the action of sodium azide on cellular respiration and on...
TRANSCRIPT

165
546.331 . 7i8 : 577 . 158 . 047
The Action of Sodium Azide on Cellular Respiration and on Some Catalytic Oxidation Reactions
By D. K eilin , F.R.S., The Molteno Institute, University, Cambridge
( Received 18 A , 1936)
In troduction
Azoimide, HN3, was prepared by Curtius in 1890. Loew (1891) studied the effect of this acid and of its sodium salt on a great variety of organisms such as bacteria, moulds, algae, seedlings of plants, ciliates, worms, Crustacea, insect larvae, and vertebrates. He found it toxic to these organisms but, while some of them died within 2 or 3 hours in 0-5% solution of this salt, other organisms were killed only after 20-40 hours or even 4-6 days by 0-1% solution of NaN3, and ciliates withstood this concentration for even 12 days.
According to Loew, the toxic effect produced by this salt is due to its decomposition accompanied by the liberation of ammonia. This interpretation was not accepted by other workers, and it is now generally accepted that the poisonous effect produced by this substance is due to the
N\action of the nitrogen anion 11 N—.n /
We owe an important study of the physiological action of azoimide to Smith and Wolf (1904). According to these authors, azoimide in intravenous injection affects both nerves and muscles which are paralysed, after a preliminary stage of increased excitability. The vapour of azoimide inhaled causes excitation of the respiratory centres with subsequent paralysis. The acid was found by them to be the most powerful compound containing the trinitrogen group. The introduction of the phenyl radical diminished the effect of the complex. The physiological effect produced by azoimide on muscles or nerves re sembles, according to these authors, the action of hydrocyanic acid. Although in vivo they did not detect any appreciable effect of azoimide on haemoglobin, in vitro they obtained clear indications that it forms a compound with methaemo- globin but not with haemoglobin or oxyhaemoglobin.
Sodium azide was tried, so far, only as a physiological poison, and no attempt has been made to analyse its effect on cellular oxidation reactions.
In 1933 it was shown for the first time (Keilin, 1933, a) that sodium azide
VOL. CXXI.—B. (2 November, 1936) O
on June 2, 2018http://rspb.royalsocietypublishing.org/Downloaded from

166 D. Keilin
inhibits cellular respiration, the oxidation of cytochrome and the indo- phenol reaction in a manner similar to that of KCN. It was also shown (Keilin and Hartree, 1934) that sodium azide strongly inhibits the activity of catalase.
In the present paper it is proposed to give a complete account of the reaction between methaemoglobin and sodium azide, and this will be followed by the study of the action of NaN3 on cellular respiration and on
0*25 0*50 0-75 1-0 1*25 1*50 1*75 2*0NaN
FeFig. 1—The formation of NaN3-methaemoglobin, showing that the experimental
curve lies very near the theoretical curve of 1 molecule of NaN3 per iron atom of methaemoglobin.
some catalytic oxidation reactions. Throughout this study azoimide was used in the form of its sodium salt (NaN3), which is very stable and safe to handle.
A zide-M ethaemoglobin C ompound
The methaemoglobin used in these experiments was prepared from horse blood by the method previously described (Keilin and Hartree, 1935). The solution of this methaemoglobin acidified with KH2P04 to pu 5 *6-6 *4 is of a brown colour and shows the characteristic absorption
on June 2, 2018http://rspb.royalsocietypublishing.org/Downloaded from

Catalytic Oxidation Reactions 167
spectrum of acid methaemoglobin. On addition of NaN3 it rapidly turns red and its absorption spectrum is replaced by two strong but diffuse bands at 575 • 0 m[x and 542 • 5 m fx.
For the determination of the minimum amount of NaN3 which is required to convert acid methaemoglobin to NaN3-methaemoglobin, the same method was adopted as was previously described in the study of H2S-methaemoglobin (Keilin, 1933, b). Acid methaemoglobin and sodium azide are mixed in different concentrations in reaction vessels and the relative concentrations of free methaemoglobin and NaN3-methaemo- globin formed are determined by comparing the absorption spectrum of the mixture with that given by the two substances; acid methaemoglobin and azide-methaemoglobin in equal compartments of a double wedge trough.
