th nvrt f n plntll ntrbtn - university of kansas
TRANSCRIPT

THE UNIVERSITY OF KANSAS
PALEONTOLOGICAL CONTRIBUTIONS
October 1, 1969 Paper 44
TAXONOMIC AND ENVIRONMENTAL STUDY OFTWO DIBUNOPHYLLID CORAL SPECIES FROM
UPPER PENNSYLVANIAN ROCKS OF KANSAS
J. M. COCKE
Kansas Geological Survey
Published by permission of Director, Kansas Geological Survey, The University of Kansas.
ABSTRACT
Two externally similar species of Upper Pennsylvanian rugose corals Dibunophyllumparvum COCKE, n. sp., from the Wyandotte Formation, and D. perplexum COCKE, n. sp.,from the Amazonia Limestone, Lawrence Formation, were studied for 1) taxonomicdifferentiation and 2) environmental significance. An objective test of numerical datafrom each species was made using discriminant analysis. Variation in dissepiment typesand characters of the columella were most useful in specific assignment. On the basis of
rock type and fossil analyses, specimens of D. parvum are considered representative ofan in situ assemblage which grew in shallow, well-lighted marine waters with numerousother organisms. In contrast, D. perplexum shows evidence of being transported andsorted.
INTRODUCTIONSmall dibunophyllid corals are abundant and
well preserved in the Argentine Limestone Mem-ber, Wyandotte Limestone and the AmazoniaLimestone Member, Lawrence Formation inKansas (Fig. 1). Collections were selected fromthese two units for detailed taxonomic study.Two species, one from each formation, can bedistinguished that are externally similar in sizeand possess septa that rise above upper limits ofthe epitheca. Except for these characters theyresemble generally other Midcontinent dibuno-phyllids noted by JEFFORDS (1948a) or describedby NEWELL (1935), jEFFORDS (1948b), FRAUN-
FELTER (1965), and COCKE (1966). In the areastudied, dibunophyllids are most abundant inthin, interbedded calcareous shale and limestone;they are less common in massive limestone, andapparently do not occur in noncalcareous shaleand sandstone. In contrast, those from the LowerCarboniferous of Scotland (HILL, 1938) andSpain (DE GRooT, 1963) are most abundant inmassive limestone low in argillaceous material.
DISCUSSIONSeveral columella-bearing corals from Mid-
continent Pennsylvanian rocks have been referredto Axophyllum MILNE-EDWARDS & HAIME (1850),a genus which HILL (1938) placed in synonomywith Carcinophyllum THOMSON (1880). Theoriginal description of Axophyllum is brief andindicates only that the type specimen is simple,trochoid, and structurally similar to Lithostrotion.Type specimens of A. rude WHITE & ST. JOHN
have been sectioned by me. Unlike dibunophyl-lids, these possess lonsdaleoid dissepiments and acolumella derived by the elongation and compli-cation of the cardinal septum. Moreover, Axo-phyllum? spp. A, B, C, E, and F described byGIRTY (1915), "Axophyllum" sp. described byDuNcAN (1962) and ?Koninckocarinia spp. A,B, described by COCKE (1965) are morphologicallysimilar to A. rude.
That dibunophyllids commonly were referredby earlier workers to Axophyllum was recognizedby JEFFORDS (1948a), DUNCAN (1962), andCOCKE (1966). In 1875, WORTHEN placed in A.

2 The University of Kansas Paleontological Contributions—Paper 44
Member
Formation
_-_ -
Lawrence
Amazonia
Sb. D
0 Zcc
0<-'---- – Haskell
-_-_--L___L.....r... estphalia-7vv<
-0
- --_-_- — - "J -0^N
,--n ":•..-..", ..._ StrangerD0latan
---. Fm.
- ----\-\ -_7.-_-,--. -, -1'-` -1 ''''• Sout Bend
DStanton 0
- # Stoner Ls. oc0
--...-\ Captain Cr .
CD-1 -_=, ==_ _-.=..-/--)'-n,-..--- - Z Z;-\--,-• -\ Vilas Sh. _ <
--/-n ',"-N, Z< 2Spring Hill Plattsburg
-.....L.
Ls. _, 0
,nMerriam onner Spr. a__ Sb. D —
"" \^
--.-.Farley 0
- 1'.7* --/1 ''..\ /eN I
,--)--,-^N'l iL ..--,-\ ArgentineWyandotte
Ls. >-1--
Frisbie li
T--±----
Fm. 1. Stratigraphie column of Upper Pennsylvanian de-posits in Kansas showing distribution of small dibunophyl-lid corals. Number (1) indicates occurrence of Dibuno-phyllum pas-tom COCKE, n. sp., and (2) shows occurrenceof D. perplex:tin COCKE, n. sp. Asterisks indicate presence
of externally similar dibunophyllids.
rude two specimens which are externally similarto both species described here, but cited only theoriginal description of the species. Efforts tolocate WORTHEN'S specimens have proved futileand their assignment to Dibunophyllum is tenta-tive. WHITE (1884) illustrated the exterior of acoral similar to those studied here and referred itto A. rude. Subsequent workers (KEyEs, 1894;BEEDE, 1900) added to the species other smallcorals which are characterized by a columella andsepta that rise above the epitheca. One interiorillustrated by BEEDE from Missourian or Virgilianunits shows definite dibunophylloid characters.ROWLEY (1901) erected A.? alleni, which shows
external affinities to both Dibunophyllum parvumCOCKE, n. sp., and D. perplexum COCKE, n. sp.GIRTY (1915) placed similar specimens from Mis-sourian rocks in Lophophyllum alleni. In sum-mary, the concept of A. rude is changed to in-clude small dibunophyllids in which septa riseabove epithecal limits of the calices and to ex-clude forms similar to the type specimens of thenew species here described.
