structural basis for telomerase catalytic subunit tert ...cwp.embo.org/glc11-02/pdf/10 mitchell nat...
TRANSCRIPT

nature structural & molecular biology VOLUME 17 NUMBER 4 APRIL 2010 513
a r t i c l e s
Telomerase is a specialized ribonucleoprotein (RNP) reverse tran-scriptase1 that extends the 3′ ends of linear chromosomes2, which would otherwise diminish in size with every cell division, eventually leading to cellular aging3. Although this process is essential for cell viability in healthy tissues early in life, the enzyme is switched off in mature cells, ensuring a limited number of mitotic events. Tumors, however, regain the ability to activate telomerase, which in turn promotes the immortalization and spread of rapidly dividing cancer cells4. Understanding the function of this enzyme at a molecular level will aid in efforts to design therapies to combat both cancer and age-related diseases3,5–7.
The catalytic subunit of telomerase is structurally similar to retro-viral reverse transcriptases, viral RNA polymerases and, to a lesser extent, the bacteriophage B-family DNA polymerases1. Like its struc-tural homologs, the core catalytic subunit of telomerase, TERT, con-tains the fingers, palm and thumb domains1 required for nucleic acid and nucleotide associations as well as catalysis. However, unlike other polymerases, telomerase exists as a stable RNP complex and contains unique domains, such as the universally conserved RNA binding domain (TRBD), which is essential for RNP assembly8–10 and repeat addition processivity11,12. A fifth domain, known as the TEN or the anchor domain of telomerase, is present in several eukaryotes and has been shown to be involved in single-stranded telomeric DNA bind-ing13,14 and processivity15–17 in these organisms. In contrast to the largely conserved protein subunit, the RNA component of telomerase varies considerably in both size and sequence across species. Despite these differences, telomerase RNAs contain conserved motifs18,19 (such as the RNA-templating region essential for nucleotide binding and selectivity), evidence that supports the existence of common mecha-nistic aspects of telomere replication across phylogenetic groups.
Telomerase is a high-fidelity enzyme in that it is able to add multiple identical repeats of DNA, a capability that is conferred in large part by its integral RNA-templating region (usually one-and-a-half telomeric repeats)20–24. Initiation of telomere replication leads to pairing of the RNA-templating region with the incoming single-stranded DNA primer to be extended20–22,24,25. Current evidence suggests that initial RNA-DNA pairing occurs between 4 or 5 bases out of the possible 7–9 bases of the templating region, thus leaving a short string of RNA nucleotides to be replicated before the 5′ end of the templating region is reached20,26. Completion of one round of telomere extension leads to a transient dissociation of the RNA-DNA hybrid, followed by repo-sitioning of the DNA end at the active site of TERT and RNA-DNA pairing at the other end of the template. Telomerase remains associ-ated with the chromosome end until several telomeric repeats have been added, a process that is highly regulated by telomere binding proteins, such as the TPIP1–POT1 complex in humans27,28.
The structure of the T. castaneum full-length catalytic subunit of telomerase, TERT, together with existing biochemical data has pro-vided significant insight into the mechanism of telomere replication by telomerase. However, the precise mechanism of RNA-template usage and DNA binding and positioning at the active site of the enzyme for nucleotide incorporation are not clear. To address these questions, we determined the high-resolution structure of the T. castaneum TERT bound to an RNA-DNA hairpin designed to resemble its RNA tem-plate and the complementary DNA sequence. The structure revealed that RNA-template binding by TERT is mediated by the conserved motifs 2 and B′ of the fingers and palm domains, respectively, both of which localize adjacent to and above the active site of the enzyme. Contacts between the protein and the backbone of the RNA tem-plate position the solvent-accessible bases in proximity to the active
1Gene Expression and Regulation Program, The Wistar Institute, Philadelphia, Pennsylvania, USA. 2Department of Integrated Biosciences, Graduate School of Frontier Sciences, University of Tokyo, Kashiwa, Japan. Correspondence should be addressed to E.S. ([email protected]).
Received 17 October 2009; accepted 20 January 2010; published online 28 March 2010; doi:10.1038/nsmb.1777
Structural basis for telomerase catalytic subunit TERT binding to RNA template and telomeric DNAMeghan Mitchell1, Andrew Gillis1, Mizuko Futahashi2, Haruhiko Fujiwara2 & Emmanuel Skordalakes1
Telomerase is a specialized DNA polymerase that extends the 3′ ends of eukaryotic linear chromosomes, a process required for genomic stability and cell viability. Here we present the crystal structure of the active Tribolium castaneum telomerase catalytic subunit, TERT, bound to an RNA-DNA hairpin designed to resemble the putative RNA-templating region and telomeric DNA. The RNA-DNA hybrid adopts a helical structure, docked in the interior cavity of the TERT ring. Contacts between the RNA template and motifs 2 and B′ position the solvent-accessible RNA bases close to the enzyme active site for nucleotide binding and selectivity. Nucleic acid binding induces rigid TERT conformational changes to form a tight catalytic complex. Overall, TERT–RNA template and TERT–telomeric DNA associations are remarkably similar to those observed for retroviral reverse transcriptases, suggesting common mechanistic aspects of DNA replication between the two families of enzymes.
© 2
010
Nat
ure
Am
eric
a, In
c. A
ll ri
gh
ts r
eser
ved
.

514 VOLUME 17 NUMBER 4 APRIL 2010 nature structural & molecular biology
a r t i c l e s
site of the enzyme for nucleotide binding, thus facilitating selectivity. TERT-DNA associations are largely mediated by the thumb loop and helix. These contacts provide the stability required for a functional telomerase elongation complex and facilitate positioning of the 3′-end nucleotides in proximity to the primer-grip region (motif E), which in turn positions the 3′-end hydroxyl of the DNA primer at the active site of the enzyme for nucleotide addition. Collectively, the data presented here highlight the detailed interactions between TERT, the RNA-templating region and the DNA primer to be extended during the elongation process, providing novel insights into the mechanism of telomere replication by telomerase.