T able IMethaemoglobin NaN3 NaNg/Fe % NaN3
as Fe, gm. atom// M. methaemoglobin2-2 x 10-4 0-38 x 10-4 0-18 202-2 x 10-4 0-55 X 10-4 0-25 272-2 x 10-4 0-83 x 10-4 0-38 332-2 x 10-4 11 x 10-4 0-50 472-2 x 10-4 1-38 X 10-4 0-63 572-2 x 10-4 1-65 x 10-4 0-75 682-2 x 10-4 1-94 x 10-4 0-88 762-2 x 10-4 2-20 x 10-4 1-00 852-2 x 10-4 2-47 x 10-4 M 2 892-2 X 10-4 2-75 x 10-4 1-25 952-2 x 10-4 3-20 x 10-4 1-45 98
The examination in this case is carried out by means of a Zeissspectroscope ocular attached to a microscope. The results of theseexperiments, which are shown in Table I and fig. 1, clearly demonstratethat the minimum amount of NaN3 required to convert acid methaemoglobin to azide-methaemoglobin corresponds to 1 molecule of NaN3 per iron atom of methaemoglobin.
The absorption spectrum of azide-methaemoglobin determined in the usual way by means of a Hilger Nutting spectrophotometer is shown infig. 2. The absorption coefficient is defined here as ̂ In j , where c
is the concentration of methaemoglobin in gram atoms of iron per cubic centimetre, d is the depth of the cell in centimetres, I0 and I are the intensities of incident and transmitted light which as log10 I0/I are read off directly on the instrument.
Methaemoglobin combines, therefore, with sodium azide in the same way as it does with KCN, NaF, H2S, H20 2, and C2H5OOH.
on June 2, 2018http://rspb.royalsocietypublishing.org/Downloaded from

168 D. Keilin
No compound has been obtained yet between azide and haematin or any other derivative of haemoglobin, apart from methaemoglobin. It may be interesting to note that among the above-mentioned six compounds, which combine with methaemoglobin, KCN alone also forms compounds with other derivatives of haemoglobin, such as haematin, parahaematin, haem, and haemochromogen.
—N aN* methb.
A cid —methb.j»H5-8
6500 AF ig. 2—Absorption spectrum of NaN3-methaemoglobin compared with that of acid
methaemoglobin. Ordinate, absorption coefficient per gram atom of methaemoglobin iron; abscissa, wave-length in A.
E ffect of A zide on the O xidation of C ysteine by H aematin
The action of sodium azide on the catalytic oxidation of cysteine by haematin compounds was tested by the oxygen uptake of 8 mg. of metal- free cysteine in presence of 1*6 X 10-4 M. haematin with and without the addition of 0-002 M. NaN3.
The oxygen uptakes in 1 hour at room temperature measured in Bar- croft differential manometers were as follows:—
c.mm.Cysteine alone .......................................................... 14Cysteine + haematin .............................................. 205Cysteine + haematin + NaN3 .............................. 162Cysteine + haematin + aa1 dipyridyl ..................... 74
on June 2, 2018http://rspb.royalsocietypublishing.org/Downloaded from

Catalytic Oxidation Reactions 169
This experiment, repeated several times with various haematin preparations, shows that the inhibition produced by NaN3 is incomplete; it varies with the haematin preparation from 20 to 50%. Considering that the haematin does not combine with azide, the question arises as to the mechanism of
, such inhibition. In our experience a haematin solution, when exposed to air, gradually undergoes some irreversible changes which are easily detected, especially when this haematin is transformed into haemo- chromogen. The absorption bands in this haemochromogen are moved towards the blue end of the spectrum and a new band appears at about 583 mg. This modification is accelerated and accompanied by a more or less complete destruction of the haematin when the latter is shaken in air with substances undergoing oxidation.
