supplementary information - media.nature.com · duf89 (3 µg) was added to one acp plus acph...
TRANSCRIPT

Supplementary Information
A diverse family of metal-dependent phosphatases implicated in metabolite
damage-control
Lili Huang1,9, Anna Khusnutdinova2,9, Boguslaw Nocek3, Greg Brown2, Xiaohui Xu2, Hong Cui2,
Pierre Petit2, Robert Flick2, Rémi Zallot4, Kelly Balmant5, Michael J. Ziemak6, John Shanklin7, Valérie
de Crécy-Lagard4, Oliver Fiehn8, Jesse F. Gregory III1, Andrzej Joachimiak3, Alexei Savchenko2,
Alexander F. Yakunin2* & Andrew D. Hanson6*
1Food Science and Human Nutrition Department, University of Florida, Gainesville, Florida, USA.
2Department of Chemical Engineering & Applied Chemistry, University of Toronto, Toronto, Canada.
3Midwest Center for Structural Genomics and Structural Biology Center, Biosciences Division,
Argonne National Laboratory, Argonne, Illinois, USA.
4Microbiology and Cell Science Department, University of Florida, Gainesville, Florida, USA.
5Plant Molecular and Cellular Biology Program, University of Florida, Gainesville, Florida, USA.
6Horticultural Sciences Department, University of Florida, Gainesville, Florida, USA.
7Biological, Environmental, and Climate Sciences Department, Brookhaven National Laboratory,
Upton, New York, USA.
8Metabolomics Core, UC Davis Genome Center, University of California Davis, California, USA.
9These authors contributed equally to this work.
*e-mail: [email protected] or [email protected]
Nature Chemical Biology: doi:10.1038/nchembio.2108
Supplementary Results
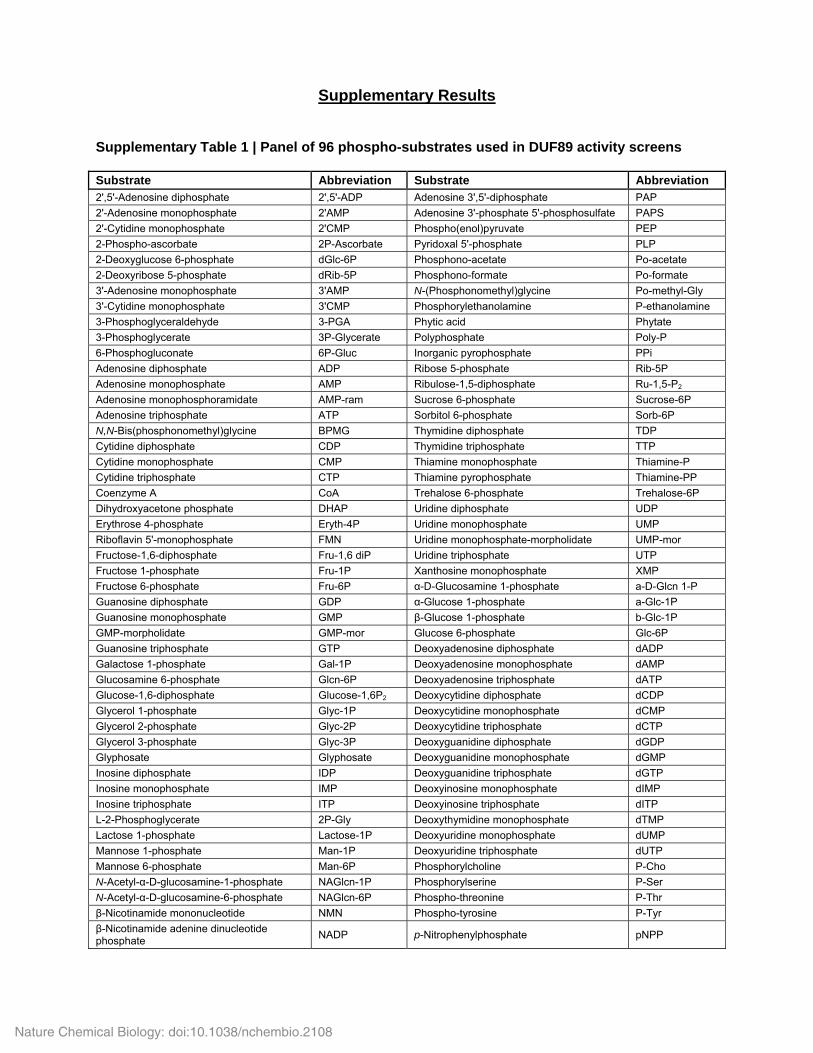
Supplementary Table 1 | Panel of 96 phospho-substrates used in DUF89 activity screens Substrate Abbreviation Substrate Abbreviation 2',5'-Adenosine diphosphate 2',5'-ADP Adenosine 3',5'-diphosphate PAP
2'-Adenosine monophosphate 2'AMP Adenosine 3'-phosphate 5'-phosphosulfate PAPS
2'-Cytidine monophosphate 2'CMP Phospho(enol)pyruvate PEP
2-Phospho-ascorbate 2P-Ascorbate Pyridoxal 5'-phosphate PLP
2-Deoxyglucose 6-phosphate dGlc-6P Phosphono-acetate Po-acetate
2-Deoxyribose 5-phosphate dRib-5P Phosphono-formate Po-formate
3'-Adenosine monophosphate 3'AMP N-(Phosphonomethyl)glycine Po-methyl-Gly
3'-Cytidine monophosphate 3'CMP Phosphorylethanolamine P-ethanolamine
3-Phosphoglyceraldehyde 3-PGA Phytic acid Phytate
3-Phosphoglycerate 3P-Glycerate Polyphosphate Poly-P
