supporting information - pnas · supporting information ... dna was stained by mounting specimens...
TRANSCRIPT

Supporting InformationGerbasi et al. 10.1073/pnas.1009781108SI Materials and MethodsGeneral Reagents and Antibodies. Drosophila S2 cells (a gift fromRichard Carthew, Northwestern University, Evanston, IL) weregrown in Schneider cell media (Invitrogen) containing 10% fetalbovine serum and 1% penicillin-streptomycin at 25 °C. Anti-Blanks serum was raised against the full-length protein fusedto an N-terminal His6-tag cloned into the pDEST17 vector (In-vitrogen) and expressed in Escherichia coli BL21(DE3). Anti-CG12493 serum was raised against an N-terminal GST fusionof amino acids 1–95 of the predicted protein sequence (to mini-mize cross-reactivity with Blanks), again expressed in E. coli BL21(DE3). Both Blanks and CG12493 rabbit antisera were raisedby Harlan Bioproducts. Polyclonal anti-Dicer2 antibodies werekindly provided by Qinghua Liu (University of Texas Southwes-tern Medical Center).
Dual-Luciferase Assay. Double-stranded RNAs corresponding toCG10630 (blanks), r2d2, dicer2, andwhitewere in vitro transcribedfrom PCR products containing T7 RNA polymerase promoters atboth ends. All Drosophila-based PCR products were amplifiedfrom cDNA generated by harvesting Canton S embryo or S2cell RNA with TRIzol (Invitrogen) and priming a SuperScript III(Invitrogen) reaction with oligo(dT) (Fermentas). PCR primersused to amplify dsRNA templates were:
10630_dsF: 5′-GCGTAATACGACTCACTATAGGGGCGGG-GAAAATAAAAGCGATGAG-3′
10630_dsR: 5′-GCGTAATACGACTCACTATAGGGTTTTCTT-TTGCTTCTTACGGCC-3′
Dcr2_dsF: 5′-GCGTAATACGACTCACTATAGGGTATCCCG-AAGACTCTGTAACCAAGA-3′
Dcr2_dsR: 5′-GCGTAATACGACTCACTATAGGGATGAAC-AACGCACGCTTTCC-3′
R2D2_dsF: 5′-GCGTAATACGACTCACTATAGGAAGATTT-CCTATTCTATCGTTGCCG-3′
R2D2_DSR: 5′-GCGTAATACGACTCACTATAGGATCTGT-GTCCGGGCACAAAA-3′
White_dsF: 5′-GCGTAATACGACTCACTATAGGGGTTTCG-TGACGAAGCTCCAAGC-3′
White_dsR: 5′-GCGTAATACGACTCACTATAGGGCCATTG-TTCAGATGCTCGGC-3′
The dsRNAs were synthesized in vitro using the T7 MEGA-script transcription kit (Ambion). The resulting dsRNAs werepurified by phenol/chloroform extraction and transfected into S2cells using Lipofectamine 2000 (Invitrogen) employing a dsRNA:Lipofectamine ratio of 3∶1. On the first day, cells were trans-fected with dsRNAs targeting control (white) or target genes(dicer2, r2d2, and CG10630). On the second day cells were trans-fected with Pp-luciferase and Renilla luciferase reporter plasmids,along with dsRNA targeting Pp-luciferase, using a 3∶1 ratio ofnucleic acid to Lipofectamine 2000 (1). On day three, cells wereharvested and assayed for Pp-luciferase and Renilla luciferaseactivity using a dual-luciferase reporter system (Promega). Pp-luciferase silencing activity was measured as the ratio of Pp-luci-ferase to Renilla luciferase activity. The average Pp-luciferaseactivity per well was calculated and standardized against thecontrol (white dsRNA-treated samples).
