telomere targeting with a new g4 ligand enhances...
TRANSCRIPT

Therapeutic Discovery
Telomere Targeting with a New G4 Ligand EnhancesRadiation-Induced Killing of Human Glioblastoma Cells
Patrick Merle3,4, Bertrand Evrard1,3,5, Anne Petitjean8,9, Jean-Marie Lehn8,Marie-Paule Teulade-Fichou10, Emmanuel Chautard3,7, Anne De Cian11, Lionel Guittat11,Phong Lan Thao Tran12, Jean-Louis Mergny11,12, Pierre Verrelle3,7, and Andreï Tchirkov2,3,6,7
AbstractThe aim of this study was to test in vitro the efficacy of TAC, an original G-quadruplex ligand,
as a potential radiosensitizing agent for glioblastoma multiforme (GBM). Two human radioresistant
telomerase-positive GBM cell lines (SF763 and SF767) were analyzed, with and without TAC treatment,
for telomere length, cell proliferation, apoptosis, cell-cycle distribution, gene expression, cytogenetic
aberrations, clonogenic survival assay, 53BP1 immunofluorescence staining, and gH2AX phosphorylation.
We found that low concentrations of TAC (0.5 and 1 mmol/L) inhibited the proliferation of GBM cells in
a concentration-dependent manner after only 1 week of treatment, with minimal effects on cell cycle
and apoptosis. TAC treatment had no visible effect on average telomere length but modified expression
levels of telomere-related genes (hTERT, TRF1, and TRF2) and induced concentration-dependent
DNA damage response and dicentric chromosomes. Survival curves analysis showed that exposure to
nontoxic, subapoptotic concentrations of TAC enhanced radiation-induced killing of GBM cells. Analysis
of DNA repair after irradiation revealed delayed repair kinetics in GBM cells treated with TAC.
Furthermore, the combined treatment (TAC and radiation) significantly increased the frequency of
chromosomal aberrations as compared with radiation alone. These findings provide the first evidence
that exposure to a G4 ligand radiosensitizes human glioblastoma cells and suggest the prospect of future
therapeutic applications. Mol Cancer Ther; 10(10); 1784–95. �2011 AACR.
Introduction
Glioblastoma multiforme (GBM) is the most commonhuman primary brain tumor with a very poor prognosis.GBM patients are treated by neurosurgery followed byradiotherapy with concomitant administration of temo-zolomide (1). This treatment is not curative, and the tumor
almost always reappears inside the irradiated tumorvolume. Increasing the sensitivity of tumor cells to radio-therapy would improve outcome in patients with GBM.
Telomeres are nucleoprotein structures at the endof chromosomes that stabilize and protect them fromnucleotide degradation and end-to-end fusion. Telome-rase is a ribonucleoprotein complex, with reverse tran-scriptase activity maintaining telomere length by addinghexameric GGTTAG repeats to the chromosomal ends(2). Telomerase activity is regulated at the expressionlevel of the human telomerase reverse transcriptase(hTERT) gene. In humans, telomerase is expressed ingerminal and stem cells but is repressed in most somaticcells. In contrast, telomerase is active in more than 85% ofcancers and allows unlimited proliferation of tumor cells(3). hTERT expression is strongly associated with GBM(4), and patients with high hTERT levels have very shortsurvival (5). Suppression of hTERT in GBM cells inducessenescence and inhibits proliferation (6). The telomeraseantagonist, imetelstat, can inhibit GBM growth, showingan increased efficacy in combination with radiation andtemozolomide (7). In addition, telomeric dysfunction dueto genetic deficiency of telomerase activity increasesradiosensitivity and decreases the capacity of DNA repair(8). Thus, there is a pathogenetic link between telomere,telomerase, and radiosensitivity.
Authors' Affiliations: 1Laboratoire d’Immunologie, 2Histologie Embryo-logie Cytog�en�etique, and 3EA 3846, Clermont Universit�e, Universit�ed’Auvergne; 4Service de Pneumologie, 5Laboratoire d’Immunologie,and 6Service de Cytog�en�etique M�edicale, CHU Clermont-Ferrand;7D�epartement de Radioth�erapie, Centre Jean Perrin, Clermont-Ferrand;8ISIS, Universit�e de Strasbourg, Strasbourg, France; 9Department ofChemistry, Queen's University, Kingston, Ontario, Canada; 10Institut Cu-rie, UMR 176, Centre Universitaire, Orsay; 11INSERM U565, Mus�eumNational d’Histoire Naturelle, CNRS UMR 7196, Paris; and 12INSERMU869, Institut Europ�een de Chimie Biologie, Universit�e de Bordeaux,Pessac, France
Note: Supplementary material for this article is available at MolecularCancer Therapeutics Online (http://mct.aacrjournals.org/).
P. Merle and B. Evrard contributed equally to this work.
Corresponding Author: Andreï Tchirkov, D�epartement de Radioth�erapie,Centre Jean Perrin, 58 rue Montalembert, BP 392, 63011 Clermont-Ferrand, Cedex 1, France. Phone: 33-4-73-27-81-42; Fax: 33-4-73-27-81-25; E-mail: [email protected]
doi: 10.1158/1535-7163.MCT-10-0664
�2011 American Association for Cancer Research.
MolecularCancer
Therapeutics
Mol Cancer Ther; 10(10) October 20111784
on July 30, 2018. © 2011 American Association for Cancer Research. mct.aacrjournals.org Downloaded from

Targeting telomeres and/or telomerase usingG-quadruplex (G4) ligands is a promising strategy forcancer treatment (9). G4 ligands bind to a family of DNAsecondary structures called G-quadruplexes, whichresult from the cation-stabilized stacking of severalG-quartets (typically 2–5), each quartet being composedof 4 coplanar guanines. Quadruplexes can be verystable under physiologic conditions. Any genomicsequence containing a repetitive motif with severalconsecutive guanines can in principle form a quadru-plex (10). This is the case for the telomeric TTAGGGmotif, which is single stranded at chromosomal ends.Thus, G4 ligands interact with telomeres (11), inducetelomere uncapping, and, indirectly, affect telomerasefunction by preventing telomerase binding on its target(12). Some of these ligands induce proliferation arrestand apoptosis in GBM cell lines (13).Here, we assessed the ability of an original G4 ligand
to inhibit tumor cell proliferation and to enhance thesensitivity to ionizing radiation of 2 human telomerase-positive GBM cell lines.
Materials and Methods
Cell linesHuman GBM cell lines, SF763 and SF767, were
obtained from the Brain Tumor Research CenterTissue Bank of the University of California. Cell lineswere Mycoplasma free (Mycoplasma Detection Kit;InvivoGen). No authentication of cell lines was doneby the authors. Normal skin fibroblasts were obtainedfrom a healthy 30-year-old female donor. Cultures werecarried out in Dulbecco’s Modified Eagle’s Media sup-plemented with 10% of FBS, 1% of nonessential aminoacids, and 1% of pyruvate of sodium and gentamicin(Gibco Invitrogen).
Chemical compoundsTAC consists of N-methylated triflate derivatives
of 4,6-bis-(6-(acrid-9-yl)-pyridin-2-yl)-pyrimidine (U.S.patent 20080119492; see Supplementary Methods forsynthesis details). Mass spectrometric analyses (liquidchromatography/mass spectrometry and electrospray-mass spectrometry; Supplementary Fig. S1–3) indicatedthat TAC is a 2:1 mixture of bis-methylated and tris-methylated products. Because of the small amount ofmaterial available, the 2 compounds were not separatedand the 2:1 mixture was used as such for biologicalexperiments. TAC was dissolved in dimethyl sulfoxideat a concentration of 2 mmol/L to produce aliquots ofstock solution. The aliquots were stored at �20�C anddiluted in culture medium immediately before use toobtain a working solution at 100 mmol/L.
Equilibrium dialysis and fluorescence resonanceenergy transfer melting assaysCompetitive dialysis is an effective method to compare
the binding of a compound to different nucleic acid
structures. Experiments were carried out as previouslydescribed on a panel of single strands, duplexes, i-motif,and quadruplexes (14). Binding was quantified by theabsorbance and fluorescence signals of TAC.