6-Phosphogluconate 6P-Gluc Inorganic pyrophosphate PPi
Adenosine diphosphate ADP Ribose 5-phosphate Rib-5P
Adenosine monophosphate AMP Ribulose-1,5-diphosphate Ru-1,5-P2
Adenosine monophosphoramidate AMP-ram Sucrose 6-phosphate Sucrose-6P
Adenosine triphosphate ATP Sorbitol 6-phosphate Sorb-6P
N,N-Bis(phosphonomethyl)glycine BPMG Thymidine diphosphate TDP
Cytidine diphosphate CDP Thymidine triphosphate TTP
Cytidine monophosphate CMP Thiamine monophosphate Thiamine-P
Cytidine triphosphate CTP Thiamine pyrophosphate Thiamine-PP
Coenzyme A CoA Trehalose 6-phosphate Trehalose-6P
Dihydroxyacetone phosphate DHAP Uridine diphosphate UDP
Erythrose 4-phosphate Eryth-4P Uridine monophosphate UMP
Riboflavin 5'-monophosphate FMN Uridine monophosphate-morpholidate UMP-mor
Fructose-1,6-diphosphate Fru-1,6 diP Uridine triphosphate UTP
Fructose 1-phosphate Fru-1P Xanthosine monophosphate XMP
Fructose 6-phosphate Fru-6P α-D-Glucosamine 1-phosphate a-D-Glcn 1-P
Guanosine diphosphate GDP α-Glucose 1-phosphate a-Glc-1P
Guanosine monophosphate GMP β-Glucose 1-phosphate b-Glc-1P
GMP-morpholidate GMP-mor Glucose 6-phosphate Glc-6P
Guanosine triphosphate GTP Deoxyadenosine diphosphate dADP
Galactose 1-phosphate Gal-1P Deoxyadenosine monophosphate dAMP
Glucosamine 6-phosphate Glcn-6P Deoxyadenosine triphosphate dATP
Glucose-1,6-diphosphate Glucose-1,6P2 Deoxycytidine diphosphate dCDP
Glycerol 1-phosphate Glyc-1P Deoxycytidine monophosphate dCMP
Glycerol 2-phosphate Glyc-2P Deoxycytidine triphosphate dCTP
Glycerol 3-phosphate Glyc-3P Deoxyguanidine diphosphate dGDP
Glyphosate Glyphosate Deoxyguanidine monophosphate dGMP
Inosine diphosphate IDP Deoxyguanidine triphosphate dGTP
Inosine monophosphate IMP Deoxyinosine monophosphate dIMP
Inosine triphosphate ITP Deoxyinosine triphosphate dITP
L-2-Phosphoglycerate 2P-Gly Deoxythymidine monophosphate dTMP
Lactose 1-phosphate Lactose-1P Deoxyuridine monophosphate dUMP
Mannose 1-phosphate Man-1P Deoxyuridine triphosphate dUTP
Mannose 6-phosphate Man-6P Phosphorylcholine P-Cho
N-Acetyl-α-D-glucosamine-1-phosphate NAGlcn-1P Phosphorylserine P-Ser
N-Acetyl-α-D-glucosamine-6-phosphate NAGlcn-6P Phospho-threonine P-Thr
β-Nicotinamide mononucleotide NMN Phospho-tyrosine P-Tyr
β-Nicotinamide adenine dinucleotide phosphate
NADP p-Nitrophenylphosphate pNPP
Nature Chemical Biology: doi:10.1038/nchembio.2108

