studies on the methane fermentation · studies on the methane fermentation ... n-butanol,...
TRANSCRIPT

STUDIES ON THE METHANE FERMENTATION
V. BIOCHEMICAL ACTIVITIES OF METHANOBACTERIUM OMELIANSKII
BY H. A. BARKER
(From the Division of Plant Nutrition of the College of Agriculture, University of California, Berkeley)
(Received for publication, April 20, 1940)
The general equation approximately describing the reactions by which methane-producing bacteria obtain their energi is
4HzA + COz -+ 4A + CHd + 2HzO (1)
where H,A represents an oxidizable compound, and A its oxidation product (1, 2). In this paper we shall consider principally three questions. (a) What oxidizable compounds can be used by Methanobacterium omelianskii and what transformations do they undergo? (b) What substances, if any, can replace COZ as an oxidizing agent? (c) How much of the substrate carbon is converted into cell material and what is its source, CO2 or H2A?
Oxidation Reactions
Qualitative Experiments-The ability of Methanobacterium omelianskii to oxidize various organic compounds has been deter- mined by growth experiments with the following medium (Medium R) made up with tap water, oxidizable compound 0.3 to 1.0 per cent, KzHPOl 0.5, KHzPOk 0.7, (NH&S04 0.03, MgS04.7Hz0 0.01, FeSOd.7Hz0 0.001, and 2 per cent by volume of a saturated CaSOp solution. Immediately after autoclaving 1.0 to 1.5 cc. of a 1 per cent NaS -9HzO solution and 5 to 8 cc. of a 5 per cent Na2C03 solution are added per 100 cc. The final pH is 7.4 to 7.7. All cultures were incubated under strictly anaerobic con- ditions at 35-38’. The details of the cultural technique and a description of the pure culture used are given elsewhere (3).
153
by guest on June 18, 2018http://w
ww
.jbc.org/D
ownloaded from

154 Methane Fermentation. V
The experiments show that the only types of compounds uti- lized by Methanobacterium omelianskii are simple primary and secondary alcohols. Ethanol, n-propanol, isopropanol, n-butanol, isobutanol, secondary butanol, and n-pentanol are oxidized. It is probable that higher alcohols of the same types can also be oxidized, though they have not been tested. Methanol and terti- ary butanol are not attacked, nor are any of the other common organic substrates tested. These include formate, acetate, propionate, n-butyrate, n-valerate, lactate, malate, succinate, glucose, glycerol, mannitol, 2,3-butylene glycol, yeast autol- ysate, and tryptone. It is particularly noteworthy that neither the fatty acids nor glucose is decomposed.
It should be mentioned here that although media containing formic acid as the sole oxidizable constituent cannot support the growth of 2Methanobacterium omelianskii, formic acid can under certain conditions be decomposed. The decomposition of formic acid will be discussed further below.
The products of the oxidation of primary alcohols are the corresponding fatty acids. Acetic acid formed by the oxidation of ethanol was identified by the sodium uranyl acetate micro- chemical reaction and was shown to be pure acetic acid by Du- claux distillation. The other volatile acids (propionic, n-butyric, isobutyric, and valeric) were both identified and shown to be 95 to 100 per cent pure by the Duclaux method.
The secondary alcohols, isopropanol and isobutanol, are oxidized to the corresponding ketones, acetone and ethyl methyl ketone. These ketones were identified by the melting points of their 2,4-dinitrophenylhydrazones.
The quantities of the various oxidation products were in all instances fairly large. Although in this particular set of experi- ments accurate determinations of the quantities of alcohol oxidized and of products formed were not made, the two were undoubtedly of the same order of magnitude.
Oxidation of Ethanol. Quantitative Experiments-It has been shown above that acetic acid is the only volatile acid formed from ethanol. This does not exclude the possibility that other prod- ucts such as carbon dioxide are also formed. Although a com- plete oxidation of ethanol appeared improbable in view of experi- ments previously carried out with enrichment cultures (l), still
by guest on June 18, 2018http://w
ww
.jbc.org/D
ownloaded from

H. A. Barker 155
it seemed highly desirable to check the point experimentally with a pure culture. Quantitative carbon balance experiments were therefore undertaken.