The iron which still may be combined with the remains of the haematin nucleus may yet have the same catalytic activity towards cysteine as haematin itself. This activity, however, can be inhibited, or completely abolished by the addition of substances which do not combine with the haematin but form catalytically less active compounds with this partly liberated iron. NaN3 is one of these substances, ocod-dipyridyl is another.* The latter combines neither with haematin nor with any other iron porphyrin compound but it forms a very stable and catalytically inactive compound with free divalent iron. This explains the strong inhibitory action (64%) which aod-dipyridyl produces on the catalytic oxidation of cysteine by haematin. In an experiment of long duration, two hours or more, most of the haematin is completely destroyed and the iron is almost completely recovered as iron-aod-dipyridyl compound.
NaN3 (0-004 M.) also inhibits by 50% the catalytic decomposition of H20 2 by haematin. KCN, on the other hand, has no effect, or if anything accelerates this reaction.
T he A ction of A zide on P eroxidase, C atalase, and O xidases
Peroxidase—Sodium azide in a concentration of 0-003 M. inhibits by 70% the activity of peroxidase prepared from horse-radish. The same peroxidase preparations are completely inhibited by small concentrations (0-0001 M.) of KCN or H 2S.
Catalase, on the other hand, prepared from horse liver, from blood, or from baker’s yeast, is very strongly inhibited by small concentrations of NaN3 (Keilin and Hartree, 1934). Liver catalase is completely inhibited by 0-002 M. azide, yeast catalase is inhibited by 94% in the
* This inhibitory effect of otod-dipyridyl was independently observed by Dr. M. Dixon.
on June 2, 2018http://rspb.royalsocietypublishing.org/Downloaded from

170 D. Keilin
presence of 0-001 M. azide at pH 6-5. This inhibition, as was shown recently by Blashko (1935), is completely reversible. The mechanism of this inhibition, as well as the action of azide on the haematin compound of the catalase preparation, will be discussed separately in a paper which follows.
Catechol oxidase prepared from cultivated mushrooms and tested with the oxidation of t//-dihydroxyphenylalanine was found not to be inhibited by 0-002 M. NaN3 at pn 7-3. The same concentration of azide at pn 5-9 inhibited the activity of the enzyme by 68%.
Indophenol oxidase of muscles (or cytochrome oxidase) is very strongly inhibited by NaN3. Thus, 0-001 M. solution of azide, as shown in Table II, inhibits the activity of this enzyme by 80-95%.
NaN3 inhibits also the oxidation of reduced cytochrome in living yeast cells, but in this case the inhibition depends upon the pH of the medium. Thus, in yeast cells suspended in phosphate buffer solutions of pn 8 to pn 7-5 the effect of azide is hardly perceptible. It begins to act only below 7 * 3 and in acid phosphate solution its inhibitory effect is as marked as that of KCN.
T able II—Effect of NaN3 on the oxygen uptake in 1 hour by />-phenylene- diamine in presence of indophenol oxidase of heart muscle, tested in Barcroft differential manometers, containing 3 cc. of solution per flask.
t Pu Oxidase P- NaN3 o 2 %c ° mg. wet
weightphenylene-
diamineM
M. uptake in c.mm.
inhibition
18 7-5 200 0-013 — 410 —
18 7-5 200 0-013 0 001 85 8021 6-5 200 0-013 — 102 — •
21 6-5 200 0013 0-001 21 8018 7-5 40 0-013 — . 68 —
18 7-5 40 0-013 0-001 3 95
A ction of NaN3 on R espiration of Y east Cells
The study of the effect of NaN3 on the respiration of yeast cells is carried out in the following way. Fresh samples of baker’s yeast are suspended and incubated for at least one hour in phosphate buffer solutions of various pH and containing 1% of glucose. After incubation, small samples of this suspension, containing from 10 to 20 mg. of wet weight of yeast cells, are put into the right-hand flasks of Barcroft differential manometers. The potash tubes of both flasks receive 0-3 cc.
