streptomyces tritolerans sp. nov., a novel actinomycete isolated from soil in karnataka, india
TRANSCRIPT

ORIGINAL PAPER
Streptomyces tritolerans sp. nov., a novel actinomyceteisolated from soil in Karnataka, India
Dastager G. Syed Æ Dayanand Agasar Æ Chang-Jin Kim Æ Wen-Jun Li ÆJae-Chan Lee Æ Dong-Jin Park Æ Li-Hua Xu Æ Xin-Peng Tian ÆCheng-Lin Jiang
Received: 10 January 2007 / Accepted: 12 March 2007 / Published online: 9 June 2007� Springer Science+Business Media B.V. 2007
Abstract A Gram-positive, nonmotile, moder-
ately halophilic, alkali and thermotolerant strain
designated DAS 165T, was isolated from a dry
land soil sample from the Gulbarga region,
Karnataka province, India. The isolate produced
yellow substrate mycelia and gray aerial mycelia
on most tested media. Strain DAS 165T showed
growth in the presence of 5 to 7% NaCl and at
45�C. The DNA G + C content was 69.7%. 16S
rRNA gene sequence analysis together with these
characteristics consistently assigned strain DAS
165T to the genus Streptomyces. The 16S rRNA
gene sequence analysis revealed that strain DAS
165T was most closely related to S. tendae ATCC
19812T (D 63873) with a sequence similarity of
99.6% (three nucleotide differences out of 1,517).
Strain DAS 165T formed a distinct clade based on
analysis of the almost complete sequence and
120-nucleotide variable c region of the 16S rRNA
gene. Despite the high sequence similarity, strain
DAS 165T was phenotypically different from
S. tendae ATCC 19812T. DNA–DNA hybridi-
zation between these strains was 47% showing
that strain DAS 165T is a distinct genomic species.
Phenetic and genetic results support the classifi-
cation of strain DAS 165T as a new species, for
which the name S. tritolerans is proposed, with
strain DAS 165T as the type strain (=DSM
41899T = CCTCCAA 206013T).
Keywords Streptomyces tritolerans sp. nov �Polyphasic taxonomy � 16S rRNA
Introduction
Streptomyces systematics has become increasingly
objective due to the application of chemotaxo-
nomic, molecular systematic and numerical taxo-
nomic methods (Kim et al.1999). The species
concept is still a difficult issue in Streptomyces
systematics due to the taxonomic complexity of
the genus and the absence of accepted minimal
D. G. Syed � C.-J. Kim � J.-C. Lee � D.-J. ParkKorea Research Institute of Bioscience andBiotechnology (KRIBB), Daejeon 305-333, SouthKorea
W.-J. Li � L.-H. Xu � X.-P. Tian � C.-L. JiangThe Key Laboratory for Microbial Resources ofMinistry of Education, Yunnan Institute ofMicrobiology, Yunnan University, Kunming, Yunnan650 091, P.R. China
D. G. Syed � D. Agasar (&)Department of Studies and Research in Microbiology,Gulbarga University, Gulbarga 585 106, Indiae-mail: [email protected]
X.-P. TianGuangdong Key Laboratory of Marine MateriaMedica, South China Sea Institute of Oceanology,Chinese Academy of Sciences, Guangzhou 510301,P.R. China
123
Antonie van Leeuwenhoek (2007) 92:391–397
DOI 10.1007/s10482-007-9166-2

standards for the delineation of species. However,
it is evident that Streptomyces species should be
circumscribed using judicious combinations of
genotypic and phenotypic data (Labeda 1993;
Labeda and Lyons 1991a, b; Goodfellow et al.