Fluorescence resonance energy transfer (FRET)meltingassays allow a semiquantitative assessment of quadru-plex stabilization. The F21T oligonucleotide was doublelabeled with fluorescein or carboxytetramethylrhoda-mine (TAMRA), and apparent melting temperatureswere determined in sodium and potassium conditions,with or without added ligand, at various concentrations(micromolar range) as previously described (15).
Proliferation assessmentCells were grown for 14 days with an intermediate
passage after 7 days of culture. After harvesting, trypanblue–negative viable cells were counted. The number ofpopulation doublings (PD) after 7 and 14 days wascalculated as follows: PD ¼ log2(Nf/N0), where Nf isthe final cell number and N0 is the initial number ofseeded cells.
Cell-cycle analysisCell membranes were lyzed by immersion in liquid
nitrogen. The cell pellet was then resuspended in 500 mLof ribonuclease A (Sigma) and 500 mL of propidiumiodide (PI; Sigma) for 20 minutes at room temperature.Cell-cycle distribution and sub-G0–G1 fraction weredetermined by flow cytometry (EPICS XL; BeckmanCoulter) using CellQuest software.
Apoptosis assessmentThe Annexin V kit of Caltag Laboratories, Inc., was
used to assess apoptosis. Cells (1 � 105) were doublelabeled with PI and anti-human Annexin V antibody andthen analyzed by flow cytometry (EPICS XL). Earlyapoptotic cells were scored as the percentage of AnnexinV–positive/PI-negative cells, and late apoptotic andnecrotic cells were scored as the percentage of cellsstaining positive for both Annexin V and PI.
Quantitative real-time reverse transcriptase PCRTotal RNA was extracted from tumor cells using
TRIzol reagent (Invitrogen). One microgram of RNAwas reverse transcribed with SuperScript II enzyme(Invitrogen). Quantitative real-time reverse transcrip-tase PCR (qRT-PCR) was carried out with the Light-Cycler system (Roche Diagnostics). The primers usedfor amplification of hTERT, TRF1, and TRF2 (telomericrepeat binding factors 1 and 2) have been describedelsewhere (16, 17). In addition, the amount of ABLtranscripts was quantified in all samples as an internalcontrol to normalize the results (18).
CytogeneticsCells were harvested after colcemide exposure
(0.01 mg/mL) for 20 hours. Chromosomal slides wereprepared and stained according to standard methods.
Telomere-Targeting G4 Ligand Radiosensitizes GBM
www.aacrjournals.org Mol Cancer Ther; 10(10) October 2011 1785
on July 30, 2018. © 2011 American Association for Cancer Research. mct.aacrjournals.org Downloaded from

The frequency of chromosomal aberrations was deter-mined by the analysis of 100 metaphases.
Cell irradiationPhoton irradiations were conducted at room
temperature with a linear accelerator PHILIPS SL75-5 (in X-ray production mode) at a dose rate of3 Gy/min.
Clonogenic survival assayClonogenic cell survival was determined by a standard
colony-forming assay. Cells (1,500) were plated in cultureflasks and were allowed to attach for 24 hours. Thefollowing day, flasks were irradiated with 0 to 8 Gy.Cells were reincubated for 10 days, then fixed withmethanol, and stained with Giemsa. Colonies containingmore than 50 cells were scored. Surviving fractions ateach dose were calculated relative to the plating effi-ciency of nonirradiated control cells.
53BP1 immunofluorescence stainingCells were fixed with 4% paraformaldehyde and
permeabilized with 0.5% Triton X-100. For immunos-taining, cells were incubated with a mouse monoclonalanti-53BP1 (p53-binding protein 1) antibody (SantaCruz Biotechnology) overnight at 4�C. Cells were thenexposed to the secondary fluorescein isothiocyanate–labeled rabbit anti-mouse antibody (Santa Cruz Bio-technology) for 1 hour. The nuclei were stained by40,6-diamidino-2-phenylindole (DAPI). Fluorescencesignals were analyzed with a Zeiss confocal laser scan-ning microscope.
Determination of gH2AX phosphorylation by flowcytometry
The level of gH2AX phosphorylation was deter-mined by flow cytometry using H2AX PhosphorylationAssay Kit (Upstate) according to the manufacturer’sinstructions. Fluorescence intensity, in arbitrary units,was expressed relative to the nonirradiated control. Foreach experiment, all the compared samples were ana-lyzed the same day.
Statistical analysisStandard errors were calculated for each mean, and
statistical differences between groups were determinedby Student’s t test, Kruskal–Wallis H test, or ANOVAusing SEM software (19). Survival data were fittedaccording to the linear quadratic model using Kaleida-Graph software (4.0).
Results
Chemical properties of TAC and its interaction withquadruplex (telomeric) DNA
The major component (70%) of TAC (TAC-Me2)comprises 2 acridinium moieties joined by a heterotriarylspacer containing 2 pyridine rings and 1 pyrimidinering in alternation (abbreviated as py-pym-py; Fig. 1A).The minor component (TAC-Me3, 30%; SupplementaryFig. S4) has a thirdmethylated site that cannot be preciselyattributed (methylated pyridine or pyrimidine). Hence,throughout the present study, all discussion of the datarefers to the bis-methylated compound. Owing to thepropensity of the py-pym-py spacer to fold in a U-shaped
DC
10.80.60.40.20
poly(dA-dT)
poly(dC-dG)
ctDNA
ds26
poly(dC)
22CT
22AG
24G20
poly(dT)
poly(dA)
poly(rU)
poly(rA)
Relative binding
Nucle
ic a
cid
str
uctu
re Single strands
Double strands
i-motifs
G4
BA
N
NN
N
NN
0
5
10
15
20
543210
TAC concentration (µmol/L)
ΔTm
(ºC
)
Figure 1. Chemical structure,selectivity, and activity of theG4-interactive molecule TAC. A,structure of TAC-Me2 majorcomponent. B, speculativeU-shaped conformation ofTAC-Me2. C, nucleic acid bindingby TAC was tested by equilibriumdialysis on a variety of nucleic acidstructures (duplexes,quadruplexes, i-motif, and singlestrands). D, FRET meltingstabilization assays with the F21Ttelomeric sequence[50-fluorescein-(GGGTTA)3GGG-TAMRA] as a function of TACconcentration in sodium (opensquares) and potassium (filledcircles) conditions.
Merle et al.
