sv2a and sv2c are not vesicular ca2+ transporters but...
TRANSCRIPT

IntroductionPancreatic hormones released by the islets of Langerhans playa pivotal role in the regulation of nutrient homeostasis.Defective insulin secretion from the �-cells leads to diabetesmellitus (Wollheim and Maechler, 2002). The moleculardetails of �-cell exocytosis, which resembles neuroexocytosis,remain to be fully clarified. However, during the last few yearsseveral key components of the machinery permitting thetargeting of secretory vesicles to the plasma membrane havebeen identified. In fact, SNARE proteins, the Ca2+ sensorssynaptotagmins and Rab GTPases with their interactingpartners, have been demonstrated to play a role in insulinrelease (Burgoyne and Morgan, 2003; Rorsman and Renstrom,2003).
Synaptic vesicle protein 2 (SV2) is a highly glycosylatedprotein also involved in exocytosis, which is present in neuronsand on secretory vesicles in endocrine cells including humanislets and HIT-T15 insulin-secreting cells (Buckley and Kelly,1985; Portela-Gomes et al., 2000). SV2 contains 12 potentialtransmembrane regions homologous to bacterial andeukaryotic transporters with cytoplasmic N- and C-terminals(Bajjalieh et al., 1992; Feany et al., 1992). However, transportsubstrates of SV2 remain to be identified.
Vertebrates have three distinct genes encoding highly
homologous proteins referred to as SV2A, -B and -C isoforms(Bajjalieh et al., 1992; Bajjalieh et al., 1993; Feany et al.,1992). SV2A and SV2C are more similar to each other than toSV2B (Janz et al., 1998). SV2A is expressed in all types ofneurons; SV2B is nearly as prevalent, whereas SV2C is onlypresent in a small subset (Bajjalieh et al., 1993; Janz andSudhof, 1999). The three SV2 isoforms are also differentiallydistributed in the mouse retina (Wang et al., 2003). Regardingthe neuroendocrine cells, SV2A is detected in the cell linePC12 whereas SV2B is associated with microvesicles of ratpinealocytes (Bajjalieh et al., 1994; Hayashi et al., 1998). Inadrenal chromaffin cells, SV2A is mainly localized to granuleswhereas SV2C is enriched on microsomes (Janz and Sudhof,1999). To date, the expression pattern and the intracellulardistribution of the three SV2 isoforms in insulin-secreting cellshave not been investigated.
Different hypotheses could explain SV2 function (Brose andRosenmund, 1999). The glycosylated intravesicular domainsof SV2 may trap soluble neurotransmitters to diminish theintravesicular osmotic pressure (Alvarez et al., 1993).Alternatively, SV2 could modulate exocytosis by interactingwith synaptotagmin I (Schivell et al., 1996; Lazzell et al., 2004)or act as a scaffold protein that regulates vesicle shape (Janz etal., 1998).
5647
Synaptic vesicle protein 2 (SV2) is expressed inneuroendocrine cells as three homologous isoforms, SV2A,SV2B and SV2C. Ca2+-dependent function in exocytosishas been attributed to SV2A and SV2B, without elucidationof the mechanism. The role of SV2C has not yet beenaddressed. Here we characterize the three SV2 isoformsand define their involvement in regulated insulin secretion.SV2A and SV2C are associated with insulin-containinggranules and synaptic-like-microvesicles (SLM) in INS-1Einsulinoma and primary ��-cells, whereas SV2B is onlypresent on SLM. Neither overexpression nor isoform-specific silencing of SV2A or SV2C by RNA interferencemodifies depolarization-triggered cytosolic [Ca2+] rises orsecretory granule [Ca2+], measured with a VAMP-2aequorin chimera. This strongly argues against anyCa2+ transport function of SV2. Moreover, up- ordownregulation of these isoforms has no influence on K+-induced insulin release suggesting that SV2 does not affect
the Ca2+-dependent step(s) of exocytosis. By contrast,glucose-elicited secretion is inhibited during the sustainedrather than the early phase, placing the action of SV2 onthe recruitment of granules from the reserve pool to theplasma membrane. This conclusion is reinforced bycapacitance measurements in glucose-stimulated SV2C-deficient cells. Like capacitance, evoked and basal hormonerelease are attenuated more by silencing of SV2C comparedwith SV2A. This indicates only partial redundancy andhighlights a key role for SV2C in the secretory process.
Supplementary material available online athttp://jcs.biologists.org/cgi/content/full/118/23/5647/DC1
Key words: INS-1E cells, Insulin release, Granular and cytosoliccalcium, Membrane capacitance, RNA interference
Summary
SV2A and SV2C are not vesicular Ca2+ transportersbut control glucose-evoked granule recruitmentMariella Iezzi1, Sten Theander1, Roger Janz2, Chantal Loze1 and Claes B. Wollheim1,*1Department of Cell Physiology and Metabolism, University Medical Center, 1 rue Michel-Servet, 1211 Geneva 4, Switzerland2W.M. Keck Center for Learning and Memory, Department of Neurobiology and Anatomy, University of Texas-Houston Medical School,6431 Fannin Street, Houston, TX 77030, USA*Author for correspondence (e-mail: [email protected])
Accepted 24 August 2005Journal of Cell Science 118, 5647-5660 Published by The Company of Biologists 2005doi:10.1242/jcs.02658
Research Article
Jour
nal o
f Cel
l Sci
ence

5648
A further study reported that SV2A and SV2A/SV2Bdouble-knockout mice suffer from strong epileptic seizuresand die postnatally, whereas mice lacking SV2B show nophenotype (Janz et al., 1999). The synaptic transmission wasincreased in double mutant hippocampal neurons and the effectwas reversed by cytosolic Ca2+ buffering suggesting thatpresynaptic terminals accumulate abnormally high Ca2+ levels.Therefore, SV2A and -B may act by binding Ca2+ or, morelikely, by pumping excess presynaptic Ca2+ into vesicles.
The epileptic seizures and the lethality of SV2A-deficientmice were reproduced in another investigation (Crowder et al.,1999). Nonetheless, a decrease in synaptic transmission wasobserved in hippocampal slices, indicating that SV2A positivelyregulates Ca2+-triggered exocytosis perhaps by binding tosynaptotagmin I. More recently, the same group analyzedchromaffin cells from SV2A and SV2A/SV2B double-knockoutmice (Xu and Bajjalieh, 2001). In these cells, the Ca2+-inducedexocytotic burst, which defines the ready releasable pool (RRP)of secretory vesicles, was diminished. Moreover, loss of SV2Awas associated with fewer SDS-resistant SNARE complexes inthe brain. These findings indicate that SV2A controls thenumber of vesicles competent for fusion.
In this study, we investigate the presence and subcellularlocalization of the three SV2 isoforms in native �-cells and inthe derived cell line INS-1E. We also examine the functionalimpact of SV2A and SV2C on Ca2+ levels and insulinexocytosis, comparing their overexpression to silencing byRNAi.
Materials and MethodsMaterials Rat SV2A, SV2B cDNAs and the monoclonal antibody against SV2were a kind gift from S. Bajjalieh (University of Washington, Seattle,WA). Rat SV2C cDNA and rabbit polyclonal antibodies against SV2A,SV2B and SV2C were prepared as described previously (Janz et al.,1999). Recombinant adenovirus expressing cytosolic aequorin underthe chicken actin promoter (AdCAcAq) was a kind gift from H. Ishihara(Tohoku University, Sendai, Japan). Chimeric cDNA encodinghemagglutinin-tagged ERaequorin fused to VAMP-2 (VAMP.Aq),generated as previously described (Mitchell et al., 2001), was suppliedby P. Pinton (University of Ferrara, Italy). The monoclonal antibodyagainst synaptotagmin IX was from BD Biosciences (Basel,Switzerland). Polyclonal antibodies against SV2A and SV2B were fromStressgen (Victoria, Canada). A rabbit polyclonal antibody directedagainst carboxypeptidase H was kindly provided by J. Parkinson(Sheffield Hallam University, Sheffield, UK). The polyclonal antibodiesagainst glucagon, somatostatin and pancreatic polypeptide weresupplied by P. Herrera (University of Geneva, Switzerland).
Cell culture and transfectionSyrian baby hamster kidney (BHK) cells were cultured in RPMI 1640medium with 10% FCS. The INS-1E clone from the insulin-secretingcell line INS-1 was cultured in RPMI 1640 medium and otheradditions as described (Merglen et al., 2004). Transient transfectionof BHK and INS-1E cells was performed using the Lipofectamine2000 reagent (Invitrogen, Gronlingen, Switzerland), according to themanufacturer’s instructions. In all the experiments, the DNA toLipofectamine ratio was 1.0:0.5.
Rat pancreatic isletsRat pancreatic islets were obtained by collagenase digestion and
purified on a Ficoll gradient as described (Pralong et al., 1990). Isletswere then homogenized and used for western blotting experiments asdescribed below. Alternatively, the islets were dissociated by trypsintreatment, seeded on poly-ornithine-coated glass coverslips andmaintained in culture 48 hours prior to the immunofluorescenceexperiments. In addition, islet cells were separated into twopopulations, non-�-cells and �-cells, by FACS and the latter wereused for RT-PCR studies.
Homogenate preparation and immunoblot analysisINS-1E cells, pancreatic islets and brain crude membranes wereprepared as described (Iezzi et al., 2004). The homogenates wereresolved by SDS-PAGE and transferred onto PVDF membrane.Immunoreactive bands were revealed by enhancedchemiluminescence (Pierce, Lausanne, Switzerland) usinghorseradish-peroxidase-coupled secondary antibodies.