RESULTSTERT structure overviewWe cocrystallized the full-length (Fig. 1a) active T. castaneum TERT (Supplementary Fig. 1a,b) with an RNA-DNA hairpin containing the putative RNA-templating region (5′-rCrUrGrArCrCrU-3′) and the complementary telomeric DNA (5′-GTCAGGT-3′) joined together with a short RNA-DNA linker (Fig. 1b and Supplementary Fig. 2). Notably, the T. castaneum TERT lacks the TEN domain required for activity and processivity in several eukaryotic telomerase genes, includ-ing those of humans17 and Tetrahymena thermophila29,30. The absence of the TEN domain from T. castaneum could explain the reduced activity observed for this enzyme when compared to those contain-ing this domain. The RNA component of T. castaneum telomerase is not known, so we used the following information to predict its templating region. The RNA-templating region of telomerase is usu-ally one-and-a-half telomeric repeats22–24, and this sequence is known for many organisms (see telomerase database at http://telomerase.asu.edu/). For example, the mammalian telomerase-templating region is ‘CUAACCCU’, and the telomeric repeat is ‘TTAGGG’. The telomeric repeat for T. castaneum is ‘TCAGG’31,32.
We designed the RNA-DNA hairpin to contain a three-nucleotide overhang at the 5′ end of the RNA template so that we could trap the enzyme in its catalytic state by cocrystallizing the protein–nucleic acid assembly with Mg2+ ions and nonhydrolyzable nucleotides. In an effort to identify a hairpin suitable for crystallographic studies, we screened a number of RNA-DNA hairpins (15-mer, 18-mer and 21-mer; Supplementary Fig. 2) in which we kept the templating region and the complementary DNA sequence the same but changed the length of the linker. Although all three hairpins tested had the same binding affinity for TERT and the same reverse transcriptase
activity (data not shown), only the 21-mer was amenable to crystallo-graphic studies. Notably, in the structure, the hairpin linker extends out of the TERT ring and is only involved in crystal contacts with adjacent molecules. We also grew the crystals in the presence of the slowly hydrolyzable nucleotide analogs dNTPαS and Mg2+ ions.
We solved the structure at 2.7-Å resolution using the method of molecular replacement, with the substrate-free TERT1 (PDB 3DU6) as a search model (Table 1). All of the TERT molecule and the RNA-DNA hybrid were interpretable in electron density maps. Unexpectedly, there was extra density for three nucleotides at the 3′ end of the telomeric DNA (Fig. 1c), suggesting that TERT had extended the 3′ end of the DNA substrate in the crystallization drop. There was no evidence for nucleotide at the active site of the enzyme, which was partially occupied by the nucleotide located at the 3′ end of the DNA substrate.
Several lines of evidence indicate that the association of the RNA-templating region and the DNA substrate with TERT in the structure presented here are biologically relevant. First, TERT is an active polymerase in the presence of the nucleic acid substrate used in this study, as determined in standard reverse transcriptase assays
Fingers Palm ThumbTRBD
CP T 12 A IFD B′
5′5′
RNAtemplating
region
DNAprimer 3′
90°
Palm
Thumb
TRBDFingers
DNA
RNA
3′
C D E
a c
b
d e
Thumbloop helix
Figure 1 Structure of the T. castaneum TERT–nucleic acid complex. (a) Primary structure of the T. castaneum TERT. The TRBD, fingers, palm and thumb domains are shown in blue, orange, tan and red, respectively. Conserved motifs are also shown. (b) Primary structure of the RNA-DNA hairpin (21-mer) cocrystallized with TERT. Blue, nucleic acid linker. (c) Simulated annealed omit map of the RNA (magenta stick) and the DNA (yellow stick) substrates at 1.0s contour level. (d) Cartoon representation of the TERT–RNA-DNA complex; colors same as in a and c. (e) TERT surface charge representation, showing the RNA-DNA hybrid (stick) docked in the highly positively charged (blue color) cavity of the ring.
Table 1 Data collection and refinement statisticsTERT–RNA-DNA complex
Data collection
Space group P21
Cell dimensions
a, b, c (Å) 77.2, 52.8, 101.6
α, b, g (°) 90, 101.9, 90
Resolution (Å) 20–2.7 (2.85–2.70)
Rsym 11.4 (45.8)
I / σI 7.9 (2.3)
Completeness (%) 93.7 (96.1)
Redundancy 2.8 (2.5)
Refinement
Resolution (Å) 20–2.7
No. reflections 19,815
Rwork / Rfree 24.2 / 28.7
No. atoms
Protein 4,982
Ligand/ion 496/1
Water 54
B-factors
Protein 45
Ligand/ion 37
Water 21
R.m.s. deviations
Bond lengths (Å) 0.006
Bond angles (°) 0.887
One crystal was used. Values in parentheses are for highest-resolution shell.
© 2
010
Nat
ure
Am
eric
a, In
c. A
ll ri
gh
ts r
eser
ved
.

nature structural & molecular biology VOLUME 17 NUMBER 4 APRIL 2010 515
a r t i c l e s
(Supplementary Fig. 3) and in the crystallization drop. Second, the RNA template makes contacts with conserved motifs that are hallmarks of this family of enzymes. Third, contacts between TERT and the RNA template position the solvent-accessible bases adjacent to and above the active site of the enzyme for nucleotide binding, thus facilitating selectivity. Moreover, TERT-RNA associations posi-tion the 5′ end of the RNA template at the entry of the TRBD RNA binding pocket and where the template boundary element located upstream of the RNA-template in many organisms is thought to bind. Fourth, interactions between the DNA substrate and the primer-grip region (a characteristic shared among telomerase and HIV reverse transcriptases (HIV RTs)) place the 3′ end of the DNA substrate at the active site of the enzyme, where it is accessible for nucleotide addition. Fifth, TERT–nucleic acid associations are strikingly similar to those observed for HIV RT, TERT’s closest structural homolog.
The four major TERT domains—the RNA binding domain (TRBD); the fingers domain, implicated in nucleotide binding and processivity1,33; the palm domain, which contains the active site of the enzyme1; and the thumb domain, implicated in DNA binding and processivity1,34—are organized into a ring configuration similar to that observed for the substrate-free enzyme1 (Fig. 1d). The arrange-ment of the TERT domains creates a highly positively charged cavity in the interior of the TERT ring (Fig. 1e), which is 22 Å wide and 21 Å deep and can accommodate seven bases of double-stranded nucleic acid. Within this cavity binds one molecule of the RNA-DNA hybrid (Fig. 1d,e), which assembles via Watson-Crick base pairing into a helical structure similar to both the DNA-DNA and RNA-DNA structures bound to HIV RT35,36.
