studies on the involvement of the yeast abc transporter pdr18 in … · yeast abc transporter pdr18...
TRANSCRIPT

Studies on the involvement of the yeast ABC transporter Pdr18 in acetic acid tolerance
at temperatures in the range 30-40°C
Catarina Alexandra Simões Prata
Thesis to obtain the Master of Science Degree in
Biotechnology
Supervisor: Prof. Dr. Isabel Maria de Sá Correia Leite de Almeida
Co-supervisor: Dr. Margarida Isabel Rosa Bento Palma
Examination Committee
Chairperson: Prof. Dr. Arsénio do Carmo Sales Mendes Fialho
Supervisor: Prof. Dr. Isabel Maria de Sá-Correia Leite de Almeida
Member of the Committee: Prof. Dr. Miguel Nobre Parreira Cacho Teixeira
November 2014

i
Acknowledgments
Firstly, I would like to acknowledge Professor Isabel Sá-Correia, as my supervisor and head of
Biological Sciences Research Group (BSRG) of the Institute for Biotechnology and Bioengineering
(IBB), Instituto Superior Técnico (IST), for giving me the opportunity to join BSRG, where this work
was developed. I am most thankful for her guidance, as well as for her prompt availability and
willingness to support me during the development of this thesis.
I would like to thank Dr. Margarida Palma for all the help throughout this work and for being always
available for me. Thank you for sharing your knowledge and experience with me.
This work was financially supported by “Fundação para a Ciência e a Tecnologia” (FCT): contract
ERA-IB/0002/2010 (INTACT – Integral Engineering of Acetic Acid Tolerance in Yeast), in the frame of
ERA-NET Industrial Biotechnology.
The following personal acknowledgements will be addressed in Portuguese:
Gostaria de agradecer a todos os membros do BSRG pelo excelente companheirismo e pelo
ambiente saudável e de entreajuda propício ao desenvolvimento de trabalhos de qualidade, o que
sem dúvida contribuiu para a realização deste trabalho. Um agradecimento muito especial à
Margarida Palma por todo o seu apoio e amizade e, principalmente, pela partilha de conhecimentos e
paciência. Não podia deixar de agradecer à Cláudia Godinho, pela sua disponibilidade e ajuda na
execução deste trabalho.
Quero também agradecer aos meus colegas de mestrado e de laboratório: Diana, José, Nicole, Pedro
e Sara; pela companhia e partilha dos bons e maus momentos.
Um agradecimento especial ao Fábio, pelo seu apoio e compreensão, que me ajudou a ultrapassar
as dificuldades que surgiram ao longo deste percurso. Por último, mas não menos importante,
agradeço aos meus pais, avó e restante família e amigos, por toda a força e ajuda incondicional que
foram preponderantes para a conclusão deste trabalho. Muito obrigada por tudo.
Catarina Prata

ii
Abstract
The occurrence of multidrug resistance (MDR) frequently relies on the action of membrane
transporters presumably involved in the active efflux of unrelated cytotoxic compounds out of the cell.
Although this is a major concern in the treatment of cancer and infection diseases, the biological
activity of multidrug transporters has also advantages, such as the improvement of the ability of
industrial strains to tolerate multiple stresses of biotechnological relevance.
Pdr18 is a plasma membrane MDR transporter of the ATP-binding cassette (ABC) superfamily
described as playing a role in ergosterol incorporation in Saccharomyces cerevisiae plasma
membrane. In this thesis, the physiological role of Pdr18 in yeast tolerance to acetic acid was
examined. PDR18 expression was found to increase yeast tolerance to acetic acid, to contribute to
counteract the dissipation of plasma membrane potential and the permeabilization of plasma
membrane in yeast cells growing in the presence of inhibitory concentrations of this weak acid.
The effect of PDR18 expression in the growth curve of yeast cells in growth medium supplemented
with increasing concentrations of acetic acid at temperatures in the range 30-40°C was also examined.
Results suggest that: i) the presence of acetic acid leads to the decrease of the optimum and
maximum temperature for growth; ii) the minimum inhibitory concentrations (MIC) for acetic acid
decreases as temperatures increases; and iii) PDR18 expression leads to the increase of yeast
robustness to the conjugated effect of these environmental stresses and that Pdr18 exerts a protective
effect against the additive deleterious action of acetic acid and supraoptimal temperatures.
Keywords: Saccharomyces cerevisiae; multidrug resistance; drug efflux pumps; ATP-binding cassette (ABC); Pdr18; ergosterol; yeast response to stress; supraoptimal temperatures.

iii
Resumo
O mecanismo de aquisição de resistência a múltiplas drogas é frequentemente dependente da ação
de transportadores membranares presumivelmente envolvidos no efluxo de diferentes compostos
citotóxicos. Deste fenómeno biológico resultam problemas graves no tratamento de cancro e doenças
infecciosas, sendo o seu estudo considerado de elevado interesse. Contudo, a atividade de
transportadores de resistência a múltiplas drogas pode também representar vantagens, tal como a
capacidade de estirpes industriais desenvolverem tolerância a diversos stresses químicos de
interesse biotecnológico.
Pdr18 é um transportador de resistência a múltiplas drogas que pertence à superfamília ATP-binding
cassette (ABC). O seu papel foi descrito na incorporação de ergosterol na membrana plasmática da
levedura Saccharomyces cerevisiae. Neste trabalho foi estudado o papel fisiológico do Pdr18 na
tolerância da levedura ao ácido acético. A expressão do gene PDR18 demonstrou ser essencial para
a maior tolerância da levedura a ácido acético, contribuindo para contrariar o efeito do ácido acético
na dissipação do potencial de membrana e no aumento da permeabilidade da membrana plasmática
em células de levedura crescidas na presença de concentrações inibitórias de ácido acético.
O efeito da expressão do gene PDR18 foi estudado no crescimento de células de levedura a
temperaturas entre 30°C e 40°C, na presença de concentrações crescentes de ácido acético. Os
resultados obtidos sugerem que: i) a presença de ácido acético leva à diminuição das temperaturas
ótima e máxima de crescimento; ii) a concentração mínima inibitória de ácido acético diminui com o
aumento da temperatura; e iii) a expressão do gene PDR18 resulta no aumento da robustez das
estirpes em relação ao efeito conjugado de ambos os stresses, e que o Pdr18 exerce um efeito
protetor contra a ação nefasta aditiva do ácido acético e de temperaturas supra-ótimas.
Palavras-chave: Saccharomyces cerevisiae; resistência a múltiplas drogas; bombas de efluxo de drogas; ATP-binding cassette (ABC); Pdr18; ergosterol; resposta da levedura a stress; temperaturas supra-ótimas.

iv
Contents
Acknowledgments .................................................................................................................. i
Abstract .................................................................................................................................. ii
Resumo ................................................................................................................................. iii
List of Figures ........................................................................................................................ vi
List of Abbreviations ............................................................................................................ viii
1. Motivation and thesis outline .......................................................................................... 1
2. Introduction .................................................................................................................... 3
2.1. The multidrug resistance (MDR) phenomenon .........................................................3
2.1.1. Multidrug resistance transporters in yeast .........................................................5
2.1.1.1. The ATP-binding cassette (ABC) transporters ............................................5
2.1.1.2. The Pdr18 multidrug resistance ABC transporter .......................................7
2.2. Effects of acetic acid stress in Saccharomyces cerevisiae .......................................8
2.2.1. Acetic acid as a stress agent .............................................................................8
2.2.2. Mechanisms of acetic acid tolerance .................................................................9
2.3. Stress associated with growth at supraoptimal temperatures .................................12
2.3.1. Cellular mechanisms associated with thermotolerance in yeast ......................13
3. Materials and methods ................................................................................................. 15
3.1. Strains, plasmids and growth conditions .................................................................15

v
3.2. Acetic acid susceptibility .........................................................................................15
3.3. Assessment of plasma membrane potential ...........................................................16
3.3.1. [14C]-Methylamine uptake experiments ............................................................16
3.3.2. DiOC6(3) (3-3’-dihexyloxacarbocianine iodide) accumulation assay ...............16
3.4. Extracellular acidification curves promoted by yeast cells cultivated in the presence or absence of acetic acid ..................................................................................................17
4. Results ......................................................................................................................... 18
4.1. Role of the ABC transporter Pdr18 in yeast tolerance to acetic acid at temperatures in the range 30-40ºC .........................................................................................................18
4.1.1. The lack of PDR18 expression leads to increased yeast susceptibility to acetic acid .....................................................................................................................18
4.1.2. Inhibitory effect of acetic acid in yeast growth at temperatures in the range 30-40ºC ........................................................................................................................19
4.2. PDR18 expression is essential to counteract the decrease of plasma membrane potential in yeast cells cultivated with inhibitory concentrations of acetic acid ...................22
4.3. Effect of acetic acid and PDR18 expression on the active efflux of H+ ....................26
5. Discussion .................................................................................................................... 28
References .......................................................................................................................... 31
Annexes .............................................................................................................................. 41
6. Annex I ......................................................................................................................... 42
7. Annex II ........................................................................................................................ 46
8. Annex III ....................................................................................................................... 48

vi
List of Figures
Figure 2.1 - Schematic representation of MDR transporters in yeast. .....................................4
Figure 2.2 - Mechanistic model for the adaptive yeast response to weak acid-induced stress. .............................................................................................................................................10
Figure 4.1 - PDR18 expression leads to increased yeast tolerance to acetic acid. ...............18
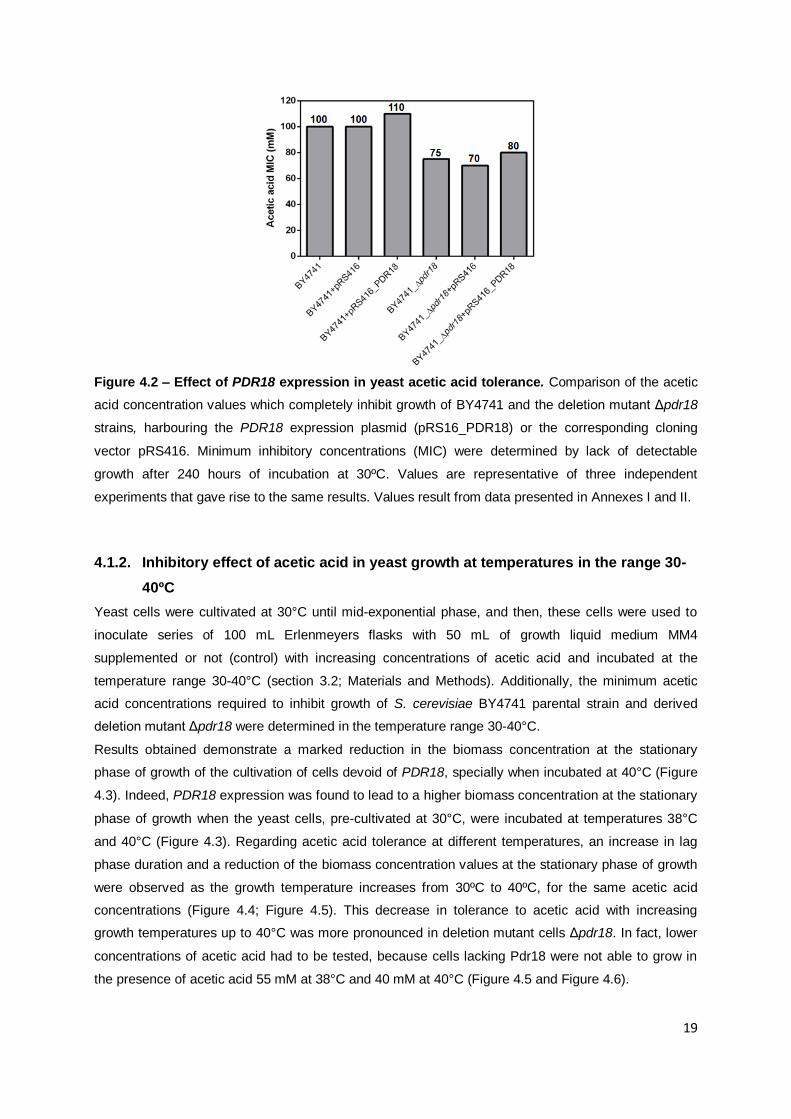
Figure 4.2 – Effect of PDR18 expression in yeast acetic acid tolerance................................19
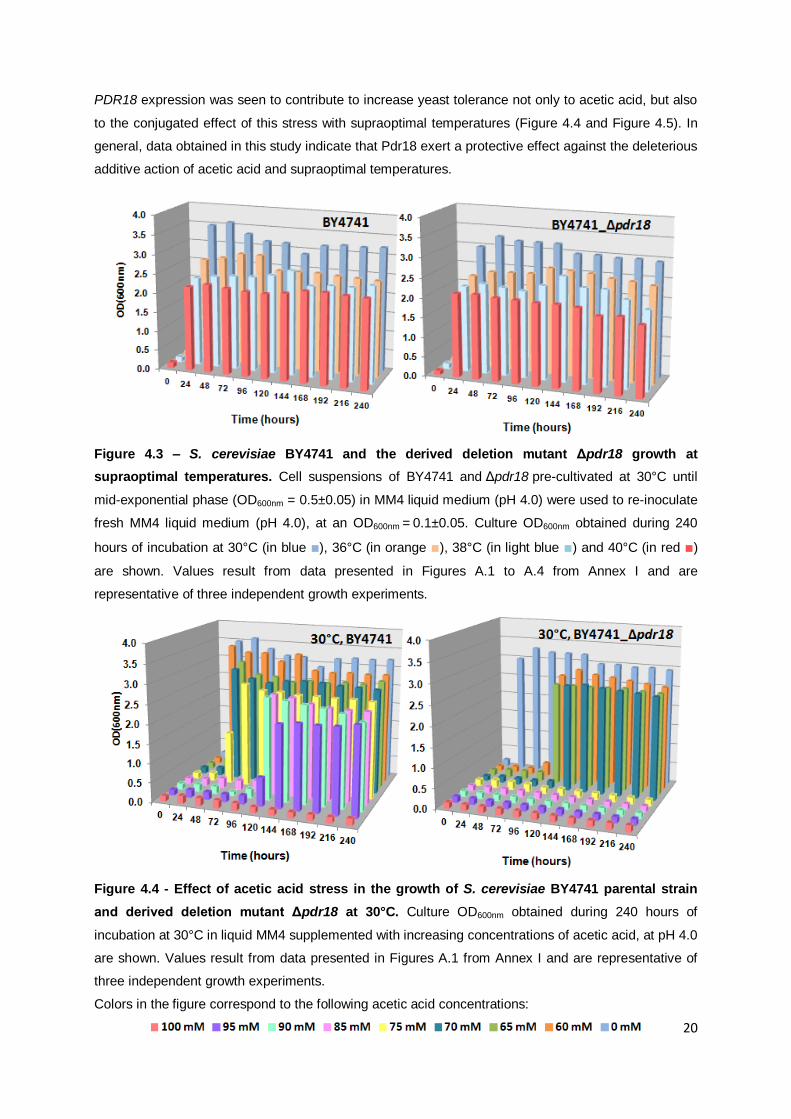
Figure 4.3 – S. cerevisiae BY4741 and the derived deletion mutant Δpdr18 growth at
supraoptimal temperatures. ..................................................................................................20
Figure 4.4 - Effect of acetic acid stress in the growth of S. cerevisiae BY4741 parental strain and derived deletion mutant Δpdr18 at 30°C. .......................................................................20
Figure 4.5 - Conjugated effect of acetic acid and supraoptimal temperatures in the growth of S. cerevisiae BY4741 parental strain and derived deletion mutant Δpdr18. ..........................21
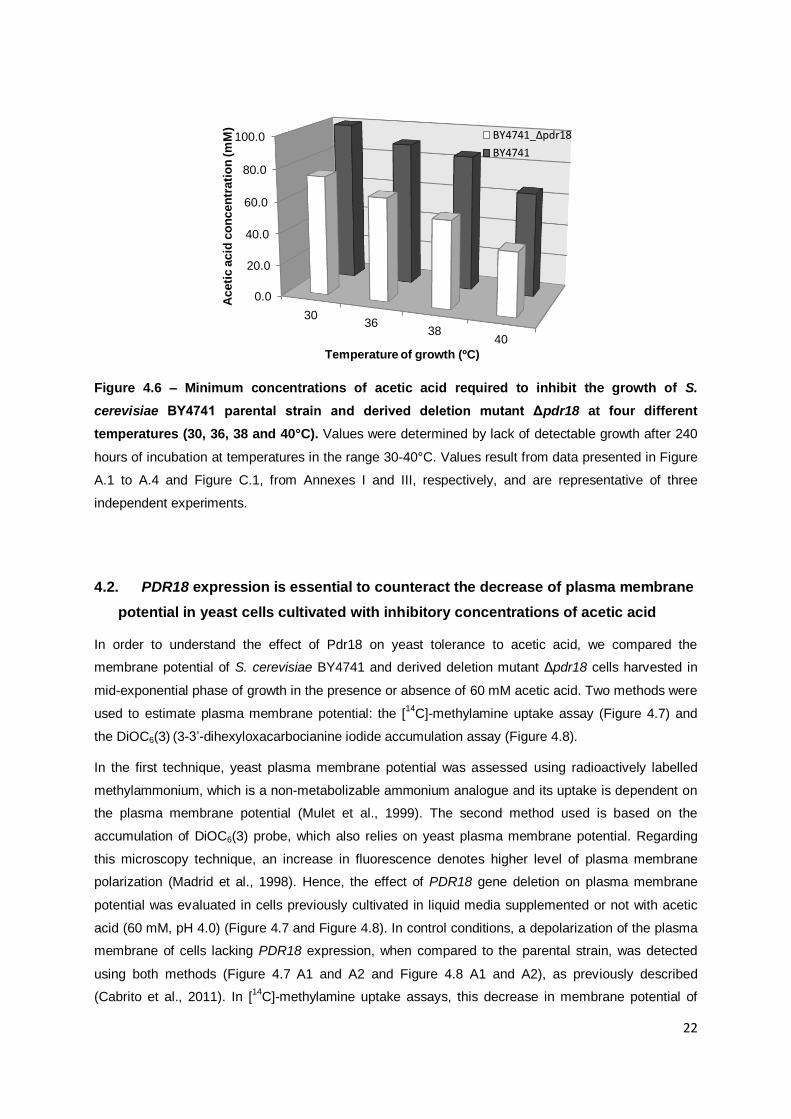
Figure 4.6 – Minimum concentrations of acetic acid required to inhibit the growth of S. cerevisiae BY4741 parental strain and derived deletion mutant Δpdr18 at four different
temperatures (30, 36, 38 and 40°C). ....................................................................................22
Figure 4.7 - PDR18 gene expression is essential to counteract the dissipation of plasma membrane potential in yeast cells cultivated in the absence or presence of inhibitory concentrations of acetic acid (60 mM, pH 4.0). .....................................................................24
Figure 4.8 – Distribution of cell population membrane potential, obtained from fluorescence microscopy using DiOC6(3) of the parental strain BY4741 (A1) and derived deletion mutant strain BY4741_Δpdr18 (A2) harvested from a mid-exponential culture growth performed in
absence (white bars) or presence (black bars) of acetic acid (60 mM)..................................25
Figure 4.9 – Effect of PDR18 gene expression in the external acidification rate in cells cultivated under control conditions or in the presence of inhibitory concentrations of acetic acid. .....................................................................................................................................27
Figure A.1 - Culture OD600nm of Saccharomyces cerevisiae parental strain BY4741 and derived deletion mutant Δpdr18 grown at 30°C in the absence and in the presence of acetic acid.. ....................................................................................................................................42