ImmunocytochemistryINS-1E and primary pancreatic endocrine cells were fixed for 10minutes in 4% paraformaldehyde in PBS and permeabilized for 1 hourin PBS containing 0.1% saponin and 0.5% BSA. The cells wereincubated with primary antibodies overnight at 4°C and then exposedto FITC- or Rhodamine-conjugated secondary antibodies for 1 hourat room temperature. Samples were analyzed using a Zeiss laserconfocal microscope (LSM 510, Zurich, Switzerland). Images weretaken with a 60� objective.
Sucrose density gradientInsulin-containing granules were separated as described (Iezzi et al.,1999). Briefly, INS-1E cells were homogenized in 5 mM HEPES pH7.4, 1 mM EGTA, 10 �g/ml leupeptin and 2 �g/ml aprotinin. Celldebris and nuclei were eliminated by centrifuging the homogenate for10 minutes at 3000 g. The supernatant was loaded on a continuoussucrose density gradient (0.45-2.00 M) and centrifuged for 18 hoursat 110,000 g. The fractions were collected from the top of the gradient.The amount of insulin was monitored by radioimmunoassay (RIA).
Reverse-transcription and PCR amplification of SV2 cDNAsTotal RNA was extracted from rat brain, rat islet �-cells purified byFACS and INS-1E cells by the use of TrizolTM Reagent (Gibco LifeSciences, Basel, Switzerland). cDNA was synthesized for 45 minutesat 48°C followed by 2 minutes at 94°C using AMV reversetranscriptase (Promega Catalys, Walisellen, Switzerland). The primersused for reverse transcription and PCR amplification corresponded tonucleotides 675-697 of SV2A, 605-627 of SV2B and 516-536 ofSV2C cDNA sequences. The PCR program involved 40 cycles ofdenaturation at 94°C for 30 seconds annealing at 60°C for 1 minuteand elongation at 68°C for 2 minutes. PCR reactions were performedusing Tfl DNA polymerase (Promega).
Adenovirus construction and infectionSV2A, SV2C and VAMP.Aq cDNAs were used for the adenovirusconstructs. Recombinant adenoviruses encoding SV2A(AdCASV2A), SV2C (AdCASV2C) and the VAMP.Aq(AdCAVAMP.Aq) under the chicken actin promoter, were generatedas previously described (Miyake et al., 1996). AdCALacZ, whichexpresses bacterial �-galactosidase, was used as a control adenovirus.INS-1E cells, spheroid clusters of INS-1E cells or isolated islets, wereinfected with adenoviruses for 90 minutes and further cultured for 24hours (AdCASV2A, AdCASV2C and AdCAcAq) or 72 hours(AdCAVAMP.Aq) before experiments. INS-1E cells were infectedwith AdCASV2A and AdCASV2C at ~50 virus particles/cell, withAdCAcAq at ~20 virus particles/cell and with AdCAVAMP.Aq at ~10
Journal of Cell Science 118 (23)
Jour
nal o
f Cel
l Sci
ence

5649Role of SV2A and SV2C in regulated exocytosis
virus particles/cell; spheroids at 2�105 virus particles/spheroid (~50virus particles/cell, assuming 4000 cells/spheroid) (Merglen et al.,2004) and islets at 3�105 virus particles/islet (~100 virus particles/cell, assuming 3000 cells/islet).
[Ca2+] measurement with the expressed Ca2+-sensitivephotoprotein aequorinFor [Ca2+]c measurement, INS-1E cells were infected with AdCAcAqadenovirus and cytosolic aequorin was reconstituted with 2.5 �Mcoelenterazine in INS-1E cell culture medium for ~2 hours. For[Ca2+]SG measurement, INS-1E cells were infected withAdCAVAMP.Aq adenovirus. Cells were then Ca2+ depleted byincubation with 10 �M ionomycin, 10 �M monensin and 10 �M ofthe SERCA inhibitor cyclopiazonic acid (CPA), in Ca2+-free Krebs-Ringer-Bicarbonate buffer (KRB; 135 mM NaCl, 3.6 mM KCl, 2 mMNaHCO3, 0.5 mM NaH2PO4, 0.5 mM MgSO4, 2.5 mM glucose and10 mM HEPES, pH 7.4) supplemented with 1 mM EGTA for 5minutes at 4°C. ER.aequorin was reconstituted in 0.1 mM EGTA, 5�M coelenterazine-n for ~3 hours at 4°C.
For both [Ca2+]c and [Ca2+]SG measurement, the coverslips were
placed in a chamber with a thermostat 5 mm from the photomultiplierapparatus (model EMI 9789; Thorn EMI, Hayes, UK) set at 37°C.Cells were perifused at a rate of 1 ml/minute in the appropriate bufferand permeabilized with 0.1 mM digitonin, 10 mM CaCl2 forcalibration. Emitted light was collected every second with a photon-counting board (EMI 660). Insulin secretion was measured in theeffluent collected every 20 seconds.
Insulin secretionAttached cells
Prior to the experiment, infected INS-1E cells were maintained at37°C for 2 hours in glucose-free culture medium. The cells werepreincubated for 30 minutes in KRB buffer (135 mM NaCl, 3.6 mMKCl, 2 mM NaHCO3, 0.5 mM NaH2PO4, 0.5 mM MgSO4, 1.5 mMCaCl2, 10 mM HEPES, pH 7.4 and 0.1% BSA) containing 2.5 mMglucose and then incubated for 15 or 30 minutes in KRBsupplemented with stimulators as indicated. Infected islets (10islets/tube) were preincubated for 60 minutes in KRB containing 2.8mM glucose and further incubated over a period of 15 or 30 minutesin KRB containing stimulators as specified.
Perifused cellsSpheroid clusters composed of INS-1E cells were prepared asdescribed (Merglen et al., 2004). Briefly, ~500 infected spheroidswere distributed per chamber and maintained at 37°C (Brandel,Gaithersburg, MD). The flow rate was set at 0.5 ml/minute andfractions were collected every minute after a 20 minute washingperiod at basal glucose concentration. Insulin contents weredetermined from acid-ethanol extraction. Insulin was measured byRIA.
Preparation of vectors for SVA and SV2C silencingMammalian expression vectors directing the synthesis ofsiRNAs targeted against SV2A and SV2C were preparedaccording to manufacturer’s guidelines (Ambion,Huntingdon, UK). cDNA fragments encoding a 19-nucleotide sequence and separated from its reverse 19-nucleotide complement by a short spacer were synthesizedby Microsynth (Balgach, Switzerland). The SV2A silencerswere generated using the sequences corresponding tonucleotides 128-150 (siRNA-1), 226-248 (siRNA-2) and326-348 (siRNA-3) of rat SV2A cDNA. The SV2C silencerswere constructed using the nucleotide sequences 301-321(siRNA-1), 336-356 (siRNA-2) and 1705-1725 (siRNA-3) of
Fig. 1. Detection of SV2 isoforms in pancreatic endocrinecells. (A) PCR amplification of SV2 cDNAs from rat brain,INS-1E cells and rat islet �-cells purified by FACS. ControlPCR reactions with no reverse transcriptase were performedfor each isoform (–). The DNA size markers (bp) are alsoindicated. (B) Homogenates of rat brain (10 �g), rat isletsand INS-1E cells (100 �g each) were separated by SDS-PAGE and subjected to immunoblotting with themonoclonal antibody against SV2 and with rabbitpolyclonal antibodies against SV2A, SV2B or SV2C.Positions of molecular size markers are indicated in kDa.(C) Pancreatic endocrine cells were analyzed by confocalmicroscopy after double immunofluorescence with anantibody against glucagon (a), somatostatin (d) or pancreaticpolypeptide (g) (revealed using FITC-coupled antibody) andwith the anti-SV2 antibody (b, e and h) (detected usingRhodamine-coupled antibody). The merged images (c, f andi) were obtained after superposition of the green and redchannels. Bars, 5 �m.
Jour
nal o
f Cel
l Sci
ence
5650
rat SV2C cDNA. The cDNA fragments were annealedand cloned in front of the U6-RNA promoter in thepSilencerTM 1.0-U6 siRNA Expression Vector (Ambion).The specificity of each sequence was verified by basiclocal alignment search tool (BLAST) search against theGenBank database.
hGH secretionINS-1E cells were co-transfected with a plasmidencoding human growth hormone (hGH), and withplasmids containing the silencers. Four days later, thecells were maintained at 37°C for 2 hours in glucose-freeculture medium before preincubation for 30 minutes inKRB buffer containing 2.5 mM glucose. The cells werethen incubated for the indicated periods under basalcondition or in the presence of stimulatoryconcentrations of glucose and KCl. Exocytosis fromtransfected cells was assessed by measuring the amountof hGH released into the medium by ELISA (Roche,Manheim, Germany).
ElectrophysiologyINS-1E cells seeded on coverslips were co-transfectedwith GFP and the silencers. For patch-clamp capacitancerecording, the coverslip was transferred to a temperature-controlled chamber and perfused with extracellularsolution containing 120 mM NaCl, 20 mM tetraethylammonium, 3.6 mM KCl, 2.6 mM CaCl2, 2 mMNaHCO3, 0.5 mM NaH2PO4, 0.5 mM MgSO4, 5 mMHEPES, 15 mM glucose, 100 �M IBMX and 1 �Mforskolin (pH 7.4 with NaOH). The pipette internalsolution contained 125 mM CsCl, 10 mM sodiumglutamate, 5 mM HEPES, 3 mM Mg-ATP, 1 mM MgCl2,0.2 mM EGTA, 0.1 mM cAMP (pH 7.10 with CsOH).Capacitance measurements (EPC 9 amplifier; HEKA,Darmstadt, Germany) were performed after applying 1kHz, 28 mV peak, to sinusoid stimulus from a DCholding potential of –70 mV (Olofsson et al., 2002). Thesine+DC mode of the software lock-in extension of thePULSE software was used to calculate membranecapacitance (Cm), membrane conductance (Gm) andaccess resistance (Ra) from the current recordings.
