tapering dose of hydrocortisone and dexamethasone...
TRANSCRIPT

Dexamethasone and hydrocortisone in neonatal rats
1
Tapering Dose of Hydrocortisone and Dexamethasone in Neonatal Rats: Effect on the
Hypothalamic-Pituitary-Adrenal Response, Somatic and Neurological Development at
Adolescence
Varsha Bhatt-Mehta 1,2
, Charles R Neal Jr3, Lidong S Huang
2 , Mercy Pawar
2,
Delia M Vazquez2
1College of Pharmacy and
2Department of Pediatrics, University of Michigan, Ann Arbor, MI, USA,
3
University of Hawaii John A. Burns School of Medicine ,Honolulu, HI,USA
Corresponding Author:
Varsha Bhatt-Mehta, MS (CRDSA), Pharm.D., FCCP
College of Pharmacy and Department of Pediatrics
F5790/0254
200 East Hospital Drive
Ann Arbor, MI 48109
734-763-4109 (Office)
734-763-7728 (Fax)

Dexamethasone and hydrocortisone in neonatal rats
2
Disclosures:
The authors have no financial relationships relevant to this article.
Grant support:
Supported by NIH grant RO1 HDDK37431

Dexamethasone and hydrocortisone in neonatal rats
3
Abstract
Exogenous administration of dexamethasone (DEX) or hydrocortisone (HC) to improve lung function and treat
refractory hypotension is common in premature infants. This occurs at a time when the central nervous system is
undergoing profound structural and functional transformations. This study was designed to evaluate the effect of
tapering doses of DEX and HC in neonatal rats with timing of exposure corresponding developmentally to times
these agents are used in clinical practice. A within litter design was used. Intramuscular DEX, HC or saline (VEH-
control) were administered to newborn pups on postnatal days (PD) 5 and 6. Neurological development and growth
was assessed weekly until pre-adolescence (PD33). At PD33 animals were submitted to novelty stress and anxiety
testing using the light-dark preference box (L-D) and Corticosterone (CORT) and ACTH levels measured after
novelty stress. Somatic growth was decreased in the DEX group compared with HC, and VEH animals at all ages.
HC animals showed increased weight gain by PD33. Neurodevelopment was appropriate for age at all times
measured. No treatment differences were appreciated between groups. CORT secretion decreased in DEX group
compared with all other groups. HC animals had significantly higher ACTH levels long after all other groups
returned to basal levels. Animals did not differ in L-D testing. A short tapering course of DEX and HC appears to be
more benign than prolonged DEX exposure in early life. The effects of HC on Limbic-Hypothalamic-Pituitary-
Adrenal (LHPA) axis, though subtle, raises concerns for altered LHPA function and potential risk for metabolic
disorders in later life.
Key words: Neonatal rats, Neurodevelopment, Dexamethasone, Hydrocortisone, ACTH, Cortisol. HPA-axis
Precie:
The present study evaluated the effects of a short course of HC on somatic growth and the developing LHPA axis. A
two day tapering dose glucocorticoid regimen was used.

Dexamethasone and hydrocortisone in neonatal rats
4
Introduction
Exogenously administered glucocorticoids, such as dexamethasone (DEX), can inhibit the stress response of the
limbic-hypothalamic-pituitary–adrenal (LHPA) axis for prolonged periods and cause adverse effects on the growth
and development in both animals and humans (1-7). In humans, DEX has been used in tapering courses lasting 28
days or more in the extremely low birth weight (ELBW) infants to reduce the morbidity of chronic lung disease
(CLD) (8-11). In practice, the timing of DEX use in ELBW infants has corresponded to corrected gestational ages
when the central nervous system (CNS) is undergoing profound structural and functional transformations, making it
vulnerable to external influences such as DEX exposure. Follow up studies in the late 1990’s provided evidence of
significant increase in neurodevelopmental dysfunction in preterm neonates who received prolonged courses of DEX
to reduce morbidity associated with chronic lung disease(12) As a result of such findings DEX use has been
significantly reduced in clinical practice and when used the courses are substantially shorter (13-15).. While the use
of DEX for chronic lung disease has been restricted substantially, few alternative treatment options are available.
In recent years, hydrocortisone (HC) has emerged as a treatment option for premature infants with respiratory
distress, chronic lung disease (CLD), and hypotension (16,17, 18,19). Hypotension can be secondary to multiple
causes (e.g. PDA, sepsis). In practice, when used for CLD in preterm infants, DEX and HC are used later and later in
chronological age (20, 17, 21,18,19, 22). The use of low-dose HC increases the likelihood of survival in premature
infants without chronic lung disease (23,24). As with early use of DEX, the long term effects of HC on the
developing CNS in preterm and term neonates remains unknown, though small studies suggest that HC may not
significantly predict adverse neurodevelopment (25, 17,18,19) .
In recent years, animal models have been developed in an attempt to better understand the neurological
consequences of perinatal glucocorticoid exposure and long term neurodevelopmental outcomes (26,27,28). When
interpreting behavioral and physiologic effects of neonatal glucocorticoids in any developmental model, it is assumed
that although timing differs significantly between species, the general sequence of brain growth is similar. An
appropriate rodent model that uses these relationships has been developed in our laboratory. Most important in our
decision to use a rodent model is the ability to take advantage of the unique neurodevelopmental profile present

Dexamethasone and hydrocortisone in neonatal rats
5
during the neonatal period. It is estimated that on postnatal day 10 the rodent brain is roughly equivalent to that of the
human brain of 38 to 40 weeks post-conception(29,30). Extrapolating from this, the brain of a rodent pup at birth
(postnatal day 1-PD1) corresponds to that of a human fetal brain at or near 21-22 weeks gestation (30, 31,32). The
brain of a PD3 pup approximates that of a 24-26 wk human and PD6 approximates a 30-32 wk human. We therefore
modeled the timing of DEX exposure in this model to that utilized in premature infants. We have demonstrated
changes in LHPA function and anxiogenic behavior in adolescent animals exposed to DEX during the first week of
life when DEX was given at PD3, 4, 5 and 6, indicating long term repercussions of such treatment (6,33)..
In view of present neonatal practices using short course DEX or HC in high risk premature infants, the
purpose of the present study was to evaluate the effects of a short course of HC on somatic growth and the
developing LHPA axis. We chose a two day tapering dose regimen starting on PD5 to capture a common clinical
scenario in the human infant where glucocorticoid exposure is given later in life to minimize chronic lung morbidity.
As in our previous DEX neonatal rat model, we started with a HC dose equivalent to that used in the human neonate,
comparing it to DEX as well as untreated controls.
Materials and Methods
Animals
Litter management and animal handling of neonatal rats in this study was similar to that reported from our
laboratories previously (6, 34) Adult Sprague-Dawley rats (Charles Rivers, Wilmington, MA) were housed and
treated according to Guide for the Care and Use of Laboratory animals. All animals were kept under constant
temperature (25 + 2oC) and photoperiodicity (14:10h light-dark cycle) and provided with food and water ad libitum.
Animals were mated using one to one mating system (1F:1M). Assuming a 21-day gestation, pregnant females were
housed separately, starting on day 18. They were then checked twice daily until pups were born. The day of the birth
of the pups was designated as postnatal day one (PD1).
Consistent with our previous studies, each litter was sexed and culled to 12 pups (6M: 6F) on the second day of life.
Pups were randomly selected as male: female pairs from 3 different dams giving birth the same day. This ensured
both genetic diversity, equality in nutrition and maternal care within litters. Within each litter, pups were randomly

