studien zur hitzeresistenz der gezeitenalgechaetomorpha cannabina (aresch.) kjellm
TRANSCRIPT

Protoplasma 67, 451--472 (1969)
Studien zur Hitzeresistenz der Gezeitenalge Chaetomorp ha cannabina (Aresch.) Kjellm.
RICHARD BIEBL
Douglas Marine Station, Douglas, Alaska, und Pflanzenphysiologisches Institut der Universitiit Wien
Mit 2 Abbildungen
Eingegangen am 26. M~irz 1969
Zusammenfassung 1. Chaetomorpha cannabina (Ares&.) Kjellm. ist durch ihre hohe osmotische Resistenz (0,1--4,0 Standortwasser), ihre osmoregulatorische Anpassungsflihigkeit an ge~inderten Salz- gehalt sowie dutch ihre Gefrierresistenz als Alge der Gezeitenzone gekennzeichnet.
2. Als Schnellmethode zur ~Sko-physiologischen Unterscheidung yon Gezeitenalgen und Algen des st~indig yore Wasser bedeckten Sublitorals wird 24stiindiges Einlegen in 0,2 Seewasser empfohlen. Gezeitenalgen iiberleben, Tiefenalgen sterben ab.
3. Der Absterbeverlauf nach kurzfristiger Erhitzung (5 Minuten, �89 Stunde) zeigt, dat~ Aussagen iJber die Resistenzgrenze friihestens 24 Stunden nach dem Erhitzen gemacht werden k6nnen (Tabelle 4 und 5).
4. Die Resistenzgrenzen (= Lebensgrenzen) yon Chaetomorpha cannabina liegen 24 Stunden nach 5 Minuten, 30 Minuten und 12 Stunden Erhitzung bei 37 ~ 36 ~ und 29 ~ C.
5. Auch bei anderen Meeresalgen (Tabelle 6) ]iegen die Lebensgrenzen nach 30 Minuten Erhitzung um 1~ ~ C tiefer als nach 5 Minuten Erw~rmung und nach 12 Stunden Erw~irmung um 6~ ~ C tiefer als nach einer Erhitzung yon 30 Minuten.
6. An ftinf aufeinanderfoigenden Tagen mehrfach t~iglich durchgefiihrte Resistenzbestimmun- gen lief~en bei in Kulturschalen gehaltener Chaetomorpha cannabina ein auffallendes Schwanken der Hitzeresistenz in einer mit den Zeitpunkten yon Ebbe und Hut zusammen- fallenden sechsstiindigen ,,Gezeitenrhythmik" erkennen (Abb. 1, 2). Die Wahrscheinlichkeit des Schwankens auch anderer physiologischer Eigenschaften sowie die M/Sgliehkeit des Auf- tretens einer solchen Rhythmik auch bei Landpfianzen werden diskutiert.
7. Vorbehandlung yon Chaetomorpha cannabina mit verdiinntem bzw. konzentriertem Seewasser fiihrt im ersten Fall zu einer Herabsetzung, im zweiten zu einer Erh~Shung der Hitzeresistenz.
8. Eine Vorbehandlung mit in Seewasser gel6ster Glukose (0,2 und 0,4mol) brachte keine Knderung der Hitzeresistenz.

452 R. BIEBL
Summary Heat Res i s t ance Studies in the In t e r t ida l A lga Chaetomorpha cannabina (Areseh.) Kjellrn.
1. Chaetomorpha cannabina (Ares&.) Kjellm. is characterized as an intertidal alga by high osmotic hardiness (0.1-4.0 times the concentration of habitat sea-water), osmoregulatory adaptation to changes in salt content, and freezing hardiness.
2. Immersion into 0.2 sea-water for 24 hours is recommended as a rapid method for eco- physiological distinction between intertidal algae and those of the sublitoral permanently covered with water. Intertidal algae survive, sublitoral algae die.
3. The course of decay after short heat exposure (5 min, �89 hour) demonstrates that valid statements on hardiness limits can be given not earlier than 24 hours after heating (Tables 4 and 5).
4. 24 hours after exposure for 5 minutes, 30 minutes and 12 hours, hardiness limits (= limits of life) of Chaetomorpha cannabina are found at 37 ~ C, 36 ~ C, and 29 ~ C, respectively.
5. Hardiness limits of other maritime seaweeds after exposure for 30 minutes are also lowered by 1-2 ~ C in comparison with exposure for 5 minutes, and lowered by 6-10 ~ C after heating for 12 hours, compared to exposure for 30 minutes (Table 6).
6. Chaetomorpha cannabina, cultivated in dishes for 5 consecutive days and repeatedly che&ed for resistance, showed conspicuous fluctuations of heat hardiness in a "tidal rhythm" coinciding with the six-hour intervals of ebb and fiood (Figs. 1 and 2). The probability is discussed of fluctuations in other physiological traits as well as the possibility that similar rhythms could occur in land plants, too.
7. Pretreatment of Chaetomorpha cannabina with dilute and concentrated sea water results in lowering of heat hardiness in the first case and in its raise in the second.
8. Pretreatment with glucose (0.2- and 0.4molar solutions in sea-water) did not change heat hardiness.
1. Einle i tung
Im Zuge vergleichender Untersuchungen der Temperaturresistenz yon Meeres- algen verschiedener Klimazonen (BIEgL 1939, 1958, 1962 a, b, c, 1968) wurden im Sommer 1968 Arbeiten in Alaska, in Friday Harbor (Wash.) und an zwei Stationen der pazifischen Ktiste yon Kalifornien durchgefiihrt. Die vorliegende Mitteilung bezieht sich im wesentlichen auf die Untersuchun- gen an der Douglas Marine Station des Institute of Marine Science der Universit~it yon Alaska. Die groi~e, gut ausgestattete Station liegt auf Douglas Island im Siidosten Alaskas. Hauptprogramm war hier, an der als Versuchsobjekt besonders geeigneten Tidepool-Alge Chaetornorpha cannabina (Ares&.) Kjellm. eine m~Sglichst viel- seitige Untersuchung ihrer Hitzeresistenz anzustellen. Zur Untersuchung stan- den die Hitzeresistenzgrenzen im Verh~iltnis zu den Standorterfordernissen und den sonstigen 6kophysiologischen Eigenschaften dieser Alge, der Absterbe- verlauf bei verschieden langer Erhitzungsdauer, die Tagesschwankungen der Hitzeempfindlichkeit und der Einflu!g yon verdiinntem und konzentriertem Seewasser sowie yon Zuckerzusatz auf die H~She der Hitzeresistenz.

Studien zur Hitzeresistenz yon Chaetomorpha cannabina 453
Ich darf an dieser Stelle Dr. K. M. RAE, Vice-President for Research and Advanced Study, fiir die Einladung und grogziigige Unterstiitzung seitens der Universit~it yon Alaska, Dr. Donald W. HOOD, Direktor des Institute of Marine Science in College, A1., far die ErmSglichung der Arbeiten an den Auigenstellen des Instituts und Dr. Y. R. NAXUDU, wissenschattlicher Leiter der Douglas Marine Station, sowie den Herren der Station ftir die iiberaus liebenswiirdige Aufnahme und tatkr~iftige Hilfe w~ihrend meines Aufent- haltes in Douglas meinen herzlichsten Dank aussprechen. Prof. Dr. RICHARD E. Nol~RIS yon der University of Washington, Seattle, hatte die grot~e Freund- lichkeit, meine Versuchsalgen zu bestimmen.
Fiir die Verleihung des A.-F.-W.-Schimper-Stipendiums 1968 fiir 5ko-physio- logische Untersuchungen in aut~ereurop~ischen L~indern habe ich der H.- und E.-Walter-Stiftung aufrichtigen Dank zu sagen.
2. Standort Douglas Island ist eine schmale, etwa 27 km lange, durch eine Briicke mit dern am Festland gelegenen St~idtchen Juneau verbundene Insel an der Siidostktiste Alaskas (58 ~ 15' 30"N, 134 ~ 16' 30" W). Sie ist dutch den Gastineau Channel vom Festland und durch die Stephens Passage yon einigen weiteren, gegen den Pazifisehen Ozean zu vorgelagerten grSgeren Inseln getrennt.
Tabelle 1. Salzgehalt und Temperatur des Was- sers in der Niihe des Standortes der Versuchs- algen. Douglas Island, ,,Fritz Cove". Gemessen 26. ]uni 1968, 17 Uhr
Tiefe (m) Salinit~it %0 Temperatur o C
Oberfl~iche 21,77 11,80
21,84 11,80 1 21,89 11,80 2 27,65 9,75 4 29,48 9,02 5 30,38 8,77
Das Wasser des Gastineau-Kanals und der Stephens-Passage ist durch Stit~wasserzufliisse, vor allem des grotgen Taku River im Osten der Insel, stark ausgesiitgt (PICHARD 1967). Dabei nimmt die SalinitS.t mit der Entfernung yon der Miindung des Flusses und mit der Tiefe rasch zu. So wies das Oberfl~ichenwasser des Gastineau-Kanals auf der H/Jhe der nahe dem 5stlichen Ende der Nordkiiste yon Douglas Island gelegenen Douglas Marine Station einen Salzgehalt yon nur 13,43%0 auf, wiihrend dieser am westlichen Ende der Nordkiiste auf der I-I/She des Standortes der Versuchsalgen schon 21,77%0 betrug. Tabelle 1 bringt Werte des Salzgehaltes und der Wassertemperatur, die Ende Juni 1968 in unmittelbarer N~ihe der klippenreichen, felsigen Kiiste am westlichen Ende der Nordk~ste gemessen wurden, aus deren Gezeitentlimpeln die Hauptversuchsalge Chaetomorpha cannabina stammte. Die Werte wurden mir in freundlicher Weise yon Mr. PEARSON, Douglas Marine Station, zur Verfiigung gestellt.

