taxonomy and evolution of the cotton genus, gossypium · taxonomy and evolution of the cotton...
TRANSCRIPT

1
Taxonomy and Evolution of the Cotton Genus, Gossypium
Jonathan F. Wendel* and Corrinne E. Grover
AbstractWe present an overview of the taxonomy of Gossypium L. (the cotton genus) and its evolutionary history. Gossypium contains more than 50 recognized species, including several recently described, distributed in arid to semiarid regions of the tropics and subtropics. Diversity in Gossypium has been promoted by two seemingly unlikely pro-cesses: transoceanic, long-distance dispersal and wide hybridization among lineages that presently are widely separated geographically. Included are four species that were independently domesticated for their seed fiber—two diploids from Africa–Asia and two allopolyploids from the Americas. This repeated domestication of different wild progenitors represents a remarkable case of human-driven parallel evolution. Mor-phological variation in Gossypium is extensive; growth forms in the genus range from sprawling herbaceous perennials to ?15-m-tall trees, representing a notable array of reproductive and vegetative characteristics. Equally impressive is the striking cyto-genetic and genomic diversity that emerged as Gossypium diversified and spread worldwide, ultimately spawning eight groups of closely related diploid (n = 13) species (i.e., genome groups A through G, and K). DNA sequence data place the origin of Gossy-pium at about 5 to 10 million years ago (mya), which rapidly diversified into these major genome groups shortly thereafter. Allopolyploid cottons appeared within the last 1 to 2 million years, a consequence of the improbable transoceanic dispersal of an A genome taxon to the New World and subsequent hybridization with an indigenous D genome diploid. Diversification of the nascent allopolyploid gave rise to three modern lineages containing seven species, including the agronomically important G. hirsutum L. and G. barbadense L.
Because of its economic importance, the cotton genus (Gossypium L.) has been of interest to agricultural scientists, taxonomists, and many other kinds of
biologists. Accordingly, a considerable amount is understood regarding the ori-gin and diversification of the genus, its basic plant biology, and its properties as a crop plant (Paterson, 2009; Stewart et al., 2010; Wendel et al., 2009, 2010, 2012). Increasingly over the past two decades, classic taxonomic questions such as the origin of the polyploid species, the relationships among species and species
Abbreviations: mya, million years ago.
Jonathan F. Wendel and Corrinne E. Grover ([email protected]), Dep. of Ecology, Evolution, and Organismal Biology, Bessey Hall, Iowa State University, Ames, IA 50011. *Corresponding author ([email protected]).doi:10.2134/agronmonogr57.2013.0020
Copyright © 2015. ASA, CSSA, and SSSA, 5585 Guilford Road, Madison, WI 53711, USA. Cotton. 2nd. ed. David D. Fang and Richard G. Percy, editors. Agronomy Monograph 57.
Published April 23, 2015

2 Wendel & Grover
groups, and the origins of the domesticated forms from their wild progenitors have been revisited using modern molecular technologies (Wendel et al., 2009, 2010, 2012; Wendel and Cronn, 2003). One of the salient features of the genus is that its history is so encompassing in scope, involving a global phylogenetic diversification and a repeated history of transoceanic dispersal. A remarkable consequence of this global spread and natural diversification has been that it pro-vided the opportunity for different ancient human cultures on several continents to independently domesticate four different cotton species—two allopolyploids from the Americas, G. hirsutum and G. barbadense, and two diploids from Africa–Asia, G. arboreum L. and G. herbaceum L. The canvas for this human history is the natural evolutionary diversification of the genus, one that involves multiple, seemingly impossible hybridization events, often between lineages that presently are geographically disjunct or even on different continents, as well as multiple, remarkable cases of transoceanic, long-distance dispersals. These twin themes of cryptic intergenomic introgression or hybridization and long-distance dispersal have repeatedly come into play as the genus diversified and colonized much of the arid to semiarid regions of the tropics and subtropics.
A wealth of recent studies have provided new insights into Gossypium tax-onomy and diversity, suggesting that it is an appropriate time to update our understanding of the evolution, taxonomy, and biogeographic history of the genus. Here we provide such an update, drawing on our earlier reviews (Wendel et al., 2009, 2010, 2012; Wendel and Cronn, 2003) and with new syntheses.
Emergence of the Genus GossypiumThe cotton genus belongs to the Gossypieae, a small taxonomic tribe that includes only nine genera, eight of which are classically recognized (Fryxell, 1968, 1979) and one that is a new segregate from Cienfuegosia (Phuphathanaphong, 2006). Five of these genera are small with restricted geographic distributions (Fryxell, 1968, 1979; Phuphathanaphong, 2006) including Lebronnecia (Marquesas Islands), Ceph-alohibiscus (New Guinea, Solomon Islands), Gossypioides (East Africa, Madagascar), Thepparatia (known only from the type locality in Northern Thailand) and Kokia (Hawaii). The tribe also includes four moderately sized genera with broader geo-graphic ranges: Hampea (21 neotropical species), the relatively diverse Cienfuegosia (25 species from the neotropics and parts of Africa), Thespesia (17 tropical species), and Gossypium, whose 50+ species (Fryxell, 1992) make it the largest and most widely distributed genus in the tribe. Thus, in aggregate this small taxonomic tribe has achieved an expansive geographic range, though many of the included taxa are quite rare and/or narrowly distributed.
Molecular phylogenetic analyses have shed light on three important aspects of the evolutionary history of the tribe (Cronn et al., 2002; Seelanan et al., 1997). First and foremost is that despite their extensive distribution and extraordinary diversity, the group of species that belong to Gossypium do constitute a single natural lineage. A second important revelation has been the identity of the clos-est relatives of Gossypium, that is, the African–Madagascan genus Gossypioides and the Hawaiian endemic genus Kokia. This is unsuspected finding, given their biogeography and lower chromosome number (n = 12), yet it is quite important in that they may serve as outgroups in phylogenetic analyses, which provides an essential context both for studying evolutionary patterns and processes within

Taxonomy and Evolution of the Cotton Genus, Gossypium 3
Gossypium and for understanding genome evolution. The third important insight gained from these molecular analyses was the provision of a temporal compo-nent to the major divergences. Using sequence divergence data as a proxy for time (i.e., a “molecular clock”), Seelanan et al. (1997) suggested that Gossypium diverged from Kokia and Gossypioides circa 12.5 mya, an estimate bolstered by a later, more extensive data set containing 10 different nuclear genes (Cronn et al., 2002). Collectively, all molecular data suggest that Gossypium diverged from its closest relatives approximately 10 to 15 mya during the Miocene and acquired its modern, worldwide distribution by multiple transoceanic dispersals combined with regional speciation.
Diversification of the Diploid Cotton SpeciesFrom the time of origin 5 to 10 mya, Gossypium experienced rapid speciation and diversification, while also achieving a nearly worldwide distribution containing several primary centers of diversity in the arid or seasonally arid tropics and sub-tropics (Table 1). Species-rich regions include Australia, especially the Kimberley region in NW Australia; the Horn of Africa and southern Arabian Peninsula; and the western part of central and southern Mexico. The taxonomy of the genus has been well studied (Cronn et al., 2002; Fryxell, 1979, 1992; Hutchinson et al., 1947; Saunders, 1961; Seelanan et al., 1997; Watt, 1907), with the taxonomic classification of Fryxell (1979, 1992) being the most modern and widely followed. In this treat-ment, species are grouped into four subgenera and eight sections (Table 1). The
Table 1. Diversity and geographic distribution of the major lineages of Gossypium. Genomic placements and taxonomic status of species enclosed by parentheses are yet to be validated by cytogenetic and other means.
Genome (no. of species) Recognized species Geographic distributionA (2) G. arboreum, G. herbaceum Africa, AsiaB (3–4) G. anomalum, G. triphyllum, G. capitis-viridis, (G.
trifurcatum)Africa, Cape Verde Islands
C (2) G. sturtianum, G. robinsonii AustraliaD (13–14) G. thurberi, G. armourianum, G. harknessii, G.
davidsonii, G. klotzschianum, G. aridum, G. raimondii, G. gossypioides, G. lobatum, G. trilobum, G. laxum, G. turneri, G. schwendimanii, (Gossypium sp. nov.)
primarily Mexico, with range extensions into Peru, Galapagos Islands, Arizona
E (5–9) G. stocksii Mast., G. somalense (Gurke) J.B. Hutch., G. areysianum Deflers, G. incanum (O. Schwartz) Hillc., G. trifurcatum, (G. benadirense), (G. bricchettii), (G. vollesenii Fryxell), (G. trifurcatum)
Arabian Peninsula, Northeast Africa, Southwest Asia
F (1) G. longicalyx East AfricaG (3) G. bickii, G. australe, G. nelsonii AustraliaK (12) G. anapoides, G. costulatum, G. cunninghamii, G.
enthyle Fryxell et al., G. exiguum Fryxell et al., G. londonderriense Fryxell et al., G. marchantii, G. nobile Fryxell et al., G. pilosum Fyxell, G. populifolium (Benth.) F. Muell. ex Tod., G. pulchellum (C.A. Gardner) Fryxell, G. rotundifolium Fryxell et al.
Northwest Australia, Cobourg Peninsula, Northern Territory, Australia
AD (7) G. hirsutum, G. barbadense, G. tomentosum, G. mustelinum, G. darwinii, G. ekmanianum, Gossypium sp. nov.†
New World tropics and subtropics, including Hawaii, the Wake Atoll, and the Galapagos Islands
† J.F. Wendel, C.E. Grover, J. Jareczek, and J.P. Gallagher, unpublished data, 2014.