Supplementary Table 2 | Kinetic parameters of wild type and mutant DUF89 proteins
Enzyme Substrate kcat
(s-1)
Km
(µM)
kcat/Km
(s-1 M-1)
Yeast YMR027W Fru-1P 11.60 0.38 310 40 3.7 104 Fru-6P 4.20 0.28 300 80 1.4 104 Man-6P 3.30 0.36 490 180 6.8 103 dRib-5P 1.90 0.14 280 80 6.7 103
Arabidopsis At2g17340 p-NPP 1.68 0.02 418 16 4.0 103
Rib-5Pa 0.94 0.02 (70 4)b -- dRib-5Pa 1.10 0.04 (90 8)b --
P. horikoshii PH1575 pNPP 1.00 0.02 110 20 9.5 103
Arabidopsis PanK2 pNPP 1.99 0.10 640 99 3.1 103
P-PaSH 0.26 0.02 18.9 3.1 1.3 104 P-PaSSO3 0.94 0.03 2.91 0.28 3.2 105 P-Pa 0.52 0.02 0.28 0.05 1.9 106
Human PanK4 pNPP 0.17 0.05 1920 189 8.9 101
P-PaSH 0.71 0.03 150 23 4.7 103 P-PaSO3 0.09 0.005 3.90 0.7 2.3 104
Assays contained 0.2 mM Ni2+ (Arabidopsis At2g17340), 0.5 mM Ni2+ (PH1575, Arabidopsis PanK2), 0.1 mM Co2+ (YMR027W), or 0.5 mM Co2+ (human PanK4). Results are means s.d. from four (YMR027W, At2g17340 and PH1575) or three (Arabidopsis PANK2 and Human PANK4) independ-ent determinations. pNPP, p-nitrophenyl phosphate. Other substrate abbreviations are given in Supplementary Table 1 and Figure 3b. aSigmoidal kinetics. bS0.5 values.
Nature Chemical Biology: doi:10.1038/nchembio.2108

Supplementary Table 3 | The Arabidopsis PanK2 DUF89 domain lacks acyl carrier protein phosphodiesterase activity
Treatment Added at 1.5 h
4'-Phosphopantetheine (pmol/assay)
Pantetheine (pmol/assay)
ACP only -- <1 <1
ACP + DUF89 -- <1 <1
ACP + AcpH -- 2131 92 <1
ACP + AcpH DUF89 2216 128 45 5
Assays (20 µl) contained ~2 nmol spinach holo-acyl carrier protein (ACP), with or without 3 µg Arab-idopsis PanK2 DUF89 domain or 1 µg E. coli acyl carrier protein phosphodiesterase (AcpH) as a positive control. The buffer was 50 mM Tris-HCl, pH 8.0 plus 1 mM Mg2+ and 0.1 mM Mn2+. Incub-ation was for 2 h at 30°C. DUF89 (3 µg) was added to one ACP plus AcpH treatment at 1.5 h to con-firm that the DUF89 preparation had the expected 4'-phosphopantetheine phosphatase activity. 4'-Phosphopantetheine and pantetheine were quantified as monobromobimane derivatives by fluoro-metric HPLC. Results are means s.d. of three replicates. Note that the Arabidopsis DUF89 domain released neither 4'-phosphopantetheine nor pantetheine from ACP, indicating that it does not cleave either of the phosphoester bonds in ACP.
Nature Chemical Biology: doi:10.1038/nchembio.2108