454 R. BIEBL
Der Salzgehalt des Wassers in den Gezeitentiimpeln kann zu Beginn ihres Freiliegens etwa jenem des Oberfl~ichenwassers gleichgesetzt werden. Infolge der niederen Lufftemperatur diirfte ein Ansteigen des Salzgehaltes w~ihrend der Ebbe nicht allzusehr ins Gewicht fallen. Zur Zeit der tiefsten Ebbe wurde in einem seichten Seewassertiimpel um 12 Uhr mittags eine Wassertemperatur yon 17 ~ C gemessen. Es ist anzunehmen, daf~ die Temperatur an besonders warmen Tagen bis gegen 20 ~ C ansteigen kann. Als Richtwerte bringt Tabelle 2 einige Angaben der Lufftemperatur, wie sie in der Wetter- station des etwa 3 km Lufflinie entfernten, am Nordufer des Gastineau-Kanals gelegenen Airport Juneau in den Tagen der Aufsammlung gemessen wurde.
Tabelle 2. Lufltemperaturen (in o C), gemessen etwa 3 km Lufllinie yore Algenstandort am Airport Juneau
Datum 1968 08.00 12.00 20.00 24.00 Min. Max.
3. Juli 15,0 20,6 17,8 11,5 6,0 24,3 4. Juli 11,2 15,5 12,5 11,1 3,0 16,0 5. Juli 9,5 10,0 10,0 9,6 8,9 11,8 6. Juli 8,0 11,8 6,8 7,2 6,0 17,0 7. Juli 8,0 16,0 11,1 10,0 4,8 18,2
3. Versuche
3.1. U k o - p h y s i o l o g i s c h e C h a r a k t e r i s t i k d e r V e r s u c h s a l g e
Chaetomorpha cannabina finder sich an den R~indern yon Gezeitentiimpeln knapp unter der Wasseroberfl~iche bis etwa 15 cm Tiefe, tells rein, teils mit anderen Algen, so etwa mit der Rotalge Odonthalia floccosa, vermengt. Dies entspricht den Angaben yon SCAGEL (1957), der als Standort dieser Alge angibt: ,,In tangled masses, often interwined with other algae in the intertidal- zone." Als Verbreitung der Alge nennt der Autor ,,Pacific Coast: Norton Sound Alaska to Northern Washington". Die Alge brachte fiir zellphysiologische Untersuchungen zwei wertvolle Eigen- schaften mit: 1. eine groi% Widerstandskra~ gegen Kulturbedingungen (14 Tage in einem Glasgef~if~ mit Standortwasser gehaltene Algen zeigten noch keinerlei Sch~idigungen) und 2. eine gute Plasmolysierbarkeit. Die grof~e Osmotische Resistenz kennzeichnet Chaetomorpha cannabina als typische Gezeitenalge (vgl. BIEBL 1958). Zur Bestimmung der osmotischen Resistenz wurde die Versuchsalge fiir 24 Stunden in weithalsigen Glasfl~isch- chen einerseits in dest. H~O, 0,i und 0,2 Seewasser (1 bzw. 2 Teile Seewasser und 9 bzw. 8 Teile dest. H~O), andererseits in dutch Eindampfen zwei-, drei- und vierfach konzentriertes Seewasser eingelegt und anschlief~end auf ihren Lebenszustand gepriJfL. Als Vitalit~tsprobe wurde bei Chaetomorpha Plas- molyse, bei anderen Algen auch Vitalf~irbung mit Neutralrot angewendet. Als ,,1,0 Seewasser" wurde Oberfl~ichenwasser des Gastineau-Kanals aus der N~ihe der Station verwendet, dessen Salinit~it, wie schon erw~hnt, .~ur 13,425%0

Studien zur Hitzeresis tenz yon Chaetomorpha cannabina 455
betrug. Die Salinitiitsbestimmungen, die in freundlicher Weise vom Biologi- schen Laboratorium des Fish and Wildlife Service in Auke Bay, Alaska, vor- genommen wurden, ergaben fiir die verwendeten LSsungen folgende Werte:
Seewasserkonzentration Salinit~it
1,0 13,425 %0 2,0 26,454 %o 4,0 53,590 %0
Die osmotische Resistenz yon Chaetomorpha cannabina gegen verdiinntes und konzentriertes Seewasser stimmt iiberein mit der tells im gleichen Tidepool, tells an benachbarten Stellen der Gezeitenzone vorkommenden Algen (Tabelle 3).
Tabelle 3. Osmotische Resistenz yon Chaetomorpha cannabina und benachbarter Algen der Gezei tenzone
Alge Verdiinntes Seewasser Konzentriertes Seewasser
dest. H=O 0,1 0,2 2,0 3,0 4,0
Chaelomorpha cannabina + 1 1 1 1 1 Cladophora spec. + 1 1 1 1 1 Ulva lacluca + 1 1 1 1 1 Enleromorpha lorta 4- 1 1 1 1 Polysiphonia paniculata + 1 1 1 1 1 Odonlhalia floccosa + 1 1 1 i 1
(-1- = tot, 1 = lebend)
Es sei daran erinnert, dafg die Hypotonieresistenz der Algen des st~indig vom Wasser bedeckten Sublitorals nach bisherigen Beobachtungen (z. B. BIEBJ. 1938, 1958) demgegeniiber kaum je eine Grenze yon 0,4 Seewasser unter- schreitet. Da Algen aus Gezeitentiimpeln manchmal schon dutch 0,1 Seewasser gesch~digt werden, kann man eine Resistenz gegen 0,2 Seewasser als allgemein kennzeichnend fiir Algen der Gezeitenzone ansprechen. Zur physiologischen Unterscheidung von Algen der Gezeitenzone und der st~indig vom Wasser bedeckten Tiefe kann man daher ein 24stiindiges Einlegen von Algenproben in 0,2 Seewasser als einfache Schnellmethode empfehlen: Gezeitenalgen iiber- leben, Tiefenalgen sterben ab. Der osmotische Wert yon Chaetomorpha cannabina entspricht bei Grenz- plasmolyse einer 3,35fach konzentrierten L5sung des verwendeten Seewassers (1,0 = 13,425%0), das heit~t einer Salinitiit yon 45,05%0. Es kommt dies etwa dem osmotischen Weft yon Cladophora-Arten in der Gezeitenzone yon Plymouth, England (BmBL 1937), gleich, bei denen Grenzplasmolyse in 1,5fach konzentriertem Standortwasser yon 33%0 Salzgehalt gefunden wurde, was einer Salinit{it yon 49,5%0 entspricht. Protoplasma 67/4 31

456 R. BIE~t
Einen Hinweis auf osmoregulatorische Anpassungsf~ihigkeit gibt die Beobach- tung der 48 Stunden lang in 0,1 und 4,0 Seewasser gelegenen Algen, die bei Versuchsbeginn in gleicher Weise Grenzplasmolyse in 3,35 Seewasser gezeigt hatten. Von den 48 Stunden in 0,1 Seewasser gelegenen Algen plasmolysierten nun schon in 2,9 S.W. 40%, in 2,8 S.W. 5% und in 2,7 S.W. noch etwa 1% der Zellen. Die in 4,0 S.W. gelegenen Algen waren hingegen durchwegs deplasmolysiert, das heii~t, ihr osmotischer Wert ist tiber 4,0 S. W. an- gesfiegen. Vgl. hiezu die Untersuchungen yon K~SS~LER (1959, 1964) an Chaet. Iinum. Schliei~lich kennzeichnet auch die Gefrierresistenz Chaetomorpha cannabina als Gezeitenalge. W~ihrend stiindig submerse Meeresalgen des Sublitorals hie- reals, auch nicht in den kalten Gew~issern von Gr~Snland (BIEBL 1968), ein Einfrieren ertragen und bei zw61fsttindiger Abktihlung bestenfalls bis zum Gefrierpunkt des Seewassers am Leben bleiben, tiberlebt Chaetomorpha, wie fast alle Gezeitenalgen temperierter und kalter Meere (KYLIN 1917, BIEBL 1958, TEe, UMOTO 1964), ein zw/Slfsttindiges Einfrieren bei einer zwischen - - 4 ~ C und - - 9 ~ C schwankenden Temperatur des Eisschrankes ohne Sch~i- digung. 24 Stunden nach dem Auftauen bei q- 5 ~ C liei~en sich die Zellen noch zu 100 % plasmolysieren.
3.2. H i t z e r e s i s t e n z u n d A b s t e r b e v e r l a u f b e i v e r s c h i e d e n e r E r w ~ i r m u n g s d a u e r
Schon SACHS (1864) wies darauf hin, daf~ die Widerstandsf~ihigkeit einer Pflanze gegen hohe Temperaturen yon der L~inge der Einwirkungsdauer ab- h~ngig ist und dai~ daher von einer bestimmten H6chsttemperatur, die eine Pflanze ertr~igt, nicht gesprochen werden kann. Dazu kommt, da~ die dutch eine gewisse Erhitzungsdauer gesetzte Sch~idigung of~ erst nach l~ingerer Zeit sichtbar wird. Angaben der Erhitzungsdauer und des Zeitpunktes der Beob- achtung der Sch~idigung sind daher zur Vergleichbarkeit yon Hitzeresistenz- grenzen unbedingt notwendig. Bei den Bestimmungen der Hitzeresistenz yon Landpflanzen hat sich eine halbst~ndige Erhitzungsdauer und Angabe der Vitalit~itsgrenzen nach 3--5 Tagen NachkuItur weitgehend durchgesetzt (LANGE 1959, BI~BL 1964, 1968, KAPPEN 1964 U. a.). Bei Untersuchungen der Hitzeresistenz yon Meeres- und Stii~wasseralgen wurde bisher noch auf keine solche Einheitlichkeit der Erhitzungsdauer und der Beobachtungszeit geachtet. Die Erw~irmung unserer Versuchsalgen wurde in weithalsigen, 1-nit 10 ml See- wasser gefiillten 50-ml-Glasfl~ischchen vorgenommen, die bis zum Hals in verschieden temperierte Wasserb~ider eingeh~ingt wurden. Die Wasserb~ider wurden durch Einh~ingethermostaten (Thermomix II der Fa. Braun, Melsun- gen, Westdeutschland) verschieden stark erw~irmt. Die Temperaturen der Mef~reihen waren auf 1~ C abgestuft. Bei fignf Minuten ianger Erhitzungsdauer wurden die Versuchsalgen direkt