4 Wendel & Grover
classification represents the accumulated scientific understanding that emerged both from basic plant exploration and taxonomic and evolutionary study. While these species groupings are based primarily on morphological and geographical evidence, most infrageneric alignments are congruent with subsequent cyto-genetic and molecular data sets (Endrizzi et al., 1985; Wendel and Albert, 1992; Wendel et al., 2009, 2010).
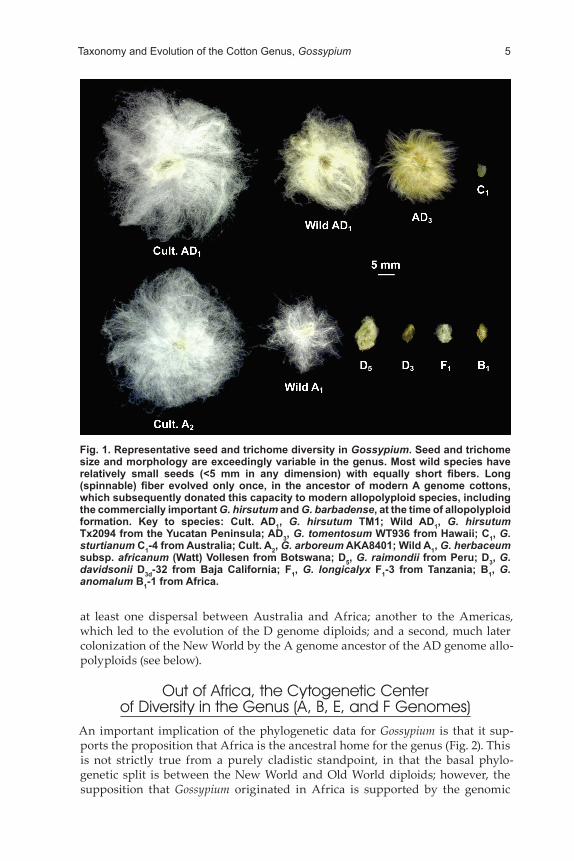
The current circumscription of Gossypium presently includes approximately 50 species (Fryxell, 1992), although new species continue to be discovered (Alva-rez and Wendel, 2006; Grover et al., 2014; Krapovickas and Seijo, 2008; Stewart et al., 2015; Ulloa et al., 2006). Morphologically, the genus is exceptionally diverse; species morphologies range from fire-adapted, herbaceous perennials with mas-sive underground rootstocks in NW Australia to trees in SW Mexico that escape the dry season by dropping their leaves. Corolla colors span a rainbow of blue to purple [G. triphyllum (Harv.) Hochr.], mauves and pinks (G. sturtianum J.H. Willis, or “Sturt’s Desert Rose,” the official floral emblem of the Northern Territory, Aus-tralia), whites and pale yellows (NW Australia, Mexico, Africa–Arabia), and even a deep sulfur-yellow (G. tomentosum Nutt. ex Seem., Hawaii). Seed coverings are equally diverse; these range from nearly glabrous to the naked eye (e.g., G. klotzs-chianum Andersson and G. davidsonii Kellogg), to short, stiff, dense, brown hairs that aid in wind dispersal (G. australe F. Muell and G. nelsonii Fryxell), to the long, fine, white fibers that characterize highly improved forms of the four cultivated species (Fig. 1). There are even seeds that produce fat bodies to facilitate ant-dis-persal (Seelanan et al., 1999).
In addition to the remarkable morphological diversity, the genus also expe-rienced extensive chromosomal evolution during diversification (Baudoin et al., 2009; Endrizzi et al., 1985). Despite sharing the same chromosome number (n = 13), genome sizes among the diploid species vary more than threefold (Hendrix and Stewart, 2005). Among closely related species, chromosome morphology is quite similar, as is demonstrated by the ability of closely related species to form hybrids that undergo normal meiotic pairing and that often experience high F1 fertility. In contrast, crosses between more distantly related species are often dif-ficult to generate, with the few successful hybrids typically experiencing meiotic abnormalities. Extensive observations concerning the pairing behavior, chro-mosome sizes, and relative fertility in interspecific hybrids ultimately led to the clustering of closely related species into “genome groups,” designated by sin-gle-letter genome symbols (Beasley, 1941). Currently, there are eight recognized diploid genome groups (A through G, plus K) (Endrizzi et al., 1985; Stewart, 1995), whose species inclusions are largely congruent with taxonomic and phylogenetic placements.
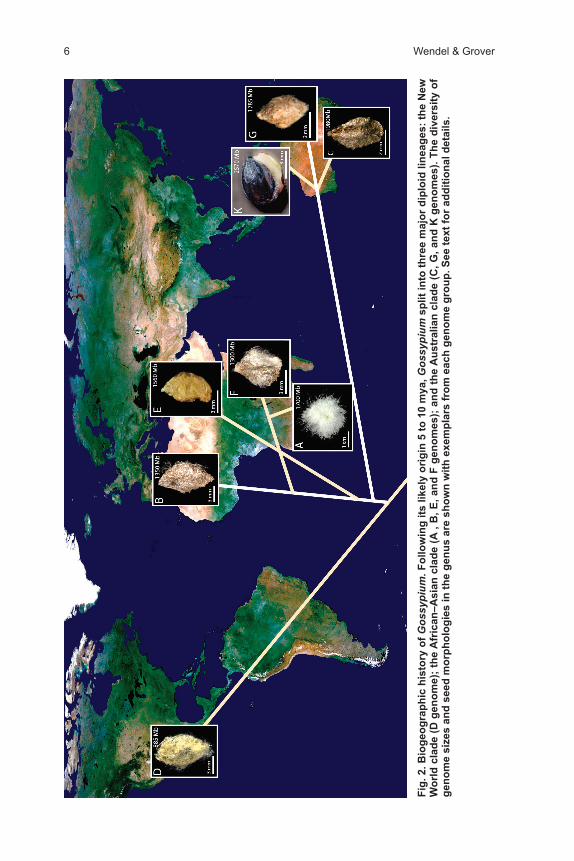
A temporal framework for the origin of Gossypium and its diversification is provided by sequence divergence data serving as a proxy for time (Senchina et al., 2003). These analyses indicate that Gossypium diverged from its closest relatives during the Miocene approximately 10 to 15 mya to originate approximately 5 to 10 mya, and these estimates of divergence times have recently been supported by an enormous amount of new sequence data derived from a global assembly of expressed sequence tags (Flagel et al., 2012). A consideration of the near world-wide distribution of species combined with their phylogenetic history (Fig. 2), relative to plate tectonic history, indicates that the evolution of Gossypium has involved multiple episodes of transoceanic dispersal. Minimally, these include
Taxonomy and Evolution of the Cotton Genus, Gossypium 5
at least one dispersal between Australia and Africa; another to the Americas, which led to the evolution of the D genome diploids; and a second, much later colonization of the New World by the A genome ancestor of the AD genome allo-polyploids (see below).
Out of Africa, the Cytogenetic Center of Diversity in the Genus (A, B, E, and F Genomes)
An important implication of the phylogenetic data for Gossypium is that it sup-ports the proposition that Africa is the ancestral home for the genus (Fig. 2). This is not strictly true from a purely cladistic standpoint, in that the basal phylo-genetic split is between the New World and Old World diploids; however, the supposition that Gossypium originated in Africa is supported by the genomic
Fig. 1. Representative seed and trichome diversity in Gossypium. Seed and trichome size and morphology are exceedingly variable in the genus. Most wild species have relatively small seeds (<5 mm in any dimension) with equally short fibers. Long (spinnable) fiber evolved only once, in the ancestor of modern A genome cottons, which subsequently donated this capacity to modern allopolyploid species, including the commercially important G. hirsutum and G. barbadense, at the time of allopolyploid formation. Key to species: Cult. AD1, G. hirsutum TM1; Wild AD1, G. hirsutum Tx2094 from the Yucatan Peninsula; AD3, G. tomentosum WT936 from Hawaii; C1, G. sturtianum C1-4 from Australia; Cult. A2, G. arboreum AKA8401; Wild A1, G. herbaceum subsp. africanum (Watt) Vollesen from Botswana; D5, G. raimondii from Peru; D3, G. davidsonii D3d-32 from Baja California; F1, G. longicalyx F1-3 from Tanzania; B1, G. anomalum B1-1 from Africa.
6 Wendel & Grover
Fig.
2. B
ioge
ogra
phic
his
tory
of G
ossy
pium
. Fol
low
ing
its li
kely
orig
in 5
to 1
0 m
ya, G
ossy
pium
spl
it in
to th
ree
maj
or d
iplo
id li
neag
es: t
he N
ew
Wor
ld c
lade
(D g
enom
e); t
he A
fric
an–A
sian
cla
de (A
, B
, E, a
nd F
gen
omes
); an
d th
e A
ustr
alia
n cl
ade
(C, G
, and
K g
enom
es).
The
dive
rsity
of
geno
me
size
s an
d se
ed m
orph
olog
ies
in th
e ge
nus
are
show
n w
ith e
xem
plar
s fr
om e
ach
geno
me
grou
p. S
ee te
xt fo
r add
ition
al d
etai
ls.