Supplementary Table 4 | Metal contents of DUF89 subfamily I, II and III proteins
Metal Metal content (g-atoms/mole)
PH1575
(subfamily I)
AtDUF89
(subfamily II)
YMR027W
(subfamily III)
Fe 0.25 0.007 -- --
Ni 0.08 0.005 0.06 0.002 0.06 0.0005
Zn 0.25 0.019 -- 0.08 0.0010
Mg 0.04 0.003 -- --
Mn 0.007 0.0005 -- --
Co -- -- --
Cu 0.08 0.002 0.005 0.0003 0.02 0.0009
Mo 0.01 0.002 -- nd
Metals were analyzed by inductively coupled plasma-mass spectrometry. Data are means s.d. of three replicates. --, below detection limit; detection limits were from <0.001 to 0.03 g-atoms/mole, depending on the metal. nd, not determined. AtDUF89 is the DUF89 domain of Arabidopsis PanK2, At4g32180.
Nature Chemical Biology: doi:10.1038/nchembio.2108

Supplementary Table 5 | Data collection and refinement statistics of YMR027W crystals
YMR027W+Mg+F6P YMR027W+Mg YMR027W+Mn YMR027W+Mna
Data collection Space group C2 C2 C2 C2 Cell dimensions a, b, c (Å) 53.3, 108.6, 88.1 53.3, 108.6, 88.1 223, 95.5,78.1 223, 95.5,78.1 α, β, γ() 90, 97, 90 90, 97, 90 90, 105.8, 90 90, 105.8, 90 Resolution (Å) 1.77 (1.81–1.77)b 1.80 (1.84–1.80)b 2.39 (2.44–2.39)b 2.70 (2.74–2.70)b Rmerge 0.048 (0.69) 0.10 (0.66) 0.054 (0.54) 0.070(0.38) I / σI 5.5 (2.0) 24.8 (2.79) 15.0 (2.0) 98.0(89.5) Completeness (%) 99.8 (99.0) 99.9 (100) 97.1(95.7) 13.72(2.33 Redundancy 3.7 (3.6) 7.4 (6.9) 2.5 (2.4) 2.9 (2.6) Refinement Resolution (Å) 29.89–1.77 34.4–1.80 30.2–2.39 No. reflections 48080 46859 53710 Rwork / Rfree 15.6/19.2 15.8/19.5 19.9/23.7 No. atoms Protein 3796 3796 10374 Ions: Mg2+/Mn2+/PO4
2- F6P
1/-/- 17
1/-/- -
-/3/30 -
Water 335 510 179 B-factors Protein 17.23 28.03 50.1 F6P/ Mg2+/Mn2+ 17.9/13.4/- -/17.8/- -/-/40.1 Water 31.81 38.7 33.87 R.m.s. deviations Bond lengths (Å) 0.012 0.013 0.003 Bond angles () 1.40 1.33 0.63
a Data collected at Mn K-edge. b Number of crystals used: one. Highest-resolution shell is shown in parentheses.
Nature Chemical Biology: doi:10.1038/nchembio.2108