RNA Affinity Chromatography and Mass Spectrometry. All biotiny-lated and nonbiotinylated small RNAs were purchased fromDharmacon. The sequences of the Pp-luciferase siRNA guideand passenger strands were 5′-UCGAAGUAUUCCGCGUAC-
GUG-3′ and 5′- CGUACGCGGAAUACUUCGAUU-3′, respec-tively (2). The 3′-biotinylated Pp-luciferase guide strand siRNAwas annealed to the nonbiotinylated passenger strand in vitro inannealing buffer [30 mM Hepes (pH 7.5), 100 mM potassiumacetate, 2 mMmagnesium acetate] at a 1∶1 (guide and passengerstrand) concentration of 40.9 μM. Annealing reactions wereincubated at 95 °C for 2 min followed by 37 °C for 60 min. Afterthe biotinylated siRNAs were annealed, 20.45 nmol of the siRNAduplex were complexed with 250 μL of streptavidin beads(Pierce) for 30 min at 4 °C with gentle agitation. Once the siRNA-avidin affinity column was formed, the beads were washed by cen-trifugation in embryo lysis buffer (ELB) [30 mM Hepes (pH 7.5),100 mM potassium acetate, 2 mM magnesium acetate, 5 mMdithiothreitol] with 20 bead volumes to remove free siRNAs.After generation of the affinity matrix, the beads were mixedwith S2 lysates (prepared as described below) for 30 min at roomtemperature. After incubation with extract, the affinity columnswere washed by centrifugation in ELB with 80 bead volumes topurify siRNA-specific factors. The purified proteins were trypsi-nized and subjected to mass spectrometry as described previously(3) (4). Briefly, raw spectra files were converted to MZXML filesusing READW and searched against the European Bioinfor-matics Institute Drosophila proteome database containing 16,496entries as of August 12, 2007. Scaffold (version Scaffold_3_00_04,Proteome Software, Inc.) was used to validate MS/MS basedpeptide and protein identifications. Peptide identifications wereaccepted if they could be established at greater than 95.0% prob-ability as specified by the PeptideProphet algorithm (5). Proteinidentifications were accepted if they could be established atgreater than 99.9% probability and contained at least fouridentified peptides. Protein probabilities were assigned by theProteinProphet algorithm (6). Proteins that contained similarpeptides and could not be differentiated based on MS/MS ana-lysis alone were grouped to satisfy the principles of parsimony.The scaffold quantitative value, which normalizes spectral countsacross samples to each protein, is displayed in Table S1. Addition-ally, the total number of spectra identified in each sample, thenumber of unique peptides, and the percent coverage for eachprotein is also displayed in Table S1.
Ion-Exchange Chromatography and Electrophoretic Mobility ShiftAssays (EMSAs). S2 cells were lysed in ELB by freezing at −80 °Covernight followed by thawing for 30 min at room temperature.S2 lysates were centrifuged at 20;000 × g for 15 min and dialyzedagainst buffer A [20 mM Tris-HCl (pH 7.95), 10 mMNaCl, 1 mMDTT, 0.1 mM EDTA, and 10% glycerol]. After dialysis, the lysatewas filtered through a 0.2 μm filter and loaded onto a HiTrap Qanion exchange column. The samples were fractionated over alinear gradient with buffer B [20 mM Tris (pH 7.95), 1 M NaCl,1 mM DTT, 0.1 mM EDTA, and 10% glycerol]. Following frac-tionation, the samples were dialyzed against ELB and analyzedfor siRNA binding by native polyacrylamide gel electrophoresis(7). Dialyzed fractions (2.52 μL) were bound to radiolabeled siR-NAs in 0.175 μL 10× reaction lysis buffer [300 mM Hepes (pH7.5), 1 M potassium acetate, 20 mM magnesium acetate, and50 mM DTT], 1 μL 5× RNAi mix [125 mM creatine phosphate,5 mM ATP, 25 mM DTT, 0.1875 μL creatine kinase, 0.125 μLRNase inhibitor (40 U∕μL, Promega)], and 1 μL 50% glycerol ina 5 μL reaction. The sequences of the RNAs used for EMSAswere as follows:
Ppluc siRNA Top: 5′-CGUACGCGGAAUACUUCGAUU-3′
Gerbasi et al. www.pnas.org/cgi/doi/10.1073/pnas.1009781108 1 of 8

Ppluc siRNA Bottom: 5′-UCGAAGUAUUCCGCGUACG-UG-3′
Ppluc (Blunt) Top: 5′-CACGUACGCGGAAUACUUCGA-3′Ppluc (Blunt) 30 Top: 5′-AUCACGUACGCGGAAUACUUC-GAAAUGUCC-3′
Ppluc (Blunt) 30 Bottom: 5′-GGACAUUUCGAAGUAUU-CCGCGUACGUGAU-3′
Let-7 miRNA (Top): 5′-UAUACAAUGUGCUAGCUUUC-UU-3′
Let-7 miRNA (Bottom): 5′-UGAGGUAGUAGGUUGUAUA-GU-3′
Let-7 siRNA (Top): 5′-UAUACAACCUACUACCUCAUU-3′Let-7 siRNA (Bottom): 5′-UGAGGUAGUAGGUUGUAUA-GU-3′
Microscopic Techniques. Adult testes were dissected in Ringer’ssolution [123 mM NaCl, 5 mM KCl, 1.5 mM CaCl2 (pH 7.4)],then fixed, permeabilized, stained, and mounted as described (8).Primary antibody dilutions were as follows: Anti-Blanks 1∶1;000;anti-CG12493, 1∶1;000; goat anti-Vasa (Santa Cruz), 1∶500. Rho-damine–phalloidin (Sigma) was used at 1∶1;000. For confocalmicroscopy, DNA was stained with TO-PRO-3 iodide (Invitro-gen) at 1∶500 in phosphate-buffered saline þ0.1% Tween-20(PBST) for 30 min. Secondary antibodies used were goat anti-rabbit Texas red (1∶1;000, Molecular Probes), donkey anti-goatAlexa Fluor 488 (1∶500, Molecular Probes).
