t a crispr/c 9 - nova science publishers
TRANSCRIPT

In: The CRISPR/Cas9 System ISBN: 978-1-53616-426-8
Editor: Alfred A. Bertelsen © 2019 Nova Science Publishers, Inc.
Chapter 3
THE APPLICATION OF CRISPR/CAS9
THERAPIES IN OPHTHALMOLOGY AND
RECENT ADVANCES FOR THE TREATMENT
OF GENETIC EYE DISEASE
C. B. Tara Moore1,, PhD, Larry A. DeDionisio2,†, PhD,
Hila Roshanravan2,‡, PhD, Connie Chao-Shern2,§
and M. Andrew Nesbit1, #, PhD 1Ulster University, Coleraine, County Londonderry,
UK/Northern Ireland 2Avellino Lab USA, Menlo Park, CA, US
Corresponding Author’s E-mail: [email protected]. † Author’s E-mail: [email protected]. ‡ Author’s E-mail: [email protected]. § Author’s Email: [email protected]. # Author’s E-mail: [email protected].
No part of this digital document may be reproduced, stored in a retrieval system or transmitted commercially in any form or by any means. The publisher has taken reasonable care in the preparation of this digital document, but makes no expressed or implied warranty of any kind and assumes no responsibility for any errors or omissions. No liability is assumed for incidental or consequential damages in connection with or arising out of information contained herein. This digital document is sold with the clear understanding that the publisher is not engaged in rendering legal, medical or any other professional services.

C. B. Tara Moore, Larry A. DeDionisio, Hila Roshanravan et al. 100
ABSTRACT
Clustered regularly interspaced short palindromic repeats (CRISPR)
and CRISPR-associated (Cas) protein systems have been used to target
chosen regions of the genome with high efficiency and specificity and,
together with the multiplexing capabilities of this technology, have
introduced a novel tool to the field of genome engineering. With a potential
for the permanent treatment of genetic ocular diseases, CRISPR based
therapeutics hold great potential for the field of ophthalmology. CRISPR
systems can repair genetic mutations in recessive diseases where both
copies of a gene carry a pathogenic mutation, or they can knock out the
pathogenic gene allele in dominant diseases where only one copy of the
gene carries the mutation leaving the functionality of the second gene copy
in place. The majority of corneal dystrophies are the result an autosomal
dominant inheritance pattern within the TGFBI gene, which presents an
ideal model suited for a CRISPR/Cas knock out methodology. This
approach can also be applied to the treatment of autosomal dominantly
inherited genetic disease in general. There are several ocular diseases that
are currently being investigated worldwide for which human gene transfer
trials have been approved. This review summarises the use of CRISPR-Cas
gene editing in ophthalmology and focuses on the advancement of gene
editing in the cornea.
1. INTRODUCTION
Vision is often considered the most important human sense. Our eyes
facilitate daily activities from basic mobility to the appreciation of art and
nature. Genetically heritable eye disease have the potential to impact those
who suffer from them in profound ways. In ophthalmology, disease-causing
genetic mutations that lead to retinal disease and the various types of corneal
dystrophy (CD) often display directly observable ocular manifestations to
the clinician. However, the considerable heterogeneity of inherited ocular
disease presents difficulty in choosing a treatment plan. Many of the
inherited, non-syndromic eye diseases are caused by the very specific spatial
expression of the mutated genes (Tucker, Mullins, and Stone 2014). To date,
more than 3000 genetic mutations in approximately 70 genes have been
associated with retinitis pigmentosa (RP) (Dias et al. 2018) and at least 8

The Application of CRISPR/Cas9 Therapies in Ophthalmology … 101
genes have been identified for diverse corneal CDs (Klintworth 2009,
Schorderet 2015). The International Committee of the Classification of
Corneal Dytrophies (IC3D) outlined genetic links to 22 different types of
CD (Weiss et al. 2015). A precision or personalised medicine that functions
to identify the genetic basis of a disease in the patient and treat it with a
tailored approach is the most effective way to treat these types of maladies.
Gene therapy is defined as either the introduction of exogenous DNA or
RNA into a patient's cells or the silencing of pathogenic gene mutations
within the patient’s DNA. Silencing endogenous gene mutations known as
gene editing is achieved either by gene knockout or mutation correction.
Genetics based therapeutic approaches include RNA interference (RNAi)
with short interfering RNA (siRNA), gene replacement and gene editing via
programmable nucleases. Gene replacement therapies, where a functional
copy of the gene to treat loss of function mutations commonly introduced
via viral transgene expression have translated to clinical trials for inherited
ocular disease of the retina. These therapies treat autosomal recessive
conditions (Parker et al. 2016, Weleber et al. 2016, Hauswirth et al. 2008,
Bainbridge et al. 2008, Bennett et al. 2016, Ghazi et al. 2016, Edwards et al.
2016, Michalakis et al. 2017, Dalkara et al. 2016). In this context, biallelic
pathogenic mutations in a disease-associated gene could be supplemented
with the wild type, functional version of the gene. However, this strategy
cannot be applied to dominant conditions, where the pathogenic gene needs
to be repaired or down-regulated. The Moore research group at Ulster
University has extensively studied the application of siRNA for treatment of
autosomal dominantly inherited genetic diseases of the skin and eye and
have clearly demonstrated the potential of therapeutic siRNA for the cornea
(Courtney, Atkinson, Allen, Moore, Walsh, Pedrioli, MacEwen, Pellegrini,
Maurizi, Serafini, et al. 2014, Courtney, Atkinson, Moore, et al. 2014, Allen
et al. 2013, Liao et al. 2011, McLean and Moore 2011, Atkinson et al. 2011,
Moore et al. 2011).
For clinical treatment of autosomal dominant diseases, gene editing
holds enormous potential. Gene editing makes use of programmable
nucleases to target DNA in a sequence specific manner inducing a double
strand break (DSB). The DSB allows the mutation within the DNA sequence

C. B. Tara Moore, Larry A. DeDionisio, Hila Roshanravan et al. 102
to either be removed by non-homologous end-joining (NHEJ) or repaired
via homology directed repair (HDR) (Rudin, Sugarman, and Haber 1989,
Plessis et al. 1992, Rouet, Smih, and Jasin 1994, Choulika et al. 1995,
Bibikova et al. 2001, Bibikova et al. 2003, Bibikova et al. 2002). Four
classes of programmable nucleases are currently used, meganucleases,
(Belfort and Bonocora 2014, Stoddard 2011) zinc finger nucleases (ZFNs),
(Urnov et al. 2010) transcription activator-like effector nucleases (TALENs)
(Bogdanove and Voytas 2011) and clustered regularly interspersed
palindromic repeats (CRISPR) associated nuclease, Cas9 (Jinek et al. 2012,
Cong et al. 2013, Mali, Yang, et al. 2013, Zetsche et al. 2015). CRISPR/Cas
systems are derived from an endogenous defence mechanism identified
within a diverse group of bacteria and archaea (Karginov and Hannon 2010,
Makarova et al. 2011, Makarova et al. 2015). Compared with the extensive
protein engineering required for the meganucleases and ZFNs,
CRISPR/Cas9 can be easily programmed with simple molecular cloning
techniques. Modifications have been engineered to enhance the
CRISPR/Cas9 complex for genome editing within mammalian cells
facilitating the use of CRISPR/CAS9 to treat various human diseases (Xu et
al. 2015, Schwank et al. 2013, Lee et al. 2015).
The ease of using the CRISPR system has resulted in it rapidly replacing
other programmable nucleases as the preferred tool for genome editing.
Gene editing with CRISPR/Cas9 also largely circumvents the limit of viral
vector capacity faced by gene-replacement therapies (Ran et al. 2015, Zuris
et al. 2015). This efficient technology is particularly attractive in
ophthalmology. The eye’s accessible anatomical position, immune privilege
and the blood-retinal barrier make it an ideal site for in vivo gene editing
(Liu, Tuo, and Chan 2011). It is foreseen that CRISPR/Cas based therapy
for ocular disease will pave the way for personalised medicine. This review
discusses the progress in the use of CRISPR/Cas systems for treating eye
disease with an emphasis on recent developments in the treatment of rare
and inherited disorders of the cornea.

The Application of CRISPR/Cas9 Therapies in Ophthalmology … 103
2. APPROACHES FOR GENE EDITING WITH CRISPR/CAS
Cas9 is a crucial component in the type II CRISPR/Cas system and
cleaves double-stranded DNA (dsDNA) in a sequence directed and specific
manner. The first step in this protein-DNA interaction is binding of the Cas9
nuclease to the DNA target site via a single guide (sg) RNA sequence that is
complementary to the target DNA and to a short segment of DNA adjacent
to the target region referred to as the protospacer adjacent motif (PAM).
Figure 1. Cas9 programmed by tracrRNA:crRNA duplex.
The PAM acts as a recognition site for Cas9 and is critical in its ability to
cleave the target DNA (Figure 1). Alteration of the 20 nucleotide spacer
sequence within the sgRNA, lying directly upstream of the PAM directs the
RNA-guided endonuclease to the desired target sequence (Jinek et al. 2012,
Xu et al. 2014). Different PAMs are recognised by Cas nucleases from
different bacterial species. The most characterised type II systems are S.

C. B. Tara Moore, Larry A. DeDionisio, Hila Roshanravan et al. 104
pyogenes Cas9, which recognises a 5–NGG–3′ PAM and S. aureus Cas9,
which recognises a 5–NNGRRT– 3’ PAM (Belfort and Bonocora 2014,
Mojica et al. 2009). The PAM-proximal region of the guide, often referred
to as the ‘seed region’ (Semenova et al. 2011) dictates the binding of Cas9
to the target site so mismatches in this region are critical in determining
specificity while mismatches in the distal region have less impact on the
specificity (Pattanayak et al. 2013, Sternberg et al. 2014, Wu et al. 2014).
There is no uniformity in the exact length of the seed region; however, the
studies suggest it is the first 5-12bp of the guide sequence that is critical.
2.1. Minimising off Target Cleavage and Improving Efficiency
There are limitations that can hinder applications of the CRISPR/Cas9
system such as inefficient delivery, dysregulation of the delivered gene, the
immune response against the CRISPR system, and off-target effects or the
unintended on-target mutations at sites that share identical or highly similar
sequence to the target locus (Cho et al. 2014, Fu et al. 2013, Hsu et al. 2013,
Pattanayak et al. 2013, Lin, Cradick, et al. 2014, Lee and Kim 2019). There
are studies that determined the mutation frequency in CRISPR/Cas9 edited
mouse and monkey models were not significantly higher than in wild type
control animals (Willi et al. 2018, Luo et al. 2018). These studies did not
rule out the possibility of rare mutations at genomic loci that possess
sequence homogeneity to the target sequence. To counteract these unwanted
genomic editing events, the sequence characteristics of the sgRNA, which
are critical for improving specificity and efficiency should be carefully
monitored. Guides with high cleavage efficiency are necessary within a
clinical setting in order to achieve therapeutic benefits. Several online tools
have been designed to allow in silico analyses of on- and off-target cleavage
of each guide sequence. Studies that utilized optimal guide design in
conjunction with genome-wide analyses have reported minimal off target
cleavage (Lin, Cradick, et al. 2014, Kim et al. 2015, Veres et al. 2014, Frock
et al. 2015, Tsai et al. 2017).

The Application of CRISPR/Cas9 Therapies in Ophthalmology … 105
Table 1. Different techniques to identify off-targets
Methods Description
linear amplification–mediated
high-throughput genome-wide
sequencing (LAM-HTGTS)
High coverage sequencing to assess the degree of
mutagenesis across the entire genome (Hu et al. 2016)
Di-genome seq in vitro Cas9-digested whole-genome sequencing (Kim et
al. 2015)
Circularization for in vitro
reporting of cleavage effects
by sequencing (CIRCLE-seq)
A highly sensitive, sequencing-efficient in vitro screening
strategy for identifying CRISPR-Cas9 genome-wide off-
target mutations (Tsai et al. 2017)
Breaks Labelling, Enrichment
on Streptavidin, and
Sequencing (BLESS)
The first method for direct labelling of DSBs in situ
followed by their genome-wide mapping at nucleotide
resolution (Crosetto et al. 2013)
Breaks Labelling In Situ and
Sequencing (BLISS)
1) Direct labelling of DSBs in fixed cells or tissue
sections on a solid surface
2) low-input requirement by linear amplification of
tagged DSBs by in vitro transcription
3) quantification of DSBs through unique molecular
identifiers
4) easy scalability and multiplexing (Yan et al. 2017)
Genome-wide Unbiased
Identification of DSBs
Enabled by Sequencing
(GUIDE-Seq)
Relies on capture of double-stranded
oligodeoxynucleotides into breaks (Tsai et al. 2015)
SITE-Seq A biochemical method, based on the selective enrichment
and identification of adapter-tagged DNA ends by
sequencing (Cameron et al. 2017)
Several methods have been devised to directly identify cleaved sites in
cells (Table 1) and these techniques have achieved good sensitivity and
resolution. However, at the present time there is no single “gold standard
method” which can accomplish unbiased, comprehensive off-target
analysis. To reduce the off-target activity of CRISPR/Cas, much focus has
been directed on improving target specificity of the system. Several
strategies to enhance Cas9 specificity have been reported (Table 2).

C. B. Tara Moore, Larry A. DeDionisio, Hila Roshanravan et al. 106
Table 2. Different strategies to enhance Cas9 specificity
Techniques Description
Reducing the amount of
active Cas9 in cells
Studies have shown increased off-target activities with high
levels of Cas9 protein (Fu et al. 2013, Hsu et al. 2013, Pattanayak
et al. 2013) and a number of techniques have been employed to
restrict the concentration of Cas9 protein such as delivering it as
a purified protein loaded with sgRNA (Cho et al. 2013, Kim et al.
2014) or limiting the duration and expression of Cas9, using
various inducible platforms (Davis et al. 2015, Zetsche, Volz,
and Zhang 2015).
Double nicking by Cas9
nickase mutants
Creates a pair of juxtaposed single-stranded DNA nicks to
enhance genome editing specificity (Ran et al. 2013, Mali, Aach,
et al. 2013).
Altering the sgRNA
sequence
Several studies showed that truncation of sgRNA leads to an
increased target specificity and reduced off target activity.
(Pattanayak et al. 2013, Fu, Reyon, and Joung 2014, Fu et al.
2014) However, it has been shown by our group that a range of
truncated guide lengths or addition of 5′-GG to the 20 nt guide
sequence did not provide a marked improvement of specificity
(Figure 2) (Christie et al. 2017).
Fusion of catalytically
inactive Cas9 to FokI
nuclease (fCas9)
Target DNA sites modified in human cells with >140-fold
higher specificity than wild-type Cas9 and the specificity of
fCas9 was around 4-fold higher than that of paired nickases at
loci with highly similar off-target sites (Guilinger, Thompson,
and Liu 2014, Tsai et al. 2014). More recent developments have
incorporated the Fokl-Cas9 system with truncated sgRNA to
further improve specificity and reduce off-target activities
(Wyvekens et al. 2015). Although each of these approaches
reduce off-target mutagenesis, they have a number of limitations:
reducing the amount of Cas9 can decrease on-target cleavage
efficiency and truncated guides can increase indel formation at
some off-target loci.
Improve the specificity
of Streptococcus
pyogenes Cas9
(SpCas9) is based on
structure-guided protein
engineering.
A study reported that neutralization of positive charges in the
non-target groove can dramatically decrease off-target indel
formation while preserving on-target activity (Slaymaker et al.
2016). In another report, engineering of additional high-fidelity
variants of CRISPR-associated nucleases by introducing
substitutions at additional non-specific DNA contacting residues
can further reduce some of the very small number of residual off-
target sites that persist for certain sgRNAs (Kleinstiver et al.
2016).

The Application of CRISPR/Cas9 Therapies in Ophthalmology … 107
In addition to the work carried out to reduce off-target effects, much
effort has been done to enhance the gene editing efficiency of the
CRISPR/Cas9 complex. The DSBs introduced by the enzyme are most
commonly repaired by the NHEJ pathway, leading to nonspecific nucleotide
insertions and deletions, referred to as ‘indels’.
This is convenient for generating gene knockouts but does not allow
introduction of specific sequence changes. The alternative path to repair,
HDR enables a recombination event between a homologous donor DNA
template and damaged DNA resulting in an accurate repair of the DSB.
Many groups have shown different strategies to improve the HDR rate of
repair over the NHEJ pathway (Zhang et al. 2017, Paquet et al. 2016, Kwart
et al. 2017, Liang et al. 2017). For example, the inhibition of DNA ligase
IV, the key enzyme utilized by NHEJ with the inhibitor Scr7 can be used to
promote HDR (Maruyama et al. 2015, Chu et al. 2015, Singh, Schimenti,
and Bolcun-Filas 2015). HDR is restricted to the G2 phase of cell division
when DNA replication is completed and sister chromatids are available to
serve as repair templates, (Lin, Staahl, et al. 2014) which limits the use of
this pathways in certain cell types such as ocular cell types where the cells
are primarily in the post-mitotic G0 state (Young 1985).
2.2. Expanding the CRISPR/Cas Protospacer Adjacent Motif
(PAM) Repertoire
Cas9 nucleases from various bacterial species have been shown to have
different PAM sequences (Table 3) including a newly characterised member
of the CRISPR programmable nuclease system, Cpf1 from the
Acidaminococcus and Lachnospiraceae bacteria, which was adapted for
genome editing in human cells (Zetsche et al. 2015). Its PAM motif, TTTN
was shown to provide the best cleavage efficiency. The expanding
CRISPR/Cas PAM repertoire will allow greater targeting across diverse
genomic regions. Moreover, it has been reported that artificially engineered
SpCas9 and SaCas9 with alternative PAM recognition motifs can be
generated (Kleinstiver, Prew, Tsai, Nguyen, et al. 2015, Kleinstiver, Prew,

C. B. Tara Moore, Larry A. DeDionisio, Hila Roshanravan et al. 108
Tsai, Topkar, et al. 2015). Thus, the application of directed evolution
technology to the development of Cas9 with varying PAM motifs will
expand the repertoire of targetable sequences throughout the human genome
and increase the range of accessible therapeutic targets.
Table 3. CRISPR/Cas Protospacer Adjacent Motif (PAM) repertoire
Source Nuclease type PAM sequence
Streptococcus
pyogenes
Cas9 NGG(Jinek et al. 2012)
Cas9 (D1135E) with
improved recognition and
speficity
NGG with (Kleinstiver, Prew,
Tsai, Topkar, et al. 2015)
Cas9 (“VQR”;
D1135V/R1335Q/T1337R)
NGAN or NGNG (Kleinstiver,
Prew, Tsai, Topkar, et al. 2015)
Cas9 (“EQR”;
D1135E/R1335Q/T1337R)
NGAG (Kleinstiver, Prew, Tsai,
Topkar, et al. 2015)
Staphylococcus aureus Cas9 NNGRRT or NNGRR(N) (Plessis
et al. 1992)
Cas9 (“KKH”;
E782K/N968K/R1015H)
NNNRRT (Kleinstiver, Prew,
Tsai, Nguyen, et al. 2015)
Neisseria meningitides Cas9 NNNNGATT (Hou et al. 2013)
Streptococcus
thermophiles
Cas9 NNAGAAW (Sapranauskas et al.
2011)
Francisella novicida Cpf1 TTN (Zetsche et al. 2015)
Acidaminococcus sp.
BV3L6
Cpf1 TTTN (Zetsche et al. 2015)
Lachnospiraceae
bacterium
Cpf1 TTTN (Zetsche et al. 2015)
Alicyclobacillus
acidoterrestris
C2c1 TTN (Shmakov et al. 2015)
IUPAC nucleotide code, where R refers to A or G; W to A or T; and N represents any nucleotide.
3. CRISPR-CAS9 GENE EDITING IN GENETIC
EYE DISEASE
The eye is an ideal site for testing of in vivo gene editing due to its
accessible position, compartmentalised anatomy and relative immune-
privilege. CRISPR-Cas9-based therapeutics provide the possibility for