Studien zur Hitzeresistenz von Chaetomorpha cannabina 457
in die bereits auf die betreffende Temperatur gebrachten Seewasserfl~ischchen eingelegt. Die Beobachtung des Absterbeverlaufes yon Chaetomorpha can- nabina zeigte innerhalb der ersten 24 Stunden eine rasche Zunahme der S&~i- digung yon den h/Schsten Temperaturen nach den niedrigeren zu. In den folgenden vier Tagen schritt aber das Absterben dann nur mehr in den schon nach 24 Stunden sichtbar gewordenen sch~idigenden Temperaturen welter, w~ihrend die Algen in den n~ichstniedrigeren Temperaturstufen ungesch~idigt blieben (Tabelle 4).
Tabelle 4. Chaetomorpha cannabina, Absterbeverlauf nach 5 Minuten Erw2irmung. Erhitzung 4. Juli, 20 Uhr
Zeit nach der Hitzeresistenz, ~ C, 5 Minuten Erwgrmung i
I 38 39
8 Minuten 3 Stunden 7 Stunden 14 Stunden 1 Tag 1 ~ Tage 2 Tage 3 Tage r Tage 5 Tage
36 37
1 I 1 1 1 1 1 l I
i 1
I 1 1 1
5% 90% 98% 99%
+ +
1 1
1% 10% 40% 9o%
+ + + +
40 I 41
1 1 2% I 5%
10% 70% 30% 90% 90% +
-4- + § + + + + + + +
(1 = alles lebend, % tot, + = alles tot)
Wurden die Algen eine halbe Stunde erhitzt, so wurden sie nicht direkt in das schon auf die Endtemperatur vorerw~irmte Wasser gebracht, sondern in den Glasfl~ischchen in 10 ml Seewasser yon Zimmertemperatur eingelegt und an- schliegend in das entsprechend temperierte, st~indig gerCihrte Wasserbad ein- geh~ingt. Dabei wurde zur halbstiindigen Erhitzungszeit fiinf Minuten ,,An- w~irmzeit" zugegeben. Temperaturkontrollen zeigten n~imlich, dag innerhalb dieser fiinf Minuten im Wasser der Fl~ischchen die jeweils zu priKende End- temperatur erreicht wurde, so daft diese dann volle 30 Minuten einwirken konnte. Auch hier zeigte sich Gleiches wie nach fiinf Minuten Erhitzungsdauer: Die. Hitzesch~idigung schreitet innerhalb der ersten 24 Stunden um mehrere Grade nach unten zu fort und fiihrt in den n~ichsten zwei Tagen in der Temperatur, in der nach 24 Stunden noch Sch~idigungen aufgetreten waren (37 ~ C), zu einem vollst~indigen Absterben der Alge. Eine um 1~ C niedrigere Erhitzungs- temperatur lieg die Algen w~ihrend der ersten drei Tage zu 100 % iiberleben. Allerdings zeigten am dritten Tag die Algen bei dieser Temperatur (36 ~ C) eine auffallend sdirkere Plasmolyse als die mit den Kontrollen bei Zimmer- temperatur iibereinstimmenden, ~A Stunde lang auf 35 ~ C erw~irmt gewesenen

458 R, B~EBL
Algen. Der osmotische Wert war sichtlich etwas abgesunken, Zellinhaltsstoffe miissen exosmiert sein. Vier Tage nach der Erw~irmung begannen dann auch die nur auf 36 ~ C erhitzten Algen abzusterben (Tabelle 5). Bezeichnet man als Resistenzgrenze jene Erw~irmungstemperatur, bei der die Algen zu 100% tiberleben, so zeigt sich, dab diese innerhalb der ersten drei Tage nach halbstiindiger Erw~irmung nur um ein geringes tiefer liegt als nach fUnf Minuren Erw~irmung. Nach fiinf Minuten Erhitzung tiberlebte Chaeto- morpha cannabina zu 100% eine Erw~irmung auf 3 7 ~ und nach halb- sttindiger Erw~irmung eine solche auf 36 ~ C. W~ihrend abet die nach ftinf Minuten Erw~irmung innerhalb von 24 Stunden erreichte Resistenzgrenze
Tabelle 5. Chaetomorpha cannabina, Absterbeverlauf nach �89 Stunde Erwiirmung
Zeit nach der Hitzeresistenz ~ C, �89 Stunde Erw~irmung
35 36 37 40
5 Minuten 7 Stunden 16 Stunden 23 Stunden 40 Stunden 2 Tage 3 Tage 4 Tage
1 = alles lebend, -- 1
1 1 1 1 1 1
--1 50%
1 1
1% 2%
90% 90% 97%
+
38 39
1 1
503/0 95 % 80% +
+ + + + + + + + + +
1 + + + + + + +
= leichte Sch~idigung, % tot, + = alles tot)
auch in den folgenden vier Tagen unver~indert erhalten blieb, verschob sich diese nach halbstiindiger Erhitzungsdauer am vierten Tag noch um 1 Grad nach unten (35 ~ C). Wesentlich tiefer liegt die Resistenzgrenze, wenn die Erhitzungsdauer auf zw?Jlf Stunden ausgedehnt wurde. In diesem Fall war die Resistenzgrenze schon unmittelbar na& Abschlui~ der Erhitzung zu erkennen und griff in den folgenden vier Tagen nicht mehr auf tiefere Temperaturen iiber. 24 Stunden nach Erw~irmung auf 30 ~ C waren etwa 60% der Zellen der Chaetomorpha- Probe abgestorben, w~ihrend die nur auf 28 ~ und 2 9 ~ erw~irmten Algen nach drei weiteren Tagen keine Sch~idigung aufwiesen. Die Grenzen des 100prozentigen fdberlebens innerhalb yon 24 Stunden nach 5 Minuten, 30 Minuten und 12 Stunden Erhitzung liegen somit ftir Chaeto- morpha cannabina bei 37 ~ 36 ~ und 29 ~ C. Tabelle 6 bringt zum Vergleich die Resistenzgrenzen (Lebensgrenzen) yon Algen, die einige Wochen sp~iter an der kleinen Feldstation des Institute of Marine Science der Universit~it yon Alaska an der Izembek Lagoon (Bering- see) am stidwestlichen Ende der Halbinsel Alaska in der N~ihe yon Cold Bay mit gleichen Erhitzungsdauerri bestimmt wurderi.

Studien zur Hitzeresistenz yon Chaetomorpha cannabina 459
Die Erhitzungsreihen waren bei den Versuchen an der Izembek-Station nur um 2 ~ abgestut~, so daf~ die M/Sglichkeit often ist, datg sich die Resistenz- grenzen nach 5 Minuten und 30 Minuten Erhitzung auch nur um 1 ~ C unter- scheiden. Jedenfalls zeigt sich auch hier, wie bei Chaetomorpha cannabina, ein nur geringer Unterschied in der Hitzeresistenz nach 5 Minuten und 30 Minuten Hitzeeinwirkung, w~ihrend die Lebensgrenzen nach 12 Stunden Hitzeeinwir- kung ebenfalls um 6--10~ C tiefer als nach 30 Minuten Erw~irmung liegen. Auch die nach �89 und 12 Stunden Erw~irmung an einigen Algen Westgr~Sn- lands beobachteten Lebensgrenzen liegen um 6 ~ bis 10~ voneinander entfernt (BIEBL 1968).
Tabelle 6. Algen yon Cape Gleznap (Beringsee), 24 Stunden nach ver- schieden langer Erhitzung (Abstufung der Erhitzungstemperaturen um 2 ~ C)
Alge Hitzeresistenz, o C, 24 St&a. n. d. Erhitzung
5 Min. 30 Min. 12 Stdn,
Ceramium doaciflcum 36 34 24
fFufhora frul#ulosa 34 32 24
Pliloia asp]enioide~ 34 32 24
Areodilsea spec. 34 32 24
Alaria spec. 34 32 --
Laminaria saccharina 34 32 24
Desmerestia intermedia 34 32 24
Znleromorpha linza 42 42 26
Eine Sonderstellung scheint die extreme Gezeitenalge Enteromorpha linza einzunehmen. Hier liegen die Lebensgrenzen nach 30 Minuten Erw~irmung (42 ~ C) und nach 12 Stunden Erw~irmung (26 ~ C) um 16~ auseinander. Die Lebensgrenze nach 5 Minuten Erw~irmung diirfte bei Staftelung der Versuchstemperaturen um 1 ~ C wahrscheinlich bei 43 ~ C zu finden sein. Dies wiirde genau dem Wert entsprechen, den FELDMANN und LUTOVA (1963) auf der russischen Seite der Beringsee (bei einer Wassertemperatur yon 10 ~ C) nach 5 Minuten ErwS.rmung fiir Enteromorpha compressa mit 43,4 _+ 0,5 festgestellt haben. Wurde diese Alge bei 19 ~ C 24 Stunden lang vorkultiviert, so stieg die Resistenz sogar auf 47,2 _+ 0,5 ~ C an. L~ingere Temperaturein- wirkungen wurden yon den beiden Autorinnen nicht untersucht. Der groi% Abstand der Resistenzgrenzen yon Enteromorpha linza nach �89 Stunde Erw~irmung (42 ~ C) und nach 12 Stunden Erw~irmung (26 ~ C) k6nnte m/Sglicherweise ein Charakteristikum des Plasmas von extremen Ge- zeitenalgen sein, die an Sch6nwettertagen w~ihrend der Ebbe kurzfristig sehr hohen Temperaturen ausgesetzt sind.