1992). Comparative taxonomic studies are needed
to find rapid and reliable phenotypic methods for
delineating potentially novel species of Strepto-
myces. Alkalitolerant actinomycetes that thrive in
alkaline environments have specific nutrient
requirements, cultural conditions and physiolog-
ical properties (Collins 1985; Lechevalier and
Lechevalier 1980). There have been previous
reports on the physiology and energetics of
alkalitolerant bacteria (Kim et al. 1999). Strain
DAS 165T was recovered and characterized by a
combination of genotypic and phenotypic meth-
ods during an investigation of the extremophilic
microbial flora of the Gulbarga region, India. It
was evident that the actinomycete isolate was
sufficiently distinct from the closely related spe-
cies of Streptomyces to warrant the description of
a novel species, for which the name Streptomyces
tritolerans sp. nov., is proposed.
Material and methods
Strain DAS 165T was isolated using a standard
serial dilution plating method, from an alkaline
soil sample (pH 9–10) collected from Gulbarga,
northern Karnataka, southern India. The isola-
tion medium used was starch casein agar and
cultures were incubated at 28�C for about
2 weeks. The strain was maintained on ISP 2
[International Streptomyces project (ISP; Shirling
and Gottlieb 1966)] and ISP 5 agar slants at 4�C,
and as glycerol suspensions (20% v/v) at -20�C.
Biomass for chemical and molecular systematic
studies was obtained by growing in shake flasks in
ISP 2 broth (28�C, 1 week, 150 rpm).
Cultural characteristics were determined after
2 weeks growth at 28�C as per the methods
prescribed in the ISP (Shirling and Gottlieb
1966). Morphological properties were examined
by light microscopy (Olympus microscope) and
scanning electron microscopy following dessica-
tion of the cells and gold coating, using a JEOL
model JSM 5600LV. A range of phenotypic
properties was examined using standard proce-
dures (Williams et al. 1983). In addition, acid
production from carbohydrates was carried out
using the media and methods described by Shir-
ling and Gottlieb (1966). The utilization of sole
carbon and sole nitrogen sources was investigated
according to Shirling and Gottlieb (1966) and
Tsukamura (1966). Tolerance to temperature (10,
27, 37 and 45�C) and sodium chloride (0, 1, 3, 5, 7
and 10%) were tested using modified Bennett’s
agar (Williams et al. 1983). Resistance to antibi-
otics was examined using novobiocin (30 lg/ml),
ampicillin (10 lg/ml), amoxicillin (10 lg/ml), nali-
dixic acid (30 lg/ml), penicillin G (10 lg/ml),
norfloxacin (10 lg/ml), rifampicin (5 lg/ml) and
carbenicillin (100 lg/ml) incorporated into glu-
cose yeast extract agar (Lechevalier and Leche-
valier 1970a, b) as a basal medium. The results
were recorded following incubation at 28�C up to
14 days. Color determination was done with color
chips from the ISCC-NBS COLOR CHARTS
standard sample No. 2106 (Kelly 1964).
Chemotaxonomy
The cell wall fraction of strain DAS 165T was
purified and analyzed by the method of Lecheva-
lier and Lechevalier (1980). The procedure of
Becker et al. (1965) and Lechevalier and Leche-
valier (1980) were used to determine the whole-
cell composition. Menaquinones were determined
according to the Collins (1985) method. Biomass
for quantitative fatty acid analysis was prepared
by scraping growth from trypticase soy broth agar
plates that had been incubated for 3 days at 28�C.
The fatty acids were extracted, methylated and
analyzed using the standard MIDI (Microbial
identification) system (Kampfer and Kroppen-
stedt 1996; Sasser 1990).
16S rRNA gene sequence
The genomic DNA of strain DAS 165T was
isolated as described by Hoopwood et al. (1985).
The 16S rRNA gene was amplified by PCR, using
procedures described by Xu et al. (2003). Multi-
ple alignments with a selection of 16S rRNA gene
sequences were used and calculation of levels
of sequence similarity were carried out using
392 Antonie van Leeuwenhoek (2007) 92:391–397
123

CLUSTAL X (Thompson et al.1997). A phylo-
genetic tree was reconstructed using the neigh-
bor-joining method of Saitou and Nei (1987) from
Knuc values (Kimura 1980,1983). The topology of
the phylogenetic tree was evaluated by the
bootstrap resampling method of Felsentein
(1985) with 1,000 replicates. S. almquistii NBRC
13015T (AB184258) was used as an out group.