Statistical analysisResults are presented as mean±s.e. Differences betweengroups were analyzed by the Student’s t-test for unpaireddata.
ResultsExpression of SV2A, SV2B and SV2C in pancreaticendocrine cellsWe first determined the presence of SV2A, -B and -C mRNAsin the insulin-secreting cells INS-1E, FACS-purified rat islet �-cells and rat brain. The latter was used as a positive control.RT-PCR with primers corresponding to the N-terminal regionof SV2 isoforms, yielded products of the predicted size (SV2A,190 bp; SV2B, 137 bp; SV2C, 144 bp) (Fig. 1A). Sequencingof the amplicons confirmed that INS-1E and �-cells express allthree SV2 isoforms, SV2B being less abundant compared withexpression in the brain.
Subsequently, we analyzed the expression of SV2 proteins.Homogenates of INS-1E cells, pancreatic islets and brain were
resolved by SDS-PAGE and the proteins were detected withmonospecific polyclonal antibodies directed against SV2A, -Bor -C. Western blotting revealed that SV2 isoforms migrate at~90-100 kDa and are present in all three homogenates (Fig.1B). Similar results were obtained using commercial SV2Aand -B antibodies (not shown). Each of the SV2 isoforms wasless abundant in pancreatic islets compared with INS-1E cells.This observation was confirmed by immunoblotting performedwith the general SV2 monoclonal antibody, recognizing allthree isoforms (Fig. 1B). Note that SV2 proteins migrate as aheterogeneous set of bands reflecting their high degree ofglycosylation (Buckley and Kelly, 1985).
To further examine the expression pattern of SV2, culturedprimary pancreatic endocrine cells were immunostained with
Journal of Cell Science 118 (23)
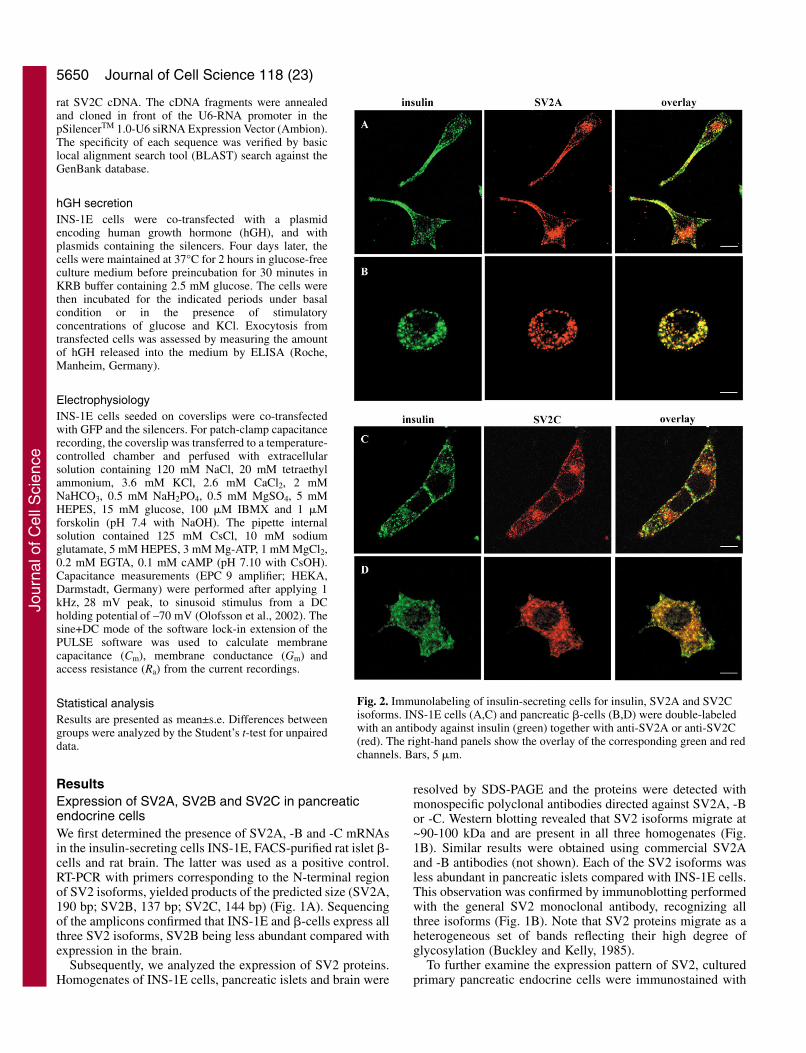
Fig. 2. Immunolabeling of insulin-secreting cells for insulin, SV2A and SV2Cisoforms. INS-1E cells (A,C) and pancreatic �-cells (B,D) were double-labeledwith an antibody against insulin (green) together with anti-SV2A or anti-SV2C(red). The right-hand panels show the overlay of the corresponding green and redchannels. Bars, 5 �m.
Jour
nal o
f Cel
l Sci
ence
5651Role of SV2A and SV2C in regulated exocytosis
the monoclonal SV2 antibody and analyzed by confocalmicroscopy. SV2 is present in the glucagon, somatostatin andpancreatic polypeptide cells (Fig. 1C).
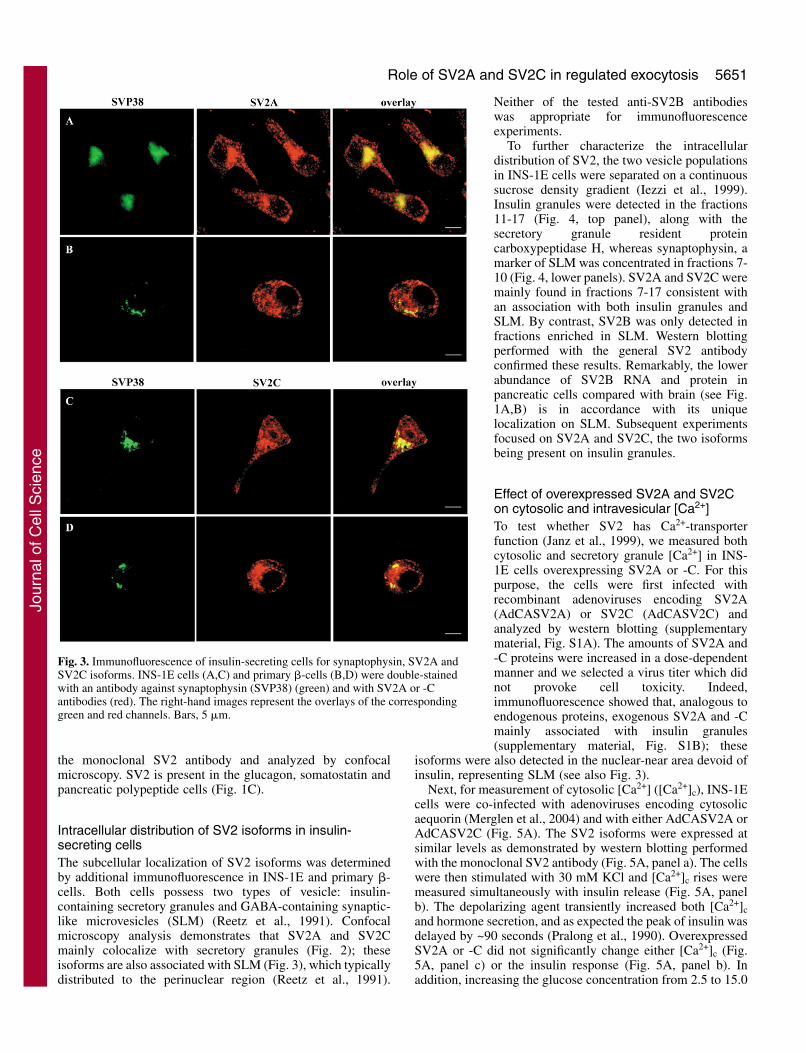
Intracellular distribution of SV2 isoforms in insulin-secreting cellsThe subcellular localization of SV2 isoforms was determinedby additional immunofluorescence in INS-1E and primary �-cells. Both cells possess two types of vesicle: insulin-containing secretory granules and GABA-containing synaptic-like microvesicles (SLM) (Reetz et al., 1991). Confocalmicroscopy analysis demonstrates that SV2A and SV2Cmainly colocalize with secretory granules (Fig. 2); theseisoforms are also associated with SLM (Fig. 3), which typicallydistributed to the perinuclear region (Reetz et al., 1991).
Neither of the tested anti-SV2B antibodieswas appropriate for immunofluorescenceexperiments.