Mol Cancer Ther; 10(10) October 2011 Molecular Cancer Therapeutics1786
on July 30, 2018. © 2011 American Association for Cancer Research. mct.aacrjournals.org Downloaded from

conformation, TAC-Me2 may possibly adopt a tweezer-type conformation inwater (Fig. 1B), similar to that observ-ed for the uncharged and monomethylated analogues inthe solid state and in organic solvents (20). Interestingly,tweezer-typemolecules holding 2 potentially intercalating
units 7 A�apart have a moderate affinity for duplex
DNA based on the neighbor exclusion principle (21, 22),which excludes bis-intercalation between contiguousbase pairs. In contrast, such compounds candisplay strongbinding affinity for non-B DNA architectures that canprovide less sterically constrained binding sites (23).In particular, G4 DNA can accommodate a large diversityof small molecules due to the presence of 3 loops and4 grooves in the vicinity of the guanines quartets.Although the first generations of G4 ligands were mainlyrestricted to planar aromatic molecules (24–26), severalrecent studies reported the design of second-generationquadruplex binders that exhibit a more complex 3-dimen-sional topology (27–31). For instance, molecular modelingstudies have suggested that ligands possessing 2 interca-lative aromatic units can establish a dual quartet–loopinteraction (32). It was thus of great interest to assessthe ability of TAC to bind the quadruplex form of humantelomeric DNA, as it is now suggested that this unusualDNA structure affects cancer cell viability. A monomericacridinium salt CH12-4 was prepared and used as control(Supplementary Fig. S5). This molecule has a planar con-formation and most likely behaves in the same manneras a classical duplex DNA intercalator.Two complementary methods were used to show that
TAC is a quadruplex ligand. Competitive dialysis experi-ments (Fig. 1D) indicate that this compound has a pref-erence for quadruplex structures. These experimentscompare the amount of dye bound to a variety of struc-tures (typically 12–20), each trapped in a small dialysischamber immerged in a large bath containing 1 mmol/LTAC. Binding to the 2 quadruplex samples [22AG and24G20, corresponding to the human telomeric motif[A(GGGTTA)3GGG] and T2G20T2 sequence, respectively]was stronger than that for duplexes and DNA andRNA single strands [poly(rA), poly(rU), poly(dT), andpoly(dU)] or i-motifs (formed with C-rich sequences).Binding to quadruplexes was not exclusive, as lower butsignificant binding was also observed for double strands.FRET melting experiments were carried out with a
fluorescently labeled oligonucleotide F21T mimicking3.5 repeats of the human telomeric motif fluorescein-d-(GGGTTA)3GGG-TAMRA. Adding increasing concen-trations of TAC led to an increase in the apparent meltingtemperature of F21T. Figure 1D shows that stabilizationreached þ20�C at 5 mmol/L ligand concentration in KCl(circles) but was systematically lower in NaCl (squares),as observed for many other ligands.Finally, TAC was active in the telomere repeat ampli-
fication protocol (TRAP) assay with an apparent IC50
value of 0.15 mmol/L (data not shown). However, aspreviously reported, this is the result of inhibitingthe amplification of G-rich sequences rather than true
telomerase inhibition (33). Under the conditions usedhere, the TRAP assay should be considered as a usefultechnique to confirm quadruplex recognition rather thanone to test its activity against telomerase.
Short-term treatment with G4 ligand TAC inhibitsGBM cell growth
SF763 and SF767 cell lines were maintained in culturefor 14 days in the presence of 0.5, 1.0, and 2.0 mmol/L ofTAC. This treatment inhibited the growth of GBM cells ina concentration-dependent way (ANOVA; P < 10�7;Fig. 2A), with a more marked effect on SF763 cells.Analysis of cellular viability indicated that the treatmentswere not toxic (<10% Trypan blue–positive cells).
In parallel, we tested the same concentrations of TACon control normal human fibroblasts. No changes inproliferation were observed (Supplementary Fig. S6),in line with previous studies with other G4 ligandsshowing that they did not affect the proliferation andviability of normal human telomerase-negative cells likefibroblasts (34) or astrocytes (13). However, the toxicityof TAC to telomerase-dependent normal human cells ashematopoietic cells or other rapidly proliferating cellsremains to be explored. In addition, the control com-pound CH12-4, a classical duplex DNA intercalator, didnot inhibit GBM cell proliferation (SupplementaryFig. S7). Taken together, these results indicate the speci-ficity of TAC effects on tumor cell proliferation.
We next analyzed the distribution of cells in differentphases of the cell cycle by flow cytometry (Fig. 2B). Ascompared with untreated controls, the distribution wasnot significantly modified in SF763 and SF767 cells trea-ted with TAC for 14 days, with only a moderate cellaccumulation in the G2–M phase.
To determine whether inhibition of tumor cell growthwas related to apoptosis induction, the fraction of earlyapoptotic cells was evaluated by flow cytometry after14 days of treatment (0.5–2 mmol/L). The apoptotic frac-tions were low (<10%) and closely similar in controls andin cells treated with 0.5 and 1 mmol/L TAC. In contrast,the early apoptotic fraction increased to 32% and 23%(Student’s t test; P < 10�3) in SF763 and SF767 cells treatedwith 2 mmol/L of TAC, respectively (Fig. 2C). There wasno significant increase in the fraction of late apoptoticand necrotic cells, which is in agreement with the lowpercentage of cells stained with Trypan blue. In addition,sub-G0–G1 analysis also detected a significant increasein apoptosis in both cell lines treated with 2 mmol/L TAC(Supplementary Fig. S8).
In the absence of significant changes in cell-cycle dis-tribution and apoptosis rate after treatment with low con-centration of TAC, the observed proliferation inhibitionmay be related to an increase in the cell-cycle duration.To address this point,weestimated the length of cell doubl-ing time in the exponential phase of cell growth, whichroughly equals the cell-cycle duration. This estimationwas done after 10 days of culturing, with an intermediatepassage of cultures after 5 days. The treatment with
Telomere-Targeting G4 Ligand Radiosensitizes GBM
www.aacrjournals.org Mol Cancer Ther; 10(10) October 2011 1787
on July 30, 2018. © 2011 American Association for Cancer Research. mct.aacrjournals.org Downloaded from

1 mmol/L TAC significantly increased the duration ofthe cell cycle from 23.2 � 1.3 to 45.4 � 2.4 hours in SF763cells (Student’s t test; P ¼ 9 � 10�4) and from 22.1 � 0.9to 34.0 � 0.9 hours in SF767 cells (P ¼ 7 � 10�4).
TAC triggers hTERT overexpression and DNAdamage but does not induce telomere shortening
SF763 and SF767 cell lines have relatively short telo-meres, with an average length for the telomeric restric-tion fragments of 4.4 and 3.3 kb, respectively. Thetreatment with 1 mmol/L TAC did not induce anysignificant reduction of total or 30-overhang telomericDNA, as radioactive signal corresponding to 30-telomericsingle-stranded DNA was roughly the same before andafter 14 days of treatment for similar DNA quantities(Supplementary Fig. S9).
The positive telomerase status of the GBM cell lineswas confirmed with qRT-PCR, which revealed hTERTexpression. We then evaluated expression levels of the
hTERT gene as well as TRF1 and TRF2 genes, whichencode the core members of the shelterin telomere pro-tection complex, in cells treated with TAC for 14 days(Fig. 3A). In comparison with untreated controls, theexpression of hTERT was greatly increased (3-fold forSF763 and 10-fold for SF767 cells) after TAC treatment(ANOVA, P ¼ 2.5 � 10�7 and P ¼ 1.7 � 10�7, respec-tively). The treatment also led to a 2-fold increase inTRF2 expression (SF763, P ¼ 6.6 � 10�4; SF767, P ¼4.4 � 10�5) whereas the expression of TRF1 gene wasdecreased (SF763, P ¼ 1.6 � 10�3; SF767, P ¼ 6.0 � 10�6).
To study the induction of DNA double-strand breaks(DSB), we counted the number of phosphorylated53BP1 foci by in situ immunofluorescence staining.G4 ligands have been reported to induce DNA damageafter only a few hours of treatment (34). Indeed, wefound that 24-hour treatment with TAC significantlyincreased the number of 53BP1 foci both in SF763cells exposed to 0.5 mmol/L TAC (P ¼ 4 � 10�4, H test),
0
2
4
6
8
10
12
1470
Days
Po
pu
lati
on
do
ub
lin
gs
NT0.5 µmol/L TAC1 µmol/L TAC2 µmol/L TAC
0
2
4
6
8
10
12
1470Days
Po
pu
lati
on
do
ub
lin
gs
NT0.5 µmol/L TAC1 µmol/L TAC2 µmol/L TAC
A
B
C
SF767SF763
0
10
20
30
40
% o
f ap
op
toti
c c
ells
*
0
10
20
30
40
2 µmol/L
TAC
1 µmol/L
TAC
0.5 µmol/L
TAC
NT 2 µmol/L
TAC
1 µmol/L
TAC
0.5 µmol/L
TAC
NT
% o
f ap
op
toti
c c
ells
*
0
10
20
30
40
50
60
70
80
G2–MSG
0–G
1
% o
f cells
NT
0.5 µmol/L TAC
1 µmol/L TAC
2 µmol/L TAC
0
10
20
30
40
50
60
70
80
G2–MSG
0–G
1
% o
f cells
NT
0.5 µmol/L TAC
1 µmol/L TAC
2 µmol/L TAC
Figure 2. Cellular effects of TAC inSF763 and SF767 cell linestreated continuously with TAC(0.5, 1, and 2 mmol/L) for 14 daysas compared with nontreated (NT)cells. Mean � SE values of 3independent experiments eachcarried out in triplicate are shown.A, proliferation assessment:population doubling of the celllines. TAC induced aconcentration-dependentproliferation inhibition in both celllines. B, cell-cycle analysis:distribution of cells in the differentcell-cycle phases. Cells exposedto TAC showed few modificationsin this distribution, with only amoderate accumulation of cells inG2–M phase. C, flow cytometricassessment of apoptosis:apoptotic cells were scored as thepercentage of Annexin V positive/PI negative cells. Cells treatedwith 2 mmol/L TAC, but not cellsexposed to a lower concentration,showed an increased apoptoticfraction. *, statistically significantdifference (P values are given inthe text).