DNA for determining the G + C content was
obtained by the method of Marmur (1962). The
DNA G + C content was determined by the
thermal denaturation method (Mandel and Mar-
mur 1968). DNA–DNA hybridization was carried
out by applying the optical renaturation method
(De Ley et al. 1970; Huss et al. 1983; Jahnke
1992) under optimal hybridization conditions,
using Escherichia coli as control.
Nucleotide sequence accession number
The 16S rRNA gene sequence of strain DAS 165T
determined in this study has been deposited in
GenBank under the accession number DQ
345779.
Results and discussion
Strain DAS 165T was Gram-positive. The isolate
developed well on several media including ISP 2,
ISP 3, ISP 4, ISP 5, ISP 7, starch casein agar,
czapek’s agar and glucose asparagine agar. Mor-
phological observation of a 14-day-old culture of
strain DAS 165T on ISP 2 showed good growth of
aerial and vegetative hyphae which were well
developed and did not exhibit fragmentation.
Smooth, oval-rod shaped spores were borne in
long straight to flexous chains (Fig. 1). DAS 165T
developed gray to yellow aerial hyphae on all
media tested. The substrate mycelium differed
from light yellow to dark yellow. Yellow colored
substrate mycelium was produced on ISP2, ISP3,
ISP4, ISP5 and Czapek’s agar; darkish yellow
substrate mycelium was produced on starch case-
in and glucose asparagine agar; greenish yellow
substrate mycelium was produced on tyrosine
agar. No diffusible pigments were produced. The
detailed physiological and biochemical properties
are given in Table 1 and the species description.
Growth of strain DAS 165T was observed at a
wide range of temperature (28–45�C), at pH 7–10
and in the presence of 7% NaCl. Whole organism
hydrolysates of strain DAS 165T were rich in
LL-diaminopimelic acid (DAP) and glycine. No
diagnostic sugars were found in the cell wall
constituents. The diagnostic phospholipid is
phosphatidylethanolamine. The predominant
menaquinones were MK-9 (H4)-8%, MK-9 (H6)-
78% and MK-9 (H8)-14%. The major cellular
fatty acids were i-C15:0(13.6%), ai-C15:0 (17.8%),
C16:0(9.6%) and i-C16:0 (28.1%).
An almost complete 16S rRNA gene sequence
of strain DAS 165T (1,517 nucleotides) was
determined. BLAST search results of the strain
DAS 165T sequence against the non-redundant
GenBank database was used to select sequence as
reference sequences. Comparison of this
sequence with those of related organisms showed
clearly that it contained all the signature nucle-
otides designated for the genus Streptomyces
(Christova et al. 1995). The value for the 16S
rRNA gene sequence similarity to closely related
members of the genus Streptomyces were 99.4–
99.7% (three nucleotide differences out of 1,517).
The phylogenetic tree (Fig. 2) from representa-
tive strains of the related species indicated that
strain DAS 165T should be placed in the genus
Streptomyces and forms a separate lineage to-
gether with its closest neighbour S. tendae (ATCC
19812T) with 99.6% 16S rRNA gene sequence
similarity. Despite this very high sequence simi-
larity, strain DAS 165T was phenetically distinct
from S. tendae ATCC 19812T (Table 1).
The DNA G + C content of strain DAS 165T
was 69.7%. DNA–DNA hybridization between
strains DAS 165T and S. tendae ATCC 19812T
was carried out and the determined DNA relat-
edness value of 47% was significantly lower than
70%, which is considered to be the threshold
value for the delineation of genomic species
(Wayne et al. 1987).