Taxonomy and Evolution of the Cotton Genus, Gossypium 7
diversity found in the region (see below), versus the relative homogeneity found in the New World. Fourteen species from Africa and Arabia are recognized in the most recent taxonomic treatment of the genus (Fryxell, 1992) and together com-pose the African–Arabian subgenus Gossypium. Recognized within the subgenus Gossypium are two taxonomic sections: section Gossypium, which contains four subsections, and another African–Arabian section (Serrata), which contains the single species G. trifurcatum Vollesen (found in desert area of eastern Somalia). The presence of dentate leaves has raised the question of whether G. trifurcatum belongs in the genus Gossypium at all, as dentate leaves are more characteristic of Cienfugosia; however, molecular work has established that this poorly known specimen is indeed a cotton species, if unusual in form (Rapp and Wendel, 2005). This example underscores the provisional nature of much of the taxonomy of the African–Arabian species of Gossypium, which are sorely in need of basic plant exploration and systematic study. Other examples include subsection Pseudopam-bak, which contains several species whose recognition and definition are based solely on limited herbarium material (e.g., G. benadirense Mattei, G. bricchettii (Ulbr.) Vollesen, G. vollesenii). As no seeds have been collected and no living spec-imens are available, there exists no information on the cytogenetic characteristics and molecular phylogenetic placement for these species. It is nearly certain that new taxonomic diversity remains to be discovered in this region of the world.
Cytogenetically, the African–Arabian species are rather diverse and account for four of the eight genome groups (A, B, E, and F), which neatly correspond to their taxonomic subsections. The A genome group contains the two cultivated cottons of subsection Gossypium, G. arboreum and G. herbaceum, while the three African species in the B genome (G. anomalum Wawra, G. capitis-viridis Mauer, and G. triphyllum) compose subsection Anomala. It bears noting that G. trifurcatum (taxonomic section Serrata) may in fact belong to the B genome, but this has not been established. The sole F genome species, G. longicalyx J.B. Hutch. & B.J.S. Lee, represents subsection Longiloba, and, of the section Gossypium, it is considered cytogenetically distinct (Phillips, 1966), morphologically isolated (Fryxell, 1971, 1992), and according to Fryxell may be better adapted to more mesic conditions than any other diploid Gossypium species. Further adding to the interest in this species is the phylogenetic placement of G. longicalyx as sister to the A genome taxa (Fig. 2), which provides a window into the evolutionary origin of “long” fiber. The remaining African–Asian species, those of subsection Pseudopambak, are con-sidered to possess E genomes, although this has yet to be verified.
To Australia, with Genomic Gigantism and New Ecological Specializations
Australia represents another center of diversity for the cotton genus. The Austra-lian cottons (subgenus Sturtia) are represented by 3 taxonomic sections containing 17 total species, including one, G. anapoides J.M. Stewart et al., nom. inval., that is newly described (Stewart et al., 2015). Cytogenetically, these taxa are divided into the C, G, and K genome groups, having 2, 3, and 12 species, respectively. DNA sequence data (Liu et al., 2001; Seelanan et al., 1997, 1999) support these groups as natural lineages, consistent with their formal alignments into the taxonomic sections Sturtia (C genome), Hibiscoidea (G genome), and Grandicalyx (K genome), although the relationships among these three are yet unresolved. Some data

8 Wendel & Grover
support a west to east radiation of Gossypium in Australia by placing G. robinsonii F. Muell. (C genome) as the basal-most branch of the Australian clade (DeJoode and Wendel, 1992). This basal placement of the C genome has not been observed in the most recent analyses (Liu et al., 2001; Seelanan et al., 1997, 1999), which provide no resolution among the Australian lineages and thereby highlight the uncertainty in the phylogenetic history of the Australian cottons.
Taxonomically, the C and G genome groups are well understood, as these are amply represented in collections and have been thoroughly studied (DeJoode and Wendel, 1992; Fryxell, 1979, 1992; Liu et al., 2001; Seelanan et al., 1997, 1999; Tiwari et al., 2014; Wendel et al., 1991). Species in the G genome are interfertile and appar-ently hybridize commonly (Tiwari et al., 2014; Wendel et al., 1991). The taxonomy of the K genome (section Grandicalyx), however, is less certain. Collecting expedi-tions to the Kimberley region have doubled the number of recognized K genome species in the past two decades through the discovery of at least seven new spe-cies (Fryxell, 1992; Stewart et al., 2015) and have expanded our understanding of the diversity in this section. The species of this section are geographically, morphologically, and ecologically distinct from other cotton species, frequently exhibiting features characteristic of fire adaptation. Specifically, these are her-baceous perennials that exhibit a biseasonal growth pattern, where vegetative growth dies back during the dry season and the plants survive as underground rootstocks until fire or the return of the wet season initiates a new cycle of growth. Also unique to section Grandicalyx are the flower and fruit morphologies. K genome species possess upright flowers (like other cotton species) that subse-quently become pendant following pollination. This change in floral positioning is thought to aid in these uniquely ant-dispersed cotton seeds. That is, at matu-rity the pendant capsules release sparsely haired seeds, which bear elaiosomes (fat bodies) that attract the ants who participate in their dispersal. Nearly all of K genome species are poorly represented in collections (Campbell et al., 2010), leading to much of this taxonomic uncertainty. These species have enormous genomes, relative to others in the genus (Fig. 2), reflecting a relatively recent pro-liferation of gypsy-like transposable elements (Hawkins et al., 2006). Phylogenetic analyses have yielded conflicting results regarding interspecific relationships in section Grandicalyx (Liu et al., 2001; Seelanan et al., 1999), although there is some support for a fundamental division of the section into a group of prostrate species and a clade of more upright plants.
One final comment about the Australian species concerns their apparent propensity to hybridize, a somewhat surprising observation given their present status as species whose populations are mostly small, scattered, and geographi-cally highly disjunct from those of related species. These geographical and often phenological barriers notwithstanding, there is abundant evidence for multiple episodes of historical introgression in the group, involving G. bickii Prokh., G. sturtianum, and G. cunninghamii Tod. (Cronn and Wendel, 2004; Seelanan et al., 1999; Tiwari et al., 2014; Wendel et al., 1991).
To the New World, Part I: The D Genome DiploidsA watershed moment in the evolutionary history of Gossypium followed a seem-ingly impossible transoceanic (either Atlantic or Pacific) dispersal of an African ancestor to the shores of Central America, possibly on what is now the western

Taxonomy and Evolution of the Cotton Genus, Gossypium 9
coast of Mexico. This dispersal event, dating to about 5 to 10 mya, led to the evo-lution and diversification of the New World D genome clade. This assemblage of species have unusually small genomes for the genus (Fig. 2), due at least in part to deletional clearance of much of the transposable element fraction (Hawkins et al., 2009), and none bear spinnable fiber (Fig. 1). Taxonomically placed into subgenus Houzingenia, the species are aligned into two sections and six subsections. These species have been more thoroughly studied than most, and consequently their taxonomy is reasonably well understood. Nonetheless, there remains sufficient cryptic variability in the group that much remains to be learned about diversity and relationships (Alvarez and Wendel, 2006; Cronn et al., 2003; Feng et al., 2011; Ulloa et al., 2006, 2013; Wendel and Cronn, 2003; Wendel et al., 1995). This is par-ticularly true for the “arborescent clade” comprising the group of small trees that includes the widespread G. aridum (Rose & Standl.) Skovst. and the more nar-rowly distributed G. laxum L. Ll. Phillips, G. lobatum Gentry, and G. schwendimanii Fryxell & S.D. Koch. Evidence indicates that cryptic variation exists within this clade, possibly including new species related to G. aridum (Alvarez et al., 2005; Feng et al., 2011) and G. laxum (Alvarez et al., 2005; Ulloa et al., 2006). There even are populations of G. aridum from Colima that have a plastid genome much more similar to those from G. klotzschianum (from the Galapagos Islands) and G. davidso-nii (from Baja California) than to all other G. aridum populations, likely reflecting historical interspecific gene flow (Alvarez and Wendel, 2006; Wendel and Albert, 1992). Gossypium gossypioides (Ulbr.) Standl. also has a somewhat mysterious intro-gressive ancestry (Cronn et al., 2003; Wendel et al., 1995), pointing out once again the significance of ancient, natural interspecific hybridization in the generation of diversity in the cotton genus (Cronn and Wendel, 2004).
The D genome cottons also have received considerable phylogenetic atten-tion (Alvarez et al., 2005; Cronn et al., 2002, 2003; Feng et al., 2011; Ulloa et al., 2006, 2013; Wendel and Albert, 1992), which provides support for the naturalness of most of the subsections. Evolutionary relationships among the apparently natu-ral subsections are less certain, however (Alvarez et al., 2005; Ulloa et al., 2013), although available evidence suggests that G. gossypioides is basal-most within the subgenus (Cronn et al., 2003; Feng et al., 2011; Yu-xiang et al., 2013). All anal-yses support the monophyly of the arborescent clade, the grouping of the Baja California species (G. harknessii Brandegee–G. armourianum Kearney–G. turneri Fryxell) as a distinct clade, and the naturalness of the G. thurberi Tod.–G. trilobum (DC.) Skovst. species pair, but the relative phylogenetic placements of these three groups with G. raimondii Ulbr. remain uncertain.
Twelve of the 14+ D genome diploid species are endemic to western Mexico, indicating this area is the center of diversity of the D genome. It seems likely, there-fore, that this lineage became established and initially diversified in this region. From this we can deduce later range extensions, probably relatively recently (dur-ing the Pleistocene), following long-distance dispersals leading to the evolution of endemics in Peru (G. raimondii) and the Galapagos Islands (G. klotzschianum). The latter species is closely related to G. davidsonii from Baja California, repre-senting a clear and unusual case of this floristic connection (Wendel and Percival, 1990). As for G. raimondii, it is interesting that the sole South American representa-tive of the New World diploid cottons is implicated as the closest ancestor of the D genome of allopolyploid cotton (more on this below); the implication, perhaps ironic, is that if allopolyploidy occurred in northern South America, as suggested