Dexamethasone and hydrocortisone in neonatal rats
6
chosen to one of three groups: VEH, DEX , or HC. Pups were permanently marked for identification according to
their treatment group by clipping the outer toe tip of the rear left paw (DEX), rear right paw (HC) and front left paw
(VEH). These permanent markings were visible throughout the experiment period negating the need for further
animal tagging and minimized handling for identification. A total of 144 animals were studied.
Drug Treatment
Pups were removed from their mothers between the hours of 1100 and 1300. Animals in the DEX group received an
intramuscular (IM) injection of DEX in a tapering dose of 0.5 mg/kg on PD5 and 0.1 mg/kg on PD6. HC animals
received an IM HC injection at 5.0 mg/kg on PD5 and 1.0 mg/kg on PD6. VEH animals received an equivalent
volume of sterile normal saline IM. The study protocol and days of treatment are depicted in Table 1.
Somatic Growth: Lengths and weights were measured for each pup in each treatment group before handling or
treatments on PD5 and PD6. Animals were also measured and weighed on PD 7, 14, 20 and prior to stress testing on
PD33. At each age, length was measured from the nose to the base of the tail (head-rump length).
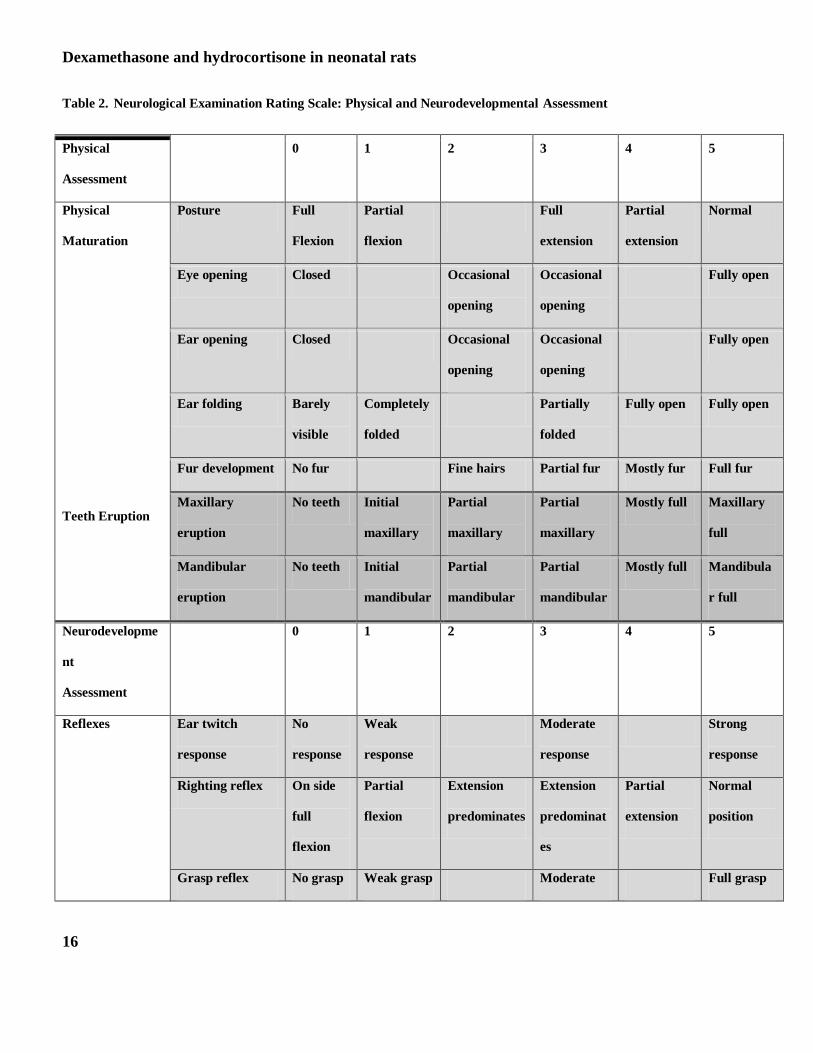
Neurological assessment: Procedures for the neurological assessment were performed on PD 7, 14 and 20 and were
adapted from work of several investigators (26,27, 28). Results using this method of neurodevelopmental assessment
have also been reported by our group (6, 34) Each pup underwent testing at the same time on each designated
neurodevelopmental assessment day. Two trained assistants measured neurodevelopmental indices in all animals
using a rating of 0-5 with a score of 5 corresponding to maturity. Neurodevelopment assessment in the neonate
included posture, righting reflex, postural flexion and extension, vibrissa placing, fore limb and hind limb placing,
geotaxis, and bar hold and cross extension. Physical maturity was measured by observing eye opening, ear opening,
ear folding, ear twitch, fur development and tooth eruption. In order to obtain meaningful analysis of physical and
neurological assessment these observations were grouped, as presented in Table 2. The grouping was done as
follows: physical maturation (posture, eye opening, ear opening and fur development); teeth eruption (maxillary and
mandibular eruption); reflexes (ear twitch, righting reflex, postural flexion and extension and fore and hind limb

Dexamethasone and hydrocortisone in neonatal rats
7
grasp reflex); sensory complex (Vibrissa placing and geotaxis); and gross motor activity (fore and hind limb placing,
cross limb and bar hold).
Brain Weight: On PD8, one pup per treatment group was sacrificed to obtain brain weights. Brain weights were also
obtained during necropsy on PD33.
Behavioral Testing: A total of 144 pups were ultimately studied at adolescence.
The light-dark preference box was used to evaluate locomotion and investigatory behavior of the animals on PD33 in
a manner previously reported by this laboratory (34). Preference for darkness and decreased activity are gross
measurements of anxiety. At age PD30, animals were acclimated to handling before the start of the procedure.
Handling consisted in transporting the cage to the test room and removal from the cage. This was done for three
consecutive days. The testing apparatus was a covered 30 x 60 x 30 cm Plexiglas shuttle-box with a computerized
monitor, two equal sized compartments (light and dark) with a 12-cm wide opening and a stainless steel grid floor
suspended above corncob bedding. The light compartment was constructed of white Plexiglas and brightly
illuminated. The dark compartment was constructed of black Plexiglas and minimally illuminated. Fluorescent lights
above the box provided the illumination. To begin the session, the animal was placed in the dark compartment and
the timer set to start. Each animal’s locomotor activity, the time spent in each compartment, the number of transitions
and the latency to leave the dark were scored. Locomotor activity as well as time spent in each compartment was
monitored by photocells located on the wall of each box, with the number of photocell beams interrupted per unit
time recorded by microprocessor. The number of transitions were recorded manually. Total testing time was 5
minutes (6).
Adrenocortical Response to Novelty Stress: After light-dark preference testing was completed, blood was collected
from the animal’s tail vein by cannulation at 15, 30 and 60 min after the beginning of the testing session. The last
blood sample was collected 90 min after the start of the test, at which time the animals were decapitated. A pre-stress
blood sample was obtained the day prior to the light-dark box as the animal was acclimated to handling. The time of
the pre-stress sample corresponded to the same time of the start of the procedure on the following day. Blood
samples were collected in pre-chilled tubes containing EDTA, placed on ice and subsequently spun at 2000 rpm for 7

Dexamethasone and hydrocortisone in neonatal rats
8
min. The plasma was separated and stored at -20 0C until assayed for corticosterone (CORT) and
adrenocorticotrophic (ACTH) hormone concentrations.
Hormonal Assays
Corticosterone levels were measured using a commercially available corticosterone I125
radioimmunoassay kit (Cat.
#07-120102, MP Biomedicals LLC, Diagnostic Division, Orangeburg, New York) after plasma samples were diluted
to 1:200 in Phosphosaline gelatin buffer (pH 7.0). The intra- and inter assay CVs for corticosterone were 4.4% and
6.5%, respectively.
Plasma adrenocorticotrophic hormone concentrations were measured using the ImmuChem™ Double antibody
hACTH 125
I RIA kit (MP Biomedicals LLC. Orangeburg, NY 10962). This assay uses un-extracted plasma with
EDTA as an anticoagulant. A standard curve was generated using specified amount of antibody reacted with
radiolabeled hormone per manufacturer’s instructions. This standard curve was then used to measure unknown
amounts of ACTH in the study samples. The intra and inter day coefficient of variation for the assay was 6.8% and
10.7%, respectively.
Statistical Analyses
Body weight and length were analyzed using repeated measures ANOVA. Total and individual neurological scores
were averaged across groups and analyzed using nonparametric test (Kruskal-Wallis). The effect of sex and HC or
DEX treatment on various neurodevelopmental outcomes on PD 7, 14 and 21 was compared. Brain weights, light-
dark behavior and plasma hormonal levels (ACTH and CORT) were analyzed using repeated measures ANOVA.
Significance was indicated by a p value p<0.05. Once significance was observed by ANOVA, the Fisher’s least
significant difference (Fisher’s PLSD) method was utilized for further pair-wise comparisons.
Results