Description of Streptomyces tritolerans
Streptomyces tritolerans (tri’to.le.rans.L.pref. tri-
ple. three. L. pres. part. tolerans. tolerating; N.L.
part. adj. tritolerans referring to the ability of the
Antonie van Leeuwenhoek (2007) 92:391–397 393
123
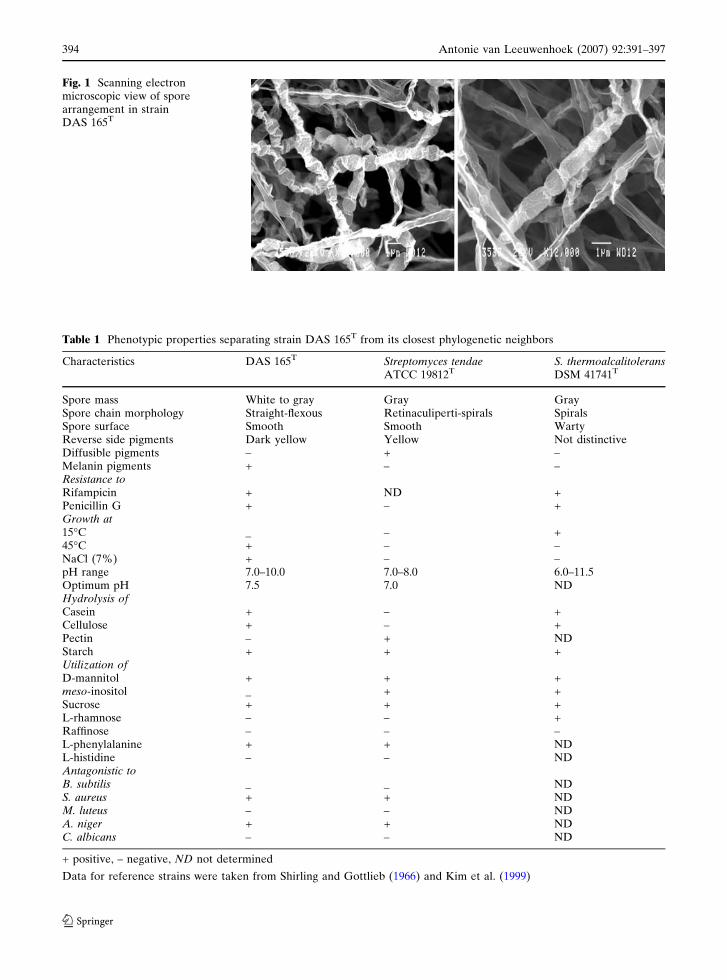
Table 1 Phenotypic properties separating strain DAS 165T from its closest phylogenetic neighbors
Characteristics DAS 165T Streptomyces tendaeATCC 19812T
S. thermoalcalitoleransDSM 41741T
Spore mass White to gray Gray GraySpore chain morphology Straight-flexous Retinaculiperti-spirals SpiralsSpore surface Smooth Smooth WartyReverse side pigments Dark yellow Yellow Not distinctiveDiffusible pigments – + –Melanin pigments + – –Resistance toRifampicin + ND +Penicillin G + – +Growth at15�C _ – +45�C + – –NaCl (7%) + – –pH range 7.0–10.0 7.0–8.0 6.0–11.5Optimum pH 7.5 7.0 NDHydrolysis ofCasein + – +Cellulose + – +Pectin – + NDStarch + + +Utilization ofD-mannitol + + +meso-inositol _ + +Sucrose + + +L-rhamnose – – +Raffinose – – –L-phenylalanine + + NDL-histidine – – NDAntagonistic toB. subtilis _ _ NDS. aureus + + NDM. luteus – – NDA. niger + + NDC. albicans – – ND
+ positive, – negative, ND not determined
Data for reference strains were taken from Shirling and Gottlieb (1966) and Kim et al. (1999)
Fig. 1 Scanning electronmicroscopic view of sporearrangement in strainDAS 165T
394 Antonie van Leeuwenhoek (2007) 92:391–397
123

organism to tolerate high salinity, alkalinity and
temperature).