The fermentations were carried out in all-glass vessels contain- ing about 250 cc. of Medium B. Incubation was at 37’; oxygen was removed by oxsorbent, an acid chromous chloride preparation. The evolved gases were collected over mercury. Samples of the medium were removed for analysis immediately after inoculation and after the fermentation had ceased (5 to 10 days). Ethanol was separated by distillation and estimated by Northrop’s method (4). Acetic acid was determined by direct steam distillation and titration after suitable corrections for carbon dioxide and in- complete recovery were applied. Dissolved carbon dioxide was estimated by the Van Slyke manometric technique (5); gaseous carbon dioxide and methane were collected over mercury and were determined by ordinary methods of gas analysis. Dissolved methane was calculated from solubility data.
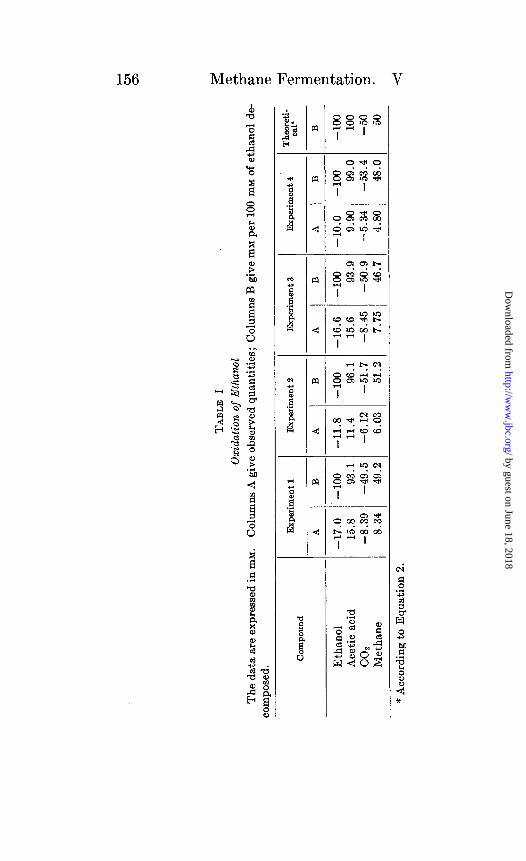
The results of four such experiments are presented in Table I. The experiments differ with respect to the substance limiting the catabolic reaction. In Experiments 1, 2, and 3, ethanol was present in excess; the quantity of carbon dioxide therefore de- termined the amount of alcohol oxidized. In Experiment 4 carbon dioxide was present in excess and ethanol was limiting.
Experiments 1,2, and 3 of Table I show that acetic acid accounts for 93 to 96 per cent of the ethanol oxidized. The discrepancy between the ethanol oxidized and acetic acid formed cannot be due to a production of carbon dioxide, since if this occurred the net disappearance of carbon dioxide would be decreased. No such effect is evident; the uptake of carbon dioxide is if anything slightly greater than is required by Equation 2. It will be shown below that the discrepancy between ethanol and acetic acid is caused by conversion of part of the alcohol into cell material.
Since in Experiments 1, 2, and 3 the oxidizing agent, carbon dioxide, was limiting while ethanol was present in excess, it is not surprizing that the oxidation of the latter compound was incom- plete. A much more rigorous test of the ability of Methanobacte- rium omelianskii to oxidize ethanol to carbon dioxide is provided by Experiment 4 in which carbon dioxide was present in excess while ethanol was limiting. Even under these conditions, how-
by guest on June 18, 2018http://w
ww
.jbc.org/D
ownloaded from
TABL
E I
Oxid
atio
n of
Eth
anol
Th
e da
ta
are
expr
esse
d in
rnw
Co
lum
ns
A gi
ve
obse
rved
qu
antit
ies;
Co
lum
ns
B gi
ve
mM
per
10
0 m
M
of e
than
ol
de-
kc
com
pose
d.
Exwr
imen
t 1
Expe
rimen
t 2
Comp
ound
A B
A __
____
Etha
nol
-17.
0 -1
00
-11.
8 Ac
etic
acid
15
.8
93.1
11
.4
co2
-8.3
9 -4
9.5
-6.1
2 M
etha
ne
8.34
49
.2
6.03
* Ac
cord
ing
to
Equa
tion
2.
B
Expe
rimen
t 3
Expe
rimen
t 4
1 B
A 1
B A
- 10
0 -1
6.6
96.1
15
.6
-51.
7 -8
.45
51.2
7.
75
-___
-1
00
-10.
0 -
100
-100
$
93.9
9.
