structure–function constraints of tracheid-based xylem: a...
TRANSCRIPT

Structure–function constraints of tracheid-based xylem:a comparison of conifers and ferns
Jarmila Pittermann1, Emily Limm2, Christopher Rico1 and Mairgareth A. Christman3
1Department of Ecology and Evolutionary Biology, University of California, Santa Cruz, CA 95064, USA; 2Save the Redwoods League, 114 Sansome
St Suite 1200, San Francisco, CA 94104, USA; 3Institute for Ecohydrology Research, 1111 Kennedy Place Suite 4, Davis, CA 95616, USA
Author for correspondence:Jarmila Pittermann
Tel: +1 831 459 1782
Email: [email protected]
Received: 20 April 2011
Accepted: 6 June 2011
New Phytologist (2011) 192: 449–461doi: 10.1111/j.1469-8137.2011.03817.x
Key words: cavitation, gas exchange,hydraulic conductivity, primary xylem,sporophytes, xylem evolution.
Summary
• The ferns comprise one of the most ancient tracheophytic plant lineages, and
occupy habitats ranging from tundra to deserts and the equatorial tropics. Like
their nearest relatives the conifers, modern ferns possess tracheid-based xylem but
the structure–function relationships of fern xylem are poorly understood.
• Here, we sampled the fronds (megaphylls) of 16 species across the fern phylo-
geny, and examined the relationships among hydraulic transport, drought-induced
cavitation resistance, the xylem anatomy of the stipe, and the gas-exchange
response of the pinnae. For comparison, the results are presented alongside a
similar suite of conifer data.
• Fern xylem is as resistant to cavitation as conifer xylem, but exhibits none of the
hydraulic or structural trade-offs associated with resistance to cavitation. On a
conduit diameter basis, fern xylem can exhibit greater hydraulic efficiency than
conifer and angiosperm xylem.
• In ferns, wide and long tracheids compensate in part for the lack of secondary
xylem and allow ferns to exhibit transport rates on a par with those of conifers. We
suspect that it is the arrangement of the primary xylem, in addition to the intrinsic
traits of the conduits themselves, that may help explain the broad range of cavita-
tion resistance in ferns.
Introduction
The evolution of tracheid-based xylem in the LowerDevonian led to profound shifts in plant size and structure,and marked the first appearance of tracheophytes, theso-called true vascular plants (Pittermann, 2010; Kenrick& Crane, 1997; Niklas, 1992; Bateman et al., 1998; Sperry,2003). Tracheids preceded the widespread appearance ofvessels by an estimated 150 million yr and served as the fun-damental water transport tissue for some of the earliest landplants, including Rhynia and Psilophyton, horsetails, fernsand the extinct arborescent lineages of the Late Devoniansuch as the lycopod Lepidodendron and pro-gymnospermssuch as Archaeopteris (Cichan, 1985; Stewart & Rothwell,1993; Taylor et al., 2009). Generally, the evolution of trac-heids is characterized by increasing length and diameter(particularly during the Devonian), greater deposition ofsecondary cell wall material and progressive specialization inthe inter-tracheid pit membranes, the complexity of which
peaked with the appearance of the torus-margo pit mem-brane found in conifers, Gingko and some angiosperms(Niklas, 1985; Sperry, 2003; Pittermann, 2010; Pittermannet al., 2005; Jansen et al., 2004). The subsequent specializa-tion of tracheids into fibers and vessels that characterizedthe evolution of angiosperm wood allowed for a division oflabor whereby short, narrow fibers provide mechanical sup-port while multicellular vessels function solely for watertransport (Bailey & Tupper, 1918; Carlquist, 1988).Although vessels may confer water transport efficiencies thatare well over three orders of magnitude greater than those ofconifers (Tyree & Zimmermann, 2002; McCulloh et al.,2010), it is remarkable that tracheid-based xylem continuesto serve as the primary transport tissue for two abundant anddiverse plant lineages, the conifers and the ferns. We knowthat, on a xylem area basis, conifers and angiosperms can exhi-bit similar hydraulic efficiencies (Pittermann et al., 2005),but how does the performance of the tracheid-based xylem offerns compare with the more derived xylem of conifers?
NewPhytologist Research
� 2011 The Authors
New Phytologist � 2011 New Phytologist Trust
New Phytologist (2011) 192: 449–461 449www.newphytologist.com

Despite the extraordinary diversity and world-wide abun-dance of terrestrial and epiphytic ferns (Moran, 2008;Schuettpelz & Pryer, 2009), our understanding of the vas-cular performance of these primitive plants is just gainingmomentum (Calkin et al., 1985; Veres, 1990; Brodribbet al., 2005; Watkins et al., 2010). Early work on watertransport in several fern species showed that the main axisof the frond exhibits progressively lower hydraulic conduc-tivities along its length as a result of a decrease in conduitabundance and conduit size, especially from the start of theleafy rachis to the tip of the frond (Gibson et al., 1985;Schulte et al., 1987). These seemingly low rates of watertransport were again reported in a broad sampling of tropi-cal pteridophytes, a finding that was mirrored in theconcurrently low rates of gas exchange (Brodribb &Holbrook, 2004, Brodribb et al., 2007, Watkins et al.,2010). Low transpiration rates are consistent with the pref-erence of these tropical ferns for the low-light habitatscharacteristic of forest understories and dense canopies:because understory plants may only experience brief photo-synthetic peaks during sunflecks, selection places a lowerpremium on the evolution of high vascular and gas-exchange capacity in favor of a smaller sized, nonwoody,slow-growing life form with reduced metabolic costs. Thereare, however, some exceptions to these generalizations, mostnotably in the form of tree ferns, desert-dwelling ferns andthe many temperate species that appear to thrive in a broadvariety of temperate high-light habitats, such as Pteridiumaquilinum and Blechnum chilense (Page, 2002; Saldanaet al., 2007). Considering that several species of ferns, suchLygodium microphyllum and P. aquilinum, can be highlyinvasive (Robinson et al., 2010), it is not unreasonable tohypothesize that ferns are capable of high rates of watertransport and photosynthesis.
The vascular system in both fern fronds and rhizomesconsists of tracheids that tend to be longer and wider thanthose of conifers, with scalariform pitting extending alongthe entirety of at least one side of the tracheid wall (Gibsonet al., 1985; Veres, 1990; Carlquist & Schneider, 2001).Cryptic vessels have been reported in the rhizomes of severalspecies on the basis of what appeared to be scalariform per-foration plates in the terminal ends of vessel elements, buttheir frequency in the ferns is now presumed to be muchlower than originally thought (Carlquist & Schneider,2001, 2007). Because water transport in ferns occurs exclu-sively through primary vascular tissue, the xylem andphloem of ferns are encased in discrete bundles that mayspan the length of the frond and bifurcate at each pinna.The bundles are arranged in a variety of stelar patterns rang-ing from the simple protostele of whisk fern (Psilotumnudum) to the more complex siphonostele- and dictyostele-like arrangements found in more derived fern species.Secondary xylem is absent in extant ferns though extinctplants such as Medullosa, which bear some resemblance to
tree ferns, produced secondary xylem from a polystelicarrangement of several bifacial vascular cambia (Cichan,1986; Wilson et al., 2008; Taylor et al., 2009). Interestingly,a recent examination of two Botrychium species excludedthe possibility that these ferns exhibit true cambial-derivedsecondary growth, although the developmental pattern oftheir rhizomes is regarded as a departure from standarddefinitions of primary and secondary growth (Rothwell &Karrfalt, 2008).
Tracheid-based xylem is common to both conifers andferns, but key differences in xylem architecture have a pro-found effect on the overall structure of these plants, as wellas the physical principles that guide the shape and size ofthe xylem conduits. Most importantly, the evolution of abifacial vascular cambium and the resultant secondaryxylem in conifers and woody angiosperms marked a radicaldeparture in the evolution of xylem function as well as over-all plant structure because it allowed plants to have thearchitectural flexibility to vary the height and horizontal dis-play of their foliage, an otherwise impossible endeavor inplants limited by a unifacial cambium or simple strands ofprimary xylem (Rowe & Speck, 2005; Spicer & Groover,2010). In conifers and pro-gymnosperms, the xylemacquired the capacity not only to transport water to the leafcanopy, but also to structurally support it (Meyer-Berthaudet al., 1999; Tyree & Zimmermann, 2002; Pittermannet al., 2006a,b; Sperry et al., 2006; Pittermann, 2010).