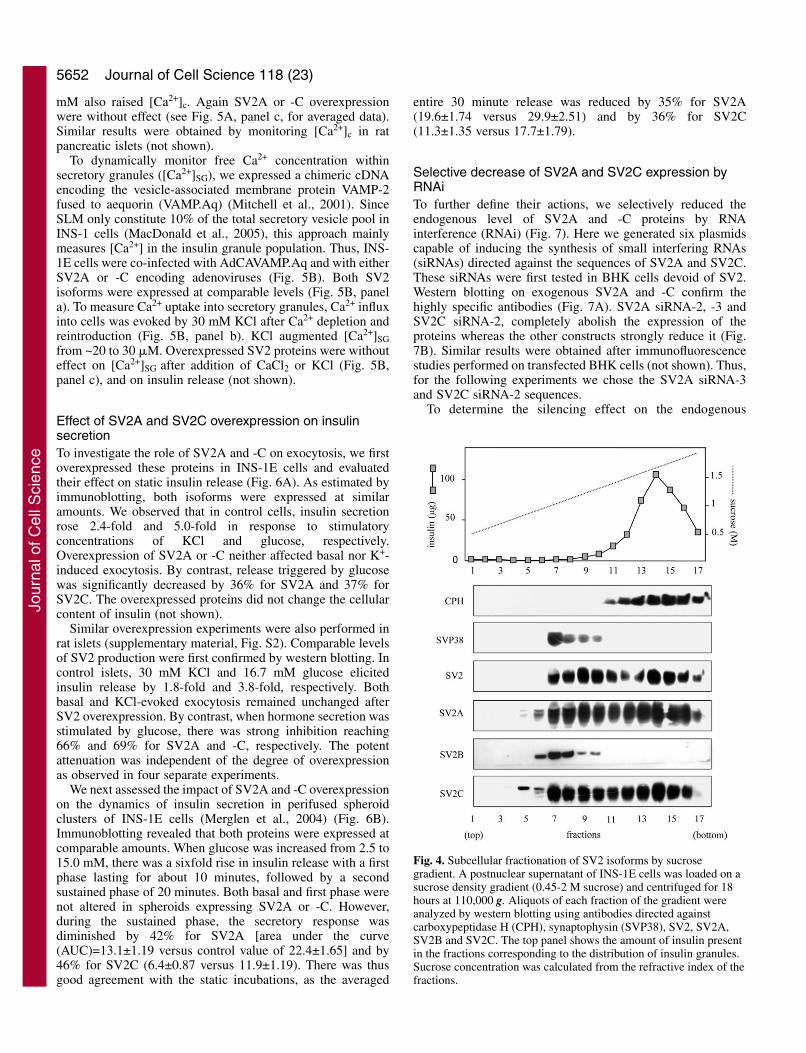
To further characterize the intracellulardistribution of SV2, the two vesicle populationsin INS-1E cells were separated on a continuoussucrose density gradient (Iezzi et al., 1999).Insulin granules were detected in the fractions11-17 (Fig. 4, top panel), along with thesecretory granule resident proteincarboxypeptidase H, whereas synaptophysin, amarker of SLM was concentrated in fractions 7-10 (Fig. 4, lower panels). SV2A and SV2C weremainly found in fractions 7-17 consistent withan association with both insulin granules andSLM. By contrast, SV2B was only detected infractions enriched in SLM. Western blottingperformed with the general SV2 antibodyconfirmed these results. Remarkably, the lowerabundance of SV2B RNA and protein inpancreatic cells compared with brain (see Fig.1A,B) is in accordance with its uniquelocalization on SLM. Subsequent experimentsfocused on SV2A and SV2C, the two isoformsbeing present on insulin granules.
Effect of overexpressed SV2A and SV2Con cytosolic and intravesicular [Ca2+]To test whether SV2 has Ca2+-transporterfunction (Janz et al., 1999), we measured bothcytosolic and secretory granule [Ca2+] in INS-1E cells overexpressing SV2A or -C. For thispurpose, the cells were first infected withrecombinant adenoviruses encoding SV2A(AdCASV2A) or SV2C (AdCASV2C) andanalyzed by western blotting (supplementarymaterial, Fig. S1A). The amounts of SV2A and-C proteins were increased in a dose-dependentmanner and we selected a virus titer which didnot provoke cell toxicity. Indeed,immunofluorescence showed that, analogous toendogenous proteins, exogenous SV2A and -Cmainly associated with insulin granules(supplementary material, Fig. S1B); these
isoforms were also detected in the nuclear-near area devoid ofinsulin, representing SLM (see also Fig. 3).
Next, for measurement of cytosolic [Ca2+] ([Ca2+]c), INS-1Ecells were co-infected with adenoviruses encoding cytosolicaequorin (Merglen et al., 2004) and with either AdCASV2A orAdCASV2C (Fig. 5A). The SV2 isoforms were expressed atsimilar levels as demonstrated by western blotting performedwith the monoclonal SV2 antibody (Fig. 5A, panel a). The cellswere then stimulated with 30 mM KCl and [Ca2+]c rises weremeasured simultaneously with insulin release (Fig. 5A, panelb). The depolarizing agent transiently increased both [Ca2+]cand hormone secretion, and as expected the peak of insulin wasdelayed by ~90 seconds (Pralong et al., 1990). OverexpressedSV2A or -C did not significantly change either [Ca2+]c (Fig.5A, panel c) or the insulin response (Fig. 5A, panel b). Inaddition, increasing the glucose concentration from 2.5 to 15.0
Fig. 3. Immunofluorescence of insulin-secreting cells for synaptophysin, SV2A andSV2C isoforms. INS-1E cells (A,C) and primary �-cells (B,D) were double-stainedwith an antibody against synaptophysin (SVP38) (green) and with SV2A or -Cantibodies (red). The right-hand images represent the overlays of the correspondinggreen and red channels. Bars, 5 �m.
Jour
nal o
f Cel
l Sci
ence
5652
mM also raised [Ca2+]c. Again SV2A or -C overexpressionwere without effect (see Fig. 5A, panel c, for averaged data).Similar results were obtained by monitoring [Ca2+]c in ratpancreatic islets (not shown).
To dynamically monitor free Ca2+ concentration withinsecretory granules ([Ca2+]SG), we expressed a chimeric cDNAencoding the vesicle-associated membrane protein VAMP-2fused to aequorin (VAMP.Aq) (Mitchell et al., 2001). SinceSLM only constitute 10% of the total secretory vesicle pool inINS-1 cells (MacDonald et al., 2005), this approach mainlymeasures [Ca2+] in the insulin granule population. Thus, INS-1E cells were co-infected with AdCAVAMP.Aq and with eitherSV2A or -C encoding adenoviruses (Fig. 5B). Both SV2isoforms were expressed at comparable levels (Fig. 5B, panela). To measure Ca2+ uptake into secretory granules, Ca2+ influxinto cells was evoked by 30 mM KCl after Ca2+ depletion andreintroduction (Fig. 5B, panel b). KCl augmented [Ca2+]SGfrom ~20 to 30 �M. Overexpressed SV2 proteins were withouteffect on [Ca2+]SG after addition of CaCl2 or KCl (Fig. 5B,panel c), and on insulin release (not shown).
Effect of SV2A and SV2C overexpression on insulinsecretionTo investigate the role of SV2A and -C on exocytosis, we firstoverexpressed these proteins in INS-1E cells and evaluatedtheir effect on static insulin release (Fig. 6A). As estimated byimmunoblotting, both isoforms were expressed at similaramounts. We observed that in control cells, insulin secretionrose 2.4-fold and 5.0-fold in response to stimulatoryconcentrations of KCl and glucose, respectively.Overexpression of SV2A or -C neither affected basal nor K+-induced exocytosis. By contrast, release triggered by glucosewas significantly decreased by 36% for SV2A and 37% forSV2C. The overexpressed proteins did not change the cellularcontent of insulin (not shown).
Similar overexpression experiments were also performed inrat islets (supplementary material, Fig. S2). Comparable levelsof SV2 production were first confirmed by western blotting. Incontrol islets, 30 mM KCl and 16.7 mM glucose elicitedinsulin release by 1.8-fold and 3.8-fold, respectively. Bothbasal and KCl-evoked exocytosis remained unchanged afterSV2 overexpression. By contrast, when hormone secretion wasstimulated by glucose, there was strong inhibition reaching66% and 69% for SV2A and -C, respectively. The potentattenuation was independent of the degree of overexpressionas observed in four separate experiments.
We next assessed the impact of SV2A and -C overexpressionon the dynamics of insulin secretion in perifused spheroidclusters of INS-1E cells (Merglen et al., 2004) (Fig. 6B).Immunoblotting revealed that both proteins were expressed atcomparable amounts. When glucose was increased from 2.5 to15.0 mM, there was a sixfold rise in insulin release with a firstphase lasting for about 10 minutes, followed by a secondsustained phase of 20 minutes. Both basal and first phase werenot altered in spheroids expressing SV2A or -C. However,during the sustained phase, the secretory response wasdiminished by 42% for SV2A [area under the curve(AUC)=13.1±1.19 versus control value of 22.4±1.65] and by46% for SV2C (6.4±0.87 versus 11.9±1.19). There was thusgood agreement with the static incubations, as the averaged
entire 30 minute release was reduced by 35% for SV2A(19.6±1.74 versus 29.9±2.51) and by 36% for SV2C(11.3±1.35 versus 17.7±1.79).