The Application of CRISPR/Cas9 Therapies in Ophthalmology … 109
permanent treatment of genetic disease. Inherited eye diseases affect
millions of people worldwide and have an immense socioeconomic impact.
With our increasing understanding of the underlying molecular mechanisms
of ocular diseases, gene therapy, especially combined with adeno-associated
viruses (AAV) delivery, has been proposed as an effective approach. With
the advantages of CRISPR/Cas over alternate genome editing technologies,
the stage is set for a new wave of gene-based clinical trials. The benefits of
using CRISPR/Cas9 systems to study eye diseases have been highlighted by
several successful applications in a variety of cell types and animal species.
3.1. Applications for Retinal Eye Disease
Research into CRISPR/CAS9 gene editing as applied to ocular diseases
was initially focused on diseases that affect the retina. Hereditary retinal
diseases comprise the leading cause of blindness among working age adults
in the United Kingdom (Liew, Michaelides, and Bunce 2014). Retinal
diseases like proliferative diabetic retinopathy (PDR), retinopathy of
prematurity (ROP), and wet age-related macular degeneration (wAMD) are
known to occur from abnormal angiogenesis, the process by which new
blood vessels grow from pre-existing vessels (Abhinand et al. 2016).
Vascular endothelial growth factor (VEGFR) plays a critical role in
angiogenesis, and experiments utilizing CRISPR/Cas9 to edit VEGFR have
shown promise. Editing VEGFR2 in mouse models for oxygen-induced
retinopathy (OIR) and laser-induced choroid neovascularization (CNV) was
shown to abrogate angiogenesis (Huang et al. 2017). In addition, another
study showed the knockout of Vegfa and Hif1a genes by CRISPR-Cas9 to
block angiogenesis in a CNV model in mouse (Kim, Park, et al. 2017).
Subretinal injection of sgRNA- ribonucleoproteins (RNPs) complexes into
the adult mouse eye gave rise to mutagenesis at the target Vegfa gene in the
retinal pigment epithelium resulting in reduced CNV in an AMD mouse
model (Kim, Park, et al. 2017).
Targeted genomic deletion using the CRISPR/Cas9 system presents a
promising therapeutic approach for the treatment of patients with Leber's

C. B. Tara Moore, Larry A. DeDionisio, Hila Roshanravan et al. 110
congenital amaurosis 10 (LCA10) bearing the CEP290 splice mutation
(Ruan et al. 2017b). Loss of cone photoreceptors in RP leads to blindness,
and preservation of cone function is a major therapeutic goal. Sub-retinal
injection of AAV-delivered CRISPR/Cas9 prevented retinal degeneration in
mice by Nrl knockdown in rod cells (Yu et al. 2017). Furthermore, gene
editing using CRISPR/Cas9 has been shown to correct retinal dystrophy in
a rat model of autosomal dominant RP by generating allele-specific
disruption of RhoS334 (Bakondi et al. 2016).
3.2. Corneal Autosomal Dominant Genetic Eye Disease–
Allele Specificity
The majority of corneal dystrophies are the result of an autosomal
dominant inheritance pattern (Klintworth 2009). Others exhibit autosomal
recessive inheritance, while a few corneal dystrophies present X-linked
inheritance. To date, 23 distinct corneal dystrophies have been described.
Table 4 describes each corneal dystrophy, including; associated inheritance
pattern, gene locus and causative genes. Corneal dystrophies are
predominantly monogenic and highly penetrant. The accessibility of the
cornea, make the corneal dystrophies an ideal starting point for genome
editing therapies where concepts of personalized medicine can be put to use
(Moore et al. 2018).
Research into the use of CRISPR/Cas9 for the treatment of rare inherited
corneal disease has progressed during the past few years. We were the first
to demonstrate an allele specific CRISPR/Cas9 approach, cleaving the
mutant DNA at a SNP-derived PAM in the KRT12 gene in the mouse cornea
(Courtney et al. 2016). For this approach, we took advantage of the
discriminating nature of the PAM, the recognition site for the CAS enzyme,
which requires a precise sequence composition for the DSB to occur. When
a DNA mutation or single nucleotide polymorphism (SNP) is known to
cause disease also creates a PAM where none existed before, this mutation

The Application of CRISPR/Cas9 Therapies in Ophthalmology … 111
Table 4. Corneal dystrophies: associated inheritance pattern, gene
locus and causative genes
Corneal
layer
Disease Inheritance
Pattern
Genetic
Locus
Gene
known
Gene/s
Affected
IC3D Category
Epithelial and
Sub-Epithelial
Dystrophies
EBMD Minority of
cases, mostly
sporadic
5q13 Some cases TGFBI Some C1
ERED Autosomal
Dominant
Unknown Unknown N/A C4
(Smolandiensis
variant = C3)
SMCD Autosomal
Dominant
Unknown Unknown Unknown C4
MECD Autosomal
Dominant
12q13 and
17q12
Yes KRT3 and
KRT12
(Stocker–
Holt variant)
C1
LECD X-chromosomal
dominant
Xp 22.3 Unknown Unknown C2
GDCD Autosomal
Recessive
1p32. Yes TACSTD2,
previously
M1S1
C1
Bowman
Layer
Dystrophies
RBCD Autosomal
Dominant
5q13 Yes TGFBI C1
TBCD Autosomal
Dominant
5q13 Yes TGFBI C1
10q24 Unknown Unknown C2
GWCD Autosomal
Dominant
Unknown Unknown Unknown C4
Stromal
Dystrophies
LCD Autosomal
Dominant
5q13 Yes TGFBI C1
Autosomal
Dominant
9q34 Yes GSN C1
GCD Autosomal
Dominant
5q31 Yes TGFBI C1
MCD Autosomal
Recessive
16q22 Yes CHST6 C1
SCD Autosomal
Dominant
1p36 Yes UBIAD1 C1
CSCD Autosomal
Dominant
12q21.33 Yes DCN C1
FCD Autosomal
Dominant
2q35 Yes PIP5K3 C1
PACD Autosomal
Dominant
12q21.33 Yes KERA,
LUM,DCN,
EPYC
C3
CCDF Unknown Unknown Unknown Unknown C4
PDCD Reported AD Unknown Unknown Unknown C4
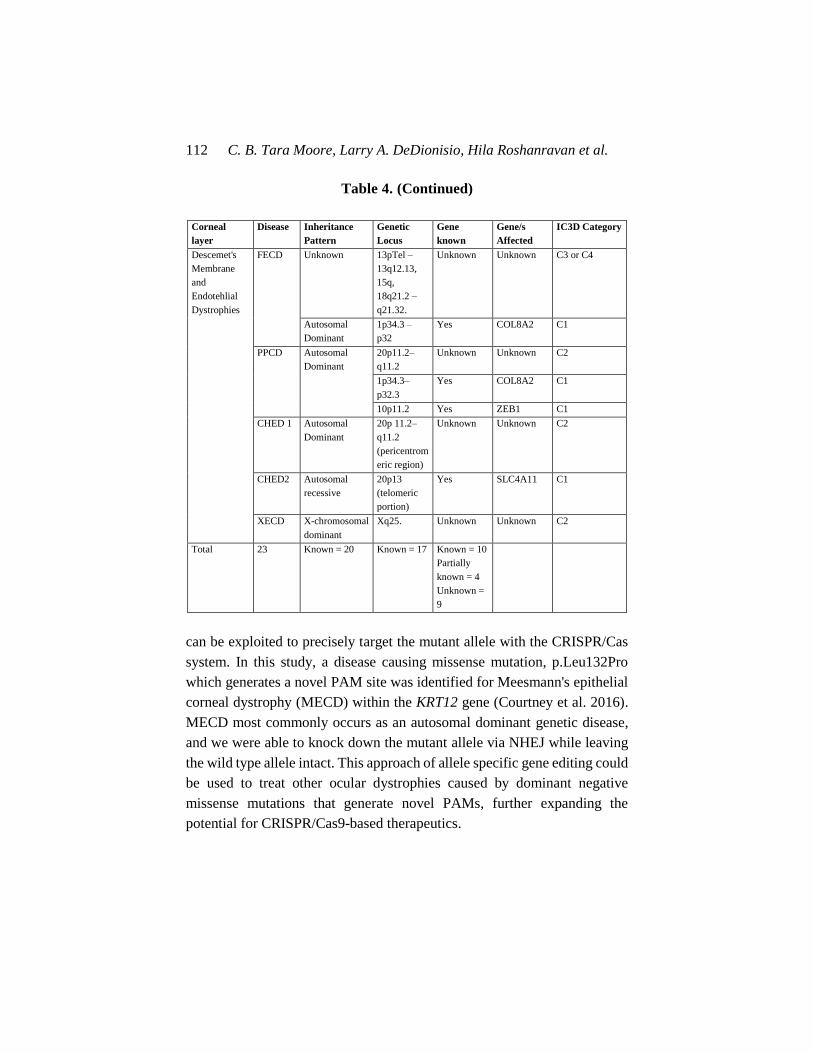
C. B. Tara Moore, Larry A. DeDionisio, Hila Roshanravan et al. 112
Table 4. (Continued)
Corneal
layer
Disease Inheritance
Pattern
Genetic
Locus
Gene
known
Gene/s
Affected
IC3D Category
Descemet's
Membrane
and
Endotehlial
Dystrophies
FECD Unknown 13pTel –
13q12.13,
15q,
18q21.2 –
q21.32.
Unknown Unknown C3 or C4
Autosomal
Dominant
1p34.3 –
p32
Yes COL8A2 C1
PPCD Autosomal
Dominant
20p11.2–
q11.2
Unknown Unknown C2
1p34.3–
p32.3
Yes COL8A2 C1
10p11.2 Yes ZEB1 C1
CHED 1 Autosomal
Dominant
20p 11.2–
q11.2
(pericentrom
eric region)
Unknown Unknown C2
CHED2 Autosomal
recessive
20p13
(telomeric
portion)
Yes SLC4A11 C1
XECD X-chromosomal
dominant
Xq25. Unknown Unknown C2
Total 23 Known = 20 Known = 17 Known = 10
Partially
known = 4
Unknown =
9
can be exploited to precisely target the mutant allele with the CRISPR/Cas
system. In this study, a disease causing missense mutation, p.Leu132Pro
which generates a novel PAM site was identified for Meesmann's epithelial
corneal dystrophy (MECD) within the KRT12 gene (Courtney et al. 2016).
MECD most commonly occurs as an autosomal dominant genetic disease,
and we were able to knock down the mutant allele via NHEJ while leaving
the wild type allele intact. This approach of allele specific gene editing could
be used to treat other ocular dystrophies caused by dominant negative
missense mutations that generate novel PAMs, further expanding the
potential for CRISPR/Cas9-based therapeutics.

The Application of CRISPR/Cas9 Therapies in Ophthalmology … 113
3.3. Mutant Allele Targeting, a Dual Cut Approach
There are at least 70 known missense mutations located in the coding region
of the TGFBI gene that can lead to various forms of corneal dystrophy. It is
neither possible nor realistic to design a sgRNA for each of these mutations,
and since at least 44 of the 70 mutations occur adjacent to a naturally
occurring PAM, stringent fidelity is required to achieve complete allele-
specificity to avoid the off target site that exists in the form of the wild-type
allele for these mutations (Christie et al. 2017). There have been studies
which demonstrated that truncating the sgRNA leads to an increased target
specificity and reduced off target activity (Pattanayak et al. 2013, Fu, Reyon,
and Joung 2014, Fu et al. 2014). However, it has been shown by our group
that a range of truncated guide lengths or addition of 5′-GG to the 20 nt
guide sequence did not provide a marked improvement of specificity for
a sub group of the TGFBI mutations (Figure 2) (Christie et al. 2017). We
therefore have conceived of a dual cut approach that in theory would achieve
knock down for all the TGFBI missense mutations applicable to
heterozygous genotypes (Christie KA 2019). With a dual cut approach, the
treatment of autosomal disease like TGFBI corneal dystrophy is not
constrained by the specific mutation responsible for any given phenotype.
Our approach utilises natural occurring variants in the target region
characterized by SNP-derived PAMs, which allow for the design of allele-
specific SNP-derived PAM sgRNAs (Christie KA 2019). Utilising natural
variants in the target region that harbour a PAM which also lie in cis with
the causative mutation allows this method to discriminate between the wild-
type and mutant alleles. Premature termination of translation via NHEJ
brings on permanent disruption of the target allele. A dual guide approach
to CRISPR/Cas9 editing was shown to be a promising therapeutic approach
for the treatment of patients with LCA10 bearing the CEP290 splice
mutation, (Ruan et al. 2017a) and in a human cell line it was demonstrated
that DNA deletions up to 10 kb coupled to precise NHEJ effectively knocked
down a mutant allele (Zheng et al. 2014). In order to generate a therapeutic
outcome for the TGFBI mutation targeted in our work, an in cis dual guide
approach was required (Christie KA 2019).


Figure 2. Guide-length screen to determine the effect on specificity of a guide-specific system. Reports have indicated that truncating the
length of the matching sequence within the guide from 20 to 18 nucleotides can reduce genome-wide off-target cutting, while
maintaining on-target efficiencies. An assessment of the effect of guide-length upon specificity between the wild-type and mutant
alleles, using a dual-luciferase assay, was conducted for the 5 most prevalent TGFBI mutations. Reports have shown that guide lengths
<16nt abolish cleavage activity. For each mutation a range of guide lengths from 16 to 22 nucleotides were tested, each guide was
targeted to the wild-type and respective mutant sequence and the firefly luciferase activity was measured as an indicator of specificity.
For all mutations investigated, the truncated guides did not provide a marked improvement of specificity; for most cases maximal
discrimination occurred with guides 20 or 19 nucleotides in length. For R124C, a 20nt guide seemed to confer allele-specificity,
however, no other guide length offered any adequate discrimination (Figure 2a). In the case of the R555Q mutation, guides in the 18–
20nt range did not offer sufficient discrimination, although, interestingly, the 21nt guide provided convincing allele-specificity (Figure
2d). R555W did not offer any considerable allele-specificity for any length tested (Figure 2e). R124H and R124L displayed clear allele-
specific cleavage, especially in the 18–20nt sgRNA range, with minimal cutting of the wild-type sequence (Figure 2b and c).
Interestingly for the R124 mutations guide lengths of 21nt seemed to impair cleavage activity in all cases.

C. B. Tara Moore, Larry A. DeDionisio, Hila Roshanravan et al. 116
While our study focussed on the issue of on-target allele-specificity in
relation to the TGFBI corneal dystrophies, for translation to the clinic a
number of key hurdles will need to be overcome, including genome wide
specificity and efficiency of delivery to the correct cells in the cornea. Potent
targeting of the correct cell population must be achieved. In corneal tissue,
the majority of TGFBI protein is produced in the epithelium, and the
epithelium is continually turned over and repopulated via the limbal
epithelial stem cells (LESCs). Thus, in order to permanently correct the
TGFBI corneal dystrophies efficient delivery to the LESCs must be achieved
(Christie KA 2019).
Another application for utilizing a dual cut CRISPR/Cas9 approach in
the context of the cornea involved a study aimed at understanding the
function of the PAX6 gene in maintaining the identity of human corneal
epithelial cells (CECs) (Kitazawa et al. 2017). PAX6 is a transcription factor
and is necessary for eye development. To investigate how PAX6 regulates
the gene expression profile of CECs, this study designed a two guided RNA
system targeting specific genomic loci to make a DSB that resulted in a
nonsense mutation upon NHEJ. This resulted in a complete knock down
PAX6 expression. The PAX6 knockout via the dual cut CRISPR/Cas9 system
resulted in a loss of CEC identity through the downregulation of corneal
epithelial-related genes and the upregulation of epidermis and
keratinization-related genes. Specifically, PAX6-depleted CECs resulted in
down-regulated genes that were primarily CEC-specific and included
keratin 12, keratin 3, clusterin (CLU), aldehyde dehydrogenase 3 family
member A1 (ALDH3A1), angiopoietin-like 7 (ANGPTL7) and transketolase
(TKT), while up-regulated genes were primarily epidermis-related and
included keratin 10, keratin 1, involucrin (IVL) and filaggrin (FLG). The
findings suggested that PAX6 maintains CEC identity by regulating
differentiation (Kitazawa et al. 2017).