The strain is Gram-positive, non motile, alka-
litolerant and thermotolerant with good growth
without fragmentation. Aerial mycelium is white
to gray. Diffusible pigments are not formed.
Spores are smooth, 0.6–1.0 lm in diameter
arranged in straight to flexous chains of 10–15
spores. Arabinose, fructose, galactose, glucose,
D-mannitol, rhamnose, raffinose, sucrose and
xylose are utilized as sole carbon and energy
sources but not inositol. Acid is not formed from
these carbon sources. L-asparagine, L-alanine,
L-cysteine (weak), L-histidine and L-phenylala-
nine are used as sole nitrogen sources, but not
L-arginine. Starch, casein and cellulose are
hydrolyzed. Urea is not hydrolyzed. Nitrate
reduction and H2S production are negative.
Tests for gelatin hydrolysis and melanin produc-
tion are positive. Growth occurs from 28 to
45�C, pH 7–10, and in presence of sodium
chloride (7% w/v). Resistance to novobiocin,
ampicillin, amoxycillin, nalidixic acid, penicillin
G, norfloxacin, rifampicin and carbencillin but
sensitive to ciprofloxacin, gentamicin, neomycin,
erythromycin, streptomycin, tetracycline, chl-
oramphenicol and vancomycin. The cell wall of
strain DAS 165T contains LL-DAP, but no
diagnostic sugars. MK-9 (H6)-78% is the major
menaquinone and minor amount of MK-9 (H4)-
DAS 16598
Streptomyces tendae ATCC 19812T (D63873)67
Streptomyces (coelicolor )violaceoruber DSM 41007T(X60514)Streptomyces pseudogriseolus NRRL 3985T(X80827)
Streptomyces ambofaciens ATCC 23877T(M27245) Streptomyces caelestis NRRL 2418T(X80824)64
Streptomyces glaucescens DSM40716T(X79322) Streptomyces thermocarboxydus DSM 44293T (U94490)
Streptomyces eurythermus ATCC 14975T (D63870)Streptomyces sampsonii ATCC 25495T (D63871)72
Streptomyces diastaticus subsp. Disataticus DSM 40496T (X53161)Streptomyces brasiliensis DSM 43159T (X53162)
Streptomyces bluensis DSM 40564T (X79324)Streptomyces espinosus NRRL 5729T (X80826)
Streptomyces thermocarboxydovorans DSM 44296T(U94489)Streptomyces. thermonitrificans DSM 40579T (Z68 98)0
Streptomycs thermovulgaris DSM 40444T (Z68094)100
100
Streptomyces thermoalkalitolerans DSM 41741T(AJ000284) Streptomyces. thermodiastaticus DSM 40573T (Z68101)7459
Streptomyces thermolineatus DSM 41451T (Z68097)Streptomyces thermoviolaceus subsp. apingens DSM 41392T (Z68095)
Streptomyces thermoviolaceus subsp. thermoviolaceus DSM 40443 (Z68096)Streptomyces mashuensis DSM 40221T (X79323)
Streptomyces rimosus DSM 40260T (X62884)Streptomyces albus DSM 40313T (X53163)
Streptoverticillium baldacii DSM 40039T (X53164)88
Streptoverticillium olivoreticuli subsp. cellulophilum DPDU 0278T (X53166)Streptoverticillium cinnamoneum subsp.cinnamoneus DPDU 0093T(X53171)Streptoverticillium salmonis DPDU 0098T(X53169)
Streptomyces mobaraensis DSM 40587T(X53167)Streptomyces griseocarneus DSM 40004T(X99943)
Streptoverticillium abikoense DSM 40831T (X53168)50
Streptomyces galbus DSM 40089T(X79852)61
Streptomyces lincolnensis NRRL 2936T (X79854)Streptomyces acidiscabies ATCC 49003T(D63865)
Streptomyces scabies ATCC 49173T (D63862)Streptomyces neyagawaensis ATCC 27449T (D63869)53
Streptomyces bottropensis ATCC 25435T(D63868)97