90
99.0
10
0 -5
0.9
-5.3
4 -5
3.4
-50
g r 46
.7
4.80
48
.0
50
* -4
,,,,,/
,,,~,
, ,,,
,, ,,,
,,,
.,
by guest on June 18, 2018http://w
ww
.jbc.org/D
ownloaded from

H. A. Barker 157
ever, the organism appears to be unable to oxidize ethanol beyond the stage of acetic acid.
From the above experiments it may be concluded that the over- all catabolic reaction carried out by Methanobacterium omelianslcii is closely described by the equation,
2CeHsOH + COz + BCH&OOH + CHa (2)
This is in agreement with the conclusion reached previously (1) from experiments with enrichment cultures.
Oxidation of B&anol--In order to see whether the oxidation of butanol goes beyond the stage of butyric acid, a quantitative fermentation experiment was undertaken in which butanol was limiting, carbon dioxide being present in excess. The medium and conditions were essentially the same as in the experiments described above except that butanol (0.5 volume per cent) was substituted for ethanol. The fermentation of butanol being much slower than the fermentation of ethanol, the culture (1000 cc.) was allowed to incubate for 21 days at 37”. At the end of this period gas production had practically ceased.
The analytical data showed that butyric acid (17.0 mM) ac- counts for 97 per cent of the butanol decomposed (17.5 mM). It may be concluded that the oxidation of butyric acid to 2 molecules of acetic acid, previously thought on the basis of experi- ments with enrichment cultures to be brought about by Methano- bacterium omelianskii (l), is probably due to another organism.
Quantitative fermentation experiments with other alcohols have not been undertaken. It may be expected, however, on the basis of the qualitative results reported above that higher alcohols are in general oxidized (without regard to ‘synthetic reactions) only as far as the corresponding fatty acids or ketones.
Reduction Reactions
The second main problem undertaken was to find out what compounds, if any, can replace carbon dioxide as an oxidizing agent for Methanobacterium omelianskii. The compounds that have been tested are methyl alcohol, formic acid, nitrate, sulfate, and oxygen.
A qualitative cxperimcnt to test the reducibility of the above compounds under normal physiological conditions was carried
by guest on June 18, 2018http://w
ww
.jbc.org/D
ownloaded from

158 Methane Fermentation. V
out with growing cultures. To Medium I3 containing an excess of ethanol as the oxidizable constituent, a small, limiting amount of carbon dioxide and 0.3 per cent of the various compounds to be tested (except oxygen) were added. Oxygen was supplied by exposing the cultures (test-tubes) to air; all other cultures were incubated under strictly anaerobic conditions. When all activity had ceased, the media were analyzed for acetic acid. It was expected that any substance acting as an oxidizing agent would increase the yield of acetic acid over that amount equivalent to the carbon dioxide available.
The results showed that oxygen and nitrate are toxic, since no acid production occurred in their presence. Sulfate and methanol appear to be completely inert, since they did not significantly in- crease or decrease the formation of acetic acid as compared with the control. Formate is the only substance utilized. Not only was the amount of acetic acid approximately doubled in the formate cultures but formate had almost completely disappeared.
Decomposition of Formic Acid-The decomposition of formic acid in the above experiment may appear to contradict earlier results (see above) which indicated that formic acid is not attacked. The contradiction, however, is only apparent, for the conditions of the two experiments were quite different. In the former formic acid was the sole oxidizable constituent of the medium, while in the latter ethanol was present as well.
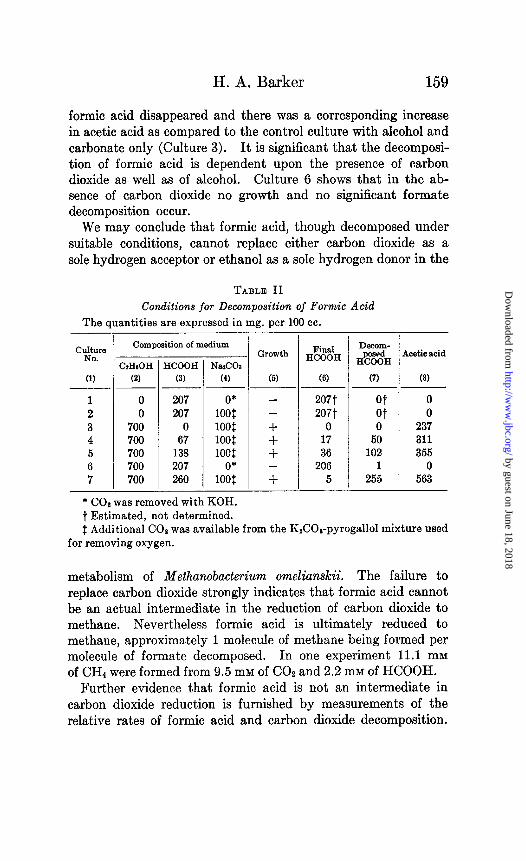
In order further to clarify the conditions necessary for the decomposition of formic acid, the growth experiment summarized in Table II was set up. The essential features of the media used are given in Columns 2, 3, and 4. Column 5 shows whether or not growth occurred during 20 days incubation. The remaining columns are self-explanatory. The data given are averages of duplicate experiments.