Recent work has shown that, in north-temperate conifers,the combined requirements for canopy support alongwith reinforced, implosion-resistant tracheids constrain themaximum hydraulic efficiency of conifer xylem because ofthe necessity to build a strong, secondary cell wall coupledwith a narrower lumen diameter (Pittermann et al., 2006a;Sperry et al., 2006). Both the cell size and the volume ofthe conduit wall are limited by the metabolic output of thedeveloping xylem cell over the growing season, so it isimpossible for conifer tracheids to be long and wide andalso sufficiently fortified to offer structural support(Pittermann et al., 2006a; Sperry et al., 2006). By contrast,fern xylem is released from the structural support require-ment by virtue of the hypodermal sterome, a ring ofschlerenchyma fibers that surrounds the main axis of thefrond as well as the thicker secondary axes in some species(Rowe & Speck, 2004; Rowe et al., 2004). This tissuesupports the frond and provides it with a high degree offlexural stiffness (Niklas, 1992; Rowe et al., 2004).Consequently, without a support function, fern tracheidscan occupy a much broader morphospace in the form oflonger and wider tracheids that have little need for excep-tionally reinforced cell walls. However, fern tracheids lackthe torus-margo pitting particular to conifer tracheids andinstead possess the ancestral, homogenous pit membranethat confers much greater pit area resistance than the torus-margo arrangement, especially in derived angiosperms
450 Research
NewPhytologist
� 2011 The Authors
New Phytologist � 2011 New Phytologist Trust
New Phytologist (2011) 192: 449–461
www.newphytologist.com

(Sperry et al., 2005; Pittermann et al., 2005; but see Hackeet al., 2007).
Without torus-margo pit membranes, secondary xylemand a canopy support function, just how does the trans-port efficiency of the tracheid-based xylem of fernscompare to that of conifers? The goal of this study was toexamine the structure and function of fern xylem across abroad phylogenetic sampling scheme that includes self-supporting ferns, climbers, and perennial and seasonallydeciduous ferns as well as the desert-dwelling ferns inorder to capture the broadest possible variation in tracheidstructure. Because water transport occurs under negativepressure and xylem conduits are vulnerable to the entry air(cavitation), we examined how the cavitation resistance offern xylem compares to that of conifers and whether fernsexhibit any of the trade-offs associated with cavitationresistance such as reduced hydraulic efficiency andincreased fortification of tracheids against implosion, asobserved in north-temperate conifers (Pittermann et al.,2006a,b; Pittermann et al., 2010; Hacke et al., 2001;Sperry et al., 2006).
Materials and Methods
Plant material was gathered locally from campus forests at theUniversity of California in Santa Cruz (UCSC) or from glass-house collections at UCSC and UC Berkeley, USA. Frondswere collected from a minimum of four individuals (but onlythree individuals for the Lygodium species) from 16 speciesbelonging to several families and exhibiting a variety of lifehistory strategies, statures, and native habitats (Table 1).Species were sampled in May–August of 2009 and 2010.
Gas-exchange measurements
Gas-exchange measurements were performed on the samefronds used for the hydraulics and anatomical studiesdescribed below using the Li-Cor LI-6400XT portable gas-exchange system (Li-Cor Biosciences Inc., Lincoln, NE,USA) fitted with a standard opaque 2 · 3 cm LED cham-ber. All plants were watered to saturation the day before thegas-exchange measurements were collected such that thelowest midday water potentials were only )0.75 MPa, asmeasured using a standard pressure chamber (PMSInstruments, Corvallis, OR, USA). Each measurement wasobtained on mid-rachis pinnae, requiring between 2 and5 min for a stable reading, with leaf temperature at 21�C,the flow rate at 300 ml min)1 and the CO2 mixer set to400 lmol m)2 s)1. The chamber humidity ranged from 70to 80%, only slightly higher than ambient glasshousehumidity. For glasshouse-grown species, the minimumsaturating light intensity was 500 lmol m)2 s)1, but it wasincreased to 700 lmol m)2 s)1 to ensure that the pinnaewere indeed light saturated. Outdoor-growing, sun-exposed T
able
1A
list
of
spec
ies
use
din
this
study
and
thei
rco
llect
ion
site
s,gro
wth
hab
itan
dnat
ive
hab
itat
feat
ure
s
Spec
ies
Fam
ilySy
mbol
Colle
ctio
nsi
teH
abit
Endem
ichab
itat
Clim
ate
Adia
ntu
mca
pillu
s-veneri
sL.
Pte
ridac
eae
AC
Gla
sshouse
colle
ctio
nEr
ect
Nort
hA
mer
ican
war
m-t
emper
ate
fore
sts
Mes
icA
sple
niu
mbulb
ifer
um
G.Fo
rst
Asp
lenia
ceae
AB
Gla
sshouse
colle
ctio
nEr
ect
South
ern-h
emis
pher
ete
mper
ate
fore
sts
Mes
icC
heil
anth
es
bonari
ensi
s(W
illd)
Pte
ridac
eae
CB
Bota
nic
algar
den
Erec
tC
entr
alA
mer
ican
des
erts
Xer
icD
ickso
nia
anta
rcti
ca(L
abill
)D
isck
sonia
ceae
DA
Gla
sshouse
colle
ctio
nEr
ect
South
ern-h
emis
pher
ete
mper
ate
fore
sts
Mes
icD
ryopte
ris
arg
uta
(Kau
lf.)
Wat
tD
rypte
ridac
eae
DR
Coas
talR
edw
ood
fore
stEr
ect
Wes
tern
Nort
hA
mer
ican
tem
per
ate
fore
sts
Mes
icLygodiu
mflexu
osu
m(L
.)Sw
.Ly
godia
ceae
LFG
lass
house
colle
ctio
nC
limbin
gSo
uth
east
Asi
atr
opic
alfo
rest
sM
esic
Lygodiu
mja
ponic
a(T
hunb.)
Sw.
Lygodia
ceae
LJG
lass
house
colle
ctio
nC
limbin
gSo
uth
east
Asi
atr
opic
alfo
rest
sM
esic
Mic
role
pia
stri
gosa
(Thunb.)
C.Pre
slD
ennst
aedtiac
eae
MS
Gla
sshouse
colle
ctio
nEr
ect
Haw
aiia
nIs
land
tropic
alan
dsu
btr
opic
alfo
rest
sM
esic
Osm
unda
regalis
L.O
smundac
eae
OR
Bota
nic
algar
den
Erec
tEu
ropea
nN
ort
hA
fric
an,an
dEa
ster
nN
ort
hA
mer
ican
bogs
and
swam
ps
Mes
ic
Poly
podiu
maure
um
(L.)
J.Sm
.Poly
podia
ceae
PA
Gla
sshouse
colle
ctio
nEr
ect
Nort
han
dSo
uth
Am
eric
antr
opic
alan
dsu
btr
opic
alfo
rest
sM
esic
Poly
stic
hum
munit
um
(Kau
lf.)
C.Pre
slD
ryopte
ridac
eae
PM
Coas
talR
edw
ood
fore
stEr
ect
Wes
tern
Nort
hA
mer
ican
tem
per
ate
fore
sts
Mes
icPsi
lotu
mnudum
(L.)
Bea
uvo
isPsi
lota
ceae
PN
Gla
sshouse
colle
ctio
nEr
ect
Haw
aiia
nIs
land
tropic
alan
dsu
btr
opic
alre
gio
ns
Mes
icPte
ridiu
maquil
inum
(L.)
Kuhn
Pte
ridac
eae
PQ
UC
SCC
ampus
Erec
tTem
per
ate
and
subtr
opic
alre
gio
ns
Mes
icPte
ris
creti
ca(L
.)Pte
ridac
eae
PC
Gla
sshouse
colle
ctio
nEr
ect
South
ern
and
Nort
her
nH
emis
pher
ete
mper
ate
regio
ns
Mes
icPyrr
osi
ali
ngua
(Thunb.)
Farw
ell
Poly
podia
ceae
PL
Gla
sshouse
colle
ctio
nEr
ect
Asi
ante
mper
ate
regio
ns
Mes
icW
oodw
ard
iafim
bri
ata
Sm.
Ble
chnac
eae
WF
Gla
sshouse
colle
ctio
nEr
ect
Wes
tern
Nort
hA
mer
ican
tem
per
ate
fore
sts
Mes
ic
NewPhytologist Research 451
� 2011 The Authors
New Phytologist � 2011 New Phytologist Trust
New Phytologist (2011) 192: 449–461
www.newphytologist.com

species such as Cheilanthes bonariensis and P. aquilinumrequired the equivalent of full sun (2000 lmol m)2 s)1) toachieve saturation, while the understory fern Woodwardiafimbriata saturated at 700 lmol m)2 s)1. An average of 10measurements were made on glasshouse species (one to twomeasurements per frond), and that was doubled for out-door-grown ferns.