The Application of CRISPR/Cas9 Therapies in Ophthalmology … 117
3.4. Multiplexing in the CRISPR System for Multigene
Ocular Disease
Complex genetic ocular diseases like primary open angle glaucoma
(POAG) or AMD where dysregulation at multiple loci contribute to the
overall disease phenotype could be treated by multiplexing the CRISPR
system. Likewise, retinal dystrophies are genetically heterogeneous diseases
(Berger, Kloeckener-Gruissem, and Neidhardt 2010, Roosing et al. 2014,
Zeitz, Robson, and Audo 2015). An advantage of the CRISPR/Cas system
over protein recognition-based methods is its ability to perform multiplex
genome editing at high efficiencies by expressing or providing multiple
guide RNAs (Cong et al. 2013, Mali, Yang, et al. 2013). Table 5 describes
current approaches for providing multiple gRNAs in vivo. More recently,
multiplexing was achieved with AAV-delivered Cas9 nuclease. A
quadruplex gRNAs/SaCas9 vector consisting of SaCas9 and multiplex
sgRNAs was successfully delivered using AAV-DJ/8 for in vivo excision of
HIV-1 proviral DNA in various solid tissues/organs via a single intravenous
injection in humanized bone marrow/liver/thymus (BLT) mice with chronic
HIV-1 infection (Yin, Zhang, et al. 2017).
A construct referred to as CRISPR-on was used in a study where an
exogenous reporter gene in mouse and human cells was activated enabling
the simultaneous activation of multiple endogenous genes (Cheng et al.
2013). The system utilized a nuclease-dead Cas9 (dCas9) protein fused with
a transcriptional activation domain and a single guide RNAs (sgRNAs) with
complementary sequence to gene promoters. The ability a system like
CRIRPR-on to regulate endogenous genes is valuable for the study of gene
functions and has great potential for therapeutic applications.
Another study found that by introducing different Cas9 proteins they
could mediate DNA cleavage or regulate transcriptional activity
simultaneously (Esvelt et al. 2013). Catalytically active Cas9 and distinct
sgRNA constructed with 14-15-bp target sequences and MS2 binding loops
activated gene expression without inducing DSBs. Co-expression of full

Table 5. Current approaches for providing multiple gRNAs
Multiplex CRISPR System Description
Multigene cassettes to express several
gRNAs
A lentiviral system was used to express a Cas9 variant, a reporter gene and up to four sgRNAs from
independent RNA polymerase III promoters incorporated in the vector with the Golden Gate cloning method.
Each sgRNA was efficiently expressed and was able to mediate multiplex gene editing (Kabadi et al. 2014).
The expression cassettes of gRNAs were tandemly ligated into a single vector using the Golden Gate cloning
method. Additionally, Cas9 nickase-mediated genome editing was constructed with a vector expressing Cas9
nickase and six gRNAs targeting three genomic loci (Sakuma, Sakamoto, and Yamamoto 2017).
Guide RNAs post-transcriptionally
processed from a single transcript using
Csy4-based excision
An RNA-based regulation and CRISPR/Cas transcription factors (CRISPR-TFs) tool utilizing multiple RNA
regulatory strategies, including RNA-triple-helix structures, introns, microRNAs, and ribozymes, with Cas9-
based CRISPR-TFs and Cas6/Csy4-based RNA processing gives the ability to modulate endogenous
promoters including multistage cascades and RNA-dependent networks (Nissim et al. 2014).
tRNA-dependent cleavage of gRNAs A general strategy to produce numerous gRNAs from a single polycistronic gene was utilized to boost the
targeting and multiplex editing capability of a CRISPR/Cas9 system. The study demonstrated that synthetic
genes with a tandemly arrayed tRNA–gRNA architecture, efficiently and precisely processed gRNAs to
desired 5′ targeting sequences in vivo and directed Cas9 to edit multiple chromosomal targets (Xie,
Minkenberg, and Yang 2015).
Arrays to express multiple crRNAs for Cpf1 Using an artificial CRISPR pre-crRNA array consisting of four spacers separated by direct repeats (DRs) from
the CRISPR locus of Francisella novicida (FnCpf1) and two Cpf1 orthologs with activity in mammalian cells,
Acidaminococcus Cpf1 (AsCpf1) and Lachnospiraceae Cpf1 (LbCpf1) multiplex gene editing was observed
(Zetsche et al. 2017).
Self-cleaving ribozyme flanked gRNAs Multiple gRNAs linked with self-cleaving ribozymes and/or tRNA could be simultaneously expressed from a
single U6 promoter to exert genome editing. The study provided a novel strategy to enable tissue-specific
expression of more than one gRNAs for multiplex gene editing (Xu et al. 2017).
Direct introduction of Cas9 proteins
preloaded with different gRNAs Cas9 RNPs
A method for the rapid synthesis of gRNA and for delivery of Cas9 protein/gRNA ribonucleoprotein
complexes (Cas9 RNPs) into a variety of mammalian cells through liposome-mediated transfection or
electroporation was described. This approach for multigene targeting in Jurkat cells resulted in two-locus and
three-locus indels in approximately 93% and 65% of the resulting isolated cell lines with a reduced off-target
cleavage rate (Liang et al. 2015).

The Application of CRISPR/Cas9 Therapies in Ophthalmology … 119
length normal sgRNA and truncated sgRNA fused with MS2 binding loops
can simultaneously cleave or activate transcription at the respective target
genes (Dahlman et al. 2015).
4. DELIVERY OF CRISPR/CAS TO THE EYE -
IN VIVO AND EX VIVO
Efficient and effective delivery of the CRISPR/Cas machinery to the
tissues or cells of interest is one of the most important aspects for translating
the therapy to the clinic. The physiology of the eye presents challenges for
the pharmacologist and formulation scientist for the design of efficient drug
delivery systems. Delivery of drugs to the targeted ocular tissues is restricted
by various precorneal, dynamic and static ocular barriers (Patel et al. 2013).
Figure 3. Delivery of Cas9-sgRNA complex. A. Viral vector based delivery B.
Physical method of microinjection based delivery C. Nanoparticle based delivery.
The outermost surface of the eye has many barriers that prevent entry of a
foreign substance. Biological barriers must be considered for any
CRISPR/Cas system to be delivered to the eye. Many avenues of delivery
have been explored, broadly sub-divided into viral and non-viral approaches

C. B. Tara Moore, Larry A. DeDionisio, Hila Roshanravan et al. 120
(Figure 3). An ideal vehicle for gene therapy should have a large capacity to
package all the CRISPR/Cas9 machinery, cell tropism for the specific cell
population of interest, high nucleus targeting efficiency along with minimal
immune response and cytotoxicity.
4.1. In Vivo Systems
A range of viral systems including adenoviruses, AAVs and
retroviruses, specifically lentivirus, has been investigated for their potential
to deliver genetic payloads to transduce various types of tissues, including
the eye (Kay 2011). Adenovirus has the advantage of a large packaging
capacity but initiates a substantial immune response; (Borras et al. 2001,
Tsubota et al. 1998) whereas, AAV provides efficient transduction with a
minimal immune response and persistence in the nucleus but has a limiting
packaging capability (Sharma et al. 2010, Zinn and Vandenberghe 2014).
Lentivirus has a considerable packaging capacity but can cause unwanted
integration (Athanasopoulos, Munye, and Yanez-Munoz 2017). Non-
integrating viruses like adenoviruses transiently express the transgene
making them ideal for applications with CRISPR/Cas system, as limited
Cas9 expression reduces the off target cleavages.
AAV has been the leading vector of choice for gene therapy, particularly
ocular gene therapy. Recently AAV technology has been deployed for
delivering CRISPR/Cas9 components to investigate gene function and
disease treatment in the mammalian eye (Huang et al. 2017, Ruan et al.
2017b, Yu et al. 2017, Kim, Koo, et al. 2017). AAV has been shown to
transfer genes successfully into photoreceptor cells and retinal pigment
epithelium (RPE) of the mouse retina with low cytotoxicity (Day et al.
2014). Sub-retinal injection of an AAV vector has been shown to be safe and
effective in animal studies, (Acland et al. 2001, Pang et al. 2006) and human
trials (Boye et al. 2013, Cideciyan et al. 2009, Jacobson et al. 2012,
MacLaren et al. 2014, Testa et al. 2013, Trapani et al. 2015, Pierce and
Bennett 2015). The use of AAV-2 based gene augmentation therapy for
RPE65-associated retinal degeneration gene therapeutics has been

The Application of CRISPR/Cas9 Therapies in Ophthalmology … 121
extensively tested in clinical trials (Pierce and Bennett 2015). However,
AAV-2 was found to be not as effective as other AAV serotypes for
transduction within the cornea, and as such a variety of other AAV vectors
have been shown to successfully transduce the cornea with varying success
(Sharma et al. 2010, Liu et al. 2008). A study reported that a hybrid serotype,
AAV-2/8, delivered by an intrastromal injection was able to transduce long
term expression of the transgene in the stromal keratocytes in both ex vivo
human and in vivo mouse models (Hippert et al. 2012). An intracameral
injection of the AAV hybrid serotype vector, AAV-2/9 was shown to
transduce corneal endothelial cells in mouse (O'Callaghan et al. 2017). In
our hands AAV- 2/9 was effective in transducing both the corneal epithelium
and corneal endothelium cells (Unpublished Data). Lastly, a novel synthetic
AAV has been reported to efficiently transduce corneal stroma and
endothelial cells following intrastromal injection in mouse (Wang et al.
2017).
With AAV vectors being the leading choice of gene therapy in
ophthalmology, different strategies have been applied to circumvent their
limiting packaging capacity. A dual AAV vector system was utilized to
deliver the the CRISPR components, the sgRNA and SpCas9 in two separate
AAV vectors and delivered in vivo to post mitotic photoreceptor cells in
experiments utilizing retinal degeneration mouse models (Yu et al. 2017).
Since the crystal structure of at least two Cas9 proteins complexed with
guide RNA and target DNA have been determined, (Nishimasu et al. 2014,
Yamano et al. 2016) the way has been paved for designing modifications to
the complex which could be used to overcome the payload restrictions of the
AAV vectors. Also, to limit an AAV mediated gene therapy’s persistent
transgene expression, the use of tissue-specific promoters or self-cleaving
vectors have been investigated. A tissue-specific reporter ensures expression
is localised to only the desired target tissue. In 2 clinical trials where LCA
was treated with gene replacement of RPE65, an AAV-2 vector with a
human RPE65 promoter, hRPE65 was used to minimise unwanted
expression in other tissues (Bainbridge et al. 2008). With respect to gene
editing with a CRISPR system, minimising expression of Cas9 may help to
reduce off-target effects (Moore et al. 2018). The use of a self-cleaving

C. B. Tara Moore, Larry A. DeDionisio, Hila Roshanravan et al. 122
system which involves introducing both a sgRNA targeted to the gene of
interest and a sgRNA targeted to the delivery vector could also be used to
minimise the period of transgene expression and off-target effects (Chen et
al. 2017, Moore et al. 2014, Petris et al. 2017).
As an alternative to non-viral delivery systems, broadly classified into
two sub categories are physical and chemical systems (Ramamoorth and
Narvekar 2015). Unlike viral vectors, non-viral systems are not limited by
packaging restrictions, and they have lower immunogenicity and toxicity
(Pack et al. 2005, Mintzer and Simanek 2008). Physical methods including
injection of naked DNA, electroporation, gene gun, ultrasound and
magnetofection utilise force to increase cell permeability (Mohan et al.
2005, Oliveira, Rosa da Costa, and Silva 2017). Chemical delivery methods
employ the principal of electrostatic interactions between the negatively
charged nucleic acid and either a natural or induced positive charge of the
carrier, generating a complex with an overall positive charge (Jin et al.
2014). Through carrier design alteration and by targeting the ligand
decoration on the cell types, non-viral vectors can deliver therapeutic nucleic
acids to target tissues or cells. Several naturally occurring and synthetic
cationic polymers have been used for the delivery of nucleic acids. A novel
hyaluronic acid-chitosan nanoparticle was used to transfect efficiently the
corneal and conjunctival cells in rabbits in vivo and in a human in vitro
model, (de la Fuente, Seijo, and Alonso 2008a, b) and ultrapure chitosan
oligomers were used as carriers for transferring a luciferase plasmid to
keratocytes in rat cornea (Klausner et al. 2010). Interestingly, another report
developed an artificial virus using a nanoparticle, liposome-protamine-DNA
complex (LPD), modified with a cell permeable peptide and a nuclear
localization signaling (NLS) peptide, to deliver a functional Rpe65 gene to
mouse eyes improving vision in vivo (Rajala et al. 2014). Thus, LPD
nanoparticles provide a promising, efficient, non-viral method of gene
delivery with clinical applications.
Other promising delivery systems utilising the advantages of non-viral
delivery are CRISPR/Cas9 RNP complexes (Zuris et al. 2015) and CRISPR-
Gold which is able to simultaneously deliver RNPs and donor DNA into
cells resulting in successful in vivo HDR (Lee et al. 2017). A recent work

The Application of CRISPR/Cas9 Therapies in Ophthalmology … 123
demonstrated that a structure-guided chemical modification of guide RNA
delivered using a non-viral lipid nanoparticle resulted in a >80% gene
editing of Pcsk9 (Yin, Song, et al. 2017). The advantage of using a direct
protein-packaged approach is that components of the CRISPR complex have
a short half-life and are not being continuously synthesized in the cell once
the genomic modification has occurred. As no delivery system on its own
meets all requirements, therapeutic genome editing could be best achieved
by combining a viral coupled to non-viral delivery. For example, Cas9
mRNA delivered using lipid nanoparticles along with AAV encoding a
sgRNA and a repair template has been shown to induce repair of Fah in a
mouse model (Yin et al. 2016). These studies pave the way for more efficient
use of alternative in vivo delivery systems for gene editing especially in the
eye.
4.2. Ex Vivo Systems
Stem cell therapies for the treatment of AMD and other degenerative
retinal disorders including inherited retinal disease are positioned to make
an impact on clinical care (Wood et al. 2019). Stem-cell-based photoreceptor
cell replacement has been investigated in different cell types and in animal
models. The CRISPR/Cas9 system was successfully employed in embryonic
stem cells (ESCs) where efficient gene correction was demonstrated in a
mouse cataract model (Wu et al. 2013). A study utilizing human embryonic
stem cells (hESCs) demonstrated the ability of a hESC-derived retina to
form structured mature photoreceptor layers after transplantation into nude
rats and two different monkey models (Shirai et al. 2016). Human retinal
progenitor cells (hRPCs) from fetal neural retina were transplanted into a rat
model where at 12 weeks after treatment, hRPC-grafted eyes had
significantly superior visual acuity compared to non-treated eyes (Luo et al.
2014). For the study of optic nerve biology and disease, a CRISPR
engineered reporter cell line was able to demonstrate the differentiation of
hESCs to retinal ganglion cells (RGCs) (Sluch et al. 2015). Although hESCs
offer extensive differentiation capabilities allowing for the study of very

C. B. Tara Moore, Larry A. DeDionisio, Hila Roshanravan et al. 124
early stage development of life as well as providing a source of cells for the
development of novel therapeutic molecules and cell-based therapies, their
use is controversial. Due to the ethical and practical reasons of using cells
derived from embryonic or fetal tissue, much work and effort has been
conducted with patient-derived induced pluripotent stem cells (iPSCs),
which offer the best possible immunologic match to the patient (Burnight et
al. 2017).
The use of iPSCs in gene editing technologies hold great promise in the
treatment of inherited disease. CRISPR/Cas tools enable precise
manipulation of these stem cells for ophthalmic applications providing a
platform for correction of deleterious mutations in a patient's own cells. For
three different types of variants that lead to retinal diseases, CRISPR/Cas9
mediated genome editing strategies were used to target and correct the
mutations in patient-derived iPSCs. A HDR strategy targeting an Alu
insertion in the MAK gene was able to restore the retinal transcript and
protein; the CRISPR-Cas9-mediated NHEJ approach was able to excise the
IVS26 cryptic-splice mutation in CEP290 demonstrating a correction of the
transcript and protein in patient iPSCs; and lastly, with allele-specific
CRISPR guides, the Pro23His rhodopsin (RHO) allele was selectively
targeted which, following delivery to both patient iPSCs in vitro and pig
retina in vivo, created a frameshift and premature stop preventing
transcription of the disease-causing variant (Burnight et al. 2017).
Additionally, in patient-specific iPSCs an RPGR point mutation that causes
X-linked RP was precisely repaired (Bassuk et al. 2016).
While iPSC driven ex vivo therapy for inherited corneal disease has not
yet been realized, the potential and the development of cell-based therapies
using stem cells represent a significant breakthrough in treatment options in
regenerative medicine for the ocular surface. There have been several studies
focused on differentiating iPSCs into corneal epithelial cells for the
treatment of limbal stem cell deficiency (LSCD) (Saghizadeh et al. 2017).
One such study was able to successfully induce corneal epithelial cells from
both human adult corneal limbal epithelial cells (HLEC)-derived iPS cells
and human adult dermal fibroblast (HDF)-derived iPS cells by the stromal
cell-derived inducing activity (SDIA) differentiation method (Hayashi et al.

The Application of CRISPR/Cas9 Therapies in Ophthalmology … 125
2012). Another study was able to derive iPSCs from human primary LSCs
and re-differentiate these iPSCs back into the limbal corneal epithelium
(Sareen et al. 2014). Our group previously reported the ability to culture
LESCs from a patient with MECD. The causative mutation, p.Leu132Pro in
the KRT12 gene in these cultures was effectively silenced using siRNA
(Courtney, Atkinson, Allen, Moore, Walsh, Pedrioli, MacEwen, Pellegrini,
Maurizi, and Serafini 2014).
On-going research with corneal cell models has advanced the potential
for the CRISPR/Cas9 complex to be delivered ex vivo for the treatment of
inherited corneal disease like the TGFBI corneal dystrophies. Efficient
corneal transduction resulting in transgene expression via intra-stromal
injection of an AAV2/8 vector was successful in human corneal explants
(Hippert et al. 2012) An AAV8/9 chimeric capsid was also shown to
transduce human corneal explants in a gene replacement study for the
treatment of cornea clouding brought on by mucopolysaccharidosis type 1
(MPS1) (Vance et al. 2016). While an AAV approach to cellular
transduction has shown promise, there are alternative methods to AAV and
viral transduction. The variant within the TGFBI gene that causes granular
corneal dystrophy type 2 (GCD2) was repaired in human corneal keratocytes
via a CRISPR/Cas9-induced HDR mechanism (Taketani et al. 2017). In this
work a plasmid vector, px458 which enabled bicistronic expression of
SpCas9 and green fluorescence protein (GFP) was co-transfected into
primary GCD2 mutant human corneal keratocytes with a single stranded 100
base oligonucleotide (ODN) that served as donor repair template. This study
was the first to demonstrate in vitro gene correction in mutant human
primary corneal cells using CRISPR/Cas9 and HDR (Taketani et al. 2017).
The importance of an ex vivo approach to the delivery of therapeutic
cells containing the CRISPR/Cas9 complex (Figure 4) for the treatment of
autosomal dominant disease such as GCD2 and other TGFBI corneal
dystrophies is crucial when NHEJ resulting in a knock down of the mutant
allele is not a viable option. We have established that the TGFBI protein is
necessary for the wound healing process (Christie KA 2019). As such,
individuals who carry homozygous mutations for an autosomal dominant
TGFBI corneal dystrophy cannot be treated with allele specific

C. B. Tara Moore, Larry A. DeDionisio, Hila Roshanravan et al. 126
CRISPR/Cas9 NHEJ gene editing since, with this method both alleles
expressing the TGFBI transcript would be silenced. HDR within a corneal
stem cell model could potentially be used to replace mutant corneal cells
within an individual who carries the mutation in both copies of the TGFBI
gene. However, ex vivo stem cell transplantation of gene edited corrected
cells via HDR is dependent on removing all resident mutations carrying
LESCs, and for those TGFBI mutations resulting in an intrastromal
pathology, stem cell transplantation will not undo existing stromal
pathology. As such, a corneal dystrophy such as MECD which affects the
anterior corneal epithelium (McLean and Moore 2011) is well suited for ex
vivo stem cell replacement. This approach would be most advantageous for
targeting the MECD SNP-derived PAM mutation, p.Leu132Pro in the
KRT12 gene (Courtney et al. 2016).
Figure 4. CRISPR therapeutic approach; transition to clinics.
5. ANIMAL MODELS FOR EYE DISEASE
The generation of new model systems with genome-editing tools, in
particular those based on CRISPR/Cas9, are opening new avenues for both
research into, and for, the treatment of eye disease. Genome-editing
technologies are allowing researchers to take full advantage of both animal
and cellular models, and to work more easily with non-traditional model
organisms for eye research. In ophthalmology, these animal models are