The data show that Methanobacterium omelianskii cannot live with formic acid alone (Culture 1) or with formic acid plus carbon dioxide (Culture 2). In the absence of ethanol no growth oc- curred. These results corroborate the conclusion reached pre- viously that formic acid cannot replace ethanol as the oxidizable substrate. Cultures 4, 5, and 7, all of which contain ethanol, carbonate, and formate, show, however, that formic acid can be decomposed. In all three cultures considerable quantities of
by guest on June 18, 2018http://w
ww
.jbc.org/D
ownloaded from
H. A. Barker 159
formic acid disappeared and there was a corresponding increase in acetic acid as compared to the control culture with alcohol and carbonate only (Culture 3). It is significant that the decomposi- tion of formic acid is dependent upon the presence of carbon dioxide as well as of alcohol. Culture 6 shows that in the ab- sence of carbon dioxide no growth and no significant formate decomposition occur.
We may conclude that formic acid, though decomposed under suitable conditions, cannot replace either carbon dioxide as a sole hydrogen acceptor or ethanol as a sole hydrogen donor in the
yEz HCOOH (7)
TABLE II Conditions for Decomposition of Formic Acid
The quantities are expressed in mg. per 100 cc.
%P Composition of medium
Growth Final HCOOH
C,HrOH HCOOH NaEOs
(1) (2) (3) (4) (5) (6) ----~~
1 0 207 o* - 207t 2 0 207 1001 - 207t 3 700 0 1001 + 0 4 700 67 lOO$ + 17 5 700 138 1003 + 36 6 700 207 o* - 206 7 700 260 1001 + 5
* GOa was removed with KOH. t Estimated, not determined.
Acetic acid
(8)
ot 0 ot 0 0 237
50 311 102 355
1 0 255 563
$ Additional CO1 was available from the K&OS-pyrogallol mixture used for removing oxygen.
metabolism of Methanobacterium omelianskii. The failure to replace carbon dioxide strongly indicates that formic acid cannot be an actual intermediate in the reduction of carbon dioxide to methane. Nevertheless formic acid is ultimately reduced to methane, approximately 1 molecule of methane being formed per molecule of formate decomposed. In one experiment 11.1 mM of CH4 were formed from 9.5 mM of CO2 and 2.2 mM of HCOOH.
Further evidence that formic acid is not an intermediate in carbon dioxide reduction is furnished by measurements of the relative rates of formic acid and carbon dioxide decomposition.
by guest on June 18, 2018http://w
ww
.jbc.org/D
ownloaded from
160 Blcthane k‘e.rmentation. V
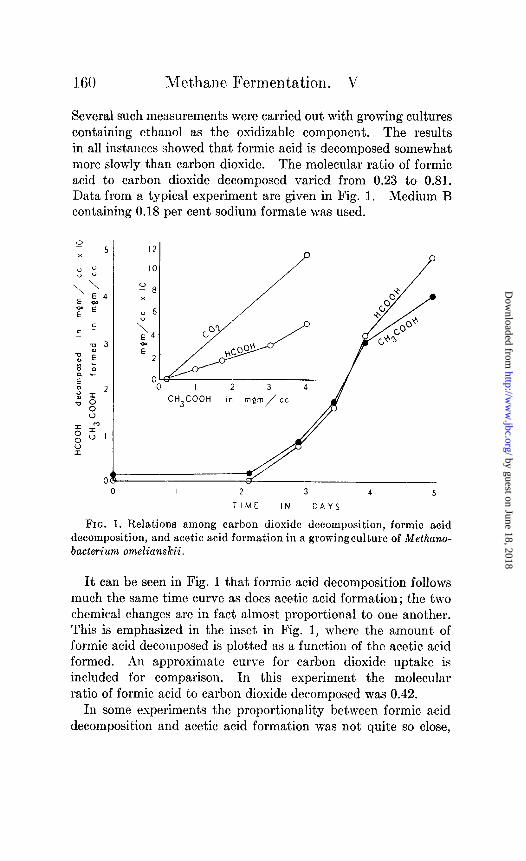
Several such measurements were carried out with growing cultures containing ethanol as the oxidizable component. The results in all instances showed that formic acid is decomposed somewhat more slowly than carbon dioxide. The molecular ratio of formic acid to carbon dioxide decomposed varied from 0.23 to 0.81. Data from a typical experiment are given in Fig. 1. Medium I3 containing 0.18 per cent sodium formate was used.