vii
Figure A.2 - Culture OD600nm of Saccharomyces cerevisiae parental strain BY4741 and derived deletion mutant Δpdr18 grown at 36°C in the absence and in the presence of acetic
acid. .....................................................................................................................................43
Figure A.3 - Culture OD600nm of Saccharomyces cerevisiae parental strain BY4741 and derived deletion mutant Δpdr18 grown at 38°C in the absence and in the presence of acetic
acid. .....................................................................................................................................44
Figure A.4 - Culture OD600nm of Saccharomyces cerevisiae parental strain BY4741 and derived deletion mutant Δpdr18 grown at 40°C in the absence and in the presence of acetic
acid. .....................................................................................................................................45
Figure B.1 - Culture OD600nm of Saccharomyces cerevisiae parental strain BY4741 harbouring a PDR18 expression plasmid or the corresponding cloning vector pRS416 in the
absence and in the presence of acetic acid. .........................................................................46
Figure B.2 - Culture OD600nm of Saccharomyces cerevisiae derived deletion mutant Δpdr18 harbouring a PDR18 expression plasmid or the corresponding cloning vector pRS416 in the absence and in the presence of acetic acid. .........................................................................47
Figure C.1 - Conjugated effect of acetic acid and temperature in the growth of Saccharomyces cerevisiae parental strain BY4741 and derived deletion mutant Δpdr18. ....48

viii
List of Abbreviations
2,4-D 2,4-dichlorophenoxyacetic acid
ABC ATP Binding Cassette
ATP Adenosine triphosphate
EUROSCARF European Saccharomyces cerevisiae archive for functional analysis
HMF Hydroxymethyl furfural
HSR Heat shock response
MDR Multidrug resistance
MFS Major Facilitator Superfamily
MM Minimal growth medium
NBD Nucleotide Binding Domain
OD600nm Optical density at 600nm
PDR Pleiotropic Drug Resistance
PDRE Pdr-responsive elements
P-gp Mammalian Glicoprotein
pHext Extracellular pH
pHi Intracellular pH
pKa -log(Ka), where Ka is the acid dissociation constant
Rpm Rotations per minute
S. cerevisiae Saccharomyces cerevisiae
SSF Simultaneous saccharification and fermentation
TMD Transmembrane Domain
TMS Transmembrane-spanning Segments
YRRE Yrr-responsive element

1
1. Motivation and thesis outline
Bio-ethanol produced by yeast cells from renewable substrates is an ecological and renewable liquid
fuel whose production has increased during the last years and is expected to continue to grow, as an
alternative to fossil fuels (Hahn-Hagerdal et al., 2006). Lignocellulosic biomass is widely available,
relatively inexpensive, and can be non-competitive with food products, leading to a cost-effective,
reproducible and sustainable large-scale production of bio-ethanol (Greetham et al., 2014, Pereira et
al., 2011). In the preparation of lignocellulosic hydrolysates for bio-ethanol production, a pretreatment
step is needed to release all the sugar monomers from the lignocellulosic biomass and make them
available for fermentation (Laluce et al., 2012, Wimalasena et al., 2014), which leads to the
appearance of sugar degradation products, such as weak acids, furan derivatives and phenolic
compounds (Galbe and Zacchi, 2002). These products are inhibitors that affect both cell growth and
ethanol biosynthesis (Lu et al., 2012, Palmqvist et al., 1999). Acetic acid is the most common of the
many weak organic acids that occur in lignocellulose hydrolysates (Ingram et al., 1999), and is formed
when the acetyl groups present in lignin and linked to the hemicelluloses chains are released,
depending on the pre-treatment type and amount of hemicelluloses present in the raw material
(Ingram et al., 1999, Palmqvist et al., 1999). Furthermore, for the production of ethanol from
lignocelluloses, a process economically advantageous is the simultaneous saccharification and
fermentation (SSF) (Lu et al., 2012). This process is less affected by product inhibitors due to the use
of enzymatic hydrolysis; but has also disadvantages, due to the optimal temperature for cellulase
activity (45 - 50°C), which is higher compared to the yeast optimal temperature for fermentation
(~30°C) (Dogan et al., 2014), affecting yeast viability and growth (Eklund et al., 1997, Olofsson et al.,
2008, Pereira et al., 2011).
In order to be economically feasible, bio-ethanol production requires yeast strains with specific
desirable traits, namely tolerance to high temperatures, acetic acid and others inhibitors present in
hydrolysates of lignocellulosic materials, and the efficient production of ethanol with high production
yields (Benjaphokee et al., 2012). Multidrug resistance (MDR) is a biological phenomenon in which
organisms become less sensitive to multiple structurally and functionally unrelated cytotoxic
compounds (Balzi and Goffeau, 1994). Even though MDR represents an alarming clinical problem in
the treatment of cancer and infectious diseases (Balzi and Goffeau, 1994), the acquisition of tolerance
to multiple chemical stresses also leads to the improvement of strain robustness for industrial
processes (Sa-Correia et al., 2009). Pdr18 is a MDR plasma membrane transporter of the ATP-
binding cassette (ABC) superfamily and its physiological role in mediating ergosterol incorporation in
the yeast plasma membrane was previously proposed by Biological Sciences Research Group
(BSRG) (Cabrito et al., 2011). Pdr18 was described as a determinant of resistance to an extensive
variety of chemical stress agents, such as the herbicides 2,4-dichlorophenoxyacetic acid (2,4-D), the
agricultural fungicide mancozeb (Cabrito et al., 2011), and also to toxic concentrations of ethanol
(Teixeira et al., 2012), which represents an advantage in industrial bio-ethanol production processes.
In this thesis, we intend to scrutinize the involvement of the MDR plasma membrane transporter Pdr18

2
in yeast response to acetic acid stress and to another environmental challenge affecting plasma
membrane lipid organization: a range of supraoptimal temperatures. For this, growth experiments in
the presence of increasing concentrations of acetic acid at temperatures in the range 30-40°C were
performed. Given Pdr18 effect in the incorporation of ergosterol in the membrane, we were interested
in studying the physiological role of PDR18 expression in the yeast plasma membrane properties of
cells cultivated in the presence of inhibitory concentrations of acetic acid, such as membrane potential
and permeability. Overall, the results of this thesis are a contribution to the characterization of the
physiological role of Pdr18 as a MDR determinant.

3
2. Introduction
2.1. The multidrug resistance (MDR) phenomenon
Multidrug resistance (MDR) is a phenomenon in the study of cellular simultaneous resistance to a
number of unrelated citotoxic compounds (Balzi and Goffeau, 1994). The occurrence of MDR was
registered in a broad range of organisms, from bacteria to mammals (Sa-Correia et al., 2009), and the
main mechanism is the active transport of structurally and pharmacologically distinct drugs out of the
cell (Higgins, 2007). In medicine the emergence of this phenomenon in therapy has led to severe
difficulties in the treatment of tumors and several infectious diseases. In agriculture, multidrug
mechanisms have also negative outcomes in the control of resistance of plant pathogens to natural
plant defence toxins and common fungicides (Balzi and Goffeau, 1994). Nevertheless, the ability of
industrial strains to tolerate different chemical stresses represents an advantage arising from the MDR
phenomenon (Sa-Correia et al., 2009).
The most important MDR membrane transporters are members of the ATP-binding cassette (ABC)
superfamily or the major facilitator superfamily (MFS) (Figure 2.1) (Del Sorbo et al., 2000). The best
characterized are the ABC transporters, which are primary active transporters, as they are involved in
the active transport of solutes using ATP hydrolysis, as the energy source. In contrast, in MFS
transporters, transport is driven by the proton-motive force, being classified as secondary active
transporters (Pao et al., 1998, Del Sorbo et al., 2000).
The study of multidrug resistance in the yeast Saccharomyces cerevisiae became very important due
to the fact that some yeast species are involved in pathogenicity for men and other animals and in
plants (Balzi and Goffeau, 1994). Moreover, yeast is considered an universal, easy-to-manipulate
model system for the study of more complex eukaryotic cells (Sa-Correia and Tenreiro, 2002),
assisting in the understand of the mechanisms underlying their cytotoxic insults (Sa-Correia et al.,
2009).

4
Figure 2.1 - Schematic representation of MDR transporters in yeast. The simplified topologies
are as predicted by the Saccharomyces Genome Database (SGD) of the (a) multidrug
transporters of the ABC superfamily and (b) the MFS-MDR transporters from the 12-spanner DHA1
(light blue) and 14-spanner DHA2 (dark blue) families (From Sá-Correia et al., 2009).