Merle et al.
Mol Cancer Ther; 10(10) October 2011 Molecular Cancer Therapeutics1788
on July 30, 2018. © 2011 American Association for Cancer Research. mct.aacrjournals.org Downloaded from

1 mmol/L TAC (P ¼ 7 � 10�5), and 2 mmol/L TAC(P < 10�7) and in SF767 cells exposed to 1 mmol/L TAC(P ¼ 0.04) and 2 mmol/L TAC (P ¼ 9 � 10�6; Fig. 3Band C). This effect was more pronounced in SF763cells (3-fold increase at 2 mmol/L) than in SF767 cells(1.5-fold increase at 2 mmol/L).To assess the induction of chromosomal aberrations,
we evaluated untreated and treated (0.5 mmol/L TAC,14 days) cultures for the presence of dicentric chromo-somes. Average frequencies of dicentrics per cell were0.11 in SF763 and 0.06 in SF767 cells, as compared with0.02 and 0 for untreated controls, respectively. Thus, thetreatment induced dicentric chromosomes, which canresult from end-to-end chromosome fusions and repre-sent signatures of telomere dysfunction.These observations indicated that short-term TAC
treatment of GBM cells modified the level of hTERT,TRF1, and TRF2 gene expression without a visible effecton average telomere length. The treatment inducedconcentration-dependent DNA damage and dicentricchromosomes.
TAC enhances radiation-induced killing of GBMcellsThe intrinsic radiosensitivity of SF763 and SF767 cells
was first studied in triplicate experiments by clonogenic
survival assay after increasing doses of radiation(0–8 Gy). Both GBM lines were found to be radioresistant,as previously observed (35). The survival fraction at2 Gy was higher for SF763 cells (86%) than for SF767cells (65%), indicating an even stronger resistance ofSF763 cells to radiation.
Subapoptotic, nontoxic concentrations of TAC(0.5 and 1 mmol/L) were selected for combinationexperiments. Of note, these concentrations did notreduce plating efficiency (data not shown). SF763and SF767 cells were treated with TAC for 14 days,the time point where TAC had shown an appreciableantiproliferative effect in both cell lines. These cellswere then irradiated and clonogenic survival assaywas conducted. During incubation, TAC was presentin the culture medium. The comparison between sur-vival curves obtained after irradiation alone and afterthe combined treatment showed a strong concentra-tion-dependent enhancement of radiation sensitivityafter TAC treatment (Fig. 4; ANOVA, P < 10�7), show-ing a synergistic effect in both cell lines. Survival (S)data after irradiation dose (D) were fitted accordingto the linear quadratic model as follows: S(D)/S(0) ¼exp(�aD � bD2). The a and b parameters were usedto calculate the dose enhancement factor at 10% sur-vival (DEF10). The DEF10 values for 0.5 and 1 mmol/L
Figure 3. TAC effects at RNA andDNA levels. A, qRT-PCR resultsfor the expression of hTERT andtelomere-related genes TRF1 andTRF2. In both cell lines, thetreatment with TAC led to anincrease in the expression ofhTERT and TRF2 genes whereasthe expression of TRF1 wasdecreased after TAC treatment. Band C, DNA breaks in GBM cells,nontreated (NT) or treated withTAC, detected with 53BP1immunofluorescence.Immunofluorescence staining ofSF763 cells was carried out aftertreatment for 24 hours with TAC.Nuclei were stained with DAPI(blue) and 53BP1 (green). TACincreased the number of 53BP1foci per cell in a concentration-dependent manner (B). Thisincrease in the number of 53BP1foci per cell was significant inSF763 cells exposed to 0.5, 1, and2 mmol/L TAC and for SF767 cellsexposed to 1 and 2 mmol/L TAC(C). *, statistically significantdifference (P values are given inthe text).
B C
Nontreated
1 µmol/L TAC
0.5 µmol/L TAC
2 µmol/L TAC
0
100
200
300
400
500
TRF2TRF1hTERT
No
rmalized
mR
NA
exp
ressio
n
*
**
0
100
200
300
400
500
TRF2TRF1hTERT
No
rmalized
mR
NA
exp
ressio
n
NT
0.5 µmol/L TAC
1 µmol/L TAC
2 µmol/L TAC
NT
0.5 µmol/L TAC
1 µmol/L TAC
2 µmol/L TAC
*
*
*
SF767SF763A
0
1
2
3
4
5
Nu
mb
er
of
53B
P1
foci p
er
cell
**
0
1
2
3
4
5
2 µmol/L
TAC
1 µmol/L
TAC
0.5 µmol/L
TAC
NT
2 µmol/L
TAC
1 µmol/L
TAC
0.5 µmol/L
TAC
NT
Nu
mb
er
of
53B
P1
foci p
er
cell
**
*SF 763
SF 767
Telomere-Targeting G4 Ligand Radiosensitizes GBM
www.aacrjournals.org Mol Cancer Ther; 10(10) October 2011 1789
on July 30, 2018. © 2011 American Association for Cancer Research. mct.aacrjournals.org Downloaded from

TAC were 1.68 and 2.32 in SF763 cells and 1.96 and 2.92in SF767 cells, respectively.
TAC modifies the repair of DNA DSBs induced byradiation
To investigate possible radiosensitization mechan-isms of TAC, we assessed the induction and repair ofDSB after irradiation through the study of phosphory-lated 53BP1 and gH2AX foci that colocalize with DNADSB.
Despite extensive analysis (at least 100 cells), we didnot detect any consistent increase in DNA break induc-tion after combined TAC (0.5 mmol/L) and radiation(2 Gy) treatment by in situ immunofluorescence ana-lysis for 53BP1. Initial levels of foci at 0.5 hours afterirradiation were around 30 per cell. The repair wasapparently efficient after this dose, as the numbers ofresidual foci 24 hours following irradiation were verylow (<3 foci per cell).