Streptomyces diastatochromogenes ATCC 12309T(D 3867)6Streptomyces seoulensis IMSNU 21266T(Z71365)
Streptomyces bikiniensis DSM 40581T(X79851)Streptomyces purpureus DSM 43460T(X53170)
Streptomyces griseus KCTC 9080T(X61478)100
Streptomyces setonii ATCC 25497T(D63872)Streptomyces subrutilus DSM 40445T(X80825)70
Streptomyces virginiae IFO 3729T(D85119)97
Streptomyces thermolineatus DSM 41451T(Z68097)Streptomyces macrosporus DSM 41449T(Z68099)84
Streptomyces megasporus DSM 41476T(Z68100)98
0.005
Fig. 2 Phylogeneticdendrogram obtained bydistance matrix analysis of16S rRNA gene sequence,showing the position ofstrain DAS 165T amongits phylogeneticneighbors. Numbers atthe branch nodes are bootstrap values, expressed asa percentage of 1,000replicates (only valuesabove 50% are shown).Bar 0.005 substitutionsper nucleotide
Antonie van Leeuwenhoek (2007) 92:391–397 395
123

8% and MK-9(H8)-14% are also present. The
diagnostic phospholipid is phosphatidylethanol-
amine. The major cellular fatty acids are
isoC15:0(14%), anteisoC15:0(18%), C16:0(10%)
and isoC16:0(28%). The G + C content of the
genomic DNA is 69.7 mol%. The strain DAS
165T was isolated from alkaline soil in Gulbarga,
Karnataka province, India.
Acknowledgments This work was supported by the 21CFrontier Microbial Genomics and Application Centreprogram. Ministry of Science and Technology, Republicof Korea. And Wen-Jun Li was supported by Program forNew Century Excellent Talent in University
References
Becker B, Lechevalier MP, Lechevalier HA (1965)Chemical composition of cell-wall preparations fromstrains of various form-genera of aerobic actinomy-cetes. Appl Microbiol 13:236–243
Christova K, Sholeva Z, Chipeva V (1995) Application ofmolecular biological methods in taxonomy of thegenus Streptomyces. J Culture Coll 1:3–10
Collins MD (1985) Isoprenoid quinine analysis in classifi-cation and identification. In: Goodfellow M, MinnikinDE (eds) Chemical methods in bacterial systematics.Academic, London, pp 267–287
De Ley J, Cattoir H, Reynaerts A (1970) The quantitativemeasurement of DNA hybridization from renatur-ation rates. Eur J Biochem 12:133–142
Felsentein J (1985) Conference limits on phylogenies: anapproach using the bootstrap. Evolution 39:783–789
Goodfellow M, Ferguson EV, Sanglier JJ (1992) Numeri-cal classification and identification of Streptomycesspecies-a review. Gene 115: 225–233
Hoopwood DA, Bill MJ, Charter KF, Kieser T, Bruton CJ,Kieser HM, Lydiate DJ, Smith CP, Ward JM, Sch-rempf H (1985) Genetic manipulation of Streptomy-ces: a laboratory manual. John. Innes Foundation,Norwich
Huss VAR, Festl H, Schlefier KH (1983) Studies on thespectrophotometric determination of DNA hybrid-ization from renaturation rates. Syst Appl Microbiol4:184–192
Jahnke KD (1992) BASIC computer program for evalua-tion of spectroscopic DNA renaturation data fromGILFORD SYSTEM 2600 spectrophotometer on aPC/XT/AT type personal computer. J MicrobiolMethods 15:61–73
Kampfer P, Kroppenstedt RM (1996) Numerical analysisof fatty acid patterns of coryneform bacteria and re-lated taxa. Can J Microbiol 42:989–1005
Kelly KL (1964) Inter-society color council-national bu-reau of standards color-name charts illustrated withcentriod colors. US Government Printing Office,Washington