Phosphate-buffered saline þ0.1% Tween-20 (PBST) was usedfor all antibodies, washes, and stains. For conventional lightmicroscopy, DNA was stained by mounting specimens in VEC-TASHIELDmedium containing DAPI (Vector Labs) and imagedon a Zeiss Axiophot microscope. For phase contrast images,freshly dissected testes were placed on a glass slide in 40 μL Ring-er’s solution. A glass coverslip was gently applied and surfacetension provided pressure to squash open the tissue. A Zeiss 510Meta/ConfoCor3 laser scanning confocal microscope was usedfor confocal images.
Blanks, R2D2, and Loqs-RA cDNAs were directionally clonedinto pENTR/TEV/D-TOPO (Invitrogen) according to the man-ufacturer’s protocol. Entry vectors were confirmed by sequencingand recombined into the pAGW vector from the DrosophilaGateway collection (Terence Murphy, Carnegie Institute ofWashington, Baltimore, MD). S2 cells were transfected withEffectene reagent (Qiagen) according to the manufacturer’sprotocol. After approximately 48 h, cells were plated on glass-bot-tomed culture dishes (MatTek) treated with poly-L-lysine at a
density of 1 × 105 cells∕mL and imaged on a Zeiss Axiovert200 M microscope.
P Element Excision. The blanksKG00804 P element insertion mutantchromosomes balanced over TM3 were crossed to Δ2–3 transpo-sase flies balanced over TM6B. This cross resulted in blanksmutant heterozygous males that contained the Δ2–3 transposaseon the opposing third chromosome. These male flies werecrossed with Dr/TM3 virgin female flies. From this cross, weselected blanks/TM3 males that were crossed again to Dr/TM3virgin females. The resulting cross yielded candidate blanksKG00804
P element excision chromosomes balanced over TM3.
Quantitative PCR. Triplicate independent samples of total RNAwere harvested by dissecting 10 pairs of testes per replicate inice-coldRinger’s solution followed by extraction with 1mLTRIzolreagent. OnemicrogramRNAwas treated with RQ1DNase (Pro-mega) and reverse transcribed with SuperScript III (Invitrogen)using oligo(dT) (Fermentas). One microliter of reverse transcrip-tion product was used in a quantitative real-time PCR reactionusing the QuantiTect SYBR Green kit (Qiagen) in an iQ5 Real-Time PCRDetection System (BioRad). Dilution series were usedto determine the linear amplification range, and mRNA abun-dance was normalized to RpL32. Primer sequences, 5′–3′, wereas follows:
RpL32_QF: CCAAGGACTTCATCCGCCACCRpL32_QR: GCGGGTGCGCTTGTTCGATCC10630_QF: AAAATAAAAGCGATGAG10630_QR: TTTTCTTTTGCTTCTTACGGCC12493_QF: CAGAAAACAGCTCGGGAGGC12493_QR: GGGCAACTGTCCAACTGACGDiver_QF: ATGTCGAAATTTGATCAACTGGDiver QR: CGCCTGATGAAATTCTTTGTTAAACOsvaldo QF: ATGGGCGTTGGAGCTGCAACOsvaldo QR: CCTTGCTGCTCTCTCGGTTGAttD QF: ATGGAATGTCAGGCTTCAGGAttD QR: GCTCCATAAACTCCCTTGGTGTotC QF: ATGAATGCCTCCATTTCTCTACTTotC QR: TCTTGGGTTCGATTGATCTTCMtK QF: ATGCAACTTAATCTTGGAGCGMtK QR: TGGTTAGGATTGAAGGGCGAGypsy primers, as described by Czech et al. (9).Copia primers, as described by Klenov et al. (10).Stellate primers, as described by Vagin et al. (11).
1. Saleh MC, et al. (2006) The endocytic pathway mediates cell entry of dsRNA to induceRNAi silencing. Nat Cell Biol 8:793–802.
2. Nykanen A, Haley B, Zamore PD (2001) ATP requirements and small interfering RNAstructure in the RNA interference pathway. Cell 107:309–321.
3. Gerbasi VR, Golden DE, Hurtado SB, Sontheimer EJ (2010) Proteomic identification ofDrosophila siRNA-associated factors. Mol Cell Proteomics 9:1866–1872.
4. Link AJ, et al. (1999) Direct analysis of protein complexes using mass spectrometry.Nat Biotechnol 17:676–682.
5. Keller A, Nesvizhskii AI, Kolker E, Aebersold R (2002) Empirical statistical model toestimate the accuracy of peptide identifications made by MS/MS and databasesearch. Anal Chem 74:5383–5392.