Supplementary Table 6 | Oligonucleotide primers used in this study
Primer name Sequence 5' to 3'a
AtDUF89-F ATCGTCATGAGATCTGACGCCAGTG AtDUF89-R ATATAAGCTTAAGCGAGGGAGGCTCGTA HsDUF89-F ATGCCCATGGAAATGGACCGGCT HsDUF89-R TATACTCGAGCTCGGCTGGGACCTC PH1575-Fb TTGTATTTCCAGGGCATGAAAGTTCAATATGAATG PH1575-Rb CAAGCTTCGTCATCATAGTTTAAATTTATTCCTCATGC YMR027W-Fb TTGTATTTCCAGGGCATGACTATCCCTGGAAGATTTATG YMR027W-Rb CAAGCTTCGTCATCATTTATGTATACCAGAGCAGAAAC AF1104-F AF1104-R MTH1744-F MTH1744-R
GCGGCGGCCCATATGAAAATTTCACCCCTCTGC GCGCGGATCCTTCGACCACCTTTGCCACC GCGGCGGCCCATATGAAGGTATACTATGAATGCG GCGCGGATCCGATCTTAACAGCAACATTGTC
aRestriction sites are underlined and bolded; F, forward; R, reverse. bPrimers for genes cloned using a ligation-independent protocol (LIC, New England Biolabs).
Nature Chemical Biology: doi:10.1038/nchembio.2108

Supplementary Table 7 | Database identifiers of DUF89 sequences for phylogenetic analysis
Species SEED Identifier GenBank Identifier
Spirochaeta smaragdinae fig|573413.5.peg.369
Chlorobium limicola fig|290315.4.peg.2622
Clostridium beijerinckii fig|290402.34.peg.3479
Thermotoga maritima fig|243274.5.peg.177
Alkaliphilus oremlandii fig|350688.3.peg.2716
Thiocystis violascens fig|765911.4.peg.4447
Wolinella succinogenes fig|765911.4.peg.4447
Korarchaeum cryptofilum fig|374847.3.peg.317
Pyrobaculum islandicum fig|384616.5.peg.562
Aquifex aeolicus fig|224324.8.peg.694
Desulfatibacillum alkenivorans fig|439235.4.peg.1861
Pyrococcus horikoshii fig|70601.1.peg.1543
Archaeoglobus fulgidus fig|224325.1.peg.1092
Methanothermobacter sp. CaT2 fig|187420.1.peg.1709
Yeast_YMR027W fig|559292.3.peg.4170
Plasmodium falciparum fig|36329.1.peg.330
Agaricus bisporus XP_006454137
Emiliania huxleyi XP_005789805
Nostoc punctiforme fig|63737.4.peg.6166
Salinibacter ruber fig|309807.25.peg.1392
Streptomyces avermitilis fig|227882.1.peg.2168
Nitrosomonas europaea fig|228410.6.peg.350
Rhodothermus marinus fig|518766.7.peg.2690
Human_C6orf211 fig|9606.3.peg.27930
Human PanK4 fig|9606.3.peg.104
Danio rerio_C6orf211 NP_001014353
Danio rerio_PanK XP_005155853
Apis mellifera XP_623407
Apis mellifera_PanK XP_006561827
Caenorhabditis elegans fig|6239.3.peg.22181
Caenorhabditis elegans_PanK fig|6239.3.peg.23236
Arabidopsis_AT2G17340 fig|3702.11.peg.17056
Arabidopsis_AT4G32180_PanK2 fig|3702.11.peg.9804
Maize_GRMZM2G151414 NP_001149554
Maize_PanK2 XP_008652990
Physcomitrella patens XP_001752572
Physcomitrella patens_PanK2 XP_001757682
Micromonas sp. RCC299 XP_002499881
Micromonas sp. RCC299_PanK XP_002507494
Micromonas sp. RCC299 XP_002500562
Nature Chemical Biology: doi:10.1038/nchembio.2108

Supplementary Figure 1 | Isolation of recombinant DUF89 proteins. The AF1104, PH1575, MTH1744,YMR027W, and Dalk1756 proteins, and the At2g17340-MBP fusion protein, were purified by Ni2+-affinitychromatography alone. The Arabidopsis PanK2 (At) and human PanK4 (Hs) DUF89 domains were furtherpurified by ion-exchange. The indicated quantities of purified proteins were analyzed by SDS-PAGE withCoomassie staining. Molecular mass markers are shown by each gel. The preparations used for enzymeassays were judged to be ≥95% homogeneous, except for Dalk1756.
At 10 µg
Hs10 µg
72 -
56 -
43 -
34 -
kDa
26 -
17 -
M
64 -
45 -
37 -
26 -
82 -
115 -
kDa M 5 µg 5 µg5 µg 5 µg
72 -
56 -
43 -
34 -
26 -
17 -
10 µgkDa M
70 -
55 -
40 -
35 -
25 -
15 -
130 -100 -
3 µg kDa M
Nature Chemical Biology: doi:10.1038/nchembio.2108