Other larger corals assigned to Axophyllumseemingly are related to ?Bothrophyllum kan-sasense (MILLER & GURLEY, 1893), originally as-signed to Cam pophyllum but changed to its pres-ent designation by EASTON (1944). On the basisof lithic and morphologic characteristics, it isevident that type specimens of ?B. kansasense arefrom the Westerville Limestone (Missourian) ofthe Kansas City area. The description and illus-trated exteriors of A. infundibulum WORTHEN
(1875) suggest affinities to ?B. kansasense.Polished interiors of A. infundibulum and ?Axo-phyllum sp. D, discussed by GIRTY (1915), dis-tinctly indicate relationship to ?B. kansasense.Because they lack cyathopsid septal thickening incardinal quadrants which typify Bothrophyllum,assignment of the corals remains uncertain.
SYSTEMATIC DESCRIPTIONSThe classification of rugose corals proposed
by HILL (1956) is adopted in this paper. Nochanges in terminology are proposed.
Family AULOPHYLLIDAE Dybowski, 1873
Subfamily AULOPHYLLINAE Dybowski, 1873
Genus DIBUNOPHYLLUM Thomson &Nicholson, 1876
TYPE SPECIES. — Clisiophyllum bipartitumM'CoY, 1849, Lower Carboniferous, British Isles.
DIAGNOSIS.—Small to large solitary and bud-ding corals that possess regular dissepimentariumand columella composed of median plate joinedto radiating lamellae comprise this genus. Minorsepta are degenerate to strongly developed. Cardi-nal fossulae are well developed in some species,but obscure or missing in others.
DiscussioN.—European Lower Carboniferouscorals typified by Dibunophyllum bipartitum(M'CoY, 1849) differ in several respects fromNorth American Upper Pennsylvanian forms as-
2
1

Cocke—Upper Pennsylvanian Dibunophyllid Corals 3
signed to the genus. Most importantly, the formerpossesses a distinct cardinal fossula, degenerateminor septa, and numerous major septa. Penn-sylvanian dibunophyllids in North America havefewer major septa, show little evidence of fos-sulae, and possess strongly developed minor septawhich commonly attain more than one-half thelength of major septa. Older forms are largerand many possess planar to peripherally concavetabellae in transverse section. Several genera havebeen erected to include forms with dibunophyl-loid characters. Arachnolasma GRABAU (1922),from the Lower Carboniferous of China, is similarto other solitary dibunophyllids but differs inhaving thinner axial structure with a thickenedmedian plate. Cortvenia SMITH & RYDER (1926),Lower Carboniferous, is a fasciculate dibuno-phyllid with numerous thin major septa and shortminor septa, dibunophylloid columella, and littleevidence of an inner wall. YABE (1950) erectedHeritschioides to include Lower Permian orUpper Pennsylvanian Corwenia-like fasciculatecorals which differ from Cortvenia in possessingdilated septa. In 1941, HERITSCH erected Aman-dophyllum for solitary Middle and Upper Car-boniferous corals which closely resemble Cor-wenia. Dibunophylloides FOMICHEV (1953), fromthe Middle Carboniferous, includes solitary toweakly fasciculate dibunophyllids. In addition,FOMICHEV assigned corals which differed fromDibunophylloides in possessing lonsdaleoid dis-sepiments to Sestrophyllum. In a comprehensivestudy of Spanish Carboniferous corals, DE GROOT
(1963) placed Amandophyllum, Heritschioides,Dibunophylloides, and Sestrophyllum in synon-omy with Cortvenia. Corals studied here closelyresemble Dibunophylloides in size, habit, numberof major septa and length of minor septa, butfurther study of dibunophyllids is necessary todetermine validity of the cited genera. However,septal dilation in corals examined in the presentstudy may indicate a relationship to species ofHeritschioides. However, complete or partial lackof columella in individuals of Dibunophyllumpart/urn COCKE, n. sp., may exclude this speciesfrom all recognized dibunophylloid genera.
DIBUNOPHYLLUM PARVUM Cocke, n. sp.
Figure 2,1-3
DIAGNosts.—Small, turbinate dibunophyllids inwhich septa generally rise above epitheca. Maxi-mum diameter and length are 9.5 and 12 mm.,
respectively. A few specimens have one to threesmall corallites in calices. Ephebic stages havewell-developed dibunophylloid columella and reg-ular dissepimentarium; major septa number 22to 24 and are slightly thickened in cardinal quad-rants. Innermost tabulae blend with more steeplyinclined tabellae of columella.
EXTERNAL CHARACTERISTICS.—This species Com-
prises small turbinate corals. Diameters rangefrom 2 to 11.5 mm. The epitheca is crossed byfine transverse growth lines and larger smoothwrinkles. Several corallites have three or fourrejuvenescent stages, and a number have asmany as three small corallite buds. Calices aredeep, with septa commonly rising as much as0.5 mm. above upper limit of the epitheca; aprominent, seemingly solid axial boss commonlyrises to a slightly lower position. The columellais bladelike, with the plane of its long axis ap-proximately bisecting the attachment area. Flat-tened triangular to quadrate attachment areasare slightly less than one-half the length of thecorallites and the maximum widths comprise one-third the diameter.