460 R. BIEBL
3.3. , , G e z e i t e n r h y t h m i k " d e r H i t z e r e s i s t e n z v o n Chaetomorpha cannabina
Die Versuchsalge wurde am 2. Juli aus einem Tidepool des eingangs be- schriebenen Standortes um 12 Uhr mittags bei tiefer Ebbe und einer Wassertemperatur im Gezeitenfiimpel von 17~ gesammelt und am Fen- ster des Laboratoriums in Glasgef~if~en mit nicht durchlfiftetem Standort- wasser bei einer zwischen 18 ~ und 25~ schwankenden Zimmertemperatur gehalten. Ab 20 Uhr desselben Tages wurden durch f~inf Tage in bestimmten Zeit- abst~inden Proben entnommen, 5 Minuten lang auf 37 ~ 38 ~ 39 ~ und 40 ~ C erhitzt und in den nachfolgenden Tagen auf ihren Lebenszustand unter- sucht. In der Abb. 1 a und b sind die Zeitpunkte der Erhitzung und die Zeiten des jeweils am Standort herrschenden niedersten (N. W.) und h6chsten (H. W.) Wasserstandes angegeben. Die Kurven der Abb. 1 a zeigen die gesch~itzten Prozente iiberlebender Zellen 24 Stunden nach 5 Minuten Erhitzung auf 38 ~ 39 ~ und 40 ~ C. Aus dem Kurvenverlauf lassen sich zwei Erscheinungen ablesen: 1. In den ersten zwei Tagen nach der Einbringung der Algen vom Standort ist ihre Hitzeresistenz geringer als in den Tagen nachher. Erhitzung auf 39 ~ und 40~ fiihrt in den ersten zwei Tagen innerhalb yon 24 Stunden zu einem vollsfiindigen Absterben, w~ihrend in den nachfolgenden Tagen die Oberlebensrate fiir 38 ~ und 39 ~ C auf fast 100% ansteigt. Sogar yon den auf 40 ~ C erw~irmten Algen iiberleben innerhalb von 24 Stunden zu bestimmten Zeiten noch hohe Prozent- zahlen. 2. Die auf 38 ~ C erw~irmten Algen zeigen in den ersten zwei Tagen charakteristische Resistenzschwankungen, die mit dem Ansteigen der Resistenz in den folgenden Tagen verschwinden, nun aber in den auf 39 ~ und 40~ erhitzten Algen - - wenn auch nicht so regelm~if~ig - - sichtbar werden. Auch dies ist ein Ausdruck der gr~Sf~eren Hitzeempfindlichkeit der standortfrischen Algen. Die Zunahme der Hitzeresistenz nach l~ingerer Kultur im Laboratorium l~it~t sich als eine Anpassung an die wesentlich h/Shere Zimmertemperatur erkl~iren. Eine Herabsetzung der Hitzeresistenz dutch Vorkultur bei niederer Tempera- tur und eine Steigerung durch Vorkultur bei h6herer Temperatur wurde an Meeresalgen schon mehrfach nachgewiesen (ScHwENKE 1959, L~JTOVA und FELDMANN 1960, FELDMANN, ZAVADSKAYA und LUTOVA 1963, FELDMANN und LUTOVA 1963, LUTOVA, ZAVADSKAYA, LUKNITSKAYA und FELDMANN 1967, fiir Sfii~wasseralgen: SCH6LM 1968). Neu und interessanter war jedoch das Auftreten der periodischen Schwan- kungen der Hitzeresistenz yon Chaetomorpha cannabina im Tagesverlauf, wie sie in Abb. 1 a 24 Stunden und in Abb. 1 b vier Tage nach 5 Minuten Erhitzung auf 38 ~ C besonders deutlich sichtbar werden.

Studien zur Hi tzeres i s tenz y o n Chaetomorpha cannabina 461
\ \ )
/
\\ \\
(,,. \\\\
/ I
l.a
~ ~
~ o r tq o o
N
�9 N
Fa . . ~
~ z ~
~o~
~ o
_d

462 R. BIEBL
Die Uberlebenskurven in Abb. 1 b beginnen zwei Tage nach dem Einholen der Algen vom Standort und zeigen den Lebenszustand vier Tage nach der Erhitzung. Zu diesem Zeitpunkt lebten die 5 Minuten auf 37~ erhitzten Algen noch zu 100%, w~ihrend die auf 40 ~ C erw~irmten Algen ausnahmslos abgestorben waren. Die auf 39 ~ C erw~irmten Algen waren nach vier Tagen auch schon zum Teil schwer und unregelm~it~ig gesch~idigt, w~ihrend die auf 38~ erhitzten Algen die Schwankungen der Hitzeresistenz besonders Mar erkennen liegen. Der si& fiber fiinf Tage erstre&ende Versu&, in dem viermal t~igli& die Hitzeresistenz yon immer neuen Algenproben bestimmt wurde, hatte die Auffindung eines vermuteten Tag-Nacht-Rhythmus zum Ziel. Es zeigten jedoch s&on die an den ersten zwei Tagen 24 Stunden ha& der Erhitzung auf 38~ auftretenden Resistenzschwankungen mit Maxima der Resistenz um 12 Uhr und um 24 Uhr und mit Minima in den Zeiten dazwischen, dafg bier kein 12-Stunden-Rhythmus, sondern ein 6-Stunden-Rhythmus vorliegt (Abb. 1 a). Glei&es gilt au& ffir die vier Tage nach der Erhitzung beoba&teten l[lber- lebensraten der auf 38~ erw~irmten Algen an den darauffolgenden Tagen (Abb. 1 b). Das vollst~indige Absterben der am 4. Juli um 12 Uhr und 20 Uhr erhitzten Algen f~illt noch in die oben erw~ihnte Zeit der anfangs grSt~eren Hitzeempfindlichkeit. Erst ab 24 Uhr dieses Tages ma&t si& der Eintritt in die dur& die hShere Temperatur des Kulturraumes bedingte Abh~irtungsphase bemerkbar. Bis zum 6. Juli um 20 Uhr wurde der Rhythmus der Erhitzungszeiten (8 Uhr [9 Uhr], 12 Uhr, 20 Uhr, 24 Uhr) beibehalten. Da die Resistenz- schwankungen dabei keine befriedigende Regelm~if~igkeit aufwiesen, wurden ab 6. Juli um 17.30 Uhr die Erhitzungsversuche zu den w~ihrend dieser Tage am natiirlichen Standort giiltigen Zeiten yon Hoch- und Niedrigwasser vor- genommen. Nun zeigte sich, dab die Punkte der hSchsten W~irmeresistenz genau mit der Zeit des Hochwassers und jene der niedersten Resistenz mit der des Niedrigwassers zusammenfielen. Die hochgelegenen Minima der zwei vorangegangenen Tage (5. und 6. Juli, jeweils 8 Uhr) lassen si& nun zwang- los dadur& erkl~iren, datg zu diesen Zeiten die Resistenz bereits wiederum im Ansteigen begriffen war. Die auf den Zustand vier Tage nach 5 Minuten Erhitzung auf 38~ bezogenen l[Iberlebenskurven (Abb. 1 b) zeigen somit eine klare Beziehung zu den Gezeiten: zum Zeitpunkt des Ho&wassers grSt~te und zum Zeitpunkt des Niedrigwassers geringste W~irmeresistenz. In auffallendem Gegensatz dazu steht die Lage der glei&falls mit den Zeit- punkten yon Ebbe und Flut wechselnden Maxima und Minima der Hitze- resistenz in den ersten zwei Tagen nach dem Einbringen der Algen yore Standort (Abb. 1 a). Hier fielen die gew~ihlten Erhitzungszeiten am 2. Juli um 20 Uhr, am 3. Juli um 9 Uhr und um 20 Uhr zuf~illig fast genau mit den Zeiten des hSchsten Wasserstandes zusammen. In allen drei F~illen zeigten