Conifer gas-exchange data were collected in May to lateJune in 2009 and 2010 from 20 individuals from thePinaceae, Cupressaceae, Podocarpaceae and Araucariaceaein the context of another project, and supplemented by datafrom the literature (Pittermann et al., 2006a,b). The gas-exchange measurements were performed by J.P. and E.L.on well-hydrated branches (the average midday waterpotential was )0.4 MPa) sampled from trees growing at theUC Santa Cruz Arboretum, San Francisco BotanicalGarden at Golden Gate Park and the UC Botanical Gardenin Berkeley, all locations in coastal, central California. Themeasurements were made as described in the previous para-graph, with a leaf temperature of 21�C, a flow rate of300 ml min)1, 400 lmol m)2 s)1 ambient CO2, chamberhumidity between 40 and 60% and a light intensity of2000 lmol m)2 s)1. Six measurements were made on fourto six stems collected from at least two trees, depending onavailability at the local arboreta.
Hydraulic measurements
The stipes represent the leafless portion of the fern frond, andunlike the frond rachis, the total xylem area is generallyinvariable along this segment. Stipes were collected from thebottom 18–20 cm of the frond for a sample size of n = 6–10.Stipe diameters ranged from 2 to 8 mm, with Lygodium sp.and Adiantum capillus-veneris possessing the narrowest stipesand P. aquilinum and W. fimbriata the widest. Stipe sampleswere re-cut under water to a length of 142 mm, and the distalends shaved smooth with a razor blade. Although all plantswere well hydrated before collection, any remaining embo-lism was removed by submersing the segments in distilledand filtered 20 mM KCl solution (0.22 lm; E-Pure filtra-tion system; Barnstead International, Dubuque, Iowa, USA)and degassing overnight under vacuum. Stipes were degassedrather than flushed in order to minimize handling damageand wound effects arising from compressed cortex tissue.
Hydraulic conductivity (k) was measured according tothe method of Sperry (1993) and calculated as the flow ratefor a given pressure gradient standardized per unit of stemlength. The stipes were mounted on a tubing apparatuswhere k was measured gravimetrically under a pressure of6–8 kPa using filtered 20 mM KCl solution. The flow ratethrough the segments was determined without a pressurehead before and after each gravimetric flow measurement.These background flows were averaged and subtracted fromthe pressure-induced flow in order to improve accuracy.
Functional xylem area was measured on samples perfusedfor several hours with basic fuchsin whereby hand-cut cross-sections were photographed and analyzed using IMAGEPRO
software (Media Cybernetics, Carlsbad, CA, USA). Xylemspecific conductivity (ks) and leaf specific conductivity (kL)represent k standardized for functional xylem area and distalleaf area, respectively.
Several workers have previously reported the presence ofmucilage that often inconvenienced hydraulic measure-ments on ferns, but with the exception of P. aquilinum andD. antarctica we did not experience this problem. In theaforementioned species, the stipe ends were carefully shavedto expose the xylem, and following a mild )0.5 MPa spinin the centrifuge (see below), the mucilage was eliminatedfrom the cut ends. All hydraulic measurements were madepromptly to minimize artifacts caused by wounding effects.
We used the centrifuge method to determine species’ vul-nerability to cavitation in response to a range of xylempressures (Pockman et al., 1995; Alder et al., 1997). Stemswere secured in a custom rotor designed to fit a Sorvall RC-5C centrifuge (Thermo Fisher Scientific, Waltham, MA,USA) and spun for 3 min at speeds that induce a knownxylem pressure (Px). The per cent loss of conductivity (PLC)caused by centrifugation at each Px was calculated from the kmeasured after spinning, relative to the maximum conduc-tivity after degassing (kmax) at Px = 0 Mpa, such that
PLC ¼ 100� ð1� k=kmaxÞ; Eqn 1
where kmax was determined at Px = 0 MPa following de-gassing. The segments were spun to progressively morenegative Px at )1 MPa increments until the PLC exceeded90%, or alternatively until Px = )10 MPa, which is themost negative Px that can be achieved using the centrifuge.A Weibull function was used to fit the vulnerability curvesfrom each stipe (Neufeld et al., 1992), and the xylem pres-sure at which segments exhibited a 50% loss of conductivity(P50) was computed as an average ± 1 SD per species.
Ferns such P. nudum and A. capillus-veneris had frondsthat were either too short or too weak to endure centrifuga-tion, so these species were subjected to the bench-drymethod of obtaining vulnerability curves on at least 15fronds. Intact fronds were cut and dehydrated down to arange of desired water potentials measured on mid-rachispinnae, at which point the stipes were removed and thehydraulic conductivity and PLC calculations were made asdescribed above. The data were fitted with a Weibull func-tion that was used to compute a single P50..
Similarly, we used the bench-dry method to verify thevalidity of the centrifuge-generated PLC data onP. aquilinum, a species known to possess vessels longer than142 mm in both the rhizome and the stipes (Carlquist& Schneider, 2007). Recent work on vines and ring-porous species suggests that the centrifuge method may
452 Research
NewPhytologist
� 2011 The Authors
New Phytologist � 2011 New Phytologist Trust
New Phytologist (2011) 192: 449–461
www.newphytologist.com
overestimate vulnerability to cavitation (Choat et al., 2010;Cochard et al., 2010), but we found that, in P. aquilinum,the two methods yielded vulnerability curves that wereindistinguishable from one another (data not shown).Similarly, we observed no differences in kmax onP. aquilinum stipes ranging in length from 14 to 22 cm,22 cm approximating the length of the longest infrequentvessel. We would expect kmax in the 14.2-cm segment to bemuch higher if the removal of conduit end walls reduced asignificant proportion of end-wall resistance.
Anatomical measurements
In north-temperate woody plants, the conduit double-wall-to-lumen-span ratio, (t ⁄ b)h
2, is positively correlated withcavitation resistance and reflects the potential of the xylemconduits to withstand implosion caused by negative waterpotentials (Hacke et al., 2001). This metric has since beenapplied to woody plants across a variety of habitats(Pittermann et al., 2006a; Jacobsen et al., 2007). Conduitdiameters and double-wall thickness:lumen span measure-ments were determined on xylem located in the center ofthe stipe previously used for hydraulic measurementsaccording to Pittermann et al. (2006a,b) and Hacke et al.(2001). The sections were treated with phloroglucinol tohighlight lignified tissue such as xylem, rinsed, andmounted in glycerin. All of the xylem was photographedunder 200–400· magnification with a digital cameramounted on a Motic BA400 compound microscope (JHTechnologies, San Jose, CA, USA). Because of the limitedamount of xylem present in the stipes, it was possible tomeasure all conduits located in each cross-section. Conduitfeatures were measured using IMAGEPRO analysis software.
Tracheid lumen areas were converted to equivalent circlediameters and the hydraulic mean diameter was calculatedfrom tracheid diameter distributions as Dh =
PD5 ⁄
PD4
according to Kolb & Sperry (1999). All conduits were mea-sured, with the sample size ranging from from 70 to 300conduits depending on species xylem area. The thick-ness:span ratio was measured on adjacent conduits where atleast one cell (although usually both) was within 10% ofDh. The double-wall thickness was determined from theshared walls of at least 40 and up to 100 of these cells.
Conduit length and diameter were measured on individ-ual tracheids obtained from macerations according to themethods of Mauseth & Fujii (1994). Individual vascularbundles at least 5 cm in length were excised from the stipeand submersed in a 50 : 50 solution of 30% hydrogen per-oxide and 80% glacial acetic acid (Sigma-Aldrich, St. Louis,MO, USA), and heated to at least 110�C for c. 2–3 d.IMAGEPRO was used to measure at least 50 tracheids per spe-cies at ·200–400.
Anatomical data from conifer root and shoot xylem werecollected from previously published work by Pittermann
et al. (2006a,b), as well as unpublished data collected in2008–2010 by J. P.
Conduit length measurements
We used the silicone injection method of Hacke et al.(2007) and Christman et al. (2009) to obtain conduitlength distributions in species of ferns suspected to possesslong conduits such as Lygodium flexuosum and P. aquilinum(Carlquist & Schneider, 2007). Five fronds of each specieswere injected basipetally from the base of the stipe with asilicone-fluorescent dye-hardener mix at a pressure of50 kPa, and left overnight (Christman et al., 2009). Thesilicone was hardened over 3 d, after which the stipes weresectioned at regular intervals starting at 5 mm from the baseof the injection site. In P. aquilinum, which is known topossess vessels, the increments were progressively increasedfrom 5 mm to 5 cm. The fraction of silicone-filled conduitsat each length increment was counted and the data analyzedaccording to Christman et al. (2009).