TRANSVERSE SECTION —The epitheca is thin,commonly with local internal ridges correspond-ing in position to peripheral ends of septa. Inephebic sections, major septa number 22 to 24;they are thin and crooked near the outer wall,increase gradually to maximum thickness nearthe inner wall, and thin into the tabularium. Themajor septal zone occupies two-thirds of corallitediameter; few major septa connect to radiatinglamellae of the columella. Minor septa are one-half to four-fifths length of major septa; theyresemble major septa but are slightly thinner. Aslight thickening of both major and minor septain the cardinal region can be noted in mostspecimens. No fossulae are present. Dissepi-mentaria comprise 3 to 5 thin, commonly com-plete dissepiments which are generally convexoutward; complete dissepiments which are planaror curve inward are common. Particularly inareas of increased septal thickening, dissepimentsare irregular, incomplete, or have both endsaffixed to a single septum. One-third to one-halfof the corallite diameter is occupied by the dis-sepimentarium. Innermost dissepiments arethickened locally to form an obscure inner wall.The tabularium occupies less than one-fourth ofthe diameter and is composed of thin tabulaewhich commonly are planar or slightly convex
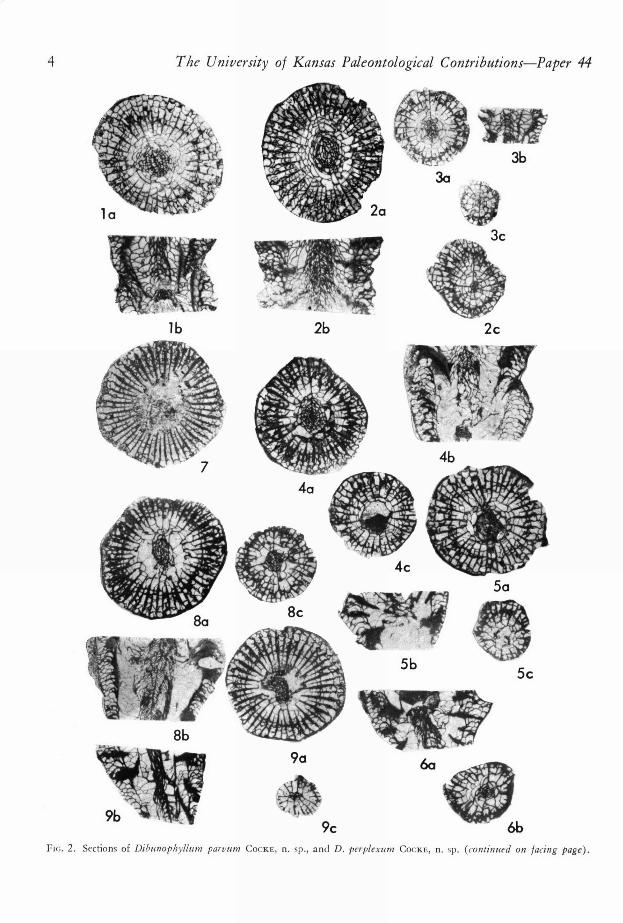
4 The University of Kansas Paleontological Contributions—Paper 44
nc. 2. Sections of Dibunophyllum parvum COCKE, n. sp., and D. perplexum COCKE, n. sp. (continued on facing page).

Cocke—Upper Pennsylvanian Dibunophyllid Corals 5
peripherally. A median plate, radiating lamellae,and tabellae comprise an oval to subcircularcolumella which comprises one-fifth to one-fourth of the corallite diameter. The median
plate is thick and usually rimmed by large tabel-
lae. Except in rare instances, the radiating lamel-
lae are short and are isolated among tabellae,neither connecting to major septa nor abuttingagainst the median plate. At diameters of 2.5 to30 mm., 12 to 14 major septa are present; theyare thin, slightly thickened in the cardinal quad-
rant, and extend into the tabularium. A cardinal-counter septum crosses the corallite and minor
septa are thin and equal to less than one-halfmajor septal length. The dissepimentarium is
composed of a row of peripherally convex plateswhich are structurally similar to both tabulae andtabellae. The central area is filled with globularplates which blend with the tabulae. No radiat-ing lamellae have been observed at this stage.
LONGITUDINAL SECTION.—A peripheral bandof three to six rows of globose dissepiments isterminated inwardly by an obscure inner wall.Short subhorizontal tabulae rim more steeplyinclined long plates which merge with the closelypacked, gently inclined tabellae of the axialstructure. A locally sinuous median lamella oc-cupies the central position.
Environmental AnalysisSAMPLING PROCEDURE.—The area studied Con-
sists of a small limestone hummock measuringapproximately 15 x 20 feet and rising 1 to 1.5
feet above a quarry floor. A total of 288 speci-mens were collected from the limestone and over-lying thin calcareous shale in the lower few feetof the Argentine Limestone. Originally, the pur-
pose of collecting these corals was to acquire asufficiently large number of adult forms fortaxonomic study. The corals were picked fromslightly weathered shale or chiseled from lime-stone. Later, bag samples of shale were taken toacquire juvenile forms. The shale was washed;juveniles of Dibunophyllum parvum were col-lected and other faunal elements were retainedfor generic identification.
LITHOLOGY.—The limestone is a fossiliferous
calcilutite formed by lithification of carbonatemud. Phylloid algae are moderately common inthe limestone and invertebrates are more com-
mon and diversified. The overlying shale containslittle silt and is highly calcareous. Above theshale, algal limestone is more common and con-tains abundant invertebrates.
FAUNAL ANALYSIS.—The preserved fauna thataccompanies the corals is diversified (Table 1).Brachiopods dominate the assemblage, with pro-ductids particularly abundant. Crinoid stemfragments also are common. Shapes of individuals
are highly variable, with planar, cylindrical,
conical, and irregular constituents present. Sizeranges vary from microscopic ostracodes to brach-iopods that attain maximum dimensions of 60
mm. Abrasion is slight. No organisms are re-
placed by silica or pyrite. Several fossils arebroken, but this feature was ignored because
(All sections x4, cardinal septum at top of transversesection.)1. Dibunophyllum parvum COCK E, n. sp., KUMIP no.
500515, Argentine Limestone Member, WyandotteFormation; la, transv. sec. slightly below base ofcalyx; lb, long. sec. perpendicular to counter-cardinalplane.
2. Dibunophyllum parvuM COCKS, n. sp., holotype,
KUMIP no. 500516; 2a, transv. sec, slightly belowbase of calyx; 2b, long. sec. perpendicular to counter-cardinal plane; 2c, transv. sec., late neanic stage.
3. Dibunophyllum parvum COCKE, n. sp., KUMIP no. 7.
500517; 3a, transv. sec, of small specimen slightly
below calyx; 3b, long. sec, perpendicular to counter- 8.
cardinal plane; 3e, transv. sec. of neanic stage.4. Dibunophyllum perplexum COCKE, n. sp., holotype,
KUMIP no. 500518; 4a, transv. sec. slightly belowbase of calyx, note dibunophyllid columella; 4b, long.
sec. perpendicular to counter-cardinal plane, showing 9.
lack of columella in lower part; 4e, transv. sec. ofearly ephcbic stage showing absence of columella.