on June 2, 2018http://rspb.royalsocietypublishing.org/Downloaded from
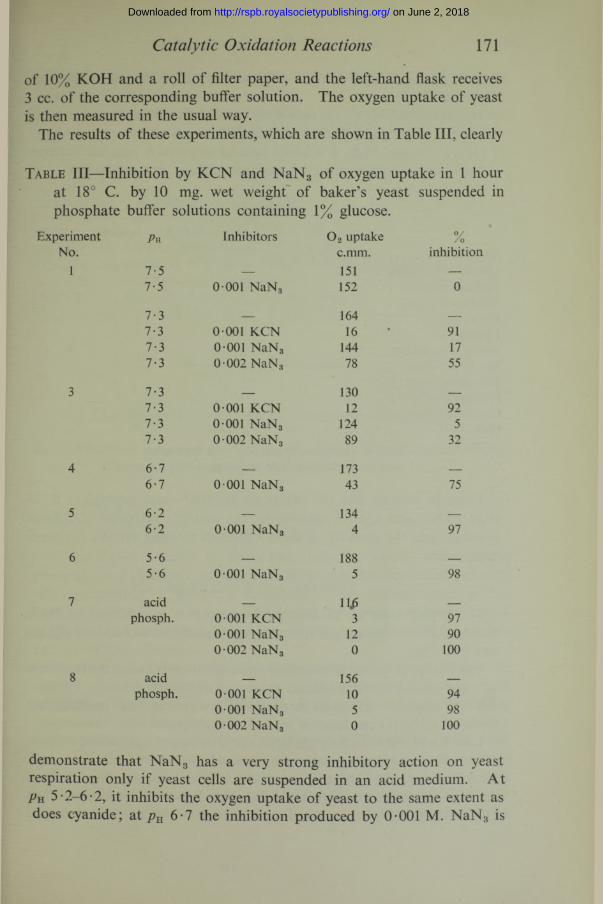
Catalytic Oxidation Reactions 171
of 10% KOH and a roll of filter paper, and the left-hand flask receives 3 cc. of the corresponding buffer solution. The oxygen uptake of yeast is then measured in the usual way.
The results of these experiments, which are shown in Table III, clearly
Table III—Inhibition by KCN and NaN3 of oxygen uptake in 1 hour at 18° C. by 10 mg. wet weight of baker’s yeast suspended in phosphate buffer solutions containing 1% glucose.
Experiment Pn Inhibitors 0 2 uptake %No. c.mm. inhibition
1 7-5 — 151 —
7-5 0-001 NaNg 152 0
7-3 ____ 164 —
7-3 0-001 KCN 16 917-3 0-001 NaN3 144 177-3 0-002 NaN3 78 55
3 7-3 ____ 130 —
7-3 0-001 KCN 12 927-3 0-001 NaN3 124 57-3 0-002 NaN3 89 32
4 6-7 — 173 —
6-7 0-001 NaN3 43 75
5 6-2 — 134 —
6-2 0-001 NaN3 4 97
6 5-6 — 188 —
5-6 0-001 NaN3 5 98
7 acid — m —
phosph. 0-001 KCN 3 970-001 NaN3 12 900-002 NaN3 0 100
8 acid ____ 156 ____
phosph. 0-001 KCN 10 940-001 NaN3 5 980-002 NaN3 0 100
demonstrate that NaN3 has a very strong inhibitory action on yeast respiration only if yeast cells are suspended in an acid medium. At Pu 5-2-6-2, it inhibits the oxygen uptake of yeast to the same extent as does cyanide; at pH 6-7 the inhibition produced by 0-001 M. NaN3 is
on June 2, 2018http://rspb.royalsocietypublishing.org/Downloaded from

only 75%; at pn 7 • 3, it is only 5-17%; and at 7 • 5 the respiration is not inhibited at all.
The action of NaN3 on respiration and different biological catalytic systems is very similar to that of cyanide.
Like KCN and H 2S, it inhibits cellular respiration and the activities of cytochrome oxidase, catechol oxidase, catalase, and peroxidase.