Results
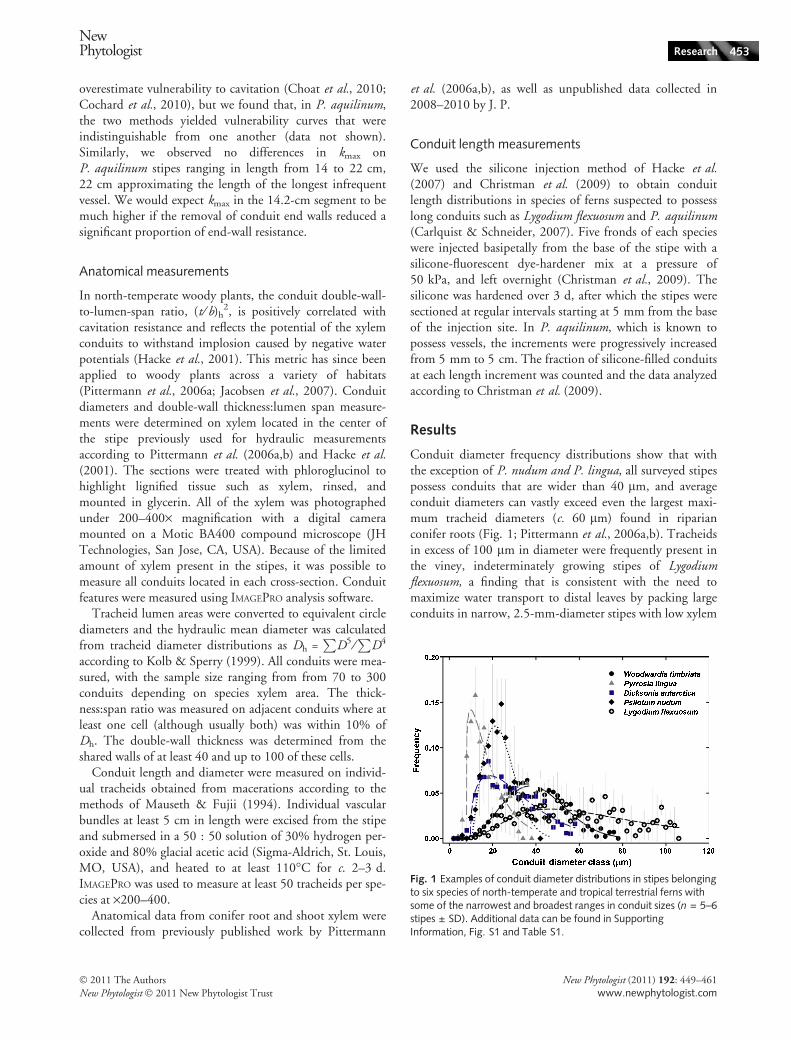
Conduit diameter frequency distributions show that withthe exception of P. nudum and P. lingua, all surveyed stipespossess conduits that are wider than 40 lm, and averageconduit diameters can vastly exceed even the largest maxi-mum tracheid diameters (c. 60 lm) found in riparianconifer roots (Fig. 1; Pittermann et al., 2006a,b). Tracheidsin excess of 100 lm in diameter were frequently present inthe viney, indeterminately growing stipes of Lygodiumflexuosum, a finding that is consistent with the need tomaximize water transport to distal leaves by packing largeconduits in narrow, 2.5-mm-diameter stipes with low xylem
Fig. 1 Examples of conduit diameter distributions in stipes belongingto six species of north-temperate and tropical terrestrial ferns withsome of the narrowest and broadest ranges in conduit sizes (n = 5–6stipes ± SD). Additional data can be found in SupportingInformation, Fig. S1 and Table S1.
NewPhytologist Research 453
� 2011 The Authors
New Phytologist � 2011 New Phytologist Trust
New Phytologist (2011) 192: 449–461
www.newphytologist.com

areas. The observed wide range in conduit diameterfrequencies explains the significant standard deviations asso-ciated with mean and hydraulic tracheid diameter valuesreported in figures below.
We generated vulnerability curves on stipes from all spe-cies examined to determine how susceptible ferns are todrought-induced cavitation given the tremendous variationobserved in conduit dimensions and xylem stele arrange-ments (Fig. 2). Fig. 3 shows vulnerability curves in sixspecies that span a broad range of cavitation resistances,from a P50 of –0.68 MPa in P. nudum to a surprising)11.8 ± 7.6 MPa (mean ± SD) in Dryopteris arguta. Inseveral fronds of D. arguta, the P50 was extrapolated fromthe Weibull fits assigned to each vulnerability curve.Surprisingly, several D. arguta stipes showed < 60% loss ofhydraulic conductivity at )9 MPa but had a tendency toshred when subjected to )10 MPa xylem pressures in thecentrifuge, so data at this pressure are not reported.
We observed no hydraulic or structural xylem trade-offsassociated with species’ cavitation resistance, in contrast totrends evident across a broad sampling of the conifer xylemresponse (Fig. 4). Indeed, cavitation resistance in coniferscomes at the cost of reduced conduit diameters and thicker
conduit walls, both of which tend to lower hydraulicconductivity, especially within the Pinaceae and north-temperate Cupressaceae (Pittermann et al., 2006a,b), but
(a) (b) (c)
(d) (e) (f)
Fig. 2 Mid-segment cross-sections of six fern stipes, stained with toluidine blue, mounted in glycerol and photographed under a compoundmicroscope. Bars, 1 mm. These species were chosen on the basis of their cavitation resistance and the arrangement of their primary xylem,beginning with species that exhibit the least cavitation resistant and integrated xylem and ending with those with the most resistant andsectored xylem: (a) Psilotum nudum, P50 = )0.69 MPa; (b) Pteridium aquilinum, P50 = )1.05 ± 0.53 MPa; (c) Dicksonia antarctica,P50 = )0.96 ± 0.45 MPa; (d) Cheilanthes bonariensis, P50 = )3.18 MPa; (e) Lygodium flexuosum, P50 = )3.97 ± 1.88 MPa; and (f)Dryopteris arguta, P50 = )11.8 ± 7.6 MPa. The arrangement of L. flexuosum tracheids within the stele is unusual because parenchyma cellsseparate the conduits from one another. A thick ring of schlerenchyma fibers known as the hypodermal sterome (Rowe & Speck, 2005) islocated just below the epidermis in all cross-sections. In addition to its mechanical role, the sterome may also aid in preventing water loss.
Fig. 3 The hydraulic response to increasingly negative waterpotential in six ferns representing the most vulnerable and the mostresistant species to drought-induced cavitation. Because of its shortstature, the vulnerability curve in Psilotum nudum stipes wasobtained using the bench-dry method while other species wereevaluated using the centrifugal method of Alder et al. (1997).
454 Research
NewPhytologist
� 2011 The Authors
New Phytologist � 2011 New Phytologist Trust
New Phytologist (2011) 192: 449–461
www.newphytologist.com

no such costs were apparent in fern xylem. We can, how-ever, make the generalization that cavitation resistance inferns is on par with cavitation resistance in conifers.
Despite showing a broad range of P50 values, fern tracheidallometry is surprisingly different from conifer tracheids(Fig. 5). In conifers, the tracheid double-wall thickness totracheid lumen diameter ratio (t ⁄ b)h
2 is a proxy of tracheidresistance to implosion, and this trait strongly correlateswith cavitation resistance in the north-temperate Pinaceaeand Cupressaceae. In these two families, higher (t ⁄ b)h
2 ratios
are found in increasingly drought-resistant species, indicat-ing greater conduit strength (Fig. 5; Hacke et al., 2001;Pittermann et al., 2006a). However, conifer tracheids alsotend to exhibit more reinforcement than strictly necessary, atrait interpreted as a safety factor protecting against implo-sion, which is shown by the relationship between the (t ⁄ b)h
2
ratio and species’ P50 (Fig. 5). By contrast, (t ⁄ b)h2 ratios in
fern conduits are decoupled from cavitation resistance andexhibit no safety factor from implosion. Thus, fern conduitsmay show some degree of mechanical flexibility. The outlierin Fig. 5 is P. nudum, a basal filicopsid fern, which, despiteshowing modestly reinforced conduits, is the least cavita-tion-resistant species sampled.