Dexamethasone and hydrocortisone in neonatal rats
9
Somatic Growth
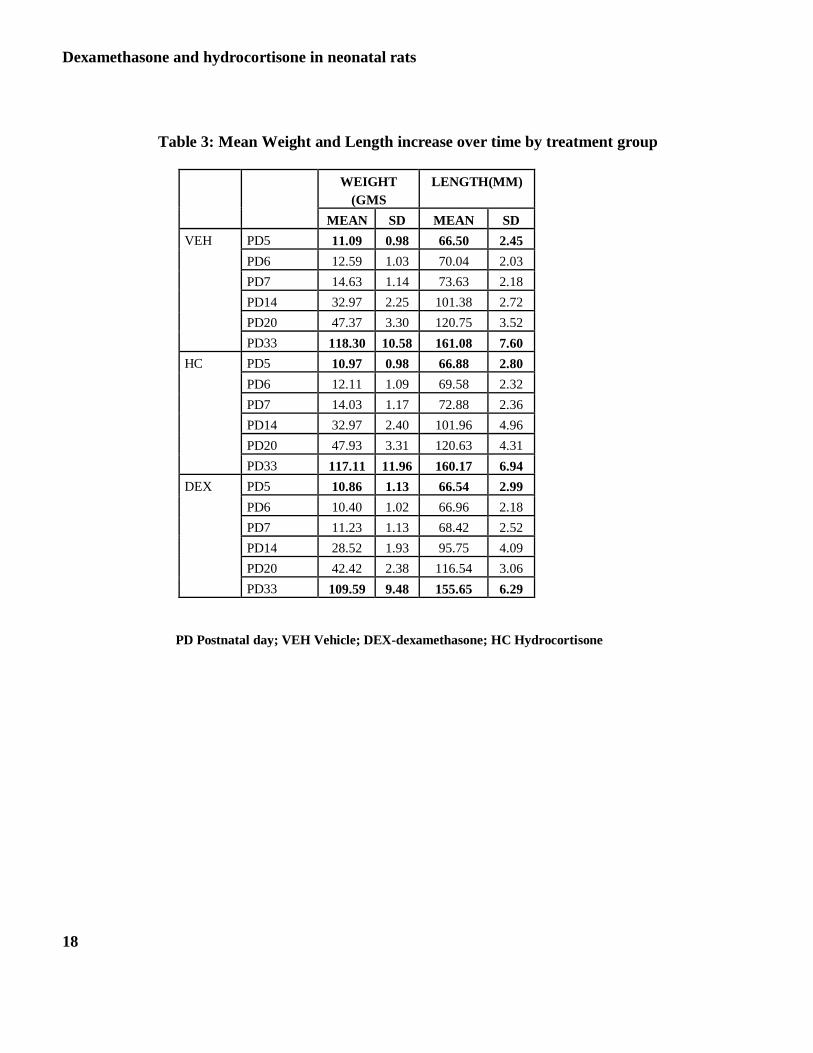
The statistically significant relationships for the parameters measured under somatic growth are presented in Table 3
and figures 1and 2.
Weight: The overall change in weight amongst the three treatment groups on days PD5-PD20 is presented in Figure
1-Panel A. As shown, weight gain in the DEX treated animals was significantly lower than in the HC and VEH
groups on days PD6 - PD20 (P<0.05). There were no sex differences during this period. The interaction between
sex and treatment was significant for PD33. Further analysis did not result in any difference in the male animals in
the three treatment groups (Panel B) but DEX treated female animals were significantly smaller than the VEH group
on PD33 (Panel C; P<0.05).
Length: The results of effect of the three treatments on length are presented in Figure 2-Panels A and B. Initial
examination revealed a significant interaction between sex by time (P<0.0002) and treatment by time (P< 0.0018).
Thus, males and females were analyzed separately. DEX treated female and male animals were significantly shorter
compared with HC and VEH on PD 6, PD7, PD14 and PD20 (P<0.0001). On PD33 males in all 3 treatment groups
had similar length. However, DEX treated females were significantly shorter than those in the VEH or HC group on
PD33 (P< 0.001).
Neurological Development
Figure 3 -Panels A-E depict the results of the neurological development. Animals were studied on three different
postnatal time points: PD 7, PD14 and PD21. Following repeated measure ANOVA, we did not observe significant
effect of sex on any neurodevelopment exams within treatment groups. Male and female data was therefore collapsed
for further analysis. There were neurological functioning differences across time with a significant age by
neurological assessment interaction (p < 0.0001). The post hoc analysis indicated that the pace of development was
not significantly different for the DEX group compared with VEH or HC for all neurological measurements across
the time of assessment with the exception of physical maturation on PD7 and teeth eruption on PD21. The teeth
eruption score was significantly lower for DEX animals compared with HC and VEH on PD21 (Panel A; P<0.05)

