
DNA by the membrane filter method (Tourova and
Antonov 1987) and experiment was repeated twice.
Determination of antimicrobial activity
For determining antimicrobial activity, 2AT and type
strains of related Streptomyces species were grown in
starch casein nitrate broth at 28 �C on a rotary shaker
at 180 rpm. Fermentation was terminated on 4th day
and culture broths were centrifuged at 10,000 rpm for
30 min at 4 �C to separate the mycelium. Antimicro-
bial activity against various test organisms was
performed using agar well diffusion assay (Bauer
et al. 1996). The plates containing Mueller–Hinton
agar, yeast malt agar and potato dextrose agar were
seeded with test bacteria, yeasts and fungi, respec-
tively. Wells of 6 mm diameter were cut out from agar
plates, filled with supernatant (filtered through
0.22 lm filter, Pall Life Sciences) and inhibition
zones were recorded after 24–48 h of incubation.
Various test organisms used in the study included
Bacillus subtilis (MTCC 619), Mycobacterium
smegmatis (MTCC 6), Staphylococcus epidermidis
(MTCC 435), Escherichia coli (MTCC 1885),
Klebsiella pneumoniae sub sp. pneumoniae (MTCC
109), Enterobacter aerogenes (MTCC 111), Salmo-
nella typhi (MTCC 733), multi-drug resistant E. coli
(resistant to cephotaxime, ciprofloxacin, rifampicin
and cefoperazone), MRSA, VRE, Candida albicans
(MTCC 3017), Rhodotorula rubra (MTCC 248),
Colletotrichum acutatum (MTCC 1037), Cercospora
beticola (KJ461435), Fusarium oxysporum f.sp. dian-
thi (MTCC 6659), and Alternaria brassicicola (MTCC
2102).
Results and discussion
Isolate 2AT showed a range of chemotaxonomic and
morphological properties consistent with its classifi-
cation in the genus Streptomyces (Kampfer 2012).
Strain 2AT formed rectiflexibilis-type spore chains
with smooth spore surface ornamentation (Fig. 1,
Supplementary Fig. S1). The organism grew well on
most of the media, except ISP 5. No distinctive color
of substrate mycelium was recorded. The color of
aerial and substrate mycelia did not show much
variation on different media (Table 1). Strain 2AT did
not produce melanin pigment on tyrosine agar (ISP 7)
but produced blackish brown pigment on peptone
yeast iron agar (ISP 6). It hydrolyzed starch; but did
not hydrolyze casein, gelatin or urea. It degraded
esculin, did not produce indole from tryptophan and
gave negative results for MR-VP tests. It could neither
reduce nitrate nor utilize citrate. The strain produced
H2S and catalase and tolerated NaCl up to 2 %. Good
Fig. 1 Electron microscopic view of spores having smooth
surface
Antonie van Leeuwenhoek (2014) 105:943–949 945
123
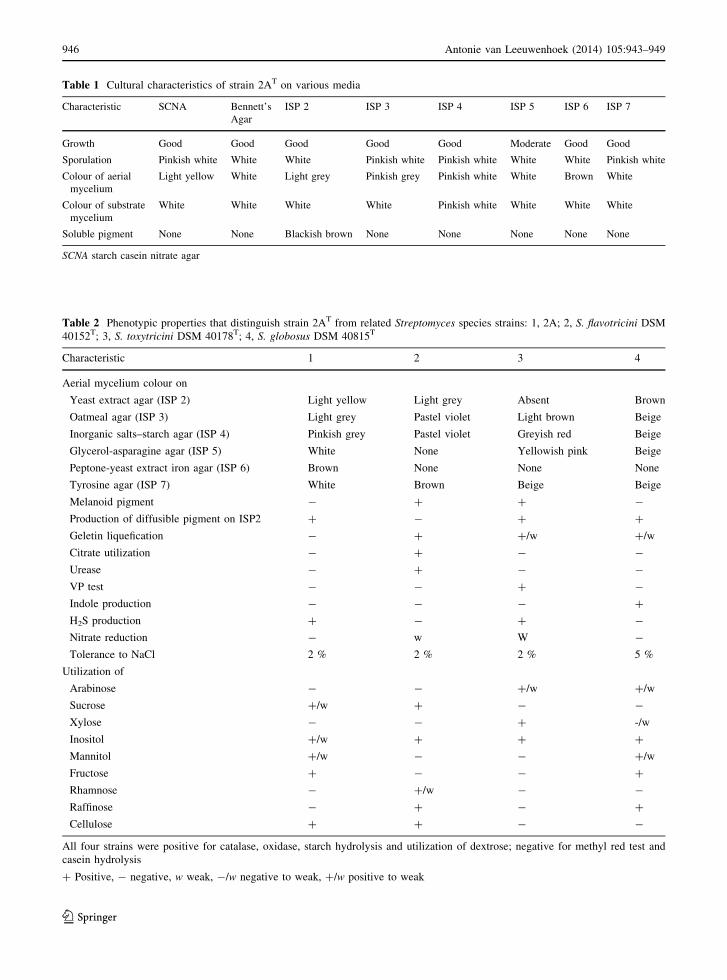
Table 1 Cultural characteristics of strain 2AT on various media