TERT nucleic acid associationsInteractions between the protein and the RNA-templating region are mediated by the fingers, palm and thumb domains. The 5′-end RNA cytosine (rC1) and uracil (rU2) are located at the interface of the fingers and palm domains and are involved in a network of inter-actions with conserved residues (Supplementary Fig. 4) of motifs 2 and B′, both of which are located in proximity to the active site of the enzyme (Fig. 2a,b). In particular, the 2′-OH of rC1 is within hydrogen bonding distance of the backbone carbonyls of Val197 of motif 2 and Gly309 of motif B′, and the pyrimidine base sits over the otherwise solvent-exposed hydrophobic side chain of the
conserved Ile196, which also forms part of motif 2 (Fig. 2b). Contacts between rU2 and the protein are mediated by the short aliphatic side chain of Pro311 and the ribose group (Fig. 2b). Interactions between rC1 and rU2 with motifs 2 and B′ place the cytosine base in proximity to the active site of the enzyme, where it is well positioned for Watson-Crick base pairing with the incoming nucleotide sub-strate. Stabilization and placement of the 5′-end bases of the templat-ing region above the active site of the enzyme are further facilitated by the interactions of the remaining five ribonucleotides with the incom-ing DNA primer. Limited contacts between this part of the RNA and the protein are mediated via a water molecule, which coordinates the 2′-OH of guanosine (rG3) with the backbone of helix α15 (Fig. 2b). Notably, the structural organization of helix α15 is influenced by the IFD motif, a long insertion consisting of two helices (α13 and α14), between motifs A and B′ (Fig. 2a), which explains why mutations in this motif lead to loss of telomerase function1,37.
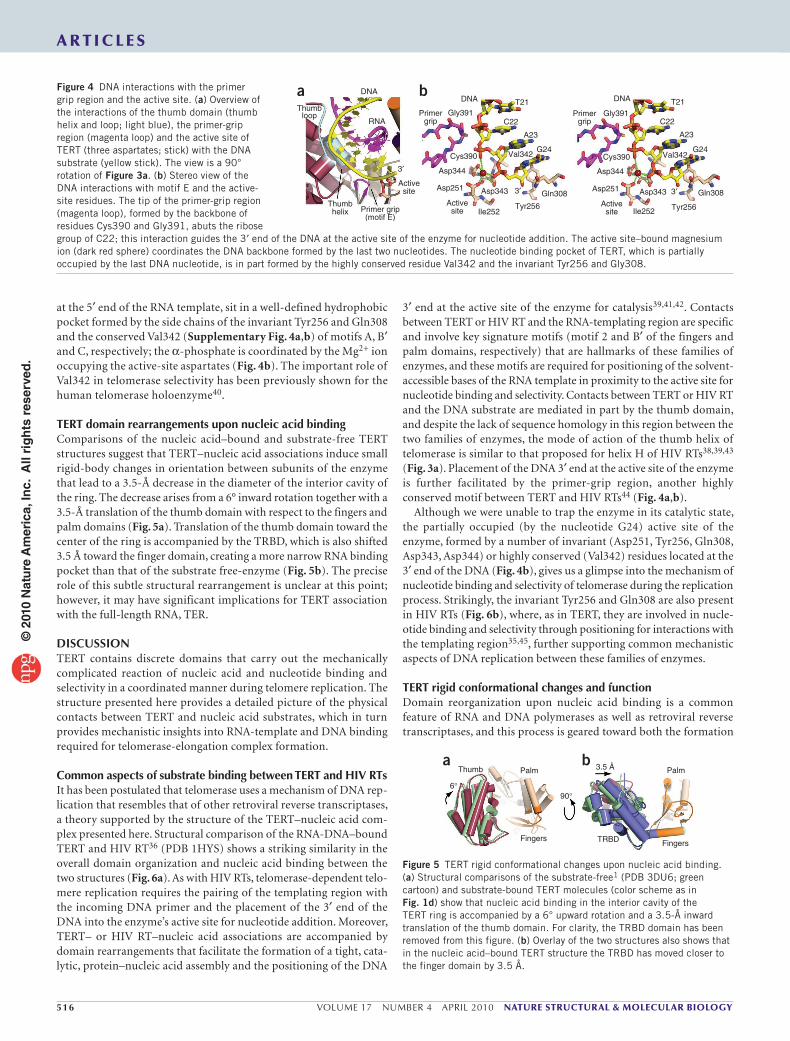
Contacts between TERT and the DNA substrate are mostly mediated via backbone interactions with the thumb loop and helix (Fig. 3a). The thumb helix1 sits in the minor groove of the RNA-DNA heteroduplex, making extensive contacts with the phosphodiester backbone and the ribose groups of the RNA-DNA hybrid. The mode of action of the thumb helix of telomerase appears to be similar to that proposed for the equivalent helix (helix H) in retroviral reverse transcriptases38,39. Another conserved element of the thumb domain, known as the thumb loop1, runs almost parallel to the curvature of the DNA primer, and the two are involved in a network of backbone- and solvent-mediated interactions (Fig. 3b). Interactions between the DNA and the thumb loop include the side chains of Lys416 and Asn423, both of which extend toward the center of the ring and are within hydrogen bonding distance of the DNA backbone. Contacts between the thumb domain and the DNA position the nucleotides located at its 3′ end within coordinating distance of the primer-grip region, a short, rigid loop located at the interface of the palm and thumb domains and in proximity to the active site of the enzyme (Fig. 4a,b). The tip of this loop, formed by the backbone of conserved residues Cys390 and Gly391 (Supplementary Fig. 4b), abuts the ribose group of C22. This interaction guides the 3′-end DNA nucleotides toward the active site of the enzyme (Fig. 4b). The active site of the enzyme, where the incoming nucleotide is projected to bind1, is partially occupied
by the nucleotide (G24) located at the 3′ end of the DNA (Fig. 4b). The ribose group and, to a certain extent, the guanosine base of G24, which makes Watson-Crick pairing interactions with the rC1 located
DNARNA
Palm
Activesite
MotifT
Fingers
Motif 2Motif B′
5′
5′
α13
α15α15
α10 α14
Motif IFD
Motif2
MotifB′
Val197
Gly309
Ile196
Pro311rC1
rU2
rG3
rA4RNA
Wat18
a b
Figure 2 TERT–RNA template associations. (a) RNA template (magenta sticks) binding in the interior of the TERT ring is mediated by motifs 2 (cyan), B′ (purple) and helix α15 via a water molecule (Wat18). Contacts between the RNA template and the protein place its 5′ end at the entry of the RNA binding pocket (motif T, blue) of TRBD. (b) The nucleotide located at the 5′ end of the RNA template (rC1) is coordinated by Ile196 and Val197 of motif 2 and Gly309 of motif B′. rU2 interacts with Pro311 of motif B′, and rG3 coordinates the backbone of helix α15 via a water molecule (Wat18; red sphere).