Dexamethasone and hydrocortisone in neonatal rats
10
and the physical maturation score was significantly lower for DEX compared with VEH on PD7 (Panel D; P<0.05).
By day 21 the physical maturation was similar in all three treatment groups.
Behavioral Assessment
On testing for place-preference in the light-dark box, there was no difference between treatment groups or between
males and females on latency to enter the light box when placed in the dark compartment (p>0.05). Similarly, the
locomotor activity and time spent in dark or light compartment was similar for males or females across groups. Thus,
all Light-Dark Box Behavioral measures obtained were not significantly different between treatment groups
irrespective of sex (data not shown).
Brain Weights
On PD33 the brain weights of males and females were significantly different within the DEX and HC treatment
groups (ANOVA, p<0.0001). The post hoc analysis showed that DEX treated animals were different from those
treated with HC. Males had greater brain weights when compared to females in both groups. However, when brain
weights were normalized for body weight the ratio of brain:body weight was not significantly different between
treatment groups or for males vs females within each treatment group (data not shown).
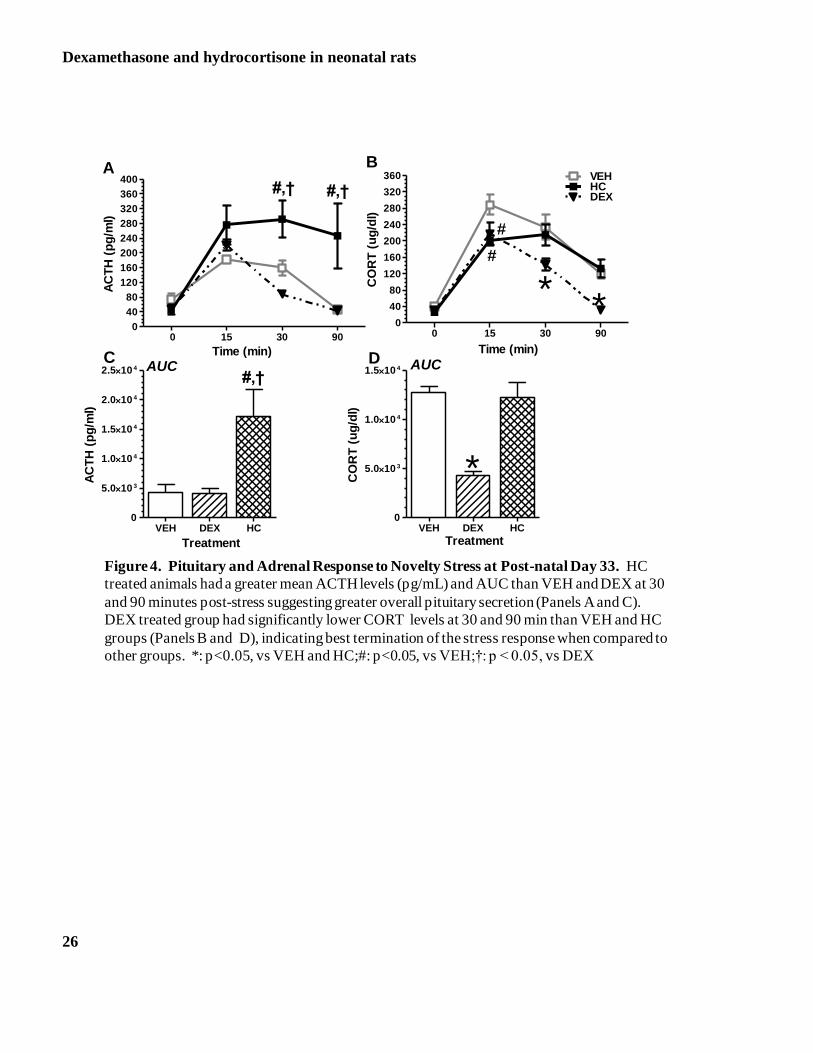
Pituitary Response to Novelty Stress
Results of novelty stress are presented in Figure 4-Panels A to D. Repeated measures ANOVA of ACTH response to
novelty stress revealed no differences between males and females, therefore the data were collapsed across this
variable. We observed significant differences among the groups with HC treated animals having greater mean ACTH
levels when compared to DEX animals (p=0.04). Analysis of the individual profiles of ACTH release at each time
point showed a significant difference across groups at 30 and 90 min (Figure 4, Panel A). At 30 min, HC treated
animals had elevated ACTH levels when compared to DEX and VEH animals. HC treated animals also had
significantly higher ACTH levels at 90 min, a time that all other groups had returned close to basal ACTH levels.
Consistent with this profile, the AUC analysis showed significantly greater overall ACTH secretion by HC treated
animals (Figure 4 –Panel C).
Adrenocortical Response to Novelty Stress

Dexamethasone and hydrocortisone in neonatal rats
11
Repeated measures ANOVA revealed that the adrenocortical activation by novelty stress was similar in both male
and females, therefore the analysis was collapsed across this variable and revealed significance across groups
(p=0.02). A significant CORT by group interaction was also present (p=0.002). The DEX treated animals had a lower
response when compared to all other groups. When we analyzed the group profiles using factorial measure ANOVA,
all groups showed peak CORT levels at 15 min, with the VEH group having the greatest response (p<0.05) All
groups returned to baseline CORT levels by 90 minutes post-stress. However, the HC and VEH treated animals
remained with significantly higher CORT levels at 90 min when compared to DEX groups. Animals treated with
DEX had significantly lower CORT levels much earlier than the other groups, at 30 min post stress (p<0.05)(Figure 4
–Panel B). These data were consistent with the AUC analysis which revealed that the overall secretion of CORT in
response to novelty stress was decreased for the DEX treated animals when compared to all other groups (p<0.001;
Figure 4-panel D).
Discussion
Long term DEX therapy in the newborn period has been shown to lead to undesirable neurological sequela in
extremely low birth weight infants (35, 36). Consequently, the use of DEX in current clinical practice has decreased
markedly over the past decade. Moreover, the comparative safety of other therapeutic alternative, such as HC or short
course DEX, has been minimally explored (16, 38, 23, 24).
The present study has used an animal model to ascertain early and long term effects of DEX and HC, at doses
commonly used in clinical practice, given during first week of life. The timing of drug administration was important
in our study because this is a period of time that captures a common clinical scenario in the older extremely
premature infant that is developing severe chronic lung disease. It is also a time when the neonatal brain appears to
be most vulnerable to insults (29,30).. It is important to note that this rat model in no way is attempting to mimic the
shorter 48-h glucocorticoid bursts used for refractory hypotension or prolonged postnatal treatment with other
glucocorticoids. Keeping in mind these differences, our findings suggest that a short course of DEX or HC during the
neonatal period may still have long-lasting consequences, though mild in general.
Somatic Growth

Dexamethasone and hydrocortisone in neonatal rats
12
Animals given DEX on PD5 and PD6 had a slower rate of weight and length gain when compared to HC
treated animals and control groups (VEH) resulting in a smaller animal in the DEX group for both weight and length
during the entire study period, up to pre-adolescence (PD33). As in our previous DEX study, we presume that
observed differences in somatic weight are due to a direct effect of DEX on catabolism and tissue accretion (31).. It
has been shown that DEX prevents adequate growth, rapidly inducing protein catabolism beyond the capacity for an
anabolic state and resulting in reduced growth and lean body mass(37,38, 39,40).. Alternatively, inadequate
nutritional intake during the postnatal period due to inability to attach to the mother’s nipples or poor suckling is a
possibility we cannot exclude. It has been shown that the dam spends more time providing nutrition, stimulation, and
warmth to a litter that is perceived to have poor health (41-44). Therefore, we favor the explanation that DEX at the
given doses altered the anabolic environment that is known to be present in the stress hyporesponsive period in the
rat (4,45).
The effect on the handled pups is not surprising, since ‘touch’ during the neonatal period has been shown to
be critical for normal growth and development both in animal models (46,47) and in humans( 48) Increased somatic
growth at pre-adolescence observed in those animals that were treated with HC early in life is difficult to explain. It
is possible that the potency of HC at the given doses was significantly less than that of DEX. If so, it is possible that
the known negative effects of glucocorticoids on bone growth plates was not fully achieved (49). However, this does
not explain the enhanced growth. A possible explanation is an effect of HC at the given doses given early in life on
insulin action. Glucocorticoid levels achieved by the early treatment could induce an insulin resistant state that
promotes adiposity (50). Elevated insulin can act as a growth factor because of its structural similarity to insulin-like-
growth factors(51,52). The important distinction here is that the detrimental effects of the more potent DEX
treatment given early in life is not observed in animals treated with the less potent glucocorticoid during the same
developmental period. While this may indicate that HC is a ‘better’ glucocorticoid agent in early life, there are other
concerns such as the catch-up growth observed in our HC treated animals which favored visceral fat deposition
which could increase the risk for insulin resistance and subsequent diabetes later in life by promoting obesity in
adulthood (53,54). This will probably be addressed in future studies.

Dexamethasone and hydrocortisone in neonatal rats
13
Neurodevelopment and physical maturation
We did not observe any gross neurological deficits in the animals that received glucocorticoid treatment early
in life. Primitive reflexes appeared and disappeared in a defined sequence during development, with an absence or
persistence of any reflex beyond the expected time period indicating significant brain dysfunction (55). Since DEX or
HC did not have any significant effect on these reflexes nor had only a transient effect, it appears that these
treatments do not have any long term neurological sequelae. Clinical animal data suggest a defect in myelination in
neonatal pups treated with long term DEX (55-59). We did not perform neuroanatomical studies to determination the
extent of myelination in these pups. Evidently, our functional observations do not support this effect. However, it
remains unclear at present whether permanent CNS changes have occurred that are too subtle to detect by our
assessment.
DEX-treated animals also had decreased gross brain weights. However, when corrected for body weight,
brain weights did not differ between groups or sex of the animal. Similar findings have been reported in human
clinical studies where a 30% reduction in cerebral tissue volume is observed in premature infants treated with DEX
compared with untreated age-matched controls(56). Therefore, even though DEX effect can be corrected by body
weight, the brain is smaller and it is possible that DEX affected neurogenesis, gliogenesis, or myelination within
specific vulnerable brain structures.
Stress Reactivity to a Novel Environment
When exposed to the novel environment of the place preference box at pre-adolescence, the glucocorticoid
treated animals did not display anxiety like behaviors. It is possible that the age we tested may be too early to see
behavioral effects. However, in a previous study, we found that animals treated with DEX from postnatal day 3 to 6
were less active in the light-dark environment of the place preference box on PD33(6) The place preference box is a
less threatening environment when compared to the elevated plus maze (EPM) or open field tests (OF) (33). More
rigorous testing may have uncovered behavioral disturbances not evident with the less stressful place preference
testing.