5
2.1.1. Multidrug resistance transporters in yeast
2.1.1.1. The ATP-binding cassette (ABC) transporters
ATP-binding cassette (ABC) proteins represent one of the largest membrane transporter superfamily
(Jungwirth and Kuchler, 2006). ABC transporters structure is divided in four core domains: two
transmembrane domains (TMD) and two nucleotide binding domains (NBD). The nucleotide binding
domains are located in the cytosol and contain the ATP-binding cassette (Dean et al., 2001), as well
as a variable number of predicted transmembrane-spanning segments (TMS) (Higgins, 2001), which
determine different substrate specificities of individual ABC proteins (Jungwirth and Kuchler, 2006).
They are conserved in all living organisms and use ATP as a source of energy to translocate an
extensive variety of substrates across the plasma membrane (Del Sorbo et al., 2000). One of the roles
of these transporters is as importers, transporting nutrients and ions into the cell (Higgins, 1992).
Others are involved in the efflux of a wide variety of xenobiotic compounds, such as the human P-
glycoprotein (P-gp), which functions as an ATP-dependent efflux pump for drugs (Juranka et al., 1989,
Nourani et al., 1997) and is responsible for multidrug resistance in cancer cells (Balzi and Goffeau,
1994, Juranka et al., 1989). Nevertheless, besides ABC functions as transporters, they also might be
involved in the regulation of heterologous channels and other membrane proteins (Higgins, 1995).
In yeast, many protein transporters belonging to the ABC superfamily are localized at the plasma
membrane or of important organelles, such as Golgi, mitochondria, endoplasmic reticulum, etc, and
one considered to have a wide range of functions, like drug transport, ion homeostasis, heavy metal
detoxification and sterol homeostasis (Prasad et al., 2002). The multidrug-resistance phenotype in
yeast is also referred as pleiotropic drug resistance (PDR) (Balzi and Goffeau, 1994, Balzi and
Goffeau, 1995, Nourani et al., 1997) and frequently results from the overexpression of plasma
membrane pumps from the ABC superfamily (Wendler et al., 1997). They represent the first line of
defense and assist in yeast detoxification, essential for the drug resistance observed in the multidrug-
tolerant cells (Balzi and Goffeau, 1994, Del Sorbo et al., 2000).
MDR regulation entails a critical regulatory network of transcription factors, including the homologous
Zn(II)2Cys6 zinc cluster transcription factors Pdr1 and Pdr3, defined as the major regulators of the PDR
network (Balzi and Goffeau, 1995), and other transcription regulators, such as Pdr7, Pdr9, Yap1 and
Yrr1 (Balzi and Goffeau, 1995, Wendler et al., 1997, Zhang et al., 2001, Dexter et al., 1994). Pdr1 and
Pdr3 have a common sequence identity and drug resistance effects (Katzmann et al., 1994). They
form homo- and heterodimers, which bind to the same DNA element designated as Pdr1/Pdr3
response elements (PDRE) (Mamnun et al., 2002), and regulate the expression of SNQ2, YOR1,
PDR5 (ABC transporters genes) (Nourani et al., 1997) and YRR1 (Zhang et al., 2001). According to
Zhang et al., Yrr1 binds to Yrr1 responsive elements (YRRE), an element different than PDRE, leading
to an increased activation of Pdr1 and Pdr3, which suggests interactions between Pdr1, Pdr3, and
Yrr1 activities.

6
The main conceivable physiological function of the PDR network appeared to be detoxification, since
PDR transporters apparently export hundreds of structurally and functionally unrelated cytotoxic
compounds, and potentially toxic metabolites (Kolaczkowska and Goffeau, 1999, Jungwirth and
Kuchler, 2006). Consequently, this hypothesize active transport may result in hypertolerance to
several unrelated exogenous drugs or xenobiotics and cell protection from unwanted side effects of
endogenous toxic metabolites (Jungwirth and Kuchler, 2006, Gbelska et al., 2006).
Moreover, some MDR transporters of the ABC superfamily, regulated by the PDR network
transcription factors Pdr1/3, have been related with lipid bilayer homeostasis, lipid uptake and control
of membrane permeability (Jungwirth and Kuchler, 2006). These include the ABC transporters Pdr5,
Pdr10 and Pdr15 which may control the membrane lipid bilayer distribution in the plasma membrane
(Schuller et al., 2007, Rockwell et al., 2009). Aus1 and Pdr11 were also found to increase the
availability of sterol in the plasma membrane for esterification in the endoplasmic reticulum, thus,
contributing to the intracellular sterol transport (Li and Prinz, 2004). Also, Yor1 together with Pdr5 may
be involved in the transport of phospholipids through the plasma membrane (Decottignies et al.,
1998). All these facts indicate that PDR pathway has a role in phospholipid and sterol composition,
controlling in part the distribution of lipid components in inner and outer leaflets of the plasma
membrane (Gulshan and Moye-Rowley, 2007). Furthermore, in S. cerevisiae plasma membrane, high
levels of ergosterol are present (Eisenkolb et al., 2002, Gulshan and Moye-Rowley, 2007). The
association of sphingolipids with ergosterol is crucial to form lipid rafts, a membrane microdomain with
important functions in the stable delivery of specific proteins, such as Pma1, to the plasma membrane
and signal transduction (Bagnat et al., 2000, Dickson and Lester, 2002). These high levels of
ergosterol and sphingolipids may be intrinsically involved in the protection of the cell against
environmental stress conditions, because of their role in the reduction of the membrane permeability
(Eisenkolb et al., 2002). This membrane lipids biosynthesis may also be directly correlated with the
PDR pathway. Therefore, the PDR pathway is required not only for the regulation of the membrane
transporters activity, but also for the lipid composition of cell membranes, which exerts an important
role in the activity of membrane embedded proteins (Gulshan and Moye-Rowley, 2007).

7
2.1.1.2. The Pdr18 multidrug resistance ABC transporter
Pdr18 is a protein plasma membrane transporter of the ABC superfamily found to be related with
plasma membrane sterol incorporation (Cabrito et al., 2011). Its physiological role in plasma
membrane sterol composition and membrane potential may affect transport across cell membrane and
drug partition between the cell interior and the extracellular medium, which was proposed to contribute
to the multidrug resistance phenotype related with its increased expression (Cabrito et al., 2011).
Pdr18 was described to determine yeast resistance to chemical stress agents, including the herbicides
2,4-dichlorophenoxyacetic acid (2,4-D) and barban, the agricultural fungicide mancozeb and cadmium,
copper, manganese and zinc (Cabrito et al., 2011). Also, PDR18 expression displays a role in
decreasing ethanol-induced plasma membrane permeabilization and reducing intracellular ethanol
concentration, which results in increased yeast tolerance to toxic concentrations of ethanol (Teixeira et
al., 2012). The effect of Pdr18 in the incorporation of ergosterol in the yeast plasma membrane
(Cabrito et al., 2011) can be explored in the construction of more robust strains for improvement of
high gravity alcoholic fermentations.

8
2.2. Effects of acetic acid stress in Saccharomyces cerevisiae
2.2.1. Acetic acid as a stress agent
Acetic acid is a by-product of the alcoholic fermentation carried out by S. cerevisiae. S. cerevisiae may
use acetic acid as a carbon source by the activity of the anapleurotic glyoxylate cycle and
gluconeogenesis, which pathways are regulated by glucose repression (Gancedo, 1998). In the
presence of glucose there is a repression of these metabolic pathways, leading to the accumulation of
acetic acid in the growth medium (Casal et al., 1996). During alcoholic fermentation by yeast, acetic
acid concentration increases, and the concentrations of ethanol attained and other toxic metabolites
produced result in growth and fermentation rate inhibition (Mira et al., 2010b, Graves et al., 2006).
Acetic acid is also one of the inhibitors of the microbial fermentation of lignocellulosic hydrolysates,
non-feedstock substrates considered important alternative for a sustainable production of bioethanol
(Lu et al., 2012).
In addition, organic acids, such as acetic acid, are weak acids used in food industry as preservatives
against microbial spoilage (Fernandes et al., 2005, Ullah et al., 2012). Although these food
preservatives inhibit most of yeast and of mould species, a number of these microorganisms have
developed the ability to grow at low pH in the presence of high acid concentrations leading to food
spoilage (Stratford et al., 2013). The antimicrobial effect of carboxylic acids is mostly determined by
their chemical properties, in particular, hydrophobicity, volatility, and pKa. Acetic acid toxicity is strongly
dependent on the pH of the medium, as it is a weak monocarboxylic acid with a pKa of 4.76 (Mira et
al., 2010c) and at neutral pH, acid acetic as well as other carboxylic acids are completely dissociated.
Thus, the antimicrobial potential of acetic acid is mainly at low pH values (below pKa), where the
protonated form predominates, since in its undissociated form, acetic acid has biophysical properties
that enable it to passively diffuse into the cell through the lipid bilayer (Piper et al., 2001). The
undissociated acid, being uncharged, readily diffuses across the plasma membrane lipid bilayer and
dissociates in the higher pH environment close to neutrality of the cytosol, resulting in the
accumulation of protons and acetate counter-ion (CH3COO-) in the interior (Piper et al., 2001). This
negatively charged form will accumulate intracellularly to very high levels due to its inability to diffuse
out of the cell. Consequently, intracellular pH decreases, affecting internal pH homeostasis and many
cellular processes such as glycolytic enzymes (Pampulha and Loureiro-Dias, 1990), acetic-acid-
mediated inhibition of NADH dehydrogenase (Zhao et al., 2008) and other metabolic processes (Krebs
et al., 1983, Piper et al., 2001). In addition, the intracellular acidification may also lead to the
generation of high turgor pressure, as well as the production of free radical that will induce severe
oxidative stress (Piper et al., 2001).

9
2.2.2. Mechanisms of acetic acid tolerance
Several mechanisms of response to acetic acid in S. cerevisiae are part of the global yeast response
to weak acids in general. After the addition of a weak acid to the culture medium, yeast cells usually
exit the cell cycle and enter a long period of stasis. During a period of several hours, which depends
on the weak acid concentration and the level of growth inhibition, a range of molecular responses are
activated and functional changes take place in yeast (Mira et al., 2010c). This stress response acts in
order to counteract weak acid effects, reducing the accumulation of high, potentially toxic levels of
acids within yeast cells (Piper et al., 2001). Therefore, in order to avoid the dissipation of plasma
membrane potential induced by lipophilic weak acids, and maintain the internal pH within physiological
values (Mira et al., 2010c), protons released are pumped out of the cell by the activation of the H+-
ATPase Pma1 protein (PM-H+-ATPase) by acetic acid (Figure 2.2) (Carmelo et al., 1996) and other
weak acids (Viegas and Sa-Correia, 1991, Holyoak et al., 1996). Consistent with the importance of
this cell response, a lowered expression of the gene encoding the major H+-ATPase (PMA1) leads to
the increase of the sensitivity of cells to weak acids (Holyoak et al., 1996), confirming its role in weak
acid adaptation. In addition, PMA2 gene is homologous to the PMA1 gene and also encodes a H+-
ATPase identical to the one encoded by PMA1 gene. However, while PMA1 gene is highly expressed,
PMA2 gene is expressed at a much lower level and is considered not essential (Fernandes and Sa-
Correia, 2001).
Furthermore, the H+-ATPase in the vacuolar membrane (V-ATPase) is also necessary for intracellular
pH homeostasis under weak acid stress, also revealing an increased activity in response to numerous
weak acids (Serrano et al., 1986, Fernandes et al., 2003, Makrantoni et al., 2007, Carmelo et al.,
1996). This V-ATPase pumps protons into the lumen of the vacuole, contributing to the recovery of
more physiological cytosolic pH and counteracting the acid-induced dissipation of the transmembrane
potential across the vacuolar membrane (Desmoucelles et al., 2002, Fernandes et al., 2003,
Makrantoni et al., 2007, Mira et al., 2009). Nevertheless, the active expulsion of weak acid anions from
the cell interior is energetically expensive (Mira et al., 2010c) and for yeast adaptation to weak acids,
specifically to acetic acid, the restriction of the passive diffusion and re-entrance of the undissociated
form of this weak acid is essential. One of the mechanisms proposed to reduce the diffusion rate of
weak acids is the reduction in porosity of the cell wall structure and the reconfiguration of membrane
lipid composition (Figure 2.2), which leads to a decrease of the internal concentration of weak acids
(Mira et al., 2009).

10
Figure 2.2 - Mechanistic model for the adaptive yeast response to weak acid-induced stress.
Representation of the activity of H+-ATPases in the plasma and vacuolar membranes for the
intracellular pH (pHi) recovery and reconfiguration of the cellular envelope (From Mira et al. 2010c).
In addition, the plasma membrane ABC transporter Pdr12, which is related to multidrug resistance
(MDR), has also a role in the reduction of the intracellular pool of the weak acid counter-ions in S.
cerevisiae (Piper et al., 1998, Holyoak et al., 1996). PDR12 is strongly induced by weak acid
preservatives, such as sorbate, benzoate, and certain other moderately lipophilic carboxylate
compounds, but not by high levels of acetate and organic alcohols (Hatzixanthis et al., 2003). Its
induction is essential for the development of weak acid tolerance and growth in the presence of food
preservatives, as it functions as an efflux pump, exporting the organic acid anion from the cell (Piper et
al., 1998, Piper et al., 2001).
Another response to acetic acid in yeast is the activation of the TOR (Target-of-Rapamycin) pathway
(Almeida et al., 2009), a regulatory system of response to nutrient starvation, such as the reduced
concentration of amino acids in weak acid challenged cells. This mechanism was proposed to
counteract nutrient limitation conditions induced by weak acid stress (Teixeira et al., 2005, Almeida et
al., 2009). Weak acid-stressed yeast cells show also modifications on their carbohydrate metabolism,
by the up-regulation of several genes encoding enzymes of the Krebs cycle and of glycolysis in
response to acetic acid (Mira et al., 2010a), such as phosphofructokinase (Pfk2) and fructose 1,6-
bisphosphate aldolase (Fba1) (Almeida et al., 2009). This response is related with the severe

11
depletion of ATP observed in these yeast cells and may be caused by the inhibition of the activity of
glycolytic enzymes (Krebs et al., 1983, Holyoak et al., 1996). In addition, several genes involved in
ATP synthesis were identified as determinants of resistance to multiple weak acids, including acetic
acid (Kawahata et al., 2006, Mira et al., 2010a). This is related with the demonstrated activation of
energy-consuming defense mechanisms, such as PM-H+-ATPase, V-H
+-ATPase and drug efflux
pumps (Mira et al., 2010c).