Thumbloop
Thumbloop
Thumbhelix
DNA
RNA
Lys416
T21
G20
G19
A18C17
DNA Asn423
Wat6
Wat9
a b
Figure 3 TERT–telomeric DNA associations. (a) TERT-DNA (yellow stick) binding is mediated by the thumb loop (light blue) and helix (light blue). The thumb helix docks itself into the minor groove of the RNA-DNA hybrid, and the thumb loop adopts the geometry of double-stranded nucleic acid and is involved in a network of interactions with the DNA. (b) Interactions between the thumb loop and the DNA are mostly mediated by backbone and solvent. Also, the side chains of Lys416 and Asn423 (light blue stick) that form part of the thumb loop extend toward the center of the ring and coordinate the DNA backbone.
© 2
010
Nat
ure
Am
eric
a, In
c. A
ll ri
gh
ts r
eser
ved
.
516 VOLUME 17 NUMBER 4 APRIL 2010 nature structural & molecular biology
a r t i c l e s
at the 5′ end of the RNA template, sit in a well-defined hydrophobic pocket formed by the side chains of the invariant Tyr256 and Gln308 and the conserved Val342 (Supplementary Fig. 4a,b) of motifs A, B′ and C, respectively; the α-phosphate is coordinated by the Mg2+ ion occupying the active-site aspartates (Fig. 4b). The important role of Val342 in telomerase selectivity has been previously shown for the human telomerase holoenzyme40.
TERT domain rearrangements upon nucleic acid bindingComparisons of the nucleic acid–bound and substrate-free TERT structures suggest that TERT–nucleic acid associations induce small rigid-body changes in orientation between subunits of the enzyme that lead to a 3.5-Å decrease in the diameter of the interior cavity of the ring. The decrease arises from a 6° inward rotation together with a 3.5-Å translation of the thumb domain with respect to the fingers and palm domains (Fig. 5a). Translation of the thumb domain toward the center of the ring is accompanied by the TRBD, which is also shifted 3.5 Å toward the finger domain, creating a more narrow RNA binding pocket than that of the substrate free-enzyme (Fig. 5b). The precise role of this subtle structural rearrangement is unclear at this point; however, it may have significant implications for TERT association with the full-length RNA, TER.
DISCUSSIONTERT contains discrete domains that carry out the mechanically complicated reaction of nucleic acid and nucleotide binding and selectivity in a coordinated manner during telomere replication. The structure presented here provides a detailed picture of the physical contacts between TERT and nucleic acid substrates, which in turn provides mechanistic insights into RNA-template and DNA binding required for telomerase-elongation complex formation.
Common aspects of substrate binding between TERT and HIV RTsIt has been postulated that telomerase uses a mechanism of DNA rep-lication that resembles that of other retroviral reverse transcriptases, a theory supported by the structure of the TERT–nucleic acid com-plex presented here. Structural comparison of the RNA-DNA–bound TERT and HIV RT36 (PDB 1HYS) shows a striking similarity in the overall domain organization and nucleic acid binding between the two structures (Fig. 6a). As with HIV RTs, telomerase-dependent telo-mere replication requires the pairing of the templating region with the incoming DNA primer and the placement of the 3′ end of the DNA into the enzyme’s active site for nucleotide addition. Moreover, TERT– or HIV RT–nucleic acid associations are accompanied by domain rearrangements that facilitate the formation of a tight, cata-lytic, protein–nucleic acid assembly and the positioning of the DNA
3′ end at the active site of the enzyme for catalysis39,41,42. Contacts between TERT or HIV RT and the RNA-templating region are specific and involve key signature motifs (motif 2 and B′ of the fingers and palm domains, respectively) that are hallmarks of these families of enzymes, and these motifs are required for positioning of the solvent-accessible bases of the RNA template in proximity to the active site for nucleotide binding and selectivity. Contacts between TERT or HIV RT and the DNA substrate are mediated in part by the thumb domain, and despite the lack of sequence homology in this region between the two families of enzymes, the mode of action of the thumb helix of telomerase is similar to that proposed for helix H of HIV RTs38,39,43 (Fig. 3a). Placement of the DNA 3′ end at the active site of the enzyme is further facilitated by the primer-grip region, another highly conserved motif between TERT and HIV RTs44 (Fig. 4a,b).
Although we were unable to trap the enzyme in its catalytic state, the partially occupied (by the nucleotide G24) active site of the enzyme, formed by a number of invariant (Asp251, Tyr256, Gln308, Asp343, Asp344) or highly conserved (Val342) residues located at the 3′ end of the DNA (Fig. 4b), gives us a glimpse into the mechanism of nucleotide binding and selectivity of telomerase during the replication process. Strikingly, the invariant Tyr256 and Gln308 are also present in HIV RTs (Fig. 6b), where, as in TERT, they are involved in nucle-otide binding and selectivity through positioning for interactions with the templating region35,45, further supporting common mechanistic aspects of DNA replication between these families of enzymes.
TERT rigid conformational changes and functionDomain reorganization upon nucleic acid binding is a common feature of RNA and DNA polymerases as well as retroviral reverse transcriptases, and this process is geared toward both the formation
Palm PalmThumb
TRBD
3.5 Å
6°90°
FingersFingers
a b
Figure 5 TERT rigid conformational changes upon nucleic acid binding. (a) Structural comparisons of the substrate-free1 (PDB 3DU6; green cartoon) and substrate-bound TERT molecules (color scheme as in Fig. 1d) show that nucleic acid binding in the interior cavity of the TERT ring is accompanied by a 6° upward rotation and a 3.5-Å inward translation of the thumb domain. For clarity, the TRBD domain has been removed from this figure. (b) Overlay of the two structures also shows that in the nucleic acid–bound TERT structure the TRBD has moved closer to the finger domain by 3.5 Å.