6. Nesvizhskii AI, Keller A, Kolker E, Aebersold R (2003) A statistical model for identifyingproteins by tandem mass spectrometry. Anal Chem 75:4646–4658.
7. Pham JW, Pellino JL, Lee YS, Carthew RW, Sontheimer EJ (2004) A Dicer-2-dependent
80 S complex cleaves targeted mRNAs during RNAi in Drosophila. Cell 117:83–94.
8. Singh SR, Hou SX (2008) Immunohistological techniques for studying the Drosophila
male germ line stem cell. Methods Mol Biol 450:45–59.
9. Czech B, et al. (2008) An endogenous small interfering RNA pathway in Drosophila.
Nature 453:798–802.
10. Klenov MS, et al. (2007) Repeat-associated siRNAs cause chromatin silencing of
retrotransposons in the Drosophila melanogaster germ line. Nucleic Acids Res
35:5430–5438.
11. Vagin VV, et al. (2006) A distinct small RNA pathway silences selfish genetic elements in
the germ line. Science 313:320–324.
Gerbasi et al. www.pnas.org/cgi/doi/10.1073/pnas.1009781108 2 of 8

6xm
yc-B
lanks
6xm
yc-C
G1249
3
Vecto
r
WB: anti-CG12493
75
50
37
WB: anti-Blanks
WB: anti-Myc
75 -
50 -
37 -
25 -
75 -
50 -
6xm
yc-B
lanks
6xm
yc-C
G1249
3
Vecto
r
A B
C
250150
3725
sdaeB
ediuG A
NRis
regnessaP + ediuG A
NRis
IB: Dicer-2
IB: Blanks
D
0
0.2
0.4
0.6
0.8
1
1.2
1.4
White dsRNA
Blanks dsRNA
Fol
d Lu
cife
rase
Act
ivity
Fig. S1. (A) S2 cells were transfected with empty expression vector, myc6-tagged CG12493, or myc6-tagged blanks. Immunoblotting the S2 cell extracts foranti-Blanks shows specificity for the tagged (◂) and endogenous (⊲) Blanks but not tagged CG12493. The same samples were probed with anti-myc to ensurethat both recombinant proteins were expressed (Lower). (B) Samples in Awere run on a separate gel and probed with anti-CG12493 antibodies. (C) The siRNAaffinity chromatography and immunoblotting for Dicer2 (Upper) and CG10630/Blanks (Lower). S2 cell extracts were subject to affinity chromatography witheither streptavidin (SA) beads (Beads), SA beads in complex with Pp-luciferase (luc) guide strand alone (siRNA guide), or SA beads in complex with Pp-luc siRNAduplex (siRNA Guideþ Passenger). (D) Dual-luciferase assay in S2 cells. Cells were transfected with dsRNAs targeting either white or CG10630. Cells were latertransfected with plasmids expressing Pp luciferase and Renilla luciferase. The standardized Pp-luciferase activity was plotted as the average fold differencebetween white gene knockdowns and the CG10630 knockdowns. Error bars indicate the standard deviation of the mean.
Gerbasi et al. www.pnas.org/cgi/doi/10.1073/pnas.1009781108 3 of 8

DIC GFP Overlay
GFP-R2D2
GFP-Loqs-PA
GFP-Blanks
GFP-CG12493
a a’ a’’
b b’ b’’
c c’ c’’
d d’ d’’
nuc
nuc
nuc
nuc
α-tubulin pre-immune merge
α-tubulin Blanks merge
e
f
Fig. S2. Blanks and CG12493 localize to the nucleus. Four Drosophila double-stranded RNA-binding domain proteins [R2D2 (A), Loqs-PA (B), Blanks (C), andCG12493 (D)] were fused to GFP at their N termini and expressed in S2 cells to ascertain subcellular localization. Nuclei are discernible as large “indented”features in the differential interference contrast images in the left column (A–D), and are indicated at right. GFP channels are portrayed in A′–D′. Mergedimages (A″–D″) reveal that R2D2 and Loqs-PA localize primarily to the cytoplasm, whereas Blanks and CG12493 are only detected in nuclei. (E and F) Localiza-tion of Blanks to the nucleus of cultured cells. S2 cells were fixed and stained with anti-α-tubulin, which stains the nucleus and cytoplasm (E), and anti-Blanksprimary antibodies (F). Blanks is distributed between the nucleus and cytoplasm of S2 cells, with the majority residing within the nucleus.