12
2.3. Stress associated with growth at supraoptimal temperatures
Yeasts growing at supraoptimal temperatures are exposed to heat stress, which has negative effects
at the level of several cellular processes, such as the inhibition of cell division, problems in protein
homeostasis because of protein aggregation, and membrane lipids disorganization (Verghese et al.,
2012). Several studies have demonstrated a great influence of temperature on the viability of yeast
(van Uden and Madeira-Lopes, 1970). Usually, there is a reduction in the growth rate of yeast cell
population within the superoptimal temperature range, the temperatures between the optimum and the
maximum temperature for growth (van Uden and Madeira-Lopes, 1970, Van Uden and Madeira-
Lopes, 1976). S. cerevisiae is a mesophilic yeast, and when growing within its superoptimal
temperature range (from about 36°C to 44°C) (van Uden, 1984), simultaneous growth and death
occur, displaying an associative profile (van Uden and Madeira-Lopes, 1970). Nevertheless, a
dissociative profile in other yeasts was also proposed, in which temperature range of exponential
growth was separated from the temperature range of thermal death (Madeira-Lopes and van Uden,
1979). Moreover, the specific growth and death rates depend on the temperature (Simoes-Mendes et
al., 1978) and, the presence of antimicrobial agents affect negatively the temperature profiles of
growth. Regarding acetic acid, Ramos and Madeira-Lopes (1990) demonstrated that acetic acid has
negative effects in the temperature profile of growth with ethanol of S. cerevisiae. In the absence of
ethanol, increasing concentrations of acetic acid narrowed the temperature range of growth, by the
progressive increase of the minimum temperature and the reduction the maximum temperature. Acetic
acid also enhanced yeast thermal death, as well as decreased tolerance to ethanol, since the
temperature range in the presence of ethanol became shorten. Nonetheless, acetic acid did not alter
S. cerevisiae associative growth profile (Ramos and Madeira-Lopes, 1990).
Membranes are affected by lipophilic compounds and by high temperatures, resulting in the damage
of membrane proteins and increase of membrane non-specific permeability and modifications in
fluidity, due to changes in the lipid organization (Salgueiro et al., 1988, Dogan et al., 2014).
Temperature effect in plasma membrane affects yeast growth and cell viability, by decreasing
plasma membrane H+-ATPase activity (Piper, 1995) and others transport systems involved in nutrient
transport (Viegas et al., 1995, Salgueiro et al., 1988), as well as by changing the fatty acid and sterol
composition of cell membranes (Beltran et al., 2008). Heat stress has also consequences in
mitochondrial electron transport chain in S. cerevisiae, leading to oxidative stress (Morano et al.,
2012). In addition, membrane fluidity is also strongly influenced by the ratio, composition, and
structure of sterols that are found in membranes (Caspeta et al., 2014) and changes in fluidity may
also have as consequences, alterations in ion transport (Verghese et al., 2012). Indeed, (Cyert, 2003)
demonstrated that cells lacking calcineurin activity are highly stress sensitive, which might be related
with the fact that the Ca2+
regulated protein phosphatase calcineurin function is the upregulation of
genes involved in several mechanisms, in response to stress, such as the biosynthesis of membrane
lipids and ergosterol (Verghese et al., 2012).

13
Despite the detrimental effects associated with supraoptimal temperature in yeasts, fermentation
temperatures higher than 35°C have several advantages, namely the reduction in the fermentation
costs because of the reducing cooling costs between pretreatment and fermentation, the reduction of
the risk of contamination input and lower energy required for ethanol separation (easy recovery of
ethanol) (Rajoka et al., 2005). Specifically in SSF, temperature for enzymatic hydrolysis in
saccharification (45-50°C) is usually higher than the optimal temperature of fermentation processes
(around 30°C) (Eklund, 1995, Olofsson et al., 2008, Dogan et al., 2014), which represents a
disadvantage of this process. Nevertheless, the use of yeast strains with higher thermotolerance,
capable of growth and of remaining active during fermentation at higher temperatures, similar to
optimal cellulase and hemicellulase activities (Lu et al., 2012, Pereira et al., 2011), could be an
approach to increase ethanol yield. Therefore, thermotolerance is undoubtedly an important issue for
these fermentation processes.
2.3.1. Cellular mechanisms associated with thermotolerance in yeast
At temperatures above 36-37°C, yeast cells activate a protective transcriptional program, the heat
shock response (HSR). Under these conditions, heat shock proteins (HSP) are expressed resulting in
the modification of other components of cells physiology, including membrane composition and
carbohydrate flux (Morano et al., 2012), in order to increase yeast thermotolerance (Yamamoto et al.,
2008). Several Hsp proteins work as molecular chaperones in the synthesis, folding, trafficking,
maturation and degradation of proteins (Mayer and Bukau, 1998, Bukau and Horwich, 1998).
In the HSR, the transcription of HSP genes with protective functions in heat-challenged yeast cells is
regulated by Hsf1, the primary regulator of HSR that is important for the recovery from a brief
exposure to extreme temperature, and the transcription factors Msn2 and Msn4, essential for long-
term survival at high temperatures (Morano et al., 2012, Yamamoto et al., 2008). Hsf1, Msn2 and
Msn4 are positive regulators of Hsp104, an antistress chaperone with a biological role in cellular
protein homeostasis (Yamamoto et al., 2008). This chaperone is essential for thermotolerance and cell
survival, since the expression of Hsp104, as well as other Hsp is required for the refolding of heat-
denatured proteins in cells recovering from severe heat shock (Parsell et al., 1994, Yamamoto et al.,
2008). Hence, thermotolerance requires an accurate equilibrium in the amounts and activities of
several cellular components before and after severe heat shock (Yamamoto et al., 2008), which is
achieved by changes in gene expression co-regulated by Hsf1, Msn2 and Msn4 (Morano et al., 2012).
Although heat shock proteins have received the major attention in the subject of thermotolerance,
there are alternative pathways to promote cell survival under temperature stress that are independent
of heat-shock protein synthesis (De Virgilio et al., 1994). Disaccharide trehalose capacities to stabilize
proteins and repress aggregation of misfolded proteins provide evidences for its role in
thermotolerance, for the maintenance of the structural integrity of cell membrane (De Virgilio et al.,

14
1994, Verghese et al., 2012). In addition, since Hsp104 chaperone role is similar to trehalose in high
temperature, both Hsp104 and trehalose may have complementary but not overlapping functions in
thermotolerance (Elliott et al., 1996).
Membrane responses to changes in temperature are also strictly dependent on the degree of
unsaturation of fatty acids of the membrane lipids (Carratu et al., 1996). Indeed, membrane fatty-acyl
composition in the yeast membrane changes with temperature: lower temperatures result in a more
unsaturated fatty-acyl composition of the membrane (Beltran et al., 2008). Moreover, sphingolipids
identified as long-chain bases (LCBs) are lipid mediators and manage intracellular protein aggregation
and translation in heat response to protect cells (Verghese et al., 2012). In fact, a strain lacking the
capacity to synthesize LCBs is heat shock sensitive, and this phenotype may be reversed by genetic
or chemical supplementation with sphingolipids, confirming these molecules role in thermotolerance
(Jenkins et al., 1997, Verghese et al., 2012).
For the maintenance of membrane fluidity and functionality, which are negatively affected at
supraoptimal temperatures (Verghese et al., 2012), yeast cells undergo changes in membrane lipid
composition (Suutari et al., 1990). Ergosterol is the most abundant sterol in the yeast plasma
membrane (Zinser et al., 1993) and studies on sterol auxotrophs demonstrated that ergosterol exerts
an effect on the biosynthesis of the different phospholipid species (Parks and Casey, 1995). Indeed,
ergosterol strongly determines the dynamic characteristics of the lipids and proteins in the yeast
membranes (Beney and Gervais, 2001). In addition, the degree of lipid saturation and the presence of
ergosterol as modulators of membrane fluidity in the S. cerevisiae plasma membrane were proposed
as a determinant factor to stress tolerance, independently of heat shock proteins and trehalose (Swan
and Watson, 1998). Thus, cells survival is strongly related with the role of ergosterol as a stabilizer of
yeast plasma membrane (Swan and Watson, 1998), affecting membrane rigidity, fluidity, and
permeability (Parks and Casey, 1995).

15
3. Materials and methods
3.1. Strains, plasmids and growth conditions
Saccharomyces cerevisiae parental strain BY4741 (MATa, his3Δ1, leu2Δ0, met15Δ0, ura3Δ0) and the
derived deletion mutant strain BY4741_Δpdr18 used in this work were obtained from the
EUROSCARF collection. The plasmid pRS416_PDR18, expressing the PDR18 gene from its natural
promoter, and the corresponding cloning vector, pRS416, were both obtained from the EUROSCARF
collection and were used for phenotypic complementation tests.
Cells were batch-cultured at 30°C with orbital agitation (250 rpm) in liquid minimal growth medium
(MM4) containing 1.7 g/L yeast nitrogen base without amino acids (Difco), 20 g/L glucose (Merck),
2.65 g/L (NH4)2SO4 (Panreac AppliChem), 20 mg/l L-methionine (Merck), 20 mg/L L-histidine (Merck),
60 mg/L L-leucine (Sigma) and 20 mg/L L-uracil (Sigma), adjusted to pH 4.0 with HCl. Cells
harbouring pRS416 or derived plasmids were grown in the same medium but without uracil (MM4-U
medium) and were preserved in MM4-U. Yeast cells were supplemented with 30% glycerol, and
stored at −80°C.
3.2. Acetic acid susceptibility
The susceptibility of the parental strain BY4741 and the derived deletion mutant Δpdr18 to acetic acid
was assessed as follows. Yeast cells were cultivated until mid-exponential phase (OD600nm=0.5±0.05)
in liquid medium MM4 (BY4741 and Δpdr18), adjusted to pH 4.0, at 30°C, and then re-inoculated by
filtration (Membrane filters white, 0.2 μm, WhatmanTM
, ME24/21ST) at an OD600nm of 0.1±0.05, in
100 mL Erlenmeyer flasks containing 50 mL of fresh medium, either or not supplemented with 60 mM
of acetic acid. Growth was followed by measuring culture OD600nm during batch cultivation at 30ºC with
an orbital agitation of 250 rpm.
In order to study the simultaneous effect of acetic acid and supraoptimal temperatures in yeast cell
growth, cell suspensions cultivated until mid-exponential phase (OD600nm=0.5±0.05) in liquid medium
MM4 (BY4741 and Δpdr18) or MM4-U (BY4741 and Δpdr18 harbouring the plasmid pRS416_PDR18
and the corresponding cloning vector, pRS416), adjusted to pH 4.0, at 30°C, were used to re-inoculate
by filtration (Membrane filters white, 0.2 μm, WhatmanTM
, ME24/21ST) at an OD600nm of 0.1±0.05
series of 100 mL Erlenmeyer flasks prepared with 50 mL liquid MM4 or MM4-U supplemented or not
(control) with progressively higher concentration of acetic acid (35-120 mM). Erlenmeyer flasks were
incubated with orbital agitation (250 rpm) at four different temperatures: 30, 36, 38 and 40°C, and
growth was followed by measuring culture OD600nm. In this work, minimum inhibitory concentration
(MIC) of acetic acid value was also assessed. MIC value was defined as the lowest concentration of
acetic acid at which no growth was detected after 10 days of incubation.

16
3.3. Assessment of plasma membrane potential
To compare the plasma membrane potential of the parental strain BY4741 and the derived deletion
mutant Δpdr18, two complementary methods were used: the [14
C]-Methylamine uptake assay
(Hoeberichts et al., 2010, Teixeira et al., 2011) and the DiOC6(3) (3-3’-dihexyloxacarbocianine iodide)
accumulation assay (Madrid et al., 1998).
3.3.1. [14C]-Methylamine uptake experiments
The uptake of [14
C]-methylamine in the parental strain BY4741 and the mutant strain Δpdr18 was
estimated as described previously (Hoeberichts et al., 2010, Teixeira et al., 2011). The parental and
mutant strains were grown in liquid MM4 medium (pH 4.0) until an OD600nm=0.5±0.1, and then re-
inoculated at an OD600nm of 0.1±0.05 into fresh liquid MM4 medium (pH 4.0) in the absence or
presence of acetic acid (60 mM). Cells grown until mid-exponential phase were harvested by
centrifugation (8600g, 5 minutes, at 4°C) and washed three times with deionized water and
resuspended in deionized water. Cell suspension (0.1 ml) was added to 0.9 ml of MES glucose buffer
[10 mM MES, 0.1 mM MgCl2 and 20 g/L glucose (pH 4.0)]. After 5 minutes of incubation at 30°C, 3 µl
[14
C]-methylamine (Biotrend) was added to a final concentration of 55 mM. At the indicated times (0, 5,
10, 15 and 20 minutes), samples (200 μl) were diluted in 10 ml cold water to stop the transport
reaction, filtered through pre-wetted glass microfiber filters (25 mm; Filter-Lab, MFV3) and washed
four times with 5 ml cold water. Subsequently, filters were put into tubes containing 7 mL scintillation
liquid (Ultima Gold™, Beckman) and the radioactivity was measured in a Beckman LS 5000TD
scintillation counter. Non-specific [14
C]-methylamine adsorption to the filters and to the cells (less than
5 % of the total bound radioactivity) was assessed and taken into consideration. Relative levels of
[14
C]-methylamine are the average of at least three independent experiments and the error bars
correspond to standard deviation. In order to confirm the significance of the differences between cell
populations after 20 minutes of incubation in MES glucose (30°C), we performed a one-way analysis
of variance for the repeated independent measures. All the comparisons between BY4741 and Δpdr18
under different conditions were done using an unpaired t test. Statistical significance was established
at P < 0.05.
3.3.2. DiOC6(3) (3-3’-dihexyloxacarbocianine iodide) accumulation assay
Plasma membrane potential was estimated using fluorescence microscopy. In order to perform the
DiOC6(3) accumulation assay, cells were harvested as described above (Section 3.3.1) and
resuspended in MES/glucose buffer [10 mM MES, 0.1 mM MgCl2 and 20 g/L glucose (pH 4.0)]. Cell
suspensions were supplemented with DiOC6(3) (Molecular Probes) at a final concentration of 0.25 nM
and incubated in the dark for 30 minutes at 30°C with orbital agitation (250 rpm). After centrifugation
(8600g during 5 minutes, at 4°C), cells were washed three times and immediately observed with a
Zeiss Axioplan microscope equipped with adequate epifluorescence filters (Zeiss BP450-490 and

17
Zeiss LP520). Fluorescence emission was collected using a CCD (charge-coupled device) camera
(Cool SNAPFX, Roper Scientific Photometrics) and MetaMorph 3.5 was used for image analysis.
Fluorescence images were background-corrected and the intensity values emitted by each individual
cell were measured pixel-by-pixel in a minimum of 80 cells per experiment. Fluorescence intensity
levels given by the software were expressed as a percentage of the value obtained for BY4741 cells in
control conditions. Only living cells were analyzed and dead cells were differentiated by bright-field
analysis of cell morphology. Levels of fluorescence are means for at least three independent
experiments and error bars represent standard deviation. To confirm the significance of the
differences between cell populations we performed a one-way analysis of variance for repeated
independent measures, as explained above (section 3.3.1).
3.4. Extracellular acidification curves promoted by yeast cells cultivated in the
presence or absence of acetic acid
The parental strain BY4741 and the derived deletion mutant Δpdr18 were grown in liquid MM4
medium (pH 4.0) until an OD600nm=0.5±0.05, and then re-inoculated at an OD600nm of 0.1±0.05 into
fresh liquid MM4 medium (pH 4.0) in the absence or presence of acetic acid (60 mM). BY4741 and
Δpdr18 strains grown until mid-exponential phase were collected by filtration (Membrane filters white,
0.2 μm, WhatmanTM
, ME24/21ST), washed twice with deionized water, and incubated in a sorbitol
solution (20 g/L, Sigma-Aldrich®) for 30 minutes at 30°C, with orbital agitation of 250 rpm. After this
incubation, deenergized cells were then harvested by filtration and resuspended in distilled water, in
order to obtain a dense cell suspension (OD600nm of 20.0 ± 2.0). The acidification assays were carried
out in a water-jacketed cell of 10 mL capacity, at 30°C, containing 5 mL of the cellular suspension
described above. The pH of the resulting suspension was adjusted to 4.0±0.1, and 1 ml of 100 g/L
glucose (pH 4.0) was added (to obtain a final concentration of 20 g/L). The external medium pH was
followed during 10 minutes by potentiometry using a pH microelectrode (Metrohm 6.0204.000)
attached to a pH meter (Metrohm 605). Comparisons of the pHextracellular variation between conditions
tested were perfomed using a one-way analysis of variance for repeated independent measures.
Experiments were done in triplicate. The comparisons between BY4741 and Δpdr18 cells under
different conditions were done using an unpaired t test and the statistical significance was established
at P < 0.05.