Supplementary Figure 2 | Metal dependence of DUF89 phosphatase activity. Except where otherwisenoted, metal salt and EDTA concentrations were 0.5 mM, assay temperature was 30°C, and incubationtime was 30 min. (a) The subfamily II standalone Arabidopsis At2g17340 protein and DUF89 domainsfrom Arabidopsis PanK2 and human PanK4. The substrate was p-nitrophenyl phosphate (1 mM forAt2g17340 and 2 mM for the other proteins). EDTA concentration for At2g17340 was 2 mM. Results aremeans and s.d. for four replicates (At2g17340) or three replicates (PanK2 and PanK4). (b) Subfamily Iprotein PH1575 from P. horikoshii. The substrate was 0.5 mM p-nitrophenyl phosphate. MgCl2concentration was 5 mM, EDTA concentration was 2 mM, and assay temperature was 55°C. Data aremeans of duplicates. (c) Subfamily III protein YMR027W from yeast. The substrate was 0.25 mM fructose6-phosphate. MgCl2 concentration was 5 mM, EDTA concentration was 2 mM, and incubation time was 15min. Data are means of duplicates.
Nature Chemical Biology: doi:10.1038/nchembio.2108

Supplementary Figure 3 | Kinetic behavior of the Arabidopsis At2g17340 enzyme with naturalaldopentose phosphate substrates or the artificial substrate p-nitrophenyl phosphate. Assays wererun at pH 7.0 with 0.2 mM Ni2+ as cofactor; incubation was for 20 min at 30°C. Results are means s.d.for four replicates. Kinetic parameters are shown in each plot.
Nature Chemical Biology: doi:10.1038/nchembio.2108

Supplementary Figure 4 | Evidence linking DUF89 subfamily I to purine synthesis. (a) Simplifiedscheme of the purine synthesis pathway showing steps governed by the genes that cluster with subfamilyI genes in various genomes. (b) Examples of chromosomal clustering patterns of DUF89 subfamily Igenes (red). Purine synthesis genes are blue; other genes are gray. Arrowheads show the direction oftranscription. Letters in parentheses after species names denote the major taxon they belong to: E,Euryarchaeota; P, ε-Proteobacteria; A, Aquificae; F, Firmicutes.
NitratiruptorSB155-2 (P) DUF89 purE Caminibacter
mediatlanticus (P) DUF89 purE
Methanosphaerastadtmanae (E) DUF89 purF
Helicobactercinaedi (P) DUF89 purE
Thermovibrioammonificans (A) DUF89 purE
Methanococcusvannieli (E) DUF89 purM
Methanocaldococcusjannaschii (E) DUF89 purFpurM
Lachnospiraceae bacterium (F) AICARFIMPC DUF89
Methanococcusmaripaludis (E)
DUF89 guaB purM
b
a
Ribose-5P PRPP Ribosyl-amine-5P GAR FGAR FGAM
AIR CAIR SAICAR AICAR FAICAR IMP
XMP GMP
AMP
purFpurM
CAPRI
purE
guaB
purP purO
Nature Chemical Biology: doi:10.1038/nchembio.2108