Gerbasi et al. www.pnas.org/cgi/doi/10.1073/pnas.1009781108 4 of 8

Pro
be A
lone
Pro
be +
S2
extr
act
Pro
be +
IEX
Pro
be A
lone
Pro
be +
S2
extr
act
Pro
be +
IEX
21-nt Pp-luc blunt dsRNA 30-nt Pp-luc blunt dsRNA
Pro
be +
S2
extr
act
Pro
be +
IEX
Probe: Let-7 miRNA
RDI/RLC RDI/RLC
BlanksComplex Blanks
Complex
A B C
Fig. S3. (A) Whole S2 cell lysates or ion-exchange fractions from Fig. 2Bwere incubated with the radiolabeled let-7 microRNA duplex. (B) Whole S2 cell lysatesor ion-exchange fractions from Fig. 2B were incubated with the radiolabeled Pp-luc 21-nt duplex blunt RNA (without overhangs). (C) Whole S2 cell lysates orion-exchange fractions from Fig. 2B were incubated with the radiolabeled Pp-luc 30-nt duplex blunt RNA (without overhangs).
DAPI anti-CG12493 merge
TO-PRO-3 anti-CG12493 merge
C C’ C’’
D D’ D’’
E E’ E’’
SV
SC
GSC
SC
GSC
ON
ON
A A’ A’’
B B’ B’’
Fig. S4. (A) Blanks is present in nuclei of “onion”-stage round spermatids following meiosis. Condensed nuclei can be seen staining with both DAPI (A) andanti-Blanks (A′; merge, A″). (B) Blanks is expelled from nucleus during spermatid differentiation. Elongating spermatid nuclei stained by DAPI (B) no longercontain Blanks, which is instead seen in puncta at the base of the preindividualized nuclear bundle. Staining is not seen in later stages of differentiation. (C–E)Anti-CG12493 staining of wild-type adult testes reveals a localization pattern identical to that of Blanks (Fig. 4). (C–C″) Low magnification view of a testis. GSC,germ line stem cells; SC, spermatocytes; SV, seminal vesicle; and ON, onion-stage spermatids. Note staining of enlarged nuclei of primary spermatocytes (D–D″)and condensed nuclei of onion-stage round spermatid (E–E″) prior to spermatid differentiation.
Gerbasi et al. www.pnas.org/cgi/doi/10.1073/pnas.1009781108 5 of 8

IH
B
A
J
D
E F
G
C
K L
M N
O
Fig. S5. A P element-induced allele suppresses Blanks expression and disrupts sperm individualization. (A) Map of SUPor-P insertion line blanksKG00804. Exons,shaded boxes; UTRs, white boxes. The P element is inserted 29 bp downstream of the annotated transcription start site. (B and C) Quantitative real-time PCRmeasurements of blanks and CG12493mRNA harvested from adult blanks testes. The blanksmRNA is reduced >10-fold in the homozygous mutant (B), whereasCG12493 mRNA levels are unchanged (C). Normalization is relative to rpl32 mRNA. (D) Architecture of the CG12493 locus and P element insertion site. (E)Quantitative RT-PCR of CG12493 and blanksmRNA levels in homozygous and heterozygous CG12493G4506 mutant testes. (F) Immunoblot of CG12493 and Blanksprotein levels in CG12493 homozygous and heterozygous mutant testes. (G) DAPI staining of heterozygous (Left) and homozygous (Right) CG12493G4506 mutantseminal vesicles. Note the reduced volume of the CG12493 homozygous seminal vesicle, consistent with impaired sperm production. (H) Magnified phase