18
4. Results
4.1. Role of the ABC transporter Pdr18 in yeast tolerance to acetic acid at
temperatures in the range 30-40ºC
4.1.1. The lack of PDR18 expression leads to increased yeast susceptibility to acetic
acid
The deletion mutant Δpdr18 strain was found to be less tolerant to acetic acid stress (60 mM, pH 4.0),
compared to the parental strain BY4741. In fact, an extended lag-phase (approximately 40 hours,
compared to 10 hours for the parental strain) was observed for Δpdr18 cells when cultivated at 30ºC in
liquid minimal medium (pH 4.0) supplemented with 60 mM acetic acid (Figure 4.1). In addition, yeast
cell population from both strains attained a reduced final biomass value when cultivated under this
stress condition, and this reduction was more significant for the Δpdr18 culture (Figure 4.1).
The higher susceptibility of the deletion mutant Δpdr18 to acetic acid was also confirmed by the
determination of the minimum inhibitory concentration (MIC) of acetic acid, which corresponds to the
concentration at which a complete inhibition of each yeast strain growth is observed (Figure 4.2).
Hence, BY4741 cells were able to grow at acetic acid concentration of 95 mM and unable to cope with
acetic acid at 100 mM. Cells lacking PDR18 expression were unable to grow in the presence of 75
mM acetic acid (Figure 4.2). Furthermore, the expression of PDR18 from a centromeric plasmid in
Δpdr18 was found to partially rescue acetic acid susceptibility phenotype exhibited by this mutant,
since the acetic acid concentration in which there was no detectable growth of the
Δpdr18+pRS416_PDR18 strain, expressing the PDR18 gene from its natural promoter, was higher
than MIC value attained by the deletion mutant Δpdr18 cells harbouring the empty plasmid pRS416
(Figure 4.2). Also, BY4741 cells harbouring the plasmid pRS416_PDR18 were found to reach a MIC
value of 110 mM, being able to grow in the presence of higher acetic acid levels than the parental
strain BY4741 (Figure 4.2).
Figure 4.1 - PDR18 expression leads to increased yeast tolerance to acetic acid. Growth curves
of S. cerevisiae parental strain BY4741 and the derived deletion mutant Δpdr18 strain at 30ºC. Cell
suspensions of BY4741 (○, ●) and Δpdr18 (□, ■), grown in MM4 liquid medium (pH 4.0) without acetic
acid supplementation were used to inoculate MM4 liquid medium in the absence (open symbols) or
presence (closed symbols) of 60 mM acetic acid. Growth curves were followed measuring culture
OD600nm and are representative of at least three independent growth experiments.
19
Figure 4.2 – Effect of PDR18 expression in yeast acetic acid tolerance. Comparison of the acetic
acid concentration values which completely inhibit growth of BY4741 and the deletion mutant Δpdr18
strains, harbouring the PDR18 expression plasmid (pRS16_PDR18) or the corresponding cloning
vector pRS416. Minimum inhibitory concentrations (MIC) were determined by lack of detectable
growth after 240 hours of incubation at 30ºC. Values are representative of three independent
experiments that gave rise to the same results. Values result from data presented in Annexes I and II.
4.1.2. Inhibitory effect of acetic acid in yeast growth at temperatures in the range 30-
40ºC
Yeast cells were cultivated at 30°C until mid-exponential phase, and then, these cells were used to
inoculate series of 100 mL Erlenmeyers flasks with 50 mL of growth liquid medium MM4
supplemented or not (control) with increasing concentrations of acetic acid and incubated at the
temperature range 30-40°C (section 3.2; Materials and Methods). Additionally, the minimum acetic
acid concentrations required to inhibit growth of S. cerevisiae BY4741 parental strain and derived
deletion mutant Δpdr18 were determined in the temperature range 30-40°C.
Results obtained demonstrate a marked reduction in the biomass concentration at the stationary
phase of growth of the cultivation of cells devoid of PDR18, specially when incubated at 40°C (Figure
4.3). Indeed, PDR18 expression was found to lead to a higher biomass concentration at the stationary
phase of growth when the yeast cells, pre-cultivated at 30°C, were incubated at temperatures 38°C
and 40°C (Figure 4.3). Regarding acetic acid tolerance at different temperatures, an increase in lag
phase duration and a reduction of the biomass concentration values at the stationary phase of growth
were observed as the growth temperature increases from 30ºC to 40ºC, for the same acetic acid
concentrations (Figure 4.4; Figure 4.5). This decrease in tolerance to acetic acid with increasing
growth temperatures up to 40°C was more pronounced in deletion mutant cells Δpdr18. In fact, lower
concentrations of acetic acid had to be tested, because cells lacking Pdr18 were not able to grow in
the presence of acetic acid 55 mM at 38°C and 40 mM at 40°C (Figure 4.5 and Figure 4.6).
20
PDR18 expression was seen to contribute to increase yeast tolerance not only to acetic acid, but also
to the conjugated effect of this stress with supraoptimal temperatures (Figure 4.4 and Figure 4.5). In
general, data obtained in this study indicate that Pdr18 exert a protective effect against the deleterious
additive action of acetic acid and supraoptimal temperatures.
Figure 4.3 – S. cerevisiae BY4741 and the derived deletion mutant Δpdr18 growth at
supraoptimal temperatures. Cell suspensions of BY4741 and Δpdr18 pre-cultivated at 30°C until
mid-exponential phase (OD600nm = 0.5±0.05) in MM4 liquid medium (pH 4.0) were used to re-inoculate
fresh MM4 liquid medium (pH 4.0), at an OD600nm = 0.1±0.05. Culture OD600nm obtained during 240
hours of incubation at 30°C (in blue ■), 36°C (in orange ■), 38°C (in light blue ■) and 40°C (in red ■)
are shown. Values result from data presented in Figures A.1 to A.4 from Annex I and are
representative of three independent growth experiments.
Figure 4.4 - Effect of acetic acid stress in the growth of S. cerevisiae BY4741 parental strain
and derived deletion mutant Δpdr18 at 30°C. Culture OD600nm obtained during 240 hours of
incubation at 30°C in liquid MM4 supplemented with increasing concentrations of acetic acid, at pH 4.0
are shown. Values result from data presented in Figures A.1 from Annex I and are representative of
three independent growth experiments.
Colors in the figure correspond to the following acetic acid concentrations:

21
Figure 4.5 - Conjugated effect of acetic acid and supraoptimal temperatures in the growth of S.
cerevisiae BY4741 parental strain and derived deletion mutant Δpdr18. Culture OD600nm obtained
during 240 hours of incubation at 36°C, 38°C and 40°C in liquid MM4 at pH 4.0, supplemented with
increasing concentrations of acetic acid, are shown. Values result from data presented in Figures A.2
to A.4 from Annex I and are representative of three independent growth experiments.
Colors in the figure correspond to the following acetic acid concentrations:
22
Figure 4.6 – Minimum concentrations of acetic acid required to inhibit the growth of S.
cerevisiae BY4741 parental strain and derived deletion mutant Δpdr18 at four different
temperatures (30, 36, 38 and 40°C). Values were determined by lack of detectable growth after 240
hours of incubation at temperatures in the range 30-40°C. Values result from data presented in Figure
A.1 to A.4 and Figure C.1, from Annexes I and III, respectively, and are representative of three
independent experiments.
4.2. PDR18 expression is essential to counteract the decrease of plasma membrane
potential in yeast cells cultivated with inhibitory concentrations of acetic acid
In order to understand the effect of Pdr18 on yeast tolerance to acetic acid, we compared the
membrane potential of S. cerevisiae BY4741 and derived deletion mutant Δpdr18 cells harvested in
mid-exponential phase of growth in the presence or absence of 60 mM acetic acid. Two methods were
used to estimate plasma membrane potential: the [14
C]-methylamine uptake assay (Figure 4.7) and
the DiOC6(3) (3-3’-dihexyloxacarbocianine iodide accumulation assay (Figure 4.8).
In the first technique, yeast plasma membrane potential was assessed using radioactively labelled
methylammonium, which is a non-metabolizable ammonium analogue and its uptake is dependent on
the plasma membrane potential (Mulet et al., 1999). The second method used is based on the
accumulation of DiOC6(3) probe, which also relies on yeast plasma membrane potential. Regarding
this microscopy technique, an increase in fluorescence denotes higher level of plasma membrane
polarization (Madrid et al., 1998). Hence, the effect of PDR18 gene deletion on plasma membrane
potential was evaluated in cells previously cultivated in liquid media supplemented or not with acetic
acid (60 mM, pH 4.0) (Figure 4.7 and Figure 4.8). In control conditions, a depolarization of the plasma
membrane of cells lacking PDR18 expression, when compared to the parental strain, was detected
using both methods (Figure 4.7 A1 and A2 and Figure 4.8 A1 and A2), as previously described
(Cabrito et al., 2011). In [14
C]-methylamine uptake assays, this decrease in membrane potential of
0.0
20.0
40.0
60.0
80.0
100.0
30 36
38 40
Aceti
c a
cid
co
ncen
trati
on
(m
M)
Temperature of growth (ºC)
BY4741_Δpdr18
BY4741

23
Δpdr18 cells was considered statistically not significant (Figure 4.7 B). However, in DiOC6(3)
accumulation assays, this decrease in membrane potential of deletion mutant Δpdr18 cells was more
pronounced and the difference from the membrane potential observed in BY4741 cells was
considered extremely significant (P < 0.0001) (Figure 4.8 B).
In yeast cells grown in the presence of inhibitory concentration of acetic acid, methylammonium
accumulation assay demonstrated that PDR18 expression lead to a significantly higher plasma
membrane potential (P < 0.05), than the observed in the same cells in the absence of stress (Figure
4.7 A1 and B). In fluorescence microscopy results, this difference in plasma membrane potential
between BY4741 cells grown in the absence and in the presence of acetic acid was seen to be
extremely significant (P < 0.0001) (Figure 4.8 A1 and B). On the contrary, in cells devoid of the PDR18
gene cultivated in the presence of inhibitory concentrations of acetic acid, there is a decrease in
membrane potential (Figure 4.7 A2 and B and Figure 4.8 A2 and B). This difference in membrane
potential between cells expressing or not the PDR18 gene, cultivated in the presence of acetic acid,
was found to be extremely significant (P < 0.0001) in DiOC6(3) accumulation assays (Figure 4.8 B)
and very significant (P < 0.01) in [14
C]-methylamine uptake assays (Figure 4.7 B). Therefore, results
obtained show that Pdr18 is responsible for an increase in membrane potential of yeast cells grown in
the presence of 60 mM acetic acid (Figures 4.7 and 4.8), in order to counteract the dissipation of the
plasma membrane potential caused by acetic acid.
Although [14
C]-methylamine uptake assays have shown differences less significant than in DiOC6(3)
accumulation assays, results obtained in both methods have a similar tendency regarding changes in
plasma membrane potential. In DiOC6(3) accumulation assays, significant differences were displayed
in the statistical tests. This is probably due to the differences in the two approaches, as in
fluorescence microscopy each cell is considered an individual experiment and it is thus, possible to
notice differences within the yeast population. At the same time, in [14
C]-methylamine uptake assay,
the population is considered as a whole and an average value is obtained for all yeast cells in the
population. Indeed, the distribution of fluorescence among a population of BY4741 and deletion
mutant Δpdr18 cells loaded with DiOC6(3) in control conditions was shown to be well described by
a Gaussian distribution (Figure 4.8 A1 and A2). Nevertheless, BY4741 cell population harvested from
a mid-exponential phase culture carried out in the presence of acetic acid, proved to be equally
distributed among the different fluorescence intensity classes, which is indicative of a highly
heterogeneous population (Figure 4.8 A1). For the deletion mutant Δpdr18 cells, a Gaussian
distribution of cell population with a translation of the average intensity to lower values of fluorescence
was registered (Figure 4.8 A2), suggesting that the lack of PDR18 leads to a decrease of membrane
potential in cells cultivated in the presence of acetic acid (Figure 4.8 A2).

24
Figure 4.7 - PDR18 gene expression is essential to counteract the dissipation of plasma
membrane potential in yeast cells cultivated in the presence of inhibitory concentrations of
acetic acid (60 mM, pH 4.0). A: Time-course accumulation of [14
C]-methylammonium during
incubation at 30ºC of S. cerevisiae parental strain BY4741 (A1) and derived deletion mutant strain
BY4741_Δpdr18 (A2) yeast cells in MES buffer with glucose (20 g/L) supplemented with the
radiolabelled methylammonium. These yeast cells were cultivated until mid-exponential phase in the
absence (open symbols) or presence (closed symbols) of 60 mM acetic acid, at 30ºC, as described in
section 3.3.1 . Relative levels of [14
C]-methylammonium were assessed during 20 minutes, as
described in section 3.3.1, and are the average of at least three independent experiments. The error
bars represent standard deviation. B: Accumulation of [14
C]-methylammonium in BY4741 and deletion
mutant Δpdr18 cells after 20 minutes, as previously described. Values are the average of at least
three independent experiments and the error bars represent standard deviation. Ns indicates P > 0.05;
a single asterisk (*) indicates P < 0.05; double asterisks (**) indicate P < 0.01.

25
Figure 4.8 – Distribution of cell population membrane potential, obtained from fluorescence
microscopy using DiOC6(3) of the parental strain BY4741 (A1) and derived deletion mutant strain
BY4741_Δpdr18 (A2) harvested from a mid-exponential culture growth performed in absence (white
bars) or presence (black bars) of acetic acid (60 mM). B: Comparison of the average membrane
potential using the fluorescent probe DiOC6(3). Values of membrane potential are set as the
percentage of the value obtained for BY4741 cells in control conditions and are means for at least
three independent experiments. Error bars represent the correspondent standard deviation. Quadruple
asterisks (****) indicate P < 0.0001.