Dexamethasone and hydrocortisone in neonatal rats
14
We found that both HC and DEX treated animals had an altered adrenocortical response to novelty stress.
DEX treated animals had an adequate corticosterone response but low when compared to control and HC treated
animals. Despite this low corticosterone response, DEX animals successfully terminated their ACTH response.
Similarly, ACTH inhibition was present in VEH treated group.
In contrast, the ACTH response to novelty was significantly greater in the HC treated animals when compared to all
other groups. This indicates slow termination to baseline. Therefore, treatment with HC did not alter the adrenal
response to stress but lead to enhanced neuroendocrine response evidenced by increased ACTH levels. This suggests
the possibility of altered LHPA related brain circuitry resulting in a faulty feedback mechanism as a consequence of
this early life treatment. Previously, we have reported decreased rat glucocorticoid receptor mRNA expression in the
hippocampus but no change in mineralocorticoid activity in adult rats that were exposed to DEX from PD3 to PD6
(60). This combination of corticoid receptor expression is linked to faulty feedback and altered behavior (61).
Glucocorticoid and mineralocorticoid receptor expression has been shown to develop primarily in the postnatal
period in neonatal rat brains (4).. This developmental phase corresponds to a very critical neurodevelopmental period
in the human premature infant (29, 62). It is during this equivalent period that the brains of premature infants are
exposed to these exogenous steroids(63,64). Therefore, adverse effects on the brain remain a serious concern when
using corticosteroids in the postnatal period.
In conclusion, exposure to a short course of DEX or HC that mimics clinical practice in neonatal intensive
care units has long-lasting effects on somatic growth and neuroendocrine function in the rat. One could interpret that
the HC treatment is benign when compared to DEX exposure in the premature infant. The effects of HC, though
subtle, raise concerns about altered LHPA function and metabolic alterations that may place the infant at risk for
future metabolic disorders depending on the postnatal environment (65,66, 67,68, 69).. Premature infants treated with
HC early in life may be vulnerable to maladaptive endocrine and behavioral strategies that may not be recognizable
until later in life. Detailed clinical research is needed to clarify these possibilities that may have important
implications on learning, mood and ultimately quality of life in long-term survivors of prematurity.

Dexamethasone and hydrocortisone in neonatal rats
15
Table 1: Schematic of the Procedures and Age of the animals when these were performed
Postnatal Day PD 2 PD5
PD6
PD7 PD8 PD14 PD20 PD33
Procedures
Drug Regimen
A*
Tests
Culled
Sexed
Body
Weight
Length
DEX
0.5mg/kg
Body
Weight
Length
DEX
0.1mg/kg
Body
Weight
Length
Neuro
Brain
Weight
Body
Weight
Length
Neuro
Body
Weight
Length
Neuro
Brain Weight
Light-Dark
Preference
Stress
Response
(ACTH &
Cort)
Procedures
Drug Regimen
B *
Tests
Culled
Sexed
Body
Weight
Length
HC
5mg/kg
Body
Weight
Length
HC
1mg/kg
Body
Weight
Length
Neuro
Brain
Weight
Body
Weight
Length
Neuro
Body
Weight
Length
Neuro
Brain Weight
Light-Dark
Preference
Stress
Response
(ACTH &
Cort)
Animals receiving drug treatment were matched within litter with animals that were treated with vehicle injections.
Dexamethasone and hydrocortisone in neonatal rats
16
Table 2. Neurological Examination Rating Scale: Physical and Neurodevelopmental Assessment
Physical
Assessment
0 1 2 3 4 5
Physical
Maturation
Teeth Eruption
Posture Full
Flexion
Partial
flexion
Full
extension
Partial
extension
Normal
Eye opening Closed Occasional
opening
Occasional
opening
Fully open
Ear opening Closed Occasional
opening
Occasional
opening
Fully open
Ear folding Barely
visible
Completely
folded
Partially
folded
Fully open Fully open
Fur development No fur Fine hairs Partial fur Mostly fur Full fur
Maxillary
eruption
No teeth Initial
maxillary
Partial
maxillary
Partial
maxillary
Mostly full Maxillary
full
Mandibular
eruption
No teeth Initial
mandibular
Partial
mandibular
Partial
mandibular
Mostly full Mandibula
r full
Neurodevelopme
nt
Assessment
0 1 2 3 4 5
Reflexes
Ear twitch
response
No
response
Weak
response
Moderate
response
Strong
response
Righting reflex On side
full
flexion
Partial
flexion
Extension
predominates
Extension
predominat
es
Partial
extension
Normal
position
Grasp reflex No grasp Weak grasp Moderate Full grasp

Dexamethasone and hydrocortisone in neonatal rats
17
A score of 0 represents complete immature physical development or immature response on exam. A score of 5 represents
completely mature physical maturation or response on exam.
Sensory Complex
Gross Motor
forelimb grasp
Grasp reflex hind
limb
No grasp Weak grasp Moderate
grasp
Full grasp
Cross extensor Absent Weak Moderate Strong
Vibrissa placing No
response
Weak
response
Moderate
response
Strong
response
Forelimb placing No
response
Weak
response
Moderate
response
Strong
response
Hind limb
placing
No
response
Weak
response
Moderate
response
Strong
response
Geotaxis No
response
Pivoting
predominat
es
Turns 1800 Strong
response
Bar hold No grasp Weak grasp Moderate
grasp
Full grasp
Postural
extension
Absent Partial Moderate Present
Postural flexion Absent Partial Moderate Present
Dexamethasone and hydrocortisone in neonatal rats
18
Table 3: Mean Weight and Length increase over time by treatment group
WEIGHT
(GMS
LENGTH(MM)
MEAN SD MEAN SD
VEH PD5 11.09 0.98 66.50 2.45
PD6 12.59 1.03 70.04 2.03
PD7 14.63 1.14 73.63 2.18
PD14 32.97 2.25 101.38 2.72
PD20 47.37 3.30 120.75 3.52
PD33 118.30 10.58 161.08 7.60
HC PD5 10.97 0.98 66.88 2.80
PD6 12.11 1.09 69.58 2.32
PD7 14.03 1.17 72.88 2.36
PD14 32.97 2.40 101.96 4.96
PD20 47.93 3.31 120.63 4.31
PD33 117.11 11.96 160.17 6.94
DEX PD5 10.86 1.13 66.54 2.99
PD6 10.40 1.02 66.96 2.18
PD7 11.23 1.13 68.42 2.52
PD14 28.52 1.93 95.75 4.09
PD20 42.42 2.38 116.54 3.06
PD33 109.59 9.48 155.65 6.29
PD Postnatal day; VEH Vehicle; DEX-dexamethasone; HC Hydrocortisone

Dexamethasone and hydrocortisone in neonatal rats
19
Figure 1: Weights obtained from PD5 to PD 20 (panel A) and PD 33 (panels B and C) in the males vs
females in the three treatment groups. Weight gain in the DEX treated animals was significantly
lower than in the HC and VEH groups on PD6-PD20(P<0.05) .There were no sex differences during
this period.(Panel A). DEX treated male animals were not significantly different from VEH or HC on
PD33 (Panel B). DEX treated female animals were significantly smaller compared with VEH (P<0.05)
but not to HC (Panel C).