Several studies indicate that photosynthesis in pterido-phytes is inherently constrained by the low hydraulicefficiency characteristic of this group (Brodribb et al., 2005;Watkins et al., 2010). Although our photosynthesis and
(a)
(b)
(c)
Fig. 4 The relationship between the xylem pressure causing a 50%loss of hydraulic conductivity as a result of cavitation (P50) andxylem specific conductivity (a), conduit hydraulic diameter (b) andconduit double-wall thickness (c) in conifers (circles) and ferns(letters; see Table 1 for species abbreviations). In contrast toconifers, fern xylem showed no hydraulic or structural trade-offsassociated with P50. ns, not significant.
(a)
(b)
Fig. 5 Conduit double-wall thickness plotted as a function ofconduit hydraulic diameter in conifers (circles) and ferns (letters; seeTable 1 for species abbreviations) (a). In contrast to conifers, weobserved in fern conduits a strong, positive correlation betweenconduit hydraulic diameter and double-wall thickness. Surprisingly,this scaling results in a low (t ⁄ b)h
2 ratio (b) that exhibits no safetyfactor from implosion, even under very negative xylem pressures(ns, not significant).
NewPhytologist Research 455
� 2011 The Authors
New Phytologist � 2011 New Phytologist Trust
New Phytologist (2011) 192: 449–461
www.newphytologist.com

stomatal conductance rates agree with previous studies (seealso Watkins et al., 2010), it is important to point out that,across a broad sampling of ferns including climbing as wellas north-temperate species, photosynthesis and stomatalconductance rates can equal or exceed those of evergreenconifers (Fig. 6). We attribute this result in part to theferns’ much wider and longer conduits which account forthe surprisingly high hydraulic conductivity of fern xylem,and which can, in turn, support a broad range of stomatalconductance rates. Indeed, it is likely that the ferns’ largeconduit volumes compensate for the ferns’ small functionalxylem areas that are two orders of magnitude smaller thanthose of conifers, despite serving a similar amount of distalfoliage (Figs 7, 8). If evolution acted as expected to increasehydraulic efficiency in the xylem-limited ferns, thenP. aquilinum represents the pinnacle of hydraulic selectionin fronds as it is the one species in our pool known to con-tain vessels in both fronds and rhizomes (not shown), withsome vessels exceeding 20 cm in length (Fig. 7).
The hydraulic consequences of the high-volume tracheidsof ferns are readily apparent when xylem conductivity isexpressed per leaf and functional xylem area, kL and kx,respectively, and compared with equivalent data in conifers(Fig. 9). The similar kL values in conifers and ferns supportthe finding that leaf-level function in both plant groups canexhibit a similar range of assimilation and conductance rates(Fig. 6).
Lastly, our data indicate that fern xylem can exhibit lowertransport resistivity (rx, the inverse of kx) for a given conduitdiameter than the xylem of the more derived conifer
(a)
(b)
Fig. 6 Photosynthesis (a) and stomatal conductance (b) as afunction of hydraulic conduit diameter in conifers (circles) and ferns(letters; see Table 1 for species abbreviations). Cheilanthes
bonariensis (CB), an outdoor-grown desert fern, exhibited thehighest rates of gas exchange, consistent with its thicker pinnaewhich presumably provide some degree of capacitance. The highestphotosynthesis and conductance rates in the conifers belong tothree deciduous species of Cupressaceae. ns, not significant.
Fig. 7 Average conduit length plotted as a function of conduitdiameter in conifers (circles) and ferns (letters; see Table 1 forspecies abbreviations). Assuming a cylindrical geometry, ferntracheid volumes are 8 to 60· greater than those of conifers. Thisfactor would be considerably higher in Pteridium aquilinum, wherevessel length exceeds 20 cm.
Fig. 8 Fern xylem supports similar leaf areas as conifer xylem despiterelying on primary xylem with a total area that is two orders ofmagnitude smaller than that of conifers. The tight scaling betweenxylem and leaf area in ferns is likely to reflect the limited venationand low capacitance of fern leaves (Brodribb et al., 2005, 2007), aresult consistent with the constant scaling between stipe length andleaf area observed in Polystichum munitum (Limm & Dawson,2010). In the absence of the indeterminately growing fronds ofLygodium japonica, the r2 of the regression increases to 0.61 andP < 0.001. Conifers are indicated by circles and ferns by letters (seeTable 1 for species abbreviations).
456 Research
NewPhytologist
� 2011 The Authors
New Phytologist � 2011 New Phytologist Trust
New Phytologist (2011) 192: 449–461
www.newphytologist.com

and angiosperm lineages (Fig. 10; additional data fromPittermann et al., 2005). Put another way, when wecompared the area-specific xylem resistivities of fern, conifer,vine, and ring- and diffuse-porous species with equivalentmean conduit sizes, fern xylem exhibited higher transportefficiencies than the xylems of the remaining plant groupswith the exception of vines, which had a mean conduit diam-eter of 150 lm and an rx of 13 MPa s m)2 (Fig. 10).
Discussion
The greater xylem specific conductivity observed in thexylem of fern stipes relative to conifer xylem can be attrib-uted in part to the ferns’ longer and wider tracheids. Largelumen diameters reduce the frictional resistance to watertransport that arises from conduit walls, while increasingconduit length reduces the frictional resistance attributableto pit membrane crossings from one conduit to the next(Sperry et al., 2006). In ferns, high-volume conduits werepresumably free to arise because the xylem serves solely in ahydraulic capacity, in contrast to the double-duty xylem ofconifers where tracheids not only deliver water to the can-opy but also support it (Pittermann et al., 2006a,b). Thissupport function reduces conifer tracheid diameter andlength, and frequently calls for extra wall thickening, asobserved in compression wood (Spicer & Gartner, 2002).In ferns, however, the presence of the hypodermal sterome(Fig. 2) relaxes the support constraints imposed upon thetracheids, such that high-volume conduits function solely inthe efficient transport of water. An analogous situationoccurs in the xylem of woody angisperms, where thick-walled fibers constitute the structural matrix that supportsthe canopy, leaving the hydraulic function to the muchlonger vessels (Hacke et al., 2001; Sperry et al., 2006), aswell as in leaves, which are supported by turgor pressureand collenchyma. In conifers, root xylem is also releasedfrom the support function and characteristically exhibitstracheids that are longer, wider and more conductive(Pittermann et al., 2006a). The primary xylem of coniferseedlings and juvenile shoots is developmentally most anal-ogous to fern xylem, but whether or not the structure–function patterns of primary tracheids resemble those ofconifer roots or ferns is unknown.
Fern xylem may be hydraulically efficient, but it is pri-mary xylem nonetheless and is subject to limits imposed bya basal developmental program. First, the overall structureof ferns is constrained by the lack of a vascular cambium,which gives rise to lateral branching and secondary xylem(Rowe & Speck, 2004; Spicer & Groover, 2010). Theabsence of wood imposes a fundamental limit on fern sizeand canopy development even in tree ferns, which raisetheir relatively modest canopy by virture of anomalous sec-ondary growth courtesy of roots and fibers. Secondly, fernspossess much smaller xylem areas in the absence of a cam-
Fig. 10 Xylem specific resistivity plotted as a function of hydraulicconduit diameter in conifers (closed circles), angiosperms (opencircles) and ferns (letters; see Table 1 for species abbreviations).Some conifer and angiosperm data were re-plotted from the originalcomparison of tracheid vs vessel transport efficiency in Pittermannet al. (2005).
(a)
(b)
Fig. 9 Leaf (a) and xylem (b) specific conductivities plotted as afunction of hydraulic conduit diameter in conifers (circles) and ferns(letters; see Table 1 for species abbreviations). Species’ transportefficiency is a function of xylem conduit diameter in both conifersand ferns. The large discrepancy in kL values between Lygodium
flexuosum and Lygodium japonica can be attributed to theindeterminately growing habit of the fronds, which varied between0.5 and 1.5 m in length.
NewPhytologist Research 457
� 2011 The Authors
New Phytologist � 2011 New Phytologist Trust
New Phytologist (2011) 192: 449–461
www.newphytologist.com

bium than conifers, despite supporting an equivalentamount of foliage and similar rates of gas exchange(Figs 8,9). Hence, the observed frond allometry in Fig. 8 isthe result of a limited vascular system that supplies water insufficient quantities for transpiration, but probably lacks thebuffer against fluctuations in water availability that capaci-tance provides in woody plants. The reliance on primaryxylem for water transport may explain why ferns exhibitmore rapid stomatal closure than angiosperm leaves(Brodribb & Holbrook, 2004).