Kim B, Sahin N, Minnikin DE, Zakrzewska-Czerwinska J,Mordarski M, Goodfellow M (1999) Classification ofthermophilic actinomycetes, including the descriptionof Streptomyces thermoalcalitolerans sp. nov. Int JSyst Bacteriol 49:7–17
Kimura M (1980) A simple method for estimating evolu-tionary rates of base substitutions through compara-tive studies of nucleotide sequence. J Mol Evol16:111–120
Kimura M (1983) The neutral theory of molecular evolu-tion. Cambridge University Press, Cambridge
Labeda DP (1993) DNA relatedness among strains of theStreptomyces lavendulae phenotypic cluster group. IntJ Syst Bacteriol 43:822–825
Labeda DP, Lyons AJ (1991a) Deoxyribonucleic acidrelatedness among species of the Streptomycs cyaneuscluster. Syst Appl Microbiol 14:158–164
Labeda DP, Lyons AJ (1991b) The Streptomyces viola-ceusniger cluster is heterogeneous in DNA related-ness among strains: emendation of the description ofS. violaceusniger and Streptomyces hygroscopicus. IntJ Syst Bacteriol 41:398–401
Lechevalier HA, Lechevalier MPA (1970a) Critical eval-uation of genera of aerobic Actinomycetes. In: Pra-suer H (eds) The actinomycetes. Glustal Fischer,Verlag, Jena, pp 393–405
Lechevalier MPA, Lechevalier HA (1970b) Chemicalcomposition as a criterion in the classification of aer-obic actinomycetes. Int J Syst Bacteriol 20:435–443
Lechevalier MPA, Lechevalier HA (1980) The chemo-taxonomy of actinomycetes. In: Dietz A, Thayer DW(eds) Actinomycete taxonomy. Society for IndustrialMicrobiology, Arlington, pp 22–291
Mandel M, Marmur J (1968) Use of ultraviolet absorbancetemperature profile for determining the gunine pluscytosine content of DNA. Methods Enzymol12B:195–206
Marmur J (1962) A procedure for the isolation of deoxy-ribonucleic acid from microorganisms. J Mol Biol3:208–218
Saitou N, Nei M (1987) The neighbor-joining method: anew method for reconstructing phylogenetic tree. MolBiol Evol 4:406–425
Sasser M (1990) Identification of bacteria by gas chroma-tography of cellular fatty acids. USFCC Newsl 20:1–6
Shirling EB, Gottlieb D (1966) Methods for characteriza-tion of Streptomyces species. Int J Syst Bacteriol16:313–340
Thompson JD, Gibson TJ, Plewniak F, Jeanmougin F,Higgins DG (1997) The Clustal X windows interface:flexible strategies for multiple sequence alignmentaided by quality analysis tools. Nucleic Acids Res24:4876–4888
Tsukamura M (1966) Adansonian classification of myco-bacteria. J Gen Microbiol 45:253–273
Wayne LG, Brenner DJ, Colwell RR, Grimont PAD,Kandle O, Krichevsky MI, Moore WIC, Murry RGE,Stackerbrandt E, Starr MP, Truper HG (1987) Reportof the ad hoc committee on reconciliation of ap-proaches to bacterial systematics. Int J Syst Bacteriol37:463–464
396 Antonie van Leeuwenhoek (2007) 92:391–397
123

Williams ST, Goodfellow M, Alderson G, WellingtonEMH, Sneath PHA, Sackin MJ (1983) Numericalclassification of Streptomyces and related genera. GenMicrobiol 129:1743–1813
Xu P, Li WJ, Xu LH, Jiang CL (2003) A microwave-basedmethod for genomic DNA extraction from actino-mycetes. Microbiology (Chinese) 30:82–84
Antonie van Leeuwenhoek (2007) 92:391–397 397
123