The Application of CRISPR/Cas9 Therapies in Ophthalmology … 127
important first steps for examining genetic conditions as well as assessing
therapeutics for such genetic diseases. For generating a transgenic animal
model, the CRISPR/Cas9 system has multiple advantages over the ZFN or
TALEN approach to gene editing. The gRNA determines the specificity of
CRISPR systems and not the nuclease; therefore, the CAS protein can
remain unchanged between target sites and between different organisms.
Adjusting the target with a change to the gRNA sequence allows for higher
efficiencies when compared to systems that require protein alterations or
changes to embryonic stem cells (ES). Since homologous recombination is
generally required to insure a site‐specific edit within ES cells, and given
that a DSB can increase the probability of homologous recombination,
CRISPR/Cas9 increases specificity and site‐specific genome targeting
frequency in view of the system’s effectiveness in producing DSBs (Zarei et
al. 2019). The emergence of this technology has allowed the genome editing
in mouse embryos to be quicker and more cost-efficient (Yang et al. 2013).
In addition, CRISPR/Cas9 has revolutionized the generation of a wide
variety of transgenic animals such as zebrafish, (Ablain et al. 2015) rats,
(Bakondi et al. 2016) monkeys, (Zuo et al. 2017) pigs, (Yan et al. 2018) and
rabbits (Yuan et al. 2016).
Zebrafish represent a powerful model for studying vertebrate eye
development. Easy access to eye tissue for imaging and drug/gene delivery,
short lifecycle, and genetic similarities to higher vertebrates and mammals
gives zebrafish many advantages over other model organisms to explore the
applications of CRISPR/Cas gene editing in the study of important pathways
and regulators in the development of ocular tissue (Richardson et al. 2017).
Several studies demonstrate the power of CRISPR/Cas genome editing in
zebrafish. CRISPR/Cas gene editing was applied in the zebra fish model to
understand the role of membrane frizzled related protein (MFRP) in the
development of nanopthalmic-hyperopia; (Collery et al. 2016) SpCas9-
generated knockouts and morpholinos to show the importance of NeuroD in
photoceptor regeneration; (Taylor et al. 2015) and to show retina
regeneration and Müller glia de-differentiation (Yin et al. 2015, Serifi et al.
2016). In another non-mammalian animal model, an amphibian, Xenopus
tropicalis was used to create a highly penetrant and rapid retinoblastoma

C. B. Tara Moore, Larry A. DeDionisio, Hila Roshanravan et al. 128
(Rb) model. CRISPR/Cas9 gRNAs were injected into X. tropicalis embryos
for rapid identification of targets for therapeutic intervention (Naert et al.
2016). These studies demonstrate the important role of CRISPR/Cas genome
editing in animal models like the zebrafish for investigating diseases of the
human eye.
CRISPR/Cas systems are also applied to mammalian models of the eye.
The following examples focus on research conducted with mouse models
for the treatment of different eye diseases. In a study of a particular LCA
model, CRISPR-engineered mosaicism reveals that the loss of Kcnj13
function in mice mimics the human disease phenotypes (Zhong et al. 2015).
RP is a challenging retinal dystrophy for ophthalmologists. A mouse model
mimicking clinical phenotypes of RP was generated using CRISPR like
knock-in with the p.Leu135Pro, an RP-associated variant (Arno et al. 2016).
The most commonly studied preclinical model of RP called the “rodless”
(rd1) mouse is homozygous for two mutations, a nonsense point mutation
(Y347X) and an intronic insertion of a leukemia virus (Xmv-28) (Wu et al.
2016). Utilizing a CRISPR/Cas9 system, it was demonstrated that the
Y347X mutation is the causative variant in this form of RP. This group was
the first to demonstrate homology-directed recombination–mediated gene
correction in the visual system using the CRISPR/Cas9 system (Wu et al.
2016). Our research on MECD utilized a humanized mouse containing a
mutant KRT12 allele mutation, p.Leu132Pro (Courtney et al. 2016).
Through CRISPR/Cas9 directed NHEJ repair resulting in frame-shifting
deletions within the mutant KRT12 allele, we were able to demonstrate
reduction of the mutant KRT12 mRNA expression. The CRISPR/Cas9
system has also been successfully employed in stem cell lines where
efficient gene correction was demonstrated in a mouse cataract model (Wu
et al. 2013).
Pre-clinical studies in other CRISP/Cas9 edited mammalian models
include rabbits and pigs. A group studying GJA8, a gene known to be
involved in maintaining lens opacity and proper lens development, created
a gene knockout via a co-injection of Cas9/sgRNA mRNA into rabbit
zygotes. Their results showed that gene mutation efficiency in the GJA8
locus reached 98.7% in the rabbit embryos and that impaired GJA8 function

The Application of CRISPR/Cas9 Therapies in Ophthalmology … 129
caused microphthalmia, small lens size and cataracts; knocking out GJA8 in
rabbit via CRISPR/Cas9 system caused human-like cataracts (Yuan et al.
2016). In research involving neurodegenerative Huntington’s disease (HD),
a large CAG repeat (150 CAGs) via somatic cell nuclear transfer (SCNT) in
combination with CRISPR/Cas9, was inserted into the endogenous pig HTT
gene generating a HD knockin (KI) pig model that expressed full-length
mutant HTT at the endogenous level (Yan et al. 2018). HD results from this
monogenetic CAG repeat expansion located in exon1 of the HTT gene.
There are advantages to using large mammals such as pigs over rodent and
rabbit models. Pigs are genetically, anatomically, and physiologically closer
to humans, and their fast breeding period and large litter size hold
advantages over non-human primates when considering the time line of
generating large animal models of human diseases (Yan et al. 2018).
6. RECENT PROGRESS IN CLINICAL GENE THERAPY
TRIALS FOR OCULAR DISEASES
Research for transitioning effective gene therapy strategies for ocular
disease to the clinic has rapidly grown in the past 15 years, particularly for
the treatment of retinal diseases. Clinical trials involving retinal gene therapy
are creating hope for future therapies for afflicted patients. They all employ
gene augmentation for loss of functions diseases, rather than gene editing
strategies. The most successful example of ocular gene therapy is by gene
replacement using AAV delivery (Pierce and Bennett 2015). It has been used
successfully for RPE65, Leber's congenital amaurosis 2 (LCA2), an early
onset form of autosomal recessive retinal degeneration caused by mutations
in the RPE65 (RPE-specific 65 kDa protein) gene. Ground breaking trials
utilising AAV vectors to deliver RPE65 did show safety and initial vision
improvements (Bennett et al. 2016, Jacobson et al. 2015). These studies were
the first to demonstrate proof of concept for gene therapy in human retinal
disease. Three separate phase I–II clinical trials were initiated, which yielded

C. B. Tara Moore, Larry A. DeDionisio, Hila Roshanravan et al. 130
promising results after subretinal administration of AAV2-hRPE65 vectors
(Bainbridge et al. 2008, Hauswirth et al. 2008, Weleber et al. 2016).
The three studies differed in terms of dose, inclusion criteria, type of
promoter, location of the injection and outcome measures. All the studies
reported high levels of safety and demonstrated efficacy up until the 3-year
follow-up. In 2018 subsequent results were reported for the trial involving
the AAV vector expressing RPE65 (rAAV2-CB-hRPE65) (Weleber et al.
2016, Pennesi et al. 2018). The outcome after 5 years was found to be small
and not sustained; however, there were no clinically significant adverse
events. The group concluded that treating patients at a younger age is
associated with better visual function outcomes during 5 years after
treatment (Pennesi et al. 2018). In a follow up study for the trial
administering the rAAV2/2 RPE65 vector, (Bainbridge et al. 2008) results
indicated that the amount of rAAV2/2 RPE65 required to drive the visual
cycle in affected persons was not met to the extent required for a durable,
robust effect (Bainbridge et al. 2015). Gene therapy for LCA is the first
ocular gene therapy to complete a phase III clinical trial for patients with
confirmed biallelic RPE65 mutation-associated retinal dystrophy (Jiang, Xu,
and Tsang 2018).
Another gene therapy presently at the pre-clinical trial stage, targets
wAMD with an AAV vector encoding aflibercept (ADVM-022) (Grishanin
et al. 2019). Vascular endothelial growth factor (VEGFA) plays a key role
in the development of wAMD, and as such VEGFA is targeted in the
treatment of the disease. Administering recombinant anti-VEGFA proteins
such as aflibercept with intravitreal (IVT) injections every 4 to 8 weeks is
an approved therapy for this disease. Aflibercept is a recombinant chimeric
protein consisting of the VEGFA binding portion of human VEGFR-1 and
VEGFR-2 fused to the Fc portion of human IgG1 immunoglobulin (Ashraf
and Souka 2017). ADVM-022 utilizes a variant of the AAV2 capsid as a
vector for delivering and encoding aflibercpt. It was reported that the
administration of ADVM-022 in non-human primates was tolerated with no
serious adverse safety-related findings and that more than one year past a
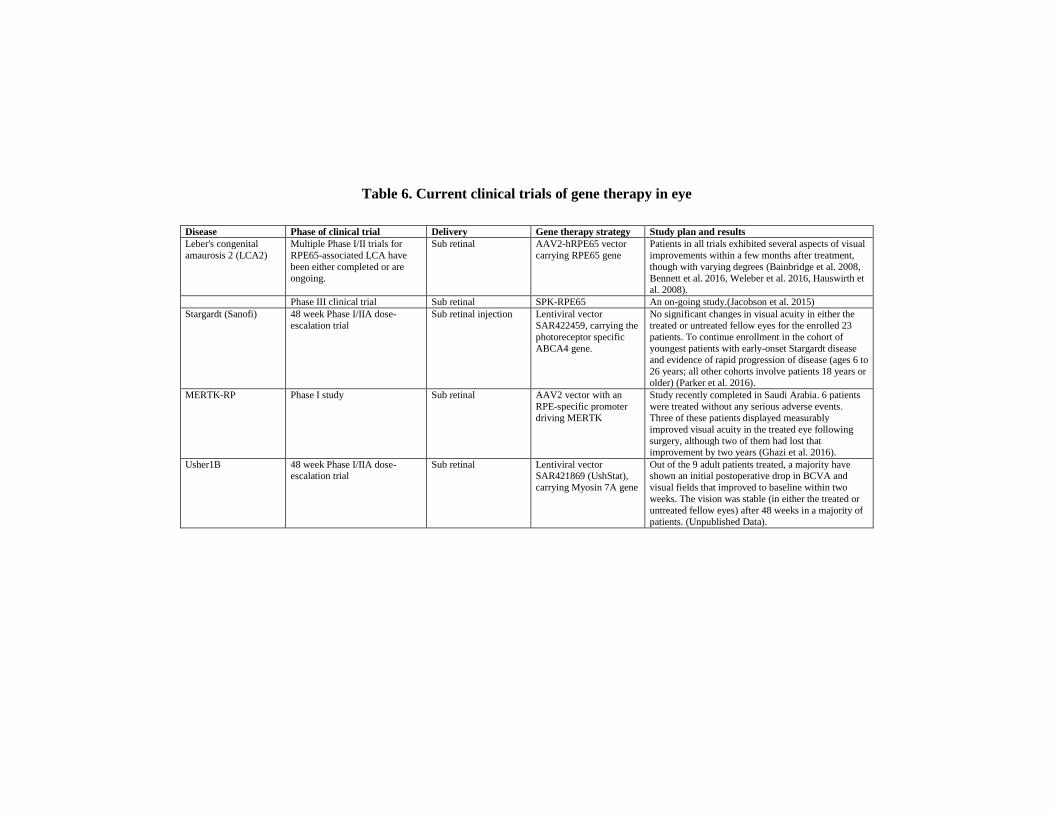
Table 6. Current clinical trials of gene therapy in eye
Disease Phase of clinical trial Delivery Gene therapy strategy Study plan and results
Leber's congenital
amaurosis 2 (LCA2)
Multiple Phase I/II trials for
RPE65-associated LCA have
been either completed or are
ongoing.
Sub retinal AAV2-hRPE65 vector
carrying RPE65 gene
Patients in all trials exhibited several aspects of visual
improvements within a few months after treatment,
though with varying degrees (Bainbridge et al. 2008,
Bennett et al. 2016, Weleber et al. 2016, Hauswirth et
al. 2008).
Phase III clinical trial Sub retinal SPK-RPE65 An on-going study.(Jacobson et al. 2015)
Stargardt (Sanofi) 48 week Phase I/IIA dose-
escalation trial
Sub retinal injection Lentiviral vector
SAR422459, carrying the
photoreceptor specific
ABCA4 gene.
No significant changes in visual acuity in either the
treated or untreated fellow eyes for the enrolled 23
patients. To continue enrollment in the cohort of
youngest patients with early-onset Stargardt disease and evidence of rapid progression of disease (ages 6 to
26 years; all other cohorts involve patients 18 years or
older) (Parker et al. 2016).
MERTK-RP Phase I study Sub retinal AAV2 vector with an
RPE-specific promoter
driving MERTK
Study recently completed in Saudi Arabia. 6 patients
were treated without any serious adverse events.
Three of these patients displayed measurably
improved visual acuity in the treated eye following
surgery, although two of them had lost that improvement by two years (Ghazi et al. 2016).
Usher1B 48 week Phase I/IIA dose-escalation trial
Sub retinal Lentiviral vector SAR421869 (UshStat),
carrying Myosin 7A gene
Out of the 9 adult patients treated, a majority have shown an initial postoperative drop in BCVA and
visual fields that improved to baseline within two
weeks. The vision was stable (in either the treated or
untreated fellow eyes) after 48 weeks in a majority of
patients. (Unpublished Data).

Table 6. (Continued)
Disease Phase of clinical trial Delivery Gene therapy strategy Study plan and results
X-linked Retinoschisis
(XLRS)
48 week Phase I/IIA safety and
efficacy trial
Intravitreal AAV8-RS1 vector
carrying the gene
retinoschisin 1(RS1)
A study evaluating three different increasing dose
levels of an AAV8-RS1 vector in 9 participants with
pathogenic RS1 mutations with VA of 20/63 or worse
in one eye (NCT02317887) (Cukras et al. 2018). The second study is evaluating an AAV-RS1 vector in
up to 27 patients. It involves three initial groups of
adult patients receiving increasing dose levels of the
vector and will also evaluate the maximum tolerated
dose level in patients 6 years and older
(NCT02416622).
Choroideremia Multiple Phase I and II trials of
the AAV.REP1 vector are
ongoing at several sites
Sub retinal AAV-REP1 vector
carrying RAB escort
protein 1 gene
In all patients, the increase in retinal sensitivity over
six months in the treated eyes correlated with the
vector dose administered per area of surviving retina.
(Cideciyan et al. 2009). The early improvement
observed in two of the six patients was sustained at
3.5 years after treatment despite progressive
degeneration in the control eyes. (Edwards et al.
2016). Other trials of subretinal placement of the AAV.REP1 vector are ongoing, including a Phase I/II
trial (NCT02341807).
Achromatopsia Phase I/II dose-escalation study Sub retinal AAV-CNGB3 vector
carrying CNGB3 gene
The study is going on in patients with CNGB3
achromatopsia at four sites in the United States
(NCT02599922).
Age-related Macular
Degeneration (AMD)
Phase 1 study Intravitreal AAV2-sFLT01 The treatment was safe with no dose-limiting toxicity
(Heier et al. 2017).
Leber Hereditary Optic
Neuropathy (LHON)
Two Phase 3 trials, in Europe
and the U.S evaluating the
effectiveness of GS010 in early
onset LHON patients
Intravitreal rAAV2/2-ND4 (GS010) A study for patients with vision loss of six months or
less (NCT02652767), and another study is assessing
patients whose vision loss began more than six
months but less than a year ago (NCT02652780).

The Application of CRISPR/Cas9 Therapies in Ophthalmology … 133
single IVT injection, ADVM-022 continued to provide robust aflibercept
expression. The effects of the ADVM-022 treatment were found to be
similar to a bolus treatment of aflibercept protein (Grishanin et al. 2019). A
single dose injection has clear advantages over a frequent dosing schedule
where lack of patient compliance may result in recurrence of active wAMD.
Gene-based therapy for wAMD has the potential to overcome such
compliance issues by ideally providing a life-long supply of the anti-
VEGFA protein following a single vector administration.
The remarkable success of the RPE65 and ADVM-022 studies has
paved the way for other clinical trials, particularly using AAV-mediated
gene delivery; however, there are challenges in delivering an effective and
enduring dose to stop cell death or degeneration. Active clinical gene
replacement trials are underway targeting other retinal diseases such as
Stargardt's disease, RP, Usher syndrome, X-linked retinoschisis (XLRS),
choroideremia, achromatopsia, AMD and Leber hereditary optic neuropathy
(LHON) (Table 6). These trials use either AAV or lentivirus as viral vectors.
7. CRISPR/CAS GENE EDITING TRANSITION TO CLINICS
CRISPR/Cas9 gene editing has undoubtedly shown great promise for
applications in gene correction therapy. The most imminent reason for
transitioning CRISPR/Cas research to clinics is direct therapeutic gene
editing in human cells, an advantage over viral-mediated gene therapy.
While no ocular gene therapy trials using CRISPR are yet underway, a
CRISPR gene editing technique has been approved by the National Institutes
of Health, US (NIH) to help extend cancer therapies that rely on enlisting a
patient’s T cells. Early in 2018 doctors at the University of Pennsylvania
were in the final steps of preparing for a clinical trial, and in 2019 two cancer
patients who are enrolled in this study were treated (Dearment 2019). This
trial is small and designed to investigate whether CRISPR is safe for use in
people. In the study, doctors will remove people’s blood cells, modify them
with CRISPR in the lab, and then infuse them back into the patients. The
study involves three different types of cancer: multiple myeloma, sarcoma,

C. B. Tara Moore, Larry A. DeDionisio, Hila Roshanravan et al. 134
and melanoma and proposes to remove T cells from at least 18 patients. Led
by Dr. Edward Stadtmauer, the protocol reprograms a person’s immune cells
to find and attack tumors. University of Pennsylvania scientists intend to use
CRISPR to edit two genes in patients’ T cells to make them better cancer
fighters. One of the genes to be edited, PD-1 makes a molecule that cancer
cells utilize to block the immune system from fighting the cancer. An
additional edit removes a natural T-cell protein that could interfere with this
process and in its place, an engineered receptor will steer the immune cells
toward particular tumors (Baylis and McLeod 2017) (Figure 4).
Similar studies have been under way in China where another trial is
making the first-ever attempt to use CRISPR to edit cells in the human body
infected with the human papillomavirus (HPV) while leaving the DNA of
normal cells untouched. (Figure 4). CRISPR would target and destroy the
viral genes of HPV that cause tumour growth. This study is to begin at the
First Affiliated Hospital of Sun Yat-Sen University in China (Le Page 2017).
In addition to this study, a research team in China edited the embryos of twin
girls born in November, 2018. In a highly controversial undertaking, a team
at the Southern University of Science and Technology in Shenzhen
purportedly eliminated the gene CCR5 via CRISPR editing in the hopes of
rendering the offspring resistant to HIV, smallpox, and cholera. If proven
successful, this effort would mark a stunning medical achievement;
however, this study and any future studies of this kind are ethically charged
because changes to an embryo could eventually affect the entire gene pool
if future generations were to inherit the edited gene. It goes without saying,
researchers and medical practitioners should adhere to ethical guidelines
such as those put forward by the Academy of Sciences in the US. Currently,
the use of genetically engineered embryos to establish a pregnancy is
prohibited in much of Europe, the United States and China (Regalado 2018).
The use of CRISPR is also under development for treating ocular
diseases. Editas Medicine and Allergan Pharmaceuticals International
Limited (Allergan) plan to initiate patient screening for clinical trials to test
the efficacy of EDIT-101, a CRISPR genome editing drug for the treatment
of Leber congenital amaurosis 10 (LCA10). They plan to enroll 10-20
patients in the U.S. and Europe. Utilizing an AAV5 viral vector that encodes