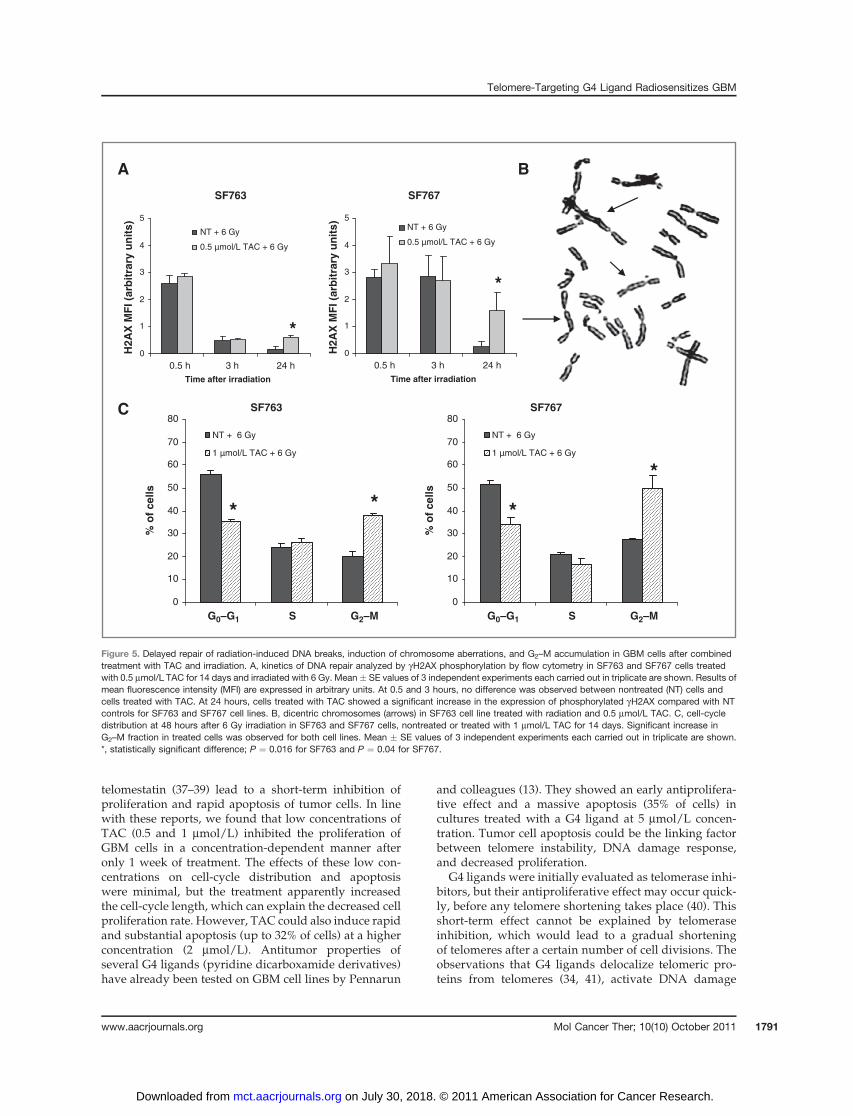
Because the radiosensitizing effect of TAC was stron-ger at higher radiation doses, we applied a differentapproach with flow cytometric analysis of phosphorylat-ed gH2AX (36), which enabled us to use the radiationdose of 6 Gy. We measured the initial peak at 0.5 hoursand signal reduction at 3 and 24 hours after irradiation.No significant difference was noted at 0.5 and 3 hoursbetween treated (0.5 mmol/L) and nontreated cells. Incontrast, a higher residual level in the presence ofTAC was observed at 24 hours (P ¼ 0.016 for SF763and P ¼ 0.04 for SF767; Fig. 5A), indicating a significantdeceleration of DNA repair.
These results suggest that TAC affected DNA repairafter irradiation. To further investigate this hypothesis,we analyzed the frequency of radiation-induced chromo-somal aberrations after irradiation with 4 Gy. The treat-ment with 0.5 mmol/L TAC for 14 days significantlyincreased the frequency of both chromosome-type andchromatid-type aberrations (Table 1). Of note, the highestincrease was observed in the frequency of dicentric chro-mosomes (Fig. 5B) after subtraction of the backgroundincidence of dicentrics induced by TAC alone.
In addition, we conducted cell-cycle analysis at48 hours after irradiation with 6 Gy in untreated cul-tures and in cultures treated with 1 mmol/L of TAC for14 days. Both GBM cell lines treated with TAC exhibiteda significant increase in percentage of cells in the G2–Mphase (P < 0.01), reflecting a more important radiation-induced G2-phase block (Fig. 5C).
To address the issue of apoptosis induction after TACand radiation, we conducted Annexin V/PI staining andflow cytometric analysis of cells harvested 3, 6, 24, 48, and72 hours after treatments with either radiation (6 Gy)alone or radiation (6 Gy) in combination with 1 mmol/LTAC. During this time course, both early and late apopto-sis/necrosis was slightly but not significantly inducedby the treatments (data not shown).
Discussion
TAC is a quadruplex ligand active on telomeric DNAin vitro. Telomere damage and DNA damage responseinduced by G4 ligands such as RHPS4, BRACO19, and
SF767SF763
0.01
0.1
1
10
100
86420
Radiation dose (Gy)
Su
rviv
al fr
acti
on
(%
)
NT
0.5 µmol/L TAC
1 µmol/L TAC
P < 10-7
Radiation dose (Gy)
0.01
0.1
1
10
100
86420
Su
rviv
al fr
acti
on
(%
)
NT
0.5 µmol/L TAC
1 µmol/L TAC
P < 10-7
βαNT 0.0170.034
0.5 µmol/L TAC 0.0240.216
1 µmol/L TAC 0.1010.043
βαNT 0.0230.166
0.5 µmol/L TAC 0.0500.464
1 µmol/L TAC 0.1580.578
S(D)/S(0) = exp (–αD – βD 2) S(D)/S(0) = exp (–αD – βD2)
Figure 4. TAC radiosensitizesGBM cells. Clonogenic survivalcurves of SF763 and SF767 cells,nontreated (NT) or treatedcontinuously for 14 days with0.5 and 1 mmol/L TAC, after X-rayirradiation with doses rangingfrom 0 to 8 Gy. Mean � SE valuesof 3 independent experiments,each carried out in triplicate areshown. Survival (S) data after aradiation dose (D) were fitaccording to the linear quadraticmodel. The linear parameter a andthe quadratic parameter b aregiven for each experimentalcondition. Radiation-inducedkilling of TAC-treated cells wassignificantly enhanced in aconcentration-dependent manner(ANOVA; P < 10�7).
Merle et al.
Mol Cancer Ther; 10(10) October 2011 Molecular Cancer Therapeutics1790
on July 30, 2018. © 2011 American Association for Cancer Research. mct.aacrjournals.org Downloaded from
telomestatin (37–39) lead to a short-term inhibition ofproliferation and rapid apoptosis of tumor cells. In linewith these reports, we found that low concentrations ofTAC (0.5 and 1 mmol/L) inhibited the proliferation ofGBM cells in a concentration-dependent manner afteronly 1 week of treatment. The effects of these low con-centrations on cell-cycle distribution and apoptosiswere minimal, but the treatment apparently increasedthe cell-cycle length, which can explain the decreased cellproliferation rate. However, TAC could also induce rapidand substantial apoptosis (up to 32% of cells) at a higherconcentration (2 mmol/L). Antitumor properties ofseveral G4 ligands (pyridine dicarboxamide derivatives)have already been tested on GBM cell lines by Pennarun
and colleagues (13). They showed an early antiprolifera-tive effect and a massive apoptosis (35% of cells) incultures treated with a G4 ligand at 5 mmol/L concen-tration. Tumor cell apoptosis could be the linking factorbetween telomere instability, DNA damage response,and decreased proliferation.
G4 ligands were initially evaluated as telomerase inhi-bitors, but their antiproliferative effect may occur quick-ly, before any telomere shortening takes place (40). Thisshort-term effect cannot be explained by telomeraseinhibition, which would lead to a gradual shorteningof telomeres after a certain number of cell divisions. Theobservations that G4 ligands delocalize telomeric pro-teins from telomeres (34, 41), activate DNA damage
BA
SF767SF763
0
1
2
3
4
5
24 h3 h0.5 h
Time after irradiation
H2
AX
MF
I (a
rbit
rary
un
its
)
NT + 6 Gy
0.5 µmol/L TAC + 6 Gy
*0
1
2
3
4
5
24 h3 h0.5 h
Time after irradiation
H2A
X M
FI (a
rbit
rary
un
its)
NT + 6 Gy
0.5 µmol/L TAC + 6 Gy
*
SF767SF763C
0
10
20
30
40
50
60
70
80
G2–MSG0–G1
% o
f c
ell
sNT + 6 Gy
1 µmol/L TAC + 6 Gy
*
*
0
10
20
30
40
50
60
70
80
G2–MSG0–G1
% o
f c
ell
s
NT + 6 Gy
1 µmol/L TAC + 6 Gy
**
Figure 5. Delayed repair of radiation-induced DNA breaks, induction of chromosome aberrations, and G2–M accumulation in GBM cells after combinedtreatment with TAC and irradiation. A, kinetics of DNA repair analyzed by gH2AX phosphorylation by flow cytometry in SF763 and SF767 cells treatedwith 0.5 mmol/L TAC for 14 days and irradiated with 6 Gy. Mean � SE values of 3 independent experiments each carried out in triplicate are shown. Results ofmean fluorescence intensity (MFI) are expressed in arbitrary units. At 0.5 and 3 hours, no difference was observed between nontreated (NT) cells andcells treated with TAC. At 24 hours, cells treated with TAC showed a significant increase in the expression of phosphorylated gH2AX compared with NTcontrols for SF763 and SF767 cell lines. B, dicentric chromosomes (arrows) in SF763 cell line treated with radiation and 0.5 mmol/L TAC. C, cell-cycledistribution at 48 hours after 6 Gy irradiation in SF763 and SF767 cells, nontreated or treated with 1 mmol/L TAC for 14 days. Significant increase inG2–M fraction in treated cells was observed for both cell lines. Mean � SE values of 3 independent experiments each carried out in triplicate are shown.*, statistically significant difference; P ¼ 0.016 for SF763 and P ¼ 0.04 for SF767.