5. Dibunophyllurn perplexum COCKE, n. sp., KUMIP no.
500519; 5a, transv. sec. in calyx showing absence ofcolumella; 5b, long. sec. (relation to counter-cardinalplane not known) showing dissepimentlike vesicles inupper part; Sc, transv. sec, showing neanic stage aboveattachment area.Dibunophyllum perplexum COCKS, n. sp., KUM1P no.500520; 6a, long. sec. perpendicular to counter-cardinalplane showing dibunophylloid columclla in lower partreplaced vertically by cystose structures; 6b, transv. sec.of early ephebic stage showing dibunophylloid colu-mella.Dibzinophyllum perplexunz COCKS, n. sp., KUMIP no.500521; transv. sec, of calyx.Dibunophyllum perplexum COCKE, n. sp., KUMIP no.500522 showing dibunophylloid columella in all sec-tions; 8a, transv. sec, slightly below calyx; 8b, long.sec, perpendicular to counter-cardinal plane; 8e, transv.sec. of early ephebic stage.Dibunophyllum perplexum COCKS, n. sp., KUMIP no.500523; 9a, transv. sec. in lower part of calyx; 9b,
long. sec. perpendicular to counter-cardinal plane; 9c,
transv. sec. of late ncanic stage.
6.
2 4 6 8 10 12
6
The University of Kansas Paleontological Contributions—Paper 44
TABLE 1. Faunal list of organisms collected withD. parvum Cocke, n. sp., from Wyandotte For-mation, Argentine Member, in SW SW sec. 25,
T. 12 S., R. 23 E., Johnson County, Kansas.
ALGAE
Archaeolithophyllum? sp.
Anchicodium? sp.
FORAMINIFERA
Fusulinids
Arenaceous forms
RUCOSA (corals)
Lophophyllidium (=Stereostylus) sp.Lophamplexus sp.
Dibunophyllum pareurn CocKE, n. sp.Geyerophyllum sp.
BRYOZOA
Rhomhopora sp.
Penestrellina sp.
Tabulipora sp.
BRACHIOPODA
Hustedia sp.
Corn posita spp.
Punctospirifer sp.
Neospnifer sp.
Lissochonetes sp.
Hystrictilina sp.
Reticulatia sp.
Cancrinella sp.
GASTROPODA
Anomphalus sp.
BIVALVIA
Myalina sp.
ECHINODERMATA
Crinoids, 3 genera
Crinoid stems, abundant
Echinoid plates and spinesARTIIROPODA
Trilobita
?Ditomopyge sp.
Ostracoda
Bairdia sp.
QUESTIONABLE FORMS
Encrusting ForaminiferaWorm borings
Worm tubes
breakage probably was due to quarry activity.All bivalves observed are articulated. The epi-fauna consists largely of encrusting bryozoanswhich are distributed evenly along the circum-ference of crinoid sterns and other moderate-sized organisms. On large specimens of thebrachiopod Neospirifer, bryozoans are presentonly on the brachial valve and along the marginsof both valves. Burrows were not observed; bor-ing occurs in a few crinoid stems and brachio-pods. The general fauna recorded was not studiedin detail. However, lack of abrasion, shape varia-
tion of constituents, wide size range, occurrenceof articulated shells, and concentration of epi-fauna on a single slide of large specimens stronglysuggest that the assemblage is essentially in situ.
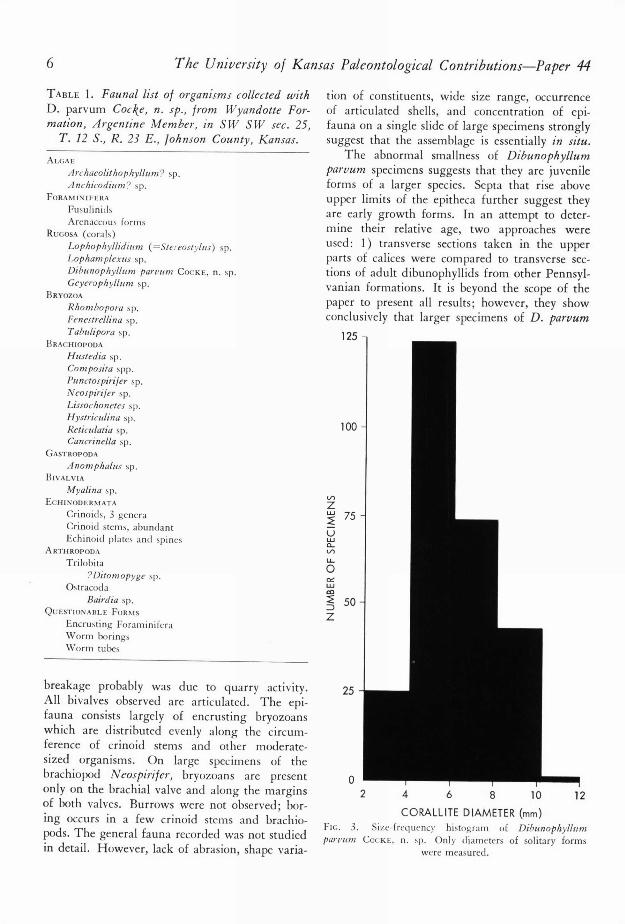
The abnormal smallness of Dibunophyllumparvum specimens suggests that they are juvenileforms of a larger species. Septa that rise aboveupper limits of the epitheca further suggest theyare early growth forms. In an attempt to deter-mine their relative age, two approaches wereused: 1) transverse sections taken in the upperparts of calices were compared to transverse sec-tions of adult dibunophyllids from other Pennsyl-vanian formations. It is beyond the scope of thepaper to present all results; however, they showconclusively that larger specimens of D. parvum
125 -
CORALLITE DIAMETER (mm)
FIG. 3. Size-frequency histogram of Dibunophyllumparent?: COCKE, n. sp. Only diameters of solitary forms
were measured.