Gerbasi et al. www.pnas.org/cgi/doi/10.1073/pnas.1009781108 6 of 8
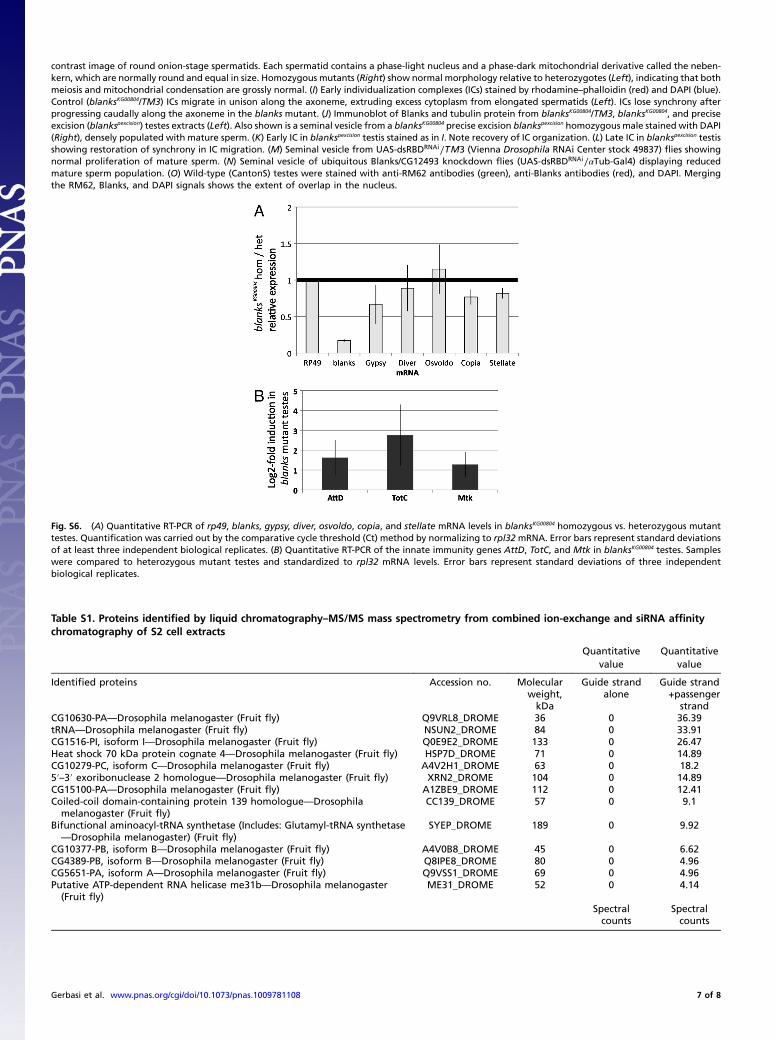
contrast image of round onion-stage spermatids. Each spermatid contains a phase-light nucleus and a phase-dark mitochondrial derivative called the neben-kern, which are normally round and equal in size. Homozygous mutants (Right) show normal morphology relative to heterozygotes (Left), indicating that bothmeiosis and mitochondrial condensation are grossly normal. (I) Early individualization complexes (ICs) stained by rhodamine–phalloidin (red) and DAPI (blue).Control (blanksKG00804/TM3) ICs migrate in unison along the axoneme, extruding excess cytoplasm from elongated spermatids (Left). ICs lose synchrony afterprogressing caudally along the axoneme in the blanks mutant. (J) Immunoblot of Blanks and tubulin protein from blanksKG00804/TM3, blanksKG00804, and preciseexcision (blankspexcision) testes extracts (Left). Also shown is a seminal vesicle from a blanksKG00804 precise excision blankspexcision homozygous male stained with DAPI(Right), densely populated with mature sperm. (K) Early IC in blankspexcision testis stained as in I. Note recovery of IC organization. (L) Late IC in blankspexcision testisshowing restoration of synchrony in IC migration. (M) Seminal vesicle from UAS-dsRBDRNAi∕TM3 (Vienna Drosophila RNAi Center stock 49837) flies showingnormal proliferation of mature sperm. (N) Seminal vesicle of ubiquitous Blanks/CG12493 knockdown flies (UAS-dsRBDRNAi∕αTub-Gal4) displaying reducedmature sperm population. (O) Wild-type (CantonS) testes were stained with anti-RM62 antibodies (green), anti-Blanks antibodies (red), and DAPI. Mergingthe RM62, Blanks, and DAPI signals shows the extent of overlap in the nucleus.
Fig. S6. (A) Quantitative RT-PCR of rp49, blanks, gypsy, diver, osvoldo, copia, and stellate mRNA levels in blanksKG00804 homozygous vs. heterozygous mutanttestes. Quantification was carried out by the comparative cycle threshold (Ct) method by normalizing to rpl32mRNA. Error bars represent standard deviationsof at least three independent biological replicates. (B) Quantitative RT-PCR of the innate immunity genes AttD, TotC, and Mtk in blanksKG00804 testes. Sampleswere compared to heterozygous mutant testes and standardized to rpl32 mRNA levels. Error bars represent standard deviations of three independentbiological replicates.