The extent to which pit membrane characteristics con-tribute to fern hydraulic efficiency is largely undetermined.However, the few studies that addressed the relative contri-butions of pit and lumen resistance in fern xylem indicatethat pit membranes in fern tracheids account for 36 to 47%of total tracheid resistance (Calkin et al., 1985; Schulteet al., 1987). By contrast, conifer and angiosperm pitmembranes confer considerably greater resistance to flow,accounting for c. 64 and 56% of conduit resistance, respec-tively (Wheeler et al., 2005; Pittermann et al., 2006b;Sperry et al., 2006). Whether or not reduced pit membraneresistance in ferns is attributable to greater porosity and ⁄ ora higher fraction of conduit pit area remains to be seen, butsome evidence suggests that fern pit membranes not onlymay be quite porous, but may extend along the full lengthof the tracheid wall (Carlquist & Schneider, 2007).Selection has been shown to act on pit membrane area aswell as porosity, and both traits are thought to have a pro-found effect on hydraulic efficiency and cavitation by theentry of air, otherwise known as ‘air-seeding’ (Choat et al.,2008; Jansen et al., 2009; Lens et al., 2011). Certainly,greater pit area and membrane porosity should compensatefor xylem comprised of single-celled conduits lacking thelow-resistance torus-margo pit membrane (Pittermannet al., 2005; Sperry et al., 2006). Interestingly, the homo-geous pit membranes found in the tracheids of vessellessangiosperms including the Amborellaceae, Winteraceae andTrochodendrales exhibit considerably higher porosity thanthose of eudicots, a finding that is consistent with the low,conifer-like area-specific pit resistance in this basal lineage(Hacke et al., 2007). Similar to these basal dicots, fernxylem exhibits hydraulic conductivities that are at least 38·greater than predicted for tracheids lacking conifer-typetorus-margo pits (Pittermann et al., 2005; Hacke et al.,2007). Whether Botrichium dissectum, an Ophioglossid fernwith torus-margo pit membranes (Morrow & Dute, 1998),exhibits comparable hydraulic efficiencies that are closer toconifers or ferns remains to be seen.
Recent work indicates that pit membrane features controlcavitation resistance in both conifers and angiosperms(Christman et al., 2009; Pittermann et al., 2010; Lenset al., 2011; Delzon et al., 2010), but what controls cavita-tion resistance in ferns? Currently, we can entertain one or acombination of two possibilities: the fern pit membrane
may exhibit variable porosity that corresponds to species’resistance to air entry air-seeding, as previously mentioned;and the integration of the vascular bundles, specifically thefrequency of conduit contact between two or more vascularbundles, affects the rate at which air spreads through thexylem. Empirical and theoretical work on woody plants hasshown that hydraulically integrated vascular networks areassociated with increased hydraulic conductivity, but at thecost of more rapid spread of air-seeding-induced embolism(Zanne et al., 2006; Loepfe et al., 2007; Schenk et al.,2008). Looking across the stelar patterns of the sampledfern species (Fig. 2), we suspect that cavitation resistance inthese plants is controlled by a similar, spatially determinedvascular arrangement that may inhibit or facilitate thespread of air through the conduit network dependingon conduit-to-conduit or bundle-to-bundle contact. Forexample, hydraulically integrated xylem belonging toP. aquilinum and P. nudum was the most vulnerable tocavitation, while xylem belonging to the cavitation-resistantD. arguta and P. cretica was packaged in a limited numberof spatially separated (sectored) vascular bundles (Fig. 2).Variation in P50 values was greater in species with sectoredxylem, presumably because of irregular and as of yet un-characterized bundle-to-bundle connectivity over the lengthof the sampled stipe segments. A combination of methodsincluding dye perfusions and high-resolution computedtomography (HRCT) could be used to construct three-dimensional renderings of variable xylem patterns along thelength of the frond and as well as in different species(Schulte et al., 1987; Brodersen et al., 2010).
Perhaps a more fundamental question than what controlsair-seeding in ferns is what selects for cavitation resistancein these overwhelmingly mesophylic plants? Watkins et al.(2010) show that tropical epiphytic ferns are more cavita-tion resistant than terrestrial species, a result consistent withthe epiphytes’ lower midday water potentials. In our assort-ment of terrestrial ferns, the issue may also be complicatedby ecology and life history strategy. For example,P. aquilinum is a drought-deciduous fern, and exhibitsxylem with high hydraulic conductivity and low cavitationresistance. This hydraulic ‘boom and bust’ strategy is similarto the phenology and physiology of deciduous ring-porousor riparian trees. By comparison, D. arguta is a semi-perennialspecies that retains fronds for at least 2 yr, during which itendures the dry, mediterranean summer climate of coastalCalifornia. In addition to drought stress, cavitation resist-ance in ferns is probably associated with frond longevity.An extreme example of this is L. flexuosum, the fronds ofwhich grow indeterminately to lengths well beyond 2 m.Although this species is native to mesic, subtropical habi-tats, its stipe is surprisingly cavitation resistant, presumablyto ensure water transport to distal pinnae. Whether or notreproductive, spore-bearing fronds show alternative hydrau-lic strategies is an open question.
458 Research
NewPhytologist
� 2011 The Authors
New Phytologist � 2011 New Phytologist Trust
New Phytologist (2011) 192: 449–461
www.newphytologist.com

Despite the high cavitation resistance exhibited by anumber of fern species, the tracheid allometry of ferns isconsiderably different from conduits in conifers and woodyangiosperms. In north-temperate conifers, the trend towardincreased implosion resistance comes courtesy of wall thick-ness:lumen diameter ratios that increase with P50, but doso by reducing tracheid diameter rather than fortifyingthe tracheid walls (Pittermann et al., 2006a; Fig. 5).Consequently, reduced transport efficiency constitutes thefundamental hydraulic trade-off associated with cavitationresistance in north-temperate conifers (Pittermann et al.,2006a,b; Fig. 4). These safety–efficiency trade-offs are alsoapparent at the level of the pit, whereby increased cavitationresistance is associated with reduced pit conductivity acrossa range of conifer species and within a single tree(Pittermann et al., 2006b, 2010; Domec et al., 2008). Bycontrast, we found no hydraulic or structural ‘investment’trade-offs in vulnerable vs cavitation-resistant ferns, similarto the results of Watkins et al. (2010). This is consistentwith the weaker relationship between vessel diameter andP50 observed in angiosperms, and the probabilistic role thatpit porosity plays in controlling cavitation resistance in di-cots (Wheeler et al., 2005; Hacke et al., 2007; Christmanet al., 2009; Lens et al., 2011). Across the pteridophytes,the costs of drought resistance may be reflected in theincreased production of osmolytes and proteins that protectliving tissues from low water potentials rather than xylemstructure per se (Proctor & Tuba, 2002).
In ferns, larger conduit diameters scale directly withgreater wall thickness (Fig. 5). Interestingly, despite theunusually thick tracheid walls found in several ferns, therelationship between species’ P50 and the conduit (t ⁄ b)h
2
ratio falls directly on the predicted conduit implosion line –a dramatic departure from the pattern observed in conifertracheids. Conduit implosion thresholds are computedwithout k, the safety factor from cell collapse, so our dataindicate two things: first, ferns do not allocate any moreresources to their xylem than is necessary to maintain acircular geometry, and second, the conduit (t ⁄ b)h
2 ratiocannot be used to infer cavitation resistance in these plants.Fern tracheids may exhibit some degree of flexibility to tol-erate bending or torsion stress associated with life in theunderstory or to facilitate the climbing habit. To wit, thelargest tracheids in the climber L. flexuosum are located inthe stele center, that is, the neutral axis region, which expe-riences minimal tensile and compressive stress (Niklas,1992). Also, flexible tracheids may accommodate shapechange in response to the tension imposed on them by thewater column. A good example of this phenomenon istransfusion tracheids in conifer leaves and needles. Theseunlignified tracheids flex and collapse under negative xylempressure, presumably to avoid cavitation and to facilitaterapid refilling (Cochard et al., 2004; Brodribb &Holbrook, 2005). Similarly distorted tracheids have been
observed in the xylem of variably dehydrated fronds(J. Pittermann & E. Limm, unpublished), and it is notunreasonable to suspect that, in the very cavitation-resistantspecies, the reductions in hydraulic conductivity withprogressively more negative water potentials may beattributable to conduit distortion, rather than embolism.The aforementioned HRCT method is nondestructive, andcould provide a window into the intricacies of fern vascularfunction in response to drought and rewatering in a liveplant (Brodersen et al., 2010).