Characteristic SCNA Bennett’s
Agar
ISP 2 ISP 3 ISP 4 ISP 5 ISP 6 ISP 7
Growth Good Good Good Good Good Moderate Good Good
Sporulation Pinkish white White White Pinkish white Pinkish white White White Pinkish white
Colour of aerial
mycelium
Light yellow White Light grey Pinkish grey Pinkish white White Brown White
Colour of substrate
mycelium
White White White White Pinkish white White White White
Soluble pigment None None Blackish brown None None None None None
SCNA starch casein nitrate agar
Table 2 Phenotypic properties that distinguish strain 2AT from related Streptomyces species strains: 1, 2A; 2, S. flavotricini DSM
40152T; 3, S. toxytricini DSM 40178T; 4, S. globosus DSM 40815T
Characteristic 1 2 3 4
Aerial mycelium colour on
Yeast extract agar (ISP 2) Light yellow Light grey Absent Brown
Oatmeal agar (ISP 3) Light grey Pastel violet Light brown Beige
Inorganic salts–starch agar (ISP 4) Pinkish grey Pastel violet Greyish red Beige
Glycerol-asparagine agar (ISP 5) White None Yellowish pink Beige
Peptone-yeast extract iron agar (ISP 6) Brown None None None
Tyrosine agar (ISP 7) White Brown Beige Beige
Melanoid pigment - ? ? -
Production of diffusible pigment on ISP2 ? - ? ?
Geletin liquefication - ? ?/w ?/w
Citrate utilization - ? - -
Urease - ? - -
VP test - - ? -
Indole production - - - ?
H2S production ? - ? -
Nitrate reduction - w W -
Tolerance to NaCl 2 % 2 % 2 % 5 %
Utilization of
Arabinose - - ?/w ?/w
Sucrose ?/w ? - -
Xylose - - ? -/w
Inositol ?/w ? ? ?
Mannitol ?/w - - ?/w
Fructose ? - - ?
Rhamnose - ?/w - -
Raffinose - ? - ?
Cellulose ? ? - -
All four strains were positive for catalase, oxidase, starch hydrolysis and utilization of dextrose; negative for methyl red test and
casein hydrolysis
? Positive, - negative, w weak, -/w negative to weak, ?/w positive to weak
946 Antonie van Leeuwenhoek (2014) 105:943–949
123

growth was recorded at temperature range of
15–37 �C and at pH range of 6.0–10.0. It utilized the
following sugars: D-glucose and fructose as sole
carbon sources, but did not utilize L-arabinose, xylose,
L-rhamnose, D-raffinose and cellulose (Table 2).
Strain 2AT had cell wall chemotype I which is
characteristic of the Streptomyces genus. The cell wall
contained LL-diaminopimelic acid and glycine.
Whole cell hydrolysates contained no characteristic
sugar. The menaquinones detected were MK-9(H6)
(58.2 %), MK-9(H8) (28.3 %) and MK-9(H10)
(13.5 %). The major phospholipids detected were
phosphatidylethanolamine, diphosphatidylglycerol,
phosphatidylglycerol (phosholipid type II). In addi-
tion, five unknown phospholipids and one unknown
glyco lipid were also present (Supplementary Fig. S2).