IO
CH3COOH in mgm/cc
0 0 I 2 3 4 5
TIME IN DAYS
FIG. 1. Relations among carbon dioxide decomposition, formic acid decomposition, and acetic acid formation in a growingculture of Methano- bacterium omelianskii.
It can be seen in Fig. 1 that formic acid decomposition follows much the same time curve as does acetic acid formation; t,he two chemical changes are in fact almost proportional t,o one another. This is emphasized in the inset in Fig. 1, where the amount of formic acid decomposed is plotted as a function of the acetic acid formed. An approximat,e curve for carbon dioxide uptake is included for comparison. In this experiment the molecular ratio of formic acid to carbon dioxide decomposed was 0.42.
In some experiments the proportionality between formic acid decomposition and acetic acid formation was not quite so close,
by guest on June 18, 2018http://w
ww
.jbc.org/D
ownloaded from

H. A. Uarker 161
the formate tending to decompose relatively more rapidly in the later stages of the fermentation.
It should be emphasized that although the above experiments show that formic acid can be converted into methane, they do not provide any evidence as to the nature of the primary reaction which formate undergoes. This could be either an oxidation or a reduction. The quest,ion of the mechanism of formate decomposi- tion will be considered in a later communication.
Toxicity of Nitrate-It has already been mentioned that the development of Methanobacterium omelianskii is prevented by 0.3 per cent potassium nitrate. In order to confirm this result and to find the lowest concentration of nitrate that will just inhibit growth, two further experiments were carried out.
In t,hc first experiment, the following KN03 concentrations were added to Medium B, 0, 0.05, 0.10, 0.20, and 0.4 per cent. Rapid and abundant growth occurred without nitrate. With nitrate, inhibition was very marked at 0.05 per cent and was almost complete at 0.10 per cent’. At higher concentrations there was no development after 7 days.
A second experiment showed that the limiting KNO, concentra- tion for growth is about 0.08 to 0.10 per cent. Below 0.02 per cent nitrate no inhibition was observed.
Assimilation of Carbon
It has been shown that the oxidation of ethanol by Methano- bacterium omelianskii may be represented approximately by Equation 2. As has previously been mentioned, this equation is not an exact description of the over-all reaction carried out by this organism because the synthesis of cell material is disregarded. From the point of view of the vital activities of the organism, the synthesis of cell material is naturally of the first importance.
Because of the physiological peculiarities of the methane bacteria, their synthetic or assimilatory activities have a special interest. Microorganisms are commonly divided into two large physiological groups on the basis of the types of carbon compound from which they build their cell material. One group, including the chemoautotrophic and photoautotrophic bacteria, synthesizes all cell materials from carbon dioxide. The other group, compris-
by guest on June 18, 2018http://w
ww
.jbc.org/D
ownloaded from

162 Methane Fermentation. V
ing all chemoheterotrophic organisms, appears’ to assimilate mainly the carbon of organic compounds. There is also a third group of organisms, the photoheterotrophic bacteria (Athior- hodaceae and Thiorhodaceae), that probably assimilates directly2 large amounts of both organic carbon.and carbon dioxide carbon. Now the methane-producing bacteria, like the photoheterotrophic bacteria, combine certain characteristics of both heterotrophic and autotrophic metabolism in that they are able both to oxidize organic compounds and to reduce carbon dioxide. It must be emphasized at once, however, that the ability to utilize both organic compounds and carbon dioxide is not in itself any proof that these bacteria assimilate carbon from both sources. It is quite possible that only one of the two substances serves as a source of cell material, the other fulfilling some other essential function, such as acting as an oxidizing or reducing agent.