Table IV
172 Catalytic Oxidation Reactions
KCN h 2s NaN3 CORespiration................................. . . . . . . + + + +Cytochrome oxidase.................. .......... + 1 + +Catechol oxidase ..................... .......... + + + +Catalase ..................................... .......... + + + +Peroxidase ................................. + + —Uricase ..................................... .......... + — + —Amino acid oxidase ................. .......... — + — —Xanthine oxidase ....................... .......... + — — —
Methaemoglobin (Fe--).............. .......... + + + —Haemoglobin (Fe”) ................... .......... — — — +Haematin (Fe—) ..................... .......... + ? — —Haem (Fe--) ......................................... + — — +
+ signifies inhibition or reaction with the system.— signifies no inhibition or no reaction with the system.
Table IV summarizes the effects of the four main inhibitors; KCN, H2S, NaN3, and CO on respiration, the catalytic activities of different enzymes, and the capacity of these inhibitors to react with Fe porphyrin compounds.
Summary and Conclusions
The brown solution of acid methaemoglobin on addition of NaN3 turns red and its absorption spectrum is replaced by two bands at 575 mp. and 542-5 mp..
The change in colour and in absorption spectrum is due to the formation of a compound between sodium azide and methaemoglobin.
The formation of azide-methaemoglobin compound requires one molecule of NaN 3 per atom of iron of methaemoglobin.
Like KCN and H2S, NaN3 inhibits the activities of indophenol oxidase, of catechol oxidase, of catalase, and of peroxidase. Like KCN, it strongly inhibits the oxygen uptake by yeast cells, but this inhibition depends upon the pu of the medium. In acid solution at 5 • 2-6 • 7 the inhibitory effect produced by NaN3 is as marked as that produced by KCN; it decreases
on June 2, 2018http://rspb.royalsocietypublishing.org/Downloaded from

with increase in pn of the solution and at 7 • 5 the respiration of yeast cells is not inhibited at all.
The inhibitory effect produced by NaN3 on the catalytic oxidation of cysteine by haematin is probably due to inactivation of some iron derivative of haematin.
Properties o f Catalase Haematin 173
R eferences
Blashko (1935). ‘ Biochem, J.,’ vol. 29, p. 2303.Curtius (1890). ‘ Ber. deuts. chem. Ges.,’ vol. 23, p. 3023.Keilin, D. (1933, a). ‘ Proc. Roy. Soc.,’ B, vol. 113, p. 393.----- (1933, b). ‘ Ergebn. Enzymforsch.,’ vol. 2, p. 239.Keilin, D., and Hartree, E. F. (1934). ‘ Nature,’ vol. 134, p. 933.----- (1935). ‘ Proc. Roy. Soc.,’ B, vol. 117, p. 1.Loew (1891). ‘ Ber. deuts. chem. Ges.,’ vol. 24, p. 2947.Smith. L., and Wolf, C. G. L. (1904). ‘ J. Med. Res.,’ vol. 7, p. 451.
577 • 158.7 : 547 • 979On Some Properties of Catalase Haematin
ByD. K eilin , F.R.S., and E. F. FIartree, Ph.D., The Molteno Institute,University of Cambridge
{Received 18 thAugust, 1936)
Introduction
It was shown recently by Zeile and Hellstrom (1930) that a strong preparation of catalase obtained from horse liver has the distinct absorption spectrum of a haematin compound. The concentration of this haematin compound in various fractions of their preparations, estimated as pyridine haemochromogen, was found to be proportional to the enzyme activity of these fractions.
They have also shown that the factors which abolish or inhibit the activity of the enzyme, such as high temperature, alkali, acids, or the addition of small amounts of KCN or H2S, modify also the absorption spectrum of the haematin.
The haematin nucleus in the catalase preparation was found by them to be similar to that of haemoglobin in forming also a complex compound with a protein. The identity of the haematin nucleus in this compound with protohaematin obtained from haemoglobin was ultimately established by Stern (1936, a). According to Zeile and Hellstrom
on June 2, 2018http://rspb.royalsocietypublishing.org/Downloaded from