The Application of CRISPR/Cas9 Therapies in Ophthalmology … 135
for the Staphylococcus aureus Cas9 along with two different gRNAs
targeted to either side of the IVS26 mutation between exon 26 and exon 27,
EDIT-101 is hypothesised to delete or invert the mutation such that
functional CEP290 expression is restored. Recently published results detail
the development of EDIT-101 and summarizes in vitro experiments in
human cells and retinal explants (Maeder et al. 2019). Subretinal delivery of
EDIT-101 was well tolerated and achieved sustained CEP290 editing in
photoreceptor cells in mice and non-human primates, which met or exceeded
the target therapeutic level. This study also determined that the 2 gRNAs
chosen for this treatment were highly specific, and no off-target sites could
be verified in either multiple human cell lines or human retinal cells.
Furthermore, the results presented in this study were encouraging since
editing rates were above the therapeutic threshold for both mice and non-
human primates, thus providing the basis for dose extrapolation to first-in-
human trial for a CRISPR/Cas9 gene therapy. These clinical trials would
pave the way for a new era for treating the inherited ocular diseases for
which current disease prognosis is poor.
CONCLUSION
Gene editing holds enormous potential for the clinical treatment of
inherited eye diseases. The eye, being an ideal target for gene editing puts
ophthalmology at the forefront of the emerging CRISPR/Cas technology.
Improvements in the development of more efficient delivery methods and
more precise CRISPR/Cas components will allow these therapies to become
a truly personalized option for medical treatment. Although a barrier to the
widespread clinical application of these new therapies is their financial cost,
CRISPR technology has the potential to disrupt pathology and cure genetic
disease.
In terms of cost, projections range from 500,000 US dollars to $1.5
million. The cost could limit the availability of the technology to lower
socioeconomic groups, but with future advances, the cost is likely to fall. To
prepare the insurance industry, many leaders in the field are seeking to

C. B. Tara Moore, Larry A. DeDionisio, Hila Roshanravan et al. 136
address the challenges, working with payers, policy makers, and other key
experts in regulatory affairs and law to recommend payment programs
(Salzman et al. 2018). There have been various models proposed to handle
the high cost of gene therapy such as a value-based payment model which
would link payments to actual outcomes observed over an extended time
period. One possibility would be for the pharmaceutical company to
reimburse the insurance provider the cost of the treatment if, after three years
there is no clinical improvement. Visual science researchers and clinicians
continue to conduct ground breaking studies that will soon translate into new
therapeutic options.
REFERENCES
Abhinand, Chandran S., Rajesh Raju, Sasikumar J. Soumya, Prabha S. Arya,
and Perumana R. Sudhakaran. 2016. “VEGF-A/VEGFR2 signaling
network in endothelial cells relevant to angiogenesis.” Journal of cell
communication and signaling 10 (4):347-354.
Ablain, Julien, Ellen M. Durand, Song Yang, Yi Zhou, and Leonard I. Zon.
2015. “A CRISPR/Cas9 vector system for tissue-specific gene
disruption in zebrafish.” Developmental cell 32 (6):756-764.
Acland, G. M., G. D. Aguirre, J. Ray, Q. Zhang, T. S. Aleman, A. V.
Cideciyan, S. E. Pearce-Kelling, V. Anand, Y. Zeng, A. M. Maguire, S.
G. Jacobson, W. W. Hauswirth, and J. Bennett. 2001. “Gene therapy
restores vision in a canine model of childhood blindness.” Nat Genet 28
(1):92-5. doi: 10.1038/88327.
Allen, E. H. A., S. D. Atkinson, H. Liao, J. E. Moore, D. M. L. Pedrioli, F.
J. D. Smith, W. H. I. McLean, and C. B. T. Moore. 2013. “Allele-
specific siRNA silencing for the common keratin 12 founder mutation
in Meesmann epithelial corneal dystrophy.” Invest Ophthalmol Vis Sci
54 (1):494-502. doi: 10.1167/iovs.12-10528.
Arno, G., S. A. Agrawal, A. Eblimit, J. Bellingham, M. Xu, F. Wang, C.
Chakarova, D. A. Parfitt, A. Lane, T. Burgoyne, S. Hull, K. J. Carss, A.
Fiorentino, M. J. Hayes, P. M. Munro, R. Nicols, N. Pontikos, G. E.

The Application of CRISPR/Cas9 Therapies in Ophthalmology … 137
Holder, C. Asomugha, F. L. Raymond, A. T. Moore, V. Plagnol, M.
Michaelides, A. J. Hardcastle, Y. Li, C. Cukras, A. R. Webster, M. E.
Cheetham, and R. Chen. 2016. “Mutations in REEP6 Cause Autosomal-
Recessive Retinitis Pigmentosa.” Am J Hum Genet 99 (6):1305-1315.
doi: 10.1016/j.ajhg.2016.10.008.
Ashraf, M., and A. A. R. Souka. 2017. “Aflibercept in age-related macular
degeneration: evaluating its role as a primary therapeutic option.” Eye
31 (11):1523.
Athanasopoulos, T., M. M. Munye, and R. J. Yanez-Munoz. 2017.
“Nonintegrating Gene Therapy Vectors.” Hematol Oncol Clin North Am
31 (5):753-770. doi: 10.1016/j.hoc.2017.06.007.
Atkinson, S. D., V. E. McGilligan, H. Liao, I. Szeverenyi, F. J. Smith, C. B.
Moore, and W. H. McLean. 2011. “Development of allele-specific
therapeutic siRNA for keratin 5 mutations in epidermolysis bullosa
simplex.” J Invest Dermatol 131 (10):2079-86. doi: 10.1038/
jid.2011.169.
Bainbridge, J. W., M. S. Mehat, V. Sundaram, S. J. Robbie, S. E. Barker, C.
Ripamonti, A. Georgiadis, F. M. Mowat, S. G. Beattie, P. J. Gardner, K.
L. Feathers, V. A. Luong, S. Yzer, K. Balaggan, A. Viswanathan, T. J.
de Ravel, I. Casteels, G. E. Holder, N. Tyler, F. W. Fitzke, R. G.
Weleber, M. Nardini, A. T. Moore, D. A. Thompson, S. M. Petersen-
Jones, M. Michaelides, L. I. van den Born, A. Stockman, A. J. Smith, G.
Rubin, and R. R. Ali. 2015. “Long-term effect of gene therapy on
Leber's congenital amaurosis.” N Engl J Med 372 (20):1887-97. doi:
10.1056/NEJMoa1414221.
Bainbridge, James W. B., Alexander J. Smith, Susie S. Barker, Scott Robbie,
Robert Henderson, Kamaljit Balaggan, Ananth Viswanathan, Graham
E. Holder, Andrew Stockman, and Nick Tyler. 2008. “Effect of gene
therapy on visual function in Leber's congenital amaurosis.” New
England Journal of Medicine 358 (21):2231-2239.
Bakondi, B., W. Lv, B. Lu, M. K. Jones, Y. Tsai, K. J. Kim, R. Levy, A. A.
Akhtar, J. J. Breunig, C. N. Svendsen, and S. Wang. 2016. “In Vivo
CRISPR/Cas9 Gene Editing Corrects Retinal Dystrophy in the S334ter-

C. B. Tara Moore, Larry A. DeDionisio, Hila Roshanravan et al. 138
3 Rat Model of Autosomal Dominant Retinitis Pigmentosa.” Mol Ther
24 (3):556-63. doi: 10.1038/mt.2015.220.
Bassuk, A. G., A. Zheng, Y. Li, S. H. Tsang, and V. B. Mahajan. 2016.
“Precision Medicine: Genetic Repair of Retinitis Pigmentosa in Patient-
Derived Stem Cells.” Sci Rep 6:19969. doi: 10.1038/srep19969.
Baylis, Françoise, and Marcus McLeod. 2017. “First-in-human phase 1
CRISPR gene editing cancer trials: are we ready?” Current gene therapy
17 (4):309-319.
Belfort, Marlene, and Richard P. Bonocora. 2014. “Homing endonucleases:
from genetic anomalies to programmable genomic clippers.” In Homing
Endonucleases, 1-26. Springer.
Bennett, Jean, Jennifer Wellman, Kathleen A. Marshall, Sarah McCague,
Manzar Ashtari, Julie DiStefano-Pappas, Okan U. Elci, Daniel C.
Chung, Junwei Sun, and J. Fraser Wright. 2016. “Safety and durability
of effect of contralateral-eye administration of AAV2 gene therapy in
patients with childhood-onset blindness caused by RPE65 mutations: a
follow-on phase 1 trial.” The Lancet 388 (10045):661-672.
Berger, W., B. Kloeckener-Gruissem, and J. Neidhardt. 2010. “The
molecular basis of human retinal and vitreoretinal diseases.” Prog Retin
Eye Res 29 (5):335-75. doi: 10.1016/j.preteyeres.2010.03.004.
Bibikova, Marina, Kelly Beumer, Jonathan K Trautman, and Dana Carroll.
2003. “Enhancing gene targeting with designed zinc finger nucleases.”
Science 300 (5620):764-764.
Bibikova, Marina, Dana Carroll, David J. Segal, Jonathan K. Trautman, Jeff
Smith, Yang-Gyun Kim, and Srinivasan Chandrasegaran. 2001.
“Stimulation of homologous recombination through targeted cleavage
by chimeric nucleases.” Molecular and cellular biology 21 (1):289-297.
Bibikova, Marina, Mary Golic, Kent G. Golic, and Dana Carroll. 2002.
“Targeted chromosomal cleavage and mutagenesis in Drosophila using
zinc-finger nucleases.” Genetics 161 (3):1169-1175.
Bogdanove, Adam J., and Daniel F. Voytas. 2011. “TAL effectors:
customizable proteins for DNA targeting.” Science 333 (6051):1843-
1846.

The Application of CRISPR/Cas9 Therapies in Ophthalmology … 139
Borras, T., B. T. Gabelt, G. K. Klintworth, J. C. Peterson, and P. L.
Kaufman. 2001. “Non-invasive observation of repeated adenoviral GFP
gene delivery to the anterior segment of the monkey eye in vivo.” J Gene
Med 3 (5):437-49. doi: 10.1002/jgm.210.
Boye, S. E., S. L. Boye, A. S. Lewin, and W. W. Hauswirth. 2013. “A
comprehensive review of retinal gene therapy.” Mol Ther 21 (3):509-
19. doi: 10.1038/mt.2012.280.
Burnight, E. R., M. Gupta, L. A. Wiley, K. R. Anfinson, A. Tran, R.
Triboulet, J. M. Hoffmann, D. L. Klaahsen, J. L. Andorf, C. Jiao, E. H.
Sohn, M. K. Adur, J. W. Ross, R. F. Mullins, G. Q. Daley, T. M.
Schlaeger, E. M. Stone, and B. A. Tucker. 2017. “Using CRISPR-Cas9
to Generate Gene-Corrected Autologous iPSCs for the Treatment of
Inherited Retinal Degeneration.” Mol Ther 25 (9):1999-2013. doi:
10.1016/j.ymthe.2017.05.015.
Cameron, P., C. K. Fuller, P. D. Donohoue, B. N. Jones, M. S. Thompson,
M. M. Carter, S. Gradia, B. Vidal, E. Garner, E. M. Slorach, E. Lau, L.
M. Banh, A. M. Lied, L. S. Edwards, A. H. Settle, D. Capurso, V. Llaca,
S. Deschamps, M. Cigan, J. K. Young, and A. P. May. 2017. “Mapping
the genomic landscape of CRISPR-Cas9 cleavage.” Nat Methods 14
(6):600-606. doi: 10.1038/nmeth.4284.
Chen, Janice S., Yavuz S. Dagdas, Benjamin P. Kleinstiver, Moira M.
Welch, Alexander A. Sousa, Lucas B. Harrington, Samuel H. Sternberg,
J. Keith Joung, Ahmet Yildiz, and Jennifer A. Doudna. 2017. “Enhanced
proofreading governs CRISPR–Cas9 targeting accuracy.” Nature 550
(7676):407.
Cheng, A. W., H. Wang, H. Yang, L. Shi, Y. Katz, T. W. Theunissen, S.
Rangarajan, C. S. Shivalila, D. B. Dadon, and R. Jaenisch. 2013.
“Multiplexed activation of endogenous genes by CRISPR-on, an RNA-
guided transcriptional activator system.” Cell Res 23 (10):1163-71. doi:
10.1038/cr.2013.122.
Cho, S. W., S. Kim, Y. Kim, J. Kweon, H. S. Kim, S. Bae, and J. S. Kim.
2014. “Analysis of off-target effects of CRISPR/Cas-derived RNA-
guided endonucleases and nickases.” Genome Res 24 (1):132-41. doi:
10.1101/gr.162339.113.

C. B. Tara Moore, Larry A. DeDionisio, Hila Roshanravan et al. 140
Cho, S. W., J. Lee, D. Carroll, J. S. Kim, and J. Lee. 2013. “Heritable gene
knockout in Caenorhabditis elegans by direct injection of Cas9-sgRNA
ribonucleoproteins.” Genetics 195 (3):1177-80. doi: 10.1534/genetics.
113.155853.
Choulika, Andre, Arnaud Perrin, Bernard Dujon, and Jean-Francois Nicolas.
1995. “Induction of homologous recombination in mammalian
chromosomes by using the I-SceI system of Saccharomyces cerevisiae.”
Molecular and cellular biology 15 (4):1968-1973.
Christie, K. A., D. G. Courtney, L. A. DeDionisio, C. C. Shern, S. De
Majumdar, L. C. Mairs, M. A. Nesbit, and C. B. T. Moore. 2017.
“Towards personalised allele-specific CRISPR gene editing to treat
autosomal dominant disorders.” Sci Rep 7 (1):16174. doi:
10.1038/s41598-017-16279-4.
Christie K. A., Blighe K., Lukkassen M., Ali Z., DeDionisio L. A., Conway
C., Lennikov A., De Majumdar S., Chao Shern C., Irwin R. E., Turnbull
D., Marshall J., Jensen L. D., Nesbit M. A., Moore C. B. T. 2019.
“ASNIP-CRISPR enables mutation-1 independent allele-specific
editing by Cas9 to treat autosomal dominant disease.” Hum Mol Genet
in print.
Chu, V. T., T. Weber, B. Wefers, W. Wurst, S. Sander, K. Rajewsky, and R.
Kuhn. 2015. “Increasing the efficiency of homology-directed repair for
CRISPR-Cas9-induced precise gene editing in mammalian cells.” Nat
Biotechnol 33 (5):543-8. doi: 10.1038/nbt.3198.
Cideciyan, A. V., W. W. Hauswirth, T. S. Aleman, S. Kaushal, S. B.
Schwartz, S. L. Boye, E. A. Windsor, T. J. Conlon, A. Sumaroka, J. J.
Pang, A. J. Roman, B. J. Byrne, and S. G. Jacobson. 2009. “Human
RPE65 gene therapy for Leber congenital amaurosis: persistence of
early visual improvements and safety at 1 year.” Hum Gene Ther 20
(9):999-1004. doi: 10.1089/hum.2009.086.
Collery, Ross F., Peter J. Volberding, Jonathan R. Bostrom, Brian A. Link,
and Joseph C. Besharse. 2016. “Loss of Zebrafish MFRP causes
nanophthalmia, hyperopia, and accumulation of subretinal
macrophages.” Investigative ophthalmology & visual science 57
(15):6805-6814.

The Application of CRISPR/Cas9 Therapies in Ophthalmology … 141
Cong, Le., F. Ann Ran, David Cox, Shuailiang Lin, Robert Barretto, Naomi
Habib, Patrick D. Hsu, Xuebing Wu, Wenyan Jiang, and Luciano A.
Marraffini. 2013. “Multiplex genome engineering using CRISPR/Cas
systems.” Science 339 (6121):819-823.
Courtney, D. G., S. D. Atkinson, E. H. Allen, J. E. Moore, C. P. Walsh, D.
M. Pedrioli, C. J. MacEwen, G. Pellegrini, E. Maurizi, C. Serafini, M.
Fantacci, H. Liao, A. D. Irvine, W. H. McLean, and C. B. Moore. 2014.
“siRNA silencing of the mutant keratin 12 allele in corneal limbal
epithelial cells grown from patients with Meesmann's epithelial corneal
dystrophy.” Invest Ophthalmol Vis Sci 55 (5):3352-60. doi: 10.1167/
iovs.13-12957.
Courtney, D. G., S. D. Atkinson, J. E. Moore, E. Maurizi, C. Serafini, G.
Pellegrini, G. C. Black, F. D. Manson, G. H. Yam, C. J. Macewen, E. H.
Allen, W. H. McLean, and C. B. Moore. 2014. “Development of allele-
specific gene-silencing siRNAs for TGFBI Arg124Cys in lattice corneal
dystrophy type I.” Invest Ophthalmol Vis Sci 55 (2):977-85. doi:
10.1167/iovs.13-13279.
Courtney, D. G., J. E. Moore, S. D. Atkinson, E. Maurizi, E. H. Allen, D. M.
Pedrioli, W. H. McLean, M. A. Nesbit, and C. B. Moore. 2016.
“CRISPR/Cas9 DNA cleavage at SNP-derived PAM enables both in
vitro and in vivo KRT12 mutation-specific targeting.” Gene Ther 23
(1):108-12. doi: 10.1038/gt.2015.82.
Courtney, David G., Sarah D. Atkinson, Edwin H. A. Allen, Johnny E.
Moore, Colum P. Walsh, Deena M. Leslie Pedrioli, Caroline J.
MacEwen, Graziella Pellegrini, Eleonora Maurizi, and Chiara Serafini.
2014. “siRNA silencing of the mutant keratin 12 allele in corneal limbal
epithelial cells grown from patients with Meesmann's epithelial corneal
dystrophy.” Investigative ophthalmology & visual science 55 (5):3352-
3360.
Crosetto, N., A. Mitra, M. J. Silva, M. Bienko, N. Dojer, Q. Wang, E.
Karaca, R. Chiarle, M. Skrzypczak, K. Ginalski, P. Pasero, M. Rowicka,
and I. Dikic. 2013. “Nucleotide-resolution DNA double-strand break
mapping by next-generation sequencing.” Nat Methods 10 (4):361-5.
doi: 10.1038/nmeth.2408.