Dexamethasone and hydrocortisone in neonatal rats
20
P5 P6 P7 P14 P200
10
20
30
40
50
60VEHDEXHC
A
* **
*
Bo
dy
We
igh
t (g
)
VEH DEX HC0
30
60
90
120
150B Male
Bo
dy
We
igh
t (g
)
VEH DEX HC0
30
60
90
120
150
#
FemaleC
Bo
dy
We
igh
t (g
)
Figure 1: Weight measurement
separated by treatment groups from
PD5 to PD 20 (panel A) and PD 33
(panels B and C) in the males vs
females in the three treatment groups.
Weight gain in the DEX treated animals
was significantly lower than in the HC
and VEH groups on PD6-PD20(P<0.05)
.There were no sex differences during
this period.(Panel A). DEX treated male
animals were not significantly different
from VEH or HC on PD33 (Panel B).
DEX treated female animals were
significantly smaller compared with
VEH (P<0.05) but not to HC (Panel C).

Dexamethasone and hydrocortisone in neonatal rats
21
Figure 2: Length measurements in animals separated by treatment and sex. from PD5 to 33 in the
males vs females in the three treatment groups. males vs females in the three treatment groups.
DEX treated female animals were significantly shorter in length compared with HC and VEH on
PD6- PD33(P<0.0001) (Panel A). DEX treated male animals were significantly shorter than VEH
and HC on PD6-PD20 (P< 0.001) but on PD33 there was no difference in length among the three
treatment groups.(Panel B)

Dexamethasone and hydrocortisone in neonatal rats
22
P5 P6 P7 P14 P20 P330
50
100
150
200VEHDEXHC
A
* ** *
#
Female
Bo
dy
Len
gth
(mm
)
P5 P6 P7 P14 P20 P330
50
100
150
200B
* ** *
Male
Days
Bo
dy
Len
gth
(mm
)
Figure 2: Length measurements in animals separated by treatment and sex. from
PD5 to 33 in the males vs females in the three treatment groups. males vs females in
the three treatment groups. DEX treated female animals were significantly shorter in
length compared with HC and VEH on PD6- PD33(P<0.0001) (Panel A). DEX treated
male animals were significantly shorter than VEH and HC on PD6-PD20 (P< 0.001) but
on PD33 there was no difference in length among the three treatment groups.(Panel B)

Dexamethasone and hydrocortisone in neonatal rats
23
Figure 3: Comparison of neurobehavioral outcomes (Physical maturation) between treatment groups
on day PD7, PD14 and PD21. There was no significant effect of sex or treatment on any
neurodevelopment exams within treatment groups (p< 0.0001) with the exception of physical
maturation (Panel D) and reflexes (Panel E); these two domains had limited change across the times
of assessments.
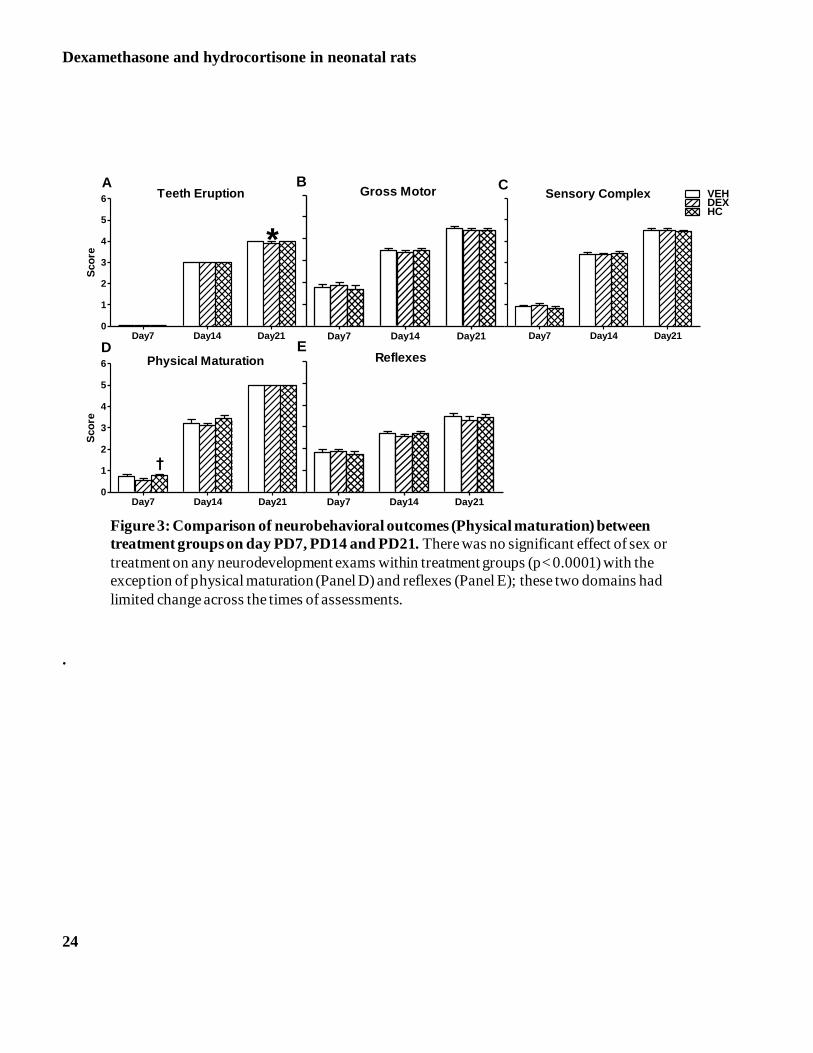
Dexamethasone and hydrocortisone in neonatal rats
24
Day7 Day14 Day210
1
2
3
4
5
6 Teeth Eruption
*
A
Sco
re
Day7 Day14 Day21
Gross MotorB
Day7 Day14 Day21
Sensory ComplexC
VEHDEXHC
Day7 Day14 Day210
1
2
3
4
5
6 Physical Maturation
†
D
Sco
re
Day7 Day14 Day21
ReflexesE
Figure 3: Comparison of neurobehavioral outcomes (Physical maturation) between
treatment groups on day PD7, PD14 and PD21. There was no significant effect of sex or
treatment on any neurodevelopment exams within treatment groups (p< 0.0001) with the
exception of physical maturation (Panel D) and reflexes (Panel E); these two domains had
limited change across the times of assessments.
.

Dexamethasone and hydrocortisone in neonatal rats
25
Figure 4. Pituitary and Adrenal Response to Novelty Stress at Post-natal Day 33. HC treated
animals had a greater mean ACTH levels (pg/mL) and AUC than VEH and DEX at 30 and 90
minutes post-stress suggesting greater overall pituitary secretion (Panels A and C). DEX treated
group had significantly lower CORT levels at 30 and 90 min than VEH and HC groups (Panels B
and D), indicating best termination of the stress response when compared to other groups. *:
p<0.05, vs VEH and HC; #: p<0.05, vs VEH; †: p < 0.05, vs DEX
Dexamethasone and hydrocortisone in neonatal rats
26
Figure 4. Pituitary and Adrenal Response to Novelty Stress at Post-natal Day 33. HC
treated animals had a greater mean ACTH levels (pg/mL) and AUC than VEH and DEX at 30
and 90 minutes post-stress suggesting greater overall pituitary secretion (Panels A and C).
DEX treated group had significantly lower CORT levels at 30 and 90 min than VEH and HC
groups (Panels B and D), indicating best termination of the stress response when compared to
other groups. *: p<0.05, vs VEH and HC;#: p<0.05, vs VEH;†: p < 0.05, vs DEX
0 15 30 900
40
80
120
160
200
240
280
320
360
400A
Time (min)
#,† #,†
AC
TH
(p
g/m
l)
0 15 30 900
40
80
120
160
200
240
280
320
360 VEHHCDEX
B
Time (min)
#
* *
#
CO
RT
(u
g/d
l)
VEH DEX HC0
5.010 3
1.010 4
1.510 4
2.010 4
2.510 4C
AUC
Treatment
#,†
AC
TH
(p
g/m
l)
VEH DEX HC0
5.010 3
1.010 4
1.510 4D AUC
Treatment
*CO
RT
(u
g/d
l)