In the special case of Methanobacterium omelianskii there are at least three possibilities for the synthesis of cell material. The carbon for cell synthesis might come from ethanol alone, from carbon dioxide alone, or from both compounds.
Before the evidence in favor of one or the other of these sources of carbon is considered, it will be desirable to present data showing the magnitude of cell synthesis, and the relation between synthesis and the dissimilation reaction expressed by Equation 2.
To determine the quantity of cell synthesis, Medium 33, con- taining ammonium sulfate as the sole nitrogen source, was used. As growth occurs in this medium, ammonia nitrogen is converted into organic nitrogen in the cells. This cell nitrogen can be meas- ured as the difference between total (Kjeldahl) nitrogen and am- monia nitrogen. The nitrogen determinations were carried out directly on 10 cc. aliquots of the fermented culture medium by micromethods. Suitable blank corrections were made on the reagents and the culture medium.
Several series of cell nitrogen determinations were carried out. In each series the amount of bacterial growth in individual cultures
1 Recent work by Wood and Werkman (6) and others indicates that car- bon dioxide may be utilized by various heterotrophic bacteria. Ruben et al. (7, 8) have proved with the aid of radioactive carbon that small amounts of CO2 a.re assimilated by many heterotrophic organisms.
* Private communication from Professor C. B. van Niel.
by guest on June 18, 2018http://w
ww
.jbc.org/D
ownloaded from
H. A. Barker 163
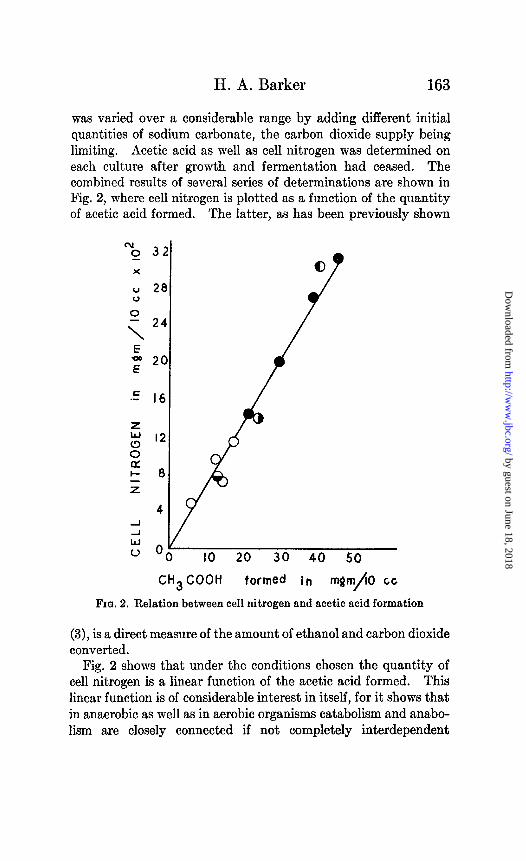
was varied over a considerable range by adding different initial quantities of sodium carbonate, the carbon dioxide supply being limiting. Acetic acid as well as cell nitrogen was determined on each culture after growth and fermentation had ceased. The combined results of several series of determinations are shown in Fig. 2, where cell nitrogen is plotted as a function of the quantity of acetic acid formed. The latter, as has been previously shown
“g 32 X
v 28 u
’ \
24
E ; 20
2 16
Z
k-ii I2 0 cc c 8 z
4 i ooi IO 20 30 40 50
CH3 COOH formed in mgm/iO cc.
FIG. 2. Relation between cell nitrogen and acetic acid formation
(3), is a direct measure of the amount of ethanol and carbon dioxide converted.
Fig. 2 shows that under the conditions chosen the quantity of cell nitrogen is a linear function of the acetic acid formed. This linear function is of considerable interest in itself, for it shows that in anaerobic as well as in aerobic organisms catabolism and anabo- lism are closely connected if not completely interdependent
by guest on June 18, 2018http://w
ww
.jbc.org/D
ownloaded from

164 Methane Fermentation. V
processes. From the slope of the line it can be calculated that 6.75 mg. of cell nitrogen are formed per gm. of acetic acid. Since the acetic acid in such a fermentation accounts for only about 94 per cent of the alcohol oxidized (Table I), 8.2 mg. of cell nitrogen are formed per gm. of ethanol decomposed.