Cocke—Upper Pennsylvanian Dibunophyllid Corals 7
are adults; 2) a larger dibunophyllid species froma different interval of the Wyandotte Formationwas sectioned at diameters within the upperrange of D. parvum diameters. Other dibuno-phyllid species from underlying and overlyingformations were similarly sectioned; all exhibitedjuvenile or early mature characteristics. Furtherevidence of adult forms is the occurrence ofbudding individuals. In rugose coral species thatare typically colonial, budding may occur invery early stages; however, dibunophyllids rarelybud and in all Pennsylvanian species known tome, budding occurs in late maturity or adultstages. Thus with growth stages of larger D.parvum reasonably established, the next problemto be solved is: Do these specimens represent ameaningful sample of a previously existing popu-lation or community?
Size-distribution analyses indicate small speci-mens of Dibunophyllum parvum are morenumerous (Fig. 3) than large ones, which yieldsa positively skewed diagram indicative of highjuvenile mortality (FAGERsTRom, 1964). ROWELL
(1960), and PERCIVAL (1944) believed that suchmortality rates are indicative of many marineorganisms. The extant coelenterate Hydra has asimilar curve (FoRREsT, 1963). The curve of D.parvum differs from the ideal curve in having norepresentatives in the 0 to 2 mm.-diameter classand relatively few in the 2 to 4 mm.-diameterclass. This apparent anomaly can be explained inat least three ways. First, most individuals sur-vived during 0-4 mm. growth in diameter, per-haps by very rapid early growth. Growth linesare faint below diameters of 2 mm., which per-haps indicates rapid growth, although othermechanisms are possible. Many specimens werefound in upright positions in the limestone, sug-gesting that they are preserved in growth posi-tion. In calcareous shales, corals were either up-right or lying subhorizontally.
Second, the condition may be due to samplingerror. This seems unlikely, because small speci-mens are easily detected in washed shale samples.Third, selective preburial and postburial destruc-tion of the small fraction may have occurred.Thus, rapid growth, selective removal of juve-niles, or a combination of these factors offer ade-quate explanation.
Many specimens of Dibunophyllum parvtimare found attached to skeletal material with agreat variety of sizes. This is accepted here as
suggesting lack of transportation by currents. Inspite of the dearth of knowledge of hydrody-namics of skeletal material, it seems unlikely thatspecimens with the observed diameter ranges ofD. parvum would be simultaneously transportedand deposited with crinoids and brachiopods thathave dimensions up to 60 mm. Coral specimens,as well as other faunal elements, are es9entiallyunabraded. Further, the restriction of these coralsto a single hummock, practically eliminates thepossibility that the forms were transported intothe area by currents.
In summary, evidence garnered from lithicand biotic relationships indicates that specimensof Dibunophyllum parvum are representative ofan in situ population or populations, which, be-cause of the small number of corals in the col-lected assemblage, suggests that the number ofgenerations was relatively small. Size distribu-tion, analysis of juvenile and adult sections, andpresence of budding individuals have been usedto establish that the assemblage represents 1) adiminutive species, not juveniles of a largerspecies, and 2) relatively few generations of thisspecies as shown by positively skewed size distri-butions. Each line of evidence is subjected tomore than one logical alternative and accordinglythe conclusions remain somewhat open to ques-tion. The highly variable accompanying fauna,as well as analogues with present-day coral oc-currences, suggests normal salinity. Depth waswell within the photic zone as shown by pro-liferation of algae. Turbulence was not great, forboth carbonate and aluminosilicate muds arecommon. That little transportation occurred issuggested by lack of abrasion and by poor sortingof fossil debris.
MATERIAL STUDIED.—Nearly 300 specimenswere collected from basal beds of the ArgentineLimestone in an active quarry in SW SW sec. 25,T. 12 S., R. 23 E., Johnson County, Kansas; a fewspecimens were collected from equivalent bedsexposed in Sunflower Quarry in SE NW sec. 28,T. 11 S., R. 23 E., Wyandotte County, Kansas.Approximately 100 transverse and 25 longitudinalsections were studied. All specimens are depositedin The University of Kansas Museum of Inverte-brate Paleontology. Holotype KUMIP no. 500516.
DIBUNOPHYLLUM PERPLEXUM Cocke, n. sp.
Figure 2, 4a-c; Sa-c; oui'; 7; Sa-c; 9a-c
DIAGNosts.—Small ceratoid corals that are

8 The University of Kansas Paleontological Contributions—Paper 44
generally solitary. A few individuals have twoor three buds. The epitheca is smooth andcrossed by small growth lines. Rejuvenescentforms are rare. Major septa number 22 to 26and are irregular near the periphery, thickenabruptly, and thin in the tabularium. Dissepi-ments are more highly varied in the cardinalquadrant than in others. The axial region ishighly variable and may be occupied by 1) atypical dibunophylloid columella, 2) widelyspaced irregular central tabulae rimmed bysteeply inclined clinotabellae, or 3) combinationsin single corallites of 1) and 2).
EXTERNAL CHARACTERISTICS. — Specimens aresmall, ranging from 8 to 11 mm. in diameterand 7 to 20 mm. in length. They are commonlyceratoid. Two orders of transverse elements, fi negrowth lines and larger, smooth wrinkles, arepresent. Few specimens show rejuvenescence orbudding. Calices are deep, with septa rising ashort distance above the epitheca. A bladelikecolumella extends to the upper limit of the calyxin some specimens. Attachment areas are triangu-lar and small, rarely attaining dimensions of2 mm.