Studien zur Hitzeresistenz yon Chaetomorpha cannabina 463
jedoch hier die Algen ein Minimum ihrer Hitzeresistenz, w~ihrend die Maxima etwa mit den Zeiten der Ebbe zusammenfielen. Dies wi~rde gut mit Beob- achtungen am natfirlichen Standort ~ibereinstimmen, wo bei Ebbe an Pelvetia canaliculata und Fucus vesiculosus um 0,6o--0,7 ~ C h/Shere Hitzeresistenzen gefunden wurden als bei Flut (LuTOVA, ZAVADSKAYA, LUtINITSKAYA und FELDMANN 1967). Die in unseren Versuchen nach etwa zwei Tagen Vorkultur bei erh6hter Zim- mertemperatur (18~ ~ C) aufgetretene Umstimmung der mit den Gezei- ten zusammenfallenden periodischen Schwankungen der Hitzeresistenz yon Resistenzminimum zu Resistenzmaximum zur Zeit des Hochwassers und yon Resistenzmaximum zu Resistenzminimum zur Zeit des Niedrigwassers ist eine Erscheinung, die noch weiterer Untersuchung bedarf. Eine enge Beziehung zwischen Schwankungen der Hitzeresistenz und den Zeitpunkten yon Ebbe und Flut scheint aber, zumindest fiir die Tidepool-Alge Chaetomorpha can- nabina, aus diesen Versuchen mit Sicherheit hervorzugehen. Besonders hervor- zuheben ist, daf~ diese Beziehung au& na& Entnahme der Alge yon ihrem natiirlichen Standort bei mehrt~igiger Kultur unter Laborbedingungen aus- gepr~igt erhalten blieb.
3.4. B e e i n f l u s s u n g der H i t z e r e s i s t e n z d u r c h V o r k u l t u r in v e r s c h i e d e n k o n z e n t r i e r t e m S e e w a s s e r u n d b e i Z u c k e r z u s a t z
Untersuchungen fiber den Einflut~ verschieden zusammengesetzter N~ihrmedien oder einer Vorbehandlung mit verschiedenen Salz- oder Zu&erl/Ssungen auf die H/She der Hitzeresistenz liegen an Moosbl~ittchen und Gewebeschnitten yon Blfitenpflanzen schon in grofger Zahl vor (DE VRIES 1871, KAHO 1921, 1924, SAPPER 1935, REUTER 1937, SCHEIBMAIR 1938, BOGEN 1949, FELDMANN 1962 U. a.). An Chaetomorpha cannabina wurde der Einflufl verschiedener Seewasserkon- zentrationen auf die H/She der Hitzeresistenz dutch zweit~igige Vorbehand- lung mit verdtinntem und konzentriertem Seewasser und nachfolgende halb- stfindige Erhitzung auf Temperaturen zwischen 34 ~ und 40~ geprii~. Als ,1,0 Seewasser" galt dabei wieder das Wasser des Gastineau-Kanals n~ichst der Douglas Marine Station mit einer Salinit~it yon 13,425%0. Zur Unter- suchung einer Zuckerwirkung wurde diesem ,,1,0 S.W." 0,2 bzw. 0,4 tool Glukose zugesetzt. Mit diesen Zuckerl/Ssungen wurden die Algen nur 24 Stun- den vorbehandelt. Die Erhitzung wurde in den betreffenden L/Ssungen vor- genommen, und auch nach der Erhitzung wurden sie in den gleichen Seewasser- konzentrationen bzw. Zuckerl/Ssungen welter kultiviert und 24 Stunden sp~ter auf ihren Lebenszustand gepr[i&. Der osmotische ~'ert der Versuchsalgen entsprach beiVersuchsbeginn 3,35 S. W. Die in 4,0 S. W. eingetretenen leichten Abhebungen der Protoplasten waren innerhalb der zwei Tage Vorbehandlung dutch Endosmose zuriickgegangen.

In 0,1 S.W. war der osmotische Wert bis zum Zeitpunkt der Erhitzung auf 2,7--2,9 S.W. abgesunken. In 1,0 S.W. plus 0,4 mol Glukose war keine Plasmolyse eingetretem Das Ergebnis der Versuche ist in Tabelle 7 zusammengefaflt.
Tabelle 7. Chaetomorpha cannabina. Beeinflussung der Hitzeresistenz durch Vorbehandlung mit verdiinntem und konzentriertem Seewasser und dutch Zusatz yon Glukose (tool). Lebens- zustand 24 Stunden nach �89 Erhitzung
Vorbehandlung 2 Tage
34
0,1 Seewasser 0,3 Seewasser 0,5 Seewasser 0,7 Seewasser 1,0 Seewasser 2,0 Seewasser 3,0 Seewasser 4,0 Seewasser
Vorbehandlung 1 Tag
1,0 Seewasser 1,0 S.W. § 0,2 G1ukose 1,0 S.W. + 0,4 Glukose
35
1 1 1
36
Hitzeresistenz, ~ C, ~ Stunde
37
90% 50% 2%
~ r 1
1
90% 90% lO%
464 R. BIEBL
38
+ + +
98% 95% 2%
1 90% 1 95% 1 90%
39
7-
+ _ + +
§ § +
(1 alles lebend, % tot, + = alles tot)
40
§
+ + + + + +
70%
+ § 7-
Aus der in Tabelle 7 gegebenen Ubersicht ist abzulesen, daft zunehmende Ver- diinnung des Seewassers eine stufenweise Herabsetzung der Hitzeresistenz und steigende Seewasserkonzentration eine Zunahme der Hitzeresistenz be- wirkt. Die Vorbehandlung mit zu&erhaltigem Seewasser blieb hingegen ohne Einflufl auf diese.
4. B e s p r e c h u n g d e r V e r s u c h e
Die Untersuchung der Hitzeresistenz und des Absterbeverlaufs na& ver- schieden langer Erhitzungsdauer war yon Interesse, um die in der Literatur vorliegenden, mit unterschiedlichen Erhitzungszeiten erhaltenen Resistenz- werte miteinander vergleichen zu kiSnnen. Den bisher durchgefiihrten eigenen Versu&en zur Hitzeresistenz yon Meeresalgen (Ioc. cir.) lag fast ausnahmslos eine zw61fstiindige Erhitzungsdauer zugrunde. Sctt6LM (1966) beniitzte bei ihren Bestimmungen der Hitzeresistenz yon Tiefenalgen in La Jolla (Cali- fornien), wie dies bei Landpflanzen iiblich ist (LANGE 1959, 1961, BIEBL 1964, 1968 a u. a.), eine halbstiindige Erhitzungsdauer, w~ihrend russische Autoren
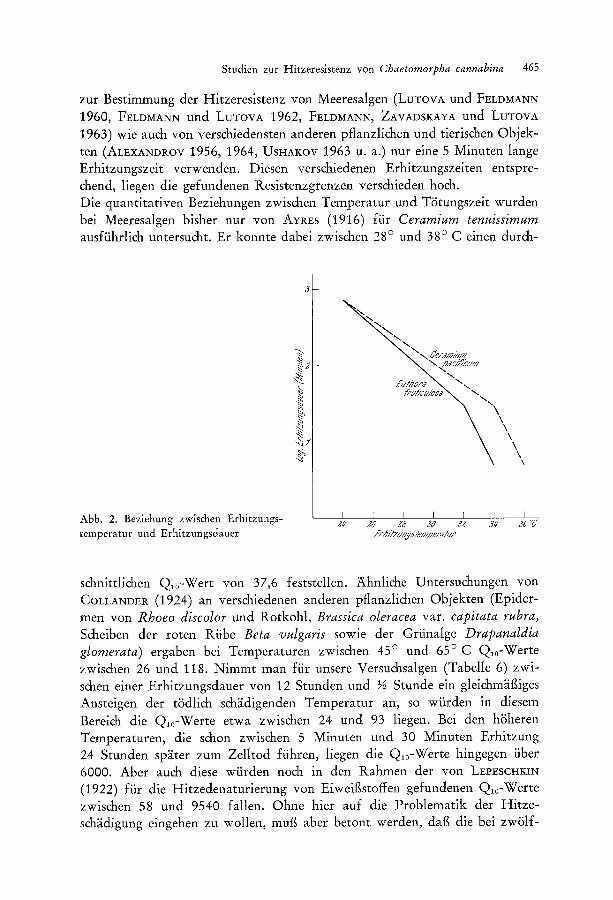
Studien zur Hitzeresistenz yon Chaetomorpha cannabina 465
zur Bestimmung der Hitzeresistenz yon Meeresalgen (LuTovA und FELDMAI'~N 1960, FELDMA~N und LUTOVA 1962, FELDMANN, ZAVADSKAYA und LUTOVA 1963) wie auch yon verschiedensten anderen pflanzlichen und tierischen Objek- ten (ALExANDROV 1956, 1964, USHAKOV 1963 U. a.) nur eine 5 Minuten lange Erhitzungszeit verwenden. Diesen verschiedenen Erhitzungszeiten entspre- &end, liegen die gefundenen Resistenzgrenzen verschieden hoch. Die quantitativen Beziehungen zwischen Temperatur und T~Stungszeit wurden bei Meeresalgen bisher nur yon AYREs (1916) fiir Ceramium tenuissimum ausftihrlich untersucht. Er konnte dabei zwischen 28 ~ und 38 ~ C einen durch-
Abb. 2. Beziehung zwischen Erhitzungs- temperatur und Erhitzungsdauer
J/ Zulkora ~ \ \
/bu/~-i//os J ~ x . .
\ \
I l I I I I I zr zd z8 30 gd 34 3s ~
{,,'ll/Tz~injJe/np~r<~tur
schnittlichen Q10-Wert yon 37,6 feststellen. Ahnliche Untersuchungen yon COLLANDER (1924) an verschiedenen anderen pflanzlichen Objekten (Epider- men yon Rhoeo discolor und Rotkohl, Brassica oleracea var. capitata rubra, Scheiben der roten Riibe Beta vulgaris sowie der Grfinalge Drapanaldia glomerata) ergaben bei Temperaturen zwischen 45 ~ und 65~ Q10-Werte zwischen 26 und 118. Nimmt man ftir unsere Versuchsalgen (Tabelle 6) zwi- schen einer Erhitzungsdauer yon 12 Stunden und �89 Stunde ein gleichm~it~iges Ansteigen der t~Sdlich sch~idigenden Temperatur an, so wfirden in diesem Bereich die Q10-Werte etwa zwischen 24 und 93 liegen. Bei den h?Sheren Temperaturen, die schon zwischen 5 Minuten und 30 Minuten Erhitzung 24 Stunden sp~iter zum Zelltod fiihren, liegen die Q10-Werte hingegen fiber 6000. Aber auch diese wfirden noch in den Rahmen der yon LEPESCHKIN (1922) ffir die Hitzedenaturierung yon Eiweif~stoffen gefundenen Q10-Werte zwischen 58 und 9540 fallen. Ohne bier auf die Problematik der Hitze- sch~idigung eingehen zu wollen, muf~ aber betont werden, daf~ die bei zw~51f-