Supplementary Figure 5 | Activity of DUF89 subfamily I protein PH1575 against various nucleotidesubstrates. Screening assays contained 0.25 mM substrate and 0.5 mM Ni2+; temperature was 75°C.Kinetic constants with ADP or 8-oxo-2dGTP as substrate are shown in boxes. Screening data are meansof duplicates; kinetic data are means of four replicates s.d. Abbreviations: dATP, 2'-deoxyadenosine-5'-triphosphate; 2-aminopurine-2'-dTP, 2-aminopurine-2'-deoxyriboside-5’-triphosphate; UTP, uridine 5'-triphosphate; dCTP; 2'-deoxycytidine-5'-triphosphate; IDP, inosine-5'-diphosphate; 5-OH-2'-dCTP, 5-hydroxy-2'-deoxycitidine-5'-triphosphate; dGDP, 2'-deoxyguanosine 5'-diphosphate; 8-oxo-GTP, 8-oxo-guanosine-5’-triphosphate; GDP, guanosine 5'-diphosphate; TDP, thymidine 5'-diphosphate; 7-Deaza-2'-dGTP, 7-deaza-2'-deoxyguanosine-triphosphate; N4-Me-2'-dCTP, N4-methyl-2'-deoxycytidine-triphos-phate; CDP, cytidine 5'-diphosphate; O6-methyl-2'dGTP, O6-methyl-2'-deoxyguanosine-triphosphate; 8-oxo-2dATP, 8-oxo-2'-deoxyadenosine-5'-triphosphate; UDP, uridine-5'-diphosphate; ADP, adenosine-5'-diphosphate; N6-methyl-2-dATP; N6-methyl-2'-deoxyadenosinetriphosphate; dCDP, 2'-deoxycytidine-5'-diphosphate; 8-oxo-2dGTP; 8-oxo-2'-deoxyguanosine-5'-triphosphate; 5-Me-UTP, 5-methyluridine-5'-triphosphate; pNPP, p-nitrophenyl phosphate. The purine synthesis intermediate AICAR was also testedbut was not a substrate.
Nature Chemical Biology: doi:10.1038/nchembio.2108

Supplementary Figure 6 | Optical absorption spectra of DUF89 subfamily I proteins. (a) Spectra ofMethanothermobacter thermautotrophicus MTH1744, Pyrococcus horikoshii PH1575, and Archaeoglobusfulgidus AF1104 proteins. Protein concentrations were 2.2, 2.0, and 2.6 mg/ml, respectively. (b) Spectrumof Desulfatibacillum alkenivorans Dalk1756. Protein concentration was 0.4 mg/ml.
0
0.1
0.2
0.3
0.4
250 350 450 550 650
Abs
orba
nce
Wavelength (nm)
280 nm
330 nm420 nm
0.0
0.5
1.0
1.5
250 350 450 550 650
Abs
orba
nce
Wavelength (nm)
MTH1744 PH1575 AF1104
280 nm
330 nm
420 nm
b Desulfatibacillum alkenivorans Dalk1756
a Methanothermobacter thermautotrophicus MTH1744, Pyrococcus horikoshiiPH1575, and Archaeoglobus fulgidus AF1104
Nature Chemical Biology: doi:10.1038/nchembio.2108

Supplementary Figure 7 | Overall structures of DUF89 protomers. The protomer structures are shownin two orientations related by 90° rotation. The protein core domains are colored in gray (helices) andcyan (strands); the cap domains are colored in magenta. (a) PH1575 (PDB code 2G8L); (b) AF1104 (PDBcode 2FFJ); (c) At2g17340 (PDB code 2Q40).
Nature Chemical Biology: doi:10.1038/nchembio.2108

Supplementary Figure 8 | Close-up views of the active site of the AF1104 and At2g17340 DUF89proteins. (a) AF1104 with bound sulfates (S1 and S2); (b) At2g17340 with bound Mg2+; the W symbolsare water molecules.
b
a
Nature Chemical Biology: doi:10.1038/nchembio.2108

Supplementary Figure 9 | Phosphatase activities of mutant yeast YMR027W and At2g17340proteins. Data for the wild type (WT) proteins are included. Phosphatase assays contained 0.5 mM p-nitrophenyl phosphate plus 0.5 mM Ni2+ (YMR027W) or 1.0 mM p-nitrophenyl phosphate plus 0.2 mM Ni2+
(At2g17340) and were run at 30°C for 15 or 20 min. YMR027W data are means of four replicates ands.d. At2g17340 data are means of duplicates.
Nature Chemical Biology: doi:10.1038/nchembio.2108