Internal Description
TRANSVERSE SECTION.—The epitheca is thinbut thickens to form faint internal ridges atjunctions of septal ends. Typically, 22 to 26major septa cross two-thirds of the corallitediameter. A typical septal formula is C3A7K7A3(C, cardinal septum; A, alar septum; K, counter-septum). Few septa connect to radiating lamel-lae. Both orders of septa are thin and crookedperipherally; a few terminate against peripheraldissepiments. Septal thickness increases sharplya short distance from the epitheca and de-creases gradually in the tabularium. In a fewcorallites, small spines on both major and minorsepta simulate carinae. Minor septa are approxi-mately four-fifths of major septal length. Septaof cardinal quadrants are somewhat thicker thanthose of counter quadrants. A few "perforate"septa are formed by coincident thickening andpartial merging of septa and adjacent dissepi-ments. In the cardinal quadrants the dissepi-rnentarium is composed of 1) irregular completedissepiments that are peripherally convex, 2) in-complete dissepiments, and 3) dissepiments inwhich both ends terminate against a single
septum. Dissepiments are slightly thicker incardinal quadrants. Planar dissepiments arethinner near the epitheca. The dissepimentariumoccupies approximately two-fifths of the corallitediameter. Stereoplasmic thickening on innermostdissepiments creates an obscure inner wall.
The species is divided into two gradationalgroups: 1) individuals that possess a continuousdibunophylloid colurnella, and 2) others that haveno columella but possess widely spaced, irregulartabulae. Gradational forms are common. Inspecimens possessing a columella throughout, thetabularium consists of an outer zone of planarto curved tabulae which generally parallel thedissepiments and an inner area comprised ofplanar to curved tabellae inclined on the colu-mella. The median plate is thick and commonlyobscured by stereoplasmic deposits, and radiatinglamellae are either prominently or poorly de-veloped. Tabellae are thick and cystlike. Theyare largest adjacent to the median lamellae andconsiderably thickened at the outer edge of thecolumella. In specimens lacking a columella, thetabularium consists of two or three rows ofthick, axially convex to planar clinotabellae; com-monly the central area is either empty or tra-versed by a few tabulae.
Early ephebic stages of columellate forms show10 to 12 major septa that are slightly thicker incardinal quadrants than elsewhere. The cardinaland counter septa form a continuous bar which issomewhat thickened medially. Minor septa havelengths less than one-half those of major ones.A few dissepiments are slightly thickened andresemble the adjacent tabulae. The columella isformed by a thickened median portion of thecounter-cardinal cross bar, irregular tabellae, andmajor septal ends. Tabulae which surround theaxial area of noncolurnellate forms are convexaxially and somewhat thicker than normal.
LONGITUDINAL SECTION.—Peripherally, 2 to 8rows of globose dissepiments comprise the dis-sepimentarium; an obscure inner wall marks itsboundary with the tabularium. Steeply inclinedaxially convex tabulae are common at the outerlimit of tabularium and broad, distally curvedtabulae generally occupy its inner edge. Somespecimens have a combination of cystlike anasto-mosing plates and subhorizontal to faintly curvedtabellae. If present, the columella consists of asinuous median plate surrounded by steeply in-clined tabellae. In corals without a columella, the
2
4
6
16
18
20
Cocke-Upper Pennsylvanian Dibunophyllid Corals
9
'C,10-,:o>To'et±m8
_..1
22
24
26
28
30
.36 -.78
.71
.39 .54
.42
.42 .68 -.37-.43 -.44
-.50.40 .57 .61
22 24
Longitud
26
ina I Variables
28 30.40 .45 .36
.44
.38 .64-.54 -.63 .39 -.42
.35.53 .57 .35 .43
-.40 -.52 -.40-.35 -.37 .37 -.36
.49-.38 .52
2
3
4
5
7 8 9 10 11 12 13 14 15 16 17 18 19 20 21Transverse Variables
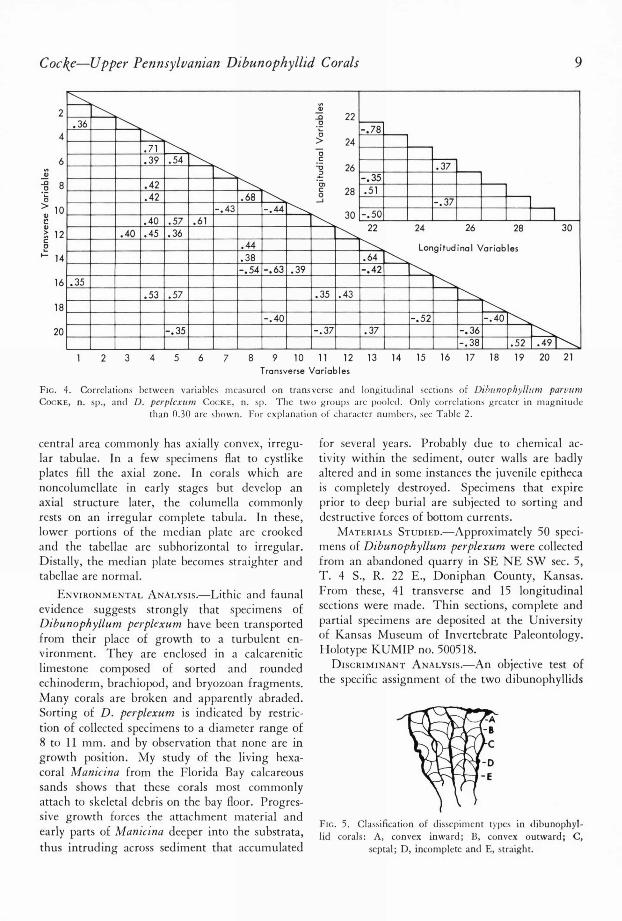
FIG. 4. Correlations between variables measured on transverse and longitudinal sections of Dibunophynum parcumCOCKE, n. sp., and D. perplexum COCKE, n. sp. The two groups are pooled. Only correlations greater in magnitude
than 0.30 are shown. For explanation of character numbers, see Table 2.
central area commonly has axially convex, irregu-lar tabulae. In a few specimens flat to cystlikeplates fill the axial zone. In corals which arenoncolumellate in early stages but develop anaxial structure later, the columella commonlyrests on an irregular complete tabula. In these,lower portions of the median plate are crookedand the tabellae are subhorizontal to irregular.Distally, the median plate becomes straighter andtabellae are normal.