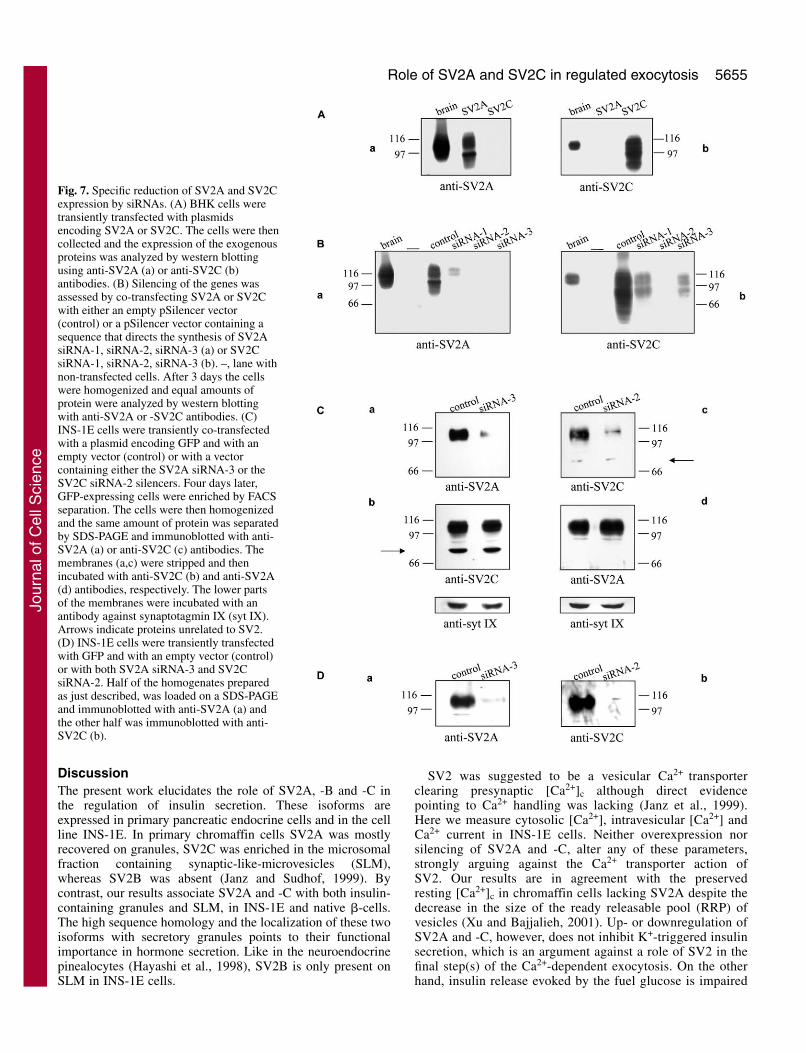
Selective decrease of SV2A and SV2C expression byRNAiTo further define their actions, we selectively reduced theendogenous level of SV2A and -C proteins by RNAinterference (RNAi) (Fig. 7). Here we generated six plasmidscapable of inducing the synthesis of small interfering RNAs(siRNAs) directed against the sequences of SV2A and SV2C.These siRNAs were first tested in BHK cells devoid of SV2.Western blotting on exogenous SV2A and -C confirm thehighly specific antibodies (Fig. 7A). SV2A siRNA-2, -3 andSV2C siRNA-2, completely abolish the expression of theproteins whereas the other constructs strongly reduce it (Fig.7B). Similar results were obtained after immunofluorescencestudies performed on transfected BHK cells (not shown). Thus,for the following experiments we chose the SV2A siRNA-3and SV2C siRNA-2 sequences.
To determine the silencing effect on the endogenous
Journal of Cell Science 118 (23)
Fig. 4. Subcellular fractionation of SV2 isoforms by sucrosegradient. A postnuclear supernatant of INS-1E cells was loaded on asucrose density gradient (0.45-2 M sucrose) and centrifuged for 18hours at 110,000 g. Aliquots of each fraction of the gradient wereanalyzed by western blotting using antibodies directed againstcarboxypeptidase H (CPH), synaptophysin (SVP38), SV2, SV2A,SV2B and SV2C. The top panel shows the amount of insulin presentin the fractions corresponding to the distribution of insulin granules.Sucrose concentration was calculated from the refractive index of thefractions.
Jour
nal o
f Cel
l Sci
ence

5653Role of SV2A and SV2C in regulated exocytosis
proteins, INS-1E cells were co-transfected with the selectedsiRNAs and GFP. Homogenates of FACS-enriched GFP-expressing cells were analyzed by western blotting (Fig. 7C).SV2A siRNA-3 potently diminished the amount of SV2A(Fig. 7C, panel a) but did not affect SV2C (panel b).Similarly, SV2C siRNA-2 drastically decreased SV2C (panelc) without changing the level of SV2A (panel d). The effectof the chosen siRNAs was specific because the expression ofsynaptotagmin IX or other endogenous proteins (arrows) wasnot modified. Since we required suppression of both isoformsfor subsequent secretion experiments, we demonstrated thatcombination of SV2A siRNA-3 and SV2C siRNA-2 stronglyreduced the corresponding proteins (Fig. 7D). Neither theoverall distribution of insulin granules nor the organizationof the cortical actin cytoskeleton revealed by phalloidin
staining was influenced by silencing of SV2 isoforms (notshown).
Effect of SV2A and SV2C suppression on [Ca2+]c and[Ca2+]SG
We next examined whether [Ca2+]c and [Ca2+]SG were alteredin cells devoid of SV2A or -C (Fig. 8). For this purpose, INS-1E cells coexpressing GFP and the selected silencers wereinfected with AdCAcAq (Fig. 8A) or AdCAVAMP.Aq (Fig.8B) and subsequently separated by FACS. Perifusion of thecells with 30 mM KCl transiently raised [Ca2+]c (Fig. 8A, paneli). This response remained unchanged after suppression ofSV2A or SV2C (see Fig. 8A, panel ii, for averaged data).Regarding Ca2+ uptake into secretory granules, the
depolarizing agent increased [Ca2+]SGfrom ~20 to 30 �M (Fig. 8B, panel i).SV2 depletion did not modify[Ca2+]SG after addition of CaCl2 orKCl to the perifusion medium (Fig.8B, panel ii).
Impact of SV2A and SV2Csilencing on hormone secretionand membrane capacitanceTo study the effect of SV2A and -Csilencing on INS-1E exocytosis, thesiRNAs constructs were co-transfected with a plasmid encodinghuman growth hormone (hGH) (Fig.9). Exogenous hGH is targeted to
Fig. 5. Effect of SV2A and SV2Coverexpression on cytosolic and vesicular[Ca2+]. (A) INS-1E cells were co-infectedwith AdCAcAq and with adenovirusesexpressing the control LacZ, SV2A orSV2C. (a) Immunoblotting analysis of theoverexpressed proteins using the SV2monoclonal antibody. (b) After cytosolicaequorin reconstitution cells wereperifused with KRB containing 2.5 mMglucose and stimulated, as indicated, withthe same buffer supplement with 30 mMKCl. Insulin secretion was assessed byRIA in the effluent. (c) Peak of [Ca2+]cafter KCl or 15 mM glucose (glc)stimulation. Values represent themean±s.e. of seven independent traces.(B) INS-1E cells were co-infected withAdCAVAMP.Aq and with LacZ, SV2A orSV2C encoding adenoviruses. (a) Westernblotting of the overexpressed proteinsusing anti-SV2 antibody. (b) After Ca2+
depletion and ER.aequorin reconstitution,cells were exposed to Ca2+-free KRBcontaining 1 mM EGTA. Whereindicated, EGTA was replaced with 1.5mM CaCl2 and the perifusion mediumsupplemented with 30 mM KCl. (c) Peakof [Ca2+]SG. Data are the mean±s.e. of sixindependent experiments.
Jour
nal o
f Cel
l Sci
ence

5654
secretory granules and can be used to monitor secretion as aninsulin substitute in the subpopulation of transfected cells(Iezzi et al., 2000). Under control conditions, hGH releaseincreased 2.9-fold after KCl (Fig. 9A) and 7.3-fold afterglucose stimulation (Fig. 9B). Loss of SV2C andSV2A/SV2C inhibited by 69% and 61% respectively basalsecretion within 15 minutes (Fig. 9A), and by 51% and 50%within 30 minutes (Fig. 9B). By contrast, repression of SV2Ahad no effect. KCl-induced exocytosis was also unchanged inSV2A or SV2C deficient cells. However, the glucose-triggered release was diminished by 32%, 71% and 69% inthe absence of SV2A, SV2C and SV2A/SV2C respectively.Remarkably, for SV2C and SV2A/SV2C the secretion wasreduced to the level attained with KCl (only evoking early-
phase release), suggesting preferential reduction of thesustained phase.
To analyze whether the two phases of insulin exocytosiswere affected by the reduction of a given SV2 isoform, wemeasured hGH secreted during the first 10 minutes (earlyphase) and between 10 and 30 minutes (sustained phase) (Fig.9C). Glucose elicited a 7.2- and 7.1-fold release during the first10 and last 20 minutes, respectively. Again, basal secretion wassignificantly inhibited by loss of SV2C but not of SV2A.Silencing of SV2A and -C decreased the early phase by 25%and 41%, while strongly reducing the sustained phase by 39%and 84%, respectively. Thus, repression of SV2A and -Ccaused an impairment of both secretory phases, the defectbeing more marked on the sustained phase. In all theseexperiments, the silencers did not modify the cellular contentof hGH (not shown).
To investigate the granule movement also at high temporalresolution we used patch-clamp capacitance recordings. Thus,we assessed the impact of SV2 depletion on membranecapacitance changes in conditions optimizing the recruitment
of granules to the readily releasablepool (RRP) (Fig. 10). Ten trains ofdepolarizing current pulses wereapplied in glucose-stimulated SV2A or-C deficient cells (Fig. 10A). WithoutSV2C, the capacitance was reduced by77% and 76% after the first and 10thpulse, respectively (see Fig. 10B foraveraged data), indicating that themaximal Ca2+ influx was not capableof normalizing the refilling of RRP.However, only a tendency of inhibitionwas observed in the absence of SV2A.The silencers did not change the Ca2+-current density (Fig. 10C). These dataare in agreement with those obtainedfor glucose-stimulated secretion (Fig.9C) and further emphasize the role ofSV2 in the translocation of granules tothe plasma membrane.