Telomere-Targeting G4 Ligand Radiosensitizes GBM
www.aacrjournals.org Mol Cancer Ther; 10(10) October 2011 1791
on July 30, 2018. © 2011 American Association for Cancer Research. mct.aacrjournals.org Downloaded from

response at telomeres, and induce chromosomal end-to-end fusions (38) strongly suggest that their direct targetis the telomere and not the telomerase enzyme.
In agreement with these reports, we found that TACcaused DNA damage without significant telomere short-ening. This damage was, in particular, revealed by anincrease in chromosomal fusions in both GBM cell lines.Pennarun and colleagues (13) also found in GBM cellstreated with G4 ligands a very clear increase in telomericfusions in metaphase and anaphasic bridges, evidenceof telomeric instability not related to telomere attrition.
The chromosome end fusions are formed when telo-meres are dysfunctional and are recognized as DSBsby repair machinery, which joins chromosomal ends.Dysfunctional (uncapped) telomeres, resulting fromquadruplex DNA stabilization by G4 ligand, associatedirectly with many DNA damage response proteins.This DNA damage response may be evidenced by theappearance of phosphorylated 53BP1 or gH2AX foci incells exposed to G4 ligands, of which about 70% arelocated at telomeres (34). In the present study, we founda rapid and significant increase in the number of 53BP1foci after TAC treatment.
The structure and function of telomeres are controlledboth by telomerase and by the shelterin complex. Pre-vious studies showed that tumor cells overexpressinghTERT or TRF2, a shelterin complex component, becomeresistant to treatments with G4 ligands (34, 42). Thesechanges were considered to represent compensatorydefense mechanisms activated in response to G4 ligandtreatment, which is consistent with the view that telomereis a direct target of G4 ligands (34, 42). In our study, theeffect of TAC on telomeres was also evidenced by
changes in the expression of telomere-related genes intreated cells. In particular, we observed a strong increasein the expression of hTERT, which might counteractthe deleterious effects of the stabilization of G4 DNAby a ligand through the extension of the single-strandedG-rich telomeric portion, which is termed 30 overhang(42). We also found increased TRF2 expression, whichcould facilitate the formation of a loop structure (t-loop),masking the 30 overhang against G4 ligand binding,and/or the recruitment and stimulation of RecQ heli-cases, which resolve G4 DNA structures (32). In addition,we observed a concomitant decrease in the expression ofTRF1, which may contribute to hTERT-dependent telo-mere elongation. TRF1 controls the access of telomeraseto telomeric DNA, and its downregulation was shown toresult in the elongation of telomeres (43).
Thus, our results indicate that TAC targets telomeres,thereby increasing telomeric instability in tumor cells,and this inhibits tumor growth and, at higher doses, leadsto tumor cell apoptosis. As telomeric dysfunction wasreported to increase radiosensitivity and to decreaseDNA repair capacity (8), we then investigated the impactof telomeric instability induced by TAC on the radio-sensitization of GBM cells.
Applying low concentrations of TAC, which itself didnot induce apoptosis or cell-cycle changes, we found astrong concentration-dependent radiosensitizing effecton GBM cell lines within a wide dose range, includingthe dose corresponding to the daily therapeutic radiationdose (2 Gy) administered to GBM patients in clinicalpractice. This result is significant, because the adminis-tration of temozolomide, the new standard chemothera-py (1) given concomitantly with radiotherapy, is based on
Table 1. Chromosome aberration frequencies in GBM cell lines after irradiation and after combinedtreatment with radiation and G4 ligand TAC
Frequency of chromosomal aberrations
Cellsanalyzed
Acentricsper cell(95% CI)
Dicentricsper cell(95% CI)
Chromatidfragmentsper cell(95% CI)
Chromatidexchangesper cell(95% CI)
SF763 cellsIrradiated (4 Gy) 100 0.34 (0.21–0.47) 0.32 (0.19–0.44) 1.12 (0.89–1.34) 0.31 (0.19–0.43)Irradiated (4 Gy) and treatedwith 0.5 mmol/L TAC
100 0.85 (0.62–1.08) 1.02 (0.83–1.21)a 1.72 (1.45–1.99) 0.81 (0.58–1.04)
P (H test) 0.011 <10�6 0.0035 0.023SF767 cells
Irradiated (4 Gy) 100 0.46 (0.32–0.59) 0.38 (0.24–0.52) 0.86 (0.63–1.09) 0.24 (0.11–0.37)Irradiated (4 Gy) and treatedwith 0.5 mmol/L TAC
100 0.86 (0.64–1.07) 0.80 (0.62–0.98)a 1.42 (1.12–1.72) 0.58 (0.39–0.77)
P (H test) 0.043 0.00076 0.0093 0.0068
aThe background frequencies of dicentric chromosomes induced by TAC alone in nonirradiated cells (0.09 and 0.06 for SF763 andSF767 cells, respectively) were subtracted.
Merle et al.
Mol Cancer Ther; 10(10) October 2011 Molecular Cancer Therapeutics1792
on July 30, 2018. © 2011 American Association for Cancer Research. mct.aacrjournals.org Downloaded from

preclinical data (44, 45), which suggested that a combi-nation of temozolomide with radiation was essentiallyadditive.Radiosensitivity in an animal model with dysfunc-
tional telomeres correlated with a deceleration of DNArepair kinetics and persistence of chromosomal aberra-tions (8). We investigated whether TAC induced theseeffects in GBM cell lines. There is a correspondencebetween the formation and subsequent loss of phos-phorylated gH2AX or 53BP1 foci and the recognitionand subsequent repair of DNA damage. On the basis ofthis, MacPhail and colleagues (36) and Olive and Banath(46) proposed that the half-loss time of phosphoryl-ated gH2AX and the residual rate could be significantfactors in the evaluation of cell response to ionizingradiation, with more rapid loss and less retention asso-ciated with more radioresistant cell lines. In our study,analysis of DNA repair after irradiation by a similarapproach revealed delayed repair kinetics in GBMcells treated with TAC. The persistence of significantamounts of residual phosphorylated gH2AX was ob-served in both cell lines, indicating that the GBM cellshad become more sensitive to radiation.An increase in radiosensitivity was also correlatedwith
a higher frequency of radiation-induced chromosomalaberrations after combined treatment with radiation andTAC as compared with radiation alone. Dysfunctionaltelomeres, which act as DNA breaks, might join radia-tion-induced DNA DSB, thus increasing radiation sensi-tivity (47). As TAC induces telomere dysfunction, wesuggest that this mechanism was involved in the radio-sensitization of GBM cells observed after TAC treatment.Notably, the highest increase was seen in the frequency ofdicentric chromosomes, which could be due to DSB–telomere joinings.Cells exposed to ionizing radiation show delay
or arrest in the cell-cycle progression. In particular,G2-phase block prevents G2 cells with damaged DNAentering mitosis. The DNA damage can be reduced bymeans of DNA repair, but heavily damaged cells areunable to reach mitosis and are arrested in the G2 phase.G2–M accumulation after irradiation in GBM cells trea-ted with TAC compared with untreated cells may reflectthe presence of cell population with increased DNAdamage. This cell-cycle arrest can explain, at least inpart, the radiosensitizing effect of TAC at the cellularlevel.We did not observe any significant apoptosis induc-
tion up to 72 hours after treatment with radiation andTAC. This result may be explained by the fact that thecells were blocked in the G2–M phase after radiation,which was particularly marked after the combinationof radiation and TAC. This blockage delays celldeath, which could occur much later, after 1 or morecell divisions (48).Here, we showed efficacy of TAC combined with
radiation on GBM cells in vitro. Next step will be totest the radiosensitizing effect of intratumoral injections
of TAC in vivo, using subcutaneous human GBM xeno-grafts in animals. Because one of the major problems inbrain tumor therapy remains the difficulty of drugdelivery across the blood–brain barrier, it will also beessential to evaluate in an orthotopic model whetherTAC is capable of crossing the blood–brain barrier toreach and radiosensitize GBM tumors. Another possi-bility can be the direct injection of G4 ligand into thetumor-infiltrated brain parenchyma under positivepressure using convection-enhanced delivery. Con-versely to conventional diffusion, convection-enhanceddelivery creates a homogenous drug concentrationextending over several centimeters in diameter, allow-ing to target brain tumor tissue as well as the areasurrounding the tumor, which may contain tumor cells.This approach is successfully used to test novel antitu-mor agents in animal glioma models and in clinicaltrials with acceptable safety (49).