466 R. BIEBL
sfiindiger Einwirkungsdauer gefundenen T6tungstemperaturen von 24 ~ bis 26 ~ C, ja auch noch jene urn 30 ~ C, viel zu niedrig sind, um an eine Protein- koagulation denken zu k6nnen. Es mul~ sich hier wohl um sch~idigende Ein- wirkungen auf h~Sher geordnete Strukturen, etwa Lipoidkomplexe in den Plasmamembranen oder ~ihnliches, handeln. In Abb. 2 sind f~ir zwei der in Tabelle 6 angef~ihrte Algen die Versuchs- temperaturen auf der Abszisse und die Logarithmen der zugeh/Srigen T~Stungs- zeiten auf der Ordinate eines Koordinatensystems aufgetragen. Sie erlauben die direkte Ablesung der Q10-Werte, das heifer jener Faktoren, um die die Reaktionsgeschwindigkeit, in unserem Fall die T6tungsgeschwindigkeit, bei Temperaturerh6hung um 10~ zunimmt. F~ir Algen der Gezeitenzone, in der Ebbe und Flut in einem Rhythmus yon etwas fiber 6 Stunden wechseln, ist eine zw/Slfstfindige Erhitzungsdauer lange genug, um ihre maximale Hitzeresistenz w~ihrend Niedrigwasser zu erfassen. Auch die Tiefenalgen lassen nach 12 Stunden Erhitzung 8kologisch unmittel- bar auswertbare Unterschiede der Temperaturresistenz erkennen. Der Vorteil der zw~Slfsti~ndigen Erhitzungszeit ist somit, dai~ sie direkte R~ickschl[isse auf am Standort noch tragbare Wassertemperaturen zulassen. Der Nachteil liegt aber aul%r in der langen Versuchsdauer auch in der UnmSglichkeit, dadurch irgendwelche kurzfristige Resistenzschwankungen erkennen zu k~Snnen. Die durch halbsti~ndige Erhitzung gefundenen Resistenzgrenzen liegen bei Meeresalgen im allgemeinen 6 ~ bis 10~ hSher als nach 12 Stunden Er- w~rmung (Tabelle 6). Auch sie sind fiJr ~Skologische Fragestellungen gut ver- wendbar. Sie erlauben einen Vergleich der graduellen Unterschiede der Hitzeresistenz yon Algen verschieden temperierter Standorte. R[ickschli~sse auf die am natiJrlichen Standort tragbaren H~Schsttemperaturen lassen sie aber nicht mehr zu. Der groi~e Vorteil der halbsttindigen Erhitzungsdauer liegt in der MSglichkeit, in kurzer Zeit eine gr/513ere Erhitzungsreihe durch- zuftihren. Dieser Vorteil trifft in noch h6herem Mat~e auf eine Erhitzungsdauer von nur 5 Minuten zu. Hier liegen die Resistenzgrenzen durchschnittlich wieder um 1 ~ bis 2 ~ h6her als nach halbsttindiger Erhitzung (Tabelle 6). Zur Auf- deckung yon kurzfristigen Schwankungen der Hitzeresistenz im Tagesablauf ist eine so kurze Erhitzungsdauer sogar die einzig brauchbare. Ftir die Festsetzung der Resistenzgrenze (= Lebensgrenze) ist es wichtig, eine minimale Nachwirkungsdauer der Erhitzung verstreichen zu lassen. Bei den Hitzeresistenzbestimmungen an Landpflanzen hat sich eine drei- bis fiinf- t~igige Nachkultur bewihrt. Eine so lange Nachkultur stS~t bei Meeresalgen vielfach dadurch auf Schwierigkeiten, dab empfindlichere Algen nach dieser Zeit schon in den unerhitzten Kontrollen abzusterben beginnen. Die Beobach- tung der Absterbeverl~iufe (Tabelle 4 und 5) zeigt aber, dat~ bei strengem Mai~stab, wobei schon ein Absterben yon wenigen Prozent der erhitzten Zellen als Elberschreiten der Resistenzgrenze angesehen wird, ein Bezug auf den

Studien zur Hitzeresistenz yon Chaetomorpha cannabina 467
Lebenszustand 24 Stunden nach der Erhitzung im allgemeinen ausreicht. Zu- mindest bei 5 Minuten langer Erhitzung (Tabelle 4) zeigt es sich, dafg die nach 24 Stunden nur geringffigig gesch~idigten Algen in den nachfolgenden vier Tagen noch vollst~indig absterben, w~hrend die nach 24 Stunden zu 100% lebenden Algen auch in den nachfolgenden vier Tagen ungesch~digt bleiben. Diese minimale Nachwirkungsdauer yon 24 Stunden wird jedoch unbedingt einzuhalten sein, urn einigermal~en zuverl~issige und vergleichbare Resistenz- grenzen zu erhalten. Beobachtungen tiber Anderungen der Hitzeresistenz in Abh~ingigkeit yon ver- schiedenen inneren und ~iut~eren Faktoren liegen schon in groi%r Zahl vor (Zusamrnenfassungen LEVITT 1956, BIrBL 1962 a, LANCE 1965, 1967). Jahreszeitliche Schwankungen der Hitzeresistenz wurden nachgewiesen an Bliitenpflanzen (LANG~ 1961), an Farnen (KAVVrN 1964), an Moosen (LANGr 1955, DIRCI~SrN 1964), an Meeresalgen (FrLDlVIANN und LuTOVA 1963, LuvovA, ZAVADSlIAYA, LUXNITSKAYA und FELDlvIANN 1967) und an Siif~- wasseralgen (ScHoLM 1968 a). An Blfitenpflanzen (BIrBL 1967) und an Si/J~- wasseralgen (SCH6LM 1968 a) konnten im Sommer bei gleichbleibender Tem- peratur durch ffinf bis zehn Tage Kurztagbehandlung herbstliche Resistenz- verh~iltnisse ausgel~Sst werden. Tagesschwankungen der Hitzeresistenz wurden demgegenfiber erst seltener beobachtet. So konnten SCHWEMMrE und LANGr (1959 a, b) einen endogen- tagesperiodischen Verlauf der Hitzeresistenz der Bl~itter von KalanchoF blossfeldiana feststellen. Nach 30 Minuten Erhitzung der Sprosse in einem Warmbad yon 46~ zeigten sich im 12 : 12-Stunden-Licht-Dunkel-Wechsel Unterschiede im Ausmaf~ der in Prozenten der Blattfl~iche ausgedrfickten Sch~digung. Die Zeiten geringerer Hitzeresistenz fielen dabei immer mit an der gleichen Pflanze beobachteten Zeiten h~Sherer Stoffwechselaktivit~it zusam- men (Fermentaktivitiit: EHRrNBrr, G 1954, Wuchsstoffgehalt der Bl~itter: B~clIEl~ 1953, Guttationsrhythmik: I-IEIMANN 1950). SCH6L~ land tagesperi- odische Schwankungen der Hitzeresistenz an Spirogyra und Rhizoclonium (1968 a) und gleichlaufend damit _Knderungen der sichtbaren Fl~iche der Zell- kerne von Spirogyra (1968 b). Die Zeiten des Resistenzminimums stimmten dabei ann~ihernd mit jenen der Maxima der Kerngr/Sgen zusammen, denen wahrscheinlich wieder Zeiten gr61~erer Stoffwechselaktivit~it in den Zellen entsprechen. Die an Chaetomorpha cannabina in einer sich fiber fiinf Tage erstreckenden Versuchsreihe gefundene, im zeitlichen Abstand yon etwa 6 Stunden schwan- kende Hitzeresistenz (Abb. 1 a und b) kann nicht auf den Wechsel yon Tag und Nachr zurfickgeffihrt werden. Bei dieser sechsstiindigen Periodizit~it han- delt es sich im Gegensatz zur ,,Circadianen Rhythmik" (HArBrRG 1959, 1963) um den Ausdruck einer typischen ,,Gezeitenrhythmik", wie solche an tieri- sdaen marinen Organismen schon in verschiedener Hinsicht beobachtet werden konnte (B/2NNING 1960, 1963, FINGERMANN 1960).