From the above data on nitrogen assimilation, the quantities of carbon assimilated can be calculated provided the ratio of carbon to nitrogen in the bacterial cells is known. This ratio has been estimated approximately by determining the nitrogen con- tent of cells grown under conditions comparable to those existing in the above experiments. The nitrogen content was found to be 10.6 per cent on a dry weight, ash-free basis. Assuming a carbon content of 50 per cent (which can hardly differ by more than 2 to 3 per cent from the true value), the C:N ratio would be 4.7. The carbon assimilated is therefore 38.6 mg. per gm. of ethanol decomposed, 74 mg. per gm. of ethanol carbon, or 59 mg. per gm. of substrate (ethanol plus carbon dioxide) carbon.
Because of the rather large quantity of cell material formed, it appeared possible to decide by analytical methods which of the three sources of cell carbon mentioned above is actually utilized. For if the cell material originates exclusively from ethanol, the carbon of the recovered acetic acid should be approximately 7 per cent less than the carbon of the alcohol decomposed, while the carbon dioxide and methane carbons should balance. If, on the contrary, the cell material is derived only from carbon dioxide, the methane carbon should be about 28 per cent less than the carbon dioxide carbon, while the ethanol and acetic acid carbons should balance. Or, finally, if both ethanol and carbon dioxide contributed to the cell material, there should be a carbon dis- crepancy in both the oxidation and the reduction reactions.
In order to obtain significant evidence on the source of cell carbon by the method indicated, it was necessary to carry out analyses of a rather high order of accuracy. The over-all analyt- ical error should, if possible, not exceed 2 to 4 per cent, whereas in most analyses of the type required the over-all error is consider- ably greater, usually 4 to 8 per cent. Fortunately in the present case, the medium to be analyzed was sufficiently simple so that nearly the desired accuracy could be attained by giving careful attention to details of procedure.
by guest on June 18, 2018http://w
ww
.jbc.org/D
ownloaded from

H. A. Barker 165
The results of three carbon balance experiments will be pre- sented. Analytical data on the transformations of ethanol, acetic acid, carbon dioxide, and methane in these experiments have al- ready been given in Table I (Experiments 1, 2, and 3). The data show that the yield of acetic acid is always too low to account for the ethanol oxidized, whereas in two of the three experiments (Nos. 1 and 2) the quantity of methane corresponds very closely to the carbon dioxide reduced. In Table III the quantities of cell carbon formed are given and are compared with the differences between ethanol carbon and acetic acid carbon and between carbon dioxide carbon and methane carbon.
When the data given in Table III are evaluated, it must be remembered that the figures, except those for cell carbon, represent
TABLE III Carbon Assimilation by Methanobacterium omelianskii
Carbon fraction Experiment 1 Experiment 2 Experiment 3 -.-.~-.-__
wl. per cent mg. per cent mg. pet cen.t
Cell C.. 113 100 70 100 115 100
C&OH-C less HOAc-C. 110 97 41 59 95 82
Con-C less CHa-C 2 2 4 6 33 29 __ ~___~_______
Total cell C accounted for. 112 99 45 65 I 128 111 --.
small differences between values of a higher order of magnitude and are therefore subject to considerable errors. The data neverthe- less strongly indicate that the major portion of the cell carbon is derived from ethanol. In Experiments 1, 2, and 3, 97,59, and 82 per cent of the cell carbon, respectively, is accounted for as unrecovered ethanol carbon. The percentage of cell carbon ap- parently derived from carbon dioxide in Experiments 1 and 2 is very small, 2 and 6 per cent, while in Experiment 3 it is 29 per cent.
It seems reasonably safe to conclude from the results of these experiments that approximately 60 per cent or more of the cell carbon is derived from ethanol, while only 30 per cent or less is derived from carbon dioxide. In other words, cell carbon defi- nitely is not derived from carbon dioxide alone. The experiments are not sufficiently accurate to determine whether cell carbon
by guest on June 18, 2018http://w
ww
.jbc.org/D
ownloaded from

166 Methane Fermentation. V
originates exclusively from ethanol or mainly from ethanol and to a lesser extent from carbon dioxide.