26
4.3. Effect of acetic acid and PDR18 expression on the active efflux of H+
Yeast cells under weak acid stress conditions undergo a strong activation of plasma membrane H+-
ATPase in the plasma membrane, resulting in the efflux of the excess amounts of the protons to the
outside, counteracting the toxic effects of weak acids (Mira et al., 2010c). Extracellular acidification
depends on the plasma membrane H+-ATPase proton pumping activity and on the plasma membrane
permeability, which influences the passive diffusion of protons into yeast cells (Rosa and Sá-Correia,
1994). In order to examine the effect of PDR18 expression in the extracellular acidification of cells
grown until mid-exponential phase in liquid media supplemented or not (control conditions) with acetic
acid (60 mM, pH 4.0), the final extracellular pH and the extracellular acidification rate obtained were
compared between each strain at different experimental conditions (Figure 4.9). Results show that
yeast cells devoid of the PDR18 gene do not have an external acidification significantly different from
the one exhibithed by the parental strain BY4741, when cultivated in the absence of acetic acid
(pHextracellular ≈ 3.2 for Δpdr18 and pHextracellular ≈ 2.9 for BY4741) (Figure 4.9 A1 and A2). When yeast
cells were cultivated in the presence of an inhibitory concentration of acetic acid (60 mM, pH 4.0),
although differences regarding final extracellular pH, as well as external acidification rate, between
cells expressing or not the PDR18 gene were observed, statistical tests considered them as not
significant (Figure 4.9 B and C).
Deletion mutant Δpdr18 cells cultivated in the presence of acetic acid exhibited a significant decrease
(P < 0.001) in the acidification rate and, consequently, an increase in the final extracellular pH
(pHextracellular ≈ 3.5) (Figure 4.9 B and C), compared to the same cells grown under control conditions
(Figure 4.9 A2). Furthermore, differences in the extracellular acidification between cells devoid of
PDR18 cultivated in control conditions and in the presence of acetic acid were more significant, than
among cells expressing PDR18 gene. This is probably due to the previous results showing that yeast
cell plasma membrane permeabilization caused by acetic acid is extremely pronounced in cells
lacking PDR18 expression (Godinho et al., unpublished results). Nevertheless, a significant reduction
of the rate of extracellular acidification (P < 0.05) was observed in BY4741 cells previously cultivated
in the presence of acetic acid (Figure 4.9 A1 and C), which led to an increase in the less acidic final
pHextracellular of 3.2, rather than the 2.9 obtained when cells were harvested from control conditions
(Figure 4.9 A1).

27
Figure 4.9 – Effect of PDR18 gene expression in the external acidification rate in cells
cultivated under control conditions or in the presence of inhibitory concentrations of acetic
acid. A: The extracellular acidification of energized cells was assessed for cells of the parental strain
BY4741 (A1) and the derived deletion mutant Δpdr18 (A2) harvested from mid-exponential culture
grown in the absence (open symbols) or presence (closed symbols) of acetic acid (60 mM, pH 4.0), at
30ºC. B: Final extracellular pH after the 10 minutes following the addition of glucose. C: External
acidification rate during the 10 minutes after addition of glucose. Values are the average of three
independent experiments and error bars represent the correspondent standard deviation. Ns indicates
P > 0.05; a single asterisk (*) indicates P < 0.05; double asterisks (**) indicate P < 0.01; and quadruple
asterisks (****) indicate P < 0.0001.

28
5. Discussion
The involvement of the Saccharomyces cerevisiae multidrug resistance (MDR) ABC transporter Pdr18
in yeast tolerance to the conjugated effect of acetic acid and supraoptimal temperatures was
examined. Results indicate that PDR18 expression contributes to increased tolerance of S. cerevisiae
BY4741 strain to acetic acid, being part in the global response that allow yeast to overcome acetic
acid induced stress. Based on the relevance of PDR18 gene expression in acetic acid stress
tolerance, and considering the role of ergosterol as a stabilizer of membrane components in heat and
ethanol tolerance (Swan and Watson, 1998), the effect of temperature in yeast growth in the presence
of inhibitory concentrations of acetic acid was evaluated in the parental S. cerevisiae BY4741 strain
and in the derived deletion mutant Δpdr18. Our results showed that yeast growth of both the parental
and the derived deletion mutant Δpdr18 strains at temperatures in the range 30-40°C could be
arrested by acetic acid, although at significantly different concentrations. Furthermore, when yeast
cells were incubated at the supraoptimal temperatures 38°C and 40°C, in the presence of increasing
concentrations of acetic acid, a marked decrease in tolerance towards this weak acid in the deletion
mutant Δpdr18 was observed. Indeed, PDR18 deletion proved to lead to extended lag-phase period
and reduced values of biomass concentration at the stationary phase of growth, when compared to
the same parameters in the parental strain BY4741. These may be indicative of Pdr18 contribution to
counteract the additive deleterious effect of this weak acid and supraoptimal temperatures on the
organization and functionality of biological membranes that lead to increased permeability and
modifications in fluidity (Salgueiro et al., 1988, Verghese et al., 2012). Survival depends on the
protection of the optimal level of the plasma membrane fluidity (Turk et al., 2011). In fact, ergosterol
incorporation in the yeast plasma membrane, in which Pdr18 is described to be involved (Cabrito et
al., 2011), is a mechanism that contributes to the maintenance of adequate fluidity and physiological
functionality of plasma membrane (Beney and Gervais, 2001, Dupont et al., 2011, Klose et al., 2012).
Therefore, these results are consistent with the need of ergosterol as a stabilizer of yeast plasma
membrane (Swan and Watson, 1998), confirming Pdr18 role in the yeast protective response to these
environmental stresses.
Furthermore, a recent study hypothesizes the decrease of sterol functions at high temperature and
observes the presence of “bended” fecosterol, rather than “flat” ergosterol in the membrane of S.
cerevisiae strains that developed thermotolerance (Caspeta et al., 2014). Therefore, since membrane
fluidity is influenced by the ratio, composition, and structure of sterols that are found in membranes
(Caspeta et al., 2014), a comparison of the relative amounts of ergosterol and other sterols in yeast
plasma membrane of the BY4741 strain and the derived deletion mutant Δpdr18 both cultivated in the
absence or presence of acetic acid at supraoptimal temperatures is proposed as future work, in order
to elucidate the specific function of Pdr18 in response to these conjugated stresses.
In order to clarify the physiological role of Pdr18 in the protective response against acetic acid stress,
the effect of PDR18 expression in yeast plasma membrane potential of cells grown in the presence of
inhibitory concentrations of acetic acid was examined. To achieve more confident results, two

29
methods were used to compare the differences in plasma membrane potential between parental
BY4741 cells and deletion mutant BY4741_Δpdr18. Results from both techniques are in agreement
and demonstrated that in cells cultivated with 60 mM acetic acid, PDR18 expression lead to an
increase of yeast plasma membrane potential, counteracting the proposed effect of this weak acid in
the dissipation of plasma membrane potential (Mira et al., 2010c). The deletion of PDR18 gene leads
to a reduction of the plasma membrane potential in cells cultivated with acetic acid, which is
hypothesized to be related with the fact that in cells lacking Pdr18, acetic acid-induced stress was
found to increase membrane permeabilization (Godinho et al., unpublished results). Given that
ergosterol plays an essential role in the normal physiological function of the plasma membrane by
regulating its fluidity and the delicate balance among other membrane components such as lipids and
proteins (Aguilera et al., 2006), membrane depolarization and increased permeabilization observed in
Δpdr18 cells may be related with low levels of ergosterol in the plasma membrane, corroborating
Pdr18 contribution against acetic acid stress in yeast. In addition, since cells used in these
experiments were previously cultivated in the presence of acetic acid (60 mM) and membrane
potential measurements were performed in the absence of stress and in the presence of glucose,
these results indicate that when acetic acid is present in the growth medium, a proper modification of
plasma membrane composition might have occurred in cells expressing PDR18, resulting in a less
harmful plasma membrane permeabilization. Thus, these results suggest that the proposed
modifications in plasma membrane lipid configuration and cell wall structure of cells in response to
weak acids (Mira et al., 2010c, Viegas et al., 2005, Simoes et al., 2006) may also involve PDR18
expression. Therefore, we hypothesize that Pdr18 involvement in ergosterol incorporation in the
membrane may contribute to the remodeling of yeast plasma membrane, leading to the reduction of
the permeabilizing effect of acetic acid and, consequently, preventing its harmful effects, as a
protective response to acetic acid stress.
Additionally, results obtained in plasma membrane potential assays using fluorescence microscopy
demonstrated that BY4741 and derived deletion mutant Δpdr18 strains cultivated in the absence of
acetic acid, displayed a Gaussian distribution of cell population. However, BY4741 cell population
presented an equally distribution among the different fluorescence intensity classes, when harvested
from a mid-exponential phase culture carried out in the presence of acetic acid. This distribution
showed that some cells displayed higher levels of fluorescence, and thus increased plasma
membrane potential; indicative of the proposed plasma membrane remodeling in cells expressing
PDR18, and decreased membrane permeabilization (Godinho et al., unpublished results). Cells
lacking Pdr18 cultivated in the presence of acetic acid proved to have a Gaussian distribution of cell
population, as in control conditions, although with a higher number of cells with lower membrane
potential, which is correlated with the increased membrane permeabilization of these cells (Godinho et
al., unpublished results). These differences in cell population distribution corroborate the protective
role of Pdr18 against acetic acid deleterious effects in yeast plasma membrane.
Moreover, since Pdr18 was proposed to have a role in reducing plasma membrane permeabilization,
as a response to acetic acid-induced stress, the effect of PDR18 expression in the extracellular

30
acidification of cells cultivated under control conditions and in the presence of inhibitory concentrations
of acetic acid, was compared. Extracellular acidification in deletion mutant Δpdr18 cells grown under
acetic acid stress conditions was significantly lower, than Δpdr18 cells grown in the absence of stress,
as expected, regarding plasma membrane potential results. This confirms that in these cells, there is
an increase of the rate of passive protons influx, which cannot be counterbalanced by the ATPase
activity, because of the enhanced permeabilizing effect of acetic acid in the plasma membrane of cells
devoid of Pdr18 (Godinho et al., unpublished results). In cells expressing the PDR18 gene, previously
cultivated in the presence of acetic acid, a reduction in the rate of extracellular acidification was
observed which is also mainly related with the passive proton influx, due to the permeabilization of the
plasma membrane caused by acetic acid (Godinho et al., unpublished results). In this case, results
obtained in extracellular acidification do not illustrate the proposed effect of PDR18 expression in the
reduction of the plasma membrane permeabilization in cells cultivated in the presence of acetic acid
observed in plasma membrane potential assays, which may be explained by limitations associated
with the experimental methodology used. Since it is an in vivo method, there are other phenomenon
and mechanisms occurring in the cell, despite the in vivo activity of the H+-ATPase and the passive H
+
influx through the plasma membrane that were roughly evaluated by the extracellular acidification
curves. In fact, this result may also be related with different availability of ATP in yeast cells, despite of
the activation of H+-ATPase, disabling the proton pumping activity, and, consequently, the acidification
of the extracellular media. Nevertheless, although statistical tests displayed differences not significant
in the final extracellular pH and extracellular acidification rate between cells expressing or not PDR18,
cultivated in the presence of acetic acid, the deletion of PDR18 was found to lead to a lower
extracellular acidification rate and a higher final extracellular pH, than cells expressing PDR18, which
demonstrate that these results have a similar tendency as those obtained in membrane potential
assays.
Altogether, results support the hypothesis that Pdr18 may be essential for the maintenance of the
membrane function and to prevent membrane permeabilization in yeast cells growing in the presence
of inhibitory concentrations of acetic acid. This work contributes to improve our understanding of the
ABC transporter Pdr18 biological function in the context of multidrug resistance, in the eukaryotic
model S. cerevisiae. The importance of the combined tolerance to both of the stress factors examined
is relevant for the improvement of the fermentation performance envisaging more efficient production
of bioethanol using lignocellulosic hydrolysates.

31
References
AGUILERA, F., PEINADO, R. A., MILLAN, C., ORTEGA, J. M. & MAURICIO, J. C. 2006. Relationship between ethanol tolerance, H+ -ATPase activity and the lipid composition of the plasma membrane in different wine yeast strains. Int J Food Microbiol, 110, 34-42.
ALMEIDA, B., OHLMEIER, S., ALMEIDA, A. J., MADEO, F., LEAO, C., RODRIGUES, F. & LUDOVICO, P. 2009. Yeast protein expression profile during acetic acid-induced apoptosis indicates causal involvement of the TOR pathway. Proteomics, 9, 720-32.
BAGNAT, M., KERANEN, S., SHEVCHENKO, A., SHEVCHENKO, A. & SIMONS, K. 2000. Lipid rafts function in biosynthetic delivery of proteins to the cell surface in yeast. Proc Natl Acad Sci U S A, 97, 3254-9.
BALZI, E. & GOFFEAU, A. 1994. Genetics and biochemistry of yeast multidrug resistance. Biochim Biophys Acta, 1187, 152-62.
BALZI, E. & GOFFEAU, A. 1995. Yeast multidrug resistance: the PDR network. J Bioenerg Biomembr, 27, 71-6.
BELTRAN, G., NOVO, M., GUILLAMON, J. M., MAS, A. & ROZES, N. 2008. Effect of fermentation temperature and culture media on the yeast lipid composition and wine volatile compounds. Int J Food Microbiol, 121, 169-77.
BENEY, L. & GERVAIS, P. 2001. Influence of the fluidity of the membrane on the response of microorganisms to environmental stresses. Appl Microbiol Biotechnol, 57, 34-42.
BUKAU, B. & HORWICH, A. L. 1998. The Hsp70 and Hsp60 chaperone machines. Cell, 92, 351-66.
CABRITO, T. R., TEIXEIRA, M. C., SINGH, A., PRASAD, R. & SA-CORREIA, I. 2011. The yeast ABC transporter Pdr18 (ORF YNR070w) controls plasma membrane sterol composition, playing a role in multidrug resistance. Biochem J, 440, 195-202.
CARMELO, V., BOGAERTS, P. & SA-CORREIA, I. 1996. Activity of plasma membrane H+-ATPase and expression of PMA1 and PMA2 genes in Saccharomyces cerevisiae cells grown at optimal and low pH. Arch Microbiol, 166, 315-20.
CARRATU, L., FRANCESCHELLI, S., PARDINI, C. L., KOBAYASHI, G. S., HORVATH, I., VIGH, L. & MARESCA, B. 1996. Membrane lipid perturbation modifies the set point of the temperature of heat shock response in yeast. Proc Natl Acad Sci U S A, 93, 3870-5.