468 R. BIEBL
Ein Ansteigen der Hitzeresistenz w~ihrend der Ebbe und ein Absinken bei Flut wurde an Gezeitenalgen auch schon am natiirlichen Standort beobachtet (LUTOVA, ZAVADSKAYA, LUKNITSKAYA und FELDMANN 1967), jedoch bracMe man diese Schwankungen mit der w~ihrend der Ebbe grSfteren Erw~irmung und dem Wasserverlust der Algen am naffirlichen Standort in Zusammenhang. Unsere Versuche wurden jedoch mit einer Tidepool-Alge ausgefiihrt, die wiihrend der Versuchstage den am Standort im Verlauf der Gezeiten wech- selnden Auftenbedingungen durch die Kultur in Glasgef~iften im Laboratorium entzogen waren. Die Algen erfuhren nach den ersten zwei Tagen in Anpas- sung an die hShere Zimmertemperatur eine Erh/Shung ihrer Hitzeresistenz. Abet unabh~ingig davon blieb ein ungef~ihr sechsstiindiger Rhythmus des Wechsels der Hitzeresistenz erhalten. Will man nicht eine so lange andauernde Na&wirkung der sich am natiirlichen Standort bei Ebbe und Hut ~indernden Auftenbedingungen annehmen, so mug man die unver~indert anhaltenden und zur Erscheinung der Gezeiten fiihrenden kosmischerl Bedingungen selbst fiir diese Resistenzschwankungen verantwortlich machen. Es ist dabei sehr wahr- scheinlich, daft auger der W~irmeempfindlichkeit auch andere physiologische Eigenschaften der Alge Schwankungen im gleichen Rhythmus mitmachen. Zur Sicherung dieses an Chaetomorpha cannabina beobachteten Ph~irtomens w~ire es vorerst nStig, mit dem bisher gefundenen Test der W~irmeresistenz Untersuchungen an anderen Gezeitenalgen vorzunehmen. Sollte sich diese Erscheinung bei den Gezeitenalgen allgemein finden und sollte sich diese im Rhythmus der Gezeiten wechselnde Hitzeresistenz nicht allein durch die sich mit Ebbe und Flut ~indernden Standortfaktoren erkl~iren lassen, so w~ire zu erwarten, daft auch die st~indig yore Wasser bedeckten Algen des Sublitorals eine gleiche Gezeitenrhythmik ihrer Hitzeresistenz zeigen. Es w~ire welters auch zu prtifen, ob nicht manche an Landpflanzen beobachtete Schwankungen physiologischer Eigenschaften, die nicht mit dem Tag-Nacht-Rhythmus iiber- einstimmen, einer solchen Gezeitenrhythmik folgen oder dutch eine solche gestSrt werden (vgl. BONNINO 1963). Die Kr~ifte des Mondes, die an den Meereskiisten zu der auffallenden Erscheinung yon Ebbe und Flut fiihren, sind ja im Binnenland in gleicher Weise wirksam. Es mug noch hervorgehoben werden, daft der beobachteten ,,Gezeitenrhythmik der W~irmeresistenz" yon Chaetornorpha cannabina kaum 5kologische Bedeu- tung zukommen diirfte. Dazu sind die Schwankungen zu gering (Abb. 1 a und b). Wenn bei Gezeitenalgen, die bei Ebbe vollkommen freiliegen, grtSftere und tSkologisch bedeutungsvolle Schwankungen der W~irmeresistenz auftreten, so wird dies wahrscheinlich auf andere Faktoren, wie Wiirmeh~irtung durch erhShte Standorttemperatur, Austrocknung oder vielleicht auch ansteigenden Salzgehalt, zur~ickzuftihren sein. Dies leitet zu einer letzten Versuchsgruppe iiber, in der der Einfluf~ verd[inn- ten und konzentrierten Seewassers sowie yon Glukosezusatz zum Seewasser

Studien zur Hi tze res i s t enz y o n Chaetomorpha cannabina 469
auf die H6he der W~irmeresistenz yon Chaetornorpha cannabina untersucht wurde (Tabelle 7). Zwei Tage Vorkultur in verd/inntem Seewasser fiihrte zu einer mit dem Ver- d~innungsgrad stufenweise abnehmenden und gleich langen Vorkultur, in ver- schieden konzentriertem Seewasser zu einer mit der Konzentration zuneh- menden Hitzeresistenz. LEVITT (1956) sieht die Ursache der yon verschie- denen Autoren beobachteten Erh6hung der Hitzeresistenz durch Vorbehand- lung mit konzentrierten Salzl6sungen in der dadurch bedingten Entw~isserung des Protoplasmas und einer damit verbundenen Inaktivierung yon Enzymen oder Stoffwechselprozessen. SCH~iEP~R und KowA~sliI (1956) fanden die Hitzeresistenz des Kiemengewebes der Nordsee-Miesmuschel in Meerwasser yon 30%o Salzgehalt etwa doppelt so groi~ als die der Ostsee-Miesmuschel in Brackwasser yon 15%o Salzgehalt. In weiteren Versuchen konnte festgestellt werden, dal~ es dabei wesentlich auf den Calciumgehalt des Seewassers an- kommt. Hielt man bei einer Verd~nnung des Seewassers auf die H~ilfte der urspriinglichen Salzkonzentration den Calciumgehalt konstant, so blieb die Hitzeresistenz des Kiemengewebes praktisch unver~indert. Diese resistenzf~Srdernde Wirkung des Calciums wurde auch an pflanzlichcn Objekten beobachtet. SCH~IBMAIR (1938) konnte zeigen, daf~ Bl~ittchen yon Plagiochila asplenioides durch Vorbehandlung mit KC1-L6sungen eine Herab- setzung, durch Vorbehandlung mit CaCl_~-L6sungen hingegen eine Erh6hung ihrer Hitzeresistenz erfuhren. BOGEN (1949) beobachtete an Blattepidermis- schnitten yon Rhoeo discolor in hypo- und hypertonischen KC1-L6sungen gleichfalls eine Herabsetzung, in MgSO4-L6sungen hingegen eine Steigerung der Hitzeresistenz. Wiisserung der Schnitte bedingte stets eine Verminderung der Hitzeresistenz. Es wird daher auch in unseren Versuchen miSglicherweise nicht so sehr auf die H6he der Gesamtkonzentration der L6sungen als auf die Vermehrung oder Verminderung bestimmter Ionen, etwa des Calciums und Magnesiums, innerhalb der L6sungsreihe ankommen. Diese Erh6hung der Hitzeresistenz durch zunehmenden Salzgehalt k6nnte fiir Gezeitenalgen bei Ebbe auch yon 6kologischer Bedeutung sein. Ein Tag Vorbehandlung yon Chaetomorpha mit Seewasser, dem 0,2 bzw. 0,4 mol Glukose zugesetzt war, hatte keinen Einflul~ auf die H6he ihrer Hitzeresistenz. Dies steht im Gegensatz zu Beobachtungen yon FELDMANN (1962), die an Epidermiszellen yon Tradescantia flurninensis und Blatt- scheidenzellen von Dactylis glomerata und Elyrnus arenarius nach Einlegen der Blattstiickchen in hyper- oder isotonische L6sungen yon Saccharose, Glukose oder Laktose eine Erh6hung der Hitzeresistenz feststellen konnte. Da eine Schutzwirkung des Zuckers nur eintreten kann, wenn dieser in das Protoplasma einzudringen vermag, so k6nnte die fehlende Schutzwirkung des Zuckers bei Chaetomorpha durch eine Impermeabilitiit des Plasmas ffir Glukose erkl~irt werden.