10 Wendel & Grover
(but not shown) by this observation, the possibility exists that they never would have evolved had colonization of the New World by an A genome propagule not followed closely on the heels of an earlier interhemispheric North American to South American dispersal of the ancestor of what we now recognize as G. raimon-dii. This aspect of the origin of the allopolyploids makes an improbable story all the more remarkable and yet again points to the recurring theme of the evolution-ary significance of long-distance dispersal in cotton diversification.
To the New World, Part II: Origin and Diversification of the Polyploid Cottons
A rich body of cytogenetic and experimental evidence now has demonstrated convincingly that the tetraploid cotton species, which are entirely New World in their distribution, are allopolyploids containing two coresident genomes: one from an African or Asian A genome species and the other from a species similar to the American D genome diploids (Endrizzi et al., 1985; Wendel, 1989; Wendel and Cronn, 2003; Wendel et al., 2012). The hemisphere-scale allopatry of these two diploid genome groups led to many decades of mystery surrounding the timing and parentage of the New World allopolyploids. The question of “when” has been convincingly addressed through gene sequence data that indicate allopolyploid Gossypium originated approximately 1 to 2 mya in the mid-Pleistocene, before the evolution of modern humans but relatively recently in geological terms (Flagel et al., 2012; Senchina et al., 2003; Wendel, 1989). The question of parentage has also been clarified through a wealth of DNA sequence data that indicate: (i) both extant A genome species (G. arboreum, G. herbaceum) are equally distant from the allopolyploid A genome and (ii) G. raimondii is the closest extant relative of the actual D genome donor of the allopolyploid (Endrizzi et al., 1985; Li et al., 2014; Wendel, 1989; Wendel and Cronn, 2003; Wendel et al., 2012; Yu-xiang et al., 2013). Additionally, all allopolyploids contain an A genome cytoplasm, as evidenced from analysis of both mitochondrial and plastidial genomes (Galau and Wilkins, 1989; Li et al., 2014; Small and Wendel, 1999; Wendel, 1989; Xu et al., 2012). Finally, the studies mentioned above, and additional, multilocus DNA sequence data (Grover et al., 2012), support a single origin for allopolyploid cotton.
As noted above, one truly remarkable feature of the origin of allopolyploid cottons is that their genesis likely was dependent on the following series of highly improbable events, which when considered in their totality amount to a biologi-cal marvel: (i) an ancient colonization of the New World by the ancestor of the D genome diploids, following an initial transoceanic dispersal; (ii) a more recent interhemispheric North American to South American dispersal of the ancestor of modern G. raimondii; (iii) a third long-distance dispersal of an African–Asian A genome ancestor to the New World, likely in the mid-Pleistocene; and (iv) the chance biological encounter between the immigrant A genome taxon and the endemic D genome entity, giving rise to hybridization and genome doubling, and hence the genomic reunion among genomes that had been isolated on different continents for 5 to 10 million years.
Given this now-justified scenario of a Pleistocene origin, the morphologi-cal diversification and spread of the allopolyploid cotton species must have been relatively rapid following polyploidization. Classically, five allopolyploid species were widely recognized, but a sixth species (G. ekmanianum Wittm.) was recently

Taxonomy and Evolution of the Cotton Genus, Gossypium 11
resurrected (Krapovickas and Seijo, 2008) and subsequently validated (Grover et al., 2014) by molecular sequence data. In addition, ongoing studies provide jus-tification for recognition of a seventh species that is relatively closely related to G. hirsutum, from two islands (Wake, Peale) in the Wake Atoll in the Pacific Ocean, Gossypium sp. nov. (J.F. Wendel, C.E. Grover, J. Jareczek, and J.P. Gallagher, unpublished data, 2014). Included among the classically recognized species are two island endemics, G. darwinii and G. tomentosum, that have differing popula-tion structures. Gossypium darwinii, a native of the Galapagos Islands, forms large and continuous populations in some areas (Wendel and Percy, 1990), whereas the Hawaiian endemic, G. tomentosum, tends to be more sparse, primarily occur-ring as scattered individuals and small populations on several islands. The latter species, G. tomentosum, has recently received some attention with respect to its reproductive biology and diversity (DeJoode and Wendel, 1992; Hawkins et al., 2005; Pleasants and Wendel, 2010). A third classical allopolyploid, G. mustelinum Miers ex G. Watt, is restricted to a relatively small region (Bahia state) of northeast Brazil, where it survives in apparently relictual and highly homozygous scattered populations (de Menezes et al., 2014; Wendel et al., 1994). In addition to these three truly wild species, there exist two species with cultivated forms (G. barbadense and G. hirsutum). Both species are characterized by large indigenous ranges that in the aggregate encompass a wealth of morphological forms spanning the wild-to-domesticate continuum (Brubaker et al., 1999; Brubaker and Wendel, 1993, 1994; Fryxell, 1979; Hutchinson, 1951, 1959; Hutchinson et al., 1947; Lubbers and Chee, 2009; Percy and Wendel, 1990; Stephens, 1963). Gossypium hirsutum is widely distributed in Central America and northern South America, the Caribbean, and even reaches distant islands in the Pacific (Solomon Islands, Marquesas). Natural diversity in this species is underscored by its complex nomenclatorial and taxo-nomic history (Fryxell, 1976, 1979). It is therefore unsurprising that a new species from the Dominican Republic was recently discovered both in natural settings and within the Gossypium germplasm collection (Grover et al., 2014; Krapovickas and Seijo, 2008) and that other undescribed species likely are encompassed by the diversity presently subsumed under the name G. hirsutum, as exemplified also by Gossypium sp. nov., as noted above. Gossypium hirsutum has a more northerly indigenous distribution than G. barbadense, with wild populations occurring as far north as Tampa Bay, Florida (27°38¢ N) (J.M. Stewart, personal observation). Gossypium barbadense has an indigenous range centered in the northern third of South America but with a large region of range overlap with G. hirsutum in the Caribbean (Percy, 2009).
Consideration of the ecological and geographic distribution of the allopoly-ploid species has led to the suggestion that polyploidy created the opportunity to invade an ecological niche that is distinct from that occupied by most dip-loids. Fryxell (1965, 1979) noted that the truly wild forms of the allopolyploid species typically occur in coastal habitats, in contrast to most of their diploid rela-tives. Three species (G. darwinii, Gossypium sp. nov., and G. tomentosum) are island endemics that typically reside near coastlines, and two of the other species (G. barbadense and G. hirsutum) possess wild forms that typically occupy littoral habi-tats tracing the Gulf of Mexico, northwest South America, and reaching distant Pacific Islands. Fryxell speculated that the adaptations that allowed the newly evolved allopolyploid to survive, and even thrive, in littoral habitats ultimately facilitated the establishment of new polyploid species and provided a means for