3′
3′
DNA
RNA
Thumbloop
Thumbhelix Primer grip
(motif E)
Activesite
Activesite Ile252
Tyr256
Gln308Asp343Asp251
Asp344
Cys390 Val342G24
A23
C22
T21DNA
Gly391Primergrip
3′Active
site Ile252 Tyr256
Gln308Asp343Asp251
Asp344
Cys390 Val342G24
A23
C22
T21DNA
Gly391Primergrip
a bFigure 4 DNA interactions with the primer grip region and the active site. (a) Overview of the interactions of the thumb domain (thumb helix and loop; light blue), the primer-grip region (magenta loop) and the active site of TERT (three aspartates; stick) with the DNA substrate (yellow stick). The view is a 90° rotation of Figure 3a. (b) Stereo view of the DNA interactions with motif E and the active-site residues. The tip of the primer-grip region (magenta loop), formed by the backbone of residues Cys390 and Gly391, abuts the ribose group of C22; this interaction guides the 3′ end of the DNA at the active site of the enzyme for nucleotide addition. The active site–bound magnesium ion (dark red sphere) coordinates the DNA backbone formed by the last two nucleotides. The nucleotide binding pocket of TERT, which is partially occupied by the last DNA nucleotide, is in part formed by the highly conserved residue Val342 and the invariant Tyr256 and Gly308.
© 2
010
Nat
ure
Am
eric
a, In
c. A
ll ri
gh
ts r
eser
ved
.

nature structural & molecular biology VOLUME 17 NUMBER 4 APRIL 2010 517
a r t i c l e s
of a tight, catalytic protein–nucleic acid assembly and the positioning of the DNA 3′ end at the active site of the enzyme for catalysis39,41,42. Unlike HIV RTs, telomerase seems to exist, at least in the absence of the full-length integral RNA component, in a closed-ring configuration, an arrangement mediated by extensive contacts between the TRBD and the thumb domains1. Comparison of the nucleic acid–bound and substrate-free TERT structures suggests that TERT–nucleic acid associations induce subtle rigid-body changes in orientation between subunits of the enzyme, leading to a 3.5-Å decrease in the diameter of the interior cavity of the ring.
These observations are unexpected because, in most polymerases, including the HIV RT, the fingers and thumb domains undergo sig-nificant conformational changes required for substrate binding and function42,46,47. For example, the fingers domain, which is known to bind and position the nucleotide at the active site of the enzyme, undergoes significant conformational changes referred to as the ‘open’ and ‘closed’ states46. It is therefore possible that the interactions between the TRBD and the thumb domain lock the fingers domain in place, thus preventing the conformational rearrangements observed in other polymerases, which would suggest the possibility of a preformed active site. A preformed active site has been previously observed for the hepatitis C viral RNA polymerase48, a close structural homolog of TERT, and also for the Y-family DNA polymerases49. Another possi-bility is that we trapped the substrate-free TERT enzyme in the closed fingers conformation. Assuming this is true, significant movement of the fingers domain of TERT would most likely require the TRBD and the thumb domain to be splayed apart. Contacts between the TRBD and the thumb domain are extensive—thus, significant energy would be required to force them apart1. This could be accomplished by accessory proteins that possibly act in a similar manner to that of the sliding clamp loader of DNA polymerases50.
Repeat addition processivityTelomerase, unlike most polymerases, has the ability to add multiple identical repeats of DNA to the ends of chromosomes, a process known as repeat addition processivity. This unique characteristic of telomerase has been attributed in part to the association of the N-terminal portion of TERT with TER and the telomeric overhang as well as the IFD motif 37. The TEN domain, present in several organ-isms, and its weak interaction with the DNA substrate is thought to be a determinant of repeat addition processivity15–17; most recently, the TRBD and its stable association with TER has also been shown to be involved in this process11,12. In the complex presented here,
the RNA does not directly engage the RNA binding pocket of TRBD. The structure shows that TERT-RNA contacts position the 5′ end of the templating region at the entry of the RNA binding pocket of TRBD (Fig. 2a). This arrangement would place either the template boundary element51–53, present in most organisms, or the short oligonucleotide overhang of rodent TER51 within the RNA binding pocket of TRBD. The stable associa-tion of TER with the TRBD would force the enzyme to stall when reaching the nucleotide located at the 5′ end of the RNA template, thus preventing replication beyond this point. Stalling of the enzyme for extended periods would lead to destabilization and dissociation of the RNA-DNA heteroduplex and initiation of another round of telomere replication.
Collectively, the data presented here, together with previous struc-tural and biochemical evidence, support common mechanistic aspects of substrate binding and DNA replication between telomerase and HIV RTs, suggesting an evolutionary link between these families of enzymes. These data also provide novel insights into the basic mecha-nisms of telomere replication and length homeostasis by telomerase, which in turn enriches our understanding of the mechanism of DNA replication by polymerases in general. Moreover, the structure presented here provides a detailed picture of the physical contacts between TERT and its nucleic acid substrates, information that is invaluable in our effort to design small-molecule inhibitors of telo-merase that may be of therapeutic value for cancer and other diseases associated with cellular aging.
METHODSMethods and any associated references are available in the online version of the paper at http://www.nature.com/nsmb/.
Accession codes. Protein Data Bank: Coordinates for the TERT–RNA-DNA complex have been deposited under accession code 3KYL.
Note: Supplementary information is available on the Nature Structural & Molecular Biology website.
AcknowlEdGMEntSWe would like to thank S.J. Gamblin and S. Pennell for helpful discussions. Funding for this project was generously provided by the Ellison Medical and the V Foundations as well as the Pennsylvania Department of Health.
AUtHoR contRIBUtIonSE.S. designed the experiment plan, analyzed the data and wrote the manuscript; A.G. prepared the recombinant telomerase and carried out the reverse transcriptase assays; M.M. carried out the TRAP assays; H.F. and M.F. provided advice with the T. castaneum TRAP assays.
coMPEtInG FInAncIAl IntEREStSThe authors declare no competing financial interests.
Published online at http://www.nature.com/nsmb/. Reprints and permissions information is available online at http://npg.nature.com/reprintsandpermissions/.
1. Gillis, A.J., Schuller, A.P. & Skordalakes, E. Structure of the Tribolium castaneum telomerase catalytic subunit TERT. Nature 455, 633–637 (2008).
2. Greider, C.W. & Blackburn, E.H. Identification of a specific telomere terminal transferase activity in Tetrahymena extracts. Cell 43, 405–413 (1985).