C. B. Tara Moore, Larry A. DeDionisio, Hila Roshanravan et al. 142
Cukras, C., H. E. Wiley, B. G. Jeffrey, H. N. Sen, A. Turriff, Y. Zeng, C.
Vijayasarathy, D. Marangoni, L. Ziccardi, S. Kjellstrom, T. K. Park, S.
Hiriyanna, J. F. Wright, P. Colosi, Z. Wu, R. A. Bush, L. L. Wei, and P.
A. Sieving. 2018. “Retinal AAV8-RS1 Gene Therapy for X-Linked
Retinoschisis: Initial Findings from a Phase I/IIa Trial by Intravitreal
Delivery.” Mol Ther 26 (9):2282-2294. doi: 10.1016/j.ymthe.2018.
05.025.
Dahlman, James E., Omar O. Abudayyeh, Julia Joung, Jonathan S.
Gootenberg, Feng Zhang, and Silvana Konermann. 2015. “Orthogonal
gene knockout and activation with a catalytically active Cas9 nuclease.”
Nature biotechnology 33 (11):1159.
Dalkara, Deniz, Olivier Goureau, Katia Marazova, and Jose-Alain Sahel.
2016. “Let there be light: gene and cell therapy for blindness.” Human
gene therapy 27 (2):134-147.
Davis, K. M., V. Pattanayak, D. B. Thompson, J. A. Zuris, and D. R. Liu.
2015. “Small molecule-triggered Cas9 protein with improved genome-
editing specificity.” Nat Chem Biol 11 (5):316-8. doi: 10.1038/
nchembio.1793.
Day, T. P., L. C. Byrne, D. V. Schaffer, and J. G. Flannery. 2014. “Advances
in AAV vector development for gene therapy in the retina.” Adv Exp
Med Biol 801:687-93. doi: 10.1007/978-1-4614-3209-8_86.
de la Fuente, M., B. Seijo, and M. J. Alonso. 2008a. “Bioadhesive
hyaluronan-chitosan nanoparticles can transport genes across the ocular
mucosa and transfect ocular tissue.” Gene Ther 15 (9):668-76. doi:
10.1038/gt.2008.16.
de la Fuente, M., B. Seijo, and M. J. Alonso. 2008b. “Novel hyaluronic acid-
chitosan nanoparticles for ocular gene therapy.” Invest Ophthalmol Vis
Sci 49 (5):2016-24. doi: 10.1167/iovs.07-1077.
Dearment, A. 2019. UPenn study of CRISPR/Cas9 cell therapy in cancers
doses first patients. In MedCity News
Dias, Marina França, Kwangsic Joo, Jessica A Kemp, Silvia Ligorio Fialho,
Armando da Silva Cunha Jr, Se Joon Woo, and Young Jik Kwon. 2018.
“Molecular genetics and emerging therapies for retinitis pigmentosa:

The Application of CRISPR/Cas9 Therapies in Ophthalmology … 143
basic research and clinical perspectives.” Progress in retinal and eye
research 63:107-131.
Edwards, T. L., J. K. Jolly, M. Groppe, A. R. Barnard, C. L. Cottriall, T.
Tolmachova, G. C. Black, A. R. Webster, A. J. Lotery, G. E. Holder, K.
Xue, S. M. Downes, M. P. Simunovic, M. C. Seabra, and R. E.
MacLaren. 2016. “Visual Acuity after Retinal Gene Therapy for
Choroideremia.” N Engl J Med 374 (20):1996-8. doi: 10.1056/
NEJMc1509501.
Esvelt, Kevin M., Prashant Mali, Jonathan L. Braff, Mark Moosburner,
Stephanie J. Yaung, and George M. Church. 2013. “Orthogonal Cas9
proteins for RNA-guided gene regulation and editing.” Nature methods
10 (11):1116.
Frock, R. L., J. Hu, R. M. Meyers, Y. J. Ho, E. Kii, and F. W. Alt. 2015.
“Genome-wide detection of DNA double-stranded breaks induced by
engineered nucleases.” Nat Biotechnol 33 (2):179-86. doi: 10.1038/
nbt.3101.
Fu, Y., J. A. Foden, C. Khayter, M. L. Maeder, D. Reyon, J. K. Joung, and
J. D. Sander. 2013. “High-frequency off-target mutagenesis induced by
CRISPR-Cas nucleases in human cells.” Nat Biotechnol 31 (9):822-6.
doi: 10.1038/nbt.2623.
Fu, Y., D. Reyon, and J. K. Joung. 2014. “Targeted genome editing in human
cells using CRISPR/Cas nucleases and truncated guide RNAs.” Methods
Enzymol 546:21-45. doi: 10.1016/b978-0-12-801185-0.00002-7.
Fu, Y., J. D. Sander, D. Reyon, V. M. Cascio, and J. K. Joung. 2014.
“Improving CRISPR-Cas nuclease specificity using truncated guide
RNAs.” Nat Biotechnol 32 (3):279-284. doi: 10.1038/nbt.2808.
Ghazi, Nicola G., Emad B. Abboud, Sawsan R. Nowilaty, Hisham Alkuraya,
Abdulrahman Alhommadi, Huimin Cai, Rui Hou, Wen-Tao Deng,
Sanford L. Boye, and Abdulrahman Almaghamsi. 2016. “Treatment of
retinitis pigmentosa due to MERTK mutations by ocular subretinal
injection of adeno-associated virus gene vector: results of a phase I
trial.” Human genetics 135 (3):327-343.
Grishanin, Ruslan, Brian Vuillemenot, Pallavi Sharma, Annahita Keravala,
Judith Greengard, Claire Gelfman, Mark Blumenkrantz, Matthew

C. B. Tara Moore, Larry A. DeDionisio, Hila Roshanravan et al. 144
Lawrence, Wenzheng Hu, and Szilárd Kiss. 2019. “Preclinical
Evaluation of ADVM-022, a Novel Gene Therapy Approach to Treating
Wet Age-Related Macular Degeneration.” Molecular Therapy 27
(1):118-129.
Guilinger, J. P., D. B. Thompson, and D. R. Liu. 2014. “Fusion of
catalytically inactive Cas9 to FokI nuclease improves the specificity of
genome modification.” Nat Biotechnol 32 (6):577-582. doi:
10.1038/nbt.2909.
Hauswirth, William W., Tomas S. Aleman, Shalesh Kaushal, Artur V.
Cideciyan, Sharon B. Schwartz, Lili Wang, Thomas J. Conlon, Sanford
L. Boye, Terence R. Flotte, and Barry J. Byrne. 2008. “Treatment of
leber congenital amaurosis due to RPE65 mutations by ocular subretinal
injection of adeno-associated virus gene vector: short-term results of a
phase I trial.” Human gene therapy 19 (10):979-990.
Hayashi, Ryuhei, Yuki Ishikawa, Miyuki Ito, Tomofumi Kageyama, Kuniko
Takashiba, Tsuyoshi Fujioka, Motokazu Tsujikawa, Hiroyuki Miyoshi,
Masayuki Yamato, and Yukio Nakamura. 2012. “Generation of corneal
epithelial cells from induced pluripotent stem cells derived from human
dermal fibroblast and corneal limbal epithelium.” PLoS One 7
(9):e45435.
Heier, Jeffrey S., Saleema Kherani, Shilpa Desai, Pravin Dugel, Shalesh
Kaushal, Seng H. Cheng, Cheryl Delacono, Annie Purvis, Susan
Richards, and Annaig Le-Halpere. 2017. “Intravitreous injection of
AAV2-sFLT01 in patients with advanced neovascular age-related
macular degeneration: a phase 1, open-label trial.” The Lancet 390
(10089):50-61.
Hippert, C., S. Ibanes, N. Serratrice, F. Court, F. Malecaze, E. J. Kremer,
and V. Kalatzis. 2012. “Corneal transduction by intra-stromal injection
of AAV vectors in vivo in the mouse and ex vivo in human explants.”
PLoS One 7 (4):e35318. doi: 10.1371/journal.pone.0035318.
Hou, Zhonggang, Yan Zhang, Nicholas E Propson, Sara E Howden, Li-Fang
Chu, Erik J Sontheimer, and James A Thomson. 2013. “Efficient
genome engineering in human pluripotent stem cells using Cas9 from

The Application of CRISPR/Cas9 Therapies in Ophthalmology … 145
Neisseria meningitidis.” Proceedings of the National Academy of
Sciences 110 (39):15644-15649.
Hsu, P. D., D. A. Scott, J. A. Weinstein, F. A. Ran, S. Konermann, V.
Agarwala, Y. Li, E. J. Fine, X. Wu, O. Shalem, T. J. Cradick, L. A.
Marraffini, G. Bao, and F. Zhang. 2013. “DNA targeting specificity of
RNA-guided Cas9 nucleases.” Nat Biotechnol 31 (9):827-32. doi: 10.
1038/nbt.2647.
Hu, J., R. M. Meyers, J. Dong, R. A. Panchakshari, F. W. Alt, and R. L.
Frock. 2016. “Detecting DNA double-stranded breaks in mammalian
genomes by linear amplification-mediated high-throughput genome-
wide translocation sequencing.” Nat Protoc 11 (5):853-71. doi:
10.1038/nprot.2016.043.
Huang, Xionggao, Guohong Zhou, Wenyi Wu, Yajian Duan, Gaoen Ma,
Jingyuan Song, Ru Xiao, Luk Vandenberghe, Feng Zhang, and Patricia
A D’Amore. 2017. “Genome editing abrogates angiogenesis in vivo.”
Nature communications 8 (1):112.
Jacobson, S. G., A. V. Cideciyan, R. Ratnakaram, E. Heon, S. B. Schwartz,
A. J. Roman, M. C. Peden, T. S. Aleman, S. L. Boye, A. Sumaroka, T.
J. Conlon, R. Calcedo, J. J. Pang, K. E. Erger, M. B. Olivares, C. L.
Mullins, M. Swider, S. Kaushal, W. J. Feuer, A. Iannaccone, G. A.
Fishman, E. M. Stone, B. J. Byrne, and W. W. Hauswirth. 2012. “Gene
therapy for leber congenital amaurosis caused by RPE65 mutations:
safety and efficacy in 15 children and adults followed up to 3 years.”
Arch Ophthalmol 130 (1):9-24. doi: 10.1001/archophthalmol.2011.298.
Jacobson, S. G., A. V. Cideciyan, A. J. Roman, A. Sumaroka, S. B.
Schwartz, E. Heon, and W. W. Hauswirth. 2015. “Improvement and
decline in vision with gene therapy in childhood blindness.” N Engl J
Med 372 (20):1920-6. doi: 10.1056/NEJMoa1412965.
Jiang, David J., Christine L. Xu, and Stephen H. Tsang. 2018. “Revolution
in gene medicine therapy and genome surgery.” Genes 9 (12):575.
Jin, L., X. Zeng, M. Liu, Y. Deng, and N. He. 2014. “Current progress in
gene delivery technology based on chemical methods and nano-
carriers.” Theranostics 4 (3):240-55. doi: 10.7150/thno.6914.

C. B. Tara Moore, Larry A. DeDionisio, Hila Roshanravan et al. 146
Jinek, Martin, Krzysztof Chylinski, Ines Fonfara, Michael Hauer, Jennifer
A Doudna, and Emmanuelle Charpentier. 2012. “A programmable dual-
RNA–guided DNA endonuclease in adaptive bacterial immunity.”
Science 337 (6096):816-821.
Kabadi, A. M., D. G. Ousterout, I. B. Hilton, and C. A. Gersbach. 2014.
“Multiplex CRISPR/Cas9-based genome engineering from a single
lentiviral vector.” Nucleic Acids Res 42 (19):e147. doi: 10.1093/nar/
gku749.
Karginov, Fedor V., and Gregory J. Hannon. 2010. “The CRISPR system:
small RNA-guided defense in bacteria and archaea.” Molecular cell 37
(1):7-19.
Kay, Mark A. 2011. “State-of-the-art gene-based therapies: the road ahead.”
Nature Reviews Genetics 12 (5):316.
Kim, D., S. Bae, J. Park, E. Kim, S. Kim, H. R. Yu, J. Hwang, J. I. Kim, and
J. S. Kim. 2015. “Digenome-seq: genome-wide profiling of CRISPR-
Cas9 off-target effects in human cells.” Nat Methods 12 (3):237-43, 1 p
following 243. doi: 10.1038/nmeth.3284.
Kim, E., T. Koo, S. W. Park, D. Kim, K. Kim, H. Y. Cho, D. W. Song, K. J.
Lee, M. H. Jung, S. Kim, J. H. Kim, J. H. Kim, and J. S. Kim. 2017. “In
vivo genome editing with a small Cas9 orthologue derived from
Campylobacter jejuni.” Nat Commun 8:14500. doi: 10.1038/
ncomms14500.
Kim, K., S. W. Park, J. H. Kim, S. H. Lee, D. Kim, T. Koo, K. E. Kim, J. H.
Kim, and J. S. Kim. 2017. “Genome surgery using Cas9
ribonucleoproteins for the treatment of age-related macular
degeneration.” Genome Res 27 (3):419-426. doi: 10.1101/gr.219089.
116.
Kim, Sojung, Daesik Kim, Seung Woo Cho, Jungeun Kim, and Jin-Soo Kim.
2014. “Highly efficient RNA-guided genome editing in human cells via
delivery of purified Cas9 ribonucleoproteins.” Genome research 24
(6):1012-1019.
Kitazawa, K., T. Hikichi, T. Nakamura, C. Sotozono, S. Kinoshita, and S.
Masui. 2017. “PAX6 regulates human corneal epithelium cell identity.”
Exp Eye Res 154:30-38. doi: 10.1016/j.exer.2016.11.005.

The Application of CRISPR/Cas9 Therapies in Ophthalmology … 147
Klausner, E. A., Z. Zhang, R. L. Chapman, R. F. Multack, and M. V. Volin.
2010. “Ultrapure chitosan oligomers as carriers for corneal gene
transfer.” Biomaterials 31 (7):1814-20. doi: 10.1016/j.biomaterials.
2009.10.031.
Kleinstiver, B. P., V. Pattanayak, M. S. Prew, S. Q. Tsai, N. T. Nguyen, Z.
Zheng, and J. K. Joung. 2016. “High-fidelity CRISPR-Cas9 nucleases
with no detectable genome-wide off-target effects.” Nature 529
(7587):490-5. doi: 10.1038/nature16526.
Kleinstiver, B. P., M. S. Prew, S. Q. Tsai, N. T. Nguyen, V. V. Topkar, Z.
Zheng, and J. K. Joung. 2015. “Broadening the targeting range of
Staphylococcus aureus CRISPR-Cas9 by modifying PAM recognition.”
Nat Biotechnol 33 (12):1293-1298. doi: 10.1038/nbt.3404.
Kleinstiver, Benjamin P., Michelle S. Prew, Shengdar Q. Tsai, Ved V.
Topkar, Nhu T. Nguyen, Zongli Zheng, Andrew P. W. Gonzales,
Zhuyun Li, Randall T. Peterson, and Jing-Ruey Joanna Yeh. 2015.
“Engineered CRISPR-Cas9 nucleases with altered PAM specificities.”
Nature 523 (7561):481.
Klintworth, G. K. 2009. “Corneal dystrophies.” Orphanet J Rare Dis 4:7.
doi: 10.1186/1750-1172-4-7.
Kwart, D., D. Paquet, S. Teo, and M. Tessier-Lavigne. 2017. “Precise and
efficient scarless genome editing in stem cells using CORRECT.” Nat
Protoc 12 (2):329-354. doi: 10.1038/nprot.2016.171.
Le Page, Michael. 2017. “Boom in human gene editing as 20 CRISPR trials
gear up.” New Scientist, https://www.newscientist.com/article/
2133095-boom-in-human-gene-editing-as-20-crispr-trials-gear-up/.
Lee, Daniel W., James N. Kochenderfer, Maryalice Stetler-Stevenson,
Yongzhi K. Cui, Cindy Delbrook, Steven A. Feldman, Terry J. Fry,
Rimas Orentas, Marianna Sabatino, and Nirali N Shah. 2015. “T cells
expressing CD19 chimeric antigen receptors for acute lymphoblastic
leukaemia in children and young adults: a phase 1 dose-escalation trial.”
The Lancet 385 (9967):517-528.
Lee, K., M. Conboy, H. M. Park, F. Jiang, H. J. Kim, M. A. Dewitt, V. A.
Mackley, K. Chang, A. Rao, C. Skinner, T. Shobha, M. Mehdipour, H.
Liu, W. C. Huang, F. Lan, N. L. Bray, S. Li, J. E. Corn, K. Kataoka, J.

C. B. Tara Moore, Larry A. DeDionisio, Hila Roshanravan et al. 148
A. Doudna, I. Conboy, and N. Murthy. 2017. “Nanoparticle delivery of
Cas9 ribonucleoprotein and donor DNA in vivo induces homology-
directed DNA repair.” Nat Biomed Eng 1:889-901. doi: 10.1038/
s41551-017-0137-2.
Lee, Minyoung, and Hyongbum Kim. 2019. “Therapeutic application of the
CRISPR system: current issues and new prospects.” Human Genetics:1-
28.
Liang, X., J. Potter, S. Kumar, N. Ravinder, and J. D. Chesnut. 2017.
“Enhanced CRISPR/Cas9-mediated precise genome editing by
improved design and delivery of gRNA, Cas9 nuclease, and donor
DNA.” J Biotechnol 241:136-146. doi: 10.1016/j.jbiotec.2016.11.011.
Liang, X., J. Potter, S. Kumar, Y. Zou, R. Quintanilla, M. Sridharan, J. Carte,
W. Chen, N. Roark, S. Ranganathan, N. Ravinder, and J. D. Chesnut.
2015. “Rapid and highly efficient mammalian cell engineering via Cas9
protein transfection.” J Biotechnol 208:44-53. doi: 10.1016/j.jbiotec.
2015.04.024.
Liao, H., A. D. Irvine, C. J. Macewen, K. H. Weed, L. Porter, L. D. Corden,
A. B. Gibson, J. E. Moore, F. J. Smith, W. H. McLean, and C. B. Moore.
2011. “Development of allele-specific therapeutic siRNA in Meesmann
epithelial corneal dystrophy.” PLoS One 6 (12):e28582. doi: 10.1371/
journal.pone.0028582.
Liew, Gerald, Michel Michaelides, and Catey Bunce. 2014. “A comparison
of the causes of blindness certifications in England and Wales in
working age adults (16–64 years), 1999–2000 with 2009–2010.” BMJ
open 4 (2):e004015.
Lin, S., B. T. Staahl, R. K. Alla, and J. A. Doudna. 2014. “Enhanced
homology-directed human genome engineering by controlled timing of
CRISPR/Cas9 delivery.” Elife 3:e04766. doi: 10.7554/eLife.04766.
Lin, Y., T. J. Cradick, M. T. Brown, H. Deshmukh, P. Ranjan, N. Sarode, B.
M. Wile, P. M. Vertino, F. J. Stewart, and G. Bao. 2014. “CRISPR/Cas9
systems have off-target activity with insertions or deletions between
target DNA and guide RNA sequences.” Nucleic Acids Res 42
(11):7473-85. doi: 10.1093/nar/gku402.