Supplementary Figure 10 | Phosphatase activities and optical absorption spectra of mutant P.horikoshii PH1575 proteins. Data for the wild type (WT) protein are included. (a) Phosphatase activitiesof all mutants. Assays contained 0.5 mM p-nitrophenyl phosphate plus 0.5 mM Ni2+ and were run at 75°Cfor 15 min. Data are means of duplicates. (a) Spectra of conserved cysteine mutants. Protein concentrat-ions (mg/ml) were: WT, 3.6; C7A, 1.4; C10A, 1.6; C264A, 1.25. (c) Spectra of non-cysteine mutants.Protein concentrations (mg/ml) were: WT, 3.6; Q14A, 2.3; N108A, 2.0; D111A, 1.6; D156A, 3.0; N157A,2.3; E160A, 2; D191A, 2.4, K263A, 1.7.
b
0
0.5
1.0
1.5
2.0
250 350 450 550 650
Abs
orba
nce
Wavelength (nm)
PH1575 Cys mutants
C7AC10AC264A
WT
a
0
0.5
1.0
1.5PH1575 all mutants
µm
olm
in-1
mg-
1pr
otei
n
c
0
0.5
1.0
1.5
2.0
250 350 450 550 650Wavelength (nm)
PH1575 non-Cys mutants
Abs
orba
nce
WTQ14A
D156AN157AD191AE160AK263A
N108AD111A
Nature Chemical Biology: doi:10.1038/nchembio.2108

Supplementary Figure 11 | DNA sequence of the recoded Desulfatibacillum alkenivorans DUF89protein. The recoded DNA sequence is 81% identical to the native sequence.
ATGGAATTTTACCTGGATTGCTATGCCTGCTTTGTCCGTCAGACCATTAACGCTGCCCGCCACGCTGGCTGTGATGACCCGGTTCAACACAAAATTATGATCAAAGTGCTGGAACTGCTGCAGAACCTGGATCCGGAAACCACGCCGCCGCATGTTGCAAAAGCTGTCCACGGCATTATCCGTGAATATAGTAACAATCCGGATCCGTACAAAGAAGCGAAAGCGCGCTGCACCCGCGCGGCGATGGAACTGTATGATGACCTGAAAGCAATGGTTGCAGCTAGCGATGACCCGCTGGGTACCGCTCTGCGTCTGGCAATTGCTGGCAACATTATCGATCTGGGTGTCTCTGATGAACATGAAGACCTGTGGCACACCGTCGATCGCGTGCTGGCACAAGACTATGCTGTTGATGACGAAGGTCTGCTGCGTCGCGAAATGGAAAAAGCGGATCATATTCTGTACCTGGGCGACAATGCCGGTGAAACCGTGTTTGATCGCGTTCTGATCGAAACGATGGAAAAACCGGTGGTTTATGCAGTCCGTGGCCGCCCGGTGCTGAACGATGCAATTATGGAAGACGCTATCGCGGCCGGTCTGGATAAAGTGGCCACCCTGATTGATAATGGTGCCGATGCACCGGGTACCATCCTGAGCCTGTGTAAACCGGAATTTGTTGAAGCGTTCAACGAAGCCCCGGTCATTATCGCGAAAGGCCAGGGTAATTACGAAACCCTGAACAATGCCGGTCCGAAAGTGTTCTGCCTGCTGCAAATCAAATGTCCGGTTATTGAACGTGATATGGGTGCCCCGATGGGTGGTATTATTCTGCGTCAGTCAACGGCTCTG
Nature Chemical Biology: doi:10.1038/nchembio.2108

Supplementary Dataset 1. Untargeted metabolomics analysis of yeast wild type and YMR027W deletion strains. Cells were cultured on minimal medium with glucose or fructose as carbon source and harvested at OD600 = 0.5 or 1.0. Data represent six biological replicates; differences between wild type and deletant that are significant at P <0.05, according to Student's t-test, are highlighted in rose.
Nature Chemical Biology: doi:10.1038/nchembio.2108