3. Harley, C.B., Futcher, A.B. & Greider, C.W. Telomeres shorten during ageing of human fibroblasts. Nature 345, 458–460 (1990).
Gln308/Gln151
Gly309/Gly152
MotifB′Tyr256/Tyr115
Val342/Met184Cys390/Met230
MotifC
ActivesiteHIV RT
TERT
Primergrip
Fingers
Thumb
RNA DNAPalm
Gly391/Gly231
MotifA
a b
Figure 6 Structural comparison of the RNA-DNA–bound TERT and HIV RT. (a) Structural comparison of TERT (yellow) bound to RNA-DNA (orange) and HIV RT (light blue) bound to RNA-DNA (blue) (PDB 1HYS)36. The TRBD has been removed for clarity. (b) Overlay of the palm domains of the RNA-DNA–bound structures36 of TERT and HIV RT (PDB 1HYS) shows a strong conservation in the sequence and organization of the primer grip and active-site residues. Active-site residues of TERT (left of slash) and HIV RT (right of slash) are labeled.
© 2
010
Nat
ure
Am
eric
a, In
c. A
ll ri
gh
ts r
eser
ved
.

518 VOLUME 17 NUMBER 4 APRIL 2010 nature structural & molecular biology
4. Kim, N.W. et al. Specific association of human telomerase activity with immortal cells and cancer. Science 266, 2011–2015 (1994).
5. Harley, C.B., Vaziri, H., Counter, C.M. & Allsopp, R.C. The telomere hypothesis of cellular aging. Exp. Gerontol. 27, 375–382 (1992).
6. Harley, C.B. & Villeponteau, B. Telomeres and telomerase in aging and cancer. Curr. Opin. Genet. Dev. 5, 249–255 (1995).
7. Harley, C.B. Telomerase and cancer therapeutics. Nat. Rev. Cancer 8, 167–179 (2008).
8. Bosoy, D., Peng, Y., Mian, I.S. & Lue, N.F. Conserved N-terminal motifs of telomerase reverse transcriptase required for ribonucleoprotein assembly in vivo. J. Biol. Chem. 278, 3882–3890 (2003).
9. Bryan, T.M., Goodrich, K.J. & Cech, T.R. Telomerase RNA bound by protein motifs specific to telomerase reverse transcriptase. Mol. Cell 6, 493–499 (2000).
10. Lai, C.K., Mitchell, J.R. & Collins, K. RNA binding domain of telomerase reverse transcriptase. Mol. Cell. Biol. 21, 990–1000 (2001).
11. Drosopoulos, W.C. & Prasad, V.R. Telomerase-specific T motif is a restrictive determinant of repetitive reverse transcription by human telomerase. Mol. Cell. Biol. 30, 447–459 (2010).
12. Rouda, S. & Skordalakes, E. Structure of the RNA-binding domain of telomerase: implications for RNA recognition and binding. Structure 15, 1403–1412 (2007).
13. Hammond, P.W., Lively, T.N. & Cech, T.R. The anchor site of telomerase from Euplotes aediculatus revealed by photo-cross-linking to single- and double-stranded DNA primers. Mol. Cell. Biol. 17, 296–308 (1997).
14. Jacobs, S.A., Podell, E.R. & Cech, T.R. Crystal structure of the essential N-terminal domain of telomerase reverse transcriptase. Nat. Struct. Mol. Biol. 13, 218–225 (2006).
15. Wyatt, H.D., Lobb, D.A. & Beattie, T.L. Characterization of physical and functional anchor site interactions in human telomerase. Mol. Cell. Biol. 27, 3226–3240 (2007).
16. Moriarty, T.J., Marie-Egyptienne, D.T. & Autexier, C. Functional organization of repeat addition processivity and DNA synthesis determinants in the human telomerase multimer. Mol. Cell. Biol. 24, 3720–3733 (2004).
17. Wyatt, H.D., Tsang, A.R., Lobb, D.A. & Beattie, T.L. Human telomerase reverse transcriptase (hTERT) Q169 is essential for telomerase function in vitro and in vivo. PLoS One 4, e7176 (2009).
18. Chen, J.L. & Greider, C.W. An emerging consensus for telomerase RNA structure. Proc. Natl. Acad. Sci. USA 101, 14683–14684 (2004).
19. Lin, J. et al. A universal telomerase RNA core structure includes structured motifs required for binding the telomerase reverse transcriptase protein. Proc. Natl. Acad. Sci. USA 101, 14713–14718 (2004).
20. Gilley, D., Lee, M.S. & Blackburn, E.H. Altering specific telomerase RNA template residues affects active site function. Genes Dev. 9, 2214–2226 (1995).
21. Greider, C.W. & Blackburn, E.H. A telomeric sequence in the RNA of Tetrahymena telomerase required for telomere repeat synthesis. Nature 337, 331–337 (1989).
22. Lee, M.S. & Blackburn, E.H. Sequence-specific DNA primer effects on telomerase polymerization activity. Mol. Cell. Biol. 13, 6586–6599 (1993).
23. Lingner, J., Hendrick, L.L. & Cech, T.R. Telomerase RNAs of different ciliates have a common secondary structure and a permuted template. Genes Dev. 8, 1984–1998 (1994).
24. Shippen-Lentz, D. & Blackburn, E.H. Functional evidence for an RNA template in telomerase. Science 247, 546–552 (1990).
25. Autexier, C. & Greider, C.W. Boundary elements of the Tetrahymena telomerase RNA template and alignment domains. Genes Dev. 9, 2227–2239 (1995).
26. Harrington, L.A. & Greider, C.W. Telomerase primer specificity and chromosome healing. Nature 353, 451–454 (1991).
27. Greider, C.W. Telomerase is processive. Mol. Cell. Biol. 11, 4572–4580 (1991).28. Wang, F. et al. The POT1–TPP1 telomere complex is a telomerase processivity
factor. Nature 445, 506–510 (2007).29. Zaug, A.J., Podell, E.R. & Cech, T.R. Mutation in TERT separates processivity from
anchor-site function. Nat. Struct. Mol. Biol. 15, 870–872 (2008).30. Finger, S.N. & Bryan, T.M. Multiple DNA-binding sites in Tetrahymena telomerase.
Nucleic Acids Res. 36, 1260–1272 (2008).