12 Wendel & Grover
rapid dispersal of the saltwater tolerant seeds. These ideas gain additional sup-port from recent ecological niche modeling studies (d’Eeckenbrugge and Lacape, 2014), which provide an elegant framework for understanding past and present ecogeographic distributions of wild polyploid cottons.
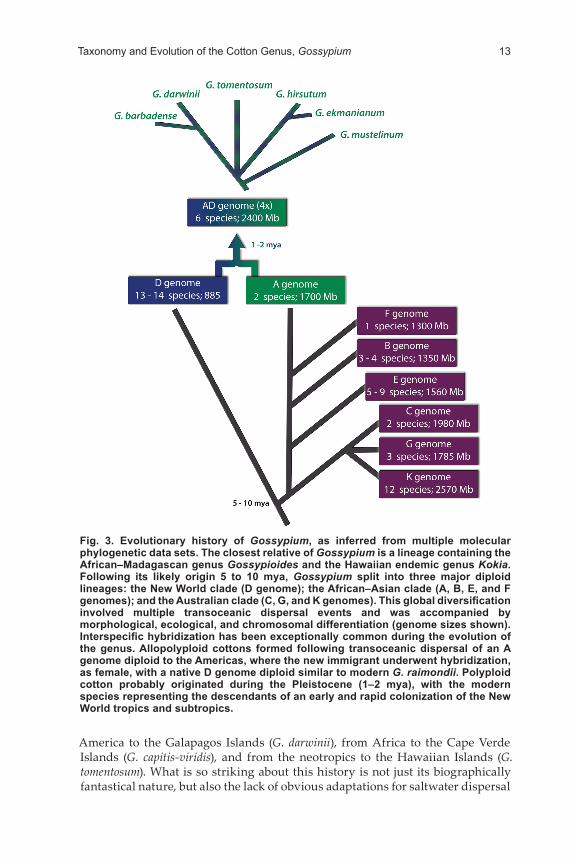
Phylogenetic Relationships in the GenusThe genealogy of Gossypium has been evaluated by multiple molecular phyloge-netic investigations (reviewed in Wendel and Cronn, 2003; Wendel et al., 2012), each of which demonstrated that the major clades of species (i.e., genealogical lineages) are largely consistent with geographical distributions, cytogenetic rela-tionships (Baudoin et al., 2009; Endrizzi et al., 1985), and genome designations. Thus, each taxonomically and cytogenetically established genome group corre-sponds to a single natural lineage, which also, in most cases, is geographically cohesive. This information is summarized in a depiction of our present under-standing of these relationships (Fig. 2 and 3).
Gossypium phylogenetic history has several broad aspects worth highlight-ing. First, four major diploid lineages exist that correspond to three continents: Australia (C, G, and K genomes), the Americas (D genome), and Africa/Arabia (itself containing two lineages: one comprising the A, B, and F genomes, and a second solely containing the E genome species). Second, the earliest divergence event separated the New World D genome lineage from that containing the ances-tor of all Old World taxa, approximately 5 to 10 mya, making the New World and Old World diploids phylogenetic sister groups. Subsequently, the Old World lin-eage itself diversified into three groups, namely, the Australian cottons (C, G, and K genome species), the African–Arabian E genome species, and the African A, B, and F genome cottons. Third, analyses identify the sole F genome species (G. longi-calyx, Africa) as sister to the A genome clade (G. arboreum and G. herbaceum), where the first emergence of fiber suitable for domestication evolved. This information diagnoses the wild forms that best represent the ancestral lintless condition, and provides a foundation for ultimately understanding the genetic basis of the ori-gin of useful, long lint (cf. Fig. 2). Fourth, the major lineages of Gossypium are characterized by rapid radiation and diversification that occurred shortly after the genus originated and diverged from the Kokia–Gossypioides clade. Fifth, and finally, the phylogenetic depiction summarizes our current understanding and uncertainties regarding relationships among the seven allopolyploid species.
From Dispersal Ecology to Human OpportunityAs emphasized in this review, a consideration of the phylogeny of Fig. 2 and 3 in a temporal context leads to the realization that the genus has had an inor-dinate fondness for transoceanic travel. As already noted, this includes at least one dispersal between Australia and Africa, another to the Americas (probably Mexico) leading to the evolution of the D genome diploids, and a second, much later, colonization of the New World by the A genome ancestor of the AD genome allopolyploids. Long-distance dispersal played a role not only in diversification of major evolutionary lines but also in speciation within Gossypium genome groups. Examples include dispersals from southern Mexico to Peru (G. raimondii), from northern Mexico to the Galapagos Islands (G. klotzschianum), from western South
Taxonomy and Evolution of the Cotton Genus, Gossypium 13
America to the Galapagos Islands (G. darwinii), from Africa to the Cape Verde Islands (G. capitis-viridis), and from the neotropics to the Hawaiian Islands (G. tomentosum). What is so striking about this history is not just its biographically fantastical nature, but also the lack of obvious adaptations for saltwater dispersal
Fig. 3. Evolutionary history of Gossypium, as inferred from multiple molecular phylogenetic data sets. The closest relative of Gossypium is a lineage containing the African–Madagascan genus Gossypioides and the Hawaiian endemic genus Kokia. Following its likely origin 5 to 10 mya, Gossypium split into three major diploid lineages: the New World clade (D genome); the African–Asian clade (A, B, E, and F genomes); and the Australian clade (C, G, and K genomes). This global diversification involved multiple transoceanic dispersal events and was accompanied by morphological, ecological, and chromosomal differentiation (genome sizes shown). Interspecific hybridization has been exceptionally common during the evolution of the genus. Allopolyploid cottons formed following transoceanic dispersal of an A genome diploid to the Americas, where the new immigrant underwent hybridization, as female, with a native D genome diploid similar to modern G. raimondii. Polyploid cotton probably originated during the Pleistocene (1–2 mya), with the modern species representing the descendants of an early and rapid colonization of the New World tropics and subtropics.

14 Wendel & Grover
(Stephens, 1958, 1966). Not only that, but the life history bauplan for Gossypium is of arid-zone adapted plants that one might assume at first blush do not possess any capacity for overseas journeys. This aspect of the natural history of the cotton genus represents a fascinating conundrum, one that lends itself more to specu-lation than to experimentation. Yet it also highlights the potential evolutionary significance of the seemingly most improbable and rare events.
Given the absence of obvious adaptations for water dispersal, what can we say about the evolutionary significance of the single-celled epidermal trichomes for which cotton is justifiably so famous? The seeds and their coverings are extraordinarily diverse in Gossypium, as illustrated in Fig. 1, which provides a visual comparison of the various lineages of wild species and domesticated cot-ton species. Even though wild and cultivated cottons both produce fiber on the seed coat, there are striking morphological and structural differences between these fibers, the most obvious of which is their size. In contrast to the canoni-cal (domesticated) cotton fiber, some D genome species (G. thurberi, G. trilobum, G. davidsonii, and G. klotzschianum) do not possess obvious seed hairs; however, the presence of these developmentally repressed structures is apparent under microscopy (Applequist et al., 2001). Similarly, while the three D genome species of subsection Caducibracteolata appear hairless to the unaided eye, these species do in fact have seed hairs that are simply tightly appressed to the seed. Culti-vated lint fiber is a model for cell wall developmental biology, as it is single cell of almost pure cellulose that experiences remarkable elongation during develop-ment in cultivated species, achieving a final length up to 6 cm (Arpat et al., 2004; Haigler et al., 2012; Kim and Triplett, 2001). In contrast, the wild-type fiber cell is composed mostly of a combination of cellulose and suberin, which elongates to <1 cm (Applequist et al., 2001; Haigler et al., 2009; Ryser and Holloway, 1985). These and many other differences reflect both natural evolutionary processes as well as human-mediated selection during domestication. The duration of the elongation phase and the timing of onset of secondary wall synthesis appear to be key determinants of the final length of the fiber in both wild and cultivated plants (Applequis et al., 2001). Phylogenetic analysis of growth rates has shown that the evolutionary innovation of prolonged elongation arose in the F genome/A genome lineage, which may have facilitated the original domestication of the A genome cottons. This trait of prolonged elongation was passed on to the allo-polyploids, which in turn was a key component of their eventual domestication. Relatively little is known about the developmental and genetic underpinnings of the diverse morphologies illustrated in Fig. 2, but an improved understanding of the changes that occurred during evolution and domestication may have impli-cations for crop improvement (Hovav et al., 2007, 2008; Hu et al., 2013; Rapp et al., 2010; Yoo and Wendel, 2014).
In nature, seed dispersal in Gossypium often follows a “shaker” model, where erect, mature capsules dehisce along the sutures and the seeds are distributed near the parental plant as wind shakes the branches. This likely is the ancestral method of seed dispersal in the genus, as it occurs in all species of the B, C, E, F, and (in part) D genome groups and in one species (G. bickii) of the G genome group. The A, AD, (some) D, G, and K genome species have evolved other mecha-nisms of seed dispersal, with perhaps the most noteworthy innovation being the fat bodies to facilitate ant dispersal on the seeds of the K genome species (Seela-nan et al., 1999). The development of spinnable fibers apparently has occurred