Dexamethasone and hydrocortisone in neonatal rats
27
REFERENCES
1. Alkalay AL, Klein AH, Nagel RA, and Pomerance JJ. 1996 Evaluation of hypothalamic-pituitary-
adrenal axis in premature infants treated with dexamethasone. Am J Perinatol 13: 473-477
2. Bakker JM, Kavelaars A, Kamphuis PJ, Zijlstra J, van Bel F, and Heijnen CJ. 2001 Neonatal
dexamethasone treatment induces long-lasting changes in T-cell receptor vbeta repertoire in rats. J
Neuroimmunol 112: 47-54
3. Dachir S, Kadar T, Robinzon B, and Levy A. 1993 Cognitive deficits induced in young rats by long-
term corticosterone administration. Behav Neural Biol 60: 103-109
4. De Kloet ER, Rosenfeld P, Van Eekelen JA, Sutanto W, and Levine S. 1998 Stress, glucocorticoids
and development. Prog Brain Res 73: 101-120
5. Duncan DF. 1997 Life stress as a precursor to adolescent drug dependence. Int J Addict 12: 1047-1056
6. Flagel SB, Vazquez DM, Watson SJ, Jr., and Neal CR, Jr. 2002 Effects of tapering neonatal
dexamethasone on rat growth, neurodevelopment, and stress response. Am J Physiol Regul Integr
Comp Physiol 282: R55-63
7. Henry C, Kabbaj M, Simon H, Le Moal M, and Maccari S. 1994 Prenatal stress increases the
hypothalamo-pituitary-adrenal axis response in young and adult rats. J Neuroendocrinol 6: 341-345
8. Avery GB, Fletcher AB, Kaplan M, and Brudno DS. 1985 Controlled trial of dexamethasone in
respirator-dependent infants with bronchopulmonary dysplasia. Pediatrics 75: 106-111
9. Cummings JJ, D'Eugenio DB, and Gross SJ. 1989 A controlled trial of dexamethasone in preterm
infants at high risk for bronchopulmonary dysplasia. N Engl J Med 320: 1505-1510
10. Kazzi NJ, Brans YW, and Poland RL. 1990 Dexamethasone effects on the hospital course of infants
with bronchopulmonary dysplasia who are dependent on artificial ventilation. Pediatrics 86: 722-727
11. Kothadia JM, O'Shea TM, Roberts D, Auringer ST, Weaver RG, 3rd, and Dillard RG. 1999
Randomized placebo-controlled trial of a 42-Day tapering course of dexamethasone to reduce the
duration of ventilator dependency in very low birth weight infants. Pediatrics 104: 22-27

Dexamethasone and hydrocortisone in neonatal rats
28
12. Yeh TF, Lin YJ, Huang CC, Chen YJ, Lin CH, Lin HC, Hsieh WS, and Lien YJ. 1998 Early
dexamethasone therapy in preterm infants: a follow-up study. Pediatrics 101: E7
13. Halliday HL, Ehrenkranz RA, and Doyle LW. 2003 Delayed (>3 weeks) postnatal corticosteroids for
chronic lung disease in preterm infants. Cochrane Database Syst Rev CD001145
14. Halliday HL, Ehrenkranz RA, and Doyle LW. 2003 Early postnatal (<96 hours) corticosteroids for
preventing chronic lung disease in preterm infants. Cochrane Database Syst Rev CD001146
15. Halliday HL, Ehrenkranz RA, and Doyle LW. 2003 Moderately early (7-14 days) postnatal
corticosteroids for preventing chronic lung disease in preterm infants. Cochrane Database Syst Rev
CD001144
16. Baker CF, Schumacher RE, Barks J, Vazquez DM, and Bhatt-Mehta V. 2008 Evaluation of
Hydrocortisone in Neonates with Pressor Resistant Hypotension. J Perinatol 28: 412-419
17. Lodygensky GA, Rademaker K, Zimine S, Gex-Fabry M, Lieftink AF, Lazeyras F, Groenendaal F, de
Vries LS, and Huppi PS. 2005 Structural and functional brain development after hydrocortisone
treatment for neonatal chronic lung disease. Pediatrics 116: 1-7
18. Rademaker KJ, Rijpert M, Uiterwaal CS, Lieftink AF, van Bel F, Grobbee DE, de Vries LS, and
Groenendaal F. 2006 Neonatal hydrocortisone treatment related to 1H-MRS of the hippocampus and
short-term memory at school age in preterm born children. Pediatr Res 59: 309-313
19. van der Heide-Jalving M, Kamphuis PJ, van der Laan MJ, Bakker JM, Wiegant VM, Heijnen CJ,
Veen S, and van Bel F. 2003 Short- and long-term effects of neonatal glucocorticoid therapy: is
hydrocortisone an alternative to dexamethasone? Acta Paediatr 92: 827-835
20. Doyle LW, Davis PG, Morley CJ, McPhee A, and Carlin JB. 2007 Outcome at 2 years of age of infants
from the DART study: a multicenter, international, randomized, controlled trial of low-dose
dexamethasone. Pediatrics 119: 716-721

Dexamethasone and hydrocortisone in neonatal rats
29
21. Ohlsson A, Calvert SA, Hosking M, and Shennan AT. 1992 Randomized controlled trial of
dexamethasone treatment in very-low-birth-weight infants with ventilator-dependent chronic lung
disease. Acta Paediatr 81: 751-756
22. Vincer MJ, Allen AC 1998 Double blind randomized controlled trial of 6-day pulse of dexamethasone
for very low birth weight infants (VLBW < 1500 grams) who are ventilator dependent at 4 weeks of
age Pediatr Res 43: 201A
23. Watterberg KL, Gerdes JS, Cole CH, Aucott SW, Thilo EH, Mammel MC, Couser RJ, Garland JS,
Rozycki HJ, Leach CL, Backstrom C, and Shaffer ML. 2004 Prophylaxis of early adrenal
insufficiency to prevent bronchopulmonary dysplasia: a multicenter trial. Pediatrics 114: 1649-1657
24. Watterberg KL, Gerdes JS, Gifford KL, and Lin HM. 1999 Prophylaxis against early adrenal
insufficiency to prevent chronic lung disease in premature infants. Pediatrics 104: 1258-1263
25. Lattimore KA, Vazquez DM, Barks JD, and Bhatt-Mehta V. 2008 Developmental outcomes of very
low birth weight infants treated with hydrocortisone for refractory hypotension. J Neonat-Perinat Med
1: 225-232
26. Altman J, and McCrady B. 1972 The influence of nutrition on neural and behavioral development. IV.
Effects of infantile undernutrition on the growth of the cerebellum. Dev Psychobiol 5: 111-122
27. Fox WM. 1965 Reflex-ontogeny and behavioural development of the mouse. Anim Behav 13: 234-241
28. Wahlsten D. 1974 A developmental time scale for postnatal changes in brain and behavior of B6D2F2
mice. Brain Res 72: 251-264
29. Halliday HL, Ehrenkranz RA, and Doyle LW. 2003 Moderately early (7-14 days) postnatal
corticosteroids for preventing chronic lung disease in preterm infants. Cochrane Database Syst Rev
CD001144
30. Dobbing J. Scientific Foundations of Paediatrics London: Heinemann Medical Books, 1981, p. 744-759.
31. Morgane PJ, Mokler DJ, and Galler JR. 2002 Effects of prenatal protein
malnutrition on the hippocampal formation. Neurosci Biobehav Rev 26: 471-483