31. Osanai, M., Kojima, K.K., Futahashi, R., Yaguchi, S. & Fujiwara, H. Identification and characterization of the telomerase reverse transcriptase of Bombyx mori (silkworm) and Tribolium castaneum (flour beetle). Gene 376, 281–289 (2006).
32. Richards, S. et al. The genome of the model beetle and pest Tribolium castaneum. Nature 452, 949–955 (2008).
33. Bosoy, D. & Lue, N.F. Functional analysis of conserved residues in the putative “finger” domain of telomerase reverse transcriptase. J. Biol. Chem. 276, 46305–46312 (2001).
34. Hossain, S., Singh, S. & Lue, N.F. Functional analysis of the C-terminal extension of telomerase reverse transcriptase. A putative “thumb” domain. J. Biol. Chem. 277, 36174–36180 (2002).
35. Huang, H., Chopra, R., Verdine, G.L. & Harrison, S.C. Structure of a covalently trapped catalytic complex of HIV-1 reverse transcriptase: implications for drug resistance. Science 282, 1669–1675 (1998).
36. Sarafianos, S.G. et al. Crystal structure of HIV-1 reverse transcriptase in complex with a polypurine tract RNA:DNA. EMBO J. 20, 1449–1461 (2001).
37. Lue, N.F., Lin, Y.C. & Mian, I.S. A conserved telomerase motif within the catalytic domain of telomerase reverse transcriptase is specifically required for repeat addition processivity. Mol. Cell. Biol. 23, 8440–8449 (2003).
38. Jacobo-Molina, A. et al. Crystal structure of human immunodeficiency virus type 1 reverse transcriptase complexed with double-stranded DNA at 3.0 Å resolution shows bent DNA. Proc. Natl. Acad. Sci. USA 90, 6320–6324 (1993).
39. Kohlstaedt, L.A., Wang, J., Friedman, J.M., Rice, P.A. & Steitz, T.A. Crystal structure at 3.5 Å resolution of HIV-1 reverse transcriptase complexed with an inhibitor. Science 256, 1783–1790 (1992).
40. Drosopoulos, W.C. & Prasad, V.R. The active site residue Valine 867 in human telomerase reverse transcriptase influences nucleotide incorporation and fidelity. Nucleic Acids Res. 35, 1155–1168 (2007).
41. Rodgers, D.W. et al. The structure of unliganded reverse transcriptase from the human immunodeficiency virus type 1. Proc. Natl. Acad. Sci. USA 92, 1222–1226 (1995).
42. Steitz, T.A. DNA and RNA polymerases: structural diversity and common mechanisms. Harvey Lect. 93, 75–93 (1997).
43. Beese, L.S., Derbyshire, V. & Steitz, T.A. Structure of DNA polymerase I Klenow fragment bound to duplex DNA. Science 260, 352–355 (1993).
44. Tantillo, C. et al. Locations of anti-AIDS drug binding sites and resistance mutations in the three-dimensional structure of HIV-1 reverse transcriptase. Implications for mechanisms of drug inhibition and resistance. J. Mol. Biol. 243, 369–387 (1994).
45. Cases-Gonzalez, C.E., Gutierrez-Rivas, M. & Menendez-Arias, L. Coupling ribose selection to fidelity of DNA synthesis. The role of Tyr-115 of human immunodeficiency virus type 1 reverse transcriptase. J. Biol. Chem. 275, 19759–19767 (2000).
46. Ding, J. et al. Structure and functional implications of the polymerase active site region in a complex of HIV-1 RT with a double-stranded DNA template-primer and an antibody Fab fragment at 2.8 Å resolution. J. Mol. Biol. 284, 1095–1111 (1998).
47. Steitz, T.A. DNA polymerases: structural diversity and common mechanisms. J. Biol. Chem. 274, 17395–17398 (1999).
48. Bressanelli, S., Tomei, L., Rey, F.A. & De Francesco, R. Structural analysis of the hepatitis C virus RNA polymerase in complex with ribonucleotides. J. Virol. 76, 3482–3492 (2002).
49. Ling, H., Boudsocq, F., Woodgate, R. & Yang, W. Crystal structure of a Y-family DNA polymerase in action: a mechanism for error-prone and lesion-bypass replication. Cell 107, 91–102 (2001).
50. Jeruzalmi, D., O’Donnell, M. & Kuriyan, J. Clamp loaders and sliding clamps. Curr. Opin. Struct. Biol. 12, 217–224 (2002).
51. Chen, J.L. & Greider, C.W. Template boundary definition in mammalian telomerase. Genes Dev. 17, 2747–2752 (2003).
52. Lai, C.K., Miller, M.C. & Collins, K. Template boundary definition in Tetrahymena telomerase. Genes Dev. 16, 415–420 (2002).
53. Tzfati, Y., Fulton, T.B., Roy, J. & Blackburn, E.H. Template boundary in a yeast telomerase specified by RNA structure. Science 288, 863–867 (2000).
a r t i c l e s©
201
0 N
atu
re A
mer
ica,
Inc.
All
rig
hts
res
erve
d.

nature structural & molecular biologydoi:10.1038/nsmb.1777
ONLINE METHODSProtein expression and purification. We overexpressed the wild-type and mutant (D251A) T. castaneum, full-length TERT proteins with some subtle modifications to the protocol used previously1 that proved useful in increasing the protein yield. We now overexpressed the protein in Escherichia coli Rosetta (DE3) pLysS (Novagen) at 30 °C for 5 h. We dialyzed stock protein (10 mg ml−1) in 10 mM Tris-HCl, 100 mM KCl, 1 mM TCEP, pH 7.5, before crystallization trials.
Preparation of T. castaneum extracts and total RNA isolation. We ground 20 T. castaneum larvae or pupae in liquid N2, homogenized them with 200 µl extrac-tion buffer (25 mM Tris-HCl, 5 mM b-mercaptoethanol (b-ME), 1 mM EGTA, 0.1 mM benzamidine, 200 mM KCl, 10% (w/v) glycerol, 10 mM imidazole and RNasin (Promega), pH 7.5) and placed them on ice for 30 min. After we centri-fuged the homogenate at 12,000g at 4 °C for 20 min, we collected the superna-tant, flash-froze it in liquid nitrogen and stored it at −80 °C before use. We then extracted the total RNA from the T. castaneum homogenate using the RNeasy Protect Mini Kit from Qiagen.