Taxonomy and Evolution of the Cotton Genus, Gossypium 15
only once in the history of Gossypium, in the ancestor of the two A genome species that became the progenitor of the A genome of the tetraploids.
The evolutionary “purpose” of epidermal seed hairs, or trichomes, is a mat-ter of speculation. Fryxell (1979) suggested that elongated fibers aid dispersal by birds, a hypothesis that gains credibility from sporadic mention of cotton seeds in birds’ nests as well as a collection of G. darwinii from a finch’s nest in the Gala-pagos Islands (Wendel, unpublished data, 1990). One might also speculate that fibers serve to inhibit germination unless there is sufficient moisture to saturate the fibers; should germination occur following a light rain, there might not be sufficient water for subsequent survival of the seedling. In this respect the waxy coating of the fibers would repel water, to a point, and thus prevent premature germination. A related possibility is that seed hairs function as “biological incu-bators” to facilitate germination only when ecological conditions are appropriate, by recruiting particular microbial communities under appropriate moisture regimes. Finally, two G genome species (G. australe, G. nelsonii) have evolved stiff straight seed hairs that facilitate wind dispersal in that the stiffening hairs func-tion to extrude seeds from the locules of the dehisced capsule.
It is against this natural history backdrop that ancient peoples discovered that the unique properties of cotton fibers made them useful for ropes, textiles, and other applications. This history has involved several cultures on different continents and a parallel plant improvement process starting from divergent and geographically isolated wild ancestors. Each of these crop species (G. hirsutum, G. barbadense, G. arboreum, G. herbaceum) has its own history of domes-tication, diversification, and utilization (Brubaker et al., 1999; Hutchinson, 1951, 1959; Hutchinson et al., 1947; Lubbers and Chee, 2009; Percival et al., 1999; Percy, 2009; Percy and Wendel, 1990; Tyagi et al., 2014; Ulloa et al., 2013; Wendel et al., 1992, 2009, 2010). Cotton domestication, thus, has been a replicated, and in many respects parallel experiments for the four domesticated species. Many papers describe various aspects of patterns of genetic diversity, the shape and severity of genetic bottlenecks that accompanied the development of landraces and cultivars, and the influence of recent human history on geographic patterns of cultivation (Brubaker et al., 1999; Brubaker and Wendel, 1994, 2001; Fang et al., 2013; Hutchin-son, 1951, 1954, 1959; Hutchinson et al., 1947; Percy and Wendel, 1990; Tyagi et al., 2014; Wendel et al., 1989, 1992). It is worth reiterating that this history involved human shaping and molding of naturally occurring diversity that originated through the process of evolutionary diversification over a period of millions of years, a legacy we continue to exploit today through deliberate introgression of exotic germplasm from diverse, wild gene pools (Mehetre, 2010; Stelly, 2014; Wu et al., 2010; Chapala et al., 2012; He et al., 2011a, 2011b; McCarty and Percy, 2001; Nacoulima et al., 2012; Niles and Feaster, 1984; Zhang et al., 2011, 2014). Moreover, new species of cotton and previously unrecognized taxonomic diversity continue to be discovered today (Alvarez and Wendel, 2006; Feng et al., 2011; Grover et al., 2014; Krapovickas and Seijo, 2008; Stewart et al., 2015; Ulloa et al., 2006), a remarkable realization when one considers that Gossypium ranks among our most important and hence frequently studied plant genera. The wild species of cotton, consequently, represent an ample genetic repository for potential exploitation by cotton breeders. The status of much of this diversity in the world’s germplasm collections was recently reviewed (Campbell et al., 2010). Given current threats to

16 Wendel & Grover
global ecosystems, it has never been more critical than it is today to recognize this legacy of speciation and diversification for humankind.
AcknowledgmentsResearch in the Wendel lab has largely been funded by the NSF Plant Genome Program, with additional support from other NSF programs, the USDA NRI, and Cotton Incorporated.
ReferencesAlvarez, I., R. Cronn, and J.F. Wendel. 2005. Phylogeny of the New World diploid cottons
(Gossypium L., Malvaceae) based on sequences of three low-copy nuclear genes. Plant Syst. Evol. 252:199–214. doi:10.1007/s00606-004-0294-0
Alvarez, I., and J.F. Wendel. 2006. Cryptic interspecific introgression and genetic differen-tiation within Gossypium aridum (Malvaceae) and its relatives. Evolution 60:505–517. doi:10.1111/j.0014-3820.2006.tb01132.x
Applequist, W.L., R.C. Cronn, and J.F. Wendel. 2001. Comparative development of fiber in wild and cultivated cotton. Evol. Dev. 3:3–17. doi:10.1046/j.1525-142x.2001.00079.x
Arpat, A., M. Waugh, J.P. Sullivan, M. Gonzales, D. Frisch, D. Main, et al. 2004. Functional genomics of cell elongation in developing cotton fibers. Plant Mol. Biol. 54:911–929. doi:10.1007/s11103-004-0392-y
Baudoin, J.-P., A. D’Hont, and G. Mergeai. 2009. Bridging classical and molecular cytogenet-ics of Gossypium. In: A.H. Paterson, editor, Genetics and genomics of cotton. Springer, New York. p. 257–281.
Beasley, J.O. 1941. Hybridization, cytology, and polyploidy of Gossypium. Chron. Bot. 6:394–395.
Brubaker, C.L., F.M. Bourland, and J.F. Wendel. 1999. The origin and domestication of cot-ton. In: C.W. Smith and J.T. Cothren, editors, Cotton; origin, history, technology and production. John Wiley & Sons, New York. p. 3–31.
Brubaker, C.L., and J.F. Wendel. 1993. On the specific status of Gossypium lanceolatum Tod-aro. Genet. Resour. Crop Evol. 40:165–170. doi:10.1007/BF00051121
Brubaker, C.L., and J.F. Wendel. 1994. Reevaluating the origin of domesticated cotton (Gossypium hirsutum; Malvaceae) using nuclear restriction fragment length polymor-phisms (RFLPs). Am. J. Bot. 81:1309–1326. doi:10.2307/2445407
Brubaker, C.L., and J.F. Wendel. 2001. RFLP diversity in cotton. In: J.N. Jenkins and S. Saha, editors, Genetic improvement of cotton: Emerging technologies. Science Publishers, Inc., Enfield, NH. p. 81–102.
Campbell, B., S. Saha, R. Percy, J. Frelichowski, J. Jenkins, W. Park, et al. 2010. Sta-tus of the global cotton germplasm resources. Crop Sci. 50:1161–1179. doi:10.2135/cropsci2009.09.0551
Chapala, M.M., D.B. Weaver, B.T. Campbell, E. van Santan, and R.R. Sharpe. 2012. Exotic germplasm introgression effect on agronomic and fiber properties of Upland cotton. In: Proceedings of the National Cotton Council Beltwide Cotton Conference, NCC, Orlando, FL. 3–6 Jan. 2012. NCC, Memphis, TN. p. 803–807.
Cronn, R.C., R.L. Small, T. Haselkorn, and J.F. Wendel. 2002. Rapid diversification of the cotton genus (Gossypium: Malvaceae) revealed by analysis of sixteen nuclear and chlo-roplast genes. Am. J. Bot. 89:707–725. doi:10.3732/ajb.89.4.707
Cronn, R.C., R.L. Small, T. Haselkorn, and J.F. Wendel. 2003. Cryptic repeated genomic recombination during speciation in Gossypium gossypioides. Evolution 57:2475–2489. doi:10.1111/j.0014-3820.2003.tb01493.x
Cronn, R., and J.F. Wendel. 2004. Cryptic trysts, genomic mergers, and plant speciation. New Phytol. 161:133–142. doi:10.1111/j.1469-8137.2004.00947.x
d’Eeckenbrugge, G.C., and J.M. Lacape. 2014. Distribution and differentiation of wild, feral, and cultivated populations of perennial Upland cotton (Gossypium hirsutum L.) in mesoamerica and the Caribbean. PLoS ONE 9:e107458. doi:10.1371/journal.pone.0107458