The major fatty acids found were anteiso-C15:0
(29.2 %), C16: 0 (16.0 %) and iso-C16: 0 (12.1 %).
The DNA G?C content of strain was estimated to be
70.5 ± 1.0 mol%.
Almost complete sequence (1463 bp) of 16S rRNA
gene of strain 2AT was determined (GenBank accession
no GQ906975) and compared with those of other closely
related taxa retrieved from the EzTaxon database.
Phylogenetic trees were constructed using neighbour-
joining (Fig. 2) and maximum-parsimony methods
(Supplementary Fig. S3). No significant differences
were observed in topologies generated by both neigh-
bour-joining and maximum-parsimony analyses. The
analyzed strain 2AT formed a distinct cluster in both
phylogenetic trees. However, low bootstrap values
(\50 %) do not reliably indicate the specific relatedness
between the species. The taxonomic integrity of the
isolate was also supported by the DNA–DNA related-
ness data as it shared DNA–DNA homology of
37.6 ± 0.6 %, 34.4 ± 0.5 % and 33.1 ± 0.4 % with
Table 3 Antimicrobial activity of strain 2AT and related Streptomyces species strains: 1, 2A; 2, S. flavotricini DSM 40152T; 3, S.
toxytricini DSM 40178T; 4, S. globosus DSM 40815T
Test organisms Reference number Inhibition zone (in mm)
1 2 3 4
Gram-positive bacteria
Bacillus subtilis MTCC 619 20 ± 0.1 - 11 ± 0.5 20 ± 0.1
Mycobacterium smegmatis MTCC 6 18 ± 0.5 - - 20 ± 0.5
Staphylococcus epidermidis MTCC 435 16 ± 0.5 - - 15 ± 0.5
Gram-negative bacteria
Escherichia coli MTCC1885 14 ± 0.5 - - -
Klebsiella pneumoniae sub sp. pneumoniae MTCC 109 10 ± 0.5 - - -
Enterobacter aerogenes MTCC 111 14 ± 0.5 - - -
Salmonella typhi MTCC 733 16 ± 0.5 - - -
Clinical isolates
MRSA 18 ± 0.1 - - 14 ± 0.5
E. coli 12 ± 0.5 - - -
VRE 11 ± 0.5 - - -
Yeasts
Candida albicans MTCC 3017 11 ± 0.5 - - -
Rhodotorula rubra MTCC 248 15 ± 0.5 -/w 12 ± 0.5 15 ± 0.5
Fungus
Colletotrichum acutatum MTCC 1037 12 ± 0.5 -/w 13 ± 0.5 ?/w
Cercospora beticola KJ461435 16 ± 0.5 -/w 12 ± 0.5 11 ± 0.5
Alternaria brassicicola MTCC 2102 16 ± 0.5 -/w 16 ± 0.5 ?/w
Fusarium oxysporum f.sp. dianthi MTCC 6659 11 ± 0.5 -/w 10 ± 0.0 10 ± 0.0
MTCC-Microbial Type Culture Collection, IMECH, Chandigarh (India)
? Positive, - negative, w weak, -/w negative to weak, ?/w positive to weak
Antonie van Leeuwenhoek (2014) 105:943–949 947
123

the type strains, S. flavotricini DSM 40152T, S. globosus
DSM 40815T and S. toxytricini DSM 40178T, respec-
tively. These values are clearly well below the 70 % cut-
off point recommended for the assignment of strains to
the same genomic species (Wayne et al. 1987).
Strain 2AT demonstrated potent antimicrobial
activity against all the test microorganisms whereas
type strains of related Streptomyces species showed
weak activity (Table 3). The difference in antimicro-
bial activity further distinguishes 2AT from type
strains of related Streptomyces species.
Description of Streptomyces amritsarensis sp. nov
Streptomyces amritsarensis (am.rit.sar.en’sis. N.L.
masc. adj. amritsarensis, pertaining to Amritsar, a
holy city of Punjab state in India, where the type strain
was isolated).