In conclusion, we showed that TAC, an original G4ligand, could enhance the sensitivity to ionizing radia-tion of 2 human hTERT-positive GBM cell lines. Apossible mechanism of radiosensitization is related todelayed DNA break repair after irradiation, whichincreases the number of chromosomal abnormalitiessuch as dicentrics, leading to cell death. Our workthus provides the first evidence that exposure to a G4ligand radiosensitizes radioresistant human GBM cells.Radiosensitizing effects will be investigated withother G4 ligands. Many elements concerning molecularmechanisms of altering tumor growth with TAC haveto be elucidated. In particular, it will be interestingto study the effects of TAC on G4 ligands locatedoutside telomeric regions, in promoters of humangenes, which are involved in cell proliferation (50).Further studies in animal models using human GBMtumor xenografts are necessary to explore the therapeu-tic potential of this radiosensitizing approach.
Disclosure of Potential Conflicts of Interest
No potential conflicts of interest were disclosed.
Acknowledgments
The authors thank M. Bombled and D. Monchaud for liquid chroma-tography/mass spectrometry analysis of chemical compounds, N. Saettelfor help in the drawings, and Y. Communal, R. Veyrat-Masson, and J.Chassagne for help in flow cytometric analysis. The authors are alsograteful to Aurore Gu�edin (U869).
Grant Support
This work was supported by grants from the R�egion Aquitaine, Fondationpour la Recherche M�edicale, and ANR (G4-TOOLBOX, ANR-Blan-09-355) to J.-L.Mergny.
The costs of publication of this article were defrayed in part by thepayment of page charges. This article must therefore be hereby markedadvertisement in accordance with 18 U.S.C. Section 1734 solely to indicate thisfact.
Received July 13, 2010; revised July 22, 2011; accepted August 1, 2011;published online October 10, 2011.
Telomere-Targeting G4 Ligand Radiosensitizes GBM
www.aacrjournals.org Mol Cancer Ther; 10(10) October 2011 1793
on July 30, 2018. © 2011 American Association for Cancer Research. mct.aacrjournals.org Downloaded from

References1. Stupp R, Mason WP, van den Bent MJ, Weller M, Fisher B, Taphoorn
MJ, et al. Radiotherapy plus concomitant and adjuvant temozolomidefor glioblastoma. N Engl J Med 2005;352:987–96.
2. Morin GB. The human telomere terminal transferase enzyme is aribonucleoprotein that synthesizes TTAGGG repeats. Cell 1989;59:521–9.
3. KimNW, PiatyszekMA, Prowse KR, Harley CB,West MD, Ho PL, et al.Specific association of human telomerase activity with immortal cellsand cancer. Science 1994;266:2011–5.
4. Langford LA, PiatyszekMA, Xu R, Schold SC Jr, Shay JW. Telomeraseactivity in human brain tumours. Lancet 1995;346:1267–8.
5. Tchirkov A, Rolhion C, Kemeny JL, Irthum B, Puget S, Khalil T, et al.Clinical implications of quantitative real-time RT-PCR analysis ofhTERT gene expression in human gliomas. Br J Cancer2003;88:516–20.
6. Lin PC, Lin SZ, Chen YL, Chang JS, Ho LI, Liu PY, et al. Butylide-nephthalide suppresses human telomerase reverse transcriptase(TERT) in human glioblastomas. Ann Surg Oncol 2011 May 7. [Epubahead of print].
7. Marian CO, Cho SK, McEllin BM, Maher EA, Hatanpaa KJ, MaddenCJ, et al. The telomerase antagonist, imetelstat, efficiently targetsglioblastoma tumor-initiating cells leading to decreased proliferationand tumor growth. Clin Cancer Res 2010;16:154–63.
8. Wong KK, Chang S, Weiler SR, Ganesan S, Chaudhuri J, Zhu C, et al.Telomere dysfunction impairs DNA repair and enhances sensitivity toionizing radiation. Nat Genet 2000;26:85–8.
9. Alberti P, Lacroix L, Guittat L, Helene C, Mergny JL. Nucleic acidsas targets for antitelomerase agents. Mini Rev Med Chem 2003;3:23–36.
10. Neidle S, Parkinson GN. The structure of telomeric DNA. Curr OpinStruct Biol 2003;13:275–83.
11. Granotier C, Pennarun G, Riou L, Hoffschir F, Gauthier LR, De Cian A,et al. Preferential binding of a G-quadruplex ligand to human chro-mosome ends. Nucleic Acids Res 2005;33:4182–90.
12. Kelland L. Targeting the limitless replicative potential of cancer: thetelomerase/telomere pathway. Clin Cancer Res 2007;13:4960–3.
13. PennarunG, Granotier C, Gauthier LR, Gomez D, Hoffschir F, MandineE, et al. Apoptosis related to telomere instability and cell cyclealterations in human glioma cells treated by new highly selectiveG-quadruplex ligands. Oncogene 2005;24:2917–28.
14. Rosu F, De Pauw E, Guittat L, Alberti P, Lacroix L, Mailliet P, et al.Selective interaction of ethidium derivatives with quadruplexes: anequilibrium dialysis and electrospray ionization mass spectrometryanalysis. Biochemistry 2003;42:10361–71.
15. De Cian A, Guittat L, Kaiser M, Sacca B, Amrane S, Bourdoncle A,et al. Fluorescence-based melting assays for studying quadruplexligands. Methods 2007;42:183–95.
16. Ohyashiki JH, Hisatomi H, Nagao K, Honda S, Takaku T, Zhang Y,et al. Quantitative relationship between functionally active telomeraseand major telomerase components (hTERT and hTR) in acute leukae-mia cells. Br J Cancer 2005;92:1942–7.
17. Lin X, Gu J, Lu C, Spitz MR, Wu X. Expression of telomere-associatedgenes as prognostic markers for overall survival in patients with non-small cell lung cancer. Clin Cancer Res 2006;12:5720–5.
18. Tchirkov A, Chaleteix C, Magnac C, Vasconcelos Y, Davi F, Michel A,et al. hTERT expression and prognosis in B-chronic lymphocyticleukemia. Ann Oncol 2004;15:1476–80.
19. Kwiatkowski F, Girard M, Hacene K, Berlie J. Sem: a suitable statis-tical software adaptated for research in oncology. Bull Cancer2000;87:715–21.
20. Petitjean A, Khoury RG, Kyritsakas N, Lehn JM. Dynamic devices.Shape switching and substrate binding in ion-controlled nanomecha-nical molecular tweezers. J Am Chem Soc 2004;126:6637–47.
21. Crothers DM. Calculation of binding isotherms for heterogenouspolymers. Biopolymers 1968;6:575–84.
22. McGhee JD, von Hippel PH. Theoretical aspects of DNA-proteininteractions: co-operative and non-co-operative binding of largeligands to a one-dimensional homogeneous lattice. J Mol Biol1974;86:469–89.