The Application of CRISPR/Cas9 Therapies in Ophthalmology … 149
Liu, J., M. Saghizadeh, S. S. Tuli, A. A. Kramerov, A. S. Lewin, D. C.
Bloom, W. W. Hauswirth, M. G. Castro, G. S. Schultz, and A. V.
Ljubimov. 2008. “Different tropism of adenoviruses and adeno-
associated viruses to corneal cells: implications for corneal gene
therapy.” Mol Vis 14:2087-96.
Liu, Melissa M., Jingsheng Tuo, and Chi-Chao Chan. 2011. “Republished
review: Gene therapy for ocular diseases.” Postgraduate medical
journal 87 (1029):487-495.
Luo, J., P. Baranov, S. Patel, H. Ouyang, J. Quach, F. Wu, A. Qiu, H. Luo,
C. Hicks, J. Zeng, J. Zhu, J. Lu, N. Sfeir, C. Wen, M. Zhang, V. Reade,
S. Patel, J. Sinden, X. Sun, P. Shaw, M. Young, and K. Zhang. 2014.
“Human retinal progenitor cell transplantation preserves vision.” J Biol
Chem 289 (10):6362-71. doi: 10.1074/jbc.M113.513713.
Luo, Xin, Yaoxi He, Chao Zhang, Xiechao He, Lanzhen Yan, Min Li, Ting
Hu, Yan Hu, Jin Jiang, and Xiaoyu Meng. 2018. “Trio deep-sequencing
does not reveal unexpected mutations in Cas9-edited monkeys.”
bioRxiv:339143.
MacLaren, R. E., M. Groppe, A. R. Barnard, C. L. Cottriall, T. Tolmachova,
L. Seymour, K. R. Clark, M. J. During, F. P. Cremers, G. C. Black, A.
J. Lotery, S. M. Downes, A. R. Webster, and M. C. Seabra. 2014.
“Retinal gene therapy in patients with choroideremia: initial findings
from a phase 1/2 clinical trial.” Lancet 383 (9923):1129-37. doi:
10.1016/s0140-6736(13)62117-0.
Maeder, M. L., M. Stefanidakis, C. J. Wilson, R. Baral, L. A. Barrera, G. S.
Bounoutas, D. Bumcrot, H. Chao, D. M. Ciulla, J. A. DaSilva, A. Dass,
V. Dhanapal, T. J. Fennell, A. E. Friedland, G. Giannoukos, S. W.
Gloskowski, A. Glucksmann, G. M. Gotta, H. Jayaram, S. J. Haskett, B.
Hopkins, J. E. Horng, S. Joshi, E. Marco, R. Mepani, D. Reyon, T. Ta,
D. G. Tabbaa, S. J. Samuelsson, S. Shen, M. N. Skor, P. Stetkiewicz, T.
Wang, C. Yudkoff, V. E. Myer, C. F. Albright, and H. Jiang. 2019.
“Development of a gene-editing approach to restore vision loss in Leber
congenital amaurosis type 10.” Nat Med 25 (2):229-233. doi:
10.1038/s41591-018-0327-9.

C. B. Tara Moore, Larry A. DeDionisio, Hila Roshanravan et al. 150
Makarova, Kira S., Daniel H. Haft, Rodolphe Barrangou, Stan J. J. Brouns,
Emmanuelle Charpentier, Philippe Horvath, Sylvain Moineau,
Francisco J. M. Mojica, Yuri I. Wolf, and Alexander F. Yakunin. 2011.
“Evolution and classification of the CRISPR–Cas systems.” Nature
Reviews Microbiology 9 (6):467.
Makarova, Kira S., Yuri I. Wolf, Omer S. Alkhnbashi, Fabrizio Costa,
Shiraz A. Shah, Sita J. Saunders, Rodolphe Barrangou, Stan J. J. Brouns,
Emmanuelle Charpentier, and Daniel H. Haft. 2015. “An updated
evolutionary classification of CRISPR–Cas systems.” Nature Reviews
Microbiology 13 (11):722.
Mali, P., J. Aach, P. B. Stranges, K. M. Esvelt, M. Moosburner, S. Kosuri,
L. Yang, and G. M. Church. 2013. “CAS9 transcriptional activators for
target specificity screening and paired nickases for cooperative genome
engineering.” Nat Biotechnol 31 (9):833-8. doi: 10.1038/nbt.2675.
Mali, Prashant, Luhan Yang, Kevin M. Esvelt, John Aach, Marc Guell,
James E. DiCarlo, Julie E. Norville, and George M. Church. 2013.
“RNA-guided human genome engineering via Cas9.” Science 339
(6121):823-826.
Maruyama, T., S. K. Dougan, M. C. Truttmann, A. M. Bilate, J. R. Ingram,
and H. L. Ploegh. 2015. “Increasing the efficiency of precise genome
editing with CRISPR-Cas9 by inhibition of nonhomologous end
joining.” Nat Biotechnol 33 (5):538-42. doi: 10.1038/nbt.3190.
McLean, W. H., and C. B. Moore. 2011. “Keratin disorders: from gene to
therapy.” Hum Mol Genet 20 (R2):R189-97. doi: 10.1093/hmg/ddr379.
Michalakis, Stylianos, Christian Schön, Elvir Becirovic, and Martin Biel.
2017. “Gene therapy for achromatopsia.” The journal of gene medicine
19 (3):e2944.
Mintzer, Meredith A., and Eric E. Simanek. 2008. “Nonviral vectors for
gene delivery.” Chemical reviews 109 (2):259-302.
Mohan, Rajiv R., Ajay Sharma, Marcelo V. Netto, Sunilima Sinha, and
Steven E Wilson. 2005. “Gene therapy in the cornea.” Progress in
retinal and eye research 24 (5):537-559.
Mojica, F. J., C. Diez-Villasenor, J. Garcia-Martinez, and C. Almendros.
2009. “Short motif sequences determine the targets of the prokaryotic

The Application of CRISPR/Cas9 Therapies in Ophthalmology … 151
CRISPR defence system.” Microbiology 155 (Pt 3):733-40. doi:
10.1099/mic.0.023960-0.
Moore, C. B. T., K. A. Christie, J. Marshall, and M. A. Nesbit. 2018.
“Personalised genome editing - The future for corneal dystrophies.”
Prog Retin Eye Res 65:147-165. doi: 10.1016/j.preteyeres.2018.01.004.
Moore, Richard, Alec Spinhirne, Michael J. Lai, Samantha Preisser, Yi Li,
Taek Kang, and Leonidas Bleris. 2014. “CRISPR-based self-cleaving
mechanism for controllable gene delivery in human cells.” Nucleic acids
research 43 (2):1297-1303.
Moore, Tara C., Haihui Liao, Alan Irvine, Louise F. Porter, Bethany Gibson,
Laura D. Corden, Carrie J. MacEwen, Kathryn H. Weed, Jonathan E.
Moore, and William H. McLean. 2011. “Development of allele-specific
therapeutic siRNA in Meesmann epithelial corneal dystrophy.”
Investigative Ophthalmology & Visual Science 52 (14):1113-1113.
Naert, T., R. Colpaert, T. Van Nieuwenhuysen, D. Dimitrakopoulou, J.
Leoen, J. Haustraete, A. Boel, W. Steyaert, T. Lepez, D. Deforce, A.
Willaert, D. Creytens, and K. Vleminckx. 2016. “CRISPR/Cas9
mediated knockout of rb1 and rbl1 leads to rapid and penetrant
retinoblastoma development in Xenopus tropicalis.” Sci Rep 6:35264.
doi: 10.1038/srep35264.
Nishimasu, H., F. A. Ran, P. D. Hsu, S. Konermann, S. I. Shehata, N.
Dohmae, R. Ishitani, F. Zhang, and O. Nureki. 2014. “Crystal structure
of Cas9 in complex with guide RNA and target DNA.” Cell 156 (5):935-
49. doi: 10.1016/j.cell.2014.02.001.
Nissim, L., S. D. Perli, A. Fridkin, P. Perez-Pinera, and T. K. Lu. 2014.
“Multiplexed and programmable regulation of gene networks with an
integrated RNA and CRISPR/Cas toolkit in human cells.” Mol Cell 54
(4):698-710. doi: 10.1016/j.molcel.2014.04.022.
O'Callaghan, J., D. E. Crosbie, P. S. Cassidy, J. M. Sherwood, C. Flugel-
Koch, E. Lutjen-Drecoll, M. M. Humphries, E. Reina-Torres, D.
Wallace, A. S. Kiang, M. Campbell, W. D. Stamer, D. R. Overby, C.
O'Brien, L. C. S. Tam, and P. Humphries. 2017. “Therapeutic potential
of AAV-mediated MMP-3 secretion from corneal endothelium in

C. B. Tara Moore, Larry A. DeDionisio, Hila Roshanravan et al. 152
treating glaucoma.” Hum Mol Genet 26 (7):1230-1246. doi: 10.1093/
hmg/ddx028.
Oliveira, A. V., A. M. Rosa da Costa, and G. A. Silva. 2017. “Non-viral
strategies for ocular gene delivery.” Mater Sci Eng C Mater Biol Appl
77:1275-1289. doi: 10.1016/j.msec.2017.04.068.
Pack, Daniel W., Allan S. Hoffman, Suzie Pun, and Patrick S. Stayton. 2005.
“Design and development of polymers for gene delivery.” Nature
reviews Drug discovery 4 (7):581.
Pang, J. J., B. Chang, A. Kumar, S. Nusinowitz, S. M. Noorwez, J. Li, A.
Rani, T. C. Foster, V. A. Chiodo, T. Doyle, H. Li, R. Malhotra, J. T.
Teusner, J. H. McDowell, S. H. Min, Q. Li, S. Kaushal, and W. W.
Hauswirth. 2006. “Gene therapy restores vision-dependent behavior as
well as retinal structure and function in a mouse model of RPE65 Leber
congenital amaurosis.” Mol Ther 13 (3):565-72. doi: 10.1016/j.ymthe.
2005.09.001.
Paquet, D., D. Kwart, A. Chen, A. Sproul, S. Jacob, S. Teo, K. M. Olsen, A.
Gregg, S. Noggle, and M. Tessier-Lavigne. 2016. “Efficient
introduction of specific homozygous and heterozygous mutations using
CRISPR/Cas9.” Nature 533 (7601):125-9. doi: 10.1038/nature17664.
Parker, Maria A., Dongseok Choi, Laura R. Erker, Mark E. Pennesi, Paul
Yang, Elvira N. Chegarnov, Peter N. Steinkamp, Catherine L.
Schlechter, Claire-Marie Dhaenens, and Saddek Mohand-Said. 2016.
“Test–retest variability of functional and structural parameters in
patients with Stargardt disease participating in the SAR422459 gene
therapy trial.” Translational vision science & technology 5 (5):10-10.
Patel, Ashaben, Kishore Cholkar, Vibhuti Agrahari, and Ashim K Mitra.
2013. “Ocular drug delivery systems: an overview.” World journal of
pharmacology 2 (2):47.
Pattanayak, V., S. Lin, J. P. Guilinger, E. Ma, J. A. Doudna, and D. R. Liu.
2013. “High-throughput profiling of off-target DNA cleavage reveals
RNA-programmed Cas9 nuclease specificity.” Nat Biotechnol 31
(9):839-43. doi: 10.1038/nbt.2673.
Pennesi, Mark E., Richard G. Weleber, Paul Yang, Chris Whitebirch,
Beverly Thean, Terence R. Flotte, Margaret Humphries, Elvira

The Application of CRISPR/Cas9 Therapies in Ophthalmology … 153
Chegarnov, Kathleen N. Beasley, and J. Timothy Stout. 2018. “Results
at 5 years after gene therapy for RPE65-deficient retinal dystrophy.”
Human gene therapy 29 (12):1428-1437.
Petris, Gianluca, Antonio Casini, Claudia Montagna, Francesca Lorenzin,
Davide Prandi, Alessandro Romanel, Jacopo Zasso, Luciano Conti,
Francesca Demichelis, and Anna Cereseto. 2017. “Hit and go CAS9
delivered through a lentiviral based self-limiting circuit.” Nature
communications 8:15334.
Pierce, E. A., and J. Bennett. 2015. “The Status of RPE65 Gene Therapy
Trials: Safety and Efficacy.” Cold Spring Harb Perspect Med 5
(9):a017285. doi: 10.1101/cshperspect.a017285.
Plessis, Anne, Arnaud Perrin, J. E. Haber, and Bernard Dujon. 1992. “Site-
specific recombination determined by I-SceI, a mitochondrial group I
intron-encoded endonuclease expressed in the yeast nucleus.” Genetics
130 (3):451-460.
Rajala, A., Y. Wang, Y. Zhu, M. Ranjo-Bishop, J. X. Ma, C. Mao, and R.
V. Rajala. 2014. “Nanoparticle-assisted targeted delivery of eye-specific
genes to eyes significantly improves the vision of blind mice in vivo.”
Nano Lett 14 (9):5257-63. doi: 10.1021/nl502275s.
Ramamoorth, M., and A. Narvekar. 2015. “Non viral vectors in gene
therapy- an overview.” J Clin Diagn Res 9 (1):Ge01-6. doi: 10.7860/
jcdr/2015/10443.5394.
Ran, F. Ann, Le Cong, Winston X. Yan, David A. Scott, Jonathan S.
Gootenberg, Andrea J. Kriz, Bernd Zetsche, Ophir Shalem, Xuebing
Wu, and Kira S. Makarova. 2015. “In vivo genome editing using
Staphylococcus aureus Cas9.” Nature 520 (7546):186.
Ran, F. A., P. D. Hsu, C. Y. Lin, J. S. Gootenberg, S. Konermann, A. E.
Trevino, D. A. Scott, A. Inoue, S. Matoba, Y. Zhang, and F. Zhang.
2013. “Double nicking by RNA-guided CRISPR Cas9 for enhanced
genome editing specificity.” Cell 154 (6):1380-9. doi: 10.1016/j.cell.
2013.08.021.
Regalado, Antonio. 2018. Chinese scientists are creating CRISPR babies. In
MIT Technology Review. USA.

C. B. Tara Moore, Larry A. DeDionisio, Hila Roshanravan et al. 154
Richardson, R., D. Tracey-White, A. Webster, and M. Moosajee. 2017. “The
zebrafish eye—a paradigm for investigating human ocular genetics.”
Eye 31 (1):68.
Roosing, S., A. A. Thiadens, C. B. Hoyng, C. C. Klaver, A. I. den Hollander,
and F. P. Cremers. 2014. “Causes and consequences of inherited cone
disorders.” Prog Retin Eye Res 42:1-26. doi: 10.1016/j.preteyeres.2014.
05.001.
Rouet, Philippe, Fatima Smih, and Maria Jasin. 1994. “Introduction of
double-strand breaks into the genome of mouse cells by expression of a
rare-cutting endonuclease.” Molecular and cellular biology 14
(12):8096-8106.
Ruan, G. X., E. Barry, D. Yu, M. Lukason, S. H. Cheng, and A. Scaria.
2017a. “CRISPR/Cas9-Mediated Genome Editing as a Therapeutic
Approach for Leber Congenital Amaurosis 10.” Mol Ther 25 (2):331-
341. doi: 10.1016/j.ymthe.2016.12.006.
Ruan, Guo-Xiang, Elizabeth Barry, Dan Yu, Michael Lukason, Seng H.
Cheng, and Abraham Scaria. 2017b. “CRISPR/Cas9-mediated genome
editing as a therapeutic approach for Leber congenital amaurosis 10.”
Molecular Therapy 25 (2):331-341.
Rudin, Norah, Elaine Sugarman, and James E. Haber. 1989. “Genetic and
physical analysis of double-strand break repair and recombination in
Saccharomyces cerevisiae.” Genetics 122 (3):519-534.
Saghizadeh, Mehrnoosh, Andrei A. Kramerov, Clive N. Svendsen, and
Alexander V. Ljubimov. 2017. “Concise review: stem cells for corneal
wound healing.” Stem Cells 35 (10):2105-2114.
Sakuma, T., T. Sakamoto, and T. Yamamoto. 2017. “All-in-One CRISPR-
Cas9/FokI-dCas9 Vector-Mediated Multiplex Genome Engineering in
Cultured Cells.” Methods Mol Biol 1498:41-56. doi: 10.1007/978-1-
4939-6472-7_4.
Salzman, R., F. Cook, T. Hunt, H. L. Malech, P. Reilly, B. Foss-Campbell,
and D. Barrett. 2018. “Addressing the Value of Gene Therapy and
Enhancing Patient Access to Transformative Treatments.” Mol Ther 26
(12):2717-2726. doi: 10.1016/j.ymthe.2018.10.017.

The Application of CRISPR/Cas9 Therapies in Ophthalmology … 155
Sapranauskas, Rimantas, Giedrius Gasiunas, Christophe Fremaux,
Rodolphe Barrangou, Philippe Horvath, and Virginijus Siksnys. 2011.
“The Streptococcus thermophilus CRISPR/Cas system provides
immunity in Escherichia coli.” Nucleic acids research 39 (21):9275-
9282.
Sareen, Dhruv, Mehrnoosh Saghizadeh, Loren Ornelas, Michael A. Winkler,
Kavita Narwani, Anais Sahabian, Vincent A Funari, Jie Tang, Lindsay
Spurka, and Vasu Punj. 2014. “Differentiation of human limbal‐derived
induced pluripotent stem cells into limbal‐like epithelium.” Stem Cells
Translational Medicine 3 (9):1002-1012.
Schorderet, Daniel. 2015. “Corneal dystrophies: Overview and summary.”
In Progress in molecular biology and translational science, 73-78.
Elsevier.
Schwank, Gerald, Bon-Kyoung Koo, Valentina Sasselli, Johanna F.
Dekkers, Inha Heo, Turan Demircan, Nobuo Sasaki, Sander Boymans,
Edwin Cuppen, and Cornelis K. van der Ent. 2013. “Functional repair
of CFTR by CRISPR/Cas9 in intestinal stem cell organoids of cystic
fibrosis patients.” Cell stem cell 13 (6):653-658.
Semenova, E., M. M. Jore, K. A. Datsenko, A. Semenova, E. R. Westra, B.
Wanner, J. van der Oost, S. J. Brouns, and K. Severinov. 2011.
“Interference by clustered regularly interspaced short palindromic
repeat (CRISPR) RNA is governed by a seed sequence.” Proc Natl Acad
Sci U S A 108 (25):10098-103. doi: 10.1073/pnas.1104144108.
Serifi, I., E. Tzima, K. Soupsana, Z. Karetsou, D. Beis, and T. Papamarcaki.
2016. “The zebrafish homologs of SET/I2PP2A oncoprotein: expression
patterns and insights into their physiological roles during development.”
Biochem J 473 (24):4609-4627. doi: 10.1042/bcj20160523.
Sharma, A., J. C. Tovey, A. Ghosh, and R. R. Mohan. 2010. “AAV serotype
influences gene transfer in corneal stroma in vivo.” Exp Eye Res 91
(3):440-8. doi: 10.1016/j.exer.2010.06.020.
Shirai, H., M. Mandai, K. Matsushita, A. Kuwahara, S. Yonemura, T.
Nakano, J. Assawachananont, T. Kimura, K. Saito, H. Terasaki, M.
Eiraku, Y. Sasai, and M. Takahashi. 2016. “Transplantation of human
embryonic stem cell-derived retinal tissue in two primate models of