Aerobic, Gram-positive, non-motile actinobacte-
rium which forms an extensively branched substrate
mycelium that bears aerial hyphae which carry
rectiflexibilis-type spore chains with smooth
surfaced spores (0.5 9 0.9 lm). Growth occurs from
15 to 37 �C, between pH 6–10 and can tolerate salt
concentration up to 2 % (w/v). Blackish brown
pigment is formed on peptone-yeast extract-iron
agar. Hydrolyses starch, but not urea or casein.
Gelatin is liquefied. Dextrose and fructose are used
as sole carbon sources for energy (at 1 %, w/v).
Additional phenotypic properties are cited in the text
and in Tables 1 and 2. Chemotaxonomic properties
are typical of the genus Streptomyces. Whole-cell
hydrolysate does not contain any characteristic sugar
and cell wall contains LL-diaminopimelic acid and
glycine. Major menaquinones are MK-9(H6), MK-
9(H8) and MK-9(H10). The phospholipid pattern is
PII type containing phosphatidylethanolamine (PE),
diphosphatidylglycerol (DPG), phosphatidylglycerol
(PG), five unknown phospholipids and one unknown
glyco lipid. The three most abundant fatty acids are
anteiso-C15: 0, C16: 0 and iso-C16: 0 and G?C content
of the genomic DNA of the type strain is
70.5 ± 1.0 mol%. Produces broad spectrum antimi-
crobial compounds active against various Gram-
S. sporoverrucosus NBRC 15458T (AB184684)
S .nojiriensis LMG 20094T (AJ781355)
S. xanthophaeus NBRC 12829T (AB184177)
S. vinaceus NBRC 13425T (AB184394)
S. cirratus NRRL B-3250T (AY999794)
S. lavendulae NBRC 12789T (AB184146)
S. spororaveus LMG 20313T (AJ781370)
S. colombiensis NRRL B-1990T (DQ026646)
S. avidinii NBRC 13429T (AB184395)
S. subrutilus DSM 40445T(X80825)
S. cinnamonensis NBRC 15873T (AB184707)
S. virginiae NBRC 12827T (AB184175)
S. toxytricini DSM 40178 T (AB184173)
S. globosus DSM 40815 T (AJ781330)
S. flavotricini DSM 40152 T (AB184132)
Strain 2AT (GQ906975)
S. katrae NBRC 13447T (AB184409)
S. racemochromogenes NRRL B-5430T (DQ026656)
S. polychromogenes NBRC 13072T (AB184292)
S. tanashiensis LMG 20274T (AJ781362)
S. nashvillensis NBRC 13064T (AB184286)
S. rubiginosohelvolus IFO 12912T (AY999864)40
100
100
60
64
46
94
46
98
99
87
80
62
6191
36
41
37
0.001
Fig. 2 Neighbour-joining tree based on 16S rRNA (1463 bp)
sequences, showing the phylogenetic relationship between strain
2AT and the related type strains obtained from the database of
type strains with validly published prokaryotic names at
EzTaxon-e identification service. Bootstrap values (expressed
as percentage of 1,000 replications) are given at the nodes
948 Antonie van Leeuwenhoek (2014) 105:943–949
123

positive and Gram-negative bacteria, drug resistant
bacteria and fungi.
The type strain, 2AT (=MTCC 11845T=JCM
19660T) was isolated from a soil sample collected
from Guru Nanak Dev University, Amritsar (India).
The species description is based on a single strain and
hence serves as a description of the type strain.
Acknowledgments The authors are thankful to Department of
Biotechnology, New Delhi, India for providing the financial
assistance.
References
Antony-Babu S, Goodfellow M (2008) Biosystematics of al-
kaliphilic streptomycetes isolated from seven locations
across a beach and dune sand system. Antonie Van Leeu-
wenhoek 94:581–591
Bauer AW, Kirby WMM, Sherris JC, Turck M (1996) Antibiotic
susceptibility testing by a standardized single disk method.