Dexamethasone and hydrocortisone in neonatal rats
30
32. Whitelaw A, and Thoresen M. 2000 Antenatal steroids and the developing brain. Arch Dis Child Fetal
Neonatal Ed 83: F154-157
33. Neal CR, Jr., Weidemann G, Kabbaj M, and Vazquez DM. 2004 Effect of neonatal dexamethasone
exposure on growth and neurological development in the adult rat. Am J Physiol Regul Integr Comp
Physiol 287: R375-385
34. Neal CR, Jr., VanderBeek BL, Vazquez DM, and Watson SJ, Jr. 2003 Dexamethasone exposure
during the neonatal period alters ORL1 mRNA expression in the hypothalamic paraventricular
nucleus and hippocampus of the adult rat. Brain Res Dev Brain Res 146: 15-24
35. Nelson KB, and Grether JK. 1999 Causes of cerebral palsy. Curr Opin Pediatr 11: 487-491
36. O'Shea TM, Kothadia JM, Klinepeter KL, Goldstein DJ, Jackson BG, Weaver RG, 3rd, and Dillard
RG. 1999 Randomized placebo-controlled trial of a 42-day tapering course of dexamethasone to
reduce the duration of ventilator dependency in very low birth weight infants: outcome of study
participants at 1-year adjusted age. Pediatrics 104: 15-21
37. Berry MA, Abrahamowicz M, and Usher RH. 1997 Factors associated with growth of extremely
premature infants during initial hospitalization. Pediatrics 100: 640-646
38. Leitch CA, Ahlrichs J, Karn C, and Denne SC. 1999 Energy expenditure and energy intake during
dexamethasone therapy for chronic lung disease. Pediatr Res 46: 109-113
39. Shrivastava A, Lyon A, and McIntosh N. 2000 The effect of dexamethasone on growth, mineral
balance and bone mineralisation in preterm infants with chronic lung disease. Eur J Pediatr 159: 380-
384
40. Lynch A. 1976 Postnatal undernutrition: an alternative method. Dev Psychobiol 9: 39-48
41. Brunelli SA, Shair HN, and Hofer MA. 1994 Hypothermic vocalizations of rat pups (Rattus
norvegicus) elicit and direct maternal search behavior. J Comp Psychol 108: 298-303
42. Lynch A. 1976 Postnatal undernutrition: an alternative method. Dev Psychobiol 9: 39-48
43. Stern JM. 1997 Offspring-induced nurturance: animal-human parallels. Dev Psychobiol 31: 19-37

Dexamethasone and hydrocortisone in neonatal rats
31
44. Wiener SG, Fitzpatrick KM, Levin R, Smotherman WP, and Levine S. 1977 Alterations in the
maternal behavior of rats rearing malnourished offspring. Dev Psychobiol 10: 243-254
45. Vazquez DM. 1998 Stress and the developing limbic-hypothalamic-pituitary-adrenal axis.
Psychoneuroendocrinology 23: 663-700
46. Johnson EO, Kamilaris TC, Calogero AE, Gold PW, and Chrousos GP. 1996 Effects of early parenting
on growth and development in a small primate. Pediatr Res 39: 999-1005
47. Schanberg SM, Ingledue VF, Lee JY, Hannun YA, and Bartolome JV. 2003 PKC alpha mediates
maternal touch regulation of growth-related gene expression in infant rats. Neuropsychopharmacology
28: 1026-1030
48. Field TM, Schanberg SM, Scafidi F, Bauer CR, Vega-Lahr N, Garcia R, Nystrom J, and Kuhn CM.
1986 Tactile/kinesthetic stimulation effects on preterm neonates. Pediatrics 77: 654-658
49. Zhang HF, Wang WP, Yang Y, Zhang YL, Pi YL, Tian XY, and Jiang XF. 2007 Dexamethasone up-
regulates the expression of glucocorticoid receptor in growth plate and inhibits the longitudinal
growth of bone: experiment with rats]. Zhonghua Yi Xue Za Zhi 87: 2575-2577
50. Grino M. 2005 Prenatal nutritional programming of central obesity and the metabolic syndrome: role
of adipose tissue glucocorticoid metabolism. Am J Physiol Regul Integr Comp Physiol 289: R1233-1235
51. Dupont J, and LeRoith D. 2001 Insulin and insulin-like growth factor I receptors: similarities and
differences in signal transduction. Hormone research 55 Suppl 2: 22-26
52. Johnson EO, Kamilaris TC, Calogero AE, Gold PW, and Chrousos GP. 1996 Effects of early parenting
on growth and development in a small primate. Pediatr Res 39: 999-1005
53. Eriksson JG, Forsen T, Tuomilehto J, Osmond C, and Barker DJ. 2003 Early adiposity rebound in
childhood and risk of Type 2 diabetes in adult life. Diabetologia 46: 190-194
54. Langley-Evans SC. 2006 Developmental programming of health and disease. Proc Nutr Soc 65: 97-105
55. Ferguson SA, and Holson RR. 1999 Neonatal dexamethasone on day 7 causes mild hyperactivity and
cerebellar stunting. Neurotoxicol Teratol 21: 71-76

Dexamethasone and hydrocortisone in neonatal rats
32
56. Ferguson SA, and Holson RR. 1999 Neonatal dexamethasone on day 7 causes mild hyperactivity and
cerebellar stunting. Neurotoxicol Teratol 21: 71-76
57. Noma K, Tamai H, Shimada S, and Funato M. 1991 Myelination in very low birth weight infants--
evaluation by MRI. No To Hattatsu 23: 336-341
58. Sechzer JA, Folstein SE, Geiger EH, Mervis RF, and Meehan SM. 1984 Development and maturation
of postural reflexes in normal kittens. Exp Neurol 86: 493-505
59. Valkama AM, Paakko EL, Vainionpaa LK, Lanning FP, Ilkko EA, and Koivisto ME. 2000 Magnetic
resonance imaging at term and neuromotor outcome in preterm infants. Acta Paediatr 89: 348-355
60. Neal CJ, Banderbeek B, Watson SJ, and Vazquez DM. 2002 Effect of a prolonged course of
dexamethasone on corticotropin releasing hormone, mineralocorticoid receptor and glucocorticoid
receptor gene expression in the adult rat. Soc Neurosci Abstr 28: 274, 279
61. Kabbaj M, Devine DP, Savage VR, and Akil H. 2000 Neurobiological correlates of individual
differences in novelty-seeking behavior in the rat: differential expression of stress-related molecules. J
Neurosci 20: 6983-6988
62. Whitelaw A, and Thoresen M. 2000 Antenatal steroids and the developing brain. Arch Dis Child Fetal
Neonatal Ed 83: F154-157
63. Hack M, Friedman H, and Fanaroff AA. 1996 Outcomes of extremely low birth weight infants.
Pediatrics 98: 931-937
64. Halliday HL, Ehrenkranz RA, and Doyle LW. 2009 Early (< 8 days) postnatal corticosteroids for
preventing chronic lung disease in preterm infants. Cochrane Database Syst Rev CD001146
65. Barker DJ, Bagby SP, and Hanson MA. 2006 Mechanisms of disease: in utero programming in the
pathogenesis of hypertension. Nature clinical practice 2: 700-707
66. Gardner DS, Bell RC, and Symonds ME. 2007 Fetal mechanisms that lead to later hypertension.
Current drug targets 8: 894-905

Dexamethasone and hydrocortisone in neonatal rats
33
67. MohanKumar SM, King A, Shin AC, Sirivelu MP, MohanKumar PS, and Fink GD. 2007
Developmental programming of cardiovascular disorders: focus on hypertension. Reviews in endocrine
& metabolic disorders 8: 115-125
68. Seckl JR. 2008 Glucocorticoids, developmental 'programming' and the risk of affective dysfunction.
Prog Brain Res 167: 17-34
69. Silveira PP, Portella AK, Goldani MZ, and Barbieri MA. 2007 Developmental origins of health and
disease (DOHaD). Jornal de pediatria 83: 494-504