The extent to which we understand variation in xylemstructure and function across tracheophytes will affect theinferences we draw about the evolution of transport tissueacross both extinct and extant lineages, including theLycopodiales, Sellaginales and perhaps even some bryo-phytes. For example, empirically based biophysical models ofxylem performance have provided important insights into thexylem function of the earliest land plants as well as the morederived seed ferns, pro-gymnosperms and other lineages withtracheid-based xylem (Wilson et al., 2008; Wilson andFischer, 2010; Wilson & Knoll, 2010). These modelssuggest that, far ahead of the appearance of heteroxylousangiosperm xylem, xylem evolution in ancient plants wasconsistently guided by selection for hydraulic efficiency,cavitation resistance and, eventually, mechanical support.Our data show that extant ferns have succeeded in master-ing these three essentials, although not in the manner onewould predict based on the structure–function modelderived from woody plants. The physiological resilienceof ferns may go a long way toward explaining why theyhave persisted across nearly all parts of the globe despiteseveral mass extinctions and the intense competitive pres-sure exerted by angiosperms (Schneider et al., 2004;Schuettpelz & Pryer, 2009; Watkins et al., 2010).
Acknowledgements
We are grateful to the NSF (IOS-1027410; J.P.) and Savethe Redwoods League (E.L. and J.P.) for supporting thisresearch. C.R. received funding through an NSF REU grantand from the Save the Redwoods league. M.A.C. was sup-ported by NSF-IBN-0743148 to John Sperry (University ofUtah). The conifer data were collected with support fromthe Miller Institute for Basic Research (UC Berkeley; J.P.)and assistance from Todd Dawson. We wish to thank LucyLynn, Stephanie Ko and Shervin Bastami for contributingto the xylem anatomical measurements, and Duncan Smithand John Sperry for kindly helping with the silicone injec-tion method. Holly Forbes, Mona Bourrell and the staff atthe University of California Botanical Garden (Berkeley,CA) and the San Francisco Botanical Garden kindly pro-vided access to plant material, as did Jim Velzy and DenisePolk (UC Santa Cruz), who also assisted with propagationand glasshouse space. We also thank three anonymous
NewPhytologist Research 459
� 2011 The Authors
New Phytologist � 2011 New Phytologist Trust
New Phytologist (2011) 192: 449–461
www.newphytologist.com

reviewers whose insightful comments helped improve themanuscript.
References
Alder NN, Pockman WT, Sperry JS, Nuismer S. 1997. Use of centrifugal
force in the study of xylem cavitation. Journal of Experimental Botany 48:
665–674.
Bailey IW, Tupper WW. 1918. Size variation in tracheary cells: I. A
comparison between the secondary xylems of vascular cryptogams,
gymnosperms, and angiosperms. American Academy of Arts SciencesProceedings 54: 149–204.
Bateman RM, Crane PR, DiMichele WA, Kenrick PR, Rowe NP, Speck
T, Stein W. 1998. Early evolution of land plants: phylogeny, physiology
and ecology of the primary terrestrial radiation. Annual Review ofEcology, Evolution, and Systematics 29: 263–292.
Brodersen CR, McElrone AJ, Choat B, Matthews MA, Shackel KA.
2010. The dynamics of embolism repair in xylem: in vivo visualizations
using high-resolution computed tomography. Plant Physiology 154:
1088–1095.
Brodribb TJ, Feild TS, Jordan GJ. 2007. Leaf maximum photosynthetic
rate and venation are linked by hydraulics. Plant Physiology 144:
1890–1898.
Brodribb TJ, Holbrook NM. 2004. Stomatal protection against hydraulic
failure: a comparison of co-existing ferns and angiosperms. NewPhytologist 162: 663–670.
Brodribb TJ, Holbrook NM. 2005. Water stress deforms tracheids
peripheral to the leaf vein of a tropical conifer. Plant Physiology 137:
1139–1146.
Brodribb TJ, Holbrook NM, Zwieniecki MA, Palma B. 2005. Leaf
hydraulic capacity in ferns, conifers and angiosperms: impacts on
photosynthetic maxima. New Phytologist 165: 839–846.
Calkin HW, Gibson AC, Nobel PS. 1985. Xylem water potentials and
hydraulic conductances in eight species of ferns. Canadian Journal ofBotany 63: 632–637.
Carlquist S. 1988. Comparative wood anatomy. Berlin, Germany: Springer-
Verlag.
Carlquist S, Schneider EL. 2001. Vessels in ferns: structural, ecological
and evolutionary significance. American Journal of Botany 88: 1–13.
Carlquist S, Schneider EL. 2007. Tracheary elements in ferns: new
techniques, observations and concepts. American Fern Journal 97:
199–211.
Choat B, Cobb AR, Jansen S. 2008. Structure and function of bordered
pits: new discoveries and impacts on whole-plant hydraulic function.
New Phytologist 177: 608–626.
Choat B, Drayton WM, Brodersen C, Matthews MA, Shackel KA, Wada
H, McElrone AJ. 2010. Measurement of vulnerability to water stress-
induced cavitation in grapevine: a comparison of four techniques
applied to a long-vesseled species. Plant, Cell & Environment 33:
1502–1512.
Christman MA, Sperry JS, Adler F. 2009. Testing the ‘rare pit’ hypothesis
for xylem cavitation resistance in three species of Acer. New Phytologist182: 664–674.
Cichan MA. 1985. Vascular cambium and wood development in
Carboniferous plants. I. Lepidodendrales. American Journal of Botany72: 1163–1176.
Cichan MA. 1986. Vascular cambium and wood development in
Carboniferous plants. IV. Seed plants. Botanical Gazette 147: 227–235.
Cochard H, Froux F, Mayr S, Coutand C. 2004. Xylem wall collapse in
water-stressed pine needles. Plant Physiology 134: 401–408.
Cochard H, Herbette S, Barigah T, Badel E, Ennajeh M, Vilagrosa A.
2010. Does sample length influence the shape of xylem embolism
vulnerability curves? A test with the Cavitron spinning technique Plant,Cell & Environment 33: 1543–1552.
Delzon S, Douthe C, Sala A, Cochard H. 2010. Mechanism of water-
stress induced cavitation in conifers: bordered pit structure and function
support the hypothesis of seal capillary-seeding. Plant, Cell &Environment 33: 2101–2111.
Domec J-C, Lachenbruch B, Meinzer F, Woodruff D, Warren JM,
McCulloh K. 2008. Maximum height in a conifer is associated with
conflicting requirements for xylem design. Proceedings of the NationalAcademy of Sciences, USA 105: 12069–12074.
Gibson AC, Calkin HW, Raphael DO, Nobel PS. 1985. Water relations
and xylem anatomy of ferns. Proceedings of the Royal Society of Edinburgh86B: 81–92.
Hacke UG, Sperry JS, Feild TS, Sano Y, Sikkema EH, Pittermann J.
2007. Water transport in vesselless angiosperms: conducting efficiency
and cavitation safety. International Journal of Plant Sciences 168:
1113–1126.
Hacke UG, Sperry JS, Pockman WT, Davis SD, McCulloh KA. 2001.
Trends in wood density and structure are linked to prevention of xylem
implosion by negative pressure. Oecologia 126: 457–461.
Jacobsen AL, Pratt RB, Ewers FW, Davis SD. 2007. Cavitation resistance
among 26 chaparral species of southern California. EcologicalMonographs 77: 99–115.
Jansen S, Choat B, Pletsers A. 2009. Morphological variation of
intervessel pit membranes and implications to xylem function in
angiosperms. American Journal of Botany 96: 409–419.
Jansen S, Choat B, Vinckier S, Lens F, Schols P, Smets E. 2004.
Intervascular pit membranes with a torus in the wood of Ulmus(Ulmaceae) and related genera. New Phytologist 163: 51–59.
Kenrick P, Crane PR. 1997. The origin and early evolution of plants on
land. Nature 389: 33–39.
Kolb KJ, Sperry JS. 1999. Differences in drought adaptation
between subspecies of sagebrush (Artemisia tridentata). Ecology 80:
2373–2384.
Lens F, Sperry JS, Christman MA, Choat B, Rabaey D, Jansen S. 2011.
Testing hypotheses that link wood anatomy to cavitation resistance
and hydraulic conductivity in the genus Acer. New Phytologist 190:
709–723.
Limm EB, Dawson TE. 2010. Polystichum munitum (Dryopteridaceae)
varies geographically in its capacity to absorb fog water by foliar uptake
within the redwood forest ecosystem. American Journal of Botany 97:
1121–1128.
Loepfe L, Martinez-Vilalta J, Pinol J, Mencuccini M. 2007. The relevance
of xylem network structure for plant hydraulic efficiency and safety.
Journal Theoretical Biology 247: 788–803.
Mauseth JD, Fujii T. 1994. Resin-casting: a method for investigating
apoplastic spaces. American Journal of Botany 81: 104–110.