Am J Clin Pathol 45:493–496
Bligh EG, Dyer WJ (1959) A rapid method of total lipid
extraction and purification. Can J Biochem Physiol
37:911–917
Cowan ST, Steel KJ (1965) Manual for the identification of
medical bacteria. Cambridge University Press, London
Euzeby JP (2012) List of bacterial names with standing in
nomenclature: a folder available on the Internet (Last full
update: 13 September 2013). http://www.bacterio.cict.fr/s/
streptomycesa.html
Jones KL (1949) Fresh isolates of actinomycetes in which the
presence of sporogenous aerial mycelia is a fluctuating
characteristic. J Bacteriol 57:141–145
Kampfer P (2012) Genus I. Streptomyces Waksman and Henrici
1943, 339AL emend. Witt and Stackebrandt 1990, 370,
emend. Wellington, Stackebrandt, Sanders, Wolstrup and
Jorgensen 1992, 159. In: Goodfellow M, Kampfer P, Busse
H-J, Trujillo ME, Suzuki K-I, Ludwig W, Whitman WB
(eds) Bergey’s manual of systematic bacteriology, Part B,
vol 5, 2nd edn. Springer, New York, pp 1455–1517
Kim OS, Cho YJ, Lee K, Yoon SH, Kim M, Na H, Park SC, Jeon
YS, Lee JH, Yi H, Won S, Chun J (2012) Introducing
EzTaxon-e: a prokaryotic 16S rRNA Gene sequence
database with phylotypes that represent uncultured species.
Int J Syst Evol Microbiol 62:716–721
Komagata K, Suzuki K (1987) Lipid and cell-wall analysis in
bacterial systematics. Methods Microbiol 19:161–207
Lanyi B (1987) Classical and rapid identification methods for
medically important bacteria. Methods Microbiol 19:1–67
Lechevalier MPA, Lechevalier HA (1980) The chemotaxonomy
of actinomycetes. In: Dietz A, Thayer DW (eds) Actino-
mycete taxonomy. Society for Industrial Microbiology,
Arlington, pp 22–291
Mandel M, Marmur J (1968) Use of ultraviolet absorbance
temperature profile for determining the guanine plus
cytosine content of DNA. Methods Enzymol 12B:195–206
Mayilraj S, Kroppenstedt RM, Suresh K, Saini HS (2006)
Kocuria himachalensis sp. nov., an actinobacterium iso-
lated from the Indian Himalayas. Int J Syst Evol Microbiol
56:1971–1975
Pandey KK, Mayilraj S, Chakrabarti T (2002) Pseudomonas
indica sp. nov., a novel butane-utilizing species. Int J Syst
Evol Microbiol 52:1559–1567
Saha P, Mondal AK, Mayilraj S, Krishnamurthi S, Bhattacharya
A, Chakrabarti T (2005) Paenibacillus assamensis sp. nov.,
a novel bacterium isolated from a warm spring in Assam,
India. Int J Syst Evol Microbiol 55:2577–2581
Sasser M (1990) Identification of bacteria by gas chromatog-
raphy of cellular fatty acids, MIDI Technical Note 101.
MIDI Inc, Newark
Sharma D, Kaur T, Chadha BS, Manhas RK (2011) Antimi-
crobial activity of actinomycetes against multidrug resis-
tant Staphylococcus aureus, E. coli and various other
pathogens. Trop J Pharm Res 10:801–808
Shirling EB, Gottlieb D (1966) Methods for characterization of
Streptomyces species. Int J Syst Bacteriol 16:313–340
Smibert RM, Krieg NR (1994) Phenotypic characterization. In:
Gerhardt P, Murray RGE, Wood WA, Krieg NR (eds)
Methods for general and molecular bacteriology. American
Society of Microbiology, Washington, pp 607–654
Tamura K, Stecher G, Peterson D, Filipski A, Kumar S (2013)
MEGA6: molecular evolutionary genetics analysis version
6.0. Mol Biol Evol 30:2725–2729
Tourova TP, Antonov AS (1987) Identification of microorgan-
isms by rapid DNA–DNA hybridization. Methods Micro-
biol 19:333–355
Waksman SA, Henrici AT (1943) The nomenclature and clas-
sification of the actinomycetes. J Bacteriol 46:337–341
Wayne LG, Brenner DJ, Colwell RR, Grimont PAD, Kandler O,
Krichevsky MI, Moore LH, Moore WEC, Murray R-G-E
et al (1987) International committee on systematic bacte-
riology. Report of the ad hoc committee on reconciliation
of approaches to bacterial systematics. Int J Syst Bacteriol
37:463–464
Antonie van Leeuwenhoek (2014) 105:943–949 949
123