23. Bacolla A, Wells RD. Non-B DNA conformations as determinantsof mutagenesis and human disease. Mol Carcinog 2009;48:273–85.
24. Monchaud D, Teulade-Fichou MP. A hitchhiker's guide to G-quad-ruplex ligands. Org Biomol Chem 2008;6:627–36.
25. Huppert JL. Four-stranded nucleic acids: structure, function andtargeting of G-quadruplexes. Chem Soc Rev 2008;37:1375–84.
26. Neidle S. The structures of quadruplex nucleic acids and their drugcomplexes. Curr Opin Struct Biol 2009;19:239–50.
27. Carrasco C, Rosu F, Gabelica V, Houssier C, De Pauw E, Garbay-Jaureguiberry C, et al. Tight binding of the antitumor drug ditercali-nium to quadruplex DNA. Chembiochem 2002;3:1235–41.
28. Granzhan A, Monchaud D, Saettel N, Guedin A, Mergny JL, Teulade-Fichou MP. "One Ring to Bind Them All"-part II: identification ofpromising G-quadruplex ligands by screening of cyclophane-typemacrocycles. J Nucleic Acids 2010;2010:460561.
29. Kaiser M, De Cian A, Sainlos M, Renner C, Mergny JL, Teulade-Fichou MP. Neomycin-capped aromatic platforms: quadruplex DNArecognition and telomerase inhibition. Org Biomol Chem 2006;4:1049–57.
30. Teulade-Fichou MP, Carrasco C, Guittat L, Bailly C, Alberti P,Mergny JL, et al. Selective recognition of G-qQuadruplex telomericDNA by a bis(quinacridine) macrocycle. J Am Chem Soc 2003;125:4732–40.
31. Xu L, Zhang D, Huang J, DengM, ZhangM, Zhou X. High fluorescenceselectivity and visual detection of G-quadruplex structures by a noveldinuclear ruthenium complex. Chem Commun (Camb) 2010;46:743–5.
32. Gabelica V, Baker ES, Teulade-Fichou MP, De Pauw E, Bowers MT.Stabilization and structure of telomeric and c-myc region intramolec-ular G-quadruplexes: the role of central cations and small planarligands. J Am Chem Soc 2007;129:895–904.
33. De Cian A, Cristofari G, Reichenbach P, De Lemos E, Monchaud D,Teulade-Fichou MP, et al. Reevaluation of telomerase inhibition byquadruplex ligands and their mechanisms of action. Proc Natl AcadSci U S A 2007;104:17347–52.
34. Salvati E, Leonetti C, Rizzo A, ScarsellaM, MottoleseM, Galati R, et al.Telomere damage induced by the G-quadruplex ligand RHPS4 has anantitumor effect. J Clin Invest 2007;117:3236–47.
35. Chautard E, Loubeau G, Tchirkov A, Chassagne J, Vermot-DesrochesC, Morel L, et al. Akt signaling pathway: a target for radiosensitizinghuman malignant glioma. Neuro Oncol 2010;12:434–43.
36. MacPhail SH, Banath JP, Yu TY, Chu EH, Lambur H, Olive PL.Expression of phosphorylated histone H2AX in cultured cell linesfollowing exposure to X-rays. Int J Radiat Biol 2003;79:351–8.
37. Leonetti C, Amodei S, D’Angelo C, Rizzo A, Benassi B, Antonelli A,et al. Biological activity of the G-quadruplex ligand RHPS4 (3,11-difluoro-6,8,13-trimethyl-8H-quino[4,3,2-kl]acridinium methosulfate)is associated with telomere capping alteration. Mol Pharmacol2004;66:1138–46.
38. Incles CM, Schultes CM, Kempski H, Koehler H, Kelland LR, Neidle S.A G-quadruplex telomere targeting agent produces p16-associatedsenescence and chromosomal fusions in human prostate cancercells. Mol Cancer Ther 2004;3:1201–6.
39. Gomez D, Paterski R, Lemarteleur T, Shin-Ya K, Mergny JL, Riou JF.Interaction of telomestatin with the telomeric single-strand overhang.J Biol Chem 2004;279:41487–94.
40. De Cian A, Lacroix L, Douarre C, Temime-Smaali N, Trentesaux C,Riou JF, et al. Targeting telomeres and telomerase. Biochimie2008;90:131–55.
41. Gomez D, O’Donohue MF, Wenner T, Douarre C, Macadre J,Koebel P, et al. The G-quadruplex ligand telomestatin inhibitsPOT1 binding to telomeric sequences in vitro and induces GFP-POT1 dissociation from telomeres in human cells. Cancer Res2006;66:6908–12.
42. Gomez D, Aouali N, Renaud A, Douarre C, Shin-Ya K, Tazi J, et al.Resistance to senescence induction and telomere shortening by aG-quadruplex ligand inhibitor of telomerase. Cancer Res 2003;63:6149–53.
Merle et al.
Mol Cancer Ther; 10(10) October 2011 Molecular Cancer Therapeutics1794
on July 30, 2018. © 2011 American Association for Cancer Research. mct.aacrjournals.org Downloaded from

43. Smogorzewska A, van Steensel B, Bianchi A, Oelmann S, SchaeferMR, Schnapp G, et al. Control of human telomere length by TRF1 andTRF2. Mol Cell Biol 2000;20:1659–68.
44. Wedge SR, Porteous JK, Glaser MG, Marcus K, Newlands ES. In vitroevaluation of temozolomide combined with X-irradiation. AnticancerDrugs 1997;8:92–7.
45. van Rijn J, Heimans JJ, van den Berg J, van der Valk P, Slotman BJ.Survival of human glioma cells treated with various combination oftemozolomideandX-rays. Int JRadiatOncolBiolPhys2000;47:779–84.
46. Olive PL, Banath JP. Phosphorylation of histone H2AX as a measureof radiosensitivity. Int J Radiat Oncol Biol Phys 2004;58:331–5.
47. Genesca A, Martin M, Latre L, Soler D, Pampalona J, Tusell L.Telomere dysfunction: a new player in radiation sensitivity. Bioessays2006;28:1172–80.
48. Shinomiya N. New concepts in radiation-induced apoptosis: ‘premi-totic apoptosis’ and ‘postmitotic apoptosis’. J Cell Mol Med2001;5:240–53.
49. Lopez KA, Waziri AE, Canoll PD, Bruce JN. Convection-enhanceddelivery in the treatment of malignant glioma. Neurol Res2006;28:542–8.
50. Huppert JL, Balasubramanian S. G-quadruplexes in promotersthroughout the human genome. Nucleic Acids Res 2007;35:406–13.
Telomere-Targeting G4 Ligand Radiosensitizes GBM
www.aacrjournals.org Mol Cancer Ther; 10(10) October 2011 1795
on July 30, 2018. © 2011 American Association for Cancer Research. mct.aacrjournals.org Downloaded from

2011;10:1784-1795. Mol Cancer Ther Patrick Merle, Bertrand Evrard, Anne Petitjean, et al. Radiation-Induced Killing of Human Glioblastoma CellsTelomere Targeting with a New G4 Ligand Enhances
Updated version
http://mct.aacrjournals.org/content/10/10/1784
Access the most recent version of this article at:
Material
Supplementary
http://mct.aacrjournals.org/content/suppl/2011/10/05/10.10.1784.DC1
Access the most recent supplemental material at:
Cited articles
http://mct.aacrjournals.org/content/10/10/1784.full#ref-list-1
This article cites 49 articles, 11 of which you can access for free at:
E-mail alerts related to this article or journal.Sign up to receive free email-alerts
Subscriptions
Reprints and
To order reprints of this article or to subscribe to the journal, contact the AACR Publications Department at
Permissions
Rightslink site. Click on "Request Permissions" which will take you to the Copyright Clearance Center's (CCC)
.http://mct.aacrjournals.org/content/10/10/1784To request permission to re-use all or part of this article, use this link
on July 30, 2018. © 2011 American Association for Cancer Research. mct.aacrjournals.org Downloaded from