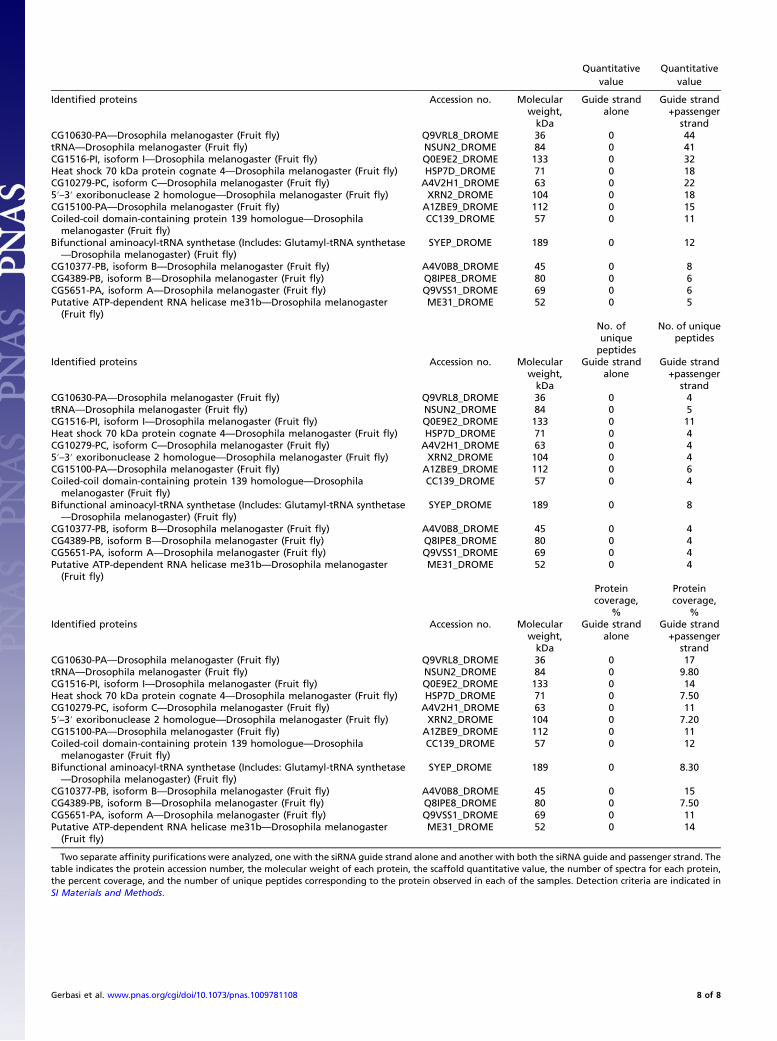
Table S1. Proteins identified by liquid chromatography–MS/MS mass spectrometry from combined ion-exchange and siRNA affinitychromatography of S2 cell extracts
Quantitativevalue
Quantitativevalue
Identified proteins Accession no. Molecularweight,kDa
Guide strandalone
Guide strand+passenger
strandCG10630-PA—Drosophila melanogaster (Fruit fly) Q9VRL8_DROME 36 0 36.39tRNA—Drosophila melanogaster (Fruit fly) NSUN2_DROME 84 0 33.91CG1516-PI, isoform I—Drosophila melanogaster (Fruit fly) Q0E9E2_DROME 133 0 26.47Heat shock 70 kDa protein cognate 4—Drosophila melanogaster (Fruit fly) HSP7D_DROME 71 0 14.89CG10279-PC, isoform C—Drosophila melanogaster (Fruit fly) A4V2H1_DROME 63 0 18.25′–3′ exoribonuclease 2 homologue—Drosophila melanogaster (Fruit fly) XRN2_DROME 104 0 14.89CG15100-PA—Drosophila melanogaster (Fruit fly) A1ZBE9_DROME 112 0 12.41Coiled-coil domain-containing protein 139 homologue—Drosophila
melanogaster (Fruit fly)CC139_DROME 57 0 9.1
Bifunctional aminoacyl-tRNA synthetase (Includes: Glutamyl-tRNA synthetase—Drosophila melanogaster) (Fruit fly)
SYEP_DROME 189 0 9.92
CG10377-PB, isoform B—Drosophila melanogaster (Fruit fly) A4V0B8_DROME 45 0 6.62CG4389-PB, isoform B—Drosophila melanogaster (Fruit fly) Q8IPE8_DROME 80 0 4.96CG5651-PA, isoform A—Drosophila melanogaster (Fruit fly) Q9VSS1_DROME 69 0 4.96Putative ATP-dependent RNA helicase me31b—Drosophila melanogaster
(Fruit fly)ME31_DROME 52 0 4.14
Spectralcounts
Spectralcounts
Gerbasi et al. www.pnas.org/cgi/doi/10.1073/pnas.1009781108 7 of 8
Quantitativevalue
Quantitativevalue
Identified proteins Accession no. Molecularweight,kDa
Guide strandalone
Guide strand+passenger
strandCG10630-PA—Drosophila melanogaster (Fruit fly) Q9VRL8_DROME 36 0 44tRNA—Drosophila melanogaster (Fruit fly) NSUN2_DROME 84 0 41CG1516-PI, isoform I—Drosophila melanogaster (Fruit fly) Q0E9E2_DROME 133 0 32Heat shock 70 kDa protein cognate 4—Drosophila melanogaster (Fruit fly) HSP7D_DROME 71 0 18CG10279-PC, isoform C—Drosophila melanogaster (Fruit fly) A4V2H1_DROME 63 0 225′–3′ exoribonuclease 2 homologue—Drosophila melanogaster (Fruit fly) XRN2_DROME 104 0 18CG15100-PA—Drosophila melanogaster (Fruit fly) A1ZBE9_DROME 112 0 15Coiled-coil domain-containing protein 139 homologue—Drosophila
melanogaster (Fruit fly)CC139_DROME 57 0 11
Bifunctional aminoacyl-tRNA synthetase (Includes: Glutamyl-tRNA synthetase—Drosophila melanogaster) (Fruit fly)
SYEP_DROME 189 0 12
CG10377-PB, isoform B—Drosophila melanogaster (Fruit fly) A4V0B8_DROME 45 0 8CG4389-PB, isoform B—Drosophila melanogaster (Fruit fly) Q8IPE8_DROME 80 0 6CG5651-PA, isoform A—Drosophila melanogaster (Fruit fly) Q9VSS1_DROME 69 0 6Putative ATP-dependent RNA helicase me31b—Drosophila melanogaster
(Fruit fly)ME31_DROME 52 0 5
No. ofuniquepeptides
No. of uniquepeptides
Identified proteins Accession no. Molecularweight,kDa
Guide strandalone