Taxonomy and Evolution of the Cotton Genus, Gossypium 17
DeJoode, D.R., and J.F. Wendel. 1992. Genetic diversity and origin of the Hawaiian Islands cotton, Gossypium tomentosum. Am. J. Bot. 79:1311–1319. doi:10.2307/2445059
de Menezes, I.P.P., F.A. Gaiotto, L.V. Hoffmann, A.Y. Ciampi, and P.A.V. Barroso. 2014. Genetic diversity and structure of natural populations of Gossypium mustelinum, a wild relative of cotton, in the basin of the De Contas River in Bahia, Brazil. Genetica (The Hague) 142:99–108.
Endrizzi, J.E., E.L. Turcotte, and R.J. Kohel. 1985. Genetics, cytology, and evolution of Gos-sypium. Adv. Genet. 23:271–375. doi:10.1016/S0065-2660(08)60515-5
Fang, D.D., L.L. Hinze, R.G. Percy, P. Li, D. Deng, and G. Thyssen. 2013. A microsatel-lite-based genome-wide analysis of genetic diversity and linkage disequilibrium in Upland cotton (Gossypium hirsutum L.) cultivars from major cotton-growing countries. Euphytica 191:391–401. doi:10.1007/s10681-013-0886-2
Feng, C., M. Ulloa, C. Perez-M, and J.M. Stewart. 2011. Distribution and molecular diversity of arborescent Gossypium species. Botany 89:615–624. doi:10.1139/b11-042
Flagel, L.E., J.F. Wendel, and J.A. Udall. 2012. Duplicate gene evolution, homoeologous recombination, and transcriptome characterization in allopolyploid cotton. BMC Genomics 13:302. doi:10.1186/1471-2164-13-302
Fryxell, P.A. 1965. Stages in the evolution of Gossypium. Adv. Frontiers Plant Sci. 10:31–56.Fryxell, P.A. 1968. A redefinition of the tribe Gossypieae. Bot. Gaz. 129:296–308.
doi:10.1086/336448Fryxell, P.A. 1971. Phenetic analysis and the phylogeny of the diploid species of Gossypium
L. (Malvaceae). Evolution 25:554–562. doi:10.2307/2407355Fryxell, P.A. 1976. A nomenclator of Gossypium: The botanical names of cotton. USDA Agri-
cultural Research Service, Washington DC.Fryxell, P.A. 1979 The natural history of the cotton tribe Texas A&M Univ. Press, College
Station, TX.Fryxell, P.A. 1992. A revised taxonomic interpretation of Gossypium L. (Malvaceae). Rheedea
2:108–165.Galau, G.A., and T.A. Wilkins. 1989. Alloplasmic male sterility in AD allotetraploid Gossy-
pium hirsutum upon replacement of its resident A cytoplasm with that of D species G. harknessii. Theor. Appl. Genet. 78:23–30. doi:10.1007/BF00299748
Grover, C.E., K.K. Grupp, R.J. Wanzek, and J.F. Wendel. 2012. Assessing the mono-phyly of polyploid Gossypium species. Plant Syst. Evol. 298:1177–1183. doi:10.1007/s00606-012-0615-7
Grover, C.E., X. Zhu, K. Grupp, J. Jareczek, J.P. Gallagher, E. Szadkowski, et al. 2014. Molec-ular confirmation of species status for the allopolyploid cotton species, Gossypium ekmanianum. Genet. Resour. Crop Evol. doi:10.1007/s10722-014-0138-x (in press)
Haigler, C.H., L. Betancur, M.R. Stiff, and J.R. Tuttle. 2012. Cotton fiber: A powerful single-cell model for cell wall and cellulose research. Front. Plant Sci. 3:104.
Haigler, C.H., B. Singh, G. Wang, and D. Zhang. 2009. Genomics of cotton fiber second-ary wall deposition and cellulose biogenesis. In: A.H. Paterson, editor, Genetics and genomics of cotton. Springer, New York. p. 385–417.
Hawkins, J.S., H. Kim, J.D. Nason, R.A. Wing, and J.F. Wendel. 2006. Differential lin-eage-specific amplification of transposable elements is responsible for genome size variation in Gossypium. Genome Res. 16:1252–1261. doi:10.1101/gr.5282906
Hawkins, J.S., J. Pleasants, and J.F. Wendel. 2005. Identification of AFLP markers that dis-criminate between cultivated cotton and the Hawaiian island endemic, Gossypium tomentosum. Genet. Resour. Crop Evol. 52:1069–1078. doi:10.1007/s10722-004-6115-z
Hawkins, J.S., S.R. Proulx, R.A. Rapp, and J.F. Wendel. 2009. Rapid DNA loss as a counter-balance to genome expansion through retrotransposon proliferation in plants. Proc. Natl. Acad. Sci. USA 106:17811–17816. doi:10.1073/pnas.0904339106
He, S., J. Sun, and X. Du. 2011. Genetic and epigenetic status of triple exotic consanguinity cotton introgression lines. Genet. Mol. Res. 10:4063–4072. doi:10.4238/2011.October.3.1

18 Wendel & Grover
He, S.-P., J.-L. Sun, C. Zhang, and X.-M. Du. 2011. Identification of exotic genetic compo-nents and DNA methylation pattern analysis of three cotton introgression lines from Gossypium bickii. Mol. Biol. 45:204–210. doi:10.1134/S002689331102018X
Hendrix, B., and J.M. Stewart. 2005. Estimation of the nuclear DNA content of Gossypium species. Ann. Bot. (Lond.) 95:789–797. doi:10.1093/aob/mci078
Hovav, R., J.A. Udall, B. Chaudhary, E. Hovav, L. Flagel, G. Hu, et al. 2008. The evolution of spinnable cotton fiber entailed prolonged development and a novel metabolism. PLoS Genet. 4:E25. doi:10.1371/journal.pgen.0040025
Hovav, R., J. Udall, E. Hovav, R. Rapp, L. Flagel, and J. Wendel. 2007. A majority of cot-ton genes are expressed in single-celled fiber. Planta 227:319–329. doi:10.1007/s00425-007-0619-7
Hu, G., J. Koh, M.J. Yoo, K. Grupp, S. Chen, and J.F. Wendel. 2013. Proteomic profiling of developing cotton fibers from wild and domesticated Gossypium barbadense. New Phy-tol. 200:570–582. doi:10.1111/nph.12381
Hutchinson, J.B. 1951. Intra-specific differentiation in Gossypium hirsutum. Heredity 5:161–193. doi:10.1038/hdy.1951.19
Hutchinson, J.B. 1954. New evidence on the origin of the Old World cottons. Heredity 8:225–241. doi:10.1038/hdy.1954.20
Hutchinson, J.B. 1959. The application of genetics to cotton improvement. Cambridge Univ. Press, Cambridge, UK.
Hutchinson, J.B., R.A. Silow, and S.G. Stephens. 1947. The evolution of Gossypium and the differentiation of the cultivated cottons.Oxford Univ. Press, London.
Kim, H.J., and B.A. Triplett. 2001. Cotton fiber growth in planta and in vitro. Models for plant cell elongation and cell wall biogenesis. Plant Physiol. 127:1361–1366. doi:10.1104/pp.010724
Krapovickas, A., and G. Seijo. 2008. Gossypium ekmanianum (Malvaceae), algodon silvestre de la Republica Dominicana. Bonplandia 17:55–63.
Li, P., Z. Li, H. Liu, and J. Hua. 2014. Cytoplasmic diversity of the cotton genus as revealed by chloroplast microsatellite markers. Genet. Resour. Crop Evol. 61:107–119. doi:10.1007/s10722-013-0018-9
Liu, Q., C.L. Brubaker, A.G. Green, D.R. Marshall, P.J. Sharp, and S.P. Singh. 2001. Evolution of the FAD2-1 fatty acid desaturase 5’ UTR intron and the molecular systematics of Gossypium (Malvaceae). Am. J. Bot. 88:92–102. doi:10.2307/2657130
Lubbers, E.L., and P.W. Chee. 2009. The worldwide gene pool of G. hirsutum and its improve-ment. In: A.H. Paterson, editor, Genetics and genomics of cotton. Springer, New York. p. 23–52.
McCarty, J.C., and R.G. Percy. 2001. Genes from exotic germplasm and their use in cultivar improvement in Gossypium hirsutum L. and G. barbadense L. In: J.N. Jenkins and S. Saha, editors, Genetic improvement of cotton: Emerging technologies. Science Publishers, Inc., Enfield, NH. p. 65–80.
Mehetre, S.S. 2010. Wild Gossypium anomalum: A unique source of fibre fineness and strength. Curr. Sci. 99:58–71.
Nacoulima, N.L., J.-P. Baudoin, and G. Mergeai. 2012. Introgression of improved fiber fine-ness trait in G. hirsutum L. from G. longicalyx Hutch. & Lee. Commun. Agric. Appl. Biol. Sci. 77(1):207–211
Niles, G.A., and C.V. Feaster. 1984. Breeding. In: R.J. Kohel and C.E. Lewis, editors, Cotton. ASA, Madison, WI. p. 201–231.
Paterson, A.H., editor. 2009. Genetics and genomics of cotton. Springer, New York.Percival, A.E., J.M. Stewart, and J.F. Wendel. 1999. Taxonomy and germplasm resources. In:
C.W. Smith and J.T. Cothren, editors, Cotton; origin, history, technology and produc-tion. John Wiley and Sons, New York. p. 33–63.
Percy, R.G. 2009. The worldwide gene pool of Gossypium barbadense L. and its improvement. In: A.H. Paterson, editor, Genetics and genomics of cotton. Springer, New York. p. 53–68.
Percy, R.G., and J.F. Wendel. 1990. Allozyme evidence for the origin and diversification of Gossypium barbadense L. Theor. Appl. Genet. 79:529–542. doi:10.1007/BF00226164