ENVIRONMENTAL ANALYSIS.-Lithic and faunalevidence suggests strongly that specimens ofDibunophyllum perplexum have been transportedfrom their place of growth to a turbulent en-vironment. They are enclosed in a calcareniticlimestone composed of sorted and roundedechinoderm, brachiopod, and bryozoan fragments.Many corals are broken and apparently abraded.Sorting of D. perplexum is indicated by restric-tion of collected specimens to a diameter range of8 to 11 mm. and by observation that none are ingrowth position. My study of the living hexa-coral Manicina from the Florida Bay calcareoussands shows that these corals most commonlyattach to skeletal debris on the bay floor. Progres-sive growth forces the attachment material andearly parts of Manicina deeper into the substrata,thus intruding across sediment that accumulated
for several years. Probably due to chemical ac-tivity within the sediment, outer walls are badlyaltered and in some instances the juvenile epithecais completely destroyed. Specimens that expireprior to deep burial are subjected to sorting anddestructive forces of bottom currents.
MATERIALS STUDIED.-Approximately 50 speci-mens of Dibunophyllum perplexum were collectedfrom an abandoned quarry in SE NE SW sec. 5,T. 4 S., R. 22 E., Doniphan County, Kansas.From these, 41 transverse and 15 longitudinalsections were made. Thin sections, complete andpartial specimens are deposited at the Universityof Kansas Museum of Invertebrate Paleontology.Holotype KUMIP no. 500518.
DISCRIMINANT ANALYSIS.-An objective test ofthe specific assignment of the two dibunophyllids
FIG. 5. Classification of dissepiment types in dibunophyl-lid corals: A, convex inward; B, convex outward; C,
septal; D, incomplete and E, straight.

10 The University of Kansas Paleontological Contributions-Paper 44
was made using discriminant analysis. Twenty-one characters were measured on transverse sec-tions of the corals, as well as nine characters fromlongitudinal section (Fig. 4). Means of variablesare listed in Table 2. Some variables show weakto moderate intercorrelation, indicating some re-dundancy but the overall efficiency of the varia-bles selected for characterization is high. Dissepi-
ment types proved to be particularly meaningful(Fig. 5).
Separation of the two groups was tested usinga discriminant analysis program developed byWOLLEBEN, PAUKEN & DEARIEN (1967). Trans-verse and longitudinal sections proved to beequally effective in separating the two species, al-though transverse sections were used primarily inthe original qualitative definition of each species.MAHALANOBIS ' distance (D 2 ) between the twospecies in transverse was 77.5, yielding an F-value
of 5.9, with 21 and 6 degrees of freedom fortesting the significance of the discrimination. Theseparation produced by the discriminant functionis significant at the 95 percent level. Four mis-classifications were produced by the function, ofwhich three are misclassification of Dibunophyl-lum parvum as D. perplexum.
Longitudinal sections supplied effective vari-ables, although fewer were utilized for characteri-zation. MAHALANOBIS ' distance between Dibuno-phyllum perplexum and D. parvum was 4.7,
with an associated F-value of 3.1 and 9 to 22 de-grees of freedom. The discrimination is sig-nificant at the 95 percent level. Three misclas-sifications were produced, all D. parvum as D.perplexum.
REFERENCES
BEEDE, J. W., 1900, Carboniferous invertebrates: KansasGeol. Survey, v. 6, pt. 2, 187 p.
COCKE, J. M., 1965, Occurrence of a Carboniferous coralnot previously reported from North American rocks:The Compass, v. 42, no. 3, p. 180-188.
, 1966, New species of Dibunophyllum from theDewey Formation: Oklahoma Geol. Survey, Circular72, pt. 2, P. 35-56.
DE GROOT, G. E., 1963, Rugose corals from the Car-boniferous of northern Palencia (Spain): Leidse Geo-logische Mecledelingen, Dee! 29, 123 p.
DUNCAN, HELEN, 1962, Phylum Coelenterata: in MUDGE,M. R., & YOCHELSON, E. L., Stratigraphy and paleon-tology of the uppermost Pennsylvanian and lower-most Permian rocks in Kansas, U.S. Geol. Survey,Prof. Paper 323, P. 64-66.
TABLE 2. Means of Variables Measured inDibunophyllum perplexum and D. parvum.
TRANSVERSE S ECTIONS
D. per-plcrum
D.parvum
I. Average diameter 7.85 7.272. Wall thickness 0.10 0.093. Minor septal length 1.74 1.914. Maximum septal thickness
(cardinal quadrants) 0.24 0.175. Maximum sepia! thickness
(counter quadrants) 0.19 0.11Cardinal quadrants
6. Dissepiment A 5.85 3.737. Dissepirnent B 3.08 3.138. Dissepiment C 6.08 4.279. Dissepiment D 10.54 7.00
10. Dissepiment E 2.92 3.87Counter quadrants
11. Dissepiment A 8.08 4.4012. Dissepimcnt B 4.23 2.2313. Dissepiment C 2.08 2.7314. Dissepiment D 5.23 4.0715. Dissepiment E 2.92 4.6716. Major septal number 24.00 23.0017. Dissepiment thickness 0.06 0.0318. Tabulae thickness 0.03 0.0219. Tabellae thickness 0.03 0.0320. Tabellae along median lamellae 5.46 11.40
Longitudinal Sections21. Number radiating lamellae 5.46 7.6022. Number dissepiments
along wall 5.80 4.3823. Number dissepiments laterally 3.69 4.0724. Thickness of inner wall 0.07 0.0725. Number tabulae against
dissepimentarium 4.30 4.7126. Number tabulae against
columella 3.67 3.5027. Number tabellae against
median lamellae 6.07 7.1228. Dissepimental thickness 0.05 0.0329. Tabulae thickness 0.02 0.0230. Outer wall thickness 0.07 0.08
DYBOWSKI, W. N., 1873, Monographie der ZoanthariaSclerodermata Rugosa ans der Silurformation Estlands,Nord-Livlands und der Insel Gotland: Arch. Naturk.Liv-, Esth-u. Kurl. [1], v; Lief. 3, p. 257-414.