Guide strand+passenger
strandCG10630-PA—Drosophila melanogaster (Fruit fly) Q9VRL8_DROME 36 0 4tRNA—Drosophila melanogaster (Fruit fly) NSUN2_DROME 84 0 5CG1516-PI, isoform I—Drosophila melanogaster (Fruit fly) Q0E9E2_DROME 133 0 11Heat shock 70 kDa protein cognate 4—Drosophila melanogaster (Fruit fly) HSP7D_DROME 71 0 4CG10279-PC, isoform C—Drosophila melanogaster (Fruit fly) A4V2H1_DROME 63 0 45′–3′ exoribonuclease 2 homologue—Drosophila melanogaster (Fruit fly) XRN2_DROME 104 0 4CG15100-PA—Drosophila melanogaster (Fruit fly) A1ZBE9_DROME 112 0 6Coiled-coil domain-containing protein 139 homologue—Drosophila
melanogaster (Fruit fly)CC139_DROME 57 0 4
Bifunctional aminoacyl-tRNA synthetase (Includes: Glutamyl-tRNA synthetase—Drosophila melanogaster) (Fruit fly)
SYEP_DROME 189 0 8
CG10377-PB, isoform B—Drosophila melanogaster (Fruit fly) A4V0B8_DROME 45 0 4CG4389-PB, isoform B—Drosophila melanogaster (Fruit fly) Q8IPE8_DROME 80 0 4CG5651-PA, isoform A—Drosophila melanogaster (Fruit fly) Q9VSS1_DROME 69 0 4Putative ATP-dependent RNA helicase me31b—Drosophila melanogaster
(Fruit fly)ME31_DROME 52 0 4
Proteincoverage,
%
Proteincoverage,
%Identified proteins Accession no. Molecular
weight,kDa
Guide strandalone
Guide strand+passenger
strandCG10630-PA—Drosophila melanogaster (Fruit fly) Q9VRL8_DROME 36 0 17tRNA—Drosophila melanogaster (Fruit fly) NSUN2_DROME 84 0 9.80CG1516-PI, isoform I—Drosophila melanogaster (Fruit fly) Q0E9E2_DROME 133 0 14Heat shock 70 kDa protein cognate 4—Drosophila melanogaster (Fruit fly) HSP7D_DROME 71 0 7.50CG10279-PC, isoform C—Drosophila melanogaster (Fruit fly) A4V2H1_DROME 63 0 115′–3′ exoribonuclease 2 homologue—Drosophila melanogaster (Fruit fly) XRN2_DROME 104 0 7.20CG15100-PA—Drosophila melanogaster (Fruit fly) A1ZBE9_DROME 112 0 11Coiled-coil domain-containing protein 139 homologue—Drosophila
melanogaster (Fruit fly)CC139_DROME 57 0 12
Bifunctional aminoacyl-tRNA synthetase (Includes: Glutamyl-tRNA synthetase—Drosophila melanogaster) (Fruit fly)
SYEP_DROME 189 0 8.30
CG10377-PB, isoform B—Drosophila melanogaster (Fruit fly) A4V0B8_DROME 45 0 15CG4389-PB, isoform B—Drosophila melanogaster (Fruit fly) Q8IPE8_DROME 80 0 7.50CG5651-PA, isoform A—Drosophila melanogaster (Fruit fly) Q9VSS1_DROME 69 0 11Putative ATP-dependent RNA helicase me31b—Drosophila melanogaster
(Fruit fly)ME31_DROME 52 0 14
Two separate affinity purifications were analyzed, one with the siRNA guide strand alone and another with both the siRNA guide and passenger strand. Thetable indicates the protein accession number, the molecular weight of each protein, the scaffold quantitative value, the number of spectra for each protein,the percent coverage, and the number of unique peptides corresponding to the protein observed in each of the samples. Detection criteria are indicated inSI Materials and Methods.
Gerbasi et al. www.pnas.org/cgi/doi/10.1073/pnas.1009781108 8 of 8