Taxonomy and Evolution of the Cotton Genus, Gossypium 19
Phillips, L.L. 1966. The cytology and phylogenetics of the diploid species of Gossypium. Am. J. Bot. 53:328–335. doi:10.2307/2439872
Phuphathanaphong, L. 2006. Thepparatia (Malvaceae), a new genus from Thailand. Thai For. Bull. (Bot.) 34:195–200.
Pleasants, J.M., and J.F. Wendel. 2010. Reproductive and pollination biology of the endemic Hawaiian cotton, Gossypium tomentosum (Malvaceae). Pac. Sci. 64:45–55. doi:10.2984/64.1.045
Rapp, R., C. Haigler, L. Flagel, R. Hovav, J. Udall, and J. Wendel. 2010. Gene expression in developing fibres of Upland cotton (Gossypium hirsutum L.) was massively altered by domestication. BMC Biol. 8:139. doi:10.1186/1741-7007-8-139
Rapp, R.A., and J.F. Wendel. 2005. Epigenetics and plant evolution. New Phytol. 168:81–91. doi:10.1111/j.1469-8137.2005.01491.x
Ryser, U., and P. Holloway. 1985. Ultrastructure and chemistry of soluble and polymeric lipids in cell walls from seed coats and fibres of Gossypium species. Planta 163:151–163. doi:10.1007/BF00393501
Saunders, J.H. 1961. The wild species of Gossypium and their evolutionary history.Oxford Univ. Press, London.
Seelanan, T., C.L. Brubaker, J.M. Stewart, L.A. Craven, and J.F. Wendel. 1999. Molecular systematics of Australian Gossypium section Grandicalyx (Malvaceae). Syst. Bot. 24:183–208. doi:10.2307/2419548
Seelanan, T., A. Schnabel, and J.F. Wendel. 1997. Congruence and consensus in the cotton tribe (Malvaceae). Syst. Bot. 22:259–290. doi:10.2307/2419457
Senchina, D.S., I. Alvarez, R.C. Cronn, B. Liu, J.K. Rong, R.D. Noyes, et al. 2003. Rate varia-tion among nuclear genes and the age of polyploidy in Gossypium. Mol. Biol. Evol. 20:633–643. doi:10.1093/molbev/msg065
Small, R.L., and J.F. Wendel. 1999. The mitochondrial genome of allotetraploid cotton (Gos-sypium L.). J. Hered. 90:251–253. doi:10.1093/jhered/90.1.251
Stelly, D. 2014. Facilitating the use of wild species germplasm for cotton improvement via CS lines. In: Proceedings of the National Cotton Council Beltwide Cotton Conference, NCC, New Orleans, LA. 6–8 Jan. 2014. NCC, Memphis, TN.
Stephens, S.G. 1958. Salt water tolerance of seeds of Gossypium species as a possible factor in seed dispersal. Am. Nat. 92:83–92. doi:10.1086/282014
Stephens, S.G. 1963. Polynesian cottons. Ann. Mo. Bot. Gard. 50:1–22. doi:10.2307/2394914Stephens, S.G. 1966. The potential for long range oceanic dispersal of cotton seeds. Am. Nat.
100:199–210. doi:10.1086/282413Stewart, J.M. 1995. Potential for crop improvement with exotic germplasm and genetic
engineering. In: G.A. Constable and N.W. Forrester, editors, Challenging the future: Proceedings of the World Cotton Research Conference, Brisbane Australia. 14–17 Feb. 1994. CSIRO, Melbourne, Australia. p. 313–327.
Stewart, J.M., L.A. Craven, C.L. Brubaker, and J.F. Wendel. 2015. Gossypium anapoi-des (Malvaceae), a new species from Western Australia. Novon 23(4):447–451. doi:10.3417/2007140
Stewart, J.M., D. Oosterhuis, J.J. Heithholt, and J.R. Mauney. 2010. Physiology of cotton. Springer, Dordrecht, The Netherlands.
Tiwari, R.S., J. Zhang, and J.M. Stewart. 2014. Molecular diversity based on AFLP markers and possible natural hybridization among the Australian arid zone Gossypium species. Aust. J. Crop Sci. 8:674–679.
Tyagi, P., M.A. Gore, D.T. Bowman, B.T. Campbell, J.A. Udall, and V. Kuraparthy. 2014. Genetic diversity and population structure in the US Upland cotton (Gossypium hirsu-tum L.). Theor. Appl. Genet. 127:283–295. doi:10.1007/s00122-013-2217-3
Ulloa, M., I.Y. Abdurakhmonov, C. Perez-M, R. Percy, and J.M. Stewart. 2013. Genetic diver-sity and population structure of cotton (Gossypium spp.) of the New World assessed by SSR markers. Botany 91:251–259. doi:10.1139/cjb-2012-0192
Ulloa, M., J.M. Stewart, E.A. Garcia-C, S. Godoy-A, A. Gaytan-M, and N.S. Acosta. 2006. Cotton genetic resources in the western states of Mexico: In situ conservation status

20 Wendel & Grover
and germplasm collection for ex situ preservation. Genet. Resour. Crop Evol. 53:653–668. doi:10.1007/s10722-004-2988-0
Watt, G. 1907. The wild and cultivated cotton plants of the world. Longmans, Green and Co., London.
Wendel, J.F. 1989. New World tetraploid cottons contain Old World cytoplasm. Proc. Natl. Acad. Sci. USA 86:4132–4136. doi:10.1073/pnas.86.11.4132
Wendel, J.F., and V.A. Albert. 1992. Phylogenetics of the cotton genus (Gossypium L.): Charac-ter-state weighted parsimony analysis of chloroplast DNA restriction site data and its systematic and biogeographic implications. Syst. Bot. 17:115–143. doi:10.2307/2419069
Wendel, J.F., C.L. Brubaker, J.P. Alvarez, R.C. Cronn, and J.M. Stewart. 2009. Evolution and natural history of the cotton genus. In: A.H. Paterson, editor, Genetics and genomics of cotton. Springer, New York. p. 3–22.
Wendel, J.F., C.L. Brubaker, and A.E. Percival. 1992. Genetic diversity in Gossypium hirsutum and the origin of Upland cotton. Am. J. Bot. 79:1291–1310. doi:10.2307/2445058
Wendel, J.F., C.L. Brubaker, and T. Seelanan. 2010. The origin and evolution of Gossypium. In: J.M. Stewart, D. Oosterhuis, J.J. Heithholt, and J.R. Mauney, editors, Physiology of cotton. Springer, Dordrecht, The Netherlands. p. 1–18.
Wendel, J.F., and R.C. Cronn. 2003. Polyploidy and the evolutionary history of cotton. Adv. Agron. 78:139–186. doi:10.1016/S0065-2113(02)78004-8
Wendel, J.F., L.E. Flagel, and K.L. Adams. 2012. Jeans, genes, and genomes: Cotton as a model for studying polyploidy. In: P.S. Soltis and D.E. Soltis, editors, Polyploidy and genome evolution. Springer-Verlag, Heidelberg. p. 181–207.
Wendel, J.F., P.D. Olson, and J.M. Stewart. 1989. Genetic diversity, introgression and inde-pendent domestication of Old World cultivated cottons. Am. J. Bot. 76:1795–1806. doi:10.2307/2444478
Wendel, J.F., and A.E. Percival. 1990. Molecular divergence in the Galapagos Island–Baja California species pair, Gossypium klotzschianum and G. davidsonii (Malvaceae). Plant Syst. Evol. 171:99–115. doi:10.1007/BF00940598
Wendel, J.F., and R.G. Percy. 1990. Allozyme diversity and introgression in the Galapagos Islands endemic Gossypium darwinii and its relationship to continental G. barbadense . Biochem. Syst. Ecol. 18:517–528. doi:10.1016/0305-1978(90)90123-W
Wendel, J.F., R. Rowley, and J.M. Stewart. 1994. Genetic diversity in and phylogenetic rela-tionships of the Brazilian endemic cotton, Gossypium mustelinum (Malvaceae). Plant Syst. Evol. 192:49–59. doi:10.1007/BF00985907
Wendel, J.F., A. Schnabel, and T. Seelanan. 1995. An unusual ribosomal DNA sequence from Gossypium gossypioides reveals ancient, cryptic, intergenomic introgression. Mol. Phylogenet. Evol.. 4:298–313. doi:10.1006/mpev.1995.1027
Wendel, J.F., J.M. Stewart, and J.H. Rettig. 1991. Molecular evidence for homoploid reticulate evolution in Australian species of Gossypium. Evolution 45:694–711. doi:10.2307/2409921
Wu, J., J. McCarty, J. Jenkins, and W. Meredith. 2010. Breeding potential of introgressions into Upland cotton: Genetic effects and heterosis. Plant Breed. 129:526–532.
Xu, Q., G. Xiong, P. Li, F. He, Y. Huang, K. Wang, et al. 2012. Analysis of complete nucleotide sequences of 12 Gossypium chloroplast genomes: Origin and evolution of allotetra-ploids. PLoS ONE 7:E37128. doi:10.1371/journal.pone.0037128
Yoo, M.-J., and J.F. Wendel. 2014. Comparative evolutionary and developmental dynam-ics of the cotton (Gossypium hirsutum) fiber transcriptome. PLoS Genet. 10:E1004073. doi:10.1371/journal.pgen.1004073
Yu-xiang, W., C. Jin-hong, H. Qiu-ling, and Z. Shui-jin. 2013. Parental origin and genomic evolution of tetraploid Gossypium species by molecular marker and GISH analyses. Caryologia 66:368–374. doi:10.1080/00087114.2013.857830
Zhang, J., R.G. Percy, and J.C. McCarty, Jr. 2014. Introgression genetics and breeding between Upland and Pima cotton: A review. Euphytica 198:1–12. doi:10.1007/s10681-014-1094-4
Zhang, Z., J. Rong, V.N. Waghmare, P.W. Chee, O.L. May, R.J. Wright, et al. 2011. QTL alleles for improved fiber quality from a wild Hawaiian cotton, Gossypium tomentosum. Theor. Appl. Genet. 123:1075–1088. doi:10.1007/s00122-011-1649-x