Ensrox, W. H., 1944, Revision of Campophyllum inNorth America: Jour. Paleontology, v. 18, p. 119-132.
FAGERST ROM, J. A., 1964, Fossil communities in paleo-ecology: their recognition and significance: Geol. Soc.America, Bull., v. 75, p. 1197-1216.
FoRREsT, H., 1963, Immortality and Pearl's survivorshipcurve for Hydra: Ecology, v. 44, p. 609-610.
FOMICHEV, V. D., 1953, Koralli Rugosa i stratigrafiyaSredne i verchnekamen- nouyolnich i Perm skich otlo-chenii Donetzkogo Bassuna [Rugose corals and strati-graphy of middle and upper Carboniferous and Per-mian deposits of the Donets basin]: Trudy Vses.

Cocke-Upper Pennsylvanian Dibunophyllid Corals
11
Nauch.-Issl. Geol. Inst. (VSEGEI) Min. Geol. Mos-cow, 1953, p. 1-622.
v FRAUNFELTER, G. H., 1965, A new Pennsylvanian dibuno-phyllid coral from Missouri: Missouri Geol. Survey,Rept. of Invest. no. 32, 8 p.
GIRTY, G. H., 1915, Invertebrate paleontology (of thePennsylvanian of Missouri): Missouri Bur. GeologyMines, Scr. 2, v. 13, p. 263-376.
GRABAU, G. W., 1922, Paleozoic corals of China. Part I,Tetraseptata: Paleont. Sinica (Geol. Surv. China) ser.B., y. 2, fasc. 1, 76 p.
HERITSCH, F., 1941, Polythecalis and Dibunophyllum ansdem Perm von Attika.: Sitz. ber. Akad. Wiss. Wien,Math.-naturw. KI., I, 150, p. 37-44.
Him., DOROTHY, 1938, The Carboniferous rugose coralsof Scotland: Palaeontograph. Soc. Mon. (London),213 p.
, 1956, Rugosa; in Coelenterata, pt. F of MOORE,
R. C. (ed.), Treatise on invertebrate paleontology:Geol. Soc. America and Univ. Kansas Press, p. 233-323.
N. JEFFORDS, R. M., 1948a, New Pennsylvanian dibunophyllidcorals (Oklahoma-Kansas): Jour. Paleontology, v. 22,p. 617-623.
, 1948b, The occurrence of corals in the latePaleozoic rocks of Kansas: Kansas Geol. Survey, Bull.76, pt. 3, p. 31-52.
K E Y ES, C. R., 1894, Paleontology of Missouri. Pt. 1: Mis-souri Geol. Survey, v. 4, 271 p.
M'Cov, F., 1849, On some new genera and species ofPaleozoic corals and Foraminifera: Ann. Mag. Nat.Hist., scr. 2, v. 3, p. 1-20, 119-136.
MILLER, S. A., & GURLEY, W. F. E., 1893, Descriptions ofsome new species of invertebrates from the Paleozoicrocks of Illinois and adjacent states: Illinois StateMuseum Nat. Hist., Bull. 3, 81 p.
MILNE-EDWARDS, H., & HAIME, J., 1850-54, British fossilcorals: London, Palaeontograph. Soc., Mon. Introduc-tion and pt. 1 (1850), p. i-lxxxv, 7-71; pt. 2 (1851),p. 73-145; pt. 3 (1852), p. 147-210; pt. 4 (1853), P.211-244; pt. 5 (1854), p. 245-299.
NEWELL, N. D., 1935, Some mid-Pennsylvanian inverte-brates from Kansas and Oklahoma; Pt. 2, Stromato-poroidea, Anthozoa, and Gastropoda: Jour. Paleon-tology, v. 9, no. 4, p. 341-355.
PERCIVAL, E., 1944, A contribution to the life-history ofthe brachiopod Terebratella inconspicua Sowerby: RoyalSoc. New Zealand Trans., v. 74, p. 1-23.
ROWELL, A. J., 1960, Some early stages in the develop-ment of the brachiopod Crania anomala (Muller):Annals Mag. Nat. History, ser. 13, v. 3, p. 35-52.
ROWLEY, R. R., 1901, Two new genera and some newspecies of fossils from the upper Paleozoic rocks ofMissouri: Am. Geologist, v. 27, p. 343-355.
SMITH, S., & RYDER, T. A., 1926, The genus Corwcnia,gen. nov.: Ann. Mag. Nat. Hist. (9), xvii, p. 149-159.
THOMSON, J., 1880, Contributions to our knowledge ofrugose corals from the Carboniferous limestone ofScotland: Phil. Soc. Glasgow, Proc., v. 12, p. 225-261.
, & NICHOLSON, H. A., 1876, Contributions to thestudy of the chief generic types of the Paleozoic corals:Ann. Map. Nat. Hist., Ser. 4, Vol. 17, no. I, P. 60-70;no. 2, p. 123128; no. 4, p. 290-305; no. 6, p. 451-461; Vol. 18, no. 7, P. 68-73.
WHITE, C. A., 1884, The fossils of the Indiana rocks, no.3: Indiana Dept. Geol. Nat. History, Ann. Rept. 13(1883), pt. 2, p. 107-180.
, & ST. JOHN, O. H., 1867, Description of newsub-Carbonifffous and coal measures fossils collectedupon the geological survey of Iowa: together with anotice of new generic characters observed in twospecies of brachiopods: Chicago Acad. Sciences, Trans.,v. I, p. 115-127.
WOLLFIBEN, J. A., PAUKEN, R. J., & DEARIN, J. A., 1967,FORTRAN II , program for multivariate paleontologicanalysis using an IBM 360-40 computer: in Computerprograms for multivariate analysis in geology, D. F.MERRIAM (Cd.), Kansas Geol. Survey ComputerContrib. 20, p. 1-12.
WORTHEN, A. H., 1875, Coal Measures: Illinois Geol.Survey, v. 6, 8 p.
YABE, H., 1950, Permian corals resembling Waagenophyl-lum, and Corwcnia: Proc. Japan Acad., v. 26, no. 3.