Journal of Cell Science 118 (23)
Fig. 6. Effect of SV2A and SV2C overexpression on insulinsecretion in INS-1E cells. (A) Western blotting analysis ofoverexpressed SV2A and -C in INS-1E cells using theantibody against SV2 (top panel). INS-1E cells infectedwith the control AdCALacZ (LacZ), AdCASV2A (SV2A)or AdCASV2C (SV2C) adenoviruses, were incubated atbasal 2.5 mM glucose (glc) and stimulated for 15 minuteswith 30 mM KCl, or for 30 minutes with 15 mM glucose.Data are the mean±s.e. of five independent experiments.Asterisks indicate significant differences from levels in theLacZ control at P<0.05. (B) Western blotting of exogenousSV2A and -C in spheroid clusters of INS-1E cells withanti-SV2 antibody (left-hand panel). Spheroids of INS-1Ecells infected with LacZ and SV2A, or SV2C encodingadenoviruses, were perifused with 2.5 mM glucose beforestimulation with 15 mM glucose for 30 minutes. Valuesrepresent the mean±s.e. (n=3 per group) of one out of fiveindependent experiments. Asterisks indicate a significantdifference from levels in the LacZ control forcorresponding time points at P<0.05.
Jour
nal o
f Cel
l Sci
ence
5655Role of SV2A and SV2C in regulated exocytosis
DiscussionThe present work elucidates the role of SV2A, -B and -C inthe regulation of insulin secretion. These isoforms areexpressed in primary pancreatic endocrine cells and in the cellline INS-1E. In primary chromaffin cells SV2A was mostlyrecovered on granules, SV2C was enriched in the microsomalfraction containing synaptic-like-microvesicles (SLM),whereas SV2B was absent (Janz and Sudhof, 1999). Bycontrast, our results associate SV2A and -C with both insulin-containing granules and SLM, in INS-1E and native �-cells.The high sequence homology and the localization of these twoisoforms with secretory granules points to their functionalimportance in hormone secretion. Like in the neuroendocrinepinealocytes (Hayashi et al., 1998), SV2B is only present onSLM in INS-1E cells.
SV2 was suggested to be a vesicular Ca2+ transporterclearing presynaptic [Ca2+]c although direct evidencepointing to Ca2+ handling was lacking (Janz et al., 1999).Here we measure cytosolic [Ca2+], intravesicular [Ca2+] andCa2+ current in INS-1E cells. Neither overexpression norsilencing of SV2A and -C, alter any of these parameters,strongly arguing against the Ca2+ transporter action ofSV2. Our results are in agreement with the preservedresting [Ca2+]c in chromaffin cells lacking SV2A despite thedecrease in the size of the ready releasable pool (RRP) ofvesicles (Xu and Bajjalieh, 2001). Up- or downregulation ofSV2A and -C, however, does not inhibit K+-triggered insulinsecretion, which is an argument against a role of SV2 in thefinal step(s) of the Ca2+-dependent exocytosis. On the otherhand, insulin release evoked by the fuel glucose is impaired
Fig. 7. Specific reduction of SV2A and SV2Cexpression by siRNAs. (A) BHK cells weretransiently transfected with plasmidsencoding SV2A or SV2C. The cells were thencollected and the expression of the exogenousproteins was analyzed by western blottingusing anti-SV2A (a) or anti-SV2C (b)antibodies. (B) Silencing of the genes wasassessed by co-transfecting SV2A or SV2Cwith either an empty pSilencer vector(control) or a pSilencer vector containing asequence that directs the synthesis of SV2AsiRNA-1, siRNA-2, siRNA-3 (a) or SV2CsiRNA-1, siRNA-2, siRNA-3 (b). –, lane withnon-transfected cells. After 3 days the cellswere homogenized and equal amounts ofprotein were analyzed by western blottingwith anti-SV2A or -SV2C antibodies. (C)INS-1E cells were transiently co-transfectedwith a plasmid encoding GFP and with anempty vector (control) or with a vectorcontaining either the SV2A siRNA-3 or theSV2C siRNA-2 silencers. Four days later,GFP-expressing cells were enriched by FACSseparation. The cells were then homogenizedand the same amount of protein was separatedby SDS-PAGE and immunoblotted with anti-SV2A (a) or anti-SV2C (c) antibodies. Themembranes (a,c) were stripped and thenincubated with anti-SV2C (b) and anti-SV2A(d) antibodies, respectively. The lower partsof the membranes were incubated with anantibody against synaptotagmin IX (syt IX).Arrows indicate proteins unrelated to SV2.(D) INS-1E cells were transiently transfectedwith GFP and with an empty vector (control)or with both SV2A siRNA-3 and SV2CsiRNA-2. Half of the homogenates preparedas just described, was loaded on a SDS-PAGEand immunoblotted with anti-SV2A (a) andthe other half was immunoblotted with anti-SV2C (b).
Jour
nal o
f Cel
l Sci
ence

5656
in both INS-1E cells and islets overexpressing the twoisoforms.
Fuel-stimulated insulin secretion involves recruitment of
granules from a reserve pool to the plasma membrane, docking,priming to achieve release competence, and formation of thereadily and immediately releasable pools (IRP). Granules in
Journal of Cell Science 118 (23)
A Bi i
0
250
500
750
1000
1250
1500
0 60 120 180 240
KCl 30 mM
[Ca2+
]nM
Control
0
5
10
15
20
25
30
35
1 60 120 180 240 300 360 420 480 540 600 660
1.5 mM CaCl2 30 mM KCl
[Ca
2+]
μM
Control
0
250
500
750
1000
1250
1500
0 60 120 180 240
KCl 30 mM
[Ca2+
]nM
SV2A
0
5
10
15
20
25
30
35
1 60 120 180 240 300 360 420 480 540 600 660
1.5 mM CaCl2 30 mM KCl
[Ca
2+]
μM
SV2A
0
250
500
750
1000
1250
1500
0 60 120 180 240
KCl 30 mM
[Ca2+
]nM
SV2C
0
5
10
15
20
25
30
35
1 60 120 180 240 300 360 420 480 540 600 660
1.5 mM CaCl2 30 mM KCl
[Ca
2+]
μM
SV2C
Time (seconds) Time (seconds)
ii ii
Control SV2A SV2C0
200
400
600
800
1000
1200
1400
�[C
a2+]
nM
Control SV2A SV2C
�[C
a2+]
μM
CaCl2 KCl CaCl2 KClCaCl2 KCl0
5
10
15
20
25
Fig. 8. Effect of SV2A and SV2C silencing on cytosolic and vesicular [Ca2+]. (A) INS-1E cells were transiently co-transfected with GFP and anempty vector (Control), or with either the SV2A siRNA-3 (SV2A) or the SV2C siRNA-2 (SV2C). Three days later, the cells were infected withAdCAcAq, further cultured for 24 hours and separated by FACS for enrichment of transfected cells. (i) After cytosolic aequorin reconstitution,the cells were perifused with 2.5 mM glucose and stimulated with 30 mM KCl as indicated. (ii) Peak of [Ca2+]c. Data show the mean±s.e. offour independent traces. (B) INS-1E cells were co-transfected as above. The day after, the cells were infected with AdCAVAMP.Aq, furthercultured for 72 hours and separated by FACS. (i) The cells were perifused as detailed in Fig. 5B, panel b. (ii) Peak of [Ca2+]SG. Values representthe mean±s.e. of four independent experiments.
Jour
nal o
f Cel
l Sci
ence

5657Role of SV2A and SV2C in regulated exocytosis
the IRP will undergo exocytosis after Ca2+ influx (Straub andSharp, 2002). Insulin release in response to glucose is biphasicwith a first phase (early phase) peaking after about 5 minutesand a second phase (sustained phase) beginning after 10minutes and lasting for hours (Wollheim and Sharp, 1981;Ishihara et al., 2003). The release of RRP granules (<5%)mainly accounts for first-phase secretion, and its end marks thedepletion of this pool. The subsequent supply of new granulesby mobilization/priming from a reserve pool (>95%) accountsfor second-phase secretion (Rorsman and Renstrom, 2003).The latter is also referred to as the amplifying pathway and isobserved with glucose but not with K+ stimulation (Henquin,2000).
In perifused INS-1E spheroids overexpressing the SV2isoforms, the secretory response is inhibited only during thesecond phase of insulin release. Thus, SV2 is likely to exert itsfunction mainly on the translocation of granules from a reservepool by interacting with proteins involved in this process. SV2
modulates the SNARE complex formation (Xu and Bajjalieh,2001) and associates with synaptotagmin (Lazzell et al., 2004;Schivell et al., 1996); the latter in turn, undergoes Ca2+-dependent binding to several components including syntaxin,SNAP-25, calmodulin, Na2+ and Ca2+ channels (Fukuda, 2003;Sudhof, 2004). Overexpression of SV2 may cause competitionwith these synaptotagmin partners thereby altering theirregulatory activity. Similarly, overexpression of the t-SNARE-interacting protein complexin I, impaired insulin exocytosis byaltering the formation of functional SNARE complexes(Abderrahmani et al., 2004). An excess of SV2 couldalso perturb secretion by decreasing the availability ofsynaptotagmin. Therefore, results based on overexpressionshould be complemented by suppression of the SV2 proteinsto further elucidate their function.