Fortunately, a more sensitive method was available for de- termining whether or not carbon dioxide is assimilated by Methano- bacterium omelianskii; namely, a method involving the use of radioactive carbon. If suspensions of cells are provided with ethanol and radioactive carbon dioxide, it should be possible to find out whether and to what extent carbon dioxide is assimilated by measuring the radioactivities of the methane evolved and of the residual cell material. This has been done and it has been found (2) that a small part (about 1.5 per cent) of the carbon dioxide reduced is converted into a non-volatile substance associ- ated with the cells. The quantity of this non-volatile material is too small to have been detected with certainty in the analytical experiments described above. Though the percentage of carbon dioxide carbon converted into cells appears to be definitely lower than the percentage of ethanol carbon so converted, still the two are of approximately the same order of magnitude. The organism appears therefore to have no very decided preference as to which form of carbon it uses.
SUMMARY
1. The ability of a pure culture of Methanobacterium omelianskii to oxidize a number of organic compounds (with carbon dioxide as the ultimate oxidizing agent) has been tested. Of the compounds studied only simple primary and secondary alcohols are attacked, these being oxidized to the corresponding fatty acids and ketones, respectively. Carbon dioxide is not produced in detectable amounts in these oxidations but is, on the contrary, consumed in an amount required by Equation 1, 4HzA + CO2 -+ 4.4 + CH* + 2HzO.
2. When the behavior of the organism toward various possible oxidizing agents was examined, it was found that oxygen and nitrate are toxic, sulfate and methanol are inert, and only formate is decomposed. .
In growth experiments formate is only utilized when both ethanol and carbon dioxide are simultaneously present; i.e., formate cannot replace either ethanol as a hydrogen donor or carbon dioxide as a hydrogen acceptor. It is concluded that
by guest on June 18, 2018http://w
ww
.jbc.org/D
ownloaded from

H. A. Barker
formate is not a normal intermediate in the reduction of carbon dioxide to methane in spite of the fact that formate is converted into methane. The primary reaction of formate is not considered here.
3. A study of the assimilation of carbon by Methanobacterium omelianskii shows that there is a linear relation between the amount of ethanol oxidized to acetic acid and the amount of cell material formed. For each gm. of substrate (ethanol plus carbon dioxide) carbon converted about 60 mg. of carbon are assimilated. By means of carbon balance experiments and by the use of radioactive carbon dioxide it is shown that both ethanol carbon and carbon dioxide carbon are converted into cell material. Most of the cell carbon is derived from ethanol. But the percentage of carbon dioxide carbon converted into cells appears to be of the same order of magnitude as the percentage of ethanol carbon so converted.
Clerical assistance was furnished by the personnel of the Works Progress Administration, Official Project No. 65-l-08-91, Unit BlO.
BIBLIOGRAPHY
1. Barker, H. A., Arch. Mikrobiol., 7,404 (1936). 2. Barker, H. A., Ruben, S., and Knmen, M. D., Proc. Nat. Acad. SC., 26,
426 (1940). 3. Barker, H. A., Ant,onic van Leeuwcnhoek, in press. 4. Northrop, J. H., Ashe, L. H., and Senior, J. K., J. Biol. Chem., 39, 1
(1919). 5. Peters, J. P., and Van Slyke, D. D., Quantitative clinical chemistry,
Methods, Baltimore, 210 (1932). 6. Wood, I-1. C., and Werkman, C. H., Rep. Proc. 3rd Internat. Cong. Micro-
biol., New Pork, 246 (1940). 7. Ruben, S., and Kamcn, M. D., Proc. Nat. dead. SC., 26, 418 (1940). 8. Carson, S., and Ruben, S., Proc. Nat. Acad. SC., 26, 422 (1940).
by guest on June 18, 2018http://w
ww
.jbc.org/D
ownloaded from

H. A. BarkerOMELIANSKIIMETHANOBACTERIUM
ACTIVITIES OFFERMENTATION: V. BIOCHEMICAL
STUDIES ON THE METHANE
1941, 137:153-167.J. Biol. Chem.
http://www.jbc.org/content/137/1/153.citation
Access the most updated version of this article at
Alerts:
When a correction for this article is posted•
When this article is cited•
alerts to choose from all of JBC's e-mailClick here
tml#ref-list-1
http://www.jbc.org/content/137/1/153.citation.full.haccessed free atThis article cites 0 references, 0 of which can be by guest on June 18, 2018
http://ww
w.jbc.org/
Dow
nloaded from