470 R. BIEBL
L i t e r a t u r
ALrXANDI~OV, W. Y., 1956: Cytophysiologische Analyse der W~irmeresistenz yon Pflanzen- zellen und einige Probleme der CytoSkologie (russisch). Bot. J. (Moskau) 41, 939--961.
- - 1964: Cytophysiological and cytoecological investigations of resistance of plant cells toward the action of high and low temperature. Quart. Rev. Biol. 39, 35--77.
AYRES, A. H., 1916: The Temperature Coefficient of the Duration of Life of Ceramium tenuissimum. Bot. Gaz. 62, 65--69.
B~CKeR, T., 1953: Wuchsstoff- und S~iureschwankungen bei Kalancho8 blossfeldiana in ver- schiedenem Licht-Dunkel-Wechsel. Planta 43, 1--24.
BIEBL, R., 1937: Ukologische und zellphysiologische Studien an Rotalgen der englischen Sfidktiste. Beih. Bot. Centralbl. 57, Abt. A, 381--424.
- - 1 9 3 9 : f3ber die Temperaturresistenz yon Meeresalgen verschiedener Klimazonen und verschieden tiefer Standorte. Jahrb. wiss. Bot. 88, 389--420.
- - 1958: Temperatur- und osmotische Resistenz yon Meeresalgen der bretonischen Kiiste. Protoplasma 50, 217--242.
- - 1962 a: Protoplasmatische (Skologie der Pflanzen. I. Wasser und Temperatur. Protoplas- matologia, Handb. d. Protoplasmaforsch. 12/1, 344 S., Wien: Springer.
- - 1962 b: Protoplasmatisch-~Skologische Untersuchungen an Mangrovealgen yon Puerto Rico. Protoplasma 55, 572--606.
- - 1 9 6 2 c : Temperaturresistenz tropischer Meeresalgen (verglichen mit jener yon Atgen temperierter Meeresgebiete). Bot. mar. 4, 241--254.
- - 1964: Temperaturresistenz tropischer Pflanzen auf Puerto Rico. Protoplasma 59, 133--156. - - 1 9 6 7 : Kurztag-Einflfisse auf arktische Pflanzen w~hrend der arktischen Langtage.
Planta 75, 77--84. - - 1968: Uber W~irmehaushalt und Temperaturresistenz arktischer Pflanzen. Flora, Abt. B,
157, 327--354. BoorN, H. J., 1949: Untersuchungen fiber Hitzetod und Hitzeresistenz pflanzlicher Proto-
plaste. Planta 36, 298--340. BISNNING, F,., 1960: Biological clocks, Opening address. In: Cold Spring Harbor Symposia
on Quantitative Biology. Vol. 25, 1--9. - - 1963: Die physiologische Uhr, 2. Auflage, 153 S., Berlin-GtSttingen-Heidelberg: Springer-
Verlag. COLLANDEt~, R., 1924: Beobachtungen fiber die quantitativen Beziehungen zwischen T6tungs-
geschwindigkeit und Temperatur beim Wiirmetod pflanzlicher Zellen. Soz. Scient. Fenn., Comlnentationes Biologicae 1, 1--12.
DIRCKSEN, A., 1964: Vergleichende Untersuchungen zur Frost-, Hitze- und Austrocknungs- resistenz einheimischer Laub- und Lebermoose unter besonderer Berficksichtigung jahres- zeitlicher Ver~tnderungen. Dissertation GSttingen.
Em~eNBEI~G, M., 1954: Einflut~ verschiedenen Licht-Dunkel-Wechsels auf die Rhythmik der PhosphataseaktivitS.t in den Bl~ittern von KalanchoF blossfeldiana. Planta 43, 528--536.
F~LDMANN, N. J., 1962: Der Einfiuf~ der Zucker auf die Resistenz einiger h/Sherer Pflanzen- zellen gegen Erwiirmung und hohen hydrostatischen Druck. Cytologia (Moskau) 4, 633--643 (russisch).
- - et M. I. LuTova, 1963: Variations de la thermostabilit6 cellulaire des Algues en fonctlon des changements de la temperature du milieu. Cahiers de Biologic marine 4, 435--458.
- - I. G. ZAVADSKAYa, and M. I. LUTOVA, 1963: A Study of Temperature Resistance of some Marine Algae under Natural and Experimental Conditions. Cytology (U.S.S.R.) 5, 125--134.
FINGER, AN, M., 1960: Tidal Rhythmicity in Marine Organisms. In: Cold Spring Harbor Symposia on Quantitative Biology. Vol. 25, 481--489.

Studien zur Hitzeresistenz yon Chaetomorpha cannabina 471
HAL~Et~ G F., 1959: Physiologic 24-hour periodicity; General and procedural considerations with reference to the adrenal cycle. Z. Vitamin-, Hormon- und Fermentforsch. 10, 225--296.
- - Y . L. TONG, and E. A. JOHNSON, 1965: Circadian System Phase--An Aspect of Temporal Morphology; Procedures and Illustrative Examples. The Cellular Aspects of Biorhythms. Symposium on Rhythmic Research, Wiesbaden 1965, 19--48.
HEIMANN, M., 1950: Einflut~ periodischer Beleuchtung auf die Guttationsrhythmik. Planta 38, 157--195.
KAHO, H.,~ 1921: ~ber -d ie Beeinflussung der Hitzekoagu~ation des Pflanzenplasmas durch Neutraisalze. Biochem. Z. 117, 87--95.
- - 1924: l~ber die Beeinflussung der Hitzekoagulation des Pflanzenplasmas durch Salze der Erdalkalien. Biochem. Z. 151, 102--1ll.
KAPP~N, L., 1963: Untersuchungen tiber den Jahresverlauf der Frost-, Hitze- und Austrock- nungsresistenz yon Sporophyten einheimischer Polypodiaceen (Filicinae). Flora 155, 123--166.
K~ss~L~R, H., 1959: Mikrokryoskopische Untersuchungen zur Turgorregulation "con Chaetomorpha Iinum. Kieler Meeresforsch. 15, 51--73.
- - 1964: Die Bedeutung einiger anorganischer Komponenten des Seewassers ftir die Turgor- regulation yon Chaetornorpha linum (Cladophorales). Helgol. Wiss. Meeresunters. 10, 73--90.
K~CLIN, H., 1917: Uber die K~ilteresistenz der Meeresalgen. Ber. dtsch, bot. Ges. 35, 370--384.
LANGG O. L., 1955: Untersuchungen iiber die Hitzeresistenz der Moose in Beziehung zu ihrer Verbreitung. I. Die Resistenz stark ausgetrockneter Moose. Flora 142, 381--399.
- - 1959: Untersuchungen fiber den W~irmehaushalt und die Hitzeresistenz mauretanischer Wiisten- und Savannenpflanzen. Flora 147, 595--651.
- - 1961: Die Hitzeresistenz einheimischer immer- und wintergriiner Pflanzen im Jahres- ablauf. Planta 56, 666--683.
- - 1965: The Heat Resistance of Plants, its Determination and Variability. Methodology of Plant-Ecophysiology. Proceedings of the Montpellier Symposium. Unesco 1965, 399--405.
- - 1967: Investigations on the Variability of Heat-Resistance in Plants. In: A. S. TROSCmN, The Cell and Environmental Temperature, 1967, 131--141.
LEV~SCHKIN, W. W., 1922: Uber die Denaturation der Eiweif~stoffe bei der Hitzekoagulation. Kolloidzeitschr. 31, 342--345.
LEVITT, j., 1956: The Hardiness of Plants. 278 S. New York: Acad. Press.
LUTOVA, M. I., and N. L. F~LD~A~N, 1960: A Study of the Ability to Temperature Adap- tation in some Marine Algae. Cytologia (Moskau) 2, 699--709 (russisch).
- - I. G. Z~VADSKAYA, A. F. LUKmTSKAY& and N. L. F~LD~ANN, 1967: Temperature Adap- tation of Cells of Marine and Freshwater Algae. In: A. S. Tt~oscmN, The Cell and Environmental Temperature, 166--172.
P~CXACD, G. L., 1967: Some Oceanographic Characteristics of the Larger Inlets of Southeast Alaska. J. Fish. Res. Bd. Canada 24 (7), 1475--1506.
REUTER, L., 1937: Protoplasmatik vergilbender Bl~itter. Protoplasma 27, 270--279.
SAPPEt~, I., 1935: Versuche znr Hitzeresistenz der Pflanzen. Planta 23, 518--556.
SCAGEL, R. F., 1957: An Annotated List of the Marine Algae of British Columbia and Northern Washington. Canada Dept. of Northern Affairs and Nat. Resources. Nat. Museum of Can. Bulletin 150, Biolog. Ser. 52, 289 pp.
SCH~IB~AI~, G., 1938: Hitzeresistenz-Studien an Moos-Zellen. Protoplasma 29, 394--424.
ScHbc~, H. E., 1966: Untersuchungen zur W~rmeresistenz yon Tiefenalgen. Bot. mar. 9, 54--61.
Protoplasma 67/4 32

472 R. BIEBL: Studien zur Hitzeresistenz yon Chaetomorpha cannabina
SCHOLM, H. ]~., 1968 a: Untersuchungen zur Hitze- und Frostresistenz einheimischer Stif~- wasseralgen. Protoplasma 65, 97--118.
- - 1 9 6 8 b : l]ber tagesperiodische .Anderungen der sichtbaren FP, tche yon Nucleus und Nucleolus einheimischer Stif~wasseralgen und ihre Beeinfluf~barkeit. Protoplasma 66, 393--4O4.
SCHWEMML~, B., und O. L. LANOZ, 1959 a: Neue Beobachtungen tiber die endogene Tages- rhythmik (_Anderungen der Hitzeresistenz bei KaIanchoF blossfeldiana). Nachr. Akad. Wiss. G&tingen II, math.-phys. K1., 29--35.
1959b: Endogen-tagesperiodische Schwankungen der Hitzeresistenz bei KalanchoF blossfeldiana. Planta 53, 134--144.
SCH\WNKZ, H., 1959: Untersuchungen zur Temperaturresistenz mariner Algen der westlichen Ostsee. I. Das Resistenzverhalten yon Tiefenrotalgen bei tSkologischen und nicht-~Skologi- schen Temperaturen. Kieler Meeresforsch. 15, 34--50.
T~RUMOTO, J., 1964: Frost Resistance in some Marine Algae from the Winter Intertidal Zone. Low Temperature Science, Ser. B, 22, 19--28 (japan. with english summary).
Us~tA~ov, B. P., 1968: Cellular Resistance Adaptation to Temperature and Thermostability of Somatic Cells with Special Reference to Marine Animals. Marine Biology 1, 153--160.
Vt~izs, H. D~:, 1871: Sur le mort des cellules v6g&ales par l'effet d'une temp6rature 61ev6e. Arch. n6erl. Sei. exact et natur. 6, 245.
Anschriflc des Verfassers: Prof. Dr. R. BIEBL, Pflanzenphysiologisches Institut der Universit~it Wien, Dr. Karl Lueger-Ring 1, A-1010 Wien.