We found that specific silencing of SV2A attenuatesglucose-induced secretion, which is consistent with thediminished neurotransmitter release in SV2A-knockout mice
hGH
secr
etio
n (n
g/m
l)
Control siRNASV2A
siRNASV2C
siRNAsSV2A/SV2C
0
10
20
30
40
50
60
70
**
A
hGH
secr
etio
n (n
g/m
l)
ControlsiRNASV2A
siRNASV2C
siRNAsSV2A/SV2C
0
20
40
60
80
100
120
140
*
**
**
B
C
hGH
secr
etio
n (n
g/m
l)
siRNASV2A
siRNASV2C
Control0
5
10
15
20
25
30
35
*
*
0-10 min
*0
10
20
30
40
50
60
70
hGH
secr
etio
n (n
g/m
l)
siRNASV2A
siRNASV2C
Control
10-30 min
*
*
*
Fig. 9. Impact of SV2A and SV2Csuppression on hormone secretion. INS-1Ecells were transiently co-transfected with aplasmid encoding hGH together with theempty vector (Control), or the SV2AsiRNA-3 (siRNA SV2A), or the SV2CsiRNA-2 (siRNA SV2C) or with bothsilencing vectors (siRNAs SV2A/SV2C).Four days later, the cells were incubatedwith basal 2.5 mM glucose (A-C, openbars) and stimulated for 15 minutes with30 mM KCl (A, filled bars) or 30 minutes(B, filled bars) or for the indicated period(C, filled bars) with 15 mM glucose. After10 minutes, an aliquot of the medium wascollected (C, left-hand panel) and theincubation was continued for an additional20-minute period. Secretion between 10and 30 minutes was calculated bysubtracting the amount of hGH releasedduring the first 10 minutes from that forthe entire 30-minute incubation (C, right-hand panel). Data represent the mean±s.e.of three or four independent experiments.The conditions indicated by asterisks aresignificantly different from levels in thecontrol at P<0.05.
Jour
nal o
f Cel
l Sci
ence

5658
(Crowder et al., 1999) and demonstrates that SV2 acts aspositive modulator of exocytosis. Likewise, the RRP ofvesicles was impaired in SV2A-depleted chromaffin cells,despite increased expression of SV2C, suggesting that theseisoforms do not perform entirely redundant functions (Xu andBajjalieh, 2001). Here, INS-1E cells devoid of SV2A do notexpress more SV2C and vice-versa. In the secretionexperiments, the small inhibition in SV2A-knockout cellscould be compensated by SV2C whereas the strong reductionin SV2C-deficient cells could be only partially (or not at all)
compensated by SV2A. Thus, SV2A and -C are not completelyredundant, SV2C being functionally more relevant than SV2A.The lack of additive effect in the absence of both isoforms alsospeaks against an important role of SV2A.
In vitro SV2A and -B interaction with synaptotagmin I(Lazzell et al., 2004; Schivell et al., 1996) is modulated by SV2phosphorylation (Pyle et al., 2000). In INS-1 cells, the insulingranule-associated synaptotagmin I, II, V or IX (Iezzi et al.,2004; Lang et al., 1997) could also bind SV2A or -C. Thus,isoform-specific interactions and/or phosphorylation changes,
Journal of Cell Science 118 (23)
A
-200
20406080
100120140
�C
m(f
F)
-200
20406080
100�
Cm
(fF)
250 ms
100 pA
250 ms
100 pA
siRNASV2A
Control Control
siRNASV2A
�C
m(f
F)
-200
20406080
100 siRNASV2C
siRNASV2C
250 ms
100 pA
-70 mV
0 mV
1 second
CB
0
5
10
15
20
25
30
Ca2+
-cu
rren
tden
sity
(pA
/pF)
siRNASV2A
siRNASV2C
Control
1 pulse10 pulses
(pF/
pA)x
105
siRNASV2A
siRNASV2C
Control
*
*
0
10
20
30
40
5060
70
80
90
100
Fig. 10. Effect of SV2A and SV2Cdepletion on membranecapacitance changes. INS-1E cellswere transiently co-transfectedwith GFP and an empty vector(Control), or with either the SV2AsiRNA-3 (siRNA SV2A) or theSV2C siRNA-2 (siRNA SV2C).Four days later, recordings wereperformed on GFP-expressingcells. (A) Single cell capacitancetraces evoked by ten depolarizingpulses to 0 mV from a holdingpotential of –70 mV (left panels),and the corresponding Ca2+-currenttraces of the first depolarization inthe pulse train (right panels). Thepulse train was started 60 secondsafter establishing the whole-cellconfiguration (bottom). Cm,membrane capacitance; fF,femtoFarad. (B) Capacitancechanges normalized to the Ca2+
current of the first pulse for everycell, after one and ten pulses.Values are the mean±s.e. of 8 to 12cells. *P<0.05 vs. levels in thecontrol. pA, picoAmpere; pF,picoFarad. (C) Ca2+-currentdensity measured from the firstdepolarization in the pulse train.The Ca2+ current was evaluated 3milliseconds after the onset ofdepolarization to avoidcontamination of fast Na+ current.
Jour
nal o
f Cel
l Sci
ence

5659Role of SV2A and SV2C in regulated exocytosis
might explain the extent to which secretion is inhibited afterSV2 depletion. Like complexin I (Abderrahmani et al., 2004),the similar effects of up- and downregulation of SV2 isoforms,indicate that correct endogenous levels are required forappropriate protein stoichiometry.
In addition to stimulated conditions, silencing of SV2Calso inhibits basal secretion. This may either correspond todiminished transport of insulin vesicles from the Golgiapparatus to the cell periphery or to impaired endocytosis.INS-1 cells express synaptotagmin I and IX participating notonly in exocytosis (Iezzi et al., 2004; Lang et al., 1997) butalso in endocytosis (Haberman et al., 2003; Jarousse et al.,2001). Accordingly, SV2C may be involved in synaptotagminendocytosis, as suggested for retinal photoreceptor synapsesof SV2B-knockout mice after reduced expression ofsynaptotagmin I (Lazzell et al., 2004). Since SV2A and -Binteract with synaptotagmin I in a Ca2+-independent manner(Lazzell et al., 2004; Schivell et al., 1996), during basalrelease, a Ca2+-independent binding of synaptotagmins toSV2C would be operative. Interestingly, if SV2 were avesicular Ca2+ transporter, increased rather decreased basalsecretion would be expected.
Suppression of SV2A and -C led to an inhibition in thesustained phase of glucose-induced exocytosis. The earlyphase is also diminished but the effect is much weaker.Consistent with these data, capacitance recordings reveal thatloss of SV2C strongly impairs glucose-triggered granulerecruitment to the RRP, whereas there is only a tendency forSV2A. This is in accordance with the decreased pool of fusion-competent vesicles in SV2A-deficient chromaffin cells (Xu andBajjalieh, 2001). Of note, the unaltered K+-evoked hGHsecretion seems at variance with the capacitance results.However, during K+-depolarization, vesicular translocationdoes not occur (Olofsson et al., 2002). Taken together, thesefindings demonstrate that SV2 is mostly involved in theglucose-dependent progression of granules to the plasmamembrane.
In addition to synaptotagmin isoforms, SNARE proteins aswell as different Rab GTPases with their effectors andregulatory proteins have been implicated in �-cell exocytosis(Burgoyne and Morgan, 2003; Lang, 1999; Rorsman andRenstrom, 2003). Dynamic association of granules with theactin cytoskeleton is also required for the final step of glucose-dependent insulin secretion (Thurmond et al., 2003).Moreover, glucose but not high K+ stimulates the mobilizationof insulin granules to the plasma membrane, probably becauseof ATP generation (Varadi et al., 2002). Thus, it is possible thatSV2 regulates the reorganization of the actin cytoskeleton torecruit granules, modulates the binding with synaptotagmins orcontrols the formation of the release machinery.
In conclusion, we show that SV2A, -B and -C are expressedwith distinct locations in insulin-secreting cells. The secretory-granule-associated SV2A and SV2C are not Ca2+ transporters,but regulate the vesicular recruitment to the plasma membrane,without affecting the final step of exocytosis. Finally, SV2C isfunctionally more important than SV2A, which emphasizes anovel role for this isoform in endocrine secretion.
We are grateful to S. Mouche and D. Brighouse for technicalassistance. We are indebted to T. Pozzan for guidance in the use ofaequorin. This work was supported by the Swiss National Science
Foundation Grant (32-66907.01) to C.B.W., the Human Frontiers ofScience Program (RG-197/98) and a generous donation fromHubertus Spierings (Geneva, Switzerland) through the Fondation deRecherche Médicale.
ReferencesAbderrahmani, A., Niederhauser, G., Plaisance, V., Roehrich, M. E.,
Lenain, V., Coppola, T., Regazzi, R. and Waeber, G. (2004). ComplexinI regulates glucose-induced secretion in pancreatic �-cells. J. Cell Sci.. 117,2239-2247.
Alvarez de Toledo, G., Fernandez-Chacon, R. and Fernandez, J. M. (1993).Release of secretory products during transient vesicle fusion. Nature 363,554-558.
Bajjalieh, S. M., Peterson, K., Shinghal, R. and Scheller, R. H. (1992). SV2,a brain synaptic vesicle protein homologous to bacterial transporters.Science 257, 1271-1273.
Bajjalieh, S. M., Peterson, K., Linial, M. and Scheller, R. H. (1993). Braincontains two forms of synaptic vesicle protein 2. Proc. Natl. Acad. Sci. USA90, 2150-2154.
Bajjalieh, S. M., Frantz, G. D., Weimann, J. M., McConnell, S. K. andScheller, R. H. (1994). Differential expression of synaptic vesicle protein 2(SV2) isoforms. J. Neurosci. 14, 5223-5235.