32
CASAL, M., CARDOSO, H. & LEAO, C. 1996. Mechanisms regulating the transport of acetic acid in Saccharomyces cerevisiae. Microbiology, 142 ( Pt 6), 1385-90.
CASPETA, L., CHEN, Y., GHIACI, P., FEIZI, A., BUSKOV, S., HALLSTROM, B. M., PETRANOVIC, D. & NIELSEN, J. 2014. Biofuels. Altered sterol composition renders yeast thermotolerant. Science, 346, 75-8.
CYERT, M. S. 2003. Calcineurin signaling in Saccharomyces cerevisiae: how yeast go crazy in response to stress. Biochem Biophys Res Commun, 311, 1143-50.
DE VIRGILIO, C., HOTTIGER, T., DOMINGUEZ, J., BOLLER, T. & WIEMKEN, A. 1994. The role of trehalose synthesis for the acquisition of thermotolerance in yeast. I. Genetic evidence that trehalose is a thermoprotectant. Eur J Biochem, 219, 179-86.
DECOTTIGNIES, A., GRANT, A. M., NICHOLS, J. W., DE WET, H., MCINTOSH, D. B. & GOFFEAU, A. 1998. ATPase and multidrug transport activities of the overexpressed yeast ABC protein Yor1p. J Biol Chem, 273, 12612-22.
DEL SORBO, G., SCHOONBEEK, H. & DE WAARD, M. A. 2000. Fungal transporters involved in efflux of natural toxic compounds and fungicides. Fungal Genet Biol, 30, 1-15.
DESMOUCELLES, C., PINSON, B., SAINT-MARC, C. & DAIGNAN-FORNIER, B. 2002. Screening the yeast "disruptome" for mutants affecting resistance to the immunosuppressive drug, mycophenolic acid. J Biol Chem, 277, 27036-44.
DEXTER, D., MOYE-ROWLEY, W. S., WU, A. L. & GOLIN, J. 1994. Mutations in the yeast PDR3, PDR4, PDR7 and PDR9 pleiotropic (multiple) drug resistance loci affect the transcript level of an ATP binding cassette transporter encoding gene, PDR5. Genetics, 136, 505-15.
DICKSON, R. C. & LESTER, R. L. 2002. Sphingolipid functions in Saccharomyces cerevisiae. Biochim Biophys Acta, 1583, 13-25.
DOGAN, A., DEMIRCI, S., AYTEKIN, A. O. & SAHIN, F. 2014. Improvements of Tolerance to Stress Conditions by Genetic Engineering in Saccharomyces Cerevisiae during Ethanol Production. Appl Biochem Biotechnol.
DUPONT, S., BENEY, L., FERREIRA, T. & GERVAIS, P. 2011. Nature of sterols affects plasma membrane behavior and yeast survival during dehydration. Biochim Biophys Acta, 1808, 1520-8.
EISENKOLB, M., ZENZMAIER, C., LEITNER, E. & SCHNEITER, R. 2002. A specific structural requirement for ergosterol in long-chain fatty acid synthesis mutants important for maintaining raft domains in yeast. Mol Biol Cell, 13, 4414-28.

33
EKLUND, R. Z., G. 1995. Simultaneous saccharification and fermentation of steam-pretreated willow. Enzyme and Microbial Technology
EKLUND, S. A., PITTMAN, J. L. & SMITH, R. C. 1997. Trends in dental care among insured Americans: 1980 to 1995. J Am Dent Assoc, 128, 171-8.
ELLIOTT, B., HALTIWANGER, R. S. & FUTCHER, B. 1996. Synergy between trehalose and Hsp104 for thermotolerance in Saccharomyces cerevisiae. Genetics, 144, 923-33.
FERNANDES, A. R., DURAO, P. J., SANTOS, P. M. & SA-CORREIA, I. 2003. Activation and significance of vacuolar H+-ATPase in Saccharomyces cerevisiae adaptation and resistance to the herbicide 2,4-dichlorophenoxyacetic acid. Biochem Biophys Res Commun, 312, 1317-24.
FERNANDES, A. R., MIRA, N. P., VARGAS, R. C., CANELHAS, I. & SA-CORREIA, I. 2005. Saccharomyces cerevisiae adaptation to weak acids involves the transcription factor Haa1p and Haa1p-regulated genes. Biochem Biophys Res Commun, 337, 95-103.
FERNANDES, A. R. & SA-CORREIA, I. 2001. The activity of plasma membrane H(+)-ATPase is strongly stimulated during Saccharomyces cerevisiae adaptation to growth under high copper stress, accompanying intracellular acidification. Yeast, 18, 511-21.
GALBE, M. & ZACCHI, G. 2002. A review of the production of ethanol from softwood. Appl Microbiol Biotechnol, 59, 618-28.
GANCEDO, J. M. 1998. Yeast carbon catabolite repression. Microbiol Mol Biol Rev, 62, 334-61.
GBELSKA, Y., KRIJGER, J. J. & BREUNIG, K. D. 2006. Evolution of gene families: the multidrug resistance transporter genes in five related yeast species. FEMS Yeast Res, 6, 345-55.
GRAVES, T., NARENDRANATH, N. V., DAWSON, K. & POWER, R. 2006. Effect of pH and lactic or acetic acid on ethanol productivity by Saccharomyces cerevisiae in corn mash. J Ind Microbiol Biotechnol, 33, 469-74.
GREETHAM, D., WIMALASENA, T., KERRUISH, D. W., BRINDLEY, S., IBBETT, R. N., LINFORTH, R. L., TUCKER, G., PHISTER, T. G. & SMART, K. A. 2014. Development of a phenotypic assay for characterisation of ethanologenic yeast strain sensitivity to inhibitors released from lignocellulosic feedstocks. J Ind Microbiol Biotechnol, 41, 931-45.
GULSHAN, K. & MOYE-ROWLEY, W. S. 2007. Multidrug resistance in fungi. Eukaryot Cell, 6, 1933-42.
HAHN-HAGERDAL, B., GALBE, M., GORWA-GRAUSLUND, M. F., LIDEN, G. & ZACCHI, G. 2006. Bio-ethanol--the fuel of tomorrow from the residues of today. Trends Biotechnol, 24, 549-56.

34
HATZIXANTHIS, K., MOLLAPOUR, M., SEYMOUR, I., BAUER, B. E., KRAPF, G., SCHULLER, C., KUCHLER, K. & PIPER, P. W. 2003. Moderately lipophilic carboxylate compounds are the selective inducers of the Saccharomyces cerevisiae Pdr12p ATP-binding cassette transporter. Yeast, 20, 575-85.
HIGGINS, C. F. 1992. ABC transporters: from microorganisms to man. Annu Rev Cell Biol, 8, 67-113.
HIGGINS, C. F. 1995. The ABC of channel regulation. Cell, 82, 693-6.
HIGGINS, C. F. 2001. ABC transporters: physiology, structure and mechanism--an overview. Res Microbiol, 152, 205-10.
HIGGINS, C. F. 2007. Multiple molecular mechanisms for multidrug resistance transporters. Nature, 446, 749-57.
HOEBERICHTS, F. A., PEREZ-VALLE, J., MONTESINOS, C., MULET, J. M., PLANES, M. D., HUESO, G., YENUSH, L., SHARMA, S. C. & SERRANO, R. 2010. The role of K(+) and H(+) transport systems during glucose- and H(2)O(2)-induced cell death in Saccharomyces cerevisiae. Yeast, 27, 713-25.
HOLYOAK, C. D., STRATFORD, M., MCMULLIN, Z., COLE, M. B., CRIMMINS, K., BROWN, A. J. & COOTE, P. J. 1996. Activity of the plasma membrane H(+)-ATPase and optimal glycolytic flux are required for rapid adaptation and growth of Saccharomyces cerevisiae in the presence of the weak-acid preservative sorbic acid. Appl Environ Microbiol, 62, 3158-64.
INGRAM, L. O., ALDRICH, H. C., BORGES, A. C., CAUSEY, T. B., MARTINEZ, A., MORALES, F., SALEH, A., UNDERWOOD, S. A., YOMANO, L. P., YORK, S. W., ZALDIVAR, J. & ZHOU, S. 1999. Enteric bacterial catalysts for fuel ethanol production. Biotechnol Prog, 15, 855-66.
JENKINS, G. M., RICHARDS, A., WAHL, T., MAO, C., OBEID, L. & HANNUN, Y. 1997. Involvement of yeast sphingolipids in the heat stress response of Saccharomyces cerevisiae. J Biol Chem, 272, 32566-72.
JUNGWIRTH, H. & KUCHLER, K. 2006. Yeast ABC transporters-- a tale of sex, stress, drugs and aging. FEBS Lett, 580, 1131-8.
JURANKA, P. F., ZASTAWNY, R. L. & LING, V. 1989. P-glycoprotein: multidrug-resistance and a superfamily of membrane-associated transport proteins. FASEB J, 3, 2583-92.
KATZMANN, D. J., BURNETT, P. E., GOLIN, J., MAHE, Y. & MOYE-ROWLEY, W. S. 1994. Transcriptional control of the yeast PDR5 gene by the PDR3 gene product. Mol Cell Biol, 14, 4653-61.

35
KAWAHATA, M., MASAKI, K., FUJII, T. & IEFUJI, H. 2006. Yeast genes involved in response to lactic acid and acetic acid: acidic conditions caused by the organic acids in Saccharomyces cerevisiae cultures induce expression of intracellular metal metabolism genes regulated by Aft1p. FEMS Yeast Res, 6, 924-36.
KLOSE, C., SURMA, M. A., GERL, M. J., MEYENHOFER, F., SHEVCHENKO, A. & SIMONS, K. 2012. Flexibility of a eukaryotic lipidome--insights from yeast lipidomics. PLoS One, 7, e35063.
KOLACZKOWSKA, A. & GOFFEAU, A. 1999. Regulation of pleiotropic drug resistance in yeast. Drug Resist Updat, 2, 403-414.
KREBS, H. A., WIGGINS, D., STUBBS, M., SOLS, A. & BEDOYA, F. 1983. Studies on the mechanism of the antifungal action of benzoate. Biochem J, 214, 657-63.
LALUCE, C., SCHENBERG, A. C., GALLARDO, J. C., CORADELLO, L. F. & POMBEIRO-SPONCHIADO, S. R. 2012. Advances and developments in strategies to improve strains of Saccharomyces cerevisiae and processes to obtain the lignocellulosic ethanol--a review. Appl Biochem Biotechnol, 166, 1908-26.
LI, Y. & PRINZ, W. A. 2004. ATP-binding cassette (ABC) transporters mediate nonvesicular, raft-modulated sterol movement from the plasma membrane to the endoplasmic reticulum. J Biol Chem, 279, 45226-34.
LU, Y., CHENG, Y. F., HE, X. P., GUO, X. N. & ZHANG, B. R. 2012. Improvement of robustness and ethanol production of ethanologenic Saccharomyces cerevisiae under co-stress of heat and inhibitors. J Ind Microbiol Biotechnol, 39, 73-80.
MADEIRA-LOPES, A. & VAN UDEN, N. 1979. Thermal association and dissociation in thermosensitive mutants of Saccharomyces cerevisiae. Z Allg Mikrobiol, 19, 303-5.
MADRID, R., GOMEZ, M. J., RAMOS, J. & RODRIGUEZ-NAVARRO, A. 1998. Ectopic Potassium Uptake in trk1 trk2 Mutants ofSaccharomyces cerevisiae Correlates with a Highly Hyperpolarized Membrane Potential. Journal of Biological Chemistry, 273, 14838-14844.
MAKRANTONI, V., DENNISON, P., STARK, M. J. & COOTE, P. J. 2007. A novel role for the yeast protein kinase Dbf2p in vacuolar H+-ATPase function and sorbic acid stress tolerance. Microbiology, 153, 4016-26.
MAMNUN, Y. M., PANDJAITAN, R., MAHE, Y., DELAHODDE, A. & KUCHLER, K. 2002. The yeast zinc finger regulators Pdr1p and Pdr3p control pleiotropic drug resistance (PDR) as homo- and heterodimers in vivo. Mol Microbiol, 46, 1429-40.

36
MAYER, M. P. & BUKAU, B. 1998. Hsp70 chaperone systems: diversity of cellular functions and mechanism of action. Biol Chem, 379, 261-8.
MIRA, N. P., BECKER, J. D. & SA-CORREIA, I. 2010a. Genomic expression program involving the Haa1p-regulon in Saccharomyces cerevisiae response to acetic acid. OMICS, 14, 587-601.
MIRA, N. P., LOURENCO, A. B., FERNANDES, A. R., BECKER, J. D. & SA-CORREIA, I. 2009. The RIM101 pathway has a role in Saccharomyces cerevisiae adaptive response and resistance to propionic acid and other weak acids. FEMS Yeast Res, 9, 202-16.
MIRA, N. P., PALMA, M., GUERREIRO, J. F. & SA-CORREIA, I. 2010b. Genome-wide identification of Saccharomyces cerevisiae genes required for tolerance to acetic acid. Microb Cell Fact, 9, 79.
MIRA, N. P., TEIXEIRA, M. C. & SA-CORREIA, I. 2010c. Adaptive response and tolerance to weak acids in Saccharomyces cerevisiae: a genome-wide view. OMICS, 14, 525-40.
MORANO, K. A., GRANT, C. M. & MOYE-ROWLEY, W. S. 2012. The response to heat shock and oxidative stress in Saccharomyces cerevisiae. Genetics, 190, 1157-95.
MULET, J. M., LEUBE, M. P., KRON, S. J., RIOS, G., FINK, G. R. & SERRANO, R. 1999. A novel mechanism of ion homeostasis and salt tolerance in yeast: the Hal4 and Hal5 protein kinases modulate the Trk1-Trk2 potassium transporter. Mol Cell Biol, 19, 3328-37.
NOURANI, A., WESOLOWSKI-LOUVEL, M., DELAVEAU, T., JACQ, C. & DELAHODDE, A. 1997. Multiple-drug-resistance phenomenon in the yeast Saccharomyces cerevisiae: involvement of two hexose transporters. Mol Cell Biol, 17, 5453-60.
OLOFSSON, K., BERTILSSON, M. & LIDEN, G. 2008. A short review on SSF - an interesting process option for ethanol production from lignocellulosic feedstocks. Biotechnol Biofuels, 1, 7.
PALMQVIST, E., GRAGE, H., MEINANDER, N. Q. & HAHN-HAGERDAL, B. 1999. Main and interaction effects of acetic acid, furfural, and p-hydroxybenzoic acid on growth and ethanol productivity of yeasts. Biotechnol Bioeng, 63, 46-55.
PAMPULHA, M. E., LOUREIRO-DIAS, M. C. 1990. Activity of glycolytic enzymes of Saccharomyces cerevisiae in the presence of acetic acid. Applied Microbiology and Biotechnology 34, 375-380.
PAO, S. S., PAULSEN, I. T. & SAIER, M. H., JR. 1998. Major facilitator superfamily. Microbiol Mol Biol Rev, 62, 1-34.