C. B. Tara Moore, Larry A. DeDionisio, Hila Roshanravan et al. 156
retinal degeneration.” Proc Natl Acad Sci U S A 113 (1):E81-90. doi:
10.1073/pnas.1512590113.
Shmakov, Sergey, Omar O. Abudayyeh, Kira S. Makarova, Yuri I. Wolf,
Jonathan S. Gootenberg, Ekaterina Semenova, Leonid Minakhin, Julia
Joung, Silvana Konermann, and Konstantin Severinov. 2015.
“Discovery and functional characterization of diverse class 2 CRISPR-
Cas systems.” Molecular cell 60 (3):385-397.
Singh, P., J. C. Schimenti, and E. Bolcun-Filas. 2015. “A mouse geneticist's
practical guide to CRISPR applications.” Genetics 199 (1):1-15. doi:
10.1534/genetics.114.169771.
Slaymaker, I. M., L. Gao, B. Zetsche, D. A. Scott, W. X. Yan, and F. Zhang.
2016. “Rationally engineered Cas9 nucleases with improved
specificity.” Science 351 (6268):84-8. doi: 10.1126/science.aad5227.
Sluch, Valentin M., O. Davis Chung-ha, Vinod Ranganathan, Justin M.
Kerr, Kellin Krick, Russ Martin, Cynthia A. Berlinicke, Nicholas
Marsh-Armstrong, Jeffrey S. Diamond, and Hai-Quan Mao. 2015.
“Differentiation of human ESCs to retinal ganglion cells using a
CRISPR engineered reporter cell line.” Scientific reports 5:16595.
Sternberg, S. H., S. Redding, M. Jinek, E. C. Greene, and J. A. Doudna.
2014. “DNA interrogation by the CRISPR RNA-guided endonuclease
Cas9.” Nature 507 (7490):62-7. doi: 10.1038/nature13011.
Stoddard, Barry L. 2011. “Homing endonucleases: from microbial genetic
invaders to reagents for targeted DNA modification.” Structure 19 (1):7-
15.
Taketani, Yukako, Kohdai Kitamoto, Toshihiro Sakisaka, Mikiko
Kimakura, Tetsuya Toyono, Satoru Yamagami, Shiro Amano,
Masahiko Kuroda, Tara Moore, and Tomohiko Usui. 2017. “Repair of
the TGFBI gene in human corneal keratocytes derived from a granular
corneal dystrophy patient via CRISPR/Cas9-induced homology-
directed repair.” Scientific reports 7 (1):16713.
Taylor, Scott M, Karen Alvarez-Delfin, Carole J Saade, Jennifer L Thomas,
Ryan Thummel, James M Fadool, and Peter F Hitchcock. 2015. “The
bHLH transcription factor NeuroD governs photoreceptor genesis and

The Application of CRISPR/Cas9 Therapies in Ophthalmology … 157
regeneration through delta-notch signaling.” Investigative
ophthalmology & visual science 56 (12):7496-7515.
Testa, F., A. M. Maguire, S. Rossi, E. A. Pierce, P. Melillo, K. Marshall, S.
Banfi, E. M. Surace, J. Sun, C. Acerra, J. F. Wright, J. Wellman, K. A.
High, A. Auricchio, J. Bennett, and F. Simonelli. 2013. “Three-year
follow-up after unilateral subretinal delivery of adeno-associated virus
in patients with Leber congenital Amaurosis type 2.” Ophthalmology
120 (6):1283-91. doi: 10.1016/j.ophtha.2012.11.048.
Trapani, I., S. Banfi, F. Simonelli, E. M. Surace, and A. Auricchio. 2015.
“Gene therapy of inherited retinal degenerations: prospects and
challenges.” Hum Gene Ther 26 (4):193-200. doi: 10.1089/hum.
2015.030.
Tsai, S. Q., N. T. Nguyen, J. Malagon-Lopez, V. V. Topkar, M. J. Aryee,
and J. K. Joung. 2017. “CIRCLE-seq: a highly sensitive in vitro screen
for genome-wide CRISPR-Cas9 nuclease off-targets.” Nat Methods 14
(6):607-614. doi: 10.1038/nmeth.4278.
Tsai, S. Q., N. Wyvekens, C. Khayter, J. A. Foden, V. Thapar, D. Reyon, M.
J. Goodwin, M. J. Aryee, and J. K. Joung. 2014. “Dimeric CRISPR
RNA-guided FokI nucleases for highly specific genome editing.” Nat
Biotechnol 32 (6):569-76. doi: 10.1038/nbt.2908.
Tsai, S. Q., Z. Zheng, N. T. Nguyen, M. Liebers, V. V. Topkar, V. Thapar,
N. Wyvekens, C. Khayter, A. J. Iafrate, L. P. Le, M. J. Aryee, and J. K.
Joung. 2015. “GUIDE-seq enables genome-wide profiling of off-target
cleavage by CRISPR-Cas nucleases.” Nat Biotechnol 33 (2):187-197.
doi: 10.1038/nbt.3117.
Tsubota, K., H. Inoue, K. Ando, M. Ono, K. Yoshino, and I. Saito. 1998.
“Adenovirus-mediated gene transfer to the ocular surface epithelium.”
Exp Eye Res 67 (5):531-8. doi: 10.1006/exer.1998.0557.
Tucker, B. A., R. F. Mullins, and E. M. Stone. 2014. “Stem cells for
investigation and treatment of inherited retinal disease.” Hum Mol Genet
23 (R1):R9-r16. doi: 10.1093/hmg/ddu124.
Urnov, Fyodor D., Edward J. Rebar, Michael C. Holmes, H. Steve Zhang,
and Philip D Gregory. 2010. “Genome editing with engineered zinc
finger nucleases.” Nature Reviews Genetics 11 (9):636.

C. B. Tara Moore, Larry A. DeDionisio, Hila Roshanravan et al. 158
Vance, Melisa, Telmo Llanga, Will Bennett, Kenton Woodard, Giridhar
Murlidharan, Neil Chungfat, Aravind Asokan, Brian Gilger, Joanne
Kurtzberg, and R Jude Samulski. 2016. “AAV gene therapy for MPS1-
associated corneal blindness.” Scientific reports 6:22131.
Veres, A., B. S. Gosis, Q. Ding, R. Collins, A. Ragavendran, H. Brand, S.
Erdin, C. A. Cowan, M. E. Talkowski, and K. Musunuru. 2014. “Low
incidence of off-target mutations in individual CRISPR-Cas9 and
TALEN targeted human stem cell clones detected by whole-genome
sequencing.” Cell Stem Cell 15 (1):27-30. doi: 10.1016/j.stem.2014.
04.020.
Wang, L., R. Xiao, E. Andres-Mateos, and L. H. Vandenberghe. 2017.
“Single stranded adeno-associated virus achieves efficient gene transfer
to anterior segment in the mouse eye.” PLoS One 12 (8):e0182473. doi:
10.1371/journal.pone.0182473.
Weiss, J. S., H. U. Moller, A. J. Aldave, B. Seitz, C. Bredrup, T. Kivela, F.
L. Munier, C. J. Rapuano, K. K. Nischal, E. K. Kim, J. Sutphin, M.
Busin, A. Labbe, K. R. Kenyon, S. Kinoshita, and W. Lisch. 2015.
“IC3D classification of corneal dystrophies--edition 2.” Cornea 34
(2):117-59. doi: 10.1097/ico.0000000000000307.
Weleber, Richard G., Mark E. Pennesi, David J. Wilson, Shalesh Kaushal,
Laura R. Erker, Lauren Jensen, Maureen T. McBride, Terence R. Flotte,
Margaret Humphries, and Roberto Calcedo. 2016. “Results at 2 years
after gene therapy for RPE65-deficient Leber congenital amaurosis and
severe early-childhood–onset retinal dystrophy.” Ophthalmology 123
(7):1606-1620.
Willi, Michaela, Harold E. Smith, Chaochen Wang, Chengyu Liu, and
Lothar Hennighausen. 2018. “Mutation frequency is not increased in
CRISPR-Cas9-edited mice.” Nature methods 15 (10):756-758.
Wood, E. H., P. H. Tang, I. De la Huerta, E. Korot, S. Muscat, D. A.
Palanker, and G. A. Williams. 2019. “STEM CELL THERAPIES,
GENE-BASED THERAPIES, OPTOGENETICS, AND RETINAL
PROSTHETICS: Current State and Implications for the Future.” Retina
39 (5):820-835. doi: 10.1097/iae.0000000000002449.

The Application of CRISPR/Cas9 Therapies in Ophthalmology … 159
Wu, W. H., Y. T. Tsai, S. Justus, T. T. Lee, L. Zhang, C. S. Lin, A. G.
Bassuk, V. B. Mahajan, and S. H. Tsang. 2016. “CRISPR Repair
Reveals Causative Mutation in a Preclinical Model of Retinitis
Pigmentosa.” Mol Ther 24 (8):1388-94. doi: 10.1038/mt.2016.107.
Wu, X., D. A. Scott, A. J. Kriz, A. C. Chiu, P. D. Hsu, D. B. Dadon, A. W.
Cheng, A. E. Trevino, S. Konermann, S. Chen, R. Jaenisch, F. Zhang,
and P. A. Sharp. 2014. “Genome-wide binding of the CRISPR
endonuclease Cas9 in mammalian cells.” Nat Biotechnol 32 (7):670-6.
doi: 10.1038/nbt.2889.
Wu, Y., D. Liang, Y. Wang, M. Bai, W. Tang, S. Bao, Z. Yan, D. Li, and J.
Li. 2013. “Correction of a genetic disease in mouse via use of CRISPR-
Cas9.” Cell Stem Cell 13 (6):659-62. doi: 10.1016/j.stem.2013.10.016.
Wyvekens, N., V. V. Topkar, C. Khayter, J. K. Joung, and S. Q. Tsai. 2015.
“Dimeric CRISPR RNA-Guided FokI-dCas9 Nucleases Directed by
Truncated gRNAs for Highly Specific Genome Editing.” Hum Gene
Ther 26 (7):425-31. doi: 10.1089/hum.2015.084.
Xie, K., B. Minkenberg, and Y. Yang. 2015. “Boosting CRISPR/Cas9
multiplex editing capability with the endogenous tRNA-processing
system.” Proc Natl Acad Sci U S A 112 (11):3570-5. doi:
10.1073/pnas.1420294112.
Xu, L., L. Zhao, Y. Gao, J. Xu, and R. Han. 2017. “Empower multiplex cell
and tissue-specific CRISPR-mediated gene manipulation with self-
cleaving ribozymes and tRNA.” Nucleic Acids Res 45 (5):e28. doi:
10.1093/nar/gkw1048.
Xu, Peng, Ying Tong, Xiu-zhen Liu, Ting-ting Wang, Li Cheng, Bo-yu
Wang, Xiang Lv, Yue Huang, and De-pei Liu. 2015. “Both TALENs
and CRISPR/Cas9 directly target the HBB IVS2–654 (C> T) mutation
in β-thalassemia-derived iPSCs.” Scientific reports 5:12065.
Xu, T., Y. Li, J. D. Van Nostrand, Z. He, and J. Zhou. 2014. “Cas9-based
tools for targeted genome editing and transcriptional control.” Appl
Environ Microbiol 80 (5):1544-52. doi: 10.1128/aem.03786-13.
Yamano, T., H. Nishimasu, B. Zetsche, H. Hirano, I. M. Slaymaker, Y. Li,
I. Fedorova, T. Nakane, K. S. Makarova, E. V. Koonin, R. Ishitani, F.
Zhang, and O. Nureki. 2016. “Crystal Structure of Cpf1 in Complex

C. B. Tara Moore, Larry A. DeDionisio, Hila Roshanravan et al. 160
with Guide RNA and Target DNA.” Cell 165 (4):949-62. doi:
10.1016/j.cell.2016.04.003.
Yan, Sen, Zhuchi Tu, Zhaoming Liu, Nana Fan, Huiming Yang, Su Yang,
Weili Yang, Yu Zhao, Zhen Ouyang, and Chengdan Lai. 2018. “A
huntingtin knockin pig model recapitulates features of selective
neurodegeneration in Huntington’s disease.” Cell 173 (4):989-1002.
e13.
Yan, W. X., R. Mirzazadeh, S. Garnerone, D. Scott, M. W. Schneider, T.
Kallas, J. Custodio, E. Wernersson, Y. Li, L. Gao, Y. Federova, B.
Zetsche, F. Zhang, M. Bienko, and N. Crosetto. 2017. “BLISS is a
versatile and quantitative method for genome-wide profiling of DNA
double-strand breaks.” Nat Commun 8:15058. doi: 10.1038/
ncomms15058.
Yang, Hui, Haoyi Wang, Chikdu S Shivalila, Albert W Cheng, Linyu Shi,
and Rudolf Jaenisch. 2013. “One-step generation of mice carrying
reporter and conditional alleles by CRISPR/Cas-mediated genome
engineering.” Cell 154 (6):1370-1379.
Yin, C., T. Zhang, X. Qu, Y. Zhang, R. Putatunda, X. Xiao, F. Li, W. Xiao,
H. Zhao, S. Dai, X. Qin, X. Mo, W. B. Young, K. Khalili, and W. Hu.
2017. “In Vivo Excision of HIV-1 Provirus by saCas9 and Multiplex
Single-Guide RNAs in Animal Models.” Mol Ther 25 (5):1168-1186.
doi: 10.1016/j.ymthe.2017.03.012.
Yin, H., C. Q. Song, J. R. Dorkin, L. J. Zhu, Y. Li, Q. Wu, A. Park, J. Yang,
S. Suresh, A. Bizhanova, A. Gupta, M. F. Bolukbasi, S. Walsh, R. L.
Bogorad, G. Gao, Z. Weng, Y. Dong, V. Koteliansky, S. A. Wolfe, R.
Langer, W. Xue, and D. G. Anderson. 2016. “Therapeutic genome
editing by combined viral and non-viral delivery of CRISPR system
components in vivo.” Nat Biotechnol 34 (3):328-33. doi:
10.1038/nbt.3471.
Yin, Hao, Chun-Qing Song, Sneha Suresh, Qiongqiong Wu, Stephen Walsh,
Luke Hyunsik Rhym, Esther Mintzer, Mehmet Fatih Bolukbasi, Lihua
Julie Zhu, and Kevin Kauffman. 2017. “Structure-guided chemical
modification of guide RNA enables potent non-viral in vivo genome
editing.” Nature biotechnology 35 (12):1179.

The Application of CRISPR/Cas9 Therapies in Ophthalmology … 161
Yin, Linlin, Lisette A. Maddison, Mingyu Li, Nergis Kara, Matthew C.
LaFave, Gaurav K. Varshney, Shawn M. Burgess, James G. Patton, and
Wenbiao Chen. 2015. “Multiplex conditional mutagenesis using
transgenic expression of Cas9 and sgRNAs.” Genetics 200 (2):431-441.
Young, R. W. 1985. “Cell differentiation in the retina of the mouse.” Anat
Rec 212 (2):199-205. doi: 10.1002/ar.1092120215.
Yu, Wenhan, Suddhasil Mookherjee, Vijender Chaitankar, Suja Hiriyanna,
Jung-Woong Kim, Matthew Brooks, Yasaman Ataeijannati, Xun Sun,
Lijin Dong, and Tiansen Li. 2017. “Nrl knockdown by AAV-delivered
CRISPR/Cas9 prevents retinal degeneration in mice.” Nature
communications 8:14716.
Yuan, L., T. Sui, M. Chen, J. Deng, Y. Huang, J. Zeng, Q. Lv, Y. Song, Z.
Li, and L. Lai. 2016. “CRISPR/Cas9-mediated GJA8 knockout in
rabbits recapitulates human congenital cataracts.” Sci Rep 6:22024. doi:
10.1038/srep22024.
Zarei, Ali, Vahid Razban, Seyed Ebrahim Hosseini, and Seyed Mohammad
Bagher Tabei. 2019. “Creating Cell and Animal Models of Human
Disease by Genome Editing Using CRISPR/Cas9.” The journal of gene
medicine:e3082.
Zeitz, C., A. G. Robson, and I. Audo. 2015. “Congenital stationary night
blindness: an analysis and update of genotype-phenotype correlations
and pathogenic mechanisms.” Prog Retin Eye Res 45:58-110. doi:
10.1016/j.preteyeres.2014.09.001.
Zetsche, B., M. Heidenreich, P. Mohanraju, I. Fedorova, J. Kneppers, E. M.
DeGennaro, N. Winblad, S. R. Choudhury, O. O. Abudayyeh, J. S.
Gootenberg, W. Y. Wu, D. A. Scott, K. Severinov, J. van der Oost, and
F. Zhang. 2017. “Multiplex gene editing by CRISPR-Cpf1 using a single
crRNA array.” Nat Biotechnol 35 (1):31-34. doi: 10.1038/nbt.3737.
Zetsche, B., S. E. Volz, and F. Zhang. 2015. “A split-Cas9 architecture for
inducible genome editing and transcription modulation.” Nat Biotechnol
33 (2):139-42. doi: 10.1038/nbt.3149.
Zetsche, Bernd, Jonathan S Gootenberg, Omar O Abudayyeh, Ian M
Slaymaker, Kira S Makarova, Patrick Essletzbichler, Sara E Volz, Julia
Joung, John Van Der Oost, and Aviv Regev. 2015. “Cpf1 is a single

C. B. Tara Moore, Larry A. DeDionisio, Hila Roshanravan et al. 162
RNA-guided endonuclease of a class 2 CRISPR-Cas system.” Cell 163
(3):759-771.
Zhang, J. P., X. L. Li, G. H. Li, W. Chen, C. Arakaki, G. D. Botimer, D.
Baylink, L. Zhang, W. Wen, Y. W. Fu, J. Xu, N. Chun, W. Yuan, T.
Cheng, and X. B. Zhang. 2017. “Efficient precise knockin with a double
cut HDR donor after CRISPR/Cas9-mediated double-stranded DNA
cleavage.” Genome Biol 18 (1):35. doi: 10.1186/s13059-017-1164-8.
Zheng, Q., X. Cai, M. H. Tan, S. Schaffert, C. P. Arnold, X. Gong, C. Z.
Chen, and S. Huang. 2014. “Precise gene deletion and replacement using
the CRISPR/Cas9 system in human cells.” Biotechniques 57 (3):115-24.
doi: 10.2144/000114196.
Zhong, H., Y. Chen, Y. Li, R. Chen, and G. Mardon. 2015. “CRISPR-
engineered mosaicism rapidly reveals that loss of Kcnj13 function in
mice mimics human disease phenotypes.” Sci Rep 5:8366. doi:
10.1038/srep08366.
Zinn, E., and L. H. Vandenberghe. 2014. “Adeno-associated virus: fit to
serve.” Curr Opin Virol 8:90-7. doi: 10.1016/j.coviro.2014.07.008.
Zuo, Erwei, Yi-Jun Cai, Kui Li, Yu Wei, Bang-An Wang, Yidi Sun, Zhen
Liu, Jiwei Liu, Xinde Hu, and Wei Wei. 2017. “One-step generation of
complete gene knockout mice and monkeys by CRISPR/Cas9-mediated
gene editing with multiple sgRNAs.” Cell research 27 (7):933.
Zuris, John A., David B. Thompson, Yilai Shu, John P. Guilinger, Jeffrey L.
Bessen, Johnny H. Hu, Morgan L. Maeder, J. Keith Joung, Zheng-Yi
Chen, and David R. Liu. 2015. “Cationic lipid-mediated delivery of
proteins enables efficient protein-based genome editing in vitro and in
vivo.” Nature biotechnology 33 (1):73.