In vitro reconstitution of T. castaneum telomerase. The telomerase RNA of T. castaneum is unknown, so we used the total RNA isolated from T. castaneum larvae and the recombinant TERT to assemble the telomerase complex in vitro. We mixed 20 µg of the His-tagged TERT (25 mM Tris, 200 mM KCl, 10% (w/v) glycerol, 5 mM b-ME and 10 mM imidazole, pH 7.5) with 50 µl of T. castaneum larvae total RNA, and we incubated the two in T. castaneum lysate for 2 h at 22 °C in the presence of RNasin. We then purified the telomerase complex over a nickel–nitrilotriacetic acid column and tested it for activity using a modified version of the TRAP assay54 as described below.
Telomerase repeat amplification protocol assays. We tested the activity of the in vitro–reconstituted T. castaneum telomerase using the following TRAP assay. We carried out the telomerase-elongation step in a 50-µl reaction mixture composed of 20 mM Tris-HCl (pH 8.3), 7.5 mM MgCl2, 63 mM KCl, 0.05% (w/v) Tween 20, 1 mM EGTA, 0.01% (w/v) BSA, 0.5 mM of each dNTP, 1 µM DNA primer (5′-AAGCCGTCGAGCAGAGTC-3′) (Tcas-TS)) and 4 µg of nickel–nitrilotriacetic acid–purified T. castaneum telomerase. After incubation at 30 °C for 60 min, we extracted each reaction mixture with phenol-chloroform and precipitated each with ethanol. We resuspended each sample in 50 µl PCR reaction buffer (10 mM Tris-HCl (pH 8.0), 50 mM KCl, 2 mM MgCl2, 100 µM dNTPs (dATP, dTTP and dGTP), 10 µM [32P]dCTP (80 Ci mmol−1), 1 µM Tcas-CX primer (5′-GTGTGACCTGACCTGACC-3′) and HotStar Taq DNA polymerase (Qiagen)). We resolved the PCR products on Tris-borate-EDTA (TBE)–polyacrylamide gels (Supplementary Fig. 1). We also confirmed the presence of multiple telomeric repeats (TCAGG)n by subcloning and sequencing the TRAP products.
TERT, reverse transcriptase assays. We carried out standard reverse transcriptase assays using the recombinant T. castaneum TERT and the RNA-DNA hairpin used in this study to test TERT’s ability to replicate the end of the DNA substrate that comprises part of the RNA-DNA hairpin. We carried reverse transcriptase assays out in telomerase buffer (50 mM Tris-HCl, 100 mM KCl, 1.25 mM MgCl2,
5 mM DTT, 5% (w/v) glycerol, pH 8 at room temperature), 100 µM dNTPs (dATP, dTTP and dGTP), 10 µM [32P]dCTP (80 Ci mmol−1), 5 µM RNA-DNA hairpin and 1 µM recombinant TERT. We incubated the samples for 2 h at room temperature and then extracted and precipitated them with phenol-chloroform and ethanol, respectively. We resuspended the DNA pellet in a solution com-posed of 90% (w/v) formamide and 10% (w/v) glycerol and ran the sample on a 12% (w/v) polyacrylamide–7 M urea gel in 1× TBE at 220 V for 70 min at 4 °C (Supplementary Fig. 3).
Protein crystallization and data collection. We prepared the binary complex by adding to the dialyzed protein 1.2 M excess nucleic acid (RNA-DNA hair-pin purchased from Integrated DNA Technologies), 5 mM dNTPαS (Jena Biosciences GmbH) and 5 mM MgCl2. Crystals of the monoclinic space group P21 that diffracted to 2.7-Å resolution appeared in 3 d and grew to final size in 2 weeks. We grew crystals by the vapor-diffusion, sitting-drop method by mixing one volume of the ternary complex with one volume of reservoir solu-tion containing 0.1 M HEPES (pH 7.5) and 12% 1,6-hexanediol or PEG 4K and 0.2 M KCl. We transferred crystals into cryoprotectant solution that contained 0.1 M HEPES (pH 7.5), 15% (w/v) 1,6-hexanediol or PEG 4K, 15% (w/v) glycerol, 0.2 M KCl and 1 mM TCEP and harvested them by flash freezing in liquid nitrogen. We collected X-ray data at the NSLS beamline X25 and processed it with MOSFILM as implemented in WEDGER-ELVES55 (Table 1). There is one monomer in the asymmetric unit.
Structure determination and refinement. We calculated phases by molecular replacement with PHASER56, using the substrate-free TERT structure1 (PDB 3DU6) as a search model. Maps calculated after one cycle of refinement by CNS-SOLVE57 revealed clear 2Fo − Fc density for all 596 residues of TERT and Fo − Fc density for the nucleic acid substrate at 3.0s contour level. We carried out model building in Coot58 and refined the model using both CNS-SOLVE and REFMAC559. We carried out the last cycles of refinement with TLS restraints as implemented in REFMAC5. We refined the structure to good stereochemistry, with 84.3, 14.3 and 1.4 of the residues in the most favorable, additional allowed and generously allowed of the Ramachandran plot, respectively.
54. Sasaki, T. & Fujiwara, H. Detection and distribution patterns of telomerase activity in insects. Eur. J. Biochem. 267, 3025–3031 (2000).
55. Holton, J. & Alber, T. Automated protein crystal structure determination using ELVES. Proc. Natl. Acad. Sci. USA 101, 1537–1542 (2004).
56. Potterton, E., Briggs, P., Turkenburg, M. & Dodson, E. A graphical user interface to the CCP4 program suite. Acta Crystallogr. D Biol. Crystallogr. 59, 1131–1137 (2003).
57. Brunger, A.T. et al. Crystallography & NMR system: A new software suite for macromolecular structure determination. Acta Crystallogr. D Biol. Crystallogr. 54, 905–921 (1998).
58. Emsley, P. & Cowtan, K. Coot: model-building tools for molecular graphics. Acta Crystallogr. D Biol. Crystallogr. 60, 2126–2132 (2004).
59. Murshudov, G.N., Vagin, A.A. & Dodson, E.J. Refinement of macromolecular structures by the maximum-likelihood method. Acta Crystallogr. D Biol. Crystallogr. 53, 240–255 (1997).
© 2
010
Nat
ure
Am
eric
a, In
c. A
ll ri
gh
ts r
eser
ved
.