37
PARKS, L. W. & CASEY, W. M. 1995. Physiological implications of sterol biosynthesis in yeast. Annu Rev Microbiol, 49, 95-116.
PARSELL, D. A., KOWAL, A. S., SINGER, M. A. & LINDQUIST, S. 1994. Protein disaggregation mediated by heat-shock protein Hsp104. Nature, 372, 475-8.
PEREIRA, F. B., GUIMARAES, P. M., GOMES, D. G., MIRA, N. P., TEIXEIRA, M. C., SA-CORREIA, I. & DOMINGUES, L. 2011. Identification of candidate genes for yeast engineering to improve bioethanol production in very high gravity and lignocellulosic biomass industrial fermentations. Biotechnol Biofuels, 4, 57.
PIPER, P., CALDERON, C. O., HATZIXANTHIS, K. & MOLLAPOUR, M. 2001. Weak acid adaptation: the stress response that confers yeasts with resistance to organic acid food preservatives. Microbiology, 147, 2635-42.
PIPER, P., MAHE, Y., THOMPSON, S., PANDJAITAN, R., HOLYOAK, C., EGNER, R., MUHLBAUER, M., COOTE, P. & KUCHLER, K. 1998. The pdr12 ABC transporter is required for the development of weak organic acid resistance in yeast. EMBO J, 17, 4257-65.
PIPER, P. W. 1995. The heat shock and ethanol stress responses of yeast exhibit extensive similarity and functional overlap. FEMS Microbiol Lett, 134, 121-7.
PRASAD, R., PANWAR, S. L. & SMRITI 2002. Drug resistance in yeasts--an emerging scenario. Adv Microb Physiol, 46, 155-201.
RAJOKA, M. I., FERHAN, M. & KHALID, A. M. 2005. Kinetics and thermodynamics of ethanol production by a thermotolerant mutant of Saccharomyces cerevisiae in a microprocessor-controlled bioreactor. Lett Appl Microbiol, 40, 316-21.
RAMOS, M. T., MADEIRA-LOPES, A. 1990. Effects of acetic acid on the temperature profile of ethanol tolerance in Saccharomyces cerevisiae. Biotechnology Letters, 12, 229-234.
ROCKWELL, N. C., WOLFGER, H., KUCHLER, K. & THORNER, J. 2009. ABC transporter Pdr10 regulates the membrane microenvironment of Pdr12 in Saccharomyces cerevisiae. J Membr Biol, 229, 27-52.
ROSA, M. F., SÁ-CORREIA, I. 1994. Limitations to the use of extracellular acidification for the assessment of plasma membrane H+-ATPase activity and ethanol tolerance in yeasts. Enzyme Microb Technol, 16, 5.
SA-CORREIA, I., DOS SANTOS, S. C., TEIXEIRA, M. C., CABRITO, T. R. & MIRA, N. P. 2009. Drug:H+ antiporters in chemical stress response in yeast. Trends Microbiol, 17, 22-31.

38
SA-CORREIA, I. & TENREIRO, S. 2002. The multidrug resistance transporters of the major facilitator superfamily, 6 years after disclosure of Saccharomyces cerevisiae genome sequence. J Biotechnol, 98, 215-26.
SALGUEIRO, S. P., SA-CORREIA, I. & NOVAIS, J. M. 1988. Ethanol-Induced Leakage in Saccharomyces cerevisiae: Kinetics and Relationship to Yeast Ethanol Tolerance and Alcohol Fermentation Productivity. Appl Environ Microbiol, 54, 903-9.
SCHULLER, C., MAMNUN, Y. M., WOLFGER, H., ROCKWELL, N., THORNER, J. & KUCHLER, K. 2007. Membrane-active compounds activate the transcription factors Pdr1 and Pdr3 connecting pleiotropic drug resistance and membrane lipid homeostasis in saccharomyces cerevisiae. Mol Biol Cell, 18, 4932-44.
SERRANO, R., KIELLAND-BRANDT, M. C. & FINK, G. R. 1986. Yeast plasma membrane ATPase is essential for growth and has homology with (Na+ + K+), K+- and Ca2+-ATPases. Nature, 319, 689-93.
SIMOES-MENDES, B., MADEIRA-LOPES, A. & VAN UDEN, N. 1978. Kinetics of petite mutation and thermal death in Saccharomyces cerevisiae growing at superoptimal temperatures. Z Allg Mikrobiol, 18, 275-9.
SIMOES, T., MIRA, N. P., FERNANDES, A. R. & SA-CORREIA, I. 2006. The SPI1 gene, encoding a glycosylphosphatidylinositol-anchored cell wall protein, plays a prominent role in the development of yeast resistance to lipophilic weak-acid food preservatives. Appl Environ Microbiol, 72, 7168-75.
STRATFORD, M., STEELS, H., NEBE-VON-CARON, G., NOVODVORSKA, M., HAYER, K. & ARCHER, D. B. 2013. Extreme resistance to weak-acid preservatives in the spoilage yeast Zygosaccharomyces bailii. Int J Food Microbiol, 166, 126-34.
SUUTARI, M., LIUKKONEN, K. & LAAKSO, S. 1990. Temperature adaptation in yeasts: the role of fatty acids. J Gen Microbiol, 136, 1469-74.
SWAN, T. M. & WATSON, K. 1998. Stress tolerance in a yeast sterol auxotroph: role of ergosterol, heat shock proteins and trehalose. FEMS Microbiol Lett, 169, 191-7.
TEIXEIRA, M. C., CABRITO, T. R., HANIF, Z. M., VARGAS, R. C., TENREIRO, S. & SA-CORREIA, I. 2011. Yeast response and tolerance to polyamine toxicity involving the drug : H+ antiporter Qdr3 and the transcription factors Yap1 and Gcn4. Microbiology, 157, 945-56.
TEIXEIRA, M. C., GODINHO, C. P., CABRITO, T. R., MIRA, N. P. & SA-CORREIA, I. 2012. Increased expression of the yeast multidrug resistance ABC transporter Pdr18 leads to increased

39
ethanol tolerance and ethanol production in high gravity alcoholic fermentation. Microb Cell Fact, 11, 98.
TEIXEIRA, M. C., SANTOS, P. M., FERNANDES, A. R. & SA-CORREIA, I. 2005. A proteome analysis of the yeast response to the herbicide 2,4-dichlorophenoxyacetic acid. Proteomics, 5, 1889-901.
TURK, M., PLEMENITAS, A. & GUNDE-CIMERMAN, N. 2011. Extremophilic yeasts: plasma-membrane fluidity as determinant of stress tolerance. Fungal Biol, 115, 950-8.
ULLAH, A., ORIJ, R., BRUL, S. & SMITS, G. J. 2012. Quantitative analysis of the modes of growth inhibition by weak organic acids in Saccharomyces cerevisiae. Appl Environ Microbiol, 78, 8377-87.
VAN UDEN, N. 1984. Temperature profiles of yeasts. Adv Microb Physiol, 25, 195-251.
VAN UDEN, N. & MADEIRA-LOPES, A. 1970. Concurrent exponential growth and death of cell populations of Saccharomyces cerevisiae at superoptimal growth temperatures. Z Allg Mikrobiol, 10, 515-26.
VAN UDEN, N. & MADEIRA-LOPES, A. 1976. Yield and maintenance relations of yeast growth in the chemostat at superoptimal temperatures. Biotechnol Bioeng, 18, 791-804.
VERGHESE, J., ABRAMS, J., WANG, Y. & MORANO, K. A. 2012. Biology of the heat shock response and protein chaperones: budding yeast (Saccharomyces cerevisiae) as a model system. Microbiol Mol Biol Rev, 76, 115-58.
VIEGAS, C. A., CABRAL, M. G., TEIXEIRA, M. C., NEUMANN, G., HEIPIEPER, H. J. & SA-CORREIA, I. 2005. Yeast adaptation to 2,4-dichlorophenoxyacetic acid involves increased membrane fatty acid saturation degree and decreased OLE1 transcription. Biochem Biophys Res Commun, 330, 271-8.
VIEGAS, C. A. & SA-CORREIA, I. 1991. Activation of plasma membrane ATPase of Saccharomyces cerevisiae by octanoic acid. J Gen Microbiol, 137, 645-51.
VIEGAS, C. A., SEBASTIAO, P. B., NUNES, A. G. & SA-CORREIA, I. 1995. Activation of plasma membrane H(+)-ATPase and expression of PMA1 and PMA2 genes in Saccharomyces cerevisiae cells grown at supraoptimal temperatures. Appl Environ Microbiol, 61, 1904-9.
WENDLER, F., BERGLER, H., PRUTEJ, K., JUNGWIRTH, H., ZISSER, G., KUCHLER, K. & HOGENAUER, G. 1997. Diazaborine resistance in the yeast Saccharomyces cerevisiae reveals a link between YAP1 and the pleiotropic drug resistance genes PDR1 and PDR3. J Biol Chem, 272, 27091-8.

40
WIMALASENA, T. T., GREETHAM, D., MARVIN, M. E., LITI, G., CHANDELIA, Y., HART, A., LOUIS, E. J., PHISTER, T. G., TUCKER, G. A. & SMART, K. A. 2014. Phenotypic characterisation of Saccharomyces spp. yeast for tolerance to stresses encountered during fermentation of lignocellulosic residues to produce bioethanol. Microb Cell Fact, 13, 47.
YAMAMOTO, N., MAEDA, Y., IKEDA, A. & SAKURAI, H. 2008. Regulation of thermotolerance by stress-induced transcription factors in Saccharomyces cerevisiae. Eukaryot Cell, 7, 783-90.
ZHANG, X., CUI, Z., MIYAKAWA, T. & MOYE-ROWLEY, W. S. 2001. Cross-talk between transcriptional regulators of multidrug resistance in Saccharomyces cerevisiae. J Biol Chem, 276, 8812-9.
ZHAO, J., WANG, Z., WANG, M., HE, Q. & ZHANG, H. 2008. The inhibition of Saccharomyces cerevisiae cells by acetic acid quantified by electrochemistry and fluorescence. Bioelectrochemistry, 72, 117-21.
ZINSER, E., PALTAUF, F. & DAUM, G. 1993. Sterol composition of yeast organelle membranes and subcellular distribution of enzymes involved in sterol metabolism. J Bacteriol, 175, 2853-8.

41
Annexes

42
6. Annex I
Acetic acid concentrations required to inhibit growth of Saccharomyces cerevisiae
parental strain BY4741 and the derived deletion mutant strain Δpdr18 incubated at
temperatures in the range 30-40°C.
Figure 6.1 - Culture OD600nm of Saccharomyces cerevisiae parental strain BY4741 and derived
deletion mutant Δpdr18 grown at 30°C in the absence and in the presence of acetic acid. Cell
suspensions of BY4741 and Δpdr18 grown at 30°C were used to inoculate MM4 liquid medium (pH
4.0) supplemented or not with increasing concentrations of acetic acid (50 – 120 mM). Growth at 30°C
was followed during 240 hours.

43
Figure 6.2 - Culture OD600nm of Saccharomyces cerevisiae parental strain BY4741 and derived
deletion mutant Δpdr18 grown at 36°C in the absence and in the presence of acetic acid. Cell
suspensions of BY4741 and Δpdr18 grown at 30°C were used to inoculate MM4 liquid medium (pH
4.0) supplemented or not with increasing concentrations of acetic acid (50 – 120 mM). Growth at 36°C
was followed during 240 hours.

44
Figure 6.3 - Culture OD600nm of Saccharomyces cerevisiae parental strain BY4741 and derived
deletion mutant Δpdr18 grown at 38°C in the absence and in the presence of acetic acid. Cell
suspensions of BY4741 and Δpdr18 grown at 30°C were used to inoculate MM4 liquid medium (pH
4.0) supplemented or not with increasing concentrations of acetic acid (50 – 110 mM). Growth at 38°C
was followed during 240 hours.

45
Figure 6.4 - Culture OD600nm of Saccharomyces cerevisiae parental strain BY4741 and derived
deletion mutant Δpdr18 grown at 40°C in the absence and in the presence of acetic acid. Cell
suspensions of BY4741 and Δpdr18 grown at 30°C were used to inoculate MM4 liquid medium (pH
4.0) supplemented or not with increasing concentrations of acetic acid (35 – 90 mM). Growth at 40°C
was followed during 240 hours.

46
7. Annex II
Acetic acid concentrations required to inhibit growth at 30°C of Saccharomyces cerevisiae
parental strain BY4741 and the derived deletion mutant strain Δpdr18 harbouring a PDR18
expression plasmid or the corresponding cloning vector pRS416.
Figure 7.1 - Culture OD600nm of Saccharomyces cerevisiae parental strain BY4741
harbouring a PDR18 expression plasmid or the corresponding cloning vector pRS416 in
the absence and in the presence of acetic acid. Cell suspensions grown at 30°C were used
to inoculate MM4 liquid medium (pH 4.0) supplemented or not with increasing concentrations
of acetic acid (30 – 110 mM). Growth at 30°C was followed during 240 hours
47
Figure 7.2 - Culture OD600nm of Saccharomyces cerevisiae derived deletion mutant Δpdr18
harbouring a PDR18 expression plasmid or the corresponding cloning vector pRS416 in the
absence and in the presence of acetic acid. Cell suspensions grown at 30°C were used to inoculate
MM4 liquid medium (pH 4.0) supplemented or not with increasing concentrations of acetic acid
(30 – 110 mM). Growth at 30°C was followed during 240 hours.

48
8. Annex III
Tolerance of Saccharomyces cerevisiae parental strain BY4741 and derived deletion
mutant Δpdr18 to acetic acid.
Figure 8.1 - Conjugated effect of acetic acid and temperature in the growth of Saccharomyces
cerevisiae parental strain BY4741 and derived deletion mutant Δpdr18. Cell suspensions of
BY4741 and Δpdr18 grown at 30°C were used to inoculate MM4 liquid medium (pH 4.0) supplemented
or not with acetic acid. Minimum inhibitory concentration (MIC) values attained are shown during 240
hours of incubation at 30°C, 36°C, 38°C and 40°C in minimal medium supplemented with different
acetic acid concentrations (35 - 120 mM, at pH 4.0). Values presented in Y axis are the lowest
concentration (mM) of acetic acid to completely inhibit growth.