Brose, N. and Rosenmund, C. (1999). SV2: SVeeping up excess Ca2+ ortranSVorming presynaptic Ca2+ sensors? Neuron 24, 766-768.
Buckley, K. and Kelly, R. B. (1985). Identification of a transmembraneglycoprotein specific for secretory vesicles of neural and endocrine cells. J.Cell Biol. 100, 1284-1294.
Burgoyne, R. D. and Morgan, A. (2003). Secretory granule exocytosis.Physiol. Rev. 83, 581-632.
Crowder, K. M., Gunther, J. M., Jones, T. A., Hale, B. C., Zhang, H. Z.,Peterson, M. R., Scheller, R. H., Chavkin, C. and Bajjalieh, S. M. (1999).Abnormal neurotransmission in mice lacking synaptic vesicle protein 2A(SV2A). Proc. Natl. Acad. Sci. USA 96, 15268-15273.
Feany, M. B., Lee, S., Edwards, R. H. and Buckley, K. M. (1992). Thesynaptic vesicle protein SV2 is a novel type of transmembrane transporter.Cell 70, 861-867.
Fukuda, M. (2003). Synaptotagmins, Ca2+-and phospholipid-binding proteinsthat control Ca2+-regulated membrane trafficking. Recent Res. Dev. Chem.Phys. Lipids 37, 15-51.
Haberman, Y., Grimberg, E., Fukuda, M. and Sagi-Eisenberg, R.(2003). Synaptotagmin IX, a possible linker between the perinuclearendocytic recycling compartment and the microtubules. J. Cell Sci. 116,4307-4318.
Hayashi, M., Yamamoto, A., Yatsushiro, S., Yamada, H., Futai, M.,Yamaguchi, A. and Moriyama, Y. (1998). Synaptic vesicle protein SV2B,but not SV2A, is predominantly expressed and associated withmicrovesicles in rat pinealocytes. J. Neurochem. 71, 356-365.
Henquin, J. C. (2000). Triggering and amplifying pathways of regulation ofinsulin secretion by glucose. Diabetes 49, 1751-1760.
Iezzi, M., Escher, G., Meda, P., Charollais, A., Baldini, G., Darchen, F.,Wollheim, C. B. and Regazzi, R. (1999). Subcellular distribution andfunction of Rab3A, B, C, and D isoforms in insulin-secreting cells. Mol.Endocrinol. 13, 202-212.
Iezzi, M., Kouri, G., Fukuda, M. and Wollheim, C. B. (2004).Synaptotagmin V and IX isoforms control Ca2+-dependent insulinexocytosis. J. Cell Sci. 117, 3119-3127.
Iezzi, M., Regazzi, R. and Wollheim, C. B. (2000). The Rab3-interactingmolecule RIM is expressed in pancreatic �-cells and is implicated in insulinexocytosis. FEBS Lett. 26, 66-70.
Ishihara, H., Maechler, P., Gjinovci, A., Herrera, P. L. and Wollheim, C.B. (2003). Islet beta-cell secretion determines glucagon release fromneighbouring alpha-cells. Nat. Cell Biol. 5, 330-335.
Janz, R. and Sudhof, T. C. (1999). SV2C is a synaptic vesicle protein withan unusually restricted localization: anatomy of a synaptic vesicle proteinfamily. Neuroscience 94, 1279-1290.
Janz, R., Hofmann, K. and Sudhof, T. C. (1998). SVOP, an evolutionarilyconserved synaptic vesicle protein, suggests novel transport functions ofsynaptic vesicles. J. Neurosci. 18, 9269-9281.
Janz, R., Goda, Y., Geppert, M., Missler, M. and Sudhof, T. C. (1999).SV2A and SV2B function as redundant Ca2+ regulators in neurotransmitterrelease. Neuron 24, 1003-1016.
Jarousse, N. and Kelly, R. B. (2001). The AP2 binding site of synaptotagmin
Jour
nal o
f Cel
l Sci
ence

5660
1 is not an internalization signal but a regulator of endocytosis. J. Cell Biol.154, 857-866.
Lang, J. (1999). Molecular mechanisms and regulation of insulinexocytosis as a paradigm of endocrine secretion. Eur. J. Biochem. 259, 3-17.
Lang, J., Fukuda, M., Zhang, H., Mikoshiba, K. and Wollheim, C. B.(1997). The first C2 domain of synaptotagmin is required for exocytosis ofinsulin from pancreatic beta-cells: action of synaptotagmin at lowmicromolar calcium. EMBO J. 16, 5837-5846.
Lazzell, D. R., Belizaire, R., Sherry, D. M. and Janz, R. (2004). SV2Bregulates synaptotagmin I by direct interaction. J. Biol. Chem. 279, 52124-52131.
MacDonald, P. E., Obermuller, S., Vikman, J., Galvanovskis, J., Rorsman,P. and Eliasson, L. (2005). Regulated exocytosis and kiss-and-run ofsynaptic-like microvesicles in INS-1 and primary rat beta-cells. Diabetes 54,736-743.
Merglen, A., Theander, S., Rubi, B., Chaffard, G., Wollheim, C. B. andMaechler, P. (2004). Glucose sensitivity and metabolism-secretion couplingstudied during two-year continuous culture in INS-1E insulinoma cells.Endocrinology 145, 667-678.
Mitchell, K. J., Pinton, P., Varadi, A., Tacchetti, C., Ainscow, E. K.,Pozzan, T., Rizzuto, R. and Rutter, G. A. (2001). Dense core secretoryvesicles revealed as a dynamic Ca2+ store in neuroendocrine cells with avesicle-associated membrane protein aequorin chimaera. J. Cell Biol. 155,41-51.
Miyake, S., Makimura, M., Kanegae, Y., Harada, S., Sato, Y., Takamori,K., Tokuda, C. and Saito, I. (1996). Efficient generation of recombinantadenoviruses using adenovirus DNA-terminal protein complex and a cosmidbearing the full-length virus genome. Proc. Natl. Acad. Sci. USA 93, 1320-1324.
Olofsson, C. S., Gopel, S. O., Barg, S., Galvanovskis, J., Ma, X., Salehi,A., Rorsman, P. and Eliasson, L. (2002). Fast insulin secretion reflectsexocytosis of docked granules in mouse pancreatic �-cells. Pflugers Arch.444, 43-51.
Portela-Gomes, G. M., Lukinius, A. and Grimelius, L. (2000). Synapticvesicle protein 2, a new neuroendocrine cell marker. Am. J. Pathol. 157,1299-1309.
Pralong, W. F., Bartley, C. and Wollheim, C. B. (1990). Single islet beta-
cell stimulation by nutrients: relationship between pyridine nucleotides,cytosolic Ca2+ and secretion. EMBO J. 9, 53-60.
Pyle, R. A., Schivell, A. E., Hidaka, H. and Bajjalieh, S. M. (2000).Phosphorylation of synaptic vesicle protein 2 modulates binding tosynaptotagmin. J. Biol. Chem. 275, 17195-17200.
Reetz, A., Solimena, M., Matteoli, M., Folli, F., Takei, K. and De Camilli,P. (1991). GABA and pancreatic �-cells: colocalization of glutamic aciddecarboxylase (GAD) and GABA with synaptic-like microvesicles suggeststheir role in GABA storage and secretion. EMBO J. 10, 1275-1284.
Rorsman, P. and Renstrom, E. (2003). Insulin granule dynamics inpancreatic beta cells Diabetologia 46, 1029-1045.
Schivell, A. E., Batchelor, R. H. and Bajjalieh, S. (1996). Isoform-specific,calcium-regulated interaction of the synaptic vesicle proteins SV2 andsynaptotagmin J. Biol. Chem. 271, 27770-27775.
Straub, S. G. and Sharp, G. W. (2002). Glucose-stimulated signaling pathwaysin biphasic insulin secretion. Diabetes Metab. Res. Rev. 18, 451-463.
Sudhof, T. C. (2004). The synaptic vesicle cycle. Annu. Rev. Neurosci. 27,509-547.
Thurmond, D. C., Gonelle-Gispert, C., Furukawa, M., Halban, P. A. andPessin, J. E. (2003). Glucose-stimulated insulin secretion is coupled to theinteraction of actin with the t-SNARE (target membrane soluble N-ethylmaleimide-sensitive factor attachment protein receptor protein)complex. Mol. Endocrinol. 17, 732-742.
Varadi, A., Ainscow, E. K., Allan, V. J. and Rutter, G. A. (2002).Involvement of conventional kinesin in glucose-stimulated secretory granulemovements and exocytosis in clonal pancreatic beta-cells. J. Cell Sci. 115,4177-4189.
Wang, M. M., Janz, R., Belizaire, R., Frishman, L. J. and Sherry, D. M.(2003). Differential distribution and developmental expression of synapticvesicle protein 2 isoforms in the mouse retina. J. Comp. Neurol. 460, 106-122.
Wollheim, C. B. and Sharp, G. W. (1981). Regulation of insulin release bycalcium. Physiol. Rev. 61, 914-973.
Wollheim, C. B. and Maechler, P. (2002). �-cell mitochondria and insulinsecretion: messenger role of nucleotides and metabolites. Diabetes 51, S37-S42.
Xu, T. and Bajjalieh, S. M. (2001). SV2 modulates the size of the readilyreleasable pool of secretory vesicles. Nat. Cell Biol. 3, 691-698.
Journal of Cell Science 118 (23)
Jour
nal o
f Cel
l Sci
ence