McCulloh K, Sperry JS, Lachenbruch B, Meinzer FC, Reich PB, Voelker
S. 2010. Moving water well: comparing hydraulic efficiency in twigs and
trunks of coniferous, ring-porous, and diffuse-porous saplings from
temperate and tropical forests. New Phytologist 186: 439–450.
Meyer-Berthaud B, Scheckler SE, Wendt J. 1999. Archaeopteris is the
earliest known modern tree. Nature 398: 700–701.
Moran R. 2008. Diversity, biogeography and floristics. In: Ranker TA,
Haufler CH, eds. Biology and evolution of ferns and lycophytes.Cambridge, UK: Cambridge University Press, 367–383.
Morrow AC, Dute RR. 1998. Development and structure of pit
membranes in the rhizome of the woody fern Botrichium dissectum.
IAWA Journal 19: 429–441.
Neufeld HS, Grantz DA, Meinzer FC, Goldstein G, Crisosto GM,
Crisosto C. 1992. Genotypic variability in vulnerability of leaf xylem to
cavitation in water-stressed and well-irrigated sugarcane. Plant Physiology100: 1020–1028.
460 Research
NewPhytologist
� 2011 The Authors
New Phytologist � 2011 New Phytologist Trust
New Phytologist (2011) 192: 449–461
www.newphytologist.com

Niklas KJ. 1985. The evolution of tracheid diameter in early vascular plans
and its implications on the hydraulic conductance of the primary xylem
strand. Evolution 39: 1110–1122.
Niklas KJ. 1992. Plant biomechanics: an engineering approach to plant formand function. Chicago, IL, USA: University of Chicago Press.
Page CN. 2002. Ecological strategies in fern evolution: a neopteridological
overview. Review of Palaeobotany and Palynology 119: 1–33.
Pittermann J. 2010. The evolution of water transport in plants: an
integrated approach. Geobiology 8: 112–139.
Pittermann J, Choat B, Jansen S, Stuart SA, Lynn L, Dawson T. 2010.
The relationships between xylem safety and hydraulic efficiency in the
Cupressaceae: the evolution of pit membrane form and function. PlantPhysiology 153: 1919–1931.
Pittermann J, Sperry JS, Hacke UG, Wheeler JK, Sikkema EH. 2005.
The torus-margo pit valve makes conifers hydraulically competitive with
angiosperms. Science 310: 1924.
Pittermann J, Sperry JS, Hacke UG, Wheeler JK, Sikkema EH. 2006a.
Mechanical reinforcement against tracheid implosion compromises the
hydraulic efficiency of conifer xylem. Plant, Cell & Environment 29:
1618–1628.
Pittermann J, Sperry JS, Wheeler JK, Hacke UG, Sikkema EH. 2006b.
Inter-tracheid pitting and the hydraulic efficiency of conifer wood: the
role of tracheid allometry and cavitation protection. American Journal ofBotany 93: 1265–1273.
Pockman WT, Sperry JS, O’Leary JW. 1995. Sustained and significant
negative water pressure in xylem. Nature 378: 715–716.
Proctor MC, Tuba Z. 2002. Poikilohydry and homoihydry: antithesis or
spectrum of possibilities? New Phytologist 156: 327–349.
Robinson RC, Sheffield E, Sharpe JM. 2010. Problem ferns: their impact
and management. In: Mehltreter K, Walker LR, Sharpe JM, eds. Fernecology. Cambridge, UK: Cambridge University Press, 255–310.
Rothwell GW, Karrfalt EE. 2008. Growth, development and systematics
of ferns: does Botrychium S.L. (Ophioglossales) really produce secondary
xylem? American Journal of Botany 95: 414–423.
Rowe N, Isnard S, Speck T. 2005. Diversity of mechanical architectures in
climbing plants: an evolutionary perspective. Journal of Plant GrowthRegulation 23: 108–128.
Rowe N, Speck T. 2004. Plant evolutionary forms; an ecological and
evolutionary perspective. New Phytologist 166: 61–72.
Saldana A, Lusk CH, Gonzales WL, Gianoli E. 2007. Natural selection on
ecophysiological traits of a fern species in a temperate rainforest.
Evolutionary Ecology 21: 651–662.
Schenk HJ, Espino S, Goedhart CM, Nordenstahl M, Martinze-Cabrera
HI, Jones CS. 2008. Hydraulic integration and shrub growth form
linked across continental aridity gradients. Proceedings of the NationalAcademy of Sciences, USA 105: 11248–11253.
Schuettpelz E, Pryer KM. 2009. Evidence for a Cenozoic radiation of
ferns in an angiosperm-dominated canopy. Proceedings of the NationalAcademy of Sciences, USA 106: 11200–11205.
Schneider H, Schuettpelz E, Pryer KM, Cranfill R, Magallon S, Lupia R.
2004. Ferns diversified in the shadow of angiosperms. Nature 428:
553–557.
Schulte PJ, Gibson AC, Nobel P. 1987. Xylem anatomy and hydraulic
conductance of Psilotum nudum. American Journal of Botany 74:
1438–1445.
Sperry JS. 1993. Winter xylem embolism and spring recovery in Betulacordifolia, Fagus grandifolia, Abies balsamea, and Picea rubens. In:
Borghetti M, Grace J, Raschi A, eds. Water Transport in Plants underClimatic Stress. Cambridge, UK: Cambridge University Press, 86–98.
Sperry JS. 2003. Evolution of water transport and xylem structure.
International Journal of Plant Sciences 164: S115–S127.
Sperry JS, Hacke UG, Pittermann J. 2006. Size and function in conifer
tracheids and angiosperm vessels. American Journal of Botany 93:
1490–1500.
Sperry JS, Hacke UG, Wheeler JK. 2005. Comparative analysis of end
wall resistivity in xylem conduits. Plant, Cell & Environment 28:
456–465.
Spicer R, Gartner BL. 2002. Compression wood has little impact on the
water relations of Douglas-fir (Pseudotsuga menziessii) seedlings despite a
large effect on shoot hydraulic properties. New Phytologist 154:
633–640.
Spicer R, Groover A. 2010. Evolution of development of vascular cambia
and secondary growth. New Phytologist 186: 577–592.
Stewart WN, Rothwell G. 1993. Paleobotany and the evolution of plants.Cambridge, UK: Cambridge University Press.
Taylor TN, Taylor EL, Krings M. 2009. Paleobotany: the biology andevolution of fossil plants. Oxford, UK: Academic Press.
Tyree MT, Zimmermann MH. 2002. Xylem structure and the ascent of sap.
Berlin, Germany: Springer.
Veres JS. 1990. Xylem anatomy and hydraulic conductance of Costa Rican
Blechnum ferns. American Journal of Botany 77: 1610–1625.
Watkins JE, Holbrook NM, Zwieniecki MA. 2010. Hydraulic properties
of fern sporophytes: consequences for ecological and evolutionary
diversification. American Journal of Botany 97: 2007–2019.
Wheeler JW, Sperry JS, Hacke UG, Hoang N. 2005. Intervessel pitting
and cavitation in woody Rosaceae and other vesseled plants: a basis for a
safety vs. efficiency trade-off in xylem transport. Plant, Cell &Environment 28: 800–812.
Wilson JP, Fischer WW. 2010. Hydraulics of Asteroxylon mackei, an early
Devonian vascular plant, and the early evolution of water transport
tissue in terrestrial plants. Geobiology 9: 121–130.
Wilson JP, Knoll AH. 2010. A physiologically expicit morphospace for
tracheid-based water transport in modern and extinct seed plants.
Paleobiology 36: 335–355.
Wilson JP, Knoll AH, Holbrook NM, Marshall CR. 2008. Modeling
fluid flow in Medullosa, an anatomically unusual Carboniferous seed
plant. Paleobiology 34: 472–493.
Zanne AE, Sweeney K, Sharma M, Orians CM. 2006. Patterns and
consequences of differential vascular sectoriality in 18 temperate tree and
shrub species. Functional Ecology 20: 200–206.
Supporting Information
Additional supporting information may be found in theonline version of this article.
Fig. S1 Additional vulnerability curves (bench dry (BD)and centrifuge method).
Table S1 Additional conduit diameter data (minimum andmaximum diameter in lm) for the species surveyed in thestudy
Please note: Wiley-Blackwell are not responsible for thecontent or functionality of any supporting informationsupplied by the authors. Any queries (other than missingmaterial) should be directed to the New Phytologist CentralOffice.
NewPhytologist Research 461
� 2011 The Authors
New Phytologist � 2011 New Phytologist Trust
New Phytologist (2011) 192: 449–461
www.newphytologist.com